
THE ITALIAN DELEGATION OF THE INTERNATIONAL COUNCIL FOR GAME AND WILDLIFE CONSERVATION

THE ITALIAN DELEGATION OF THE INTERNATIONAL COUNCIL FOR GAME AND WILDLIFE CONSERVATION
2024
by Stefano Mattioli
Layout and graphics: Elisa Bossotto
Printed in Italy at Tipolitografia Botalla s.r.l. - Gaglianico (BI)
Photographic contribution: Andrea Dal Pian
© 2024 Italian Delegation of the CIC
All rights reserved. No part of this publication may be reproduced, stored or transmitted in any form or by any means medium, electronic, mechanical, photocopying, recording, or otherwise without the prior permission of either the copyright owner or the publisher of this book. This includes text and images of the book, which may not be reproduced in any form without prior consent written by the Italian Delegation of the CIC.
Unauthorized reproduction and use will result in legal proceedings.

Premise of the Chief of the Italian Delegation of the CIC, Nicolò Amosso.............
Preface of Stefano Mattioli .....................................................................................
Red Deer in Apennine............................................................................................
References ............................................................................................................
Acknowledgements ............................................................................................... pag. 4 pag. 5 pag. 7 pag. 53 pag. 55
by Nicolò Amosso Chief Of The Italian Delegation Of The CIC
Among the aims of the Italian Delegation of the CIC is increasingly important the protection of the environment, biodiversity and ecosystems in the interest of future generations and we wish to support and contribute to the popularization of applied scientific research.
From this perspective, after the publication of the “Practical Guide to the CIC Handbook for the Measurement and Evaluation of Hunting Trophies”, of the “The Sardinian Red Deer” and of the “The Apennine Chamois”, it is now the turn of “Red deer in the Apennine, or plasticity in action.”
This latest study is inspired by an interesting conference that Dr. Stefano Mattioli held on the occasion of obtaining the Certificate of Excellence, a recognition that our Delegation assigns to those who have particularly distinguished themselves in the defense, study or otherwise in favor of nature and hunting.
In his speech, the scholar explained the origin of the Acquerino red deer population, the period of palmated super-trophies, the phase of the “normalisation” of antlers, the first censuses by monitoring roaring stags and the different patterns of body growth recorded on the two slopes of the Apennine ridge, the Tuscan and the Emilian one.
This publication shows the great versatility of the red deer with respect to climatic and dietary variations in terms of body growth and antler investment, which can be used as an indicator of the health of the population.
I thank Stefano Mattioli, a zoologist involved both in the direct management of ungulates and inresearch, author of many scientific and popular publications, lecturer and CIC Expert of our Delegation for his important scientific contribution first in our project on the Sardinian Red Deer and now with “Red Deer in Apennine, or plasticity in action”.
Special thanks to Andrea Dal Pian for the splendid photographic contribution that accompanies the study.

Foreword by Stefano Mattioli
and CIC Expert
At the invitation of the Italian delegation of the C.I.C., in the following pages I wish to tell the story of one of the first groups of deer reintroduced in the northern Apennine. In the early days, this group was mainly known as the Acquerino population, from the name of the forest rangers’ station where the release of the founding animals was carried out. Here, we retrace the story of this population for more than sixty years, and follow its development, in terms of distribution and abundance, against the backdrop of major changes in the economy and landscape of the Apennine mountains. Through the analysis of the antler size and morphology of red deer it was possible to reconstruct the physical changes of the stags over the decades. The reintroduction of modest-sized Alpine red deer with antlers of simplified structure into Apennine environments, rich in trophic resources and without competition, allowed the animals to thrive and “flourish”, and produce antlers of truly peculiar shape during the 1970s and 1980s. With the start of hunting in 2000, it was possible to monitor the physical performance of the population in more detail and to relate the somatic data to environmental changes in a broad sense. This confirmed the great plasticity of the red deer, a species capable of responding quickly to changes in climatic conditions and food sources. This booklet is also an opportunity to discuss the initial research on this population, which began in 1993 (including: the analysis of antlers; reconstruction of the distribution range; estimates of population size through censuses of roaring stags), as well as the most striking results of the most recent studies (including: body growth of stags on both sides of the Apennine chain; antler investment; interactions between body size; climate and environment in hinds and calves). Finally, I demonstrate that these studies, apparently of strictly scientific interest, can actually have practical implications in wildlife and landscape management.
I wish to dedicate this book to Franco Odorici (1942-2018), a kind and generous friend, and selective hunter, who for years had been responsible for a hunting district. He was an enthusiastic biometric surveyor, passionate about trophies, and a key figure in the red deer management in the Bologna Apennine.


Red Deer in the Apennine or the plasticity in action
In each species, animals interact with food resources by adjusting their body size depending on the productivity of the environment, i.e. the number of prey for carnivores, the quantity and quality of plants for herbivores. Red deer are one of the most adaptable ungulate species in terms of body size, a characteristic known to biologists as “phenotypic plasticity”. Normally in Central and Eastern Europe an adult stag can weigh an average of 160 to 220 kg after the rut season, while an adult hind weighs an average of 90 to 120 kg. However, in marginal environments, such as the Mediterranean scrubland (e.g. Sardinia) or sub-mediterranean coastal woods on sandy soil (e.g. Mesola) or the Scottish moorlands, red deer tend to get smaller in order to survive, with adult stags averaging 95110 kg and females of 60-80 kg. Conversely, in rich environments, such as the Carpathian mountains or the Pannonian plain, cases of red deer stags with post-rut weights of up to 340-350 kg and hinds weighing up to 150-160 kg are documented. If we doubt the incredible size flexibility of the red deer, let us remember that in the Upper Pleistocene on the Island of Jersey in the English Channel lived miniature red deer of around 36 kg and in the islands of Malta deer of just 25 kg (Mattioli et al. 2022). It is hard to imagine red deer of the size of a roe deer, but it was exactly like that.
The famous pioneer of field ethology V. Geist, a great expert on caprine and Cervids, was among the first to describe how animal species can fluctuate between the two extremes of the so-called “maintenance or efficiency phenotypes” (i.e. small-sized frugal ecotypes adapted to low-productive environments) and “luxuriant or dispersal phenotypes” (i.e. large-sized strong ecotypes adapted to environments with an abundance of resources) (Geist 1987). J. Huxley (1931) had already reported how the small red deer of the Scottish Highlands, moved in the second half of the nineteenth century to the vast and rich pastures of New Zealand, soon became large. We must therefore always think of red deer as a species that responds quickly and consistently to variations in trophic and climatic resources by adapting its size.
Most of the current European red deer populations have been in some way shaped by anthropogenic environmental changes. If today Scottish red deer are generally smallsized, this is mainly due to the profound landscape alterations caused by humans since 3000-4000 years ago, when in the Bronze Age the vast Caledonian forests were gradually dismantled, leaving room for heathlands. If medium-sized deer with relatively modest antlers prevail in Central Europe, this is due to the changes made in forest management in the second half of the Eighteenth century, when humans preferred to replace oaks, maples and beeches with even-aged plantations of fast-growing conifers, creating poorer habitats for red deer. A visit to the Baroque Moritzburg Castle near Dresden in Saxony (Germany), can allow us to admire gigantic trophies weighing up to 20 kg dating back to the late Seventeenth and early Eighteenth centuries - evidence of what red deer could have looked like before humans intervened heavily on the environment. In the chronicles of those times we read about stags that in Saxony and in other regions of Central Europe reached pre-reproductive weights of 400-450 kg (cf Geist 1986).
The red deer was quite common in Italy until the first half of the Middle Ages. Then, competition with livestock, deforestation and direct persecution began to drastically reduce its range and numbers. By the beginning of the 18th century it had almost completely disappeared from the plains and hilly areas of the Peninsula. Between the end of the same century and the beginning of the next, the red deer also became extinct in most of the mountains (Alps and Apennines).
Around 1840, the Grand Duke of Tuscany, Leopold II Lorraine, had the red deer reintroduced into the Casentino forests, in the Apennine mountains above Arezzo, probably using animals from Bohemia. At the beginning of the 20th century the species survived only in a few enclosures (La Mandria, the royal estate of the Savoy family near Turin), in the Mesola Wood near the Po river delta and in Sardinia. Around 1925, the first red deer entered Val Venosta (Vinschgau Valley), South Tyrol, from the Swiss Engadin through natural expansion. In the years following the Second World War, due to poaching, the Mesola red deer population (now recognized as a distinct subspecies, Cervus elaphus italicus) was reduced to just a dozen animals. While in the central and eastern Italian Alps the recovery of the red deer depended above all on spontaneous recolonization starting from neighboring countries (Switzerland, Austria and Slovenia), in the western Alps the deer returned exclusively thanks to reintroductions. The first liberation was carried out in 1962-64 in the Gran Bosco di Salbertrand, not far from Turin, mainly with animals from the Slovenian Alps. From 1989 to 2005 more than 200 red deer were reintroduced in Piedmont Alps (P.G. Meneguz, pers. com.). Apennines were also repopulated exclusively by reintroductions. Most of the releases in northern Apennine took place between 1950 and 1998, in central Apennine between 1972-2012, and in southern Apennine from 1993 until today. In Sardinia red deer reached the minimum numbers in 1970, when only three small separate populations survived in the south of the island, for a total of around one hundred individuals. Thanks to the creation of protected areas, breeding programs in large enclosures managed by the Regional Forestry Corps, and reintroduction projects, Sardinian red deer (C.e. corsicanus) recovered and now numbers around 11,000 individuals.
