

1 September 2004
Dr Kurt Grove
Research Coordinator
UPR Sea Grant College Program PO Box 9011
Mayaguez PR 00681-9011
Dear Dr Grove,
I am pleased to enclose my final report for my grant entitled “The demographics of changing coral community structure in St. John, USVI” (Grant No. R-101-2-02). The report consists of (a) 17 pages of text, (b) 9 pages of figures, (c) two reprint (≈50 copies each), and (d) acknowledgement from the thesis of G Horst Please note that the graphics and tables should be considered as “drafts” as final data screening and editing still needs to be completed with a view to publication. Our sponsored projects office should be providing a summary of the financial aspects of this grant.
As I indicate in the proposal, there are several publications arising from this work that are in various stages of publication. I will be sure to acknowledge the UPR/Sea Grant award as you require.
Please contact me should you need more details Many thanks for supporting my work.
Yours sincerely,
Peter J. Edmunds Ph.D. Professor cc Rafe Boulon, Chief, Resource Management (VINP)
University of Puerto Rico/Sea Grant – Final Report
The demographics of changing coral community structure in St. John, USVI
Submitted by: Peter J Edmunds Ph D ,
Principle Investigator Department of Biology
California State University 18111 Nordhoff Street Northridge, CA 91330
e-mail: peter edmunds@csun.edu phone: 818.677.2502
Date: 1 September 2004
Summary
The goals of this project were to: (1) describe the demographic processes contributing to the decline of near-shore habitats in St. John, USVI, and (2) establish management and conservation recommendation that will contribute to the health and viability of these systems. To achieve these goals, this project was conceived as a 2-year addition to an existing 15-year project Specifically the work sought to: (a) quantify the trajectories of change on shallow coral reef habitats, (b) identify the demographic processes underlying these changes, and (c) test the potential role of seawater temperature as a physical forcing function for the changes in community structure At the conclusion of the grant, the funding has supported approx. 32 person-weeks in the field (one trip in each of 2002 and 2003) and the analyses of both new and backlogged data The results demonstrate further decline in coral cover at one site, with slight increases at a near-by location; at six other sites, coral cover changed in an idiosyncratic manner but did not amount to more than about 10% cover.
The demographic analyses focused on three systems: (a) a Montastraea annularis reef at Yawzi Point, (b) a P orites astreoides system off Yawzi Point and Cabritte Horn, and (c) the population of juvenile corals (i e , colonies ≤ 4 cm diameter) These analyses provided new insights into the dynamics of the reefs in this location, notably demonstrating that: (i) the M annularis reef is continuing to suffer demographic collapse even though the cover appears to have stabilized; the most recent losses are driven by fission of the few remaining larger colonies together with high mortality of the smallest colonies, (ii) shallow reefs on consolidated granite surfaces will likely become dominated (ca. 45% cover) by P . astreoides within the next 50 years, and (c) that juvenile coral populations are highly dynamic based on their rates of growth, recruitment and mortality Analyses of the associated between the dynamics of juvenile corals and seawater temperature have provided evidence of a strong correlative link. In brief, it appears that warm water increases the recruitment of coral larvae, but then reduces the growth rates and elevates mortality in a pattern that creates subtle changes in species assemblages. The establishment of conservation and management recommendations has been impaired by the realization that such recommendations should encompass a spatial extent that is
beyond the scope of the VINP and the USVI. Nevertheless, frequent meetings with NPS personnel and peer-reviewed publications have been used to provide a deeper understanding of the resources within the VINP and their long-term trajectories of changes. The creation of the capacity to project reef structure over 50-100 years has substantial potential to facilitate resource management goals
One of the principle goals of this proposal was to add further to an existing data set in order to provide a more ecologically relevant time scale across which trajectories of change in reef structure can be assessed. Not only has the project been successful in this regard – extending the continuous data set to the 17th year – it also has been used successfully to leverage 5 more years of support from the National Science Foundation. Thus, the integrity of this data set now is insured through to the 22nd year
Project Objectives
Near-shore, shallow coral reefs provide essential habitat for coastal fish, and a potentially renewable resource that sustains tourism and local economies. However, coral reefs have suffered more than two decades of large-scale declines in “health” that now threatens their survival Monitoring of benthic habitats is an important response to these events, but cannot alone identify the demographic causes of the changes. That is, monitoring cannot pinpoint the cause of change, nor can its potential be exploited fully to make management decisions to mitigate the damage. The current project augmented a longterm monitoring project that has been carried out in cooperation with the VI National Park to create a 14-year photographic record of coral community structure around St John. The working relationship with the VI National Park has been used to assist in the development of management strategies leading to the conservation of near-shore coastal habitats. Thus, this project meets the UPRSGC Strategic Goals of conservation and sustainable use of marine and coastal resource s, by focusing on essential fish habitat and benthic habitats. The broad goals of this research were:
1. Identification of the demographic processes leading to the decline of near-shore, coral habitats.
2. Establishing management and conservation recommendations (based on the results of the modeling) to ensure the long term health and viability of benthic habitats
To achieve these goals, three questions were addressed:
I. What are the trajectories of long-term change on the shallow coral reefs of the US Virgin Islands?
R ationale: Anthropogenic disturbances are the leading cause of biome decline and biota loss (Tilman & Lehman 2001, Woodruff 2001), and the damage affects all ecosystem components ranging from primary produces (e.g., macroalgae and algal turf), to reef corals, to commercially and ecologically important fish populations. Such extensive damage now requires management at the ecosystem level to mitigate the effects (Christensen et al. 1996, Novecek & Cleland 2001, Steneck & Carlton 2001) and this can
be achieved only with explicit management goals that are realized through policies, protocols and practices operating at multiple scales (Christensen et al. 1996).
