Sea Grant Final Report
Long-term passive acoustic tracking of juvenile blacktip and lemon sharks: an investigation into the spatiotemporal dynamics and connectivity of shark nursery habitat in St. John, USVI
Gregory B. Skomal, PhD
Massachusetts Marine Fisheries, 838 S. Rodney French Blvd., New Bedford, MA, 02744; phone: 508-910-6305; University of Massachusetts, School for Marine Science and Technology, New Bedford, MA; e-mail: Gregory.Skomal@state.ma.us
Bryan M. DeAngelis
The Nature Conservancy, North America Region, South Ferry Rd., Narragansett, RI, 02882; phone: 401-874-6030; email: bdeangelis@tnc.org
EXECUTIVE SUMMARY
Project title: Long-term passive acoustic tracking of juvenile blacktip and lemon sharks: an investigation into the spatiotemporal dynamics and connectivity of shark nursery habitat in St. John, USVI.
Date: May 21, 2013
Project Number: R-31-1-10
Investigators and affiliation:
Gregory B. Skomal, PhD, Massachusetts Marine Fisheries, 838 S. Rodney French Blvd., New Bedford, MA, 02744; phone: 508-910-6305; University of Massachusetts, School for Marine Science and Technology, New Bedford, MA; e-mail: Gregory.Skomal@state.ma.us
Bryan M. DeAngelis, The Nature Conservancy, North America Region, South Ferry Rd., Narragansett, RI, 02882; phone: 401-874-6030; email: bdeangelis@tnc.org
Dates Covered: February 1, 2010 – January 31, 2013.
Summary of Impacts and Contributions
1. Objectives
The overall objective of the study was to provide multi-year information on the movements and habitat utilization of blacktip (Carcharhinus limbatus) and lemon (Negaprion brevirostris) sharks in USVI coastal nurseries using acoustic tracking. Specifically, our objectives were to:
(i) Determine the degree to which young sharks are utilizing near-shore coastal mangrove habitats, including how they partition the area between species and how long they remain in the area. This objective has been met. We quantified residency, site fidelity, and home range for young-of-the-year (YOY) and juvenile blacktip and lemon sharks in two important shark nurseries on St. John, USVI. We examined the degree to which these two species exhibit habitat partitioning.
(ii) Assess emigration rates. This objective has been met. In quantifying habitat use in Fish and Coral Bays by YOY blacktip and lemon sharks, we assessed residency and the subsequent rates at which they emigrate from these nurseries.
(iii) Examine levels of connectivity between nursery habitats. This objective has been met. Both Fish Bay and Coral Bay have been identified as important nurseries for blacktip and lemon sharks. Despite extensive tagging and tracking efforts over six years, very few sharks utilized both embayments.
(iv) Determine if these areas are isolated “pockets” of productivity. This objective has been met. Given the lack of movement exhibited by YOY blacktip and lemon sharks between multiple
embayments along the coast of St. John and St. Thomas, USVI, Fish Bay and Coral Bay appear to be isolated pockets of productivity worthy of protection.
(v) Increase community awareness and education about the ecological importance of sharks to a healthy coral reef ecosystem. This objective has been met. Working with the Coral Bay Community Council, Maho Bay Camps and Concordia Eco-Resort, and the University of the Virgin Islands, we disseminated this information to the general public and local newspapers over the course of this study.
2. Advancement of the field
Using passive acoustic telemetry, this research provides the first detailed examination of nursery habitat utilization by YOY and juvenile blacktip and lemon sharks in the Caribbean. In doing so, we quantified the temporal and spatial aspects of habitat use in terms of residency, site fidelity, and home range. As a result, we were able to delineate essential nursery habitat for these two species within Coral Bay and Fish Bay on St. John, USVI. We provide evidence that these species utilize this habitat for the first several months of their lives, a period when they are most vulnerable to predation. Some sharks remain in these embayments throughout the year. Based on our findings, we now know that shark nursery habitat for these two species in this region is limited in size and represents isolated pockets of productivity. Therefore, these nurseries are critical to the health of blacktip and lemon shark populations in the region. As such, management measures should be not only be taken to protect this habitat from anthropogenic impacts, but also to protect the young sharks from exploitation in these areas.
3. Problems encountered
The only problem encountered during this study involved sample size. We proposed to tag a total of 80 young sharks comprising 40 blacktip and 40 lemon sharks. However, we were unable to capture the latter and our resulting sample size was 52 blacktips and 29 lemons. Although this did not impact our assessment of residency and site fidelity, we were unable to estimate home range for the lemon shark due to low sample size in each bay.
4. Research impacts Management
The results of this research will be disseminated to the appropriate local, regional, and federal management bodies so that measures to protect this essential habitat can be implemented. This information will provide local fisheries managers and other regulatory bodies with the tools necessary to establish effective conservation measures such as fishing closures, habitat protection, additional marine protected areas, and limits on coastal development. This work has already proven useful in protecting juvenile sharks and shark nursery habitat because it was used by USVI coastal management agencies to decline development requests to build submerged docks and piers within Coral Bay, one of the shark nurseries documented by this study The information collected by this study has also been beneficial to local stakeholders. The Coral Bay Community Council, a local group, has incorporated our data into their comprehensive
environmental database for Coral Bay. These data are used to demonstrate the ecological value and importance of the bay.
Outreach
This project was also designed to increase community awareness and education about the ecological importance of sharks to a healthy coral reef ecosystem. Through public presentations and local media coverage, we actively increased public awareness of the importance of protecting, conserving, and managing nearshore essential fish habitat for sharks, as well as other species in the marine community. By incorporating community volunteers into the project, we elevated the importance of coastal mangrove habitat.
5. Other important impacts or products
UVI graduate student Bryan Legare (Bryan.Legare@tpwd.state.tx.us) was funded by this study. He completed and defended his master’s thesis, which was centered on this research, in December, 2011. His thesis title is: Juvenile blacktip shark (Carcharhinus limbatus) and lemon shark (Negaprion brevirostris) movements within two nursery areas of St. John, United States Virgin Islands. Over the duration of the study, a total of $48,051.80 was funded by Sea Grant to support Bryan’s participation. The work contained in the thesis is currently being edited for formal publication in peer-reviewed journals. It is anticipated that this will result in at least two publications highlighting the fine and broad-scale behavior of these two species in USVI nurseries. Moreover, additional analyses are being conducted on these data and those collected in 2012; the results of this work will also be published.
6. Sources of matching funds
The source of matching funds was the time and fringe benefits associated with the participation of G. Skomal, principal investigator. Total match was $27,022.71, which was $2,122.71 in excess of the original commitment.
7. New extramural funds in addition to match
Not applicable.
8. Additional time and effort attributed to PI’s, Co-PI’s and associates
See item 6.
9. Additional benefits
Noted above.
Background and Project Relevance
Sharks represent an important component of the Caribbean marine ecosystem as apex predators and keystone indicators of coral reef productivity and health (Bascompte et al. 2005). Sharks are typically slow growing, k-selected species with low intrinsic rates of reproduction (Holden 1974). It has been hypothesized that shark population growth is limited by the amount of suitable nursery habitat (Springer 1967). Given these life history characteristics, the identification and understanding of shark nursery areas are critical for the proper protection and management of these species. Recent research indicates that shark nursery habitat in St. Thomas and St. John, USVI is limited to a handful of relatively small bays, estuaries, and reef lagoons (DeAngelis 2006; DeAngelis et al. 2008). The Essential Fish Habitat (EFH) Generic Amendment to the Fishery Management Plans of the U.S. Caribbean states that the waters and substrates that comprise EFH of the USVI are readily susceptible to a number of human activities including overexploitation, sedimentation, pollution, marinas, and commercial and industrial development (Caribbean Fishery Management Council 1998). These anthropogenic activities may pose a risk to shark EFH in USVI coastal waters.
