







Por su demostrada eficacia y seguridad, LIDERFEED es el ÚNICO GENUINO PROMOTOR DE CRECIMIENTO español aprobado por EFSA para la UE

(Register of Feed Additives pursuant to Regulation (EC) Nª 1831/2003 – Annex I.List of Additives https://ec.europa.eu/food/sites/food/files/safety/docs/ animal-feed_additives_eu-register_1831-03.pdf) Plaza García Lorca, 15 Bajos Tfno: (+34) 977 552316 43006 Tarragona (Spain) email: lidervet@lidervet.com
Es cierto que vivimos en un tiempo bastante movido. La situación de los diferentes mercados de productos ganaderos, en su conjunto, es bastante más que buena. Precios relativamente altos, márgenes razonables, buenas perspectivas internacionales para algunos productos…
Curiosamente, esto debería significar una cierta tranquilidad empresarial, pero, al contrario, estamos viendo una gran cantidad de movimientos de compras de empresas productoras, en prácticamente todas las especies.
No deja de ser curioso que, precisamente en este periodo de relativa (o no tan relativa) bonanza, se produzcan todos estos procesos de compras, fusiones o readaptaciones. Quizá para empresas que han pasado momentos muy difíciles en el pasado sea un buen momento de conseguir, al fin, beneficios. Puede que por medio haya cambios generacionales, puede que sea casualidad, pero no deja de ser notable el enorme movimiento de mercado de estos meses.
Y es evidente que aún veremos nuevos movimientos, como para no dejar de estar entretenidos.

Por otra parte, aun en situaciones buenas de mercado tenemos riesgos ciertos. Desde posibles movimientos en el plano internacional, por aranceles USA a nuevos acuerdos comerciales de la UE, a procesos patológicos (véase la lengua azul, la PPA o la influenza) que pueden hacer peligrar las producciones o la comercialización de algunos productos. Necesariamente hay que mantener la guardia alta. Cuando estás arriba, la dirección probable en el futuro es el descenso.
En cualquier caso, como me explicaron en catequesis, el Señor te lo da, el Señor te lo quita, Bendito sea el nombre del Señor… En este caso, que el Señor nos da, sólo queda agradecerle por su merced, y, como mucho, pedirle que dure todo lo posible… y que seamos prudentes para no estropear el regalo.

José Ignacio
Barragán Consultor avícola independiente
EDITOR
GRUPO DE COMUNICACIÓN AGRINEWS S.L.
PUBLICIDAD
Laura Muñoz +34 629 422 552 laura@mediatarsis.com
Luis Carrasco +34 605 09 05 13 lc@agrinews.es
DIRECCIÓN TÉCNICA
José Ignacio Barragán (aves)
Guillermo Fondevila (porcinos)
Lourdes Cámara (porcinos)
Fernando Bacha (rumiantes)
COORDINACIÓN TÉCNICA
Facundo Apecetche
REDACCIÓN
Amanda Abalo
Daniela Morales
COLABORADORES
Juan Acedo-Rico (Consultor)
Joaquim Brufau (IRTA)
Lorena Castillejos (SNiBA)
Carlos De Blas (UPM)
Gonzalo Glez. Mateos (UPM)
Xavier Mora (Consultor)
Edgar O. Oviedo-Rondón
Alba Cerisuelo (CITA-IVIA)
Carlos Fernández (UPV)
ADMINISTRACIÓN
Mercè Soler
Tel: +34 93 115 44 15
info@grupoagrinews.com www.nutrinews.com
Precio de suscripción anual: España 30 €
Extranjero 90 €
GRATUITA PARA FABRICANTES DE PIENSO, EMPRESAS DE CORRECTORES Y NUTRÓLOGOS
Depósito Legal Nutrinews B-17990-2015
Impreso - ISSN 2696-8053
Digital - ISSN 2696-8045
se hace responsable de las opiniones de los autores. Todos los derechos reservados. Imágenes: Dreamstime, Freepik, Adobe Stock.
EDICIÓN TRIMESTRAL
Aplicación de carbohidrasas en la alimentación animal: Eficiencia nutricional y sostenibilidad
Rafael Durán Giménez-Rico y Félix Calvo Falcón Danisco Animal Nutrition & Health

Tablas nutriNews Actualizaciones 2025 PREBIÓTICOS, PROBIÓTICOS, Y OTROS
Tablas nutriNews Actualizaciones 2025 ÁCIDOS ORGÁNICOS Y/O GRASOS
El papel de la fibra en la alimentación de cerdas reproductoras

Guillermo Fondevila1 Lourdes Cámara2
1Universidad de Zaragoza
2Universidad Politécnica de Madrid

El arte de alimentar cerdas reproductoras: inteligencia artificial al servicio de la nutrición y productividad

Gustavo Márquez Esteban1 , Pablo Fuentes Pardo2 y Sara Crespo Vicente 3

1Técnico veterinario Cefusa especialista en maternidad Técnico veterinario Cefusa I+D+i especialista en nutrición
3Técnico veterinario Cefusa & Departamento de Fisiología de la Universidad de Murcia
Destetando al lechón, un mal de siempre ¿Cómo lo facilitamos?
Lorena Pérez Esteruelas Servicio técnico de porcino en Elanco. PM-ES-25-0389.
46
Análisis bromatológico y solubilidad en KOH 0,2% de muestras de harina de soja obtenidas en Venezuela con procedencia de EE.UU.

Manuel Vázquez
REVEEX NUTRICION C.A.
56
La ciencia oculta de la vitamina A en las aves de corral: ¿Por qué la fuente del producto, su estabilidad y biodisponibilidad son importantes?
Yauheni Shastak, Wolf Pelletier, Adriana Barri
BASF SE, División Nutrición y Salud
64
El nuevo complejo zinc(II)betaína permite reducir un 50% el cinc en los piensos para broilers sin afectar su productividad
Dr. Rubén Crespo Sancho1 y Dr. Susanne Rothstein2
1Technical Manager, Biochem Zusatzstoffe GmbH, Alemania
2R&D Product Manager Organic Minerals, Biochem Zusatzstoffe GmbH, Alemania

68
Un pequeño repaso de la importancia de la fibra neutro detergente en rumiantes

Fernando Bacha Baz Nacoop S.A., Director técnico rumiantes nutriNews España
80
Efecto de la subnutrición durante el último tercio de gestación y la raza en el equilibrio pro- y anti-oxidante de la sangre en terneros recién nacidos de aptitud cárnica








88
Escalera-Moreno N.1 , ÁlvarezRodríguez J. 3 , Martín-Alonso M.J.1 , Villalba D.1 , Molina E.1 , López de Armentia L. 2 , Sanz A. 2 , Serrano-Pérez B.1
1Dpto. Ciencia Animal, UdL, Lleida, España;
2CITA Aragón - IA2 (UNIZAR), Zaragoza, España;
3Escuela politécnica superior-IA2, UNIZAR, Huesca, España.
Los carbohidratos en raciones para gatos ¿alternativa energética o enemigo silencioso? Parte II

Ada Lugo Médico Veterinario y Nutricionista, Asesora independiente
94
Informe materias primas ASFAC
Rafael Durán Giménez-Rico y Félix Calvo Falcón Danisco Animal Nutrition & Health
Los carbohidratos, que incluyen los azúcares más simples, el almidón y varios de los polisacáridos no amiláceos (PNA estructurales, y de almacenamiento), la fuente de energía más importante para monogástricos y rumiantes.
Los PNA y la lignina (completamente indigestible) son los dos componentes principales de las paredes celulares; bien es cierta la presencia de PNA también de naturaleza intracelular.


Arabinosa y xilosa (pentosas) y las hexosas, glucosa (la principal)
Galactosa
Manosa
Las paredes celulares además se asocian a proteínas con más o menos cuerpos fenólicos e incluso con lignina, lo que hace de estos PNA, una fracción realmente difícil de digerir, si no fuera con el uso de enzimas exógenas.
A su vez, los polisacáridos de mayor presencia en los ingredientes vegetales son la celulosa (polímero de glucosas), los arabinoxilanos (polímeros de arabinosa y xilosa) y los beta-glucanos (polímeros de glucosas unidas por distintos enlaces glicosídicos).


Cabe añadir que la lignina en unión a los PNA comprende lo que se denomina fibra dietética (FD).
Los nutrientes de mayor digestibilidad en los ingredientes, en mayor o menor medida se hallan recubiertos por los diferentes PNA, según la zona en la que se encuentren.
Por tanto, son utilizados ineficazmente, ya que ni cerdos ni aves poseen las actividades enzimáticas que pudieran, eventualmente, digerir/degradar eficazmente los PNA.
Consecuentemente, con el uso de las actividades enzimáticas exógenas adecuadas, podemos asegurar que el valor intrínseco de esos ingredientes utilizados para formular los piensos, aumentará de forma directa con la calidad de aquellas.
No todas las carbohidrasas servirían de igual manera. Con este breve trabajo pretendemos resaltar la que creemos, basándonos en años de experiencia
Las carbohidrasas, especialmente xilanasas y beta-glucanasas, se utilizan ampliamente en la alimentación animal para degradar polisacáridos no amiláceos (PNA) presentes en cereales (trigo, cebada, centeno, incluso maíz).
Estas enzimas, sobre todo en el caso de los cereales “viscosos”, reducen la viscosidad del contenido intestinal, sobre todo en animales jóvenes, mejorando la interacción entre enzimas digestivas y los sustratos diana; de forma inmediata, se favorece la absorción de nutrientes.
Además, y debido a un efecto de degradación de estructuras fibrosas más complejas, liberan otros nutrientes contenidos en el endospermo de dichos cereales.

Asimismo, en el caso de las proteínas de origen vegetal, las estructuras más complejas ya no son solo arabinoxilanos y beta-glucanos, sino también galactomananos, pectinas y otros carbohidratos más complejos sin valor nutricional y difícilmente aprovechables,

En dietas basadas en maíz y soja, los efectos con carbohidrasas exógenas, pueden resultar más variables, aunque se observan mejoras en digestibilidad ileal y retención de nutrientes, también en función de qué actividades se elijan.

La beta-mananasa también ha mostrado efectos positivos en aves y cerdos, mejorando la ganancia diaria, conversión alimenticia y cierta mejoría del metabolismo energético (Kiarie et al., 2021). Sin embargo, en nuestra experiencia, no es una actividad enzimática que muestre un porcentaje de pruebas con monogástricos satisfactorias, como para considerarla candidata a utilizarse junto a otras.
En relación con el apartado anterior, según Ravindran (XXVI Curso de Especialización FEDNA 2010), la información científica disponible sugiere que las respuestas a las enzimas son dependientes, entre otros, de la calidad de los ingredientes; a menor calidad, mayor será la magnitud de las mejoras por la suplementación enzimática.
Ahondando en esta dirección, y dado que convendremos que el nivel de PNA -fibra dietética en general- es un factor anti nutricional (por tanto, determinante de “la calidad” como fuente potencial de energía de un ingrediente), los diferentes niveles de aquellos y sus características en cada uno de las materias primas usadas, suponen una referencia para predecir una mayor o menor eficacia de las carbohidrasas exógenas elegidas como más relevantes.
La Tabla 1 reúne a las carbohidrasas más empleadas y a sus actividades principales (Menezes-Blackburn et al., Functional Polymers in Food Science: From Technology to Biology capítulo 4, “Enzymes used in Animal Feed”, marzo 2015).
Enzima Número EC Sustrato
EC 3.2.1.39
Beta-glucanasa (celulasa)
Xilanasa
Mananasa
Galactosidasa
Pectinasa
EC 3.2.1.4
EC 3.2.1.6
EC 2.1.73
Producto degradación
Beta-glucanos y Celulosa Glucosa
EC 3.2.1.8 Arabinoxilanos Xilosa
EC 3.2.178
EC 3.2.1.25
EC 3.2.1.22
EC 3.1.1.11
EC 4.2.2.10
Galacto-mananos Heterosacáridos
PNA con grupos galactósidos
EC 3.2.1.15 Sustancias pécticas
Diversos complejos
Mono, di y oligo galacturanos
Tabla 1: Enzimas exógenas seleccionadas como aditivos y actividad principal.
Desde nuestro punto de vista y basándonos en todo el trabajo de investigación y desarrollo de actividades enzimáticas que se realiza de forma continua en IFF Danisco Aninal Nutrition, la combinación de xilanasa y beta-glucanasa es la que, de forma muy superior respecto a las demás, mayores y más estables respuestas confirman a través de pruebas experimentales.
Incluso, en nuestra experiencia, otras actividades diferentes a éstas, como las incluidas en la Tabla 1, son de mayor inconsistencia.
Por otro lado, y tomando como referencia la diferencia en presencia de sustrato entre distintos PNA (abajo se observa en tres dietas sencillas, Tabla 2), la diferencia existente entre la suma de AX/BG frente a la de mananos es sustancial.
Dieta 1
Tabla 2: Contenido en PNA y mananos de 3 dietas simples, cereales y HS48. Los mananos fueron calculados por medio de la tabla ß-Mannan & Xylan Contents (www.ChemGen.com), los PNA, base de datos IFF-Danisco Animal Nutrition. Ingredientes
Dieta 2
Dieta 3
El hecho de que las enzimas exógenas “encuentren” un sustrato suficiente, es de importancia máxima , de cara a obtener una respuesta en forma de mejoras de digestibilidad (de todos los nutrientes que los PNA encapsulan) y de rendimientos como resultado de esa mejoría de la digestibilidad. De ahí que la preferencia por nuestro lado es seguir desarrollando las actividades principales, xilanasa/ beta-glucanasa, incluso en el desarrollo de nuevos productos.
Llegados a este punto, habiendo subrayado la importancia de las xilanasas/betaglucanasas, no queremos olvidar la eficacia que las amilasas (no mencionadas en la tabla) pueden aportar en dietas de monogástricos, esencialmente en dietas de avicultura (basados en nuestra experiencia en pollos).

En trabajos realizados durante los últimos años en busca de mejoras en el desarrollo de las carbohidrasas más adecuadas a cada tipo de dieta, encontramos respuestas de relevancia con el empleo de solo una amilasa (Tabla 3).

Mejora calórica vs CN,
a los 42 días (1-42)
Tabla 3. Mejora calórica e IC en dietas de pollos de 1-42 días (maíz /harina soja 48). TR-XAP.NZ.B.30 (IFF, Danisco Animal Nutrition). ab: (P<0,05). (ab): (P<0,10)
El uso individual de una amilasa mejorada, desarrollada recientemente por IFF, Danisco Animal Nutrition, nos muestra el potencial de ésta, mejorando incluso la conversión calórica del tratamiento positivo (maíz/HS48) a los 42 días, en 14 kcal/kg de ganancia.
La mejora de la amilasa mencionada queda reflejada en un metaanálisis mostrado en la siguiente figura (IFF, Danisco Animal Nutrition),
Figura 1: Mejora calórica con el uso de la nueva amilasa. EMAn, kcal/kg MS, en metaanálisis de 4 pruebas a 21 días de vida con dietas maíz/HS48/DDGS.
En dos pruebas (Figura 2), realizadas con dietas a base de maíz/HS48, DDGS de maíz y harina de colza, la combinación xilanasa/ beta-glucanasa mejoró significativamente el IC y la digestibilidad ileal de la energía a los 21 días.
En las mismas, se observó una mejora de ambas digestibilidades ileales, la del almidón -de manera significativa- y la de la grasa (P<0,10).

Por otra parte, estudios más recientes, en los que se aplica la combinación xilanasa/ beta-glucanasa en dietas con niveles elevados de PNA, demuestran la eficacia de esta combinación. Abajo en la figura 3 se muestra la eficacia del complejo X/BG mediante el efecto sobre el IC a 42 días de vida de los pollitos.
+0,37MJ/kg (2,6%)
La enzima XBG añadida 100g/t (1220 U xilanasa / 152 U beta-glucanasa), mejoró el IC significativamente en todas las dietas; interesante observar que, a medida que el nivel total de PNA aumentaba, 10,8 – 11,6 –12,0 la respuesta fue superior, 1,8% - 3,0% - 3,9%.
14,41b
P<0,05
Figura 2: Energía digestible aparente ileal (MJ/kg MS). ab: valores con diferente índices son diferentes significativamente (P<0,05)
Bajaharina Colza Bajaharina ColzaXBG BajaharinaColzaGirasol AltaharinaColzaGirasol BajaharinaColzaGirasol XBG AltaharinaColzaGirasol XBG
Figura 3: IC de pollos a 42 días, alimentados dietas de diferentes niveles de ingrdientes fibrosos. TR. XB.NL.B.32 (Tipo de dieta P<0,01; XGB P<0,05; Dieta * XBG NS).