Environmental monitoring plays a central role in this process by furnishing the data and ecological context necessary to develop and support ongoing management efforts (Christensen et al 1996) Coral reefs provide an excellent example of an ecosystem with an immediate need for large-scale management (e.g., Christensen et al. 1996, Oliver 1999), because widespread downward trajectories of change (Hughes 1994, Brown 1997, Hoegh-Guldberg 1999, Wilkinson 1999) indicate a gloomy outlook for the their continued survival (Hoegh-Guldberg 1999, Knowlton 2001) Apart from a few studies incorporating large temporal and/or spatial scales (e g., Hughes 1994, Connell et al 1997, Chiappone et al. 1997, Murdoch & Aronson 1999, Bythell et al. 2000, Ninio et al. 2000), most monitoring programs still are too limited in duration and geographic extent (Connell 1997) to provide sufficient data to craft effective management strategies for coral reefs.
My ongoing research in St. John now is attaining considerable value because the trends in community structure are documented over 15 years. Such long time scales are essential to reliable describe changes in populations of long lived organisms (such as corals), and to build effective management programs based on ecologically significant trends. Continuation of this research for 2 more years (expanding the study to its 16th and 17th year) will help to ensure that the consequences of coastal development and urban impacts can be predicted with higher confidence than currently is possible.
II. What are the demographic processes underlying the long term changes in coral community structure?
R ationale: Terry Hughes and his colleagues in Australia recently have underscored the importance of demographic analyses in understanding the significance of changes in coral community structure (Hughes & Tanner 2000) Phrased another way, coral community data can be misleading without an indication of demographic trends, such that a population increasing in coral cover might mask an imminent demographic collapse. This portion of the research will be used to construct demographic models of three representative and ecologically important coral species in order to better understand the likely fate of near shore coral communities
III. What is the relationship between coral demographics and seawater temperature, and does this vary among genera?
R ationale: A correlative analysis of the effects of seawater temperature on coral demographics will be central to understanding how global climate change (i e , increasing temperatures) might affect coral populations and degrade coastal resources in the VI. This approach assumes that temperature characteristics will vary among sampling intervals (justified below), and effect the demographics of corals. This approach does not discount the possible effects of other environmental parameters such as irradiance and sedimentation, but it assumes the profound relationship between temperature and calcification (Lough & Barnes 1997) will result in temperature having the greatest effect. This hypothesis will be tested using growth and mortality data collected for corals in Lameshur Bay since 1996. The proposed research will provide two more years of data [from 2001 (already funded) to 2002, and 2002 to 2003] that will provide six data points
to test the association with seawater temperature (daily seawater temperature are available from the VINP from 1987).
A
brief history of the study reefs (1987-2002)
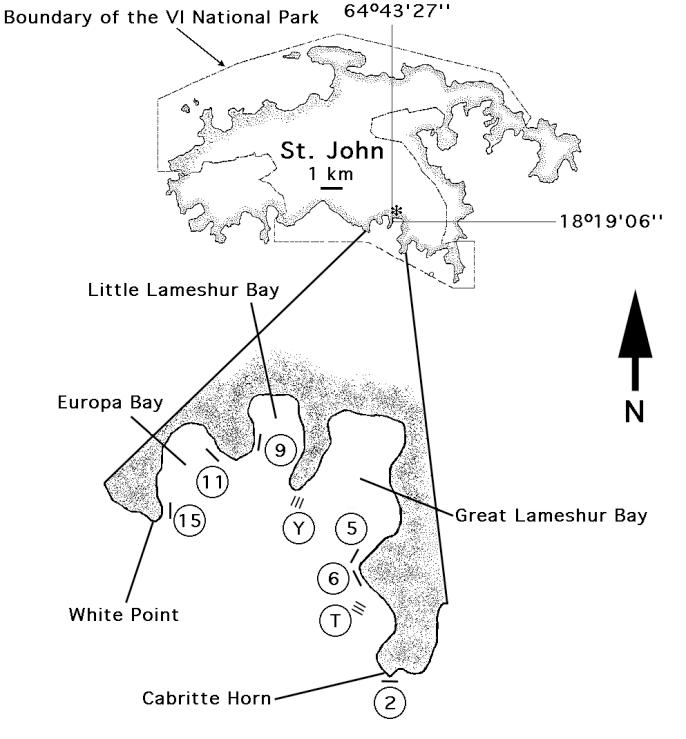
Fig 1 Map showing the lo cation of the study sites in Great L am eshur Bay and its env irons Y and T r efer to the Yaw zi and T ektite s ites that w ere es tab lished in 1987, and th e numbers ref er to th e f ive random sites established in 1992. S ix additional s ites were established at ≈5 m depth in 1994 for the purpose of quantifying coral r ecruitm ent (not shown)
The fringing coral reefs that are the focus of this study are located on the south coast of St. John, US Virgin Islands, and lie within the VI National Park and Biosphere Reserve They are relatively free of local anthropogenic disturbances including excessiv e fishing [although fishing is permitted (Rogers & Beets 2001)], terrestrial development and anchoring, and are patrolled by NPS rangers. Between Cabritte Horn and White Point - two headlands that delineated the spatial extent of much of this project (Fig. 1) -- the coral reefs in shallow water (<9 m depth) typically grow as a thin veneer on an igneous rock substratum In deeper water (10-18 m), large patches of reef are dominated by stands of Montastraea annularis (20-40% cover), and rich growths of seagrasses (Thallasia and Syringodium) fill the sandy bays. Historically, well-developed mangrove communities characterize the shoreward margin of the bays, often providing a seawater margin to a brackish salt pond.
