
DELEGAZIONE ITALIANA DEL CONSIGLIO INTERNAZIONALE
DELLA CACCIA E DELLA SALVAGUARDIA DELLA FAUNA

DELEGAZIONE ITALIANA DEL CONSIGLIO INTERNAZIONALE
DELLA CACCIA E DELLA SALVAGUARDIA DELLA FAUNA
Stefano Mattioli
Impaginazione e grafica: Elisa Bossotto Stampato in Italia presso Tipolitografia Botalla s.r.l. - Gaglianico (BI)
Contributo fotografico: Andrea Dal Pian
© 2024 Delegazione Italiana del CIC
Tutti i diritti riservati. Nessuna parte di questa pubblicazione può essere riprodotta, archiviata o trasmessa in qualsiasi forma o con qualsiasi mezzo, elettronico, meccanico, fotocopia, registrazione o altro senza la previa autorizzazione sia del proprietario del copyright che dell’editore di questo libro. Ciò include il testo e le immagini di questo manuale che non possono essere riprodotti in alcuna forma senza il previo consenso scritto della Delegazione Italiana del CIC.
La riproduzione e l’utilizzo non autorizzati comporteranno procedimenti legali.

Premessa del Capo Delegazione Italiana del CIC Nicolò Amosso........................
Prefazione di Stefano Mattioli ................................................................................
Cervi in Appennino.................................................................................................
Riferimenti Bibliografici...........................................................................................
Ringraziamenti......................................................................................................... pag. 4 pag. 5 pag. 7 pag. 50 pag. 52
Tra le finalità della Delegazione Italiana del CIC sempre più centrale è la tutela dell’ambiente, della biodiversità e degli ecosistemi nell’interesse delle future generazioni supportando e contribuendo alla divulgazione della ricerca scientifica applicata.
In quest’ottica, dopo la pubblicazione della “Guida Pratica al Manuale CIC per la Misurazione e la Valutazione dei Trofei di Caccia”, del “Il Cervo Sardo”, del “Il Camoscio Appenninico” è ora la volta di “Cervi in Appennino, ovvero la plasticità in azione”.
Quest’ultimo studio prende spunto da una interessante conferenza che il dott. Stefano Mattioli tenne in occasione del conseguimento dell’Attestato di Eccellenza, riconoscimento che la nostra Delegazione assegna a chi si è distinto particolarmente nella difesa, studio o altro a favore della natura e della caccia.
Nell’intervento lo studioso spiegò l’origine della popolazione di cervi dell’Acquerino, il periodo dei super-trofei palmati, il periodo della “normalizzazione” dei palchi, i primi censimenti al bramito e i diversi accrescimenti corporei registrati sui due versanti appenninici quello toscano e quello emiliano.
Questa pubblicazione mette in mostra la grande versatilità del cervo rispetto alle variazioni climatiche e alimentari in termini di crescita corporea e investimento nel palco, utilizzabile quest’ultimo come indice dello stato di salute della popolazione.
Ringrazio Stefano Mattioli, zoologo impegnato sia nella gestione diretta degli ungulati sia nella ricerca, autore di numerose pubblicazioni scientifiche e divulgative, docente nonchè CIC Expert della nostra Delegazione per il Suo importante contributo scientifico prima nello studio sul “Cervo Sardo” e ora con “Cervi in Appennino, ovvero la plasticità in azione”.
Un ringraziamento particolare ad Andrea Dal Pian per lo splendido contributo fotografico che accompagna lo studio.

Su invito della delegazione italiana del C.I.C. nelle pagine che seguono racconto la storia di uno dei primi nuclei di cervo reintrodotti nell’Appennino Settentrionale, noto soprattutto nei primi tempi come la popolazione dell’Acquerino, dal nome della stazione della Forestale dove vennero effettuate le liberazioni degli esemplari fondatori. Un racconto che ha permesso di ripercorrere più di sessant’anni, seguendo lo sviluppo della popolazione in termini di areale e consistenza nello sfondo di grandi modificazioni dell’economia e del paesaggio della montagna appenninica. Attraverso l’analisi delle dimensioni e della morfologia dei palchi di cervo è stato possibile anche ricostruire i cambiamenti fisici dei maschi nel corso dei decenni. La reintroduzione di cervi alpini di taglia modesta e di palchi dalla struttura semplificata in ambienti appenninici ricchi di risorse trofiche privi di competizione permise agli animali di prosperare e “fiorire”, con la produzione negli anni Settanta e Ottanta di palchi davvero peculiari per architettura. Con l’inizio del prelievo venatorio nel 2000 si è potuto monitorare il rendimento fisico della popolazione in modo più dettagliato e mettere in relazione i dati somatici con i cambiamenti ambientali in senso lato, confermando la grande plasticità del cervo, una specie in grado di rispondere velocemente alle variazioni delle condizioni climatiche e delle fonti alimentari. E’ stata anche l’occasione per raccontare le prime indagini iniziate nel 1993 (analisi dei palchi, ricostruzione degli areali, stime di consistenza mediante censimenti al bramito) e i risultati più eclatanti degli studi più recenti (accrescimento corporeo dei cervi maschi sui due versanti della catena appenninica, investimento nel palco, interazioni tra taglia, clima e ambiente nelle femmine e nei piccoli). Ho cercato infine di dimostrare che questi studi apparentemente di stretto interesse scientifico in realtà possono avere ricadute pratiche nella gestione faunistica e del paesaggio.
Voglio dedicare questo libro a Franco Odorici (1942-2018), un amico gentile e generoso, cacciatore di selezione, per anni responsabile di un distretto di caccia, rilevatore biometrico entusiasta, appassionato di trofei e trofeistica, tra i protagonisti della gestione del cervo nell’Appennino bolognese.


Cervi in Appennino ovvero la plasticità in azione
In ogni specie gli animali interagiscono con le risorse alimentari aggiustando le dimensioni corporee a seconda della produttività dell’ambiente e cioè del numero di prede per i carnivori, della quantità e qualità delle piante per gli erbivori. Il cervo è uno degli ungulati più adattabili in termini di taglia corporea, una caratteristica nota ai biologi come “plasticità fenotipica”. Di norma in Europa centrale e orientale un maschio adulto può pesare dopo gli amori in media dai 160 ai 220 kg, mentre una femmina adulta in media dai 90 ai 120 kg. Ma in ambienti marginali come la macchia mediterranea (per es. Sardegna), i boschi su terreno sabbioso come la Mesola o le brughiere scozzesi, i cervi pur di sopravvivere tendono a rimpicciolire, con maschi adulti in media di 95-110 kg e femmine di 60-80 kg. E d’altra parte in ambienti particolarmente ricchi nei Carpazi o nella pianura pannonica sono documentati casi di cervi di anche 340-350 kg di peso post-riproduttivo e femmine di anche 150-160 kg. E se dubitassimo dell’incredibile flessibilità dimensionale del cervo, ricordiamoci che nel Pleistocene Superiore nell’Isola di Jersey nel Canale della Manica vivevano cervi nobili miniatura di circa 36 kg e nelle isole di Malta cervi di appena 25 kg (Mattioli et al. 2022). Difficile immaginare dei cervi della taglia di un capriolo, ma fu proprio così.
Il famoso pioniere dell’etologia di campagna V. Geist, grande esperto di caprini e Cervidi, fu tra i primi a descrivere come le specie possano fluttuare tra i due estremi dei cosiddetti “fenotipi di mantenimento o di efficienza” (cioè ecotipi frugali adattati ad ambienti poco produttivi) e dei “fenotipi lussureggianti o di dispersione” (ecotipi di grossa taglia adattati ad ambienti con abbondanza di risorse) (Geist 1987). E già J. Huxley (1931) aveva riferito dei piccoli cervi delle brughiere scozzesi presto diventati di enormi dimensioni nei vasti e ricchi pascoli della Nuova Zelanda dove erano stati traslocati nella seconda metà dell’Ottocento. Dobbiamo quindi pensare sempre al cervo come una specie che risponde in maniera rapida e consistente alle variazioni delle risorse trofiche e climatiche adattando la propria taglia.
Gran parte delle popolazioni odierne europee di cervo sono state in qualche modo “forgiate” dai cambiamenti ambientali di origine antropica. Se oggi i cervi scozzesi sono molto spesso di piccole dimensioni, ciò è dovuto principalmente ai profondi mutamenti di paesaggio causati dall’uomo fin da 3000-4000 anni fa, quando nella prima Età del Bronzo le vaste foreste caledoni furono via via smantellate lasciando il posto a brughiere. E se in Europa centrale prevalgono cervi di medie dimensioni con palchi relativamente modesti lo si deve ai cambiamenti apportati nella gestione forestale nella seconda metà del Diciottesimo secolo, quando l’uomo preferì sostituire farnie, aceri e faggi con impianti coetanei di conifere a rapida crescita, creando habitat più poveri per i cervi. Una visita al castello barocco di Moritzburg presso Dresda, in Sassonia (Germania), ci consente di ammirare trofei giganteschi di anche 20 kg risalenti alla fine del Diciasettesimo e gli inizi del Diciottesimo secolo, testimoni di come potevano essere i cervi prima che l’uomo intervenisse pesantemente sull’ambiente. E nelle cronache di quei tempi si legge di cervi maschi che nella stessa Sassonia e in altre regioni d’Europa Centrale raggiungevano pesi pre-riproduttivi di 400-450 kg (cfr Geist 1986).