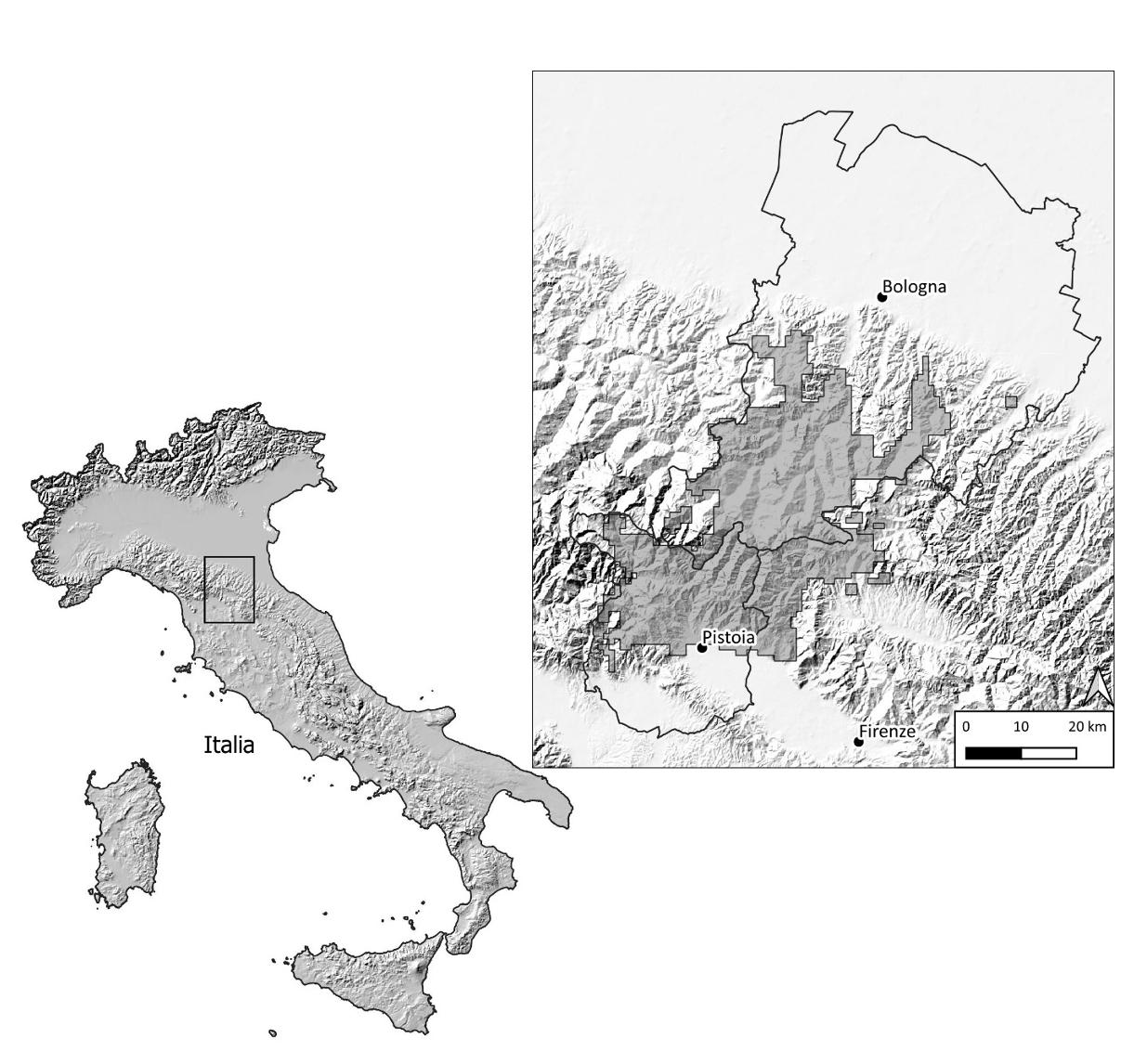
The total population of red deer in Italy grew from 3,500 in 1970, to 7,800 in 1980, 16,000 in 1990 and 44,000 in 2000 (Mattioli et al. 2001, Pedrotti et al. 2001). Unfortunately the last official estimate, provided by ISPRA (National Institute of Environmental Protection), dates back only to 2010, with around 68,000 animals (Raganella Pelliccioni et al. 2013). Northern Apennine population size increased from just about 200 red deer in 1970 to 3,000 in 2000 and 11,000 today. Currently we can guess for Italy a total population of around 100,000 individuals. Hunting is permitted in the Alps and in northern Apennine.

At the end of the last World War, the stretch of the Apennine between the towns of Pistoia, Florence and Bologna was very different from how it appears today. The wood cover was much more limited and showed all the signs of overexploitation. For decades, the coppice woods had undergone frequent cutting cycles and often looked more like shrubs than woods. Many areas were barren due to deforestation and overgrazing by sheep and cattle. On the Bologna side of the mountains there were more open areas cultivated with cereals and forage plants, while on the Tuscan side there was a subsistence agricultural economy with small fields of rye, oats, wheat and forage crops in the middle of the wooded areas.The wildlife had been extremely impoverished for some time: large herbivores (wild boars, red deer, roe deer) had already disappeared by the end of the 18th century, and the predators (especially wolves, wild cats, beech martens, otters) were almost extinct or very rare. The State Forestry Corps was called in to restore and protect the Apennine woods: the coppices were partly converted into high forests, and many open areas were reforested. The administrator of the State Forests of the Pistoia province, G. Premuda, who arrived in 1950, did not stop simply at the silvicultural management of the Apennine, but focused on revitalizing those woods through reintroductions and introductions. In 1952 he asked official permission from the National Agency for State Forests in Rome to plan the release of several species. With determination and enthusiasm, together with some approximation, the administrator built acclimatization enclosures and later released wild boars, red deer, roe deer, hares, partridges, as well as exotic species, such as fallow deer, mouflons, marmots and black grouse, in order to enrich those mountains with wildlife (Premuda 1975). Such naiveties and errors we can understand, especially considering they occurred in those pioneering years, with no national terms of reference regarding appropriate wildlife management, with the Italian academic community being completely uninterested.
In 1958, four red deer (one adult stag, two hinds and a calf) from the State Forests of Tarvisio, in the Friulian Alps, where the forest engineer and inspector had worked at the beginning of his career, were released at the Acquerino forestry station, in the State Forests of Pistoia province. Another three animals from Tarvisio were added in 1965 by his successor Pettinà (Mazzarone and Mattioli 1996). The Tarvisio forest rangers had a fenced area with a group of red deer, donated by their Austrian colleagues from the neighboring Carinthia. Therefore seven red deer from the eastern Alps were transferred to an area of the northern Apennine in total. These deer, which had previously adapted to living in one of the harshest climate areas in Italy, with long, snowy winters and short, cold springs, were now inhabiting an area with a much milder climate, with shorter, less snowy winters and less extreme temperatures. The deer had also moved from a coniferous habitat to an area of rather open forests, with a prevalence of broad-leaved trees and many young reforested patches. We cannot know how much the deer living in Tarvisio weighed then, but we know the average weights in the same area in more recent times. A few years ago adult males weighed just 148 kg whilst females weighed 86 kg - both rather modest values even for the Alps (Mattioli et al. 2022). Harsh winters force the Alpine animals to decrease their movements to save their fat reserves, and to expend energies to keep their body temperature constant. Cold and rainy springs also risk negatively affecting the survival and body growth of newborns. Furthermore, coniferous forests offer few food resources, and the fir and spruce needles themselves are not very nutritious or digestible. For those first red deer founders, the translocation from the Alps to the state forests of Pistoia certainly led to a clear improvement in their physical conditions, as they now inhabited an environment rich in trophic sources and with a decidedly less severe climate (see tabs. 1 and 2). The reintroduction of red deer in the Acquerino Forest was not the only case in the northern Apennine: in the period 1950-60, the Foresters reintroduced the species in Casentino (mountains of Forlì and Arezzo provinces) and in 1966-72, red deer were reintroduced in Orecchiella (mountains of Lucca province) (Mattioli 1992, Mattioli et al. 2001).
Tab. 1: Average meteorological data from the Tarvisio weather station (Alps, Udine province), 780 m a s l (the mean number of frost days is 70% higher than in Acquerino)
Tab. 2: Average meteorological data from the Acquerino weather station (Apennine, Pistoia province), 890 m a s l
Data from the National Meteorological Service
We do not know how the passage from the Alps to the Apennines, with the consequent improvement in climatic and environmental conditions, affected deer body weight. This is partly due to the fact that the Acquerino population remained protected until 1999 and therefore the biometric monitoring, typical of hunting management, was absent. For four decades we can only follow the physical conditions of this group of red deer indirectly, through the collected cast antlers and the trophies of recovered dead animals. The dimensions and some characteristics of the antlers reflect the size and physical state of the stags which produced them: healthy stags living in richer environments grow more and can invest more in the antlers, while stags living in less productive environments tend to be smaller in size and to invest less in the antlers. The antler size therefore represents a good index of the overall health state of a population for all deer species (Brown 1990).
Red deer stags from Tarvisio Forest have a modest body size and relatively small antlers with a rather simplified structure. The deer which were translocated to Acquerino, and their direct descendants, immediately produced antlers of considerable size and with a complex structure. This allowed us to deduce that, thanks to the greater quality of the Apennine environments and the more favorable climate compared to the eastern Alps, as well as the species’ ability to respond promptly to environmental factors, the animals had soon reached respectable physical conditions.
Tales were told for decades about big and extraordinary antlers and finally, between 1993 and 1995, some technicians from the DREAm Italia company, and myself, were called by the EmiliaRomagna and Tuscany regional Administrations to carry out a first study on the Acquerino red deer population. It was a wide-ranging investigation that had to document the populations’ characteristics, reconstruct its distribution and estimate its abundance. One of my tasks was to collect morphometric data on antlers (i.e. measuring them and noting their conformation), which involved going to visit local collectors to measure a representative sample sufficiently large to “characterize” the population (i.e. to describe its peculiar traits). This task was not always easy because some of the most avid collectors were secretive about the stored antlers and therefore not particularly happy to show them to strangers.
Thus, 211 antler beams were examined (60 pairs and 91 single ones), of which 171 belonged to adult stags (i.e. 5 or more years old). These were collected mainly in the provinces of Prato and Pistoia, and could be dated to a broad range of years, between 1975 and 1995 (tab. 3).
Tab. 3: Biometric characteristics of the antlers of adult stags from northern Apennine, 1975-95
(from Mazzarone and Mattioli 1996; mean, standard deviation and sample size)
Tab. 4: Comparison between the mean antler beam lengths of adult stags from some European populations, put in ascending order (Mattioli et al. 2022, Ciccotti 2022)
If the average values and the variability of antler dimensions of Apennine red deer were compared with those of other European populations, on a continental scale, antlers from Acquerino were found to be of medium-high size. They were certainly superior to those from marginal areas such as Scottish moorlands (Rum) and the Mediterranean maquis scrub (Sardinia, Spain), as well as to the antlers from most of Central Europe (tab. 4). The translocation from eastern Alps had certainly favoured an increase in body size (not yet documented at that time) and a greater antler investment (fortunately documented with our survey).
The interest in the antlers from Acquerino of that period lies more in their conformation than in their considerable size. First of all, the terminal crown and the bez tine were very common features, missing in only 18% and 17.5% of adult antlers respectively. Despite being two peculiar characteristics of the red deer, the crown and bez tine occur infrequently in many European populations. More generally, the antlers in our sample tended to have a rather high number of branches, with average values of 13 total tines in adults. In the more extreme cases, several antlers, especially those dating back to the period 1975-1985, have more than 10 tines, with recorded maximums of 12, 13 and 16 tines. This means that some antler pairs had 24, 25 and 26 tines (observed, see photos 3-6), or even 32 tines (not observed). The 16-point antler beam, given its extraordinary nature, was recently acquired by the La Specola Museum of Natural History in Florence, which exhibits it in the Skeleton Hall (photos 5-6). Sometimes, the overproduction of branches not only affected the terminal crown, which appeared “arborized”, but also involved tines of the lower half of the beam, such as the brow, the bez and the trez tines, which in rare cases could be bifid (photos 3-6). The 16-point antler beam, found around 1980, has 12 crown tines and a forked trez tine.