The reefs of Lameshur Bay first were described in the late 1950’s and 1960’s by John Randall (Randall 1961), and the scientists in the Tektite undersea laboratory (Collette & Earle 1972). Thereafter, few investigators have worked in this location (Rogers & Teytaud 1988, Rogers & Beets 2001) with the only consistent efforts (other than this project) organized through the Virgin Islands National Park (VINP) (Rogers 1985, 1999) Despite this attention, other than the present research there only are published long-term data describing coral reef community structure at one site on the south coast of St John, beginning in 1989 (Rogers et al. 1991, 1997).
The current project began with a description of the basic ecology of the shallow reefs (Edmunds et al. 1990) and an analysis of the impacts of Black Band Disease (Edmunds 1991) However, it was the conspicuous bleaching phenomenon that affected reefs throughout the Caribbean in 1987 (Roberts 1987) that prompted the start of rigorous monitoring. Two photographic study areas were established at 10 m and 14 m depth at Yawzi and Tektite, respectively, and since have become the core areas of the study. Initially, coral cover at Yawzi (mostly M. annularis) was > 44% in 1987 and was higher than the 32% found at Tektite. Bleaching in 1987 ultimately had little effect on coral cover, but Hurricanes Gilbert (1988) and Hugo (1989) initiated changes that have affected coral reef community structure for more than a decade Coral cover at Yawzi began a decline in 1989 that resulted in a >50% loss by 1998; at Tektite, <1 km away,
Fig 2 Coral com munity stru cture at the Yaw zi and Tektite sites over the firs t 12 y ears of th e s tudy (mean ± SE shown for n = 30 qu adrats per tim e interval) (Edmunds 2002). Arrows show th e timing of Hurricanes Gilb ert (I), Hugo (II) and Marilyn (III) Repeated measures ANOVA shows th at th e per cen tage cover of coral, m acro alg ae and CT B d iffer ed s ignifican tly ov er time Reefs in bo th of thes e locations ar e sp atially dominated w ith dens e growths of co lumnar lobate colonies of Mo n ta s tra ea a n n u la ris and the ch anges in coral cover have been m atched by a s trik ing abs ence of recruits of this sp ecies
coral cover avoided the damaging effects of the Hugo-generated waves, and increased slowly by 35% to end the 1987-98 period with 43% coral cover (Fig. 2) In response to these changes, macroalgae increased to exploit the vacant space preempted from the crustose coralline algae, turf and bare space category (CTB), and supplemented by the death of coral. Thus, surprisingly in an international MPA, coral cover has declined at a rate comparable to that seen on the heavily impacted reefs of Jamaica (Hughes 1994), yet on a spatial scale of kilometers, patches of healthy reef remain. The inferences from these trends are that: 1) the reefs in St. John are likely influenced by events acting on a scale larger than the VI National Park, and 2) that patches of “healthy” reef (like Tektite) might be critical for the regrowth (i.e., “reseeding”) of denuded areas Strikingly, during the 16-years of this study, few recruits of M annularis have been seen, yet this species is spatially dominant in many deeper areas Together with similar results from other locations (Bak & Engel 1979), this findings suggests that M. annularis may exploit a “storage effect” (sensu Warner & Chesson 1985) to maintain coexistence with other species
In response to the limitations of not having selected the Yawzi and Tektite sites at random, in 1992 five new sites were added using random coordinates, together with the restriction that the substratum should be at similar depths and on coral reef (Fig. 1). The coral reef community structure at these locations turned out to be different from that found at Yawzi and Tektite (Fig. 3), namely in that coral cover was low (<10%) and changed idiosyncratically and in a directionless manner (Fig 3)
In response to evidence that the St. John reefs were variable across a spatial scale of kilometers (e.g., compare Figs. 2 and 3), in 1994 I began to quantify this variation (Edmunds & Bruno 1996) with the goal of exploiting it to better understand the factors creating this spatial mosaic of reef structure To test the hypothesis that such patterns might be related to recruitment, I started to quantify juvenile corals (colonies ≤ 40 mm
diameter) and to relate their distribution to that of adults. Initial efforts focused on shallow areas along much of the south coast (i.e., beyond the area shown in Fig. 1) and revealed the anticipated patchy distribution of adult corals (Fig 4) Juvenile corals also were common and, although distributed in a patchy manner (Fig. 5), were not significantly correlated with the percentage coral cover (i.e., the adult corals) at each site (P>0.05) Thus, it appears that while juvenile corals are variable in their distribution, they are not related to the contemporaneous distribution of adults
