
46 minute read
Las células de cáncer hacen trampa con su dieta


Por Marisa Alonso Nuñez
Todas las células tienen que comer para generar energía y así poder llevar a cabo todas sus funciones. Simple. Sí. Simple y complicado a la vez. Todos los procesos implicados en el proceso de crecimiento y división de una célula son tan importantes que están muy controlados. Imaginaos que cada célula de nuestro cuerpo decidiera crecer y dividirse de forma incontrolada… sería un caos. De hecho existen muchos mecanismos en nuestras células que, cuando algo va mal en ellas, provocan su propia muerte para que esos defectos no pasen a células nuevas. Sin embargo, las células que componen los cánceres parece que escapan a estos mecanismos de control y son capaces de dividirse incontroladamente. Y es que las células cancerígenas son demasiado listas y modifican una serie de cosas en su funcionamiento para que los mecanismos de control de las propias células no detecten que son células anómalas y evitar así su propia muerte. Un ejemplo muy claro lo tenemos en la forma en que las células de cáncer obtienen su energía. Las células en condiciones normales y en presencia de oxígeno transforman la glucosa en energía (en forma de una molécula llamada ATP) a través de varios procesos. El primero se llama glucolisis que no ocurre en ningún compartimento específico de la célula, sino en su citoplasma. Este proceso da lugar a un compuesto que se llama piruvato y que en presencia de oxígeno será transformado en el ciclo de Krebs para producir una alta cantidad de ATP. El ciclo de Krebs ocurre en las mitocondrias, que son las centrales energéticas de las células. Sin embargo, cuando no hay suficiente cantidad de oxígeno el piruvato es transformado en lactato, a través de la glucolisis anaeróbica o fermentación, en el citoplasma dando lugar a ATP pero en mucha menos cantidad. Todos estos
Advertisement
procesos están muy bien regulados y cuando algo no va bien en las mitocondrias la célula lo nota y activa un mecanismo de autodestrucción llamado apoptosis para evitar problemas mayores.
Durante los años 30 Otto Warburg sugirió la idea de que las células de cáncer apagaban sus mitocondrias y obtenían la energía que necesitaban a través de la glucolisis en el citoplasma incluso en presencia de oxígeno (glucolisis aeróbica). El efecto Warburg, como se le llama a esta teoría, se ha aceptado durante muchos años pero nunca se ha sabido cómo ocurre. Sin embargo, recientemente el grupo del profesor Michael Lisanti (en donde trabaja el investigador Stephanos Pavlides, que ha ayudado con sus conocimientos sobre el tema a la elaboración de este artículo) ha realizado una serie de estudios que ponen en entredicho esta teoría. Las células cancerígenas producen radicales libres y viven rodeadas de un estrés oxidativo debido a esto. Los investigadores del grupo de Lisanti estudiaban las células normales que se encuentran rodeando a las células de cáncer de mama, llamadas fibroblastos, y al cultivarlas junto con las células de cáncer de mama vieron que el estrés oxidativo producido por las células de cáncer afectaba a dichos fibroblastos. Estos radicales
libres producidos por las células cancerígenas estimulan el que los fibroblastos digirieran sus componentes por un proceso llamado autofagia. Al digerir sus componentes crean moléculas energéticas (piruvato y lactato) que las células de cáncer captan para alimentarse y poder llevar a cabo sus funciones. Así los fibroblastos estarían alimentando a las células de cáncer promoviendo su crecimiento y haciendo que estas células vuelvan a encender sus mitocondrias de nuevo (y escapando de la apoptosis por problemas en las mitocondrias). Ellos han llamado “efecto Warburg inverso” a este hecho de que la glucolisis aeróbica no ocurra en las células de cáncer, sino en los fibroblastos. Además, la autofagia en fibroblastos produce más radicales libres que a su vez amplifican el efecto de los producidos por las células del cáncer fomentando también el daño en el ADN de las células y generando su inestabilidad (inestabilidad genómica), uno de los procesos que hacen que las células se vuelvan cancerígenas. En definitiva, el que las células de cáncer hagan que los fibroblastos se comporten así promueve que ésFigura 2. División de las células normales y tos a su vez las alimenten (producicancerígenas. Fuente Wikipedia endo por medio de la autofagia los Y todas estas conclusiones vienen de compuestos energéticos), las ayuden unos estudios previos realizados en a que evolucionen (produciendo el ese y otros laboratorios en los que se daño en el ADN y la inestabilidad descubrió que niveles bajos de una genómica) y las protejan contra la proteína llamada caveolina1 o Cav1 muerte celular apoptótica (haciendo estaban relacionados con los cánceque las células de cáncer vuelvan a res de mama más agresivos y de peor usar sus mitocondrias de una forma evolución. Al principio no sabían muy normal). bien porqué pero descubrieron que en los tumores donde no existe apenas caveolina1 había una alta cantidad de proteínas implicadas en la glucolisis aeróbica, es decir, indicio de que estas células tenían una alta actividad autofágica. Y esto pasaba concretamente en las células que rodeaban a los cánceres (fibroblastos) y no en las células cancerígenas propiamente dichas. De hecho esta baja cantidad de caveolina1 en los fibroblastos que rodean al tumor predice la recurrencia del cáncer, su metástasis y la resistencia al tamoxifeno (fármaco común para tratar el cáncer de mama) en muchos de los cánceres de mama y de páncreas entre otros.
Como podéis ver este tema es muy complicado pero muy prometedor.
De hecho este efecto puede explicar varios hechos relacionados con el tratamiento de ciertos cánceres. Un tipo de tratamiento que se ha intentado hacer sin mucho éxito es la terapia que busca suprimir la formación de nuevos vasos sanguíneos en el tumor o terapia antiangiogénica. Estos fármacos producen una deficiencia de oxígeno o hipoxia en los alrededores del cáncer, promoviendo a su vez estrés oxidativo y autofagia. Estas son las condiciones ideales para que las células de cáncer crezcan y se reproduzcan alimentándose de los nutrientes que producen los fibroblastos que se encuentran en sus alrededores al llevar a cabo la autofagia.
Por otra parte, también se han hecho estudios con antioxidantes en el tratamiento de cáncer como ayuda a la