Although highly productive areas of shark nursery habitat have been identified in this region, these areas are too limited spatially to support large numbers of young coastal sharks (DeAngelis 2006; DeAngelis et al. 2008). Therefore, the protection of existing shark nursery habitat is critical to the long term conservation of these ecologically important species. However, little is known about the spatial and temporal dynamics of habitat use. Specifically, information on movements, site fidelity, immigration and emigration between and within habitats, and associations with National Park boundaries is crucial for coast-wide habitat protection and management. Additionally, the extent to which these “pockets” of productivity are interconnected is unknown. Our initial tracking data indicate that there is no exchange between these isolated pockets of productively, but our sample sizes are limited to date. If this is indeed the case, enhanced protection of these areas is warranted to sustain local shark populations. Clearly, shark nurseries in this region need further investigation to characterize the importance of these areas to individual species, to quantify, in a spatial and temporal sense, the use of these areas by young sharks, and to assess the protection afforded by marine reserves. The use of passive acoustic telemetry is currently the most efficient method to monitor the long-term movements of fishes (Heupel et al. 2006). This technology has been used extensively in the shark nursery areas of western Florida to study movement patterns of juvenile sharks (Heupel and Hueter 2002, Heupel and Simpfendorfer 2002). The objective of the current study was to provide multi-year information on the movements and habitat utilization of sharks in USVI coastal shark nurseries using this technology. Long term acoustic tracking allowed us to: (i) determine the degree to which young sharks are utilizing near-shore coastal mangrove habitats, including how they partition the area between species and how long they remain in the area; (ii) assess emigration rates; (iii) examine levels of connectivity between nursery habitats; and (iv) determine if these areas are isolated “pockets” of productivity. This information has provided local fisheries managers and other regulatory bodies with the tools necessary to establish effective conservation measures such as fishing closures, habitat protection, additional marine protected areas, and limits on coastal development. Moreover, this project was designed to increase community awareness and education about the ecological importance of sharks to a healthy coral reef ecosystem.
Methods
Study Sites
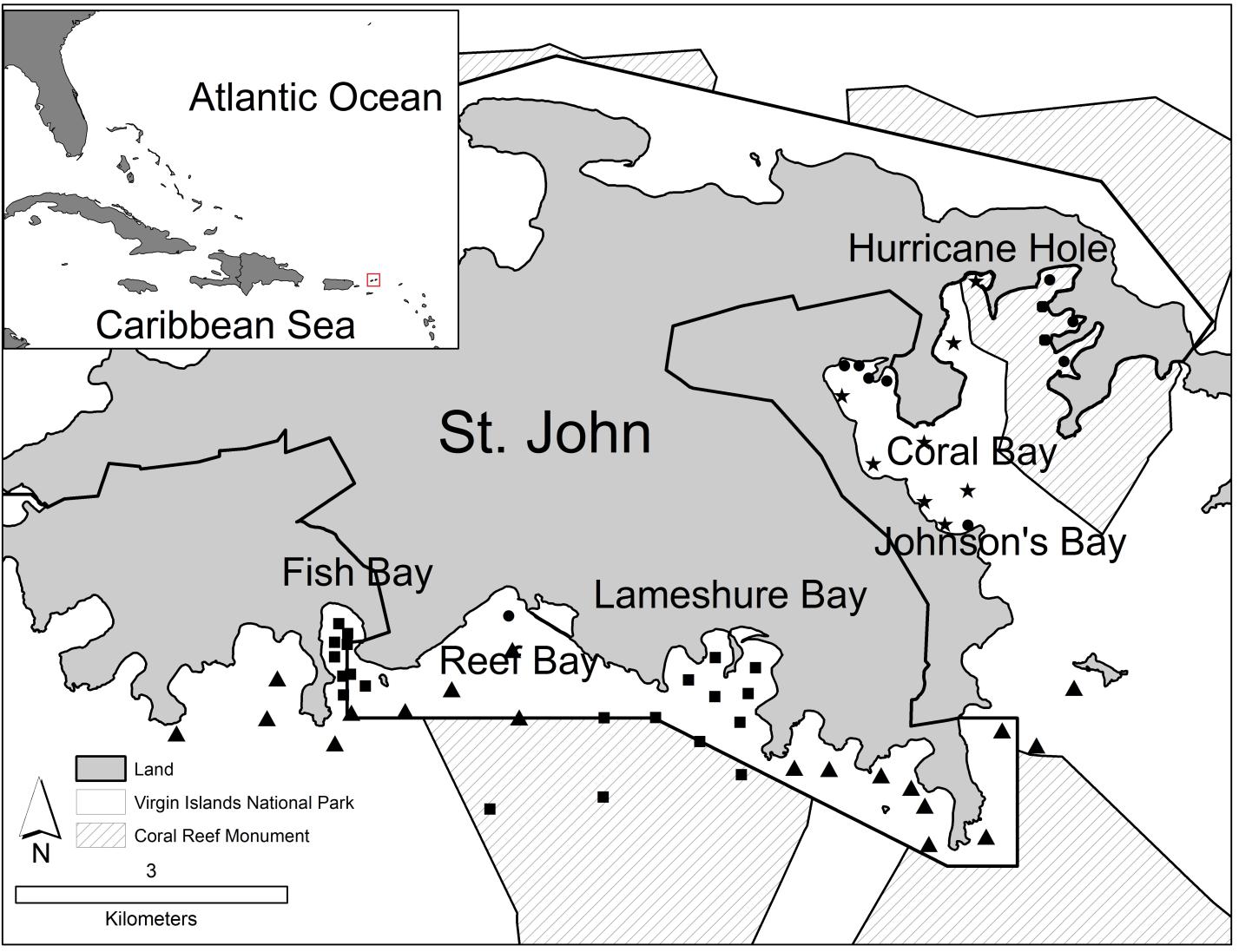
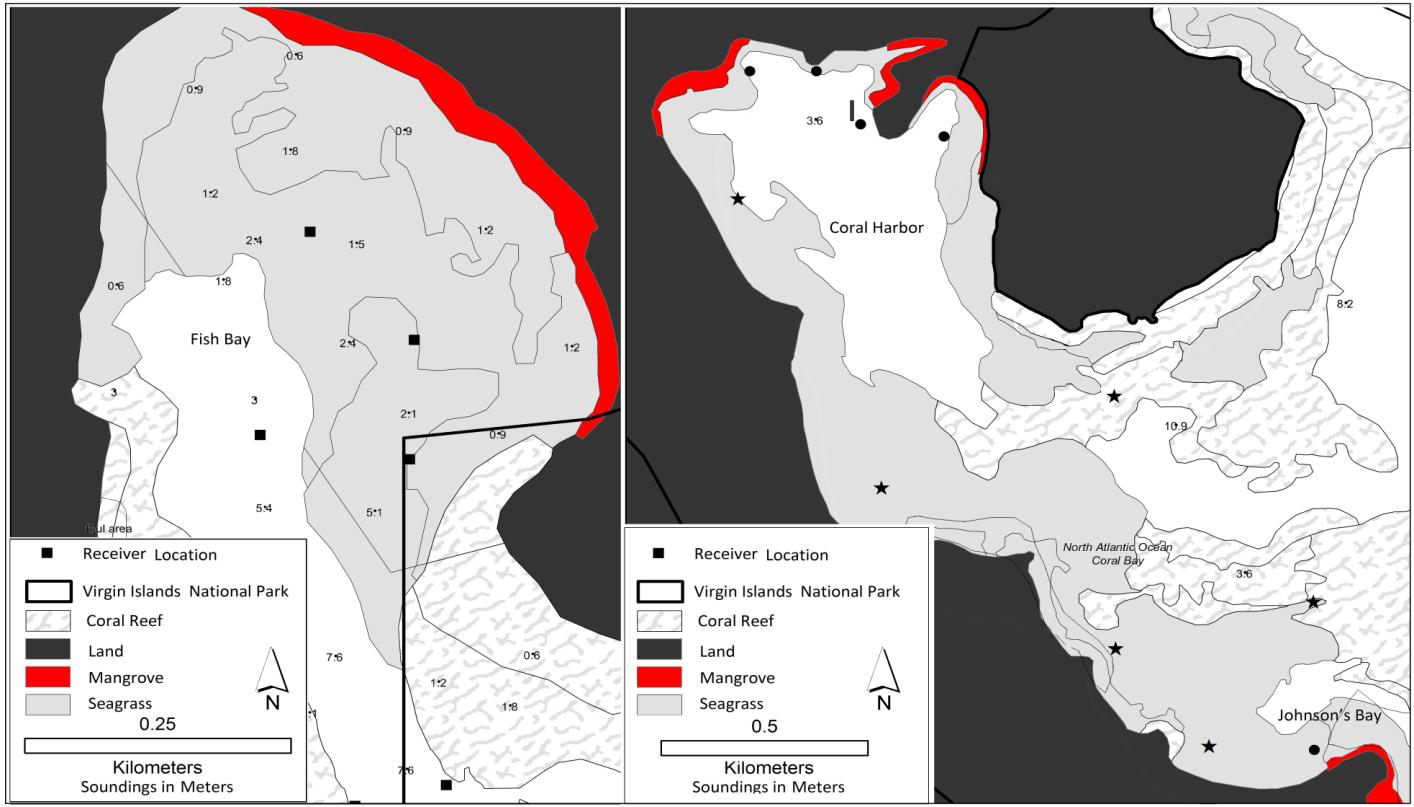
St. John is characterized by steep, rugged terrain, with two-thirds of St. John’s land area designated as a National Park. A large portion of St. John’s coastal waters lies within both the National Park and the Virgin Islands Coral Reef National Monument (Figure 1). St. John is home to several regionally important bays with seagrass, coral reefs, macroalgae beds, and mangrove habitats with high densities of neonate and juvenile blacktip and lemon sharks (DeAngelis et al. 2008). This research was conducted in two of these nurseries, Fish Bay and Coral Bay (Figure 2). Fish Bay, located on the south side of St. John at 18.318°N, 64.763°W, is approximately 0.34 km2 (Figure 1). The eastern side of Fish Bay, which is partially within the Virgin Islands National Park, is bordered with red ,black and white mangroves (Rhizophora mangle, Avicennia germinans and Laguncularia racemosa) and a shallow (<1.5m) seagrass (mainly Thalassia testudinum) bed. The latter thickens (50-70% cover) and expands to the west into deeper water (2-3m), giving way to sparse patches of macroalgae toward the southern mouth of the bay, which is bordered by fringing reefs (Figure 2). Neonatal and young-of-the-year (YOY) blacktip and lemon sharks have been surveyed in Fish Bay, which is thought to provide important nursery habitat for this species during the summer months with juvenile shark abundance peaking in June and July (DeAngelis 2006, DeAngelis et al. 2008). Anthropogenic impacts to Fish Bay include a relatively small number (<3) of live aboard vessels, a semi-developed shoreline, and large amounts of sediment run-off from the watershed (Gray et al. 2008).
