
Executive Summary
Project Title: Character and Timing of a Reef Give-Up Event on the Southwest Puerto Rico Shelf
Date: 23 April 2012
Project Number: R-111-1-08
Investigators:Clark Sherman, UPRM Department of Marine Sciences
Wilson Ramírez, UPRM Department of Geology
Project Period: 1 February 2008 – 30 April 2012
Summary of Impacts and Contributions
Objectives
Objective 1 – Document the Chronology and Paleoecologic Character of Reef Accretion
This objective has been met. Five separate outer-shelf hummock reefs were cored to determine their age and accretion history. Radiocarbon ages of corals indicate that the upper 1-2 m of these reefs are recent (<1200 yr BP; yr BP = years before present) and actively accreting upward. This is contrary to our original hypothesis that these reefs were largely relict features formed prior to ~6 ka (ka = “kilo-annum”, thousands of years before present). The primary frame builder of these reefs is the Montastraea annularis species complex, with secondary contributions from Acropora cervicornis and Acropora palmata. These are all common reef builders in the Caribbean, though the depth of the reefs (~14-19 m) is somewhat deeper than typically expected for A. palmata (Aronson et al., 2002; Montaggioni and Braithwaite, 2009) Internally, the reefs are poorly cemented and unconsolidated coral rubble is a major constituent within the framework.
Objective 2 – Establish Coral-Based Oxygen and Carbon Isotopic Records of Paleoenvironmental Conditions
This objective has not yet been met. Meaningful coral-based stable isotope records typically require larger corals with massive growth morphology so that records of several years to decades may be established. Isolated measurements are generally not as useful. While the M. annularis complex is the primary frame builder in these reefs, coral rubble is also a major component. Thus, recovery of large, intact massive corals was problematic due to the nature of the reefs. Additionally, larger corals were often heavily bioeroded disrupting the resultant record. Still, over the course of the coring several larger, massive coral samples suitable for paleoenvironmental work were eventually recovered. These samples range in age from ~200 to 1200 yr BP and are now being analyzed for Sr/Ca and stable oxygen isotope composition to establish seasonal paleotemperature records.
Objective 3 – Correlate Reef-Accretion and Isotopic Records with Other Paleoclimatic Records
Thus far, this objective cannot be met due to the nature of the record recovered. An overriding hypothesis of the stated objectives was that the outer shelf reefs to be studied were relict give-up reefs. This was based on previous coring studies in the area (Hubbard et al., 1997). Our research shows that the reefs are not relict, but, rather, are recent features that are actively accreting. Their recent upward accretion is consistent with the availability of accommodation space and minimal sea-level changes over the last millennia. Upon completion of Objective 2 described above, seasonal paleotemperature records from recovered corals will allow for a
comparison of seasonal temperature patterns from intervals prior to anthropogenic influence with modern instrumental records.
Advancement of the Field
Based on a cross-shelf coring study, Hubbard et al. (1997, 2008) proposed a two-phase model of Holocene reef accretion on the insular shelf off La Parguera, southwest Puerto Rico (the Parguera Shelf). Phase-I reef accretion began at the shelf edge ~10 ka and lasted until ~6.56.8 ka when, quite inexplicably, vertical accretion by these reefs stopped. Phase-II reef accretion began along the inner and middle shelf ~8 ka and has continued up to the present. We had hypothesized that a series of submerged, oval-shaped, low-relief reefs (herein referred to as hummock reefs) along the outer shelf were part of the Phase-I shelf-edge reef accretion. However, coring and radiocarbon dating of the hummock reefs does not support this hypothesis. Rather, the new data suggest a third phase of reef accretion starting along the outer shelf sometime after ~4 ka and continuing up through the present. Additionally, this study identifies an area of very active vertical reef accretion that may represent a new locus of reef growth on the Parguera Shelf.