Figura 4. Esquema de las características de los fibroblastos en un cáncer

quimioterapia. La base de esto sería que los antioxidantes eliminarían los radicales libres producidos por las células cancerígenas evitando la autofagia en los fibroblastos y cortando la vía de alimentación alternativa del cáncer. Sin embargo, al administrarse conjuntamente con quimioterapia su efecto se vería bastante mermado debido a que muchos tipos de quimioterapia producen una alta cantidad de radicales libres para matar a las células cancerígenas y los antioxidantes no podrían eliminarlos todos. Así, ciertos estudios que se han llevado a cabo con esta combinación de quimioterapia y antioxidantes no han dado muy buenos resultados todavía.
Otra posibilidad es el uso de fármacos que bloqueen la autofagia, evitando que los fibroblastos de digieran y liberen los nutrientes que las células de cáncer tomarían para alimentarse. De hecho, el fármaco que se utiliza en el tratamiento de la malaria, la cloroquina, actúa de esta forma y sería interesante estudiar su efecto contra el cáncer. Además, los fármacos que suprimen la habilidad de la mitocondria para quemar lactato y otros productos de la glucolisis podrían ayudar a cortar el abastecimiento de energía a las células de cáncer. De hecho, uno de estos fármacos, la metformina, que se utiliza en el tratamiento de la diabetes, parece que tiene un efecto sobre el cáncer haciendo que enfermos que la están tomando tengan menos riesgo de desarrollar cáncer, aunque no se ha demostrado científicamente que esto sea cierto. En definitiva, el estudio de los procesos de producción y consumo de energía, o metabolismo, en las células de cáncer y en las células de su entorno se está convirtiendo en un campo de investigación cada vez más prometedor a la vez que polémico, ya que últimamente se están publicando bastantes artículos científicos sobre estudios con fármacos bien conocidos y utilizados en otras enfermedades, mostrando el efecto que tienen en el cáncer. Esto abre una línea nueva de posibles tratamientos para los diferentes cánceres que esperemos tengan resultados positivos y ayuden a disminuir la mortalidad que produce este grupo de enfermedades llamadas “cáncer”.
Marisa Alonso Núñez es de Astorga (León) y estudió las licenciaturas de Farmacia y Bioquímica en Salamanca, doctorándose en Microbiología y Genética Moleculares por la Universidad de Salamanca en 2007. Después de terminar la tesis se marchó a Manchester (Reino Unido) donde lleva trabajando como investigadora postdoctoral en el Paterson Institute for Cancer Research durante 4 años.En sus ratos libres se dedica a la divulgación científica. De hecho puedes seguirla en twitter en su cuenta @lualnu10 y en su blog Caja de Ciencia en español o Science Box en inglés. En estos momentos es la Coordinadora del Secretariat de Eurodoc, el Consejo Europeo de Jóvenes Investigadores y le apasiona la música desde hace 24 años siendo ésta y los 4 instrumentos musicales que toca muy importantes para ella.
En el suelo las plantas no viven solas. A todos nos han dicho alguna vez de pequeños después de jugar en la tierra, “¡No te lleves las manos a la boca! o “lávate las manos” al llegar a casa. Qué gran verdad. El suelo está habitado por toda una variedad de organismos, muchos de ellos beneficiosos para las plantas que comparten el mismo territorio. Así, además de pequeños bichitos, encontramos bacterias y hongos. El suelo como veis, es un gran ecosistema. El caso es que aunque a muchas personas no les suene este tipo de simbiosis, lo cierto es que no es nada nueva. El término de “micorriza” fue empleado por primera vez por Albert Berhhard Frank en 1885. Las plantas y sus micorrizas tienen una historia evolutiva común ya que los registros fósiles de plantas más antiguos que se conocen presentan en sus primitivas raíces unas estructuras similares a las de las actuales micorrizas. En la Fig. 3 podéis ver el parecido tan asombroso entre el fósil Rhynie y las actuales micorrizas. Datan de hace unos 460 millones de años (Ordovícico), momento en el cual establecerían relaciones con los ancestros de los
COMENSALISMO