On the eastern side of St. John (18.337°N, 64.763°W), Coral Bay is considerably larger that Fish Bay at 1.3 km2 (Figure 1). Coral Bay is partially bordered by mangroves along the northern shoreline and contains extensive shallow (<1.5m) seagrass beds (>70% coverage; T. testudinum; Figure 2). Seagrass beds extend south along the western side of Coral Bay. The southern end of Coral Bay contains a smaller embayment called Johnson’s Bay, which is characterized by shallow (<1m) seagrass beds and large patches of mangroves; during extreme low tides, much of Johnson’s Bay is exposed. The center of Coral Bay contains expansive soft bottom mud, macroalgae, sand, and patch reefs scattered at depths of 5-10m. Like Fish Bay, this embayment provides nursery habitat to juvenile and YOY blacktip and lemon sharks (DeAngelis 2006). Anthropogenic impacts in Coral Bay may exceed those in Fish Bay as there are consistently more live-aboard vessels (50+), more riparian development, considerable road runoff and sedimentation, and no protection from the National Park (Brooks et al. 2007, Smith et al. 2007, Gray et al. 2008)
Shark Sampling
Juvenile and YOY blacktip and lemon sharks were captured on benthic longlines set in Fish Bay and Coral Bay. Prior to Sea Grant funding, sampling was conducted from 2006-2009; these data were included in all subsequent analyses. With Sea Grant funding, additional sampling was conducted in 2010 (June 8-11th) and 2011 (January 14-16th, May 30th-June 2nd). The gear and sampling methods used to survey sharks in these areas were modeled after the methodology of the NMFS Cooperative Atlantic States Shark Pupping and Nursery Survey (Kohler and Turner 2001). The longline was comprised of a 305 m, 0.64 cm diameter braided nylon mainline with 25 gangions (100 cm of 0.64 cm diameter braided nylon line connected to 50 cm of 0.16 cm diameter stainless cable) attached to circle hooks (size 12/0) with barbs depressed (DeAngelis
2006). Hooks were baited with mackerel (Scomber scombrus), Atlantic bonito (Sarda sarda), little tunny (Euthynnus alletteratus), and barracuda (Sphyraena barracuda). The line was allowed to set between 0.5 to 1.0 hours to minimize mortality of young sharks. Sample sites were determined from catch data obtained from DeAngelis (2006) and DeAngelis et al. (2008)
Acoustic Tagging and Tracking
Individually coded transmitters (models V9-2L-R64K, V13-1L, Vemco Ltd., Nova Scotia) were surgically implanted into blacktip and lemon sharks using the methodology of Heupel and Heuter (2001). In addition, all sharks were fitted with NMFS conventional tags (Dalton-Henly, Oxford, England) through the dorsal fin after information was collected on sex and fork length.
Shark movements were monitored using passive acoustic receivers (Models VR2, VR2W, Vemco, Nova Scotia, Canada) placed in Fish Bay and Coral Bay as well as throughout the coastal waters of St. John (Figure 1). In the spring of 2006, eight receivers were deployed in Fish Bay by scientists with the National Marine Fisheries service (Galveston, Texas) and nine VR2 receivers were deployed in Lameshure Bay by the NOAA Biogeography Branch (Monaco et al. 2007). From 2007 to 2009, 25 additional receivers were deployed along the south coast of St. John by the NOAA Biogeography branch. In 2008, eight additional receivers were deployed within Coral Bay. In June 2010, 11 additional receivers were deployed in Coral Bay (Figure 1). Receivers were attached to the bottom with sand screws or suspended on a subsurface float. Range testing was performed on all receivers within Coral Bay and Fish Bay. Transmitters were activated and submerged at incremental distances along transects in at least three cardinal directions from each receiver. Locations derived using a Global Positioning System (Garmin 76, Olathe, KS, USA) were used to create detection ranges for each receiver. Ambient water temperature was measured and recorded every 30 minutes at each receiver location using temperature loggers (Hobo pendant and Hobo Pro temperature loggers, Onset Computer Corp , Bourne, MA).
Data Analyses
Blacktip and lemon shark movements and habitat use were quantified using three metrics: residency time, site fidelity, and home range. Residency time was calculated as the number of days each blacktip and lemon shark was present within each bay. Presence was defined as two or more detections in a 24 hour period. Residency time was log transformed to adjust for variance and normality. A One-Way ANOVA was used to compare bays and years and a linear regression assessed how size (fork length) influenced the length of residency
Site fidelity was quantified using a site fidelity index, which measures the level of site attachment to each receiver, or an array of receivers, within each bay (i.e. within the boundaries of Fish Bay or Coral Bay) (Meyer et al. 2007, Bellquist et al. 2008). Daily site fidelity is the number of half hours present in 24 hours as shown in the equation:
The site fidelity index ranges from 0.0, indicating no site attachment, to 1.0, indicating a high degree of site attachment (Bellquist et al. 2008). The mean of the daily site fidelity indices was calculated over the course of residency for each shark.
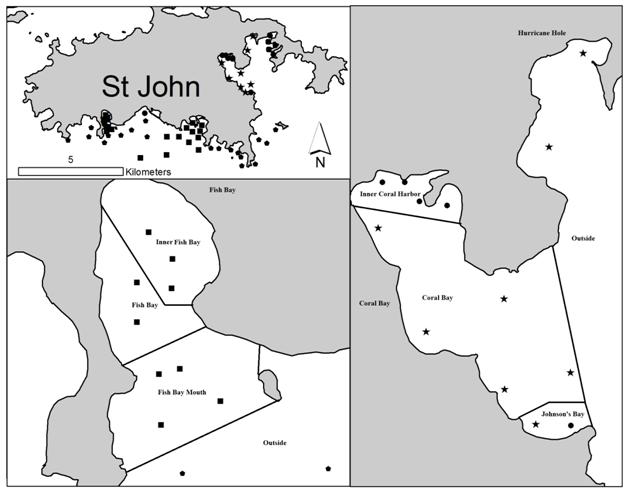
To assess areas of high use, adjacent receivers were grouped into zones representing similar areas within each bay (Figure 3). These zones were used to test the preference of sharks to a particular area or areas. The proportion of days that sharks were present in each zone was used to rank each zone in importance. Zones were compared using a Kruskal-Wallis one way ANOVA and a post hoc Tukeys test.
Given the irregular boundaries and the deployment pattern of the acoustic receivers in both bays, conventional estimators of animal home range and activity space, such as minimum convex polygons and kernel analyses, were not suitable for the robust estimation of these ecological parameters. Instead, a lattice-based density estimator (Barry & McIntyre 2011) was used to generate estimates of activity space for all tagged fish. This approach allows the home range to be quantified using the probability of moving within the defined area. As the receiver array changed over the course of this study, home range analysis was only conducted for years 2008 and 2009 in Fish Bay and 2010 and 2011 in Coral Bay when the array covered the majority of the study sites. Shark locations were calculated using a 30 minute center of activity equation:
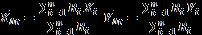
where n is the number of receivers in the array, Ri is the number of detection on the ith receiver during time t; Xi is the X geoposition of the ith receiver, and Yi is the Y coordinate of the ith receiver (Simpfendorfer et al. 2002). Activity spaces (50, 90, and 95%) were calculated daily, weekly, as well as for day and night periods for each shark. Traditional animal movement theory states that all movement does not constitute a home range, rather 95% and 99% probabilities are said to model normal daily activity space while 50% identifies core areas of site attachment; 100% will encompass all random movements of an animal (Ford & Krumme 1979, Anderson 1982). A linear regression for each daily home range was used to detect trends in habitat use over time. Mean day and night home range area estimates were compared using a student’s t test. The areas were square root transformed for analysis to adjust for normality and variance.