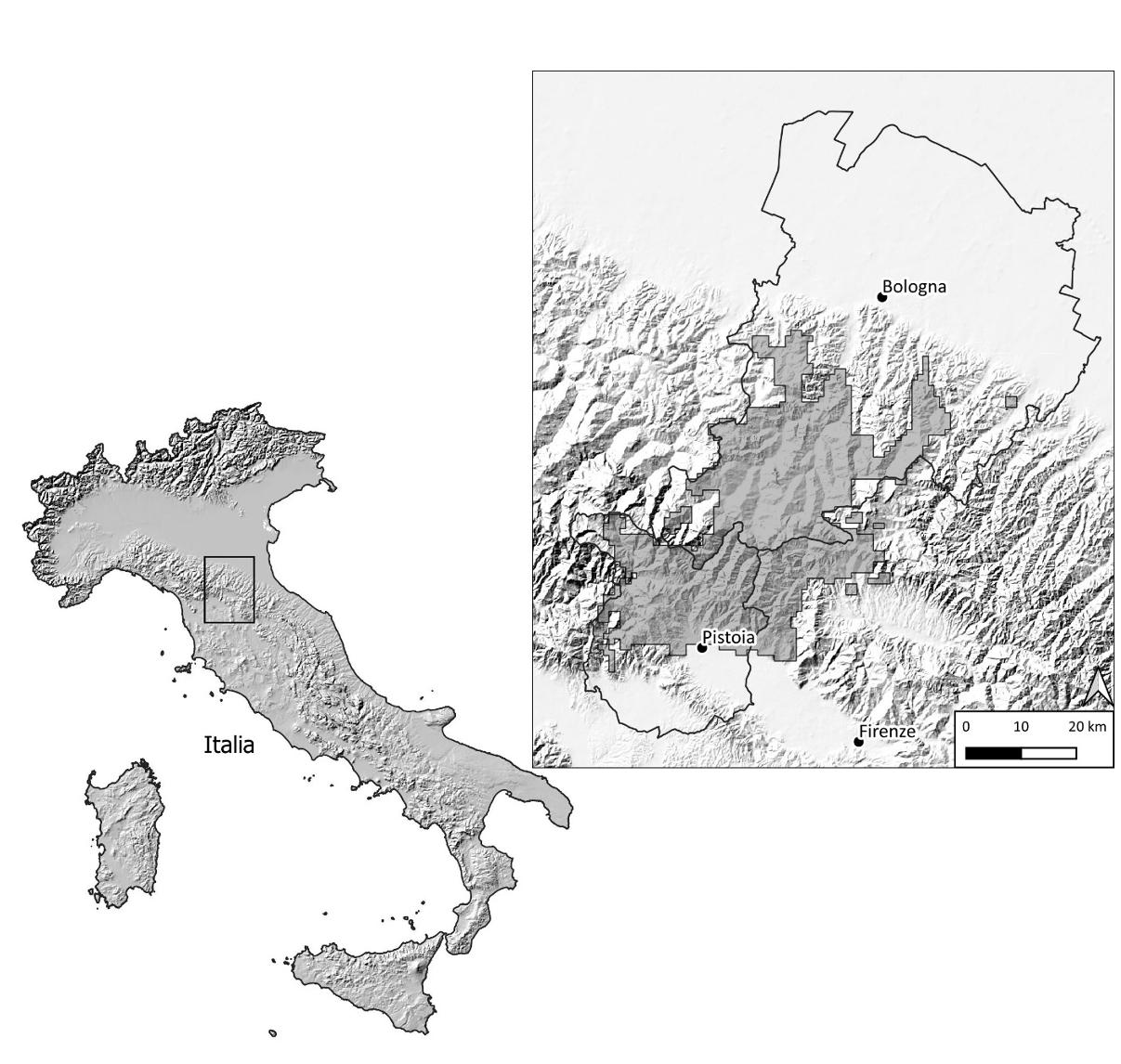
Origine del nucleo di cervi dell’Acquerino
Alla fine dell’ultimo conflitto mondiale il tratto di Appennino tra Pistoia, Firenze e Bologna era molto diverso da come appare oggi. La copertura boscata era molto più limitata e mostrava tutti i segni del sovrasfruttamento. I boschi cedui avevano per decenni subito cicli di taglio ravvicinati e spesso sembravano più arbusteti che boschi. Molte aree erano brulle per i disboscamenti e il sovrapascolo di ovini e bovini. Sul versante bolognese c’erano più aree aperte coltivate a cereali e foraggere mentre sul versante toscano c’era una economia agricola di sussistenza con campi e campetti a segale, avena, grano e foraggere in mezzo alle aree boscate. La fauna selvatica da tempo si era estremamente impoverita: i grossi erbivori (cinghiale, cervo, capriolo) erano scomparsi addirittura già dalla fine del XVIII secolo, e i predatori (soprattutto lupo, gatto selvatico, martora, lontra) erano quasi estinti o rari. Il Corpo Forestale dello Stato fu chiamato ad un immane sforzo per ripristinare e proteggere i boschi appenninici: i cedui furono riconvertiti ad alto fusto, molte aree aperte furono rimboschite. L’amministratore delle Foreste Demaniali di Pistoia G. Premuda, arrivato nel 1950, non si fermò alla gestione selvicolturale dell’Appennino, ma si preoccupò di rivitalizzare quei boschi attraverso reintroduzioni e introduzioni. Nel 1952 chiese ufficialmente il permesso alla direzione dell’Azienda di Stato per le Foreste Demaniali a Roma di pianificare liberazioni di parecchie specie. Con determinazione ed entusiasmo accompagnati da approssimazione, l’amministratore fece costruire recinti e liberare cinghiali, cervi, caprioli, lepri, starne, ma anche specie esotiche come daini, mufloni, marmotte, galli forcelli pur di arricchire quelle montagne di fauna selvatica (Premuda 1975). Ingenuità ed errori comprensibili se inseriti in quell’epoca pionieristica, priva di punti di riferimento per quanto riguarda la gestione faunistica, con il mondo accademico italiano completamente assente.
Nel 1958 furono liberati presso la stazione forestale dell’Acquerino, nell’alto Pistoiese, 4 cervi (un maschio adulto, due femmine e un piccolo) provenienti dalle Foreste Demaniali di Tarvisio, nelle Alpi Friulane, dove l’ispettore aveva lavorato da giovane, e altri 3 esemplari sempre da Tarvisio furono aggiunti nel 1965 dal suo successore Pettinà (Mazzarone e Mattioli 1996). I forestali di Tarvisio avevano un’area recintata con un nucleo di cervi, regalati dai colleghi austriaci della vicina Carinzia. In tutto quindi 7 cervi delle Alpi orientali, adattati a vivere in una delle zone a clima più rigido d’Italia, con inverni lunghi e nevosi, primavere brevi e fredde, furono trasferiti in un’area dell’Appennino settentrionale dal clima più mite, con inverni più brevi, meno nevosi e dalle temperature meno estreme, passando inoltre da un ambiente a conifere a foreste piuttosto aperte a prevalenza di caducifoglie, allora con molti giovani rimboschimenti. Non possiamo sapere quanto pesassero allora i cervi che vivevano a Tarvisio, ma conosciamo i pesi medi nella stessa area in tempi più recenti, pochi anni fa: appena 148 kg per i maschi adulti e 86 kg per le femmine, valori piuttosto modesti anche per l’arco alpino (Mattioli et al. 2022). Gli inverni rigidi mettono alla prova gli animali, che devono cercare di muoversi poco per consumare il meno possibile le riserve di grasso accumulate, che devono spendere parecchie energie per mantenere inalterata la propria temperatura corporea; inoltre le primavere fredde e piovose rischiano di influire negativamente sulla sopravvivenza e l’accrescimento corporeo dei nuovi nati. I boschi di conifere inoltre offrono poche risorse alimentari e gli stessi aghi di abete sono poco nutrienti e digeribili. Per quei primi fondatori alpini il trasferimento nelle foreste demaniali pistoiesi rappresentò sicuramente un netto miglioramento delle condizioni fisiche, in un ambiente ricco di fonti trofiche e con clima decisamente meno severo (cfr tabb. 1 e 2). La reintroduzione dei cervi all’Acquerino non fu l’unico caso nell’Appennino settentrionale: nel periodo 1950-60 i Forestali immisero la specie in Casentino e nel 1966-72 all’Orecchiella, nell’Alto Lucchese (Mattioli 1994, Mattioli et al. 2001).
Tab. 1: Dati medi meteorologici della stazione di Tarvisio (UD), 780 m slm
Tab. 2: Dati medi meteorologici della stazione dell’Acquerino (PT), 890 m slm
Dati del Servizio Meteorologico dell’Aeronautica Militare.
Non sappiamo come il passaggio dalle Alpi all’Appennino con il conseguente miglioramento di condizioni climatiche e ambientali si concretizzò in termini di pesi corporei, anche perché la popolazione dell’Acquerino rimase protetta fino al 1999 e quindi priva dei monitoraggi biometrici tipici della gestione venatoria. Per ben quattro decenni possiamo seguire le condizioni fisiche di questo nucleo di cervi solo indirettamente, attraverso i palchi caduti e raccolti e i trofei di animali morti recuperati. Le dimensioni e alcune caratteristiche dei palchi rispecchiano la taglia e lo stato fisico di chi li produce: cervi maschi in salute, che vivono in ambienti più ricchi, crescono di più e possono investire di più nel palco, mentre cervi in difficoltà, che abitano in ambienti poco produttivi, tendono ad avere taglia minore e a investire meno nel palco. Il palco rappresenta quindi un buon indice dello stato di salute complessivo di una popolazione, per tutti i Cervidi (Brown 1990).
I cervi maschi tarvisiani, oltre ad avere una taglia modesta hanno palchi di dimensioni e peso altrettanto modesti, con una struttura piuttosto essenziale. I cervi traslocati all’Acquerino e i loro diretti discendenti hanno subito prodotto palchi di dimensioni notevoli e dalla struttura complessa, il che ci permette di dedurre che, grazie alla maggior ricchezza degli ambienti appenninici e al clima più favorevole rispetto alle Alpi orientali, gli animali avevano presto raggiunto condizioni fisiche di tutto rispetto. E tutto questo grazie alle capacità della specie di rispondere prontamente ai fattori ambientali.
Di questi palchi grandi e fuori dall’ordinario si favoleggiava per decenni e finalmente, dal 1993 e il 1995 alcuni tecnici della società DREAm e il sottoscritto fummo chiamati dalle Regioni EmiliaRomagna e Toscana a svolgere uno primo studio sulla popolazione di cervi dell’Acquerino. Un’indagine ad ampio raggio che doveva documentare le sue caratteristiche, ricostruire l’areale di distribuzione e stimare la consistenza. Tra i miei compiti c’era quello di raccogliere dati morfometrici sui palchi (cioè misurandoli e annotandone la conformazione), andando a visitare i collezionisti locali per misurare un campione sufficientemente ampio da “caratterizzare” la popolazione, cioè da descriverne in modo rappresentativo i tratti peculiari: un compito non sempre facile perché alcuni dei raccoglitori più accaniti erano gelosi dei palchi raccolti e quindi non particolarmente felici di mostrarli ad estranei.
Furono così esaminate 211 stanghe (60 coppie e 91 stanghe spaiate), delle quali 171 appartenenti ad esemplari adulti (cioè di 5 o più anni), raccolte prevalentemente nelle province di Prato e Pistoia e risalenti ad un arco temporale piuttosto ampio, compreso tra il 1975 e il 1995 (tab. 3).
Tab. 3: Caratteristiche biometriche del campione di cervi adulti dell’Appennino tosco-emiliano (media e deviazione standard) (da Mazzarone e Mattioli 1996)
Lunghezza stanga
Lunghezza oculare
5,0 168
Lunghezza ago 17,7 9,3 110
Lunghezza mediano
Circonferenza rosa
Tab. 4: Confronto tra le lunghezze medie della stanga negli adulti di cervo in alcune popolazioni europee, ordinate in modo crescente (Mattioli et al. 2022, Ciccotti 2022)
I. di Rum (UK)
Norvegia
Sierra Morena (Spagna)
Harz (Germania)
Alpi piemontesi
Masuria (Polonia)
Appennino settentrionale (PO)
Carpazi polacchi 88,6
Appennino settentrionale (PT + BO)
Baranja (Croazia)
Appennino abruzzese
Potendo confrontare i valori medi e la variabilità delle stanghe di cervi dell’Appennino con quelli di altre popolazioni europee, le dimensioni dei palchi appenninici dell’Acquerino risultarono su scala continentale medio-alte, certamente superiori a quelle dei palchi di aree marginali come le brughiere scozzesi e la macchia mediterranea (Rum, Sardegna, Spagna) ma anche dei palchi centro-europei conosciuti (tab. 4). Il trasferimento dalle Alpi orientali aveva sicuramente favorito nei cervi un aumento di taglia (allora ancora non documentabile) e un maggiore investimento nel palco (fortunatamente documentato con quella prima indagine).
Ma l’interesse dei palchi dell’Acquerino di quel periodo risiede non tanto nelle considerevoli dimensioni ma nella conformazione. Innanzitutto la corona terminale e l’ago sono tratti molto comuni, che mancano solo rispettivamente nel 18% e nel 17,5% delle stanghe degli adulti. Pur essendo due caratteristiche peculiari del cervo nobile, in molte popolazioni europee corona e ago sono tutt’altro che frequenti. Più in generale i palchi del campione appenninico tendono ad avere un numero piuttosto elevato di ramificazioni, con valori medi di 13 punte totali negli adulti e casi estremi addirittura eccezionali. Diverse stanghe, soprattutto risalenti al periodo 1975-1985, hanno più di 10 punte con massimi registrati di 12, 13 e 16 punte, che rendevano possibili in quell’epoca palchi a 24, 25, 26 (osservati, vedi foto 2-4) e addirittura a 32 punte (non documentato). La stanga a 16 punte, data la sua straordinarietà, è stata recentemente acquisita dal Museo di Storia Naturale La Specola di Firenze che la espone nel Salone degli Scheletri (foto 5-6). La sovraproduzione di punte talvolta non interessava solo la corona terminale che appariva “arborizzata” ma coinvolgeva anche punte della metà inferiore della stanga come l’oculare, l’ago e il mediano che in rari casi potevano essere bifidi (foto 2-6). La stanga a 16 punte, ritrovata intorno al 1980, ha ben 12 punte di corona e un mediano biforcuto.
Per dare un’idea concreta dell’eccezionalità di queste iper-ramificazioni, possono servire alcuni confronti con altre aree d’Europa. In Ungheria, su 28.790 trofei esaminati tra il 1884 e il 1957, solo 3 raggiungevano le 26-28 punte (Szunyoghy 1959). Su 17.731 cervi maschi abbattuti nell’Harz (Germania) tra il 1959 e il 1985, solo 2 possedevano palchi a 21 cime totali e nessuno aveva palchi di 22 o più cime (Reulecke 1987). In epoche più recenti su 195.358 trofei di cervi adulti esaminati e valutati tra il 1990 e il 2020 in Ungheria, 5 sono arrivati a 29-35 punte totali; nei 10 trofei più ramificati 7 stanghe avevano da 16 a 22 cime (S. Csanyi, comunicazione personale).