Problems Encountered
Initial problems were related to unforeseen delays in procuring necessary equipment (specifically the coring system) due to the UPRM purchasing process. This delay did, however, allow us the opportunity for a short complementary study of an inner-shelf shoal (see Project Narrative). Once coring equipment was obtained and field work began, the only problems were related to the nature of the recovered record. The reefs were not the age that was hypothesized and much of the material recovered was not suitable for isotopic analyses. This required altering the objectives somewhat to accommodate the unforeseen nature of the reef record. Still, this work is providing important new information on reef accretion patterns across the Parguera Shelf and altering our view of preexisting models of Holocene reef accretion history in the area.
Other Impacts and Products
Students Supported
Student Role
Francisco Soto franciscoj_soto@yahoo.com
Graduate
Duane Sanabria duane.sanabria@upr.edu
Graduate
Marianela Mercado marianela.mercado@upr.edu
Graduate
Diver and field assistant
Diver and field assistant
Lab assistant and Masters student
$1200 Aug 2009-Dec 2010
$1000 Jan 2011-Jun 2011
$1375 Jan 2012-Apr2012
MS: Reef accretion during the last 700 years in the outer insular shelf of southwest Puerto Rico (May 2012)
Edgardo Quiñones ejqc_15@hotmail.com
Graduate
Lab assistant and Masters student
MS: Using corals from southwest Puerto Rico to determine variations in sea surface temperature during the last millennium (In progress))
Presentations & Publications
Presentations
Sherman, C., Ramírez, W. and Mercado, M., Reef accretion on the outer shelf of southwest Puerto Rico. 12th International Coral Reef Symposium, Cairns, Queensland, Australia, July 9-13, 2012.
Sherman, C., Ramírez, W. and Mercado, M., Submerged outer-shelf hummock reefs, southwest Puerto Rico: Record of a regional reef give-up event? 35th Scientific Meeting of the Association of Marine Laboratories of the Caribbean, San Jose, Costa Rica, May 23-27, 2011.
Sherman, C. and Ramírez, W., Character and Timing of a Reef Give-Up Event on the Southwest Puerto Rico Shelf. Simposio de investigación, University of Puerto Rico Sea Grant Program, Mayagüez, Puerto Rico, October 29, 2010.
Sherman, C. and Ramírez, W., Character and Timing of a Reef Give-Up Event on the Southwest Puerto Rico Shelf. Conversatorio Marino, University of Puerto Rico Sea Grant Program, Mayagüez, Puerto Rico, April 20, 2010
Manuscripts in preparation
Sherman C. et al., In prep. Intrinsic and extrinsic controls on the late Holocene depositional history of an inner-shelf mangrove-capped shoal, southwest Puerto Rico
Sherman, C., Mercado, M. and Ramírez, W., In prep. Recent reef accretion on the outer insular shelf of southwest Puerto Rico.
New Extramural Funds
Building upon this study, Sherman has submitted a proposal to National Geographic Society to continue and expand upon Holocene reef accretion studies in Puerto Rico.
Title: Structure and development of mesophotic coral reefs, Southwest Puerto Rico (PendingPre-Proposal has been approved, invited to submit full proposal)
PI: Clark Sherman
Requested Amount: $20,000
Agency: National Geographic Society
Proposed Duration: 12 months
Report Narrative
Statement of Problem
Coral reefs represent a critically important ecologic, economic, social, and recreational; resource. However, reefs worldwide are in a state of serious decline with an estimated 30% severely damaged and the possibility of 60% being lost by the year 2030 (Hughes, 1994; Hughes et al., 2003; Wolanski et al., 2003). The causes of this recent decline are a result of both natural factors such as storms, bleaching, and disease and anthropogenic influences including active mining of reef limestone, overfishing and destructive fishing practices, coastal land-use changes leading to increased terrestrial runoff, and anthropogenically induced global-warming effects (ibid.). A persistent problem in understanding the recent decline in reefs and establishing effective management strategies is differentiating between natural variability in reef systems and anthropogenically induced change. Coring of reefs and examining the recent fossil record of reef accretion provides an important geological backdrop against which recent coral-reef decline may be assessed (e.g., Aronson et al., 2002; Hubbard et al., 2005). In particular, examining the Holocene record can help us to establish long-term patterns of variability in reef accretion and identify periods of reef “give-up” and rapid ecological turnover. These natural trends and events can then be compared to the recent decline in reefs to help us better understand the relative magnitude of the recent decline and its likely primary causes.
