sampling-free measurement of glucose and lactate in culture medium. Visualize real-time changes in cell metabolism. The PHCbi Live Cell Metabolic Analyzer will open the door to new discoveries.
• In-Line sensors: the key to continuous measurements of glucose and lactate
• Measure cells in their usual culture environment
• Evaluate glycolytic changes directly
IncuSafe CO2 Incubators offer the most precise and regulated environment. The Direct Heat System achieves accurate, uniform, and highly responsive temperature control within the chamber. The high quality Dual IR Sensor regulates the CO2 levels, providing exceptional uniformity. During cell culturing, the InCu-saFe Germicidal Interior and SafeCell UV Lamp continuously prevent contamination.
• Precise & Regulated Environment
• Increasing work efficiency
• Time-Saving Decontamination
MCO-170 AIC-PE, CO2 Incubator www.phchd.com/eu/biomedical
Los transposones, componentes esenciales y motores de la evolución de los genomas eucariotas
Josep M. Casacuberta y Elena Casacuberta
Los elementos móviles: ADN repetitivo relevante para la función, estructura y evolución de los genomas
Josefa González
Elementos transponibles en humanos y su impacto en el desarrollo de enfermedades
Layla Díaz Portal y Bernardo Rodríguez Martín
El Origami de la cromatina: organización estructural y regulación
génica
Jorge Mañes García y Leonardo Beccari
ENTREVISTA
Jordi Camí
Presidente del Comité Español de Ética de la Investigación
Ismael Gaona Pérez
POLÍTICA CIENTÍFICA
Ciencia, ética y estética: el equilibrio necesario en la investigación
Ismael Gaona Pérez
A FONDO
El modelo de investigación en el futuro de la Unión Europea
Alonso Rodríguez Navarro
EDUCACIÓN UNIVERSITARIA
Es hora de llevar al aula la predicción del plegamiento de proteínas
Ángel Herráez
Cinética hiperbólica y cinética sigmoide: la forma de la curva importa, ¡y mucho!
Vicente Rubio, Félix Goñi, Carlos Gancedo
INVESTIGACIÓN
Joaquim Ros
IN MEMORIAM
Antonio Ballesteros Olmo (1940-2025)
Joan Guinovart (1947-2025)
Federico Mayor Zaragoza (1934-2024)
Eugenio Vilanova Gisbert (1948-2024)
SOCIEDAD
Noticias de actualidad de la SEBBM
RESEÑA DE LIBROS
Plantas que nos ayudan Rosa Porcel
PUBLIRREPORTAJES
Antonio Ferrer
Presidente
SEBBM
La semilla, transformación y presente de la SEBBM
Comienzo esta tribuna recordando a nuestros compañeros, y expresidentes de la SEBBM, Federico Mayor Zaragoza y Joan J. Guinovart Cirera que, tristemente, nos dejaron hace apenas unas semanas. Federico y Joan han sido promotores y pilares esenciales de la SEBBM, que trabajaron intensamente para que fuera una sociedad científica relevante e influyente a nivel nacional y a nivel internacional. Sin duda, un hito que se ha conseguido y del que hemos de estar altamente agradecidos a los dos. Federico contribuyó a la constitución de la Sociedad Española de Bioquímica (SEB), participando primero en la reunión de Bioquímica de 1961 celebrada en Santander y, posteriormente en la II reunión de Bioquímicos Españoles de 1963 celebrada en Santiago de Compostela y que culminó con la fundación de la SEB. Implicado en la SEB desde su creación, fue su presidente en el periodo 1972-1976, contribuyendo a su desarrollo e implantación nacional e internacional. En 2018, Federico fue nombrado Presidente de Honor por su incuestionable compromiso, apoyo y trabajo con y por la SEBBM.
Joan fue un embajador muy activo de la SEBBM que potenció la influencia de la SEBBM a nivel global. Con su carisma, su presidencia (1996-2000) promovió un impresionante impulso de la SEBBM coronado, además, por su magnífica labor como editor de la Revista SEBBM a la que proporcionó un nuevo diseño, tanto en contenido como en continente, altamente valorados y apreciados. Joan fue el motor de diversos programas científicos, formativos y divulgativos que incrementaron el impacto social de la SEBBM. No menos destacable, Joan fue el promotor en la creación de la Fundación SEBBM cuya motivación principal fue su deseo de contribuir a potenciar las acciones y actividades de la SEBBM a través de una donación. Fue precisamente esta voluntad de Joan la que motivó a la junta directiva a contemplar la creación de una entidad de interés público, que se materializó con la constitución de la Fundación SEBBM cuyo objetivo es canalizar las donaciones de particulares e instituciones en programas sociales, ofreciendo deducciones fiscales a los donantes. Como inspirador de la Fundación, Joan ha sido su primer donante. Todo un ejemplo de socio amante de la SEBBM, comprometido con su futuro y con el destino de la bioquímica y la biología molecular española. Todo nuestro afectuoso agradecimiento por este magnífico gesto que confío sea un modelo
que imitar para potenciar los programas sociales y educativos de la Fundación y aumentar el interés e influencia de la SEBBM.
Sin duda, Federico y Joan han dejado una impronta en la SEBBM que debemos cuidar y mantener como un ejemplo, y trasmitirla a las nuevas generaciones como modelos de bioquímicos comprometidos con el desarrollo de esta disciplina a nivel nacional, y en su proyección internacional. Han sido excelentes embajadores de ello, y les tenemos que reconocer y agradecer su contribución al espectacular avance de la SEBBM. ¡Gracias de corazón a los dos!
Y un evento que ambos potenciaron durante sus periodos de presidencia fue el congreso anual como punto de encuentro de los socios, y como evento para fomentar la incorporación de nuevos socios. El congreso anual continúa siendo el acontecimiento más relevante de la SEBBM que cada año organiza a lo largo de la geografía española. En 2025, el congreso se celebrará en el Complejo Cultural San Francisco en Cáceres, un antiguo monasterio franciscano fundado en el siglo XV por fray Pedro Ferrer. Una maravilla de sede en la que disfrutar de la ciencia, la cultura y la gastronomía. Una preocupación que puede surgir con Cáceres es su limitada accesibilidad en comparación con otras ciudades que gozan de mejores vías de comunicación. Los organizadores y la SEBBM conscientes de ello, van a poner autobuses en Madrid que faciliten el traslado de los congresistas a Cáceres. Con ello esperamos aumentar las alternativas de comunicación, y que llegar a la ciudad extremeña no sea un obstáculo para no venir a disfrutar del congreso.
Un reto que presenta el Complejo Cultural San Francisco es el número de salas que limita la cantidad de grupos que se pueden reunir simultáneamente. Conscientes de la relevancia de las reuniones de grupo, la organización del congreso ha remodelado la agenda del congreso para asegurar que todos los grupos SEBBM puedan reunirse, sin necesidad de fusionar reuniones. Para ello, las reuniones se han repartido a lo largo de los cuatro días que dura el congreso. Además, esta distribución extendida permite una mejor disposición temporal de los grupos, minimizando la programación simultanea de grupos afines y promocionando la participación cruzada entre sus miembros, así como una mayor asistencia a estas sesiones. Estoy convencido de que esta organización contribuirá a promover y fortalecer la interacción de los grupos SEBBM, aumentando el interés y atractivo por el congreso. Además, permitirá contener el coste cuando haya que recurrir a usar palacios de congresos como sede.
Otro desafío que impone el Complejo Cultural San Francisco es la localización de los expositores junto con los pósteres y el cáterin. De nuevo, se han considerado soluciones que acomodan todas las necesidades de forma adecuada, aunque con algún mínimo sacrificio espacial que espero sea aceptable y aceptado por todos. Aprovecho para insistir en la importancia de visitar los pósteres y discutir con los jóvenes autores que los presentan, así como interaccionar con los expositores, tanto en sus presentaciones como en los estand en que presenten sus productos. Ambos jóvenes y expositores son pilares fundamentales para el presente y futuro de la SEBBM.
He limitado esta tribuna a comentar algunos retos que nos presenta la sede en Cáceres, pero quiero recalcar que son trastornos menores superados ampliamente por el excelente programa científico y la historia, cultura y hospitalidad de la ciudad. Mayor desafío es competir con la cantidad de congresos afines que intentan atraernos. Confío que el gran trabajo realizado por Guadalupe Sabio y su equipo ofreciendo un magnífico programa científico y una impresionante sede sean motivos para escoger participar en el 47º Congreso de la SEBBM del 2 al 5 de septiembre de 2025 en Cáceres. ¡Os esperamos!
Pensar la creatividad
Inmaculada
Yruela Guerrero
Directora Revista
SEBBM
La creatividad no es exclusiva de la expresión artística en sus diferentes facetas – artes audiovisuales, escénicas, gráficas, literarias, plásticas…- sino que también es propia de la actividad científica. El trabajo científico requiere creatividad para explorar nuevos caminos que nos lleven a imaginar las soluciones que resuelvan los problemas que se plantean. Federico Mayor Zaragoza, recientemente fallecido, escribía en un artículo publicado en la revista Limbo en 2018 –
“Si no existiera un ser dotado de estas facultades inusitadas y distintivas – reflexionar, imaginar, anticiparse, innovar, ¡crear! – si no hubiera este «sens de la vie», la inmensidad y belleza del universo, la indescriptible perfección de lo colosal y de lo ínfimo, serían percibidas pero no conocidas y admiradas. […] ¡Saber que sabe!: esta conciencia refleja estar dotado no sólo para describir sino para imaginar, para innovar, constituye el fundamento del «sentido» de la vida humana.”
(Limbo, 38, 2018, p. 41).
Para imaginar e innovar hay que tener tiempo para pensar y reflexionar, además de tener profundos conocimientos. En algunos ámbitos educativos no se da el suficiente valor a esto último. Se piensa que la adquisición de conocimientos ya no es tan importante porque todo está accesible en internet. Se defiende que lo importante es aprender a resolver problemas, desarrollar el pensamiento crítico y fomentar la creatividad. Pero se olvida que el tener, o no, estas habilidades depende de los conocimientos adquiridos. Sin conocimiento no puede haber creatividad.
¿Se dan las circunstancias favorables para la creatividad en la actividad científica? El artículo “Scientists need more time to think” publicado el 25 de julio de 2024 en la revista Nature destaca la idea de que el tiempo invertido para pensar suele estar infravalorado. Nos evalúan por la productividad que se traduce en materiales medibles –conferencias, informes, patentes, proyectos, publicaciones, trabajos dirigidos, talleres, etc.- pero el tiempo dedicado a pensar y reflexionar se escapa de las métricas. ¿Es posible valorar el tiempo dedicado a las tareas reflexivas y de creación en la actividad científica?
Un intento de valorar aspectos cualitativos del trabajo científico se ha puesto en marcha con la iniciativa europea CoARA (Coalition for Advancing Research Assessment), impulsada por la Europe Science y European Universities Association (EUA), que cuenta con el apoyo de la Comisión Europea. España es el país con más universidades y organismos de investigación adheridos a esta coalición, y desde 2022 se trabaja en una propuesta de evaluación flexible y multidimensional que proporcione herramientas útiles y metodología viable para este ambicioso cambio. La tarea no es fácil. Me pregunto si hay cultura en la comunidad científica para integrar esta reforma de la evaluación tan necesaria. Si no cambiamos de manera responsable los sesgos adquiridos se quedará en “papel mojado”.
Habremos perdido una oportunidad.
El artículo de Nature, anteriormente referido, también señala que la ciencia es cada vez menos disruptiva, a pesar de que se publican más artículos y se invierte más en ciencia. La era digital y la Inteligencia artificial nos proporciona valiosas herramientas para desarrollar el trabajo científico, nos da unas ventajas indiscutibles; pero surgen voces de alarma sobre si el uso de los dispositivos digitales y sus aplicaciones restan tiempo para concentrarnos y pensar. ¿Limitan la creatividad en el trabajo científico? El neurocientífico
Ardem Patapoutian, premio Nobel de Fisiología o Medicina en 2021, en sus charlas suele abogar por la mejora del mundo académico y de los investigadores; y con este propósito ha publicado 13 reglas sobre cómo hacer ciencia. En la lista destaca: (1) “Don’t be too busy” -No excuses. If you’re too busy, you’re not being creative-“; (2) “Learn to say no”. El tiempo necesario para concentrarse sin interrupciones siempre ha sido fundamental para la investigación. Las listas de correos interminables, el goteo e inmediatez de los anuncios de convocatorias, las tareas burocráticas que se solapan… son algunos de los obstáculos que nos abruman y tenemos que esquivar. ¿Cómo elegir lo prioritario para no perder la capacidad de reflexionar y pensar? ¿Cuál es el impacto de la pérdida de tiempo en la ciencia, no sólo en la estructura sino también en el contenido y en la calidad de la investigación? Otro aspecto no menor es la presión por publicar para conseguir una óptima evaluación en las acreditaciones, evaluación del currículum, solicitud de proyectos, etc.
Sería conveniente que nuestras instituciones, tanto académicas como científicas, reflexionaran sobre ello y disminuyeran el ruido que ocasiona la avalancha de comunicaciones, que en muchas ocasiones es prescindible o se puede minorar. Los anuncios y las comunicaciones institucionales se han multiplicado por cinco o por diez en los últimos años, en gran medida por un ejercicio de transparencia y de estar presente. Pero debería evaluarse, por contrapartida, el ruido que generan y el efecto negativo que producen. Es necesario realizar estudios sobre el efecto de estas prácticas en el trabajo científico, así como estudios sobre cómo proteger el tiempo para pensar y reflexionar en un mundo de comunicación instantánea. Deberíamos reivindicar una “ciencia lenta”, con más tiempo para equivocarnos y no aspirar continuamente al éxito.
Es necesario crear una cultura de investigación más favorable. Proteger la creatividad y, como nos recordaba Federico Mayor Zaragoza, proteger el «sens de la vie», ¡Saber que sabe!
No puedo cerrar estas líneas sin recordar a Joan J. Guinovart, ex-editor jefe y editor honorario de la Revista SEBBM desde 2013, que nos dejó al comenzar el año.
EL GENOMA NO CODIFICANTE
Crisanto Gutiérrez
Director del Programa de Dinámica y función del genoma
Centro de Biología Molecular ‘Severo Ochoa’ (CBM), CSIC-UAM, Madrid
https://doi.org/10.18567/sebbmrev_223.202503.dc1
Toda la información necesaria para la vida de los organismos está almacenada en su genoma. Cuando se habla sobre su importancia frecuentemente se piensa solamente en las regiones del genoma que codifican proteínas como las más relevantes desde el punto de vista funcional y de organización. Sin embargo, los datos de secuenciación de cientos de genomas han revelado que la fracción del genoma ocupado por secuencias codificantes es en realidad minoritaria. Es de resaltar que tras demostrarse la enorme cantidad del genoma ocupado por secuencias repetidas no codificantes se pasó a considerar estas regiones como “ADN basura”. Si bien se puede encontrar ~75% de
regiones del genoma humano que se transcriben, solamente un 2% se traducen a proteínas mientras que el resto corresponde a ARNs no codificantes (ncRNAs, por sus siglas en inglés). De hecho, la presencia de grandes cantidades de ADN no codificante, que pueda transcribirse o no, es una característica de los genomas de todas las especies estudiadas, incluidos hongos, plantas y animales. Descifrar la relevancia estructural y funcional de dichas regiones no ha sido, ni está siendo, una tarea fácil no solo debido a la cantidad sino también a la diversidad de elementos genómicos incluidos en esta categoría. Dado el potencial y retos a los que se enfrenta este ámbito de
investigación, el Consejo Superior de Investigaciones Científicas (CSIC) promovió la redacción de una monografía sobe Genoma y epigenoma, uno de cuyos capítulos versó sobre el genoma no codificante. El desarrollo de estrategias experimentales avanzadas está permitiendo aumentar enormemente nuestra capacidad de estudio y de entender su relevancia. El genoma no codificante se compone de diversos elementos que globalmente pertenecen a dos grandes grupos: las secuencias repetidas y las secuencias reguladoras. Las regiones genómicas que contienen secuencias repetidas incluyen no solo las regiones teloméricas y centroméricas sino también los transposones o elementos móviles del genoma, cuya existencia fue propuesta por Barbara McClintock en las décadas de 1940 y 1950, y cuya excepcional y pionera investigación mereció la concesión del Premio Nobel en 1983. El resto del genoma no codificante con funciones reguladoras está constituido por intrones y regiones intergénicas que se transcriben para producir ARNs en los enhancers (eRNAs, por sus siglas en inglés), ARNs no codificantes largos (long ncRNAs, por sus siglas en inglés), microARNs (miRNAs, por sus siglas en inglés) o ARNs circulares (cirRNAs, por sus siglas en inglés). En el presente dosier se incluyen cuatro artículos que tratan sobre la organización, función y relevancia de los transposones y de los enhancers para darnos una visión de estos elementos genómicos.
El Dr. Josep Casacuberta y la Dra. Elena Casacuberta nos describen los transposones, su organización estructural y relevancia en la respuesta a estrés, y la producción de mutaciones tanto en plantas como en animales. Conocer el contexto histórico de la investigación sobre transposones es también fundamental para
entender la controversia inicial sobre su existencia así como la relevancia funcional en la dinámica de los genomas que con los años han llevado a definirlos como elementos cruciales en la organización y función de los genomas eucarióticos. Un aspecto fundamental es la capacidad de los transposones de poder modificar el desarrollo de los organismos, en particular de las plantas, para adaptar su crecimiento a las cambiantes condiciones ambientales.
La Dra. Josefa González abunda en la discusión de dos aspectos muy relevantes de la biología de los transposones, derivados de su capacidad de movilizarse de una región del genoma a otra. Por un lado, su función como reguladores de la expresión génica al insertarse en zonas próximas al inicio de la transcripción de algunos genes, y poseer promotores y secuencias de unión de factores de transcripción. Por otro, la movilidad de los transposones contribuye a la generación de nuevos transcritos que pueden modificar la adaptabilidad y la evolución de las especies.
El Dr. Bernardo RodríguezMartín y Layla Díaz-Portal, estudiante de su laboratorio, nos iluminan sobre las consecuencias de la transposición de los elementos móviles en salud humana. Su artículo nos ilustra sobre las consecuencias de las inserciones de
transposones en la línea germinal así como su implicación en enfermedades como el cáncer.
Por último, El Dr. Leonardo Beccari, junto con su estudiante Jorge Mañes García, enfoca su contribución a otros elementos del genoma no codificante fundamentales en la regulación de la expresión génica, los enhancers y cómo su función depende de la organización estructural del genoma dentro del núcleo eucariótico en dominios topológicamente asociados (TADs, por sus siglas en inglés).
Estos cuatro artículos son solamente una selección de temas para empezar a entender la situación actual de la investigación del genoma no codificante. Una descripción exhaustiva de todos los temas relevantes requeriría más artículos para tratar la diversidad funcional de los ARNs no codificantes que pueden servir de base para otro dosier de esta revista. Lo que queda patente es la necesidad de dedicar mucha atención al antes conocido como “ADN basura” ya que, muy al contrario, se trata de un componente del genoma muy diverso y con funciones cruciales en el control de la expresión génica, en la organización del genoma, tanto en su integridad como en su dinamismo, con enormes implicaciones en el desarrollo de los organismos, su adaptabilidad y evolución.
Para leer más
Gutiérrez C y Hernández-Munaín C. The non-coding genome, Chapter 4, pp. 79-100. En: CSIC Scientific Challenges: Towards 2030; Vol 3: Genome and epigenetics. Montoliu L y Rada-Iglesias A, eds. (2021). e-ISBN: 978-84-00-10738-3e.
Fox Keller E. A feeling for the organism. The life and work of Barbara McClintock. Henry Holt & Company, New York (1983). ISBN: 0-8050-7458-9.
LOS TRANSPOSONES, COMPONENTES ESENCIALES Y MOTORES DE LA EVOLUCIÓN DE LOS GENOMAS EUCARIOTAS
Josep M. Casacuberta
Centro de Investigación en Agrigenómica (CRAG), CSIC-IRTA-UAB-UB, Cerdanyola del Vallès, Barcelona
Elena Casacuberta
Instituto de Biología Evolutiva (IBE), CSIC-UPF, Barcelona
https://doi.org/10.18567/sebbmrev_223.202503.dc2
Evolución genómica. Últimos 20 años
En estos últimos 25 años los laboratorios de genética o biología molecular se han transformado radicalmente y muchas poyatas de laboratorio se han convertido en mesas con ordenadores conectados a internet. Las comparaciones de miles de secuencias para buscar mutaciones en un gen concreto, la búsqueda de posibles elementos reguladores comparando centenares de genomas secuenciados o la predicción de la estructura de proteínas que nunca han sido purificadas se han
convertido en rutina. Y obtenemos respuestas casi instantáneas a preguntas que serían impensables hace pocos años. Hace menos de 25 años, en el año 2000, se publicó el primer genoma completo de una planta, Arabidopsis thaliana, y un año después se publicó el primer borrador del genoma humano. Y supuso una revolución. A muchos les costará de creer, pero poco antes de la publicación del genoma humano había apuestas en muchos laboratorios sobre el número de genes que finalmente tendría ¿40.000, 50.000, o quizás 80 o 100.000 genes? Era de
esperar que los humanos tuviéramos muchos más genes que una mosca o que una planta, porque además de ser organismos más complejos, tenemos un genoma más grande. Y sin embargo… La publicación del genoma humano nos deparó auténticas sorpresas. La primera, que el número de genes anotados no superaba los 30.000, un número que dos años más tarde se rebajó a unos 20.000; y la segunda, que estos genes sólo ocupan una pequeñísima parte del genoma. Por ello, El País, dando a conocer estos resultados, publicó un artículo en
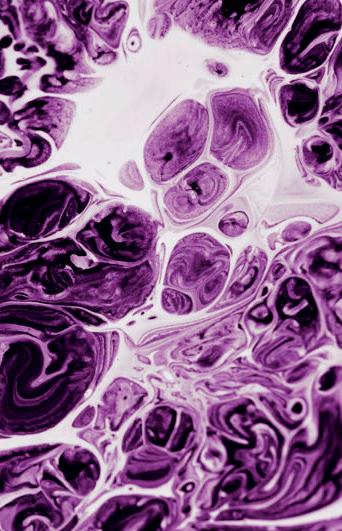
Figura 1
Barbara McClintock (1902-1992), Department of Genetics, Carnegie Institution at Cold Spring Harbor, New York, trabajando en su laboratorio (fuente Smithsonian Institution, http://commons.wikimedia.org/).
febrero de 2001 con el expresivo título de “El 95% del genoma humano es basura”. Esa basura genómica está formada, en su mayor parte, por transposones. El papel de los transposones ha sido siempre controvertido desde que Barbara McClintock propusiera su existencia en los años 50 del siglo pasado, y han sido considerados durante mucho tiempo como basura genómica. Sin embargo, de forma paulatina se han ido acumulado evidencias de su importancia en la evolución de los genomas y en la regulación génica. Sólo 10 años después del
artículo mencionado, en septiembre de 2012, el mismo periódico El País publicaba un artículo titulado “La basura genómica ya es medicina”, donde se explicaba que muchos de los elementos reguladores humanos se encuentran en secuencias de elementos móviles. En este artículo proponemos un breve viaje en la historia para entender hasta qué punto las ideas de McClintock fueron revolucionarias en su tiempo y como fueron confirmadas con el avance de la genómica, para después hacer un resumen de lo que conocemos actualmente sobre el
impacto de los transposones en los genomas eucariotas.
Los transposones, una idea visionaria de Barbara McClintock
Barbara McClintock (Figura 1) desarrolló con gran habilidad la citogenética de maíz a finales de los años 40 del siglo pasado. McClintock recopiló datos de numerosos cruces genéticos y observó detalladamente los diferentes caracteres de los granos de las mazorcas y sus coloraciones en las plantas hijas. En estos estudios encontró las primeras pruebas físicas de la recombinación
génica, cuya existencia habían sugerido Sturtevant y Morgan en los años veinte, además de las primeras pruebas de la transposición. McClintock descubrió que en los cromosomas de maíz había determinantes genéticos (el concepto de gen era aún muy debatido) que producían roturas en los cromosomas, de aquí el nombre Disociador, (Dissociator, Ds por sus siglas en inglés) y vio que éstos podían cambiar posiciones en el genoma. También encontró que había otros determinantes genéticos que parecían influenciar (controlar) el comportamiento de Ds, a los que llamó Activadores (Activators, Ac por sus siglas en inglés). Estos estudios dieron lugar a dos ideas importantes: (1) los determinantes genéticos podían moverse de sitio en los cromosomas (transposición); (2) estos determinantes controlaban la expresión génica.
En 1951, en el congreso anual de Cold Spring Harbour, Barbara McClintock presentó por primera vez las evidencias experimentales de la existencia de los transposones. En su exposición McClintock relacionó también este descubrimiento con la idea del control génico de la expresión al bautizar los elementos móviles como elementos controladores “controlling elements” del desarrollo de la planta. Es difícil hacerse una idea del salto conceptual que significaba el descubrimiento de la transposición hoy en día. Que el ADN era la molécula encargada de transmitir la herencia se demostró un año después (1952, Hersey y Chase) y la estructura de la molécula del ADN dos años más tarde (1953, Watson y Crick). Tuvieron que pasar aún diez años, hasta 1961, para que se asociara por primera vez un triplete de bases a un codón, y cinco más, hasta 1966, para descifrar todo el código genético. Todos estos descubrimientos fueron muy rápidos y
Figura 2
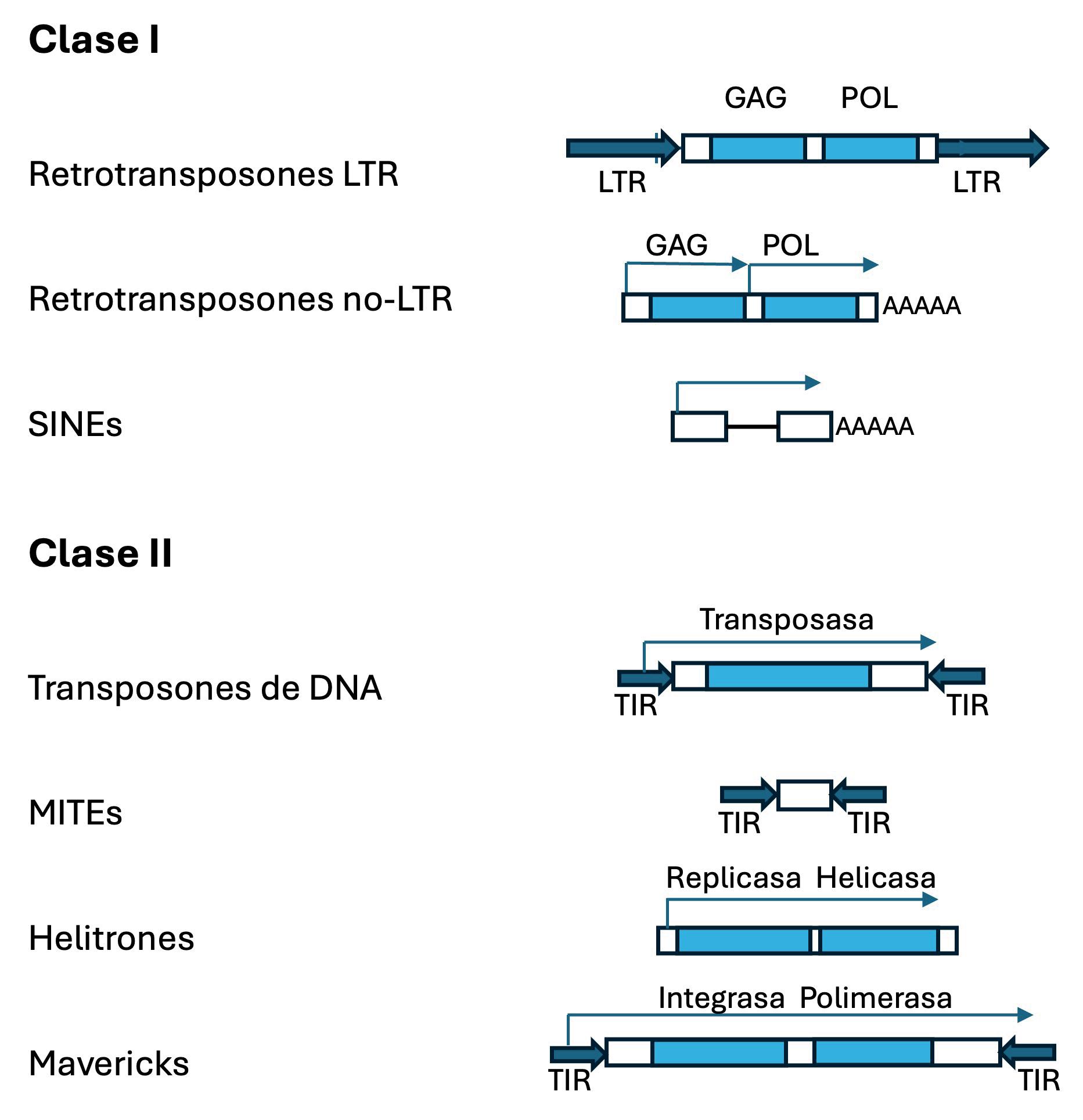
Clasificación general de los Transposones: clases y superfamílias principales. Las cajas azules representan las regiones codificantes para las distintas actividades necesarias para la transposición. Las flechas sólidas indican las repeticiones terminales directas (Long Terminal Repeat, LTR por sus siglas en inglés), o invertidas (Terminal Inverted Repeat, TIR por sus siglas en inglés). Las flechas finas indican las unidades transcripcionales.
pedían tiempo para su asimilación. Incluso el concepto de gen estaba en debate. Probablemente añadir la idea de movilidad dentro del ADN creaba demasiada confusión para que la transposición pudiera ser rápidamente aceptada.
Durante las siguientes décadas, se descubrieron nuevos elementos transponibles en maíz (en 1955 ya se habían identificado, Spm, Dotted, Modulator y Enhancer), en Drosophila (1965), en fagos (1972), bacterias (1966) y finalmente en levadura (1979) demostrando que los transposones eran parte intrínseca de los
genomas procariotas y eucariotas, y mostrando la gran variedad estructural de estos elementos (Figura 2) que se corresponde con distintos mecanismos de transposición (Figura 3). En 1983, Barbara McClintock recibió el Premio Nobel de Fisiología o Medicina por el descubrimiento de los elementos móviles.
Aunque fue muy polémica su definición de los elementos móviles como elementos controladores, cuando Jacob y Monod presentaron en 1960 su modelo del operón en E. coli, McClintock saltó de alegría al ver los paralelismos
Figura 3
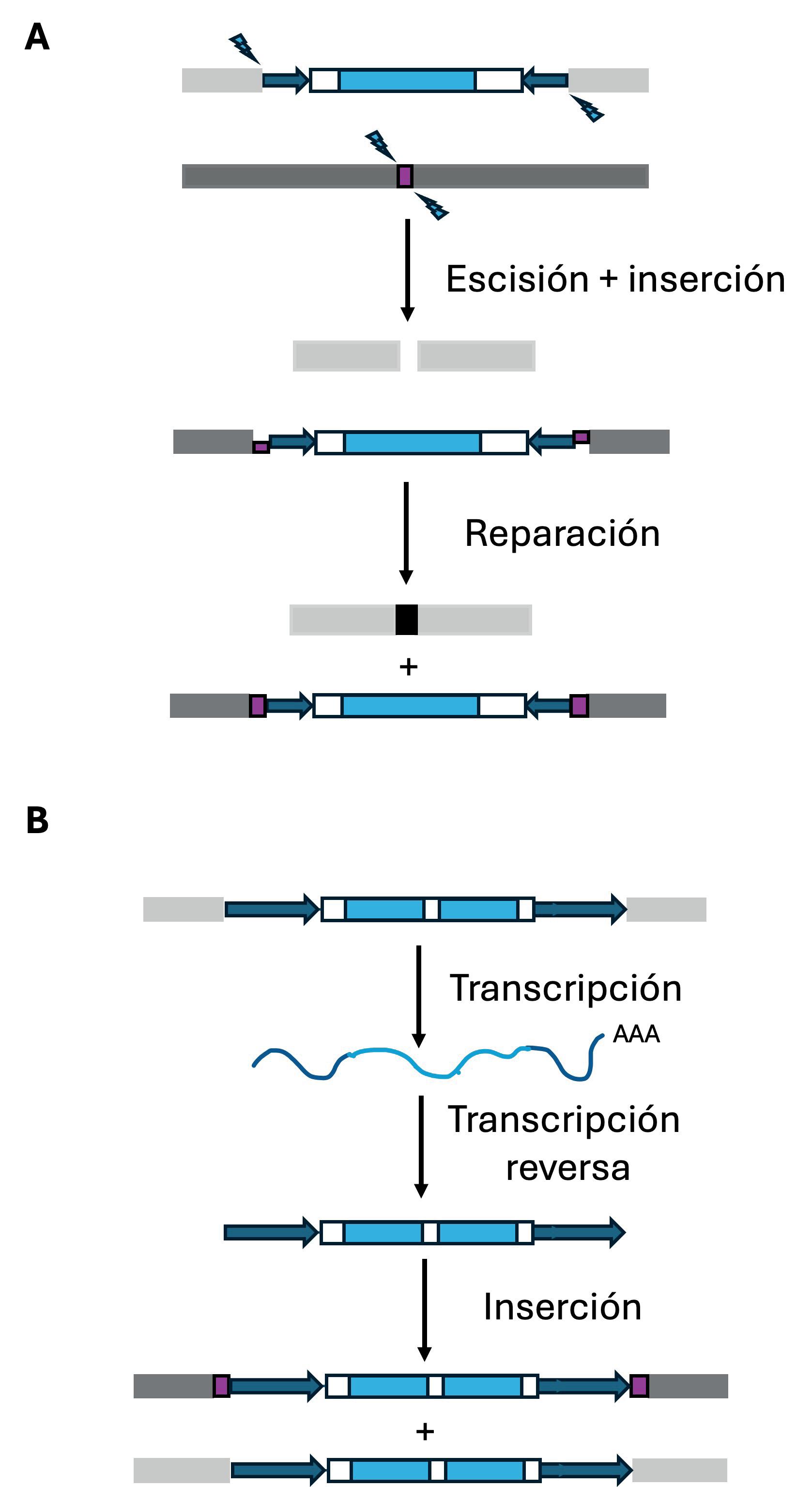
Principales mecanismos de transposición. Los transposones se representan siguiendo el mismo código que en la Figura 2. (A) Transposición de los elementos móviles de clase II, o transposones de ADN. Mecanismo conocido como “cortar y pegar”. Los sitios de corte de la transposasa en el locus dador (cajas gris claro) y aceptor (cajas gris oscuro) se muestran con flechas azules. El sitio de inserción (target site) que será duplicado a ambos lados de la inserción se muestra como una caja violeta. (B) Transposición de los elementos móviles de clase I, o retrotransposones. Mecanismo conocido como “copiar y pegar”. El sitio de inserción, que se repara como en el caso de los transposones de clase II y se verá duplicado como resultado de la inserción, se muestra como una caja violeta.
con su idea de “elementos controladores”. La idea de que existían los operones en bacterias y los elementos controladores en maíz, demostraba, según McClintock, que este sistema de regulación era extensible a todo organismo vivo. Es más, para McClintock, el control sobre la expresión génica debía de estar conectado con una causa celular (algo en el citoplasma), o en el ambiente en el que se desarrolla el organismo. Poder responder frente a cambios en el ambiente (estrés) regulando la expresión génica permite a los organismos hacer frente a estos cambios, adaptarse, evolucionar. Como veremos en el apartado siguiente, la intuición de McClintock de la relación de los tranposones con la adaptación al estrés ha resultado certera.