Ma oltre che ad essere iper-ramificati, i palchi dell’Acquerino di quelli anni avevano una tendenza alla palmatura, cioè a punte collegate tra loro a formare vistose parti appiattite che si accompagnano all’arborizzazione della porzione apicale delle stanghe (foto 2-8). Il fenomeno è molto raramente documentato per altre popolazioni europee, ma in questo caso interessava il 10% del nostro campione di stanghe dei cervi adulti dell’Acquerino; in realtà, dato che parecchie stanghe risalivano ad anni più recenti, è possibile che la frequenza della palmatura negli “anni d’oro” fosse più elevata.
Quindi, soprattutto tra il 1975 e il 1985, i cervi maschi di questo nucleo appenninico, soprattutto quelli della seconda e terza generazione, forse solo 12-30 esemplari, approfittando della bassa densità, dell’ambiente ricco di risorse e del clima mite, mostrarono la loro esuberanza, la loro florida salute, tendendo a produrre palchi lussureggianti, che privilegiavano il numero di ramificazioni e la palmatura al peso e alla lunghezza delle stanghe, palchi in qualche modo più vistosi che imponenti. Altre popolazioni reintrodotte in Appennino, come quella fondata negli anni Settanta
nel Parco Nazionale d’Abruzzo, nel mostrare tutta la propria vitalità hanno puntato più su palchi lunghi e pesanti, non sperimentando in alcun modo l’iper-ramificazione e solo raramente la palmatura (Ciccotti 2022). Molto difficile risulta trovare una spiegazione soddisfacente per questa differenza di risposta tra i due nuclei reintrodotti; forse hanno inciso le caratteristiche genetiche di quel piccolo gruppo di cervi capostipiti (il cosiddetto “effetto fondatore”), che -non più frenate dai rigori alpini- finalmente in condizioni ambientali ottimali hanno potuto esprimersi appieno e anzi si sono amplificate. D’altra parte i grandi palchi del Parco d’Abruzzo hanno probabilmente potuto approfittare non solo della compresenza di vaste praterie e foreste ma anche della diversa dinamica delle reintroduzioni, con un elevato numero di fondatori provenienti da molte fonti, in qualche modo una miscela vincente (cfr Mattioli et al. 2001).
L’iper-ramificazione dei palchi è un fenomeno piuttosto raro: anche nei famosi esperimenti di foraggiamento artificiale di F. Vogt degli anni 1927-1941 (presso Sněžnik, nella cosiddetta Svizzera Boema, nell’attuale Cechia) i cervi, sottoposti ad alimentazione con prodotti ad alto contenuto proteico e minerale, ebbero un sorprendente aumento della taglia e delle dimensioni dei palchi, con pesi pre-riproduttivi fino ai 350 kg e trofei fino ai 14 kg, ma con un numero massimo di 20-22 punte totali (Vogt 1948, Geist 1986).
Quanto alla palmatura, se è vero che in epoca attuale è molto rara, in epoche lontane era sicuramente più comune. La presenza in Italia di cervi con palchi grandi e palmati tra il Tardo Pleistocene e la prima metà dell’Olocene (fino all’Età del Bronzo 4000-3000 anni fa) (Abbazzi 1995, Wilkens 1989) fece addirittura pensare al paleontologo G. De Stefano all’esistenza di una sottospecie particolare che nel 1911 chiamò Cervus elaphus palmidactyloceros. Un graffito su frammento di calcare rinvenuto nell’atrio della Grotta Paglicci, presso Rignano Garganico in Puglia ritrae la testa di un cervo dal maestoso palco palmato e ricco di punte terminali che viveva circa 15 000 anni fa (Palma di Cesnola 1988, fig. 34 a pagina 67). In realtà, come si è visto all’Acquerino, la formazione di palchi palmati può semplicemente indicare basse densità di popolazione e grandi disponibilità alimentari di qualità. I resti fossili e le raffigurazioni rupestri di cervi con palchi palmati forse fotografano momenti favorevoli ricchi di fonti trofiche, come animali che hanno da poco colonizzato ambienti caratterizzati da abbondanza di cibo e nessuna competizione di altri erbivori. Una lezione utile anche ai paleontologi, che non devono necessariamente pensare a ceppi o a sottospecie o a “cronospecie” particolari per spiegare resti di cervi iper-ramificati e palmati, ma più semplicemente a periodi molto propizi per clima e risorse trofiche in presenza di basse densità di popolazione. Ma all’Acquerino quella stagione di palchi complessi e palmati fu un periodo destinato a terminare.







Già quando iniziammo la nostra indagine su quel tratto di Appennino conformazioni del genere stavano gradualmente diventando sempre più rare. In realtà nulla di sorprendente. Se i palchi sono in qualche modo lo specchio delle condizioni ambientali, senza che noi ci rendessimo conto di quanto stesse accadendo, l’Appennino tosco-emiliano stava diventando un ambiente un po’ meno ospitale per i cervi. Il superamento della soglia di densità di 1-1,5 capi per kmq (relativamente bassa, ma un valore evidentemente sensibile per la specie), la progressiva chiusura dei boschi cedui non avviati ad alto fusto, e sul lato toscano la fine definitiva dell’agricoltura di sussistenza con l’abbandono dei campetti coltivati e la conseguente conquista dei ginestreti e dei felceti, cominciarono sottilmente ad influire sul vigore dei cervi e quindi anche sui palchi. I palchi non erano più quelli del periodo d’oro tra il 1975 e il 1985, ma ancora non era facile accorgersene perché i valori medi erano sostanzialmente simili: solo le stanghe iper-ramificate o palmate erano diventate sempre più rare. Si può dire che negli anni Novanta le condizioni dei cervi tra Firenze, Prato, Pistoia e Bologna, erano andate “normalizzandosi”, cioè densità e ambiente avevano portato ad eliminare i casi più estremi, i palchi più vistosi ed elaborati.
Due versanti, due diversi accrescimenti corporei
Nel 2000 le due Regioni Emilia-Romagna e Toscana avviarono per la prima volta il prelievo selettivo del cervo e quindi fu possibile mettere a punto un monitoraggio biometrico approfondito sia delle dimensioni corporee di tutti gli animali sia dei palchi dei maschi. E così ci si poté rendere conto che la situazione si stava evolvendo ulteriormente. Mentre sul lato toscano dell’Appennino i cambiamenti ambientali si facevano più evidenti con la progressiva scomparsa degli antichi campi, sul lato emiliano la compresenza di boschi, boschetti, arbusteti, pascoli e campi garantiva ancora una buona diversificazione delle risorse trofiche. Il monitoraggio quindi permise per la prima volta di distinguere tra le caratteristiche morfometriche dei cervi del lato toscano da quelle del lato bolognese della catena montuosa. Come ci si poteva aspettare da un animale così plastico come il cervo, la specie rispondeva prontamente ai cambiamenti ambientali e l’ambiente ne modulava l’accrescimento somatico e l’investimento del palco. Bastava nascere e abitare su un versante dello stesso rilievo per tendere ad avere una certa crescita corporea o un’altra, un tipo di palco o un altro: nessuna differenza eclatante ma sicuramente due percorsi non più sovrapponibili come nei primi decenni.
Tab. 5: Pesi medi corporei e del palco nei cervi dei due versanti dell’Appennino
(da Mattioli et al. 2021, Vannini et al. 2021, basato sui dati relativi a 1565 maschi giovani, subadulti e adulti raccolti tra il 2000 e il 2020 e a 2562 piccoli e femmine raccolti tra il 2000 e il 2017. Pesi pieni di femmine e piccoli ricostruiti a partire dal peso eviscerato)
variabili (valori arrotondati)
Bologna Pistoia
Peso corporeo medio dei maschi adulti (kg) 195 170
Peso corporeo medio delle femmine adulte (kg) 111 102
Peso corporeo medio dei maschi piccoli (kg) 71 60
Peso corporeo medio delle femmine piccole (kg) 64 51
Peso medio del palco (kg) 4,7 3,8
Peso medio del trofeo (kg) 6,1 5,1
In media i maschi adulti che vivono sul versante bolognese pesano circa il 15% in più dei maschi adulti del versante pistoiese, le femmine adulte il 9% in più, i maschi piccoli il 18% in più, i piccoli di sesso femminile il 26% in più (tab. 5). Pesi corporei simili a quelli pistoiesi vengono rilevati anche nel vicino Pratese (Becciolini et al. 2016). I palchi dei cervi sul lato emiliano in media pesano circa il 20% in più di quelli pistoiesi. Dato che le condizioni ambientali forgiano specie plastiche come il cervo, i cambiamenti occorsi negli ultimi decenni del Ventesimo secolo agli habitat appenninici in maniera diseguale sui due versanti hanno finito per incidere in modo differenziato sull’accrescimento e le dimensioni finali dei cervi delle due regioni. Nei palchi le iper-ramificazioni e le palmature si sono fatte rarissime, soprattutto sul versante toscano. Le stanghe con più di 9 punte, che nel campione del 1970-94 (n = 171) rappresentavano il 5,3%, nel periodo 2000-2020 hanno costituito appena l’1% (n = 1425; dato inedito), e in realtà erano stanghe al massimo di 11 punte e del tutto assenti sul lato pistoiese.
Quando iniziarono i primi prelievi venatori sperimentali nel 2000 pianificammo un monitoraggio biometrico estremamente approfondito, che ha prodotto una banca dati senza uguali in Europa. I dati raccolti comprendono i pesi corporei, diverse misure lineari somatiche esterne (come ad esempio la lunghezza testa-tronco e l’altezza al garrese), parecchie misure craniometriche (come la lunghezza condilo-basale e la lunghezza della mandibola) più per i maschi le misure dei palchi (peso del trofeo e misure lineari previste dalla formula CIC). I pesi corporei e le misure lineari esterne vengono registrati dai rilevamenti biometrici, cacciatori di selezione abilitati dopo un corso specifico e un esame a misurare gli esemplari abbattuti in appositi centri di controllo. Le misure del cranio sono state rilevate dal tecnico faunistico, mentre le misure lineari dei trofei sono state effettuate in prima istanza dai rilevatori biometrici e, per i casi più importanti, per lo meno in provincia di Bologna, ripetute dai valutatori CIC qualche settimana prima della mostra dei trofei. Tutti i trofei sono stati pesati almeno tre mesi dopo l’abbattimento come previsto dal CIC. L’età di ciascun esemplare è stata stimata dal tecnico faunistico sulla base dei ritmi medi di eruzione, sostituzione e usura dei denti, facendo riferimento ad un atlante fotografico (De Marinis 2013) costruito sulla base dell’analisi di microsezioni di denti di un sottocampione proveniente dall’Appennino pistoiese.
Evoluzione dell’areale e della consistenza
Una volta reintrodotto presso la stazione forestale dell’Acquerino il piccolissimo nucleo di fondatori dovette attendere qualche anno per crescere ed insediarsi stabilmente nelle foreste demaniali pistoiesi. Probabilmente i cervi traevano vantaggio dagli spostamenti altitudinali scendendo e risalendo lungo tutto il versante montano e così potendo utilizzare appieno tutte le risorse alimentari disponibili, dato che le fruttificazioni e la nascita dei germogli hanno una fenologia (cioè una tempistica) legata alla diversa altitudine. Già intorno al 1968 si verificarono i primi avvistamenti sul versante bolognese, nell’area di Monte Calvi, oggi compresa nel Parco Regionale dei laghi di Suviana e Brasimone. E probabilmente negli stessi anni alcuni esemplari cominciarono a frequentare aree dell’Alto Pratese. In realtà solo dalla fine degli anni Settanta la presenza della specie fu percepita sul versante bolognese e nella media collina del versante toscano. Un’inchiesta dell’Università di Pavia svolta tramite questionari inviati nel 1977 a tutte le stazioni forestali d’Italia restituisce una fotografia riduttiva dell’areale tosco-emiliano (Pavan e Mazzoldi 1983). Al di fuori delle aree storiche dell’Acquerino, nonostante la grossa taglia e la vistosità dei maschi, il cervo passava per lo più inosservato. In quei primi decenni gli animali dovevano essere molto mobili, tornando per il periodo riproduttivo nel cuore dell’areale intorno all’Acquerino: l’areale era quindi di tipo pulsante, con quartieri riproduttivi più ristretti e quartieri invernali-estivi più ampi e così è rimasto ancor oggi. La colonizzazione del Bolognese fu più complessa: all’espansione del nucleo storico dell’Acquerino si aggiunsero negli anni Ottanta la formazione del nucleo di Monte Sole e la fuoriuscita di esemplari da recinti di allevamento (Prada presso Grizzana, Rongrino presso S.Benedetto Val di Sambro, Azienda S. Uberto presso Monterenzio). Un recente studio genetico su un campione di cervi del Pratese ha permesso di riconoscere anche tipi di DNA mitocondriale diversi da quelli attesi per una popolazione di ceppo alpino (Giovannelli et al. 2022), probabilmente il risultato concreto della fuoriuscita di cervi dal recinto di Rongrino.