The southwest shelf of Puerto Rico represents an ideal setting to study reef accretion histories. The broad shelf has been conducive to reef accretion throughout the Holocene and has been an area of intense geologic and ecologic study for several decades. Coring studies by Hubbard et al. (1997) showed that reef accretion began at the shelf edge ~10 ka (ka = “kiloannum”, thousands of years before present) as the shelf was flooded by the deglacial rise in sea level. These reefs were dominated by Acropora palmata and actively accreted upward until ~6.5-6.8 ka when, quite inexplicably, vertical accretion by A palmata stopped, i.e., the reefs “gave up” (cf. Neumann and Macintyre, 1985). Although these shelf-edge areas have abundant live coral cover today, this represents just a thin veneer over a substrate that that is 6 to 7 thousand years old. In addition, present-day cover is dominated by massive and soft corals in contrast to the underlying substrate dominated by A. palmata. Accretion of inner and mid-shelf reefs began ~8 ka as shallower portions of the shelf were flooded. However, in contrast to the shelf-edge reefs, these shallower reefs were dominated by massive corals, only shifting to branching acroporids much later. In addition, these (massive-coral) reefs continued to continuously accrete upward to their present position, while vertical accretion at the shelf edge essentially ended more than 6,000 years ago. While rapid sea-level rise is often cited as a cause for such a landward shift in reef position, i.e., “backstepping,” widely accepted sea-level curves for the Caribbean do not show any rapid sea-level rise events at this time (e.g., Toscano and Macintyre, 2003). Thus, the causes of the give-up of Acropora palmata shelf-edge reefs remain problematic. A nearly identical history of cross-shelf reef accretion and give-up is also evident in St. Croix (Hubbard et al., 2005).
Along the outer shelf of southwest Puerto Rico are a series of submerged, oval-shaped, low-relief reefs that rise several meters above the surrounding flat, sandy substrate in water depths of ~15-20 m (Fig. 1). For sake of this discussion, these reefs will herein be referred to as outer-shelf hummock reefs. These features form the dominant habitat of the outer shelf (Prada
Triana, 2002). Water depth over the tops of the outer-shelf hummock reefs (~14 m) is similar to that over the shelf-edge reefs cored by Hubbard et al. (1997) suggesting that the hummock reefs succumbed to the same regional give-up event that affected shelf-edge reefs about 6,500 years ago. The uniform geomorphology of the hummock reefs (cf. Prada Triana, 2002) suggests that they all may have succumbed to the same give-up event, indicating that the give-up event was much broader in scale than previously reported and affected most or all of the outer shelf, not just the shelf-edge reefs. The causes of this event are unknown. A coring investigation of the outer-shelf hummock reefs builds upon the work of Hubbard et al. (1997) and provides important new information on patterns of Holocene reef accretion on the southwest Puerto Rico shelf.