Riesgos y beneficios de un genoma con transposones: mutación, plasticidad y adaptación Los transposones son elementos genéticos móviles que se encuentran repetidos en el genoma. Estas dos características, su movilidad y su repetitividad, los hacen altamente mutágenos. Y por ello, la presencia de un gran número de estos elementos en los genomas parece paradójica. ¿Cómo es posible mantener la información genética de célula a célula y de generación en generación, cuando esta información también contiene elementos que tienden a modificarla? En realidad, la información genética no se mantiene inalterable durante la vida de un organismo o la evolución de una especie, y tanto las mutaciones somáticas como las germinales son, de hecho, esenciales. La cuestión es cuánto cambio puede ser útil y cuándo empieza a ser un riesgo demasiado alto. Y esto puede depender del tipo de organismo, de la célula o el tejido en cuestión o de las condiciones en las que se encuentre el organismo. La
DOSIER CIENTÍFICO
actividad de los transposones no es igual en todas las células y, por ejemplo, suele estar fuertemente reprimida en la línea germinal, mientras que la actividad en ciertas células somáticas puede ser particularmente alta. Un ejemplo paradigmático es la activación de los retrotransposones L1 durante el desarrollo neuronal en humanos, que puede ser en parte responsable de la alta mosaicidad y plasticidad de las neuronas, características esenciales para el correcto desarrollo del cerebro.
Por otra parte, el trabajo de muchos laboratorios durante los últimos 20 años ha puesto de manifiesto que la actividad de los transposones de plantas está frecuentemente ligada a situaciones de estrés, confirmando una vieja hipótesis de McClintock que proponía a los transposones como elementos reguladores que permiten la reorganización del genoma en situaciones a las que el organismo no está bien adaptado. Muchos transposones se activan en situaciones de estrés, en particular en plantas, debido a que tienen promotores regulados por estrés. Por otra parte, trabajos recientes sugieren que algunos transposones podrían insertarse preferencialmente cerca de los genes de respuesta al estrés guiados por marcas epigenéticas específicas, lo que permitiría generar nueva variabilidad en estos genes y permitir al organismo explorar variaciones en la respuesta al estrés y adaptarse. De una manera general, la inserción de transposones, con sus promotores o elementos reguladores específicos, puede modificar la regulación de los genes que se encuentran cerca, y los datos acumulados en los últimos años sugieren la importancia de este fenómeno en la evolución de la regulación de la transcripción en eucariotas. Por ejemplo, más de la mitad de las regiones de cromatina abierta específica de primates,
que potencialmente contienen elementos regladores específicos, están en secuencias de transposones y muchos de los sitios de unión de factores de transcripción en estas especies también están en secuencias de transposones. También en plantas se ha podido demostrar que los transposones pueden amplificar y redistribuir en el genoma sitios de unión de factores de transcripción, dotando de plasticidad y capacidad de evolución a las redes transcripcionales. Los transposones han sido pues una fuente de variación genética muy importante en la evolución de los eucariotas. Por ejemplo, muchas de las mutaciones clave para la domesticación y mejora de las plantas cultivadas, son debidas a la inserción de transposones. De la arquitectura de la planta del maíz moderno a la diversidad de color de las uvas o de las naranjas sanguinas, de la forma de algunos tomates a la piel suave de las nectarinas, todos son fenotipos provocados por la inserción de transposones.
Pero, por supuesto, solo una pequeña parte de las mutaciones supone una ventaja fenotípica y los eucariotas han desarrollado sofisticados mecanismos de control para regular la actividad de los transposones. El principal mecanismo de control de los transposones es el silenciamiento génico, y en particular su metilación y heterocromatinización. De hecho, es probable que los mecanismos epigenéticos de control de la expresión génica apareciesen en la evolución por la necesidad de controlar a los elementos genéticos invasores del genoma, como los virus y los transposones. Por lo tanto, es en buena medida la presencia de transposones, a pesar del riesgo que entrañan, o en algunos casos debido a ese riesgo y a la necesidad de controlarlo, lo que ha permitido alcanzar la complejidad de los genomas eucariotas.
Además de forzar el desarrollo de sistemas de control, los transposones también han sido una fuente importante de nuevos genes y funciones. Los transposones codifican para distintas proteínas, necesarias para asegurar su movilidad, que han sido domesticadas repetidas veces durante la evolución para realizar otras funciones celulares. En animales, quizás el caso más evidente son las Sincitinas que están implicadas en el desarrollo de la placenta de los mamíferos y que derivan de genes Env de retrotransposones (retrovirus endógenos), domesticados independientemente en distintos linajes de mamíferos. En plantas, varios factores de transcripción, como FHY3 y FAR, que regulan la transducción de señal de la luz, derivan de transposasas y varias proteínas que participan en la regulación epigenética de genes y transposones, como MAIL1, MAIN y ALP1 derivan de distintas proteínas de elementos móviles.
Los transposones como componentes estructurales de los cromosomas
Además de las funciones que hemos descrito en el apartado anterior, los transposones también son esenciales para asegurar la estructura y la homeostasis de los genomas eucariotas. Los telómeros de los genomas eucariotas se mantienen en la mayoría de los casos por la acción de la telomerasa, que copia por transcripción reversa un molde de ARN, un proceso filogenéticamente relacionado con la transposición de algunos retrotransposones. Una relación todavía más evidente si se tiene en cuenta que Drosophila, que no tiene telomerasa, asegura la longitud de sus telómeros por la transposición específica de dos retrotransposones, HeT-A y TART, al extremo de sus cromosomas. Por otra parte, los transposones pueden ayudar a mantener la
integridad de los cromosomas insertándose en roturas de la doble cadena de ADN (Double-Strand Breaks, DSB por sus siglas en inglés) ayudando a reparar el ADN.
Un caso muy particular de rotura y reparación del ADN es la recombinación V(D)J que se da en los linfocitos de los vertebrados, y que permite generar una gran variedad de anticuerpos, inmunoglobulinas y receptores de células
T. Esta recombinación está mediada por las recombinasas RAG1 y RAG2 que se unen, cortan y recombinan secuencias de ADN específicas. Tanto estos enzimas como las secuencias reconocidas derivan directamente de antiguos transposones.
Por otro lado, la relación de los transposones con el centrómero, esencial para la estructura y la dinámica de los cromosomas, es también muy estrecha. Además del hecho de que la proteína CNP-B, importante para la estabilidad centromérica en mamíferos, sea una antigua transposasa, los transposones son parte integral de los centrómeros de la mayoría de eucariotas y en Arabidopsis se ha podido demostrar que el silenciamiento de los retrotransposones centroméricos mediado por pequeños ARNs es esencial para el correcto funcionamiento del centrómero. La función centromérica está muy conservada en los eucariotas, pero paradójicamente su secuencia es extraordinariamente variable. Las secuencias centroméricas suelen ser altamente repetitivas y la región funcionalmente activa se define por la presencia de CENH3, la variante centromérica de la histona H3. Sin embargo, la secuencia del ADN varía entre cromosomas del mismo individuo o entre variedades o especies cercanas. En plantas, los centrómeros suelen estar formados por repeticiones en tándem de satélites centroméricos (CEN178 en Arabidopsis), e inserciones de
retrotransposones, aunque en algunas especies los centrómeros pueden estar formados únicamente por secuencias de retrotransposones. La acumulación de retrotransposones en los centrómeros y zonas pericentroméricas puede ser debido a que esas regiones tienen una baja densidad en genes y por lo tanto, las inserciones de transposones tienen un efecto menos deletéreo, y la presión de selección para eliminarlos es menor. Por otra parte, la baja recombinación de esas regiones hace más difícil su eliminación. Pero además de estos mecanismos que podrían facilitar la acumulación de retrotransposones en el centrómero, se ha demostrado que existen retrotransposones centrofílicos que se insertan específicamente en estas regiones. Los centrómeros se caracterizan por una evolución rápida de su secuencia, dentro de la misma especie y entre especies distintas. Se ha propuesto que los centrómeros de plantas podrían alternar de forma dinámica entre estar formados
por repeticiones centroméricas o retrotransposones. Los retrotransposones centrofílicos podrían colonizar un centrómero basado en repeticiones centroméricas, pero a su vez, existen algunos datos que sugieren que la eliminación por recombinación de los retrotransposones podría dar lugar a nuevas repeticiones simples en tándem que podrían convertirse en nuevas repeticiones centroméricas, lo que proporcionaría un mecanismo para explicar la rápida evolución de estas secuencias.
Conclusión
Casi setenta y cinco años después de que McClintock propusiera la existencia de los transposones, y gracias a los datos acumulados por un gran número de laboratorios, podemos afirmar que estos elementos, lejos de ser sólo “basura genómica”, son esenciales para explicar la estructura y la dinámica de los cromosomas, y asegurar al mismo tiempo, la complejidad y la plasticidad de los genomas eucariotas.
Para leer más
Cui X, Cao X. Epigenetic regulation and functional exaptation of transposable elements in higher plants. Current Opinion in Plant Biology 21 (2014) 83–88. https://doi.org/10.1016/j.pbi.2014.07.001
Joly-Lopez Z, Bureau TE. Exaptation of transposable element coding sequences. Current Opinion in Genetics & Development 49 (2018) 34–42. https://doi.org/10.1016/j.gde.2018.02.011
Lisch D. How important are transposons for plant evolution? Nature Review Genetics 14 (2013) 49–61. https://doi.org/10.1038/nrg3374
Naish M, Henderson IR. The structure, function, and evolution of plant centromeres. Genome Research 34 (2024) 161–178. doi: 10.1101/gr.278409.123
Wells JN, Feschotte C. A field guide to eukaryotic transposable elements. Annual Review of Genetics 54 (2020) 539–561. https:// doi.org/10.1146/annurev-genet-040620-022145
LOS ELEMENTOS MÓVILES:
ADN REPETITIVO
RELEVANTE PARA LA FUNCIÓN, ESTRUCTURA Y EVOLUCIÓN DE LOS GENOMAS
Josefa González
Institut Botànic de Barcelona, CSIC, CMCNB, Barcelona
https://doi.org/10.18567/sebbmrev_223.202503.dc3
Descubiertos por la Dra. Barbara McClintock en la década de 1940, los elementos móviles fueron definidos como secuencias de ADN que pueden controlar la expresión génica y cambiar su localización en el genoma. Este descubrimiento fue recibido con escepticismo por la comunidad científica y los elementos móviles fueron considerados durante años como una peculiaridad del genoma del maíz, donde habían sido descubiertos. Sin embargo, el descubrimiento de que el ADN repetitivo representaba una fracción considerable de los genomas, basado inicialmente en el análisis
de reasociación del ADN y posteriormente en la secuenciación de los genomas completos, cambió la perspectiva sobre la potencial relevancia de estas secuencias. Actualmente, la evidencia a favor de la capacidad de algunos elementos móviles para controlar no solo la expresión de genes individuales sino también de redes génicas es incuestionable. Aun así, continúa habiendo un desconocimiento, en algunos sectores de la comunidad científica, sobre qué son exactamente los elementos móviles y cuál es su relevancia para la función, la evolución y la estructura de los genomas.
Los elementos móviles son componentes diversos, abundantes y activos de los genomas
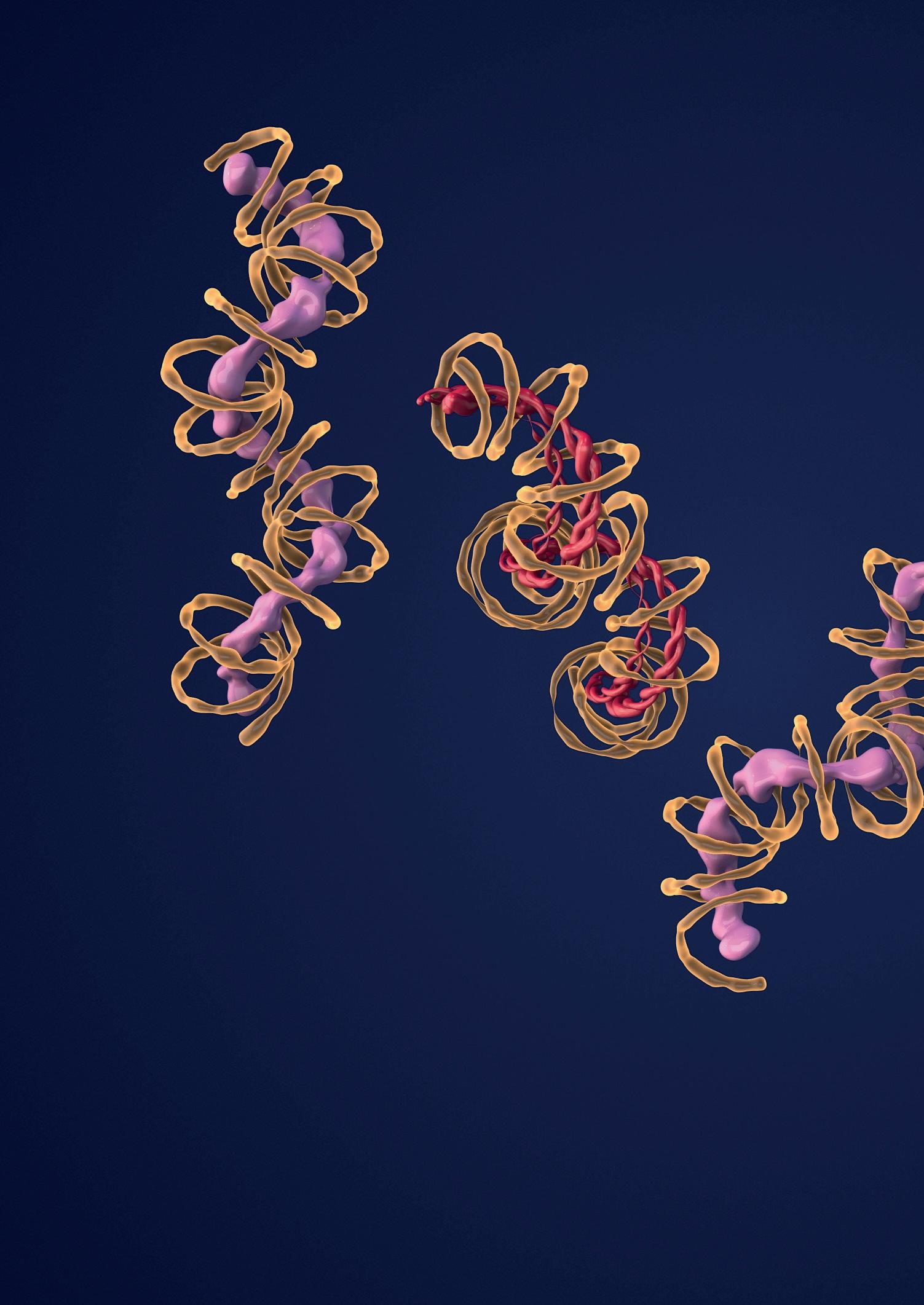
Bajo la definición de elementos móviles se agrupa una gran diversidad de secuencias de ADN que se caracterizan por su capacidad de moverse de un lugar a otro en el genoma (Figura 1). Mientras que solo algunos elementos móviles codifican los genes que catalizan este movimiento, todos los elementos móviles contienen las secuencias que son reconocidas por estos genes permitiendo así su movilización. Algunos elementos móviles están relacionados con los retrovirus, mientras que otros tienen como origen otras secuencias del genoma como los ARN de transferencia. A pesar de la gran diversidad en la secuencia de los elementos móviles, el número de dominios proteicos que catalizan su replicación y movilización es limitado. Algunos
de estos dominios contienen un motivo de reconocimiento de ARN que sugiere que el origen de los elementos móviles es anterior al origen de los eucariotas.
La capacidad de los elementos móviles de generar mutaciones en el genoma está asociada tanto a su capacidad de movimiento (mutaciones insercionales), como a su naturaleza repetitiva, ya que los elementos móviles generan nuevas copias de su secuencia como parte del proceso de movilización. Las copias de elementos móviles de la misma familia, y por tanto muy similares en su secuencia, pueden actuar como substratos para la recombinación ectópica generando reordenaciones cromosómicas como inversiones y translocaciones.
Tanto el contenido de elementos móviles de un genoma como el porcentaje de elementos móviles que potencialmente podrían
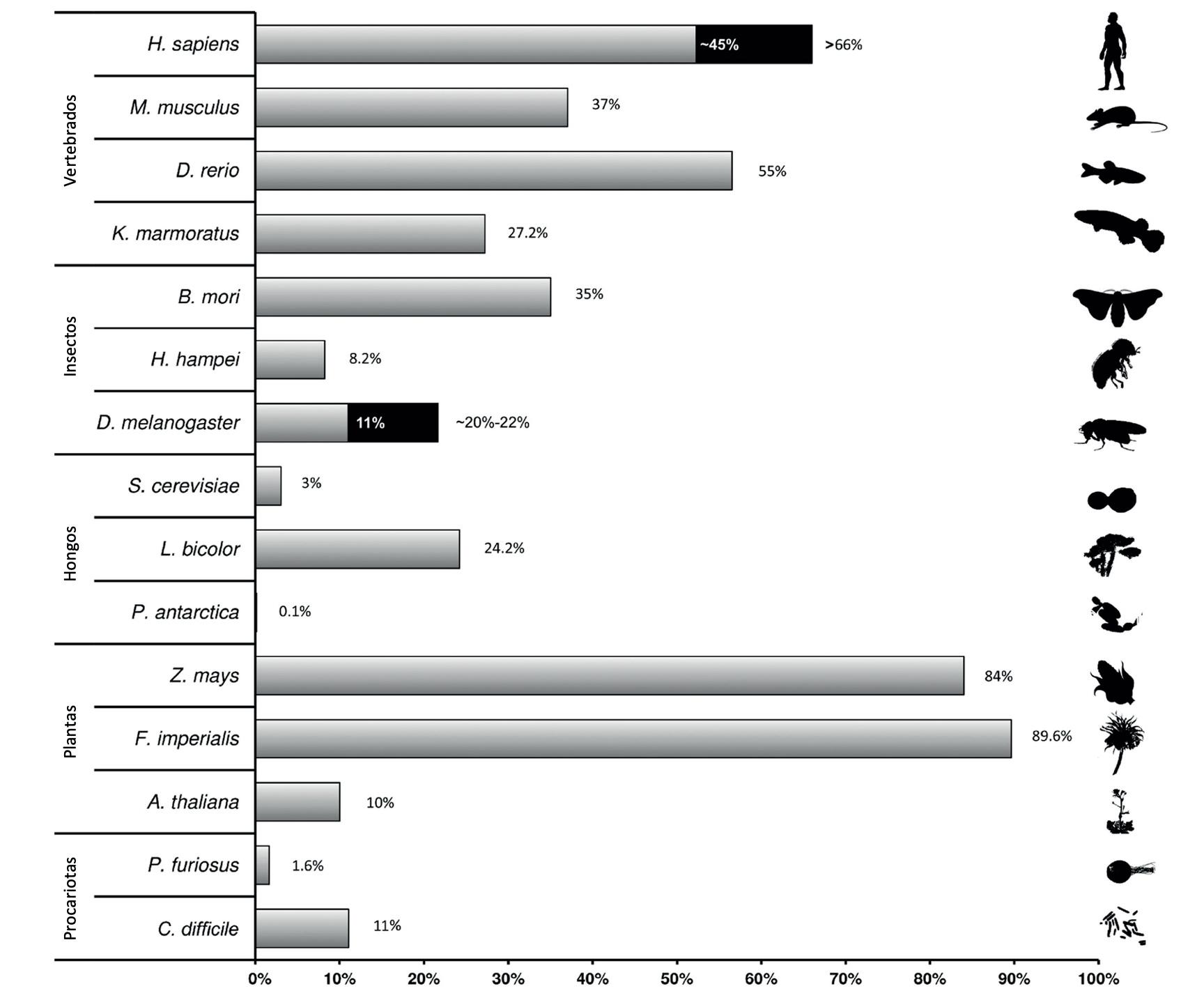
ser activos, es decir, moverse de un lugar a otro en el genoma, varía de unas especies a otras (Figura 2). En el organismo modelo Drosophila melanogaster (la mosca de la fruta), aproximadamente el 20% del genoma son elementos móviles y la mayoría de las familias son potencialmente activas. En humanos, más de la mitad del genoma son elementos móviles, y solo 3 familias, L1, Alu, y SVA, son activas. En todos los genomas, la mayoría de los elementos móviles están silenciados por la maquinaria celular, evitando por tanto su movimiento y su incremento incontrolado en número de copias. Los mecanismos que las células utilizan para silenciar a los elementos móviles son también diversos e incluyen tanto mecanismos epigenéticos (p.ej., metilación y marcas de histonas) como mecanismos de autorregulación basados por ejemplo en splicing
Figura 1
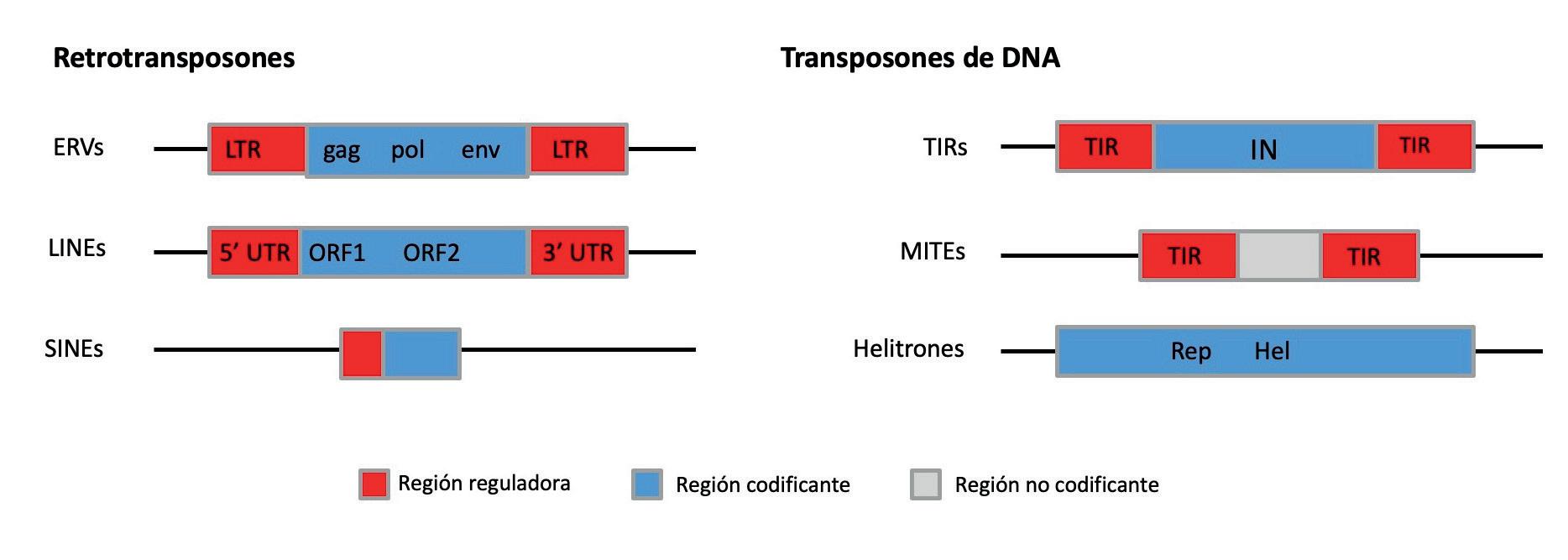
Los elementos móviles son secuencias de ADN muy diversas. Los elementos móviles se clasifican en retrotransposones y transposones de ADN, dependiendo de si utilizan o no un intermediario de ARN para su movilización. En la figura se muestran algunos de los elementos móviles que pertenecen a estos dos grandes grupos incluyendo elementos móviles que codifican los genes necesarios para su movilización (ERVs, LINEs, Transposones y Helitrones) y elementos móviles que necesitan la maquinaria enzimática de otros elementos para poder moverse (SINEs, MITEs).
LTR: long terminal repeat; gag, pol, env, IN, Rep, Hel: dominios proteicos presentes en elementos móviles; UTR: untranslated region; ORF: open reading frame; TIR: terminal inverted repeat
alternativo. La mayoría de copias de elementos móviles en el genoma son fragmentos de elementos móviles que en algún momento durante la evolución fueron copias activas. Son principalmente estos fragmentos inmóviles de elementos móviles los que pueden reutilizarse en los genomas para llevar a cabo funciones biológicas, con
algunas excepciones de elementos móviles aún activos que también han adquirido un papel funcional.
Los elementos móviles pueden controlar la expresión génica Los elementos móviles contienen regiones reguladoras, como promotores y sitios de unión de factores de transcripción, que
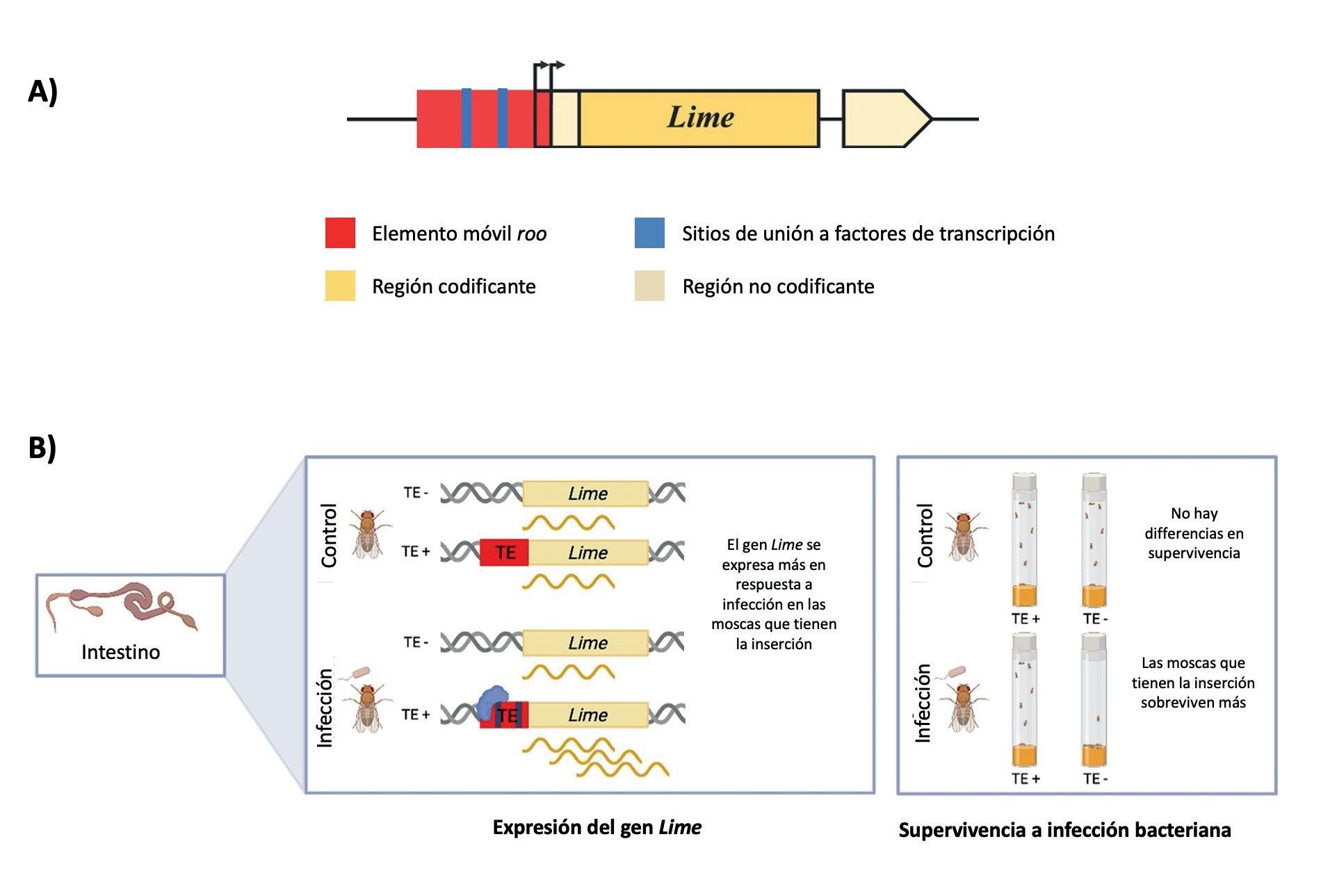
controlan la expresión de los genes que permiten su replicación y movilización (Figura 1) Sin embargo, estas secuencias reguladoras pueden afectar también la expresión de otros genes. Por ejemplo, en D. melanogaster un elemento móvil de la familia roo insertado en el promotor del gen Lime (Linking immunity and
Figura 2
Contenido de elementos móviles en diferentes organismos. La estimación del contenido de elementos móviles se representa como el porcentaje de elementos móviles respecto al total del genoma. Para humanos (H. sapiens) y D. melanogaster además de las estimaciones basadas en homología de secuencia, se dan (en negro) las estimaciones basadas en la aproximación P-cloud y en aproximaciones que identifican TEs de novo, respectivamente.
Figura modificada de Guio y González (2019). New insights on the evolution of genome content: population dynamics of transposable elements in flies and humans. En: Anisimova, M. (eds) Evolutionary Genomics. Methods in Molecular Biology, vol 1910. Humana, New York, NY. https://doi.org/10.1007/978-1-4939-9074-0_16 Licencia internacional: Creative Commons Attribution 4.0 (https://creativecommons.org/licenses/by/4.0/)
Figura 3
El elemento móvil roo afecta a la supervivencia de D. melanogaster en respuesta a una infección. (A) El elemento móvil roo está insertado en el 5’UTR del gen Lime y contiene lugares de unión a factores de transcripción. Las flechas indican lugares de inicio de la transcripción. (B) Las moscas que tienen la inserción del elemento roo en el gen Lime tienen un mayor nivel de expresión de este gen en el intestino de la mosca en respuesta a la infección. Estas diferencias de expresión son las responsables de la mayor supervivencia a la infección de las moscas que contienen este elemento móvil. Figura modificada de Merenciano y González (2023). The interplay between developmental stage and environment underlies the adaptive effect of a natural transposable element insertion. Mol Biol Evol 40: msad044, https://doi.org/10.1093/molbev/msad044 Licencia internacional: Creative Commons Attribution-NonCommercial License (https://creativecommons.org/licenses/by-nc/4.0/).
metabolism) contiene regiones reguladoras que modulan la expresión de este gen en el intestino afectando a la respuesta inmune (Figura 3). Mediante la técnica de CRISPR, se eliminó la secuencia del elemento roo del genoma y se demostró que la inserción era la responsable del incremento de la expresión del gen Lime y de la mayor supervivencia a infección bacteriana de las moscas que tienen este elemento móvil. El análisis de la secuencia del
elemento móvil reveló la presencia de varios sitios de unión a factores de transcripción implicados en la respuesta inmune y mediante mutagénesis dirigida se demostró que estas secuencias eran las responsables del incremento de la expresión del gen Lime en respuesta a infección Además de afectar a genes individuales, los elementos móviles también pueden afectar la expresión de redes génicas. Se ha demostrado, también mediante la
técnica de CRISPR, que elementos móviles de la familia MER41B, actúan como enhancers de genes de inmunidad innata en mamíferos. El análisis de varias líneas celulares permitió identificar varias familias de elementos móviles enriquecidas en sitios de unión a factores de transcripción implicados en la respuesta a interferón. La mayoría de estas familias de elementos móviles eran retrovirus endógenos y estaban localizados cerca de genes de respuesta
DOSIER CIENTÍFICO
inmune. La deleción mediante CRISPR de varias copias de una de estas familias de retrovirus endógenos, MER41B, permitió demostrar que estos elementos móviles controlan la expresión de varios genes de respuesta a interferón. Otros ejemplos de retrovirus endógenos que afectan a la regulación de múltiples genes son la familia MER130 que actúa como enhancers de genes con actividad específica en el prosencéfalo y se han relacionado con la evolución del neocórtex en mamíferos; o la familia RLTR13 específica de ratón que actúa como enhancers específicos de placenta y se ha relacionado con la diversificación morfológica de la placenta en mamíferos. Además de la reutilización de las secuencias reguladoras contenidas en los elementos móviles, la inducción de cambios epigenéticos o la generación de ARN no codificante son otros de los mecanismos por los cuales los elementos móviles pueden controlar la expresión génica.