No es la intención de este breve trabajo en extenderse explicando cuáles son los retos de la alimentación y la producción animal en relación a la sostenibilidad; para ello me permito recomendar tres referencias recientes de excelentes revisiones (Coma y col., FEDNA 2021 y FEDNA 2022; Ana Gutiérrez Portugal, Nutrinews, marzo 2023).
Lo que queremos mostrar, como resultado de una de tantas pruebas realizadas en los últimos años (Neuro Livestock Research, South Africa, TR pendiente de publicarse) es el efecto de la combinación de dos enzimas exógenas en pollos de 1 a 32 días.
Como consecuencia de la correcta aplicación de nuestra fitasa de última generación en dosis decrecientes junto a las carbohidrasas X/BG comentadas, los resultados principales son:
Reducción del coste de la alimentación (9,8%)
Mantenimiento del peso canal
Reducción de la huella de carbono (17,1%)
Huella de carbono, g CO2 eq/kg ganancia PV (1-32 dias)
La eficacia de las carbohidrasas exógenas está directamente relacionada con la presencia del sustrato adecuado/específico y en cantidad adecuada; lógicamente, queremos utilizar las carbohidrasas que tengan mayor especificidad con los PNA presentes en las dietas.
Los PNA están compuestos de diferentes componentes. Los principales en los cereales son: la celulosa, los arabinoxilanos,y los beta-glucanos; en mucha menor medida, xilanos, arabinogalactanos, galactomananos, etc.
Aplicación de carbohidrasas en la alimentación animal: Eficiencia nutricional y sostenibilidad
DESCÁRGALO EN PDF
propia pendiente de publicarse; Neuro Livestock Research, South Africa). Fitasa a 2000, 1500 y 1000 FTU/kg arranque, crecimiento y finalizador y 100 gramos X/BG/tn.
Coste alimentación (euro/kg ganancia PV)
Desde la experiencia de IFF, Danisco Animal Nutrition con el empleo de carbohidrasas en los diferentes mercados en los que trabajamos, y las distintas composiciones de dietas, la combinación de xilanasa y beta-glucanasa se destaca como la mejor opción (sin dejar de lado la eficacia mostrada en estudios diversos de la amilasa).

El empleo de fitasa de última generación junto a una combinación X/BG y sus correspondientes matrices ajustadas, permite una reformulación que, manteniendo los rendimientos, permite reducir los coste de producción y un efecto reductor de la huella de carbono del 17,1%.


Xilanasa y beta-glucanasa en su justa medida
• Mejora la digestibilidad de dietas complejas
• Reduce los costes de alimentación
• Permite dosis variables en busca del beneficio óptimo
• Estable hasta los 90oC info.animalnutrition@iff.com | animalnutrition.iff.com


“Tabla de Prebióticos” PON EN GOOGLE


“Tabla Ácidos Orgánicos” PON EN GOOGLE

Guillermo Fondevila1 y Lourdes Cámara2
1Universidad de Zaragoza 2Universidad Politécnica de Madrid


La fibra dietética desempeña un papel clave en numerosos procesos fisiológicos de las cerdas, como la sensación de saciedad, la motilidad del tracto gastrointestinal, la incidencia de estreñimiento y el comportamiento nervioso (Guillemet et al., 2007; Jarrett y Ashworth, 2018; Mateos et al., 2019).

En este sentido, los efectos de incorporar diferentes fuentes de fibra en el pienso están determinados en gran medida por sus propiedades físico-químicas (solubilidad, capacidad de retención de agua y grado de lignificación), lo que ofrece beneficios sobre la fisiología y el funcionamiento digestivo que varían según el nivel de inclusión y la fase productiva considerada.
Por ello, es esencial seleccionar cuidadosamente la fuente y nivel de fibra más adecuados en función de sus características para su incorporación en los piensos para cada fase productiva, para optimizar tanto el rendimiento productivo como el bienestar de las cerdas reproductoras.
Las cerdas hiperpolíficas actuales presentan una mayor prolificidad y menor engrasamiento que las genéticas anteriores, lo que exige ajustar la curva de ingestión para asegurar la recuperación de las reservas corporales durante la gestación y favorecer el consumo voluntario de pienso en la lactación, mejorando así la supervivencia y el crecimiento de la camada.
No obstante, la restricción alimenticia puede alterar el comportamiento y comprometer el bienestar de las cerdas gestantes. En este contexto, la inclusión de niveles elevados de fibra en el pienso de gestación estimula la peristalsis de la pared intestinal y disminuye la velocidad de tránsito de la digesta, mejorando el confort digestivo y reduciendo el riesgo de mortinatos.
En estos piensos, es recomendable priorizar el uso de fuentes de fibra soluble, como la pulpa de remolacha, por su mayor efecto saciante. Además, las fuentes de fibra soluble no lignificada fermentan fácilmente en el intestino grueso de las cerdas, generando ácidos grasos volátiles que contribuyen a la salud intestinal, reducen el pH de la digesta y aportan una fuente adicional de energía para las células epiteliales.

Por ello, la utilización de fibra soluble resulta clave para mejorar el confort digestivo y disminuir la agresividad y los comportamientos estereotípicos en las cerdas (Souza da Silva et al., 2012; 2013).
Un factor adicional a considerar es que, durante el periodo de preinseminación, la inclusión de niveles extra de fibra en la dieta puede mejorar la supervivencia prenatal, especialmente en cerdas primerizas (Jarrett y Ashworth, 2018). Aunque la causa exacta de esta mejora no se conoce con exactitud, podría relacionarse con un mejor desarrollo del ovocito dentro del folículo ovárico, lo que favorecería una mayor supervivencia embrionaria.

El periodo de transición o peri-parto comprende los 7-10 días anteriores y posteriores al parto (Theil, 2015). Desde el punto de vista nutricional, es la fase más crítica del ciclo productivo de la cerda, ya que debe afrontar profundos cambios fisiológicos y adaptarse al paso de un pienso de gestación a otro de lactación (Theil et al., 2015; Pedersen et al., 2019).
Por este motivo, el tipo de dieta suministrada a las cerdas reproductoras durante este periodo ha cobrado un creciente interés en los últimos años. Dada la breve duración de esta fase y las limitaciones logísticas de muchas granjas comerciales, se recomienda cambiar al pienso de lactación el mismo día en que las cerdas entran a las parideras.
No obstante, en algunas circunstancias, y siempre que la logística de la granja lo permita, puede resultar útil emplear un pienso específico para el peri-parto, con valores nutricionales intermedios entre los piensos de gestación y lactación (Theil et al., 2022).
En este sentido, la inclusión de fuentes de fibra en la dieta de periparto favorece el proceso del parto y acorta su duración, reduciendo la mortalidad neonatal y mejorando la viabilidad de los lechones (Darroch et al., 2008; Feyera et al., 2017).
Además, la fibra estimula el peristaltismo digestivo y disminuye el estreñimiento en los días anteriores y posteriores al parto (Jarrett y Ashworth, 2018), reduciendo también la incidencia de edema mamario, mastitis y agalaxia, lo que favorece la producción de leche durante la lactación (Oliviero et al., 2009).

Es importante considerar que tanto la incidencia de estreñimiento como la condición corporal de la cerda son factores clave que influyen en la duración del proceso del parto y en la producción de calostro (Oliviero et al., 2010; Loisel et al., 2013; Quesnel et al., 2012).
El objetivo principal del periodo de lactación es maximizar la producción de leche y el crecimiento de la camada, por lo que el manejo y la composición del pienso durante esta fase resultan fundamentales para optimizar los resultados productivos a lo largo de toda la vida reproductiva de la cerda.
Durante la primera fase de la lactación, aunque la ingesta de pienso es limitada, la movilización de reservas corporales en cerdas multíparas supone un riesgo reducido, ya que la producción de leche también es menor. Por contra, en las cerdas primíparas, las necesidades de crecimiento son todavía elevadas, por lo que un bajo consumo de pienso tras el parto puede derivar en problemas metabólicos relevantes.

Debe tenerse en cuenta que niveles excesivos de fibra dietética, especialmente cuando provienen de fuentes de fibra soluble, pueden disminuir la palatabilidad y el consumo voluntario de pienso, lo que podría convertirse en un problema a partir de los 5 días post-parto, un periodo en el que las necesidades nutricionales para la producción de leche aumentan drásticamente (Jarrett y Ashworth, 2018).

Por esta razón, aunque el consumo total de fibra sea mayor en la cerda lactante que en la gestante como consecuencia de la mayor ingestión de pienso, las dietas de lactación incluyen niveles de fibra inferiores a los de gestación para evitar pérdidas excesivas en la condición corporal y favorecer la ingesta de nutrientes.
En este contexto, la inclusión controlada de cantidades suficientes de fibra en el pienso de lactación ayuda a reducir el estreñimiento y mejora el confort digestivo de la cerda, aportando energía extra gracias a la producción de ácidos grasos volátiles en el intestino grueso (Theil et al., 2022).
Reforzamos nuestra presencia en Iberia!
Nos complace anunciar la actualización de la estr uctura de asociación comercial de nuestros productos.

Sigue siendo el socio comercial de confianza para
Para optimizar la inmunidad respiratoria

Solución ecaz para controlar la diar r hea
El factor X en la gestión de los cocci
Ahora es el socio comercial exclusivo autorizado para
Alter nativa más ecológica al clor uro de colina


Vidara gestionará en exclusiva la promoción y distribución de Kolin Plus™ FC en toda Iberia.
¿Qué significa esto para usted?
Fiabilidad ininterr mpida de s s prod ctos de confian a Soporte técnico sobre el terreno mejorado
Una cadena de s ministro n compromiso local más f ertes
Para
Para Vidara:
Gracias por seguir confiando en nosotros mientras nos fortalecemos juntos.
Para más información o cons ltas, póngase en contacto con s respec o representante local de Vidara o Vedia
Director Sol ciones Nat rales Director de Especialidades Técnicas Msc
En resumen, el uso adecuado de fuentes de fibra en las dietas de cerdas reproductoras contribuye a mejorar aspectos productivos y fisiológicos fundamentales, incluyendo el bienestar animal, la reducción del estreñimiento, la disminución de los comportamientos estereotípicos y la mejora del rendimiento reproductivo.
Las fibras solubles ejercen un efecto saciante prolongado en la cerda gestante, una respuesta que depende en gran medida de la fermentabilidad de la fuente utilizada, lo que ayuda a reducir la mortalidad prenatal y favorece el consumo voluntario de pienso durante la lactación. Asimismo, el aumento del contenido de fibra dietética en el pienso durante los periodos de peri-parto y pre-inseminación se perfila como una estrategia efectiva para optimizar los rendimientos reproductivos.
Por su parte, durante la lactación, la incorporación equilibrada de fuentes de fibra contribuye a disminuir la tasa de estreñimiento y mejora el confort digestivo de la cerda. No obstante, deben evitarse niveles excesivos que puedan comprometer el consumo de pienso y, en consecuencia, afectar tanto la condición corporal como la producción de leche.

El papel de la fibra en la alimentación de cerdas reproductoras DESCÁRGALO EN PDF



• Favorece el equilibrio de la microbiota intestinal
• Mantiene la sanidad del sistema digestivo
• Mejora la fisiología digestiva
• Mejora los parámetros productivos
• Disminuye el uso de antibióticos
Gustavo Márquez Esteban1, Pablo Fuentes Pardo2 y Sara Crespo Vicente3
1Técnico veterinario Cefusa especialista en maternidad
2Técnico veterinario Cefusa I+D+i especialista en nutrición
3Técnico veterinario Cefusa & Departamento de Fisiología de la Universidad de Murcia

En un contexto de alta exigencia productiva, la alimentación de precisión se ha consolidado como una herramienta clave para mejorar la eficiencia y el bienestar en las granjas de cerdas hiperprolíficas. La inteligencia artificial y la automatización permiten ajustar las raciones de forma individualizada, optimizando tanto la productividad como la salud de los animales.
El cambio actual de la producción implica la búsqueda de mayores rendimientos económicos mediante selección genética y estrategias productivas que permitan obtener:
Mayor productividad numérica
Mayor velocidad de crecimiento
Mayor eficiencia productiva
Estos parámetros afectan directamente a los costes de producción y a la rentabilidad de la empresa (Kyriazakis et al., 2006; Piñeiro et al., 2025).

Tradicionalmente en producción porcina, la alimentación de la cerda ha consistido básicamente en “echarle de comer” de forma que quedaran cubiertos sus requerimientos nutricionales según su estado fisiológico y etapa dentro de este, estado corporal, etc., todo ello con un criterio subjetivo.
Así, la correcta alimentación de la cerda dependía de la disponibilidad de personal en la granja, de sus horarios, días de trabajo, reducciones de plantillas y horas laborales los fines de semana y festivos, etc.
En los últimos años la alimentación de la cerda ha evolucionado a un aporte de la ración de forma automatizada, monitorizada y de precisión, tanto en la fase de gestación como en la fase de lactación.
Esto permite una alimentación más objetiva, ajustada de manera casi milimétrica a los requerimientos nutricionales, para conseguir así una óptima producción.
Otro factor que afecta directamente a la rentabilidad es la nutrición, tanto de forma directa a través del precio del pienso, como indirecta a través de su efecto en el crecimiento del lechón (Menegat et al., 2024).
La nutrición y el manejo de la alimentación para alcanzar la ingesta necesaria son pilares fundamentales para lograr el crecimiento adecuado de camadas de cerdas hiperprolíficas (Sulabo et al., 2014).
Ello también contribuye a lograr una mejor condición corporal en el momento del destete de las madres y, a su vez, a un un menor intervalo entre el destete y la siguiente gestación, mejorando los rendimientos de la siguiente camada.
Cubrir las necesidades nutricionales de las cerdas en diferentes momentos del ciclo productivo, a través del adecuado manejo de la alimentación y de la estrategia nutricional, es uno de los principales retos a los que se enfrenta el sector porcino.
En muchas ocasiones las cerdas son subalimentadas, ya que las curvas de alimentación se diseñan de una forma estándar, descuidando en cierto modo las necesidades individuales de los animales.
Los nuevos sistemas de alimentación electrónica permiten suministrar la ración diaria, sin importar el día de la semana, la presencia o ausencia de personal y la hora del día.

Una vez establecida la curva de alimentación, la cerda dispone del pienso en el momento que ella lo demande, siendo más que una alimentación ad libitum, una alimentación a demanda que se ajusta a los momentos en que la cerda tiene apetencia, se siente segura, responde a sus dictados etológicos o a su ritmo circadiano.

Para el buen funcionamiento de una alimentación de precisión, es indispensable diseñar una curva bien calculada, basada en todas las características y condicionantes de la granja y de la cerda.
Una vez establecida, deberá ser valorada y recalculada las veces que sea necesario, hasta conseguir que las curvas de pienso ofrecido y de pienso consumido como media, en el rebaño, se ajusten de forma íntimamente paralela.
Los sistemas electrónicos de alimentación de precisión trabajan, en primer lugar, en base a una curva de alimentación estándar (Gráfica 1) establecido según aspectos específicos de la granja como:
Genética de las cerdas
Valores nutricionales del pienso administrado
Clima y época del año
Ciclo reproductivo de la cerda (nulípara, primeriza o cerda adulta)
Condición corporal media en la granja
Estado sanitario general de la granja
Histórico de datos de consumo de la granja
Experiencia previa
Cantidad de pienso ofrecido (g)
8.000
7.000 6.000 5.000 4.000 3.000 2.000 1.000
Días relativos al parto de la cerda primeriza Nivel de alimentación diaria
Gráfica 1. Ejemplo de curva de periparto y lactación de cerdas primerizas en una granja comercial (Fuente propia).
A partir de la curva de alimentación estándar, se podrán hacer las modificaciones necesarias para obtener diferentes curvas de alimentación adaptadas (Gráfica 2), que se aplicarán de la forma más individualizada posible a cada cerda, teniendo en cuenta las particularidades de cada animal y las variaciones en los parámetros previamente mencionados (Aparicio et al., 2025b).
2.800
Cantidad de pienso ofrecido (g)
Días desde el inicio de la ingesta en maternidad
Nivel de alimentación diaria
Gráfica 2. Ejemplo de curva adaptada de maternidad en cerda enferma con aumento paulatino de alimentación durante los 5 primeros días de ingesta (Fuente propia).
En el caso en el que la curva no esté bien adaptada, se recomienda trabajar con un 20 % adicional de alimentación, de forma que las cerdas que queden con hambre en su última comida tengan la oportunidad de poder llegar a comer un mayor porcentaje de la curva estándar (Gráfica 3).
Si existe un amplio porcentaje de animales por encima del 15-20 % de la curva de alimentación significa que la curva está mal diseñada y se debe aumentar la cantidad de alimentación total por tomas (Gráfica 3B).
El diseño de los principales parámetros de alimentación automática se muestra en la Gráfica 3.
A Porcentaje de animales de más de 3 días sin comer.
>3 días sin comer
1,14 % animales que llevan más de 3 días sin comer.
B Porcentaje de animales con alimentación superior al umbral máximo del 100 % de ingesta diaria estimada (solicitando el 20 % adicional).
Exceso de alimentación
86 % de los animales con alimentación superior al umbral máximo del 100 % de ingesta diaria. Necesidad de adaptación de curva urgente.
C Porcentaje de animales con alimentación inferior al umbral mínimo de 50 % de ingesta diaria estimada.
< umbral mínimo de alimentación
2,28 % animales con alimentación inferior al umbral mínimo del 50 % de ingesta diaria.
Gráfica 3. Patrones de identificación de consumo en curvas de alimentación. A. Porcentaje de cerdas que llevan más de 3 días sin comer. B. Porcentaje de cerdas superior al umbral máximo de alimentación. C. Porcentaje de cerdas inferior al umbral mínimo de alimentación (Fuente propia, n=175).
El pienso de gestación está adaptado a las necesidades de las cerdas gestantes, con el objetivo de:
Asegurar un desarrollo embrionario equilibrado.
Evitar sobreengordes o carencias nutricionales que puedan afectar la salud reproductiva.

