23 minute read
Running Away from Cancer: An investigation into the dynamic metabolism of cancer cells under an increase in extracellular lactate concentration


Running Away from Cancer: An investigation into the dynamic metabolism of cancer cells under an increase in extracellular lactate concentration
Sarah Arnold – Menai High School
Abstract
A distinctive hallmark of cancer cells is a high glucose uptake and lactate production regardless of oxygen availability, known as the Warburg Effect. Emerging studies suggest the Warburg Effect could be counteracted by increasing extracellular lactate concentration, which could occur during anaerobic exercise, however research is scarce. This experiment aimed to investigate how a cumulative increase in extracellular lactate affects the ability of a cancer cell line to switch from aerobic glycolysis to oxidative phosphorylation (OXPHOS) through measuring extracellular acidification rate (ECAR) and oxygen consumption rate (OCR) respectively, using advanced Seahorse XF technology. It was found when the total lactate injected into the cancerous cell line was 15mM and 20mM, there was a significant increase in OCR compared to basal measurements, where P=0.00301 and P=0.000686 respectively. This suggests that an increase in extracellular lactate does cause cancer cells to shift to an oxidative phenotype in vitro, however further investigations involving a larger sample size and in vivo models are pivotal in assessing the role of lactate, and potentially exercise, in the metabolic processes of cancer.
Literature Review
Cancer is the leading cause of death worldwide, responsible for approximately 10 million deaths in 2020 (WHO, 2021). Additionally, it is estimated that ¼ of adults worldwide do not achieve sufficient amounts of physical exercise (Mathewson, 2018). Alarming, emerging studies suggest an intertwine between these global issues, indicating that lactate resulting from single bouts of exercise may have a direct effect on tumour intrinsic factors (Dethlefsen, 2017, Hofmann, 2018).
Cancer cells: An unusual metabolism
Noncancerous cells rely primarily on oxidative phosphorylation (OXPHOS) to generate approximately 70% of their ATP for cellular processes. One fuel for OXPHOS is pyruvate, the end product of the enzymatic breakdown of glucose, known as glycolysis (Ristow, 2006). Under aerobic conditions, pyruvate is transported to the mitochondria where it is oxidised to acetyl-CoA. Acetyl-CoA is further combined with oxaloacetate to initiate the tricarboxylic acid (TCA) cycle, leading to OXPHOS and resulting in the synthesis of ATP energy (Zheng, 2012).
Discovered by Otto Warburg (Warburg et al., 1927), a hallmark of cancerous cells is an accelerated glycolytic metabolism to convert glucose to lactate rather than through OXPHOS, even under fully oxygenated conditions (San-Millán and Brooks 2017). This theory has been supported by various studies (Fadaka et al., 2017, Hirschhaeuser et al., 2011, Jose et al., 2010, Ruiz et al., 2009, SanMillán and Brooks, 2017, Zheng, 2012) and is known as the Warburg Effect or aerobic glycolysis. This phenomenon has remained a mystery across scientific literature, as glycolysis appears inefficient to cancer cells, yielding only 2 ATP molecules, compared to 40 ATP molecules generated by OXPHOS (Fadaka et al., 2017). Consequently, cancerous cells demand a high glucose consumption to maintain homeostasis (Hanahan & Weinberg, 2011).
Whilst previous literature accepts the Warburg Effect is a consequence of defects in cellular respiration, oncogenic alterations, and an overexpression of glycolytic enzymes and metabolite transporters (Hirschhaeuser et al., 2011), the underlying mechanisms of Warburg Effect in cancer cells has been unknown for nearly a century. This may be partially due to an unparalleled focus on genomic techniques in cancer research over the recent decades, which has resulted in a neglected understanding of cancer metabolics (Hofmann, 2018). However, it was recently proposed that the purpose of the Warburg Effect is solely lactate production, known as lactagenesis (SanMillán and Brooks, 2017), implying its role beyond a waste product. During lactagenesis, pyruvate is reduced into lactate, catalysed by the enzyme lactate dehydrogenase (LDH) (Xie et al., 2014), and this reaction is reversible (Mishra and Banerjee, 2019). However, limited studies have accounted for the reversible nature of this equilibrium reaction, in regards to the underlying mechanisms of the Warburg Effect.
Challenging the Warburg paradigm
Building on Warburg’s model, the Reverse Warburg Effect proposes that not all cancer cells undergo aerobic glycolysis, but rather lactate is shuffled and used as an energy source from cancer-associated fibroblasts (CAFS) via monocarboxylate transporters (MCTs) (Wilde et al., 2017). This is supported by the findings that tumours are not exclusively hypoxic, but rather contain aerobic regions which receive shuttled lactate from other glycolytic cancer cells (Semenza, 2008). The Reverse Warburg Effect induces localised lactic acidosis in the tumour microenvironment (TME) causing an accumulation of lactate (Siska, 2020) and due to the high ionisation of lactic acid, a consequent decrease in pH which may favour metastasis, angiogenesis and immunosuppression (de la Cruz- Lopez et al., 2019) and potentially chemoresistance (Brown et al., 2019). However, it is important to note that cancers are extremely heterogeneous with individual metabolic features (Zheng, 2012, Semenza, 2008), potentially limiting the application of Reverse Warburg Effect.
It has also been outlined that the transport of lactate into cancer cells through MCTs is dependent on a concentration gradient (Hofmann 2018, Payen et al., 2019) in order to avoid intracellular acidification (Brown et al., 2019). In 2018, Hofmann further hypothesised if the blood lactate concentration surrounding the tumour can be increased, for example through exercise, this could inhibit the shuttling of lactate and hence the process of the Reverse Warburg Effect. However, it was necessitated by Hofmann that there is extremely limited research on the mechanisms of single bouts of exercise on cancer.
Reverting the Warburg Effect?
