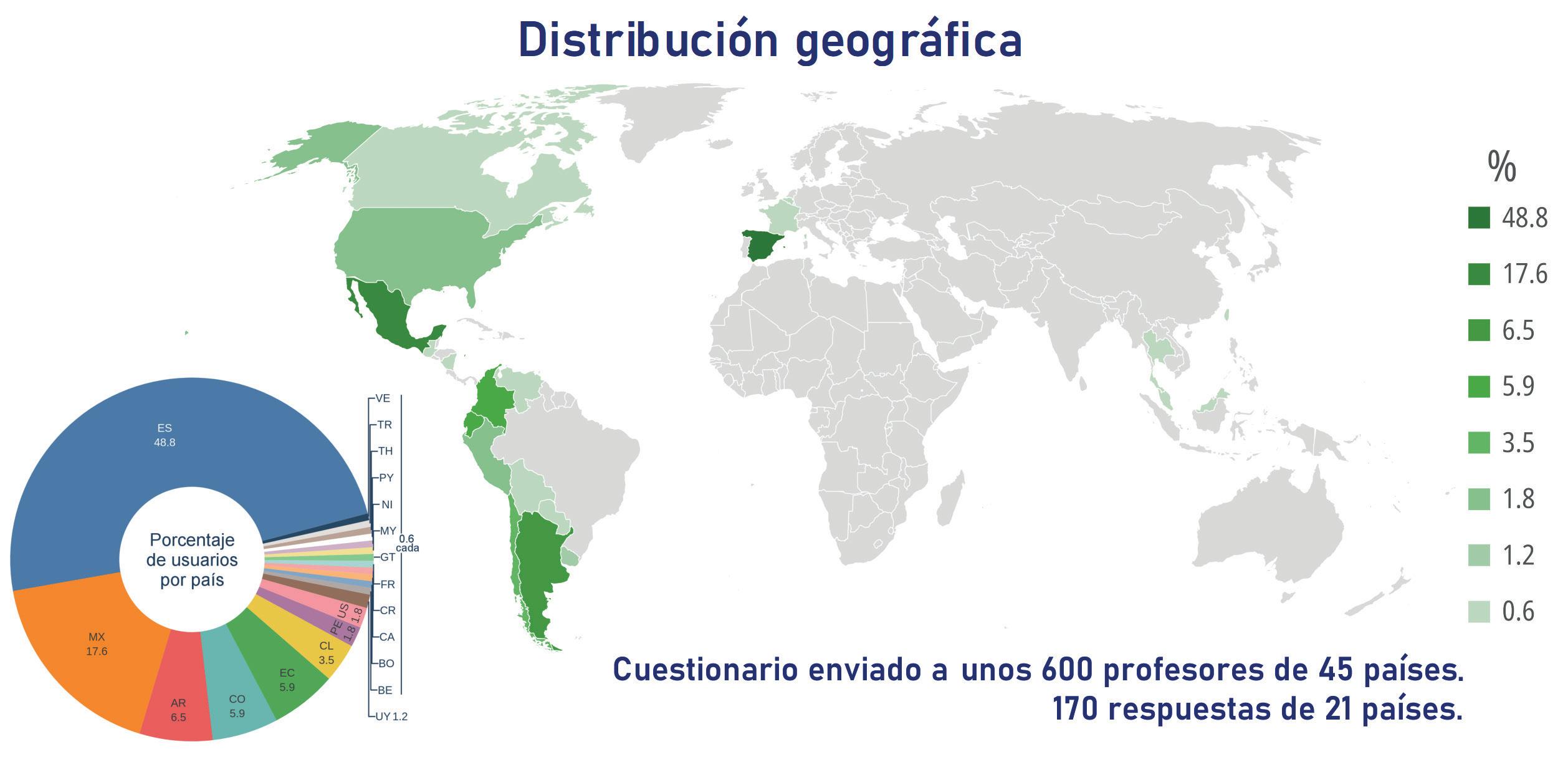
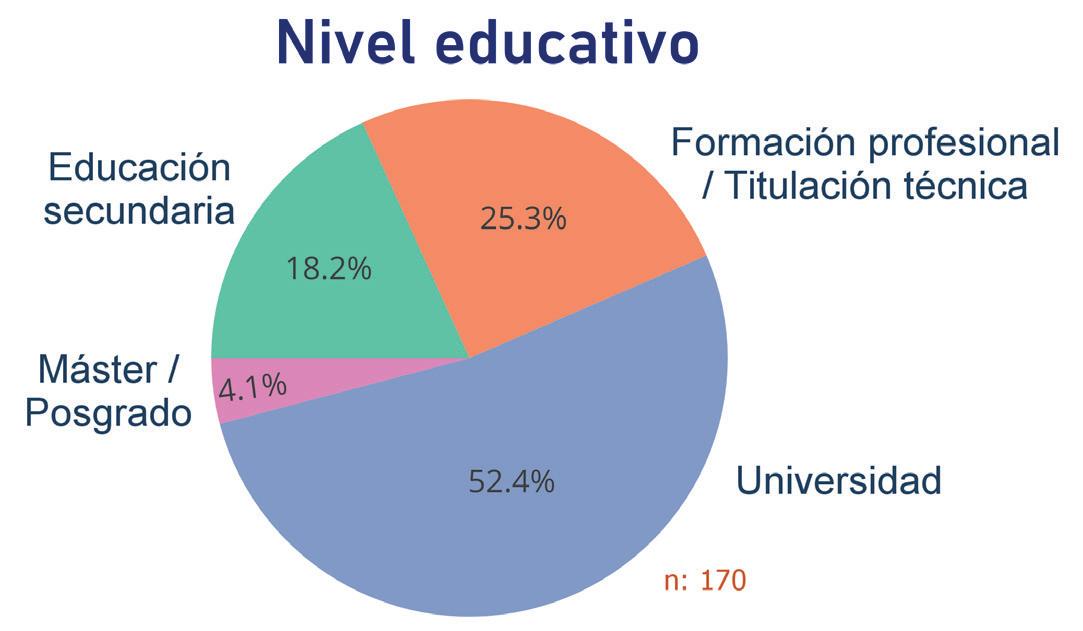
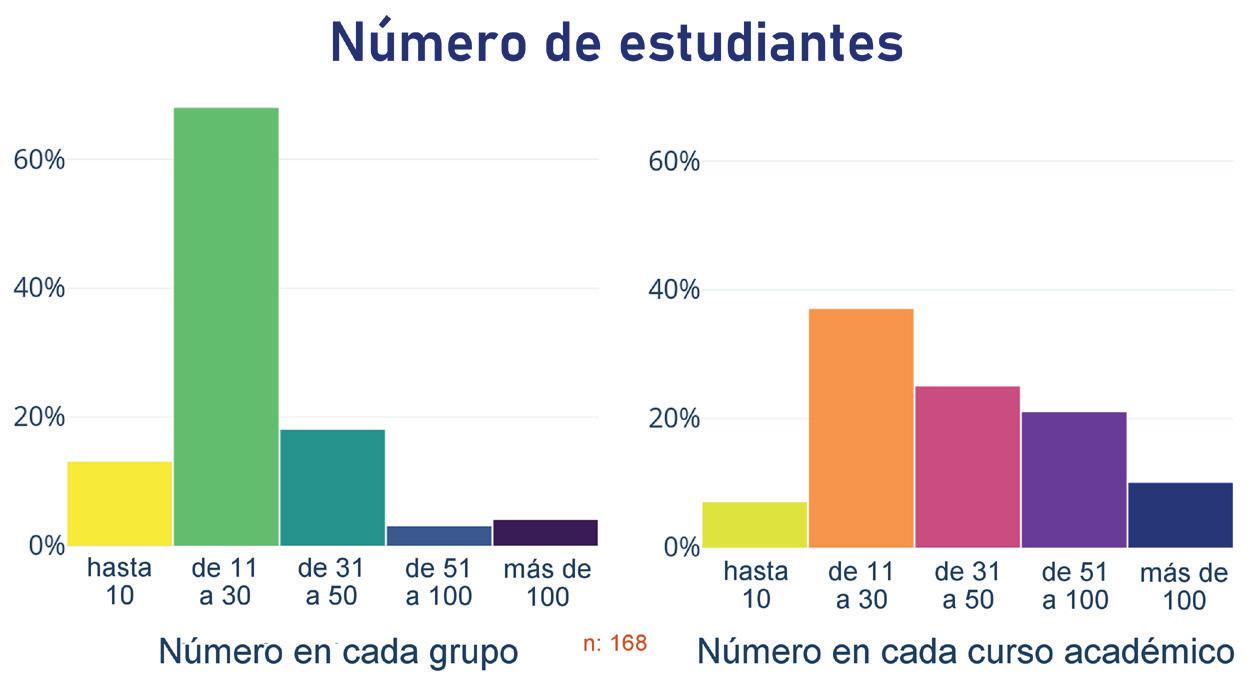
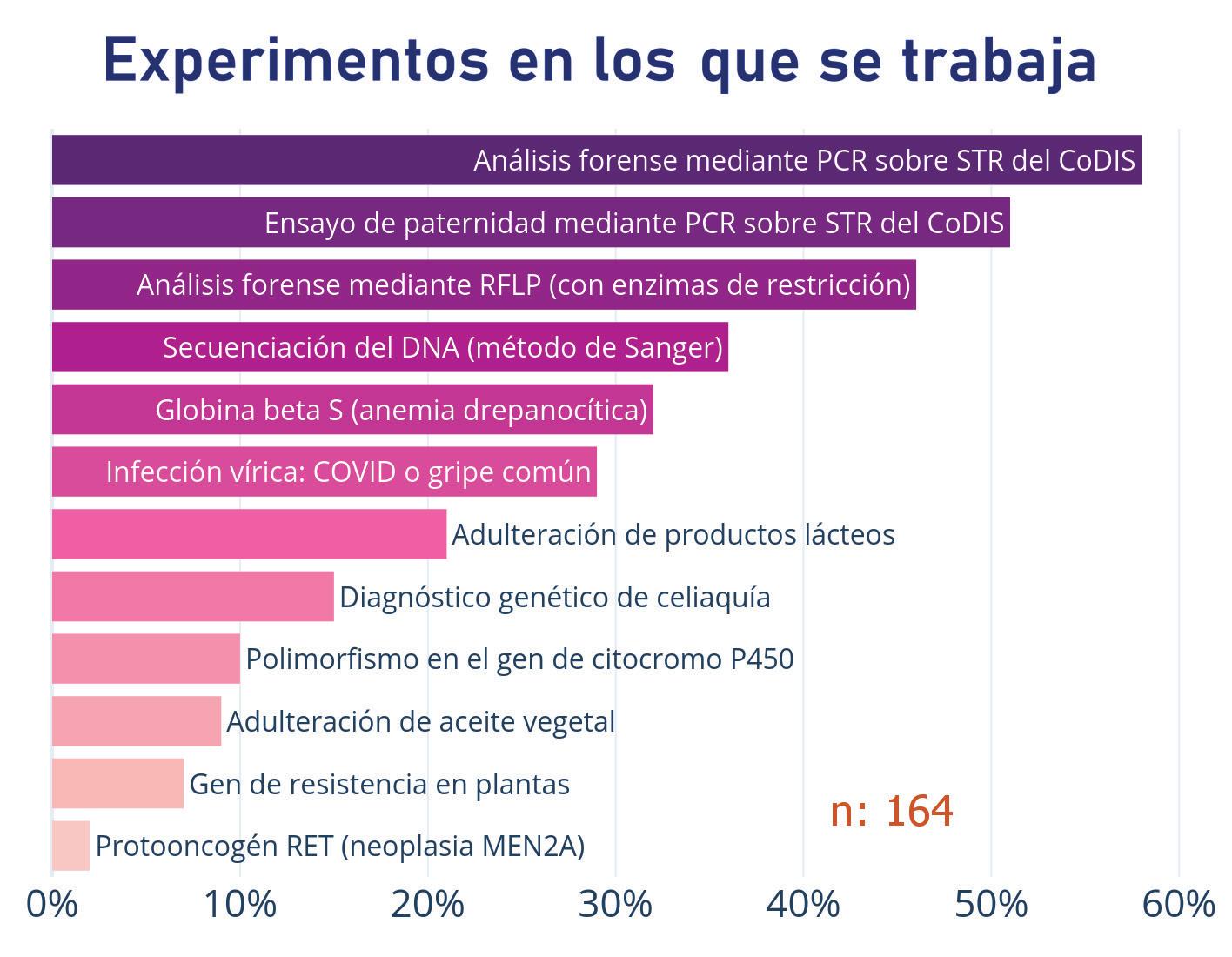
Noticias de actualidad de la SEBBM «Sobre la ciencia y la política científica»
Alonso Rodríguez Navarro
RESEÑA DE LIBROS
Órbitas. Apuntes de una vida en continua exploración
Sara García Alonso
PUBLIRREPORTAJES
El creciente impacto normativo en la I+D+I
Antonio Ferrer
Presidente SEBBM
Nos venimos enfrentando a un incremento en los trámites administrativos para solicitar, gestionar y justificar los proyectos de I+D+I. La inversión de tiempo y recursos se ha disparado de forma aplastante, siendo necesario contemplar una reducción para que los investigadores podamos centrar nuestra atención y esfuerzo en generar el conocimiento científico que aporte las soluciones a los retos sociales. No es una tarea fácil, puesto que los procedimientos se han ido complicando con el tiempo. En primer lugar, para asegurar la ejecución correcta y transparente de los presupuestos como pretende la normativa para la gestión de los fondos del Plan de Recuperación, Transformación y Resiliencia (PERTE). En segundo lugar, por la creciente necesidad de reducir el perjuicio ambiental de los proyectos, todavía asumible por estar en sus inicios, pero que se complicará con el paso del tiempo.
Vayamos por partes. Aplaudo las medidas que garantizan el correcto uso de los fondos concedidos a los proyectos de I+D+I, pero me parece descabellado que prácticamente se nos criminalice aplicando una fiscalización irrazonable en la gestión de unos presupuestos modestos, que requieren gastos moderados. Lo vivimos con la aprobación de la ley 9/2017 de contratos del Sector Público, que limitaba la compra a proveedores en base al NIF/CIF de la institución, sin considerar que en la ejecución de los proyectos de I+D+I las compras a un proveedor son realizadas por una constelación de grupos de investigación independientes y autónomos de la institución, y no por una unidad central. Esto obligó a crear Acuerdos Marco con los proveedores que nos fuerzan a comprar los productos ofertados en dichos acuerdos, a pesar de que pueden no tener la calidad requerida por el proyecto o estar obsoletos. Una medida pensada para controlar y ahorrar pero que, en ocasiones, resulta punitiva, incluso económicamente. Los investigadores seguimos asombrados por semejante torpeza de nuestras autoridades.
Actualmente, para la ejecución de los fondos PERTE, hemos de añadir el análisis de conflictos de interés (Minerva) junto con la declaración de ausencia de conflicto de interés (DACI) que debe firmar el investigador principal para obtener la autorización de la AEAT para prácticamente cada compra, independiente de su valor; simplemente, ¡impresionante! ¿Alguna autoridad se ha detenido a pensar en el tiempo y recursos invertidos en este procedimiento? Entiendo que este requisito se exija para compras importantes (infraestructuras, equipos, etc.) para evitar la corrupción que, por cierto, es incapaz de conseguir pues estamos viendo como el procedimiento falla en casos altamente gravosos, lo que ha encendido, una vez más, las alarmas en la Comisión Europea. Que se tenga que aplicar a los proyectos es una aberración legislativa promovida por nuestros gobernantes como una excusa para diluir el deficiente control de la corrupción entre sus afines. Probablemente, es una normativa necesaria para asegurar una gestión correcta, ética y transparente de grandes inversiones, pero incorrectamente aplicada a los proyectos de I+D+I sobrecargando a los investigadores y sus instituciones de procesos administrativos costosos y cuestionables. Confiemos que los últimos acontecimientos políticos no resulten en una normativa más restrictiva y dolosa para la I+D+I. A la comentada carga burocrática hay que añadir procedimientos adicionales que la incrementan más. Por una parte, el aumento de las exigencias para usar animales de experimentación, así como células y muestras humanas. No pongo en duda que tenemos el deber de cumplir con los requerimientos éticos en ambos casos, lo que cuestiono es que se tenga que solicitar la aprobación de los mismos procedimientos para cada proyecto, lo que nos obliga a cumplimentar los mismos impresos de forma recurrente. Por otra parte, se ha instaurado la declaración del principio de no causar perjuicio significativo al medio ambiente (DNSH) tanto a nivel de la solicitud como de la justificación con el fin de evaluar el impacto medioambiental que pueden generar los proyectos de I+D+I. Por el momento, el DNSH es un trámite relativamente simple que consiste en contestar unas preguntas referentes al perjuicio medioambiental que podría ocasionar el proyecto. Poco me equivoco si afirmo que la mayoría de los proyectos en biociencias y biomedicina declaran tener un impacto mínimo o nulo sobre el medio ambiente, a menos, que estén centrados en él. Esta afirmación, aparentemente cierta, resulta cada vez menos asumible por el intensivo uso de plástico en los laboratorios. Cada vez más agencias internacionales están exigiendo en las solicitudes de proyectos una estimación del uso del plástico y otros agentes contaminantes, así como un plan de medidas para su reducción. No nos extrañemos que en breve se nos exija hacer esta estimación en un formulario DNSH actualizado. En relación con este tema, otro aspecto que está surgiendo, y al que también debemos prestar atención en el futuro es la huella de carbono que producimos los investigadores con las reuniones presenciales (reuniones, congresos, etc.). Recientemente, se me ha pedido calcular la huella de carbono, así como proponer un plan de contingencia, como fomentar las videoconferencias y usar transporte no contaminante. No me sorprenderá ver en un futuro no tan lejano una exigencia similar en las solicitudes de los proyectos nacionales. En mi opinión, estos procedimientos junto a otros como el plan de gestión de datos, si bien tienen un objetivo defendible y asumible, debieran consensuarse con la comunidad científica para no sobrecargarnos de actividades que cada vez nos distraen más del planteamiento y desarrollo de proyectos innovadores de I+D+I, perjudicando así la generación de conocimiento que contribuya a resolver los retos de nuestra sociedad. Dedicar nuestro cerebro, tiempo y esfuerzo a estos aspectos, si no se controla, tendrá un importante impacto sobre la creatividad y productividad científica. Cuidar el medio ambiente, ejecutar correcta y transparentemente los fondos, y fomentar la compartición de los datos y recursos, ¡SI!, pero no a costa de distraer en exceso nuestra sustancia gris. Busquemos el equilibrio para cumplir con la ética y la sostenibilidad, sin distorsionar, ni afectar el avance científico, motor del bienestar social.
Ciencia exprés sin barreras
Inmaculada
Yruela Guerrero
Directora Revista
SEBBM
El pasado 30 de mayo asistí a la final de la cuarta edición del certamen Falling Walls Lab Spain 2025 como miembro del jurado; un concurso internacional en el que España participa seleccionando al representante que defenderá su proyecto en la final de Berlín del 6 al 9 de noviembre. Allí se darán cita los finalistas de cada país, en una fecha que rememora la caída del Muro de Berlín en 1989. El tiempo marcado para la exposición de cada proyecto es muy breve - sólo tres minutos-, y se penaliza tanto exceder el tiempo asignado como no emplearlo.
Fue ilusionante ver la motivación de los jóvenes al exponer sus innovadores proyectos, persiguiendo romper barreras al inicio de sus carreras investigadoras y/o profesionales, incluso algunos cursando aún sus estudios universitarios de grado o máster. Es de aplaudir la valentía mostrada, siendo tan jóvenes, para participar en un certamen internacional de estas características, y superar el reto de cumplir con el tiempo de tres minutos, marcado en una pantalla, para la exposición. Un formato de ciencia exprés que se ha consolidado tanto en los foros de divulgación de la ciencia como en los congresos científicos -flash talkspara dar cabida a un mayor número de participantes, con excelentes trabajos, y hacer llegar el mensaje a un público no especialista más heterogéneo, donde la interdisciplinariedad es más frecuente. El ejercicio de condensación y síntesis se ha ido imponiendo en la comunicación de la ciencia, al igual que en otros ámbitos (noticias, música, publicidad, teatro, …). Se argumenta que la capacidad de atención, concentración y comprensión de un oyente no va más allá de 15 minutos. Lejos quedan las conferencias, seminarios y exposiciones de trabajos, con el máximo detalle, de cerca de una hora. Los tiempos se han acortado, no más allá de 30, 20 o cinco minutos. Un ejemplo, las flash talks, de cinco minutos, en las sesiones de los grupos científicos de los congresos SEBBM para favorecer la presentación de un mayor número de comunicaciones científicas.
Es evidente que este formato de ciencia exprés tiene ventajas para moverse en campos interdisciplinares -cada vez más necesarios- aunque requiere un esfuerzo extra para los más jóvenes. Una fórmula que está en línea con la tendencia de «pensamiento positivo» (‘positive thinking’) -un conjunto de prácticas que los psicólogos relacionan con el optimismo y el éxito a la hora de comunicar-. Pero no hay que dejar a un lado los debates sosegados para profundizar en el fondo de los temas. No hay que olvidar que las discusiones de los detalles y la exposición de los fracasos son muy necesarias, y en esto también hay que incidir y educar.
Con este tipo de iniciativas -Falling Walls y otras- las barreras no sólo se rompen en el ámbito del conocimiento y de la comunicación, sino también en el ámbito social. Participar en estos certámenes y defender la vocación científica puede ser un hándicap para los más jóvenes al no ser reconocidos como iguales en los círculos donde se mueven. Hay barreras que siguen costando derribar. Muchas veces pienso que el círculo al que llegamos tiene un diámetro pequeño y que su expansión es limitada a pesar del gran esfuerzo realizado. La necesidad de conocimiento científico para abordar los problemas y retos que se nos plantean no parece calar lo suficiente en nuestra sociedad, a la vista de los acontecimientos. Al finalizar el certamen Falling Walls visité el espacio sobre D. Santiago Ramón y Cajal instalado en el Museo Nacional de Ciencias Naturales (MNCN), muy cerca del lugar donde estuvimos reunidos. Hice un recorrido por su biblioteca, su material de laboratorio, sus objetos personales -un telescopio y una máquina fotográfica- que «dan a conocer cómo era el hombre que había detrás del investigador y cuáles eran sus aficiones». Además, dibujos, videos y grabaciones sonoras de Cajal, fotografías, paneles explicativos y documentos de algunos de sus principales discípulos. Os animo a visitarla si no lo habéis hecho, al igual que el «Espacio Cajal» en el Edificio Paraninfo de la Universidad de Zaragoza, una exposición permanente también de interés. Seguramente a D. Santiago le hubiera costado sintetizar su investigación en una flash talk.
La exposición del MNCN, en colaboración con el Instituto Cajal del CSIC, se instaló para celebrar el centenario de su nombramiento como director del Instituto del CSIC que lleva su nombre el 12 de noviembre de 1920, a la espera de crear un museo nacional en Madrid dedicado a su persona y legado. Desde 2022 hemos estado celebrando el «Año de Investigación Ramón y Cajal», declarado de especial interés público por el Gobierno de España, que se ha extendido hasta el pasado 31 de mayo de 2025. Llegada la fecha de cierre de este periodo conmemorativo, de cerca de tres años, científicos y familiares de Cajal «gritaron» con urgencia por el Museo Cajal en un artículo en El País. Aún continuamos esperando el prometido museo nacional según dictaba el Real decreto 590/2024 de 25 de junio del pasado año. Este mes de julio, no obstante, ha saltado la noticia de un acuerdo por parte del Ministerio de Ciencia Innovación y Universidades, la familia de Cajal y representantes de varias instituciones científicas para instalar el esperado museo en un histórico caserón de la calle Atocha de Madrid, a escasos veinte metros del Museo Reina Sofía.
Confiemos que este acuerdo llegue a buen puerto sin mucha demora y haga caer este muro pronto.
Antes de terminar quiero mencionar que el dosier de este número de la Revista SEBBM va dedicado a la «Biología del ARN», un tema que ha visto y sigue viendo caer barreras de conocimiento, y que de nuevo en 2024 han sido reconocidas con la concesión del Nobel de Fisiología o Medicina a Victor Ambros y Gary Ruvkun. En la Revista SEBBM nº 223 dimos cuenta de ello.
EL ARN: UN ACTOR VERSÁTIL CON MÚLTIPLES FORMAS Y FUNCIONES
Encarnación Martínez-Salas
Centro de Biología Molecular Severo Ochoa, CSIC-UAM, Madrid
https://doi.org/10.18567/sebbmrev_226.202509.dc1
El ácido ribonucleico (ARN, también llamado RNA por sus siglas en inglés «ribonucleic acid») juega un papel esencial en la cadena de procesos de la vida. En las últimas dos décadas, investigaciones puramente básicas en diferentes campos de la Biología y la Química han descubierto funciones propias del ARN, que resaltan su papel protagonista en todos los seres vivos. Por ello, ha merecido nueve premios Nobel en los últimos veinte años. Cuántos secretos más todavía guarda el ARN es desconocido y, por otra parte, fascinante. Curiosamente, a nivel molecular el ARN existe en todas las conformaciones imaginables: moléculas de cadena sencilla de polaridad positiva (+ssARN), de polaridad negativa (-ssARN), de doble banda (dsARN), o covalentemente cerrado (circARN). Por otra parte, los distintos tipos de ARN varían en longitud, estructura, presencia
de nucleótidos atípicos (Inosina) y/o modificaciones post-transcripcionales (M7G, M6A, M5C, Ψ, etc.), que contribuyen a su estabilidad, localización, interacción con proteínas y otros factores celulares y, por lo tanto, a su función.
De hecho, el ARN es el actor principal en numerosos procesos celulares, en los que realiza distintas funciones claves: catalizador (ribozimas: RNasa P, rARN, intrones), genoma (virus ARN, viroides), partícula estructural (ribosomas, tARNs, snARNs, snoARNs, SRP), como regulador de la estabilidad del ARN y de la traducción (miARNs, lncARNs, asARNs, UTRs), como transmisor de la capacidad codificante del genoma (mARNs), como agente terapéutico (vacunas de ARN), etc. Esta diversidad de actividades es inherente a la plasticidad y dinamismo de su estructura secundaria y terciaria que, junto con su secuencia, aumenta sinérgicamente la capacidad
de reconocer distintos ligandos y efectuar distintas acciones.
La precisión y especificidad de los sistemas biológicos se atribuye a la interacción entre diferentes componentes del sistema. El ARN y las proteínas son componentes fundamentales de todos los organismos, imprescindibles para su supervivencia y propagación. El ARN ribosómico, el transferente y el mensajero, actúan coordinadamente para generar las proteínas, que forman parte de la maquinaria que realiza las funciones celulares necesarias para mantener la homeostasis. Es bien conocido que la interacción entre miARNs, mARNs y proteínas regula la función de los mismos en numerosos procesos celulares. De hecho, las partículas ribonucleoproteícas (RNPs) son complejos dinámicos, implicados en distintos pasos de la expresión génica, el tráfico y la degradación del ARN, así como el recambio de proteínas, etc.
La comprensión de la influencia de modificaciones químicas en el ARN, así como la inesperada a la vez que sorprendente capacidad codificante de lncARNs y cirARNs, está en continuo avance. De hecho, la diversidad de estructuras y modificaciones del ARN ha sido clave en el desarrollo de estrategias alternativas para regular su función. Ejemplos paradigmáticos del papel regulador de las modificaciones y la estructura del ARN son el resíduo M7GTP (conocido como «cap») del mARN, o los sitios de entrada interna del ribosoma (IRES) que dirigen la iniciación interna de la traducción. En los organismos eucariotas, la mayoría de los mARNs se traducen mediante un mecanismo dependiente de cap en un proceso regulado por varias proteínas, denominadas eIFs. Defectos, o inhibición, de la síntesis de proteínas tiene implicaciones graves en procesos fisiológicos, por ejemplo, la respuesta a diversos tipos de estrés, el
desarrollo, la proliferación celular, o la memoria a largo plazo. En estas circunstancias se dispara la traducción de mARNs atípicos, además de algunos lncARNs y circARNs, mediante mecanismos independientes de cap, un proceso en el que intervienen proteínas de unión a ARN (RBPs), dando lugar a la síntesis de proteínas específicas claves para la supervivencia en determinadas circunstancias de alerta celular.
Este dosier contiene cuatro artículos en los que se describen distintos tipos de moléculas de ARN que desempeñan funciones claves, descubiertas recientemente:
• Una forma novedosa de ARN son los denominados «obeliscos», presentados en el artículo del Dr. Marcos de la Peña (IBMPC, CSIC-UPV, Valencia): pequeñas moléculas circulares de ARN con capacidad replicante, descubiertos en numerosos seres vivos, que se repliegan predominantemente como doble banda, similares a los viroides.
• El artículo del Dr. Pablo Manavella y la Dra. Catharina
Para leer más
Merchante (IHSM «La Mayora», Universidad de Málaga - CSIC) describe el papel esencial de los microARNs en el desarrollo de las plantas, desde su síntesis y maduración hasta su implicación en la regulación postranscripcional de distintos mARNs diana.
• El Dr. Juan Carlos GarcíaSoriano y la Dra. Puri Fortes (CIMA, Pamplona), explican la capacidad codificante de algunos, quizás inapropiadamente llamados long non-coding ARNs. Estos ARNs, junto con los ARN circulares, representan un mundo de pequeñas moléculas, inadvertidas en etapas anteriores, con nuevas funciones por explorar.
• Finalmente, el capítulo dedicado a epitranscriptómica, presentado por la Dra. Sandra Blanco (CIC, Salamanca) describe la asombrosa capacidad reguladora de la expresión génica determinada por las modificaciones (metilaciones de adenina y citosina, o presencia de pseudouridina) en el ARN.
Cech TR, Steitz JA. The noncoding RNA revolution-trashing old rules to forge new ones. Cell 157 (2014) 77-94. https://doi. org/10.1016/j.cell.2014.03.008
Karikó K, Weissman D. Naturally occurring nucleoside modifications suppress the immunostimulatory activity of RNA: implication for therapeutic RNA development. Current Opinion Drug Discovery & Development 10 (2007) 523-532.