PARASITISMO


SIMBIOSIS
Figura 1. Interacciones biológicas entre organismos.
La inmensa mayoría de las plantas (un 95%) que crecen sobre la corteza terrestre viven asociadas en forma de simbiosis, normalmente mutualista, con ciertos hongos del suelo constituyendo las llamadas “micorrizas”, término que deriva del griego mykos (hongo) y riza (raíz). Esta simbiosis está tan extendida que comúnmente se dice que las plantas no tienen raíces sino micorrizas. ¿Qué es una simbiosis? En el siguiente esquema lo veréis claro (Figura 1). primitivos briófitos. La razón sobre la que se fundamenta tal aseveración es obvia: las micorrizas conferirían a las primitivas plantas una capacidad inusitada para establecerse y captar nutrientes y agua en un medio tan hostil. La biología molecular, una vez más, ha sido la encargada de confirmar la datación, filogenia y evolución de estos hongos y su asociación con las plantas.
El botánico alemán Anton de Bary a finales del siglo XIX acuñó el término “simbiosis” para describir la estrecha relación de organismos de diferente tipo. Concretamente la definió como «la vida en conjunción de dos organismos de diferente especie, normalmente en íntima asociación, y por lo general con efectos benéficos para al menos uno de ellos». Quizá el ejemplo de simbiosis que más se conoce es el de la anémona y el pez payaso (Figura 2).

Hay otros ejemplos importantísimos y de una entrega total y hermosa en la naturaleza, pero esos se quedarán para otra ocasión.
Volviendo a la simbiosis que hoy os presento, sólo en unas pocas familias botánicas (fundamentalmente crucíferas y quenopodiáceas) hay especies que no forman micorrizas… o dicho de otro modo, algunas plantas pueden vivir sin micorrizas, pero el hongo es un simbionte OBLIGADO que no puede vivir sin estar asociado a una planta. Figura 2. Ejemplo de simbiosis mutualista entre el pez payaso y la anémona de mar. El pez payaso obtiene protección frente a depredadores (no toquéis una anémona) y usa como despensa alimenticia a la anémona, que a su vez obtiene alimento y protección frente a otros peces comedores de anémonas debido al carácter de territorialidad de los peces payaso.
Rhynie sil ó F

actual Micorriza

Figura 3. Comparativa entre distintas estructuras del hongo de la micorriza perteneciente al fósil Rhynie (370 millones de años) y el hongo actual.
Antes de entrar en materia y conocer cuál es la Os presento una micorriza arbuscular… base de esa simbiosis que llevan a cabo con las plantas y la cantidad de beneficios que aportan, veamos los tipos de micorriza que encontramos. Una pista… estamos en otoño y seguramente habréis ido a buscarlas a vuestros bosques. El nombre de micorriza arbuscular viene de la forma de la estructura más representativa del hongo, el arbúsculo. Tiene forma de arbolito y es la estructura donde tiene lugar el intercambio de nutrientes. La espora, Tipos de micorrizas es la forma reproductiva y la más importante, pues es capaz de esperar latente en el suelo años hasta encontrar las condiciones idóneas para germinar, buscar una raíz compatible, penetrarla a través del apresorio y desarrollar A grandes rasgos, podemos encontrar dos grupos el micelio fúngico tanto intra como extracelularmente, tróficos que albergan cinco tipos de micorrizas: colonizando así la totalidad de la raíz. Cabe mencionar que la colonización también puede llevarse a cabo (a) Las ectomicorrizas, más conocidas por la mayoría mediante hifas presentes en el suelo o bien a través de de vosotros ya que son las que conocemos como setas fragmentos de raíz previamente micorrizados. Podéis y trufas. Se caracterizan porque no colonizan la raíz observar todas las estructuras características en la figura 6. intracelularmente sino que forman un manto externo y por eso también se le llaman micorrizas formadoras de manto (Figura 4). El suelo está ¿Y qué hacen las micorrizas? (b) El otro grupo, menos conocido habitado por y no por ello menos importante, toda una variedad son las endomicorrizas. Más del 80% de las especies de de organismos, Pues parece que la función básica de esta simbiosis es adquirir del suelo plantas, entre ellas las de interés muchos de ellos y transferir a la planta nutrientes agronómico y las características del beneficiosos para minerales (fundamentalmente matorral mediterráneo, forman las endomicorrizas llamadas “micorrizas arbusculares” (MA). las plantas que comparten el mismo fosfato, amonio) y agua. Todo ello a cambio de carbono que él es incapaz de sintetizar por sí mismo y que lo Como su nombre indica, colonizan territorio recibe gracias a la fotosíntesis de las intracelularmente la raíz y son plantas. microscópicas. (Figura 5.)
Desde este momento nos centraremos en la estructura y función biológica de las micorrizas arbusculares así que pasemos a continuación a ver cómo es una micorriza. Se ha demostrado que algunos árboles como los pinos, son incapaces de sobrevivir más de dos años si no están micorrizados y otras especies como las orquídeas ni siquiera podrían subsistir si no estuvieran colonizadas por ellos. Sin embargo, esta simbiosis representa muchísimo