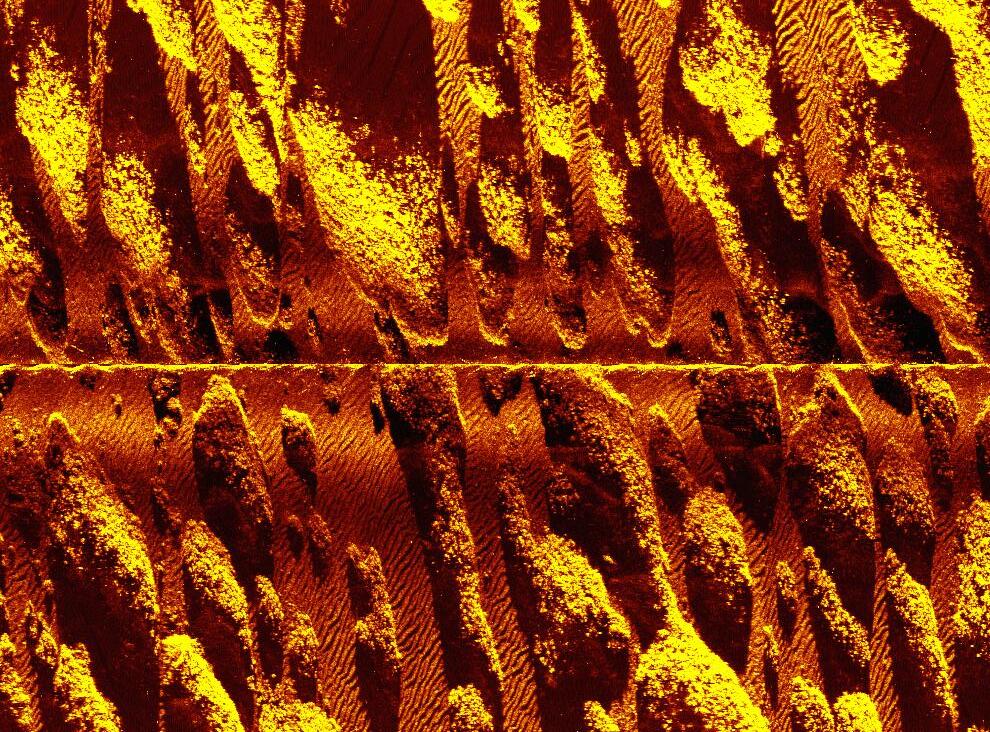
Figure 1. A) Location map showing Puerto Rico. Black square shows location of study area. B) Side-scan sonar image of outer-shelf hummock reefs. Note similar geomorphology and alignment of reefs suggesting that they have similar accretion histories. These reefs form the dominant habitat of the outer shelf. (Side-scan sonar image after Prada Triana, 2002).
Methods
To determine reef-accretion and paleoecologic histories, the outer-shelf hummock reefs were sampled via a diver-operated Tech 2000 submersible hydraulic drilling system acquired
with funds from this grant. Cores were examined on macroscopic and microscopic/petrographic scales using a facies approach. Thin sections were made of representative facies and of coral samples selected for dating. Paleoenvironmental reconstructions were made on the basis of facies’ sedimentologic and paleoecologic characteristics, including the specific makeup of primary and secondary framework assemblages, i.e., corals and coralline algae, and the taphonomic character of the framework components imparted by encrustation, cementation, and physical and biological erosion (cf. Aronson et al., 2002; Blanchon and Perry, 2004; Cabioch et al., 1999; Hubbard et al., 2005). Mineralogy of samples was determined by x-ray diffraction to aid in characterizing reef facies and to screen samples for radiometric dating. Stable oxygen isotopic composition of coral samples are being determined to serve as proxy paleotemperature indicators. Stable isotope work is still in progress. X-ray diffraction and isotopic measurements were conducted at facilities of the UPRM Department of Geology. Numerical ages of fossil corals were determined using radiocarbon (14C) methods at the National Ocean Sciences
Accelerator Mass Spectrometry Facility at Woods Hole Oceanographic Institution. Radiocarbon ages were converted to calendar years BP using the INTCAL09/MARINE09 (Calib 6.0.1) calibration curve (Reimer et al., 2009) and a reservoir deviation of 28 ±4 (ΔR).
During the early part of this study, a ship-grounding on an inner-shelf shoal close to the Magueyes Marine Laboratories, allowed for the opportunity to examine the internal structure, stratigraphy and depositional history of this feature. Techniques described above were employed to accomplish this.