Favorecer la preparación para el parto y la lactación.
Su fórmula es moderada en energía, pero rica en fibra y minerales, promoviendo la salud digestiva y el bienestar de la cerda a lo largo de toda la gestación (Bruun et al., 2023).
La cantidad de pienso suministrada en gestación depende mucho de la genética y condición corporal de la cerda, oscilando entre 2 kg y 2,6 kg
Los sistemas de alimentación automatizada en gestación permiten administrar la ración de pienso a lo largo de 24 horas, comenzando a las 14:00 para poder localizar a las cerdas rezagadas antes, dentro del horario laboral, valorarlas, instarlas a comer y así evitar que salgan en alarma al cambio de día.
Esto permite observar los patrones habituales de ingesta de las cerdas, entre los cuales destaca que un 14,7 % de los animales se alimenta después de las 0:00, aprovechando las horas de menor temperatura (n = 340) (Gráfica 5).
Media 12-14mm de grasa dorsal Nulíparas Óptimo Gorda >15 mm de grasa dorsal
Gráfica 4. Variación de la cantidad de pienso suministrado a las cerdas durante la gestación en función de su condición corporal y los días postcubrición (Fuente: Manual de Alimentación Danbred).
Número de cerdas alimentadas

Hora del día (h)
Gráfica 5. Patrón de alimentación observado en cerdas (n=340) alimentadas mediante sistemas de alimentación automática de precisión en gestación durante las 24 horas del día.
Diseñado específicamente para los días previos y posteriores al parto, esta dieta de transición:
Prepara el sistema digestivo para el cambio a pienso de lactación.
Reduce el riesgo de estreñimiento, frecuente en esta etapa.
Aporta los micronutrientes clave para un parto sin complicaciones y una recuperación más rápida.
Los piensos periparto contienen una combinación equilibrada de fibra funcional, prebióticos y minerales biodisponibles pensada para suavizar el cambio metabólico y mejorar el bienestar, tanto de la madre como de los lechones al nacimiento (Feyera et al., 2022).
En el manejo de alimentación tradicional, en el mejor de los casos, durante el periparto la cerda deberá comer la misma cantidad y tipo de pienso que ha estado comiendo en la etapa de gestación.
Los días previos y posteriores al parto, la cerda necesitará un mayor aporte de fibra para controlar y evitar la aparición del síndrome de agalaxia, también conocido como síndrome MMA (Mastitis, Metritis, Agalactia) caracterizado por la inflamación de la ubre (mastitis), la inflamación del útero (metritis) y la disminución o ausencia de producción de leche (agalactia), mejorando asimismo la calidad y cantidad del calostro (Decaluwé et al., 2014).

La incorporación de este pienso especifico (Tabla 1) (Dumniem et al., 2024) permite:
Disminuir la duración del parto.
Con la alimentación electrónica individualizada existe la posibilidad de suministrar un pienso periparto con un mayor contenido en fibra desde la entrada a lactación hasta dos o 3 días posteriores al parto, mediante la disposición de dos líneas de pienso diferente.
Disminuir el porcentaje de lechones nacidos muertos.
Mejorar la ingesta de calostro.
Reducir la mortalidad durante los 3, 7 y 21 días postparto.
Tabla 1. Producción, mortalidad de lechones e ingesta de calostro en grupo control (con dieta convencional) y grupo tratamiento (con dieta suplementada con fibra 7 días previos al parto) analizada mediante least-square means ± SEM (Dumniem et al., 2024).

Formulado para cubrir las altas exigencias nutricionales de la cerda durante la fase de lactación, este pienso está diseñado para:
Maximizar la producción de leche, asegurando una óptima nutrición de los lechones.
Evitar la pérdida excesiva de peso corporal de la cerda.
Mejorar la recuperación postparto, favoreciendo su posterior ciclo reproductivo.
Los piensos de lactación contienen un alto contenido energético y proteico, vitaminas del grupo B, minerales esenciales (como calcio y fósforo) y aditivos que favorecen la digestibilidad y el aprovechamiento de los nutrientes (Quesnel et al., 2009).
La alimentación electrónica en maternidad permite habilitar tantas ventanas de alimentación como se deseen a lo largo del día.
En nuestro caso, para que la cerda disponga de las fracciones de su ración diaria repartidas en varias tomas disponibles, ofrecemos 5 raciones (Gráfica 6) que van de más a menos cantidad:
1. 2. 3. 4. 5.
A las 7:30: 25 % de la ración diaria
A las 11:30: 20 % de la ración diaria
A las 15:30: 20 % de la ración diaria
A las 18:30: 20 % de la ración diaria
A las 21:30: 15 % de la ración diaria
Posibilidad de un 20% adicional en total repartido en las 5 tomas anteriores.
Este sistema permite aumentar o disminuir in situ desde dispositivos portátiles, de forma manual e individualizada, la ración de la cerda durante la revisión diaria, corrigiendo así casos puntuales como cerdas que van por encima de la media en cuanto a capacidad de ingesta, condición corporal o estado sanitario (Aparicio et al., 2025a).
Gráfica 6. Ejemplo de distribución horaria de ingesta de pienso en lactación y porcentaje de pienso (%) suministrado durante 1 hora y media en cada una de las 5 comidas asignadas a un animal (Fuente propia).
Habitualmente, los dosificadores cuentan con indicadores luminosos o pantallas con un código de colores (Imagen 1) que aportan, de forma ágil y visual, información sobre el nivel de consumo alcanzado por el animal respecto al objetivo diario fijado en su curva.
Estos avisos luminosos indican, mediante diferentes grados de alarma, si la cerda ha comido por debajo del nivel crítico, una cantidad baja, la cantidad establecida o ha sobrepasado esta cantidad prefijada. Así se puede evaluar qué le está pasando a la cerda y actuar en consecuencia.
En el caso de una cerda con una alta ingesta, que consuma una cantidad superior a la establecida dado el extra que ofrecen estos dosificadores, se procederá a un aumento individualizado de su ración, corrigiéndose así la subestimación de su curva de lactación a partir de ese día y en adelante.
En el caso de la cerda enferma o sobrepasada, se hará una reducción individual de la ración diaria con el fin de minimizar el desperdicio de pienso. Después se podrán recuperar de forma progresiva las cantidades hasta volver al punto de la curva que le corresponda o a la cantidad que la cerda admita.
Se puede actuar sobre cerdas individuales o de forma colectiva si se repite el mismo patrón de consumo en el grupo de cerdas lactantes

Imagen 1. Diferentes sistemas de alimentación automática (Fuentes: Piñeiro et. al; Premio Grupo Fuertes y propia).


Una ventaja adicional de estos tipos de sistemas es que permiten proporcionar un extra de pienso, que suele ser de hasta un 20 % por encima del máximo asignado a una cerda de forma automática, sin necesidad de nuestra intervención, si lo reclama (Gráfica 7).
Esto da un margen de seguridad muy valioso para cubrir con mayor precisión los requerimientos nutricionales de la cerda y asegurar que alcanza su máximo potencial de producción láctea.
En el momento en el que gran parte de las cerdas estén recurriendo a este aporte adicional de pienso, el sistema indicará que la mayoría tiene una curva de ingesta superior a la curva de pienso asignado y, por tanto, será necesario recalcular al alza la asignación (Gráfica 7).
En el caso contrario, cuando la mayor parte de las cerdas no lleguen con su ingesta a la línea de la curva de pienso asignado, se deberá recalcular a la baja la asignación.
Cantidad de pienso ofrecido y consumido (g/día)
La alimentación de precisión permite maximizar la producción láctea de la cerda, ya que su curva de ingesta, a voluntad y autorregulada, se adapta de forma paralela a la curva de producción láctea, lo que contribuye a la obtención de camadas de mayor peso al destete (Piñeiro et al., 2024).
En un estudio comparativo de campo realizado en nuestras explotaciones, comprobamos un incremento de hasta 400 gramos más de peso al destete, en comparación con los sistemas tradicionales no electrónicos, a los 28 días de lactación.
Días desde el parto
Consumo Objetivo
Gráfica 7. Ejemplo de alimentación de una cerda que se encuentra por encima de la curva, pudiendo comer un 20 % más de la ración en cada comida (Fuente propia).
Tabla 2. Estudio comparativo entre alimentación electrónica (n=668) y control (n=672) en una granja comercial de cerdas reproductoras (Fuente propia).
Durante el estudio, también se observó un menor número de bajas en lactación por aplastamientos, debido a una reducción del nerviosismo y, por tanto, de los picos de estrés en los momentos previos a la administración del pienso que se producen con la alimentación tradicional, reduciéndose asimismo el estrés del lechón en los momentos de amamantamiento (Aparicio et al., 2024a).
Otra de las apreciaciones fue que, dada la mayor producción láctea observada, se redujo la necesidad de realizar movimientos de lechones, recogidas y donaciones, con las consecuentes ventajas sanitarias que ello conlleva (Aparicio et al., 2024a).
Se necesitó un menor número de cerdas nodrizas, pasando del 12 % al 8 %.
Se observó un aumento en la capacidad de crianza (número de lechones que la cerda es capaz de amamantar hasta el final de la lactación), pasando de 13,1 a 13,2 lechones destetados por camada.

Los sistemas de alimentación automatizada no se consideran ad libitum, pero permiten la autorregulación del propio animal, ya que se le dota de libertad de elección para comer toda o parte de su ración asignada en el momento en el que ella lo desea a lo largo de las 24 horas del día.

Un racionamiento más controlado, quizás se debería ejercer en los periodos más críticos, como el periparto, en los que una alimentación excesiva podría acarrearnos problemas de edemas mamarios y, por tanto, agalaxia.
Sin embargo, en la mayoría de los casos, cuando monitorizamos la curva de ingesta, esta se adapta en gran medida a la curva de alimento disponible, lo que demuestra que la autorregulación por parte de la cerda existe desde el primer momento (Aparicio et al., 2025c).
Durante la lactación, hemos observado que, en la mayoría de los casos, durante la lactación la cerda opta por comer la mayor parte de su ración diaria durante la noche, buscando la tranquilidad, minimizando su estrés y el de su camada, por lo que será interesante introducir la primera ventana de alimentación durante la noche, incrementando la oferta hasta un total de 6 tomas diarias (Gráfica 6).
Imagen 2. La dosificación automatizada en cerdas lactantes permite adaptar la alimentación al ritmo natural de ingesta, favoreciendo la producción láctea y reduciendo problemas como el edema mamario o la disgalaxia/ agalaxia.

Limitar la ración desde la mitad hasta el final de la lactación daría lugar a la llegada a un umbral de máxima producción láctea demasiado bajo, comprometiendo la disponibilidad de leche para la camada de lechones y, por tanto, su ganancia diaria de peso y el peso al destete alcanzado a la edad prevista.
Para mejorar el rendimiento reproductivo de la cerda, es clave establecer un estricto control sobre el intervalo destete-celo (IDC), ya que ello:
Permite mantener elevado el índice de partos por cerda y año.
Posibilita la inseminación del número semanal necesario de cerdas destetadas.
(Leman, 1992)
El IDC está directamente influenciado por la presencia de anestros postdestete, considerados uno de los problemas reproductivos con mayor impacto económico en las granjas porcinas, al provocar un incremento de los días no productivos (DNP) (Rodríguez-Estévez, 2010).
En cambio, con la dosificación automatizada, la curva de alimentación durante toda la lactación, tendrá un amplio límite superior, que dará la posibilidad de la expresión máxima del potencial de producción láctea (Aparicio et al., 2025c).
Como resultado, se obtienen lactaciones más exitosas y una reducción considerable del número de cerdas con edema mamario, disgalaxia o agalaxia que, a menudo derivan en fallos de lactación completos, con la consecuente necesidad de destetes precoces y periodos de reposo para la cerda (Aparicio et al., 2025c).


Imagen 3. Control individualizado de la ingesta mediante dosificadores automáticos en maternidad. Una alimentación precisa y ajustada tras el parto contribuye a reducir el intervalo destete-celo y mejorar los índices reproductivos.
Cuanto menor sea la duración del intervalo entre partos, mayor será el número de partos por cerda y año.
Un solo día no productivo implica una reducción de 0,007 camadas por cerda y año (Rodríguez-Estévez, 2010)

El aumento del IDC no solo conlleva una mayor acumulación de días no productivos, sino también una asincronía en el momento de la ovulación que, en ausencia de un protocolo de inducción y sincronización del celo y la ovulación, puede traducirse en un descenso de la fertilidad de la cerda (Cassar et al., 2008).
Tras el parto de la cerda, debe producirse la regeneración endometrial y la involución uterina en un plazo aproximado de tres semanas para que pueda iniciarse un nuevo ciclo estral con una elevada capacidad fértil (Björkman et al., 2018).
En algunos animales, esta involución puede retrasarse debido a infecciones uterinas, a menudo acompañadas de infecciones mamarias, lo que provoca una disminución de la fertilidad y la prolificidad en el siguiente ciclo, con graves consecuencias para los rendimientos productivos y económicos.
Estas infecciones son más frecuentes en animales longevos, con un alto número de partos (Glock y Bilkei, 2005).
En un estudio realizado con 296 cerdas (149 alimentadas mediante sistema electrónico y 147 con alimentación control) en su segundo ciclo reproductivo, se observó una mejoría en el número de lechones nacidos vivos, así como una reducción de los lechones muertos y momificados, en el grupo de alimentación electrónica.
Además, los días medios de gestación fueron mayores en el grupo con alimentación electrónica, lo que podría justificarse por la mayor prolificidad asociada al incremento del tamaño de camada, en el contexto del uso de líneas hiperprolíficas (Oliviero et al., 2019).
Tabla 3. Comparativa entre nacidos vivos, muertos, momificados y días medios de gestación en cerdas hiperprolificas de segundo ciclo alimentadas con alimentación electrónica y tradicional con dosificador (Fuente propia).
El principal inconveniente de los sistemas de alimentación electrónica de precisión es la necesidad de contar con personal formado y, al menos, mínimamente cualificado en el manejo de sistemas electrónicos, informáticos, etc.
El operario que antes destacaba por su experiencia y buen ojo, ahora también debe adquirir competencias para analizar, controlar e interpretar las señales y parámetros que devuelven estos sistemas con el fin de corregir posibles desviaciones o errores y alcanzar así una alimentación eficiente que maximice la producción de la cerda.

La principal ventaja de estos sistemas es que, una vez superado el reto de la formación del personal, se disponen de herramientas útiles, prácticas, visuales y objetivas que informan en todo momento sobre cómo se está llevando a cabo la alimentación de nuestras cerdas lactantes.
Ofrecen todas las ventajas del dosificador manual (el pienso está visible, se conoce la cantidad que se va a suministrar y se puede verificar), siempre que el sistema esté correctamente calibrado, con el valor añadido de no requerir la intervención del personal fuera del horario laboral.
Es importante señalar que los sistemas automáticos no son completamente autónomos, ya que requieren una supervisión constante para detectar a tiempo cualquier incidencia que pueda producirse. No obstante, una vez identificada, su corrección puede realizarse de forma inmediata desde cualquier dispositivo habilitado para su control.
Referencias disponibles en la versión web del artículo en nutrinews.com

Basado en esporas de Clostridium butyricum:
▶ Produce ácido butírico en el ciego
▶ Previene desórdenes intestinales causados por bacterias patógenas
▶ Incrementa la biodiversidad de la flora butírica impidiendo la colonización de patógenos
▶ Estable durante el granulado y la digestión
Lorena Pérez Esteruelas
Servicio técnico de porcino en Elanco. PM-ES-25-0389.

El lechón pasa por diferentes momentos en su etapa productiva, todos ellos marcados por algún grado de estrés. Uno de los más importantes y que se podría considerar como “el mismo cuento de todos los años”, es el momento del destete.
Cuando el lechón se traslada a la siguiente fase productiva, conocida como transición o fase 2, aparece un estrés que viene desencadenado por varios factores:
Cambios sociales
Cambios ambientales
Cambios nutricionales
Desafíos sanitarios
Todos estos factores se suelen traducir en estados de ayuno de duración variable que afectan al aparato gastrointestinal.
Una de las preguntas que debemos plantear en este momento es si en el destete, el aparato gastrointestinal ya se considera maduro.
La respuesta es no. De hecho, un lechón de este tipo puede tardar hasta 12 semanas en desarrollar completamente su tracto gastrointestinal (Quiniou, 1995; Koong, 1993; Bikker, 1996).