Los elementos móviles contribuyen a la diversificación del transcriptoma y del proteoma
Además de afectar la expresión génica, los elementos móviles también pueden incorporarse a regiones codificantes de genes generando así nuevos transcritos, conocidos como transcritos quiméricos. Dos estudios recientes han explorado la contribución de los elementos móviles a los transcriptomas de D. melanogaster y humanos. En D. melanogaster, la contribución de los transcritos quiméricos a la expresión total de los genes es variable, con un 38% de genes que solo expresan el transcrito quimérico. Además, varios de los transcritos quiméricos identificados incorporan un dominio proteico del elemento móvil al gen y para al menos dos de ellos hay evidencia de que el transcrito quimérico tiene consecuencias
funcionales para el organismo. En humanos, análisis funcionales para cuatro de los transcritos quiméricos identificados en líneas celulares cancerosas, demostraron que, a pesar de que el nivel de expresión de estos transcritos es bajo, los cuatro tienen una función biológica distinta a la de los transcritos no quiméricos. Los elementos móviles contribuyen por tanto a generar diversidad en el transcriptoma que puede ser funcional. Algunas de las proteínas codificadas por los elementos móviles han sido domesticadas para realizar funciones en las células. Este es el caso por ejemplo de las proteínas conocidas como sincitinas. Las sincitinas son proteínas que tienen su origen en el gen envelope que codifican los retrovirus endógenos, un tipo de elemento móvil. El gen envelope, también presente en virus, juega un papel en la fusión de membranas que permite a los virus entrar en las células. En mamíferos, las sincitinas juegan un papel en la placenta creando una capa de células fusionadas que ayuda en el intercambio de nutrientes entre el feto y la madre. Las sincitinas han sido domesticadas de manera independiente al menos 10 veces en varios grupos de mamíferos. Los elementos móviles, por tanto, han jugado un papel relevante en la transición de organismos ovíparos a organismos vivíparos, una innovación clave que podría haber jugado un papel importante en el éxito evolutivo de los mamíferos. Recientemente se ha descrito la función en el desarrollo del pez cebra de la proteína gag de un retrovirus endógeno aún activo en el genoma de este organismo. Los autores proponen que algunas de las proteínas codificadas por los elementos móviles podrían realizar funciones redundantes con las de otras proteínas de las células. Similar a lo que ocurre con algunos genes duplicados, la
redundancia funcional introducida por la proteína del elemento móvil podría conllevar la relajación de la selección en la secuencia de la proteína celular, resultando finalmente en la proteína del elemento móvil siendo la única que lleva a cabo la función.
Los elementos móviles son componentes estructurales de los genomas
Centrómeros y telómeros son esenciales para la estructura de los cromosomas. Los elementos móviles están presentes en los centrómeros de muchos organismos eucariotas, aunque su función exacta es desconocida en la mayoría de ellos. En la planta Arabidopsis thaliana se ha demostrado recientemente que el silenciamiento epigenético del elemento móvil ATHILA5, presente en los centrómeros, es esencial para la cohesión de los cromosomas y para la segregación de los cromosomas durante la división celular. Los elementos móviles son además el origen de las proteínas de unión al centromero CENP-B-like que también juegan un papel relevante en la segregación de los cromosomas durante la división celular. Estas proteínas han evolucionado a partir de secuencias de elementos móviles al menos dos veces de manera independiente, una en mamíferos y otra en hongos.
Los telómeros de la mayoría de organismos están formados por repeticiones cortas de ADN, cuya longitud es mantenida por la proteína telomerasa. Sin embargo, en D. melanogaster no existe esta proteína y los telómeros están formados por elementos móviles. Mientras que la mayoría de domesticaciones de elementos móviles, como la de los que han dado lugar a las proteínas CENPB-like o a las sincitinas, son de fragmentos de elementos móviles, en el caso de los telómeros
de D. melanogaster se han domesticado elementos móviles que aún son activos.
Las nuevas técnicas de análisis de la estructura 3D del genoma están revelando también el papel de los elementos móviles en la estructura de dominios y bucles de la cromatina. Es precisamente gracias a los avances tecnológicos en secuenciación del genoma, transcriptoma y epigenoma, y al desarrollo de herramientas bioinformáticas para su estudio, que la relevancia de los elementos móviles en procesos diversos como el desarrollo, cáncer, o envejecimiento comienza a ser reconocida por sectores más amplios de la comunidad científica.
Para leer más
Coronado-Zamora M, González J. Transposons contribute to the functional diversification of the head, gut and ovary transcriptomes across Drosophila natural strains. Genome Research 33 (2023) 1541-1553. doi: 10.1101/gr.277565.122
Naville M, Warren IA, Haftek-Terreau Z, Chalopin D, Brunet F, Levin P, Galiana D, Volff J-N. Not so bad after all: retroviruses and long terminal repeat retrotransposons as a source of new genes in vertebrates. Clinical Microbiology and Infection 22 (2016) 312-323. https://doi.org/10.1016/j.cmi.2016.02.001
Shimada A, Cahn J, Ernst E, Lynn J, Grimanelli D, Henderson I, Kakutani T, Martienssen RA. Retrotransposon addiction promotes centromere function via epigenetically activated small RNAs. Nature Plants 10 (2024) 1304–1316. https://doi.org/10.1038/ s41477-024-01773-1
ELEMENTOS TRANSPONIBLES EN HUMANOS Y SU IMPACTO EN EL DESARROLLO DE ENFERMEDADES
Layla Díaz-Portal y Bernardo Rodríguez-Martín Centre for Genomic Regulation (CRG), The Barcelona Institute of Science and Technology, Barcelona Universitat Pompeu Fabra (UPF), Barcelona
https://doi.org/10.18567/sebbmrev_223.202503.dc4
Introducción a los elementos transponibles
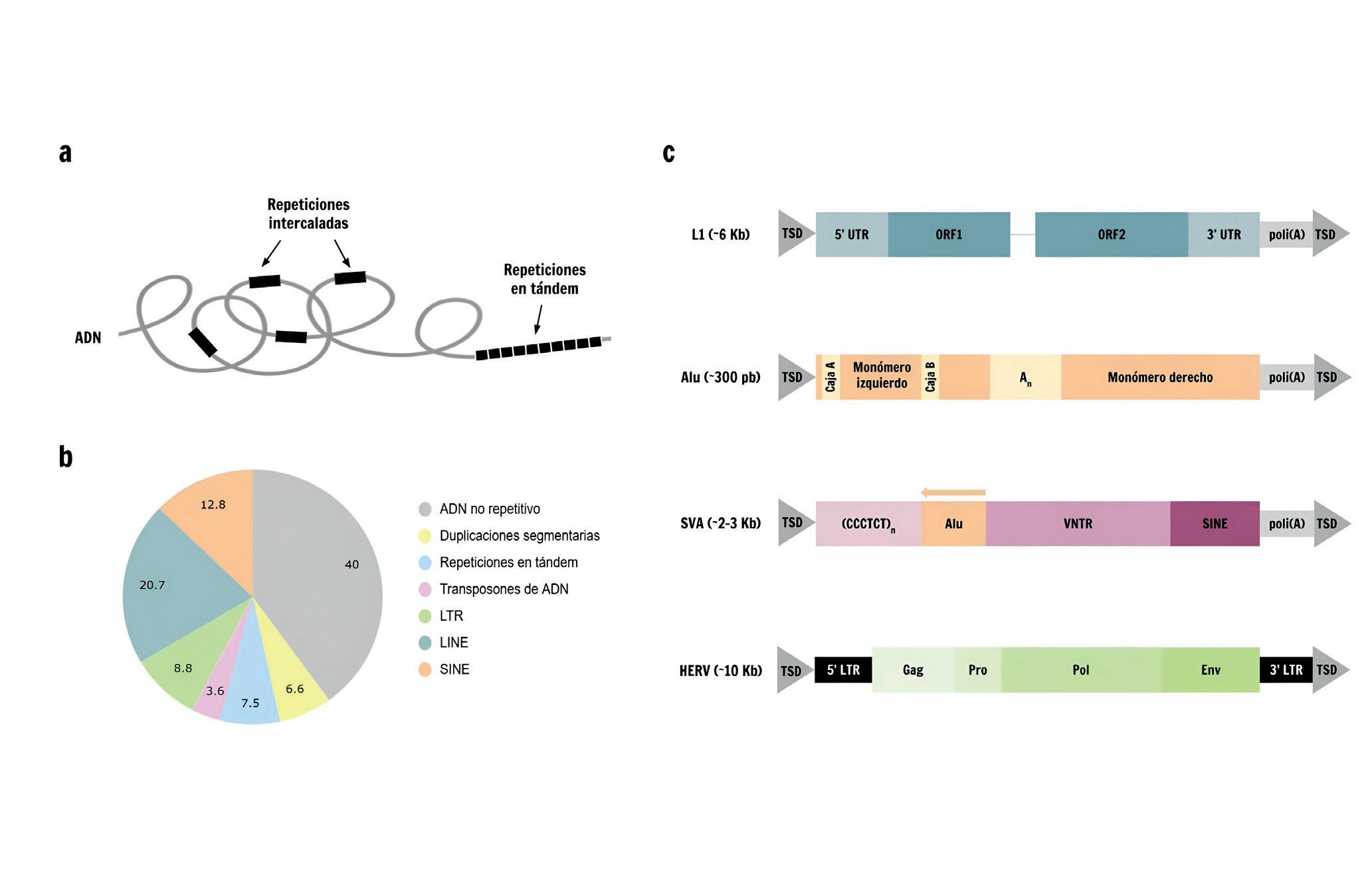
Tanto los organismos procariotas como los eucariotas contienen una proporción variable pero considerable de elementos repetidos a lo largo de su genoma. Estos pueden agruparse en dos categorías en base a su disposición: las repeticiones en tándem y las repeticiones intercaladas (Figura 1a) Mientras que las primeras son secuencias de longitud variable repetidas de forma contigua una tras
otra, las repeticiones intercaladas, incluyendo duplicaciones segmentarias y elementos transponibles (Transposable Elements, TE por sus siglas en inglés), se repiten en posiciones distantes, ocupando aproximadamente la mitad del genoma humano (Figura 1b)
Los TE, también conocidos como “genes saltarines”, son secuencias de ADN capaces de cambiar de posición en el genoma por sí mismos. En función de su mecanismo de movilización,
Figura 1
Naturaleza de los elementos transponibles y su distribución en el genoma humano. a) Las secuencias de ADN repetitivo pueden agruparse en dos categorías en base a su disposición: repeticiones intercaladas (incluyendo las duplicaciones segmentarias y los elementos transponibles) y las repeticiones en tándem. b) Porcentaje de cada tipo de elemento repetitivo en el genoma humano de referencia (versión T2T-CHM13). Los SVA no aparecen representados en el gráfico porque se encuentran en una frecuencia muy baja (~0,2%). c) Representación esquemática de los elementos transponibles más comunes en humanos con su tamaño completo indicado en kilobases (Kb) o pares de bases (pb). L1 codifica dos marcos de lectura (ORF, Open Reading Frame), el segundo con dominios de endonucleasa y transcriptasa inversa. Los elementos Alu presentan dos monómeros separados por un conector rico en A. SVA es un elemento compuesto, constituido por una repetición hexamérica CCCTCT, una secuencia tipo Alu invertida, un número variable de repeticiones en tándem (VNTR, Variable Number of Tandem Repeats) y una secuencia de tipo SINE. Tanto L1, como Alu y SVA presentan en su extremo 3’ una cola de poli(A). Los retrovirus endógenos humanos (HERV, Human Endogenous Retroviruses) codifican tres proteínas virales esenciales (Gag, Env y Pol), además de una proteasa (Pro). Todos los elementos están flanqueados en sus extremos por duplicaciones de la secuencia objetivo (TSD, Target Site Duplication).
podemos dividirlos en dos categorías: los TE de clase I, más conocidos como retrotransposones, que se movilizan mediante un mecanismo de “copia y pega” que involucra un intermediario de ARN, y los TE de clase II, denominados transposones, que emplean un mecanismo de “corta y pega”, movilizándose directamente como ADN. Los retrotransposones se pueden clasificar a su vez en dos grupos en base a la presencia de repeticiones terminales largas
(Long Terminal Repeats, LTR por sus siglas en inglés) flanqueando su secuencia o la carencia de ellas (no-LTR).
A día de hoy, sólo hay evidencias de actividad para tres clases de TE (Alu, LINE-1 y SVA), todos ellos retrotransposones no-LTR, en el genoma humano. Tanto Alu como SVA dependen de la maquinaria proteica codificada por LINE-1 (L1) para su retrotransposición, mientras que L1 se trata del único retrotransposón con capacidad
autónoma para movilizarse. Un elemento L1 de longitud completa tiene un tamaño aproximado de 6 kilobases (Kb) y dos marcos de lectura abiertos (Open Reading Frame, ORF por sus siglas en inglés), que codifican la maquinaria proteica necesaria para su movilización e integración (Figura 1c). L1 representa el tipo de TE más prevalente en el genoma humano en términos de su contribución al número total de bases, constituyendo en torno al 17% de su secuencia. En total,
Figura 2
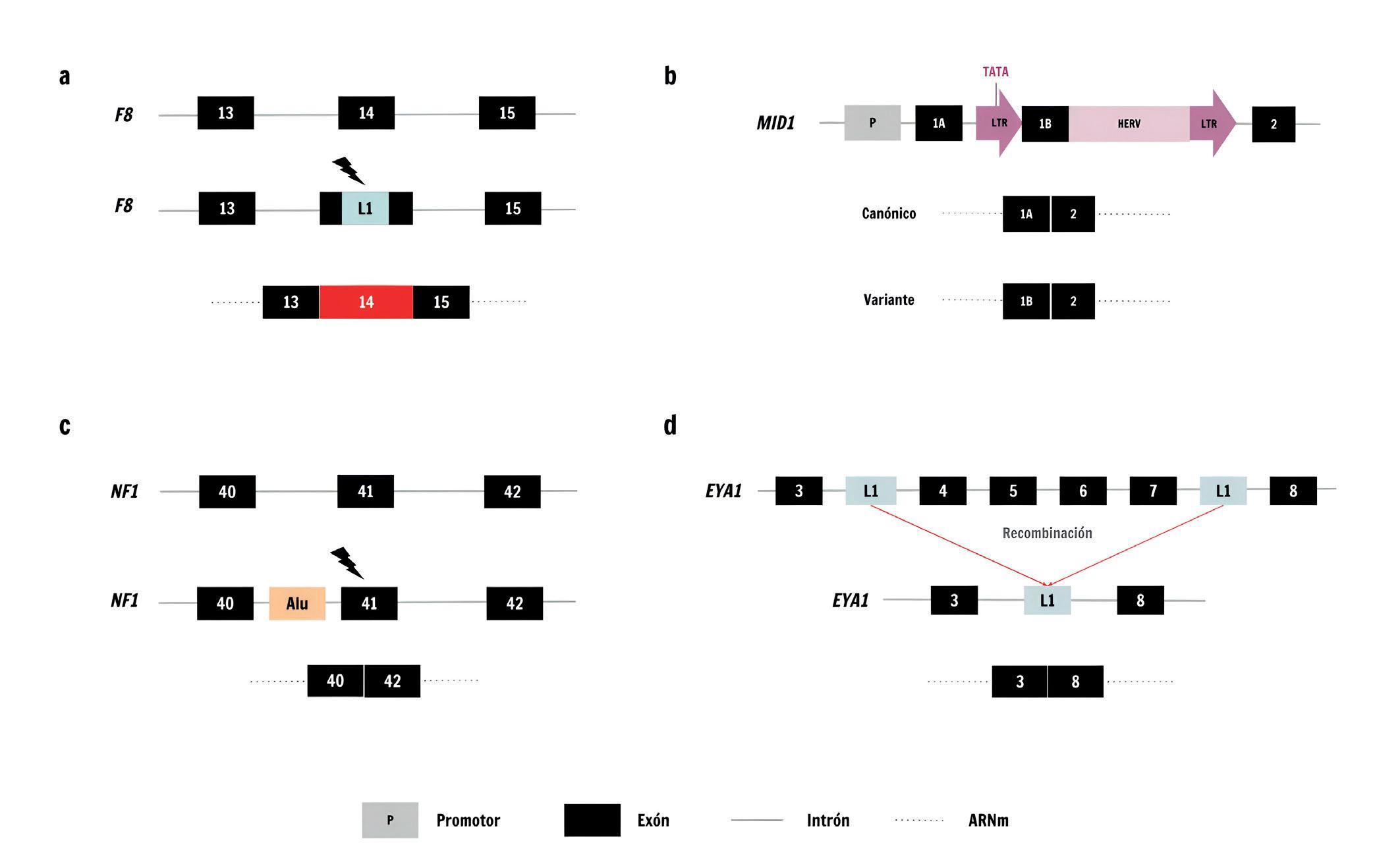
Principales mecanismos mediante los cuales los TE pueden causar enfermedades monogénicas. a) Alteración de la función génica a través de la disrupción de regiones codificantes. La inserción de novo de L1 interrumpe el exón 14 del gen para el factor de coagulación VIII (F8) en un paciente con hemofilia A. b) Alteración de la transcripción del gen MID1 debido a la introducción de sitios de unión para elementos reguladores. La inserción de un HERV en un paciente con síndrome de Opitz ligado al cromosoma X proporciona un nuevo sitio de inicio de la transcripción (TATA) y un nuevo intrón 1 que conduce a un transcrito de ARNm más corto que el original. c) Alteración de la función génica a través de la disrupción de regiones no codificantes. Una inserción Alu en un paciente con neurofibromatosis tipo I se localiza 44 pb aguas arriba del exón 41 de NF1, lo que provoca la omisión del mismo y cambios en el marco de lectura. d) Reordenamientos del material genético debidos a la recombinación entre elementos. La recombinación entre dos L1 intrónicos dentro del gen EYA1 provoca la pérdida de los exones 4 a 7 en un paciente con síndrome branquio-oto-renal.
existen más de 500.000 copias de L1 en el genoma humano de referencia, resultado de su actividad y movilización continua durante los últimos 150 millones de años. Sin embargo, la mayoría de L1 ha perdido la capacidad para propagarse debido a la acumulación de mutaciones puntuales, reordenamientos en su secuencia, o bien a que se encuentran localizados en regiones con poca actividad transcripcional, como la heterocromatina. Por lo tanto, únicamente un reducido número de elementos (se estima que
en torno a unos 100 por genoma humano) están completamente intactos, y un número aún más pequeño son altamente activos (denominados en inglés hot L1).
La retrotransposición y subsecuente inserción de TE puede afectar tanto a la expresión génica como a la estructura genómica. Si bien esto hace de los TE una importante fuente de variabilidad genética y fenotípica, impulsando la evolución de los genomas y la adaptación de las especies al medio, esta capacidad mutagénica
también puede provocar que se vean asociados al desarrollo de ciertas enfermedades.
Inserción de los TE en la línea germinal y sus consecuencias en la estructura genómica y la expresión génica
El primer caso reportado de una inserción de TE patogénica en humanos tuvo lugar en 1988, cuando un grupo de investigadores identificó, en el genoma de un paciente con hemofilia A sin antecedentes familiares, una inserción L1 de novo en el exón 14 del gen F8 en
Figura 3
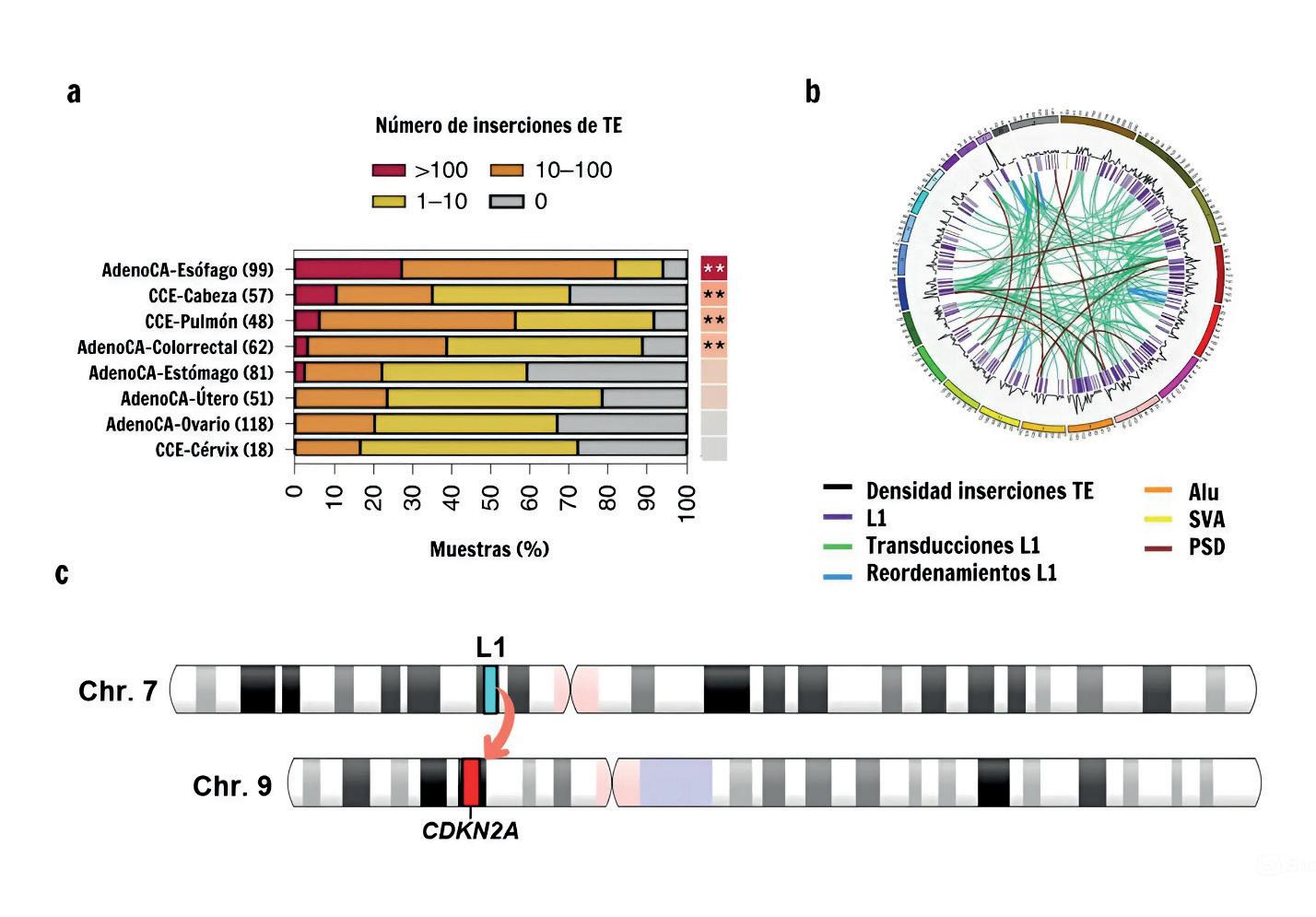
Retrotransposición somática en los cánceres humanos. a) Para los ocho tipos de cáncer del proyecto Pan-Cáncer con mayores tasas de retrotransposición somática, se representa la proporción de tumores con >100 (rojo), 10–100 (naranja), 1–10 (amarillo) y 0 (gris) retrotransposiciones somáticas. AdenoCA se refiere a los adenocarcinomas, mientras que CCE hace referencia a los carcinomas de células escamosas. El número de muestras analizadas para cada tipo de tumor se muestra entre paréntesis. b) Tumor de cabeza y cuello con una alta tasa de retrotransposición (638 inserciones somáticas). La localización genómica de cada tipo de inserción se indica con una línea de color: inserciones de L1, transducciones (movilización de ADN aguas abajo del TE) mediadas por L1, reordenamientos mediados por L1, inserciones de Alu, SVA y pseudogenes (PSD). c) Ilustración de una pérdida de ADN de 5 millones de bases, afectando al gen supresor CDKN1A, causada por un L1 altamente mutagénico en el cromosoma 7. Figura adaptada de doi: 10.1038/s41588-019-0562-0.
el cromosoma X (Figura 2a) que codifica un factor de coagulación sanguínea. Desde entonces, se han reportado más de 100 inserciones de TE en la línea germinal causantes de varias decenas de enfermedades monogénicas.
Más allá de la disrupción de regiones codificantes, se deben considerar los efectos inherentes a la propia secuencia del elemento insertado, la cual puede contener sitios de unión para elementos reguladores (p. ej. promotores) o aceptores para el corte y empalme
(en inglés splicing) del ARN mensajero (ARNm). Un ejemplo notable es el caso de las inserciones de retrovirus endógenos humanos (Human Endogenous Retroviruses, HERV por sus siglas en inglés) asociadas al síndrome de Opitz ligado al cromosoma X. Los HERV están flanqueados por repeticiones terminales largas (Long Terminal Repeats, LTR por sus siglas en inglés), las cuales pueden actuar como promotores y potenciadores, generando un sitio alternativo de inicio de la
transcripción para el gen MID1 (Figura 2b), que resulta en el desarrollo de la enfermedad. Por otro lado, las inserciones de TE en regiones intrónicas pueden alterar el proceso de corte y emplame (splicing), afectando a la función génica con efectos patogénicos. Un ejemplo es el caso de la neurofibromatosis tipo I, una enfermedad genética común vinculada al gen supresor de tumores NF1, que se caracteriza por cambios en la coloración de la piel y el crecimiento de tumores
DOSIER CIENTÍFICO
benignos a lo largo de los nervios. Se ha encontrado que la inserción de una secuencia Alu en el intrón 40 del gen NF1 provoca la exclusión del exón 41 durante la maduración del ARNm (Figura 2c) Por su parte, la distrofia muscular de Duchenne es una enfermedad caracterizada por una degeneración muscular progresiva causada por mutaciones en el gen de la distrofina (DMD), un gen de gran tamaño (~2,5 Mb) que contiene más de 60 exones y está ubicado en el cromosoma X. Se han reportado varios casos de inserciones tanto de L1 como de Alu que afectan al empalme alternativo de los transcritos del gen DMD y resultan en la omisión de alguno de sus exones, dando lugar a una distrofina no funcional. Por último, como consecuencia de su abundancia en el genoma y homología a nivel de secuencia, la recombinación de TE puede promover la formación de diversos tipos de reordenamientos cromosómicos, tales como deleciones, duplicaciones, translocaciones o inversiones de ADN. Aunque los eventos de recombinación Alu-Alu son los más frecuentes, con más de 70 casos reportados de enfermedades monogénicas, la recombinación entre dos elementos L1 o dos HERV también pueden tener consecuencias patogénicas. Un ejemplo es la pérdida de material genético en el gen EYA1, causada por la recombinación entre dos inserciones de L1 (Figura 2d), lo que da lugar en un paciente al síndrome branquio-oto-renal.
Movilización somática de los TE y su implicación en el cáncer
Debido a su capacidad mutagénica, los TE pueden ser considerados una amenaza para la integridad genómica, por lo que las especies hospedadoras han desarrollado mecanismos de defensa para silenciarlos. Estos mecanismos operan en todos los
niveles de la regulación de los TE, que van desde su silenciación epigenética hasta la degradación de los transcritos derivados de su expresión. No obstante, bajo determinadas condiciones, estos mecanismos pueden verse comprometidos, desencadenando la activación de TE y la mutagénesis derivada de su actividad. Hasta hace poco, se creía que los TE se movilizaban principalmente en las células germinales. Sin embargo, estudios recientes han mostrado actividad de TE en tejidos somáticos adultos, como el cerebro y el colon, además de altas tasas de movilización de retrotransposones de tipo L1 en cánceres epiteliales. En un estudio en el que se analizaron los genomas del cáncer para 244 individuos, se encontró que los tumores del 53% de los pacientes presentaban inserciones somáticas de L1. En un segundo estudio, utilizando una cohorte de casi 3.000 tumores de 38 tipos distintos de cáncer pertenecientes al proyecto PanCáncer, se reportaron altas tasas de retrotransposición para L1 en adenocarcinomas de esófago, cáncer colorrectal, y cabeza y cuello (Figura 3a). De hecho, en estos tres tipos de cáncer, la inserción de L1 representa uno de los tipos más frecuentes de variación estructural (p. ej. cambios en el ADN mayores de 50 bp) somática. Además, se encontró que la activación de sólo un pequeño conjunto de elementos L1 altamente mutagénicos puede desencadenar de decenas a cientos de retrotransposiciones en un sólo tumor (Figura 3b).
Por otro lado, investigadores del proyecto Pan-Cáncer hallaron un nuevo mecanismo mutacional en cáncer, que implica la generación de grandes reordenamientos cromosómicos como resultado de la integración aberrante de elementos L1. Estas integraciones pueden provocar la pérdida
de regiones cromosómicas de gran tamaño (Figura 3c), lo que puede resultar en la eliminación de genes supresores de tumores, así como la generación de translocaciones y duplicaciones a gran escala. Además, mediante este mecanismo, L1 puede desencadenar ciclos de inestabilidad genómica conduciendo a la amplificación de oncogenes. En definitiva, estos estudios remarcan la capacidad mutagénica de L1 y su impacto como fuente de variación somática e inestabilidad genómica en el genoma del cáncer, fomentando su evolución de una manera análoga a la evolución de las especies.
Desafíos y oportunidades para el estudio de los TE Los TE representan una fuente relevante de variabilidad genética, sirviendo de sustrato para la evolución de los genomas y adaptación al medio. No obstante, las mismas características que los hacen beneficiosos a nivel evolutivo también los convierten en una fuente de enfermedades, tanto mendelianas como complejas, incluido el cáncer. A pesar de su relevancia y potencial impacto en la salud, los TE continúan siendo uno de los tipos de secuencias más inexplorados del genoma humano. Debido a su alto porcentaje de homología a nivel de secuencia y a la gran cantidad de copias que se encuentran en el genoma, los TE han representado un desafío considerable para su detección y estudio durante décadas. No obstante, el desarrollo de tecnologías de secuenciación de lectura larga, como PacBio y Oxford Nanopore, cada vez más accesibles y precisas, junto con los avances en técnicas de ensamblaje de genomas, brindan una oportunidad sin precedentes para el estudio de la evolución y la diversidad de los TE y en el genoma humano.
Para leer más
Burns K. Transposable elements in cancer. Nature Reviews Cancer 17 (2017) 415–424. https://doi. org/10.1038/nrc.2017.35
Chénais B. Transposable elements and human diseases: Mechanisms and implication in the response to environmental pollutants. International Journal of Molecular Science 23 (2022) 2551. https://doi. org/10.3390/ijms23052551
Liao X, Zhu W, Zhou J. et al. Repetitive DNA sequence detection and its role in the human genome. Communications Biology 6 (2023) 954. https://doi.org/10.1038/s42003-023-05322-y
Payer LM, Burns KH. Transposable elements in human genetic disease. Nature Reviews Genetics 20 (2019) 760–772. https://doi.org/10.1038/s41576-019-0165-8
Rodríguez-Martín B, Álvarez EG, Báez-Ortega A, et al. Pan-cancer analysis of whole genomes identifies driver rearrangements promoted by LINE-1 retrotransposition. Nature Genetics 52 (2020) 306–319. https://doi.org/10.1038/s41588-019-0562-0
EL ORIGAMI DE LA CROMATINA: ORGANIZACIÓN ESTRUCTURAL
Y REGULACIÓN
GÉNICA
Jorge
Mañes García y Leonardo Beccari
Centro de Biología Molecular ‘Severo Ochoa’ (CBM), CSIC-UAM, Madrid.
https://doi.org/10.18567/sebbmrev_223.202503.dc5
La mayor parte del genoma de los vertebrados está constituido por ADN no codificante, que contiene elementos reguladores en cis (ERCs) responsables de controlar la expresión de sus genes diana mediante la interacción física con sus promotores. En el genoma humano, se ha llegado a estimar que existen más de 6 millones de ERCs putativos, de los cuales decenas de miles están activos en cada tejido o tipo celular, pudiendo localizarse a grandes distancias (hasta 1-2 Mb) de los genes que controlan. La organización 3D de la cromatina es clave para asegurar interacciones específicas entre ERCs y sus genes diana.