The shift of energy metabolism from OXPHOS to aerobic glycolysis has now been widely accepted as a quintessential feature of cancer (Hanahan & Weinberg, 2011). In 2016, Wu inquired if cancer cells can revert from the Warburg effect to OXPHOS when induced by TME pressures. Wu’s results concluded without lactic acidosis, glycolysis and OXPHOS provided 23.7% - 52.2% and 47.8% - 76.3% of total ATP generated, respectively; whilst with lactic acidosis, glycolysis and OXPHOS provided 5.7%13.4% and 86.6% - 94.3% of total ATP generated respectively. This suggested lactic acidosis could revert cancer cells from the Warburg to the OXPHOS phenotype. Furthermore, it has been demonstrated, when 4T1 cancer cell lines were induced with lactic acidosis the cells showed a non-glycolytic phenotype characterised by a high oxygen consumption rate over glycolytic rate, negligible lactate production and efficient incorporation of glucose into cellular mass, revealing the dual metabolic nature of cancer cells (Xie et al., 2014).
However, Wu’s study induced the switch between glycolysis and OXPHOS using inhibitors (oligomycin, FCCP, Rotenone/ AntimycinA), and no research appears to stimulate this metabolic switch by increasing extracellular lactate. The study conducted by Wu in 2017 was also the first to quantitatively measure such a metabolic switch and only studied cell lines in two conditions (lactic acidosis and a control group). Therefore, such metabolic switch has not been investigated under varying concentrations of lactate as a means to model the accumulation of lactate as demonstrated in anaerobic exercise. This metabolic switch was also hypothesised by SanMillán and Brooks in 2017, who suggested that aerobic exercise could contribute to counteracting such switch to a glycolytic metabolism in cancer cells by creating epigenetic responses to restorate oxidative phenotypes, however, experimentations have been conducted.
It has been further suggested, if glycolysis could be inhibited in cancer cells, OXPHOS could be restored (Zheng, 2012). This is supported by the increasing number of studies that reveal lactate released by glycolysis and/or CAFS is not discharged as cellular waste, but rather is taken up by oxygenated tumour cells as energy fuel. It has been proposed that this occurs as lactate is converted to pyruvate by LHD where it enters the mitochondria and undergoes OXPHOS (Zheng, 2012). Limited studies however investigate the implications of adding the product, i.e. lactate, to this equilibrium as a potential mechanism for lactic acidosis causing the apparent shift from glycolysis to OXPHOS. However, it was noted that circulating lactate levels are critical in dictating the status of the LHD equilibrium, and Wu in 2014, concluded no net lactate generation was due to the equal rate of pyruvate generated from glycolysis with the removal into the TCA cycle (Xie et al., 2014). However, these concepts are mechanistic explanations of the Warburg Effect, which have yet to be justified by experiment.
A potential model for the effects of exercise on cancer
Despite a history of lactate being defined as only a waste product, over the last decade research has revealed lactate produced by exercise is an active metabolite, moving between cells and capable of being oxidised as a fuel (Philp, et al., 2005). Currently, there is limited research on the intrinsic effects of exercise on cancer cell metabolism. Exercise unquestionably has a role in regulating metabolic processes, but how this consequently affects tumour growth and metastatic rate is not currently mechanistically understood (Hojman, 2018). A study by McTiernan suggested physical activity may be linked to cancer production through exercise-dependent reductions in cancer risk factors; including sex hormones, insulin growth factor (IGF), inflammatory markers and improving immune function. The large inconsistencies regarding the exercise dose which is ideal for cancer management in current research with reference to intensity, duration, frequency and type hinders compelling conclusions (Ashcraft, 2016).
Therefore, cancer cell research demands quantitative over qualitative studies to meet the increasing prevalence of disease. Lactate holds promise to play a role in cellular properties beyond purely a waste product (San-Millan, 2017). Therefore, investigations in the role of lactate on cancer cells in vitro, could provide insight into a further understanding of the potential benefits, or risks, of exercising for cancer patients.
Scientific Research Question:
Will increased extracellular lactate concentrations in a cancerous cell line cause a change in the extracellular acidification rate (ECAR) and oxygen consumption rate (OCR)?
Scientific Hypothesis:
The cancerous cell line exposed to increased lactate concentrations is expected to increase in OCR and decrease in ECAR. This is supported by studies which propose that lactic acidosis can revert glycolytic cancer cells to a dominant oxidative phenotype (Dethlefsen, 2017, Hofmann, 2018, Nijsten & van Dam, 2009, San-Millán & Brooks, 2017, Wu et al., 2016).
Methodology
The investigation consisted of measuring extracellular acidification rate (ECAR) and oxygen consumption rate (OCR) in a cancerous cell line 4T1 (ATCC) at differing concentrations of sodium lactate using the novel Seahorse Extracellular Flux 24 (XF) analyser (Seahorse Bioscience). Sodium lactate was injected to the medium at cumulative increments from 0mM to 20mM (0mM, 5mM, 10mM, 15mM, 20mM) through integrated injection ports. In the control sample, a buffered serum, without lactate, was injected at the same concentrations and increments as the lactate group. This technology was specifically selected due to its dual ability to provide an indication of glycolysis and OXPHOS by measuring ECAR and OCR respectively, through real time measurements of changing pH and oxygen concentrations in the extracellular medium.
Glycolysis will be determined through measurements of the ECAR of the surrounding cell media, caused by the excretion of lactate per unit of time after the conversion from pyruvate in cells, altering pH (Wu et al., 2006). However since the lactate injected into the cell line also has acidic properties, this will inevitably reduce the validity of glycolysis measurements by altering pH levels which may not reflect the metabolic measurements of the cells. Therefore this justifies the selection of the Seahorse technology which has the ability to measure oxygen consumption rate, which is calculated by oxygen concentration (Plitzko et al., 2017) and therefore will not be influenced by lactate injections.