To give a concrete idea of the exceptional nature of these hyper-ramifications, some comparisons with other areas of Europe may be useful. In Hungary, out of 28,790 red deer trophies examined between 1884 and 1957, only 3 reached 26-28 tines (Szunyoghy 1959). Of 17,731 adult stags harvested in the Harz (Germany) between 1959 and 1985, only 2 had antler pairs of 21 total tines and none had antler pairs of 22 or more tines (Reulecke 1987). In more recent times, out of 195,358 adult red deer trophies examined and evaluated between 1990 and 2020 in Hungary, 5 reached a total of 29-35 tines; in the 10 most branched trophies, 7 antler beams had 16 to 22 tines (S. Csanyi, personal communication).
In addition to being hyper-branched, the antlers from Acquerino of those years had a tendency to palmate, that is, where tines fuse to each other, to form conspicuous flattened parts which are accompanied by the arborization of the apical portion of the beams (photos 3-9). The phenomenon is very rarely documented for other European populations, but in this case it affected 10% of our sample of antlers of adult Acquerino red deer stags. Although, given that several antlers are from more recent years, it is possible that the frequency of palmation in the “golden years” was higher.
This demonstrates that the stags of this Apennine nucleus, especially those of the second and third generation from the reintroduction (possibly only 12-30) between 1975 – 1985 took advantage of the low density, the environment rich in resources and the mild climate, which resulted in their exuberance and florid health, and led to the production of luxuriant antlers. These elaborate antlers favoured the number of branches and the palmation over the weight and length, which made them somehow more showy than imposing. Meanwhile, other populations reintroduced in the Apennines,
such as the one founded in the 1970s in the Abruzzo National Park, have shown their vitality through the production of long and heavy antlers (photos 11-12). These deer have not experimented in any way with hyper-branching and only rarely with palmation (Ciccotti 2022). It is really difficult to find a satisfactory explanation for this difference in response between the two reintroduced populations. Perhaps the genetic characteristics of that small group of founders had an impact (the so-called “founder effect”): no longer held back by the rigors of the Alps and moving to optimal environmental conditions, they were finally able to fully develop and indeed amplify their unexpressed traits. On the other hand, the red deer with large antlers of the Abruzzo National Park have probably been able to take advantage not only of the co-occurrence of vast prairies and forests, but also of the different dynamics of reintroductions, with a high number of founders coming from many sources. This somehow represented a winning mix (cf Mattioli et al. 2001).
The hyper-branching of the antlers is a rather rare phenomenon in the red deer. Even in the famous experiments by F. Vogt during 1927-1941 (near Sněžnik, in the so-called Bohemian Switzerland, in present-day Czechia) the deer, fed with high protein and mineral-rich products, had a surprising increase in their body and antler size. Pre-rut live weights reached up to 350 kg, whilst trophies reached up to 14 kg, but with a maximum number of 20-22 total tines (Vogt 1948, Geist 1986).
As for palmation, although recently it is very rare, in historical times it was certainly more common. Red deer with large and palmated antlers are known to have occurred in Italy between the Late Pleistocene and the first part of the Holocene (until the Bronze Age, 4000-3000 years ago) (Abbazzi 1995, Wilkens 1989). These fossil antlers even made paleontologist G. De Stefano consider the existence of a particular subspecies, which in 1911 he named Cervus elaphus palmidactyloceros
A graffito on a limestone fragment found in the atrium of the Paglicci Cave, near Rignano Garganico in Apulia, southern Italy, and dating back around 15,000 years ago, portrays the head of a red deer with a majestic multi-pointed and palmated antler (Palma di Cesnola 1988, fig. 34 on page 67). In reality, as seen in the Acquerino Forest, the formation of palmated antlers may simply indicate low population densities and large availability of high-quality food. The fossil remains and rupestrian representations of palmated red deer perhaps document favorable moments rich in trophic sources, such as animals that have recently colonized environments with an abundance of food and no competition from other herbivores. This poses a useful lesson for paleontologists - they do not necessarily have to imagine distinct lineages, subspecies or “chronospecies” to explain the remains of hyper-branched and palmated red deer, but more simply the antlers could highlight very suitable periods for climate and trophic resources at low population densities. However, at the Acquerino Forest, that season of complex and palmated antlers was destined to end.







Already when we began our investigation in 1993 on that stretch of the northern Apennine, antler conformations of this kind were becoming increasingly rare. This was not surprising; if the antlers are in some way the mirror of environmental conditions, without us realizing what was happening, the Tuscan-Emilian Apennine was becoming a slightly less hospitable environment for red deer. The exceeding of the density threshold of 1-1.5 animals per square kilometer (a relatively low value, but an evidently sensitive one for the species), along with the gradual closure of the coppice woods not converted into high forest and, on the Tuscan side, the definitive end of subsistence agriculture with the abandonment of the cultivated fields and the consequent conquest by broom stands and bracken ferneries, began to subtly influence the vigor of the deer and therefore also the antlers of stags. The antlers were no longer those of the golden period between 1975 and 1985, but it was still not easy to notice it, because their average size was substantially similar: only the hyper-branched or palmated antlers had become increasingly rare. It can be said that in the 1990s the conditions of the red deer between Florence, Prato, Pistoia and Bologna had become in some way “normalised”, that is, density and environment had led to the elimination of the most extreme cases, the most showy and elaborate antlers.
In 2000, the two regions of Emilia-Romagna and Tuscany started selective hunting of red deer and it was therefore possible to develop an in-depth biometric monitoring of both the somatic dimensions of all the animals and the antlers of the stags. As a result, we realized the situation was evolving further. While on the Tuscan side of the Apennines the environmental changes became more evident with the progressive disappearance of the ancient fields, on the Emilian side the cooccurrence of woods, groves, shrubs, pastures and fields still guaranteed a good diversification of trophic resources. The biometric survey therefore allowed us, for the first time, to distinguish between the morphometric characteristics of the red deer on the south facing Tuscan slope from those on the north facing Emilian slope of the mountain range. As could be expected from such a plastic animal as the red deer, the species responded promptly to environmental changes and the environment modulated its body growth and antler investment. Being born and living on a slope of the same relief was sufficient to tend to have one pattern of body growth or another, one kind of antlers or another: no striking differences but certainly two paths no longer superimposable, as in the first decades.
Tab. 5: Mean body and antler weights in red deer of the two slopes of the Apennine
(from Mattioli et al. 2021, Vannini et al. 2021, based on the data of 1565 yearling, subadult and adult stags between 2000-2020 and of 2562 calves and hinds collected between 2000-2017.
Whole body weights of hinds and calves reconstructed from the eviscerated weight)
variable (rounded values)
Mean body weight of adult stags (kg)
body weight of adult hinds (kg)
Mean body weight of male calves (kg)
Mean body weight of female calves (kg)
Mean weight of the antler pair of adult stags (kg)
Mean weight of the trophy of adult stags (kg)
On average, adult stags living on the Bologna side weigh approximately 15% more than adult stags on the Pistoia side, adult females 9% more, male calves 18% more, and female calves 26% more (tab. 5). Body weights similar to those found in Pistoia are also detected in nearby Prato province (Becciolini et al. 2016). The antlers of adults on the Emilian side weigh on average about 20% more than those on the Tuscan side. Given that environmental conditions shape plastic species such as the red deer, the unequal changes that occurred in the last decades of the twentieth century to the Apennine habitats on the two sides ended up having different impacts on the growth and final size of the animals in the two regions. Multi-pointed and/or palmated antlers have become very rare, especially on the Tuscan side. The antler beams with more than 9 points, which in the 197094 sample (n = 171) represented 5.3%, constituted just 1% in the 2000-2020 period (n = 1425; unpublished data), and were completely absent on the Pistoia side. On the Bologna side, they reached a maximum of 11 tines.
When the first experimental hunting began in 2000, we planned an in-depth biometric monitoring program, which produced a database without equal in Europe. The data collected include body weights, various external somatic linear measures (such as head-trunk length and height at the withers), several craniometric measures (such as condyle-basal length and mandible length), and, for stags, the metrics of antlers (weight of the trophy and linear measurements foreseen by the CIC formula). Body weights and external linear measurements are recorded by biometric surveyorshunters trained with a specific course and an exam to measure the harvested animals in special check stations. The skull measurements were taken by the wildlife technician, whilst the linear measurements of the trophies were carried out initially by the biometric surveyors and, for the most important cases, at least in the Bologna province, repeated by the CIC evaluators a few weeks before the annual trophy exhibition. All trophies were weighed at least three months after killing as required by CIC. The age of each individual was estimated by the wildlife biologist on the basis of the average patterns of eruption, replacement and wear of the teeth, referring to a photographic atlas (De Marinis 2013), built on the basis of the analysis of micro-sections of teeth of a subsample coming from the Pistoia side of the Apennine.
Once reintroduced at the Acquerino forestry station, the very small nucleus of founders had to wait a few years to grow and settle permanently in the state forests of Pistoia. The red deer probably took advantage of the altitudinal movements by going down and up along the entire mountain slope. They were thus able to fully use all the available food resources, given that the birth of new shoots and fruiting have a phenology linked to the different elevation. The first sightings occurred on the Emilian side of the Apennine around 1968, in the Monte Calvi area, now included in the Regional Park of the Suviana and Brasimone lakes. Probably in the same years, some individuals began to move eastward and populate areas of the Prato province, always on the Tuscan slope. In reality, the presence of the species was perceived on the Bologna side and in the middle hills of the Tuscan side only from the end of the 1970s. A survey by the University of Pavia was carried out through questionnaires sent in 1977 to all the forestry stations in Italy. It returned a reductive image of the distribution range of red deer on the Tuscan-Emilian Apennine mountains (Pavan and Mazzoldi 1983). Outside the historical areas of the Acquerino Forest, despite the large size and conspicuousness of the stags, the red deer went mostly unnoticed. In those first decades the animals must have been very mobile, returning for the rutting period to the heart of the range around Acquerino. The range was therefore of a pulsating type, with smaller rutting quarters and larger winter-summer quarters, which remains so still to this day. The colonization of the Bologna side of the Apennine was more complex: in the 1980s, the expansion of the historical nucleus of Acquerino was accompanied by the formation of the nucleus of Monte Sole (a sort of “gemmation” from the northernmost part of the continuous range but separate by almost 30 km) and the escape of deer from breeding farms and enclosures (Prada near Grizzana, Rongrino near S. Benedetto Val di Sambro, S. Uberto hunting estate near Monterenzio). A recent genetic study on a sample of red deer from the Prato province made it possible to recognize types of mitochondrial DNA different from those expected of a population of Alpine origin (Giovannelli et al. 2022), probably the result of escapees from the Rongrino enclosure.