Results and Findings
Blacktip Sharks
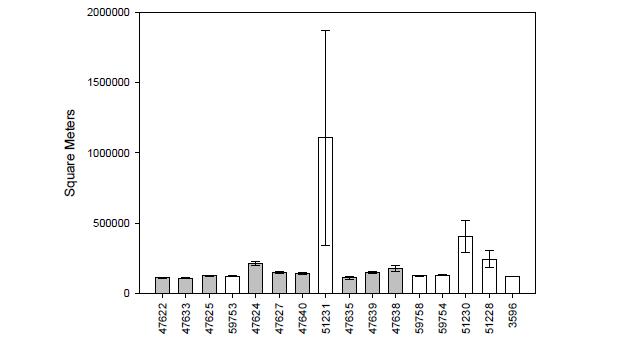
From 2006-2011, 94 YOY and juvenile blacktip sharks were captured in Coral Bay (n=50) and Fish Bay (n=44) ranging in size from 42.5-69.5 cm (mean = 50.9 cm, SD = 4.32 cm; Figure 4). Of these sharks, 52 were surgically implanted with acoustic transmitters (28=Fish Bay, 24 = Coral Bay: Table 2); two were identified as post-capture mortalities in Coral Bay and removed from subsequent analyses.
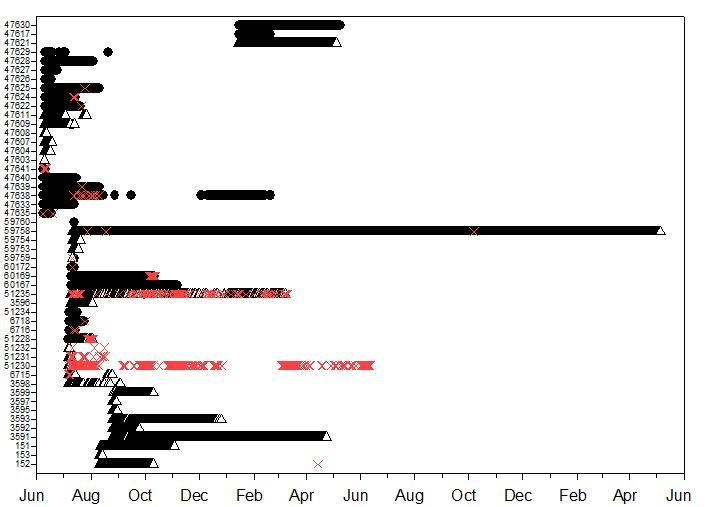
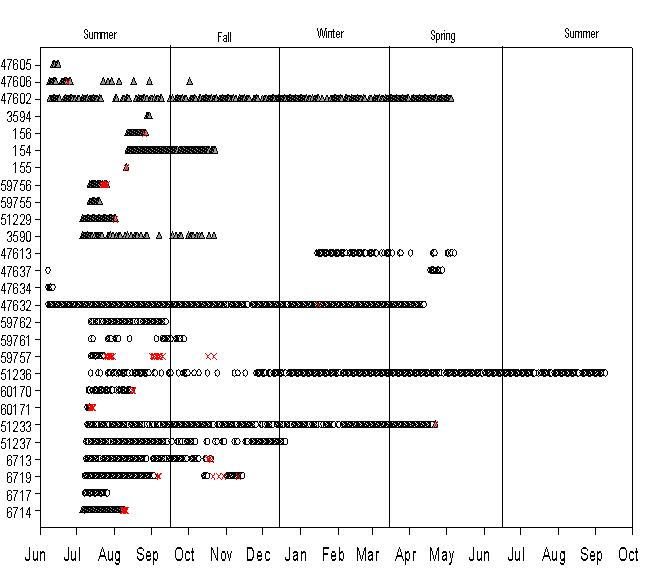
The blacktip sharks remained in Coral Bay and Fish Bay from 1-186 d (mean 53 d, median 36 d) and 2-663 d (mean 63.9 d, median 25.5 d), respectively (Table 1). Although some sharks spent a short amount of time within each bay, the majority resided in the bays until at least late August to October and some remained through the winter. Only one individual demonstrated multi-year residency within Fish Bay and was still present during the final data download, 663 days at liberty (Table 2, Figure 5). This individual (59758), which was recaptured
after 333 days at liberty, moved out of Fish Bay three times: twice into Reef Bay in 2009 and once into Lameshure Bay on October 7th 2010, the 3rd and final day of tropical storm Otto Residency did not differ significantly between bays (F=0.001, df=49, p=0.971) or across years (F=0.97, df =49, p=0.441; Figure 6). There was not a significant relationship between shark size and residency (linear regression, df=46, p=0.064).
Mean daily site fidelity indices were calculated for 50 acoustically tracked blacktip sharks from 2006-2011 (Table 1). The mean daily site fidelity index ranged from 0.069 ± 0.095 to 0.945 ± 0.028 (daily mean ± 95%CI) with an overall mean of 0.504 ± 0.0711 and median of 0.545 ± 0.0711 (median ±95% CI) (Table 1). Site fidelity for Coral Bay across all years was 0.500 ±0.112 (pooled daily mean ±95% CI, n=22) and Fish Bay across all years was 0.491±0.097 (pooled daily mean ±95% CI, n=28). There was no significant difference between bays (p=0.911, df=28, student’s t test). Twenty nine (58%) individuals tracked since 2006 had a site fidelity index > 0.50, indicating high site fidelity; the remaining 21 (42%) individuals spent less than 50% of their time within the nursery area (Figure 7).
Site preference in Coral Bay was consistent across years with the proportion of days found within Inner Coral Harbor being significantly higher when compared to all other zones: 2008 (F=69.2, df=8, mean=88.2%, p<0.001), 2009 (H=15.969, df=5, median=95.6%, p=0.007), 2010-2011 (H=40.173, df=3, median=77.8%, p=0.007; Figure 8). Site preference within Fish Bay was significantly higher than outside of Fish Bay, but varied within Fish bay between years (Figure 8). In 2006-2007, Inner Fish Bay was utilized significantly (H=26.24, df=3, median=82%, p<0.001) more than every other site, as was the case in 2008 (F=15.969, df=8, mean=50.5%, p<0.001) and 2009 (H=34.122, df=3, median 58.0%, p<0.001). In 2010-2011, Inner Fish Bay (median=58.9%), Fish Bay (median=29.2%) and Fish Bay Mouth (median= 22.9%) did not differ significantly, but were used significantly more than other sites monitored (H=28.142, df=5, median=58.9%, p=0.001).