Prior to the assay, the cells were stored in a biosafety cabinet II at 37oC and 5% CO2 to effectively replicate conditions of the human body. Both cell lines were cultured in a low glucose Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal bovine serum (FBS) as well as other nutrients to control cell growth. 10 wells of the Seahorse XF 24 well microplate were each filled with approximately 10 000 cells of the cancerous 4T1 cell line and 10 remaining wells were likewise filled and allocated as the control. 4 wells were left blank as a control for the machine and the cells were left in the biosafety cabinet (37oC, 5% CO2) overnight. Prior to the assay the next morning, the cancerous cells were examined and observed under the microscope to monitor any abnormalities (Appendices 1).
The microplate was placed into the Seahorse XF and 3 basal measurements of ECAR and OCR were recorded with 0mM lactate. The assay then recorded 15 measurements for each of the 20 wells, recording ECAR and OCR rates at each time increment. Between each measurement the cells were mixed during a 5 minute cycle. Lactate concentration was cumulatively increased and 3 measurements were recorded for each concentration increment in each well. The same procedure was completed for the control, with the buffer solution injected.
Ethical and biosafety considerations have been addressed by the completion of the experiment to be conducted at Garvan Institute of Medical Research, where the hazardous and cancerous cell lines were stored in a Biosafety cabinet II and under appropriate conditions and protocol.
Results and analysis:
Null hypothesis:
As extracellular lactate cumulatively increases, the rates of extracellular acidification rate and oxygen consumption rate will not change as extracellular lactate concentration has no influence on cancer cell metabolism.
Alternative hypothesis H1:
As extracellular lactate cumulatively increases, the rates of extracellular acidification rate and oxygen consumption rate will change as extracellular lactate concentration has an influence on cancer cell metabolism.
Inferential statistical analysis:
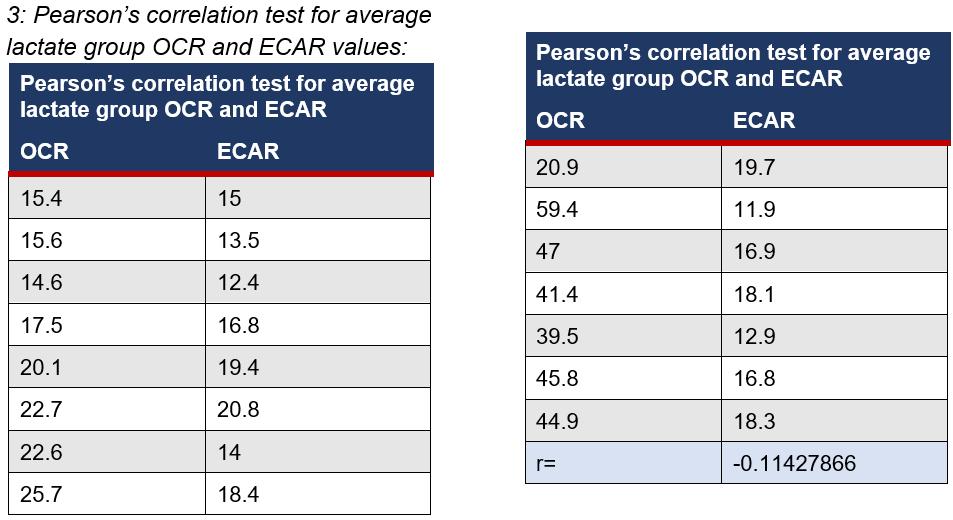
Unpaired, student t-tests were applied to determine if there was a statistical significance for each increment of lactate (5mM, 10mM, 15mM, 20mM) compared to basal measurements for both ECAR and OCR rates. The alpha value was set to p=0.05, meaning any values that were more than 5% due to chance, were deemed insignificant. The same t-tests were conducted for the control with the addition of a buffered solution. All p values for the control were insignificant (p > 0.05) and ranged from p=0.98 to p=0.40. Significant differences were measured at 15mM (Figure 1A) and 20mM for OCR (Figure 1B) and at 5mM (Figure 2A), 10mM (Figure 2B), and 20mM (Figure 2C) for ECAR.
Statistical analysis of lactate group for OCR measurements:
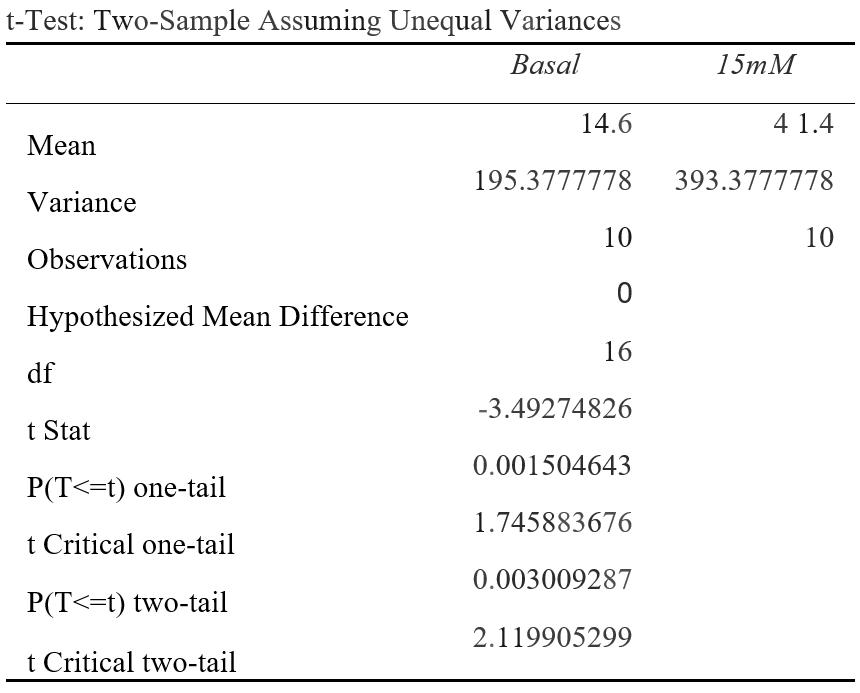
Figure 1A: P(T<=t) two-tail = 0.00301 (P<0.05) OCR (pMoles/min) was measured at a basal rate (13mins) and when 15mM was injected to the cell line (74mins). The difference between the average OCR for cancer cells at basal rates compared to 15mM were statistically significant as the results were 0.0301% due to chance. This suggests that 15mM of lactate could affect OCR.