Per una prima ricostruzione completa dell’areale e le prime stime obiettive della consistenza bisognò attendere la nostra indagine del 1993-95, le verifiche sul campo e i primi censimenti. La tecnica di censimento adottata, il conteggio notturno mediante ascolto e triangolazione dei maschi in bramito e ricostruzione della struttura demografica mediante sessioni di avvistamento era stata messa a punto nel 1988 in Casentino (Mazzarone et al. 1989). Grazie anche al fattivo volontariato dell’URCA bolognese, potemmo fin da subito campionare 67 kmq ne Pratese, Pistoiese e Bolognese e potemmo così accertare che nell’autunno 1995 la popolazione di cervi contava all’incirca 1350 capi su un areale complessivo di 655 kmq. Nel 2000 la consistenza totale era stimabile in circa 2000 capi su 810 kmq, cresciuti a 2600 capi su 1390 kmq nel 2010. Nell’autunno 2022 la stima di popolazione è stata di circa 4200 cervi su un areale totale di 1950 kmq: di questi circa 890 a Pistoia e almeno 1700 a Bologna. Nonostante l’incremento numerico, la densità di popolazione complessiva è rimasta piuttosto costante e su livelli tollerabili, intorno a 2-2,5 capi per kmq. I quartieri riproduttivi, che nel 1995 costituivano il 24% dell’areale complessivo e nel 2010 il 38%, oggi rappresentano il 61% del totale: questo significa che i cervi hanno cambiato gradualmente il loro comportamento spaziale con una tendenza
a spostamenti stagionali relativamente più ridotti. Se nel 1995 alcuni cervi bolognesi dovevano migrare per decine di chilometri per raggiungere le aree di bramito, talvolta sul lato toscano, già un decennio più tardi potevano trovare il proprio quartiere riproduttivo in un raggio più ristretto sul versante emiliano. Insomma gli areali restano di tipo pulsante ma i movimenti stagionali sono più circoscritti.
Tab. 6: Ripartizione dell’areale del cervo nei principali habitat sui due versanti appenninici
(da
Mattioli
et al. 2021)






Gli articoli scientifici del 2021
Nel 2021, utilizzando una parte considerevole dei dati raccolti, alcuni ricercatori dell’Università di Siena e dell’Università di Friburgo, più il tecnico faunistico responsabile della gestione del cervo nel Pistoiese (S. Nicoloso della società DREAm) e il sottoscritto, abbiamo cercato di conoscere meglio la plasticità fenotipica di questa popolazione appenninica di cervi pubblicando due articoli scientifici in inglese su importanti riviste di settore.
Nel primo articolo, intitolato “Spatial variation in antler investment of Apennine red deer”, cioè “Variabilità spaziale dell’investimento nel palco del cervo appenninico”, scritto da S. Mattioli, F. Ferretti, S. Nicoloso e L. Corlatti, pubblicato sulla rivista Ecology and Evolution e liberamente accessibile online, abbiamo preso in esame i pesi corporei dei maschi, i corrispondenti pesi dei palchi e l’età degli animali. L’analisi dei dati permette di ricostruire l’accrescimento corporeo e il cosiddetto “investimento nel palco”. Il nostro studio voleva rispondere a diverse domande: quanti anni dura la fase di accrescimento corporeo? Quando tende a stabilizzarsi il peso corporeo? A che età eventualmente i cervi maschi cominciano a mostrare un declino ponderale? E similmente, con quali modalità i palchi si sviluppano anno dopo anno nei cervi? A che età i palchi raggiungono il massimo peso? Quando si assiste ad un regresso? A noi interessava molto analizzare l’investimento nel palco, inteso come la quantità di tessuto osseo del palco prodotta dai cervi per unità di peso corporeo, cioè il peso relativo del palco, che misura lo sforzo nel trasferire energie per costruire il palco. La plasticità fenotipica osservata nel cervo non si esprime solo nella grande flessibilità della taglia corporea, ma anche nella diversa capacità dei maschi di trasferire energie nel produrre il proprio palco. Cervi che vivono in ambienti particolarmente ricchi di risorse tenderanno a essere di grandi dimensioni e ad avere palchi grandi e pesanti, mentre cervi che vivono in ambienti poco produttivi tendono ad avere taglia modesta e a risparmiare energie sviluppando palchi altrettanto modesti. In fondo il palco è un carattere sessuale secondario a bassa priorità di crescita, sul quale si può “fare economia” se le condizioni ambientali lo richiedono. Quando iniziammo l’analisi dei dati, ci interessava capire se l’investimento nel palco cambiava a seconda della classe d’età, come ci si aspetterebbe da strutture che hanno una funzione attiva solo tra i maschi adulti. Ma più in generale eravamo interessati a documentare le differenze nell’accrescimento corporeo, nello sviluppo del palco e nell’investimento nel palco sui due versanti appenninici. Se fin dall’inizio i due lati della stessa catena montuoso presentavano caratteristiche ambientali diverse e se negli ultimi tempi le differenze sono andate accentuandosi, quali saranno le differenze di taglia e dei palchi nelle due province?
Per analizzare in modo corretto i pesi corporei e quindi l’accrescimento dei cervi maschi è necessario utilizzare dati omogenei. Visto che i pesi variano molto durante la stagione venatoria (che nei primi anni andava da agosto a febbraio-marzo) bisognava conoscere la relazione tra peso e giorno del prelievo per ricostruire per tutti gli esemplari un peso corporeo comparabile. Così abbiamo fissato il peso al 20 ottobre, due settimane dopo la fine del periodo riproduttivo principale (“peso postriproduttivo”).
Per analizzare i pesi dei palchi il compito era meno facile. I dati disponibili infatti erano i pesi dei trofei, cioè i pesi dei palchi con cranio. Dato che il peso del cranio in media nei giovani costituisce il 70% del totale, nei subadulti il 40% e negli adulti il 25%, era necessario eliminare questo elemento di disturbo che avrebbe rischiato di interferire pesantemente sull’analisi dell’investimento. Come fare dunque per stimare il peso netto delle due stanghe se si ha a disposizione solo il peso lordo del trofeo? Noi abbiamo scelto una strada mai percorsa prima. Innanzitutto abbiamo raccolto un
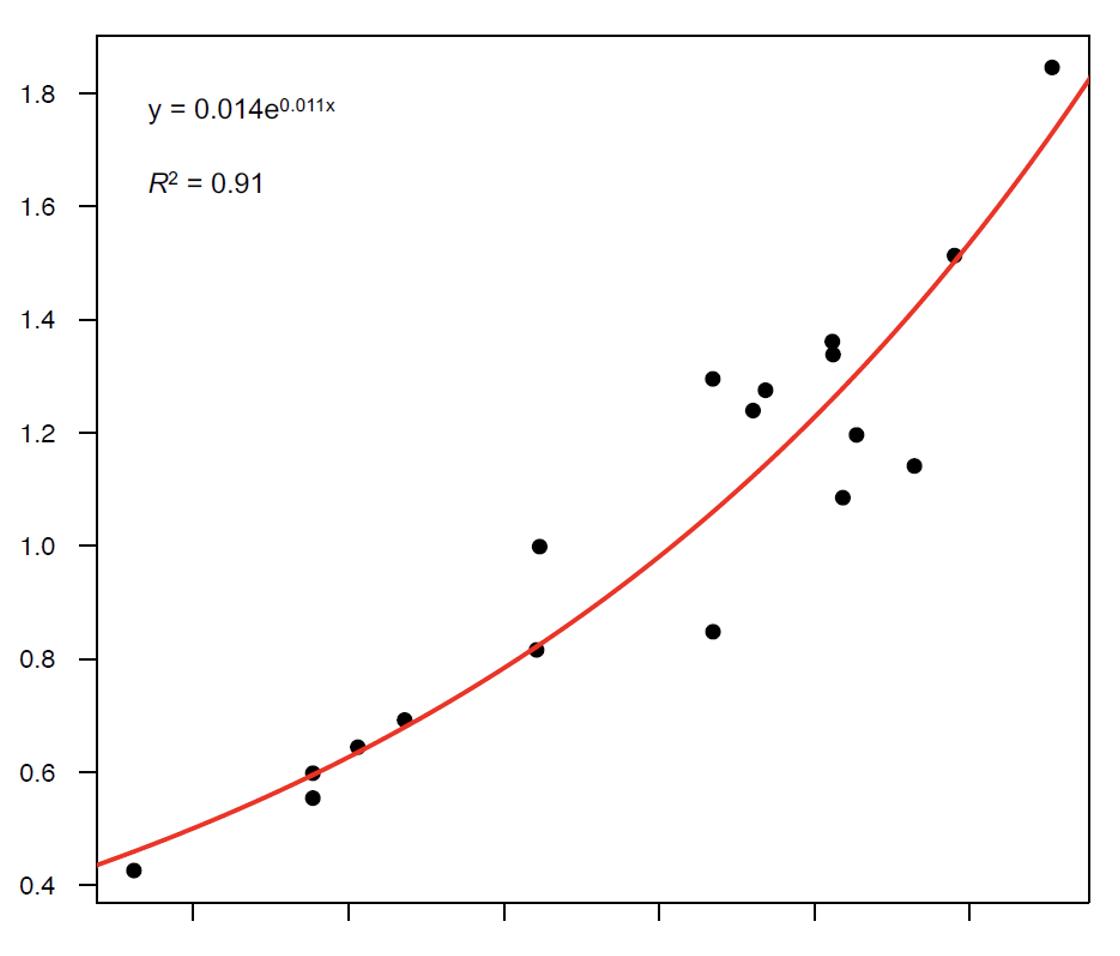
campione di crani di cervi maschi di ogni classe d’età senza palco, crani di animali che avevano perso da poco il palco (“spalcati”) oppure crani ai quali noi avevamo segato le stanghe (foto 13). Li abbiamo pesati, abbiamo misurato la loro lunghezza condilo-basale e abbiamo potuto così constatare che esiste una chiara e robusta relazione matematica tra le due variabili (grafico 1). A questo punto, se per ogni esemplare di cervo maschio della nostra banca dati abbiamo il peso lordo e la lunghezza condilo-basale del cranio, è sufficiente applicare la formula matematica per ricostruire il peso del cranio, sottrarlo al peso del trofeo e ricavare così la stima del peso del palco vero e proprio, cioè delle sole due stanghe.