Todo este desarrollo que ocurre en el periodo de lactación (cuando el lechón se encuentra con la cerda), se va a ver muy afectado en el momento del destete.
Desde antes del nacimiento hasta las primeras semanas de vida del lechón, el intestino pasa por un desarrollo muy rápido que requiere de altas cantidades de energía.
Por ejemplo, durante los primeros 10 días después del nacimiento ocurren aumentos significativos en su peso absoluto y en el peso de la mucosa intestinal (Cranwell, 1995).
Después del día 10 de vida se observa que el intestino delgado pasa por un proceso de maduración, que se caracteriza por aumentos marcados del volumen, longitud y peso. Puede llegar a equipararse con un piso de hasta 114 m².
Los cambios macroscópicos del intestino delgado están asociados con las transformaciones que sufren las vellosidades de la mucosa intestinal.
Es decir, en un periodo muy corto de tiempo el intestino del animal se desarrolla de forma exponencial y también, en otro momento de muy corta duración y con una sola acción, que es el destete, todo este proceso de maduración se pierde.
De hecho, se ha demostrado que hasta el 75% de las vellosidades intestinales se pueden atrofiar durante las primeras 24 horas posdestete (Jensen, 1997; Makkink, 1994; Effird, 1982).
Figura 1. Desarrollo del intestino delgado del lechón en los primeros días de vida.
Volumen, longitud y peso
Nacimiento
PRENATAL 3d 10d 114m2
No enterocitos se duplica
Durante las 3 semanas prenatales el volumen del ID se incrementa en un 70% con respecto al cuerpo del animal
14 días
altura de vellosidad
Profundidad y engrosamiento C. de Lieberkhün población celular, longitud y diámetro de ID
NO COMO O COMO POCO
NO CUBRO MIS
REQUERIMIENTOS DE ENERGÍA Y PROTEÍNA

DISMINUYE LA ALTURA DE LAS VELLOSIDADES INTESTINALES (hasta un 75%) en las primeras 24 horas de P.D
Figura 2. Consecuencias en el desarrollo del intestino tras el destete.
Teniendo en cuenta que el peso al destete es un predictor de la productividad del animal (López-Vergé et al., 2018), es necesario estimular la ingesta de pienso lo antes posible para que en el momento del destete ya sepa comer e intentar que gane algo más de peso cuando se le aparte de la cerda.
Un lechón que gana 250 gramos al día tomando leche en maternidad va a necesitar hasta 300 gramos por día de pienso sólido en destete para mantener el mismo nivel de crecimiento (Le Dividich, 2000).
A todo esto, hay que añadir que no todos los lechones comen por igual, hay buenos y malos comedores.
Se ha observado que lechones destetados que consumen poco pienso, es decir, los “malos comedores”, entre el día 1 y 8 posdestete tienden a comer menos (169 g/d) cuando se compara con los “buenos comedores” (245 g/d) (p<0,001) en el mismo periodo.
Esto acaba teniendo consecuencias negativas porque durante el resto de la transición siguen comiendo menos (p<0,001) (Fabà, 2023).
El peso a destete es clave para conseguir crecimientos mayores en la fase de posdestete, ya que existe una fuerte correlación entre éste y peso de salida a cebo.
Un incremento adicional en el peso al destete de tan solo 270 gramos puede traducirse en una mejora sustancial de 2,28 kg a salida a cebo (Muro, 2023). Por ello, es importante que el animal comience a tomar dieta sólida ya en lactación.


Del 60% al 80% del consumo total de pienso en esta fase se da en la última semana de lactación (Bruininx, 2002; Sulabo, 2010; Fraser, 1994). Por lo que, estimular el consumo en este periodo puede ayudar a conseguir mayor peso a destete.
Según Pluske y colaboradores (2005), el efecto del peso al destete y de la ingesta de alimento durante la primera semana posdestete son aditivos y explican aproximadamente el 80% de la variación en el peso corporal al día 20 después del destete. Por tanto, estimular el consumo de pienso en estos primeros días de posdestete también va a contribuir a alcanzar un buen peso a lo largo de la Fase 2.
PIENSO ALREDEDOR DEL DESTETE?
Tonisity Px™ es un pienso complementario en forma de polvo soluble en agua. Se administra a lechones a partir de los 2 días de vida. Contiene proteínas, aminoácidos, electrolitos, saborizantes y acidificantes.
Alta palatabilidad
Es fuente de energía rápida para los enterocitos
Textura/viscosidad similar a la leche
Gráfico 1. Ingesta de pienso en lactación en función de la edad del animal.
Con la administración de Tonisity Px™ alrededor del destete lo que se busca es:
Estimular el consumo de pienso:
En la primera toma de contacto entre lechón y pienso que suele ser a partir de la segunda semana de vida (día 8-9-10 de vida aproximadamente), ayuda el administrar Tonisity Px™ los primeros 2 a 3 días encima del pienso.
El producto se puede administrar de 3 a 4 días antes del destete directamente encima del pienso que se encuentra en los platos.
La dosis es de 40 ml por lechón (500 ml por camada si hay ≤14 lechones o 600 ml por camada si hay >14 lechones).
Ayudar a preparar el intestino para afrontar el proceso de destete. Gracias a su composición aporta energía extra directamente a los enterocitos para ayudar a la maduración del intestino.
Evitar el periodo de ayuno justo después del destete: el protocolo continúa los primeros 3 a 4 días de posdestete para continuar con el estímulo de consumo de pienso y aporte de energía extra. Para ello se administra Tonisity Px™ a razón de 50-100ml por lechón al día, directamente encima del pienso.

ml/lechón y día*
y día*
ml/lechón y día**
*500ml/camada y día si ≤14 lechones por camada 600ml/camada y día si > 14 lechones por camada **ratio 1:1 (Tonisity px:pienso)
Figura 3. Protocolo administración Tonisity Px™ alrededor del destete.
Este producto cuenta con un amplio bagaje de estudios que demuestran la eficacia en este ámbito.
En un estudio llevado a cabo en España (Study 23-016-P-P) en una granja de 400 cerdas ibéricas se siguió el siguiente protocolo con dos grupos:
Grupo control (Control) que solo recibió pienso en platos
Grupo Tonisity Px™ (Px) que recibió pienso en platos con Tonisity Px™ al 3%, a razón de 400 ml/camada.
Se observó mayor consumo de pienso en el periodo de lactación en el grupo Tonisity Px™ (24% más) en comparación con el grupo control.
Gráfico 2. Consumo de pienso en Kg pienso/cerdo y grupo en los días 8, 9 y 10 de vida. Study 23-016-P-P.
Respecto al crecimiento alrededor del destete el grupo Px se destetó con una diferencia a favor de +210 gramos/lechón destetado (7,34 kg Grupo Control vs 7,55 kg Grupo Px).
Esta diferencia de peso al destete se tradujo posteriormente en una diferencia a favor del grupo Px de +1420 gramos/cerdo salido a cebo (12,29 kg Grupo Control vs 13,71 kg Grupo Px).
Gráfico 3. Peso a destete y peso de salida a cebo para grupo
Control y grupo Px. Study 23-016-P-P.
En 2024, en un isowean localizado en Aragón (Study 24-010-P-P) se recibieron 200 lechones destetados de 26 días de edad (Danbred con finalizador Pietrain).
Se crearon 2 grupos, un grupo control (C) y un grupo tratamiento (Px).
Los animales fueron divididos en dos corrales por grupo, albergándose 50 animales por corral.
En el grupo C se administró un total de 5,4 kg de pienso en formato sólido repartido homogéneamente en platos. En el grupo Px se administraron 5,4 kg del mismo pienso junto con Tonisity Px™ al 3%, a razón de 100 ml por animal.
Cada 30 minutos y hasta un total de 2 horas se fue pesando el contenido de los platos para calcular el consumo de alimento en cada grupo.

Se observó que en los corrales del grupo Px el consumo fue de 51,3% y 46,2%, respectivamente, durante los primeros 30 minutos y fue en aumento. En el mismo periodo, el grupo C consumió 0,0% y 16,3%, respectivamente. Al cabo de las 2 horas del primer contacto con el pienso, el grupo C consumió un total de 17% de pienso, mientras que en el grupo Px el consumo total fue del 71%.
El estudio evidenció que el uso de Tonisity Px™ al 3% estimuló el consumo de pienso hasta 4,2 veces más con respecto al grupo C.

En el último metaanálisis del uso de Tonisity Px™ donde se engloban más de 32.000 animales y con el objetivo de obtener un mayor peso de destete se observa que de media se consiguen 300 gramos de ventaja en comparación con los grupos control.
Gráfico 5. Metaanálisis de diferentes estudios en el análisis de peso a destete entre grupo Px y grupo Control.
-41%, P<0,0001, 21 estudios, 26.243 lechones
Es lógico pensar que al aumentar el peso de destete de los lechones se va a conseguir una reducción del porcentaje de animales de bajo peso a destete (los conocidos como saldos). En el metaanálisis la reducción de este tipo de animales con respecto a grupos controles es del 41%.
Gráfico 6. Metaanálisis de diferentes estudios en el análisis de porcentaje de saldos a destete entre grupo Px y grupo Control.

Es importante conseguir pesos a destete altos, ya que estos están fuertemente correlacionados con el peso de salida a cebo.
Con el uso de Tonisity Px™ de acuerdo a los protocolos descritos se consigue este objetivo y, además, se reduce el porcentaje de saldos a destete que son aquellos animales que más desafío sanitario experimentan y mayor manejo necesitan en el periodo de transición.
Referencias disponibles en la versión web del artículo en nutrinews.com
Destetando al lechón, un mal de siempre, ¿cómo lo facilitamos?
DESCÁRGALO EN PDF
Elanco y la barra diagonal son marcas registradas de Elanco o sus filiales © 2025 Elanco
Para más información del valor de Tonisity Px en el nuevo entorno productivo contacta con el equipo de Elanco Spain S.L.U. Avda. de Bruselas, 13, 2ª planta 28108 Alcobendas – Madrid, España T: (+34) 91 903 3840 W: my.elanco.com/es/productos/tonisitypx

¿Quién dijo que la nutrición animal era aburrida?




El frijol de Soja (Glycine max) es una oleaginosa que se ha venido sembrando en forma intensiva, por su alto valor proteico y contenido de aceite.
Una de las formas de uso del frijol de soja en alimentación animal es en su forma integral, sin extraerle el aceite y previa desactivación con calor de los factores antinutricionales presentes en el grano.
Como todo no es perfecto, el frijol de Soja contiene una serie de factores antinutricionales con efectos negativos muy importantes sobre la digestión y absorción de nutrientes en el tracto gastrointestinal, principalmente en los monogástricos.

Manuel Vázquez REVEEX NUTRICION C.A.


Luego de la extracción del aceite estos factores permanecen en gran medida en la fracción harina de soja, sin embargo y afortunadamente la mayor parte de estos factores antinutricionales se pueden desactivar tratando la harina con calor.
Del procesamiento industrial del frijol de soja se obtienen principalmente 2 fracciones de interés para alimentación animal, aceite crudo de soja y harina de soja.
ACEITE
DESGOMADO
El aceite crudo de soja se usa en forma directa en alimentación animal. Del proceso de refinación del aceite crudo de soja se obtiene el aceite refinado de soja y la lecitina de soja, esta última se usa tanto en alimentación humana como en alimentación de diferentes especies animales incluyendo acuicultura.
La harina de soja generalmente y dependiendo del grado de eliminación de la cáscara, presenta un valor de proteína que oscila entre 44 y 48%.
La harina tratada térmicamente, se usa en alimentación animal y de la misma se pueden obtener varias fracciones proteicas aplicando tratamientos industriales adicionales para eliminar en forma más precisa los factores antinutricionales que no se desactivan o eliminan mediante el proceso térmico. Ejemplos: Harina de soja fermentada, proteína concentrada de soja, proteína aislada de soja, entre otros.
GRANOS DE SOJA
DESCASCARILLADO
ACONDICIONAMIENTO
EXTRACCIÓN POR SOLVENTES (CON HEXANO)
REFINAMIENTO ALCALINO GLICERINA
DECOLORACIÓN
DESODORIZACIÓN
ACEITE REFINADO DE SOYA
Figura 1. Diagrama general del procesamiento industrial del frijol de soja y principales subproductos que se generan.
DESGRASADA
Principalesfactoresantinutricionales presentesenelfrijoldesojacrudo quenormalmentepasanalaharina
FACTORES TERMOLÁBILES (Pueden ser desactivados con tratamiento térmico):
Inhibidores de tripsina (Kunitz y Bowman-Birk): Reducen la actividad de la enzima tripsina que es crucial para la digestión de las proteínas. Al afectarse la digestibilidad de las proteínas y tener más proteína sin digerir en el intestino, se afecta el balance de la flora intestinal con las consecuencias negativas sobre la salud intestinal que ello conlleva. Con niveles altos de estos factores se puede presentar también hipertrofia pancreática, disminución del crecimiento, etc.
Lectinas (hemaglutininas): Producen daño del epitelio intestinal disminuyendo la absorción de nutrientes, esto puede también generar gas, hinchazón, diarrea y náuseas. Todo lo anterior puede alterar el equilibrio de la flora intestinal.
Saponinas: Las saponinas producen el sabor amargo-astringente-metálico (Característico de habas crudas) lo que pueden afectar el sabor y la palatabilidad generando bajas en el consumo del alimento.
FACTORES NO TERMOLÁBILES (No se desactivan con calor, pero puede disminuirse su actividad mediante el uso de enzimas y otros procesos químicos):
Oligosacáridos (rafinosa y estaquiosa):
Causan flatulencia y reducen la digestibilidad. Disbiosis en lechones (Zhan y Cols 2003). Disbiosis en pollos, afectación de la absorción de nutrientes, pérdida de consistencia de la gallinaza (Zhu y Col, 2020). Se puede usar la enzima alfa-galactosidasa para ayudar a digerir estos compuestos.
Fitatos (ácido fítico): Acción quelante sobre minerales (P, Ca, Zn) y también sobre otros compuestos tales como aminoácidos, etc., disminuyendo su biodisponibilidad. Puede disminuir su efecto negativo con el uso de la enzima fitasa, la cual hoy es de uso común en la mayoría de alimentos para aves y cerdos.
La calidad de la harina de soja depende de diferentes factores tales como la variedad genética, la humedad del grano, contaminantes, zona geográfica donde se produce, método y tecnología usada para el procesamiento.
Generalmente en la mayoría de las referencias se establece que la harina de soja proveniente de USA es de mejor calidad que la procedente de Brasil y Argentina, para lo cual indican que depende de varios factores, pero en buena parte es consecuencia de diferencias importantes en la tecnología existente en las plantas de procesamiento.
BRASIL USA
NOTA: Los cuadros ocre claro representan el rango preferido para cada indicador; en el caso de Rlys/Lys no se especifica el rango, pero se busca que el valor sea el más alto posible. Fuente: USSEC (U.S. Soybean Export Council) 2021
Gráfico 1. Comparación entre harina de soja de Brasil Vs USA, Indicadores del procesamiento de la harina de soja 2021 (Promedios y desviación estándar).
Algunosmétodosdisponiblespara medirlacalidaddelprocesamiento térmicodelaharinadesoja
Métodos organolépticos
Sabor: En el frijol crudo o poco procesado, así como en la harina de soja poco procesada, es factible percibir el sabor característico a “habas crudas”.
Color: Tanto en frijol como en harina los colores oscuros generalmente denotan sobreprocesamiento con calor. Esto indica desnaturalización de las proteínas mediante la reacción de Maillard y afectación negativa de la biodisponibilidad del aminoácido Lisina, seguido de Cistina, Metionina, Treonina y Arginina.
Colores muy claros generalmente pueden indicar falta de suficiente procesamiento con calor, lo cual hace sospechar sobre la presencia de cantidades importantes de los factores antinutricionales termolábiles.
Métodos químicos de uso más




LISINA REACTIVA (RELACIÓN DE LISINA
REACTIVA A LISINA TOTAL (RLYS/LYS)):
Esta relación mide la lisina reactiva, disponible para la digestión en animales monogástricos, respecto al contenido total de lisina. La lisina reactiva es la cantidad de lisina total biodisponible para el animal. Una relación más alta significa menos daño por calor a la harina de soja y viceversa.
Este método requiere bastante equipamiento y recursos en laboratorio, es costoso,lento, y no mide los factores antinutricionales. Está relación debería ser lo más alta posible, nunca inferior a 85%.

Pienso complementario, fuente de emulsionantes de origen vegetal y polifenoles.

Ahorros visibles en el resultado.
Mejora los resultados zootécnicos.