Figure 1B: P(T<=t) two-tail = 0.000686 (P<0.05) OCR (pMoles/min) was measured at a basal rate (13mins) and when 20mM was injected to the cell line (94mins). The difference between the average OCR for cancer cells at basal rates compared to 20mM were statistically significant as the results were 0.00686% due to chance. This suggests that 20mM of lactate could affect OCR.
Statistical analysis of lactate group for ECAR measurements:

Figure 2A: P(T<=t) two-tail = 0.0128 (P<0.05) ECAR (mpH/min) was measured at a basal rate (13mins) and when 5mM was injected to the cell line (33mins). The difference between the average ECARfor cancer cells at basal rates compared to 5mM were statistically significant as the results were 1.28% due to chance. This suggests that 5mM of lactate could affect ECAR.

Figure 2B: P(T<=t) two-tail= 0.0294 (P<0.05) ECAR (mpH/min) was measured at a basal rate (13mins) and when 1 0mM was injected to the cell line (54mins). The difference between the average ECAR for cancer cells at basal rates compared to 10mM were statistically significant as the results were 2.94% due to chance. This suggests that 10mM of lactate could affect ECAR.
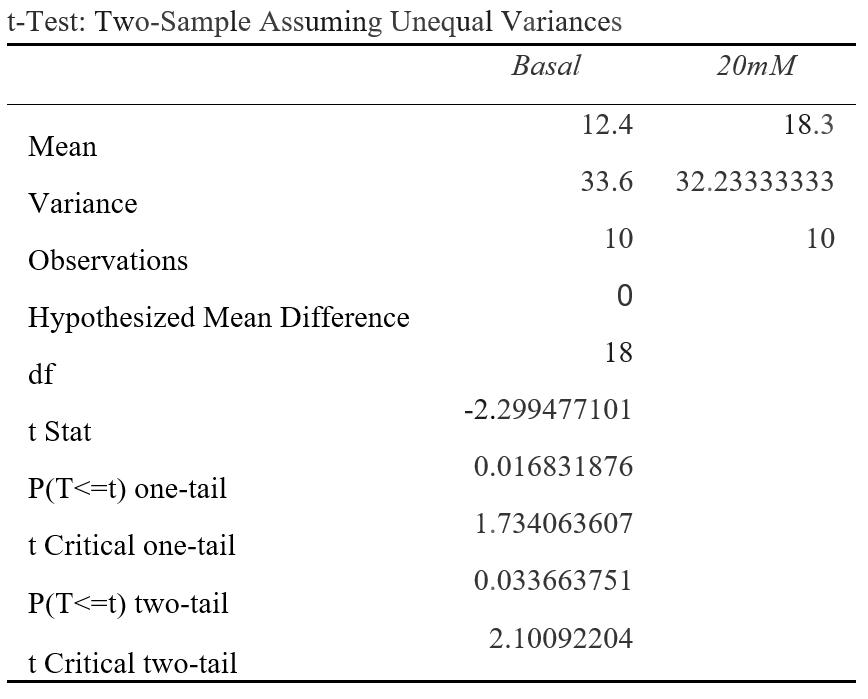
Figure 2C: P(T<=t) two-tail= 0.0337 (P<0.05) ECAR (mpH/min) was measured at a basal rate (13mins) and when 20mM was injected to the cell line (94mins). The difference between the average ECAR for cancer cells at basal rates compared to 20mM were statistically significant as the results were 3.37% due to chance. This suggests that 20mM of lactate could affect ECAR.