For a first complete reconstruction of the distribution range and the first estimates of the population size, we had to wait for our 1993-95 field survey and the first censuses. The census technique adopted, i.e. the nocturnal counts by listening from permanent points and triangulating the roaring stags, as well as the reconstruction of the demographic structure through sighting sessions, was developed in 1988 in Casentino (Mazzarone et al. 1991). Thanks also to the active volunteering of an association of selective hunters from Bologna (URCA), we were able to cover 67 km2 in the Prato, Pistoia and Bologna provinces and were thus able to estimate in 1995 an autumn population of approximately 1350 red deer for a total distribution range of 655 km2. In 2000 the total population was estimated at around 2000 deer over 810 km2, which increased to 2600 head over 1390 km2 in 2010. In autumn of 2022 the population estimate was around 4200 red deer over a total area of 1950 km2: of these, there were approximately 890 in Pistoia and at least 1700 in Bologna. Despite the numerical increase, the overall population density has remained
rather constant and at tolerable levels, around 2-2.5 animals per km2. The rutting areas, which in 1995 constituted 24% of the total range, and 38% in 2010, today represent 61%. This means that the deer have gradually changed their spatial behavior, with a tendency towards relatively more reduced seasonal movements. In 1995, some deer from the Emilian side had to migrate for dozens of kilometers to reach the rutting quarters, sometimes on the Tuscan side of the mountain - only a decade later, they could find their rutting stands in a narrower radius on the Bologna slope. In short, the range remains pulsating but the seasonal movements are more limited.
Tab. 6: Percentage of major land cover types in the red deer range in the opposing slopes of the Apennine mountains; BO = Bologna and PT = Pistoia (from Mattioli et al. 2021)






In 2021, using a considerable part of the data collected in the big trans-regional management unit, some researchers from the University of Siena and the University of Fribourg, plus the wildlife technician responsible for the management of red deer in the Pistoia province (S. Nicoloso of the DREAm Italia company) and myself, tried to learn more about the phenotypic plasticity of this Apennine deer population by publishing two scientific articles in English in important journals on biological topics.
In the first article, entitled “Spatial variation in antler investment of Apennine red deer”, written by S. Mattioli, F. Ferretti, S. Nicoloso and L. Corlatti, published in the journal Ecology and Evolution and freely accessible online, we examined the body weights of the stags, the corresponding antler weights and the age of the animals. Data analysis allows us to reconstruct the patterns of body growth of red deer males and the so-called “antler investment”. Our study aimed to answer several questions: how many years does the body growth phase last? When does the body weight tend to stabilize? At what age, if any, do stags begin to show a decline in weight? And similarly, how do antlers develop year after year? At what age do antlers reach their maximum weight? When is there a regression? We were very interested in analyzing the antler investment, i.e. the quantity of the antler bone tissue produced by the deer per unit of body weight, in other words the relative weight of the antlers, which measures the effort in transferring energy to build the antler pair. The phenotypic plasticity observed in the red deer is not only expressed in the great flexibility of body size, but also in the different ability of stags to allocate energy in producing their antler. Deer living in environments rich in food resources will tend to be large in size and have large, heavy antlers, while deer living in low-productivity environments tend to be modest in size and save energy by developing equally modest antlers. Ultimately antlers are secondary sexual character with a low growth priority, which can be “economised” if environmental conditions require it. When we began analyzing the data, we were interested in understanding whether the antler investment changed depending on the age class, as would be expected from structures that have an active function only among adults. More generally we were interested in documenting the differences in body growth, in the antler development and in the antler investment on the two slopes of the Apennine. If from the beginning the two sides of the same mountain range presented different environmental characteristics and if in recent times the differences have become more pronounced, what will be the differences in body size and antlers in the two provinces?
To correctly analyze the body weights and therefore the body growth of stags it is necessary to use homogeneous data. Since weights vary greatly during the hunting season (which in the early years ran from August to February-March) it was necessary to know the relationship between body weight and day of harvest to reconstruct a comparable body weight for all individuals. So we set the body weight on October 20, two weeks after the end of the main rutting period (“post-rut weight”).
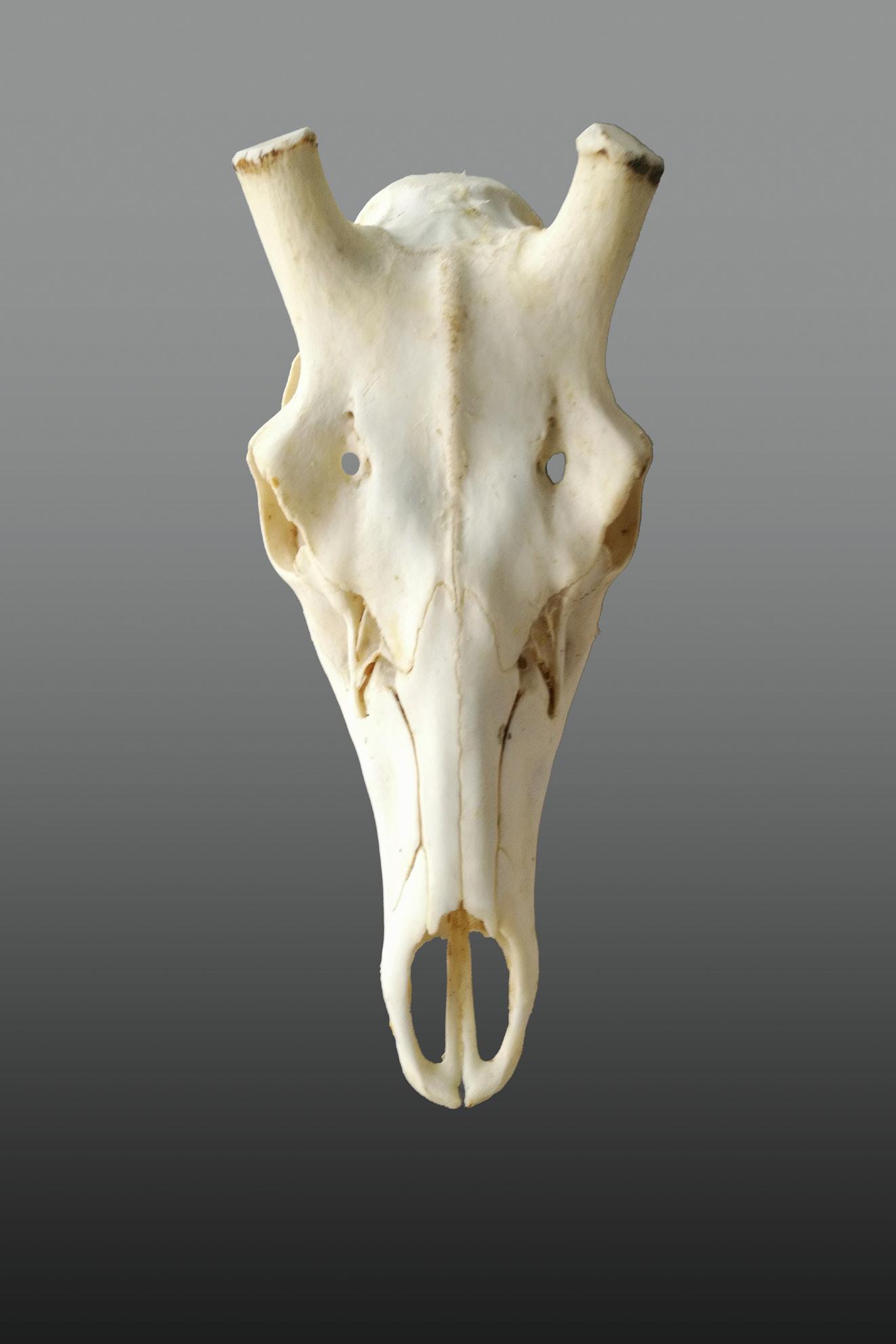
To analyze the antler weights the task was less easy. In fact, the data available were the weights of the trophies, i.e. the weights of the two antlers plus the whole skull. Given that the weight of the skull on average in yearlings constitutes 70% of the total, in subadults 40% and in adults 25%, it was necessary to eliminate this disturbing element which would have risked interfering with the analysis of the actual antler investment. So how can we estimate the net weight of the two antlers if we only have the gross weight of the trophy available? We have chosen a path never traveled before. First of all, we collected a sample of skulls from stags of each age class without antlers, i.e. skulls of animals that had recently cast their antlers or skulls from which we had sawed off the antlers (photo 14).
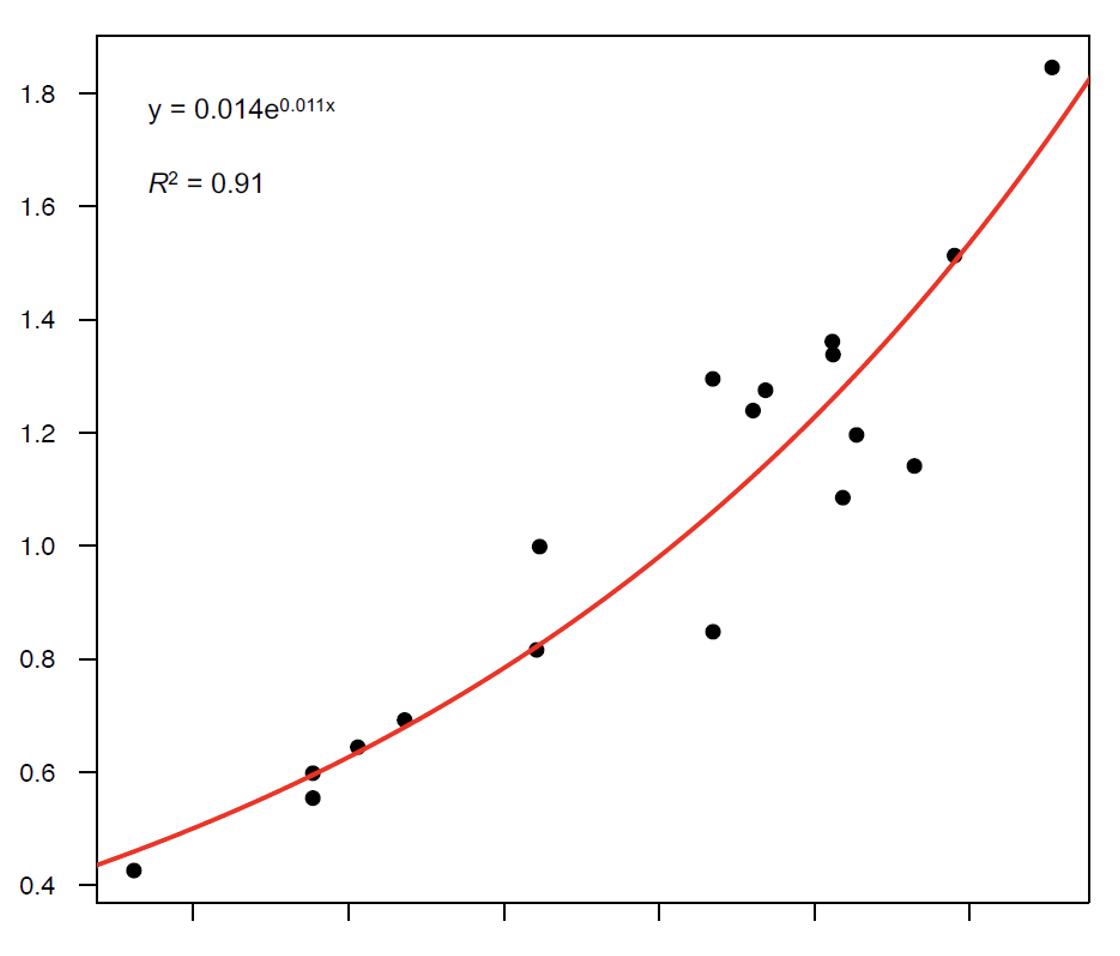
We weighed them, we measured their condyle-basal length and we found a clear and strong mathematical relationship between the two variables (graph 1). At this point, if for each stag in our database we have the gross weight and the condyle-basal length of the skull, it is sufficient to apply the mathematical formula to reconstruct the weight of the skull, to subtract it from the weight of the trophy and thus obtain the estimate of the actual weight of the antler pair.