Lozano G, Martínez-Salas E. Structural insights into viral IRESdependent translation mechanisms. Current Opinion in Virology 12 (2015) 113-120. doi:10.1016/j.coviro.2015.04.008
Sinha T, Panigrahi C, Das D, Chandra Panda A. Circular RNA translation, a path to hidden proteome. Wiley Interdisciplinary Reviews (WIREs) RNA 13 (2022) e1685. doi: 10.1002/wrna.1685
UN NUEVO
«MUNDO» DE ARN
Marcos de la Peña
Instituto de Biología Molecular y Celular de Plantas (IBMCP), CSIC-UPV, Valencia
https://doi.org/10.18567/sebbmrev_226.202509.dc2
Vida
El fenómeno natural que todos conocemos como vida, y que en general solemos contraponer a todo aquello que es inerte, es en realidad un concepto difícil de definir satisfactoriamente. Ya el origen de la palabra «Vida», del latín vita, está íntimamente relacionado con la simple idea de existencia. Y algo similar ocurre desde Life en inglés o Leben en alemán (del indoeuropeo Leip, permanecer), así como el término chino 生 (sheng, existir) o el hindi जिजीविषा (jijivisha, persistir). Obviamente, no todo aquello que simplemente exista se puede considerar como vida, y al final nuestra definición está inevitablemente ligada a la experiencia humana en la que seguimos unos criterios, en parte arbitrarios, para distinguir lo vivo de lo no vivo. Existen sistemas tanto naturales como artificiales que comparten o simulan muchos aspectos de la vida biológica, como las tormentas y otros fenómenos meteorológicos -que se autoorganizan, intercambian energía y mueren disipándose en el ambiente-, los cristales -que crecen, replican patrones y compiten- o incluso la inteligencia artificial (IA) o los virus informáticos -que replican,
mutan y evolucionan en entornos digitales-. Por eso, se han definido características exclusivas de la vida biológica, como (i) la capacidad de transformar materia y energía en crecimiento ordenado, (ii) la existencia de un programa o propósito funcional codificado en su estructura molecular que dirige mecánicamente su supervivencia y reproducción (lo que se conoce como «teleonomía interna»), y (iii) la capacidad de evolución darwiniana, en la que la información se hereda con variación y es sometida a selección a lo largo de las generaciones. Sin embargo, la vida puede ser considerada un fenómeno mucho más amplio si relajamos algunos de estos requisitos. Y esto es algo necesario, casi obligatorio, si queremos entender dos fenómenos límite en biología íntimamente relacionados: el origen de la vida (abiogénesis) y los microorganismos de genomas mínimos (virus y entidades biológicas similares), donde la frontera entre lo inerte y lo vivo se vuelve algo muy poroso.
Y si queremos reconstruir los primeros pasos en el origen de la vida, necesariamente hay que aceptar que éstos se tuvieron que dar con entidades replicantes muy
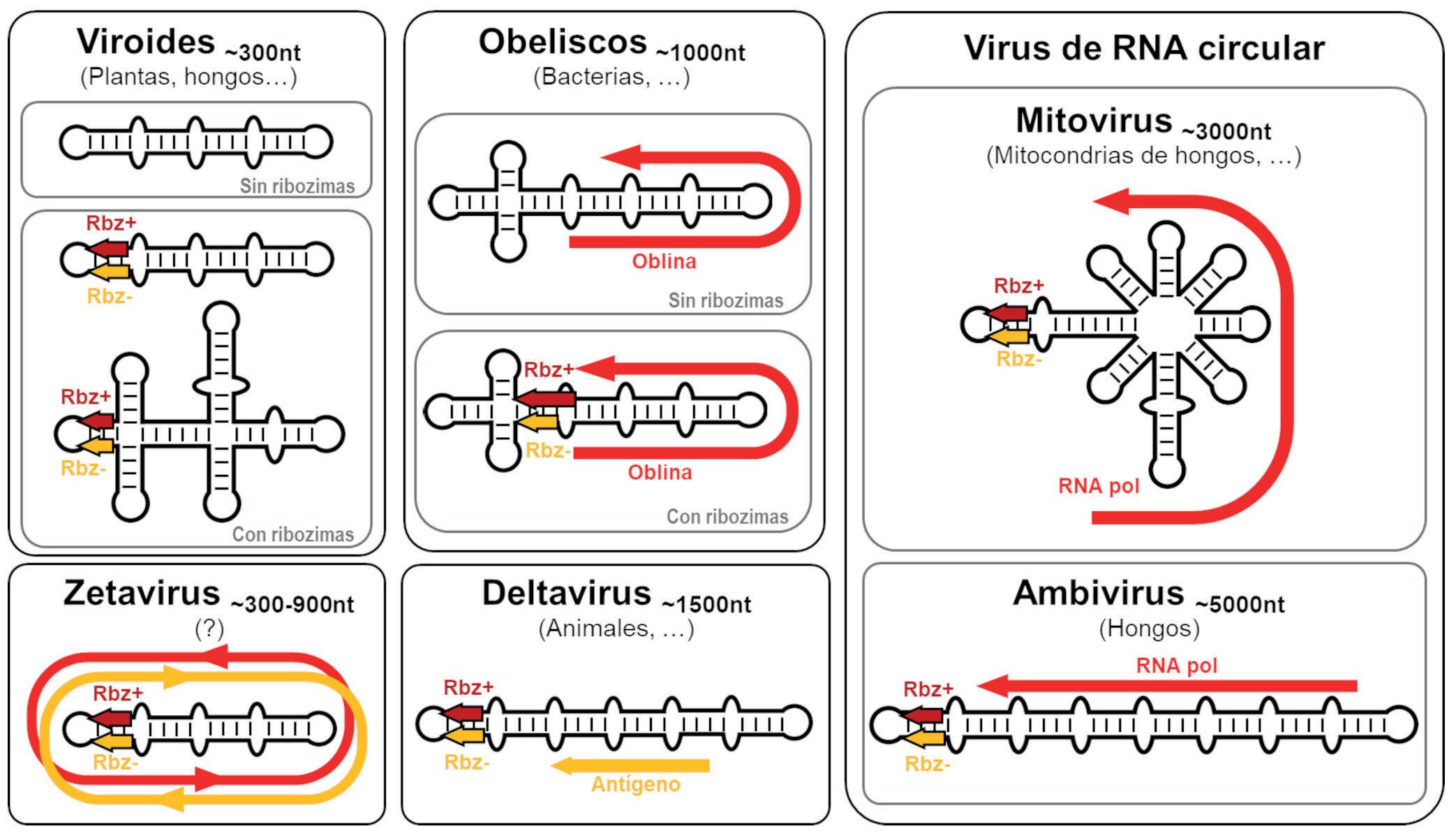
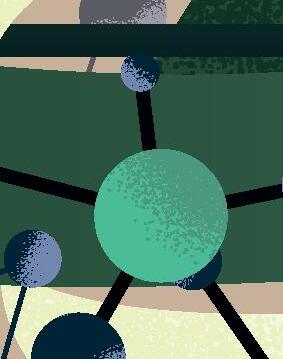
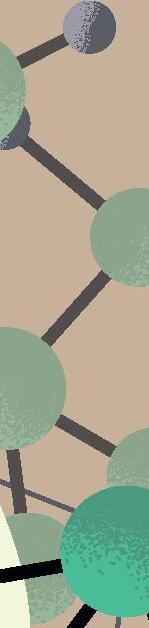
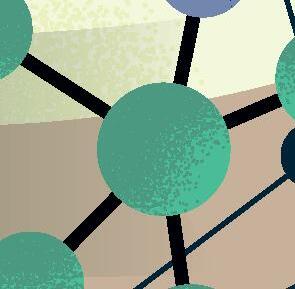
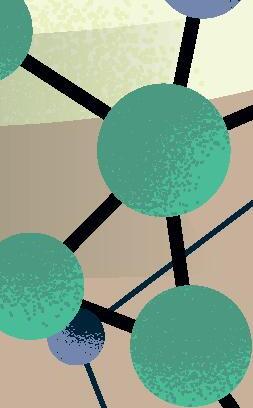
sencillas, o al menos muy alejadas de la complejidad del organismo vivo «canónico» más simple conocido actualmente (típicamente, una bacteria con cientos de genes en un genoma de cientos de miles de pares de bases). Por lógica evolutiva, y siguiendo principios de máxima parsimonia, esos primeros genomas replicantes debieron ser pequeñas moléculas, quizás con decenas o cientos de nucleótidos a lo sumo, que pudieron surgir y organizarse al azar. Solo cuando estas protoentidades adquirieron una mínima capacidad de copiarse, pudo comenzar la carrera para incrementar su complejidad de forma gradual mediante errores de replicación y selección darwiniana, una carrera para persistir en el mismo sitio que aún sigue en nuestros días. Tras al menos cuatro mil millones de años de evolución desde ese origen, el árbol de la vida actual despliega una complejidad asombrosa, desde enormes organismos multicelulares hasta entidades biológicas mínimas y tan extremadamente simples que quizás podrían emular a los primeros replicadores ancestrales. En los peldaños más básicos de esta escala encontramos a los que llamaríamos en general como «replicones de ARN», un grupo de entidades virales y subvirales con genomas de ARN que los sitúa en el límite de la definición de vida. Ejemplos extremos de dicha sencillez son los viroides de plantas descubiertos hace medio siglo (Figura 1) y que consisten en meros genomas desnudos de ARN circular de tan solo unos 300 nucleótidos. Otro ejemplo paradigmático de replicón mínimo es el virus de la Hepatitis D o Deltavirus humano, un ARN circular parecido a los viroides pero de mayor tamaño, unos 1.600 nucleótidos, y considerado el parásito molecular más sencillo conocido en humanos (Figura 1). La existencia de todos estos replicones depende
Figura 1
Representación esquemática de los principales genomas de ARN circular descritos hasta la fecha y ordenados por el tamaño de sus genomas. De izquierda a derecha, y de arriba abajo: viroides, Zetavirus, Obeliscos, Deltavirus y virus de ARN circular de hongos. Rbz: ribozima, RNA pol: RNA polimerasa.
por completo de células vivas a las que parasitan para reproducirse. Un rasgo notable de estos microorganismos es que, curiosamente, emplean ARN como material genético, el cual además llega a catalizar por sí mismo algunas reacciones químicas. Esto es algo excepcional en los seres vivos convencionales, basados en genomas de ADN y con una bioquímica basada casi exclusivamente en las proteínas.
El Mundo de ARN: la llave para el origen de la vida en la tierra
El descubrimiento de ARNs con actividad catalítica o ribozimas en los años 80 revolucionó esta narrativa. Saber que el ARN puede actuar tanto como material genético (replicones virales y subvirales) como desempeñar funciones catalíticas apoyó la hipótesis de un Mundo de ARN primigenio. Según esta idea, existió un escenario prebiótico donde moléculas de ARN autoreplicantes cumplirían simultáneamente funciones genómicas
y biocatalizadoras que pudieron evolucionar hasta organismos cada vez más y más complejos hasta finalizar en seres vivos basados en ADN y proteínas. Entre las pruebas que apoyan este hipotético Mundo tenemos el ribosoma actual, una ribozima que decodifica y cataliza la síntesis proteica en todos los seres vivos (virus incluidos) y que sería un fósil molecular clave de aquel periodo. Otra prueba apoyando una era prebiótica de ARN son los genomas mínimos de ARN actuales, que por su extrema simplicidad y la presencia habitual de ribozimas estarían emulando a los agentes replicantes primigenios. Sin embargo, y durante medio siglo, persistió una paradoja sobre estos sencillos replicones, ¿por qué apenas existían unos pocos ejemplos de agentes viroidales (aproximadamente 50 especies descubiertas en casi 50 años), y todos ellos replicándose en los organismos más complejos y recientes de la biosfera como son plantas o humanos? Todo ello
también empujaba a pensar que estos raros genomas de ARN circular quizás no tuvieran mucho que ver con entidades replicantes primigenias, sino que su origen podría ser mucho más reciente, o quizás ambas cosas.
Replicones de ARN circular en todas partes
La solución a esta paradoja ha llegado de manera progresiva y sorpresiva en los últimos años. Así, en 2018 varios trabajos desvelaron casi simultáneamente que el único Deltavirus conocido, el humano, en realidad no era único. Hasta seis nuevos genomas similares se detectaron en serpientes, patos, anfibios, peces e incluso insectos como las termitas. Curiosamente, alguno de estos nuevos Deltavirus animales no empleaban las clásicas ribozimas del Deltavirus humano sino las ribozimas descritas en viroides de plantas, lo que evolutivamente permitía conectar aún más a todos estos replicones mínimos. En 2022, el análisis
bioinformático de las ingentes bases de datos genéticos recopiladas durante décadas reveló la existencia de docenas de ejemplos de Deltavirus en mamíferos y otros animales, tanto vertebrados como invertebrados, confirmando que estos extraños agentes infecciosos están en realidad ampliamente distribuidos en el reino animal. Pero no solo eso, también se detectaron genomas de Deltavirus ancestrales en muestras medioambientales de suelos, lagos u océanos entre otros, sugiriendo que estos replicones de ARN estarían utilizando como hospedadores otros organismos más sencillos que los animales, y muy probablemente unicelulares. Más sorprendente aún fue descubrir que muchas de estas muestras medioambientales escondían también cientos de curiosos genomas mínimos (~300 a 900 nt) de ARN circular con ribozimas, a medio camino entre viroides y Deltavirus, y que fueron bautizados como Zetavirus (Figura 1). Los Zetavirus muestran unas características asombrosas, empezando por su matemático tamaño, siempre múltiplo de tres nucleótidos. Además, tienen capacidad para codificar una ribozima y una proteína por cada una de sus dos polaridades (positiva y negativa). Pero no solo eso, las dos proteínas predichas carecen de codones de parada, por lo que al tratarse de genomas circulares con un tamaño múltiplo de tres, su traducción en teoría no pararía nunca, produciendo polipéptidos «infinitos» compuestos de la repetición de la secuencia codificada por el Zetavirus. Por ahora, se desconocen los posibles huéspedes de estos intrigantes ARNs circulares, si es que en realidad son agentes replicantes. Lo que sí se deduce de sus curiosas características es que podemos estar ante entidades con un novedoso lenguaje genético implicado quizás
en parasitar los ribosomas de las células en las que habitan.
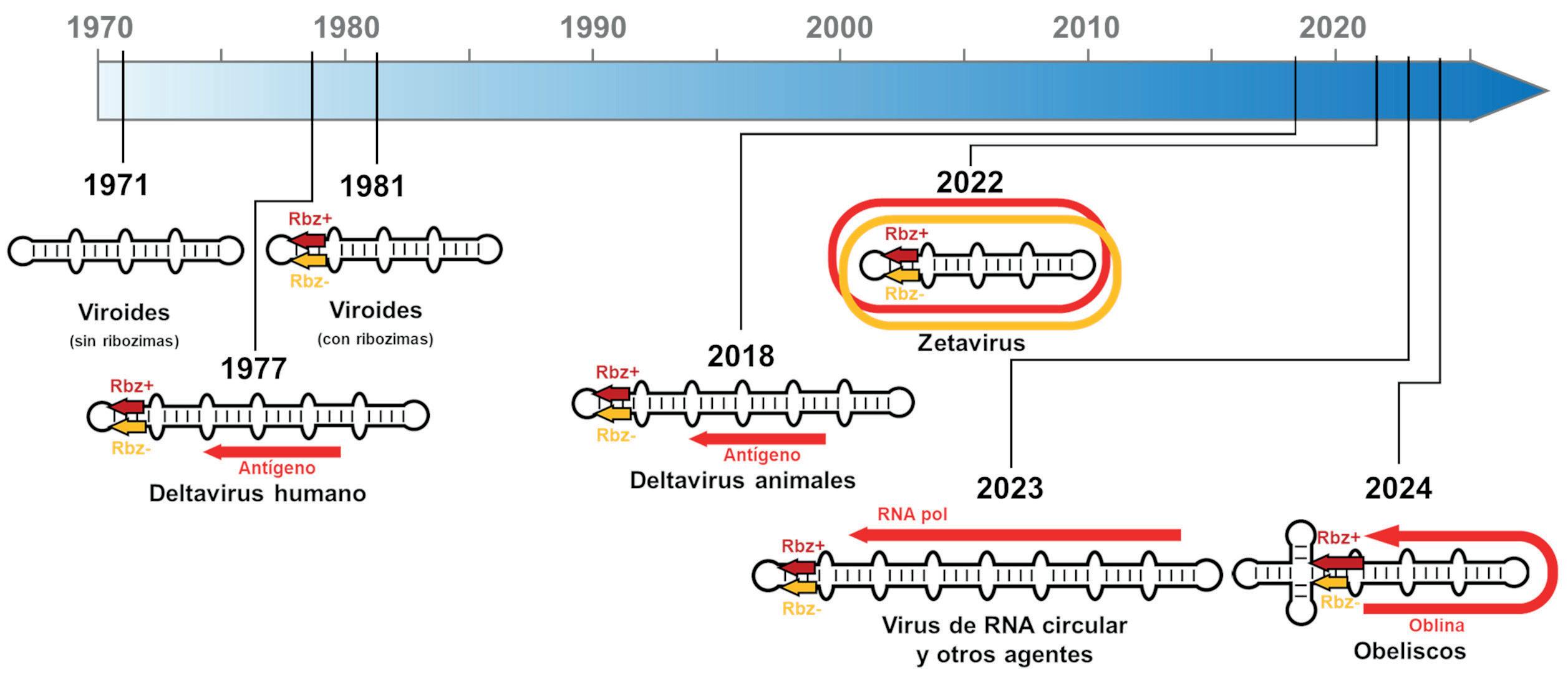
En conjunto, todos estos resultados empujaban en la misma dirección: existen muchos más ARNs circulares tipo viroide en la naturaleza que han pasado desapercibidos desde su descubrimiento en plantas allá por los años 70 del siglo pasado (Figura 2) En el año 2023, dos trabajos con fuerte componente bioinformática terminaron de confirmar estas sospechas al extender en varios órdenes de magnitud lo que hasta entonces era una minúscula familia de agentes de ARN circular. El análisis computacional de miles de estudios de metagenómica y metatranscriptómica permitió detectar decenas de miles de nuevos genomas de ARN circular con ribozimas. Prácticamente la mitad de esta colección de ARNs eran ejemplos de los pequeños Zetavirus. Sin embargo, también se detectaron en diversas especies de hongos la existencia de agentes no codificantes similares a viroides de plantas, confirmando la existencia de viroides fúngicos descrita por otros grupos. También se detectaron varias familias de ARNs de gran tamaño (entre 3.000 y 5.000 nucleótidos) que resultaron ser los primeros ejemplos de virus con genomas de ARN circular con ribozimas. Estos correspondieron a dos familias de virus de hongos: los Mitovirus (genoma de ~3.000 nt) y los Ambivirus (genoma de ~5.000 nt) (Figura 1). Como todo virus de ARN, éstos también codifican para la clásica ARN polimerasa dependiente de ARN, y a su vez, también contenían en sus dos polaridades algunas de las ribozimas típicas de viroides y Deltavirus, así como otras familias de ribozimas nunca antes descritas en agentes viroidales. Se trataba por tanto de agentes infecciosos híbridos, mitad virus de ARN mitad agente viroidal.
A pesar de la enorme cantidad de novedades encontradas en unos pocos años, estos descubrimientos parecen ser solo el principio de todo un nuevo Mundo de novedosos replicones de ARN. Los resultados desvelados indican que nos adentramos en una terra incognita compuesta por una vasta y variada colección de ARNs circulares que necesitará años de investigación para conocer con detalle, tanto sus posibles hospedadores y modos de replicación como los secretos moleculares que atesoran. En esta línea de trabajo, en 2024 se publicó la caracterización bioinformática y molecular de toda una nueva familia de genomas de ARN circular que ha resultado especialmente reveladora. Estos nuevos agentes fueron inicialmente detectados en datos de secuenciación de ARN de muestras de heces humanas, donde se comprobó que aproximadamente el 7 % de los individuos contenían unos curiosos ARNs circulares de tamaño en torno a los 1.000 nucleótidos. Dichos ARNs adoptaban una estructura de cuasi-varilla similar a la de viroides y Deltavirus, por lo que fueron bautizados como Obeliscos (Figura 1). Como característica diferenciadora de los Obeliscos se encontró que podían codificar al menos una proteína de unos 200 residuos, denominadas Oblinas, que no mostraban homología alguna con proteínas conocidas, por lo que su papel biológico es aún un completo misterio.
A través de aproximaciones bioinformáticas se detectó la existencia de cerca de 30.000 especies distintas de Obeliscos presentes en todo tipo de entornos y nichos ecológicos, pero muy notablemente en los microbiomas de animales de todo tipo. Entre todas estas especies de Obeliscos se encontró una subfamilia caracterizada por codificar en sus dos polaridades las típicas
Figura 2
Descubrimiento de los principales replicones de ARN circular por orden cronológico. En los años 70 del siglo XX se describen los primeros viroides de plantas y el Deltavirus humano. Durante 40 años únicamente se encontraron nuevas especies de viroides de plantas, hasta que en 2018 se describen nuevos Deltavirus animales, y posteriormente los Zetavirus (2022), Mitovirus y otros virus circulares (2023) y Obeliscos bacterianos (2024), entre otros.
ribozimas de los viroides de plantas, reforzando posibles conexiones evolutivas entre estos minúsculos agentes de ARN circular. Más aún, al menos una especie de Obelisco se halló en cepas de la bacteria Streptococcus sanguinis aislada de la cavidad oral humana. Este resultado permitió confirmar la abundante acumulación de Obeliscos en bacterias, lo que constituye el primer ejemplo de un agente de ARN circular tipo viroidal replicándose en un organismo procariótico. Con estos resultados se está por fin cerrando el círculo que comenzó hace ya 50 años con el descubrimiento pionero de un agente infeccioso atípico de ARN circular en plantas superiores. Ahora ya podemos decir que esos minúsculos genomas, históricamente considerados anomalías biológicas, tan solo eran una pequeña muestra de todo un nuevo «Mundo» de ARNs circulares que habitan a lo largo y ancho de este planeta, y cuyo origen muy probablemente haya que buscarlo hace miles de millones de años, tras los cuales aún parecen persistir.
Para leer más
Chang WS, Pettersson J, Le Lay C, Shi M, Lo N, Wille M, Eden JS, Holmes EC. Novel hepatitis D-like agents in vertebrates and invertebrates. Virus Evolution 5 (2019) vez021. https://doi.org/10.1093/ve/ vez021
de la Peña M, Ceprián R, Casey JL, Cervera A. Hepatitis delta virus-like circular RNAs from diverse metazoans encode conserved hammerhead ribozymes. Virus Evolution 7 (2021) veab016. https:// doi.org/10.1093/ve/veab016
Edgar RC, Taylor J, Lin V, Altman T, Barbera P, Meleshko D, Lohr D, Novakovsky G, Buchfink B, Al-Shayeb B, Banfield JF, de la Peña M, Korobeynikov A, Chikhi R, Babaian A. Petabase-scale sequence alignment catalyses viral discovery. Nature 602 (2022)1–6. https:// doi.org/10.1038/s41586-021-04332-2
Forgia M, Navarro B, Daghino S, Cervera A, Gisel A, Perotto S, Aghayeva DN, Akinyuwa MF, Gobbi E, Zheludev IN, Edgar RC, Chikhi R, Turina M, Babaian A, Di Serio F, de la Peña M. Hybrids of RNA viruses and viroid-like elements replicate in fungi. Nature Communications 14 (2023) 1–11. https://doi.org/10.1038/ s41467-023-38301-2
Lee BD, Neri U, Roux S, Wolf YI, Camargo AP, Krupovic M, RNA Virus Discovery Consortium, Simmonds P, Kyrpides N, Gophna U, Dolja VV, Koonin EV. Mining metatranscriptomes reveals a vast world of viroid-like circular RNAs. Cell 186 (2023) 646-661.e4. DOI: 10.1016/j.cell.2022.12.039
Zheludev IN, Edgar RC, Lopez-Galiano MJ, de la Peña M, Babaian A, Bhatt AS, Fire AZ. Viroid-like colonists of human microbiomes. Cell 187 (2024) 6521-6536.e18. https://doi.org/10.1016/j.cell.2024.09.033
s ,
GUARDIANES DEL EQUILIBRIO GENÉTICO VEGETAL
Catharina Merchante y Pablo A. Manavella
Instituto de Hortofruticultura Subtropical y Mediterránea «La Mayora» (IHSM «La Mayora»), Universidad de Málaga (UMA) - CSIC, Málaga
https://doi.org/10.18567/sebbmrev_226.202509.dc3
Desde su descubrimiento en plantas, los microARNs (miARNs) han emergido como protagonistas cruciales en la regulación de la expresión génica. Estas pequeñas moléculas de ARN no codificante, de alrededor de 21 nucleótidos, controlan una amplia gama de procesos biológicos esenciales, desde el desarrollo embrionario hasta la respuesta a factores ambientales. Su papel es comparable al de los directores de orquesta que aseguran la armonía de toda una sinfonía celular. Por estas características, los miARNs han captado la atención de la comunidad científica, revelando un mundo molecular de alta precisión y extraordinaria complejidad que permiten a los organismos adaptarse a su entorno. Esto es particularmente importante en plantas dado que, por su naturaleza sésil, no pueden escaparse de ambientes hostiles. Y es por ello que los procesos de síntesis y acción de los miARNs en plantas han evolucionado de forma divergente a los de animales, incluyendo muchas particularidades fascinantes aunque también muchos puntos de conexión.
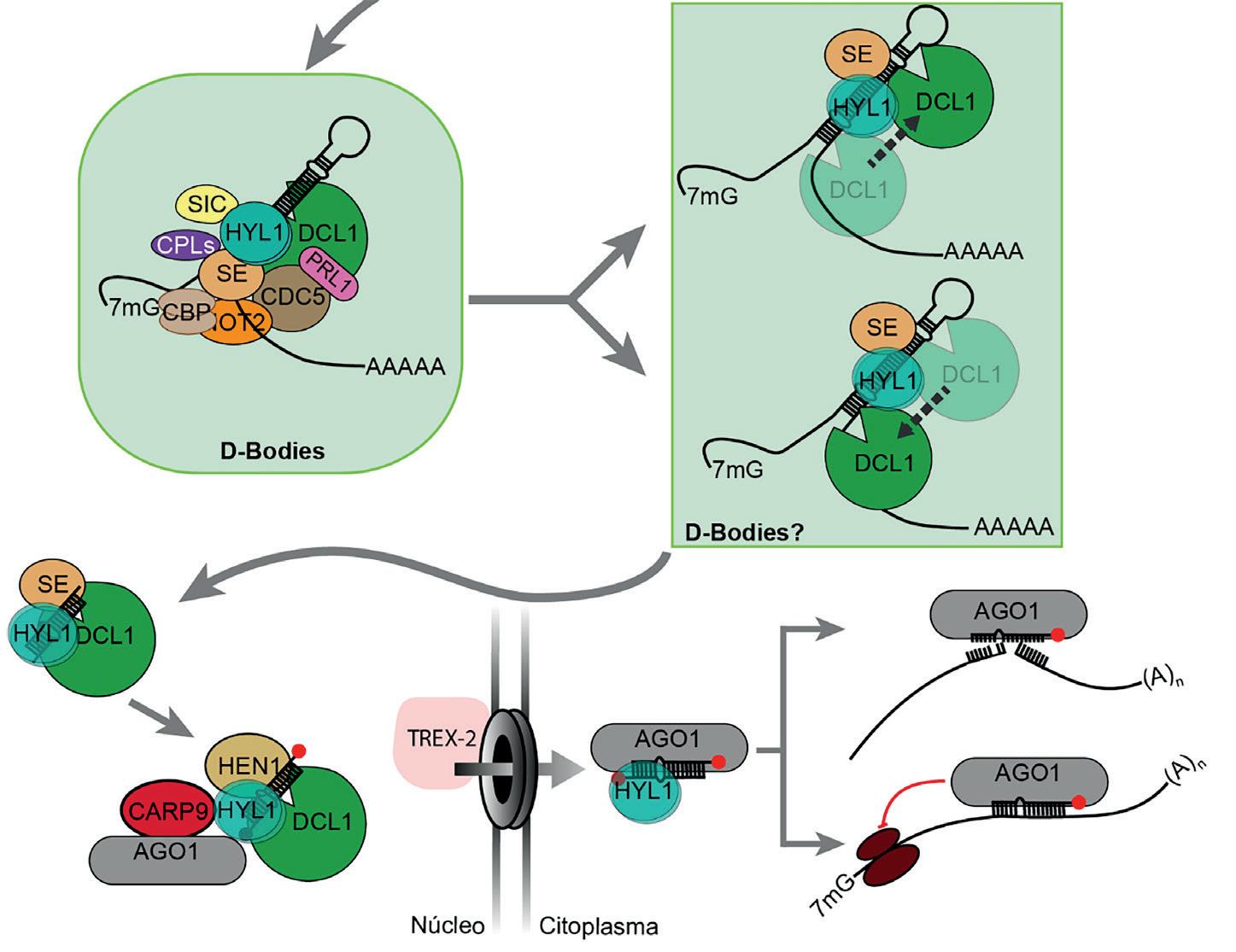
Como la mayoría de los procesos que permiten la vida eucariota, todo comienza en el núcleo celular. La producción de miARNs en plantas empieza con la transcripción de genes MIRNA por la ARN polimerasa II, que es la misma polimerasa que transcribe los genes que codifican proteínas. Estos transcritos primarios, llamados «pri-miARNs», presentan una característica muy especial, y es que se pliegan sobre sí mismos generando estructuras de ARN de doble cadena del tipo horquilla, y esto es esencial para su reconocimiento por el complejo de procesamiento. Éste es un complejo de proteínas conocido como microprocesador y cuya pieza central es la enzima DICER-LIKE1 (DCL1, por sus siglas en inglés). DCL1, asistida por los cofactores SERRATE (SE) y HYPONASTIC LEAVES1 (HYL1, por sus siglas en inglés), reconoce esta estructura en horquilla y comienza el proceso de producción de miARNs (Figura 1). A diferencia de animales, donde la producción de miARNs está compartimentada en etapas nucleares y citoplasmáticas, en plantas este proceso
ocurre íntegramente en el núcleo. El resultado de este procesamiento es la generación de un dúplex miARN que será posteriormente metilado para su estabilización. Se ha demostrado que el procesamiento de estos pri-miRNAs puede iniciarse mientras el primiARN está todavía sintetizándose, en un mecanismo denominado procesamiento cotranscripcional. Factores como el complejo ‘Elongator’ y ciertas modificaciones en la ARN polimerasa II favorecen que el microprocesador se asocie de manera temprana al pri-miARN naciente, lo que permite una regulación más eficaz y un procesamiento casi inmediato de los precursores. La producción de miARNs no ocurre de manera aislada. Existe una estrecha conexión entre la biogénesis de miARNs y otros procesos nucleares, especialmente el splicing de ARN mensajero. Proteínas como SERRATE y componentes del Complejo ‘Cap-binding’ (CBC, por sus siglas en inglés) actúan tanto en el procesamiento de miARNs como en el splicing, lo que sugiere que existen centros nucleares
multifuncionales donde ambos procesos están coordinados. Este acoplamiento permite una regulación integrada de la expresión génica, lo que optimiza la respuesta celular frente a cambios en el entorno, y contribuye al mantenimiento de la homeostasis.
El procesamiento de los pri-miARNs no es un mecanismo rígido ni universal, sino que depende de características intrínsecas de cada pri-miARN, como su estructura secundaria, modificaciones químicas, o su asociación con proteínas accesorias. Este ajuste fino permite a la célula regular, de forma específica, qué miARNs se producen en cada tipo celular o en respuesta a señales externas concretas. De hecho, aunque el procesamiento acoplado a la transcripción ofrece muchas ventajas como un menor consumo energético y una mayor eficiencia, muchos miARNs no se procesan de esta manera. En esos casos, una vez transcritos, muchos pri-miARNs se dirigen a estructuras subnucleares especializadas denominadas «Dicing bodies» o «D-bodies». En ellos se concentran componentes del microprocesador, como DCL1, HYL1 y SE, responsables de llevar a cabo el corte preciso de los pri-miARNs (Figura 1)
Aunque los D-bodies se han asociado tradicionalmente con el procesamiento de pri-miARNs, nuevas evidencias apuntan a que su papel podría ser más amplio, actuando como reservorios de miARNs o centros donde regular su maduración y estabilidad. La formación de estos D-bodies depende de la correcta interacción entre varios factores, entre los cuales destaca HYL1 como pieza clave para su ensamblaje.
