
FINAL REPORT
Spatio-Temporal Reconstruction of insular and regional climatic oscillations in the E. Caribbean using fluorescent coral banding
J. Morell, J. Corredor and A. Cabrera, Department of Marine Sciences
U.
of Puerto Rico, Mayaguez
INTRODUCTION
Credible evidence of climate change has slowly but unequivocally arisen from the observation and or reconstruction of current and past biological, chemical and physical processes. This evidence has prompted the scientific community to search for information regarding processes pertinent to the prediction of future climate changes (i.e. estimation of biochemical processes with inherent importance on global change). Furthermore, the community has recognized the need to reconstruct past climate oscillations as a mean to enhance our understanding of the processes modulating them. These needs indicate the urgency for reconstructing climate indicators, such as precipitation, across various time scales.
Water-soluble humic acids, arising from the degradation of plant material in soil, are readily transported through river runoff to the coastal ocean. Given their fluorescent nature (Zepp and Schlotzhauser, 1981), dissolved humic materials (HM) have been used extensively as river plume tracers and characteristic fluorescent signatures can in fact be used to discriminate different water masses (De Zouza Sierra et al., 1994).
Reef-building corals, long-lived organisms that constantly deposit calcium carbonate skeletons, have been analyzed by numerous researchers with the purpose of documenting recent environmental changes. The fluorescent banding along the growth axes of these organisms has been linked to their exposure to insular and continental runoff (Susic et al. 1991). Fluorescent fulvic and humic acids of terrestrial origin are argued to be sequestered in discreet bands in the skeletal material of massive corals (Isdale, 1984; Boto, and Isdale, 1985) and may thus provide a paleo-climatic record of terrestrial runoff. Freshwater flow to the coral reef might thus be reconstructed from fluorescent banding in corals subject to terrestrial runoff sources. Such reconstructions have been performed for the Florida Everglades as recorded in corals of Florida Bay (Smith III et al., 1989) and the Australian Great Barrier Reef (Susic and Isdale, 1989). More recent studies (Hendy et al. 2003) used yellow luminescent lines seen in multicentury Porites cores obtained from eight different locations on the central Great Barrier Reef, to developed "a 373-year chronology providing a good proxy for both nearby river runoff and Queensland summer rainfall”
The influence of fresh waters from the Orinoco and Amazon Rivers is apparent in the eastern Caribbean region extending as far north as the greater Antilles and as far west as 75o W in the central Caribbean Sea (Froelich et.al. 1978; Muller Karger et al., 1989;
Corredor and Morell, 2001; Hellweger and Gordon, 2002). Fluorescence of HM from both these rivers has been reported (Zika et al, 1993; Leenheer, 1980) and the spreading of colored dissolved organic matter in the Orinoco River plume (mainly humic material) throughout the eastern Caribbean has also been previously described (Blough et al., 1993). Additional evidence for such influence has been provided by salinitysilicate ratios (Froelich et.al., 1978; Corredor and Morell, 2001).