Descriptive statistical analysis:
Descriptive analysis for OCR measurements:
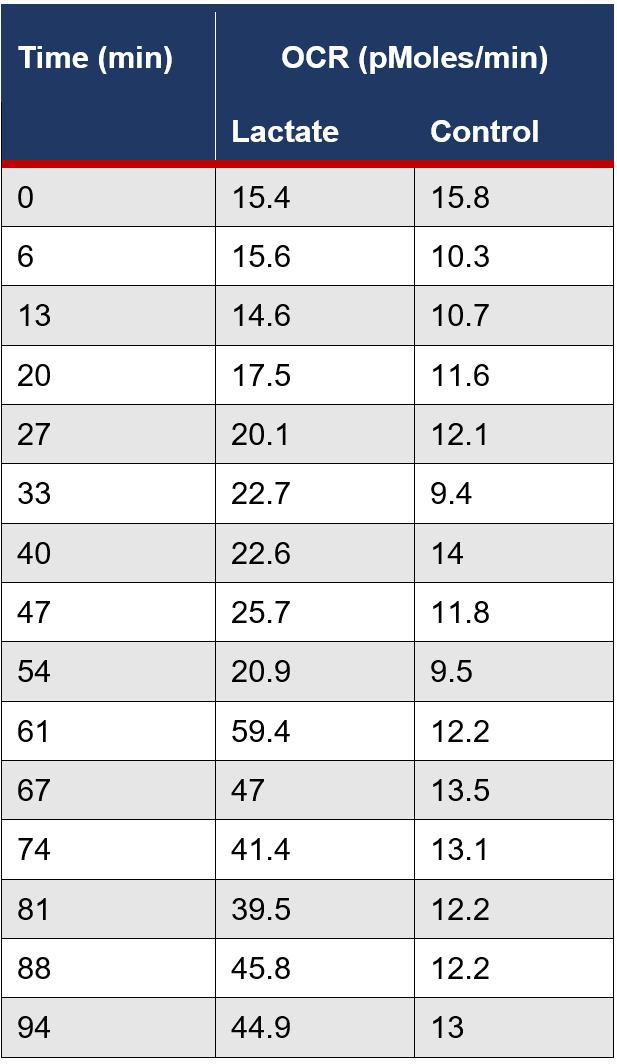
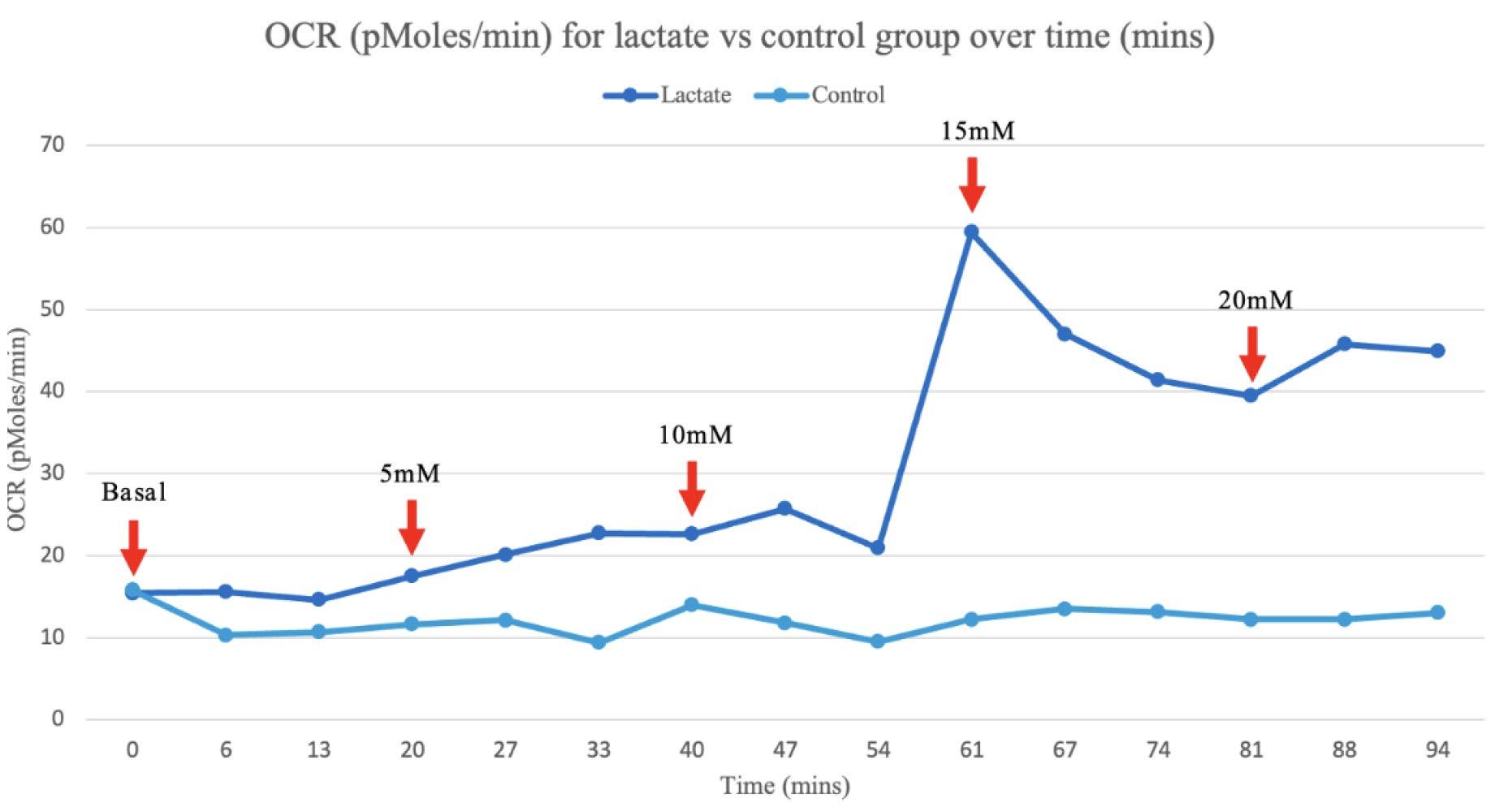
Figure 4A: The average OCR measurements were calculated for the lactate and control group for each time interval during the assay.

Figure 4B: The average OCR measurements for the lactate and control group were graphed for each recorded time interval. The red arrows signify the point at which 5mM of lactate was cumulatively injected.
Descriptive analysis for ECAR measurements:

Figure 5A: The average ECAR measurements were calculated for the lactate and control group for each time interval during the assay.