Pero ¿quién es HYL1 y por qué nombramos tanto a esta proteína?
HYL1 es una proteína de unión al ARN, o, más específicamente, es una proteína de unión a ARN de doble cadena. Sí, no es un error de
Figura 1
Representación esquemática de la vía de biogénesis de miARNs en plantas (modificada de Achkar y col., 2016).
escritura, no solo el ADN forma hebras bicatenarias, el ARN también. De hecho, el ARN de doble cadena tiene, de forma general, una mayor energía de disociación que el ADN de doble cadena, es decir, es más difícil separar sus hebras. HYL1 tiene gran afinidad por estas estructuras y se une a ellas mediante dos dominios de unión a ARN. Esta alta afinidad hace que HYL1 sea una de las primeras proteínas en reconocer a los pri-miARN nacientes, y esto permite la unión del resto de las proteínas del complejo, y guía a DCL1 hacia los sitios de corte precisos para liberar el dúplex de miARN, al que también estabiliza. En este contexto, HYL1 destaca como una proteína multifuncional clave. Además de su rol como cofactor en el procesamiento preciso de pri-miARNs, HYL1 participa en la regulación de la transcripción de genes MIRNA, protege los pri-miARNs de su degradación prematura y facilita el transporte del dúplex maduro hacia
el citoplasma. De forma sorprendente, también interviene en la inhibición de la traducción de genes diana en el retículo endoplásmico, lo que refuerza su papel como un verdadero centro regulador en la vía de los miARNs. Como es de esperar para una proteína con tanta importancia en la vía, la actividad de HYL1 no es constante, sino que está finamente regulada por mecanismos postraduccionales, principalmente por fosforilación. La forma activa de HYL1, la que es capaz de promover un procesamiento preciso de los pri-miARNs, es su versión no fosforilada. Existe un equilibrio entre ambas formas, no fosforilada (activa) y fosforilada (inactiva), que está regulado por diversas quinasa y fosfatasas, y esto permite a la planta ajustar rápidamente su actividad ante cambios en el ambiente. Un ejemplo de esta regulación se da ante variaciones de luz. En condiciones de oscuridad, HYL1 se degrada, lo que reduce
DOSIER CIENTÍFICO
la producción de nuevos miARNs. Sin embargo, una fracción de HYL1 permanece inactiva en el núcleo como «reserva», lista para ser reactivada cuando vuelva la luz, lo que asegura una rápida respuesta adaptativa.
Pero ¿qué pasa después de su síntesis y para qué sirven los miARNs? Como mencionábamos, el procesamiento de pri-miARNs da lugar a los miARNs maduros de doble hebra. Posteriormente, estos se metilan por HUA ENHANCER1 (HEN1, por sus siglas en inglés) y se cargan preferentemente en proteínas del tipo ARGONAUTE (AGO). En especies como Arabidopsis thaliana, se cargan mayoritariamente en AGO1 (Figura 1). Una vez cargado en AGO1, el miARN funcional se incorpora al complejo RISC (por sus siglas en inglés, RNA-Induced Silencing Complex), mientras que la hebra complementaria suele ser degradada. Un miARN asociado a AGO1 puede actuar en el núcleo o exportarse al citoplasma, donde ejerce su función reguladora sobre ARN mensajeros (ARNm), induciendo su silenciamiento ya sea por degradación o inhibición de la traducción.
De forma llamativa, no todos los miARNs se cargan inmediatamente en AGO1. Existe un conjunto de miARNs libres en el citoplasma que podría participar en funciones no autónomas, moviéndose de una célula a otra y coordinando respuestas a nivel de tejido u órgano. Se ha propuesto que HYL1 pudiera intervenir en esta selección de destinos, acompañando a los miARNs en su salida del núcleo y participando tanto en su transferencia a AGO1 como en su movilidad intercelular, aunque esta hipótesis requiere confirmación experimental.
Ya sea en su célula de origen o en otra actuando remotamente, el modo de acción principal de los miARNs en plantas es la represión génica post-transcripcional,
que puede manifestarse de dos formas:
Corte endonucleolítico del ARNm diana: este es el mecanismo predominante, favorecido por la alta complementariedad entre el miARN y su blanco. Este proceso está catalizado por la actividad ribonucleasa de AGO1 y, en última instancia, conlleva la desestabilización del ARNm diana, su degradación y, en consecuencia, una disminución de los niveles de la proteína codificada por el gen diana.
Represión traduccional: en algunos casos, especialmente cuando la complementariedad entre un miARN y su diana no es perfecta o hay interferencia estructural, el miARN puede inhibir la traducción del ARNm sin inducir su degradación. Aunque este mecanismo se consideró inicialmente secundario en plantas, estudios recientes han demostrado su relevancia en varios contextos biológicos, como el desarrollo o la respuesta a estrés. La inhibición puede producirse tanto a nivel de iniciación como de elongación de la traducción, y puede implicar la asociación del RISC con polirribosomas. Una particularidad de esta forma de silenciamiento es que es reversible, ya que una disociación del miARN a su diana puede restablecer su traducción, lo que es imposible si el mecanismo de silenciamiento conlleva el corte del mensajero diana.
El resultado de este tipo de regulación, sea por un mecanismo o por el otro, es muy variable y por supuesto depende de los genes diana. Sin embargo, una particularidad de los miARNs en plantas es que muestran una notable preferencia evolutiva a silenciar factores de transcripción, lo que conlleva que sus efectos regulatorios sean variados y puedan amplificarse por la desregulación de cientos de genes diana de estos factores de transcripción.
Algunos de los ejemplos mejor
caracterizados de miARNs activos en plantas incluyen:
• miR156: regula la transición de fase juvenil-adulta controlando genes de la familia SPL (por sus siglas en inglés, SQUAMOSA PROMOTER BINDING
PROTEIN-LIKE). Su acumulación es alta en etapas tempranas del desarrollo y disminuye con la edad, lo que permite la expresión de genes que promueven la floración y el desarrollo reproductivo.
• miR172: actúa en concierto con miR156 pero en sentido opuesto. Reprime genes como APETALA2 y otros reguladores de la identidad floral. Está implicado en la correcta determinación del momento de floración.
• miR160 y miR167: regulan genes de la familia AUXIN
RESPONSE FACTOR (ARF, por sus siglas en inglés), modulando respuestas al ácido indolacético (auxina), una fitohormona clave en el desarrollo radicular,formación de órganos y tropismos.
• miR393: participa en respuestas a estrés biótico regulando receptores de auxinas como TRANSPORT INHIBITOR
RESPONSE 1 (TIR1, por sus siglas en inglés), inhibiendo así vías de crecimiento que podrían ser contraproducentes durante una infección.
• miR168: tiene una función singular ya que actúa sobre el propio sistema de silenciamiento. Regula la expresión de AGO1, estableciendo un bucle de retroalimentación negativa que modula la abundancia del complejo RISC. Esto permite un equilibrio dinámico en la intensidad del silenciamiento mediado por otros miARNs.
• miR319: el primer miRNA descubierto en plantas. Regula genes de la familia TCP (por sus siglas en inglés, TEOSINTE BRANCHED/CYCLOIDEA/ PCF), que están implicados en
la proliferación celular, el crecimiento de hojas y el desarrollo floral. Su sobreexpresión produce hojas con márgenes serrados y participa en la integración entre desarrollo y señalización hormonal.
• miR396: controla la proliferación celular mediante la regulación de genes de la familia GRF (por sus siglas en inglés, GROWTH REGULATING FACTOR). Su sobreexpresión restringe el crecimiento de hojas y órganos, y se ha vinculado también a la respuesta frente al estrés y al equilibrio entre proliferación y diferenciación celular.
Además de estas funciones intracelulares, algunos miARNs son móviles y participan en la señalización a larga distancia. Por ejemplo, miR399, un regulador clave del metabolismo del fósforo, se produce en las hojas y se transporta a la raíz donde reprime a PHO2, un gen que regula la homeostasis de este nutriente. Otro ejemplo notable es miR390, que participa en la producción de ta-siRNAs (ARNs pequeños trans-activados) a partir del locus TAS3. En este caso, miR390 no causa la degradación directa de su mRNA diana, sino que guía a AGO7 para activar una vía secundaria de silenciamiento que regula los genes ARF2, ARF3 y ARF4, implicados en el desarrollo lateral y la respuesta a auxinas.
Estos ejemplos ilustran la diversidad funcional de los miARNs en plantas, actuando como nodos centrales en redes de regulación genética que integran señales ambientales, hormonales y de desarrollo.
Como es de esperar, la actividad reguladora de los miARN debe estar sujeta a regulación. Así, una vez en el citoplasma, la estabilidad de los miARNs y su actividad en el complejo RISC no son indefinidas. La degradación, o el recambio, de miARNs es un proceso regulado y dinámico que garantiza la
plasticidad en la regulación génica. En este sentido, la enzima SDN (por sus siglas en inglés, Small RNA Degrading Nuclease) participa en la degradación progresiva del extremo 3’ de los miARNs cuando dejan de estar protegidos por metilación o por AGO1. Factores como HEN1 SUPPRESSOR 1 (HESO1, por sus siglas en inglés) o RNA URIDYLYLTRANSFERASE 1 (URT1, por sus siglas en inglés) pueden añadir uridinas codificadas en el extremo 3’ del miARN, lo que los marca para su degradación. A su vez, la estabilidad del complejo RISC también puede ser modulada por la interacción con proteínas accesorias, por competencia entre miARNs por el acceso a las proteínas ARGONAUTE disponibles, por cambios en la expresión de AGO1 o directamente por su degradación. Relacionado con esto, es interesante que siendo el silenciamiento génico el principal mecanismo de defensa de las plantas contra infecciones virales, los virus de plantas empleen estrategias relacionadas con estos mecanismos para bloquear estas vías de silenciamiento para favorecer la infección. Por ejemplo, los polerovirus producen un supresor de silenciamiento llamado P0 que se une e induce la degradación de proteínas ARGONAUTE. Por su parte, el supresor de silenciamiento P19
Para leer más
de tombusvirus actúa uniéndose específicamente a ARNs pequeños de 19 a 21 nucleótidos que funcionan como ARN interferentes pequeños (siRNA) en el sistema de silenciamiento por ARN. El estudio de los miARNs en plantas pone de manifiesto un entramado molecular dinámico y sofisticado. Lejos de actuar como simples silenciadores, los miARNs y su maquinaria asociada constituyen un mecanismo capaz de integrar señales internas y externas para regular tanto el desarrollo como la adaptación de las plantas.
Desde su compleja biogénesis en el núcleo hasta su acción específica sobre genes diana, pasando por mecanismos de transporte, modulación y degradación, estas pequeñas moléculas son piezas clave en la maquinaria genética vegetal. Comprender su funcionamiento no solo amplía nuestro conocimiento sobre la biología fundamental de las plantas, sino que abre nuevas posibilidades para la mejora agrícola, la resistencia a estrés y el desarrollo de herramientas biotecnológicas innovadoras. En un mundo donde las plantas son esenciales para la vida y enfrentan condiciones cada vez más extremas, descifrar el lenguaje de los miARNs puede ser una de las llaves para garantizar nuestra sostenibilidad futura.
Achkar NP, Cambiagno DA, Manavella PA. miRNA Biogenesis: A Dynamic Pathway. Trends in Plant Science 21 (2016) 1034-1044. doi: 10.1016/j.tplants.2016.09.003
Gonzalo L, Giudicatti AJ, Manavella PA. HYL1’s multiverse: A journey through miRNA biogenesis and beyond. Current Opinion in Plant Biology 80 (2024)102546. doi: 10.1016/j.pbi.2024.102546
Mencia R, Gonzalo L, Tossolini I, Manavella PA. Keeping up with the miRNAs: current paradigms of the biogenesis pathway. Journal of Experimental Botany 74 (2022) 2213-2227. doi: 10.1093/jxb/erac322
MÁS ALLÁ DEL DOGMA CENTRAL DE LA BIOLOGÍA MOLECULAR:
EL ARN NO
CODIFICANTE Y
EL QUE CODIFICA
Juan Carlos García-Soriano y Puri Fortes Centro de Investigación Médica Aplicada (CIMA), Universidad de Navarra (UNAV). Centro de Investigación Biomédica en Red de Enfermedades Hepáticas y Digestivas (CIBERehd). Cancer Center de la Clínica Universidad de Navarra (CCUN), Pamplona https://doi.org/10.18567/sebbmrev_226.202509.dc4
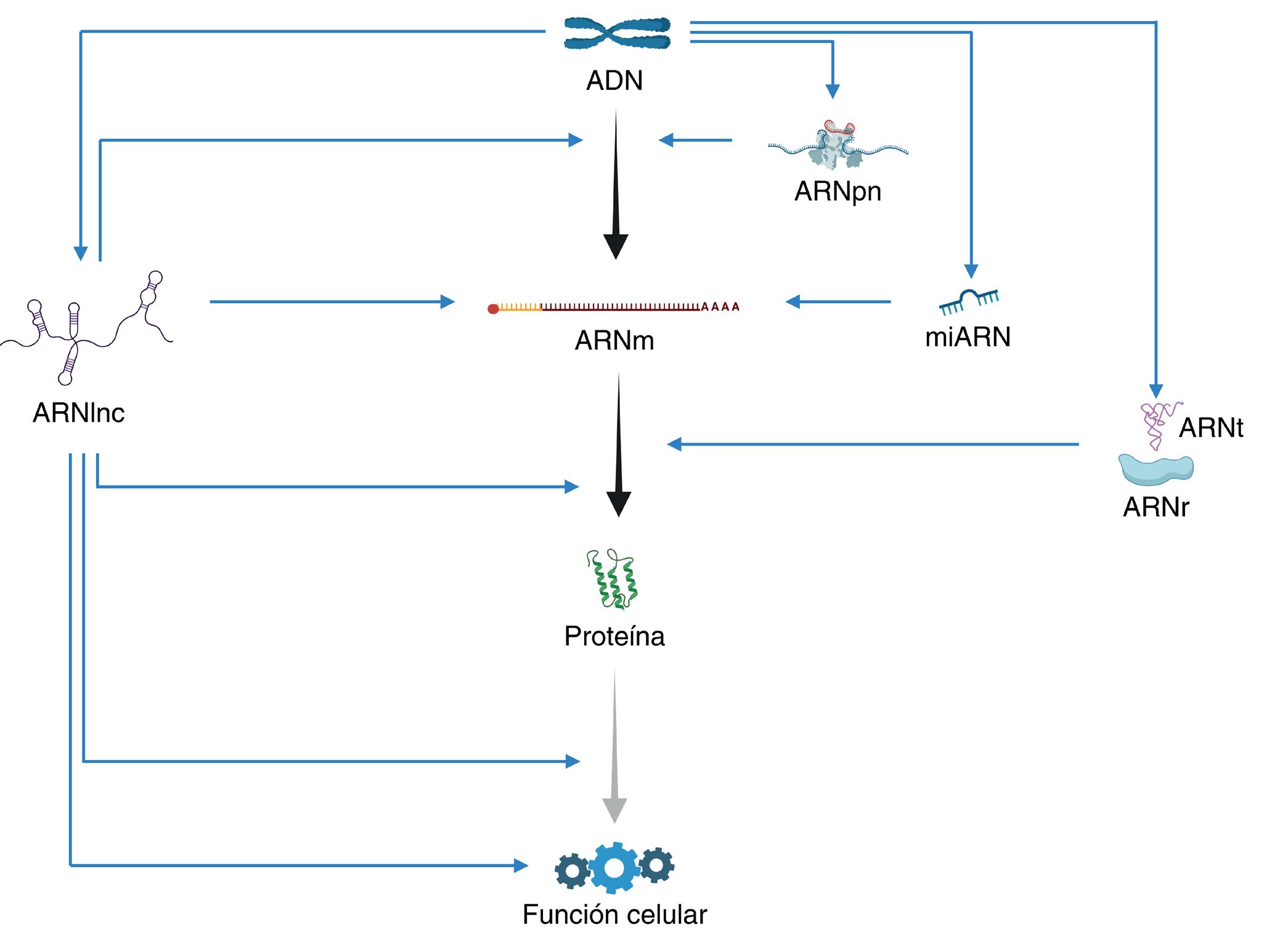
En 1958 Francis Crick postuló el Dogma central de la Biología Molecular, una idea que Jacques Monod inmortalizó diciendo: «lo que ocurre en las bacterias, ocurre en el elefante». Este dogma establece un flujo de información del ADN al ARN y de ahí a las proteínas (Figura 1). El ADN se transcribe para dar ARN mensajero (ARNm) utilizando un código similar de cuatro nucleótidos (nt) y éste se traduce a proteínas cambiando la información a 20 aminoácidos (aa). Este dogma ponía en el centro de la ecuación al ARNm e ignoraba a otros ARNs que no se traducen, y por ello son llamados «no codificantes». Estos incluyen, por ejemplo, el ARN de transferencia (ARNt), el ribosómico (ARNr) o los pequeños nucleares (ARNpn), que son necesarios para la maduración del ARNm o su traducción a proteínas. Más adelante, se descubrieron los micro ARNs (miARN). Habían pasado desapercibidos por su pequeño tamaño (algunos con tan solo 21 nt). Sin embargo, son numerosos, se expresan a altos niveles y resultaron ser extraordinariamente relevantes para controlar la expresión génica de los ARNm con los que interaccionan. Algo parecido ocurrió con ARNs más largos no codificantes (ARNlnc).
Los ARNlnc se identificaron en estudios transcriptómicos, una poderosa tecnología que permite mapear todos los ARNs producidos en una célula. Esta técnica mostró que casi todo el ADN de la célula se transcribe en ARN. Para sorpresa de la comunidad científica se encontró que, de todo este ARN, el ARNm constituye tan solo un 2 %. El 90 % restante, es ARN no codificante que incluye, además de ARNt, ARNr, o miARN, a los ARNlnc. Estos últimos son los más numerosos. Los últimos análisis del genoma humano cuantifican casi 20.000 genes que
Figura 1
Dogma Central de la Biología Molecular con ARNs diversos. En el centro, con flechas negras, se muestra el flujo de información bajo la visión tradicional del Dogma: el ADN se transcribe para dar lugar a ARN mensajero (ARNm) y este se traduce a proteína. A la derecha de esta línea se muestran los ARN pequeños nucleares (ARNpn; que participan en la maduración del ARN), los ARN de transferencia y ribosomales (ARNt y ARNr; que participan en la traducción) y los microARN (miARN; que disminuyen la estabilidad y la traducción de los ARNm con los que interaccionan). A la izquierda se muestran los ARN largos no codificantes (ARNlnc), que ejercen una gran variedad de funciones de regulación de la expresión génica. Tanto las proteínas como los ARNlnc pueden formar parte de maquinarias celulares no relacionadas directamente con la expresión génica.
codifican proteínas (se transcriben a ARNm) y unos 50.000 que producen ARNlnc. Esta cifra incluye también a los pseudogenes, antiguos genes codificantes que perdieron esa capacidad. Como la mayoría de los estudios hasta la fecha se habían enfocado en los genes codificantes (esos 20.000) y en las proteínas que estos producen, los análisis transcriptómicos indicaron que nos falta por estudiar lo que se produce en la mayor parte del genoma.
Enseguida aparecieron detractores a la idea de que esto fuese relevante. Los críticos argumentaban que los bajos niveles de producción de los ARNlnc y su escasa conservación entre especies los convertían en «basura» genética, sin funciones importantes. Esto de la conservación es fascinante. La mayoría de los genes que codifican proteínas, esos 20.000 en humanos, son muy parecidos a los de organismos tan diversos como gusanos o moscas, a pesar de las
evidentes diferencias en complejidad evolutiva. Sin embargo, los ARNlnc son muy distintos entre especies y su número aumenta con la complejidad evolutiva: hay menos en bacterias, más en moscas, muchos más en mamíferos, y los primates somos los que tenemos más genes de ARNlnc en nuestro genoma. Se postula que esta abundancia de ARNlnc en primates podría deberse, en parte, a una infección masiva de retrovirus en un ancestro común. Estos virus
DOSIER CIENTÍFICO
insertaron sus genomas en el de nuestro antepasado, colonizándolo y dejando innumerables réplicas de genes virales en prácticamente la mitad de su genoma. Lo que a primera vista parecería perjudicial, pudo haber conferido una ventaja evolutiva tan única, que hoy somos los herederos de ese primate ancestral.
Nuestro genoma no tiene mucha «basura», sino, en gran medida, restos de virus que, modificados por la evolución, nos han dotado de miles de ARNlnc diferentes. Una teoría evolutiva muy interesante, conocida como «same brick, different building» (mismo ladrillo, diferente edificio), que sugiere que las proteínas están muy conservadas porque actúan como los «ladrillos» de los «edificios» que formamos los seres vivos. Así como una caseta de perro y la catedral de León pueden construirse con ladrillos similares, la complejidad de un organismo no reside tanto en los «ladrillos» sino en el «arquitecto» o regulador que dicta su disposición específica en cada especie. Es tentador pensar que los ARNlnc podrían ser los «arquitectos», los reguladores que controlan qué ladrillo (proteína) se produce en cada tejido y dónde debe colocarse. Esto ofrece una explicación elegante a la asombrosa diversidad biológica que se obtiene con un conjunto de componentes moleculares compartidos en gran parte. Los resultados obtenidos durante los últimos años no son suficientes para validar el papel de los ARNlnc como los «arquitectos» del genoma de esta compleja teoría evolutiva. Sin embargo, sí indican que los ARNlnc estudiados no son «basura». La mayoría de los genes que producen ARNlnc analizados responden a estímulos de una forma similar a los genes codificantes. Los ARNlnc y los ARNm comparten procesos de síntesis y maduración, aunque
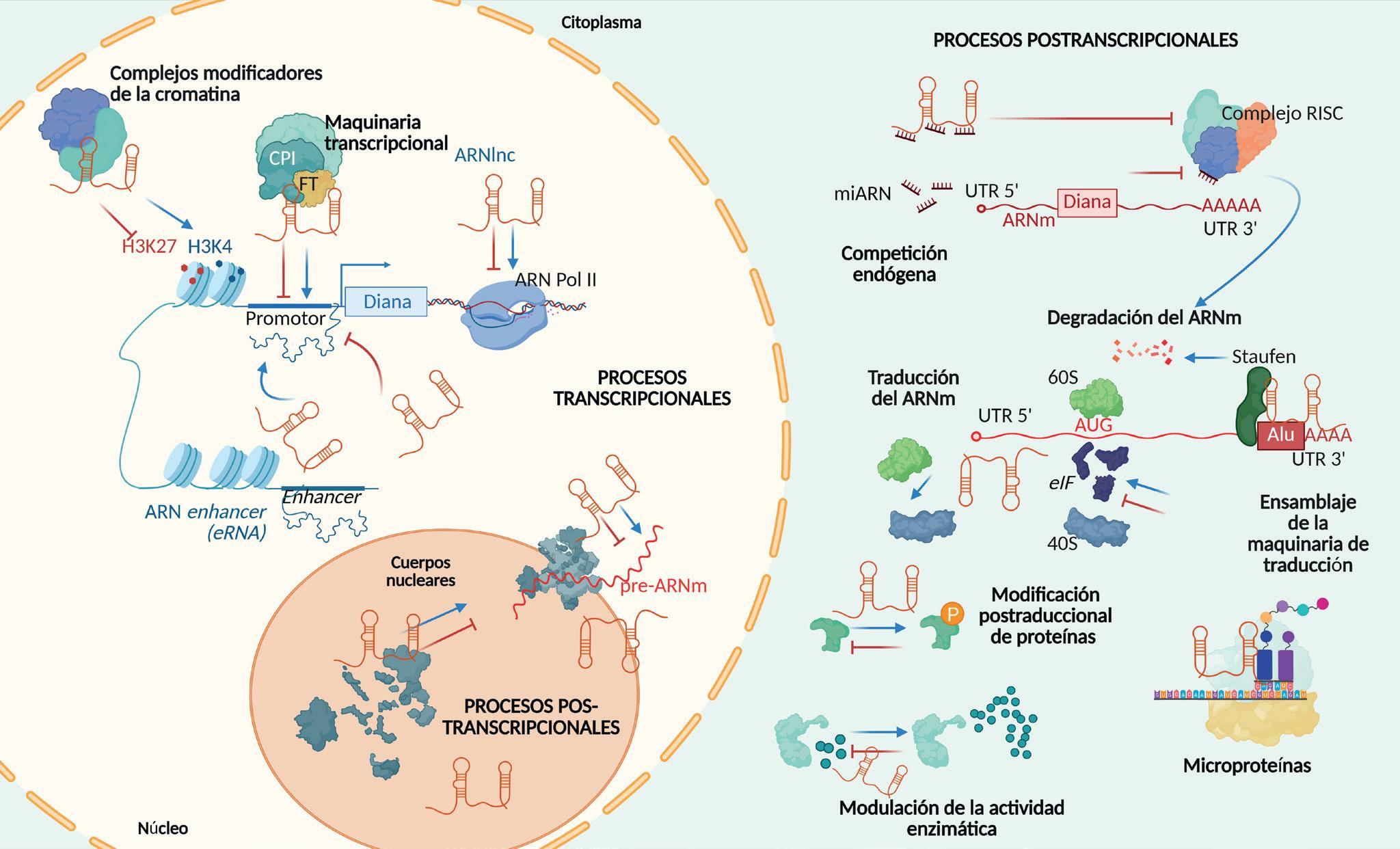
esta última parece que funciona un poco peor con los ARNlnc. Sin embargo, en general, comparados con los ARNm, los ARNlnc se localizan más preferentemente en el núcleo de la célula, son más específicos de tejido y de especie, y se producen a niveles mucho más bajos. Estas características están directamente relacionadas con la función de los ARNlnc, que en muchos casos sirven para regular la expresión de genes concretos. Ciertos ARNlnc actúan en cis, es decir, ejercen su función en el mismo sitio del genoma en el que se sintetizan, actuando sobre genes vecinos. Esta acción tan localizada explicaría una función con muy baja expresión. Otros funcionan en trans, como las proteínas, viajan por la célula hasta localizarse donde ejercen su función. Independientemente de actuar en cis o trans, los ARNlnc han de plegarse en una estructura determinada por su secuencia, como las proteínas, y unirse a ADN, proteínas o a otros ARNs para formar complejos activos. Se han descrito ARNlnc que contribuyen a modificar el genoma haciéndolo más o menos accesible a la maquinaria de transcripción, otros que modulan la transcripción de determinados genes directamente, o bien que alteran la maduración del ARNm, su transporte, su estabilidad o su traducción (Figura 2) En general, muchos de los ARNlnc estudiados son excelentes reguladores de la expresión génica. Como son específicos de tejido, contribuyen a que, aunque todas las células compartan el mismo ADN, esta información se utilice de forma específica en distintos tejidos.
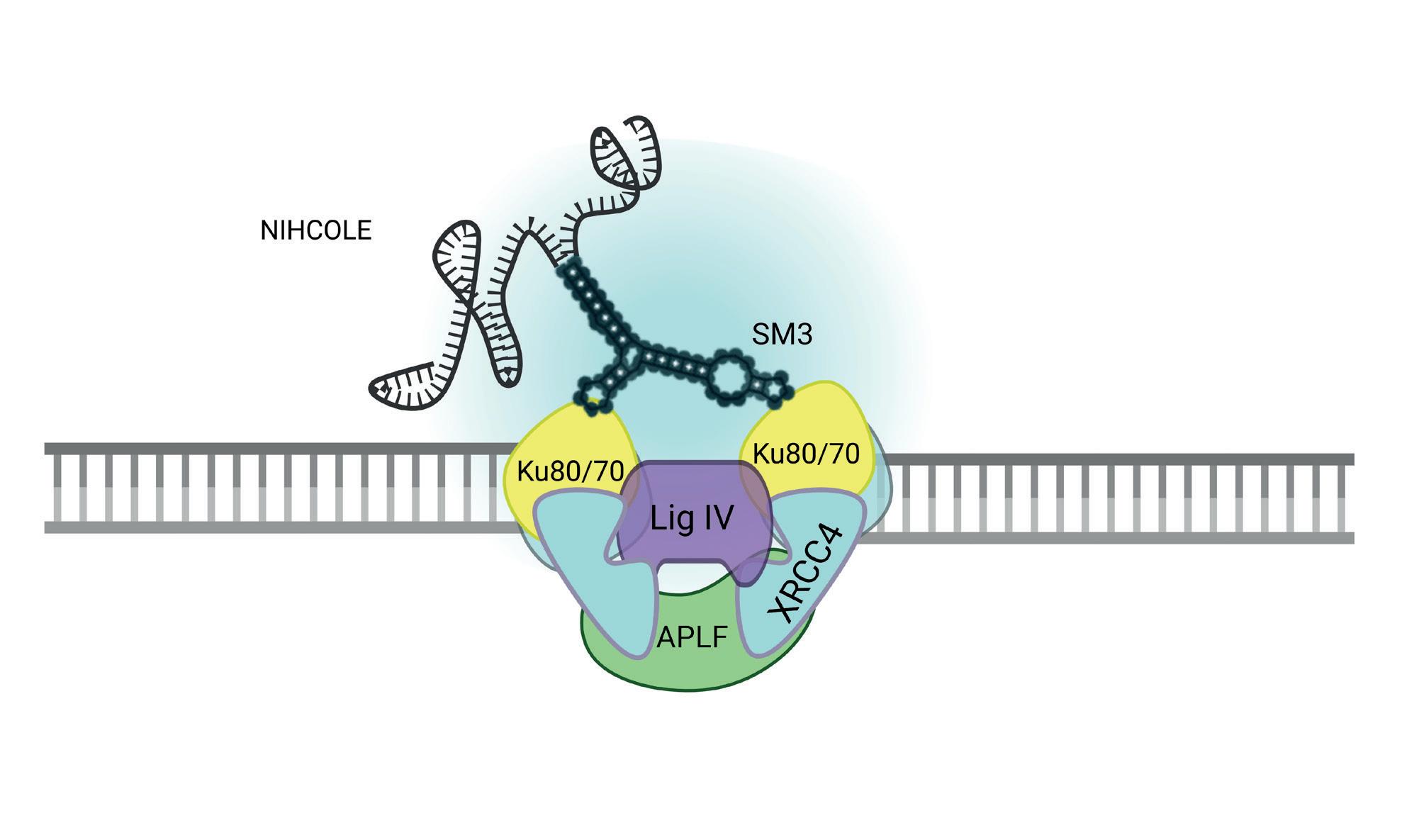
Además de funciones reguladoras, muchos ARNlnc se unen a maquinarias celulares para modular su función. Ese es el caso de NIHCOLE, que se une a la maquinaria de reparación del ADN y aumenta su eficacia (Figura 3)
Sorprendentemente, NIHCOLE no se detecta en células sanas, pero sí a niveles elevados en tumores de distinto origen: cáncer de mama, de pulmón, de colon y el cáncer de hígado más prevalente (hepatocarcinoma). En este último caso, donde NIHCOLE se ha estudiado en más detalle, los pacientes con peor pronóstico tienen niveles más elevados de NIHCOLE. Esto se debe, probablemente, a que el crecimiento tumoral y los tratamientos de radio y quimioterapia producen roturas en el ADN que se reparan más eficazmente cuando las células tumorales tienen NIHCOLE. Las células con NIHCOLE deberían seleccionarse y proliferar más, y eso las vuelve vulnerables. En el laboratorio hemos demostrado que bloquear la producción de NIHCOLE en células tumorales induce su muerte. Al igual que NIHCOLE, numerosos ARNlnc están implicados en el crecimiento tumoral o el desarrollo de otras enfermedades. Otra función ejercida por los ARNlnc es quizá la más inesperada. A pesar de haber sido bautizados como «no codificantes», algunos ARNlnc se traducen para producir proteínas no canónicas, generalmente pequeñas, y por ello llamadas microproteínas (miP). Al igual que los miARN hace un par de décadas, las miP han pasado desapercibidas hasta hace poco.