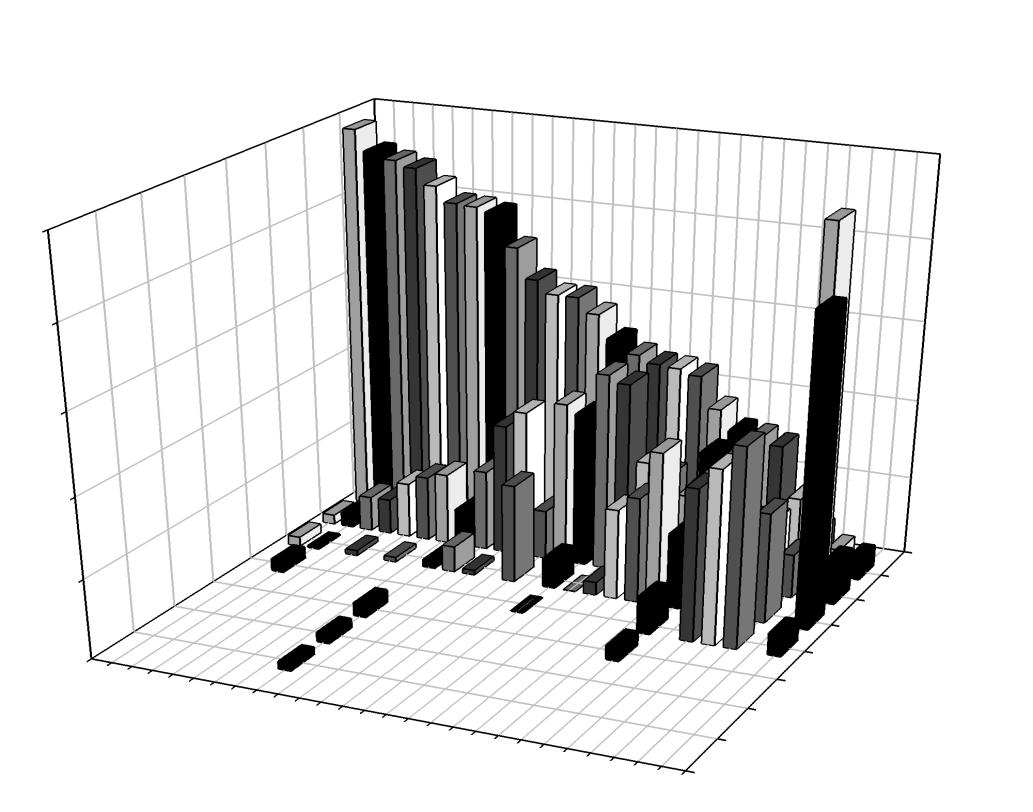
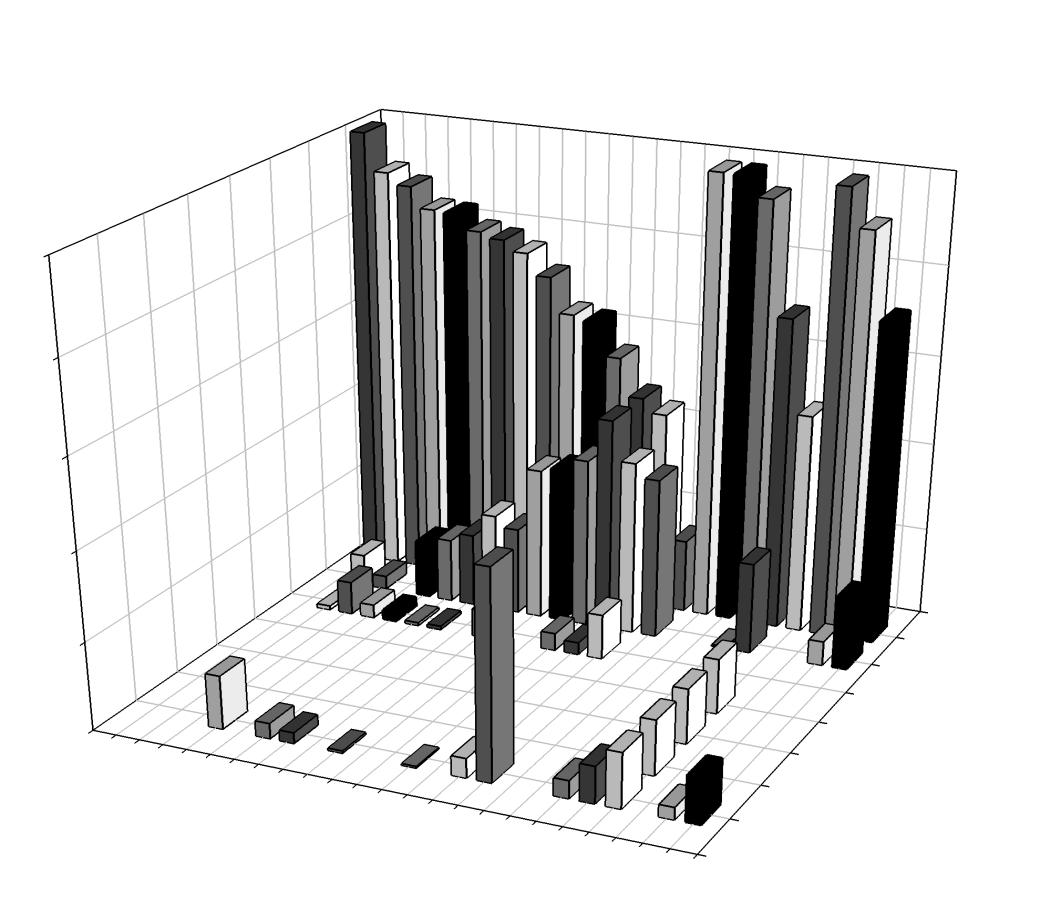
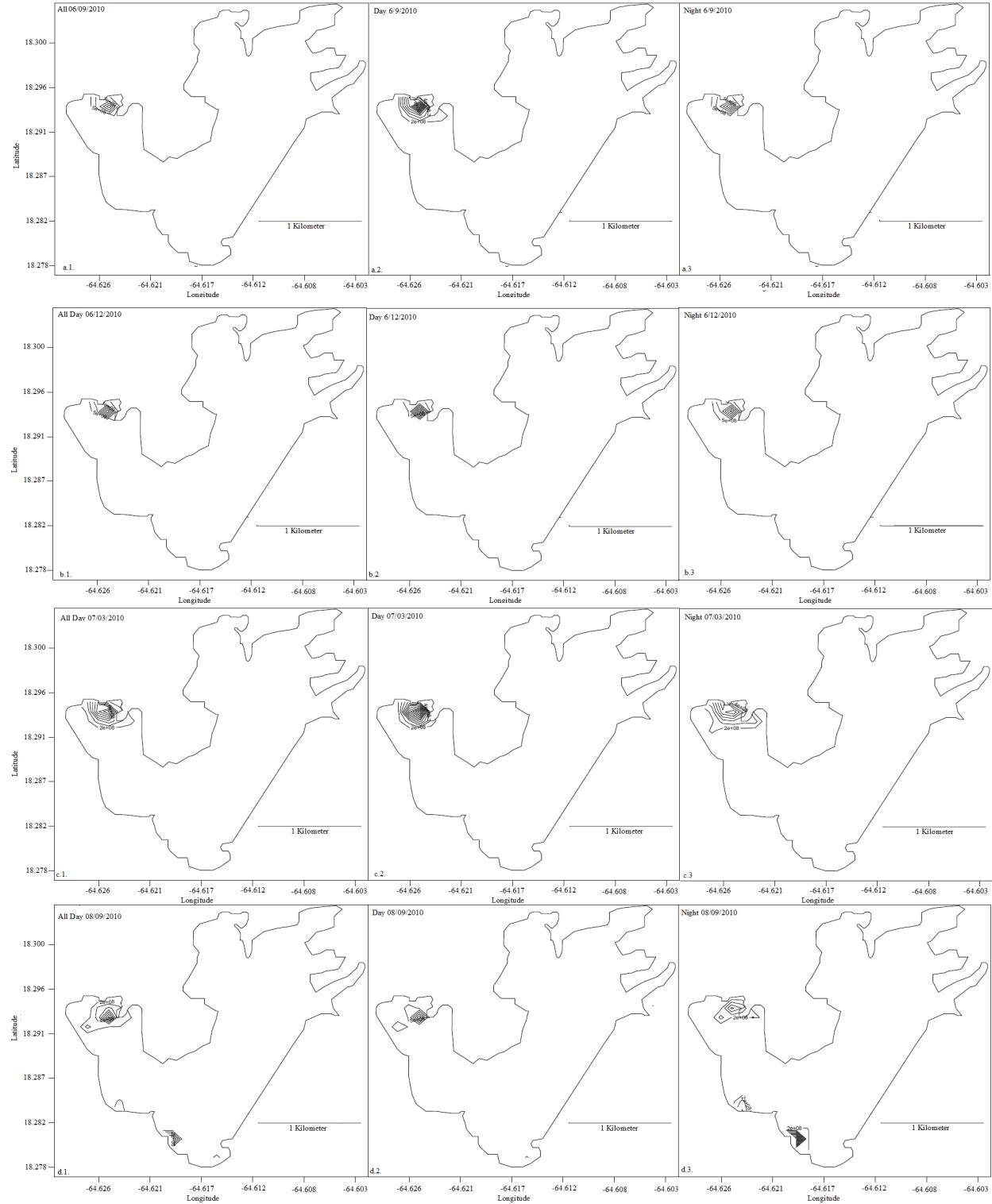
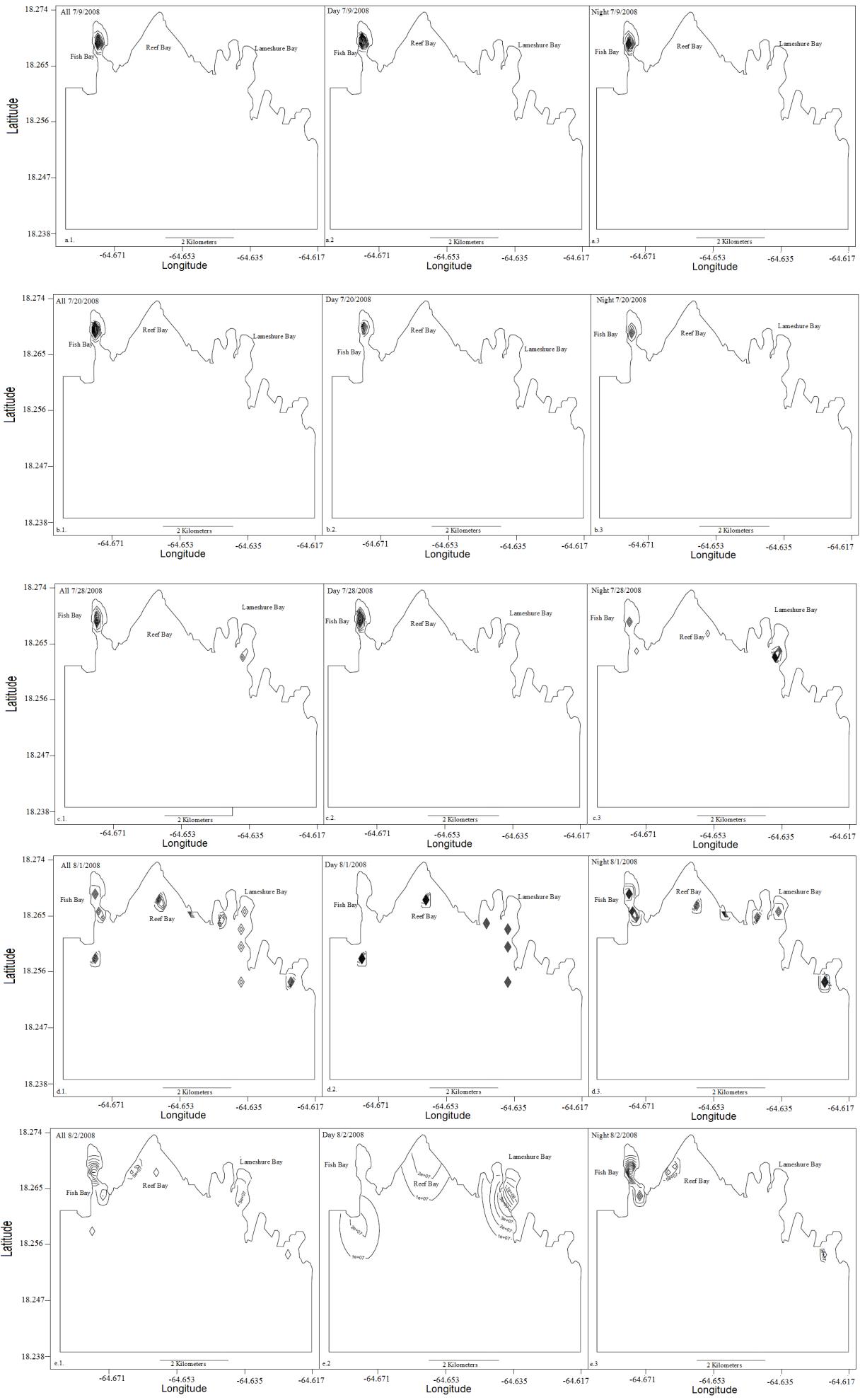
Daily home ranges varied between 0.06 km2 to 6.25 km2 (Figure 9) with individual means ranging from 0.107 km2 (± 0.0037 SE) to 1.106 km2 (± 0.7651 SE) and an overall mean of 0.221 km2 (± 0.0619 SE; Figure 10). No difference in mean home range size was found between bays (F=2.754, df = 14, p=0.121; Figure 11). Daily home range size increased in seven individuals with time in the bay, while two individuals had significantly decreased home range size while resident; the remaining individuals (n=7) demonstrated no significant relationship between home range size and residency (Table 3). Significantly larger mean home range was found during the day at both the 95 and 99% probability levels (Table 3). Upon visual inspection of the maps, movement into new areas often occurred at night with expansion into areas not visited before (Figures 12, 13). The mean home range for daytime activity is larger than night, but sharks that did foray, did so at night (Figures 14, 15).
Water temperature in Fish Bay varied between 24.2-31.7°C between June 2006 and May 2011. The water temperature in Coral Bay ranged 24.2-31.2°C between July 2008 and May 2011. Low temperatures occurred in February and high temperatures occurred in late summer from August to October (Figure 14).
From 2006- 2011, 53 lemon sharks were captured in Coral Bay (n=34, 48-70.3cm FL, 58.4 cm ± 5.7) and Fish Bay (n=19, 52-103cm FL, 61.2cm ± 13.5) (n, range, mean ± SD; Figure 15). Twenty seven of these sharks (Fish Bay: n=12, 52-81.2cm FL, 58.6cm ± 9.3; Coral Bay: n=15, 48-70.3cm FL, 59.3 cm ± 6.4) were implanted with acoustic transmitters and tracked for 1-
234 d (mean=40.3 d, median=18.5 d) within Fish Bay and 5-326 d (mean=93.6 d, median=33 d) within Coral Bay (Table 4, Figure 16). There was no significant differences in residency time between bays (F=3.521, df= 26, p=0.072; Figure 17) or between years (F=3.182, df =24, p=0.118; Figure 18). The majority of sharks vacated by early fall, although several individuals maintained long term residency up to 234 days in Fish Bay and 326 days in Coral Bay (Table 5). Mean daily site fidelity indices were calculated for 27 acoustically tracked lemon sharks from 2006-2011. The mean daily site fidelity index ranged from 0.096 ± 0.051 to 0.951 ± 0.065 (daily mean ± 95%CI) with an overall mean of 0.31 ± 0.079 (Figure 19). Lemon sharks exhibited spatial preferences for specific zones in each bay Those tagged in Coral Bay, for any year of the study, spent significantly more time in Johnson’s Bay than all other zones monitored (F=38.0, df=8, p<0.001; Figure 20). These sharks spent 68% ± 41of days monitored (mean % days ± SD) within Johnson’s Bay. This zone was followed by Inner Coral Harbor with 23% ± 39 of days monitored. Only one individual captured in Coral Bay moved to Fish Bay, where it spent only the last of 257 days.