Skull length (in mm)
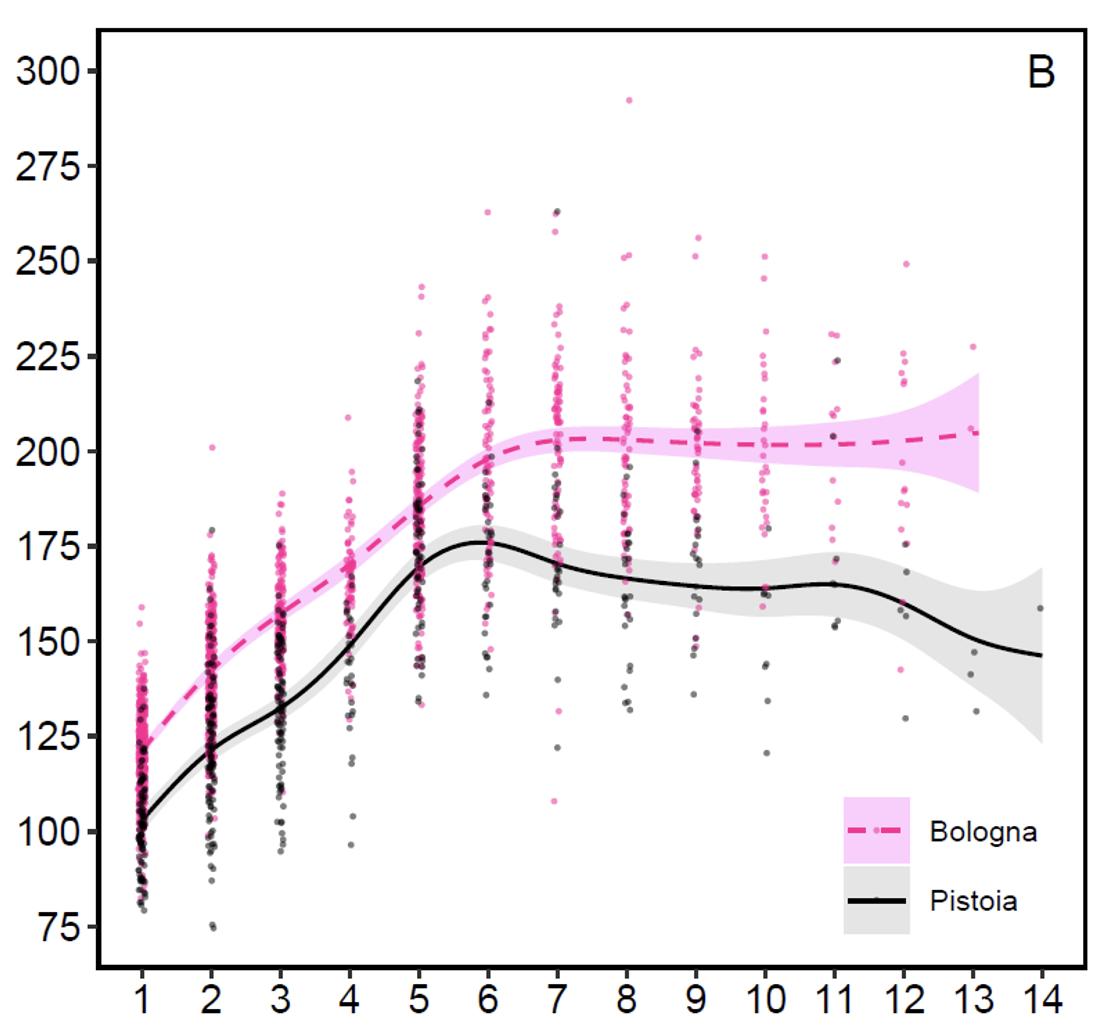
Body weight of stags (graph 2): The body growth phase lasts up to 6 years of age in Pistoia and up to 7 years in Bologna, but while on the Emilian side the weight stabilizes, surprisingly on the Tuscan side the weight immediately tends to decline gradually up to 11 years and then more decisively at 12-14 years of age. It is difficult to find a satisfactory explanation for this body weight loss in Tuscan stags 8-9 years old, at an age when the maximum physical condition is normally attained. In Bologna, young adult males aged 5-7 years weigh on average around 192 kg after the main rutting season and those aged 8 or more around 200 kg. In Pistoia for the same age groups the mean body weight goes from 173 kg to 163 kg. The highest post-rut weight recorded in the Bologna side of the mountain was 292 kg, while in the Pistoia side it was 263 kg. The differences in size between the two slopes are evident but not particularly marked and the performance on the Tuscan side is comparable to many central European populations and always far superior to the progenitor red deer from the Tarvisio Forest in the eastern Alps. ‘Reduced’ skull mass (in kg)
Body mass (in kg)
Age (in years)
Graph 1: Relationship between the antlerless skull weight (in kg) and the condyle-basal length of the skull (from Mattioli et al. 2021).
Graph 2: Post-rut body weight variation with age (in years) on the two slopes of the Apennine (from Mattioli et al. 2021)
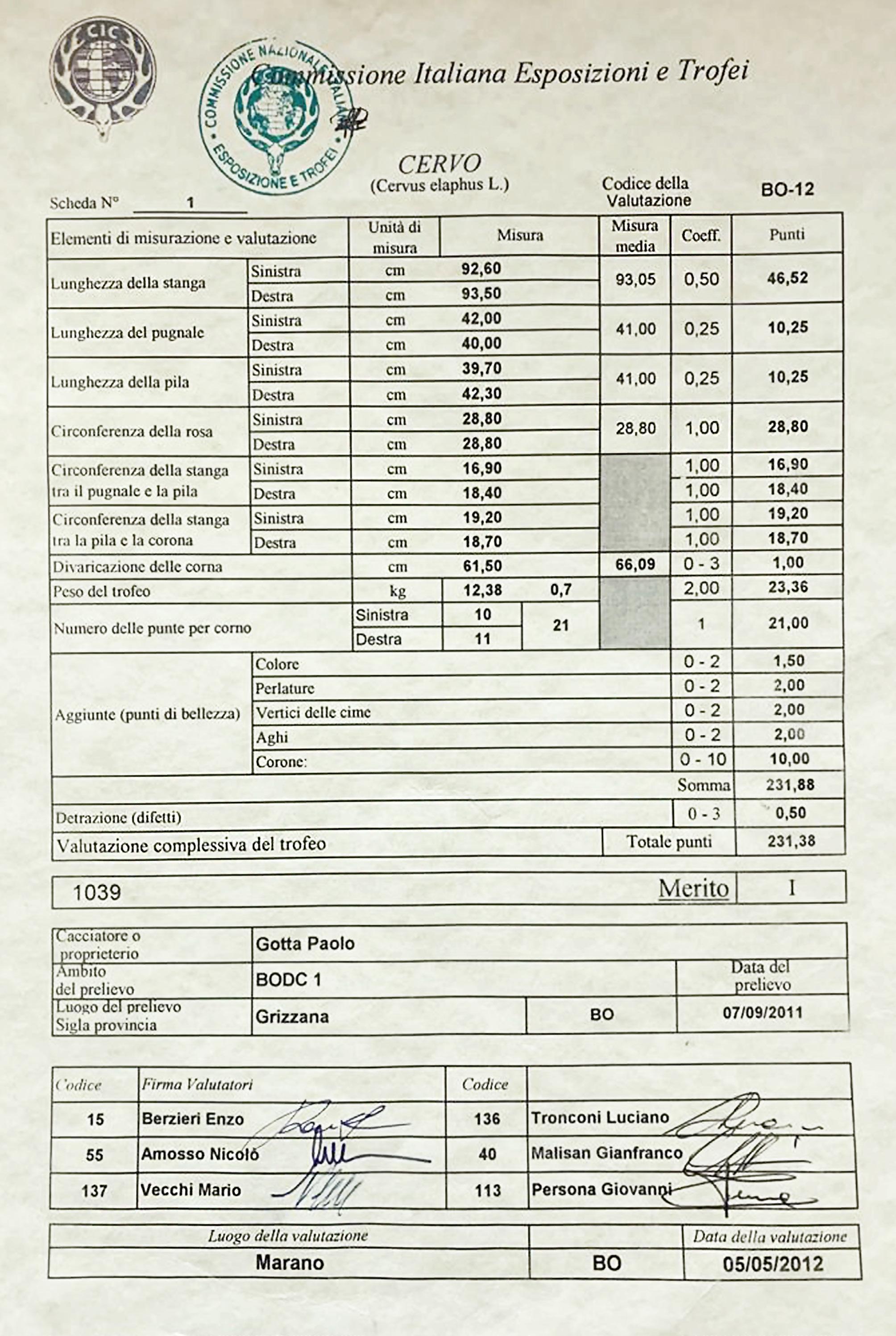
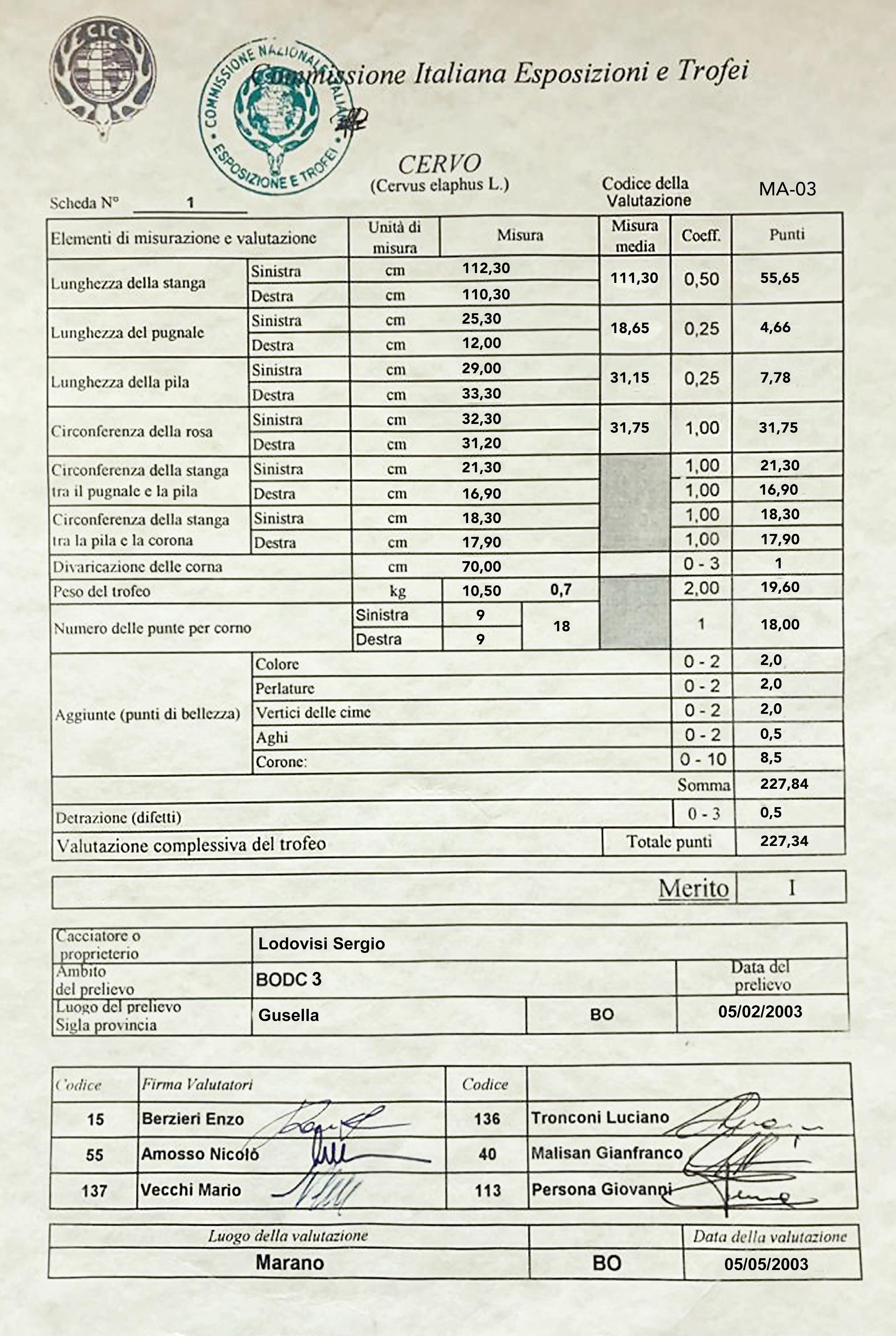
Antler weight (graph 3): The average development of antler weight has a fairly similar trend to that of body weight, growing up to 6 years of age in Pistoia and 7 in Bologna, remaining more or less stable up to 11 years and undergoing a decline or regression from age 12 onwards. The average weights of antlers stabilize at 5.3 kg in the Emilian side and at 4 kg in the Tuscan one. The highest weights detected in the 20 years of monitoring were 10.78 kg in Bologna and 6.66 kg in Pistoia. A 21-tined trophy from the Bologna side, weighing 12.38 kg with the skull, represented the Italian CIC record for 2011-12, with a score of 231.38 points (see photo 16 and the corresponding CIC trophy evaluation chart). The following year it was surpassed by a trophy from the Reggio Emilia province, also in northern Apennine, of the same