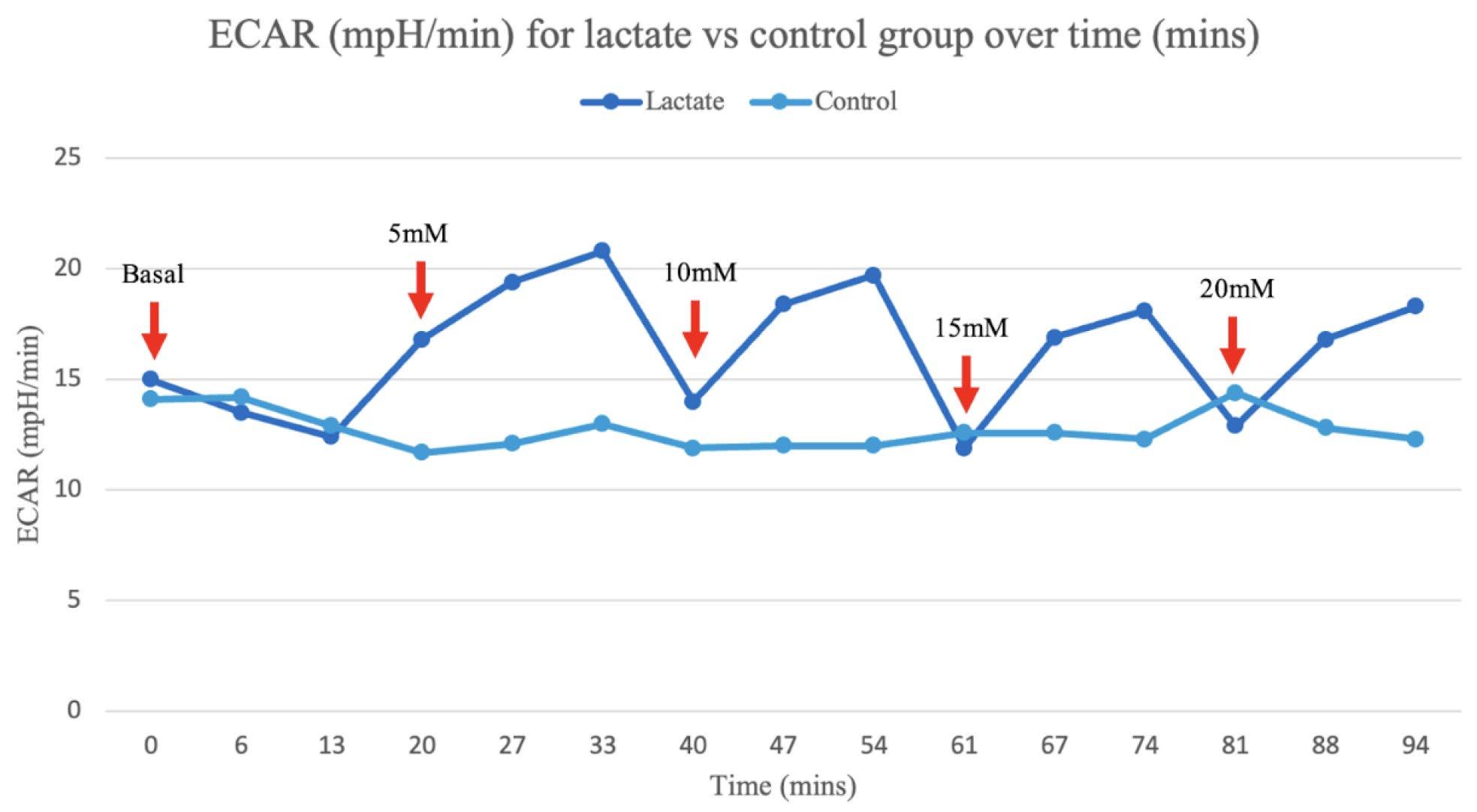
Figure 5B: The average ECAR measurements for the lactate and control group were graphed for each recorded time interval. The red arrows signify the point at which 5mM of lactate was cumulatively injected.
Discussion:
The T-tests conducted reveal lactate has a significant effect on the rates of ECAR and OCR at various concentrations when compared to basal measurements. For the independent group of cancerous cells, significant differences between the means (p<0.05) of the basal rate when compared to the interval at a set concentration (5mM, 10mM, 15mM or 20mM) suggested lactate had a profound effect on either ECAR or OCR measurements as there were a less than 5% likelihood that results were obtained by chance. This occurred at intervals 15mM (p=0.00301) and 20mM (p=0.000686) for OCR measurements (Figure 1A and B), and at 5mM (p=0.0128), 10mM (p=0.0294) and 20mM (p=0.0337) for ECAR measurements (Figure 2A, B and C). For the control group of cancerous cells, which were injected with a buffer serum without lactate, all p-values were high ranging from p=0.404 to p=0.975. This indicates that there were most likely no confounding variables significantly influencing the results, and lactate was the source of change in the experiment. Therefore, the null hypothesis that lactate does not change the ECAR and OCR measurements in a cancerous cell line can be rejected in favour of the alternate hypothesis.
An average of the independent samples indicated ECAR measurements increased over the 5 minute cycle period following an increase in lactate concentration at 13, 40, 61 and 81 minutes, then returned to a similar rate, despite the cumulative increase in extracellular lactate concentration (Figure 5B). This is most likely due to the acidic properties of the sodium lactate added causing anomalous results as ECAR measurements are dependent on pH, evident in the significant increase in ECAR at 5 and 10mM increments (Figure 2A and B). However, the OCR measurements, which are calculated from moles rather than pH, suggest that as lactate concentration increases the rate of OCR also increases, evident in the spike in the rate of OCR in the independent group after 54 minutes when compared to the control (Figure 4B).
The evident switch in the cancerous cell line to a more oxidative metabolism when an accumulation of lactate injected was 15mM and greater, corresponds with emerging literature in the field and reitarties the importance of lactate in understanding cancer metabolics (Wu et al., 2016). The significant increase in OCR in the independent group at 15mM (p=0.00301) from 14.6 to 41.4 pMoles/min and at 20mM (p=0.000686) from 14.6 to 44.9 pMoles/min, provides a point at which lactate causes a metabolic switch from aerobic glycolysis to OXPHOS, extending from current scientific literature (Wu et al., 2016).