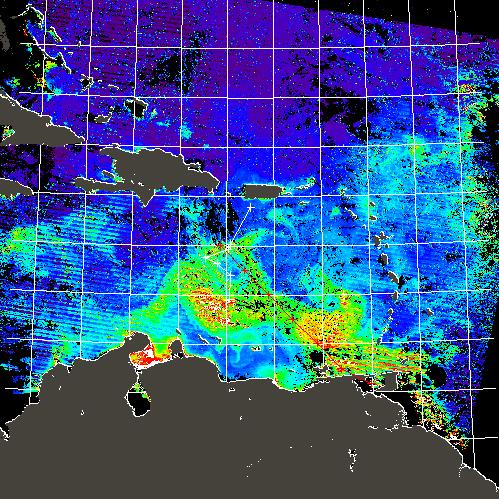

MODIS CDOM absorption coefficient
Most HM, while resistant to microbial degradation, are nonetheless subject to UV-photodecomposition in the upper layers of the ocean (Kieber et al, 1990; Skoog et al., 1996); the former authors having estimated a half life for HM in the oceanic mixed layer of 5-15 y. Other researchers have evidence for fertilization by HM through biological degradation as well as photochemically mediated release of inorganic nutrients (Amon and Benner, 199; Bushaw et. al, 1996). Susic and Isdale (1989) have developed a model for humic acid carbon export by measuring the content of this material sequestered in corals at varying distances from the Burdekin River of north-eastern Australia and assuming simple linear dilution of river water with seawater. Nevertheless, this approach disregards the well-known process of photobleaching of HM which causes exponential decay of the fluorescent signal (Zika et al. 1993; Jones and Amador 1993).Recent information regarding changes in the concentration and composition of the HM present in the a Orinoco river plume, as it is spreads throughout the Eastern Caribbean, indicate the potential use of this signature captured in coral skeletons to reconstruct climatic trends in the region. While we (Del Castillo et al., 1998) have provided evidence for the occurrence of highly refractory fluorophore (ex/em 250/425) which might constitute a robust signal for net exposure to riverine waters, we (Morell and Corredor, 2001) have also derived an empirical model for the photobleaching of more reactive fluorophores (ex/em 350/440) in the same plume. There is significant decay of the humic fluorescent signal bringing about a ten-fold reduction in humic content of Orinoco River plume waters during its transit across the Caribbean from latitude 10o N to 15o N.
Despite the apparent success of fluorescent banding studies, serious doubt has been cast on the “nature and causes “of fluorescent banding in coral and hence on its utility as a climate proxy. Such studies (Barnes and Taylor, 2001) indicate that apparent fluorescence may well be due to the structure of the coral skeleton and not to sequestered HM. In order to address this concern, we developed an extraction methodology that allows fluorescence characterization of HM in a liquid matrix, free from the confounding effects of the coral skeleton.
We examine here decadal-scale time series for sequestration of humic materials in corals from diverse sites of the eastern Caribbean reflecting a gradient of continental freshwater influence. We report on the stability, magnitude and gradient of the insular and continental riverine signals in corals along a transect across the Orinoco plume dispersal axis providing fundamental information for the reconstruction of records on the inter-annual variability of freshwater runoff and validating the use of extracted humic material as a proxy for terrestrial runoff.
METHODOLOGY

Corals from the E. Caribbean region were sampled using a pneumatic drill with a 5 cm diameter by 20 cm length steel bit (Table 1). Samples were frozen immediately until processing. In the laboratory, coral cores were slabbed using a rock saw taking care to orient the cut along the axis of growth.
Table.1

Station Localization
Grenadines
Canouan
Nevis island Nevis reef
Saba Saba Bank
coral slabs
Number of cores
M.a.3, M.a.4, D.s.1

Date of sampling
March 2001
M.f.1, M.f.2 May 1999
M.a.1, M.a.2 May, 1999
St Croix Fredericksted Sp1, sp2
Mona Island Mona Channel
Pinnacles Parguera
June 2000
M.a 1 April 2002
M.a 1,2 November 2002
Micro-cores, ca. 3 mm. in diameter, were sampled along the coral growth axis, pulverized using mortar and pestle and the coral powder dispensed in 1ml HCl 0.05N. Spectrofluorometric analysis was carried out using a Hitachi F2000 scanning spectrofluorometer. Detailed excitation/emission matrices were obtained by sequential scanning of the extracts over the emission spectrum of interest (400-600 nm) at 2 nm excitation intervals. Sclerochronology was established by X-radiography. Radiographs were then utilized for identifying density banding associated with yearly variation in skeleton growth.
The image below depicts an x-radiograph of a core sample of the reef building coral Montastrea annularis. Note the higher density annual growth bands.