La estructura organizada y heterogénea del genoma eucariota
empezó a describirse ya a principios del siglo XX, con la observación de dominios de heterocromatina y eucromatina en el núcleo mediante microscopía electrónica. No obstante, la implementación de técnicas de captura de conformación de cromatina, especialmente Hi-C y sus variantes, ha sido fundamental para revelar la compleja y jerárquica organización del genoma y su vínculo con el control de la expresión génica (Figura 1)
Dominios topológicamente asociados: la unidad básica de organización del genoma
Los estudios pioneros de Dixon y col. revelaron que los genomas se organizan en dominios 3D denominados TADs (del inglés
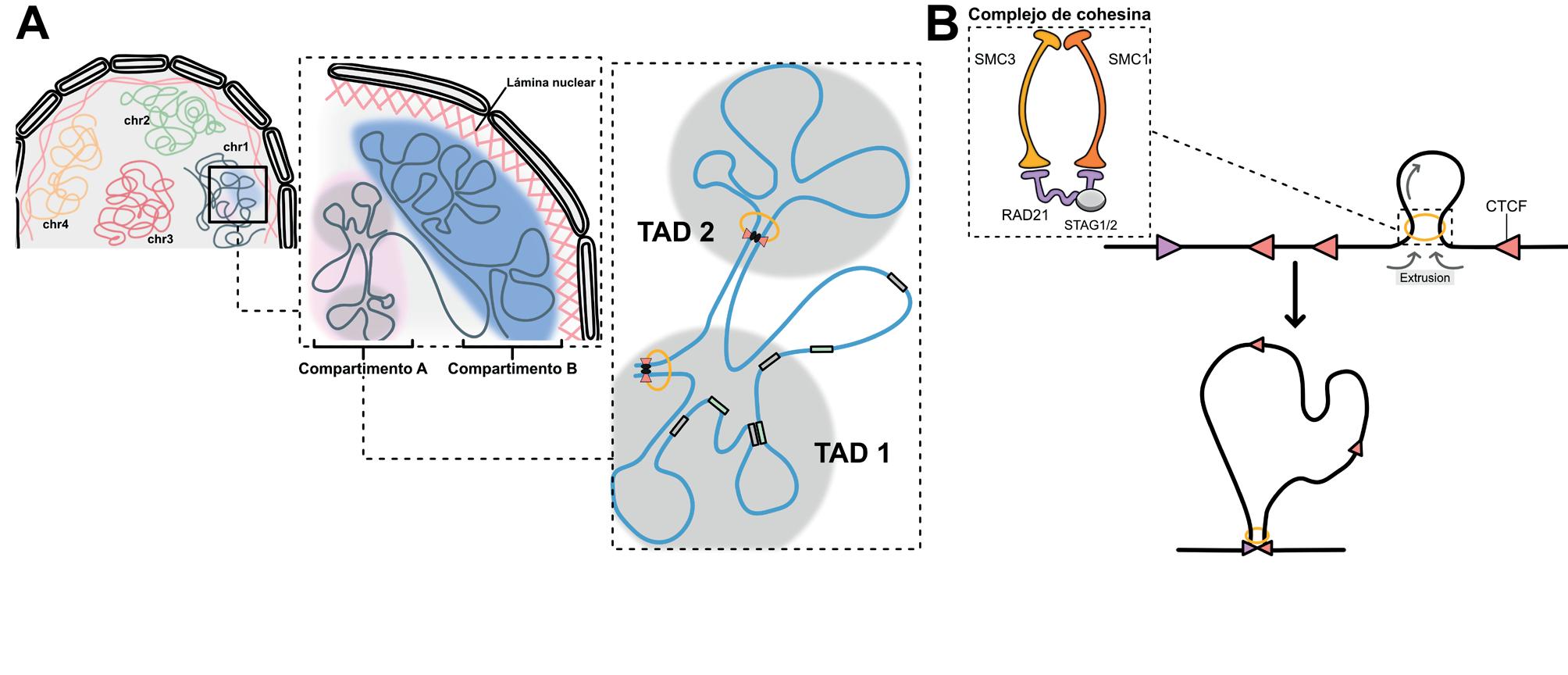
Topologicaly Associated Domains): regiones donde las secuencias de ADN interactúan preferentemente entre sí en comparación con secuencias fuera de ellas. Estos dominios, de 0,5-1 Mb en mamíferos, se han propuesto como delimitadores del entorno regulador de los genes gracias a la actividad aisladora de sus bordes. A su vez, la organización interna de los TADs se basa en el establecimiento de bucles de cromatina que corresponden, en gran medida, a interacciones entre elementos reguladores y promotores, y que tienen un tamaño medio de 100-200 kb. Diferentes estudios apuntan a que los TADs constituyen entidades en gran parte conservadas entre diferentes tipos celulares/ tejidos, mientras que los bucles de cromatina muestran una mayor plasticidad entre los mismos. Además, los TADs pueden subdividirse en dominios más pequeños (subTADs) y/o asociarse en dominios jerárquicamente superiores (superTADs, TAD cliques). A escalas más altas, los TADs se asocian dependiendo del estado transcripcional de los genes que contienen, constituyendo compartimentos que varían considerablemente entre diferentes tipos celulares y tejidos. Así, los TADs “activos” conforman los compartimentos de tipo A, que tienden a ubicarse alejados de la membrana nuclear, mientras que los TADs “reprimidos” se organizan en los compartimentos B, a menudo asociados con la lámina nuclear. Finalmente, los cromosomas se organizan en el núcleo interfásico ocupando territorios específicos y reproducibles.
Mecanismos que contribuyen a la organización de los TADs El establecimiento de los TADs está mediado por un conjunto de proteínas que incluye CTCF (CCCTC binding factor) y el complejo Cohesina. CTCF es un factor
de transcripción con dominios de dedo de zinc que reconoce una secuencia consenso de 20 pb. Sus sitios de unión están particularmente enriquecidos en los bordes de los TADs y su número y afinidad por CTCF correlaciona con el aislamiento de dichos límites. Cohesina es un complejo proteico que media la cohesión entre las cromátidas hermanas durante la división celular, pero que también ejerce importantes funciones de regulación transcripcional. En este contexto, Cohesina abraza el ADN y utiliza la hidrolisis del ATP para desplazar la fibra de cromatina. El modelo actual para el establecimiento de los TADs, se conoce como “loop extrusion”, y postula que los complejos Cohesina se cargan aleatoriamente en la cromatina donde extruden bucles de cromatina hasta colisionar con sitios CTCF ocupados por la proteína con su dominio N-terminal enfrentado al sentido de progresión
del complejo. Así, los bordes de los TADs muestran a menudo sitios CTCF orientados de forma convergente (Figura 1B) Dentro de los TADs, la interacción entre promotores y ERCs depende tanto de la actividad extrusiva de Cohesina como de la acción de enlace de diversos factores de transcripción. La primera se basa en que los promotores génicos también están enriquecidos en sitios de unión de CTCF, aunque estos suelen estar en menor número y ser de menor afinidad que los encontrados en los bordes de los TADs. Dichos sitios CTCF actúan como elementos de anclaje promoviendo la interacción con ERC distales donde Cohesina es retenida por complejos transcripcionales, modificadores epigenéticos y enhancer-RNAs/lncRNAs Entre los elementos de enlace implicados en las interacciones ERCs-promotores cabe destacar YY1, un factor de transcripción con dominios de dedos de zinc
que dimeriza promoviendo contactos entre enhancers y promotores activos. Otro actor importante es Mediator, un gran complejo de 26 subunidades que interacciona por una parte con los factores de transcripción unidos a ERCs, y por otra con el complejo de la ARN polimerasa II. De esta forma, Mediator promueve la expresión génica mediante el reclutamiento de diversos ERCs y la formación de grandes agregados proteicos que se segregan espacialmente en hubs transcripcionales mediante un mecanismo de separación de fases.
Implicaciones funcionales de los TADs Diversos estudios han abordado la cuestión de la relevancia funcional de la organización 3D del genoma en TADs mediante diversas aproximaciones genéticas. Trabajos pioneros a los que contribuyeron diferentes investigadores españoles, como los del laboratorio de José Luis Gomez Skarmeta y
Figura 1
La organización jerárquica de la cromatina. (A) Esquema de los distintos niveles de organización de la cromatina, desde los territorios cromosómicos (izquierda) hasta los TADs y bucles de la cromatina (derecha), pasando por los compartimentos. (B) Esquema del modelo de ”loop extrusion” y del complejo Cohesina. Las cabezas de flecha púrpura y rosa indican los sitios de unión de CTCF y su orientación.
DOSIER CIENTÍFICO
Figura 2
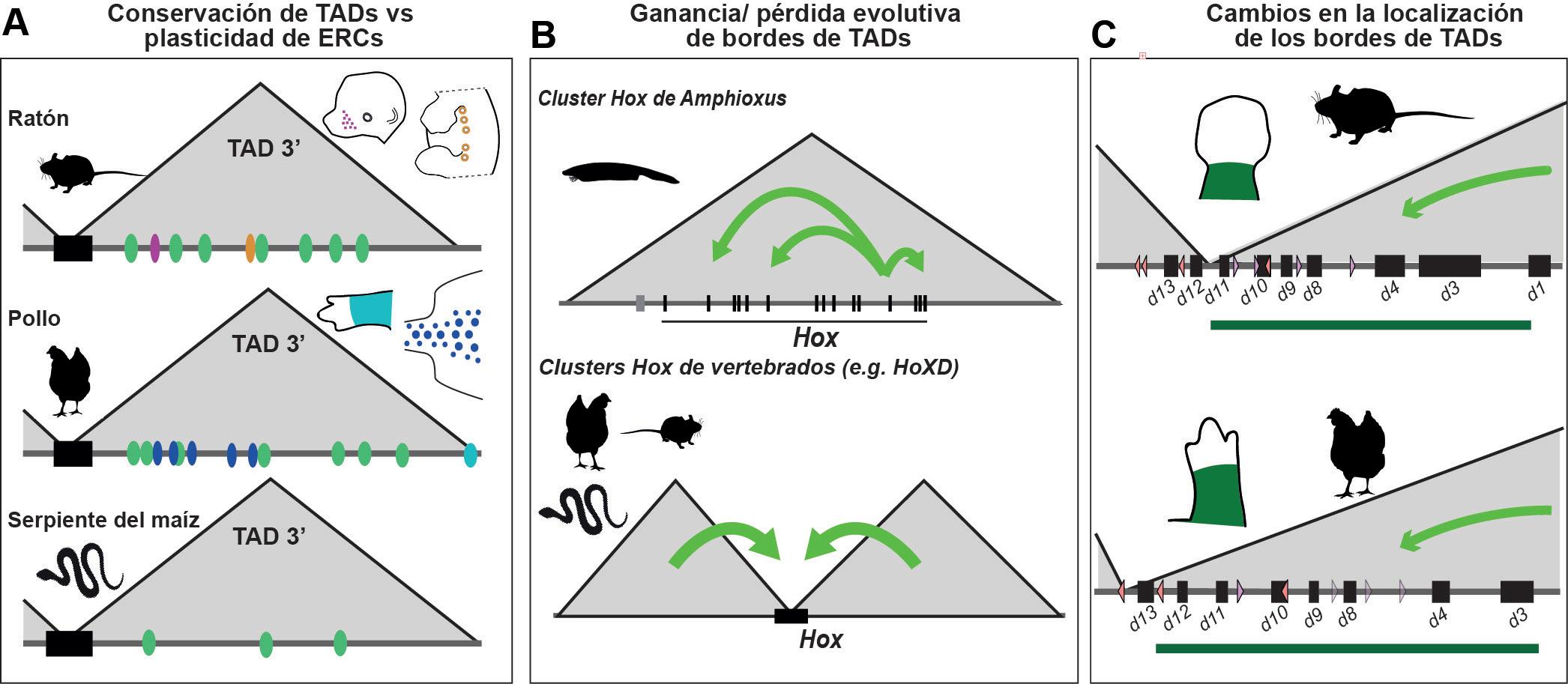
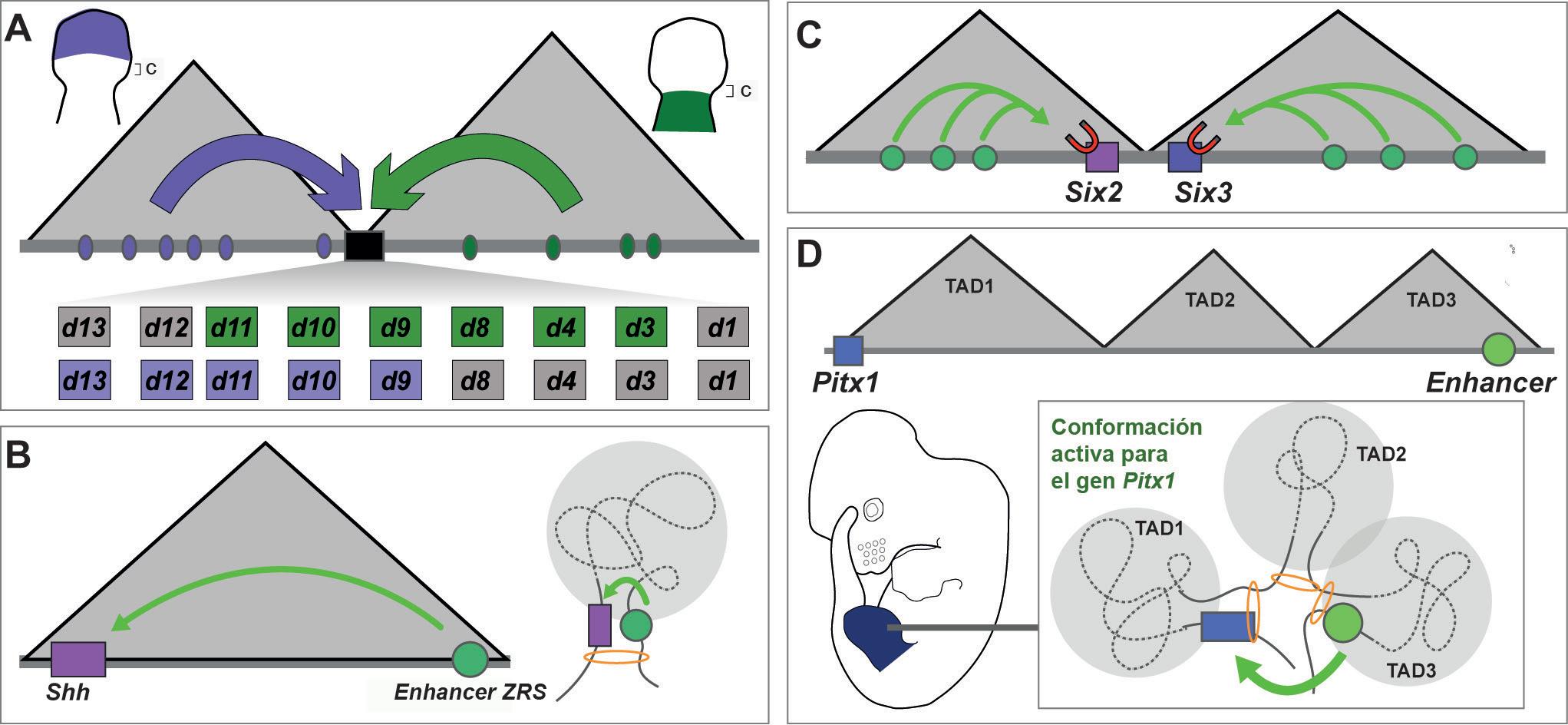
Evolución de los TADs en vertebrados. Esquemas de distintos aspectos de la evolución de los TADs en vertebrados, ejemplificados por el estudio de la organización de los genes Hox. (A) En vertebrados el clúster de genes Hox (representado por el rectángulo negro), se localiza en el borde entre dos TADs (triángulos grises). Dentro de la TAD3’ han evolucionado diferentes enhancers linaje-específicos (p. ej. enhancers para los primordios de glándulas mamarias y vibrisas en mamíferos, enhancers para primordios de plumas y algunos enhancers para extremidades específicos de aves). Incluso ERCs para estructuras evolutivamente conservadas como los somitas (círculos verdes) también muestran una gran plasticidad evolutiva. En serpientes, muchos de estos elementos se han perdido de la TAD3’ y han sido compensados por ERCs ganados dentro del clúster, pero la organización de los TADs se mantiene. (B) La estructura en dos TADs del clúster Hox de vertebrados evolucionó a partir de una estructura de un solo TAD en el clúster Hox ancestral de cordados, representado por el clúster Hox de Amphioxus. En esta especie los enhancers se localizan entre los propios genes Hox (representados individualmente como rectángulos negros). (C) A pesar de su conservación en vertebrados, se han producido cambios sutiles en la organización de los TADs del clúster HoxD de vertebrados, como el cambio de la posición del borde de los dos TADs entre ratón y pollo que se asocia con un cambio en el grupo de parálogos HoxD expresados en el dominio proximal de las extremidades de ambas especies.
de Dario Lupiañez, entre otros, demostraron que los bordes de los TADs permiten aislar la regulación de genes localizados en dominios adyacentes. De hecho, la eliminación de estas regiones, o de sus sitios CTCF, resulta en la pérdida de aislamiento entre TADs y en la ganancia de interacciones ectópicas entre enhancers y promotores. En este contexto, cabe desatacar la implementación de herramientas informáticas, como la desarrollada por el laboratorio de Álvaro Rada Iglesias, capaces de predecir el efecto de dichas variantes en función de la estructura de los TADs y la distribución de ERCs.
Dichas evidencias contrastan con la observación de que la disrupción de CTCF o elementos del complejo Cohesina, a pesar de disrumpir la organización de los
TADs solo repercute en efectos modestos en la expresión génica, abriendo un debate sobre la relevancia funcional de la organización en TADs. Una posible explicación es que estos estudios se realizaron en células terminalmente diferenciadas, que no recapitulan la compleja dinámica de expresión génica de los procesos de desarrollo. De hecho, en ensayos de diferenciación celular la depleción de Cohesina resulta en un impacto mayor sobre la expresión de genes dinámicamente regulados durante dichos procesos. También se ha demostrado que enhancers localizados a largas distancia (particularmente enriquecidos en los dominios reguladores de genes de desarrollo), suelen ser mayormente dependientes de la actividad de proteínas arquitecturales para interaccionar con sus promotores
diana en comparación con ERC proximales. Otra posibilidad es que CTCF/Cohesina contribuyan al establecimiento de las interacciones enhancer-promotor, pero no serían estrictamente necesarias para su mantenimiento. En este modelo, las modificaciones epigenéticas y la actividad de factores de transcripción serían suficientes para mantener el estado transcripcional de los genes, particularmente en células diferenciadas y no proliferativas.
La relación entre la organización de la cromatina y la transcripción también ha sido intensamente estudiada en el contexto de la activación de la expresión génica en el cigoto (Zigote Genome Activation, ZGA por sus siglas en inglés). Después de la fusión de los gametos, la organización en TADs y compartimentos del
Figura 3
Diferentes aspectos de la localización de genes de desarrollo en o cerca del borde de TADs. (A) Como se ha explicado en la Figura 2, los genes HoxD de vertebrados se localizan en el borde entre dos TADs, pudiendo ser regulados por CREs de un dominio u otro según el tejido, activando diferentes combinaciones de los mismos en cada estructura (p. ej. dígitos vs porción proximal de las extremidades). (B) La localización cerca de los bordes de los TADs favorece la interacción entre enhancers y promotores, al quedar ambos en la base del bucle formado entre los sitios CTCF que delimitan el TAD, como se ejemplifica en el caso de Shh y el enhancer de extremidades ZRS. (C) La localización cerca del borde también actúa de forma sinérgica con la actividad de los sitios CTCF para potenciar el aislamiento de los TADs, ya que los promotores de los mismos ejercen como ”atractores” de los enhancers, impidiendo que estos ”crucen” el borde. Cambios en la posición relativa de los genes resulta en una disminución del aislamiento entre TADs como queda demostrado para los genes Six3 y Six2. (D) Algunos enhancers pueden cruzar bordes de los TADs para contactar sus genes diana como demuestra el caso del gen Pitx1. En este caso el TAD intermedio queda segregado de los otros de forma que el enhancer y el promotor de este gen quedan espacialmente cerca en la base de los bucles.
genoma cigótico es muy débil y no se afirma hasta el estadio de 4-8 células (en mamíferos) correlacionando con la activación de la transcripción de los genes del cigoto. Sin embargo, hasta qué punto el establecimiento de la organización 3D de la cromatina es necesaria para la ZGA en mamíferos aún no está clara. Una discusión mucho más exhaustiva de estos aspectos puede encontrarse en la revisión de Bondarieva A y Tachibana K (2024).
La organización del genoma en perspectiva Evo-Devo Los TADs se describieron inicialmente en mamíferos y Drosophila, y posteriormente en todos los linajes eucariotas, e incluso procariotas, analizados hasta la fecha, aún con excepciones. Por ejemplo, los autosomas del nematodo
C. elegans se organizan en TADs, pero presentan bajos niveles de interacción intra-dominio y de aislamiento, coincidiendo con la pérdida secundaria de CTCF a lo largo del linaje de nematodos. Así mismo, el urocordado (el grupo hermano de los vertebrados) Ciona intestinalis y el cnidario Nematostella vectensis carecen de TADs, posiblemente por eventos de pérdida secundaria asociados a la divergencia evolutiva de sus genomas.
Ensayos funcionales y la distribución de sitios CTCF sugieren que el modelo de “loop extrusion” es clave para formar estos dominios en vertebrados y cordados. En cambio, en otros linajes, como dípteros (Drosophila y Anopheles), cnidarios (Hydra vulgaris) o plantas, su organización depende del estado epigenético y transcripcional (y, por tanto, del establecimiento de compartimentos A/B). Estas observaciones suscitan importantes reflexiones sobre la evolución de la organización de los TADs, el papel de CTCF y el control transcripcional mediante ERCs de largo alcance. Para una discusión más detallada de estos aspectos nos remitimos a la excelente revisión de Acemel RD y Lupiañez DG (2023). Enfocándonos en la organización de los TADs en vertebrados, los trabajos iniciales describieron que gran parte de estos dominios se encuentran conservados entre diferentes especies. Esto contrasta con el alto grado de plasticidad en la actividad enhancer entre especies, por lo que los TADs se han postulado como nichos regulatorios donde evolucionan y se ganan/pierden
ERCs. En este sentido, la propia organización interna de los TADs contribuye a determinar el patrón de interacciones de nuevos ERCs adquiridos durante el proceso evolutivo (Figura 2A). Así, reordenamientos genómicos que disrumpiesen TADs serían evolutivamente desfavorecidos, debido a la consiguiente desregulación de los genes contenidos en dichos dominios. En cambio, se ha visto que puntos de ruptura sinténicos localizan preferentemente en bordes de los TADs. No obstante, trabajos recientes han demostrado que existe una mayor variabilidad en la localización de los bordes de los TADs, incluso entre especies filogenéticamente próximas con respecto a lo inicialmente postulado. Uno de los ejemplos más destacados es la transición de la organización del clúster de genes Hox de Amphioxus en una única TAD a una organización de dos TADs en los clúster Hox de vertebrados, permitiendo la implementación de complejas estrategias de regulación en diferentes estructuras embrionarias (Figura 2B). Otros trabajos han relacionado cambios sutiles en la localización y/o en la capacidad de aislamiento de los bordes de los TADs con cambios evolutivos en la regulación génica (Figura 2C)
Pese a estos avances, aún se conoce poco sobre las implicaciones funcionales de la mayoría de los cambios evolutivos en la organización de los TADs. Cruzando fronteras
Desde su identificación, los bordes de los TADs han recibido gran atención como elementos aisladores. Sin embargo, trabajos recientes sugieren un escenario considerablemente más complejo. De hecho, estos bordes están enriquecidos en genes “housekeeping” y/o de desarrollo. Por una parte, los primeros dependen principalmente de elementos
reguladores proximales, sin requerir interacciones con ERCs de largo alcance, lo que contribuye al aislamiento de los bordes. En cambio, la localización de genes de desarrollo en los bordes de los TADs se ha relacionado con varios aspectos de su regulación. En el caso de los clústeres Hox de vertebrados, su localización entre dos TADs les permitiría interaccionar con ambos dominios reguladores adyacentes dependiendo del tejido, y resultaría en patrones de expresión complejos y dinámicos como los descritos para los genes HoxD durante el desarrollo de las extremidades (Figura 3A)
En otros casos, la localización de genes cerca del borde de sus respectivos TADs favorece la interacción con enhancers de larga distancia, de forma que los sitios CTCF que constituyen dichos
bordes actúan tanto como elementos aisladores como de anclaje para interacciones enhancers-promotor (p. ej. Shh, Protocadherins; Figura 3B). A su vez, la localización de estos genes cerca del borde contribuiría a incrementar su capacidad de aislamiento al actuar como atractores de los enhancers de sus propias TADs (Figuras 3B,C). Finalmente, también se han descrito enhancers capaces de interaccionar con sus genes diana a través de bordes de TADs (bystander gene activation), llegando incluso a sortear TADs completos que quedarían segregados aparte, acercando así los TADs del gen y el enhancer en cuestión (Figura 3D)
En resumen, estas evidencias ponen de manifiesto cómo los TADs y sus bordes contribuyen de forma compleja y diversa a las interacciones ERC- promotor.
Para leer más
Acemel RD, Lupiáñez DG. Evolution of 3D chromatin organization at different scales. Current Opinion in Genetics & Development 78 (2023) 102019. https://doi.org/10.1016/j.gde.2022.102019
Bondarieva A, Tachibana K. Genome folding and zygotic genome activation in mammalian preimplantation embryos. Current Opinion in Genetics & Development 89 (2024) 102268. https://doi. org/10.1016/j.gde.2024.102268
Darbellay F, Duboule D. Topological domains, metagenes, and the emergence of pleiotropic regulations at Hox loci. Current Topics in Developmental Biology, 116 (2016) 299-314. https://doi.org/10.1016/ bs.ctdb.2015.11.022
Davidson IF, Peters JM. Genome folding through loop extrusion by SMC complexes. Nature Reviews Molecular Cell Biology 22 (2021) 445-464. https://doi.org/10.1038/s41580-021-00349-7
Maeso I, Acemel RD, Gómez-Skarmeta JL. Cis-regulatory landscapes in development and evolution. Current Opinion in Genetics & Development 43 (2017) 17-22. https://doi.org/10.1016/j. gde.2016.10.004
Pachano T, Haro E, Rada-Iglesias Á. Enhancer-gene specificity in development and disease. Development 149 (2022) dev186536. https://doi.org/10.1242/dev.186536
Última tecnología en electroforesis capilar en gel con detección por fluorescencia al alcance de todos
Qsep1
Cartuchos optimizados según aplicaciones
Genotipado
RFLP
CRISPR
Purif icación de pl ásmidos
Ol igonucl eót idos cf DNA gDNA
Qsep100
Anál isis SSRc Producto de PCR (DNA/RNA)
Control de calidad NGS
JORDI CAMÍ
Presidente del Comité Español de Ética de la Investigación
SEBBM
« La etapa de doctorado es la más idónea para formar en valores éticos
Ismael Gaona Pérez /
Jordi Camí es doctor en Medicina y catedrático en la Universidad Pompeu Fabra. Ha desarrollado su actividad académica entre la Universidad Autónoma de Barcelona y la Universidad Pompeu Fabra, en las que ha ocupado diferentes cargos (delegado del rector, decano y director de departamento). De 1985 a 2005 fue director del Instituto Municipal de Investigación Médica, y ha participado en la gestión de nuevos centros e impulsado infraestructuras de investigación como el PRBB. Ha sido impulsor y primer director de la Fundación Pasqual Maragall entre 2008 y 2020, y actualmente es vicepresidente. Su actividad científica se ha centrado durante más de 35 años en el campo de las neurociencias.
Profesor, ¿cómo ve la situación actual de la investigación en España? ¿Qué cree que hacemos bien y en qué deberíamos mejorar desde el punto de vista de la ética científica?
Primero, quiero aclarar que responderé en relación con un concepto clave: la integridad científica. Este término incluye aspectos de la ética profesional, pero no abarca otros ámbitos éticos de la investigación, como la evaluación de si un proyecto es éticamente apropiado para investigación clínica o para el uso de animales. Es importante no confundir estos temas con los tratados en el campo de la integridad científica.
Todo este es un mundo -el de la ética de la investigación científica- que no lo tratamos en el campo de la integridad. Por ejemplo, el Comité Español de Ética de la Investigación tiene un nombre que, desde mi
punto de vista, puede llevar a confusión, ya que no se centra en los mismos aspectos que los comités éticos de las universidades o los que evalúan ensayos clínicos. De hecho, el Ministerio de Ciencia y Tecnología está apoyando un cambio de nombre, pasando a llamarse Comité de Integridad Científica de España, lo cual delimitará su ámbito de actuación a esta área específica.
Dicho esto, no es un problema exclusivo de España. Todo el sistema de ciencia y tecnología a nivel mundial enfrenta graves retos en cuanto a integridad. Por ejemplo, en el blog Retraction Watch, que sigue estos problemas, se notifican semanalmente entre ocho y diez nuevos casos de plagios, fraudes y otros tipos de malas prácticas científicas. España no es la excepción y también enfrenta este tipo de desafíos.
Si tuviera que puntuar la ética en la comunidad investigadora española del 0 al 10, ¿qué nota le pondría?
No disponemos de datos concretos para establecer una puntuación precisa. Sin embargo, sabemos que hay problemas. Por ejemplo, España está entre los países con mayor número de autores que publican en revistas depredadoras. Además, hemos tenido casos relevantes de integridad científica, como el conflicto de filiaciones con universidades saudíes, que ya ha sido resuelto.
En una entrevista con El País en noviembre de 2024, comentó que la autonomía universitaria debería tener ciertos límites. ¿Qué problemas pueden surgir si esa autonomía no está bien regulada?
Es necesario contextualizar este tema. Si surge un conflicto de integridad que involucra a alguien en una posición de responsabilidad, como un rector o vicerrector ¿quién tiene la autoridad para intervenir? Esto no está claramente regulado, y ahí es donde yo creo que la autonomía universitaria debería tener ciertos límites.
No se trata de restringir la autonomía en su conjunto, sino de garantizar mecanismos externos transparentes que puedan actuar en situaciones críticas. Esto no significa que no confiemos en las universidades, sino que necesitamos un sistema que sea transparente y objetivo, incluso en los casos más delicados. En otros países, existen agencias independientes que supervisan estos conflictos. En España, dependemos principalmente de la autorregulación, que en la mayoría de los casos funciona, pero no siempre es suficiente.
¿Cree que estos problemas son casos aislados o reflejan cuestiones más profundas? ¿Qué se podría hacer para recuperar la confianza de la sociedad?
Los retos éticos en el ámbito científico no sólo surgen por conductas individuales, sino también por la evolución del sistema, que no siempre prevé ciertas prácticas problemáticas. Un ejemplo paradigmático de un desafío reciente es el tema de las filiaciones científicas, que hasta ahora no había sido suficientemente reflexionado ni regulado en los códigos de buenas prácticas científicas, como el desarrollado por ALLEA, la red de academias europeas. Este documento, aunque valioso, no aborda de forma específica la necesidad de ser riguroso con las filiaciones, un aspecto crítico que plantea problemas éticos y de integridad.
En la práctica, siempre ha habido casos de manipulación de datos, currículos inflados de manera artificial y otros tipos de trampas que, aunque en ocasiones son detectados, no siempre salen a la luz. Esto revela una debilidad estructural en el sistema científico,
particularmente si consideramos uno de sus productos principales: las publicaciones. Un alto porcentaje de estas publicaciones no recibe atención ni seguimiento en el ámbito global, y solo una minoría es contrastada y validada por terceros.
El sistema actual, además, permite la proliferación de publicaciones científicas de escaso valor, que pasan inadvertidas y se integran en métricas que no reflejan el impacto real del trabajo. Lo preocupante es que este tipo de comportamiento suele coincidir con perfiles que presentan currículums extraordinarios, algunos de los cuales pueden ser legítimos en el caso de figuras creativas y excepcionales, pero otros levantan sospechas debido a la falta de solidez en sus resultados.
Esto subraya la necesidad urgente de fortalecer los mecanismos de control y reflexión ética, para garantizar que el sistema no solo premie la cantidad, sino la calidad y la integridad del trabajo científico. Además, es fundamental desarrollar herramientas para detectar y prevenir estos comportamientos antes de que afecten la credibilidad y la sostenibilidad de la ciencia como institución.
En el contexto actual de “publicar o perecer”, ¿cómo afecta esta presión a la ética en la investigación? ¿Qué cambios harían falta para mejorar esta situación?
La presión por publicar es un problema grave que ha empeorado con el tiempo, especialmente desde que se comenzó a dar mucha importancia a los indicadores bibliométricos. Esta presión se manifiesta cuando los investigadores compiten por obtener promociones profesionales o para recibir ayudas de investigación. La cantidad de trabajos publicados es un factor clave, pero más recientemente se ha añadido la influencia de los índices de impacto, el índice h y el número de citas recibidas. Estos indicadores han llevado a una competencia feroz que, en ocasiones, promueve malas prácticas, todo con el fin de mejorar la posición de las instituciones en los ranking internacionales. Esta situación ha empeorado la calidad de la investigación, ya que en algunos casos se prioriza la cantidad sobre la calidad de las publicaciones. Ya en el año 1997 propuse cambiar radicalmente la forma en que se evalúan los currículums en el ámbito académico y en las solicitudes de ayudas de investigación (Impactolatría: diagnóstico y tratamiento). Es necesario que se revise solo una selección de trabajos, en lugar de evaluar todo el historial de publicaciones de un investigador. Algunas organizaciones ya están implementando esta idea, pero yo añadiría un nuevo requisito: eliminar la posibilidad de entregar un currículum bibliográfico completo, que suele incluir una lista interminable de publicaciones. Solo permitiendo una selección cuidada de los
trabajos más relevantes se desincentivaría el comportamiento fraudulento, que en algunos casos es excesivamente obvio.
Un ejemplo relevante de esto es lo que ocurrió en los años 80 en la Harvard Medical School. Un médico clínico muy reconocido, Eugene Braunwald, que era coordinador del célebre tratado médico Harrison’s Principles of Internal Medicine, acumuló en su CV docenas de trabajos realizados por un cardiólogo colaborador suyo, John Darsee, unos trabajos que luego se demostró que estaban íntegramente inventados. En esa época, no se revisaban los trabajos de los colaboradores, y el jefe firmaba todo. El fraude que se descubrió en 1981 tuvo un impacto tan grande que motivó a la Harvard Medical School a revisar sus procesos de evaluación. A partir de ese momento, la institución decidió exigir que, en las promociones a profesor asociado o a catedrático, solamente se presentara una selec-
ción de cinco a diez trabajos de todas las publicaciones del autor. Esta catástrofe fue la causa de un cambio conceptual en la manera en que se manejan las promociones y se evalúan los currículums en el ámbito académico.
¿Cómo se puede incluir la ética en la rendición de cuentas de las universidades y centros de investigación?
Estamos hablando de valores, más que de normativa. Por ejemplo, España ha regulado temas clave, como los ensayos clínicos en seres humanos y el uso de animales en experimentación, lo que fue un gran avance. Sin embargo, la ética profesional de quien hace ciencia es difícil de regular, porque se trata de valores que deben enseñarse y ejemplarizarse. Y aquí son importantes dos aspectos. Por un lado, la etapa de doctorado, que considero la más idónea para formar en valores e integridad científica. Durante los últimos 20 años, hemos trabajado en este enfoque. Y, de otro lado, la ejemplaridad. El comportamiento
ético del jefe de grupo o líder de laboratorio es clave. Si el líder es riguroso y fomenta la formación ética de los investigadores, esto influye positivamente. Por el contrario, si el líder tolera trampas, da un mal ejemplo y afecta la cultura ética del grupo. La ejemplaridad es un antes y un después en la formación.