Peso del cranio senza palco (in kg)
Lunghezza del cranio (in mm)
Grafico 1: Relazione tra peso del cranio senza palco (in kg) e lunghezza condilobasale del cranio (in mm) (da Mattioli et al. 2021)
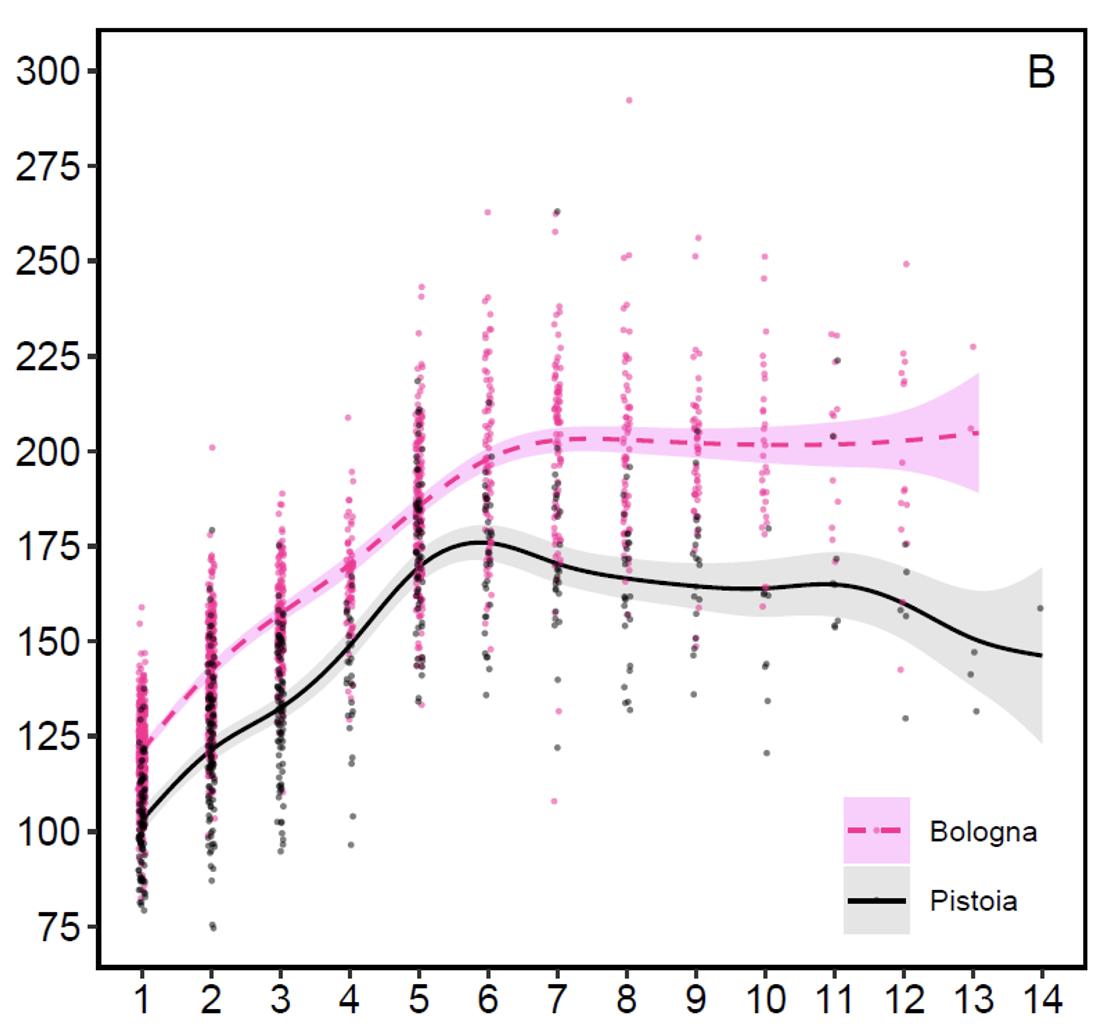
Peso corporeo dei maschi (grafico 2): La fase di accrescimento corporeo dura fino ai 6 anni a Pistoia e fino ai 7 anni a Bologna, ma mentre sul versante emiliano il peso si stabilizza, su quello toscano sorprendentemente il peso tende subito a declinare con gradualità fino agli 11 anni per poi calare con più decisione a 12-14 anni. Difficile trovare una spiegazione soddisfacente per questo calo ponderale nei cervi maschi toscani a 8-9 anni, ad un’età in cui normalmente si raggiunge il massimo della condizione. A Bologna i maschi adulti giovani di 5-7 anni dopo gli amori pesano in media circa 192 kg e quelli di 8 o più anni circa 200 kg. A Pistoia per le stesse classi d’età si passa in media dai 173 kg ai 163 kg. Il peso post-riproduttivo più elevato registrato nel Bolognese era di 292 kg, mentre nel Pistoiese era di 263 kg. Le differenze di taglia tra i versanti sono evidenti ma non marcate e le prestazioni sul lato toscano sono comunque paragonabili a molte popolazioni centroeuropee e sempre di gran lunga superiori ai cervi capostipiti di Tarvisio.
Peso corporeo (in kg)
Età (in anni)
Grafico 2: Peso corporeo post-riproduttivo in relazione all’età (in anni) sui due versanti appenninici (da Mattioli et al. 2021)
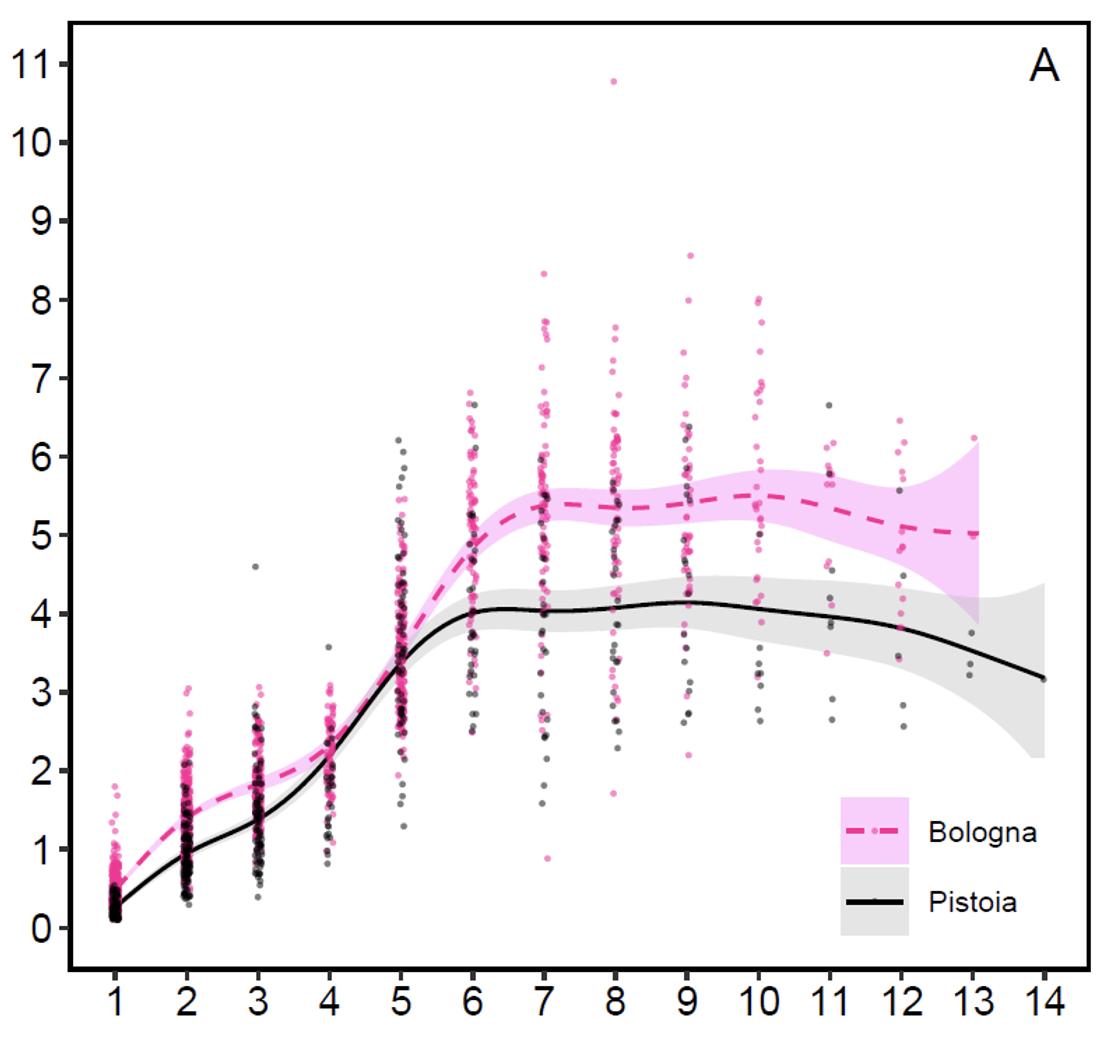
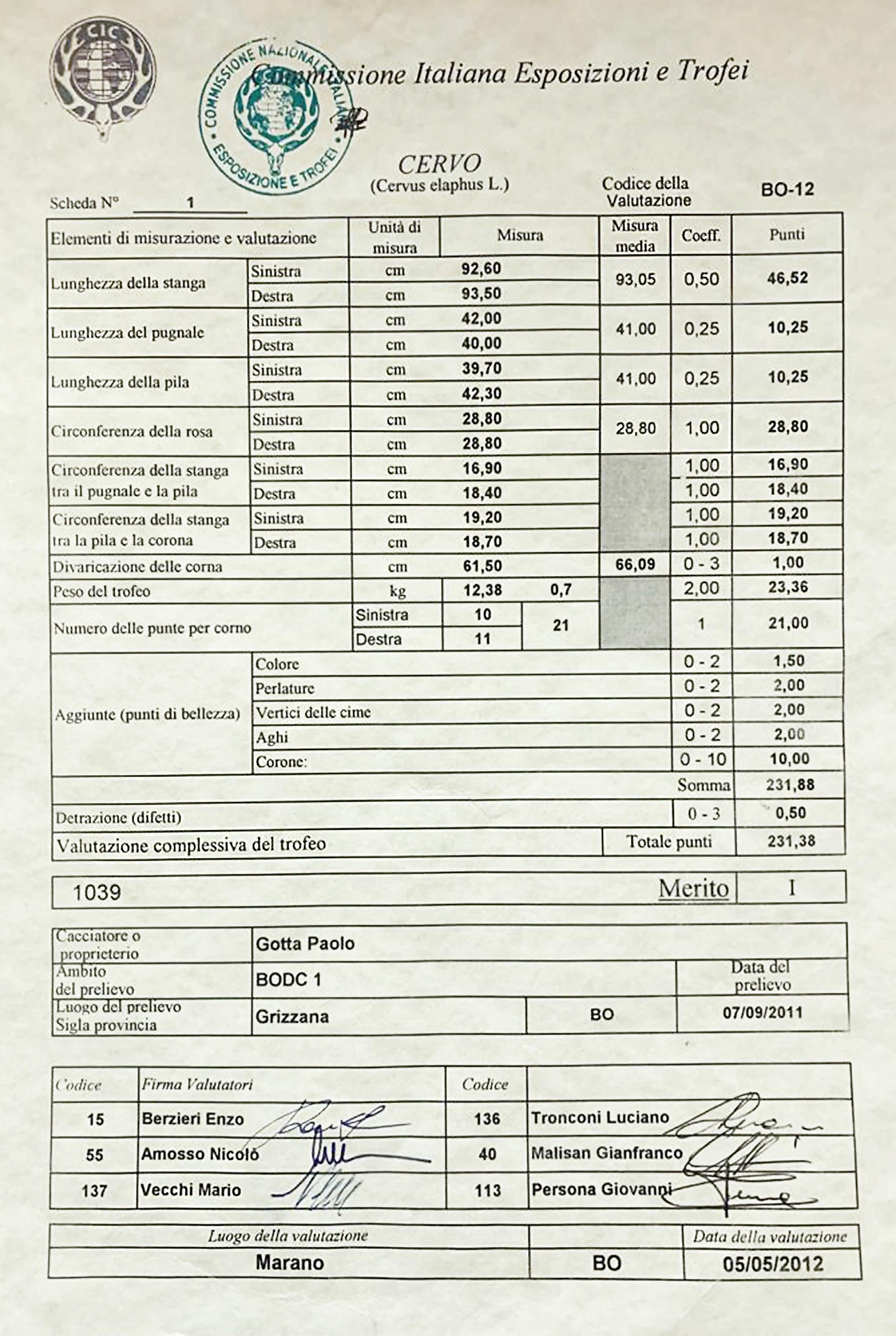
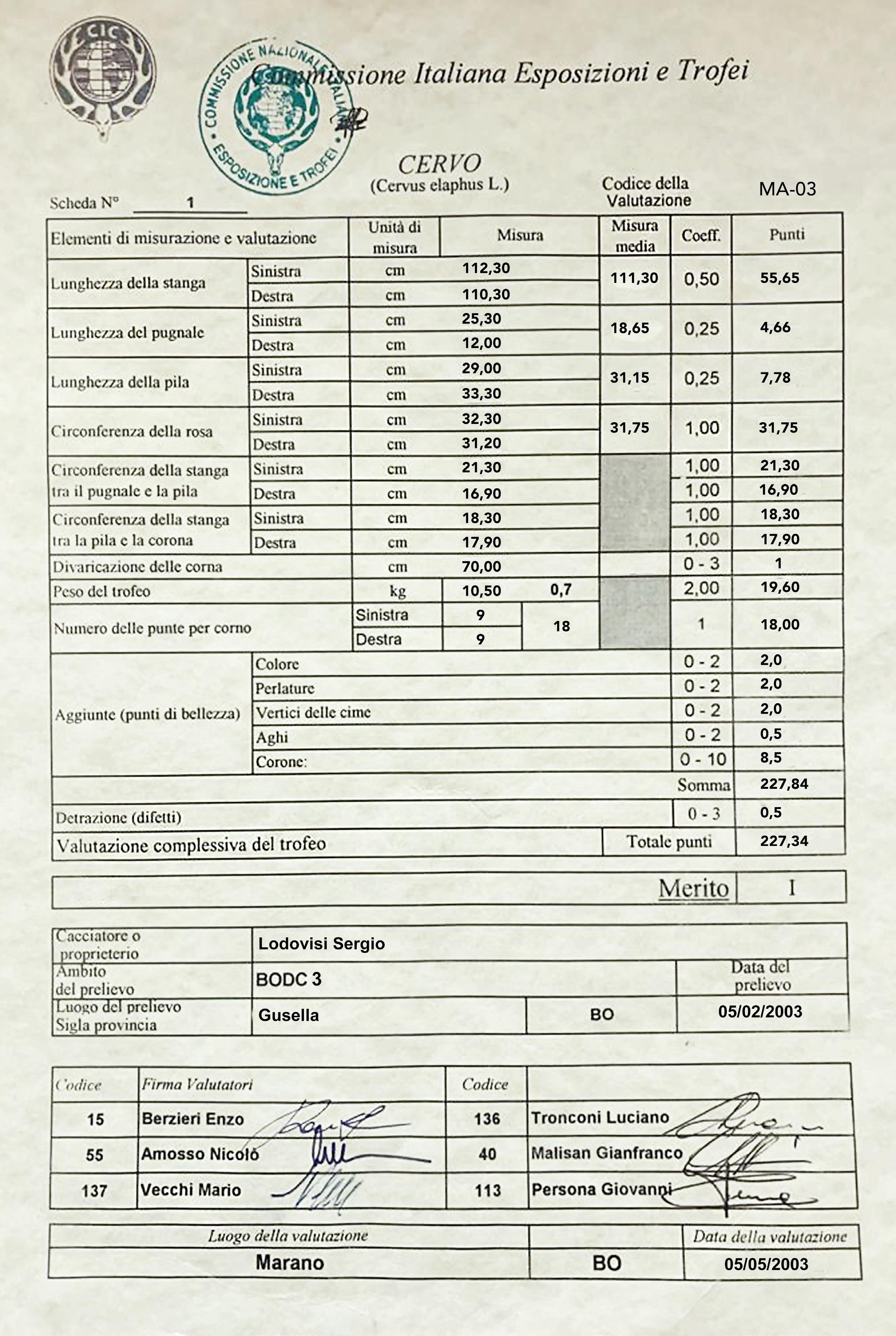
Peso dei palchi (grafico 3): Lo sviluppo medio del peso del palco ha un andamento abbastanza simile a quello del peso corporeo, crescendo fino ai 6 anni nel Pistoiese e a 7 nel Bolognese, rimanendo più o meno stabile fino agli 11 anni e subendo un declino o regresso dai 12 anni in poi. I pesi medi dei palchi si stabilizzano sui 5,3 kg nei cervi emiliani e sui 4 kg in quelli toscani. I pesi più elevati rilevati nei 20 anni di monitoraggio sono stati 10,78 kg a Bologna e 6,66 kg a Pistoia. Il trofeo bolognese, che aveva 21 cime e pesava con il cranio 12,38 kg, rappresentò per il 2011-12 il record italiano, con un punteggio di 231,38 punti CIC (foto 15). L’anno successivo fu superato da un trofeo dell’Appennino reggiano, dell’identico peso, con 16 cime ma con stanghe più lunghe e con un punteggio di 235,25 punti CIC. Il secondo trofeo di Bologna più massiccio aveva 18 punte, pesava 10,5 kg e aveva un punteggio di 227,34 punti CIC (foto 16). Quindi, se è vero che i cervi dell’Acquerino, dopo il periodo di particolare splendore tra il 1975 e il 1985, sono andati via via normalizzando il loro rendimento in termini di struttura e dimensioni dei palchi, è anche vero che sono ancora capaci di produrre palchi di grandi dimensioni, talvolta persino più pesanti di quelli del periodo d’oro, durante il quale non sono noti trofei oltre i 10,7 kg (dato inedito); la maggiore differenza rispetto al passato sta nella conformazione più essenziale, con un numero meno estremo di ramificazioni e generalmente senza palmatura. Per dare un’idea della buona qualità complessiva dei cervi del versante bolognese, ricordiamo che dal 2000-01 al 2022-23 il 50,5% dei maschi adulti prelevati è stato medagliato alle valutazioni CIC (dato inedito) e che 8 trofei erano medaglia d’oro.
Peso del palco (in kg)
Età (in anni)
Grafico 3: Peso del palco in relazione all’età sui due versanti (da Mattioli et al. 2021)



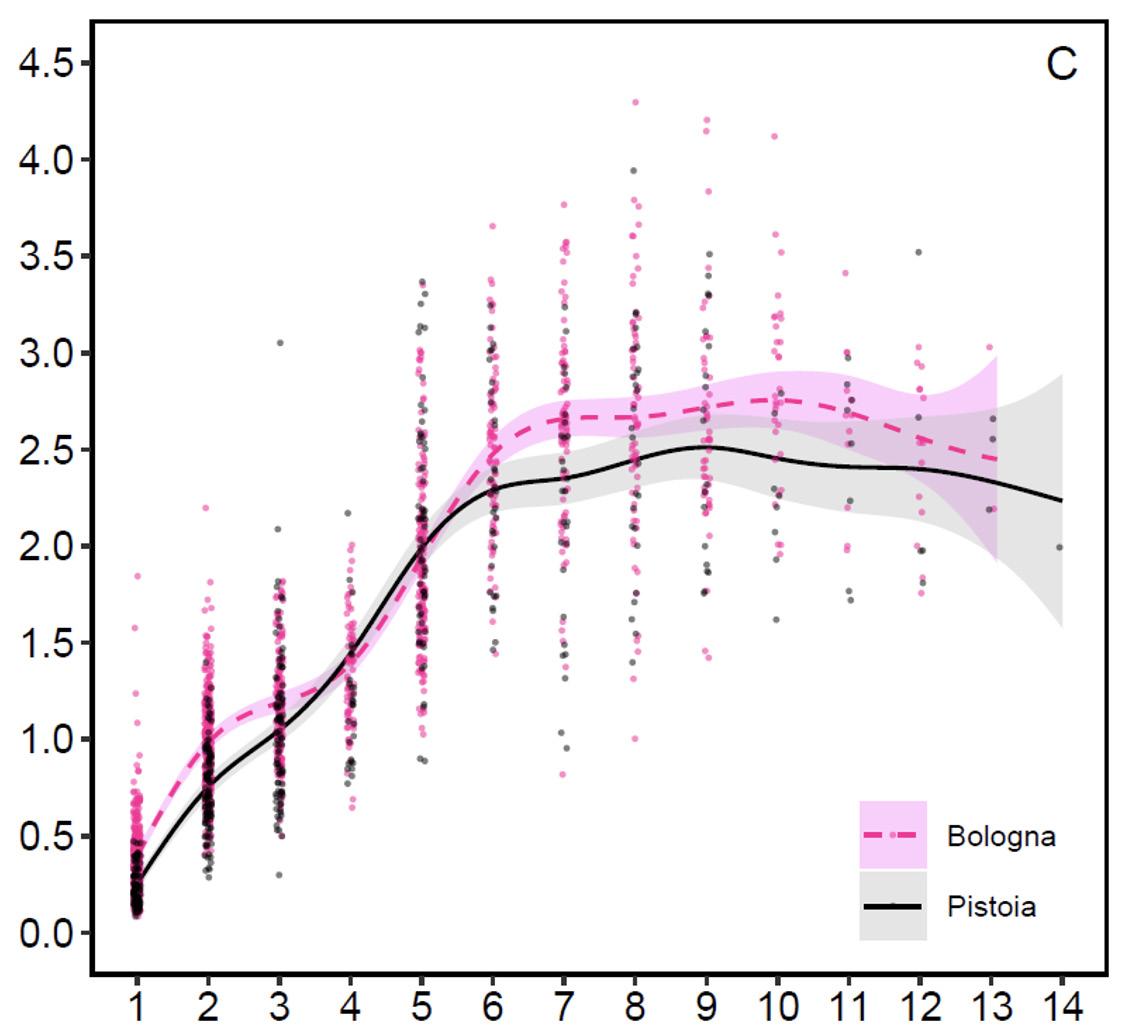