RESULTS:
Spectrofluorometric analysis of coral humic extract collected at Canouan, Grenadine Islands yielded a fluorescent signature comparable to that of Orinoco River water. A slight displacement of the emission maxima is attributable to pH differences between both samples (Fig. 1)
Excitation (nm)
Figure 1. Contour plots of synchronous excitation/emission spectrofluorometric analysis of top: subsections of sample M.annularis coral sample and bottom: Orinoco river water collected at the Gulf of Paria.
In order to test the relationship between coral extract fluorescence and riverine influence, fluorescence observations of samples collected at Canoaun, the station closest to the Orinoco river mouth, were pooled into climatology and compared with the climatologic river flow data (Figure 2, below)
Agreement between spectrofluorometric and temporal signatures is consistent with the argument linking the presence of fluorescent dissolved organic material in coral skeletons to concentrations of these organic compounds at the time of coral growth. The spatial distribution of coral fluorescence climatology is presented in figure 3 (below). The data indicates an exponential decay of the regional fluorescence signal along an axis extending from the Southeastern Caribbean (Canouan) and the Northeastern Caribbean consistent with observations by Del Castillo et al. (1999) and Morell and Corredor (2001).
Although the fluorescence signal includes a region wide component, contrasting coral fluorescence with local precipitation indicates a regional or localized response to precipitation and presumably runoff (figure 4, below) as previously reported.
Time series plots of coral fluorescence at Canouan, Mona and Parguera are included below.
Conclusions:
The apparent success of early fluorescent banding studies has been questioned as serious doubt has been cast on the “nature and causes “of fluorescent banding (Barnes and Taylor, 2001). The extraction methodology we developed, allowing fluorescence characterization of HM in a liquid matrix free from the confounding effects of the coral skeleton, has provided further proof of sequestration of dissolved organic material into coral skeleton. Furthermore, the analytical and spatiotemporal resolution of the method has allowed differentiation between regional and local process contributing to FDOM concentrations in the water column.
Observations at all sites evidence a marked variability in fluorescent organic matter concentrations. These variations indicate the need for further understanding of the dissolved organic carbon component and its responsiveness to climatic processes across temporal and spatial scales.
Other outcomes:
Research Assistantships:
Mrs. R. García, PhD student 7/2000 -12/2000
Mr. A. Cabrera, MSc. student 01/2001- 12/2002 (graduated)
Mr. A. Cabrera PhD student, 1/2003-3/2004
Publications:
Manuscript currently in preparation, submission planned for summer 2006.
REFERENCES:
Amon R.M. and R. Benner. 1996. Photochemical and microbial consumption of dissolved organic carbon and dissolved oxygen in the Amazon River system. Geochem. et. Cosmochem. Acta. 60:1783-1792.
Barnes, D.J. and Taylor, R.B. 2001. On the nature and causes of luminescent lines and bands in coral skeletons. Coral Reefs 19: 221-230.
Blough, N.V. O.C. Zafiriou and J. Bonilla. 1993. Optical absorption spectra of water from the Orinoco River outflow: Terrestrial input of colored organic matter to the Caribbean. J. Geophys. Res. 98: 22712278.
Boto, K. and P. Isdale. 1985 Fluorescent bands in massive corals result from terrestrial fulvic acid inputs to the nearshore zone. Nature: 315: 396-397.
Bushaw, K.L., R.G. Zepp, M.A. Tarr, D. Schulz-Jander, R.A. Bourbonniere, R.E. Hodson, W.E. Miller, D.A. Bronk and M.A. Moran. 1996. Photochemical release of biologically available nitrogen from dissolved organic matter. Nature 381: 404-407
De Souza Sierra, M.M., O.F.X. Donard, M. Lamotte, C. Belin and M. Ewald. 1994. Fluorescence spectroscopy of natural waters. Mar. Chem. 47: 127-144.
Del Castillo, C.E., P.G. Coble, J.M. Morell, J.M. López and J.E. Corredor. 1998. Analysis of optical properties of the Orinoco River by absorption and fluorescence spectroscopy: Changes in optical properties of the dissolved organic matter. Mar. Chem. 66: 35-51.