To explore the dynamics of juvenile coral distribution in more detail, techniques were developed to assess the growth and mortality at the same sites as used for density determinations. The shallow reefs of Lameshur Bay are well suited to such studies, as the igneous substratum provides smooth surfaces upon which is it relatively easy to identify and track coral recruits This analysis of juvenile coral success revealed an unusually high mortality rate (37% for all taxa; Edmunds 2000) that suggests that the coral communities in shallow water “turn over” rapidly. An analysis of the ecology of the encrusting hydrozoan Millepora alcicornis on the rocky substratum suggested that much of this flux is powered by the blasting of the rocky surfaces by suspended sand, cobbles and boulders during storms (Edmunds 1999). Despite the dynamic nature of these reefs, juvenile coral mortality rate was not associated with the distribution of adult corals, and therefore is unlikely to have a simple causal relationship in determining the distribution of adults
Fig 3 Coral com munity stru cture between 1992 and 1998 at fiv e s ites (7-9 m depth) selected at random with in th e Lameshur Bay ar ea ( Fig 1) (Edmunds 2002) Although mark edly different from Y awzi and T ektite in term s of community structure and dynamics, they have underscored th e importance of spatial var iation and the po tential ro le of cor al r ecru itment in establishing su ch sp atial v ariation
Fig. 4 . Per centage coral cover ( mean ± SE) at 18 sites (5 m dep th) along 10 km of the south coas t of S t. John. Coral cover diff ered s ignif icantly among s ites (P< 0 05) (Ed munds 2000)
In light of the apparent high turnover rate of the juvenile corals, recently (supported in part with the present funding) I started to explore the potential causes of mortality. To this end, correlations of temperature and recruitment have revealed a strong relationship between these factors Briefly, coral recruitment increases in years that have warmer temperatures, but the subsequent success of the recruits (measured as growth and mortality) declines (refer to attached reprint from Marine Ecology Progress Series) With a differential effect among genera, a 12-year warming trend in
Fig. 5 . Box p lot showing the d ensity of juven ile corals along 10 km of th e south co ast of S t John and at the sam e sites as shown in F ig. 4 . Although juv enile corals are dis tributed in a patchy mann er, they are not correlated w ith th e distr ibution of adults
Lameshur Bay (0.06oC/y) has resulted in a significant change in the relative abundance of genera of juvenile corals between 1994 and 2001. Thus changes in seawater temperature appear to be related (perhaps causally?) to subtle changes in the dynamics of recruiting corals
Summary of results 2002/2003 (and set in the perspective of the 19872003 data set)
Funding from this grant has supported two years of work and generated results that have been used to address the goals and questions at the core of the original proposal. Additionally, the progress achieved with the present grant has been instrumental in securing 5 more years of support from the National Science Foundation, Long Term Research in Environmental Biology program (NSF-LTREB) The results of the present award have been incorporated into one publication supported in part by the present grant, and several others in various stages of preparation Moreover, the “longterm” aspect of this work is best served by publishing the data in temporal “bites” that are ecologically meaningful, such as the most recent summary of the long term data that covered the period 1987-1998. At the time of submission of this report, the manuscripts are not ready for distribution, although the most of the results are presented in figure and table form (see below)
Goal 1:Identification of the demographic processe s leading to the decline of near-shore, coral habitats.
Questions
I. What are the trajectories of long-term change on the shallow coral reefs of the US Virgin Islands?
The “long-term” trajectories now can be described with up to 17-years of continuous data (Yawzi and Tektite sites) that last were published for the period 1987-1998 (1992-1998 for the six random sites). In short, the most recent data for percentage cover of the main benthic categories reinforces the trends already extablished in 1998 (refer to Figures 2 and 3). Namely, that the reef at Yawzi point continues to lose coral cover, while the reef at the Tektite gains coral slowly; at the random sites the coral cover changes in an idiosyncratic pattern. It is unclear why the trajectories of change are drastically different on reefs separated by less than 1 km, but it is likely that the protection of Tektite from storm waves (notably those from Hurricane Hugo in 1989) and the slightly cooler water in this location may play a critical role The demographic analyses underscore the limitations of quantifying coral community structure by percentage
cover alone, namely by (a) providing “early warning” of further declines, (b) identifying the likely mechanisms of these declines, and (c) bringing into focus the demographic changes in demographic processes that have occurred between 1988 and 2003.