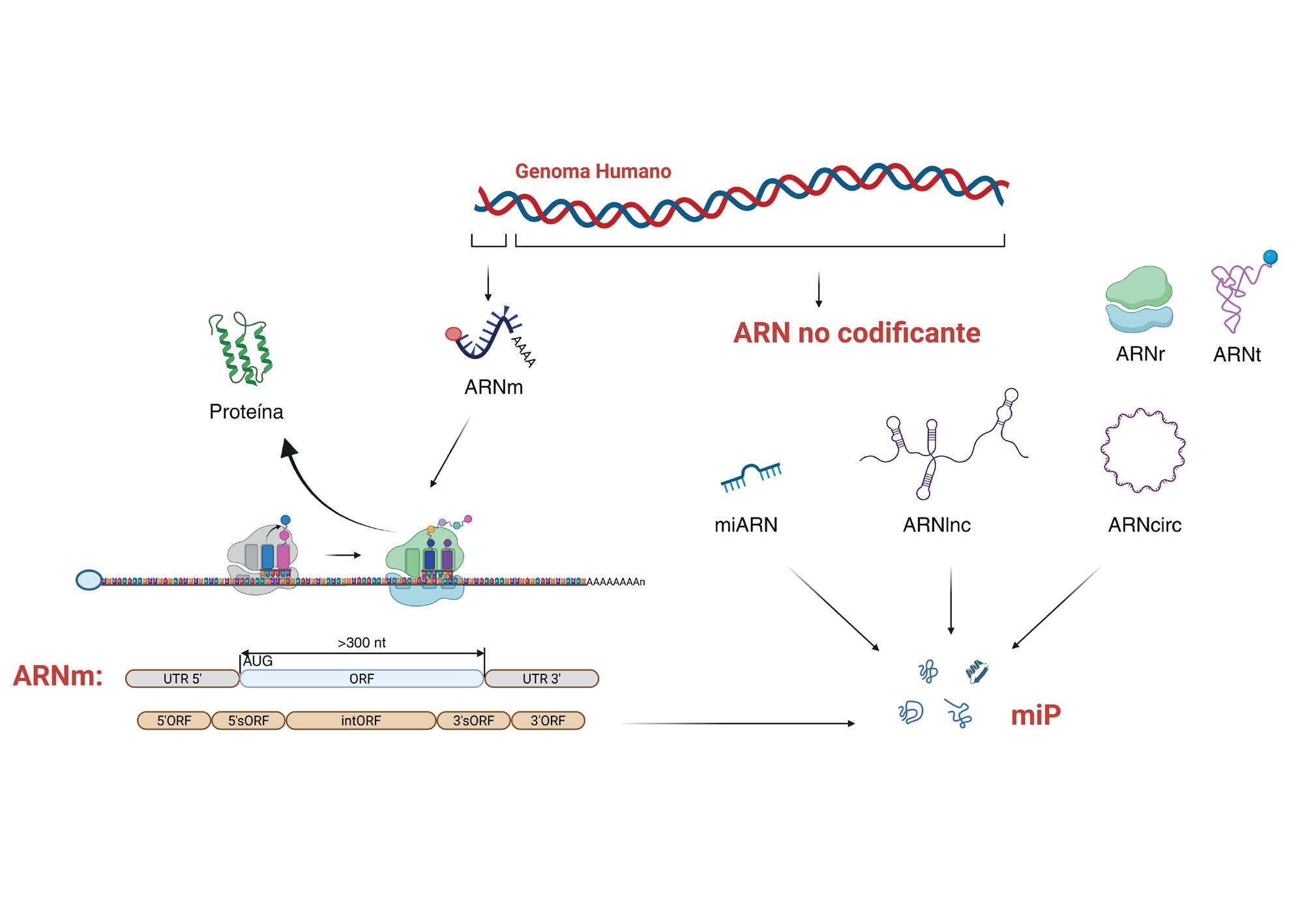
La dificultad para identificar estas miP reside en que la región de un ARN que se traduce, el marco de lectura abierto (ORF, por sus siglas en inglés open reading frame), es difícil de identificar, especialmente cuando es pequeño. Esto se debe a la complejidad de distinguir entre ORF verdaderos de los que no se traducen. Por ello, para diferenciar entre ORF genuinos y espurios, se estableció por consenso que las secuencias codificantes comienzan con los nt AUG (el codón iniciador), tienen más de 300 nt y están flanqueadas por
Figura 2
Funciones de los ARNlnc. A la izquierda se muestra un esquema del núcleo de la célula, donde los ARNlnc se esquematizan uniéndose a maquinarias implicadas en modificación de la cromatina, en transcripción y en procesos postranscripcionales. A la derecha se esquematiza el citoplasma, donde ciertos ARNlnc están implicados en varios procesos postranscripcionales relacionados con la estabilidad del ARNm (por ejemplo, compitiendo por miARN) y su traducción. Algunos ARNlnc pueden modular la modificación de proteínas o su actividad o pueden traducirse para producir microproteínas. Se indican procesos de activación (flechas azules) o inhibición (líneas rojas).
regiones no traducidas (UTR, por sus siglas en inglés untranslated region) 5’ y 3’ (Figura 4). Esto dibujó un gen modelo en el que cada ARNm producía una única proteína canónica. Sin embargo, esta visión ha sido desafiada por los datos obtenidos con el perfilado de ribosomas (RiboSeq), una tecnología novedosa que permite identificar fragmentos de ARN protegidos por los ribosomas durante la traducción. El RiboSeq ofrece una imagen precisa de toda la traducción celular con una resolución a nivel de nt. A la información de RiboSeq se suma la proteómica, que utiliza espectrometría de masas para identificar trozos de proteínas (péptidos) de extractos celulares o unidos
al complejo mayor de histocompatibilidad (CMH). Esta última aplicación, conocida como ligandoma, explota el hecho de que las proteínas celulares se fragmentan en péptidos (epítopos). Algunos de estos se unen al CMH, que los presenta al sistema inmune, controlando que la célula no exprese proteínas «extrañas» (como las virales). Si el sistema inmune detecta epítopos anómalos, desencadena la destrucción celular. Así, cualquier miP que se sintetice, incluso a bajos niveles, puede ser detectada en estudios de ligandoma si los epítopos resultantes se unen fuertemente al CMH. Los análisis de los datos de RiboSeq y proteómica han revelado una complejidad inesperada.
Algunas bases de datos de células humanas incluyen más de 500.000 miP, mientras que otras, con mayor nivel de confianza, describen más de 30.000. Las miP se han clasificado, según su ORF, en aquellas derivadas de ARN circulares (circORF, otra variante de la diversidad del ARN celular), largos o cortos no codificantes (lncORF y sncORF, por sus siglas en inglés) y aquellas traducidas desde la UTR 5’ o 3’ de ARNm (5´ORF y 3´ORF) o incluso desde las secuencias codificantes canónicas, solapando con la ORF de la proteína principal, pero en una fase de lectura diferente (5´ o 3´sORF o internas, intORF) (Figura 4). La mayoría de las miP son 5´ORF (47 %) y lncORF (21 %), seguidas de intORF (10 %). De
DOSIER CIENTÍFICO
hecho, se estima que alrededor del 75 % de todos los ARNm no solo producen una proteína canónica, sino también una o más miP.
Esto nos lleva a un nuevo paradigma donde cada ARN es capaz de generar múltiples proteínas.
Todo lo anterior modifica la imagen clásica de los genes codificantes. Primero, porque muchos ARNs codifican para más de una proteína, siendo los ARNlnc los más complejos (hasta 8 miP/ARNlnc en nuestros datos). Además, porque las secuencias codificantes pueden ser mucho más pequeñas de 300 nt, con miP de tan solo 3 nt (un solo aa).
Asimismo, a diferencia de las proteínas canónicas, cuyo ORF siempre empieza con el codón AUG
(traducido a metionina), el de las miP puede iniciar en AUG o en una mutación puntual de esta secuencia, resultando en codones que codifican para leucina, valina, treonina, isoleucina, arginina o lisina. Quizá las miP no comiencen con estos aminoácidos, sino que, en esta traducción no canónica, se permitan apareamientos incorrectos por los que se traduzca una metionina a partir de un codón que no codifica para ella. Nuestros resultados muestran que esta iniciación no requiere una secuencia lineal concreta (lo que se conoce como secuencia Kozak), sino que parece depender de estructuras secundarias de ARN alrededor del codón de inicio. Por lo tanto, las miP que
comienzan en codones distintos de AUG provienen de regiones estructuradas. Quizás, si el ribosoma se «entretiene» con estas estructuras, su menor velocidad le permite aceptar pequeños errores de inicio. Por último, comparado con las proteínas canónicas, las miP están enriquecidas en aminoácidos hidrófobos, lo que podría activar su degradación. Al igual que ocurrió con los ARNlnc, se ha sugerido que las miP podrían no ser relevantes. Esto se basa en que muchas podrían ser inestables, su tamaño pequeño dificulta que formen estructuras robustas y ordenadas, y no están conservadas. Sin embargo, el número de miP aumenta con la complejidad del desarrollo,
Figura 3
Mecanismo de acción de NIHCOLE. NIHCOLE tiene un dominio denominado SM3 que interacciona con las proteínas Ku (heterodímero de Ku80 y Ku70), que reconoce los extremos rotos del ADN. Este complejo se une a la maquinaria formada por APLF y XRCC4 unido a la ligasa IV, que une los extremos del ADN roto. En presencia de NIHCOLE, aumenta la sinapsis con la que se unen los dos extremos del ADN y se mejora la eficacia de ligación.
Figura 4
Esquema del genoma humano codificante y no codificante. La región del genoma humano catalogada clásicamente como codificante (que se transcribe a ARNm y da lugar a proteínas canónicas) es pequeña. La región no codificante es mayoritaria. A partir de ella se transcriben ARNr, ARNt, miARN, ARNcirc y ARNlnc. Sin embargo, algunos de los ARN considerados no codificantes tienen marcos de lectura abiertos (ORF, por sus siglas en inglés open reading frame) y pueden traducirse a proteínas generalmente pequeñas denominadas microproteínas (miP). Estos incluyen tanto los ARN no codificantes como las regiones no traducidas (UTR) de los ARNm. En este último caso se detectan proteínas no canónicas derivadas de UTR 5´ y 3´ (5´ y 3´ORF) o de regiones que comparten secuencia con el ORF de la proteína canónica, pero están en otra fase de lectura (5´ o 3´ sORF, cuando el ORF solapa con la zona 5´ o 3´ UTR y parcialmente con el ORF de la proteína canónica, o intORF cuando solapa completamente con éste).
siendo los humanos quienes poseen el mayor número, lo cual no es sorprendente ya que tenemos más ARNlnc capaces de producirlas (se estima que un 14-22 % de ARNlnc producen miP). Por ello, se ha sugerido que muchas de estas miP podrían ser «basura», y representar la producción de proteínas muy inestables y sin función, debido a que evolutivamente son demasiado recientes. Sin embargo, ya se han descrito unas 200 miP funcionales.
Curiosamente, las miP también pueden actuar en trans o en cis,
de manera independiente de la secuencia. La función de estas últimas es el mero acto de traducción: la traducción de algunos 3´ORF puede mejorar la expresión del ORF canónico cercano, mientras que la de ciertos 5´ORF puede disminuirla. La mayoría de las miP estudiadas funcionan en trans, se localizan en músculos (incluido el corazón), y están unidas a membranas o mitocondrias. Afectan a: (i) la función mitocondrial y el metabolismo celular; (ii) la expresión génica (procesamiento, estabilidad, modificación y traducción del
ARN, y ensamblaje y estabilidad de proteínas); (iii) la señalización celular, proliferación y diferenciación; (iv) las funciones relacionadas con el sistema inmunitario; (v) la reparación del ADN y el estrés celular. Estudios preliminares estiman que un 10-19 % de las miP son importantes para la supervivencia celular, y que la modulación de los niveles del 72 % de las miP produce cambios celulares detectables. Por último, las miP podrían ser clave en el desarrollo de terapias contra el cáncer. Muchos de los ARNlnc que se expresan en
DOSIER CIENTÍFICO
cáncer no se detectan en tejido sano. Entre ellos, se han identificado varios que se traducen a miP. Estas miP, específicas de cáncer, aun siendo no funcionales o inestables, pueden degradarse dentro de la célula y ser presentadas al sistema inmune a través de las moléculas del CMH. Si las identificamos, se podrían desarrollar vacunas contra el cáncer con las que activar a las células del sistema inmune para que reconozcan a estas miP y, tras ello, eliminen las células tumorales.
De hecho, estos factores no conservados, tanto ARNlnc como miP, de evolución tan reciente, podrían ser especialmente relevantes en la carcinogénesis y en el crecimiento tumoral. Las células cancerosas se enfrentan a un enorme estrés por la falta de oxígeno y nutrientes, vigilancia del sistema inmune, o por tratamientos como la quimio o la radioterapia, que buscan eliminarlas.
Para leer más
Para sobrevivir, solo tienen una opción: evolucionar y adaptarse usando su genoma. En células altamente proliferativas, las mutaciones, reorganizaciones genómicas y alteraciones epigenéticas pueden generar heterogeneidad celular y contribuir a la selección de células en división resistentes al estrés. Estos cambios pueden afectar la parte conservada del genoma, permitiendo la selección de células que han activado distintos oncogenes. Esto conlleva un riesgo, ya que se podrían dañar genes esenciales. Sin embargo, los cambios en la parte no conservada deberían ser más favorables, más numerosos (dado que alrededor del 95 % del genoma no está conservado) y, por lo tanto, generar una mayor diversidad de la cual se pueden seleccionar células resistentes. Nuestra hipótesis es que, en el estresante microambiente tumoral, la parte no conservada del genoma podría
generar una gran cantidad de ARNlnc, y estos, a su vez, diversas miP. Algunos de estos factores podrían ser capaces de regular la expresión de genes que alivian el estrés o que forman estructuras que mejoran los mecanismos para resolverlo y/o impulsar la progresión tumoral. Este es el caso del ARNlnc NIHCOLE. Las células que expresen estos factores, nuevos oncogenes, tenderían a ser seleccionadas por su ventaja adaptativa. Estos factores no conservados a menudo se han ignorado. Sin embargo, creemos que podrían ser dianas terapéuticas ideales, ya que las células tumorales deberían ser dependientes de su expresión. Los ARNlnc y las miP, a pesar de que en ciertos ámbitos se consideren «basura» genética, podrían estar mostrándonos el camino hacia nuevas y efectivas terapias contra el cáncer y otras enfermedades.
Baena-Angulo C, Platero AI, Couso JP. Cis to Trans: small ORF functions emerging through evolution. Trends in Genetics 41 (2025) 119-131. DOI: 10.1016/j.tig.2024.10.012
Camarena ME, Theunissen P, Ruiz M, Ruiz-Orera J, Calvo-Serra B, Castelo R, Castro C, Sarobe P, Fortes P, Perera-Bel J, Albà MM. Microproteins encoded by noncanonical ORFs are a major source of tumor-specific antigens in a liver cancer patient meta-cohort. Science Advances 10 (2024) eadn3628. DOI: 10.1126/sciadv.adn3628
Chen LL, Kim VN. Small and Long non-coding RNAs: Past, present, and future. Cell 187 (2024) 6451-85. DOI: 10.1016/j.cell.2024.10.024
Ruiz-Orera J, Villanueva-Cañas JL, Albà MM. Evolution of new proteins from translated sORFs in longnon-coding RNAs. Experimental Cell Research 391 (2020) 111940. DOI: 10.1016/j.yexcr.2020.111940
Statello L, Guo C-J, Chen LL, Huarte M. Gene regulation by long non-coding RNAs and its biological functions. Nature Review Molecular Cell Biology 22 (2021) 96-118. https://doi.org/10.1038/ s41580-020-00315-9
Unfried JP, Marín-Baquero M, Rivera-Calzada Á, Razquin N, […], Llorca O, Lees-Miller SP, Fortes P. Long Noncoding RNA NIHCOLE Promotes Ligation Efficiency of DNA Double-Strand Breaks in Hepatocellular Carcinoma. Cancer Research 81 (2021) 4910-4925. DOI: 10.1158/0008-5472.CAN-21-0463
LA EPITRANSCRIPTÓMICA, UNA CAPA REGULADORA
CODIFICADA QUÍMICAMENTE
Sandra Blanco
Centro de Investigación del Cáncer (CIC), Instituto de Biología Molecular y Celular del Cáncer, CSIC - Universidad de Salamanca
https://doi.org/10.18567/sebbmrev_226.202509.dc5
Durante décadas, el dogma central de la biología planteado por Francis Crick en 1958, nos ayudó a entender las reglas de la expresión génica: el ADN se transcribe a ARN, y éste se traduce a proteínas. Esta secuencia de procesos biológicos establecía una vía unidireccional y sencilla de cómo se descodifica el código genético. Sin embargo, tras años de investigación en biología molecular, ahora sabemos que la realidad es mucho más compleja, y aunque todavía desconocemos las reglas completas del código, sabemos que cada paso de este
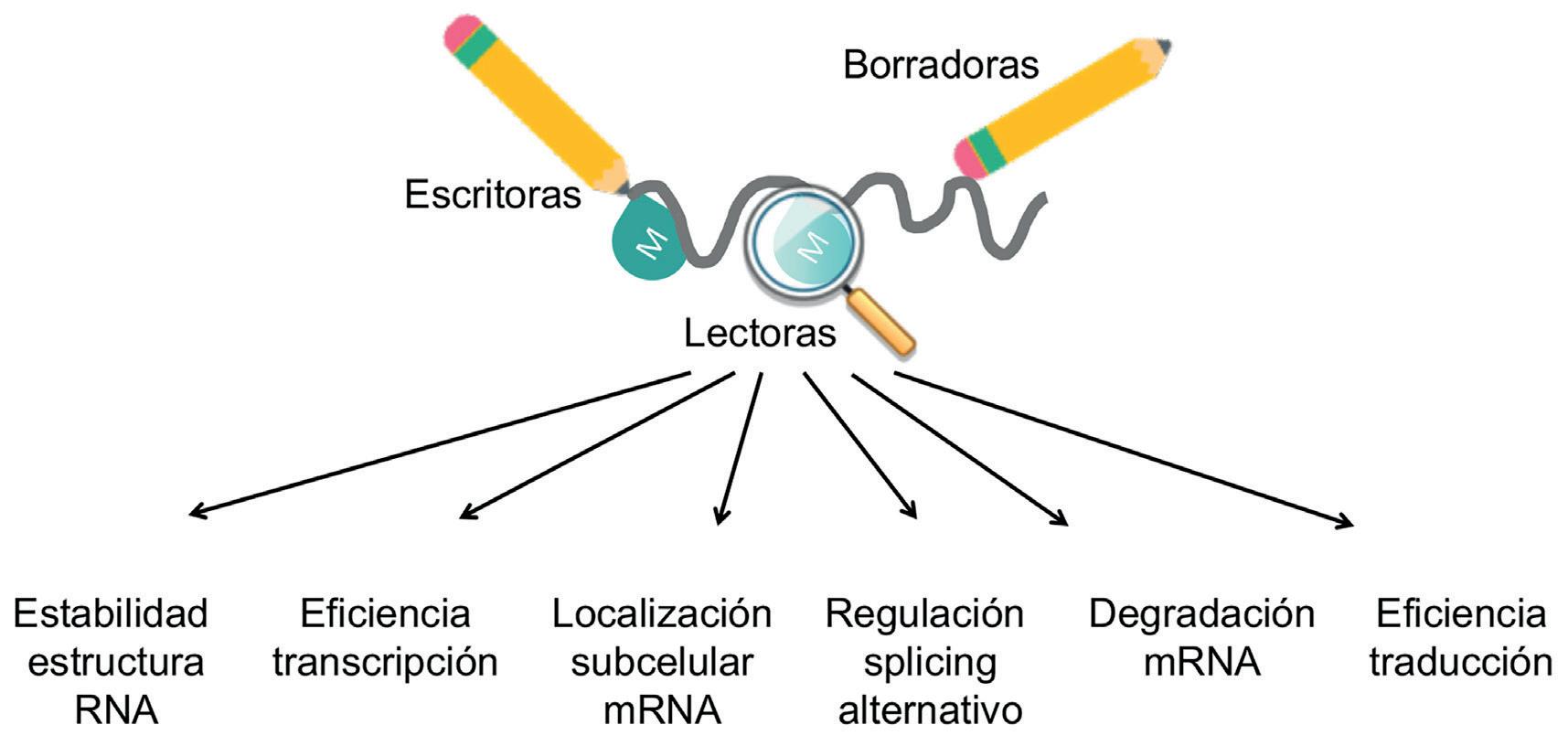
flujo de información está sujeto a varios niveles de regulación. Por ejemplo, el ADN y la expresión génica en general están regulados por modificaciones químicas que decoran el ADN, como la metilación del ADN, mecanismos que son estudiados por la epigenética. La función, estabilidad y localización subcelular de las proteínas también están reguladas por gran número de modificaciones postraduccionales, como la fosforilación o la metilación, entre otras muchas modificaciones químicas. Asimismo, el propio ARN sufre modificaciones químicas. En este
contexto ha emergido un nuevo campo de investigación revolucionario: la epitranscriptómica, que estudia las modificaciones químicas del ARN y cómo influyen en su función, expresión, estructura y localización subcelular, sin alterar su secuencia de nucleótidos.
Las modificaciones del ARN: más allá de la transcripción
El descubrimiento de las modificaciones del ARN se remonta a la década de 1950, cuando se detectaron nucleótidos modificados en ARN ribosomales (ARNr) y de transferencia (ARNt). No sorprendentemente, ya que estos tipos de ARN son los que albergan mayor número de modificaciones. Posteriormente estas modificaciones han sido detectadas en ARN mensajeros (ARNm) y en otros tipos de ARN no codificante, como microARNs (miARNs) y ARN largos no codificantes (ARNlnc), aunque en estos últimos las modificaciones son menos prevalentes. No obstante, no fue hasta 2011 cuando se comenzó a reconocer el impacto funcional de estas modificaciones gracias al avance tecnológico de las técnicas de secuenciación masiva y de biología molecular. Las investigaciones del Profesor Chuan He, en la Universidad de Chicago (EE.UU.), descubrieron que la actividad funcional del gen Fat mass and obesity-associated protein (FTO, por sus siglas en inglés), cuyas mutaciones y variaciones genéticas se habían asociado a varias enfermedades humanas neurológicas y metabólicas, consistía en desmetilar ARNs metilados en la posición N6 de la adenosina (m6A). Un año después, el Profesor Samie R. Jaffrey, de Weill Cornell Medicine, y el Profesor Gideon Rechavi, de la Escuela de Medicina Sackler de la Universidad de Tel Aviv, desarrollaron una técnica de secuenciación que permitió mapear de manera global, con resolución de
DOSIER CIENTÍFICO
nucleótido, la modificación m6A en ARNm de humanos y ratones. Estos descubrimientos revelaron que los ARNm humanos estaban metilados en adenina-6, y que esta modificación era reversible y, por tanto, regulada por proteínas específicas, cuya función era esencial para el funcionamiento del organismo y su alteración podría causar enfermedades. Este hallazgo marcó el nacimiento de la epitranscriptómica como disciplina, que fue acuñada como tal en 2015.
m6A no es la única modificación que decora el ARN. Existen más de 170 modificaciones químicas diferentes en el ARN. Muchas de ellas están muy conservadas entre diferentes organismos, pero otras son específicas de especie y del tipo de ARN. Aunque todavía no existen métodos que nos permiten mapear de forma precisa y con resolución a nivel de nucleótido todas ellas, ni conocemos la
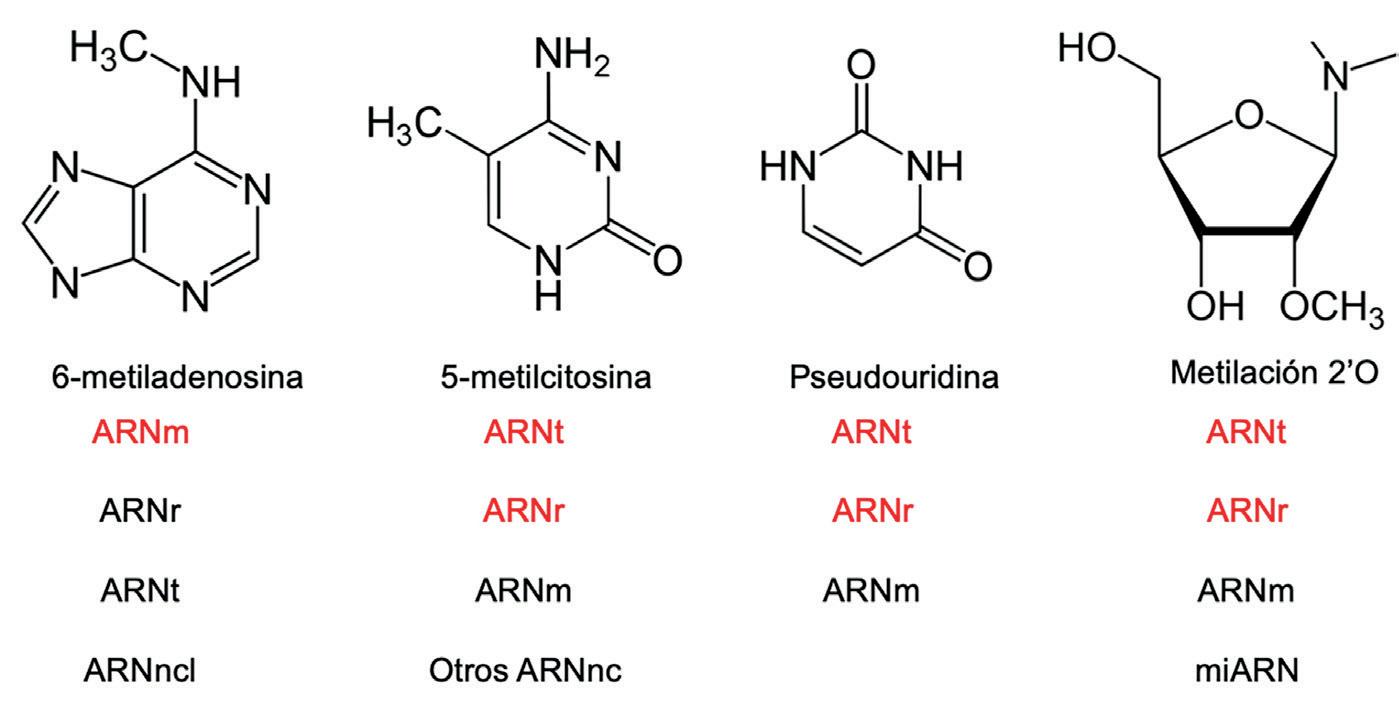
función biológica de la gran mayoría, en los últimos 14 años hemos avanzado en el conocimiento del impacto funcional y patológico de algunas de ellas. Algunas de las modificaciones más estudiadas son 6-metiladenosina, 5-metilcitosina (m5C), pseudouridina (Ψ) y metilación 2’O de la ribosa (Nm) (Figura 1)
• 6-metiladesina (m6A). Es la modificación más abundante en ARNm en mamíferos, aunque también se encuentra en ARNt y ARNr. Su formación está mediada por la metiltransferasa 3 (METTL3, por sus siglas en inglés), y es reversible mediante las demetilasas FTO y AlkB homolog 5 (ALKBH5). Su función ha sido principalmente estudiada en ARNm, en el que está vinculada a regulación de la estabilidad y la traducción de los ARNm modificados. Estas marcas no están distribuidas al azar, sino que aparecen en
contextos específicos: regiones no traducidas del ARNm (UTR, por sus siglas en inglés: región no traducida), exones codificantes, y en algunos casos, en la caperuza del extremo 5’. Además, pueden cambiar dinámicamente según el estado celular o fisiológico, lo que las convierte en una herramienta fina de regulación postranscripcional.
• 5-metilcitosina (m5C): se trata de la metilación de la citosina en posición 5. Se trata de una modificación muy abundante del transcriptoma de mamíferos, especialmente en ARNt y ARNr, aunque también se ha detectado en ARNm. En ARNt, m5C contribuye a mantener una conformación adecuada para el reconocimiento del codón por parte del anticodón, y también protege al ARNt frente a la degradación por endonucleasas. En los ARNr, esta modificación
1
Estructuras químicas de algunas modificaciones de ARN. ARNm: ARN mensajero; ARNt: ARN de transferencia; ARNr: ARN ribosómico; ARNlnc: ARN largo no codificante; ARNnc: ARN no codificante; miARN: microARN. El color rojo indica el tipo de ARN con modificaciones preferentes.
Figura
está implicada en la biogénesis ribosomal y puede influir en la precisión y eficiencia de la traducción. Su metilación en mamíferos está mediada por enzimas de la familia NOP2/ Sun domain family (NSUN1-7) y por DNA methyltransferase-2 (DNMT2). En los ARNm, esta modificación es dinámica y reversible, gracias a la acción de enzimas demetilasas de la familia ‘Ten-eleven translocation’ (TET, por sus siglas en inglés). Su función en ARNm se ha asociado con la actividad en el transporte del núcleo al citoplasma del ARNm, la estabilidad del transcrito y su eficiencia de traducción. Curiosamente, ciertos patrones de metilación en m5C también parecen estar regulados en respuesta a condiciones de estrés celular o procesos de diferenciación.
• Pseudouridina (Ψ): es una modificación muy abundante en el transcriptoma eucariótico y la primera que se descubrió en ARN. Es frecuente en ARNt y ARNr, y más recientemente también se ha detectado en ARNm, aunque en menor proporción. La pseudouridina es un isómero de la uridina, lo que confiere mayor rigidez estructural. Esta modificación no es reversible, y su incorporación está mediada por la pseudouridil sintasa Diskerina que, a su vez, es guiada por ARN cortos hacia su secuencia diana, o enzimas de la familia Pseudouridine synthase (PUS, por sus siglas en inglés) que no necesitan guía de ARN. En ARNt y ARNr, la pseudouridina desempeña funciones clave en la estabilidad estructural y la eficiencia de la traducción. Recientemente, esta modificación ha adquirido especial relevancia en el diseño de vacunas de ARN, como las vacunas frente a la COVID-19, donde la sustitución de uridinas
por pseudouridinas ha permitido mejorar la estabilidad del ARNm sintético y reducir la activación del sistema inmune innato, favoreciendo una traducción más eficiente y sostenida.