En 2023, se creó el Comité Español de Ética de la Investigación tras más de una década de espera. ¿Qué retos tiene por delante?
El principal reto del Comité es promover buenas prácticas científicas. Su vocación no es inspectora ni sancionadora ante supuestas irregularidades. Nació con el propósito de contribuir, formar, ayudar y fomentar que los cambios en la comunidad científica sean positivos. Por supuesto, también atendemos consultas, siempre que estén fundamentadas. En Europa, la mayoría de los países funcionan bajo este esquema. Por ejemplo, en algunos países como Holanda, el Comité Nacional puede estar presidido por un juez, lo que define un modelo más formal y jurídico. En Estados Unidos, la Oficina de Integridad Científica depende del Senado y tiene competencias inspectoras y sancionadoras.
Con el auge de tecnologías como la Inteligencia artificial, ¿qué retos éticos ve en este nuevo escenario?
Con la Inteligencia artificial, los retos éticos se multiplican en este nuevo escenario. Desde nuestra perspectiva, hemos emitido tres recomendaciones clave, además de comunicados sobre casos mediáticos. El concepto de “recomendación” se traduce en textos cortos y directos que sirven como referencia para la comunidad científica.
La más reciente aborda la gestión de conflictos de integridad. Si bien en España muchas instituciones han desarrollado códigos de buenas prácticas científicas, esto no es suficiente. Es un paso necesario, pero es crucial apoyar a los centros para que entiendan que, ante un conflicto de integridad, deben actuar con la mayor transparencia y cuidado. Cuando se producen denuncias, es fundamental proteger a ambas partes, ya que quienes se atreven a exponer irregularidades suelen enfrentarse a consecuencias personales muy serias.
Las dos restantes están relacionadas con las filiaciones científicas (emitida en julio de 2023 a petición de la ministra), y que surgió tras los casos polémicos relacionados con filiaciones académicas en universidades saudíes. Y la última, con la Inteligencia artificial, un tema más complejo que se centró en formular ideas y directrices éticas para científicos que desarrollan algoritmos avanzados. Este documento enfatiza
la responsabilidad profesional en el diseño y uso de la Inteligencia artificial, instando a los investigadores a priorizar principios éticos en su trabajo.
Nuestro objetivo no solo es aportar claridad en estos temas, sino también fortalecer la confianza en la comunidad científica, fomentando un entorno donde la ética sea parte integral del avance tecnológico y del conocimiento.
¿Cómo puede ayudar la ciencia abierta a reforzar la ética en la investigación?
La ciencia abierta contribuye porque expone más. Pasa de ser un mundo cerrado, entre las paredes de los laboratorios, a compartir su conocimiento con la sociedad. Una característica propia de la integridad científica es que, a medida que la sociedad evoluciona, exige conocer las consecuencias de lo que hace la comunidad científica. Los científicos no son superiores al resto de los mortales y, como en todos los colectivos, pueden existir casos de conductas inapropiadas. La ciencia abierta aumenta la exigencia y el escrutinio. Además, aunque determinados sectores de la ciencia, como aquellos enfocados a la innovación, son los principales beneficiarios de este modelo, su alcance puede extenderse para fortalecer la transparencia y la confianza pública.
Por último, una cuestión más personal (como aficionado a la magia). En su libro El cerebro ilusionista (RBA), explica cómo los magos aprovechan las limitaciones del cerebro para crear ilusiones sorprendentes. ¿Qué nos dicen esas ilusiones mágicas sobre cómo el cerebro interpreta el mundo?
La percepción de la realidad es un tema clave en la cognición. Evolutivamente, el cerebro humano ha aprendido a gestionar la realidad de forma eficiente, interpretándola con muy poca información. Hemos desarrollado la capacidad de anticiparnos porque el procesamiento completo de la realidad es lento, y el cerebro utiliza estrategias para llenar esos vacíos. A veces estas estrategias nos llevan a errores, pero en general nos permiten adaptarnos y sobrevivir.
La magia, cuyos efectos aparentan ir en contra de las leyes de la naturaleza, es un arte que explota estas estrategias cognitivas. Los magos, de manera empírica, han aprendido a “hackear” el cerebro, construyendo una realidad paralela, lógica y consistente, para que el desenlace nos sorprenda, por imposible. Es interesante cómo la magia revela las estrategias que el cerebro utiliza para interpretar el mundo, por este motivo, consideramos a la magia como una suerte de caja de herramientas que nos permite estudiar con nuevas estrategias cómo funciona el cerebro.
Master of Bioactive Molecules
Inhibitors • Screening Libraries • Proteins
MedChemExpress: Industry Leading Protein Manufacturer
Diverse Products
13,000+ commonly used recombinant protein products
Various Grades
RUO grade, GMP grade, Animal-free grade.
MCE Recombinant Proteins Include:
● 1,700+ Cytokines
● 3,200+ Enzymes & Regulators
● 4,000+ Receptor Proteins
● 120+ Fc Receptors
● 400+ CAR-T related Proteins
● GMP-grade Proteins
Superior
Biological Activity
High purity & Low endotoxin levels.
● 600+ Immune Checkpoint Proteins
● Animal-free Recombinant Proteins
● Recombinant Protein Expression Service
Upgrading Drug Development: 3D Cell Culture
Basement Membrane Matrix
A variety of basement membrane matrices, composed mainly of various growth factors and extracellular matrix components.
● Low endotoxin levels
● High security
● Strict dialysis sterilization
About MedChemExpress
Organoid Culture Medium
Dozens of organoid culture media are provided for researchers, including media tailored for tumor and normal tissue organoids.
Excellent lot-to-lot consistency
Low endotoxin levels
High cytokine activity
www.MedChemExpress.com
MedChemExpress (MCE) is a global leader in research chemicals and bioactive compounds. Our extensive product portfolio includes a wide range of active molecules, such as inhibitors, compound libraries, and recombinant proteins. Our experienced and dedicated team is committed to meeting your needs. We are a capable and reliable partner for laboratory and research applications.
Headquarter in U.S, Warehouse in China, India, and Europe
MedChemExpress Europe
30,000+ research institutions and drug discovery companies customers
50,000+ literature citations
CIENCIA, ÉTICA Y ESTÉTICA: EL EQUILIBRIO NECESARIO
EN LA INVESTIGACIÓN
Ismael Gaona Pérez
Filiaciones, falsas autorías, manipulación de datos... La ciencia afronta en la actualidad un dilema trascendental para preservar su credibilidad y legitimidad como pilar de conocimiento y bienestar social. Este escenario plantea preguntas fundamentales: ¿Dónde están los límites éticos? ¿cómo puede la comunidad científica garantizar la transparencia de los estudios? ¿Qué medidas deben implementarse para combatir las malas prácticas?
“La ciencia sin ética es ciega, pero la ética sin ciencia es estéril”. Esta frase, atribuida al bioquímico norteamericano Van Rensselaer Potter, cobra plena vigencia en un momento en el que la investigación y la innovación avanzan de forma exponencial, ya bien entrado el siglo XXI. Su autor, fallecido en 2001, reflexionó a mediados del siglo XX sobre cómo el conocimiento científico sin filtros ni límites podría convertirse en un grave problema para la credibilidad y la reputación de la comunidad investigadora.
Ejemplos históricos de fraude como el protagonizado por Andrew Wakefield, cuyo artículo publicado en The Lancet en 1998 vinculaba falsamente la vacuna contra el sarampión con el autismo, muestran cómo la falta de rigor puede provocar una crisis global de confianza, sembrando el miedo en millones de familias y poniendo en riesgo la salud pública. O el caso de Haruko Obokata, una científica japonesa autora de un artículo en Nature en 2014, en el que aseguró haber descubierto una técnica revolucionaria para reprogramar células madre. Investigaciones posteriores revelaron que había manipulado datos e imágenes, lo que derivó en el desprestigio de su trabajo y un daño a la credibilidad científica en general.
Hoy, con la inteligencia artificial, la edición genética o el análisis de datos masivos, que sostienen disciplinas como la biología, la farmacología o la epidemiología, entre otras, el control ético es más necesario que nunca. Desde los tiempos de Sócrates hasta ahora, persisten preguntas fundamentales: ¿Hasta dónde podemos llegar? ¿Quién decide qué está permitido y qué no?
La ciencia y la ética comparten un objetivo esencial: ambas buscan lo mejor para el ser humano. Pero debe haber límites que eviten que la búsqueda del conocimiento comprometa derechos fundamentales o erosione la confianza de la ciudadanía. Casos como el uso indebido de datos personales por parte de empresas tecnológicas o investigaciones no reguladas subrayan los peligros de una ciencia sin normas claras.
Garantía de confianza
La ética es un pilar fundamental para garantizar la integridad y la credibilidad del conocimiento científico. La confianza en los resultados de la ciencia depende de la transparencia, la honestidad y el rigor
con los que los investigadores gestionan los datos y resultados. El fenómeno de las malas prácticas científicas es una preocupación global, y España no es la excepción, como señala el profesor Jordi Camí, presidente del Comité Español de Ética de la Investigación. Los datos disponibles respaldan la gravedad del problema.
Un estudio difundido en 2023 por el Science Media Centre, el primero realizado en España sobre la percepción, actitudes y experiencias respecto al fraude científico en el ámbito biomédico, reveló cifras alarmantes. Según los resultados, el 43% de los 403 investigadores encuestados (27 preguntas) admitió haber incurrido en alguna forma de mala práctica científica.
Entre las malas prácticas más comunes, la falsa autoría fue reportada por el 35% de los participantes. Además, un 10% de los encuestados admitió haber omitido el consentimiento informado en investigaciones, y un 3,6% reconoció haber falsificado o manipulado datos en al menos una ocasión. El estudio, publicado en la revista Accountability in Research, también mostró que el 78,8% de los científicos entrevistados conocía al menos un caso de conducta poco ética en su entorno laboral, resultados que subrayan la necesidad de contar con estructuras que promuevan la integridad científica y detecten las malas prácticas a tiempo.
“Es preocupante constatar que, aunque estos resultados necesiten ser confirmados, existe un entorno donde las malas prácticas parecen estar, en cierta medida, consentidas. Esto demanda una respuesta urgente. Desde hace años insistimos en la necesidad de sensibilizar sobre este problema en España. También sería esencial implementar sistemas de análisis de malas prácticas y establecer medidas disciplinarias en los casos más graves”, afirmó Pere Puigdomènech, profesor de investigación emérito del Centro de Investigación en Agrigenómica (CRAG) y presidente del Comitè per a la Integritat de la Recerca a Catalunya (CIR-CAT).
Por su parte, Eduard Aibar, catedrático de Estudios de la Ciencia y Tecnología de la Universitat Oberta de Catalunya (UOC), valoró el informe señalando que, aunque la muestra no sea representativa, “los resultados son muy fiables y significativos”. Aibar destacó que “el tratamiento estadístico es riguroso, y tanto el
diseño de la investigación como la discusión de los resultados están sólidamente fundamentados en estudios internacionales previos”. Para Aibar, el principal hallazgo es “la alta prevalencia del fraude científico en España, donde cuatro de cada diez investigadores admiten haber incurrido en conductas fraudulentas”.
En cuanto al contexto, a nivel global, se estima que aproximadamente el 4% de los artículos científicos publicados contienen algún tipo de mala conducta.
El fraude no ocurre en un vacío. Las presiones por publicar, obtener financiación y avanzar en la carrera profesional son factores clave que alimentan la tentación de manipular datos o cometer plagio. La “carrera por publicar” está profundamente arraigada en el ámbito académico, ya que las publicaciones son una medida clave de éxito y un requisito para acceder a la financiación. Según la Universidad de Hong Kong, el 25 % de los investigadores reconoce haber sentido presión por publicar para progresar profesionalmente, lo que a menudo lleva a prácticas poco éticas, como la manipulación de los resultados, el plagio o la publicación de estudios incompletos.
Situación en el mundo y en España
Aunque las cifras en nuestro país no difieren significativamente de este promedio, uno de los principales problemas ha sido, históricamente, la falta de un organismo nacional dedicado a supervisar la integridad científica. En España, históricamente han existido redes colaborativas, oficinas, y protocolos dispersos, a diferencia de otros países europeos que cuentan con unidades específicas de integridad científica. Por ejemplo, el Reino Unido dispone
de la UK Research Integrity Office, que regula las prácticas científicas y fomenta la investigación ética. En Alemania, la Deutsche Forschungsgemeinschaft (DFG) establece guías detalladas sobre buenas prácticas científicas. En Estados Unidos, los National Institutes of Health (NIH) y la Office of Research Integrity (ORI) han implementado normativas claras para abordar casos de mala conducta científica, como el fraude en la publicación de resultados, la manipulación de datos, entre otros.
En cuanto a España, la Red de Comités de Ética de las Universidades y Organismos Públicos de Investigación (RCE) juega un papel clave en garantizar que los proyectos de investigación cumplan con los principios éticos más estrictos. Sus orígenes se remontan a 1995, cuando Marius Rubiralta, entonces vicerrector de Investigación de la Universidad de Barcelona (UB), y María Casado, directora del Observatorio de Bioética y Derecho, identificaron la necesidad de un organismo que evaluara los aspectos éticos de los proyectos de investigación presentados a convocatorias nacionales e internacionales. En ese momento, no existían normativas ni leyes autonómicas o nacionales que regularan estas comisiones. Así, la Universidad de Barcelona fundó la primera Comisión de Bioética en España.
Con el tiempo, esta iniciativa se extendió de manera informal, creando una red de colaboración entre universidades públicas. Inicialmente, incluyó a la Universidad Autónoma de Barcelona y la Universidad de Murcia, para luego ampliarse a otras instituciones como la Universidad de Granada y la Universidad del País Vasco. Este esfuerzo culminó en 2002 con la formalización de la Red de Comités de Ética de las Universidades Públicas Españolas (RCEUE), respaldada por el Ministerio de Educación y Ciencia.
Tras el primer encuentro en Sitges (2002), la red adquirió un carácter nacional con la incorporación de los Organismos Públicos de Investigación (OPIs). Esto permitió que la red se convirtiera en un referente más amplio, integrando universidades y centros de investigación públicos de todo el país. Bajo el liderazgo del Consejo Superior de Investigaciones Científicas (CSIC), la red adoptó el nombre Red de Comités de Ética de las Universidades y Organismos Públicos de Investigación de España (RCE), fortaleciendo la colaboración y aumentando su impacto en el ámbito científico nacional.
Hoy en día, 87 universidades y centros de investigación conforman la Red de Comités de Ética de las Universidades y Organismos Públicos de Investigación de España (RCE). Su cometido incluye, entre otros, revisar y aprobar proyectos de investigación para garantizar que cumplen con los estándares éticos establecidos.
Por otro lado, la Confederación de Sociedades Científicas de España (COSCE) también ha asumido un papel activo en el debate ético. En un artículo publicado en su web en 2023 (La ética de la ciencia), subrayan la importancia de su papel tanto para defender las posiciones científicas como para ejercer una labor pedagógica hacia los científicos y la sociedad. En sus conclusiones, destacan varios puntos clave sobre su cometido: colaborar en la toma de conciencia sobre los requerimientos éticos de la actividad científica, participar en los debates relacionados con la adopción de legislación que afecta la actividad científica, contribuir a la definición de Códigos de Buenas Prácticas Científicas en las instituciones de investigación en España, reflexionar sobre los cambios en la evaluación de la actividad científica y en la gestión de las publicaciones en el entorno digital actual, y tomar parte en los debates sobre ideas o aplicaciones científicas y sus implicaciones.
El Comité Español de Ética de la Investigación
Y, más recientemente, el Comité Español de Ética de la Investigación (CEEI), constituido en 2023 con 12 años de retraso respecto a su inclusión en el
marco de la Ley de la Ciencia, cambiará de nombre a “Comité de Integridad Científica de España”, según anunció su presidente, Jordi Camí, en una entrevista para este número. Cuando se aprobó su creación, España no contaba con un órgano respaldado por el Gobierno que garantizara que la investigación se realizara bajo los más altos estándares éticos, cumpliendo con normativas nacionales e internacionales.
Configurado como un órgano colegiado de ámbito estatal, independiente y de carácter consultivo, el CEEI tiene competencias en integridad científica, investigación responsable y ética en la investigación científica y técnica.
Entre sus funciones principales destacan: emitir informes, propuestas y recomendaciones sobre temas relacionados con la ética profesional en la investigación científica y técnica (hasta la fecha ha publicado dos informes y tres recomendaciones); establecer principios generales para la elaboración de códigos de buenas prácticas en la investigación científica y técnica, incluyendo la resolución de conflictos de intereses entre actividades públicas y privadas (estos códigos serán desarrollados por los Comités de Ética de la Investigación y el Comité de Bioética de España);
representar a España en foros y organismos supranacionales e internacionales relacionados con la ética de la investigación, salvo en materia de bioética, donde la representación corresponde al Comité de Bioética de España; e impulsar la creación de comisiones de ética vinculadas a los agentes ejecutores del Sistema Español de Ciencia, Tecnología e Innovación.
En cuanto a la estética: la retractación
Adicionalmente, las malas prácticas no solo generan una imagen pésima del sistema científico, sino que también implican una carga económica significativa. Según un informe de Science Integrity (2021), las retractaciones científicas pueden costar hasta 50.000 dólares por artículo retirado, considerando gastos como la corrección de tratamientos médicos, la invalidación de ensayos clínicos y la pérdida de confianza pública. Pese a estas cifras, el número de retractaciones científicas ha crecido notablemente en las últimas décadas.
Un estudio de la Universidad de Santiago de Compostela y la Universidad de Yale (2022) señala que las retractaciones han aumentado un 10 % anual en los últimos diez años, siendo más frecuentes en la investigación biomédica, que representa el 77 % de los casos.
¿Qué podemos hacer?
La ética es un concepto complejo que ha estado en constante evolución. Desde la promulgación de la Ley de la Ciencia en 1986, que organizó y promovió la investigación científica en España, hasta
la implementación del Real Decreto sobre Ensayos Clínicos en 2006, que marcó un hito en la regulación de los ensayos en seres humanos, la legislación ha buscado establecer las bases para una práctica científica ética. Asimismo, la experimentación con animales ha sido objeto de regulación, exigiendo a los investigadores justificar la necesidad de utilizar animales en sus estudios y cumplir estrictos protocolos de bienestar animal.
Sin embargo, más allá de la regulación técnica, existe una dimensión ética que no puede ser legislada de manera tan directa: los valores que guían la conducta de los científicos. La ética científica no se limita al cumplimiento de las normas, sino que incluye también los principios personales de quienes la practican, lo que plantea un desafío aún mayor.
“Los valores son cuestiones profundamente subjetivas y, por ello, no pueden ser fácilmente regulados. Se transmiten principalmente a través del ejemplo y la formación continua, lo que pone de manifiesto la importancia de la educación en valores a lo largo de toda la carrera científica. Es en el doctorado, etapa clave en la formación de los futuros investigadores, donde se deben cultivar no solo los conocimientos técnicos, sino también el carácter y la ética profesional. Durante este período, los jóvenes científicos aprenden no solo a hacer ciencia, sino a ser científicos responsables y comprometidos con los principios éticos”, propone Jordi Camí. Además, subraya que la ejemplaridad de los líderes de investigación es un factor determinante para fomentar una cultura ética en el entorno científico.
Qué dice la sociedad
En el ámbito sanitario, los fraudes, como el caso de Wakefield, tienen un impacto directo y grave en la salud pública. Las decisiones basadas en investigaciones fraudulentas pueden dar lugar a políticas de salud ineficaces y costosas, como ocurrió con el movimiento antivacunas, cuyas consecuencias todavía persisten en la actualidad. Además, el fraude científico erosiona la confianza pública en la ciencia, un recurso fundamental para abordar retos globales como la crisis climática, las pandemias y el desarrollo de tecnologías emergentes. Si la ciudadanía comienza a cuestionar la veracidad de los estudios científicos, se vuelve más vulnerable a la desinformación y las teorías conspirativas, lo que agrava los problemas sociales y científicos.
Pese a los continuos esfuerzos de organismos, universidades e instituciones por mejorar la percepción de la ciencia en España, hay datos que invitan a reflexionar. Según un estudio de la Fundación BBVA (2023), el porcentaje de españoles que considera que la ética no debería poner límites a los avances científicos ha aumentado del 47 % en 2012 al 57 % en 2023. Este incremento evidencia un cambio en la percepción pública que merece un análisis más profundo.
El Virus Oropuche (OROV) es un arbovirus perteneciente a la familia Peribunyaviridae, aislado por primera vez en 1955 en Trinidad y Tobago, en una región llamada Oropouche. Se transmite a animales; incluyendo humanos, principalmente a través de la picadura de la mosca de la arena Culicoides paraensis y el mosquito Culex quinquefasciatus; insectos ampliamente distribuidos en América y en regiones tropicales.
La mayoría de los casos de fiebre de Oropouche son leves y generalmente se resuelven en un lapso de 2 a 3 semanas; sin embargo, en situaciones excepcionales, el OROV puede provocar problemas neurológicos o afectar al feto provocando abortos o nacimientos con anomalías congénitas.
Lo que más preocupa actualmente es la expansión de este virus. Durante 2024 se notificaron 16.239 casos confirmados en once países y territorios de la Región de las Américas (Barbados, Bolivia, Brasil, Canadá, Colombia, Cuba, Ecuador, USA, Guyana, Islas Caimán, Panamá y Perú, incluidas 4 defunciones en Brasil.
También se han descrito 30 casos importados en Italia, España y Alemania; considerándose las primeras importaciones documentadas de OROV
a Europa continental. Este hecho es de gran importancia si tenemos en cuenta que las especies de Culicoides se encuentran entre los vectores más abundantes de arbovirus en Europa.
Las arbovirosis son difíciles de diagnosticar, especialmente durante las primeras etapas. La sospecha diagnóstica se basa en las características clínicas y antecedentes de viaje a zona endémica. El diagnóstico específico y la confirmación de casos requieren el análisis de laboratorio que en muchos casos se basa en la detección el material genético del virus (ARN) por métodos moleculares (RT-PCR) durante la fase aguda de la enfermedad.
Acorde a la estrategia establecida por la Organización Mundial de la Salud y con el objetivo de contribuir al diagnóstico y vigilancia de los casos de OROV, Certest ha desarrollado un producto basado en RT-PCR para la detección altamente sensible y especifica de OROV. Este producto ha sido verificado y validado con muestras clínicas (suero) de personas previamente confirmadas positivas a infección por OROV pudiendo contribuir significativamente a la gestión efectiva de la salud pública y la atención médica.
EL MODELO DE
INVESTIGACIÓN EN EL FUTURO DE LA UNIÓN EUROPEA
Alonso Rodríguez Navarro
Profesor
emérito, Universidad Politécnica de Madrid
La Unión Europea (UE) ha tenido un éxito incuestionable. Frente a nuestros más directos competidores, EEUU y China, la UE presenta notables ventajas en bienestar social. Pero para financiar nuestro modelo social, la UE tiene que incrementar mucho su productividad, y esto no va a ocurrir si la UE no introduce cambios que mejoren su capacidad innovadora, especialmente en las tecnologías con mayor impacto económico. En los últimos 20 o 30 años, la capacidad innovadora de la UE ha ido decayendo. Actualmente, en las tecnologías económicamente más importantes, las diferencias con EEUU y China empiezan a ser abrumadoras y amenazan a la UE con una lenta agonía económica. En esta decadencia, aunque multifactorial, una investigación científica deficiente ha jugado un papel central, por una causa que describió Peter Drucker para los negocios: “You can’t manage what you can’t measure”. Aunque en el caso que nos ocupa habría que cambiar la condición de “no puede
medir” por “no sabe medir” o “no quiere medir correctamente”.
Este artículo describe la historia de este proceso y explica las causas principales que lo han originado. También explica cómo España se adhirió a él, con escasas razones para hacerlo.
El informe Draghi
En septiembre de 2024 se publicó un importante informe del economista y político Mario Draghi: “The future of European competitiveness”. Este informe, así como otros del Fondo Monetario Internacional en su blog (IMF BLOG) y en su Regional Economic Outlook, dejan claro el déficit de innovación en la UE. Para corregir esta situación, el Informe Draghi propone medidas que conllevan inversiones mil millonarias. Pero estas inversiones y otras medidas podrían incumplir los objetivos si no se mejora la investigación. De hecho, el informe hace una advertencia crucial sobre la investigación en la UE: “insufficiently focused on disruptive innovation”.
En julio de 2012 Mario Draghi, presidente del BCE en aquel momento, salvó al euro con tres palabras que describían lo que haría el BCE para salvarlo: “whatever it takes”, seguidas de una reflexión: “and believe me, it will be sufficient”. En esta ocasión, la solución no puede ser tan inmediata, pero sería prudente considerar el informe de Mario Draghi, que es tan certero como fue su intervención en 2012, especialmente en lo relativo a las innovaciones disruptivas.
La falsa paradoja europea y su adopción en España
Algunos aspectos de la política científica en la UE han sido un total desacierto, y el más notable ha sido la falsa Paradoja Europea, basada en una ilusoria excelencia científica. El déficit en innovación que ahora es evidente la Comisión Europea (CE) ya lo intuía en un “White Paper” sobre “Growth, Competitiveness, Employment” de 1994. En el mismo año, “The First European Report on Science and Technology Indicators” observa la emergencia de una “research
paradox”, debida a una insuficiente transferencia de los resultados de la investigación al sector productivo. Al año siguiente, un “Green Paper on Innovation” bautiza esa conjetura como “European paradox” y la incorpora como base de la política científica en la UE que, de acuerdo con ella, hay que centrar en mejorar la transferencia de los resultados de una investigación que es excelente. Lamentablemente, la paradoja se sustenta en comparaciones de los números de publicaciones científicas y patentes, ignorando que el número de publicaciones no determina la excelencia científica.
En la misma línea de éxito científico, en 2017, un supuesto High Level Group, nombrado por la CE, inicia su informe, “LAB – FAB –APP. Investing in the European future we want”, redefiniendo la paradoja: “the contrast is striking between Europe’s comparative advantage in producing knowledge and its comparative disadvantage in turning that knowledge into innovation and growth”. A continuación crea el eslogan “Europe is a global scientific powerhouse”, y desde entonces, eslóganes de este tipo han sustituido a la Paradoja Europea en la política científica de la UE. Por ejemplo, un documento del Parlamento Europeo de 2019 denominado “The EU strategy for international cooperation in research and innovation” comienza con la frase: “The EU is a global leader in research, development and innovation”.
En este clima de suficiencia sin base empírica, hay que esperar hasta un documento de la CE de septiembre de 2024: “ Align, Act, Accelerate. Research, Technology and Innovation to boost European Competitiveness ”, para que la idea de “ Europe’s comparative advantage in producing knowledge ” aparezca cuestionada. Algo
increíblemente tardío, ya que las publicaciones académicas que dejan sin soporte a la Paradoja Europea y a la idea del éxito científico empiezan mucho antes. En 2006 y 2007 aparecen las primeras publicaciones de investigadores italianos, unos años después son seguidas por otras de investigadores españoles, y actualmente ya son muchas.
Siguiendo la idea de la UE, alrededor de 2008, España incorpora la idea de la excelencia investigadora sin soporte empírico, y la idea crece como una bola de nieve. Dos ejemplos son la creación de los Campus de Excelencia Internacional y la intervención del Presidente del Gobierno en la Cumbre Iberoamericana de México de 2014. El último capítulo ha sido la Ley de la Ciencia, la Tecnología y la Innovación de 2022, que se redacta a partir de la idea de que nuestra investigación “ha alcanzado estándares de excelencia investigadora perfectamente homologables a su posición económica y geopolítica en el panorama internacional”.
Como explico más adelante, las medidas rigurosas de la ciencia tienen cierta complejidad matemática y estadística, pero hay publicaciones que usan métodos sencillos para demostrar el desacierto de las manifestaciones de éxito descritas. El más sencillo es contar el número de avances científicos que merecen un premio Nobel y compararlo con el número de publicaciones. Haciendo esto se encuentra que en la UE por cada premio Nobel en Física, Química o Fisiología/Medicina el número de publicaciones es cuatro veces más alto que en EEUU. Incluso en un país tan avanzado como Alemania, la ratio es 2,5 veces mayor que en EEUU. En España es aún peor. Atendiendo al número de publicaciones, si España fuera homologable a EEUU habría tenido seis
o siete premios Nobel en el siglo XXI y habrían sido tres si fuera homologable con el Reino Unido, Alemania o Francia.
La medida de la ciencia Medir el éxito de la investigación científica no es fácil porque lo que se produce, el avance del conocimiento, es intangible. Pero hay que superar las dificultades, porque, como ya se ha dicho: “no se puede gestionar lo que no se puede medir”. Pero a las dificultades a veces se añaden errores evitables, como ilustran dos ejemplos con España y Japón.
La Figura 1 reproduce un detalle de un documento del Gobierno de España con motivo de la Semana de la Ciencia y la Innovación en 2021. El documento induce a pensar que como el 2% de nuestras publicaciones está en la élite del 1% mundial más citado, nuestra ciencia es excelente, lo que no es cierto. En cualquiera de los informes bisanuales sobre investigación de la CE, el valor de ese mismo dato estadístico está por debajo del 0,8%, como también muestra la Figura 1. No me corresponde a mi explicar el origen de ese 2% que publica el Gobierno de España y su errónea asociación con la excelencia científica, pero es fácil suponer que se trata de un error al contabilizar los trabajos de colaboraciones internacionales, haciéndose un lío con los indicadores fraccionario y multiplicativo. Algo así como utilizar un termostato con escala Fahrenheit creyendo que es Celsius, poner el climatizador de casa a 20 grados para estar confortable y ver que el agua del vaso se congela.
El caso de Japón tiene más relevancia bibliométrica. En cualquiera de las evaluaciones de la CE referidas antes (como en el panel de la derecha en la Figura 1), la evaluación de Japón es mucho peor que la de España y correspondería a un país en vías
Figura 1
Alabanza de la ciencia en España. A la izquierda, reproducción parcial de un documento del Gobierno de España; a la derecha, reproducción parcial de la Figura 3.1-22 (publicaciones en el 1% más citado) del informe de la CE: Science, Research and Innovation Performance of the EU, 2024.
de desarrollo. Este error proviene de idear un parámetro estadístico y convertirlo en indicador científico sin validarlo. Porque Japón ha tenido 17 premios Nobel y globalmente es el país con mayor número de familias de patentes triádicas en relación al PIB. Evidentemente eso es un éxito y las malas evaluaciones se deben a que una parte importante de la investigación en Japón se dirige a las innovaciones incrementales, y, en estos casos, la investigación no puede evaluarse con un solo parámetro.
El progreso en la frontera del conocimieto
Los resultados de la investigación dirigida a las innovaciones incrementales son más tangibles y
fáciles de evaluar que los resultados de la investigación que persigue innovaciones disruptivas, que hay que juzgarlos en función del progreso en la frontera del conocimiento. Aunque explicar los métodos de evaluación queda fuera de los objetivos de este artículo, hay un aspecto relativamente fácil de explicar que permite juzgar el papel de la UE y España en la investigación dirigida a empujar la frontera del conocimiento. Esta facilidad aparece porque los sistemas de investigación, países o instituciones, se comportan como sistemas complejos, más fáciles de evaluar que la simple suma de investigadores.
Un ejemplo ilustra la diferencia. Supongamos una carrera ciclista
donde se premia a los equipos y no a los corredores aislados. Terminada la carrera, para determinar el equipo vencedor, se puede utilizar un método paramétrico, sumando o haciendo alguna otra operación con los tiempos de los ciclistas, o se puede utilizar un método no paramétrico, basado en el número de orden de los ciclistas de cada equipo en la clasificación general. Con este procedimiento, los números de orden en la clasificación general de los ciclistas de un mismo equipo serán series aleatorias de números en orden creciente entre los del mejor y peor clasificado. Esto es así porque un equipo puede tener cualquier tipo de proporción de ciclistas fuertes y débiles. Como los
Competitividad de la investigación. Posición en el mundo y en el país de las publicaciones ordenadas por el número de citas, de mayor a menor. Publicaciones domésticas en 2014−2017, citas en 2019−2022.
ciclistas de cada equipo colaboran entre ellos, el equipo de ciclistas es, sin duda, un sistema algo más que los ciclistas individuales, pero solo algo más, y las clasificaciones de los ciclistas por equipos no se pueden caracterizar con ecuaciones matemáticas.
La investigación que persigue empujar la frontera del conocimiento es completamente distinta. Si ordenamos las publicaciones globales en una disciplina por el número de citas y buscamos las publicaciones de un país, sus números de orden en la clasificación general no son números
aleatorios, se ajustan a la siguiente ley de potencias:
Número de orden en la lista global = A x (Número de orden en la lista local)α (1)
donde A y α son unas constantes. Obviamente, con escalas logarítmicas, la representación de esta función es una línea recta. No obstante, esta es una ley ideal, como la de los gases perfectos, que en la práctica tiene desviaciones que no son objeto de este artículo, pero están estudiadas. Todo esto lleva a una conclusión
crucial para la política científica, el éxito de la investigación depende del país (sistema científico) y se puede evaluar.
La Figura 2 es la representación gráfica de varios casos de evaluación basados en la Ecuación (1).
En esta figura, los ejes tienen escalas logarítmicas y las variables están intercambiadas con respecto a la Ecuación (1), como se indica en los títulos de los ejes. Las dos tecnologías presentadas, células solares y grafeno, se han seleccionado por ser objeto de investigación intensa en la mayor parte de los países y porque los
Figura 2
resultados para la UE y España son representativos de muchos otros temas de investigación, incluyendo los biomédicos. Los países que se comparan parten de números de publicaciones similares, y esto simplifica la comparación, pero aunque no fuera así, la pendiente de las rectas de regresión que se pueden trazar en cada país pone de manifiesto la eficiencia de su investigación. Cuanto menor es la pendiente, la eficiencia es mayor, porque implica que con el mismo número de trabajos se está más próximo al éxito que representa estar en los primeros lugares.