II. What are the demographic processes underlying the long-term changes in coral community structure?
The development of the demographic models is an ongoing and time-consuming task that (to date) has been applied to two corals species (Montastraea annularis and P orites astreoides) and one coral functional group (juveniles). Model development has been slowed by the difficulty of collecting sufficient data in the field (principally the tagging and relocation of 100’s of coral colonies), and analyzing images for coral “condition” in pairs of multiple time points (the analytical approach applied to M. annularis). Nevertheless, the approach already has supported substantial progress towards addressing the core questions in the present proposal. Most notably, the demographic analyses have revealed that:
• coral reefs in shallow water will likely become spatially dominated by P orites astreoide s within 50 years;
• populations of juvenile corals (< 40 mm diam) are highly dynamic and exhibit changes that broadly are related to seawater temperature;
• the population of Montastraea annularis at Yawzi Point (that locally dominates coral cover in this location) will likely continue declining in size over the next 10-50 years;
• the mechanisms mediating the loss of M annularis at Yawzi Point have changed over the last 17 years, switching from broad mortality of all size classes, to extensive partial mortality of larger colonies (leading to fission) and high mortality of the smallest colonies;
• the loss of M. annularis at Yawzi Point will likely extirpate larger colonies from the reef (i e , those >150 cm2 in area) with potentially severe consequences for population viability on account of the disproportionately severe loss of reproductively mature individuals;
• the decline in cover of M. annularis at Yawzi Point are unlikely to be reserved by coral recruitment alone – even with extreme recruitment (defined by magnitude and frequency), the abundance of large colonies cannot be restored within 50 years.
III. What is the relationship between coral demographics and seawater temperature, and does this vary among genera?
Results from the St. John project that were analyzed during the present award show that the density and dynamics of juvenile coral populations track seawater temperature. In brief, recruitment increases during warm years, but the same years result in reduced growth rates and elevated mortality that may drive changes in the species composition of juvenile coral populations; the results from 2002
and 2003 further support this trend. Unfortunately, it remain challenging to complete similar analyses for individual taxa due to limited sample sizes and limited ability to increase them due to restrictions on the time spent underwater as a result of SCUBA.
Goal 2: Establishing management and conservation recommendations (based on the results of the modeling) to ensure the long term health and viability of benthic habitats.
The discovery that the coral reefs along a subset of the bay along the south coast of St John are likely affected by processes acting on a large spatial scale poses substantial challenges to the management of resources on a local scale Put simply, within the realms of geo-political and economic constraints, it is not practical to manage the reefs of St. John on the 100-10,000 km scale that likely is necessary to affect changes in trajectories of the major components of the benthos (notably of the stony corals).
However, close discussions with senior NPS personnel indicate that results of the present study have been influential in developing management strategies within the context of understanding better what needs to be protected and what the likely outcomes will be of ongoing changes Most recently (20 May, 2004), the Superintendent of the VI National Park indicated that one of the most valuable “deliverables” of the present work would be enhanced ability to project the reef structure into the future In other words, to be able to predict (within specific assumptions of consistent conditions) what the reef will look like in 50 or 100 years is invaluable to resource managers.
Fieldwork and personnel supported
Funding from this grant support two field trip to St. John and 3 graduate students as summarized below.
F ield trip 1: A ugust 2002
Location: St John, USVI
Host facility: Virgin Islands Environmental Resource Station
Duration: 4 weeks
Personnel:
Peter J Edmunds PhD (PI)
Laurie Requa (Graduate Student)
Geoffrey Horst (Graduate Student)
Robin Elahi (Graduate Student)
Elizabeth Keach (Volunteer Assistant)
F ield trip 2: A ugust 2003
Location: St. John, USVI
Host facility: Virgin Islands Environmental Resource Station
Duration: 4 weeks
Personnel:
Peter J. Edmunds PhD (PI)
Laurie Requa (Graduate Student)
Geoffrey Horst (Graduate Student)
Robin Elahi (Graduate Student)
Elizabeth Keach (Volunteer Assistant)
Graduate R esearch A ssistant ship used for data analysis at CSUN:
1. Laurie Requa (Graduate Student)
2. Geoffrey Horst (Graduate Student)
3. Robin Elahi (Graduate Student)
4. Mairead Maheigan (Graduate Student)
Graduate theses (MS) supported in part by UP R /Sea Grant support
1. Geoffrey Horst: Effects of temperature and CO2 variation on calcification and photosynthesis of two branching reef corsals (June 2004).
2. Laurie Allen-Requa: The effects of algal shading on juvenile corals (ongoing, expected completion in December 2004).
3. Robin Elahi: Size and age effects in the physiological ecology of reef corals (ongoing, expected completion in December 2005)
Outreach activities
The research station used as a field base for this research is operated by Clean Islands International and hosts multiple user groups ranging from academic researchers to K-12 “eco-camps”. The eco-camps attract children from the US mainland, Puerto Rico and Caribbean Island nations, and were ongoing during both of the field trips supported by this grant. My research team participated in these camps by providing evening presentations on reef ecology, conservation and the pros/cons of a career choice in science.
Matching Funds
2002: 1. National Park Service: ≈$3,000 (assist with field costs)
2. CSUN: ≈$5,000 (upgrade VIERS compressor facility)
2003: 3. National Park Service: ≈$3,000 (assist with field costs)
4. CSUN: ≈$5,000 (graduate research assistantship)
2004: 5. CSUN: ≈$5,000 (graduate research assistantship)
TOTAL MATCHING (2002-2004) = $21, 000
Note: based, in part on the progress attained with this grant, the data set and publications from my long term research in St. John recently were awarded 5 years of support from the National Science Foundation This funding comes from the Long Term Research in Environmental Biology program (LTREB) and will cover the costs of field work and the
graduate labor necessary to analyze the data. This new award, together with the UPR/Sea Grant award, will provide 22-years of continuous data from this site.