• Metilación 2’O de ribosa (Nm): esta modificación consiste en la metilación del oxígeno 2 de la ribosa de un nucleótido. Es una modificación frecuente en ARNr y ARNt, pero también se ha identificado en ARNm. En ARNm se localiza principalmente en la región 5’ UTR y en la caperuza del extremo 5’. Su catálisis está mediada por la metil-metilasa Fibrilarina, que está guiada a sus secuencias diana por ARNs cortos. Nm contribuye en la estabilización de la estructura tridimensional del ARN y su resistencia a la degradación por nucleasas, la eficiencia de traducción, la maduración del ARNm y la evasión del sistema inmune, especialmente en el contexto de infecciones virales o terapias con ARN sintéticos.
El nodo epitranscriptómico: enzimas escritoras, borradoras y proteínas lectoras
La regulación epitranscriptómica se basa en un trío funcional, compartido con la red epigenética:
• Enzimas escritoras: como el complejo metilasa de adenina-6 formado por METTL3 que es la unidad catalítica, y varios reguladores como Methyltransferase 14 (METTL14), WT1 Associated Protein (WTAP), Vir like m6A methyltransferase associated (VIRMA), RNA binding motif protein 15 (RBM15), zinc finger CCCH domain-containing protein 13 (ZC3H13).
• Borradoras: como las enzimas dedicadas a demetilar o eliminar el grupo metilo de las adeninas metiladas en posición 6 como FTO o ALKBH5.
• Lectoras: son proteínas que reconocen los nucleósidos
modificados como las proteínas de la familia YTH Domain Family (YTHDF1-3), que interpretan las modificaciones para ejecutar funciones celulares (Figura 2) Este sistema recuerda al que ocurre en la epigenética del ADN y demuestra que el ARN también tiene su propio «código» regulador, análogo a un código epigenético. No obstante, hay varias diferencias. Por ejemplo, algunas de las modificaciones de ARN son complejas y se deben a una reacción en cadena de varias enzimas escritoras, como por ejemplo mcm5s2U o 5-metoxicarbonilmetil-2-tiouridina, una modificación común en ARNt. Además, todavía no se han encontrado enzimas borradoras para la mayoría de las modificaciones de ARN, y se sospecha que algunas de ellas no son reversibles. Tampoco se han encontrado proteínas lectoras para la mayoría de las modificaciones de ARN.
Regulación epitranscriptómica con impacto a nivel celular
Las consecuencias funcionales de estas modificaciones son muy amplias (Figura 2). Pueden:
• aumentar o reducir la estabilidad de la estructura tridimensional del ARN,
• aumentar o reducir la estabilidad del ARN, determinando su vida media,
• modular la eficiencia de traducción, alterando la cantidad de proteína sintetizada,
• modular la eficiencia de transcripción, alterando la cantidad de ARNm transcrito,
• afectar la localización subcelular del ARN,
• regular el splicing alternativo, modificando qué isoformas de proteína se expresan. Estas funciones son relevantes para el funcionamiento normal de una célula, así como en el desarrollo embrionario, en procesos de respuesta celular al estrés o a
Figura 2
El sistema epitranscriptómico de enzimas escritoras, borradoras y proteínas lectoras, y sus implicaciones funcionales.
otros estímulos, en la diferenciación celular, en la proliferación y el crecimiento celular.
Impacto del epitranscriptoma en enfermedades
El papel del epitranscriptoma en el desarrollo de procesos patológicos es particularmente evidente en enfermedades neurológicas y cáncer. Por ejemplo, sabemos que alteraciones en la expresión o en la actividad catalítica de las proteínas escritoras, borradores y lectoras puede llevar a un desequilibrio en la cantidad de modificaciones encontradas en el ARN. Consecuentemente este desbalance, puede alterar procesos de estabilidad del ARNm, síntesis de proteína, splicing alternativo entre otros procesos en genes o transcritos específicos,
lo que puede favorecer o desfavorecer la proliferación celular descontrolada, la evasión a la muerte celular programada, la evasión del sistema inmune o la supervivencia y resistencia a terapias. Por ejemplo, en ciertos tipos de leucemia y otros tumores sólidos, una expresión incrementada de la metilasa METTL3, comparado con las células normales, promueve un aumento de la acción de esta metilasa, incrementando la metilación m6A específicamente en los trascritos que codifican para oncogenes. Este incremento de metilación en estos oncogenes induce un aumento de su traducción, dando lugar a una ventaja para las células tumorales. Estas observaciones han abierto nuevas posibilidades terapéuticas. En concreto, se están empezando a
desarrollar fármacos específicos que bloquean la acción de estas enzimas modificadoras del ARN que son más activas en las células tumorales.
El epitranscriptoma: un código postranscripcional de la regulación génica
Una de las grandes barreras en el avance de este campo ha sido las técnicas que se utilizan para poder identificar estas modificaciones en el ARN. Las modificaciones del ARN no cambian la secuencia, por lo que no son visibles en métodos tradicionales de secuenciación. No fue hasta mediados de la década de 2010, gracias a tecnologías tales como la meRIP-seq (donde se realiza la inmunoprecipitación de ARN modificados con anticuerpos que
reconocen la metilación m6A) y, más recientemente, la secuenciación directa por nanoporos, cuando ha sido posible identificar y mapear estas modificaciones a nivel de nucleótido.
Con estos avances tecnológicos se ha podido comprobar que los patrones epitranscriptómicos son diferentes entre tejidos, condiciones o tipos celulares. Los datos sugieren que el epitranscriptoma es dinámico y confiere plasticidad e identidad celular. Este nuevo paradigma del ARN, cuyo código epitranscriptómico supone una capa de información clave del código genético, se contrapone a su concepción inicial como simple mensajero del ADN, y ahora lo ha convertido en un actor principal en la regulación génica. La epitranscriptómica permite dar una respuesta celular rápida, plástica y reversible, sin que haya mutaciones o alteraciones en la arquitectura del ADN.
Para leer más
Aplicaciones biomédicas y biotecnológicas
Actualmente, la epitranscriptómica es una herramienta aplicada en biomedicina y en biotecnología. En biomedicina, sus aplicaciones se extienden desde el desarrollo de terapias que se centran en inhibir o bloquear las enzimas escritoras y borradoras implicadas en la aparición de cáncer, hasta el diseño de ARN terapéuticos modificados estables y eficaces. Ejemplos recientes de estas dos aplicaciones son la puesta a punto de inhibidores farmacológicos que ya están en ensayos clínicos para la enzima METTL3, o las vacunas de ARNm desarrolladas frente a la COVID-19 que incorporaron modificaciones como la pseudouridina para mejorar su estabilidad y reducir la respuesta inmune adversa. En el campo de la biotecnología, el control epitranscriptómico podría ser clave para la producción de proteínas recombinantes en bacterias, la
optimización de la tasa de traducción en determinadas células, o la reprogramación celular. En conclusión, la epitranscriptómica se encuentra aún en una fase relativamente joven, pero su crecimiento es vertiginoso. El descubrimiento de que la modificación m6A es abundante en ARNm en mamíferos y reversible solamente fue el inicio. Desde entonces, el desarrollo de tecnologías de secuenciación y edición de ARN en la última década han abierto una nueva frontera del conocimiento. Hoy, se considera que el ARN no solo porta información genética, sino también información reguladora codificada químicamente. Estamos ante una revolución comparable a la epigenética del ADN, pero con la ventaja de que los cambios en el ARN son más rápidos y reversibles, lo que los convierte en herramientas ideales para terapias temporales, personalizadas y de alta precisión.
Blanco S, Bandiera R, Popis M, Hussain S, Lombard P, Aleksic J, Sajini A, Tanna H, Cortes-Garrido R, Gkatza N, Dietmann S, Frye M. Stem cell function and stress response are controlled by protein synthesis. Nature 534 (2016) 335-340. https://doi.org/10.1038/nature18282
Dominissini D, Moshitch-Moshkovitz S, Schwartz S, Salmon-Divon M, Ungar L, Osenberg S, Cesarkas K, Jacob-Hirsch J, Amariglio N, Kupiec M, Sorek R, Rechavi G. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 485 (2012) 201-206. https://doi.org/10.1038/nature11112
Jia G, Fu Y, Zhao X, Dai Q, Zheng G, Yang Y, Yi C, Lindahl T, Pan T, Yang YG, He C. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nature Chemical Biology 7 (2011) 885887. DOI: 10.1038/nchembio.687
Meyer KD, Saletore Y, Zumbo P, Elemento O, Mason CE, Jaffrey SR. Comprehensive analysis of mRNA methylation reveals enrichment in 3’ UTRs and near stop codons. Cell 149 (2012) 1635-1646. DOI: 10.1016/j.cell.2012.05.003
Nombela P, Miguel-Lopez B, Blanco S. The role of m(6)A, m(5)C and Psi RNA modifications in cancer: Novel therapeutic opportunities. Molecular Cancer 20 (2021) 18. https://doi.org/10.1186/s12943-020-01263-w
Director de la AEI
JOSÉ MANUEL FERNÁNDEZ DE LABASTIDA
SEBBM/IP
« Necesitamos nuevos presupuestos para garantizar políticas científicas estables
Catedrático de Física Teórica, hasta su nombramiento como director de la Agencia Estatal de Investigación
(AEI) Fernández de Labastida desempeñaba el cargo de director de la Unidad de Apoyo al Consejo Científico del Consejo Europeo de Investigación (ERC, por sus siglas en inglés). De 2011 a 2021 dirigió el Departamento
Científico en la misma organización. Su trayectoria profesional incluye responsabilidades de gestión en la Administración General del Estado, donde ocupó el cargo de secretario general de Política Científica y Tecnológica, director general de Investigación y Gestión del Plan Nacional de I+D+i y subdirector General de Proyectos de Investigación. También fue vicepresidente de Investigación Científica y Técnica en el CSIC. Además, ha sido autor y coautor de más de 70 publicaciones científicas, ha liderado cinco proyectos de investigación nacionales y europeos, y ha dirigido cinco tesis doctorales.
El secretario de Estado de Ciencia, Innovación y Universidades, Juan Cruz Cigudosa, afirmó durante su toma de posesión que su etapa abrirá un nuevo ciclo de crecimiento para la AEI. ¿Cuál es su visión para fortalecer la Agencia, hacerla más ágil, accesible y útil para la comunidad científica?
En mi toma de posesión expuse lo que será la visión de la AEI y sus líneas estratégicas para los próximos tres años, que es el mandato que tengo. Mi objetivo es consolidar su posición como referente nacional -que ya lo es-, pero también internacional, donde sigue siendo poco conocida. He trabajado 14 años en el Consejo Europeo de Investigación (ERC) y, cuando se hizo público que iba a ser director de la AEI, pregunté a colegas internacionales si la conocían. La mayoría no sabía de su existencia, y eso se debe en gran parte a su juventud institucional.
Las agencias son organismos que median entre la comunidad científica y las políticas públicas. Para tener éxito, deben contar, por un lado, con el respaldo de la comunidad científica, que debe percibirlas como un servicio de calidad, y por otro, con la confianza de quienes hacen política, que deben verlas como instituciones fiables, con capacidad para actuar de manera independiente y eficiente. Tenemos independencia operacional.
A partir de 2028 se debatirá en España el próximo Plan Estatal de Investigación, que define las políticas a seguir, y ahí podremos influir.
Una de las ventajas de las agencias es ese contacto directo con la comunidad científica, lo que nos permite proporcionar mucha información relevante a quienes diseñan las políticas. En Europa, al delegar
El nuevo director de la AEI propone implementar el contrato de gestión para realizar una planificación anual real
parte de la implementación de sus programas a las agencias, se dieron cuenta de que no recibían suficiente información sobre lo que realmente estaba ocurriendo en el sistema científico. Ese papel de puente es una tarea pendiente también en España. Durante mi etapa en el ERC trabajé intensamente en esto, y en la AEI es un área que aún no está plenamente desarrollada. Creo que ahí podemos aportar mucho valor. Espero que esta actividad, la llamada «feedback to policy» en términos anglosajones, sea decisiva. En esta línea colaboramos con la Oficina Nacional de Asesoramiento Científico (ONAC).
Para fortalecer la AEI es fundamental mejorar su gestión, y para ello necesitamos un contrato de gestión. Las agencias, recordemos, son estructuras relativamente recientes: se crearon en 2006, pero en 2015 el Gobierno decidió suprimirlas, aunque después se recuperaron. Nunca han terminado de estar plenamente asentadas. Ese contrato de gestión es un acuerdo plurianual por el que el Gobierno se compromete a garantizar recursos estables, mientras que la agencia fija objetivos medibles, con evaluaciones periódicas sobre su cumplimiento. Esto proporciona autonomía operativa y estabilidad a medio plazo.
Contar con ese contrato nos permitiría hacer una planificación plurianual real. Mi sueño es que en el verano del año anterior podamos anunciar todas las convocatorias y actividades del año siguiente. Esto en Europa es relativamente sencillo: de hecho, una de mis últimas responsabilidades en el ERC fue coordinar el programa de trabajo anual, que se ajustaba a un calendario como el mencionado.
En su toma de posesión habló de tres aspectos casi innegociables para el desarrollo de su estrategia al frente de la AEI: calidad, transparencia y simplificación. ¿Podría desarrollar más esta triada?
Efectivamente. Además del contrato de gestión, me gustaría destacar estos tres conceptos. En primer lugar, la calidad. Es fundamental someter los procesos a un escrutinio constante, tanto por parte de la propia agencia como de los beneficiarios. Necesitamos conocer qué funciona y qué no, tanto en la evaluación científica como en la gestión admi-
nistrativa. Actualmente somos unas 250 personas trabajando en la agencia, además de unos 250 investigadores e investigadoras a tiempo parcial que colaboran en los procesos de evaluación y nos proporcionan información clave para mejorar. En segundo lugar, la transparencia. Es imprescindible que la comunidad científica comprenda con claridad cómo funcionan los procesos y cuáles son los criterios de evaluación. Debemos ser lo más transparentes posible en nuestras actuaciones, lo que implica compartir información de forma proactiva. Y, por último, la simplificación, que tiene dos vertientes. Una es la simplificación administrativa, iniciada por mi predecesor, que se empezará a notar con más intensidad próximamente, sobre todo en lo relativo al seguimiento económico de las ayudas. Estas medidas afectan principalmente a las últimas convocatorias, pero aún no están en fase de seguimiento. La otra vertiente es normativa. Queremos avanzar en la simplificación revisando la orden de bases, orientándola hacia modelos más sencillos. Ya lo dije en mi toma de posesión: en la Comisión Europea soplan vientos muy fuertes a favor de la simplificación. La CE está implantando un instrumento financiero llamado lump sums (financiación a tanto alzado), que funciona así: se concede la ayuda y, una vez otorgada, solo se exige justificar los resultados científicos y técnicos, pero no los detalles de cómo se ha gastado el dinero. Sin embargo, esto es todavía un objetivo lejano. La Ley General de Subvenciones en España no permite aplicar ese modelo. Creo que deberíamos abrir una reflexión -no desde la agencia, sino desde el propio legislador- sobre la posibilidad de alinearnos con el enfoque de la Comisión Europea.
¿Está previsto que la IA, por su relevancia estratégica, se convierta en una nueva área temática dentro de las 19 existentes?
¿Cómo aborda la AEI los desafíos éticos, regulatorios y sociales de esta tecnología, en línea con las directrices de la UE?
Mi tarea es impulsar herramientas de gestión que ayuden tanto a nuestros beneficiarios como a los propios gestores a ser más eficientes, y entre ellas está la inteligencia artificial (IA). Es una corriente relativamente nueva que empieza a consolidarse. Además, este tema está ahora mismo en el foco de todas las agencias financiadoras del mundo. Hace poco estuve en la reunión anual del Global Research Council, donde se reúnen todas las agencias de financiación del mundo, y el tema estrella fue precisamente éste: cómo integrar la IA en la gestión de la ciencia. Durante mi etapa en el Consejo Europeo de Investigación (ERC) ya implantamos herramientas basadas en IA para la identificación de evaluadores. Una buena implantación de estas herramientas permite acelerar los procesos, ofrecer más soporte a los evaluadores y
gestionar mejor el volumen creciente de propuestas. Respecto a si vamos a incluir la IA como área temática específica… Bueno, está y no está. La IA es transversal, afecta a muchísimas disciplinas. Ahora mismo en la AEI tenemos 19 áreas y 55 subáreas, una estructura de detalle similar al de otras agencias europeas. El ERC, por ejemplo, trabaja con 28 subáreas, algo más amplias, y tampoco tiene ninguna específicamente llamada «Inteligencia Artificial». Es cierto que en el ERC abrimos ese debate, y aquí lo tendremos también. Será una cuestión para abordar con nuestro Comité Científico y Técnico, que es quien nos asesora sobre la organización de las áreas de conocimiento. Está encima de la mesa y hay que discutirlo. De hecho, acabamos de lanzar una convocatoria específica sobre IA, a petición de la Secretaría de Estado de Ciencia, Innovación y Universidades. El objetivo es ver cómo la comunidad científica española está desarrollando proyectos en diferentes campos -desde ciencias naturales hasta biomedicina- aplicando IA. Y lo cierto es que ha sido un éxito: hemos duplicado el número de solicitudes respecto a la convocatoria anterior, con propuestas muy potentes. Ahora estamos justo iniciando el proceso de evaluación.
Se lo comento, claro está, porque la IA podría romper la burocratización del sistema español de I+D+i del que se queja el cuerpo investigador… He visto dos burocracias: la europea y la española. Tengo esa experiencia. Por un lado, tengo que decir que me fui a la Comisión Europea a principios de
2011 y he vuelto ahora, en 2025. Y lo primero que me ha impactado al regresar ha sido el nivel de digitalización que tiene ahora nuestra Administración General del Estado. No tiene nada que envidiarle a la Comisión Europea, al contrario, diría que estamos más avanzados en algunos aspectos, y por supuesto por delante de otros países de nuestro entorno. Hemos avanzado muchísimo en digitalización de la gestión, y me he quedado gratamente sorprendido.
Cuando me fui todavía hablábamos del portafirmas en papel. Hay burocracia, claro, tanto aquí como en la Comisión Europea, pero diría que allí es incluso más pesada. Los flujos para aprobar determinadas cosas son eternos, aquí son más simples.
Y en lo que se refiere a nuestras ayudas, los tiempos están bastante acompasados con los de la Comisión Europea y con los de otras muchas agencias. Por ejemplo, en proyectos de investigación, gestionamos unas 7.000 solicitudes por convocatoria, y conseguimos resolver -al menos de forma provisional, que es lo que se puede comparar con la CE, el momento en que los investigadores reciben noticias por primera vez- en unos cinco o seis meses. En el ERC es un año. El año no te lo quita nadie. Así que, comparado, diría que nuestros tiempos son muy razonables.
También estamos avanzando en el seguimiento de las ayudas, especialmente en el ámbito del control económico, con el ánimo de reducir cargas innecesarias sobre los beneficiarios, manteniendo la debida garantía de control de la actividad subvencionada. Desde hace unos años, la agencia cuenta con un
plan anual de actuación para la comprobación económica de las actividades subvencionadas mucho más ágil ya que no se hace sobre el 100 % de las ayudas, sino por muestreo, de forma aleatoria, aunque con ciertos factores de ponderación: por ejemplo, se otorga mayor peso a las ayudas de mayor cuantía. Este enfoque busca asegurar una revisión representativa y eficaz.
En comparación, en el ERC tienes que presentar tres informes económicos. Es bastante más complejo que lo que pedimos nosotros. ¿Que nuestra burocracia es mejorable? Sin duda. ¿Qué tenemos que simplificar? También. Pero si te comparas con países como Alemania, diría que estamos en un grado bastante avanzado de simplificación. El sueño sería llegar algún día a una financiación a «tanto alzado», pero eso no depende de mí, sino de una modificación de la Ley General de Subvenciones. Mientras tanto, todo lo que podamos simplificar lo vamos a abordar, y además queremos establecer flujos de información para generar una cultura de mejora continua. Que los beneficiarios nos cuenten qué dificultades se encuentran, qué cosas cambiarían… Esa retroalimentación de la que hablaba antes es fundamental para seguir mejorando.
Hablamos de vocaciones STEM y del papel de la mujer en la ciencia. Usted se refirió a la igualdad como uno de los pilares de su gestión. ¿Podría concretar algún aspecto? Sí. España es uno de los países que mejor está en este contexto a nivel europeo. ¿Qué quiero decir con eso? Que la participación de mujeres liderando
proyectos es de las más altas de Europa. Por darte un dato: en la última convocatoria de Proyectos de Generación de Conocimiento, el 37 % de las ayudas financiadas tiene como investigadora principal a una mujer. Si lo comparamos, por ejemplo, con el ERC, allí estamos en torno al 30 %, o sea que vamos unos siete puntos por encima.
Pero más allá de la foto fija, lo importante es ver cómo evoluciona ese porcentaje a lo largo del tiempo. El ERC mejora año a año, pero no a la velocidad que sería deseable. En el caso de la Agencia, he pedido los datos completos, pero mi impresión -y creo que se confirmará- es que la tendencia es de mejora constante. ¿Por qué? Porque es justo en las franjas de edad más jóvenes donde hay más mujeres liderando proyectos, y eso apunta a un relevo generacional que hará crecer esos porcentajes.
Esto va a ocurrir. Llegaremos a la equidad, pero es una cuestión de tiempo. Y eso también es fruto de las políticas que se han impulsado en España en los últimos 20 años para favorecer el papel de la mujer en la ciencia. De hecho, en mi toma de posesión insistí en que no basta con medir el número de proyectos o publicaciones; necesitamos medidas concretas que impulsen esa participación.
La Agencia acaba de aprobar su II Plan de Igualdad precisamente con ese objetivo: monitorizar la participación de mujeres, analizar la ratio de éxito, estudiar los datos con rigor y, sobre todo, generar medidas que faciliten su liderazgo en los proyectos. Esto es un compromiso de gestión, no solo una declaración.
Y, por último, la sostenibilidad. ¿Qué papel ocupa en su estrategia?
Efectivamente, la sostenibilidad es otro de los pilares de mis líneas estratégicas. Y me refiero no solo a que la propia Agencia sea más sostenible en sus procesos internos -que lo debe ser-, sino también a incorporar ese enfoque en la propia actividad investigadora. Ya hay grupos de trabajo en Europa, vinculados a Science Europe, que están avanzando precisamente en esa línea, y nuestra intención es alinearnos con esas directrices. Se trata de que los investigadores, cuando desarrollen sus proyectos, tengan en cuenta también el factor de sostenibilidad: que piensen en cómo minimizar el impacto ambiental de su propia actividad científica.
No hablamos solo de resultados o de publicaciones, sino de que el propio proceso de investigación sea más respetuoso con el entorno. Ese es el camino en el que queremos avanzar.
Otro de los objetivos que se ha marcado es la internacionalización de la Agencia. Ha mencionado el escaso grado de conocimiento que tiene en el exterior. ¿Cómo piensa mejorar esa percepción?
Nuestro objetivo es claro: vamos a alinearnos con Europa, tanto en el Espacio Europeo de Investigación como con las agencias homólogas a través de Science Europe. Queremos jugar un papel más relevante, participar activamente en los foros internacionales y, sobre todo, ser muy rápidos en adaptar las iniciativas que surgen allí a nuestra forma de trabajar.
Se trata de visibilizar lo que hacemos, de que se nos reconozca como un actor clave dentro del sistema europeo de financiación de la ciencia. Vamos a estar donde se toman las decisiones y vamos a traer esas dinámicas a nuestro día a día.
Pongamos el ejemplo de CoARA, la coalición internacional de organizaciones científicas que trabajan juntas para permitir una reforma sistémica de la evaluación de la investigación. ¿Qué grado de desarrollo tiene actualmente el modelo de evaluación propuesto por CoARA en el plan de evaluación de proyectos de la AEI?
No formamos parte de la coalición CoARA, al menos de momento, no sé si lo haremos en el futuro. Pero sí te puedo decir que los principios de CoARA ya los hemos ido integrando en nuestros procesos. Me refiero a lo que se conoce como evaluación responsable, que no se basa solo en métricas o números, sino que incorpora un enfoque más cualitativo. Eso en la Agencia lo aplicamos desde hace un par de años. Por ejemplo, el currículum vitae abreviado que utilizamos está muy alineado con esa filosofía. Lo que pedimos es que los investigadores expliquen el impacto real de sus publicaciones, qué aportan, por qué son relevantes. No basta con el número de artículos o las
citas: queremos saber qué significado tiene su trabajo.
¿Las métricas?
Fernández
de Labastida
subraya que para fortalecer la AEI es fundamental mejorar su gestión
Claro que tienen su valor, pero por sí solas no sirven para evaluar individualmente a una persona. Son útiles para una visión global del sistema, pero para valorar una carrera científica necesitamos contexto, impacto real y aportaciones específicas. Y esa es la tendencia en Europa. Lo hemos visto también en la reciente reunión del Global Research Council ¿Supone más trabajo para la evaluación? Sí. Pero también es una evaluación más justa, más rigurosa. Y seguiremos alineados con los grupos internacionales que están desarrollando estas buenas prácticas para integrarlas en nuestros procedimientos.
En 2025, la AEI cuenta con un presupuesto de 1.000 millones de euros. Entre 2018 y 2024, el Gobierno duplicó la inversión científica con fondos europeos y aumentó un 40 % los fondos estatales. ¿Qué balance hace del ecosistema investigador español, considerando logros como el 40 % de fondos de recuperación europeos destinados a I+D y retos como la inversión del 1,4 % del PIB en I+D y la contribución empresarial del 0,7 % del PIB? Con los presupuestos nacionales prorrogados, las nuevas directrices de la UE para aumentar el gasto en defensa y la difícil situación geopolítica, ¿la inversión científica se va a ver afectada? Presupuestariamente seguimos por debajo de la media europea, tanto en inversión pública como privada. Alcanzar el 2 % del PIB en I+D, que es el objetivo del Gobierno, todavía requiere esfuerzo, no solo público, también privado. Se han hecho avances importantes, sobre todo gracias a los fondos europeos de recuperación y resiliencia. España es, de hecho, el segundo país de Europa que más porcentaje de estos fondos ha destinado a I+D, en torno al 40 %. Eso ha supuesto un impulso claro. El reto ahora es consolidar esa financiación: que no sea un esfuerzo coyuntural, sino estructural.
En los años en los que se aprobaron presupuestos generales vimos incrementos sostenidos en la financiación pública a la ciencia. Ahora estamos funcionando con presupuestos prorrogados, lo que implica trabajar con las cuentas del año anterior. Afortunadamente, esos incrementos previos nos permiten mantener una financiación razonable, pero 2026 va a ser un reto. Necesitamos nuevos presupuestos para garantizar políticas estables, sobre todo para con-
vocatorias estratégicas, como las dirigidas a atraer talento o consolidar investigadores.
España ha mejorado claramente respecto a hace unos años -yo lo he seguido desde Bruselas-, pero el margen de mejora sigue siendo importante. Y además del aspecto presupuestario, hay otro reto igual o más relevante: necesitamos seguir avanzando en la definición de una carrera investigadora clara y atractiva, mejorar las retribuciones y estabilizar las trayectorias. No es solo un problema de recursos, es también un problema de estructura. Para parecernos a los países de nuestro entorno europeo necesitamos resolver esas cuestiones.
El contexto europeo y geopolítico complica la situación, es evidente. Las nuevas directrices sobre inversión en defensa van a tensionar los presupuestos generales. Pero si queremos un país competitivo y sostenible, invertir en ciencia e innovación no es opcional, es estratégico.
El modelo de financiación de la AEI se basa en convocatorias competitivas. ¿Considera que este sistema satisface las necesidades del ecosistema investigador?
La investigación siempre es un equilibrio entre competir y cooperar. La clave está en encontrar el balance adecuado. Competir es necesario, especialmente en contextos amplios, porque cuando se ponen a competir las mejores ideas, obtenemos los mejores resultados. Creo que la competencia por ideas y proyectos de investigación es positiva y debe mantenerse, pero al mismo tiempo hay que fomentar más la cooperación.
Un porcentaje muy alto de los proyectos que financiamos se resuelve mediante competencia, y creo que ese modelo funciona. Pero hay margen para abrir más a la cooperación, tanto nacional como internacional. España participa activamente en muchas iniciativas internacionales, pero, sinceramente, creo que aún podemos mejorar. Actualmente, la parte del presupuesto de la Agencia destinada a cooperación internacional no llega al 5 %, y uno de mis objetivos es aumentarlo. He tenido ya contactos bilaterales con otras agencias europeas y puedo afirmar que España es un país atractivo para cooperar, y lo es cada vez más.
Además, somos buenos en proyectos de cooperación. De hecho, creo que se nos dan mejor los proyectos colaborativos que los individuales. Lo vemos en el caso del ERC: al analizar la participación de investigadores e investigadoras españoles, destaca nuestro buen papel en los Synergy Grants, que son proyectos colaborativos, más que en los individuales. Y no solo participamos, también lideramos. Tenemos potencial y experiencia, y debemos seguir reforzando ese camino.
¿Qué mecanismos de evaluación ex post implementará la AEI para vincular la financiación a resultados medibles, como publicaciones, patentes, transferencia tecnológica o impacto económico y social, en línea con su mandato de transparencia?
Hay varios aspectos importantes. Por un lado, el Plan Estatal de Investigación Científica, Técnica y de Innovación cuenta con un programa propio de evaluación de convocatorias y programas. Este proceso lo coordina el Ministerio de Ciencia, Innovación y Universidades, a través de la Secretaría de Estado de Ciencia, y la Agencia colabora y contribuye activamente, aunque no lo lidera directamente.
Además, uno de mis objetivos es avanzar en la definición de indicadores concretos para medir el impacto de la investigación financiada desde la Agencia. No me refiero solo al impacto en publicaciones o al posicionamiento en el 1 % más citado, sino a un conjunto más amplio y preciso de indicadores para seguir el impacto real de nuestras actuaciones. Este trabajo está en marcha y lo haremos coordinados con el Ministerio. Para mí es fundamental publicar de manera accesible los datos sobre la financiación que concedemos. No solo por transparencia, sino porque muchos investigadores en políticas científicas pueden usar esa información para realizar estudios y de esta manera contribuir a la mejora del sistema.
De hecho, ya gestioné esta responsabilidad en el ERC, donde desarrollamos herramientas para ofrecer información sobre la ciencia financiada. Ahora quiero impulsar una iniciativa similar desde la Agencia, siempre en coordinación con la Secretaría de Estado, que tiene el mandato de seguimiento general del Plan.
Regiones como Madrid y Cataluña concentran la mayoría de los fondos de la AEI. ¿Qué medidas adopta la AEI para promover una distribución más equitativa e impulsar la capacidad científica en comunidades como Extremadura o Castilla-La Mancha?