Sharks tagged in Fish Bay showed similar affinity to specific regions of the bay. For any year measured, these sharks spent significantly more time (64 ± 25%) in Shallow Fish Bay than any other site monitored (F=4.737, df=11, p<0.001). In contrast, lemon sharks spent significantly less time in Deep Fish Bay (26 ± 20%) and at the mouth of Fish Bay (13 ± 18%). These results demonstrate that the sharks prefer the eastern, inner portion of Fish Bay, characterized by shallow seagrass beds adjacent to mangroves, as opposed to the deeper seagrass beds and the reef-lined mouth of the bay. Only one individual traveled to Coral Bay, where it spent only one of 37 monitored days.
Conclusions
Using acoustic telemetry, we gained valuable insights into the behavior of two common Caribbean shark species in important nursery habitat. Coral Bay and Fish Bay provide nursery habitat for YOY blacktip and lemon sharks during the summer months after birth, which is the most vulnerable stage of their lives. The bulk of these sharks either leave the bays throughout the fall and winter, are preyed upon by larger sharks, or both Based on our data, it appears both species use either Fish Bay or Coral Bay almost exclusively when compared to adjacent similar embayments.
Nursery areas for blacktip and lemon sharks have been identified along the east coast of the US, the Bahamas, and South America (Gruber et al. 1988, McCandless et al. 2002, Yokota & Lessa 2006, Freitas et al. 2009). Blacktip shark nurseries are typically large estuarine embayments along the east coast of the United States and along the Gulf of Mexico (Hueter 2002). For lemon sharks, long term nursery areas have been described in Bimini, where young lemons spend 2-3 years in an extensive seagrass lagoon (Chapman et al. 2009). In contrast, blacktip and lemon sharks in the U.S. Virgin Islands are utilizing small embayment’s that are significantly smaller than those along the east coast, Gulf of Mexico, and the Bahamas.
Despite these spatial differences, YOY blacktip and lemon sharks exhibit similar patterns of seasonal occupancy. However, the environmental cues or other factors that trigger emigration may not be the same. In other areas, the movements of blacktip and lemon sharks are triggered by a drop in water temperature to far below the minimum temperature in the Virgin Islands. Temperature does not appear to play a significant role in the USVI. In the current study, some blacktip and lemon sharks were detected throughout the year in Coral and Fish Bays. Therefore, it is possible that the seasonal abundance trends and residency time observed in this study are a
reflection of mortality (i.e. predation instead of emigration). In other nurseries, mortality has been estimated as high as 90% in juvenile blacktip sharks (Heupel & Simpfendorfer 2002). The extent to which natural mortality plays a role in the residency of juvenile blacktip and lemon sharks in the USVI is difficult to quantify.