Tel.: +34 91 725 08 00 www.liptosa.com liptosa@liptosa.com
...the green way of life

TIA (ACTIVIDAD DEL INHIBIDOR DE TRIPSINA):
Los inhibidores de tripsina son factores antinutricionales que se asocian negativamente con la digestión de proteínas. La actividad de tripsina en la planta de soja es un mecanismo de protección y aumenta en condiciones de estrés.
La harina de soja correctamente procesada debe tener un TIA inferior a 2,5 mg/g para primeras edades. Método costoso y lento, pero es preciso y mide en forma directa los factores antitripsina.
SOLUBILIDAD EN KOH 0,2%:
La solubilidad en KOH diferencia la harina de soja sobrecalentada de la harina correctamente procesada y de la subprocesada, si bien para la subprocesada KOH>85% se recomienda verificar con Ureasa. Es uno de los análisis más populares, es relativamente rápido y de bajo costo. Se trata de un método indirecto, ya que no mide directamente los inhibidores.
La harina de soja correctamente procesada debe tener un valor de KOH entre 75 y 85 %. En algunos países consideran rangos diferentes tales como 70-85%, 73-85%, 75-83% etc... Se ha demostrado que valores inferiores al 75% reducen el rendimiento animal debido a la pérdida de calidad proteica y de afectación de aminoácidos como la lisina por exceso de calor. Cuando los valores son superiores a 85% hay riesgo de tener cantidades importantes de factores anti nutricionales termolábiles con las consecuencias negativas que ello conlleva.
La prueba de actividad ureásica es una medida indirecta que indica, mediante la variación del pH, si ha existido un subprocesamiento térmico de la soja. También puede indicar si está sobreprocesada aunque en ese rango no es factible saber la intensidad del sobreprocesamiento porque una vez que la escala llega a cero no es factible diferenciar diferentes grados.
Cuanto mayor sea el valor en el análisis, mayor será la actividad de la ureasa presente en la muestra, y se infiere que habrá más factores antinutricionales termolábiles presentes.
La soja cruda suele presentar una variación de pH comprendida entre 2,0 y 2,5 (Butolo, 2002). Luego del tratamiento térmico si se hace en forma adecuada, la harina de soja debe presentar una variación de pH comprendida entre 0,05 y 0,20 según el NRC (USA), mientras que FEDNA (España) establece un rango más amplio en el límite superior 0,05-0,25.


ROJO FENOL:
Es una variante de la actividad ureásica. En este caso se estima la variación de pH mediante el indicador rojo fenol el cual genera una coloración roja al producirse un cambio en el pH debido a la reacción de la ureasa con la urea.
Cuanto menos procesamiento tenga la harina, el color rojo será más intenso, si no presenta nada de color rojo indica sobreprocesamiento. Es un método indirecto cualitativo, es muy rápido y de bajo costo.
ÍNDICE DE DISPERSABILIDAD
PROTEICA (PDI):
El PDI es un método que no mide los factores antinutricionales termolábiles, pero da una idea del grado de procesamiento térmico. La harina de soja con un PDI del 15-30% se considera bien procesada según el U.S. Soybean Export Council (USSEC).
Es muy rápido y barato, pero tiene el problema de que no está bien estandarizado (las variaciones en el contenido de fibra y humedad afectan los resultados).
NIR (INFRARROJO CERCANO):
El costo inicial del equipo es considerable. Las curvas de calibración son costosas al inicio, luego de esto el análisis es económico y rápido. No es destructivo ni usa compuestos contaminantes.
En cuanto a la curva de calibración debe estar calibrada para la bromatología e inhibidores de tripsina y/o KOH y/o actividad de ureasa, en ese orden.

¿Quémétododeanálisisusar?
1 2 3
NIR sería la elección, siempre y cuando se cuente con recursos suficientes para la inversión inicial del equipo y siempre que se pueda contar con una curva robusta y bien ajustada que incluya al menos uno de los siguientes parámetros y en ese orden (TIA, KOH, Ureasa).
KOH sería la elección, si no hay NIR y se cuenta con laboratorio con capacidad para determinar proteína, complementar en algunos casos con ureasa sobre todo en muestras con KOH>85%. Pudiera disponerse de TIA para verificar algunas muestras, pero es costoso y largo.
Ureasa y/o rojo fenol en caso de tener un laboratorio pequeño, pero sin capacidad de determinar proteína.
Análisisbromatológicosdela harinadesojarecibidadurante 2024hastajulio2025
A saber, todas las muestras analizadas durante el periodo 2024-2025, pertenecen a harina de soja procedente de USA.
2024 (n=218)
En el Cuadro 1 podemos observar que durante el año 2024 el comportamiento promedio de la proteína es de 45,07% y la desviación estándar de 0,49. En el caso del KOH para el mismo año obtuvimos un promedio de 81,90% con desviación estándar de 1,99.
Durante los 7 primeros meses del año 2025 obtuvimos un promedio de 44,92% para la proteína con desviación estándar de 0,59 y el promedio para el KOH en ese mismo periodo fue de 85,46% con desviación de 2,36.
La proteína descendió 0,14 puntos porcentuales respecto al año 2024 y la desviación aumentó 0,10.
Respecto al 2024 el KOH promedio se incrementó 3,56 puntos porcentuales y la desviación aumentó en 0,37.
Podemos observar que la fibra tiende a ser alta durante el 2024 incrementándose algo en 2025, esta fibra alta guarda correspondencia con el hecho de que la harina de soja es de proteína baja que suele asociarse a conservar niveles altos de cáscara. Los niveles altos de cáscara contribuyen también a acelerar el tránsito intestinal.
2025 (n=83) Análisis
Cuadro 1. Resumen de los valores bromatológicos promedio y desviación estándar años 2024 y 2025 hasta julio. Nota: Muestras analizadas por análisis químicos. Se usó método Kjeldahl para proteína. Para KOH se usó el método según Araba y Dale 1990, adoptado actualmente por la AOCS.

PROPIEDADES BRONCODILATADORAS
MEJORA LA INMUNIDAD LOCAL
ACTIVIDAD MUCOLÍTICA Y EXPECTORANTE
PROTEGE DEL ESTRÉS OXIDATIVO
REDUCCIÓN DE LA RESPUESTA INFLAMATORIA
En los Gráficos 2, 3 y 4 en general se observa que durante los 7 meses transcurridos de este año 2025 ha habido muestras que sobrepasaron en forma importante el valor de referencia máximo de 85% para el KOH.

Gráfico 2. Solubilidad KOH de la harina de soja año 2024.
Gráfico 3. Solubilidad KOH de la harina de soja, primeros 7 meses año 2025.
Observacionesacampo
Durante los siete primeros meses del año 2025, se ha observado que concomitante al aumento en los valores de KOH (Imágenes 1, 2 y 3; menor tratamiento térmico) ha habido correspondencia con observaciones de campo, donde los técnicos han reportado mayor incidencia de “tránsito rápido” y presencia de alimento sin digerir en las excretas de los pollos.

Imagen 1. Excreta de pollos con abundante alimento sin digerir.
Gráfico 4. Comparativo de la solubilidad KOH de la harina de soja años 2024 y 2025.

En algunos casos se ha visto alimento poco digerido acompañado con heces bastante líquidas (disbiosis), en otros casos se ha observado alimento poco digerido, pero sin la presencia de heces líquidas. En algunos casos también hubo presencia de erosión de mollejas de diferentes grados.
Todo esto ha ido acompañado con diferentes grados de pérdida de desempeño de las aves, y en algunos casos se observó deterioro importante de la cama, sobre todo como consecuencia de la presencia de tránsito rápido en algunas granjas (ver Imágenes 1,2 y 3 ).
El mayor impacto de esto se ha visto principalmente en las primeras edades.


Para disminuir el impacto negativo observado en campo, se adoptaron varias medidas tales como: Disminuir entre 10 a 15% la inclusión de harina de soja e incluir y/o incrementar otras fuentes proteicas de buena calidad tales como, harina de pescado, Harina de vísceras, hemoglobina, proteína concentrada de soja, gluten de maíz 60% etc. También se incrementó la inclusión de ácidos orgánicos.
Comprar harina de soja preferiblemente con proteína superior a 46% y solubilidad en KOH 0,2% entre 75 y 83%. Inhibidor de tripsina menor a 3 mg/g.
Ante situaciones similares a la descrita se sugiere:
Bajar en forma significativa la inclusión en la dieta de harina de soja, incluyendo fuentes proteicas de alta digestibilidad tanto de origen animal como vegetal.
Uso de un grupo enzimático termoestable (fitasas, proteasa, xilanasas, beta-glucanasa, alfa-galactosidasa)
Uso de acidificantes orgánicos para disminuir la incidencia de problemas de salud intestinal tales como disbiosis, pérdida de epitelio, etc… y mejorar la absorción de nutrientes.
Uso de probióticos (esporulados) que resistan el pelletizado y expander (Bacillus subtilis).
Análisis bromatológico y solubilidad en KOH 0,2% de muestras de harina de soja obtenidas en Venezuela con procedencia de EE.UU
DESCÁRGALO EN PDF
Yauheni Shastak, Wolf Pelletier, Adriana Barri
BASF SE, División Nutrición y Salud, 67063
Ludwigshafen, Alemania
En la dinámica de la industria avícola actual, donde el rendimiento y la eficiencia son fundamentales, el papel de los micronutrientes —especialmente la vitamina A— es más crucial que nunca. A medida que la genética aviar y los sistemas de producción continúan evolucionando, también debe hacerlo nuestro enfoque nutricional.


Entre todas las vitaminas, la vitamina
A destaca por sus funciones esenciales en la visión, la defensa inmunitaria, la protección antioxidante, la reproducción y el crecimiento.
Es ampliamente reconocida por nutricionistas y veterinarios como la vitamina más importante en la nutrición animal(1, 2, 3). Sin embargo, no todas las fuentes de vitamina A son iguales.
Este artículo explora los conocimientos más recientes sobre la variabilidad en las fuentes de vitamina A, centrándose en dos atributos clave que determinan su eficacia en las dietas avícolas: la estabilidad y la biodisponibilidad.
La vitamina A suele denominarse la “vitamina antiinfecciosa”, y con razón(4, 5, 6, 7). Contribuye a la función inmunitaria al mantener la integridad de los tejidos epiteliales, que constituyen la primera línea de defensa del organismo frente a los patógenos (8)
También desempeña un papel fundamental en la visión, especialmente en condiciones de poca luz, y es esencial para la salud reproductiva, el crecimiento de los tejidos y la defensa antioxidante (9)
En la nutrición avícola, la vitamina

Figura 1. Estructura general de una formulación en perlas que contiene acetato de vitamina A.

A suele suplementarse en forma de acetato de retinilo, comúnmente en forma de gránulos estabilizados (Figura 1). Estas formulaciones en polvo se añaden a las premezclas y se incorporan al alimento.
Sin embargo, los excipientes y las tecnologías de formulación utilizadas por distintos fabricantes pueden influir significativamente en la estabilidad de la vitamina durante el almacenamiento y el procesamiento, así como en su biodisponibilidad una vez que son ingeridas por las aves.
La vitamina A es una de las vitaminas más sensibles en las formulaciones de premezclas y piensos (10). Se degrada al exponerse a la luz, el calor, el oxígeno y la humedad, lo que convierte su estabilidad en una preocupación clave tanto para los fabricantes de piensos como para los nutricionistas.
Estudios recientes han demostrado que la estabilidad de la vitamina A en premezclas y piensos puede variar considerablemente entre productos comerciales.
Por ejemplo, en un ensayo de almacenamiento de 56 días a 35°C y con una humedad relativa del 60-70%, dos productos comerciales conservaron casi la mitad de su actividad de vitamina A, mientras que otros dos perdieron más del 90% (11) (Figura 2).
Estas diferencias probablemente se deben a variaciones en las técnicas de formulación, incluido el tipo de microencapsulación, los antioxidantes y materiales portadores utilizados, así como las propiedades físicas de las microesferas.
Figura 2. Estabilidad de cuatro premezclas de vitaminas y minerales (incl. cloruro de colina) para pollos de engorde que contienen cuatro fuentes comerciales diferentes de vitamina A (productos almacenados hasta por 56 días a 35°C y 60-70% h.r. (11). Los valores se presentan como media ± desviación estándar (n = 3). Las premezclas se prepararon para contener 4,4 Mio UI de vitamina A por kg. a-c En cada día (0, 28 o 56 días), los valores que no comparten un mismo superíndice son significativamente diferentes (p < 0,05).

4
Estabilidad del producto puro / vida útil bajo condiciones controladas.
Para los productores de alimento, esto significa que la elección de una fuente adecuada de vitamina A puede influir significativamente en la cantidad de vitamina A activa que permanece en el alimento al momento de llegar a las aves.

La biodisponibilidad se evalúa normalmente mediante estudios de respuesta a la dosis in-vivo, en donde se alimenta a las aves con dietas que contienen distintos niveles de vitamina A y se miden los niveles de retinol en el hígado o la sangre. El hígado es el estándar de referencia para este tipo de evaluaciones, ya que es el órgano que almacena hasta el 90% de las reservas de vitamina A dentro del organismo (3) .
Estudios recientes han demostrado que no todos los productos de vitamina A tienen la misma biodisponibilidad. Ensayos independientes que evaluaron diferentes fuentes de vitamina A en pollos de engorde revelaron diferencias sustanciales en su biodisponibilidad relativa (RBA), medida a través de la acumulación de retinol en el hígado.
En un estudio, ciertos productos superaron significativamente a otros en cuanto al rendimiento productivo y al estado de la vitamina A (12), mientras que otro estudio, utilizando un modelo de depleción-repleción encontró una variación notable en la eficacia con la que los diferentes productos restablecían los niveles de vitamina A en el hígado (13)
Estos hallazgos sugieren que las técnicas de formulación pueden influir no sólo en la estabilidad de los productos de vitamina A, sino también en su absorción y utilización en el ave (Tablas 1 y 2). En algunos casos, las formulaciones diseñadas para una alta estabilidad pueden resistir la descomposición en el tracto digestivo, limitando la bioaccesibilidad y, en última instancia, reduciendo la biodisponibilidad.
Tabla 1. Biodisponibilidad relativa (RBA, %), error estándar (EE) e intervalo de confianza (IC) del 95% de fuentes de vitamina A en un ensayo de pollos de engorde 1 (13).
de referencia
Tabla 2. Biodisponibilidad relativa (RBA, %), error estándar (EE) e intervalo de confianza (IC) del 95% de fuentes de vitamina A en un ensayo de pollos de engorde 2 (12).
¿Qué significa todo esto para la industria avícola?
1
No todos los productos de vitamina A son iguales
La calidad y la formulación de la vitamina A pueden afectar significativamente tanto su estabilidad como su biodisponibilidad. Productos que parecen similares en papel pueden comportarse de manera muy diferente en la práctica.
3
La estrategia de formulación influye en el coste y el rendimiento
Fuentes de vitamina A con mayor biodisponibilidad pueden permitir tasas de inclusión más bajas, reduciendo los costos del alimento sin comprometer la salud de las aves.
Alternativamente, utilizar una fuente más biodisponible con niveles estándar de inclusión puede ofrecer un margen de seguridad en condiciones estresantes o desafiantes.