Froelich P.N., D.K. Atwood and G.S. Giese. 1978. The influence of the Amazon River water on surface salinity and dissolved silicate concentration in the Caribbean Sea. Deep-Sea Res. 25:735-744.
Gill, I, Olson, J.J. and Hubbard, D.K., 1996. Corals, paleotemperature and the aragonite-calcite transformation: Reply. Geology, v. 24, no. 1, pp.92-93.
Gill, I., Olson, J.J., and Hubbard, D.K., 1995. Corals, paleotemperature and the aragonite-calcite transformation. Geology, v. 23, no. 4, p. 333-336.
Hendy, E.J., Gagan, M.K. and Lough, J.M. 2003. Chronological control of coral records using luminescent lines and evidence for non-stationary ENSO teleconnections in northeast Australia. The Holocene 13: 187-199.
Isdale, P.J. 1984. Fluorescent bands in massive corals record centuries of coastal rainfall. Nature 310: 578-579.
Jones R.D. and J.A. Amador. 1993. Methane and carbon monoxide production, oxidation and turnover times in the Caribbean Sea as influenced by the Orinoco River. J. Geophys. Res., 98, 2353-2359.
Kieber, D.J., J. McDaniel and K. Mopper. 1989. Photochemical source of biological substrates in seawater: Implications for carbon cycling. Nature 341: 637-639.
Hellweger, F.L. and Gordon, A.L., 2002. Tracing Amazon River water into the Caribbean Sea. Journal of Marine Research, 60(4): 537–549.
Leenheer, J.A.. 1980. Origin and nature of humic substances in the waters of the Amazoin River Basin. Acta Amazonica. 10: 512-526.
Morell J..M. and J.E. Corredor, 2001. Photomineralization of fluorescent organic matter in the Orinoco River plume: Estimation of ammonium release. J. Geophys. Res., 106 (C8) 16,807-16,813.
Muller Karger, F.E., C. R. McClain, T.R. Fisher, W.E. Esaias and R. Varela. 1989. Pigment distribution in the Caribbean Sea: Observations from space. Prog. Oceanog. 23.
Skoog, A., M. Wedborg And E. Folqvist. 1996. Photobleaching of fluorescence and organic carbon concentration in a coastal environment. Mar. Chem. 55: 333-345.
Smith, T.J. III, J.H. Hudson, M. B. Robblee G. V. N. Powell, and P.J. Isdale. 1989. Freshwater Flow From The Everglades To Florida Bay: A Historical Reconstruction Based On Fluorescent Banding In The coral Solenastrea bournoni. Bull. Mar. Sci. 44. 274-282.
Susic, M. K. Boto and P. Isdale. 1991. Fluorescent humic acid bands in coral skeletons originate from terrestrial runoff. Mar. Chem. 33: 91-104.
Susic, M. and P. Isdale. 1989. A model for humic acid carbon export from a tropical river system using coral skeletal fluorescence data. Conf. Proc. Hydraulic and Environmental Modelling of of Coastal, River and Estuarine Syustems. Bradford, UK, p. 588.
Zepp, R.G. and P.F. Schlotzhauser, 1981. Comparison of photochemical behavior of various humic substances in water. III Spectroscopic properties of humic substances. Chemosphere 10: 479-486.
Zika, R.G., P. Milne and O.C. Zafiriuo. 1993. Photochemical studies of the eastern Caribbean: An introductory overview. J. Geophys. Res. 98: 2223-2232.
Susic, M., Boto, K.G. & Isdale, P.J. Fluorescent coral skeletal bands result from terrestrial runoff. Marine Chemistry, 33, 91-104 (1991).
Susic, M. & Isdale, P. A model for humic acid carbon export froma tropical river system using coral skeletal fluorescence data. In Hydraulic and Environmental Modelling of Coastal, Estuarine and River Waters (ed. Falconer, R.A.,Goodwin, P. & Matthews, R.G.S.), PP. 588-597 (Gower, Aldershot, UK, 1989).