Investimento nel palco (grafico 4): Una volta esaminati separatamente i pesi dei palchi e i pesi corporei, è possibile analizzare il loro rapporto, cioè l’investimento percentuale che i cervi appenninici fanno nel costruire il proprio palco. Il peso dei palchi nei giovani di un anno costituisce in media lo 0,2 (Pistoia) e 0,4% (Bologna) del peso corporeo, nei subadulti lo 0,9 e 1,1%, negli adulti di 5-7 anni il 2,2% su entrambe i versanti e negli adulti di 8 e più anni il 2,7 e 2,5%. I cervi pienamente maturi di 8 e più anni investono nel palco quindi in media 7-11 volte di più rispetto ai giovani, a dimostrazione che l’investimento deve essere massimizzato solo quando i cervi hanno necessità di usare il palco nella competizione riproduttiva. Più in dettaglio i cervi pienamente maturi producono in media 24,6 grammi di tessuto osseo del palco per ogni chilogrammo di peso corporeo sul versante pistoiese e 26,7 grammi sul versante bolognese, una differenza minima per peso unitario. Questo significa che su entrambi i lati i cervi pienamente maturi si impegnano a trasferire più o meno la stessa proporzione di osso del palco e che la differenza nel peso dei palchi dipende dal diverso peso corporeo degli animali sui due versanti dovuto alla diversa produttività degli ambienti. Il massimo investimento percentuale rilevato nel Bolognese è stato del 4,3% e nel Pistoiese del 3,9%.
Rapporto peso palco peso corporeo (in %)
Età (in anni)
Grafico 4: Investimento nel palco (percentuale del peso del palco rispetto al peso corporeo) in relazione all’età sui due versanti (da Mattioli et al. 2021)
Investimento nel palco in altre popolazioni di cervo: Naturalmente per inquadrare correttamente i valori percentuali riscontrati in Appennino bisogna confrontarli con quelli di altre popolazioni di cervi note in letteratura (tab. 7). Purtroppo non è semplice perché soprattutto in Europa orientale in molti casi non vengono rilevati tutti i pesi dei trofei ma solo di quelli medagliati o comunque superiori a qualche soglia e spesso non vengono registrati sistematicamente i pesi corporei. Per quanto riusciamo a ricostruire l’investimento nel palco dei cervi dell’Appennino tosco-emiliano tra Pistoia e Bologna, questo sembra collocarsi tra quelli di popolazioni dal rendimento medio-alto, come quelle dei Carpazi polacchi (medie di 23-27 g per kg di peso corporeo), mentre in Centroeuropa sembrano prevalere valori medio-bassi (16-19 g/kg). In Sardegna, alla Mesola e nelle brughiere scozzesi l’investimento risulta decisamente più basso (10-12 g/kg), come ci si può attendere da cervi di ambienti poco produttivi che devono dosare con parsimonia la produzione di tessuto osseo. L’investimento più elevato (medie intorno a 35 g/kg) è documentato in Slavonia (Croazia) e Pannonia (Ungheria), ma probabilmente valori medi altrettanto alti o ancora maggiori si possono registrare anche nei Carpazi rumeni e nelle montagne bulgare, terre dalle quali provengono splendidi e pesanti supertrofei (di anche 16-18 kg). E probabilmente sorprese potranno venire da una migliore conoscenza dell’investimento nel palco dei cervi reintrodotti nell’Appennino abruzzese, dove sono note coppie di stanghe pesanti fino a 12 kg (peso presunto del trofeo intorno ai 14 kg) e lunghe fino ai 131 cm (Ciccotti 2022) (foto 11-12). Che l’investimento medio del cervo nobile vari in Europa da circa 10 a 35 g e più per kg di peso corporeo è un’ennesima dimostrazione della grande plasticità fenotipica di questa specie.