It has been demonstrated that other biotic and abiotic factors contribute to residency in blacktip and lemon shark nurseries, such as seasonal resource availability, inter- and intraspecific competition, and/or anthropomorphic forces. Shark habitat utilization has been linked to the seasonal abundance of resources for multiple species in an Australian shark nursery (Simpfendorfer & Milward 1993). The short residency time of blacktip sharks in this nursery has been attributed to inter-specific competition during the same season (Simpfendorfer & Milward 1993). Habitat alteration resulted in shorter residency time for lemon sharks in a Bimini nursery (Bakker 2007, Jennings et al. 2008).
We also found that blacktip and lemon sharks show site fidelity to particular zones within each of these two nurseries. These areas are characterized by shallow seagrass beds adjacent to mangroves. High site fidelity has also been described in blacktip sharks in Terra Ceia Bay, FL (Heupel et al. 2004a) and lemon sharks in the Bahamas (Morrissey & Gruber 1993a). In the former, blacktip sharks were predominantly in the northern sections of Terra Ceia Bay where they aggregated during the day, but forayed at night in search of food (Heupel & Simpfendorfer 2005a). In the Bahamas, lemon sharks used only 5% of the total nursery available and showed great homing ability back to the nursery areas when relocated kilometers away (Morrissey & Gruber 1993b). Both species have been found to exhibit natal philopatry (Feldheim et al. 2002, Keeney et al. 2003, Hueter et al. 2004, Chapman et al. 2009). In the current study, we found that blacktip sharks predominantly utilize Inner Coral Harbor and Inner Fish Bay, which have shallow (<1.5 m) seagrass beds adjacent to mangroves. For lemon sharks, we found they primarily utilize Johnson’s Bay and Inner Fish Bay, which are also expansive seagrass beds adjacent to mangroves.
The extent to which these two species compete for resources in these nurseries is unknown. Habitat partitioning may be occurring in Coral Bay as the majority of lemon sharks reside on the southern end in Johnson’s Bay, while blacktip sharks primarily use Inner Coral Harbor to the north. Although our data in the current study were too coarse, DeAngelis et al (2008) used catch and active tracking data to suggest that nursery area partitioning occurs in Fish Bay as well
Coral and Fish Bays represent essential nursery habitat for blacktip and lemon sharks in the USVI with very little movement between the bays. We found similar utilization patterns in both bays with sharks spending the bulk of their time in shallow seagrass beds adjacent to mangroves. In particular, Johnson’s Bay, Inner Coral Harbor, and Inner Fish Bay are essential fish habitat for YOY blacktip and lemon sharks. There are indications that these areas are threatened by anthropogenic stresses (Brooks et al. 2007, Gray et al. 2008, Jennings et al. 2008). Habitat alteration and chronic stress have been shown to impact shark habitat use and mortality (Gruber et al. 2001, Bakker 2007). With natural mortality estimates as high as 90% (Heupel & Simpfendorfer 2002), additional mortality will impact population growth.
Objectives Accomplished (or Not)
The overall objective of the current study was to provide multi-year information on the movements and habitat utilization of sharks in USVI coastal shark nurseries using acoustic tracking. Specifically, our objectives were to:
(1) Determine the degree to which young sharks are utilizing near-shore coastal mangrove habitats, including how they partition the area between species and how long they remain in the area. This objective has been met. We quantified residency, site fidelity, and home range for YOY and juvenile blacktip and lemon sharks in two important shark nurseries on St. John, USVI. We examined the degree to which these two species exhibit habitat partitioning.
(2) Assess emigration rates. This objective has been met. In quantifying habitat use in Fish and Coral Bays by YOY blacktip and lemon sharks, we assessed residency and the subsequent rates at which they emigrate from these nurseries.
(3) Examine levels of connectivity between nursery habitats. This objective has been met. Both Fish Bay and Coral Bay have been identified as important nurseries for blacktip and lemon sharks. Despite extensive tagging and tracking efforts over six years, very few sharks utilized both embayments.
(4) Determine if these areas are isolated “pockets” of productivity. This objective has been met. Given the lack of movement exhibited by YOY blacktip and lemon sharks between multiple embayments along the coast of St. John and St. Thomas, USVI, Fish Bay and Coral Bay appear to be isolated pockets of productivity worthy of protection.
(5) Increase community awareness and education about the ecological importance of sharks to a healthy coral reef ecosystem. This objective has been met. Working with the Coral Bay Community Council, Maho Bay Camps and Concordia Eco-Resort, and the University of the Virgin Islands, we disseminated this information to the general public and local newspapers over the course of this study.
Project Impacts and Products
Publications
In November, 2011, UVI student Bryan Legare, who was funded by this study, completed and defended his master’s thesis, which was centered on this research. His thesis title is: Juvenile blacktip shark (Carcharhinus limbatus) and lemon shark (Negaprion brevirostris) movements within two nursery areas of St. John, United States Virgin Islands. The work contained in the thesis is currently being edited for formal publication in peer-reviewed journals. It is anticipated that this will result in at least two publications highlighting the fine and broadscale behavior of these two species in their USVI nurseries. Moreover, additional analyses are being conducted on these data and those collected in 2012; the results of this work will also be published.
Management
The results of this research will be disseminated to the appropriate local, regional, and federal management bodies so that measures to protect this essential habitat can be implemented. This information can provide local fisheries managers and other regulatory bodies with the tools necessary to establish effective conservation measures such as fishing closures, habitat
protection, additional marine protected areas, and limits on coastal development. This work has already proven useful in protecting juvenile sharks and shark nursery habitat because it was used by USVI coastal management agencies to decline development requests to build submerged docks and piers within Coral Bay, one of the shark nurseries documented by this study
The information collected by this study has also been beneficial to local stakeholders. The Coral Bay Community Council, a local group, has incorporated our data into their comprehensive environmental database for Coral Bay. These data are used to demonstrate the ecological value and importance of their bay.
Outreach
This project was also designed to increase community awareness and education about the ecological importance of sharks to a healthy coral reef ecosystem. Through public presentations (such as those listed below) and local media coverage, we actively increased public awareness of the importance of protecting, conserving, and managing nearshore essential fish habitat for sharks, as well as other species in the marine community By incorporating community volunteers into the project, we elevated the importance of coastal mangrove habitat, particularly those that act as essential fish habitat. Through continued communication with community organizations such as the Coral Bay Community Council and the St. Thomas Fisherman’s Association, a dialogue about the importance of sharks in the reef ecosystem continues. Since the preliminary stages of this project in the early 2000’s (DeAngelis, 2006), the perception of sharks and other apex predators in the USVI has begun to shift. We strongly believe that this project has been vital in changing public perception on the importance of near-shore sharks and their nursery habitat.
Recommendations
Research
This research provides the first detailed examination of the nursery habitat utilization by YOY blacktip and lemon sharks in St. John, USVI. Further research is warranted to identify and examine other important nurseries for these species in the region. To augment the current study, research is needed to examine the biotic and abiotic factors influencing juvenile shark habitat utilization. Studies should be conducted to examine the trophic ecology of these sharks in this important habitat and to further explore interspecific habitat and resource partitioning. The extent to which juvenile sharks are preyed upon by larger predators is important to assessing true emigration rates from these nurseries. Studies relevant to population connectivity (throughout the greater Caribbean and beyond) are warranted. This could be accomplished through genetic sampling and additional tagging. The former would also allow researchers to explore natal philopatry in these species.
Management
This research has identified essential nursery habitat for blacktip and lemon sharks within Coral Bay and Fish Bay on St. John, USVI. We provide evidence that these species utilize this habitat for the first several months of their lives, likely for protection during the period they are most vulnerable to predation. Some sharks remain in these embayments throughout the year. Our
tracking work shows that these nurseries are limited in size and represent isolated pockets of productivity. Therefore, these nurseries are critical to the health of blacktip and lemon shark populations in the region. Given the importance of this habitat, fisheries managers should implement measures to protect these areas from anthropogenic impacts. Moreover, territorial law should become compliant with federal regulations that prohibit the harvest of sharks less than 54 inches fork length. This would afford protection to juvenile blacktip and lemon sharks as well as numerous other shark species.
Bibliography
Anderson DJ (1982) The home range: A new nonparametric estimation technique. Ecology 63:103-112.
Bakker J (2007) Effects of the loss of mangrove nursery area on the home ranges of juvenile lemon sharks (Negaprion brevirostris). MS, University of Amsterdam.
Barry RP, McIntyre J (2011) Estimating animal densities and home range in regions with irregular boundaries and holes: A lattice-based alternative to the kernel density estimator. Ecol Model. 222:1666-1672.