Results and Findings
Outer-shelf hummock reefs
Five separate outer-shelf hummock reefs were cored to determine their age and accretion history. Cores were recovered from the tops of the reefs at depths of ~14 m. Core lengths ranged from ~1.3 to 1.8 m. The primary frame builder of these reefs is the Montastraea annularis species complex, with secondary contributions from Acropora cervicornis and Acropora palmata. These are all common reef builders in Puerto Rico and the Caribbean, though the depth of the reefs (~14-19 m) is somewhat deeper than typically expected for A. palmata (Aronson et al., 2002; Montaggioni and Braithwaite, 2009). The Montastraea annularis species complex consists of M. annularis, M. faveolata and M. franksi (Budd and Klaus, 2001; Knowlton et al., 1992). These three species are not differentiated here as they a very difficult to differentiate from one another in core samples. Additionally, there is some debate as to whether they actually represent separate species (cf. Knowlton et al., 1997). Within the cores, the M annularis complex includes in situ growth-position corals that form primary framework. Acropora cervicornis and Acropora palmata occur only as secondary framework, i.e., coral clasts bound together by encrusting organisms, and loose coral rubble. The abundance of secondary framework and loose coral rubble is expected given the reefs are composed of a mixture of massive and branching coral growth forms (Hubbard et al., 2001). While A. cervicornis occurs primarily as loose rubble in the cores, its abundance is indicative of its importance as a reef builder in this setting. The M. annularis complex and A. cervicornis are both common on these reefs today. In contrast, no living A. palmata colonies were observed. Standing dead and toppled colonies of A. palmata are common on some of these reefs and A. palmata occurs in most of the cores. This indicates that over the last millennia, conditions were once favorable for A. palmata growth at these sites. Given the reefs occur at depths slightly
beyond the typical range of A. palmata, these corals may have been especially susceptible to disease and anthropogenic influences resulting in their recent disappearance. Internally, the reefs are poorly cemented and unconsolidated coral rubble is a major constituent within the framework.
Radiocarbon ages of corals (Table 1) indicate that the upper 1-2 m of these reefs are recent (<1200 yr BP; yr BP = years before present) and actively accreting upward. Accretion rates for these reefs, calculated on the basis of radiocarbon ages and recovered cores, range from ~1-3 m/ka. Which are similar to average accretion rates in the Caribbean (Hubbard et al., 2008). Ages of the reefs are contrary to our original hypothesis that these reefs were largely relict features formed prior to ~6 ka. Based on a cross-shelf coring study, Hubbard et al. (Hubbard et al., 2008; Hubbard et al., 1997) proposed a two-phase model of Holocene reef accretion on the insular shelf off La Parguera, southwest Puerto Rico (the Parguera Shelf). Phase-I reef accretion began at the shelf edge ~10 ka and lasted until ~6.5-6.8 ka when, quite inexplicably, vertical accretion by these reefs stopped. Phase-II reef accretion began along the inner and middle shelf ~8 ka and has continued up to the present. We had hypothesized that a series of submerged, oval-shaped, low-relief reefs (hummock reefs) along the outer shelf were part of the Phase-I shelf-edge reef accretion. However, coring and radiocarbon dating of the hummock reefs does not support this hypothesis. Rather, the new data suggest a third more recent phase of reef accretion. Based on radiocarbon ages, accretion rates and the morphology of the hummock reefs, we suggest that this Phase-III reef accretion started along the outer shelf sometime after ~4 ka and continues up through the present. It is, therefore, possible that shelf-edge reef accretion did not cease 6,000 years ago. Rather, the locus of active shelf-edge accretion may have retrograded landward since 6 ka. Importantly, this study identifies an area of very active vertical reef accretion that may represent a new locus of reef growth on the Parguera Shelf.
Table 1. Radiocarbon ages of corals from outer-shelf hummock reefs.