Investimento nel palco per altri Cervidi: È interessante anche inquadrare l’investimento nel palco della specie cervo all’interno della famiglia dei Cervidi (tab. 8). Il cervo nobile fa parte delle specie di Cervidi più poliginici e dimorfici, cioè le specie in cui i maschi competono maggiormente per l’accesso alle femmine e quindi hanno dimensioni corporee decisamente più grandi rispetto alle femmine e tendono ad investire di più anche nei caratteri sessuali secondari come il palco: i maschi adulti di wapiti e daino, che hanno strategie riproduttive piuttosto simili a quelle del cervo, producono in media circa 22-34 g per kg di peso corporeo. Poi esistono specie con dimorfismo sessuale intermedio (come il cervo coda bianca americano), che hanno una competizione meno intensa e quindi investono in media intorno a 11-12 g per kg di peso corporeo, e infine ci sono specie come il capriolo in cui la competizione tra maschi e il dimorfismo sessuale sono decisamente più attenuati e i maschi come sappiamo si accontentano di costruire palchi piccoli e leggeri investendo in media solo 4-7 g per kg di peso corporeo.
Tab. 7: Investimento medio nel palco (g per kg di peso corporeo) di alcune popolazioni di cervo
(da Mattioli et al. 2021)
popolazione
Baranja (Ungheria) 8-10 36,5
Baranya (Croazia) 8-10 34,3
Appennino PT-BO 8 e+ 24,6-26,7
Carpazi polacchi
(Polonia)
Bassa Sassonia (Germania)
P. N. Słowinski (Polonia)
(Regno Unito)
Glenfeshie (Regno Unito) 5-10 10,2
Tab. 8: Investimento medio nel palco (g per kg di peso corporeo) di alcune specie di Cervidi (da Mattioli et al. 2021)
specie
classe d’età (anni) Investimento nel palco (g/kg)
Daino 5 e + 26-28,3
Wapiti 7-10 22,1-34,4
Cervo Coda Bianca 5-7 11,5
Capriolo 3 e + 4,4-6,6
Implicazioni gestionali: Il nostro articolo scientifico sull’investimento nel palco a prima vista potrebbe interessare solo una ristretta cerchia di specialisti, ma in realtà ha ricadute concrete nella gestione quotidiana del cervo. Raccogliere regolarmente i dati dei pesi dei palchi e quelli corporei dei maschi, conoscere quindi il rapporto tra peso del palco e peso corporeo, cioè appunto l’investimento dei cervi nel palco, permette di stimare la condizione fisica dei singoli esemplari e dell’intera popolazione. Se una popolazione che vive in ambienti più produttivi investe mediamente di più nei palchi di una che vive in ambienti più poveri, questo dato può essere usato come misura della salute e del vigore della popolazione che stiamo seguendo, come indice generale di condizione, e i valori medi da noi trovati possono servire da riferimento. Anche la valutazione dei trofei potrebbe trarre vantaggi da un monitoraggio biometrico sistematico. Oggi spesso la valutazione dei trofei è in qualche modo del tutto separata dalla conoscenza dell’intera popolazione, e chi valuta i trofei non conosce le caratteristiche biometriche degli animali che portavano quei trofei. Conoscere invece il peso del trofeo e quello corporeo di tutti i maschi di una popolazione di cervi e quindi anche di quelli medagliati consente di capire meglio le potenzialità e la qualità di quella popolazione; monitorare nel tempo tutti gli animali attraverso questa doppia raccolta di dati permette di seguire i cambiamenti di rendimento ed eventualmente di decidere gli interventi adeguati per migliorare la situazione.
Nel secondo articolo, intitolato “Land cover and weather jointly predict biometric indicators of phenotypic quality in a large herbivore”, cioè “Copertura del suolo e clima predicono gli indicatori di qualità fenotipica in un grande erbivoro”, scritto da C. Vannini, N. Fattorini, S. Mattioli, S. Nicoloso e F. Ferretti, pubblicato sulla rivista Ecological Indicators e liberamente accessibile online, si è invece preso in esame l’influsso di fattori ambientali e climatici sui pesi corporei e su diverse misure somatiche delle femmine e dei piccoli di cervo.
Attraverso l’analisi dei pesi corporei dei maschi, dei pesi dei palchi e dell’investimento nei palchi dello studio precedente eravamo riusciti ad osservare solo differenze su grande scala spaziale, tra un versante e l’altro della catena appenninica, mentre con l’analisi della taglia di cerve e cerbiatti di questa seconda indagine siamo riusciti ad evidenziare differenze anche su piccola scala spaziale e su scala temporale. I cervi maschi hanno spazi vitali grandi e le loro condizioni fisiche sono il risultato dell’alimentazione su vaste superfici; il fatto che all’interno di ciascun versante non avevamo trovato differenze tra distretti o periodi significa che all’interno delle ampie zone frequentate i maschi nei diversi distretti hanno sempre trovato disponibilità alimentari sostanzialmente equivalenti. Le femmine e i piccoli – come documentato anche da studi preliminari di radiotelemetria in zona - tendono invece ad avere spazi vitali più ristretti (con medie intorno ai 300 ha; cfr Bocci et al. 2009, Crocetti et al. 2010) e quindi è possibile valutare le condizioni ambientali vissute dai singoli esemplari nell’area circostante il punto di prelievo. Sono state analizzate in particolare cinque variabili biometriche: peso corporeo (eviscerato), altezza al garrese, circonferenza toracica, lunghezza del garretto e lunghezza della mandibola. Le prime quattro erano state rilevate dal 2001 al 2017 da rilevatori biometrici abilitati, mentre la lunghezza della mandibola da un tecnico faunistico (S. Mattioli per il Bolognese, S. Nicoloso per il Pistoiese). Per ognuno dei cervi prelevati veniva creato su mappa informatizzata un intorno circolare a partire dal punto di abbattimento e ne veniva analizzata la copertura del suolo, cioè la proporzione dei diversi ambienti, focalizzandoci sulla percentuale di aree coltivate. Inoltre i servizi meteorologici regionali di Emilia-Romagna e Toscana ci avevano fornito i dati pluviometrici e di temperatura del periodo di indagine. Ci interessava vedere se la piovosità e le temperature dell’inverno e della stagione vegetativa (primavera ed estate) precedenti
il prelievo avevano effetti sui pesi e sull’accrescimento scheletrico degli animali.
I risultati della nostra analisi sono decisamente interessanti. Innanzitutto si conferma su grande scala che le cerve adulte e sottili e i cerbiatti che vivono sul versante Bolognese hanno taglia maggiore di quelli del Pistoiese (tab. 5). L’areale occupato dai cervi nel Bolognese ha una eterogeneità maggiore rispetto a quello del Pistoiese, che mostra tratti forestali più vasti e compatti e relativamente pochi coltivi e piccoli pascoli, spesso raggruppati. Sul versante bolognese le aree aperte sono più di tre volte più ampie di quelle all’interno dell’areale sul versante pistoiese mentre i boschi del lato emiliano rappresentano solo i due terzi di quelli del lato toscano (tab. 6).
Ma un’analisi su scala più fine ha permesso di apprezzare meglio la plasticità della specie, capace di rispondere in termini di accrescimento corporeo a seconda delle caratteristiche ambientali e di fattori climatici.
Esaminiamo separatamente i risultati per classe:
Nelle femmine adulte il peso corporeo cresce col crescere della proporzione di aree coltivate negli intorni (buffer) di 300 ettari centrati sul punto di prelievo. Secondo il modello matematico sviluppato dagli autori dell’articolo scientifico una cerva adulta che nel Pistoiese viva in un’area con grandi superfici coltivate (70-90% di coltivi nell’intorno) ha in media un peso eviscerato superiore di 5 kg rispetto ad una femmina che frequenti un’area con pochi coltivi o del tutto priva (73,9 kg contro 69 kg). Il peso subisce anche gli effetti di piovosità e temperature ed è maggiore se nella stagione vegetativa precedente il prelievo il tempo è stato più piovoso e più fresco.
Nelle femmine sottili, ma solo sul lato Pistoiese, il peso corporeo cresce col crescere della proporzione di coltivi nell’intorno. Secondo il modello matematico un esemplare di questa classe prelevato all’interno di un’area con il 90% di coltivi avrebbe un peso eviscerato 15 kg superiore rispetto ad uno prelevato in un’area senza coltivi. E per cerve ancora in accrescimento come le sottili si tratta di una percentuale estremamente significativa. Anche la lunghezza del garretto e quella della mandibola nelle sottili cresce col crescere dell’estensione delle zone coltivate.
Nei piccoli femmine di 8-10 mesi, solo nel Pistoiese, peso corporeo, garretto e mandibola crescono col crescere dei coltivi nell’intorno. Secondo il modello un piccolo prelevato in un’area con molti coltivi (70-90%) ha in media un peso eviscerato circa 5,5 kg kg superiore ad un individuo di una zona con pochi coltivi o del tutto priva (42,5 kg contro 36,9 kg). Primavere ed estati più piovose e più fresche hanno effetti positivi su peso corporeo e circonferenza toracica.
Nei piccoli maschi della stessa età, su entrambi i versanti ma in maniera più forte nel Pistoiese, si è dimostrato un effetto positivo della percentuale di coltivi nell’intorno su peso corporeo e circonferenza toracica. Secondo il modello matematico nel Pistoiese i cerbiatti maschi che vivessero in aree con il 90% di coltivi avrebbero un peso eviscerato circa 10 kg superiore a quelli di aree prive di coltivazioni. Solo nel Pistoiese è dimostrato come anche altezza al garrese e lunghezza della mandibola crescono col crescere dell’estensione dei coltivi. Anche nei cerbiatti maschi è documentabile un effetto degli eventi meteorologici nei mesi precedenti il prelievo. Il peso corporeo è maggiore negli anni in cui primavera ed estate sono stati più piovose e fresche e negli anni in cui l’inverno precedente era stato più mite (e il piccolo era solo un feto nell’utero della madre); inverni meno rigidi avevano un effetto positivo anche sulla crescita della circonferenza toracica e della mandibola.
Alcuni risultati della nostra indagine erano sicuramente attesi, altri sono sembrati almeno inizialmente meno ovvi. Che la taglia di femmine e piccoli fosse sempre maggiore sul lato emiliano non c’erano dubbi, dato lo studio precedente sui cervi maschi e viste le caratteristiche territoriali del Bolognese, con maggiore diversità ambientale e quindi relativamente più ricco di risorse. Che a scala più fine la percentuale di coltivi nell’area attorno al punto di prelievo potesse influire sull’accrescimento corporeo e la taglia degli animali era immaginabile: campi, pascoli, in genere aree aperte offrono fonti alimentari relativamente ricche di nutrienti. Ma mentre per le cerve adulte il fenomeno era osservabile su entrambi i versanti, per cerve sottili e cerbiatti in accrescimento era dimostrabile solo sul lato Pistoiese. La spiegazione sta proprio nella rarità dei coltivi nell’Appennino pistoiese: qui le aree coltivate possono effettivamente fare la differenza e determinare sorti molto diverse tra esemplari che possono goderne ed esemplari che ne vivono lontano. Lo studio preliminare sulla mobilità condotto dall’Università di Firenze su 12 cerve munite di radiocollare sul crinale appenninico di confine e sul versante toscano ha permesso di scoprire come le femmine tendono ad usare i campetti più di quanto atteso in base alla proporzione dei coltivi all’interno degli spazi vitali, come se la loro presenza fosse estremamente importante (Crocetti et al. 2010). Mentre sul versante emiliano, cerve sottili e cerbiatti che abitino sia in aree con coltivi sia senza hanno comunque a disposizione un territorio più diversificato e quindi più ricco.