2
El almacenamiento y el manejo son importantes
Incluso la mejor formulación de vitamina
A puede degradarse si no se almacena adecuadamente. Las premezclas y los piensos deben conservarse en ambientes frescos, secos y oscuros.
La inclusión de antioxidantes en las premezclas también puede ayudar a proteger la vitamina A del daño oxidativo.
4
La nutrición de precisión es el futuro
A medida que avanzamos hacia una producción avícola más sostenible y eficiente, la precisión en la suplementación de micronutrientes será cada vez más importante.
Comprender el verdadero valor de las fuentes de vitamina A -más allá de las UI por gramo- es esencial para optimizar el rendimiento y la rentabilidad en la producción.
avícola, la vitamina A sigue siendo una piedra angular de la estrategia nutricional. Sin embargo, para aprovechar realmente su potencial, debemos ir más allá de lo que se indica en la etiqueta y comprender la ciencia que hay detrás de su origen.
Los fabricantes de piensos y los productores avícolas deben trabajar en estrecha colaboración con sus proveedores para evaluar la estabilidad y la biodisponibilidad de los productos de vitamina A.
Invertir en fuentes de vitamina A de alta calidad y bien formuladas puede traducirse en beneficios para la salud de las aves, su crecimiento y la eficiencia general de la producción.
A medida que la investigación continúa avanzando, una cosa queda clara: elegir la fuente adecuada de vitamina A puede marcar una gran diferencia.
La ciencia oculta de la vitamina A en las aves de corral: ¿Por qué la fuente del producto, su estabilidad y biodisponibilidad son importantes?
DESCÁRGALO EN PDF
Referencias
1. 1. McDowell, L. R. (2000). “Vitamin A,” in Vitamins in Animal and Human Nutrition. Ed. L. R. McDowell (Iowa State University Press, USA), 5–90. doi: 10.1002/ 9780470376911
2. Silver, R. J. (2024). Liquid A Drops. Technical report for veterinarian use only. VBS Direct Ltd. 1 Mill View Close, Bulkeley, Cheshire, SY14 8DB.
3. Shastak Y and Pelletier W (2024) Vitamin A source variability: a mini review on stability and bioavailability in poultry. Front. Anim. Sci. 5:1484262. doi: 10.3389/fanim.2024.1484262
4. Green H.N., Mellanby E. 1928. Vitamin A as an anti-infective agent. Brit. Med. J. 2: 691. doi: 10.1136/bmj.2.3537.691.
5. Semba R.D. 1999. Vitamin A as “Anti-Infective” Therapy, 1920–19401. Journal of Nutrition 129 (4): 783-791. https://doi.org/10.1093/jn/129.4.783.
6. Yao, H. 2010. The anti-viral effects of retinoids in canine distemper virus infection: the missing link between measles and Vitamin A. Master-Thesis, Faculty of Medicine McGill University, Montreal Quebec, Canada.
7. Raoof, A. and Abood, A. (2022). Histopathological effects of retinol on blastocyst implantation in pregnant rats. International Journal of Health Sciences 6(S5): 11622–11631. https://doi.org/10.53730/ijhs.v6nS5.11999
8. Shastak Y and Pelletier W (2023a) The role of vitamin A in non-ruminant immunology. Front. Anim. Sci. 4:1197802. doi: 10.3389/fanim.2023.1197802
9. Shastak, Y., and Pelletier, W. (2023b). Delving into vitamin A supplementation in poultry nutrition: current knowledge, functional effects, and practical implications. World’s Poultry Science Journal, 80(1), 109–131. https://doi.org/10.1080/00439339.2023.225032
10. Galli GM, Andretta I, Martinez N, Wernick B, Shastak Y, Gordillo A and Gobi J (2024) Stability of vitamin A at critical points in pet-feed manufacturing and during premix storage. Front. Vet. Sci. 11:1309754. doi: 10.3389/fvets.2024.1309754
11. Hirai, R. A., D. De Leon, M. Randig-Biar, A. Silva, E. Sanchez, A. P. McElroy, C. A. Bailey,N. Martinez, A. Sokale, and L. Music. 2023. Evaluation of the Stability of Vitamin A Acetate Concentrates Mixed in a Vitamin-Trace Mineral Premix Over a 56-Day High Temperature and Humidity Storage Stress. In proceedings of the International Poultry Scientific Forum 2023, 112, Atlanta, Georgia, USA.

12. Sacakli, P., Calik, A., Ramay, M.S., Özen, D., Pelletier, W., Gordillo, W., Shastak, Y., 2025. Comparative evaluation of relative bioavailability among various sources of vitamin A in broilers. In the proceedings of the 24th European Symposium on Poultry Nutrition, PS3-188, p. 358, Maastricht, the Netherlands.
13. Pelletier W., C. Scharch, Y. Shastak. 2025. A pilot study on the relative bioavailability of different vitamin A sources in broiler chickens. In the proceedings of the 24th European Symposium on Poultry Nutrition, PS3-189, p. 359, Maastricht, the Netherlands.

A medida que crece la demanda de animales criados de manera más sostenible, también lo hace el rendimiento de nuestros ingredientes eficientes de máxima calidad.
Vitaminas
Carotenoides
Enzimas
Ácidos orgánicos
Aditivos anti-micotoxinas
Glicinatos
Monoglicéridos

animal-nutrition@basf.com
Dr. Rubén Crespo Sancho, Technical Manager, Biochem Zusatzstoffe GmbH, Alemania Dr. Susanne Rothstein, R&D Product Manager Organic Minerals, Biochem Zusatzstoffe GmbH, Alemania
OBJETIVO Los microminerales y la betaína juegan un papel relevante a nivel de estrés oxidativo, metabolismo energético, integridad intestinal y función inmunitaria. En concreto, el cinc (Zn) es un mineral esencial para las aves y juega un papel importante en el crecimiento, la salud y la productividad, ya que está involucrado en numerosos procesos metabólicos. Es importante optimizar la suplementación de Zn para equilibrar las necesidades nutricionales de las aves con el menor impacto ambiental, para favorecer una producción más sostenible. El nuevo betainato combina las propiedades de los minerales orgánicos con un aporte extra de grupos metilo, aportados por la betaína anhidra.
El objetivo de este estudio fue investigar si era posible reducir un 50% la cantidad de cinc suplementado sin comprometer la productividad y comparar los efectos de una fuente orgánica de Zn unida a la betaína respecto a una fuente inorgánica, sulfato de Zn, en pollos de engorde.
MATERIALES Y MÉTODOS
RESULTADOS Y DISCUSIÓN:
Animales:
300 pollitos macho, de un día de vida, Ross 308.
Alojamiento:
Prueba realizada en la University of Life Sciences de Poznań (Polonia)
Diseño experimental
10 aves por corral y 10 réplicas por tratamiento.
Tres tratamientos durante 35 días de engorde:
Control negativo: suplementados con 80 ppm de cinc en forma de sulfato de cinc (ZnSO4).
Control positivo: suplementados con 40 ppm de cinc en forma de sulfato de cinc (ZnSO4).
Tratamiento: suplementados con 40 ppm de cinc en forma de complejo zinc(II)-betaína (BetaTrace® Zn; Biochem Zusatzstoffe).
Parámetros medidos:
Consumo de pienso y peso vivo durante toda la fase experimental.
Digestibilidad ileal del cinc.
Parámetros productivos
Hubo un empeoramiento del índice de conversión en el grupo control positivo (40 ppm de sulfato de Zn) respecto al control negativo (80 ppm de sulfato de Zn), lo que podría indicar que reducir la dosis de cinc un 50% con una fuente inorgánica no sería suficiente para cubrir las necesidades de los pollos.
Sin embargo, a diferencia de lo observado en el control positivo, el uso de una fuente orgánica, grupo tratamiento, compensó la reducción del 50% cinc, mejorando numéricamente el peso vivo respecto al control negativo y sin perjudicar el índice de conversión. Este efecto puede explicarse por la mayor biodisponibilidad del betainato de cinc, que se consigue gracias al alto grado de unión entre el mineral y la betaína anhidra.
Figura 1: Índice de conversión del periodo experimental. ab diferencias significativas entre grupos (p<0,05)
vivo final
Figura 2: Peso vivo al final del periodo experimental. AB fuerte tendencia (p=0,06)
CONCLUSIÓN
Se observó un efecto similar que en los parámetros productivos. Numéricamente, comparado con el control negativo, 80 ppm de Zn, la digestibilidad del Zn en el grupo control positivo se redujo, mientras que el grupo tratamiento tuvo un valor similar. Esto puede indicar la mayor biodisponibilidad del betainato de Zn respecto a la fuente inorgánica.
Esta mayor digestibilidad numérica del betainato de Zn, respecto al sulfato de Zn, podría explicar la mejora del índice conversión a pesar de reducir a la mitad la dosis de suplementación.
Digestibilidad ileal del Zn
80 ppm Sulfato de Zn 40 ppm Sulfato de Zn
40 ppm BetaTrace® Zn
Figura 3: Digestibilidad ileal del Zn de las diferentes fuentes y niveles de Zn (%) (media, DE)
Este estudio demostró que una reducción sostenible del 50% de la dosis de cinc en el pienso sin perjudicar la productividad solo se pudo alcanzar con el uso del complejo zinc-betaína (BetaTrace® Zn).
Si está interesado en más datos e información sobre esta prueba, pónganse en contacto con el equipo de Biochem.

El nuevo complejo zinc(II)-betaína permite reducir un 50% el cinc en los piensos para broilers sin afectar su productividad DESCÁRGALO EN PDF
Fernando Bacha Nacoop, S.A.

La fibra neutro detergente (FND) es el análisis que más comúnmente se cita en las publicaciones sobre fibra en la nutrición de rumiantes. Se utiliza como base del cálculo del consumo de materia seca en el ganado lechero (NASEM, 2021).
La segunda ecuación de predicción de consumo que publica el NASEM, 2021, para predecir la ingestión media diaria (DMI) desarrollada por Allen et al. (2019) se basa en la composición del alimento principalmente el contenido en FND y los factores animales:
Dt_DMIn_Lact2 = 12,0 − (0,107 × Dt_fFND) + (8,17 × Dt_FAD/Dt_FND) + (0,0253 × ForFND48_FND) − {0,328 × [(Dt_FAD/Dt_FND) − 0,602] × (ForFND48_FND − 48,3)} + (0,225 × Milk_ProdTarget) + [0,00390 × (ForFND48_FND − 48,3) × (Milk_ProdTarget − 33,1)],
Donde:

Dt_DMIn_Lact2 = Ingestión media diaria kg/d (DMI, kg/d),
Dt_fFND = contenido de FDN del forraje de la dieta (%),
Dt_FAD = contenido de FDN de la dieta (%),
Dt_FND = contenido de FDN de la dieta (%),
ForFND48_FND = digestibilidad de la FDN del forraje (incubación in vitro de 48 h)
Milk_ProdTarget = producción de leche deseada (kg/d).
El análisis de laboratorio es un método empírico, define el analito (Nutritional Ecology of the Ruminant, 1994 Peter J. Van Soest) por lo que debe quedar bien determinado y que se cumpla estrictamente la metodología en el laboratorio, para lograr resultados aceptables.
El método de referencia, actualmente definido, para el análisis de la FND tratada con amilasa (aFND) es el método de Mertens, 2002. Las muestras se tratan con un detergente neutro, sulfito de Na y α-amilasa termoestable.
Como todos los análisis tiene desventajas que hay que tener en cuenta:
Subestima la pared celular porque solubiliza las pectinas, especialmente importante en las leguminosas.
Las proteínas que se dañan con el calentamiento en los alimentos procesados son retenidas en la FND sobreestimando el contenido.
La degradación de la pared celular de las plantas es uno de los procesos metabólicos más complejos e importantes en la nutrición, este mecanismo biológico involucra la acción cooperativa de una amplia gama de hidrolasas, la mayoría secretadas por microorganismos como bacterias y hongos.

Teniendo en cuenta que la pared celular de los vegetales es el mayor reservorio de energía química en la biosfera (Haynie, 2008), la fermentación ruminal de estos carbohidratos convierte a los rumiantes domésticos en agentes de transformación de esta energía, dentro del ciclo biogénico del carbono.
La característica de insolubilidad de la FND en medios neutros y acuosos le dan la capacidad de ocupar un espacio físico durante un tiempo dado dentro del rumen, por lo tanto, su efecto de llenado ruminal depende de su velocidad de degradación y de su tasa de tránsito a través del rumen.
La digestibilidad de la FND puede usarse como uno de los principales indicadores de la energía disponible de la celulosa y de las hemicelulosas, utilizable para el crecimiento microbiano del rumen y para satisfacer las necesidades de mantenimiento y producción del animal hospedador.
Pero, nos proporciona mucho menos información que el conjunto completo de parámetros dinámicos del rumen, separar el proceso en sus distintos componentes nos permite mejorar el entendimiento de su dinámica y facilita su descripción matemática.
La cinética de degradación ruminal de la materia seca (MS), del nitrógeno (N) y de algunos constituyentes de la pared celular (FND) pueden ser descritos a través de modelos no lineales. El modelo propuesto por Orskov y McDonald ha sido el más utilizado.
= a + b * (1 – exp–c*t)
En donde:
P = degradabilidad potencial
t = tiempo de incubación
a = intercepto con el eje en el tiempo cero. Representa el sustrato soluble y completamente degradable que sale rápidamente de las bolsitas de nylon.
b = la diferencia entre el intercepto (a) y la asíntota. Representa la fracción insoluble pero potencialmente degradable del sustrato, el cual es degradado por los microorganismos de acuerdo con un proceso cinético de primer orden.
c = tasa constante de la función b, (velocidad de degradación).
1-(a+b) = representa la fracción no degradable de la muestra.
La ecuación nos muestra la existencia de tres fracciones en el nutriente medido, una la no degradable que se calcula 1-(a+b); otra que no es soluble, pero es potencialmente degradable por los microorganismos ruminales (b); y una tercera rápidamente aprovechable de material soluble que incluye, también, pequeñas partículas que pueden salir de las “bolsitas de nylon” (a).


A estos tres parámetros le integraron la tasa de paso de la partícula a través del rumen “k” para obtener la degradabilidad efectiva:
Degradabilidad efectiva = a + ((b*c) / (c + k)).
Posteriormente, Mertens (1979) indicó que estos procesos describen tres fenómenos dinámicos:
1 2 3
“Lag-time” periodo de latencia o tiempo de colonización.
Periodo de rápida degradación.
Una lenta degradación proporcional al incremento de la fracción indigestible.
Antes de analizar el nivel de fibra “ideal” para las dietas de rumiantes, la FND se fermenta mediante un proceso de degradación que se inicia con la adhesión de las bacterias a la pared del vegetal, este proceso se realiza a una velocidad inversa al nivel de lignificación, ya adheridas se desdobla por acción de las celulasas y hemicelulasas, en este proceso se pierde un carbono que forma metano y es la razón por la que este proceso es menos eficiente energéticamente que el del resto de los nutrientes, y su principal producto es el acetato, precursor de la síntesis de la grasa en la glándula mamaria.
Otra peculiaridad de la degradación de la fibra es su importante periodo de latencia con lo cual, la degradación se ve muy afectada por la velocidad de paso a través del rumen (k), o sea, vacas con un nivel productivo muy alto y, por lo tanto, una ingesta muy elevada y una velocidad de paso de partícula muy alta, la degradación teórica de la fibra será baja, aunque la tasa de degradación (c) aumenta y compensa en parte el aumento de la velocidad de paso.
El objetivo de este ataque en el rumen, es la degradación del polímero (FND) y liberar los azúcares solubles. Este proceso lo llevan a cabo algunos microorganismos (bacterias y hongos) que poseen una “maquinaria” especializada.
En particular, algunas bacterias anaerobias han desarrollado un complejo conocido como celulosoma que contiene una proteína de elevado peso molecular no catalítica llamada escafoldina capaz de unir varias enzimas y dirigirlas al sustrato a la vez que las sostiene en la superficie celular.
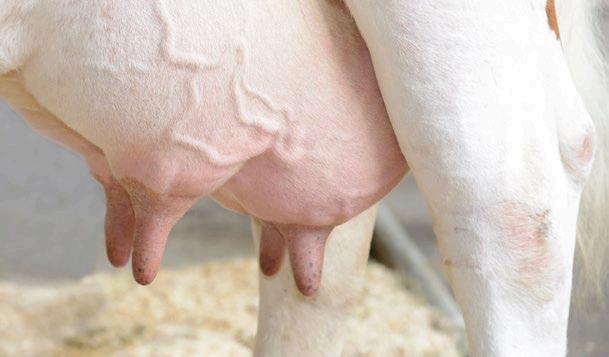

Las especies de bacterias que degradan la celulosa más importantes son:
Ruminococcus flavefaciens
Ruminococcus albus
Bacteroides succinogenes
Butyrivibrio fibrisolvens
En algunas condiciones especiales la Eubacterium cellulosolvens puede constituir la bacteria celulolítica más importante en el rumen.
Cuando los animales consumen dietas ricas en forrajes existe un elevado número de bacterias celulolíticas, sin embargo, también aparecen con dietas ricas en cereales.
Otras características, el Bacteroides succinogenes posee una celulasa extracelular que se libera y se difunde en el medio ambiente y el R. albus posee un complejo de varias enzimas con funciones específicas en la degradación escalonada de la celulosa hasta glucosa, aunque no se conoce aún el mecanismo de regulación de estas enzimas.
Incluir suficiente, pero no excesiva FDN en las dietas de las vacas lecheras de alta producción es primordial para garantizar la salud del rumen, maximizar la ingesta de materia seca y la producción láctea. En la 8ª edición de “Necesidades nutricionales de bovinos lecheros” (NASEM, 2021), el comité científico le ha dado más énfasis a la FND del forraje que a la FND del total de la dieta y recomienda una inclusión entre el 15% y el 19% de materia seca (MS) de FNDforrajera. Sin embargo, no hacen distinción con respecto a la calidad de la fibra.
A la fibra neutro detergente la podemos dividir según su comportamiento ruminal en una fracción no degradable (ndFND) y una potencialmente degradable (pdFND), de modo que:
La fracción ndFND es el residuo que queda después de 240 o 288 h de incubación ruminal. Según la bibliografía, la ndFND tiene una digestibilidad real de cero y, por lo tanto, no proporciona nutrientes al animal hospedador.
Por otro lado, la pdFND tiene potencial de proporcionar nutrientes dependiendo del equilibrio ruminal entre la tasa de degradación (c) y la velocidad de paso (k), lo que escapa de pdFND del rumen en su mayoría no es aprovechado intestinalmente por el animal y terminará en las heces, junto con la ndFND, ya que la cantidad que se fermenta en el intestino grueso es relativamente pequeña.