Current literature proposes that glycolytic cancer cells are able to sustain their metabolism through types of lactate shuttling including the Reverse Warburg Effect, metabolic symbiosis and vascular endothelial growth (Kooshki et al., 2021), as not all cancer cells necessarily produce lactate (Semenza, 2008). The increase in an oxidative metabolism from 15mM of lactate could therefore be due to disruption of the lactate shuttling concentration gradient required to transport lactate through MCTs, causing the cancer cells to rather use lactate as a fuel for oxidation, potentially indicated by the decrease plateau of OCR from 61 minutes onwards (Figure 4B). The increase in OCR at 15mM could also be attributed to an increase in lactate favouring the oxidation of lactate to pyruvate via LDH, resulting in pyruvate to be used as fuel for OXPHOS (Xie et., 2014). The increase in OCR could also be due to the increase in extracellular lactate decreasing the efficacy at which glucose can diffuse into GLUT transporters in glycolytic cells, creating an environment where glycolytic cancer cells adapt by increasing OCR.
Key limitations and future directions for scientific research:
The in vitro nature of the experiment was beneficial for providing quantitative data on a cellular level for the effects of lactate on cell metabolism, however is limited in providing a thorough understanding of the role of lactate in vivo. The variation across each individual assay (Appendices 2), highlights the need for further replications to analyse results with a large variance to mirror the diversity of ways heterogeneous cancer cells may react under experimental conditions. Despite the highly advanced Seahorse XF technology, it is limited as during the mixing phase of each cycle during the assay, as there is potential that cells could have broken off, impacting the cell count and the reliability of the results. The Seahorse XF results also indicated on various occasions 0 values for either ECAR and OCR (Appendices 2) which also limits the validity of the results as it is questionable if the cells are ever at a state of 0. The limitations of using lactate as the independent variable, which affects pH measurements, also impacts findings as no noticeable trend was observed from ECAR measurements, which could further suggest why there was an insignificant Pearson's correlation coefficient (r=-0.114) between the two measurements (Appendices 3).
Whilst the experiment provided insight to the role of lactate on a cellular level, it cannot be concluded that this directly mirrors the role of exercise, which also produces lactate, on cancer cells. Whilst the concentrations of lactate in the experiment mirror the concentration produced during exercise, further research is required to understand how lactate is transported to tumours from a bodily perspective to assess the accuracy of this experiment as a potential model. Further experimentation should be conducted with a larger sample size over multiple occasions, as well as comparing the results with a noncancerous cell line to further scientific understanding. Investigations into quantitative, in vivo experiments for the role of lactate on glycolysis and OXPHOS will also be beneficial in order to understand the role of single bouts of exercise on tumour intrinsic factors.
Conclusion:
A clear increase in OCR when lactate injected cumulated to 15mM and 20mM, where P=0.00301 and P=0.000686 respectively, indicates a significant link between the concentration of extracellular lactate and the metabolic profile of cancer cells in vitro. Therefore, despite the lack of validity of ECAR measurements due to the addition of lactate potentially impacting the pH of results, it can still be established that an increase in lactate does result in cancer cells switching to a more oxidative phenotype, and the null hypothesis can be rejected in favour of the alternate hypothesis. Hence, these results suggest that lactate could revert the Warburg Effect in cancer cells by potentially disrupting the concentration gradient required for cells to shuttle lactate intercellularly to sustain such a highly glycolytic metabolism. It is critical that further investigations explore the effects of lactate on cancer cells from an in vivo perspective, in order to determine the effects of single bouts of lactate on cancer cell metabolics, as a means of providing a foundation of the possible mechanisms of the effects of exercise on cancer.
Acknowledgements
I would like to express my gratitude to Dr Andy Philip and Garvan Institute of Medical Research for conducting the experiment and providing the Seahorse XF experimental data. I would also like to thank my teacher Ann Hanna for guidance with the data analysis, and my mentor Clara Zwack from the University of Sydney for their support with contacting academics. I would also like to extend my appreciation to my peer Emily Cliff and chemistry teacher Zoe Liley for their feedback regarding the scientific report.
References:
Ashcraft, K. A., Peace, R. M., Betof, A. S., Dewhirst, M. W., & Jones, L. W. (2016). Efficacy and Mechanisms of Aerobic Exercise on Cancer Initiation, Progression, and Metastasis: A Critical Systematic Review of In Vivo Preclinical Data. Cancer Research, 76(14), 4032–4050. https://doi.org/10.1158/00085472.can-16-0887
Brown, T. P., & Ganapathy, V. (2020). Lactate/GPR81 signaling and proton motive force in cancer: Role in angiogenesis, immune escape, nutrition, and Warburg phenomenon. Pharmacology & Therapeutics, 206, 107451. https://doi.org/10.1016/j.pharmthera.2019.107451