Molto interessante è stato anche vedere come il clima durante la stagione vegetativa primaverile ed estiva influisce sui pesi corporei e sulla crescita scheletrica degli animali, dimostrabile soprattutto per i cerbiatti e in minor misura sulle cerve adulte. Primavere ed estati più piovose e fresche aumentano la produttività della vegetazione e quindi anche maggiore produzione di latte delle madri e questo favorisce un accrescimento più rapido dei piccoli.
Altrettanto intrigante è stato vedere come la severità dell’inverno precedente la nascita influisca sulle dimensioni corporee future dei cerbiatti maschi. Temperature più basse sperimentate durante la fase fetale producono effetti sulla crescita dei piccoli di sesso maschile perché i costi della gestazione di un figlio maschio (più grande di una femmina) sono per una cerva madre maggiori rispetto a quelli che dovrebbe sopportare per la gravidanza di una figlia (Clutton-Brock et al. 1981, Landete-Castillejos et al. 2004).
Questa indagine ci ha permesso di osservare su piccola scala il lavorio continuo della plasticità fenotipica, la capacità della specie cervo di modulare l’accrescimento corporeo a seconda delle condizioni ambientali e climatiche. I fenomeni osservati ci consentono anche di guardare (con preoccupazione) al futuro. Se cerve e cerbiatti si accrescono maggiormente in presenza di ambienti aperti e di primavere ed estati più piovose e fresche, cosa potrà succedere se la tendenza in atto è di ulteriore riduzione delle superfici coltivate, di chiusura progressiva del bosco e del riscaldamento globale? Con l’aumento delle temperature e la riduzione della piovosità, saranno sempre più comuni e prolungati i periodi di siccità, con vegetazione sempre più fibrosa e più povera di nutrienti, con vistose conseguenze (anche) sulla specie cervo, destinata -a parità di densità di popolazione- ad avere accrescimenti corporei più lenti e taglia finale ridotta.
Implicazioni gestionali: I risultati di questa ricerca ci dicono comunque che si potrebbe percorrere una strada diversa, e se è vero che per sfidare i cambiamenti climatici la lotta non può che essere globale, è anche vero che per contrastare la tendenza cui sta andando incontro il cervo in Appennino basterebbe introdurre interventi sul territorio per migliorare la qualità ambientale.
Con il forte declino della pastorizia e l’abbandono della montagna, in Appennino boschi e arbusteti negli ultimi 50-70 anni hanno quasi raddoppiato di estensione e le aree aperte compreso le coltivazioni agricole si sono dimezzate (Falcucci et al. 2007). E se i cervi di ceppo alpino traslocati
all’Acquerino inizialmente trovarono ancora un paesaggio diversificato e poterono esprimere molto del loro potenziale, dopo i primi decenni ginestreti, felceti e boschi andarono sostituendosi nell’alto Pistoiese alle aree aperte, a molti dei campi un tempo coltivati e ai pascoli usati dalla pastorizia. Inoltre sia sul versante toscano sia su quello emiliano i boschi cedui invecchiando hanno chiuso sempre più la volta arborea con conseguenze negative per lo sviluppo del sottobosco che esige luce. E mentre nel Bolognese l’eterogeneità ambientale ha comunque garantito al cervo quantità e qualità di risorse trofiche di tutto rispetto, nel Pistoiese la struttura più compatta delle compagini boscate e la scarsa presenza di aree aperte coltivate hanno sicuramente inciso sulla ricettività complessiva del territorio. Quando si definisce il cervo come una specie forestale si commette un grave errore: in realtà è una specie che esige la compresenza di boschi non compatti disetanei con radure e di aree aperte a prateria (Mattioli et al. 2022). Coltivi e pascoli possono in qualche modo sostituire almeno in parte le praterie originarie, come dimostrato anche in altre parti d’Europa (Mysterud et al. 2002, 2022). Di qui la necessità di miglioramenti ambientali soprattutto (ma non esclusivamente) sul versante toscano: sfoltimenti dei boschi cedui coetanei e invecchiati ovunque e creazione di aree aperte, con risemine e sfalci periodici per mantenere attrattivo il cotico erboso (per una disamina più dettagliata delle azioni necessarie si veda Mazzarone e Mattioli 1996, pagine 108-118). Interventi abbastanza impegnativi e tutto sommato contro corrente ma che si potrebbero inizialmente applicare anche soltanto alle aree protette presenti. E se è vero che sarà quasi impossibile rivedere i maestosi cervi dai palchi iper-ramificati e palmati degli anni Settanta e Ottanta, è anche vero che miglioramenti ambientali di questa portata porterebbero non solo ad un miglioramento delle condizioni dei cervi (grazie proprio alla plasticità della specie) ma anche più in generale ad un aumento della biodiversità complessiva.


Riferimenti bibliografici
Abbazzi L. (1995) Occurrence of palmated Cervus elaphus from Italian late Pleistocene localities. Rendiconti Lincei 6(3): 189-206
Becciolini F., Bozzi R., Viliani M., Biffani S., Ponzetta M.P. (2016) Body measurements from selective hunting: biometric features of red deer (Cervus elaphus) from Northern Apennine, Italy. Italian Journal of Animal Science 15: 461-472
Bocci A., Monaco A., Brambilla P., Angelini I., Lovari S. (2009) Alternative strategies of space use of female red deer in a montainous habitat. Annales Zoologici Fennici 47: 57-67
Brown R.D. (1990) Nutrition and antler development. pp 426- 441 in: Bubenik G.A. e Bubenik A.B. (cur.) Horns, Pronghorns, and Antlers. Springer, New York
Ciccotti E. (2022) Studio preliminare su una popolazione di cervo (Cervus elaphus L. 1758) in Abruzzo e prospettive di gestione a livello regionale. Tesi Università di Firenze
Clutton-Brock T.H., Albon S.D., Guinness F.E. (1981) Parental investment in male and female offspring in polyginous mammals. Nature 289: 487-489
Crocetti C., Ponzetta M.P., Minder I., Messeri A., Cervasio F. Argenti G., Maccelli S. (2010) Analysis of land utilisation by red deer in the Apennine mountains. pp 265-267 in: Schnyder et al. (cur.) Grassland in a changing world. EGF Kiel
De Marinis A.M. (2015) Valutazione dell’età nei Cervidi tramite esame della dentatura. Guida pratica all’identificazione delle classi d’età del cervo. Manuali e Linee Guida 90.2/2013:1-136 ISPRA
Falcucci A., Maiorano I., Boitani I. (2007) Changes in land-use/land-cover patterns in Italy and their implications for biodiversity conservation. Landscape Ecology 22: 617-631
Geist V. (1986) Super antlers and pre-world war II European research. Wildlife Society Bulletin 14: 91-94
Geist V. (1987) On speciation in Ice Age mammals, with special reference to cervids and caprids. Canadian Journal of Zoology 65: 1067-1084
Giovannelli F., Mori E., Zaccaroni M., Dessì-Fulgheri F., Baratti M. (2022) Genetic insights into an Apennine population of the Italian red deer. Mammal Research 67: 399-406
Huxley J.S. (1931) The relative size of antlers in deer. Proceedings of the Zoological Society of London 101: 819-864
Landete-Castillejos T., Gortazar C., Vicente J., Fierro V., Garcia A., Gallego L. (2004) Age-related foetal sex ratio bias in Iberian red deer (Cervus elaphus hispanicus): are male calves too expensive for growing mothers? Behavioral Ecology and Sociobiology 56: 1-8
Mattioli S. (1994) Le reintroduzioni di Cervidi in Italia, con particolare riferimento all’Appennino”, pp 19-25 in: Inglisa M. (cur.) “La fauna appenninica e la sua conservazione” Atti del Convegno Nazionale Paolo Barrasso, Airone-Corpo Forestale dello Stato
Mattioli S., Meneguz P.G., Brugnoli A. e Nicoloso S. (2001) Red deer in Italy: recent changes in distribution and numbers. Hystrix, Italian Journal of Mammalogy 12: 21-35
Mattioli S., Ferretti F., Nicoloso S., Corlatti L. (2021) Spatial variation in antler investment of Apennine red deer. Ecology and Evolution 11: 7850-7864
Mattioli S., Zachos F., Rossi L., Lister A., Corlatti L. (2022) Red deer Cervus elaphus. Linnaeus, 1758. pp 51-86 in: Corlatti L. e Zachos F. (cur.) Terrestrial Cetartiodactyla. Handbook of the Mammals of Europe. Springer Nature
Mazzarone V., Apollonio M., Lovari C., Mattioli L., Pedone P., Siemoni N. (1989) Censimento di cervo al bramito montano appenninico. In: Fasola M. (cur.) Atti del II Seminario sui Censimenti Faunistici dei Vertebrati, Brescia. Supplemento alle Ricerche di Biologia della Selvaggina 26: 587-590
Mazzarone V. e Mattioli S., cur. (1996) Indagine sulla popolazione di cervo dell’Acquerino. Relazione finale 1993-1995. Regione Toscana, Firenze 1-133
Mysterud A., Langvatn R., Yoccoz N.G., Stenseth N.C. (2002) Large-scale habitat variability, delayed density effects and red deer population in Norway. Journal of Animal Ecology 71: 569-580
Mysterud A., Langvatn R., Meisingset E.L., Rivrud I.M. (2022) Agricultural grasslands buffer density effects in red deer populations. Journal of Wildlife Management 87: e22357, 1-14
Palma di Cesnola A. (1988) Paglicci, Rignano Garganico. Mostra iconografica permanente. Regione Puglia 1-88
Pavan G. e Mazzoldi P. (1983) Banca dati della distribuzione geografica di 22 specie di Mammiferi in Italia. Collana Verde 66, Ministero dell’Agricoltura e Foreste, Roma 1-279
Premuda G. (1975) Origine e consistenza attuale del patrimonio faunistico della montagna pistoiese. Diana 11: 28-31
Reulecke K. (1987) Zur Geweihentwicklung der Hirsche im Harz. Wild und Hund 24/87: 41-45
Szunyoghy J. (1959) The variability of the branch numbers od stag antlers in the Carpathian Basin. Annales Historico-Naturales Musei Nationalis Hungarici 51: 489-492
Vannini C., Fattorini N., Mattioli S., Nicoloso S., Ferretti F. (2021) Land cover and weather jointly predict indicators of phenotypic quality in a large herbivore. Ecological Indicators 128: 107818, 1-11
Vogt F. (1948) Das Rotwild. Oesterreicher Jagd- und Fischereiverlag, Vienna 1-207
Wilkens B. (1989) Il cervo dal Mesolitico all’Età del Bronzo nell’Italia centro-meridionale.
Rassegna di Archeologia 8: 63-95
Ringrazio di cuore la Delegazione italiana del C.I.C. per avermi invitato a scrivere questo testo divulgativo sull’origine e lo sviluppo della popolazione dell’Acquerino e sui recenti articoli scientifici in inglese prodotti dal nostro gruppo di ricerca. Sono molto grato ad A. Dal Pian per aver messo a disposizione le sue belle foto. La mia gratitudine va anche ai colleghi e amici L. Corlatti, N. Fattorini, F. Ferretti, S. Nicoloso e C. Vannini, co-autori dei due recenti articoli. L. Corlatti e N. Fattorini mi hanno gentilmente revisionato questo testo. È giusto menzionare anche i miei compagni della prima indagine del 1993-95 V. Mazzarone, C. Lovari, N. Siemoni I. Aiardi, S. Gualazzi e P. Pedone. S. Csanyi, dell’Università Szent István di Gödöllő (Ungheria) mi ha cortesemente messo a disposizione dati inediti sui trofei ungheresi di cervo. Voglio infine ricordare P. Agnelli, già curatore dei Mammiferi presso il Museo di Storia Naturale La Specola dell’Università degli Studi di Firenze, per l’impegno profuso nel far acquisire la stanga iper-ramificata e palmata alle collezioni del museo stesso.