weight, with 16 total tines but with longer beams and with a score of 235.25 CIC points. The second heaviest trophy from Bologna Apennine had 18 tines, weighed 10.5 kg and had a score of 227.34 CIC points (see photo 17 and the corresponding CIC trophy evaluation chart). Therefore, if it is true that the Acquerino deer, after the period of particular splendor between 1975 and 1985, have gradually normalized their performance in terms of structure and size of the antlers, it is also true that they are still capable of produce large antlers, sometimes even heavier than those of the golden period, during which no trophies weighing over 10.7 kg are known (unpublished data). The biggest difference compared to the past lies in the more essential architecture, with a less extreme number of branches and generally without palmation. To give an idea of the overall good quality of the red deer on the Bologna side, we remind that from 2000-01 to 2022-23, 50.5% of the adult stags harvested gained a medal in the CIC evaluations (unpublished data) and 8 trophies were awarded with a gold medal.
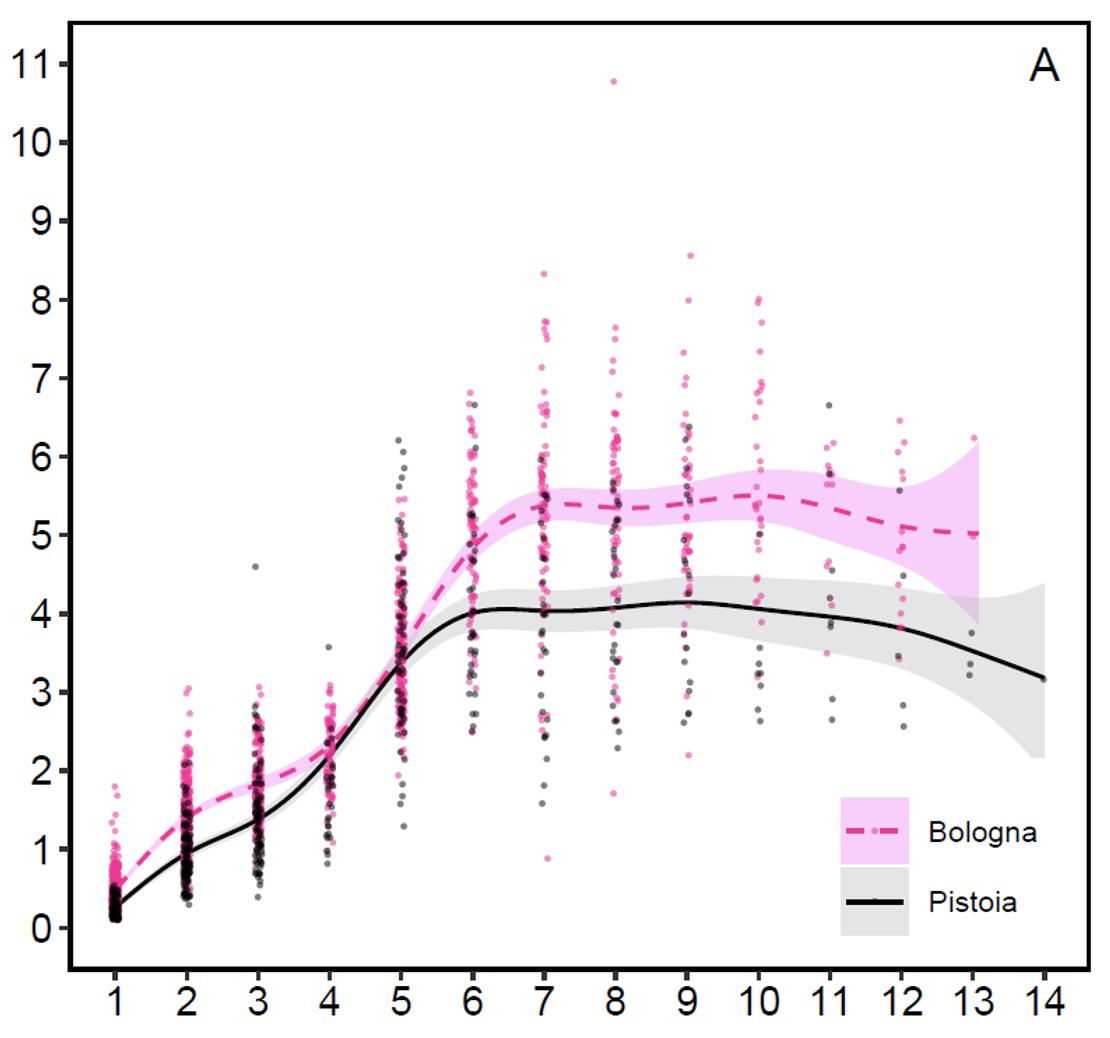



Antler investment (graph 4): Once the weights of the antlers and the body weights have been examined separately, it is possible to analyze their relationship, i.e. the percentage investment that the Apennine red deer make in building their antler. The weight of the antlers in one-year-old stags constitutes on average 0.2 (Pistoia) and 0.4% (Bologna) of the body weight, in subadults 0.9 and 1.1%, in adults aged 5-7 years 2.2% and in adults aged 8 and over 2.5-2.7%. Fully mature red deer stags aged 8 and older invest in the antler on average 7-11 times more than yearlings, demonstrating that investment should only be maximized when deer need to use the antlers in competition for breeding. In more detail, fully mature stags produce on average 24.6 grams of antler bone tissue for every kilogram of body weight on the Pistoia side and 26.7 grams on the Bologna side, a minimal difference per unit weight. This means that on both sides the fully mature stags allocate more or less the same proportion of antler bone and that the difference in the antler weight depends on the different body weight of the animals on the two sides of the Apennine due to the different productivity of the environments. The maximum percent investment recorded in Bologna was 4.3% and in Pistoia 3.9%.