de la Cruz-López, K. G., Castro-Muñoz, L. J., Reyes-Hernández, D. O., GarcíaCarrancá, A., & Manzo-Merino, J. (2019). Lactate in the Regulation of Tumor Microenvironment and Therapeutic Approaches. Frontiers in Oncology, 9. https://doi.org/10.3389/fonc.2019.01143
Dethlefsen, C., Pedersen, K. S., & Hojman, P. (2017). Every exercise bout matters: linking systemic exercise responses to breast cancer control. Breast Cancer Research and Treatment, 162(3), 399–408. https://doi.org/10.1007/s10549-017-41294
Fadaka, A., Ajiboye, B., Ojo, O., Adewale, O., Olayide, I., & Emuowhochere, R. (2017). Biology of glucose metabolization in cancer cells. Journal of Oncological Sciences, 3(2), 45–51. https://doi.org/10.1016/j.jons.2017.06.002
Hanahan, D., & Weinberg, Robert A. (2011). Hallmarks of cancer: the next generation. Cell, 144(5), 646–674. https://doi.org/10.1016/j.cell.2011.02.013
Hirschhaeuser, F., Sattler, U. G. A., & Mueller-Klieser, W. (2011). Lactate: A Metabolic Key Player in Cancer. Cancer Research, 71(22), 6921–6925. https://doi.org/10.1158/0008-5472.can11-1457
Hofmann, P. (2018). Cancer and Exercise: Warburg Hypothesis, Tumour Metabolism and High-Intensity Anaerobic Exercise. Sports, 6(1), 10. https://doi.org/10.3390/sports6010010
Hojman, P., Gehl, J., Christensen, J. F., & Pedersen, B. K. (2018). Molecular Mechanisms Linking Exercise to Cancer Prevention and Treatment. Cell Metabolism, 27(1), 10–21. https://doi.org/10.1016/j.cmet.2017.09.01 5
Kooshki, L., Mahdavi, P., Fakhri, S., Akkol, E. K., & Khan, H. (2021). Targeting lactate metabolism and glycolytic pathways in the tumor microenvironment by natural products: A promising strategy in combating cancer. BioFactors. https://doi.org/10.1002/biof.1799
Mathewson, T. (2018, October 30). More than 1 in 4 people across the world don’t get enough exercise, study says. Global Sports Matters. https://globalsportmatters.com/health/201 8/10/30/over-1-in-4-people-across-theworld-dont-getenough-exercise-studysays/
Mishra, D., & Banerjee, D. (2019). Lactate Dehydrogenases as Metabolic Links between Tumor and Stroma in the Tumor Microenvironment. Cancers, 11(6), 750. https://doi.org/10.3390/cancers11060750
Payen, V. L., Mina, E., Van Hée, V. F., Porporato, P. E., & Sonveaux, P. (2020). Monocarboxylate transporters in cancer. Molecular Metabolism, 33, 48–66. https://doi.org/10.1016/j.molmet.2019.07. 006
Philp, A., Macdonald, A. L., & Watt, P. W. (2005). Lactate – a signal coordinating cell and systemic function. Journal of Experimental Biology, 208(24), 4561–4575. https://doi.org/10.1242/jeb.01961
Plitzko, B., Kaweesa, E. N., & Loesgen, S. (2017). The natural product mensacarcin induces mitochondrial toxicity and apoptosis in melanoma cells. Journal of Biological Chemistry, 292(51), 21102–21116. https://doi.org/10.1074/jbc.m116.774836
Ristow, M. (2006). Oxidative metabolism in cancer growth. Current Opinion in Clinical Nutrition and Metabolic Care, 9(4), 339–345. https://doi.org/10.1097/01.mco.00002328
San-Millán, I., & Brooks, G. A. (2016). Reexamining cancer metabolism: lactate production for carcinogenesis could be the purpose and explanation of the Warburg Effect. Carcinogenesis, bgw127. https://doi.org/10.1093/carcin/bgw127
Semenza, G. L. (2008). Tumor metabolism: cancer cells give and take lactate. Journal of Clinical Investigation https://doi.org/10.1172/jci37373
Siska, P. J., Singer, K., Evert, K., Renner, K., & Kreutz, M. (2020). The immunological Warburg effect: Can a metabolic‐tumor‐stroma score (MeTS) guide cancer immunotherapy? Immunological Reviews, 295(1), 187–202. https://doi.org/10.1111/imr.12846 Warburg, O. (1927). THE METABOLISM OF TUMORS IN THE BODY. The Journal of General Physiology, 8(6), 519–530.
https://doi.org/10.1085/jgp.8.6.519
Wilde, L., Roche, M., Domingo-Vidal, M., Tanson, K., Philp, N., Curry, J., & Martinez-Outschoorn, U. (2017). Metabolic coupling and the Reverse Warburg Effect in cancer: Implications for novel biomarker and anticancer agent development. Seminars in Oncology, 44(3), 198–203. https://doi.org/10.1053/j.seminoncol.2017. 10.004
World Health Organization. (2022, February 3). Cancer. World Health Organization. https://www.who.int/newsroom/fact-sheets/detail/cancer
Wu, H., Ying, M., & Hu, X. (2016). Lactic acidosis switches cancer cells from aerobic glycolysis back to dominant oxidative phosphorylation. Oncotarget, 7(26). https://doi.org/10.18632/oncotarget.9746
Wu, M., Neilson, A., Swift, A. L., Moran, R., Tamagnine, J., Parslow, D., Armistead, S., Lemire, K., Orrell, J., Teich, J., Chomicz, S., & Ferrick, D. A. (2007). Multiparameter metabolic analysis reveals a close link between attenuated mitochondrial bioenergetic function and enhanced glycolysis dependency in human tumor cells. American Journal of Physiology-Cell Physiology, 292(1), C125–C136. https://doi.org/10.1152/ajpcell.00247.200 6
Xie, J., Wu, H., Dai, C., Pan, Q., Ding, Z., Hu, D., Ji, B., Luo, Y., & Hu, X. (2014). Beyond Warburg effect – dual metabolic nature of cancer cells. Scientific Reports, 4(1). https://doi.org/10.1038/srep04927
Xie, J., Wu, H., Dai, C., Pan, Q., Ding, Z., Hu, D., Ji, B., Luo, Y., & Hu, X. (2014). Beyond Warburg effect – dual metabolic nature of cancer cells. Scientific Reports, 4(1). https://doi.org/10.1038/srep04927
Zheng, J. (2012). Energy metabolism of cancer: Glycolysis versus oxidative phosphorylation (Review). Oncology Letters, 4(6), 1151–1157. https://doi.org/10.3892/ol.2012.928
Appendices:
1: The Seahorse Bioscience XF Training Manual states that cells should be examined prior to assay to:
• Confirm cell health, morphology, seeding uniformity and purity (no contamination)
• Ensure cells are adhered, and no gaps present
• Make sure no cells are plated in the background correction wells
2: Experimental data provides by Garvan Research Institute
OCR data:

ECAR data