Bascompte J, Melián CJ, Sala E (2005) Interaction strength combinations and the overfishing of a marine food web. Proc. Nat. Acad. Sci. 102:5443-5447.
Bellquist LF, Lowe CG, Caselle JE (2008) Fine-scale movement patterns, site fidelity, and habitat selection of ocean whitefish (Caulolatilus princeps). Fish Res 91:325-335
Brooks GR, Devine B, Larson RA, Rood BP (2007) Sedimentary development of coral bay, St. John, USVI: a shift from natural to anthropogenic influences. Carib J Sci 43:226-243
Caribbean-Fisheries-Management-Council (1998) Essential fish habitat generic amendment to the fishery management plan of the US Caribbean including a draft environmental assessment. San Juan, Peurto Rico
Chapman DD, Babcock EA, Gruber SH, Dibattista JDand others (2009) Long-term natal sitefidelity by immature lemon sharks (Negaprion brevirostris) at a subtropical island. Mol. Ecol. 18:3500-3507.
DeAngelis B (2006) The distribution of elasmobranchs in St. Thomas and St. John, United States Virgin Islands with an emphasis on shark nursery areas. M.S. Thesis, University of Rhode Island.
DeAngelis B, McCandless CK, NE., Recksiek C, Skomal GB (2008) First characterization of shark nursery habitat in the United States Virgin Islands: evidence of habitat partitioning by two shark species. Mar Ecol Prog Ser 358:257-271
Feldheim KA, Gruber SH, Ashley MV (2002) The breeding biology of lemon sharks at a tropical nursery lagoon. Proceedings: Biological Sciences 269:1655-1661.
Ford RG, Krumme DW (1979) The analysis of space use patterns. J Theo Biol 76:125-155
Freitas RHA, Rosa RS, Wetherbee BM, Gruber SH (2009) Population size and survivorship for juvenile lemon sharks (Negaprion brevirostris) on their nursery grounds at a marine protected area in Brazil. Neotrop. Ichthyol. 7:205-212.
Gray S, Gobbi K, Narwold N (2008) Comparison of sedimentation in bays and reefs below developed versus undeveloped watersheds on St. John, US Virgin Islands. 11th International Coral Reef Symposium, Ft. Lauderdale, Florida.
Gruber SH, de Marignac JRC, Hoenig JM (2001) Survival of juvenile lemon sharks at Bimini, Bahamas, estimated by mark–depletion experiments. Trans. Amer. Fish. Soc. 130:376-384.
Gruber SH, Nelson DR, Morrissey JF (1988) Patterns of activity and space utilization of lemon sharks, Negaprion brevirostris, in a shallow Bahamian lagoon. Bull. Mar. Sci. 43:61-76.
Heupel M, Hueter R (2002) Importance of prey density in relation to the movement patters of juvenile blacktip sharks (Carcharhinus limbatus) within a coastal nursery area. Mar Freshw Res. 53:543=550.
Heupel M, Simpfendorfer C (2005a) Quantitative analysis of aggregation behavior in juvenile blacktip sharks. Mar. Biol. 147:1239-1249.
Heupel M, Simpfendorfer C, Hueter R (2004a) Estimation of shark home range using passive acoustic monitoring techniques. Env Biol Fish 71:135-142
Heupel MR, Carlson JK, Simpfendorfer CA (2007) Shark nursery areas: concepts, definition, characterization, and assumptions. Mar Ecol Prog Ser 337:287-297
Heupel MR, Semmens JM, Hobday AJ (2006) Automated acoustic tracking of aquatic animals: scales, design and deployment of listening station arrays. Mar. Freshw. Res. 57:1-13.
Heupel MR, Simpfendorfer CA (2002) Estimation of mortality of juvenile blacktip sharks, Carcharhinus limbatus, within a nursery area using telemetry data. Can. J. Fish. Aquat. Sci. 59:624-632
Heupel MR, Simpfendorfer CA (2005b) Using acoustic monitoring to evaluate MPA’s for shark nursery areas: the importance of long-term data. Mar. Tech. Soc. J. 39:10-18.
Heupel MR, Simpfendorfer CA, Hueter RE (2004b) Estimation of shark home ranges using passive monitoring techniques. Env. Biol. Fish. 71:135-142.
Holden MJ (1974) Problems in the rational exploitation of elasmobranch populations and some suggested solutions. In: Hardin-Jones FR (ed) Sea fisheries research. John Wiley and Sons, New York, NY. 117–137
Jennings D, Gruber S, Franks B, Kessel S, Robertson A (2008) Effects of large-scale anthropogenic development on juvenile lemon shark (Negaprion brevirostris) populations of Bimini, Bahamas. Env. Biol. Fish. 83:369-377.
Keeney DB, Heupel M, Hueter RE, Heist EJ (2003) Genetic heterogeneity among blacktip sharks, Carcharhinus limbatus, continental nurseries along the U.S. Atlantic and Gulf of Mexico. Mar. Biol. 143:1039-1046.
Kohler NE, Turner PA (2001) Shark tagging: A review of conventional methods and studies. Env. Biol. Fish. 60:191-224.
McCandless C, Pratt Jr HL, Kohler N (2002) Shark nursery grounds of the Gulf of Mexico and the east coast waters of the United States: An overview, USDOC/NOAA/NMFS/NEFSC, Narragansett RI.
Meyer CG, Holland KN, Papastamatiou YP (2007) Seasonal and diel movements of giant trevally, Caranx ignobilis, at remote Hawaiian atolls: implications for the design of marine protected areas. Mar. Ecol. Prog. Ser. 333:13-25.
Monaco ME, Friedlander AM, Caldow C, Christensen JD (2007) Characterizing reef fish populations and habitats within and outside the US Virgin Islands coral reef national monument: a lesson in marine protected area design. Fish Manag Ecol 14:33-40
Morrissey J, Gruber S (1993a) Home range of juvenile lemon sharks. Copeia 2:425-434
Morrissey JF, Gruber SH (1993b) Habitat selection by juvenile lemon sharks, Negaprion brevirostris. Env. Biol Fish 38:311-319.
Simpfendorfer CA, Heupel MR, Hueter RE (2002) Estimation of short-term centers of activity from an array of omnidirectional hydrophones and its use in studying animal movements. Can. J. Fish Aquat Sci 59:23-32
Simpfendorfer CA, Milward NE (1993) Utilization of a tropical bay as a nursery area by sharks of the families Carcharhinidae and Sphyrnidae. Env. Biol. Fish. 37:337-345.
Smith TB, Blondeau J, Taylor MG, Nemeth R, Calnan JM, Tyner E (2007) Continuation and expansion of the territorial biological monitoring program. Center for Marine and Environmental Studies, University of the Virgin Islands.
Springer S (1967) Social organization of shark populations. In: Gilbert P, Mathewson R, Rall D (eds) Sharks, skates and rays. John Hopkins press, Balitimore, MD, p 149-174.
Yokota L, Lessa R (2006) A nursery area for sharks and rays in northeastern Brazil. Env Biol Fish. 75:349-360.
Appendices
Table 1. Biological and acoustic data for blacktip sharks (C. limbatus) from 2006 to 2011 (n=50); ID= transmitter number, FL= fork length (cm), Residency time = days present within Coral Bay or Fish Bay, Site Fidelity index = mean daily site attachment as a
spent within each bay (mean ±SD)
Table 2. Residency of acoustically tagged blacktip sharks within Coral Bay and Fish Bay (pooled) from 2006-2011 (n=50). Days N (%) 0-7 13 (26%) 8-30 13 (26%) 31-60 11 (22%) 61-99 5 (10%) 100-365 7 (14%) 365 + 1 (2%)
N
Table 3. Home range summary data with 95 and 99% mean daily, day, and night home range estimates (mean km2±95%CI; n=16). Results of students T-test indicated mean differences between day and night activity space at both 95% and 99% estimates. Linear regression shows trends in activity space over the course of residency (Slope trend (+/-), pValue, r2
Table 4. Biological and acoustic data for lemon sharks (N. brevirostris) from 2006 to 2011 (n=27); ID= transmitter number, FL= fork length (cm), Residency time = days present within Coral Bay or Fish Bay, Site Fidelity index = mean daily site attachment as a percent time spent within each bay (mean ±SD).
Table 5. Residency of acoustically tagged lemon sharks in Coral Bay and Fish Bay (pooled) from 2006-2011 (n=27).
Days N
0-7 5 (19%) 0-7 8-30. 9 (33%) 8-30. 31-60 5 (19%) 31-60 61-90 2 (7%) 61-90 90-180 2 (7%) 90-180 >180 4 (15%) >180