LP1-S1
LP1-S2
LP3A-S1
LP3A-S2
LP4-S1
LP4-S2 Massive coral 148-149 1670 ±25 1140-1270 1210
LP5-S1 Massive coral 147-151 1650 ±25 1110-1260 1190
1 Years BP
Inner-shelf mangrove shoal
In the early summer of 2007, a 39-m motor vessel ran aground on a small, inner-shelf mangrove-capped shoal adjacent to the Magueyes Marine Laboratories. The grounding carved a v-shaped scar into the landward margin of the shoal that extended from the seafloor at ~2.75 m water depth down to ~6 m water depth. This fresh exposure allowed for an examination of the internal structure and stratigraphy of the shoal. The seafloor adjacent to the scar is a shallow seagrass bed with carbonate mud and fine sand. From the seafloor downward, three stratigraphic units were identified. Unit I extends from the seafloor at -2.75 m down to -4 m. This unit consists of a homogeneous carbonate mud with abundant bivalves. Unit II extends from -4 to4.5 m. This unit consists of small, delicately branching corals, e.g., Porites porites and Acropora cervicornis, in a muddy sand matrix. The corals display good preservation and many are in growth position. Unit III extends from -4.5 to -6 m. This unit consists of a mixture of massive and branching corals including the M annularis complex, Porites astreoides and exceptionally large branches of A. cervicornis in a muddy sand matrix. The corals show a range of preservation and larger massive corals display ragged margin growth morphologies indicative of growth in a high-sedimentation environment (James and Bourque, 1992).
Radiocarbon ages of corals (Table 2) indicate that Unit III was accreting upward from at least ~4700 to 4300 yr BP. Unit II was deposited ~3200 yr BP. Following this, coral growth abruptly ceased and Unit I was deposited. We propose that from ~5000 to 3000 yr BP conditions at this inner-shelf site were conducive to coral growth and upward reef accretion. At this same time reefs further offshore were also actively accreting upward (Hubbard et al., 1997). The upward accretion of offshore reefs would have progressively changed the inner-shelf environment to a more restricted setting with lower wave energy. The restricted, lower energy setting resulted in less flushing more accumulation of sediments, both in situ produced carbonates sediments and terrigenous material from the adjacent coastline. This change in conditions led to a cessation of coral growth and transition from a reef to a seagrass-mangrove capped shoal. These results document an example of reef give-up and ecological turnover as a result of natural intrinsic and extrinsic factors and highlight the delicate nature of reef ecosystems even under pristine, pre-anthropogenic conditions.
Table 2. Radiocarbon ages of corals from an inner-shelf mangrove shoal.
1 Years BP
Objectives
Objective 1 – Document the Chronology and Paleoecologic Character of Reef Accretion
This objective has been met. Five separate outer-shelf hummock reefs were cored to determine their age and accretion history. Radiocarbon ages of corals indicate that the upper 1-2 m of these reefs are recent (<1200 yr BP; yr BP = years before present) and actively accreting upward. This is contrary to our original hypothesis that these reefs were largely relict features formed prior to ~6 ka (ka = “kilo-annum”, thousands of years before present). The primary frame builder of these reefs is the Montastraea annularis species complex, with secondary contributions from Acropora cervicornis and Acropora palmata. These are all common reef builders in the Caribbean, though the depth of the reefs (~14-19 m) is somewhat deeper than typically expected for A. palmata (Aronson et al., 2002; Montaggioni and Braithwaite, 2009). Internally, the reefs are poorly cemented and unconsolidated coral rubble is a major constituent within the framework.
Objective 2 – Establish Coral-Based Oxygen and Carbon Isotopic Records of Paleoenvironmental Conditions
This objective has not yet been met. Meaningful coral-based stable isotope records typically require larger corals with massive growth morphology so that records of several years to decades may be established. Isolated measurements are generally not as useful. While the M. annularis complex is the primary frame builder in these reefs, coral rubble is also a major component. Thus, recovery of large, intact massive corals was problematic due to the nature of the reefs. Additionally, larger corals were often heavily bioeroded disrupting the resultant record. Still, over the course of the coring several larger, massive coral samples suitable for paleoenvironmental work were eventually recovered. These samples range in age from ~200 to 1200 yr BP and are now being analyzed for Sr/Ca and stable oxygen isotope composition to establish seasonal paleotemperature records.