Tables
Table 1. Leslie matrices for Montastraea annularis at Yawzi Point (9 m depth) for three time periods. Size Class I ≤ 50 cm 2 , Size Class II ≤ 150 cm 2 , Size Class III ≤ 250 cm 2 , Size Class IV > 250 cm 2 ; Death = death rate, and N = sample size (no. of colonies)
Values for λ are the intrinsic rate of population change, where the rates of decline are inversely proportional to the value of λ. Thus, the population declined faster between 1998 and 2003 (λ = 0 634) than it did between 1988 and 1993 (λ = 0 660)
Interval Size Class I II
Table 2. Leslie Matrix and population vector for Porites astreoide s growing at 3-5 m depth on granite rock at Yawzi Point and Cabritte Horn. Size Class I ≤ 40 mm, Size Class II ≤ 80 mm, Size Class III > 80 mm. The matrix displays transition probability describing the probability of an even occurring. For example, the probability that a small colony will stay in size class I after 1 year is 69.6% Note that this matrix is calculated with a time interval of 1 year, whereas the matrices for M. annularis were calculated over an interval of 5 years The vector describes the number of colonies present at time 0 in a finite area (in this case 10.75 m2). The product of the matrix and the vector gives the population size at the next time interval (the vector at time t+1); iterative multiplications were used to generate the projection shown in Fig 12.
MATRIX
VECTOR
Figure Legends
Figure 1. (see above) Map of the study sites in St. John USVI.
Figure 6. Community structure at the Yawzi site (9 m depth) between 1997 and 2003 (’02-’03 supported with this grant): values display mean [±SE, n ≈ 30 (with sample size varying slightly by year)] percentage cover of the benthos of the three main components of the benthos. (A) Coral cover (all scleractinians combined), (B) Macroalgae (mostly Dictyota, Halimeda, and Lobophora), and (C) Crustose coralline algae, algal Turf and Bare space (CTB) combined (these categories are pooled as they cannot be distinguished reliably in planar images) Broadly speaking, the coral cover at this site has continued the downward trend initiated in 1987 and most recently culminating in an apparently stable cover of about 12%. In conjunction with these change, macroalgal cover has climbed consistently (although substantial inter-annual changes occur) with broadly inverse changes in the CTB category.
Figure 7. Community structure at the Tektite site (14 m depth) between 1997 and 2003 (’02-’03 supported with this grant): values display mean [±SE, n ≈ 30 (with sample size varying slightly by year)] percentage cover of the benthos of the three main components of the benthos (categories as in Fig. 6) In marked contrast to the Yawzi site, coral cover at Tektite site has maintained a slow but consistent increase to about 45% in 2003. Throughout the full study period (1987-2003), the changes in the macroalgae and CTB categories have been relatively minor with a slight increase in macrolagae and a slight decrease in CTB
Figure 8. Community structure at six sites selected at random through Lameshur Bay and its environs (see Fig. 1 for locations), and monitored from 1992 to 2003 (’02’03 supported with this grant): values display mean [±SE, n ≈ 18-40 (with sample size varying slightly by year, and increasing to 40 in 2001)] percentage cover of the benthos of the three main components of the benthos (categories as in Fig. 6) Coral cover has remained low and displayed idiosyncratic changes (i.e., varying among sites) over the 12 year period, but macroalgae and CTB hav displayed more consistent changes among sites For instances, 2001 marked a year with dramatic increased in macroalgae and concomitant declines in CTB.
Figure 9. Density (no. per 0.25 m 2 , 80-240 replicate measurements per datum) of juvenile corals (defined as colonies ≤ 4 cm diameter) at 5-9 m depth within Lameshur Bay and its environs between 1994 and 2003 (’02-’03 supported with this grant). (A) Overall density (i.e., pooled among taxa) of juvenile corals expressed for all small colonies (“pooled) and pooled, less F avia fragum and Siderastrea radians. The rationale for removing these species in a subset of the data is their attainment of sexual maturity at a small size, and therefore functioning as adults (not juveniles) at small size. Over the 12 year period,
both categories increased slightly in a response that is likely related functionally to seawater temperature (refer to attached reprint). (B) Density of juvenile corals separated by genus for taxa with sufficient replication for a finer resolution analysis. While F avia and S. radians (many of which probably were adults in the size class analyzed) accounted for a substantial fraction of upward trend in recruitment between 1994 and 2003, a similar trend was shown by P orites and A garicia
Figure 10. Growth rate (mm/y) of juvenile corals at 5- 9 m depth in Lameshur Bay and its environs between 1996 and 2003 (’02-’03 supported with this grant); means ± SE for all values. (A) Growth for all taxa combined (i.e., pooled), and (B) growth separated by major taxa with sufficient replication for a finer grain analysis. There are substantial changes in growth rates from year to year, and these probably are related to seawater temperature (refer to attached reprint).