Coste alimentación/kg de leche*
LEVUCELL SC, levadura viva específica para el rumen, le ayuda a optimizar las dietas de alimentación de precisión. Su efecto predecible sobre la liberación de energía de la fibra permite integrarla fácilmente en sus modelos de formulación para mejorar la eficiencia alimentaria. La ciencia lo demuestra.
LEVUCELL SC. Ninguna otra levadura funciona igual.

También podríamos decir que la efectividad de la fibra en una ración o ingrediente es una función del contenido de fibra neutro detergente y del tamaño de partícula. Se denomina FND físicamente efectiva, y se calcula como el producto del factor de efectividad física, o la fracción retenida en un tamiz de > de 3,35 mm, y el contenido de FND de la ración o ingrediente.

El tamaño de la partícula ingerida también tiene que ver con el tiempo de ingesta. Cuando se ofrecen solos los piensos concentrados, se consumen rápidamente, mientras que los forrajes se consumen lentamente, en bocados pequeños y frecuentes.
Sin embargo, cuando los concentrados y forrajes se dan juntos en forma de raciones integrales (TMR), el aumento de la proporción de forraje no necesariamente aumenta el tiempo de alimentación dado que la ingesta total también puede verse disminuida en función del tipo de forraje y su contenido en ndFND.

Por ejemplo: los ensilados se consumen más rápidamente que el heno de la misma planta cuando se usan como base de la alimentación, debido al menor tamaño de partícula del ensilado y a su mayor contenido de humedad, que facilitan la formación de los bolos digestivos.
El heno por su parte se mastica lentamente, y el tamaño medio de las partículas del bolo primerio tragado puede ser menor que los del ensilado.
Por lo tanto, el tamaño de partícula del alimento usado como único dato del alimento ingerido no refleja necesariamente el tamaño del bolo y su facilidad de ser atacado en el rumen.
Haciendo un ejercicio para poner ejemplo en el Cuadro 1 podemos ver la comparación de dos dietas en las que se cambia la base forrajera.
La dieta 1 se formuló utilizando ensilado maíz y la dieta 2 con paja de trigo.
Se definió una vaca tipo: un animal de 700 kg de peso vivo (PV) con una producción media de 30 litros/día (Cuadro 2).
En el Cuadro 1 vemos las dos dietas y su valoración en fibras, almidón y el nivel forraje concentrado, aunque las dietas son muy distintas, las dos cubren las necesidades de la vaca tipo (Cuadro 2). Las diferencias en la calidad de la fibra se pueden ver en el Cuadro 3 la FND potencialmente degradable (pdFND). Para esta comparación la hemos asimilado a la diferencia de la FND menos la FAD:
pdFND = FND – FAD.

harina 61% Almi.
T.C. kg: Inclusión de la materia prima en materia fresca, en kg/día
MS kg: Inclusión de la materia prima en materia seca, en kg/día
% TC: Inclusión en % de la materia prima en materia fresca
% MS: Inclusión en % de la materia prima en materia seca
MS: Materia seca
FB: Fibra bruta

FND: Fibra neutro detergente
aFNDmo: Fibra neutro detergente con amilasa y libre de cenizas
FAD: Fibra ácido detergente
pdFND: FND potencialmente degradable
ndFND: FND indegradable
Cuadro 1. Ejemplo de raciones totales con dos fuentes de fibra muy diferentes.





Número de animales n 200
Días en el ciclo días 365 Tipo de raza Lechera
Raza primaria Holstein
Producción anual media/cabeza kg
Número de lactancia n 2,5
Intervalo entre partos meses 13
Edad a primer parto meses 24
Edad media actual meses 47
Peso corporal actual. Medio kg 713
Peso corporal medio adulto kg 750
Días de lactancia días 100
Días de gestación días 0
Producción de leche kg 30
Grasa de Leche
Proteína cruda de Leche
Cambio de reservas corporales
Condición corporal objetivo (o esperado) 3

Días para alcanzar la CC objetivo (o esperado) días 100
de nacimiento kg

Cuadro 2. Definición de vaca tipo.
Cuadro 3. Comparación de la FND y FAD entre dietas.
aFNDmo: Fibra neutro detergente con amilasa y libre de cenizas
FAD: Fibra ácido detergente
Que en la dieta 1 es de 13,15 y en la dieta 2 es de 12,65 si lo expresamos en porcentaje, en la dieta 1 el 59,61% de la FND es FAD esto significa que esta dieta tiene una pdFND del 40,39%.
Haciendo la misma operación, la dieta 2 sólo tiene el 35,20% de pdFND.
Esta potencial ventaja energética de la dieta 1, unida al contenido de materia seca de las dietas (Gráfico 1) que les facilita la elaboración del bolo de la rumia a las vacas que consumen como base forrajera el ensilado de maíz, haría que estos animales tuvieran la posibilidad de incrementar su producción, simplemente aumentando su tasa de consumo, mientras que en los animales que consumieran la dieta “pienso y paja” tendríamos que reformular toda la estrategia.
Ingesta diaria

Ensilado maíz
Materia seca
Un pequeño repaso de la importancia de la fibra neutro detergente en rumiantes
DESCÁRGALO EN PDF
Paja de trigo
Materia fresca
Gráfico 1: Comparación de ingesta diaria de los dos tipos de ración.
Trabajo ganador del concurso Jóvenes Talentos de Nutriforum 2025

Escalera-Moreno N.1, Álvarez-Rodríguez J.3, Martín-Alonso M.J.1, Villalba D.1, Molina E.1, López de Armentia L.2, Sanz A.2, Serrano-Pérez B.1
1Dpto. Ciencia Animal, UdL, Lleida, España;
2CITA Aragón - IA2 (UNIZAR), Zaragoza, España;
3Escuela politécnica superior-IA2, UNIZAR, Huesca, España
Los objetivos de selección del vacuno de carne persiguen actualmente la mejora del rendimiento productivo. Esta condición incrementa sus necesidades físicas y nutricionales, lo que puede provocar un balance energético negativo durante el último tercio de la gestación.
En determinadas épocas del año, los rebaños atraviesan períodos de subnutrición, especialmente cuando la disponibilidad de pasto es escasa debido a la estación del año o a una elevada carga ganadera.

También puede deberse a una estrategia de reducción de costes en la explotación mediante la restricción de la dieta (Sanz et al.,2024). Esta situación puede afectar la capacidad de las vacas gestantes para cubrir los requerimientos homeorréticos del crecimiento fetal.
La falta de adaptación fisiológica altera la homeostasis, generando estrés metabólico y comprometiendo tanto la salud materna como la del ternero, así como su capacidad antioxidante frente a los procesos de peroxidación lipídica (Abuelo et al., 2019).
Las diferentes razas españolas pueden desarrollar una respuesta metabólica diferente, influyendo en el bienestar de los terneros. Se denomina estrés metabólico al desequilibrio en la homeostasis fisiológica causado por un uso anormal de los nutrientes, ya sea por exceso o por deficiencia de los mismos (Lacetera, 2026).
Si este estrés es intenso o prolongado, aumentará el gasto energético del animal para enfrentarlo.
El estrés metabólico se manifiesta a través de respuestas catabólicas, lo que puede llevar a una lipomovilización excesiva, disfunciones inmunitarias e inflamatorias, y estrés oxidativo (Abuelo et al., 2019), lo que a su vez puede derivar en enfermedades metabólicas o en trastornos clínicos y subclínicos, muy comunes en la ganadería de leche (Sordillo y Raphael, 2013).
Por ejemplo, los terneros nacidos de vacas que experimentaron estrés oxidativo durante el último tercio de la gestación presentaron una respuesta inmunitaria alterada y mayor estrés oxidativo, lo que se asocia con una mayor susceptibilidad a enfermedades (Ling et al., 2018).
En este sentido las razas Pirenaica y Parda de montaña, similares en su etapa adulta, podrían responder de manera diversa a este desafío metabólico en su etapa de cría, ya que las primeras muestran un peso al nacimiento más reducido que las segundas.
El objetivo del presente estudio fue evaluar el impacto de la subnutrición durante el último tercio de gestación, la raza y su interacción sobre los niveles de marcadores de peroxidación lipídica (MDA), capacidad antioxidante total (ABTS) y la expresión de genes clave en la respuesta antioxidante (SOD2, CAT, GPX1 y NRF2) en sangre de terneros recién nacidos de aptitud cárnica.

Un total de 37 vacas en el último tercio de gestación, de entre 5 y 8 años, fueron asignadas a un diseño factorial 2x2, con dos niveles de cobertura de las necesidades nutricionales mediante una mezcla completa. Un grupo cubría sus necesidades energéticas al 100% (T100%, n=10), recibiendo 10,5 kg/día y otro cubría sus necesidades al 60% (T60%, n=9), con un consumo de 7 kg/día; y dos razas, la Parda de Montaña (PA, n=10) y la Pirenaica (PI, n=9).
Tras el parto, todas las vacas recibieron la misma dieta, que cubría sus necesidades nutricionales completamente. La composición química de la mezcla unifeed era de 111 g de proteína bruta (PB)/kg de materia seca (MS), 529 g de fibra neutro detergente (FND) /kg MS, 19 g de extracto etéreo (EE) /kg MS y 161 g de almidón/kg MS.

Los datos se analizaron con el programa JMP Pro17 (SAS Institute Inc. Cary, NC, EEUU), utilizando modelos mixtos con medidas repetidas que incluyeron como efectos fijos el nivel de alimentación y la raza, así como sus interacciones. La comparación de medias se realizó con la prueba de Tukey. Los resultados se representan como medias ± error estándar.
Las muestras de sangre fueron tomadas de la vena yugular en la primera y quinta semana de vida del ternero, en tubos Tempus RNA (Applied Biosystems) para el análisis de la expresión de los genes NRF2 (nuclear factor erythroid 2-related factor 2), SOD2 (superoxide dismutase 2), CAT (catalase), y GPX1 (Glutathione Peroxidase 1), mediante qPCR y cálculo de su cuantificación relativa (RQ) respecto a genes de referencia, y en tubos con heparina para el análisis de malondialdehído (MDA) y ABTS.
La raza afectó significativamente el peso vivo (PV) del ternero al nacimiento, con mayores pesos en la raza Parda de montaña que en la Pirenaica (51,32 ± 1,13 vs 42,84 ± 1,2, respectivamente, P < 0,001).
La concentración de MDA total, marcador de actividad pro-oxidativa, fue mayor en los terneros PA-T100% que en PA-T60% (4,08±0,15 vs. 3,39±0,17 μM, respectivamente, P<0,01), sin diferencias significativas con los de raza Pirenaica, cuyos valores fueron intermedios.
Gama de antioxidantes sinérgicos que preservan el valor nutricional, el sabor, el aroma y el color de piensos, premixes y materias primas.

Antioxidante sinérgico que combina BHT/BHA con galato de propilo y quelantes de metales. Formato líquido y polvo.
Fórmula avanzada que une BHT/BHA con compuestos quelantes y potenciadores de acción. Disponible en polvo y líquido.


Gráfica 1. Interacción entre el nivel de alimentación y la raza con los niveles de expresión génica en sangre de MDA. Las medias con letras diferentes, A,B, difieren significativamente (P < 0,05).
No se observaron efectos del nivel de alimentación, la raza ni su interacción en los niveles de ABTS, indicador de capacidad antioxidante (P>0,05).
Las expresiones de SOD2, enzima que cataliza la dismutación de superóxido de oxígeno en peróxido de hidrógeno y de CAT y GPX1, enzimas que reducen el peróxido de hidrógeno a agua y oxígeno, se vieron afectadas por la interacción entre el nivel de alimentación y la raza, teniendo sus valores más altos para PI-T100% y los más bajos para las PA-T100% (SOD1: 0,76±0,1 vs 0,37±0,1 RQ, respectivamente, CAT: 0,79 ± 0,2 vs 0,07± 0,2 RQ, respectivamente, y GPX1: 0,67± 0,1 vs 0,15±0,1 RQ, respectivamente, P<0,05), siendo las vacas T60% de ambas razas, con valores intermedios, las que no mostraron diferencias con los dos grupos anteriores.
Gráfica 2. Interacción entre el nivel de alimentación y la raza con los niveles de expresión génica en sangre de ABTS. Las medias con letras diferentes, A,B, difieren significativamente (P < 0,05).
Gráfica 3. Interacción entre el nivel de alimentación y la raza con los niveles de expresión génica en sangre de SOD2. Las medias con letras diferentes, A,B, difieren significativamente (P < 0,05).
Gráfica 4. Interacción entre el nivel de alimentación y la raza con los niveles de expresión génica en sangre de CAT. Las medias con letras diferentes, A,B, difieren significativamente (P < 0,05).
Gráfica 5. Interacción entre el nivel de alimentación y la raza con los niveles de expresión génica en sangre de GPX1. Las medias con letras diferentes, A,B, difieren significativamente (P < 0,05).
Por otro lado, NRF2, un factor de transcripción clave en la regulación de la respuesta antioxidante, mostró diferencias solo para el efecto raza, donde PI tuvo una mayor expresión de este gen que PA (2,25 ± 0,2 vs 1,08 ± 0,2 RQ, respectivamente, P<0,001).

Gráfica 6. Relación entre la raza y los niveles de expresión génica en sangre de NRF2. Las medias con letras diferentes, A,B, difieren significativamente (P < 0,05).
Los resultados anteriores evidencian que los terneros de raza Pirenaica presentaron una mayor capacidad antioxidante basal o inicial, siempre que sus madres no hubieran sufrido ningún tipo de restricción nutricional durante el último tercio de gestación, con niveles más altos de expresión de genes relacionados con esta función (NRF2, SOD1, CAT y GPX1) en comparación con los de raza Parda en las mismas condiciones.

Sin embargo, estas diferencias no se reflejaron en los niveles de ABTS, donde no se observaron variaciones significativas entre grupos. Cabe destacar que los terneros PA-T60% mostraron una tendencia a una mayor capacidad antioxidante a nivel enzimático (SOD2, CAT y GPX1) en comparación con aquellos de su misma raza que provenían de madres sin restricción nutricional y también afrontan mejor este desafío que los PI-T60%.

Esto podría estar también relacionado con que los terneros PA-T60% mostraron niveles más bajos de MDA, en comparación con los PA-T100% y con los terneros de raza Pirenaica, que tuvieron niveles moderados de actividad pro-oxidativa. Esto sugiere que la raza Parda de montaña podría contar con mejores mecanismos o mayor flexibilidad metabólica para enfrentar el estrés oxidativo derivado de la subnutrición materna.
La capacidad de adaptación puede verse reflejada también en la mayor eficiencia en el uso de nutrientes durante la gestación por parte de la raza Parda de montaña, mejorando el desarrollo fetal, visible en el mayor peso al nacimiento de sus terneros.
Por otro lado, dado que estos animales son alimentados exclusivamente con leche materna, los niveles más elevados de MDA observados en los terneros de raza Parda de montaña cuyas madres recibieron la cantidad de unifeed completa podrían atribuirse, en parte, a la mayor producción láctea característica de esta raza. Esta mayor producción, por parte de la madre, implica una demanda energética más elevada, lo que, a su vez, expone a los animales a un mayor estrés metabólico, reduciendo su capacidad antioxidante y aumentando la actividad prooxidativa.
Esto podría provocar una posible transferencia de dichos efectos a la descendencia, ya que una mayor producción láctea puede conllevar una mayor ingesta diaria, por parte de los terneros, de ácidos grasos insaturados de naturaleza prooxidante.
Finalmente, la raza Pirenaica destacaría por presentar un mejor estado oxidativo en condiciones favorables. No obstante, los resultados podrían indicar una menor flexibilidad metabólica o una peor eficiencia en el uso de los nutrientes ante un desafío nutricional, ya que no se observó ninguna tendencia hacia una mayor actividad de los genes marcadores de actividad antioxidante en los terneros Pirenaicos más desfavorecidos (T60%).
Durante las primeras cinco semanas de vida, los terneros de raza Pirenaica mostraron un mejor estado oxidativo en condiciones óptimas, pero los terneros de raza Parda de Montaña presentaron una mejor respuesta al desafío nutricional materno sufrido durante el último tercio de gestación, con una aparente mayor capacidad antioxidante y menor nivel de estrés oxidativo.

Efecto de la subnutrición durante el último tercio de gestación y la raza en el equilibrio pro- y anti-oxidante de la sangre en terneros recién nacidos de aptitud cárnica DESCÁRGALO EN PDF
Ada E. Lugo
Médico Veterinario y Nutricionista
Asesora independiente
CARBOHIDRATOS EN EL GATO: PARTICULARIDADES METABÓLICAS
Debido a las presiones evolutivas, gatos desarrollaron diversas adaptaciones anatómicas y fisiológicas en los procesos de digestión, absorción y metabolismo de los carbohidratos que reflejan su verdadera naturaleza carnívora (Pekel et al., 2020; Li y Wu, 2024).
Estas particularidades comúnmente se asocian con una capacidad mucho menor para digerir y utilizar el almidón dietético en el intestino delgado, comparado con los perros (Li y Wu, 2024).


en comparación con animales omnívoros.
Por otra parte, el intestino grueso, no posee microvellosidades y el ciego es poco desarrollado, a pesar de lo cual cuenta con una buena carga bacteriana (Osorio y Cañas, 2012; Pekel et al., 2020; He et al., 2024; Li y Wu, 2024).