Respecto a la desigualdad territorial, es cierto que existe, pero hay que recordar que en España las políticas de I+D+i son competencia compartida entre Comunidades autónomas y la Administración General del Estado. Aquí debe haber un esfuerzo conjunto. Aquellas comunidades que más se han esforzado, en recursos y planificación, han obtenido más éxitos a nivel nacional y europeo. La distribución regional de las ayudas nacionales es muy similar a la de las ayudas europeas competitivas. Esto no implica que debamos abandonar a las comunidades que están menos avanzadas. Para ellas existen los fondos estructurales europeos y regionales, gestionados también, en parte, desde la Administración General del Estado, y la Agencia en particular, que ayudan a crear infraestructuras y capacidades. Desde mi punto de vista, no sería razonable modificar el sistema competitivo en programas como proyec-
tos de I+D o contratos Ramón y Cajal, ya que compiten las mejores ideas y los mejores investigadores/ as, ese sistema garantiza calidad. Pero sí debemos trabajar con las Comunidades para mejorar su capacidad competitiva. Si mejoran a nivel nacional, estarán mejor preparadas para competir en Europa. Por tanto, mi estrategia se basa en dos líneas: inversiones en infraestructuras y personal apoyadas con fondos europeos, y promover instrumentos específicos para que las Comunidades incrementen su competitividad. Uno de mis objetivos es reunirme con responsables de cada Comunidad autónoma para analizar la situación y estudiar mejoras que refuercen su participación en el sistema nacional de ciencia e innovación.
¿Cómo coordina la AEI sus actividades con el Ministerio de Ciencia, el CSIC, el CDTI y las agencias autonómicas para evitar solapamientos y maximizar el impacto, por ejemplo, a través de programas conjuntos o reuniones interinstitucionales?
Evitar solapamientos es prioritario. Algunas comunidades, como Cataluña, lo han conseguido naturalmente, focalizándose en personas e institutos sin replicar la administración estatal. La competencia es más efectiva cuanto mayor es su ámbito, siempre que evitemos solapamientos y haya complementariedad. Eso es lo que intentamos desde la AEI.
Mantenemos diálogo constante con todas las instituciones para impulsar la investigación. Con el CDTI, tenemos una relación importante en convocatorias de colaboración público-privada. Dentro del Ministerio, uno de mis primeros trabajos ha sido reunirme con las secretarías generales para establecer cooperación que evite solapamientos, especialmente en la evaluación del impacto de las investigaciones financiadas. Esta será una constante en la dirección de la Agencia.
España invierte el 1,4 % del PIB en I+D, con solo un 0,7 % proveniente de empresas. ¿Qué medidas impulsa la AEI, como los 320 millones de euros para proyectos público-privados en 2024 o los 21,1 millones del programa Torres Quevedo, para fomentar la colaboración entre universidades, centros de investigación y el sector privado en áreas como inteligencia artificial y biotecnología?
Las ayudas para la colaboración público-privada es una de nuestras convocatorias más importantes. Queremos que los resultados de la investigación lleguen al sector productivo lo antes posible. La convocatoria «Prueba de Concepto», que nació con fondos de recuperación y que quiero mantener a toda costa, es clave en este objetivo. Solo pueden acceder quienes ya han tenido un proyecto financiado. Su finalidad es que el investigador identifique el potencial de sus resultados de investigación en el ámbito de la innovación. Lo esencial es crear conciencia de que ningún resultado de la investigación con potencial para desarrollar productos o servicios que lleguen al mercado se quede en el cajón.
En el ERC, la Prueba de Concepto existe desde 2011 y ha sido un éxito: ha despertado un fuerte interés por transferir conocimiento ágilmente al tejido productivo. España es el país con más éxito en esta convocatoria del ERC, lo que demuestra nuestro talento y capacidad.
En la Agencia mantenemos esta línea y la impulsaremos siempre que los presupuestos lo permitan. Es el primer paso, hacia proyectos más ambiciosos que consoliden colaboraciones público-privadas y generen impacto real en la economía.
Por último, el programa ATRAE 2025 moviliza 45 millones de euros anuales para atraer talento internacional. ¿Qué incentivos ofrece España para competir con polos científicos como Alemania o EE. UU.?
Hemos incrementado la dotación hasta un máximo de 45 millones y hemos visto mayor demanda que en convocatorias anteriores, debido a la situación en EE. UU. Los centros de investigación nos indicaron un creciente interés por parte de investigadores en este país y la Secretaría de Estado nos pidió ampliar el presupuesto. Así lo hicimos.
ATRAE incentiva a las instituciones a crear puestos estables para investigadores e investigadoras, con solicitudes conjuntas, investigador/a e institución, que fomentan la incorporación de talento externo y su estabilidad. Hay interés en Europa, con agencias como Francia y Alemania lanzando convocatorias similares, y el ERC está considerando un tipo de ayuda de naturaleza similar.
CIENCIA ESPAÑOLA
SEBBM/IP
En las últimas décadas, España ha consolidado su posición en ciencia, tecnología e innovación gracias a la transformación de su ecosistema científico. En este contexto, la Agencia Estatal de Investigación (AEI), creada en 2015, ha desempeñado un papel clave como motor del sistema de I+D+i, gestionando cerca de 7.500 millones de euros y financiando miles de proyectos y programas. Al celebrar su décimo aniversario, la AEI destaca por su apoyo a la investigación y su contribución al desarrollo científico y tecnológico del país
Antes de 2015, los recursos para la investigación estaban dispersos entre múltiples ministerios y organismos, lo que generaba ineficiencias, duplicidades y un impacto limitado de las inversiones públicas. Esta situación, que lastraba el potencial científico del país, llevó a la comunidad investigadora a reclamar un modelo más centralizado y eficiente. El verdadero punto de inflexión para la investigación científica en España llegó con la aprobación de la Ley 14/2011, de 1 de junio, de la Ciencia, la Tecnología y la Innovación. Respaldada por las Cortes Generales, esta norma sentó las bases de un nuevo modelo de financiación y gestión de la investigación pública, al establecer la creación de la Agencia Estatal de Investigación (AEI).
Fue en su articulado donde la ley fijó un mandato claro: en el plazo máximo de un año debía aprobarse el estatuto de la nueva agencia, concebida para agilizar y profesionalizar la gestión de los fondos destinados a la I+D+i. La AEI no solo respondía a una demanda histórica del sector, sino que nacía con vocación de alinearse con las mejores prácticas internacionales, tomando como referencia modelos como el del Consejo Europeo de Investigación (ERC), la National Science Foundation (NSF) de Estados Unidos o la Deutsche Forschungsgemeinschaft (DFG) de Alemania.
Sin embargo, el camino hacia su constitución no fue sencillo. Las elecciones generales de diciembre de 2011 supusieron un cambio de gobierno que suspendió la creación de nuevas agencias estatales, lo que retrasó el proyecto. En junio de 2012, el entonces ministro de Economía y Competitividad, Luis de Guindos, impulsó una enmienda a los Presupuestos Generales del Estado para permitir, de forma excepcional, la creación de la AEI, con el objetivo de que estuviera operativa en 2013. No obstante, las trabas administrativas y políticas persistieron, y no fue hasta el 27 de noviembre de 2015 cuando se aprobó el Real Decreto 1067/2015, por el que se formalizó su creación. La agencia comenzó a operar efectivamente en 2016, con la primera reunión de su Consejo Rector, y recibió su primer presupuesto propio en 2017.
Adscrita inicialmente al Ministerio de Economía, Industria y Competitividad, la AEI asumió funciones clave que antes ejercían órganos como la Secretaría de Estado de I+D+i y las direcciones generales de Investigación Científica y Técnica y de Innovación y Competitividad. Este proceso de transición, aunque complejo, permitió centralizar la gestión de la I+D+i, eliminar redundancias y mejorar la coordinación entre los actores del sistema científico español. En 2018, con la reactivación del Ministerio de Ciencia,
POLÍTICA CIENTÍFICA
Innovación y Universidades, la AEI se integró en su estructura, consolidándose como el principal motor financiero de la investigación científica y técnica en España.
Desde su creación, la AEI ha gestionado presupuestos crecientes, lo que refleja su importancia estratégica dentro del sistema español de I+D+i. Su estructura organizativa se basa en 19 áreas y 55 subáreas temáticas, lo que permite una asignación especializada y coherente de los recursos. En 2017, su primer ejercicio con presupuesto propio, gestionó 609,9 millones de euros. En 2018, la cifra aumentó un 4,94 %, hasta los 640,1 millones. Entre 2018 y 2020, el presupuesto se mantuvo relativamente estable. Sin embargo, en 2021 experimentó un incremento del 29 %, alcanzando los 825,7 millones, impulsado por las políticas de recuperación tras la pandemia de COVID-19. En 2022, el presupuesto dio un salto del 64,48 %, situándose en 1.358,2 millones, y en 2023 alcanzó su máximo histórico: 1.420 millones, con un aumento adicional del 4,55 %. En 2024, con presupuestos prorrogados, se mantuvo en torno a los 1.300 millones, y para 2025 se estima que superará nuevamente los 1.000 millones, según las previsiones oficiales.
En total, la AEI ha gestionado cerca de 7.500 millones de euros desde su puesta en marcha. Estos fondos se han distribuido de forma equilibrada entre investigación básica y aplicada, priorizando áreas estratégicas como la salud, las ciencias físicas, la inteligencia artificial, las energías renovables y la biomedicina. Todo ello en línea con los objetivos del Plan Estatal de Investigación Científica, Técnica y de Innovación (PEICTI) y del Plan Estatal 2017–2020.
Diversidad del sistema de I+D+i
La AEI respalda a una amplia gama de beneficiarios que reflejan la diversidad del sistema científico español, incluyendo universidades, organismos públicos de investigación como el Consejo Superior de Investigaciones Científicas (CSIC), centros tecnológicos y empresas privadas. En líneas generales, un 20 % de las ayudas suelen ir para el CSIC y un 60 % para las universidades. En cuanto a la financiación de proyectos de alto impacto, los proyectos que han recibido más respaldo económico por la AEI en el área de Biociencias y Biotecnología han sido los siguientes en 2024: la Universidad de Granada lidera con dos proyectos destacados que suman más de 2 millones de euros. El primero, dotado con 1.066.129,65 €, introduce un citómetro de masas y un nanocitómetro de alta resolución en el Centro de Investigación GENYO. Estos equipos, únicos en España, permitirán análisis multiparamétricos a nivel unicelular y de nanopartículas,
con aplicaciones en vesículas extracelulares y biomedicina. El segundo, con 1.020.333,95 €, incorpora un microscopio confocal de súper-resolución para caracterizar células únicas y exosomas, fortaleciendo la investigación en el Parque Tecnológico de Ciencias de la Salud de Granada.
En Sevilla, la Universidad Pablo de Olavide (UPO) recibe 1.259.000 € para potenciar su animalario, un referente europeo en modelos animales. Este proyecto incorpora tecnologías CRISPR y biología sintética, consolidando a la UPO como un centro global en experimentación animal. Asimismo, la Universidad de Sevilla, con 1.999.500 €, establece una Unidad de Imagen y Cirugía Traslacional con resonancia magnética y una plataforma quirúrgica, única en Andalucía, que beneficiará a más de 100 grupos de investigación.
En Valencia, el Instituto de Biomedicina (IBV-CSIC) adquiere un criomicroscopio electrónico de última generación con 1.197.900 €, democratizando el acceso a la biología estructural en la Comunidad Valenciana. Este equipo, ubicado en la nueva sede Yúfera, promete reducir costes y mejorar la resolución en estudios de macromoléculas biológicas.
La Universidad de Murcia, con 1.010.178,35 €, instala un espectrómetro de masas de alta resolución para estudios avanzados posicionándose a la vanguardia en proteómica y metabolómica. Este equipamiento permitirá identificar biomarcadores clave para enfermedades como el cáncer, beneficiando a investigadores nacionales e internacionales.
Por su parte, la Universidad de Málaga recibe 1.512.495 € para una plataforma de secuenciación de alto rendimiento, combinando lecturas cortas y largas por nanoporos. Este sistema, ubicado en el Centro de Supercomputación y Bioinnovación, optimizará proyectos de transcriptómica y epigenómica, mientras que la Universidad de Córdoba, con 1.866.042,14 €, implementa una plataforma proteómica para el sector agronómico y medioambiental, alcanzando niveles de detección a escala de célula única.
El Centro Nacional de Investigaciones Oncológicas (CNIO) en Madrid obtiene 4.500.000 € para avanzar en la investigación del cáncer, mientras que el Centro de Investigación en Medicina Molecular y Enfermedades Crónicas (CIMUS) de la Universidade de Santiago de Compostela y el Departamento de Medicina y Ciencias de la Vida de la Universitat Pompeu Fabra reciben 2.250.000 € cada uno, consolidando su liderazgo en biomedicina y biología molecular.
Estas inversiones no solo fortalecen la investigación básica, sino que también fomentan la traslación al sector biosanitario y empresarial. Proyectos como los de la Universidad de Almería (1.699.999,42 €) para
estudios de metabolómica clínica mediante resonancia magnética nuclear, o los de la Universidad de Cádiz (1.035.079 €) para microscopía confocal avanzada, destacan por su potencial para mejorar diagnósticos y tratamientos, así como para colaborar con la industria.
Centros e investigación de excelencia
Uno de los programas más emblemáticos de la Agencia Estatal de Investigación (AEI) es el de acreditaciones Severo Ochoa y María de Maeztu, que reconocen la excelencia investigadora a nivel internacional. En abril de 2024, el Ministerio de Ciencia, Innovación y Universidades (MICIU), a través de la AEI, publicó la resolución de esta convocatoria, destinando 75,7 millones de euros, lo que supone un incremento del 78 % respecto a 2018, para financiar a nueve centros y ocho unidades de excelencia reconocidos por su impacto científico y liderazgo internacional. Actualmente, España cuenta con 64 centros y unidades acreditadas con este prestigioso sello de calidad.
Cada centro acreditado como Severo Ochoa recibirá 4,5 millones de euros durante cuatro años, mientras que las unidades María de Maeztu, de menor tamaño, obtendrán 2,25 millones en el mismo periodo. Esta financiación está orientada a reforzar la
organización interna, la planificación estratégica de la investigación, la implementación de buenas prácticas de gestión, la visibilidad internacional y la captación de talento joven. Además, se han previsto ayudas para la formación de 138 nuevos investigadores, mediante tesis doctorales alineadas con las líneas prioritarias de los centros y unidades acreditados. Entre los centros Severo Ochoa seleccionados en la convocatoria 2024 figuran el Centro Nacional de Investigaciones Oncológicas (CNIO), el Instituto de Ciencias del Mar (ICM), la Estación Biológica de Doñana (EBD), el Instituto de Ciencias Fotónicas (ICFO), el Institut Català d’Investigació Química (ICIQ), el Institut de Física d’Altes Energies (IFAE), el Donostia International Physics Center (DIPC), el Instituto de Ciencia de Materiales de Madrid (ICMM) y la Barcelona School of Economics (BSE).
En cuanto a las unidades María de Maeztu, han sido seleccionadas el Departamento de Medicina y Ciencias de la Vida de la Universitat Pompeu Fabra (MELIS-UPF), el Institut de Ciència i Tecnologia Ambientals (ICTA), el Centro de Investigación en Medicina Molecular y Enfermedades Crónicas (CiMUS), el Institut de Ciències del Cosmos (ICCUB), el Instituto de Ciencia Molecular (ICMOL), IMDEA
POLÍTICA CIENTÍFICA
Software, el Instituto Universitario de Investigación de Matemáticas de la Universidad de Sevilla (IMUS) y el Institut Català de Paleoecologia Humana i Evolució Social (IPHES).
Entre los acreditados, el ICMM, IMDEA Software, CiMUS e IMUS han recibido el reconocimiento por primera vez, mientras que el CNIO, ICFO y BSE han renovado su acreditación por cuarta vez consecutiva, lo que refleja su sostenido liderazgo científico. Estas acreditaciones, gestionadas bajo el Subprograma de Fortalecimiento Institucional del Plan Estatal de Investigación Científica, Técnica y de Innovación (PEICTI), no solo premian la excelencia científica, sino que promueven la colaboración con el tejido empresarial y social, aumentando así el impacto económico y social de la investigación producida en España.
Adicionalmente, la atracción y retención de talento ha constituido otro pilar clave en la política científica de la AEI. Los contratos Ramón y Cajal, que en 2025 celebran su 25º aniversario, han sido determinantes para incorporar al sistema español de ciencia y tecnología a investigadores altamente cualificados. En 2023, la AEI concedió 497 contratos Ramón y Cajal, con una dotación de 98,8 millones de euros, permitiendo que estos profesionales desarrollen proyectos propios en instituciones públicas y privadas.
En paralelo, los contratos Juan de la Cierva, dirigidos a investigadores en etapas iniciales, han facilitado la transición hacia la investigación independiente. En 2023, se otorgaron 650 contratos con una inversión total de 40,5 millones de euros. A estos se han sumado las convocatorias del Certificado R3, que reconoce a investigadores establecidos por su calidad e independencia científica conforme a la Ley 14/2011. En la convocatoria de 2024, cerca de 280 investigadores han obtenido este reconocimiento. En conjunto, estos programas han fortalecido la base investigadora del país, contribuyendo a reducir la fuga de talento y promoviendo carreras científicas estables
Por otro lado, la transferencia tecnológica constituye una prioridad estratégica para la AEI, que promueve activamente la colaboración entre el sector público y privado para acelerar el paso del conocimiento generado en laboratorios hacia el mercado. En este ámbito, destacan las ayudas Torres Quevedo y los Doctorados Industriales, que financian contratos de investigadores en empresas y proyectos de alto valor tecnológico, fortaleciendo así el tejido innovador. En la convocatoria 2025, las ayudas Torres Quevedo han incentivado la incorporación de talento consolidado en el sector privado, reforzando la competitividad empresarial mediante la ciencia. Además, la convocatoria Prueba de Concepto, dotada con 30
millones de euros, ha ofrecido respaldo económico para validar la viabilidad técnica y comercial de descubrimientos científicos, reduciendo el riesgo para la inversión privada.
Asimismo, los proyectos de Colaboración PúblicoPrivada, que en 2024 movilizaron 320 millones de euros, han fomentado alianzas estratégicas entre universidades, centros de investigación y empresas, generando soluciones innovadoras con impacto económico y social tangible. Finalmente, la convocatoria de Consolidación Investigadora 2024, con un presupuesto de 50 millones de euros, ha apoyado la creación de puestos permanentes y la renovación de equipamientos científicos en laboratorios de entidades sanitarias públicas y privadas sin ánimo de lucro, así como en institutos de investigación acreditados, fortaleciendo la sostenibilidad de las capacidades científicas nacionales.
Una agencia más global
La cooperación internacional constituye un eje fundamental para la AEI. Actualmente, España es el segundo país de la Unión Europea en participación en proyectos de Horizon Europe (2021-2027). En 2024, la AEI ha financiado 193 Proyectos de Colaboración Internacional (PCI) con un presupuesto de 46,2 millones de euros, abarcando 20 programas transnacionales, principalmente vinculados a Horizon Europe. Además, ha destinado 12 millones de euros a 53 ayudas para la Gestión y Preparación de Proyectos Europeos, facilitando así la participación española en convocatorias internacionales. Para 2025, la AEI gestionará más de 50 programas transnacionales en colaboración con la Fundación Española para la Ciencia y la Tecnología (FECYT), según un acuerdo firmado en 2024.
Entre los acuerdos bilaterales, destacan los firmados con la Deutsche Forschungsgemeinschaft (Alemania) para proyectos en ciencias e ingeniería, y con el Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) de Brasil, enfocados en biotecnología y energías renovables. La AEI también participa en programas internacionales como el Collaborative Research in Computational Neuroscience (CRCNS) con la National Science Foundation (NSF) y los National Institutes of Health (NIH) de EE.UU., centrado en neurociencia computacional; el EIG EU-CELAC, que impulsa proyectos con América Latina en biodiversidad y salud; y el EIGConcert Japan, destinado a colaboraciones en tecnologías emergentes con Japón. Asimismo, la cooperación con la Oficina Española de Ciencia y Tecnología (SOST) en Bruselas y la red EURAXESS fortalece la movilidad de investigadores, facilitando intercambios y estancias internacionales.
Visión global
José Manuel Fernández de Labastida es el cuarto director general de la AEI, tras Domènec Espriu (20222024), Enrique Playán (2019-2021) y Marina Villegas (2016-2018). En su discurso de toma de posesión desveló una ambición clara: situar a esta institución al nivel de las principales agencias de financiación de la ciencia en el mundo. Su objetivo, declarado sin ambages, es colocarla en la misma liga que la National Science Foundation (NSF) de Estados Unidos, la Deutsche Forschungsgemeinschaft (DFG) de Alemania o el prestigioso Consejo Europeo de Investigación (ERC). «No es una ambición teórica», ha afirmado, recordando su experiencia directa en el ERC, donde ha ocupado puestos de responsabilidad durante más de una década. «Nuestra Agencia tiene el potencial para estar a la altura de estas instituciones», ha insistido.
No esconde que el camino no será fácil. La Agencia Estatal de Investigación, creada en 2015, aún no goza del reconocimiento internacional que Fernández de Labastida considera que merece, un déficit que atribuye en buena parte a la juventud de la institución. Pero su compromiso es firme: transformar esa situación y lograr que la Agencia sea un referente internacional en financiación de la investigación.
Para ello, no parte de cero. Como ha recordado, España sentó las bases del actual modelo en 2006 con la aprobación de la Ley de Agencias
Estatales, un proyecto en el que el propio Fernández de Labastida participó desde el grupo de trabajo interministerial. Esta ley —derogada en 2015 pero parcialmente reintegrada en 2020 en la Ley de Régimen Jurídico del Sector Público— permitió configurar un modelo administrativo moderno, orientado a resultados, con autonomía operativa y responsabilidad por objetivos.
Desde su creación, la Agencia ha sabido consolidarse como un actor clave en la política científica española. Su estructura, el trabajo de su personal y la dirección de quienes le han precedido han permitido posicionarla en el sistema de I+D nacional. Pero el nuevo director cree que es el momento de dar un salto cualitativo.
Para ello, ha presentado seis pilares estratégicos sobre los que articular su mandato.
El primero es dotar a la Agencia de un contrato de gestión sólido. «Es urgente», ha dicho. Este contrato debe fijar objetivos plurianuales, mecanismos claros de evaluación y una orientación inequívoca hacia el servicio a la comunidad científica. «Solo así podremos consolidar nuestro prestigio nacional e internacional», ha subrayado.
El segundo pilar es reforzar una cultura organizativa basada en la calidad, la transparencia y la simplificación administrativa. La Agencia debe ir más allá del mero cumplimiento formal de los procedimientos. «Europa se está moviendo hacia la simplificación administrativa, y debemos izar la vela para que esos vientos nos empujen hacia modelos basados en la confianza», ha señalado.
El tercer eje es reforzar la cooperación, tanto a nivel nacional como internacional. Fernández de Labastida quiere que la Agencia sea un actor central en la articulación del sistema español de ciencia, tecnología e innovación. Y, al mismo tiempo, que tenga un papel protagonista en la cooperación científica internacional, en especial en el marco europeo. El cuarto pilar pasa por instaurar una cultura de mejora continua. El Comité Científico y Técnico de la AEI debe estar en el centro de la toma de decisiones estratégicas. En este ámbito, ha destacado la importancia de incorporar reformas acordadas en el Espacio Europeo de Investigación y el uso progresivo de tecnologías avanzadas —como la inteligencia artificial— para mejorar los procesos de evaluación y financiación. «Nuestro sistema de I+D está alcanzando niveles homologables a los países de nuestro entorno, y es hora de avanzar en la mejora de nuestros instrumentos, apostando por la confianza en los investigadores y en los centros», ha defendido. El quinto elemento es alinear a la Agencia con las nuevas prioridades europeas. Fernández de Labastida ha citado los informes de Mario Draghi y Enrico Letta sobre el futuro de la competitividad europea, que insisten en fortalecer las instituciones científicas, atraer talento y fomentar la transferencia de conocimiento hacia la innovación. La Agencia, ha dicho, debe ser una herramienta clave para lograr estos objetivos en España.
El sexto y último pilar es apostar decididamente por la sostenibilidad, la igualdad y la diversidad. «No es un añadido, sino un principio estructural», ha afirmado. La actividad investigadora debe contribuir a los Objetivos de Desarrollo Sostenible y avanzar en políticas activas para garantizar entornos más inclusivos y equitativos. Como prueba del compromiso en esta línea, Fernández de Labastida ha recordado que el Consejo Rector acaba de aprobar el segundo plan de igualdad de la Agencia.
Con este programa, el nuevo director asume el reto de situar a la Agencia Estatal de Investigación a la altura de los mejores. No será un camino corto ni sencillo, pero Fernández de Labastida ha dejado claro que, con trabajo, rigor y ambición, el objetivo es alcanzable.
EL RETO FARMACÉUTICO DE EUROPA: CÓMO EQUILIBRAR INNOVACIÓN Y ACCESO
Ion Arocena Director general de AseBio
La reforma de la legislación farmacéutica europea avanza con el respaldo del Consejo de la UE a una propuesta de compromiso. El texto propone cambios en protección regulatoria, incentivos como vales de exclusividad y mayor claridad en la exención hospitalaria. Busca modernizar el marco legal, fomentar la innovación y mejorar el acceso equitativo a medicamentos en la UE.
Tras meses de intensas negociaciones marcadas por diferencias técnicas y políticas, el pasado 4 de junio la Presidencia polaca del Consejo de la Unión Europea logró que el Comité de Representantes Permanentes (Coreper I) respaldara su propuesta de texto de compromiso sobre el Paquete Farmacéutico. Superado este paso, la atención se centra ahora en la fase de trílogos con el Parlamento Europeo y la Comisión que comenzó el pasado 17 de junio en Estrasburgo y que continúa bajo la Presidencia danesa del Consejo. Siendo esta una etapa decisiva con el objetivo de concluir
la reforma de la legislación farmacéutica general antes de que se produzca un nuevo ciclo político en Bruselas.
El aval del Coreper I representó un punto de inflexión. La propuesta del Consejo ofreció una oportunidad para modernizar el marco regulador actual, posicionar a la Unión Europea como referente en innovación y mejorar el acceso de los ciudadanos europeos a medicamentos innovadores. En este contexto, la industria biotecnológica juega un papel clave, como ya han subrayado las instituciones europeas.
La biotecnología ha transformado el desarrollo de terapias y
medicamentos, con más de 300 aprobaciones y un amplio pipeline de moléculas en investigación. Su impacto es fundamental en ámbitos como la medicina personalizada, las terapias avanzadas, los diagnósticos, las vacunas o la lucha contra la resistencia antimicrobiana. De hecho, en Europa, esta industria es responsable de aproximadamente dos tercios de las innovaciones biomédicas actuales.
Protección regulatoria: buscando el equilibrio entre innovación y acceso En cuanto a los aspectos regulatorios, el Consejo propuso un periodo base de Protección de Datos
Regulatorios (RDP) de ocho años, superior a los seis planteados por la Comisión y los siete y medio del Parlamento Europeo. Este plazo podría extenderse hasta un máximo de nueve años mediante el uso del vale transferible de exclusividad (TEV).
Otro punto clave en debate es la protección de mercado, que permite una exclusividad adicional tras finalizar la protección de datos del medicamento. La propuesta del Consejo establece una protección estándar de un año, ampliable a dos si se cumplen ciertos criterios, como abordar necesidades médicas no cubiertas o demostrar un beneficio clínico relevante con nuevas indicaciones. Esta posición contrasta con la de la Comisión y el Parlamento, que abogan por una duración inicial de dos años.
Esta propuesta del Consejo busca un equilibrio entre el estímulo a la innovación y la necesidad de un acceso más rápido y equitativo a los tratamientos.
Para los medicamentos huérfanos, el Consejo planteó una exclusividad comercial de diez años, con la posibilidad de ampliarla en hasta dos años adicionales (uno por cada nueva indicación relacionada con enfermedades raras), siempre que se consigan antes de que finalice el periodo original y solo en un máximo de dos ocasiones.
En términos generales, la posición del Consejo se inclinó hacia la continuidad con respecto al modelo actual, con ajustes puntuales en lugar de una transformación profunda del sistema. En este sentido, desde AseBio seguimos insistiendo en la importancia de los incentivos para atraer inversión, destacando la necesidad de un entorno estable y predecible. Frente a la visión más restrictiva de la Comisión y el Parlamento, el enfoque del Consejo puede considerarse más favorable a mantener un marco atractivo para la
inversión, aunque los trílogos son clave para definir el equilibrio final
Incentivos e innovación en antimicrobianos
Uno de los elementos más novedosos de la futura legislación que se está discutiendo es la creación de los «vales transferibles de exclusividad» (TEV), diseñados para incentivar el desarrollo de antimicrobianos prioritarios. Este mecanismo permite extender en un año la protección de datos de otro medicamento autorizado, no necesariamente el antimicrobiano desarrollado. El Consejo propuso que estos vales solo puedan utilizarse si el medicamento beneficiario no supera los 490 millones de euros en ventas anuales durante los cuatro primeros años, y solo en el quinto año de protección. Además, los TEV caducarían si no se utilizan en cinco años y su número se limita a cinco, frente a los diez inicialmente previstos. Aunque esta medida representa un enfoque interesante para fomentar la innovación en antibióticos, las limitaciones impuestas en cuanto a cantidad y condiciones de uso generan dudas sobre su efectividad real como incentivo.
Exención hospitalaria: necesidad de una definición clara
Respecto a la exención hospitalaria, una herramienta crucial para ofrecer tratamientos innovadores en contextos excepcionales fuera del procedimiento de autorización centralizado, se valora positivamente el esfuerzo por armonizar su aplicación a nivel europeo. Es esencial que esta opción mantenga su carácter de uso excepcional y no se convierta en una vía paralela de acceso con un nivel inferior de exigencia regulatoria, para productos que deberían perseguir una aprobación de mercado centralizada ante la EMA, que es necesaria para que las terapias avanzadas puedan estar disponibles en el
conjunto de la UE. Por ello, es fundamental que el concepto de «uso no rutinario» quede claramente definido en la nueva legislación, asegurando así que se preserve su finalidad original.
Conclusión: una reforma que busca equilibrio y liderazgo
El acuerdo alcanzado durante la Presidencia polaca supuso un avance significativo en la reforma de la legislación farmacéutica en la UE. La propuesta del Consejo reflejó un enfoque continuista que prioriza la estabilidad del sistema actual, buscando un delicado equilibrio entre promover la innovación —especialmente en biotecnología— y garantizar un acceso justo y rápido a los medicamentos.
La fase de negociación con el Parlamento y la Comisión es determinante para definir si Europa consigue consolidar su liderazgo global en el ámbito de la salud, sin renunciar a la equidad ni a la eficiencia. Desde AseBio esperamos que haya nuevos avances en la negociación de trílogos de este mes de septiembre.
En conclusión, y con la vista puesta en el futuro, el rumbo que adopte la Unión Europea en esta reforma marcará su papel en el contexto internacional. Un marco que incentive la inversión y proteja la innovación puede fortalecer la autonomía estratégica europea y su competitividad frente a potencias como Estados Unidos o China. Por el contrario, un enfoque que no favorezca la innovación en Europa podría frenar el desarrollo de terapias punteras y acelerar el traslado de los proyectos innovadores fuera del continente, afectando tanto al tejido industrial como al acceso de los ciudadanos europeos a nuevos tratamientos. La reforma es, por tanto, también una decisión sobre el modelo de liderazgo que Europa quiere ejercer en el ámbito de la salud global.
Redefine Functional Metabolic Analysis
Agilent Seahorse XF Flex Analyzer—The future of 3D discovery
The new Agilent Seahorse XF Flex 24-well platform is redefining what’s possible in functional metabolic analysis, empowering researchers to explore metabolism across a broader range of model systems than ever before. From traditional 2D cell cultures to cutting-edge 3D organoids, spheroids, and tissue models, this advanced platform delivers a more physiologically relevant view of metabolism in disease. By bridging the gap between in vitro research and real-world biology, the Seahorse XF Flexenhances research translatability, unlocking deeper insights into cellular function and disease mechanisms.