En la comparación de EEUU y la UE, tanto en células solares como en grafeno, EEUU tiene las pendientes menores, lo que indica la superioridad de EEUU. El caso de grafeno es interesante por la atención prestada en la UE a este material en el programa “Graphene Flagship”. En los años de estudio, en el mundo hubo 82.758 publicaciones, y las domésticas de EEUU y la UE fueron 5.874 y 7.555. Como consecuencia de la diferencia en la pendiente de las rectas de regresión en la Figura 2, las tres primeras publicaciones de EEUU tienen las posiciones 3, 5 y 9, mientras que las de la UE tienen las posiciones 18, 25 y 24. La desviación que se observa en los puntos más bajos de la gráfica de la UE, aproximadamente las 100 primeras publicaciones, probablemente indica que algunos países pequeños son más eficientes que el conjunto de la UE. En el caso de España, también es evidente que está lejos del Reino Unido en células solares y de Singapur en grafeno.
La política científica como única causa
Los casos presentados en la Figura 2 para la UE y para España son una muestra que es representativa de otras tecnologías y de las
colaboraciones internacionales. Como era esperable, la ventaja de EEUU sobre la UE que se observa con estas comparaciones es muy parecida a la realizada más arriba contando los Premios Nobel. Por ello, la pregunta que emerge es por qué EEUU tiene mejor investigación que la UE. Aunque hay notables diferencias en PIB, el nivel de ambas economías es lo suficientemente alto como para pensar que las diferencias en investigación no se deben a las diferencias en el PIB total o per cápita. La clave está en que el número de publicaciones en relación al PIB es más del doble en la UE que en EEUU y a la vez, la pendiente de las rectas de regresión en la Figura 2 son más altas en la UE que en EEUU: son dos sistemas radicalmente distintos. Evidentemente, solo una política dirigida al número de publicaciones y no a los descubrimientos puede explicar las diferencias. En otras palabras, es una política que dirige a publicar desarrollos y no a buscar avances disruptivos, como señala Draghi.
Para leer más
Conclusión
La brecha en innovación entre la UE y EEUU existe desde hace muchos años y a ella se ha sumado China más recientemente. El Informe Draghi propone inversiones mil millonarias para cerrar esta brecha, pero ese esfuerzo inversor se verá comprometido si no se mejora la investigación. Los 30 años de la falsa Paradoja Europea y juicios similares han marcado el camino de la UE a la situación actual. Sería un logro que la advertencia del Informe Draghi sobre una investigación “insufficiently focused on disruptive innovation” tuviera más éxito que lo aportado en 18 años por la investigación académica, que ha sido radicalmente ignorado. En España la situación es similar porque la excelencia científica que asume la Ley de la Ciencia, la Tecnología y la Innovación de 2022 se aprobó sin votos en contra y sin reparos de la COSCE, ignorando la investigación académica sobre la materia. Corregir esta situación es urgente para España y para la UE.
Rodríguez-Navarro A. Sound research, unimportant discoveries. Research, universities, and formal evaluation of research in Spain. Journal of the American Society for Information Science and Technology 60 (2009) 1845–1858.
Rodríguez-Navarro A, Narin F. European paradox or delusion—Are European Science and economy outdated? Science and Public Policy 45 (2018) 14–23.
Rodríguez Navarro A. Cómo medir el éxito científico. Los errores de España. McGrow Hill - Aula Magna. España (2022)
Rodríguez-Navarro A. Citation distributions and research evaluations: The impossibility of formulating a universal indicator. Journal of Data and Information Science 9 (2024) 24-48.
ES HORA DE LLEVAR AL AULA LA PREDICCIÓN DEL PLEGAMIENTO DE PROTEÍNAS
Ángel Herráez
Bioquímica y Biología Molecular, Departamento de Biología de Sistemas, Universidad de Alcalá, Alcalá de Henares, Madrid
La reciente revolución resultante de la popularización de los algoritmos de Inteligencia Artificial (IA) ha alcanzado también al campo de la biología estructural. Todos aprendimos, y así lo enseñamos en nuestras clases de bioquímica, que la secuencia de una proteína determina su plegamiento en una estructura tridimensional. También afirmamos que los principios que rigen dicha consecución de la estructura nativa se nos escapan en buena parte. Así ha sido durante muchos años: sólo algunas pinceladas de la tendencia de estos o aquellos aminoácidos a adoptar un tipo de estructura secundaria u otra, la influencia de los enlaces disulfuro –que estabilizan pero no determinan el plegamiento–, la importancia del efecto hidrófobo –no siempre correctamente descrito–, el papel de los iones metálicos para fijar determinadas estructuras... en resumen, muchas ideas que ayudan, pero continúa la incapacidad de afirmar “esta secuencia llevará a tal estructura terciaria “. A partir de 2020 aproximadamente, con la publicación de AlphaFold2, nos vimos en una nueva situación: los algoritmos de predicción de estructura proteica basados en IA y puestos a disposición de la comunidad científica
abrían una nueva vía para estimar cómo podría ser la estructura terciaria de una secuencia concreta de residuos aminoácidos, para un número ingente de proteínas. Las pruebas experimentales obtenidas en los concursos CASP [1] mostraban que estas predicciones tenían una buena fiabilidad: la estructura predicha coincide, en muchos casos, en buena medida con aquélla determinada experimentalmente. Esto ha dado una reputación de fiabilidad a los métodos de IA que, no obstante, no nos explican el cómo y por qué del plegamiento, sino que más bien actúan como una caja negra de la que conocemos lo que entra y lo que sale, pero no comprendemos la maquinaria interna. Eso sí, nos hace felices poder obtener aproximaciones a la estructura de proteínas hasta ahora inalcanzables. También es una importante ayuda para orientar el trabajo de determinación experimental. En el número previo de esta revista hemos podido leer varias contribuciones de nuestros compañeros sobre el impacto de la IA en nuestro campo de estudio. No pretendo en modo alguno alcanzar ese nivel ni reincidir en lo ya tratado, en particular visto el artículo de Pablo Chacón que cubre precisamente, a fondo, la influencia en
EDUCACIÓN UNIVERSITARIA
la biología estructural. Sí quisiera traeros algunas ideas sobre cómo podemos integrar, si bien de forma sencilla, estas nuevas herramientas en la formación de nuestros alumnos.
El planteamiento de esta contribución no es que los estudiantes averigüen la estructura de una proteína usando estas herramientas de predicción, sino que reflexionemos sobre qué pueden aprender mientras prueban la predicción. Y ello en cualquier asignatura de bioquímica general, en una situación realista, sin emplear para este aspecto mucho tiempo ni recursos de software. Dicho de otro modo, pretendo compartir herramientas sencillas de usar, tanto en el aula como en actividades propuestas para que las realicen los alumnos, sin requerir la instalación de programas ni profundizar en sus prestaciones. ¿Podemos utilizar los servicios de predicción para
que nuestros alumnos refuercen su aprendizaje de la estructura de las proteínas y, al mismo tiempo, se vayan familiarizando con estas nuevas herramientas, que no podrán sino crecer, como parte de su formación y su futuro profesional?
Consulta de las bases de datos
Tenemos la oportunidad –probablemente la obligación– de informar a los estudiantes de que pueden encontrar cuál es la estructura de una proteína en la base de datos experimentales (Protein Data Bank, PDB [2]) y asimismo la predicción de su estructura en las bases de datos generadas por IA, entre las que destacan AlphaFoldDB [3] y ESM Metagenomic Atlas [4]. La búsqueda es sencilla y será ventajoso incluirla como la primera fase de una actividad para los alumnos, que para promover un aprendizaje provechoso debe ir acompañada de algunas preguntas que deban
resolver explorando dicha estructura, bien en los propios portales de búsqueda o descargando el archivo de datos para trabajar sobre él con alguno de los programas de visualización de los que ya hemos hablado en otras ocasiones. También se podría proponer un análisis crítico comparando la estructura predicha en las distintas plataformas entre sí o bien con la estructura experimental, para evitar la percepción de verdad absoluta en la que en ocasiones se pueda caer.
Fiabilidad de los algoritmos de predicción
Es esencial transmitir a nuestros alumnos la idea de que estos métodos no proporcionan una certeza absoluta sobre la estructura resultante, y trabajar en su análisis crítico como parte de las actividades que les propongamos.
Más allá de la estructura tridimensional en sí, el parámetro
Figura 1
Estructura predicha para la secuencia de Serum amyloid A-1 en el servicio ESMfold, coloreada de acuerdo con el índice de fiabilidad pLDDT.
principal que proporcionan los servicios de predicción es el valor pLDDT, un indicador de confianza o de la fiabilidad esperada para las coordenadas espaciales de cada átomo o, según los casos, de cada residuo aminoácido en su conjunto. Hay otros indicadores pero, para los propósitos docentes, nos bastará con éste salvo que hablemos de cursos avanzados. Los valores de pLDDT inferiores a 50 (en una escala de 0 a 100) se consideran como de fiabilidad no significativa. Por encima de ese umbral se establecen varios niveles, aunque los resultados para la proteína sólo se consideran sólidos en el segmento más alto. Se suele utilizar un código de coloreado (Figura 1) para representar este parámetro sobre el modelo tridimensional y así poder identificar las regiones más fiables.
Ejercicios de predicción
¿Qué ejercicios podemos plantear a los estudiantes, sin complicarnos con la instalación de programas, procedimientos elaborados o aprendizaje del funcionamiento de plataformas? Propongo algunas soluciones simples que se pueden incorporar tanto en el tiempo de aula como en el de trabajo personal o en grupos.
Aunque el algoritmo de AlphaFold2 (2020) se liberó como código abierto, no identifiqué ninguna herramienta de uso sencillo y rápido para su utilización por usuarios noveles. La principal ventaja fue la disponibilidad de la base de datos de sus resultados (AlphaFoldDB), pero sin acceso a una fácil predicción de cualquier secuencia arbitraria. Esto ha cambiado con la versión AlphaFold3 (2024), como explicaré a continuación.
En contraste, el algoritmo de ESM sí ofrece una interfaz de uso sencillo en línea, sin instalación ni registro de usuario.
Predicción en ESM “Fold Sequence”
Este servicio es de acceso libre en línea y no requiere siquiera de registro como usuario. Sí tiene algunas limitaciones como la longitud de secuencia que podemos proporcionar y el número de peticiones que podemos enviar, pero será más que suficiente para nuestro uso docente. Primeramente, necesitamos obtener la secuencia de la proteína que queremos analizar. Una forma sencilla es dirigirnos a la base de datos
Protein en NCBI:
• Introduce en el buscador “NCBI Protein” para llegar a http://ncbi.nlm. nih.gov/protein/
• Abre una búsqueda con el nombre de la proteína.
• Copia la secuencia; el modo más rápido es accediendo al enlace “FASTA”. Guárdala en un editor de texto.
A continuación, ve al servidor de predicción:
• Introduce en el buscador “ESM Fold Sequence” para llegar a http:// esmatlas.com/resources?action=fold
• Pega la secuencia conseguida anteriormente; elimina cualquier salto de línea y los espacios que pueda haber al final.
• Pulsa la tecla Intro (Enter) o pulsa en el icono de lupa. En poco tiempo se mostrará el modelo tridimensional de la estructura. Para guardar el resultado, encontrarás un botón o enlace Download PDB file El coloreado que se muestra en la estructura es muy importante, pues nos habla de la fiabilidad de la predicción. Hay en ello mucha enseñanza y una posible reflexión sobre la validez de estos métodos, y el resultado será muy diferente dependiendo de la proteína que nos hayamos propuesto analizar. Lamentablemente, no se muestra la leyenda de colores en la misma página donde estamos viendo el resultado y, aunque el archivo que descargamos incluye los valores pLDDT, no es inmediato su aprovechamiento en muchos programas de visualizacíón. En seguida propondré una solución.
Predicción en AlphaFoldServer
Con la publicación de la versión 3 del algoritmo, AlphaFold ha puesto a disposición un servicio de predicción en línea. Este es gratuito pero requiere acceder con una cuenta de Google. Asimismo posee algunas limitaciones para evitar un uso abusivo.
Algunas ventajas de este servicio:
• Guarda las tareas que le hemos enviado anteriormente, permitiéndonos volver a ellas, descargarlas más tarde, duplicarlas para modificarlas en una nueva petición...
• Muy importante: ya no está limitado a una sola cadena polipeptídica, sino que incluye la posibilidad de predecir proteínas con varias subunidades, ácidos nucleicos, incluir cofactores o ligandos, así como residuos aminoácidos con las modificaciones más frecuentes, incluso la glicosilación.
Para el ejercicio, una vez obtenida la secuencia como se ha indicado en el apartado anterior, ve al servidor de predicción:
• Introduce en el buscador “AlphaFoldServer” para llegar a http://alphafoldserver.com/
• Usa tu cuenta de Google para acceder.
• Elige “protein” como tipo de molécula.
• Pega la secuencia conseguida anteriormente; elimina cualquier salto de línea y los espacios que pueda haber.
• Pulsa en Continuar y espera hasta que se complete el cálculo.
EDUCACIÓN UNIVERSITARIA
Pulsando en la tarea (dentro del listado de consultas enviadas) se mostrará el modelo tridimensional de la estructura. De nuevo, el coloreado es muy importante, pero no se hace patente su significado ni los datos de pLDDT.
Para guardar el resultado hay un enlace Download y también se puede acceder desde el listado de consultas enviadas (menú que se despliega en el icono de 3 puntos a la derecha). En este caso la descarga es un archivo zip que contiene varios modelos, todos en formato mmCIF. Normalmente nos bastará con extraer el que termina en model_0.cif (ésta es la predicción con mayor puntuación).
Al igual que en el servidor ESM, tendremos que procesar un poco el archivo si queremos examinar los valores pLDDT o mostrarlos coloreando el modelo tridimensional.
Visualización y análisis de los archivos descargados Como he mencionado, la mayoría de programas de visualización en modo local no son capaces de interpretar los valores pLDDT de cada átomo. Éstos se graban en la ubicación del archivo dedicada al valor del B-factor, coloquialmente denominado “temperatura”. La definición original de este dato refleja la incertidumbre en la posición del átomo obtenida del análisis de los datos experimentales (difracción con rayos X de la proteína cristalizada, o resultado de la resonancia magnética nuclear, o de otras técnicas de resolución de las coordenadas). Como tal, el valor varía entre 0 y 100 (en la práctica quizás entre 10 y 90), con mayores valores indicando mayor imprecisión, o mayor movilidad de los átomos, partes más flexibles de la proteína. Los visualizadores suelen leer este dato y colorear en alguna variante de gradiente desde azul (“frío”, poco variable) hasta rojo (“caliente”, más móvil). Desafortunadamente, los resultados pLDDT de la predicción se interpretan al contrario: valores bajos son baja fiabilidad y los altos alta fiabilidad. Para colmo, algunos servidores graban los datos en escala de 0 a 100, mientras que otros lo hacen de 0,0 a 1,0. Además, como hemos mencionado, la parte significativa está por encima de 70, por lo que el gradiente de color tradicional no permite ver bien las diferencias en pLDDT.
A la vista de todo esto, acudí al rescate preparando una utilidad [5] que lee los archivos resultantes de ESMfold y de AlphaFold (DB o Server), los muestra como modelo 3D interactivo y los colorea de acuerdo a los datos pLDTT usando el esquema de colores naranja-amarillo-cian-azul. Además, permite grabar de nuevo el archivo con factores de temperatura recalculados adecuadamente para que se puedan mostrar en otros visualizadores usando el patrón estándar de gradiente “color por temperatura”. Por último, extrae todos los valores pLDDT en una tabla que se puede copiar y guardar con facilidad para un análisis detallado (lo que ya excede el objetivo de esta comunicación).
Nota: ChimeraX tiene una opción para colorear por pLDDT, mediante código (color bfactor palette alphafold y color bfactor palette esmfold). PyMOL lo puede hacer a través de una serie de instrucciones de código (no automatizado) o instalando una extensión del programa [6]. iCn3D tiene una opción directa en el menú Color
Para quien tenga cierta familiaridad con Proteopedia, procede indicar que en el editor de escenas (SAT) se podrá colorear automáticamente por fiabilidad cualquier archivo que carguemos procedente de AlphaFold, ESMfold o RoseTTaFold, y se incluirá la leyenda de colores bajo el modelo 3D. Además, es también posible obtener una predicción de ESMfold simplemente escribiendo la secuencia dentro del propio SAT (esto es, sin necesidad de acudir al servidor ni de descargar el archivo).
Otros servicios de predicción
Comento tan solo brevemente otros servidores, en general de uso menos simple y, por ello, que serán adecuados sólo para cursos más avanzados o con más tiempo disponible para este tema.
La posibilidad de usar el modelo AlphaFold2 sólo era asequible a través de algunas interfaces llamadas Google Colabs que ejecutan código en la nube y requieren acceder con una cuenta de Google. A mi juicio, no son amigables para los no iniciados. Afortunadamente, como se ha indicado, la versión 3 sí ofrece una accesibilidad sencilla.
Otro algoritmo que se desarrolló independientemente es RoseTTaFold. Se puede utilizar en el servidor Robetta [7] pero requiere registrarse. También hay un servicio de acceso libre proporcionado por NeuroSnap [8], que ofrece la opción de varios algoritmos alternativos.
Corolario
Como ya he mencionado anteriormente, no creo que el mérito de estos métodos de predicción esté en que los estudiantes de una bioquímica básica averigüen la estructura de una proteína, sino más bien en que se acostumbren a las estructuras secundarias y terciarias, las herramientas de visualización, las bases de datos y los servicios de predicción, y acompañen todo ello de un análisis crítico de lo que encuentren. Como muestra, pueden verse los resultados presentados en la Figura 2
Debe tenerse en cuenta, además, que los servidores gratuitos a menudo limitan el tamaño máximo de la proteína o complejo proteico y, al mismo tiempo, si enviamos polipéptidos pequeños la fiabilidad de la predicción suele ser baja o moderada; esto es lógico, pues el número de interacciones necesarias para estabilizar fuertemente una estructura terciaria es difícil de alcanzar cuando su tamaño es pequeño.
Figura 2
Resultado de la predicción de estructura con diversos algoritmos para el “péptido navideño”, de secuencia MERRYCHRISTMASANDHAPPYNEWYEAR. De izquierda a derecha: ESMfold, AlphaFoldServer, RosettaFold Neurosnap Boltz y RosettaFold Neurosnap Chai. La representación de cintas está hecha con Jmol.
Agradecimientos
Me ha resultado de ayuda para esta presentación la recopilación [9] elaborada por Eric Martz, profesor emérito de la Universidad de Massachussetts Amherst y por largo tiempo experto –y maestro de muchos, incluido el que esto escribe– en la visualización de estructura molecular y su empleo como recurso educativo.
Referencias
1. CASP: Critical Assessment of protein Structure Prediction (1994-2024) Protein Structure Prediction Center, https://predictioncenter.org/
2. wwPDB, Protein Data Bank, https://pdb.org
3. AlphaFoldDB, https://alphafold.ebi.ac.uk
4. ESM Metagenomic Atlas, https://esmatlas.com
5. A. Herráez (2024) Color protein residues by pLDDT score. https://biomodel.uah.es/Jmol/fold_AI/
6. (a) Kuen Wu (2023) Color AlphaFold2’s pLDDT. https://kpwulab.com/2023/03/09/color-alphafold2s-plddt/ (b) UCSF ChimeraX User Guide, https://www.cgl.ucsf.edu/chimerax/docs/user/commands/palettes.html (c) Christian Balbin (2021) PyMOL plugin, https://github.com/cbalbin-bio/pymol-color-alphafold (d) Bob Schiffrin (2021) PyMOL script, https://github.com/BobSchiffrin/pymol_scripts
7. Robetta protein structure prediction service, using RoseTTAFold and TrRosetta. https://robetta.bakerlab.org/
9. How to predict structures with AlphaFold (2024). https://proteopedia.org/w/ How_to_predict_structures_with_AlphaFold&oldid=4278539
CINÉTICA HIPERBÓLICA Y CINÉTICA SIGMOIDE: LA FORMA DE LA CURVA IMPORTA, ¡Y MUCHO!
Vicente Rubio
Instituto de Biomedicina de Valencia (IBV), CSIC y grupo CB06/07/0077 del CIBER de Enfermedades Raras (CIBERER-ISCIII). rubio@ibv.csic.es
Félix M. Goñi
Instituto Biofisika, CSIC-UPV/EHU, Leioa, Bizkaia. felix.goni@ehu.es
Carlos Gancedo
Ex Profesor de Investigación del CSIC. carl.gancedo@gmail.com
Decir que la vida sería imposible sin enzimas es superfluo para los lectores de SEBBM, que saben cuánto aceleran las enzimas las reacciones químicas del organismo, hasta 1017 veces [1]. Los estudios de la velocidad de acción (catálisis) de las enzimas son objeto de la cinética, esa parte de la asignatura de Bioquímica que muchos estudiantes dicen sufrir y olvidar pronto; y a la que los textos hacen escasa referencia, fuera del capítulo que le dedican.
Sin embargo, la cinética está implícita y cotidianamente presente en muchas actividades profesionales (medicina, biotecnología, biofarma, etc.), habiendo sido la gran introductora de las enzimas como protagonistas biológicos, con
los pioneros estudios publicados en la primera década del siglo XX por el británico Adrian J. Brown (químico, profesor de fermentaciones cerveceras) y por el francés Victor Henri (doctor en química-física y en…¡psicología!) [2], conducentes a la ecuación principal de la cinética enzimática, que consolidarían y difundirían en 1913, Leonor Michaelis y (Miss) (sic) Maud
L(eonora) Menten, padre y madre definitivos de la ecuación que lleva sus nombres.
Para paliar el olvido de la cinética, proponemos un “enzimokit” de primera ayuda, que incluye la hipérbola que pasa por el cero, en coordenadas lineales que relacionan velocidad de reacción enzimática (v, en el eje Y)
frente a concentración de sustrato (S; eje X), y que corresponde a la ecuación de Michaelis y Menten, v=V max ×S/(Km+S), con sus magnitudes asociadas: Vmax (v a concentración infinita, o sea saturante, de S), y Km (valor de S a la que v=Vmax/2). El enzimokit también nos recuerda la existencia de inhibidores de diversos tipos, y de otros factores que influyen sobre la velocidad de las enzimas, como el pH o la temperatura. Pero el enzimokit estaría incompleto si no incluyera también la cinética sigmoide, así llamada porque la forma de la curva que la representa se desvía de la hipérbola para recordar una letra S mayúscula. El paradigma de cinética sigmoide es la representación del grado de oxigenación de la hemoglobina frente a la concentración de oxígeno, que caracterizó matemáticamente en 1910 el británico Archibald V Hill [3]. Algunas enzimas presentan cinética sigmoide, descrita por la ecuación de Hill, v=V max ×SN/(S0.5N+SN), en la que S0.5 es la concentración de sustrato a la que v=Vmax/2 (Km en cinética “michaeliana”); y N es un número que, para un valor de 1 describe cinética hiperbólica y, para valores superiores cinéticas sigmoides. Veamos dos ejemplos de lo que puede suponer en patología la diferencia entre que la cinética de una enzima sea michaeliana o sigmoide. El primero se refiere a una forma mutante de hipoxantina-guanina-fosforribosil-transferasa (HPRT) que causa síndrome de Lesch-Nyhan (síndrome neurológico con tendencia a la autoagresión mutiladora, asociado a gran producción de ácido úrico) [4]. La HPRT forma parte de la vía de recuperación de purinas, transfiriendo el fosforribosilo del pirofosfato de fosforribosilo (PRPP) a guanina e hipoxantina, dando respectivamente GMP o IMP. Aunque la cinética para el PRPP de la HPRT normal es michaeliana, en un
Figura 1
Cinéticas para PRPP y CP de HRPT (arriba) y OTC (abajo) normales y mutantes halladas en pacientes con deficiencias parciales de esas enzimas. Las curvas para las formas normales son o se aproximan a hipérbolas michaelianas, mientras que las formas mutantes son sigmoides con valores de N≈2 e igual Vmax que las formas normales. Se indican en azul los valores de K m y S0.5 respectivos. Los datos para HRPT están tomados de [4] y los de OTC del laboratorio de uno de los autores (VR).
paciente era sigmoide (Figura1) [4]. Ese hallazgo fue importante, ya que indicó (¡en 1971!) la heterogeneidad genética de esa enfermedad. Desgraciadamente no ha permitido desarrollar tratamientos para la misma. El segundo ejemplo es una variante de la ornitina transcarbamilasa (OTC), enzima que cataliza la síntesis de citrulina a partir de ornitina y de carbamil fosfato (CP)
en el ciclo de la urea [5], ciclo que convierte en urea (poco tóxica) el amonio (neurotóxico) derivado del catabolismo proteico. Una mutación puntual en la OTC de un paciente convierte la cinética de esa enzima para CP de michaeliana en sigmoide (Figura 1) (datos no publicados de VR), lo que hace a la OTC inactiva a las concentraciones normales de CP en la mitocondria hepática [6], donde se
localiza la OTC. El déficit funcional de OTC debido a esta mutación, y su posible secuela de hiperamoniemia, deben poder prevenirse o aliviarse con carbamilglutamato, un fármaco oral, activador artificial de la carbamil fosfato sintetasa 1 (CPS1), enzima que convierte el amonio en CP. Eso debe permitir aumentar la concentración intramitocondrial de CP a las concentraciones normales de amonio.
Estos ejemplos ilustran la importancia de la cinética enzimática aún en los tiempos de AlphaFold y de Next generation sequencing CG agradece a Juana M. Gancedo los comentarios a lo largo de la redacción de este artículo.
Referencias
1. Radzicka A, Wolfenden R (1995) A proficient enzyme. Science 267 (1995) 90-93.
2. Cornish-Bowden A. The origins of enzyme kinetics. FEBS Lett 587 (2013) 2725-30.
3. Hill AV. The possible effects of the aggregation of the molecules of haemoglobin on its dissociation curves. J Physiol (Lond.) 40 (1910) 4-7.
5. Häberle J et al. y Rubio V, Dionisi-Vici C. Suggested guidelines for the diagnosis and management of urea cycle disorders. Orphanet J Rare Dis 7 (2012) 32.
6. Alonso E & Rubio V. Orotic aciduria due to arginine deprivation. Changes in the levels of carbamoyl phosphate and of other urea cycle intermediates in mouse liver. J Nutr 119 (1989) 1188-1195.
INVESTIGACIÓN
Microexón de 24 nucleótidos en el gen CPEB4 y el trastorno del espectro autista
Carla Garcia-Cabau, Anna Bartomeu, Giulio Tesei, Kai Chit Cheung, Julia Pose-Utrilla, Sara Picó, Andreea Balaceanu, Berta Durán-Arqué, Marcos Fernández-Alfara, Judit Martín, Cesare De Pace, Lorena Ruiz-Pérez, Jesús García, Giuseppe Battaglia, José J Lucas, Rubén Hervás, Kresten Lindorff-Larsen, Raúl Méndez, Xavier Salvatella. 2025. Mis-splicing of a neuronal microexon promotes CPEB4 aggregation in ASD. Nature 637(8045):496-503. doi: 10.1038/s41586-024-08289-w
La inclusión de microexones mediante splicing alternativo ocurre frecuentemente en proteínas neuronales. Las funciones de estas secuencias son en gran medida desconocidas y los cambios en su grado de inclusión se asocian con trastornos del neurodesarrollo. En 2018 se descubrió que la disminución de la inclusión de un microexón específico de neurona de 24 nucleótidos en CPEB4, una proteína de unión a ARN que regula la traducción a través de cambios citoplasmáticos en la longitud de la cola poli(A), está relacionada con el trastorno del espectro autista (TEA) idiopático. De todas formas, no se sabía por qué se requiere este microexón y cómo pequeños cambios en su grado de inclusión tienen un efecto dominante negativo en la expresión de genes relacionados con el TEA. En este artículo, publicado en la revista Nature y liderado por los equipos de Raúl Méndez y Xavier Salvatella (IRB Barcelona), se demuestra que CPEB4 forma condensados que se disuelven durante la despolarización neuronal, una transición asociada con un cambio de la represión traslacional a la activación. Las interacciones heterotípicas entre el microexón y un grupo de residuos de histidina previenen la agregación irreversible de CPEB4 al competir con las interacciones homotípicas entre grupos de histidina. Con esto, se puede concluir que el microexón es necesario en CPEB4 neuronal para preservar la regulación reversible de la expresión génica mediada por CPEB4 en respuesta a la estimulación neuronal.
HuMicA: un recurso para comprender el papel de la microglía en patologías
neurodegenerativas
Ricardo Martins-Ferreira, Josep Calafell-Segura, Bárbara Leal, Javier Rodríguez-Ubreva, Elena Martínez-Saez, Elisabetta Mereu, Paulo Pinho E Costa, Ariadna Laguna, Esteban Ballestar. 2025. The Human Microglia Atlas (HuMicA) unravels changes in disease-associated microglia subsets across neurodegenerative conditions. Nature Communications 16(1):739. doi: 10.1038/s41467-025-56124-1
La microglía, el principal tipo de célula inmune del cerebro, es fundamental para el desarrollo y el buen funcionamiento de la actividad neuronal. Se activa tras la exposición a estímulos. Una activación proinflamatoria excesiva de la microglía se asocia con diversas patologías. Las tecnologías de secuenciación de ARN de célula única (scRNA-seq) han contribuido a identificar microglía activada en enfermedades (DAM, disease-activated microglia), considerada el estándar para el estudio de la microglía en enfermedades neurodegenerativas. No obstante, la DAM resulta insuficiente para representar la complejidad de la microglía en estas enfermedades. El grupo liderado por E. Ballestar publica en Nature Commun el Atlas de la Microglía Humana (HuMicA), un recurso integrado de datos de scRNA-seq compuesto por 90.716 células mieloides cerebrales. Este atlas se genera a partir de 241 muestras obtenidas de 19 bases de datos correspondientes a 6 enfermedades neurológicas. El HuMicA identificó 9 grupos celulares con distintas subpoblaciones de microglía. Entre ellas, se identifica una población de macrófagos derivados de monocitos similares a microglía, previamente descrita en ratón, que demuestra cómo de prevalente es en el cerebro humano. Además, la firma DAM se manifiesta en 4 subpoblaciones distintas, una de las cuales mostró una alta expresión de genes involucrados en el metabolismo lipídico (GPNMB-high Lipo.DAM), que aparece expandida en la enfermedad de Alzheimer y en esclerosis múltiple. Por tanto, el HuMicA representa un recurso de gran valor para comprender no solo el papel de la microglía en las patologías, sino también su biología intrínseca.
La proteína mitocondrial J controlada por metilación y su papel en la termogénesis
Beatriz Cicuéndez, Alfonso Mora, et al. 2025. Absence of MCJ/ DnaJC15 promotes brown adipose tissue thermogenesis. Nature
La obesidad es una de las mayores preocupaciones en cuanto a salud mundial, lo que hace crucial entender cómo funciona el tejido graso o adiposo y, especialmente, sus mitocondrias. Este estudio se centra en la proteína mitocondrial J controlada por metilación (MCJ) y su papel en la regulación de la producción de calor o termogénesis en el tejido adiposo marrón (BAT). La investigación, realizada por el grupo liderado por la Dra. G. Sabio en el CNIO y publicada en Nature Communications, muestra que los niveles de MCJ en el tejido adiposo están regulados por obesidad y cambios térmicos en humanos y ratones, sugiriendo un papel de MCJ en el control energético. Además, los ratones que carecen de MCJ en su BAT (MCJKO) bajo una dieta alta en grasa muestran una mayor termogénesis y disminución en peso, incluso sin la proteína UCP1, que generalmente es esencial en este proceso. De hecho, el trasplante de BAT sin MCJ fue capaz de aumentar la termogénesis y disminuir el aumento de peso en ratones bajo dieta alta en grasa. Mediante microscopía electrónica pudieron observar cambios estructurales en las mitocondrias del BAT sin MCJ, similares a los inducidos por la activación en frío, indicando mayor producción de energía. El análisis proteómico reveló que la vía de respuesta al estrés de eIF2α desempeña un papel esencial en este proceso. Al eliminar eIF2α en el BAT de ratones MCJKO, se revirtió el aumento de la termogénesis, confirmando su papel clave para la termogénesis en ausencia de MCJ. Este hallazgo posiciona a MCJ como un regulador central de la termogénesis en el BAT y un prometedor objetivo terapéutico para combatir la obesidad.