Figure 11. Mortality rates (%/y) and population growth (defined as λ and derived from the Leslie Matrices describing population changes) of juvenile corals at 5-9 m depth in Lameshur Bay and its environs between 1996 and 2003 (’02-’03 supported with this grant). (A) Mortality rates of groups (i.e., by genus and pooled among genera) varies inversely with growth rates (Fig 6) and varies markedly among years Mortality rates probably are related to seawater temperature (refer to attached reprint). (B) λ values for all juvenile corals between 1996 and 2003: for the analysis of a cohort of corals (as accomplished here), λ is ≤ 1 where 1 is a stable population (i.e., not changing in size) and values < 1 indicated rates of decline in population size. In the present analysis, the λ values show that juvenile coral populations were most stable in 1997 and 2002 and displayed a protracted period of rapid population reduction between 1998 and 2001.
Figure 12. Population projection for Porites astreoides growing at 3 m depth on rocky surfaces in Lameshure Bay (specifically Yawzi Point and Cabritte Horn) The projection uses the Leslie matrix from 2002-2003 with the rationale that the matrices varied little between 1999 and 2002 (λ =0.945, 0.980, 0 986 and 0.945, respectively) for a population with 78 recruits in an area of 10.75 m2, and 185 colonies allocated among three size classes: I (≤ 40 mm diam = 43% of the population), II (≤ 80 mm diam = 26% of the population), and III (> 80 mm diam = 31% of the population). The population assumes these conditions do not change, and projects the population structure over 100 years in 1-year increments; the dark line displays the percentage coral cover of P . astreoides assuming the colonies are circular in diameter. In brief, the projection suggests that coral reefs in shallow water on granite surfaces will become dominated with P astreoides, with colony densities increasing ca 5-fold and the percentage cover of this species increasing ca. 9-fold.
Figure 13. Population projections for Montastraea annularis at the Yawzi Point study site (9 m depth). Projections are shown using Leslie matrices derived from three
growth periods (1988-1993, 1993-1998 and 1998-2003) and prepared using four size classes of coral colonies (i.e., discrete lobes of M annularis): Size Class I (≤50 cm2), II (≤150 cm2), III (≤ 250 cm2) and IV (> 250 cm2). Areas were determined using NIH Image 1.62 software and color images recorded on Kodachrome film (1988-1998) or as digital images (2003) Projections were prepared for a population with no recruitment (first row), consistent recruitment of 25 recruits/m2 per 5 year interval (second row), and pulsed recruitment with 500 recruits/m2 every 25 years (third row; note logarithmic yaxis) Time 0 values are based on the mean lobe (= colony) density in 1987 (Edmunds 1994) In comparing the three time intervals, the relative importance of the events affecting colonies of M. annularis has changed markedly from the first to the final 5-year period Initially, mortality rates were broadly similar among size classes, but more recently, the mortality rates of the smallest colonies has increased markedly. At the same time, the rate of transitioning from big to small colonies (i.e., fission) has increased markedly reflecting the high chance that big (and old) colonies will suffer partial mortality and break into multiple smaller pieces, which then have a high chance of dying. The contrasts among the recruitment scenarios were selected to test the hypotheses that present-day population sizes could be maintained by (i) consistent recruitment (even though few recruits ever have been found on natural surfaces), or (ii) episodic recruitment occurring on a time frame exceeding the longevity of individual investigators. The most important outcomes of these projections are: (A) colonies in the two largest size classes (III and IV) effectively disappear from the populations regardless of the recruitment level or the matrix employed, (B) recruitment (at rates never recorded) can theoretically maintain populations dominated by smaller colonies, but these do not transition to larger size classes due to high rates of mortality, (C) even though the cover of M annularis at this site has stabilized over the last 5 years (Fig. 6), the population projections predict that further declines already are underway, and most significantly (D) the population decline of M annularis at Yawzi point likely do not reflect a system routinely maintained by infrequent recruitment. Thus, it is more likely that the loss of M annularis represent a “new” phenomenon driven by high mortality rates (First row, insets) and frequent fission to smaller colonies.
References cited
Bak RPM, Engel MS (1979) Distribution, abundance and survival of juvenile hermatypic corals (Scleractinia) and the importance of life history strategies in the parent community. Mar Bio 54, 341-352
Brown BE (1997) Adaptation of reef corals to physical environmental stress. Advances in Marine Biology 31: 221-299
Bythell JC, Hillis-Starr ZM, Rogers CS (2000) Local variability but landscape stability in coral reef communities following repeated hurricane impacts Mar Ecol Prog Ser 204: 93-100.
Chiappone M, Sullivan KM, Sluka R (1997) Status of reefs in the central Bahamas on a large-scale survey. Proc 8th Int Coral Reef Symp 1: 345-350.
Christensen NL, Bartuska AM, Brown JH, Carpenter S, D’Antonio C, Francis R, Franklin JF, MacMahon JA, Noss RF, Parsons DJ, Peterson CH, Turner MG, Woodmansee RG (1996). The report of the Ecological Society of America committee on the scientific basis for ecosystem management. Ecological Applications 6: 665-691.