Scan the code to learn more.
DE-006635
Research Use Only. Not for use in diagnostic procedures. This information is subject to change without notice.
OCHO AÑOS DE EXPERIENCIA CON EL LABORATORIO VIRTUAL
«CIBERTORIO»
Ángel Herráez
Bioquímica y Biología Molecular Departamento de Biología de Sistemas Universidad de Alcalá, Alcalá de Henares, Madrid
Estimados compañeros:
Como muchos sabréis, desde hace tiempo he venido desarrollando «Cibertorio» [1], un laboratorio virtual que simula en el ordenador experimentos propios de la biología molecular, basados en el análisis de secuencia del material genético. Ofrece una serie de combinaciones de muestras de ADN y reactivos, bien para preparar reacciones de fragmentación del ADN con enzimas de restricción o bien reacciones de amplificación mediante PCR. En ambos casos, el análisis de los resultados se realiza empleando electroforesis en gel de los fragmentos de restricción del ADN o los resultantes de la amplificación.
En ocasiones anteriores se ha presentado este recurso [2-7], disponible de forma libre, en español e inglés, para toda la comunidad de educadores en bioquímica y
ciencias afines a través de la sede
web Biomodel UAH. Este año decidí hacer una encuesta entre los profesores que han venido empleando el «Cibertorio» en algún momento a lo largo de los últimos siete u ocho años, recabando información sobre sus percepciones y el modo como lo han estado usando con los estudiantes a su cargo. Junto con el resultado obtenido de esta consulta quiero presentar aquí algunas reflexiones sobre la utilidad de esta herramienta y el mejor modo como estimo podemos aplicarla en la práctica docente para conseguir beneficios en el aprendizaje de nuestros alumnos.
Algunas características que entiendo importantes con respecto a la posibilidad de emplear provechosamente simulaciones y laboratorios virtuales, y que he intentado incorporar como parte
del diseño de «Cibertorio» y de otros simuladores disponibles en Biomodel, incluyen:
• Superar las limitaciones que, en muchas circunstancias, pueden afectar a los laboratorios docentes: espacio, tiempo, equipamiento, muestras, reactivos, requisitos de seguridad.
• Enriquecer la experiencia de aprendizaje, proporcionando un complemento a la experimentación real, oportunidades de repaso o refuerzo, familiarización con las técnicas fuera del laboratorio físico, o acceso a técnicas modernas no disponibles en el centro.
• Simulaciones informáticas como éstas pueden proporcionar tanto alternativas como complemento a la experimentación real.
• De modo muy importante, interesa que se evite un material cerrado, donde todo fluye como
Figura 1
Distribución geográfica de los profesores que han aportado información sobre el uso de Cibertorio con sus alumnos.
se espera y el resultado es fijo y predecible. Es mucho más adecuado que haya posibilidad de realizar verdaderos experimentos ya que se dispone de muestras diversas, parámetros modificables que llevan a resultados diferentes, acordes con el diseño que se haya elegido para realizar el experimento.
• El uso de muestras diferentes para cada usuario y en cada ocasión, asignadas aleatoriamente, proporciona resultados diferentes, incluso resultados negativos (experimentos no exitosos), tal como ocurre en la vida real. Cabe mencionar igualmente algunos beneficios que podrán conseguirse con el empleo de laboratorios virtuales bien elaborados:
• Los estudiantes llegan al laboratorio físico mejor preparados, más familiarizados con lo que van a hacer en él.
• Los estudiantes pueden repasar su trabajo práctico con posterioridad, repetir ensayos, probar el efecto de modificar las condiciones del experimento, libremente.
• La formación y entrenamiento se aproximan más a la investigación real en un laboratorio.
• Se aprende a emplear el método científico, a través de la repetición, la modificación de condiciones y la observación de resultados diversos que permitan obtener conclusiones.
• Se explora libremente, estimulando la indagación.
• Se practica más la interpretación de múltiples resultados.
• Se observan resultados realistas, tanto previsibles como inesperados.
Estamos, pues, hablando de un material que no se limita a una demostración para que los
alumnos «la vean», sino que indefectiblemente exige una actividad por su parte, un trabajo experimental –si bien simulado–que deben realizar rigurosamente para poder obtener conclusiones al analizar los resultados.
En las figuras se presenta el resultado de los cuestionarios recibidos, que resumen el impacto de este laboratorio virtual en la comunidad educativa sin necesidad de redactar una descripción explícita.
Como conclusión, y en lugar de usar mi propio punto de vista, he recogido dos comentarios especialmente detallados recibidos de sendos profesores que manifiestan su experiencia tras el uso de este laboratorio virtual con sus alumnos. Estimo que recogen varios de los aspectos clave que han inspirado la construcción de esta herramienta.
Figura 2
Niveles educativos en los que se está empleando Cibertorio.
Figura 3
Tamaño de los grupos de estudiantes y número total de estudiantes con los que se realizan actividades en este laboratorio virtual.
Figura 4
Tipos de experimentos que han alcanzado mayor aplicación con los grupos de estudiantes.
«El ‘Cibertorio’ nos ha permitido realizar prácticas que han sido una experiencia increíble para los estudiantes, ya que son técnicas que no podríamos llevar a cabo en un laboratorio escolar convencional. Lo que más destaco de estos recursos es su accesibilidad y su capacidad para acercar a los alumnos a técnicas de vanguardia de forma práctica y visual. Los estudiantes, en su mayoría muy interesados en las ciencias, disfrutan enormemente de las simulaciones, ya que logran entender procesos complejos de manera dinámica y atractiva. Además, al ser herramientas interactivas, les permite experimentar de manera segura y efectiva, lo cual no sería posible sin estos simuladores. Personalmente, me siento muy agradecida por tener la oportunidad de integrar estos recursos en mis clases. El uso de Biomodel ha enriquecido mi enseñanza y ha aumentado el nivel de motivación y comprensión de mis estudiantes. Llevo ya varios años utilizando estos simuladores, y siempre he obtenido excelentes resultados. En resumen, los recursos de Biomodel no solo son una herramienta educativa impresionante, sino que son un aliado fundamental en la enseñanza de las ciencias en el ámbito escolar. Mi experiencia ha sido más que positiva, y espero poder seguir utilizándolos en el futuro para seguir enriqueciendo la formación de nuestros estudiantes.»
— Profesora de bachillerato, España, 2025.
«Mi gratitud por permitirnos acceder a esta poderosa herramienta de enseñanza aprendizaje. Sin duda, desde que empecé a usarla en mis clases de Genética Molecular y de Herramientas Biotecnológicas, mis estudiantes han perdido el miedo a no poder comprender y se han aventurado mucho más a intentarlo en la vida real. En el laboratorio a mi cargo no dispongo de las herramientas para poder correr todos los análisis que gracias al ‘Cibetorio’ puedo realizar. Mis estudiantes se sorprenden al ver que en la práctica real se obtienen los mismos resultados y me encanta que cada uno pueda experimentar la importancia de ser minucioso en el desarrollo de sus experimentos y que obtengan distintos resultados es fantástico, porque eso los motiva a seguir hasta obtener sus propios resultados y compararlos con sus compañeros. Sin duda, el ‘Cibertorio’ ha ayudado a que muchos jóvenes de mi institución cambien su panorama acerca de la dificultad de hacer este tipo de pruebas y ha redirigido los pasos de varios de ellos, inspirando vocaciones hacia el área de la biología molecular y sus aplicaciones. Ojalá que nos permita seguir usando esta herramienta, que se ha vuelto indispensable para mí y mis estudiantes.»
— Profesor de universidad, Méjico, 2025.
EDUCACIÓN UNIVERSITARIA
Figura 5
Momento de utilización del laboratorio virtual en la práctica docente, en relación con las prácticas en laboratorio real.
Figura 6
Motivaciones que han llevado a los profesores al uso de Cibertorio con sus alumnos.
Figura 7
Características del laboratorio virtual valoradas como positivas por estudiantes y profesores.
Figura 8
Satisfacción global con la herramienta Cibertorio.
Referencias
«Cibertorio», laboratorio virtual de biología molecular. Disponible en línea: https://biomodel.uah.es/lab/ cibertorio/
Diez JC y Herráez A. Iniciación al diagnóstico genético: una aproximación a la medicina molecular. RIECS 2 (2017) 22-27. DOI:10.37536/RIECS.2017.2.1.23
Diez JC y Herráez A. De paternidad, susceptibilidad y otras individualidades.- Una ayuda para aprender diagnóstico genético y medicina molecular. Revista SEBBM 193 (2017) 32-36.
Herráez A. Virtual laboratories as a tool to support learning. Turkish Journal of Biochemistry 45 (2020) 20190146. DOI:10.1515/tjb-2019-0146
Herráez A. Alternativas virtuales: trabaja en casa (casi) como si estuvieras en el laboratorio. Revista SEBBM 206 (2020) 38-40.
Herráez A (2020) Learn by doing — risk-free and from your home: how a genetic test detects Coronavirus. FEBS Network, https://bit.ly/2U4HEkw
Avaro MG, Sturla Lompré J, Sequeiros C, Herráez A. Diseño y ampliación colaborativos de un laboratorio virtual en Biomodel desde Patagonia. Revista SEBBM 212 (2022) 30-33.
INVESTIGACIÓN
Descifrando la especificidad de la proteasa de SUMO humana SENP5
Matthew D Maletic, Anna De Bolòs, Helena Borràs-Gas, Bing Liu, Nathalia Varejão, Virginia Amador, Monique PC Mulder, David Reverter. 2025. Structural basis for the human SENP5’s SUMO isoform discrimination. Nature Communications 16, 4764. https:/ doi.org/10.1038/s41467-025-60029-4
Las modificaciones post-traduccionales de proteínas desempeñan un papel fundamental en la regulación celular, alterando la actividad biológica de las mismas e influyendo en diversos procesos fisiológicos. Entre ellas se encuentra la SUMOilación, un mecanismo en el que la proteína SUMO (Small Ubiquitin-like MOdifier) se une covalentemente a otras proteínas, modulando sus funciones. Es un proceso complejo en el que participan un gran número de proteínas, destacando las proteasas de SUMO, encargadas tanto de activar la molécula de SUMO, como de eliminar su unión a las proteínas diana. En este artículo publicado en Nature Communications se describe la caracterización estructural de la proteasa humana SENP5 con el objetivo de identificar los residuos clave para la discriminación y la especificidad de SENP5 respecto a las diferentes isoformas de SUMO, SUMO1 vs SUMO2/3. La caracterización estructural de SENP5 con las diferentes isoformas de SUMO expone una región con carga positiva directamente implicada en la preferencia de SENP5 por SUMO2/3. Estos contactos clave entre SENP5 y SUMO2/3 abren el camino a la investigación y diseño de posibles inhibidores específicos de SENP5 humana. Este trabajo ha sido realizado por el grupo de investigación del Dr. David Reverter del Instituto de Biotecnología y Biomedicina, dentro del departamento de Bioquímica y Biología Molecular de la Universidad Autónoma de Barcelona, en colaboración con el grupo de la Dr. Monique Mulder, vinculado a la Universidad de Leiden (Holanda) y con el grupo de la Dra. Virginia Amador, IDIBAPS (Barcelona).
La relación de PHD3 en
la progresión de la enfermedad de Alzheimer
Manuel A Sánchez-García, Nieves Lara-Ureña, Rosana MarchDíaz, Clara Ortega-de San Luis, Silvia Quiñones-Cañete, Bella Mora-Romero, Juan M Barba-Reyes, Daniel Cabello-Rivera, Carmen Romero-Molina, Antonio Heras-Garvin, Victoria Navarro, José López-Barneo, Marisa Vizuete, Javier Vitorica, Ana M Muñoz-Cabello , Ana B Muñoz-Manchado, Matthew E Cokman, Alicia E Rosales-Nieves, Alberto Pascual. 2025. Inactivation of the PHD3FOXO3 axis blunts the type I interferon response in microglia and ameliorates Alzheimer’s disease progression. Science Advances 11(22):eadu2244. doi: 10.1126/sciadv.adu2244
Estudios genéticos previos han determinado que la actividad del sistema inmune innato cerebral, la microglía, está relacionada con distintas enfermedades del sistema nervioso y en particular con la enfermedad de Alzheimer. Entre las actividades de la microglía que pueden contribuir al deterioro cerebral, en los últimos años ha cobrado importancia la activación de la vía de interferón tipo I, que se asocia con aumento del deterioro cognitivo. Investigadores del Instituto de Biomedicina de Sevilla, liderados por A. Pascual, han publicado en Science Advances un estudio donde se demuestra por primera vez la relación entre los niveles del enzima prolil hidroxilasa 3 (PHD3) y la expresión de la respuesta a interferón tipo I en la microglía. Esta respuesta es dañina y acelera la patología de Alzheimer. El estudio sugiere que la microglía desarrolla con el tiempo una respuesta tóxica contra el propio tejido debido al estrés por falta de oxígeno. En concreto, se ha abordado el estudio de la contribución de PHD3, una enzima que se acumula en células con niveles bajos de oxígeno, a la producción de señales mediadas por interferón tipo I. La eliminación de la expresión de PHD3 evita esta respuesta, disminuye los daños asociados a la patología de Alzheimer y mejora los defectos cognitivos observados en modelos de ratón de la enfermedad. El estudio va un paso más allá y señala a la proteína FOXO3 como responsable de mantener la respuesta a interferón tipo I inhibida, actuando como un represor transcripcional. Por tanto, este estudio define nuevas dianas terapéuticas y su mecanismo de acción para la enfermedad de Alzheimer.
La helicasa RecD2 destaca por
su
tendencia a cambiar de hebra en las horquillas de replicación
Silvia Hormeño, Cristina Ramos, Javier Mendía-García, Clara Aicart-Ramos, Silvia Ayora, Fernando Moreno-Herrero. 2025.
Bacterial RecD2 is a processive single-stranded DNA translocase with strand-switching capacity at DNA forks. Nucleic Acids Research 53(11):gkaf459. doi: 10.1093/nar/gkaf459
Las helicasas son motores moleculares esenciales en el metabolismo de los ácidos nucleicos. El mal funcionamiento de las helicasas impide la correcta reparación del ADN dañado o propicia la replicación del ADN de forma descontrolada, lo que se asocia directamente con diversas enfermedades humanas, incluido el cáncer. Un estudio realizado por los grupos de Silvia Ayora y Fernando Moreno, del Centro Nacional de Biotecnología (CNB), CSIC, publicado en Nucleic Acids Research, caracteriza en detalle las propiedades de la helicasa bacteriana RecD2, que tiene homología con la helicasa humana HELB, importante a su vez en la replicación y reparación de nuestro genoma. Los investigadores han combinado ensayos bioquímicos con técnicas de molécula individual, como las pinzas ópticas y magnéticas. De esta manera, el equipo ha estudiado la actividad ATPasa, la afinidad de unión a distintas estructuras de ADN y cómo la enzima desenrolla la doble hélice del ADN. El equipo ha demostrado que RecD2 se desplaza a gran velocidad por ADN monocatenario y ha descubierto que es capaz de cambiar de hebra al encontrarse con estructuras como las horquillas de replicación, una actividad conocida como strand switching. Además, basándose en evidencias bioquímicas y en modelos predictivos de la estructura, los autores proponen que RecD2 tiene dos sitios de unión al ADN, uno de ellos en el dominio N-terminal y que sería esencial para explicar el mecanismo molecular que permite el cambio de hebra descubierto. Los resultados de este estudio vienen a confirmar el papel de RecD2 como remodelador de las horquillas de replicación. Gracias a este trabajo ahora se sabe que RecD2 es capaz de alternar entre la hebra conductora y la retardada durante el proceso de replicación. Esta acción puede ser clave para regular la actividad de otras proteínas involucradas (SSB o RecA) y determinar el reinicio de la replicación.
Un ARNlnc regula el metabolismo de las células senescentes
Elena Grossi, Francesco P Marchese, Jovanna González, Enrique Goñi, José Miguel Fernández-Justel, Alicia Amadoz, Nicolás Herranz, Leonor Puchades-Carrasco, Marta Montes, Maite Huarte. 2025. A lncRNA-mediated metabolic rewiring of cell senescence. Cell Reports 44(6):115747. doi: 10.1016/j.celrep.2025.115747
La senescencia es un proceso en el que las células dejan de dividirse de forma permanente al enfrentarse a diferentes tipos de estrés, como el acortamiento de telómeros, la activación de oncogenes o la quimioterapia. Estas células que dejan de proliferar, se mantienen metabólicamente activas y pueden influir en procesos como la progresión tumoral o el envejecimiento, por lo que es fundamental conocer sus mecanismos de regulación. Los grupos de las doctoras Maite Huarte y Marta Montes del CIMA, Universidad de Navarra, en colaboración con L. Puchades-Carrasco (Instituto de Investigación Sanitaria La Fe, Valencia), en un trabajo publicado en Cell Reports, han identificado un ARN largo no-codificante inducido en senescencia (sin-lncRNA, por sus siglas en inglés) que desempeña un papel clave en la regulación metabólica durante la senescencia celular. Este ARN se activa cuando las células entran en senescencia e interacciona con la enzima dihidrolipoamida-S-succinil transferasa (DLST) facilitando su localización mitocondrial esencial para su actividad en el ciclo de Krebs. En ausencia de sinlncRNA, DLST se transloca al núcleo regulando epigenéticamente los programas transcripcionales de genes relacionados con la fosforilación oxidativa. Además, estos resultados ponen de relieve las implicaciones importantes en cáncer ya que la inhibición de sin-lncRNA en células de cáncer de ovario reduce el consumo de oxígeno y aumenta la sensibilidad al tratamiento con cisplatino. Estos hallazgos abren nuevas vías terapéuticas basadas en la reprogramación metabólica mediada por ARN.
INVESTIGACIÓN
Mecanismo de detección de mutaciones abásicas por la ARN polimerasa I
Alicia Santos-Aledo, Adrián Plaza-Pegueroles, Marta Sanz-Murillo, Federico M. Ruiz, Peini Hou, Jun Xu, David Gil-Carton, Dong Wang, Carlos Fernández-Tornero. 2025. Cryo-EM uncovers a sequential mechanism for RNA polymerase I pausing and stalling at abasic DNA lesions. Nature Communications 16(1): 5254. doi: 10.1038/s41467-025-60536-4
Alrededor de 10.000 lesiones abásicas aparecen diariamente en el ADN de las células eucariotas, lo que las convierte en una de las mutaciones más frecuentes. Su detección temprana es clave para el mantenimiento de la integridad genómica y para la supervivencia de la célula, pero todavía se desconocen los mecanismos subyacentes a estos procesos. La ARN polimerasa I es particularmente relevante en la detección de lesiones en el ADN, ya que es responsable de más de la mitad de la actividad transcripcional en la célula al escanear el ADN ribosómico para producir el precursor del ARN ribosómico. El laboratorio del Dr. Carlos Fernández Tornero, del Centro de Investigaciones Biológicas
Margarita Salas (CIB) del CSIC, en un trabajo publicado en Nature Communications, ha desvelado el mecanismo secuencial por el cual la ARN polimerasa I se detiene ante las lesiones abásicas. A través de nueve estructuras atómicas, obtenidas por medio de la crio-microscopía electrónica, este grupo ha revelado que la ARN polimerasa sufre una pausa inicial al detectar la lesión. Posteriormente, añade lentamente un ATP a la hebra naciente de ARN, antes de intentar translocarse sin éxito para quedar finalmente detenida. En este trabajo, en colaboración con el grupo del Dr. Dong Wang de la UCSD, también se describe cómo la ARN polimerasa I adquiere distintas conformaciones tras detectar una lesión abásica, lo que puede facilitar la resolución de las pausas transcripcionales. Este mecanismo único de detección de mutaciones contribuye al funcionamiento normal de la célula, por lo que su alteración puede modificar el equilibrio celular y poner en peligro la supervivencia.
Modelo animal dede la lipofuscinosis neuronal ceroidea autosómica dominante del adulto
Santiago López-Begines, Nozha Borjini, Ángela Lavado-Roldán, Cristina Mesa-Cruz, Fabiola Mavillard, Vera I Wiersma, Fátima Rubio-Pastor, Emanuela Tumini, Carmen Paradela-Leal, María L Chiclana-Valcárcel, Carolina Aguado, Rafael Luján, Wiep Scheper, José L Nieto-González, Rafael Fernández-Chacón. 2025. Neuronal lipofuscinosis caused by Kufs disease/CLN4 DNAJC5 mutations but not by a CSPα/DNAJC5 deficiency. Science Advances 11, eads3393 (2025). doi: 10.1126/sciadv.ads3393
La lipofuscinosis neuronal ceroidea autosómica dominante de inicio adulto (enfermedad de Kufs o CLN4) es una enfermedad neurodegenerativa devastadora causada por mutaciones en el gen DNAJC5. Este gen codifica una co-chaperona de las vesículas sinápticas denominada Cysteine String Protein a (CSPa). El equipo dirigido por el Dr. Rafael Fernández-Chacón (Instituto de Biomedicina de Sevilla IBiS, Hospital Universitario Virgen del Rocío-CSIC-Universidad de Sevilla, Dpto. de Fisiología Médica y Biofísica y CIBERNED (ISCIII)), ha desarrollado y analizado el primer modelo genético en ratones para estudiar esta enfermedad. El estudio, publicado en Science Advances, muestra que los ratones que expresan las versiones mutadas de la proteína CSPa desarrollan alteraciones neurológicas asociadas a la acumulación neuronal de la lipofuscina patológica como ocurre en los pacientes con la enfermedad de Kufs. Es interesante resaltar que, los ratones que carecen completamente de CSPa no muestran la lipofuscinosis patológica. Esto sugiere que, en vez de una simple pérdida de su función normal, la enfermedad resultaría de un efecto tóxico propio de la proteína CSPa mutada debido a una ganancia de función aberrante que podría, por ejemplo, propiciar interacciones anómalas proteína-proteína todavía por descifrar. El estudio deja claro que el nuevo modelo proporciona una herramienta muy útil para futuros estudios en la fisiopatología y terapéutica de este trastorno.
FERMÍN SÁNCHEZ DE MEDINA CONTRERAS
María Dolores Suárez Ortega Catedrática de la Universidad de Granada
El pasado 29 de marzo de 2025 nos dejó el profesor Fermín Sánchez de Medina Contreras, Catedrático emérito del Departamento de Bioquímica y Biología Molecular en la Universidad de Granada. Nació en Ubrique en una familia de, como se decía antaño, boticarios, ejerciendo su padre en Puerto Real (Cádiz). Estudió en el colegio de La Salle de Utrera, donde surgió su interés por la poesía, que mantuvo a lo largo de su vida. También comenzó a desarrollarse su pasión por la música (él se definía como un músico–bioquímico o bioquímico–músico). De esta época procede la famosa «Rosa de Cuba», una habanera que forma parte del repertorio de todas las tunas desde entonces.
Se desplazó a Granada en el curso 1957-58 para estudiar Farmacia, residiendo en el Colegio Mayor Isabel la Católica, donde colaboró en la fundación de la tuna. Posteriormente formó parte, junto con sus hermanos y algunos amigos, del grupo musical los Windys, que cosechó gran éxito y reconocimiento.
En 1963 llegó a la Universidad el Prof. D. Federico Mayor Zaragoza, quien llevó a lo largo de la siguiente década una labor investigadora y docente muy destacada. Fermín, su primer discípulo, se incorporó ese curso, seguido por Eduardo García Peregrín, Magdalena de Ugarte Pérez y María Flor Zafra Ortega. Fermín fue director del departamento en distintas etapas, así como Secretario (1973-1977) y Decano de la Facultad de Farmacia (1981-1985), con una labor muy vinculada a la elaboración de un plan de estudios acorde con la realidad de la profesión. También fue miembro y Presidente de la Academia Iberoamericana de Farmacia. Sus primeros pasos en investigación se centraron en el ciclo del g-aminobutírico, vicario del ciclo de Krebs. Fermín fue secretario del IV Congreso de Bioquímica
en 1967 en Granada, con asistencia de Severo Ochoa y otros Premios Nobel. Posteriormente (año 2000) fue presidente del Comité Organizador del XXIII Congreso de la SEBBM. Durante un año realizó una estancia de investigación en el CNRS de Marsella.
Fermín aplicó la técnica de perfusión hepática con circulación extracorpórea al estudio del papel del hígado y la influencia de las biguanidas en la regulación de la gluconeogénesis, además de la influencia del ejercicio en la gluconeogénesis hepática y renal en ratas, demostrando el papel relevante del riñón. Sus trabajos fueron muy reconocidos internacionalmente y fue invitado a un simposio de Bioquímica clínica en Reisensburg. Posteriormente cedió esta línea a sus colaboradores y comenzó a trabajar en bioquímica de la nutrición con el Dr. Ángel Gil, vinculado a la empresa PULEVA.
Desde 1981 a 1996 sus trabajos se centraron en evaluar el metabolismo proteico en los recién nacidos de bajo peso, por los que en 1983 recibió el Premio Nacional de Investigaciones Perinatológicas. Asimismo, contribuyó a evaluar los efectos de los nucleótidos libres de la leche humana en el metabolismo lipídico en los recién nacidos, que llevó en 1996 a incluir los nucleótidos en las fórmulas lácteas en la legislación europea y de EE.UU.
Organizó y participó en numerosos cursos de Formación Permanente en colegios profesionales, cooperativas, colegios mayores, etc., en los que siempre dejó patente su puesta al día en temas científicos, su amenidad y su facilidad para comunicar.
En suma, nos deja una persona extraordinaria, investigador notable y magnífico profesor, enormemente apreciado por sus compañeros, discípulos y alumnos, tanto de promociones pasadas como recientes.
Descanse en Paz
AYUDANDO A LOS JÓVENES DESDE LA SEBBM: PROGRAMA DE MENTORÍA Y
PREMIOS CAMPUS
Raúl Estevez Povedano
Vocal de mentoría y comunicación SEBBM
IDIBELL-Universidad de Barcelona
La carrera científica es un camino apasionante, pero también desafiante, lleno de incertidumbres, decisiones críticas y una constante necesidad de adaptación. Desde la SEBBM siempre se ha querido apostar por los más jóvenes en dos niveles, premiando a los estudiantes más brillantes y aconsejándoles en los primeros años de su carrera científica. En primer lugar, para ayudar a las jóvenes promesas en el campo de la Bioquímica y Biología Molecular se han convocado los premios campus SEBBM. Todas aquellas universidades interesadas podrán seleccionar a un máximo de tres estudiantes de último año por cada título de grado relacionado con la Bioquímica y Biología Molecular. El criterio de selección será definido por la propia universidad. La SEBBM recibe estas propuestas y selecciona a los premiados, los cuáles recibirán un diploma conmemorativo y un año de Socio Adherido SEBBM. El ser Socio Adherido proporciona entre otras ventajas, el tener acceso a una red de contactos con cerca de 3.000 socios, el poder inscribirse al Congreso SEBBM o a otras actividades de la SEBBM, FEBS e IUBMB a un precio reducido, o el poder participar en el programa de mentoria SEBBM.
Desde la SEBBM, pensamos que la figura del mentor puede ser un pilar fundamental para el desarrollo profesional y personal
de los investigadores, especialmente en las etapas iniciales de su trayectoria. La mentorización es una relación profesional en la que una persona con experiencia (el mentor) guía, apoya y aconseja a otra con menos experiencia (el mentee o aprendiz). En el ámbito científico, esta relación puede abarcar desde la orientación en la elección de líneas de investigación, hasta el desarrollo de habilidades transversales como la escritura científica, la gestión del tiempo, la ética en la investigación y la construcción de redes de colaboración. Es durante el pregrado, el posgrado y los primeros años de la carrera investigadora, en el que los jóvenes científicos enfrentan múltiples retos: definir
su área de especialización, publicar sus primeros artículos, obtener financiación y construir una reputación académica. Un mentor puede ofrecer una perspectiva valiosa basada en su experiencia, ayudando a evitar errores comunes y a tomar decisiones más informadas.
Los mentores son socios de la SEBBM dispuestos a orientar y asesorar de forma desinteresada a otros investigadores. La adhesión al Programa de Mentoría es una opción personal, por tanto, no se va a realizar ninguna evaluación ni exigir una carga excesiva de trabajo. Un buen mentor hace preguntas, escucha, es accesible, comparte su red de contactos, proporciona feedback. Su
cometido no es dirigir las actividades del mentee, sino únicamente tratar de dar consejo y aportar diferentes puntos de vista. Como pensamos desde la SEBBM que no es fàcil ser mentor, aunque sí muy reconfortante, cada año se realiza un taller de mentoría gratuito para los socios dirigidos por especialistas en el tema. Por otro lado, los mentees deben de ser proactivos y contactar directamente con sus posibles mentores. Este año también se ha realizado un taller de formación virtual para los mentees ¿Cómo se articula el programa de Mentoría? El proceso es tremendamente fàcil, toda la información está disponible en la página web de la SEBBM
Cada socio que quiera ser mentor simplemente ha de marcar en su ficha personal que acepta formar parte del programa de mentoría. Así, con este simple acto pasa a formar parte de un listado actualizado de mentores que se encuentra en el área privada. Así, una vez mentor y mentee se ponen de acuerdo, se firma un acuerdo de confidencialidad (formulario online ) y se produce un acuerdo durante un año. Desde la SEBBM pedimos si es posible, que al acabar este acuerdo, ambos rellenen un sencillo cuestionario de valoración, para así poder desde la SEBBM mejorar este proceso. Pensamos también que aunque tradicionalmente la relación
mentor- mentee se ha visto como una relación unidireccional, la mentorización también beneficia al mentor. Acompañar a jóvenes investigadores permite renovar perspectivas, mantenerse actualizado y desarrollar habilidades de liderazgo. Se fomenta una cultura de colaboración y solidaridad dentro de la comunidad SEBBM. En resumen, desde la SEBBM queremos seguir apostando por los jóvenes, el futuro de la investigación en nuestro país. Actividades como los premios Campus SEBBM y la mentorización fortalecen la calidad de la investigación, promueven la innovación y contribuyen a la formación de comunidades científicas más humanas y colaborativas.
Investigadora predoctoral, Instituto de Investigaciones Biomédicas ‘Sols-Morreale’ (IIBM), CSIC-UAM
Falling Walls Lab es una competición internacional de pitch en la que estudiantes y profesionales al inicio de su carrera presentan ideas innovadoras en solo tres minutos, con el objetivo de romper barreras científicas y sociales. La final de la edición española de 2025 se celebró el viernes 30 de mayo en el Salón de Actos del edificio central del CSIC, en Madrid. Durante el evento, cada participante dispuso de tres minutos para presentar su proyecto ante un jurado multidisciplinar.
El primer premio fue para Rocío Vázquez Martínez (Universidad de Zaragoza), con «Breaking the Wall of Cancer Treatment». El segundo lugar lo obtuvo Elena Barba Sarasua (Universidad Complutense de Madrid (UCM)), con «Breaking the Wall of Cancer Shield». El premio del público fue para Javier Rodríguez por «Breaking the Wall of Mental Health Suffering». Rocío representará a España en la final internacional en Berlín el 9 de noviembre de 2025.