Objective 3 – Correlate Reef-Accretion and Isotopic Records with Other Paleoclimatic Records
Thus far, this objective cannot be met due to the nature of the record recovered. An overriding hypothesis of the stated objectives was that the outer shelf reefs to be studied were relict give-up reefs. This was based on previous coring studies in the area (Hubbard et al., 1997). Our research shows that the reefs are not relict, but, rather, are recent features that are actively accreting. Their recent upward accretion is consistent with the availability of accommodation space and minimal sea-level changes over the last millennia. Upon completion of Objective 2 described above, seasonal paleotemperature records from recovered corals will allow for a comparison of seasonal temperature patterns from intervals prior to anthropogenic influence with modern instrumental records.
Discussion of Impact and Products
This work documents two new records of Holocene reef accretion on the insular shelf of southwest Puerto Rico. The record obtained from the outer-shelf hummock reefs identifies a new, more recent phase of Holocene reef accretion and alters our view of existing models of Holocene reef accretion patterns on the Parguera Shelf. Importantly, this work identifies an area that represents the current locus of active reef growth on the outer shelf. The record obtained from the inner-shelf shoal documents an example of reef give-up and ecological turnover and highlights the delicacy of reef ecosystems even under natural pristine conditions. Two separate manuscripts for publication are in preparation that will summarize these findings.
Data generated by this project forms the basis for two Masters theses in the UPRM Department of Geology under the direction of Dr. W. Ramírez. One student (Marianel Mercado) is completing her thesis and will graduate in May 2012. The other student’s thesis (Edgardo Quiñones) is still in progress. Dr. C. Sherman has given three presentations on this work, two at UPR Sea Grant symposia and one at the 2011 meeting of the Association of Marine Laboratories of the Caribbean in San Jose, Costa Rica. A fourth presentation will be given by Sherman at the 2012 International Coral Reef Symposium in Cairns, Australia.
Recommendations
The cross-shelf coring study of Hubbard et al. (1997) consisted of three roughly shoreparallel transects of cores along the inner, middle and shelf-edge reef tracts. The two-phase model of Holocene reef accretion on the Parguera Shelf is based on this record. Our work identifies a third more recent phase of reef accretion along the outer shelf that was not evident in the Hubbard et al. study. Coring sites from this work were similarly oriented parallel to the shelf margin, landward of the Hubbard sites. Given our results, it now becomes evident that a new coring study is required along the shelf margin with a transect of cores oriented perpendicular to the margin. This work would more clearly document Holocene development of the shelf-margin reefs and possible retrogradation of active reef growth across the margin. Understanding shelfedge reef accretion is especially important as Holocene accretion at the shelf-edge often exceeds that of emergent reefs further inshore and shelf-edge reefs typically have the highest live coral cover across the Parguera Shelf.
Bibliography
Aronson, R.B., Macintyre, I.G., Precht, W.F., Murdoch, T.J.T. and Wapnick, C.M., 2002. The expanding scale of species turnover events on coral reefs in Belize. Ecological Monographs, 72(2): 233-249.
Blanchon, P. and Perry, C.T., 2004. Taphonomic differentiation of Acropora palmata facies in cores from Campeche Bank reefs, Gulf of Mexico. Sedimentology, 51: 53-76.
Budd, A.F. and Klaus, J.S., 2001. The origin and early evolution of the Montastraea "annularis" species complex (anthozoa: Scleractinia). Journal of Paleontology, 75(3): 527-545.
Cabioch, G., Montaggioni, L.F., Faure, G. and Ribaud-Laurenti, A., 1999. Reef coralgal assemblages as recorders of paleobathymetry and sea level changes in the Indo-Pacific province. Quaternary Science Reviews, 18: 1681-1695.
Hubbard, D.K., Burke, R.B. and Gill, I.P., 2001. The role of framework in modern reefs and its application to ancient systems. In: G.D. Stanley (Editor), The history and sedimentology of ancient reef systems Topics in geobology. Springer, New York, pp. 351-386.