Collette BB, Earle SA (1972) Results of the Tektite Program: ecology of coral reef fishes. Bull Nat Hist Mus Los Angeles County 14: 178 pp
Connell JH, Hughes TP, Wallace CC (1997) A 30-year study of coral abundance, recruitment, and disturbance at several scales in space and time Ecological Monographs. 67: 461-488.
Edmunds PJ (1991) The extent and affect of Black Band Disease on a Caribbean reef. Coral Reefs 10: 161-166
Edmunds PJ (1999) The role of colony morphology and substratum inclination in the success of Millepora alcicornis on shallow coral reefs Coral Reefs 18: 133-140.
Edmunds PJ (2000) Patterns in the distribution of juvenile corals and coral reef community structure in St. John, US Virgin Islands. Mar Ecol Prog Ser 202:113124.
Edmunds PJ, Bruno JF (1996) The importance of sampling scale in ecology: kilometerwide variation in coral reef communities Mar Ecol Prog Ser 143: 165-171.
Edmunds PJ, Roberts DA, Singer R. (1990) Reefs of the northeastern Caribbean I. Scleractinian populations Bull Mar Sci 46: 780-789.
Hoegh-Guldberg O (1999) Climate change, coral bleaching and the future of the world's coral reefs Australian J Freshwater Marine Biology 50: 839-866.
Hughes TP (1994) Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science 265: 1547-1551.
Hughes TP, Connell JH (1999) Multiple stressors on coral reefs: a long-term perspective Limnol Oceanog 44: 932-940.
Hughes TP, Tanner JE (2000) Recruitment failure, life histories and long-term decline of Caribbean corals. Ecology (in press)
Knowlton N (2001) The future of coral reefs. Proc Natl Acad Sci USA 98: 5419-5425.
Lough JM, Barnes DJ (1997) Several centuries of variation in skeletal extension, density and calcification in massive P orites colonies from the Great Barrier Reef: A proxy for seawater temperature and a background of variability against which to identify unnatural change. J. exp. Mar. Biol. Ecol. 211: 29-67
Maragos JE, Grober-Dunsmore R (1999) Proceedings of the Hawaii Coral Reef Monitoring Workshop June 9-11, 1998. East-West Center, Honolulu, Hawaii.
Murdoch TJT, Aronson RB (1999) Scale-dependent spatial variability of coral assemblages along the Florida reef tract Coral Reefs 18: 341-351
Ninio R, Meekan M, Done T, Sweatman H (2000). Temporal patterns in coral assemblages on the Great Barrier Reef from local to large spatial scales Mar Ecol Prog Ser 194: 65-74
Novacek MJ, Cleland EE (2001) The current biodiversity extinction event: scenarios for mitigation and recovery. Proc natl Acad Sci (USA) 98: 5466-5470
Oliver J (1998) Coral reef monitoring origrams and management of the Great Barrier Reef pp 39-53 In Maragos JE, Grober-Dunsmore R (eds) Proceedings of the Hawaii coral reef monitoring workshop.
Randall JE (1961) Tagging reef fishes in the Virgin Islands. Proc Gulf and Caribbean Fisheries Institute 14: 210-241.
Roberts CM (1997) Connectivity and management of Caribbean coral reefs. Science 278: 1454-1457.
Rogers CS (1985) Degradation of Caribbean and Western Atlantic coral reefs and decline of associated fisheries Proc 5th Int Coral Reef Symp 6: 491-496.
Rogers CS (1999) Sampling may be hazardous to your monitoring program. pp 57-64 In Maragos JE, Grober-Dunsmore R (eds), Proc of the Hawai’is Coral Reef Monitoring Workshop.
Rogers CS, Teytaud R (1988) Marine and terrestrial ecosystems of the Virgin Islands National Park and Biosphere Reserve Biosphere Reserve Rep 29: 1-112.
Rogers CS Beets J (2001) Degradation of marine ecosystem s and decline of fishery resources in marine protected areas in the US Virgin Islands Environmental Conservation 28: 312-322.
Rogers CS, McLain LN, Tobias CR (1991) Effects of Hurricane Hugo (1989) on a reef in St. John, USVI. Mar Ecol Prog Ser 78: 189-199.
Rogers CS, Garrison V, Grober-Dunsmore R (1997) A fishy story about hurricanes and herbivory: seven years of research on a reef in St John, US Virgin Islands Proc 8th Int Coral Reef Symp 1: 555-560.
Steneck RS, Carlton JT (2001) Human alterations of marine communities: students beware! Pages 445-468 in MD Bertness, SD Gaines, ME Hay, editors. Marine community ecology. Sinauer Associates, Sunderland, Massachusetts, USA
Tilman D, Lehman C (2001) Human-caused environmental change: impacts on plant diversity and evolution. Proc Nat Acad Sci (USA) 98: 5433-5440.
Warner RR, Chesson PL (1985) Coexistence mediated by recruitment fluctuations: a field guide to the storage effect. Am Nat 125: 769-787.
Wilkinson C (2002) (editor) Status of coral reefs of the world: 2002 Australian Institute of Marine Science pp 377.
Woodruff DS (2001) Declines of biomes and biotas and the future of evolution. Proc natl Acad Sci (USA) 98: 5471-5476