Antler/body mass ratio (in %)
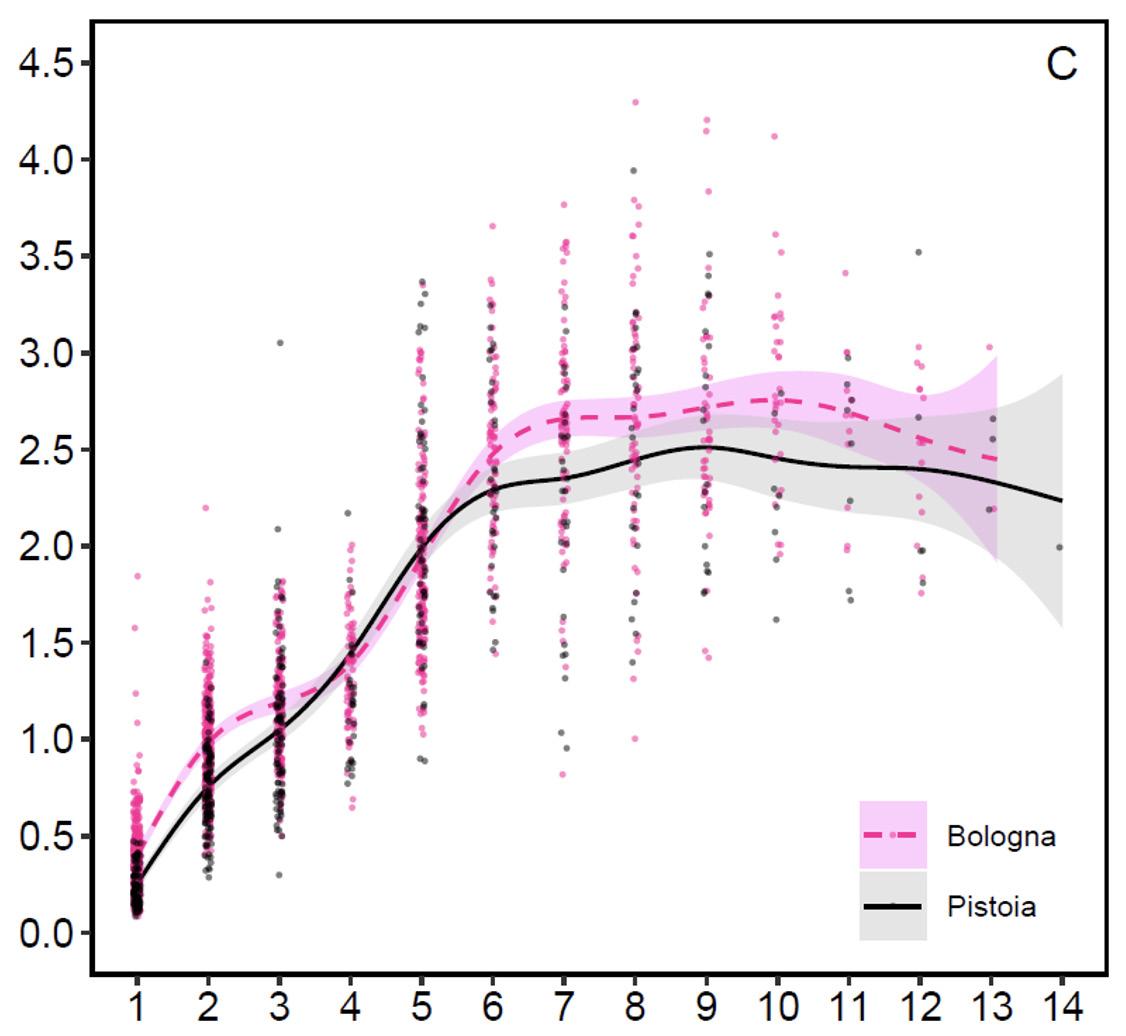
Age (in years)
Antler investment in other red deer populations: Naturally, to correctly frame the percent values found in the Apennines they must be compared with those of other red deer populations known in the literature (tab. 7). Unfortunately this is not simple because in many cases – especially in Eastern Europe - not all the trophy weights are recorded but only those that have been awarded with medals or in any case exceed some threshold, and body weights are often not systematically recorded. As far as we are able to reconstruct, the antler investment in the red deer from the Tuscan-Emilian Apennine between the provinces of Pistoia and Bologna, appears to fit among those of mediumhigh performance populations, such as those of the Polish Carpathians (averages of 23-27 g per kg of body weight), while in Central Europe medium-low values seem to prevail (16-19 g/kg). In Sardinia, Mesola and the Scottish moorlands the antler investment is decidedly lower (10-12 g/kg), as can be expected from deer living in low-productive environments which must measure sparingly the production of bone tissue. The highest investment (averages around 35 g/kg) is documented in Slavonia (Croatia) and Pannonia (Hungary), but probably equally high or even higher average values can also be recorded in the Romanian Carpathians and in the Bulgarian mountains, lands which produce splendid and heavy super-trophies (of up to 16-18 kg). And surprises will probably come from a better knowledge of the antler investment reintroduced deer in the Abruzzo Apennine, central Italy, where pairs of cast antlers weighing up to 12 kg (presumed trophy weight around 14 kg) and up to 131 cm long (Ciccotti 2022) are known (photos 11-12). That the average investment of red deer varies in Europe from around 10 to 35 g and more per kg of body weight is yet another demonstration of the great phenotypic plasticity of this species.
Antler investment in other deer species: It is also interesting to frame the antler investment in other species within the Cervidae family (tab. 8). The red deer is part of the most polygynous and dimorphic species cervids, that is, the species in which the adult males face a high competition for the access to the females and therefore have decidedly larger body size than the females and tend to invest more also in sexual secondary traits such as antlers: adult males of wapiti and fallow deer, which have reproductive strategies rather similar to those of red deer, produce on average approximately 22-34 g per kg of body weight. There are also species with intermediate sexual size dimorphism (such as the American white-tailed deer), which have less intense competition and therefore invest on average around 11-12 g per kg of body weight, and finally there are species such as the roe deer in which the competition between males and sexual dimorphism are decidedly more attenuated and, as we know, adult males content themselves to build small and light antlers, investing on average only 4-7 g per kg of body weight.
Tab. 7: Mean antler investment (g per kg of body weight) in some European populations of red deer (from Mattioli et al. 2021)
Tab. 8: Mean antler investment (g per kg of body weight) in some other Cervid species (from Mattioli et al. 2021)
Management implications: At first glance, our scientific article on antler investment might only be of interest to a small circle of specialists, but in reality it has concrete implications for the daily management of a red deer population. Regularly collecting data on the weights of antlers/trophies and body weights of all the harvested stags, therefore knowing the relationship between the antlers and the body weight, i.e. the antler investment, allows to estimate the physical condition of the single deer and of the entire population. If stags living in more productive environments tend to invest more in antlers than those living in poorer environments, this biometric data can be used as a measure of the health and vigor of the population, as a general index of condition, and the mean values found by us in the Apennine red deer can serve as a reference. Trophy evaluation could also benefit from systematic biometric monitoring. Today, trophy scoring is often somehow completely separated from the knowledge of the entire population, and those who evaluate the trophies do not know the biometric characteristics of the animals that carried those cranial appendages. However, knowing the trophy and the body weights of all the stags of a population of red deer and therefore also of those awarded with medals permits us to better understand the potential and quality of that population. Monitoring all the animals over time through this double data collection allows to follow changes in performance and possibly decide on appropriate interventions to improve the situation.
In the second scientific paper, entitled “Land cover and weather jointly predict biometric indicators of phenotypic quality in a large herbivore”, written by C. Vannini, N. Fattorini, S. Mattioli, S. Nicoloso and F. Ferretti, published in the journal Ecological Indicators and freely accessible online, the influence of environmental and climatic factors on the body weight and various somatic measurements of Apennine red deer hinds and calves was examined.
Through the analysis of the body weight of stags, the weight of their antlers and, therefore, the antler investment of the previous study we were able to observe only differences on a large spatial scale, between one side of the Apennine chain and the other one. In contrast, with the analysis of the body size of the hinds and calves of this second investigation, we were able to highlight differences even on a small spatial scale and on a temporal scale. Stags have large home ranges and their physical conditions are the result of feeding on vast surfaces; the fact that within each slope we had not found differences between districts or periods means that within the large frequented ranges the stags in the different districts have always found substantially equivalent food availability. Hinds and calves - as also documented by preliminary radiotelemetry studies in the area - tend to have smaller home ranges (around 300 ha, on average; see Bocci et al. 2009, Crocetti et al. 2010) and therefore it is possible to evaluate the environmental conditions experienced by individual animals in the area surrounding the sampling point. In particular, five biometric variables were analysed: body weight (eviscerated), shoulder height, thoracic circumference, hind foot length and mandible length. The first four had been recorded from 2001 to 2017 by authorized biometric surveyors, while the length of the jaw was recorded by a wildlife technician (S. Mattioli for Bologna, S. Nicoloso for Pistoia).
For each of the harvested deer, a buffer was created on a computerized map around the harvest point and land cover of the buffer zone were analysed, i.e. the proportion of the different main habitats, focusing on the percentage of cultivated areas. Furthermore, the regional meteorological services of Emilia-Romagna and Tuscany had provided us with rainfall and temperature data for the investigation period. We were interested in seeing if the rainfall and temperatures of the growing season (spring and summer) and the winter severity of the previous winter prior to sampling had any effects on the body weight and skeletal growth of the animals The results of our analysis are quite interesting. First of all, it is confirmed on a large scale that adult
and yearling hinds and calves living on the Bologna side of the Apennine chain are larger in size than those on the Pistoia side (tab. 5). The range occupied by deer in the Bologna area has greater heterogeneity than that of the Pistoia one, which shows larger and more compact forest tracts and relatively few cultivated fields and small pastures, often grouped together. On the Bologna side, the open areas are more than three times larger than those on the Pistoia slope, while the woods on the Emilian slope represent only two-thirds of those on the Tuscan side (tab. 6).
However, an analysis on a finer scale has allowed us to better appreciate the plasticity of the species, capable of responding in terms of body growth depending on environmental characteristics and climatic factors.
Let us look at the results by sex and age class separately:
In adult hinds (3 years of age and more), body weight increases as the proportion of cultivated areas into the buffer of 300 ha centered on the sampling point increases. According to the mathematical model developed by the authors, an adult hind that lives in an area with large cultivated surfaces in Pistoia (70-90% of crops in the surrounding areas) has a mean eviscerated weight that is 5 kg higher than a hind that frequents an area with few or no crops at all (73.9 kg versus 69 kg). The body weight is also affected by rainfall and temperatures and it is greater if in the growing season preceding the harvest the weather was rainier and cooler.
In yearling hinds, but only on the Pistoia side, body weight increases as the proportion of crops in the buffer zone increases. According to the mathematical model, an individual of this class harvested in an area with 90% of crops would have an eviscerated weight 15 kg higher than one taken from an area without crops. For hinds that are still growing - like the yearling ones - this is an extremely significant percentage. Even the hind foot length and the mandible length of the yearlings increase as the extension of the cultivated areas increases.
In female calves of 8-10 months, only in the Pistoia area, body weight, hind foot and mandible increase as the surrounding crops grow. According to the model, a calf from an area with many crops (70-90%) around has on average an eviscerated weight approximately 5.5 kg higher than an individual from an area with few crops or none at all (42.5 kg against 36.9 kg). Wetter and cooler springs and summers have positive effects on body weight and thoracic circumference.
In male calves of the same age, on both sides but more strongly in Pistoia, a positive effect of the percentage of crops on body weight and chest circumference was demonstrated. According to the mathematical model, in Pistoia, male calves living in areas with 90% of crops would have an eviscerated weight approximately 10 kg higher than those in areas without crops. Only on the Pistoia side is demonstrated that the height at the withers and the length of the mandible also increase as the cultivation area grows. Even in male calves, the effect of meteorological events in the months preceding the harvest can be documented. The body weight is greater in the years in which spring and summer were rainier and cooler and in the years in which the previous winter had been milder (and the calf was just a fetus in the mother’s womb); less harsh winters also had a positive effect on the growth of the chest circumference and mandible.
Some results of our investigation were certainly expected, others seemed at least initially less obvious. There was no doubt that the body size of hinds and calves was always larger on the Emilian side of the Apennine, given the previous study on stags and given the characteristics of the Bologna landscape, with greater environmental diversity and therefore relatively richer in
resources. That on a finer scale the percentage of crops in the area around the sampling point could influence the body growth and size of the animals was imaginable: cultivated crops and fields, pastures, generally open areas offer relatively nutrient-rich food sources. But while for adult hinds the phenomenon was observable on both sides of the mountain, for yearling hinds and growing calves it was demonstrable only on the Pistoia slope. The explanation lies precisely in the rarity of cultivations in the Pistoia Apennine: here the cultivated areas can actually make a difference and determine very different fates between individuals that can enjoy them and animals that live far away from them. The preliminary study on mobility conducted by the University of Florence on 12 hinds equipped with radio-collars on the Apennine border ridge and on the Tuscan side made it possible to discover how females tend to use the fields more than expected based on the proportion of open crops inside their home ranges, as if the presence of small open areas was extremely important (Crocetti et al. 2010). While on the Emilian side, yearling hinds and calves that live both in areas with and without crops still have a more diversified and therefore richer territory at their disposal.