La fosforegulación de Cav1 es relevante en la mecanoadaptación del adipocito
María CM Aboy-Pardal, Marta C Guadamillas, Carlos R Guerrero, Mauro Català-Montoro, Mónica Toledano-Donado, Sara Terrés-Domínguez, Dácil M Pavón, Víctor Jiménez-Jiménez, Daniel Jiménez-Carretero, Moreno Zamai, Cintia Folgueira, Ana Cerezo, Fidel-Nicolás Lolo, Rubén Nogueiras, Guadalupe Sabio, Miguel Sánchez-Álvarez, Asier Echarri, Ricardo García, Miguel A Del Pozo. 2024. Plasma membrane remodeling determines adipocyte expansion and mechanical adaptability. Nature Communications 15(1):10102. doi: 10.1038/s41467-024-54224-y
La elevada prevalencia de sobrepeso y obesidad hace esencial encontrar mecanismos para paliar el daño que el exceso de grasa ocasiona en el organismo. En este sentido, los adipocitos cobran especial interés, pues son el tipo celular especializado en gestionar las reservas lipídicas de forma eficiente y segura. En el artículo publicado en Nature Communications, liderado por M.A. del Pozo (CNIC) en colaboración con grupos de ICMM, CIMUS, CNIO, IIBM y CIB, se describe cómo las caveolas asisten al adipocito en su cometido. Durante la acumulación de grasa, el adipocito aumenta de volumen y sufre un gran aumento de tensión en su superficie. Las caveolas son sensibles a estos cambios de tensión, a lo que, en un proceso dependiente de la fosforilación de Cav1, responden aplanándose y liberando el reservorio de membrana contenido en la invaginación. Esto no sólo proporciona más superficie a la célula y le permite aumentar todavía más su volumen, sino que también compensa el aumento de tensión derivado del llenado, evitando así que la célula se rompa. Además, cuando las caveolas se aplanan, componentes moleculares de estas estructuras viajan a otros compartimentos de la célula, pudiendo actuar como un mensaje que ayuda a coordinar el metabolismo celular con el nivel de reservas de energía. Al preservar la integridad de los adipocitos y facilitar la acumulación de grasa, las caveolas protegen la salud metabólica y cardiovascular. Los adipocitos desprovistos de caveolas son frágiles y no pueden acumular energía de manera eficiente, lo que permite un exceso de grasa circulante, y su depósito en la pared vascular o el hígado.
INVESTIGACIÓN
La unión de peroxiredoxinas con proteínas; una modificación postraduccional para la protección frente al estrés
Gerhard Seisenbacher, Zrinka Raguz Nakic, Eva Borràs, Eduard Sabidó, Uwe Sauer, Eulalia de Nadal, Francesc Posas. 2025. Redox proteomics reveal a role for peroxiredoxinylation in stress protection. Cell Reports 44(2):115224. doi: 10.1016/j.celrep.2024.115224
El estrés oxidativo daña proteínas y afecta la supervivencia celular. Las peroxiredoxinas, como Tsa1 en Saccharomyces cerevisiae, son enzimas clave en la detoxificación del peróxido de hidrógeno y protegen frente al estrés oxidativo. Además de su función antioxidante, actúan como chaperonas y participan en la señalización redox. Un estudio liderado por Francesc Posas y Eulàlia de Nadal en el IRB Barcelona y la Universidad Pompeu Fabra, publicado en Cell Reports, identifica el interactoma de Tsa1 y revela que tiene un papel protector de proteínas sensibles a su estado redox. Mediante espectrometría de masas, se observó que Tsa1 se une a proteínas involucradas en traducción, metabolismo y procesamiento de carbohidratos. Esta unión, llamada “peroxiredoxinilación”, es un acoplamiento covalente que protege la actividad de las proteínas diana en situaciones de estrés. El equipo demostró esta función protectora con Gnd1 (6-fosfogluconato deshidrogenasa), una enzima clave en la ruta de las pentosas fosfato. La modificación por Tsa1 es esencial para mantener la funcionalidad de Gnd1, permitiendo a las células preservar rutas metabólicas críticas incluso bajo estrés oxidativo. El estudio sugiere que la peroxiredoxinilación regula la reactividad de grupos tiol en proteínas, manteniendo el equilibrio redox. Estos hallazgos abren nuevas vías para investigar su papel en otros procesos sensibles al estrés oxidativo, no sólo en levaduras, sino también en sistemas celulares más complejos.
El intercambiador mitocondrial de sodio/calcio NCLX participa (también) en la respuesta a hipoxia mediada por HIF
Carmen Choya-Foces, Elisa Navarro, Cristóbal de Los Ríos, Manuela G López, Javier Egea, Pablo Hernansanz-Agustín, Antonio Martínez-Ruiz. 2024. The mitochondrial Na+/Ca2+ exchanger NCLX is implied in the activation of hypoxia-inducible factors. Redox Biology 77:103364. doi: 10.1016/j.redox.2024.103364
A finales del siglo XX se descubrió el mecanismo molecular por el que muchas células animales responden a la hipoxia a través de la ruta de PHD y HIF, hallazgos por los que Kaelin, Ratcliffe y Semenza obtuvieron el Nobel de Medicina en 2019. Tanto PHD como HIF se encuentran en el citosol y en el núcleo, pero ya desde antes de identificar PHD se sabía que algunas funciones mitocondriales eran necesarias para que se diera la respuesta a hipoxia, por mecanismos que aún no se conocen bien. Previamente, el grupo liderado por A. Martínez Ruiz en el Instituto de Investigación Sanitaria Princesa (IIS-IP), ya había publicado que el intercambiador Na+/Ca2+ mitocondrial, NCLX, era necesario para la producción de una señal mitocondrial de especies reactivas de oxígeno (ROS) que está implicada en algunas respuestas agudas a hipoxia (como la vasoconstricción pulmonar), que son inmediatas e independientes de HIF. En el trabajo publicado en Redox Biology este grupo responde a la pregunta de si hay conexión entre ambos mecanismos. La respuesta es afirmativa ya que NCLX es necesario también para que las células organicen adecuadamente la respuesta a hipoxia mediada por HIF. La inhibición o interferencia de NCLX (sin alterar la estabilidad de los complejos de la cadena de transporte de electrones mitocondriales o la función respiratoria) suprime la respuesta a la hipoxia mediada por la estabilización de la subunidad HIF-α y la actividad transcripcional en diferentes tipos de células. Aunque los mecanismos de respuesta a la disminución de oxígeno no están del todo esclarecidos, estos resultados ponen de relieve el papel de NCLX en la respuesta a hipoxia.
LA PRÓXIMA GENERACIÓN DE ENZIMAS RETROTRANSCRIPTASAS
La sensibilidad a la temperatura es un desafío clave en el uso de enzimas retrotranscriptasas (RT) en investigaciones moleculares. Las RT tradicionales suelen perder actividad a altas temperaturas, lo que limita su aplicación en muestras complejas y protocolos sensibles. Esto causa resultados inconsistentes, tiempos de procesamiento más largos y retrasos en experimentos. NZYtech ha resuelto este problema con sus soluciones Supreme Reverse Transcriptase, diseñadas para mantener un rendimiento alto incluso en condiciones de alta temperatura, permitiendo a los investigadores expandir sus estudios.
Las RT son esenciales para convertir ARN en ADN complementario (ADNc), pero las tradicionales fallan a temperaturas elevadas, afectando la fiabilidad de los datos. Investigadores en secuenciación de ARN, análisis de expresión génica o detección viral se ven especialmente afectados, con resultados inconsistentes y procesos prolongados.
La Supreme Reverse Transcriptase de NZYtech, en proceso de patente, ha sido diseñada para superar las limitaciones de temperatura de las RT tradicionales. Esta enzima funciona eficazmente hasta 70oC, garantizando un rendimiento confiable incluso en experimentos
exigentes. Ideal para muestras biológicas complejas o protocolos que requieren optimización, esta enzima ofrece flexibilidad para obtener resultados de alta calidad en condiciones donde otras RT fallarían.
Además, la Supreme NZY Reverse Transcriptase permite una síntesis rápida de ARN a ADNc, completando el proceso en menos de un minuto. Su formato liofilizado y alta concentración de 1 kU/μL ofrecen estabilidad y comodidad para uso a largo plazo.
La Supreme NZY Reverse Transcriptase es ideal para investigadores que necesitan estabilidad a altas temperaturas y velocidad en sus procesos. Como una innovación en proceso de patente, han sido desarrolladas y probadas rigurosamente para asegurar el máximo rendimiento en condiciones desafiantes.
¿Listo para descubrir cómo se comparan las Supreme Reverse Transcriptases de NZYtech?
Escanee el código QR para descargar el folleto de análisis del producto y revisar datos comparativos sobre su estabilidad y eficiencia.
La transcriptasa inversa Supreme NZY se ofrece en tres formatos que sintetizan eficientemente cDNA en un amplio rango de temperaturas, hasta 70°C. Se comparó la actividad de los productos MB482 (Lyo), MB494 (1 kU/µL) y MB496 (200 U/µL) a altas temperaturas. Se utilizó una serie de diluciones de ARN total de ratón (de 1 µg a 10 pg) como plantilla para la síntesis de cDNA durante 1 minuto a 42, 50, 60 y 70 °C. El cDNA sintetizado se detectó luego utilizando cebadores y sondas TaqMan específicos para el gen Rpl27 del ratón.
ANTONIO BALLESTEROS OLMO (1940-2024)
Francisco J. Plou
Profesor de investigación
Instituto de Catálisis y Petroleoquímica (ICP), CSIC
El pasado 17 de diciembre nos dejó Antonio Ballesteros (1940-2024), Profesor de Investigación ad honorem del CSIC y pionero de la investigación en biocatálisis en España. Antonio desarrolló una destacada carrera como investigador, que le valió un amplio reconocimiento nacional e internacional.
Antonio Ballesteros se licenció en Ciencias Químicas por la Universidad de Sevilla en 1962, obteniendo el Premio Extraordinario y un Accésit al Premio Nacional Fin de Carrera. Se doctoró en Ciencias Químicas por la Universidad Complutense de Madrid (UCM) en 1966, siendo reconocido con el Premio Extraordinario. Posteriormente, en 1972, completó en la UCM la licenciatura en Farmacia.
Inició en 1969 las investigaciones en biocatálisis en el Instituto de Química Física Rocasolano del CSIC (actual IQF ‘Blas Cabrera’, CSIC), en el Departamento de Catálisis (que más tarde se transformaría en el Instituto de Catálisis y Petroleoquímica, ICP-CSIC). Fue una apuesta arriesgada, que lo llevó a invertir cinco años, a lo largo de las décadas de 1970 y 1980, en estancias en centros y universidades de renombre en Estados Unidos, Reino Unido, Japón, Alemania y Francia. Pasó dos años en el National Institute of Health trabajando con el profesor Christian Anfinsen, galardonado con el Premio Nobel de Química en 1972.
En 1973, Antonio Ballesteros inició el estudio de la nucleasa de Staphylococcus aureus, que inmovilizó sobre soportes de agarosa. En 1976, comenzó una nueva línea sobre hidrogenasas. En 1987, investigó el uso de lipasas para la acilación regioselectiva de azúcares y la preparación de aditivos alimentarios. Más adelante, gracias a un proyecto europeo, se adentró en enzimas activas en carbohidratos, como la ciclodextrina glucanotransferasa y la dextransacarasa. A partir de ahí, enfocó
la investigación sobre la obtención de oligosacáridos prebióticos. En 2006, abrió un nuevo tema dedicado a la modificación enzimática de vitaminas antioxidantes y polifenoles bioactivos, entre ellos el resveratrol.
Antonio Ballesteros fue investigador principal en numerosos proyectos de ámbito nacional e internacional, participó activamente en diversas colaboraciones entre la academia y la industria, y dirigió más de veinte tesis doctorales.
Desde la creación de la European Federation of Biotechnology (EFB) en 1978, participó en sus grupos de trabajo sobre biocatálisis. Entre 1996 y 2000, presidió la Sección de Biocatálisis Aplicada de la EFB.
Dirigió el comité organizador de dos simposios internacionales celebrados en Córdoba: Stability and Stabilization of Biocatalysts (1998) y Environmental Biocatalysis (2006). Además, fue editor ejecutivo de la revista Biocatalysis and Biotransformation entre 1996 y 2011, año en el que asumió el puesto de editor jefe. En 2015 fue nombrado académico correspondiente de la Real Academia Nacional de Farmacia; en septiembre de 2020 tomó posesión.
Pero más allá de sus méritos profesionales, Antonio siempre hizo gala de una extraordinaria calidad humana. Una persona humilde, optimista, muy trabajadora, gran conversador, a quien siempre le gustaba encontrar el lado positivo de las cosas. Su legado en la ciencia española perdurará, pues logró fundar una escuela en la que las personas y sus valores tenían tanta relevancia como la ciencia que llevaban a cabo. Hoy son numerosos los grupos de biocatálisis en España tanto en el CSIC como en las universidades; la mayoría de ellos son fruto de aquella aventura biocatalítica que inició Antonio hace ya más de 50 años.
Descanse en paz.
JOAN GUINOVART (1947-2025)
Vicente Rubio, M. A. de la Rosa y Joaquim Ros
El primer día de 2025 nos abandonó Joan Guinovart Cirera, Presidente (1996-2000) y Socio de Honor de SEBBM y ex-editor jefe de esta revista. Escribimos esta nota desde el dolor y una profunda amistad. Sencillo, próximo, de humor personalísimo, aguda inteligencia, con gran capacidad comunicativa, fue un científico muy brillante. Hizo el recorrido desde la enzimología a la terapia molecular, con contribuciones clave sobre el metabolismo del glucógeno, la diabetes, el síndrome de Lafora, y el wolframato como terapia antidiabética. Estaba exultante por haber creado (2005) y dirigido (2005-2018) el IRB Barcelona, que consolidó como centro de referencia internacional de investigación en biomedicina básica y traslacional, dejando así un legado excepcional. La deuda de SEBBM con Joan es enorme. Durante su presidencia revitalizó nuestra sociedad, ampliando el número de grupos científicos; creó la figura de los “cónsules SEBBM” para la difusión y promoción de la SEBBM. Así, con su eslogan “2000 socios en el año 2000”, se pudo superar esta cifra cuando se cumplió el milenio. Organizó para SEBBM, en Barcelona (1996), el segundo congreso en España de FEBS, y aumentó y regló las relaciones internacionales de SEBBM, especialmente con Latinoamérica; además, con las colaboraciones del periodista Xavier Pujol y del editor Jaume Estruch, hizo del Boletín SEBBM la influyente Revista SEBBM que estáis leyendo. Siempre tuvo un gran sentido de pertenencia a SEBBM, que trasladó a FEBS cuando fue miembro de su ejecutiva y Congress Counsellor
(1997-2016), y a IUBMB, que presidió de 2015 a 2018. Legítimamente podemos decir que Joan fue embajador planetario de nuestra ciencia.
Con su gran capacidad de liderazgo promovió, junto a SEBBM (2003-2004), el Pacto de Estado por la Ciencia, movilizador de medidas públicas pro-ciencia y embrión del informe CRECE de la Confederación de Sociedades Científicas Españolas (COSCE), confederación que, con fuerte apoyo SEBBM, Joan gestó y lideró. Generó asimismo el programa CONOCEROS para mejorar el conocimiento y colaboración entre científicos y parlamentarios. Hombre público de extraordinaria ejecutoria, recibió muchas distinciones como las medallas Narcís Monturiol y la Creu de Sant Jordi de la Generalitat de Catalunya, la de Académico numerario de la Real Academia Nacional de Farmacia y el Diplôme d’Honneur de FEBS (lista más completa en la web de la SEBBM).
Siempre abogó por devolver los frutos de la ciencia a la sociedad y promover una economía basada en el conocimiento. También fue paladín del cambio de la gestión científica, que, como el fútbol de éxito, debía crear cantera, que inició con los programas de atracción juvenil “¿Y tú? ...yo ¡Bioquímica!”, “Crazy about Biomedicine”, y el proyecto ENCIENDE; fichar donde haga falta; y tener un plan que prioriza el objetivo, es decir, el éxito científico. Participemos de su talante, adhirámonos a estos objetivos y estrategias como la mejor forma de rendirle tributo como gran persona, investigador brillante, hombre público visionario, gestor tenaz y líder indiscutible.
FEDERICO MAYOR ZARAGOZA
(1934-2024)
Carmen Aragón Rueda
Creo que todos los miembros de la SEBBM conocéis la triste noticia del fallecimiento del Profesor Federico Mayor Zaragoza el 19 de diciembre de 2024. Fue una muerte repentina y no esperada. Tras su desaparición se han escrito numerosas notas necrológicas muy documentadas resaltando la figura y la trayectoria del Prof. Mayor Zaragoza a las que hay poco que añadir. Cuando se me encargó que escribiese esta nota para la revista de la SEBBM pensé en la dificultad, por un lado, de no ser reiterativa y por otro poder destacar brevemente una personalidad y trayectoria científica, humanista, política y personal tan dilatada e impresionante como la del Profesor Mayor Zaragoza. Es por lo me gustaría contribuir con algo personal en estas líneas.
Con respecto a la SEBBM baste recordar que fue uno de los promotores y su presidente en los albores de la misma e impulsor muy activo, junto con miembros destacados de la SEB y por supuesto Severo Ochoa, del Congreso FEBS en Madrid en 1969 que sirvió de espaldarazo internacional a la SEB en tiempos muy convulsos y difíciles para España. Este congreso logró congregar a un número considerable de científicos de muy alto nivel incluyendo a galardonados Nobel (Ochoa, Cori, Theorell, Krebs, Sanger, Lipman) que sirvieron de un escaparate impagable para el país y para la SEB.
Mi experiencia personal junto al Prof. Mayor Zaragoza empieza en la Facultad de Farmacia de la Universidad de Granada como alumna, y se ha extendido a lo largo de cincuenta años en el Departamento de Biología Molecular de la
Universidad Autónoma de Madrid y del Centro de Biología Molecular Severo Ochoa. De su trayectoria profesional me gustaría destacar varios hitos. En primer lugar, la implantación, primero a nivel local en Granada, y más tarde a nivel nacional, del Programa Diagnóstico de Enfermedades Moleculares con la valiosa ayuda de Magdalena Ugarte. La repercusión social y sanitaria de esta iniciativa ha evitado multitud de graves enfermedades metabólicas y genéticas de consecuencias devastadoras. En segundo lugar, la creación bajo la iniciativa y tutelaje de Don Severo Ochoa del Centro de Biología Molecular como centro mixto Consejo Superior de Investigaciones Científicas-Universidad Autónoma de Madrid junto a Eladio Viñuela, David Vázquez y Antonio GarcíaBellido. Centro matriz donde a lo largo de sus casi 50 años de existencia se han formado notables científicos que pertenecen, y en algunos casos lideran, otros prestigiosos centros de investigación.
De su dilatada trayectoria política elijo dos periodos: su paso por el rectorado de la Universidad de Granada, y su largo periodo en la Unesco como director general del organismo.
Me gustaría acabar esta breve reseña diciendo que a lo largo de tantos años he aprendido de Federico Mayor muchísimas cosas difíciles de enumerar, por lo que le estoy enormemente agradecida. Y lo que si me gustaría resaltar a nivel personal, son dos de sus cualidades humanas: el enorme respeto por su maestro Don Ángel Santos, y el gran amor por su familia.
FEDERICO MAYOR ZARAGOZA Y LA ESTACIÓN EXPERIMENTAL DEL ZAIDÍN (CSIC) DE GRANADA
Luis Alfonso del Río Legazpi
Profesor de Investigación ad honorem del CSIC (jubilado)
Departamento de Estrés, Desarrollo y Señalización en Plantas
Estación Experimental del Zaidín, CSIC, Granada
Araíz del reciente fallecimiento del Prof. D. Federico Mayor Zaragoza, aparte de sus innumerables méritos e importantes trabajos realizados en distintos campos, querría destacar el papel clave que desarrolló en su día en la creación de un Departamento de Bioquímica en la Estación Experimental del Zaidín (EEZ) del CSIC, de Granada. En 1968, siendo Federico Mayor Catedrático de Bioquímica en la Facultad de Farmacia de la Universidad de Granada y Rector de dicha Universidad, el Prof. Luis Recalde Martínez (qepd), Catedrático de Fisiología Vegetal y entonces Director de la EEZ, le sugirió al Prof. Mayor Zaragoza la creación de un grupo de Bioquímica en el Centro para abordar la problemática biológica de la planta desde un punto de vista molecular.
Este requerimiento fue rápidamente aceptado por el Prof. Mayor, y en la planta semisótano de un edificio del Centro, comenzaron a trabajar los Colaboradores Cientificos Julio López Gorgé (qepd) y Ana Chueca Sancho, y los doctorandos Juan Pedro Donaire Navarro (qepd) y Luis Alfonso del Río Legazpi, y más tarde se incorporaron Juan José Lázaro Paniagua y Angel de la Torre (qepd). Con el tiempo fueron llegando nuevos becarios, y obteniéndose plazas de investigadores en plantilla del CSIC y diversificándose las líneas de investigación. Lo que comenzó en 1968 siendo una Sección de Bioquímica, con los años se transformó en el Departamento de Bioquímica, Biología Celular y Molecular de Plantas. El proceso de evolución del Departamento está descrito en el libro “Estación Experimental del Zaidín.
Medio Siglo de Historia: 1955-2005". Editado por el Ministerio de Educación y Ciencia-CSIC-EEZ, en 2005. En la actualidad, el Departamento ha evolucionado y diversificado, y se denomina “Estrés, Desarrollo y Señalización en Plantas”. Está compuesto por 5 grupos de investigación, con un total de 24 personas en plantilla del CSIC (15 científicos y 9 técnicos), y 7 becarios pre- y postdoctorales. Los grupos de investigación actuales del Departamento son:
• Antioxidantes, Radicales Libres y Óxido Nítrico en Biotecnología y Agroalimentación (ARNOBA).
• Biología Reproductiva y Microscopía Avanzada de Plantas (BReMAP).
• Fisiología Molecular del Transporte Iónico en Plantas bajo Condiciones Ambiéntales Adversas.
• Regulación Redox, Señalización por Azúcares y Fenotipado mediante Técnicas de Imagen para la Detección de Estrés Vegetal.
• Señalización dependiente de ROS y Óxido Nítrico y Dinámica Peroxisomal en Plantas.
En conclusión, aquel Grupo de Bioquímica, creado con mucha ilusión por el Prof. Federico Mayor Zaragoza en 1968, que comenzó con tan solo 4 científicos, está compuesto en la actualidad por 31 personas, y tiene un buen número de proyectos de investigación y numerosas publicaciones anuales en revistas de alto impacto, habiendo obtenido renombre internacional la labor que allí se realiza en distintas especialidades de biología vegetal.
Esta fue la cosecha de aquella modesta siembra científica que hizo el Profesor Federico Mayor Zaragoza en la EEZ hace hoy 57 años. Descanse en paz.
EUGENIO VILANOVA GISBERT
(1948-2024)
Jorge Estévez, Carmen Estevan, Miguel A. Sogorb Universidad Miguel Hernández de Elche
El pasado día 2 de diciembre de 2024 falleció Eugenio Vilanova Gisbert, socio de la Sociedad Española de Bioquímica y Biología Molecular. Eugenio Vilanova nació en Villena (Alicante) y se graduó en Ciencias Químicas por la Universidad de Valencia. De su estancia como estudiante universitario nos destacaba siempre a los que fuimos sus discípulos su especial relación con el Colegio Mayor San Juan de Ribera, relación que mantuvo hasta el final de sus días. Cuando terminó sus estudios y recién casado con Leonor, ambos se mudaron a Bilbao, donde Eugenio Vilanova fue contratado como profesor de la Universidad del País Vasco. Tras dos años regresó a Alicante, donde contribuyó a la creación de la Universidad de Alicante y donde se doctoró con una tesis centrada en el estudio de la actividad tirosina hidroxilasa en tirosina inmovilizada.
Eugenio Vilanova continuó desarrollando su carrera docente e investigadora en la Universidad de Alicante, donde alcanzó una cátedra en Bioquímica. Posteriormente fue fundador de la Universidad Miguel Hernández de Elche, donde prestó sus servicios, ya dentro del área de conocimiento de Toxicología desde su creación en 1997 hasta su jubilación en 2018, cuando fue nombrado profesor emérito, figura bajo la cual continuó desempeñando su incansable labor investigadora, literalmente, damos fe de ello, hasta una semana antes de su deceso.
Dentro de la Universidad Miguel Hernández de Elche desempeñó varias labores, pero queremos destacar su papel primero como miembro fundador y posteriormente como director del Instituto de Bioingeniería. Bajo su dirección el Instituto de Bioingeniería se consolidó y expandió llegando a ser
uno de los centros puntales en la investigación de la Universidad Miguel Hernández.
El movimiento desde el área de Bioquímica a la de Toxicología no implicó en ningún momento un abandono de la Sociedad Española de Bioquímica y Biología Molecular ni de la Bioquímica, ni de la Química, ya que mantuvo su relación con el Ilustre Colegio de Químicos y con la Asociación de Química de la Comunidad Valenciana y siempre intentó entender los efectos adversos causados por xenobióticos sobre los seres vivos a través de una perspectiva molecular y mecanicista.
Las líneas de investigación desarrolladas por Eugenio Vilanova en Toxicología fueron variadas, incluyendo los mecanismos moleculares de neurotoxicidad y toxicidad al desarrollo embrionario de diversos plaguicidas, monitorización de exposición a disolventes orgánicos volátiles y metales pesados y evaluación de riesgos de exposición a fitosanitarios. Todo ello le llevó a publicar más de 150 artículos en revistas indexadas y más de 25 capítulos de libros científicos y a dirigir 18 tesis doctorales.
Eugenio Vilanova fue presidente de la Asociación Española de Toxicología desde 1995 a 2001 y también miembro de la comisión promotora de la Red Española para el Desarrollo de Métodos Alternativos a la Experimentación Animal (REMA), y vocal de la misma desde su constitución en 1999 hasta su fallecimiento.
La labor de Eugenio Vilanova fue reconocida internacionalmente por diversas asociaciones, como lo demuestra su nombramiento como miembro honorario de EUROTOX, y que EUROTOX e IUTOX le encargaran la organización de sus respectivos Congresos
Internacionales en 1996 y 2010 en Alicante y Barcelona, respectivamente.
Los que fuimos sus discípulos podemos dar fe de la calidad humana de Eugenio Vilanova, del que siempre recibimos su apoyo personal y profesional y un trato amable, respetuoso y considerado, tanto cuando estábamos de acuerdo con sus planteamientos y actuaciones como las escasas veces en que no lo estuvimos. Pero esta no es solo una impresión de los firmantes de estas líneas, creemos que nuestra opinión queda reforzada por el bonito homenaje que recibió en nuestra universidad en 2022, cuando personas que estuvimos en contacto con él en diversas etapas de su vida profesional, desde sus primeros pasos como profesor hasta el momento de su jubilación, acudimos a expresarle públicamente nuestro respeto y admiración. Algunas de las personas intervinientes hubieron de, literalmente, recorrer Europa para poder participar en el acto.
Eugenio Vilanova fue también una persona entusiasta, siempre dispuesta a colaborar en lo que se le pidiera, lo que le llevaba a embarcarse en cualquier causa para la que se le pidiera colaboración, y a abarcar simultáneamente un sinfín de actividades y colaboraciones en docencia, investigación, transferencia, gestión universitaria, difusión e incluso cooperación internacional, ya que una de sus últimas grandes causas fue contribuir al esclarecimiento de la etiología de la llamada nefropatía mesoamericana.
Su Villena natal siempre estaba en su cabeza. Era habitual que sorprendiera a la audiencia que le escuchaba por primera vez con diapositivas de Villena (especialmente del Castillo) introducidas en la presentación con la finalidad de rebajar el ritmo del discurso o de hacer pausas para enlazar temáticas.
También su familia estuvo siempre presente en nuestras charlas y conversaciones no laborales, los que tuvimos la suerte de conocerlo y poder aprender de él muchos años le hemos visto hablar mucho primero de sus hijos, y luego, conforme la familia crecía, de sus nietos, y siempre, siempre, siempre, de su esposa y compañera Leonor.
Quedan también para siempre en nuestra memoria las veladas veraniegas que organizaba en su casa de campo donde nos juntábamos junto a su familia, los miembros del equipo y nuestras familias, compañeros de la universidad, e investigadores visitantes del año en cuestión.
Queremos ofrecer nuestras más profundas condolencias a su viuda, Leonor, a sus cuatro hijos Héctor, Emma, Inés y Marcos, así como a sus yernos, nueras, nietos y hermanos.
Con la muerte de Eugenio sentimos que hemos perdido un compañero, un maestro y un amigo.
Descansa en paz Eugenio, nunca te olvidaremos.
Fotografía cedida por la familia de Eugenio Vilanova. De izquierda a derecha: Emma Vilanova, Héctor Vilanova, Eugenio Vilanova, Leonor Blanes, Inés Vilanova y Marcos Vilanova.
V EDICIÓN DEL PREMIO A LA MEJOR TESIS DOCTORAL EN BIOQUÍMICA Y BIOLOGÍA MOLECULAR PATROCINADO POR LA FUNDACIÓN LILLY
La SEBBM, con el patrocinio de la Fundación Lilly, convoca la V edición del Premio a la Mejor Tesis Doctoral en Bioquímica y Biología Molecular. El premio reconoce los trabajos de iniciación a la carrera investigadora que destaquen por su calidad científica. La convocatoria está dirigida a las/los socias/os de la SEBBM que hayan defendido su Tesis Doctoral en 2024. Se dará preferencia a los trabajos realizados por socias y socios con una antigüedad de al menos dos años y que hayan sido dirigidos por socias/os de la SEBBM. Se concederá un Primer Premio dotado con 2.000 € y dos accésits de 500 €. Las Tesis Doctorales que opten a los premios deberán tratar temáticas vinculadas con el área de la Bioquímica y la Biología Molecular, siendo en cualquier caso el jurado quien determine el cumplimiento
de este requisito. El plazo máximo para la recepción de candidaturas finaliza el 31 de marzo de 2025
El jurado, formado por los miembros de la Comisión de Admisiones de la SEBBM, comunicará los premios a los ganadores antes del 1 de julio de 2025. El jurado se reserva el derecho de declarar desierto el Premio Fundación Lilly a la Mejor Tesis Doctoral en Bioquímica y Biología Molecular, así como los accésits. La entrega del premio tendrá lugar durante el acto de clausura del 47º Congreso de la SEBBM a celebrar en Cáceces del 2 al 5 de septiembre. Los candidatos deben enviar al correo de la secretaría técnica (sebbm@sebbm.es) la documentación que se detalla en las bases del premio publicadas en la web de la SEBBM
PREMIOS A JÓVENES INVESTIGADORES
FUNDACIÓN BBVA-SEBBM
En 2025, con el patrocinio de la Fundación BBVA, la SEBBM convoca por tercer año consecutivo los premios a la Mejor Comunicación Joven, durante el 47º Congreso de la SEBBM, para cada uno de los dieciséis grupos científicos temáticos. Se concederá un premio de 1.500€ para cada uno de ellos (la dotación total de la convocatoria es de 24.000 €). Los premios están dirigidos a las/los socias/os adheridos de la SEBBM, tanto doctorandos como doctores recientes (dos años máximo), que se inscriban en el Congreso y presenten sus comunicaciones orales y/ o póster durante el mismo. Los aspirantes deben enviar la documentación, en formato digital, a través del formulario electrónico o bien por correo electrónico a sebbm@ sebbm.es, indicando en el mensaje el listado de documentos que se adjuntan o enlazan.
La fecha límite para el envío de la documentación es el 15 de mayo de 2025
Las bases del premio se pueden consultar en la web de la SEBBM
PREMIO IBUB-SEBBM
La SEBBM, junto con el Instituto de Biomedicina de la Universidad de Barcelona (IBUB), convoca el premio IBUB-SEBBM en reconocimiento a la trayectoria investigadora de los jóvenes socios/as de la SEBBM en el área de la Bioquímica y la Biología Molecular. El premio está dotado con 2.500 €. Las solicitudes pueden ser presentadas por los propios candidatos o ser propuestas por el Comité Organizador del 47º Congreso de la SEBBM – Cáceres 2025. El plazo para presentar las candidaturas finaliza el 15 de junio de 2025. El Comité Organizador del Congreso elegirá, entre todas las solicitudes recibidas, un máximo de cinco candidatos que serán propuestos a la Junta Directiva de la Sociedad. El candidato premiado dará una conferencia durante el 47º Congreso de la SEBBM que se anunciará como Conferencia IBUB-SEBBM. La entrega del premio tendrá lugar durante el acto de clausura del 47º Congreso de la SEBBM en Cáceres el 5 de septiembre 2025. Las bases del premio se pueden consultar en la web de SEBBM.
PRESENTACIÓN DEL NUEVO GRUPO SEBBM ‘MATRIX EXTRACELULAR’
El grupo SEBBM sobre Matriz Extracelular ha surgido por iniciativa de sus coordinadores, Fernando Rodríguez Pascual (CBM ‘Severo Ochoa’) y Ana V. Villar (IBBTEC), quienes asumirán esta responsabilidad en los próximos años. Con su reciente creación, queremos darnos a conocer y establecer un espacio de interacción y debate para investigadores interesados en los diversos aspectos de la Matriz Extracelular (ECM, por sus siglas en inglés).
Este grupo nace como un foro para fomentar la discusión en torno a la ECM, un componente clave en patologías como la fibrosis, el cáncer, la medicina regenerativa y la senescencia celular, entre otras. Los socios de la SEBBM que nos visiten en el congreso de este año o a través de nuestra página web podrán descubrir cómo un conocimiento profundo de este biomaterial dinámico, esencial para la comunicación intercelular y la estructura de los tejidos, ofrece
nuevas perspectivas en el estudio de enfermedades. La ECM no solo proporciona soporte mecánico y puntos de anclaje para las células, sino que también regula interacciones biológicas clave, influye en la evolución de patologías según su grado de rigidez y modula la fisiopatología de los órganos a través del matrisoma y sus múltiples componentes. Queremos destacar dos áreas de especial interés que abordaremos en el próximo congreso de la SEBBM en Cáceres:
• La fibrosis, caracterizada por un exceso de depósito de componentes de la ECM.
• El microentorno tumoral, que actúa como soporte y contribuye a la proliferación del cáncer. Además, animamos a otros grupos a unirse a nuestros debates y a enriquecer el estudio de la ECM con nuevas perspectivas. Estamos convencidos de que esta área de investigación en España crecerá rápidamente, como ya se está
viendo con iniciativas recientes como la Red de Investigación Integrada en Fibrosis Orgánica (REDFIBRO), financiada por la Agencia Estatal de Investigación (AEI) de la que somos miembros los coordinadores de este grupo SEBBM sobre Matriz Extracelular.