El día previo, los 15 finalistas participaron en una sesión formativa en Compluemprende (UCM), donde recibieron orientación sobre comunicación, emprendimiento y desarrollo profesional. Una tarde que permitió a los participantes hacer introspección de sus trabajos, coger ideas de cómo contar su proyecto al día siguiente, y, por supuesto, conocerse entre ellos, relajar nervios, y poder empezar a crear lazos. Porque si algo nos enseña la ciencia es que el progreso se consigue a través
de conexiones, colaboraciones, es decir, trabajando de forma multidisciplinar. El evento fue organizado por el Museo Nacional de Ciencias Naturales (MNCN), CSIC, Compluemprende (UCM), la Embajada de Alemania en España, y la SEBBM. Contó con el apoyo de entidades como INCYDE, Innogate to Europe, Alsa, Iberitos y Pimentón de la Vera como patrocinadores, y la Academia Joven de España, la Universidad de Alicante y DAAD
como colaboradores.
Desde la primera edición la SEBBM ha tenido un papel clave en la organización del evento, a través de su vocal de juventud, Sara García, que fue una de las promotoras, y además desde la creación de la Sección Junior, sus coordinadores han estado implicados en la misma. Un claro ejemplo de que la SEBBM apuesta por la juventud, y apuesta por eventos que pueden impulsar la carrera científica de los más jóvenes.
Vídeo resumen: Final de Falling Walls Lab Spain 2025
Retransmisión de la final: Falling Walls Lab Spain 2025
Falling Walls Lab Spain 2025 ha vuelto a demostrar que la ciencia joven está llena de ideas potentes, con capacidad para transformar el futuro. Una edición inspiradora que, gracias al esfuerzo conjunto de múltiples instituciones, ofrece una plataforma única para visibilizar el potencial investigador emergente.
XVI EDICIÓN PREMIOS MEDES 2025
La SEBBM ha recibido el premio Medes 2025, en su XVI edición, a la Mejor Institución o Entidad destacada en la promoción del español como lengua del conocimiento biomédico. La entrega del premio tuvo lugar el 7 de julio durante la XIX Jornada Medes «El español y la ciencia multilingüe» celebrada en San Lorenzo del Escorial, Madrid, en el marco de los cursos de verano Complutense. En representación de la SEBBM recogieron el premio Lluis Montoliu, secretario de SEBBM, e Inmaculada Yruela, directora de la Revista SEBBM Los Premios MEDES, convocados por la Fundación Lilly, nacen en 2008 con el objetivo de reconocer, en el ámbito de los países de habla hispana, aquellas iniciativas que favorecen la utilización del español en la comunicación del
conocimiento biomédico y de las ciencias de la salud, así como las instituciones que hacen posible el logro de estos objetivos. La evaluación y calificación de las candidaturas presentadas se ha realizado por un jurado, integrado
por profesionales de prestigio reconocido en al ámbito de la comunicación y la publicación científicas en español.
Desde SEBBM damos las gracias por este reconocimiento.
ALBERTO HIPÓLITO CARRILLO DE ALBORNOZ, PREMIO FUNDACIÓN LILLY A LA MEJOR TESIS DOCTORAL EN BIOQUÍMICA Y BIOLOGÍA MOLECULAR 2025
El jurado, formado por la Comisión de Admisiones de la SEBBM, ha concedido un primer premio y dos accésit en la V edición del premio Fundación Lilly a la Mejor Tesis Doctoral en Bioquímica y Biología Molecular 2025. El primer premio ha sido para Alberto Hipólito Carrillo de Albornoz, por su tesis: Dinámicas evolutivas de los cassettes de resistencia en integrones. Profundizando en el modelo de integrón, realizada
en la Facultad de Veterinaria de la Universidad Complutense (UCM) de Madrid. Los accésits han sido para Álvaro de la Gándara Fernández por la tesis Mecanismos moleculares y estructurales de la transposición de ADN dependiente de ATP usando la familia IS21 como modelo, realizada en Centro de Investigaciones Biológicas ‘Margarita Salas’ (CIB), CSIC de Madrid y para Andrés Gámez García-Cervigón por la
tesis Role of sirtuins in haematopoiesis and leukaemia, realizada en el Josep Carreras Leukaemia Research Institute, Badalona, Barcelona.
La entrega de los premios ha tenido lugar el 5 de septiembre de 2025 en el Auditorio del Complejo Cultural San Francisco, Cáceres, en el marco del 47º Congreso SEBBM. Desde SEBBM felicitamos a los premiados.
EL CENTRO DE BIOLOGÍA MOLECULAR, MEDIO SIGLO EXPLORANDO LAS BASES MOLECULARES DE LA VIDA
Los más de 500 científicos y cien grupos de investigación que suma en la actualidad han afianzado el Centro de Biología Molecular Severo Ochoa como uno de los mayores de España y una referencia a nivel mundial, cuando cumple medio siglo explorando las bases moleculares de la vida. Cincuenta años durante los que se han sucedido una cascada de descubrimientos científicos en áreas como la inmunología, virolo-
gía, microbiología, neurociencias o biología celular y del desarrollo, y durante los que se han desarrollado algunas de las patentes más rentables de la historia de la
El Centro de Biología Molecular, medio siglo explorando las bases moleculares de la vida (abc.es)
ciencia en España e impulsado innovadores métodos y técnicas de investigación que forman parte ya de la práctica rutinaria en miles de laboratorios de todo el mundo.
Alberto Hipólito Carrillo de Albornoz Álvaro de la Gándara Fernández Andrés Gámez García-Cervigón
TERCERA CONFERENCIA
SEVERO OCHOA: EPIGENÉTICA, ENVEJECIMIENTO Y CÁNCER
La ciudad de Gijón acogió los días 26 y 27 de junio de 2025 la tercera conferencia Severo Ochoa, centrada en esta edición en el papel de la regulación epigenética en procesos tales como el cáncer y el envejecimiento. Estas jornadas, promovidas de manera conjunta por la SEBBM y la Fundación Carmen y Severo Ochoa (FCySO) han sido coorganizadas por Mario Fernández Fraga, Ana Gutiérrez, Agustín Fernández, Xosé Antón Suárez Puente, Juan Ramón Tejedor y José María Pérez Freije, investigadores del Instituto Universitario de Oncología del Principado de Asturias (IUOPA), del Consejo Superior de Investigaciones Científicas (CSIC) y del Instituto de Investigación Sanitaria del Principado de Asturias (ISPA). El evento ha permitido reunir a algunos de los expertos más influyentes en el campo de la epigenética para poder discutir durante dos intensas jornadas acerca de los últimos avances en esta temática. El conjunto histórico de la Universidad Laboral de Gijón fue sede de uno de los principales
eventos científicos celebrados en Asturias en 2025.
La epigenética explora cómo los factores externos pueden influir en la expresión de nuestros genes, dando lugar a rasgos observables e incluso a la susceptibilidad a determinadas enfermedades sin cambiar el propio código del ADN. Los avances tecnológicos que han surgido en los últimos años han permitido perfilar de manera minuciosa el paisaje epigenético
de nuestras células y tejidos, y el estudio de estas alteraciones epigenéticas podría ayudarnos a entender, entre otros fenómenos, por qué algunas de nuestras células se convierten en tumores o por qué se produce el proceso de envejecimiento.
La conferencia contó con la participación de más de 100 investigadores en distintas etapas de su carrera profesional. El pistoletazo de salida comenzó con
la charla inaugural del Profesor Steve Horvath (Altos lab), experto mundial en el ámbito de la epigenética e inspirador conceptual de los relojes epigenéticos, biomarcadores basados en el estado de metilación del ADN que permiten estimar la edad biológica de los organismos. El resto del programa se organizó en torno a distintas temáticas de rabiosa actualidad, con el fin de explorar las relaciones existentes entre la epigenética y el proceso de envejecimiento (Sesión 1), la sinergia que establecen la epigenética y el cáncer (Sesión 2), el uso de aplicaciones epigenómicas en el ámbito de la salud (Sesion 3) y la irrupción de los análisis computacionales en la era de las tecnologías multiómicas (Sesión 4). Las jornadas contaron también con una sesión exclusiva de charlas y otra sesión de pósteres impartidas por investigadores junior y personal investigador en formación, los cuales deleitaron a los asistentes con resultados
de una calidad científica extraordinaria. A modo de resumen, estas jornadas han dado lugar a discusiones científicas muy inspiradoras que esperamos den lugar a futuras colaboraciones para fomentar el avance y la traslación de estos resultados en el ámbito de la biomedicina. Una vez cerrado el telón de estas jornadas, cedemos el testigo a la organización de la cuarta edición de las conferencias Severo Ochoa, la cual deseamos que coseche el mejor de los éxitos.
Agradecimientos
En nombre de todo el comité organizador y científico, ha sido todo un honor poder organizar estas jornadas en homenaje a la figura de Severo Ochoa en Asturias y poder contar con la participación de expertos mundiales en el campo de la epigenética, tanto a nivel internacional (Steve Horvath, Sara Hägg, Wolfgang Wagner, Stephan Beck, Chrisoph Bock, Andrew E.
Teschendorff), como a nivel nacional (María Blasco, José Vicente Sánchez-Mut, Iñaki Martín-Subero, Sandra Blanco, Pablo Menéndez, Manel Esteller, Pedro MoralQuirós, Biola Javierre, Marta Melé, Raúl F. Pérez e Isabel Mendizabal) a los cuales nos gustaría transmitir toda nuestra gratitud. También agradecemos la participación de las empresas y entidades colaboradoras (MedchemExpress, Novogene, BMKGENE, Agilent, Thermofisher Scientific, Dismed, Asociación Española Contra el Cáncer, Ayuntamiento de Gijón y personal de organización de Laboral Ciudad de la Cultura), así como a la junta directiva de la SEBBM, el patronato de la fundación Carmen y Severo Ochoa, científicos de la comisión organizadora de estos ciclos de conferencias (M.ª Ángeles Serrano, Carlos Gancedo, Mariam Sahrawy, Emilio Lecona y Luis Blanco), y a todos los asistentes por todo su apoyo y confianza. ¡Gracias!
SOCIEDAD
4TH INTERNATIONAL MOLECULAR
BIOSCIENCES PHD AND POSTDOC
CONFERENCE
GLASGOW 2025
Este año el congreso internacional FEBS-IUBMB-ENABLE, dirigido a doctorandos y postdocs, se celebrará en Glasgow del 10 al 12 de septiembre.
CONGRESO BIANUAL DE LAS SOCIEDADES ESPAÑOLAS Y PORTUGUESAS DE MICROSCOPÍA MFS2025. CENTRO DE INVESTIGACIÓN PRÍNCIPE FELIPE (CIPF), VALENCIA
Este año se celebrará en Valencia del 23 al 26 de septiembre 2025, participarán varios miembros de la SEBBM y la microscopía de alta resolución de muestras biológicas tendrá un papel central.
PREMIOS REY JAIME I
Los Premios Rey Jaime I, en su 37ª edición, han galardonado este año la investigación sobre el origen y mutaciones del cáncer, los estudios del virus del papiloma y la labor empresarial filantrópica para ayudar al tratamiento de menores con enfermedades raras, según han dado a conocer los jurados de los galardones.
Investigación Biomédica
El Jaume I en Investigación Biomédica de 2205 es Nuria López Bigas, profesora de Investigación ICREA en el Institut de Recerca Biomèdica (IRB) de Barcelona y profesora asociada de la Universidad Pompeu Fabra. Su investigación se centra en la identificación de mutaciones y genes causantes del cáncer y en la comprensión de los procesos mutacionales que conducen a la acumulación de mutaciones en las células cancerosas. El jurado ha destacado de López Bigas «su investigación innovadora sobre el origen y la evolución de las mutaciones del cáncer y el desarrollo de herramientas para su estudio». Esta categoría es la que cuenta con más mujeres este año, con 12 científicas entre los 28 candidatos (43 %). La Dra. López Bigas es socia SEBBM y miembro del grupo Mujer y Ciencia.
Investigación Clínica y Salud
Pública
El jurado ha otorgado el premio en Investigación Clínica y Salud Pública a Silvia de Sanjosé
Llongueras, investigadora del Instituto de Investigación Biomédica de Bellvitge (Idibell) de l’Hospitalet de Llobregat. Lidera el grupo Infecciones y Cáncer y su investigación se centra en la lucha entre virus y huéspedes, desde una perspectiva evolutiva. El jurado la ha premiado «por sus estudios sobre la relación del Virus del Papiloma Humano y ciertos carcinomas en mucosas genitales y por promocionar el uso de la vacuna-HPV en jóvenes, en particular en niñas adolescentes que ha contribuido a la erradicación del carcinoma de cuello de útero, enfermedad común y, en muchos casos, fatal».
Investigación Básica
El premio en Investigación
Básica ha sido para José Luis
Mascareñas Cid, catedrático de Química Orgánica en la Universidad de Santiago de Compostela (USC) y director del Centro de Investigación en Química Biológica y Materiales Moleculares (CIQUS). El jurado ha destacado sus «trabajos pioneros en química órganometálica, especialmente por introducir reacciones catalizadas por metales en células vivas». El Dr. Mascareñas es socio SEBBM.
Estos son los siete ganadores de los premios Jaume I de 2025.
A esta edición de los premios Rey Jaime I se han presentado 226 candidaturas, más de un tercio representadas por mujeres, que han recibido cuatro de los siete galardones. Desde la SEBBM damos la enhorabuena a todos los premiados en esta edición, especialmente a nuestros socios.
Nuria López Bigas
Silvia de Sanjosé
José Luis Mascareñas
LA GENETISTA MARY-CLAIRE KING, PREMIO PRINCESA DE ASTURIAS DE INVESTIGACIÓN CIENTÍFICA Y TÉCNICA 2025
El jurado del premio Princesa de Asturias destaca la combinación de impacto científico y compromiso social de Mary-Claire King y ha valorado las contribuciones «pioneras» de King «en la aplicación de la genética al estudio y prevención del cáncer, la similitud entre especies y la defensa de los derechos humanos».
La genetista King se graduó en Matemáticas por el Carleton College (Northfield, Minnesota) en 1966 con la calificación cum laude y, posteriormente, realizó su tesis doctoral en Genética en la Universidad de Berkeley, bajo la dirección de Allan Wilson. Ha sido profesora de Genética y Epidemiología en el departamento de Biología Molecular y Celular y es desde 1995 miembro de la Asociación Estadounidense del Cáncer, en la Universidad de Washington y miembro afiliado del Fred Hutchinson Cancer Center. Además, es Doctora Honoris Causa en veinte universidades del mundo y en 2012 fue presidenta de la Sociedad Estadounidense de Genética Humana.
Grandes Contribuciones al Cáncer Junto con el grupo del Dr. Petrakis en 1974 King comenzó a investigar el cáncer de mama y en 1990 consiguió identificar un gen localizado en el cromosoma 17, denominado BRCA1, así como que las mutaciones en este gen estaban implicadas en el desarrollo de determinados tipos de cáncer de mama y ovario. Los estudios de la doctora King permitieron establecer por primera vez la estrecha relación entre un gen concreto y el riesgo de desarrollar cáncer de mama u ovario,
contribuyendo de este modo a salvar millones de vidas. «Este trabajo supuso un cambio de paradigma en el conocimiento del origen del cáncer, una enfermedad compleja de la que no se sospechaba que pudiese tener un origen genético, y en su tratamiento», señala el jurado. A partir de entonces, añaden, «el estudio de esta enfermedad y de otras enfermedades denominadas raras, como la enfermedad de Huntington o la fibrosis quística, comenzó a enfocarse desde otro punto de vista».
Además, con sus investigaciones descubrió la sorprendente similitud genética entre humanos y chimpancés y desarrolló una metodología rigurosa, tanto científica como legal, con la que se ha conseguido la reunificación familiar de personas desaparecidas, un total de 138 por el momento. Durante su labor, King trabajó con las abuelas argentinas de desaparecidos para establecer el Banco Nacional de Datos Genéticos en Argentina, la primera institución dedicada a preservar sistemáticamente la información genética para futuras identificaciones.
Asimismo, también ha realizado contribuciones clave a la comprensión de la esquizofrenia, proporcionando evidencias de que ese trastorno surge de mutaciones de novo — cambio en la secuencia de ADN de un gen que se observa por primera vez en una persona y que no ha aparecido en generaciones anteriores — que dañan los genes que regulan la neurogénesis en la corteza prefrontal fetal. Entre los múltiples premios que ha recibido destacan la Medalla de la Universidad de California (2004), el Mujeres y Ciencia del Instituto Weizmann de Israel (2006), el Lasker-Koshland de Investigación Médica (EE. UU., 2014), la Medalla Nacional de Ciencias de Estados Unidos (2016), el Shaw en Ciencias de la Vida y Medicina (Hong Kong, 2018), la Medalla Mendel de la Sociedad de Genética de Reino Unido (2018), el Premio Internacional Gairdner de Canadá (2021) y la Public Welfare Medal de la Academia Nacional de Ciencias de Estados Unidos (2025). La genetista experta en cáncer MaryClare King, Premio Princesa de Asturias 2025 en Investigación | ConSalud
CARTA DE SOCIO A REVISTA SEBBM
«SOBRE LA CIENCIA Y LA POLÍTICA CIENTÍFICA»
Alonso Rodríguez Navarro Profesor emérito, Universidad Politécnica de Madrid
En el número 225 de junio de la Revista SEBBM, el artículo sobre Política Científica (p. 40) contiene un juicio inadecuado sobre la ciencia en España basado en el número de publicaciones, un método tradicional en el nivel político, pero que contradice el consenso académico. El resumen del artículo comienza con la frase: «El sistema científico español ha logrado consolidarse como uno de los más productivos a nivel global». Esta conclusión es la primera premisa de la versión española de la paradoja europea, que se origina al confundir el número de publicaciones con el éxito científico: no es lo mismo producir publicaciones que producir avances científicos. En mi artículo en la Revista SEBBM del mes de marzo1, bajo el epígrafe «La falsa paradoja europea y su adopción en España», explico los errores de la Comisión Europea (CE) al juzgar la ciencia y el origen del eslogan «Europe is a global scientific powerhouse», conceptualmente reproducido para España en el artículo que comento. Incluso en el nivel político resulta difícil mantener que España tiene una ciencia competitiva a nivel global. Cada dos años, la CE hace un informe titulado Science, Research and Innovation Performance of the EU en los que analiza la producción científica de los países de la UE. Desde 2018, la proporción de las publicaciones españolas que están en el 1 % de los artículos más citados en el mundo se sitúa un 20–25 % por debajo de
la media global (la Figura 1 del artículo de marzo1 reproduce parcialmente uno de estos informes), lo que lleva a concluir que la investigación en España no tiene una posición global relevante. La situación es aún peor si se analizan las publicaciones disruptivas2, que son las que corresponden a las innovaciones radicales. El supuesto éxito científico que conduce a la paradoja europea ha llevado a la CE a adoptar «Wrong diagnoses and misguided
policies»3 y al estancamiento científico de la UE4; en España, la Ley de la Ciencia de 2022 reproduce los errores de la CE5. El Informe Draghi cambia el mensaje y advierte que la investigación en la UE está «insufficiently focused on breakthrough innovation»6. Esta es la valoración del mundo académico tanto para la UE como para España2,4 y las valoraciones alternativas confunden a los políticos y conducen a retrasar urgentes medidas correctoras.
Referencias
1 Rodríguez Navarro A. El modelo de investigación en el futuro de la Unión Europea. Revista SEBBM 223 (2025) 46–51.
2 Rodríguez-Navarro A. Countries pushing the boundaries of knowledge: the USA’s dominance, China’s rise, and the EU’s stagnation. Quality & Quantity 59 (2025) 1861–1878.
3 Dosi G, Llerena P, Labini MS. The relationships between science, technologies and their industrial exploitation: An illustration through the myths and realities of the so-called ‘European Paradox’. Research Policy 35 (2006) 1450–1464, p. 1461.
4 Rodríguez-Navarro A, Brito R. Might Europe one day again be a global scientific powerhouse? Analysis of ERC publications suggests it will not be possible without changes in research policy. Quantitative Science Studies (2020) 872–893.
5 Rodríguez Navarro A. A vueltas de nuevo con la I+D. Blog Nada es Gratis, 14 de octubre 2024.
6 Draghi M. The Future of European Competitiveness. Part A: A competitiveness strategy for Europe, p. 25 (2024).
UNA BIÓLOGA
MOLECULAR ESPACIAL
Juli Peretó
Departament de Bioquímica i Biologia Molecular, Universitat de València Instituto de Biología Integrativa de Sistemas I2SysBio, CSIC-UV
Órbitas.
Apuntes de una vida en continua exploración
Sara García Alonso
Penguin Random House | Barcelona (2025) | 286 p.
Sara García Alonso estudió biotecnología en la Universidad de León y se doctoró en biología molecular del cáncer con premio extraordinario en la de Salamanca. Hoy dirige su propio grupo de investigación en el CNIO. En 2022 fue seleccionada como astronauta en la reserva de la ESA y, desde entonces, combina su investigación oncológica con los entrenamientos periódicos en el Centro Europeo de Astronautas en Colonia y con una labor permanente de estímulo del interés público por la investigación. Se ha convertido en un verdadero altavoz a favor de la ciencia, ahora materializado en Órbitas, un libro aparecido a principio de este año.
Decir que Sara es una mujer excepcional no hace justicia a la realidad. Su curiosidad insaciable, desde que era una niña, le ha llevado a explorar una infinidad de mundos, desde las bases moleculares del cáncer, pasando por una autodisciplina física estricta, hasta alcanzar algo para lo que no se había preparado pero que quizá se escondía en algún rincón de sus sueños: ser astronauta. No tuvo una adolescencia fácil y se curtió a sí misma en una búsqueda permanente de respuestas, viajando en esas órbitas que nunca se detienen «en continua exploración». Ella ha encontrado en la investigación oncológica y en la aventura como astronauta una manera de mantenerse, como dice, en «aprendizaje vitalicio» y en convertirse en la mujer que siempre quiso ser «por dentro y por fuera».
El libro se estructura en seis «órbitas», cada una acompañada de una sugerente ilustración de Sara Herranz, y nos narra la evolución de la autora desde su niñez en León hasta el proceso, largo y tremendamente complejo, de la selección de astronautas por la ESA. Hay partes muy duras de su camino en busca de una personalidad propia y muy libre. Situaciones en las que Sara vive al límite de su capacidad de resistencia o asumiendo riesgos extremos –no me atrevo a desvelar aquí los detalles de esos entrenamientos casi militares, esas experiencias con el buceo, la espeleología o el paracaidismo (añadir aquí un escalofrío).
Me quedo con los mensajes reconfortantes de la Sara actual a su yo adolescente –¿alguien se puede imaginar que esta científica tan comunicativa fue una vez una «adolescente chunga», por usar su misma terminología? Me quedo con su defensa acérrima de la ciencia como parte indisociable de la cultura
humana y con su manera de aceptar la imperfección y la incertidumbre, su flexibilidad ante lo inesperado, su atrevimiento con lo erróneo, su capacidad de acción y reacción a pesar del miedo. Me quedo, cómo no, con su pasión por la música –por cierto, el libro se acompaña de una lista de reproducción de las canciones que van jalonando las órbitas.
Tuve el privilegio de acompañar a Sara durante la presentación de su libro en la pasada Fira del Llibre de València. Fueron unos minutos que disfruté mucho conversando con ella y, además de los diferentes aspectos del libro y de su experiencia como escritora, me permití plantearle lo que me parece a mí que va a ser un tema de debate cada vez más presente. Una cuestión sobre la que los científicos, en general, y los estudiosos de la vida, en particular, deberíamos tomar posiciones. Justo cuando me llegó Órbitas estaba leyendo las últimas páginas de The decline and fall of the human empire: why our species is on the edge of extinction (Picador, 2025) de Henry Gee. Se trata de una argumentación muy bien documentada de por qué, con toda probabilidad, a la especie humana le quedan unos diez milenios de existencia. La conclusión de Gee, en la línea de otros científicos como Stephen Hawking y, lo que es peor, de los multimillonarios que se han metido en el negocio del espacio, es que la especie humana solo alargará su existencia colonizando otros planetas. Personalmente creo que es una conclusión decepcionante para un libro que, por otro lado, me fascinó.
El neocolonialismo espacial me parece una falacia evolutiva disfrazada de ciencia ficción banal y que, me temo, nos enfrenta a unos escollos éticos y legales fabulosos. Órbitas no se refiere explícitamente a la cuestión del futuro de nuestra especie fuera de la Tierra. Pero no me resistía a conocer la opinión de alguien como Sara García Alonso, una bióloga que también trabaja en fortalecer la capacidad de exploración espacial de la Unión Europea. Y no pude estar más de acuerdo con ella en su defensa de la investigación científica, el desarrollo tecnológico y las innovaciones que se derivan de esa ansia sin límites de conocimiento sobre el universo, al mismo tiempo que, con humildad, reconoció que la especie humana no tiene un planeta B al que recurrir, que lo que nos quede hasta la extinción como especie lo pasaremos aquí, sin remedio, sobre esta delicada y maltratada canica azul.
congresos.sebbm.es/caceres2025
4ª generación de Espectrofotometría UV-Vis: NanoDrop-Ultra y Electroforesis Capilar en Gel para Work ows de qPCR/RT-qPCR
ULTRA
Qsep100
Análisis de la pureza de una muestra de DNA de doble cadena y detección de contaminantes, llevado a cabo con el equipo NanoDrop-Ultra. Se detecta la presencia de un contaminante, en este caso fenol (espectro naranja) y se obtiene un valor de concentración corregida de 221,6 ng/µL.
Detección de contaminantes
Calculadora qPCR integrada
La combinación perfecta en análisis cuantitativos de concentración, pureza e integridad de las muestras.
“Cómo NanoDrop-Ultra y Qsep pueden suponer un antes y un después en tu work ow de qPCR”
El control de calidad de las muestras es un paso clave en múltiples etapas de los ujos de trabajo en Biología Molecular. La guía MIQE de ne la cantidad y calidad de ácido nucleico como una información esencial para las aplicaciones downstream
Todas las técnicas de extracción de ácidos nucleicos comparten un denominador común: añaden contaminantes a las muestras. Esta contaminación residual puede in uir drásticamente en el análisis posterior ya sea por inhibición enzimática o por sobreestimación de concentración. Si bien es cierto que los ratios de pureza de absorbancia (A260/A280 y A260/A230), son útiles, no aportan toda la información de la pureza de la muestra.
La 4ª Generación de Espectrofotometría UV-VIS, NanoDrop-Ultra va más allá de las mediciones. Permite obtener resultados robustos y reproducibles gracias a su software integrado, Acclaro, de identi cación de contaminantes proporcionando a los usuarios no sólo información sobre la calidad y carga de contaminantes, sino los valores reales de concentración de su muestra. Además, esta nueva generación, incorpora una calculadora qPCR integrada para un ujo de trabajo e ciente.
Resulta esencial tener un control de calidad atendiendo a la pureza, cantidad y calidad de las muestras:
Pureza: impurezas como fenol y sales inter eren en la sensibilidad y e ciencia de las reacciones enzimáticas, además de afectar a los valores de concentración.
Cantidad/concentración: una estimación errónea de DNA/RNA puede alterar las interacciones muestra-enzima y dar lugar a workflows fallidos.
Integridad: una alta proporción de fragmentos degradados in uye signi cativamente, en los valores de concentración de las muestras, y en etapas posteriores del protocolo.
Los bioanalizadores de fragmentos Qsep1 y Qsep100 (Bioptic Inc.), basados en electroforesis capilar en gel combinada con detección por uorescencia, y los espectrofotómetros UV-Vis de microgota NanoDrop-Ultra, representan equipos de alta calidad que, combinados, proveen una solución simple y efectiva para determinar pureza, concentración e integridad del DNA/RNA y asegurar el éxito de cualquier ujo de trabajo de Biología Molecular.
A continuación, se recogen algunos ejemplos de control de calidad de muestras involucradas en pasos clave de workflows moleculares, cuyos análisis se han llevado a cabo con los equipos NanoDrop-Ultra y Qsep100.
Análisis de pureza de las muestras y detección de contaminantes
El software Acclaro detecta la presencia de contaminantes en las muestras de ácidos nucleicos y proporciona información acerca de su identidad, su contribución en la absorbancia de la muestra y los valores de concentración originales (con contaminante) y corregidos (sin contaminante). Entre los contaminantes que puede predecir el equipo se encuentran fenol, sales de guanidina, proteínas y RNA. Además, la calculadora de qPCR integrada permite saber rápidamente la cantidad precisa de muestra necesaria para las reacciones de qPCR posteriores.
Análisis de la integridad del DNA genómico (gDNA)
Con los cartuchos de kilobases y de alta sensibilidad, es muy sencillo detectar qué muestras presentan un gDNA intacto y cuáles presentan degradación. Q-Analyzer, además, facilita el índice de calidad del DNA (DQN), que permite determinar su nivel de fragmentación (Fig. A) También genera información relativa al tamaño de los fragmentos y sus valores de concentración, entre otros.
Control de calidad del RNA total
Q-Analyzer asigna automáticamente las regiones correspondientes a las subunidades 18S/28S para facilitar el análisis. Además, proporciona el índice de calidad de RNA (RQN), que permite seleccionar las muestras de mayor calidad (Fig. B).
Fragmentación y validación de librerías
La proporción de cada uno de los fragmentos de DNA y su concentración son dos factores determinantes para obtener librerías de calidad. Los bioanalizadores Qsep, permiten realizar robustos análisis cualitativos rápidamente y generar resultados como los de la Fig. C, recogiendo los valores de DQN, tamaño y concentración promedio y una tabla resumen de cada fragmento.
Resultados obtenidos tras el análisis de muestras de DNA/RNA destinadas a NGS, empleando el equipo Qsep100 (Bioptic Inc.).
A) Análisis de la integridad de gDNA. Electroferograma y tabla de resultados: tamaño y concentración promedio y DQN = 8,5 (muy buena calidad). B) Control de calidad del RNA total. Electroferograma y tabla de resultados: ratio 28S/18S,RQN = 9,84 y DV200 = 98,3% (muy buena calidad). C) Fragmentación y validación de una librería de DNA. Electroferograma, gel y tablas resumen.
Resumen
El control de calidad de las muestras es paso crítico en workflows moleculares. NanoDrop-Ultra y Qsep1/Qsep100 son dos plataformas óptimas que permiten llevar a cabo el QC de las muestras de DNA/RNA, contribuyendo así mismo al ahorro de tiempo y recursos de sus usuarios al identi car de primera mano las muestras ideales para el protocolo de Biología Molecular.
Con más de 3.000 socios y 19 grupos científicos, la SEBBM es la principal agrupación científica en su campo.
Contribuimos a que la Bioquímica y la Biología Molecular tengan un papel relevante en nuestra sociedad, favoreciendo el contacto entre investigadores e impulsando a los jóvenes científicos.
Pero no estamos solos...
Los socios protectores contribuyen al progreso de la SEBBM
Porque son de los nuestros*
Más información sobre la figura de socio protector en: sebbm@sebbm.es o llamando al +34 681 916 770
*Serán socios protectores aquella entidades que quieran contribuir al sostenimiento y desarrollo de la SEBBM y sean aceptadas como tales. Tendrán derecho a voto en las asambleas, pero no podrán ser elegibles para cargos directivos.