It was also very interesting to see how the climate during the spring and summer growing seasons affects the body weight and skeletal growth of the animals; this was demonstrable especially for calves and to a lesser extent on adult hinds. Rainier and cooler springs and summers increase the productivity of the vegetation and therefore trigger a greater milk production of the mothers which in turn favours faster growth of the young.
It was equally intriguing to see how the severity of the winter preceding birth affects the future body size of male calves. Lower temperatures experienced during the fetal stage produce effects on the growth of male offspring because the costs of gestation of a male offspring (larger than a female) are greater for a mother hind than those if she would have to bear a female daughter (Clutton-Brock et al. 1981, Landete-Castillejos et al. 2004).
This investigation allowed us to observe on a small scale the continuous working of phenotypic plasticity, the ability of the red deer species to modulate body growth depending on environmental and climatic conditions. The observed phenomena also allow us to look (with concern) to the future. If hinds and calves grow more in the presence of open environments and rainier and cooler springs and summers, what could happen if the current trend is one of further reduction in cultivated areas, progressive closure of the forest and shrub encroachment, as well as global warming? With the increase in temperatures and the reduction in rainfall, periods of drought will be increasingly common and prolonged, with increasingly fibrous and nutrient-poor vegetation, with evident consequences (also) on the red deer species, destined - given the same level of population density - to have slower body growth and reduced final size.
Management implications: The results of this research tell all of us, however, that we could take a different path, and if it is true that to challenge climate change the fight can only be global, it is also true that to counter the trend that red deer in the Apennines are facing it would be enough to introduce interventions in the area to improve environmental quality. With the strong decline of pastoralism and the abandonment of the mountains, woods and shrublands in the Apennines have almost doubled in size over the last 50-70 years and the open areas including agricultural crops have halved (Falcucci et al. 2007). And if the red deer of Alpine stock translocated to the Acquerino Forest initially still found a diversified landscape and were able to express much of their potential, after the first decades broom stands, bracken ferneries and woods in the upper Pistoia slope of the Apennine replaced the open areas, many of the fields once cultivated and pastures used by sheep farming. Furthermore, both on the Tuscan and Emilian sides, as the coppice woods age, they have increasingly closed the tree canopy with negative
consequences for the development of the undergrowth which requires light. And while in the Bologna side the environmental heterogeneity has nevertheless guaranteed the deer a respectable quantity and quality of trophic resources, in the Pistoia one the more compact structure of the woods and the scarce presence of open cultivated areas have certainly affected the overall suitability of the territory. When the red deer is defined as a forest species, a serious mistake is made: in reality it requires the co-occurrence of uneven-aged, non-compact woods and open prairie areas (Mattioli et al. 2022). Crops and pastures can somewhat replace - at least in part - the original grasslands, as demonstrated also in other parts of Europe (Mysterud et al. 2002, 2022). Hence the need for environmental improvements, above all (but not exclusively) on the Tuscan side: the thinning of even-aged and aged coppice woods everywhere and the creation of open areas, with reseeding and periodic mowing to keep the grass attractive (for a more detailed examination of the actions necessary see Mazzarone and Mattioli 1996, pages 108-118), as well as interventions that are quite demanding and all in all against the general trend but which could initially be applied even only to the existing protected areas. And if it is true that it will be almost impossible to see the majestic stags with the hyper-branched and palmated antlers of the 1970s and 80s again, it is also true that habitat improvements of this magnitude would lead not only to an enhancement in the conditions of the red deer (thanks precisely to the plasticity of species) but also more generally to an increase in overall biodiversity.



Abbazzi L. (1995) Occurrence of palmated Cervus elaphus from Italian late Pleistocene localities. Rendiconti Lincei 6(3): 189-206
Becciolini F., Bozzi R., Viliani M., Biffani S., Ponzetta M.P. (2016) Body measurements from selective hunting: biometric features of red deer (Cervus elaphus) from Northern Apennine, Italy. Italian Journal of Animal Science 15: 461-472
Bocci A., Monaco A., Brambilla P., Angelini I., Lovari S. (2009) Alternative strategies of space use of female red deer in a montainous habitat. Annales Zoologici Fennici 47: 57-67
Brown R.D. (1990) Nutrition and antler development. pp 426- 441 in: Bubenik G.A. e Bubenik A.B. (eds.) Horns, Pronghorns, and Antlers. Springer, New York
Ciccotti E. (2022) Studio preliminare su una popolazione di cervo (Cervus elaphus L. 1758) in Abruzzo e prospettive di gestione a livello regionale. Bachelor thesis, University of Florence
Clutton-Brock T.H., Albon S.D., Guinness F.E. (1981) Parental investment in male and female offspring in polyginous mammals. Nature 289: 487-489
Crocetti C., Ponzetta M.P., Minder I., Messeri A., Cervasio F. Argenti G., Maccelli S. (2010) Analysis of land utilisation by red deer in the Apennine mountains. pp 265-267 in: Schnyder et al. (eds.) Grassland in a changing world. EGF Kiel
De Marinis A.M. (2015) Valutazione dell’età nei Cervidi tramite esame della dentatura. Guida pratica all’identificazione delle classi d’età del cervo. Manuali e Linee Guida 90.2/2013:1-136 ISPRA
Falcucci A., Maiorano I., Boitani I. (2007) Changes in land-use/land-cover patterns in Italy and their implications for biodiversity conservation. Landscape Ecology 22: 617-631
Geist V. (1986) Super antlers and pre-world war II European research. Wildlife Society Bulletin 14: 91-94
Geist V. (1987) On speciation in Ice Age mammals, with special reference to cervids and caprids. Canadian Journal of Zoology 65: 1067-1084
Giovannelli F., Mori E., Zaccaroni M., Dessì-Fulgheri F., Baratti M. (2022) Genetic insights into an Apennine population of the Italian red deer. Mammal Research 67: 399-406
Huxley J.S. (1931) The relative size of antlers in deer. Proceedings of the Zoological Society of London 101: 819-864
Landete-Castillejos T., Gortazar C., Vicente J., Fierro V., Garcia A., Gallego L. (2004) Age-related foetal sex ratio bias in Iberian red deer (Cervus elaphus hispanicus): are male calves too expensive for growing mothers? Behavioral Ecology and Sociobiology 56: 1-8
Mattioli S. (1994) Le reintroduzioni di Cervidi in Italia, con particolare riferimento all’Appennino”, pp 19-25 in: Inglisa M. (ed.) “La fauna appenninica e la sua conservazione” Atti del Convegno Nazionale Paolo Barrasso, Airone-Corpo Forestale dello Stato
Mattioli S., Meneguz P.G., Brugnoli A. e Nicoloso S. (2001) Red deer in Italy: recent changes in distribution and numbers. Hystrix, Italian Journal of Mammalogy 12: 21-35
Mattioli S., Ferretti F., Nicoloso S., Corlatti L. (2021) Spatial variation in antler investment of Apennine red deer. Ecology and Evolution 11: 7850-7864
Mattioli S., Zachos F., Rossi L., Lister A., Corlatti L. (2022) Red deer Cervus elaphus. Linnaeus, 1758. pp 51-86 in: Corlatti L. e Zachos F. (eds.) Terrestrial Cetartiodactyla. Handbook of the Mammals of Europe. Springer Nature
Mazzarone V., Siemoni N., Lovari C., Pedone P., Mattioli L. (1991) A method of red deer (Cervus elaphus L.) census during the roaring period in a forested area of the northern Apennines. pp 140145 in: Csanyi S. and Ernhaft J. (eds.) Transactions of the 20th I.U.G.B. Congress, Gödöllő
Mazzarone V. and Mattioli S., eds. (1996) Indagine sulla popolazione di cervo dell’Acquerino. Relazione finale 1993-1995. Regione Toscana, Firenze 1-133
Mysterud A., Langvatn R., Yoccoz N.G., Stenseth N.C. (2002) Large-scale habitat variability, delayed density effects and red deer population in Norway. Journal of Animal Ecology 71: 569-580
Mysterud A., Langvatn R., Meisingset E.L., Rivrud I.M. (2022) Agricultural grasslands buffer density effects in red deer populations. Journal of Wildlife Management 87: e22357, 1-14
Palma di Cesnola A. (1988) Paglicci, Rignano Garganico. Mostra iconografica permanente. Regione Puglia 1-88
Pavan G. e Mazzoldi P. (1983) Banca dati della distribuzione geografica di 22 specie di Mammiferi in Italia. Collana Verde 66, Ministero dell’Agricoltura e Foreste, Roma 1-279
Pedrotti L. Dupré E., Preatoni D., Toso S. (2001) Banca dati Ungulati. Biologia e Conservazione della Fauna 109: 1-132
Premuda G. (1975) Origine e consistenza attuale del patrimonio faunistico della montagna pistoiese. Diana 11: 28-31
Raganella Pelliccioni E., Riga F., Toso S. (2013) Linee guida per la gestione degli Ungulati: Cervidi e Bovidi. Manuali e Linee Guida ISPRA 91: 1-225
Reulecke K. (1987) Zur Geweihentwicklung der Hirsche im Harz. Wild und Hund 24/87: 41-45
Szunyoghy J. (1959) The variability of the branch numbers od stag antlers in the Carpathian Basin. Annales Historico-Naturales Musei Nationalis Hungarici 51: 489-492
Vannini C., Fattorini N., Mattioli S., Nicoloso S., Ferretti F. (2021) Land cover and weather jointly predict indicators of phenotypic quality in a large herbivore.
Ecological Indicators 128: 107818, 1-11
Vogt F. (1948) Das Rotwild. Oesterreicher Jagd- und Fischereiverlag, Vienna 1-207
Wilkens B. (1989) Il cervo dal Mesolitico all’Età del Bronzo nell’Italia centro-meridionale.
Rassegna di Archeologia 8: 63-95
I sincerely thank the Italian Delegation of the C.I.C. for inviting me to write this text on the origin and development of the Acquerino red deer population and on the recent scientific papers produced by our research group. I am very grateful to A. Dal Pian for making his beautiful photos available. My gratitude also goes to colleagues and friends L. Corlatti, N. Fattorini, F. Ferretti, S. Nicoloso and C. Vannini, co-authors of the two recent articles. L. Corlatti and N. Fattorini kindly revised this text. It is also right to mention my companions from the first investigation of 1993-95 V. Mazzarone, C. Lovari, N. Siemoni I. Aiardi, S. Gualazzi and P. Pedone. S. Csanyi, of the Szent István University of Gödöllő (Hungary) kindly made unpublished data available to me on Hungarian red deer trophies. Finally, I would like to mention P. Agnelli, former curator of Mammals at the La Specola Natural History Museum of the University of Florence, for his efforts in having the hyper-branched and palmated antler acquired by the museum itself. H. Puleston (The Zoological Society of London) kindly reviewed parts of my English version.