III CONFERENCIA SEVERO OCHOA: «EPIGENÉTICA, ENVEJECIMIENTO
Y CÁNCER»
Los días 26 y 27 de junio de 2025 tendrá lugar, en la Laboral Ciudad de la Cultura de Gijón, la III Conferencia Severo Ochoa, organizada por la SEBBM en colaboración con la Fundación Carmen y Severo Ochoa (FCySO). El programa cuenta con un fantástico grupo de ponentes invitados: Steve Horvath (UCLA, Altos Labs), Sara Haag (Karolinska Institutet), Andrew Teschendorff (Shanghai Institute of Nutrition and Health), Maria Blasco (CNIO), Iñaki Martin-Subero (Hospital Clinic Barcelona), Christophe Bock (Medical University of Vienna) y Manel Esteller (Josep Carreras Leukemia Research Institute). Además, habrá sesiones de pósteres y se seleccionarán charlas breves entre ellos.
La III Conferencia Severo Ochoa está organizada por Mario F. Fraga (CINN-CSIC, IUOPA, ISPA), Ana Gutiérrez (Universidad de Oviedo, IUOPA, ISPA) y Agustín F. Fernández (CINN-CSIC, IUOPA, ISPA). La inscripción permanecerá abierta hasta el 30 de abril, con un número limitado de plazas disponibles. Los miembros de SEBBM disfrutarán de una tarifa de inscripción reducida. Para más información consultar nuestra web
III
«EPIGENETICS, AGING AND CANCER»
Invited Speakers
Steve Horvath (UCLA, Altos Labs)
Sara Haag (Karolinska Institutet)
Andrew Teschendorff (Shanghai Institute Nutrition Health)
Maria Blasco (CNIO)
Iñaki Martin-Subero (Hospital Clinic Barcelona)
Christophe Bock (Medical University of Vienna)
Manel Esteller (Josep Carreras Leukemia Research Institute)
Local organizers:
Mario F Fraga (CINN-CSIC, IUOPA, ISPA)
Ana Gutierrez (University of Oviedo, IUOPA, ISPA)
Agustín F. Fdez. (CINN-CSIC, IUOPA, ISPA)
Scientific committee:
Xose Puente (University of Oviedo, IUOPA, ISPA)
Ramón Tejedor (CINN-CSIC, IUOPA, ISPA)
Jose María Pérez Freije (University of Oviedo, IUOPA, ISPA)
SEVERO OCHOA CONFERENCE:
LA SEBBM COLABORA EN LA I SEMANA SEVERO OCHOA
Isabel
Varela Nieto, Regina Revilla y Antonio Ferrer
La I Semana Severo Ochoa 2024 se celebró del 4 al 7 de noviembre de 2024 en Madrid, organizada por la Real Academia Nacional de Medicina de España (RANME) en colaboración con la Fundación Carmen y Severo Ochoa (FCySO), la Universidad de Nebrija y la Sociedad Española de Bioquímica y Biología Molecular (SEBBM). Durante esta semana, se llevaron a cabo actividades para honrar la figura y el legado del Dr. Severo Ochoa, premio Nobel de Fisiología o Medicina en 1959. Aparte de las reseñas introducidas por Federico Mayor y Vicente Rubio sobre la figura de Severo Ochoa y el desarrollo de la bioquímica y la biología molecular en España, se realizó la lectura de la autobiografía de Severo Ochoa con la participación de estudiantes de bachillerato y Universidad. Además, se hizo entrega del Premio Carmen y Severo Ochoa de Investigación en Biología Molecular a María Llorens-Martín, y María Blasco impartió la conferencia conmemorativa 2024. La agenda de actividades también incluyó la visita al Centro de Biología Molecular Severo Ochoa en el que intervinieron su directora, Paola Bovolenta, Miguel Manzanares, José A. Esteban y Ana Ortega. Entre los eventos destacados, se incluyó una exposición titulada “Severo Ochoa: un científico apasionado”, continuación de la que, diseñada por Inmaculada Yruela, se inauguró en el museo Domus de A Coruña durante el 46º Congreso de la SEBBM y permaneció expuesta durante el mes de septiembre de 2024. Esta muestra, organizada conjuntamente por la SEBBM y
la FCySO, ofreció una visión detallada de la vida y contribuciones científicas del Dr. Ochoa, resaltando su influencia en la modernización y promoción de la ciencia en España. Además, se destacó su papel en la constitución de la Sociedad Española de Bioquímica (SEB) y en su incorporación a la Federación Europea de Sociedades de Bioquímica (FEBS). En el marco de la I Semana Severo Ochoa, la RANME y la SEBBM firmaron un acuerdo de colaboración, suscrito por el Dr. Eduardo Díaz-Rubio García, presidente de la RANME y el Dr. Antonio V. Ferrer Montiel, presidente de la SEBBM. Este convenio tiene como objetivo promover la cooperación en la organización de actividades de formación y difusión, participación en conferencias, foros, cursos de especialización y seminarios de actualización, así como el apoyo a actividades de divulgación científica. Gracias a este acuerdo, se facilitará el intercambio de publicaciones y la colaboración técnica en la realización de informes, estudios, proyectos y otras iniciativas conjuntas. Además, permitirá la difusión de actividades, premios y becas organizadas por ambas instituciones a través de sus respectivos canales de comunicación. Siendo una actividad central la colaboración en la organización anual de la Semana Ochoa. La I Semana Severo Ochoa 2024 representó una oportunidad única para reflexionar sobre la importancia de la ciencia y la investigación en la sociedad actual, tomando como referente la figura del Dr. Severo Ochoa, cuyo legado continúa inspirando a nuevas generaciones
de científicos y profesionales de las biociencias y la biomedicina en España y en el mundo. En esta misma línea, destacar que en paralelo se estaba preparando el libro “Severo Ochoa: una vida por la ciencia”, publicado por Editorial Planeta y el CSIC en diciembre del 2024. Esta obra biográfica profundiza en los logros científicos y personales de Ochoa, destacando su trabajo pionero en la síntesis de ARN y su papel en la creación de la SEBBM, consolidando su legado como un referente de la ciencia moderna. La obra ha contado con la estrecha colaboración entre la SEBBM y la FCySO, que comparten el compromiso de divulgar el legado del científico. La participación de la SEBBM en este proyecto editorial refleja su compromiso con la difusión de la historia de la bioquímica y la biología molecular en España, así como con el reconocimiento de figuras clave como Severo Ochoa, cuyo trabajo ha sido fundamental para el desarrollo de estas disciplinas en el país. Queremos destacar que el libro está prologado por Don Federico Mayor Zaragoza, en la que posiblemente haya sido su última contribución literaria. La I Semana Severo Ochoa 2024 no solo celebró a un científico de renombre, sino que también invitó a reflexionar sobre la importancia de la ciencia en la sociedad actual. El ejemplo de Ochoa, basado en la pasión por el conocimiento y el rigor científico, continúa inspirando a nuevas generaciones, consolidándose como un modelo de excelencia y compromiso que trasciende fronteras y generaciones.
PREMIO PRINCESA DE ASTURIAS 2024 DE LA INVESTIGACIÓN CIENTÍFICA
Y TÉCNICA
Los cinco creadores de los fármacos Ozempic y Wegovy, han sido premiados con el Premio Princesa de Asturias de Investigación Científica y Técnica 2024 Daniel J. Drucker, médico, (Montreal, Canadá, 1956); Jeffrey M. Friedman, biólogo molecular, (Orlando, EE. UU, 1954); Joel F. Habener, endocrinólogo, (Estados Unidos 1937); Jens Juul Holst, químico, (Copenhague, Dinamarca, 1945); y Svetlana Mojsov, química, (Skopie, Macedonia del Norte y Estados Unidos 1947) son líderes mundiales en el campo de la endocrinología.
Las investigaciones de los científicos galardonados han establecido las bases endocrinas de la diabetes y la obesidad, patologías
prominentes que son un problema global de salud pública sin tratamiento efectivo hasta la fecha. Jeffrey M. Friedman ha sido pionero en establecer la base genética de la hormona que regula el apetito. Daniel J. Drucker, Joel F. Habener, Jens Juul Holst y Svetlana Mojsov han estudiado el efecto de determinadas hormonas que regulan la secreción de insulina y consecuentemente los niveles de glucosa. Estas investigaciones han permitido por primera vez el desarrollo de fármacos eficaces para combatir la diabetes y la obesidad. Además, permiten mitigar patologías asociadas como las cardiovasculares. Estos tratamientos, ya disponibles, están mejorando la calidad de vida de cientos de millones de personas en todo el mundo.
En los últimos años se ha producido un gran avance en el tratamiento de la diabetes tipo 2, con la aparición de fármacos que utilizan como principio activo la semaglutida, un péptido semejante a una hormona, el glucagón-1 o GLP-1, que juega un papel de contrapeso de la insulina en el equilibrio del azúcar en sangre. Cuando el nivel de azúcar baja, el glucagón induce al hígado a liberar glucosa y cuando sube se genera más insulina, que se encarga de reducir el exceso. Además, la semaglutida produce una notable reducción del apetito, lo que ha convertido en un éxito al Ozempic, uno de los fármacos producidos con este principio activo. La revista Science designó a estos fármacos contra la obesidad
Figura 1
Daniel J. Drucker, Jeffrey M. Friedman, Joel F. Habener, Jens Juul Holst y Svetlana Mojsov - Premio Princesa de Asturias (premiosprincesa.es)
como el mayor avance científico de 2023. Drucker, Habener, Holst y Mojsov comparten el reconocimiento de haber iniciado y desarrollado esta investigación desde los años setenta del pasado siglo. Desde sus diferentes laboratorios estudiaron las hormonas que intervienen en el proceso y regulan el metabolismo digestivo, como la somatostatina, que inhibe la producción de glucagón y de insulina, y variantes del glucagón, denominadas GLP-1 y GLP-2, y comprobaron que este sistema de homeostasis podía ser una diana terapéutica efectiva contra la diabetes de tipo 2. La semaglutida (como algunas otras moléculas semejantes) funciona como agonista del receptor del glucagón GLP-1, por lo que inhibe la producción de esta hormona, reduciendo los niveles de azúcar en sangre y mejorando el crecimiento de
las células beta-pancreáticas, responsables de la producción y liberación de insulina. Además, se ha comprobado que tiene efectos protectores de accidentes vasculares en adultos con obesidad, indicación autorizada recientemente por la Administración de Alimentos y Medicamentos estadounidense (FDA, por sus siglas en inglés). Por su parte, Friedman descubrió en 1994 otra hormona, la leptina, que se genera en las células grasas o adipocitos y actúa sobre la región cerebral que controla el apetito. Se trata de un sistema en equilibrio: cuanta más grasa hay más leptina se produce, lo que disminuye el apetito, reduciendo la grasa del organismo y por tanto la producción de leptina. En el caso de los obesos este mecanismo está desequilibrado. También han estudiado la predisposición genética a la obesidad.
PREMIO FRONTERAS DEL CONOCIMIENTO
DE LA FUNDACIÓN BBVA EN BIOMEDICINA
Cuatro de los anteriores galardonados con el Premio Princesa de Asturias, Daniel J. Drucker, Joel Habener, Jens Juul Holst y Svetlana Mojsov han recibido también el Premio que concede la Fundación BBVA por lograr descubrimientos biológicos para el desarrollo de una nueva generación de fármacos eficaces contra la obesidad y la diabetes. Además, sus descubrimientos han mostrado un potencial más allá de estos objetivos y hoy se investiga
su impacto en enfermedades neurológicas como el párkinson y el alzhéimer, así como trastornos de la adicción, como el alcoholismo. “Este es una de las áreas más fascinantes en la que vamos investigar cómo funcionan y cuál es el impacto. Exploramos los mecanismos que inciden en los procesos de inflamación en las enfermedades neurodegenerativas”, ha comentado Daniel J. Drucker, del Hospital Monte Sinaí Toronto (Canadá). Jens Juul Holst, de
la Universidad de Copenhague (Dinamarca) ha señalado los estudios que se realizan sobre “el centro de recompensa del cerebro y su impacto en las conductas adictivas que derivan en la dependencia del alcohol y otras sustancias”.
Los padres del 'Ozempic' reciben el Premio Fronteras del Conocimiento de la Fundación BBVA en Biomedicina | Salud (elmundo.es)
WORKSHOP
“PLANTS
UNDER ENVIRONMENTAL STRESS: OVERCOMING CURRENT CLIMATE CHANGE”
Mónica Balsera y Francisco Javier Cejudo Socios SEBBM
Los días 4 al 7 de noviembre de 2024 se celebró en el campus de la Universidad Internacional de Andalucía (UNIA), Palacio de Jabalquinto en Baeza (Jaén), el workshop internacional “Plants under Environmental Stress: Overcoming Current Climate Change”. Organizado por Francisco Javier Cejudo (IBVF, US-CSIC, Sevilla, España), Mónica Balsera (IRNASA-CSIC, Salamanca, España) y José M. Estévez (Instituto Leloir, Buenos Aires, Argentina), este workshop fue financiado por la UNIA dentro de su programa “Environment Workshops 2024” y contó con el apoyo de la Agencia Estatal de Investigación a través de la Red de Excelencia “Redox in Plants”, de las sociedades científicas SEBBM y SEBP, y los institutos IBVF e IRNASA.
El objetivo central del workshop fue el de discutir estrategias biológicas para abordar la crisis climática provocada por el calentamiento global, dado que los organismos fotosintéticos constituyen un mecanismo natural de disminución de CO2 atmosférico. Para ello, el workshop reunió a destacados investigadores en biología vegetal de diez países en un entorno excepcional para el intercambio de ideas y la discusión de los últimos avances en biología de plantas en relación con el cambio climático. Para abordar los objetivos científicos, el workshop se estructuró en 16 conferencias invitadas, charlas cortas y presentaciones en forma de póster, que se organizaron en tres sesiones:
La primera sesión abordó la optimización del metabolismo
fotosintético y la señalización del cloroplasto, explorando temas como la regulación redox, la biología sintética y la adaptación de las plantas al estrés ambiental. Se presentaron investigaciones sobre la maquinaria fotosintética como sensor del estrés ambiental, avances en optogenética, procesos de captura de carbono por microalgas y la comunicación plastidio-núcleo, además de respuestas mitocondriales y la dinámica de los fotosistemas del cloroplasto bajo estrés abiótico.
La segunda sesión se enfocó en la aclimatación de las plantas a cambios ambientales. Se discutieron temas como el papel de los peroxisomas en la dinámica redox, los mecanismos moleculares de la señalización del sulfuro, las respuestas a la disponibilidad de nitrato, y la regulación del desarrollo de pelos radiculares en ambientes fríos y deficientes en nutrientes. Además, se abordó la función de hemoglobinas en leguminosas y el análisis
bioinformático del estrés salino en olivo y cítricos.
La última sesión se centró en los avances biotecnológicos en plantas y algas para mejorar la productividad y la sostenibilidad agrícola. Se discutieron temas como estrategias de bioingeniería para la fijación de nitrógeno en plantas, la optimización de la fotosíntesis en cultivos sostenibles, y la biotecnología de musgos con aplicaciones industriales. También se discutió sobre factores de transcripción, el estrés nitro-oxidativo y las respuestas a fitófagos.
La sesión concluyó con una charla sobre señales oxidativas derivadas de la fotosíntesis.
Los debates generaron nuevas ideas para mejorar la tolerancia de las plantas al estrés ambiental y optimizar su uso biotecnológico y fomentaron colaboraciones futuras. El éxito del workshop subraya la importancia de estos encuentros para avanzar en la investigación y consolidar la cooperación internacional en biología vegetal.
47º CONGRESO DE LA SOCIEDAD ESPAÑOLA DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR (SEBBM)
Guadalupe Sabio Buzo Presidenta del 47º Congreso SEBBM
Estimados asistentes, Es un verdadero honor darles la bienvenida al 47º Congreso de la Sociedad Española de Bioquímica y Biología Molecular (SEBBM), que tendrá lugar en la histórica y encantadora ciudad de Cáceres del 2 al 5 de septiembre de 2025. Este evento representa un hito significativo, ya que por primera vez nuestra reunión anual se celebra en Extremadura, una región que combina una riqueza cultural única, tradiciones y paisajes únicos.
El programa de este año refleja nuestro compromiso inquebrantable con la excelencia científica y la diversidad temática. Estamos encantados de contar con destacados conferenciantes plenarios que nos inspirarán con sus descubrimientos y perspectivas innovadoras:
• Dr. Roger Davis (University of Massachusetts Medical School, USA), quien abrirá el congreso con la conferencia inaugural titulada “Molecular Biology and Inflammation: New Insights”.
• Dra. Alexandra Newton (University of California, San Diego, USA), quien impartirá la prestigiosa conferencia IUBMB sobre “Phosphoinositide Signaling in Health and Disease”.
• Dr. Alejandro Villa (Universidad Nacional de Córdoba, Argentina), quien protagonizará la conferencia Luis F. Leloir, cortesía de la Sociedad Argentina de Investigación Bioquímica y Biología Molecular.
• Dra. Paola Pizzo (University of Padua, Italia), que ofrecerá la FEBS National Lecture titulada “Calcium Signaling in Cell Death and Survival”.
• Dra. Ana María Cuervo (Albert Einstein College of Medicine, USA), quien cerrará el congreso con la conferencia de clausura “Selective Autophagy: Healthy Aging One Protein at a Time”.
Además, contaremos con un variado programa de simposios, talleres y actividades diseñados para fomentar el intercambio de ideas y fortalecer nuestra comunidad científica. Algunos de los simposios destacados incluyen:
• Hispano-Luso Symposium: Aging, Cancer and Telomeres: Mechanisms and Therapeutic Frontiers.
• The Heart at the Core: Novel Insights into Cardiovascular Pathophysiology.
• Autophagy in Human Diseases.
• Nutrient Signaling: From Signaling Transduction to Disease.
• DNA Break Repair in the CRISPR Era.
• Application of Cutting-edge Proteomics in Biomedical and Clinical Research.
• Advances and Challenges in Structure-Based Drug Discovery.
El programa también incluirá eventos sociales que brindarán a los asistentes la oportunidad de disfrutar de la exquisita gastronomía y el patrimonio cultural de Cáceres, declarada Patrimonio de la Humanidad por la UNESCO Quiero expresar mi más sincero agradecimiento a los patrocinadores, colaboradores y al comité organizador por su dedicación y esfuerzo para hacer posible este congreso. Espero que este encuentro sea una experiencia enriquecedora y memorable para todos.
¡Bienvenidos a Cáceres y al 47º Congreso SEBBM!
CONGRESO DE LA SOCIEDAD ESPAÑOLA DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR 47º
CÁCERES
congresos.sebbm.es/caceres2025
DEL 2 AL 5 DE SEPTIEMBRE 2025
UNA REIVINDICACIÓN NECESARIA DE LA BIOQUÍMICA VEGETAL
J. M. Mulet
Instituto de Biología Molecular y Celular de Plantas IBMCP, CSIC-UPV
Plantas que nos ayudan
La ciencia detrás del conocimiento ancestral de las plantas medicinales Rosa Porcel | Destino | Barcelona (2024) | 356 páginas
Cuando vemos un título donde aparecen “plantas” y “salud” en la misma línea, tendemos a pensar en programas de televisión como “La botica de la abuela” o en personajes tan poco rigurosos como Txumari Alfaro o Josep Pàmies. Por suerte ha salido una visión del lado de la ciencia. Plantas que nos ayudan de Rosa Porcel asume un desafío considerable: hablar de plantas medicinales sin caer en la pseudociencia y, al mismo tiempo, ofrecer una visión accesible y entretenida para el lector, y lo consigue. El gran mérito de este libro radica en su capacidad para equilibrar historias antropológicas que recogen tradiciones de diferentes culturas relacionadas con las plantas medicinales con el rigor científico. Es evidente que la autora ha dedicado tiempo a investigar los usos históricos de las plantas, desde el poder del ajo como antimicrobiano hasta las propiedades antiinflamatorias de la cúrcuma. Sin embargo, lo que realmente destaca es la habilidad para contextualizar estas afirmaciones en el marco de lo que sabemos hoy en día, sobre la base de estudios fiables y, lo más importante, poniendo límites claros a las promesas milagrosas y, a la vez, haciendo la lectura entretenida. Por ejemplo, cuando se aborda el tan cacareado tema de los antioxidantes, la autora no se pierde en mitos modernos ni deja que el marketing de la industria alimentaria dicte el guión. En lugar de eso, explica de manera sencilla cómo estos compuestos funcionan realmente en el cuerpo humano, sin ofrecer ilusiones de eterna juventud.
Otro de los aciertos es que el libro es claramente de divulgación científica. No se plantea como un recetario o un libro de autoayuda. No estamos ante una lista de recetas mágicas ni de pócimas imposibles de preparar. En su lugar, se ofrece una visión sensata sobre los efectos que pueden tener ciertas plantas en la vida diaria, ya sea a través de la alimentación o en forma de complementos ocasionales, pero sin olvidar las advertencias necesarias sobre dosis y posibles contraindicaciones. Y lo más importante, si alguien tiene un problema de salud… que vaya al médico.
Este equilibrio entre lo útil y lo riguroso es raro en libros de esta temática, y aquí es donde Plantas que nos ayudan realmente brilla. Sí, hay plantas que pueden ayudarte con las digestiones pesadas, como el hinojo, pero no esperes que cure un virus. Si algo queda claro tras leer este libro, es que la autora es
ante todo científica, y que pertenece a un departamento de bioquímica. A lo largo de sus páginas, se citan estudios relevantes y se desmontan mitos de forma sistemática pero amable, sin caer en un tono soberbio o arrogante. Por ejemplo, cuando trata la eterna cuestión de si el aloe vera es, de verdad, el santo grial de los remedios naturales, la autora explica sus propiedades reales (hidratante, antiinflamatorio tópico) sin inflar su reputación ni menospreciarla. Es un enfoque equilibrado que, a diferencia de otros textos del género, no hace que el lector termine confuso entre exageraciones y escepticismo extremo. Uno de los grandes logros del libro es su capacidad para comunicar ideas complejas de manera sencilla. La autora no solo conoce bien su materia, sino que sabe cómo transmitirla de forma que cualquier lector pueda seguirla, ya sea un profesional de la salud o alguien que simplemente quiere saber si añadir cúrcuma a su dieta tiene algún sentido. Además, el estilo es fresco, con toques de humor y una pizca de ironía que hacen que la lectura sea agradable. En lugar de ser un manual pesado, Plantas que nos ayudan se presenta como una conversación amena en la que aprendes mientras disfrutas. A veces tienes la sensación de que la autora te está contando cosas al oído.
En resumen, este es un libro que destaca por su rigor, su enfoque práctico y su capacidad para entretener. En un panorama saturado de libros que glorifican propiedades que las plantas no tienen hasta niveles absurdos, o que adoptan enfoques pseudocientíficos esta obra se erige como un faro de sentido común. Es una lectura recomendable para cualquier persona interesada en la relación entre plantas y salud, sin importar su nivel de conocimiento previo. Y lo mejor de todo: cuando terminas el libro, te quedas con la sensación de haber aprendido algo útil, sin la necesidad de ir corriendo a ninguna herboristería.
Así que, si estás buscando un libro que te ayude a entender las plantas desde un punto de vista científico pero accesible, con una base bioquímica rigurosa, este es el indicado. Solo un consejo: lee con una taza de té al lado. ¿De qué planta? De la que más te guste. Que no existan las curaciones mágicas no quiere decir que una infusión caliente no te pueda hacer un rato de lectura más agradable. Sin necesidad de promesas mágicas. Ni caso a Pàmies.
Análisis de datos NGS en proyectos de investigación biomédica
Dreamgenics lleva más de una década siendo una de las empresas de referencia en España en el análisis bioinformático de datos provenientes de secuenciación masiva (NGS). Cada año ayudan a decenas de investigadores de universidades, hospitales y centros de investigación a obtener una mayor y mejor comprensión de las bases moleculares de las enfermedades humanas a través del estudio del ADN y del ARN, un proceso de gran complejidad que consiguen llevar a cabo gracias al trabajo en equipo y a la elevada formación de todos sus profesionales.
¿Qué tipo de servicios ofrecéis en Dreamgenics? Somos especialistas en análisis bioinformático de datos NGS y la gran mayoría de los análisis que realizamos a nivel de investigación son exomas y RNA-Seq, aunque recientemente hemos añadido también a nuestra cartera de servicios el análisis Single cell RNA-Seq y la integración de datos multiómicos. Además, este año hemos comprado un robot de preparación de librerías y un secuenciador para poder ofrecer internamente el servicio de secuenciación y reducir los tiempos de entrega de resultados al no depender de proveedores externos. Ha sido una inversión muy importante y estamos seguros de que servirá para dar un mejor servicio a nuestros clientes.
¿Qué diferencia vuestro servicio del ofrecido por otras empresas?
Destacaría dos aspectos; nuestra adaptabilidad a las necesidades del cliente y nuestra plataforma Genome One Reports. Respecto al primero, tenemos la capacidad de ofrecer a nuestros clientes un servicio completo que incluye desde la recepción de las muestras biológicas en nuestro laboratorio hasta la entrega de los resultados, lo que nos permite ofrecer siempre un servicio adaptado a sus necesidades. En segundo lugar, está nuestra plataforma web Genome One Reports, a través de la que entregamos los resultados de nuestros análisis. Este es un aspecto diferenciador muy importante, ya que mientras otras empresas entregan hojas Excel y PDF, nosotros utilizamos una plataforma interactiva que facilita a los investigadores trabajar con sus resultados. Por ejemplo, en un análisis de exoma se obtienen de media unas 250.000 variantes, por lo que es mucho más práctico para el cliente utilizar nuestra plataforma Genome One Reports para hacer filtrados, revisiones de variantes, etc. que trabajar con hojas de Excel. Además, el uso de esta plataforma no tiene ningún sobrecoste, ya que está incluido en el precio del análisis.
Carlos Martínez, CEO de Dreamgenics
¿Cómo es vuestra manera de trabajar con el cliente cuando os solicitan un servicio?
Normalmente nos contactan por email o a través del formulario de nuestra página web. Hay ocasiones en las que el cliente ya tiene claras las características del experimento que quiere realizar y, en otras, tiene una idea general de lo que está buscando, pero necesita asesoramiento técnico. Este asesoramiento inicial es una parte fundamental para nosotros en cualquier nuevo servicio, ya que existen muchas opciones posibles a nivel de secuenciación (por ejemplo; tipo de RNA que se quiere estudiar, librerías, profundidad de lectura, etc.) que es necesario elegir adecuadamente para intentar obtener los mejores resultados posibles y, además, nos gusta hacer una demo de nuestros entregables en Genome One Reports para cada tipo de análisis para que el cliente conozca la plataforma y se vaya familiarizando con ella. Una vez realizados los análisis, les damos unas credenciales de acceso y hacemos una nueva reunión para comentar con ellos los resultados obtenidos, si lo desean.
Actualmente tiene cada vez mayor importancia el aseguramiento de la Calidad por parte de las empresas sobre los productos y servicios que ofrecen, ¿cómo es en vuestro caso?
Desde 2019 estamos certificados por AENOR en las normas ISO 9001, ISO 13485 de productos sanitarios para nuestro software Genome One, y en la norma ISO 27001 de seguridad de la información. Además, nuestro software también tiene marcado CE-IVD para diagnóstico in vitro, al igual que el robot de preparación de librerías y el secuenciador que utilizamos en el laboratorio. También participamos cada año en programas externos de Calidad a través de diversos esquemas EQA de EMQN y GenQA en los que obtenemos excelentes resultados. Estamos convencidos de que todo ello es una buena demostración de nuestro compromiso con desarrollar nuestro trabajo con el máximo nivel de exigencia posible y bajo estrictos estándares de Calidad. Al final, se trata de llevar a cabo un proceso de mejora continua a todos los niveles, buscando siempre avanzar y mejorar para ofrecer a nuestros clientes una cartera de productos y servicios cada vez más amplia y que se ajuste a sus necesidades.
Monitoreo del cambio metabólico de las células inmunes durante la diferenciación
Introducción
Los monocitos y macrófagos son componentes cruciales del sistema inmunitario innato. Los monocitos, producidos en la médula ósea, circulan en la sangre y se diferencian en macrófagos al infiltrarse en los tejidos. Cuando los macrófagos se activan en un fenotipo proinflamatorio, dependen de la glucólisis para obtener energía, un fenómeno conocido como el efecto Warburg, que también se observa en las células cancerosas. Sin embargo, en las células inmunes, este efecto puede indicar envejecimiento y muerte celular en lugar de supervivencia y proliferación. El perfil metabólico de las células inmunes varía según su estado de actividad, con los monocitos y los macrófagos M2 mostrando un aumento en la oxidación de ácidos grasos y la fosforilación oxidativa, mientras que los macrófagos M0 y M1 exhiben una glucólisis aumentada. Comprender estos perfiles metabólicos puede proporcionar información sobre la diferenciación de las células inmunes.
Materiales y Métodos
Células THP-1, una línea celular de monocitos, se cultivaron en medio RPMI 1640 con 10% de FBS durante 96 horas. Las concentraciones de glucosa y lactato en el medio de cultivo se midieron continuamente utilizando el LiCellMo™ de PHCbi, un analizador metabólico en tiempo real de células vivas. Se añadió PMA (forbol 12-miristato 13-acetato) para inducir la diferenciación en macrófagos M0. Los grupos de control recibieron solo el medio (DMSO). Los niveles de glucosa y lactato se midieron al inicio y al final del período de cultivo. La morfología celular se examinó utilizando microscopía de contraste de fases y la expresión de CD11b. Se utilizó citometría de flujo para evaluar un marcador de macrófagos durante los experimentos.
Resultados
El analizador metabólico de células vivas proporcionó mediciones precisas de las concentraciones de glucosa y lactato, con una desviación máxima del 14% en comparación con el método colorimétrico (Figura A). Tras la adición de PMA a las 48 horas, las tasas de consumo de glucosa y producción de lactato aumentaron, indicando una glucólisis mejorada (Figura B). La duración de la medición fue de 96 horas. El grupo de control tuvo un mayor recuento celular al final del cultivo, lo que sugiere un mayor consumo de glucosa por célula y producción de lactato debido al PMA (Figura C). La microscopía confirmó que los monocitos, inicialmente no adherentes, se convirtieron en macrófagos adherentes tras la adición de PMA (Figura D). La citometría de flujo mostró un aumento en la expresión de CD11b, confirmando la diferenciación en macrófagos (Figura E).
Discusión
El estudio demostró que los monocitos, que inicialmente dependen de la oxidación de ácidos grasos y la fosforilación oxidativa, cambian su metabolismo a la glucólisis al diferenciarse en macrófagos. Sin embargo, 12 horas después de la adición de PMA, la tasa metabólica comenzó a disminuir, probablemente debido al aumento de la concentración de lactato en el medio, lo que suprime la glucólisis y reduce la función de las células inmunes. Este fenómeno se observa en el microambiente tumoral, donde los altos niveles de lactato inhiben las respuestas inmunitarias. El estudio sugiere que monitorear los cambios metabólicos puede proporcionar información valiosa sobre la diferenciación y función de las células inmunes.
Conclusión
Utilizando el LiCellMo™ de PHCbi, un analizador metabólico en tiempo real de células vivas, este estudio midió cronológicamente los cambios metabólicos durante la diferenciación de monocitos a macrófagos. El perfil metabólico de las células inmunes está estrechamente relacionado con su estado de diferenciación, y observar estos perfiles puede ayudar a evaluar la diferenciación de las células inmunes. Esta técnica permite el monitoreo en tiempo real de los cambios metabólicos, ofreciendo nuevas perspectivas sobre la inmunometabolismo y posibles aplicaciones en la investigación metabólica.
Nota: Los resultados mostrados y el equipo LicellMo son únicamente para fines de investigación.
Para más información o preguntas sobre el artículo o el equipo LiCellMo™, por favor contacte a: Adrian Baumann adrian.baumann@eu.phchd.com
Anna Tomas anna.tomas@eu.phchd.com
Referencias
1. Warburg O. On the origin of cancer. Science 1956;123(3191):309-314.
2. Dietl K, Renner K, Dettmer K, et al. Lactic Acid and Acidization Inhibit TNF Secretion and Glycolysis of Human Monocytes. J Immunol 2010;184(3):1200-1209.
3. Kempner W. The nature of leukemic blood cells as determined by their metabolism. J Clin Invest 1939;18 (3):291-300.
4. Nagy C, Haschemi A. Time and demand are two critical dimensions of immunometabolism: The process of macrophage activation and the pentose phosphate pathway. Front Immunol 2015;6(164):1-8.
5. Cao J, Zeng F, Liao S, Cao L, Zhou Y. Effects of glycolysis on the polarization and function of tumour-associated macrophages (Review). Int J Oncol 2023;62(70):1-12.
6. Abuawad A, Mbadugha C, Ghaemmaghami AM, Kim DH. Metabolic characterisation of THP-1 macrophage polarisation using LC–MS-based metabolite profiling. Metabolomics 2020;16(33):1-14.
7. Palmer CS, Cherry CL, Sada-Ovalle I, Singh A, Crowe SM. Glucose Metabolism in T Cells and Monocytes: New Perspectives in HIV Pathogenesis. EBioMedicine. 2016;6:31-41.
8. Fischer K, Hoffmann P, Voelkl S, et al. Inhibitory effect of tumour cell-derived lactic acid on human T cells. Blood 2007;109 (9):3812-3819.
Con mas de 2.900 socios y 19 grupos científicos, la SEBBM es la principal agrupación científica en su campo.
Contribuimos a que la Bioquímica y la Biología Molecular tengan un papel relevante en nuestra sociedad, favoreciendo el contacto entre investigadores e impulsando a los jóvenes científicos.
Pero no estamos solos...
Los socios protectores contribuyen al progreso de la SEBBM
Porque son de los nuestros*
Más información sobre la figura de socio protector en: sebbm@sebbm.es o llamando al +34 681 916 770
*Serán socios protectores aquella entidades que quieran contribuir al sostenimiento y desarrollo de la SEBBM y sean aceptadas como tales. Tendrán derecho a voto en las asambleas, pero no podrán ser elegibles para cargos directivos.