En relación a los procesos digestivos, amilasa salival está ausente o es muy escasa en la saliva felina.
De igual forma, la amilasa pancreática y disacaridasas como la maltasa, poseen actividad reducida, mientras que la digestión enzimática de la lactosa y la fructosa es prácticamente nula en gatos adultos (Osorio y Cañas, 2012; Verbrugghe y Hesta, 2017; Pekel et al., 2020; Li y Wu, 2024).

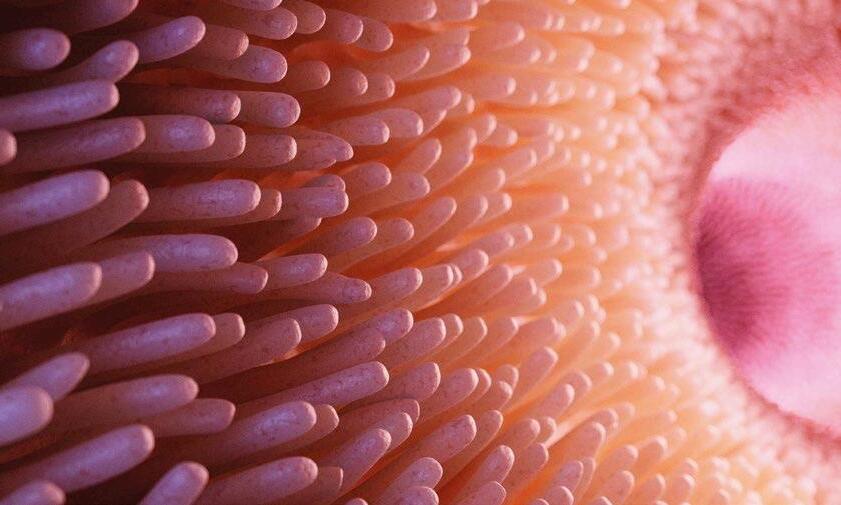
Por otra parte, el hígado felino tiene una mínima e incluso ausente actividad de la enzima glucoquinasa, quien interviene en la fosforilación de la glucosa hepática, así como una reducida actividad de la enzima glucógenosintetasa, encargada de la conversión de la glucosa a glucógeno (Osorio y Cañas, 2012; Verbrugghe y Hesta, 2017; Li y Wu, 2024)
Los aspectos anatómicos, digestivos y metabólicos mencionados, determinan una capacidad mucho menor de digestión y utilización del almidón
los CHO no hacen parte de la dieta habitual del felino doméstico, la mayoría de los alimentos comerciales destinados a esta especie actualmente disponibles en el mercado contienen como ingrediente principal cereales ricos en almidón (maíz, arroz, entre otros), a un nivel de inclusión que puede superar el 40% (Case et al., 2011; ANFAAC, 2020; Pekel et al., 2020; Laflamme et al., 2022; Singh y Singh, 2024).
Volvemos entonces al cuestionamiento inicial:
¿Pueden los gatos consumir carbohidratos en la dieta regular, o estos se convierten en enemigos

En líneas generales, los CHO no son esenciales y no hacen parte de la dieta habitual del felino doméstico, por lo que no hay requisitos establecidos de carbohidratos dietéticos para gatos (Kirk, 2011; Fascetti, 2013; Verbrugghe y Hesta, 2017; Laflamme et al., 2022; Kayser et al., 2024; Li y Wu 2024).
Sin embargo, la glucosa sí cumple un rol fisiológicamente indispensable para esta especie, al existir diversos órganos y tejidos como el cerebro, los glóbulos rojos, los leucocitos, células especializadas en la médula renal, los testículos y los ojos, que dependen exclusivamente de este carbohidrato para cubrir sus necesidades energéticas (Case et al., 2011; Verbrugghe y Hesta, 2017; ANFAAC, 2020; Laflamme et al., 2022; Kayser et al., 2024; Li y Wu, 2024; Singh y Singh, 2024).
De tal forma que es indispensable para el gato mantener una concentración estable de glucosa en sangre (3,9-6,7 mmol/L) (Osorio y Cañas, 2012; Pekel et al., 2020; Li y Wu 2024).
Esta alta demanda endógena de glucosa no puede ser satisfecha únicamente por los escasos carbohidratos presentes en la dieta natural felina, basada en presas, por lo que el gato ha desarrollado una alta capacidad de síntesis de glucosa vía gluconeogénesis, utilizando como combustibles principales los aminoácidos y las grasas presentes en su dieta natural, produciendo glucosa de forma continua (Osorio y Cañas, 2012; Verbrugghe y Hesta, 2017; Pekel et al., 2020K; Kayser et al., 202;4 Li y Wu, 2024).
Es necesario resaltar que, aunque la vía gluconeogénica podría ser suficiente para suplir las demandas energéticas de glucosa a partir de las proteínas de la dieta, esto no implica que los gatos no puedan utilizar los carbohidratos en la dieta como fuente de energía (McKenzie, 2022),
lo que es especialmente válido durante estados fisiológicos energéticamente demandantes como la gestación y la lactancia en gatas adultas 2006; Kirk, 2011; Kayser et al., 2024).

Bajo estas premisas, y aun siendo carnívoros estrictos, múltiples investigaciones avalan la posibilidad de uso de CHO en la dieta de los gatos domésticos (Danks, 2016; NRC, 2006; Kirk, 2011), fundamentando esta afirmación en elementos evolutivos, genéticos, metabólicos y tecnológicos (Fascetti, 2013; Danks, 2016; Hu et al., 2014; Kim et al., 2023; Osorio y Cañas, 2012; Pekel et al., 2020; McKenzie, 2022; He et al., 2024; Li y Wu, 2024 ;Verbugghe y Hesta, 2017).
En este sentido, se cita la presencia de modificaciones evolutivas en el genoma del gato doméstico que han determinado cambios morfológicos en el intestino y en el metabolismo de los nutrientes, lo que pudiera deberse al consumo de restos de alimentos (Kim et al., 2023) que incluían cereales como el mijo o el arroz en gatos que ya acompañaban los asentamientos humanos (Hu et al., 2014).
Por otra parte, la menor longitud del tracto gastrointestinal del gato, la carencia de la enzima amilasa salival, la baja actividad de las enzimas amilasa intestinal y pancreática y la prácticamente ausente glucoquinasa hepática, en comparación con el perro, podría ser compensado con una mayor área de superficie de absorción mucosa (Osorio y Cañas, 2012; Pekel et al., 2020; He et al., 2024; Li y Wu, 2024)
y por la existencia de mecanismos enzimáticos alternativos como la presencia de las enzimas hexoquinasa I, fosfofructoquinasa y piruvatoquinasa.
De igual forma, estudios han demostrado que la fracción de CHO en la dieta a base de presas consumida por gatos en su ambiente natural puede alcanzar de 10 a 15%, lo que implica que incluso en la naturaleza los gatos podrían tolerar una fracción mayor de CHO en su alimentación (Fascetti, 2013; Danks, 2016; Laflamme et al., 2022).
Al existir estas 3 enzimas en el hígado felino en concentraciones mayores en comparación con el perro, potencialmente compensan la ausencia de la glucoquinasa y posibilitan la fosforilación y oxidación de la glucosa (Osorio y Cañas, 2012; Li y Wu 2024), reflejando la capacidad de adaptación de las enzimas digestivas felinas (Verbugghe y Hesta 2017).
Por otra parte, aunque los gatos solo producen entre 2-5% de la amilasa pancreática producida por el perro, pueden digerir CHO complejos como el almidón con alta eficiencia equivalente, alcanzando hasta el 90% o 100% en el gato adulto (Osorio y Cañas, 2012, Verbugghe y Hesta, 2017, Li y Wu, 2024).
Adicionalmente, el patrón de consumo de múltiples comidas pequeñas a lo largo del día favorece una ingesta reducida pero regular de CHO lo que puede complementar su fisiología enzimática (Fascetti, 2013).
Este proceso permite que el almidón se gelatinice, lo que favorece su hidrólisis por enzimas intestinales mejorando su digestibilidad y evitando efectos adversos.
El almidón gelatinizado también proporciona integridad estructural a la croqueta, evitando que se desmorone (Case et al., 2011; Williams, 2019; ANFAAC, 2020; Singh y Singh, 2024) y al interactuar con las proteínas presentes contribuye a mejorar la textura y el sabor (Danks, 2016; Verbugghe y Hesta, 2017; Corsato et al. 2021).
En los alimentos húmedos, la proporción de CHO digeribles es menor, pero igualmente funcionan como agentes gelificantes al procesarse (Verbugghe y Hesta 2017).
Las consideraciones anteriores permiten sustentar que, a pesar de las limitantes descritas, los gatos pueden digerir, absorber y utilizar CHO de forma eficiente, incluso en las cantidades comúnmente contenidas en los alimentos comerciales para felinos.
Sin embargo, para que esto sea posible, el almidón utilizado en los alimentos secos formulados debe necesariamente ser sometido a procesamiento y cocción (Kirk, 2011; Fascetti, 2013; Verbugghe y Hesta 2017).
Sin embargo, es importante señalar que aún con este procesamiento y los ajustes metabólicos descritos, el gato no puede hacer frente a altas concentraciones de carbohidratos en la dieta.

Existe evidencia de que al suministrar dietas que superan el 40-50% de carbohidratos, incluso procesados y cocidos, los gatos pueden limitar voluntariamente su ingesta, comprometiendo la capacidad de la ración para cubrir todos los demás requisitos de nutrientes, especialmente las proteínas.
con el consiguiente aumento del peristaltismo, heces blandas y diarrea (Kirk, 2011; Fascetti, 2013; Verbugghe y Hesta, 2017; Laflamme et al.,2022; Ly y Wu, 2024).
Es notoria la tendencia que hoy apunta a definir a los CHO como innecesarios e incluso perjudiciales en la alimentación de los gatos domésticos, premisa que se basa fundamentalmente en su naturaleza carnívora y en las particularidades anatómicas y metabólicas que condicionan el uso de esta fracción en la dieta.
Por otra parte, el uso creciente de CHO en el alimento balanceado comercial felino, se cita con frecuencia como poco saludable e incluso responsable de patologías específicas, por lo que existe la controversia en relación a si estos son realmente beneficiosos como fuente de energía o si por el contrario son un enemigo silencioso que afecta negativamente
La evidencia analizada señala que los gatos sanos, aun con las especificaciones pueden utilizar los CHO de forma tan eficiente como el perro, siempre que estén debidamente procesados y cocidos y formen parte de una dieta nutricionalmente completa y equilibrada,
convirtiéndose en fuente de energía que permite satisfacer de manera eficiente el requerimiento celular de glucosa y ahorrando proteínas que, al no ser utilizadas como combustible gluconeogénico, son dirigidas a la síntesis de tejido.
Sin embargo, las investigaciones también indican que los CHO (procesados y cocidos) no deben predominar y/o ser excesivos en la dieta regular del gato, garantizando así que se cubran los requerimientos de otros nutrientes, particularmente proteínas.

En síntesis, es posible utilizar los CHO en la dieta de los gatos domésticos siempre que se procesen adecuadamente, se respeten las proporciones recomendadas y se garanticen los parámetros de calidad y cantidad de este ingrediente, sin olvidar que, a pesar los procesos adaptativos, el gato debe ser alimentado siguiendo las especificaciones que exige su naturaleza de carnívoro estricto.

Referencias disponibles en la versión web del artículo nutrinews.com
Los carbohidratos en raciones para gatos ¿alternativa energética o enemigo silencioso?- Parte II DESCÁRGALO EN PDF



Según un informe reciente de la Red de Información Agrícola Mundial (GAIN) del Servicio de Agricultura Exterior del Departamento de Agricultura de EE.UU. (USDA), se espera que la superficie sembrada de cereales en la UE para el ciclo 2025/26 ascienda a 49,2 Mha, superando los 48,9 Mha de la campaña anterior. De cumplirse esta estimación, finalizará la tendencia a la baja en cuanto al área cultivada de granos que inició en 2021/22.
Asimismo, se prevé que la producción total de cereales de la UE en la temporada 2025/26 supere los niveles de 2024/25 (257,6 Mt), alcanzando la cifra de 279,2 Mt.
Este aumento de la producción se explica por una mayor superficie dedicada a los cereales junto a una mejora generalizada de los rendimientos en casi toda la UE (especialmente en Francia, Rumanía y Alemania, donde las condiciones meteorológicas adversas redujeron los rendimientos de la temporada anterior).
En el caso de España, el incremento de las cosechas estará impulsado por condiciones meteorológicas cercanas a las ideales, haciendo crecer los rendimientos a niveles posiblemente cercanos a los máximos históricos.
Por otra parte, aunque el impacto final de las elevadas temperaturas en los cultivos de verano está pendiente de evaluación, tanto en Italia como en Bulgaria se proyecta un ascenso sustancial de la producción para 2025/26.
Por el contrario, en Hungría, el calor extremo junto a la escasez de lluvias perjudicó el desarrollo de los cultivo, mientras que, en Polonia, después de un clima adverso en primavera, las condiciones climatológicas en verano fueron favorables, por lo que se estima una producción similar a la de la campaña anterior.



Se prevé que la cosecha de maíz de Brasil de la temporada 2024/25 alcance niveles récord gracias a una mayor superficie plantada y a las excelentes condiciones meteorológicas.
En este sentido, el área cultivada se estima ahora en 22 Mha, cifra superior a los 21,6 Mha de la campaña anterior.
De esta forma, las previsiones apuntan a una cosecha de maíz para 2024/25 de 134 Mt, creciendo 15 Mt con respecto al período anterior.
Por lo que respecta al comercio, las exportaciones podrían no crecer significativamente debido al aumento proyectado del consumo interno para la fabricación de piensos y etanol.
Mientras tanto, para el ciclo 2025/26, se estima que la superficie dedicada al maíz crezca aún más, alcanzando los 22,6 Mha. Las proyecciones de producción estiman una disminución del 2% en comparación con los máximos históricos del 2024/25, totalizando 131 Mt.

Según un informe reciente de la GAIN, para la campaña 2025/26 se prevé que la superficie dedicada al cultivo de trigo no presente variaciones desde el ciclo anterior, manteniéndose en 5,2 Mha.
Asimismo, el área sembrada de maíz crecería un 10% interanual hasta los 4,5 Mha, mientras que la sembrada de cebada se estima que se contraiga un 10%, hasta 1,4 Mha.
La superficie dedicada al centeno rondará las 60.000 hectáreas, un 14% menor que durante el período anterior. Asimismo, la producción de trigo alcanzará las 22,8 Mt, cifra muy similar a la de 2024/25, mientras que la cosecha de maíz se estima que crezca un 29% hasta los 34,7 Mt.

La cosecha de cebada se contraerá un 10% llegando a los 4,9 Mt y la de centeno alcanzará un volumen de 200.000 toneladas (-8% interanual).


En cuanto a las exportaciones, una vez finalizada la cosecha 2024/25 las estimaciones con respecto al trigo apuntan a un volumen enviado de 15,8 Mt (-15% interanual), mientras que para la cebada la cifra alcanzaría los 2,3 Mt (-9%), y para el centeno las exportaciones crecerían un 550%, alcanzando las 11.000 toneladas.


Los datos publicados recientemente por el Departamento de Medio Ambiente y Asuntos Rurales del Reino Unido apuntan para 2025 a un incremento de la superficie dedicada al trigo, que alcanzaría los 1,53 Mha. La cifra representa un ascenso interanual del 9%, aunque todavía se sitúa por debajo de los 1,58 Mha del 2023.
Asimismo, el área sembrada de avena alcanzará las 162.000 hectáreas, suponiendo una expansión interanual del 9% y 7.000 hectáreas por debajo del máximo registrado en 2020.
Por el contrario, se estima que la superficie cultivada de cebada caiga un 13% respecto al año anterior, siendo la más baja desde 2014. En este sentido, para la cebada de invierno la contracción sería del 7% interanual, con 303.000 hectáreas, mientras que en el caso de la cebada de primavera la reducción alcanzaría el 16% en comparación con 2024 (439.000 hectáreas).


Por otra parte, con 204.000 hectáreas, el área sembrada de colza en Reino Unido se situaría un 16% por debajo de la superficie del año pasado, siendo la más baja desde 1983.
Materias primas ASFAC DESCÁRGALO EN PDF



3 & 4 DICIEMBRE 2025


Palacio de Congresos y Exposiciones de León (CONELE)

Unidos por la salud animal, comprometidos con la calidad lechera