Hubbard, D.K., Burke, R.B., Gill, I.P., Ramirez, W.R. and Sherman, C.E., 2008. Coral-reef geology: Puerto Rico and the US Virgin Islands. In: B.M. Riegl and R.E. Dodge (Editors), Coral Reefs of the USA. Springer, pp. 263-302.
Hubbard, D.K., Gill, I.P., Burke, R.B. and Morelock, J., 1997. Holocene reef backsteppingsouthwestern Puerto Rico shelf. In: H.A. Lessios, I.G. Macintyre and M. McGee (Editors), Proceedings of the Eighth International Coral Reef Symposium, pp. 1779-1784.
Hubbard, D.K., Zankl, H., van Heerden, I. and Gill, I.P., 2005. Holocene reef development along the northeastern St. Croix shelf, Buck Island, U.S. Virgin Islands. Journal of Sedimentary Research, 75(1): 97-113.
Hughes, T.P., 1994. Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science, 265: 1547-1551.
Hughes, T.P., Baird, A.H., Bellwood, D.R., Card, M., Connolly, S.R., Folke, C., Grosberg, R., Hoegh-Guldberg, O., Jackson, J.B.C., Kleypas, J., Lough, J.M., Marshall, P., Nystrom, M., Palumbi, S.R., Pandolfi, J.M., Rosen, B. and Roughgarden, J., 2003. Climate change, human impacts, and the resilience of coral reefs. Science, 301: 929-933.
James, N.P. and Bourque, P.-A., 1992. Reefs and mounds. In: R.G. Walker and N.P. James (Editors), Facies models: response to sea level change. Geological Association of Canada, pp. 323-348.
Knowlton, N., Maté, J.L. and Guzmán, H.M., 1997. Direct evidence for reproductive isolation among the three species of the Montastraea annularis complex in Central America (Panamá and Honduras). Marine Biology, 127(4): 705-711.
Knowlton, N., Weil, E., Weigt, L.A. and Guzmán, H.M., 1992. Sibling Species in Montastraea annularis, Coral Bleaching, and the Coral Climate Record. Science, 255(5042): 330-333. Montaggioni, L.F. and Braithwaite, C.J.R., 2009. Quaternary Coral Reef Systems: History, Development Processes and Controlling Factors. Developments in Marine Geology, Volume 5. Elsevier, 532 pp. Neumann, A.C. and Macintyre, I.G., 1985. Reef response to sea-level rise: keep-up, catch-up, or give-up, Fifth International Coral Reef Symposium, Tahiti, pp. 105-110.
Prada Triana, M.C., 2002. Mapping benthic habitats on the south west of Puerto Rico as determined by side scan sonar, University of Puerto Rico-Mayaguez, Mayaguez, 176 pp. Reimer, P.J., Baille, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Blackwell, P.G., Bronk Ramsey, C., Buck, C.E., Burr, G.S., Edwards, R.L., Friedrich, M., Grootes, P.M., Guilderson, T.P., Hajdas, I., Heaton, T.J., Hogg, A.G., Hughen, K.A., Kaiser, K.F., Kromer, B., McCormac, G., Manning, S.W., Reimer, R.W., Richards, D.A., Southon, J.R., Talamo, S., Turney, C.S.M., van der Plicht, J. and Weyhenmeyer, C.E., 2009. INTCAL09 and MARINE09 radiocarbon age calibration curves, 0–50,000 yrs Cal BP. Radiocarbon, 51: 1111-1150.
Toscano, M.A. and Macintyre, I.G., 2003. Corrected western Atlantic sea-level curve for the last 11,000 years based on calibrated 14C dates from Acropora palmata framework and intertidal mangrove peat. Coral Reefs, 22: 257-270.
Wolanski, E., Richmond, R., McCook, L. and Sweatman, H., 2003. Mud, marine snow and coral reefs. American Scientist, 91: 44-51.