Figura 4. Estructura y morfología de las ectomicorrizas.
más que un “simple” intercambio de nutrientes, y es aquí es donde empieza el interés para la biotecnología, ecología, medio ambiente y todas las áreas de la ciencia preocupadas por mantener la diversidad y la salud en el ecosistema. Los beneficios ecológicos que aportan las micorrizas a las plantas quedan reflejados en la figura 7.
MA desempeñan un papel crucial en la supervivencia y desarrollo de las plantas, sobre todo en suelos sometidos a condiciones de estrés (sequía, salinidad, deficiencia de nutrientes), como los que caracterizan a los ecosistemas mediterráneos, así como en suelos degradados por procesos erosivos, incendios forestales, laboreo excesivo y contaminación. No os asustéis. No voy a desarrollar cada una de las funciones biológicas porque darían para capítulos individuales, pero sí daremos una pincelada de los más destacables.
Incrementan la estabilidad del suelo
Como podréis imaginar, se considera que las
Durante el desarrollo de la MA, el micelio del hongo crece rápidamente expandiendo sus hifas y colonizando el suelo de una forma totalmente invasiva (Figura 8). Se han encontrado 30 m de micelio fúngico por gramo de suelo. La consecuencia es una aumento de la superficie de captación de agua y nutrientes para la planta y por supuesto, favorecer la estabilidad del sustrato.
La inmensa mayoría de las plantas ... viven asociadas en forma de simbiosis, normalmente mutualista, con ciertos hongos del suelo constituyendo las llamadas “micorrizas”
Además del micelio, una proteína insoluble en agua producida por el hongo, la glomalina, tiene un color típico café-rojizo y está implicada en la formación de agregados del suelo. Tanto el micelio como la glomalina conducen a incrementar la estabilidad y calidad del suelo.
Algunas prácticas de agricultura como los monocultivos, el arado o la fertilización tienen efectos negativos tanto en la cantidad como en la diversidad de hongos MA presentes en el suelo. El hecho de que se reduzca la biomasa fúngica hace que se produzca un efecto

Figura 5. Estructura y morfología de las endomicorrizas. negativo en la estabilidad del suelo y como consecuencia, aumentará la erosión. En Reino Unido, las pérdidas de las cosechas debido a la erosión del suelo agrícola se han estimado en 9.99 millones de euros/año. Y esto es algo a tener en cuenta, ya que en muchas ocasiones, el impacto de la erosión es acumulativo y la mayoría de las veces irreversible.
Incrementan la tolerancia frente a estreses abióticos
Principalmente sequía, salinidad y metales pesados son los estreses abióticos que más afectan a nuestros cultivos españoles. Pero no son los únicos. También entran en juego el frío, el calor, el pH, el viento y un sin fin de situaciones ambientales a las que las plantas, por su condición de organismos sésiles no pueden escapar.
Reducen la necesidad de P para la planta
Los hongos MA son unos aliados importantísimos
El fosfato, que es un nutriente esencial para la planta, es uno de los tres principales nutrientes que se aplican en agricultura. Las fuentes de fosfato de roca son limitadas y se calcula que desaparecerán en 100 años. El exceso de aplicación de P es una importante causa de eutrofización del agua, es decir, de un enriquecimiento de este nutriente, lo que podría originar un crecimiento masivo de organismos como algas, alterando por tanto la estabilidad del ecosistema. en este campo. Les proporcionan estrategias bioquímicas, filológicas y moleculares para evitar y paliar los efectos originados por estos estreses que cada año causan pérdidas astronómicas.
Tenemos que tener en cuenta que sólo un porcentaje muy bajo (entre 0,1-0,3 partes por millón) se encuentra realmente en solución, plenamente disponible para plantas y microorganismos. Como ya hemos mencionado, otra de las ventajas del gran desarrollo del micelio fúngico es aumentar en varios órdenes de magnitud el volumen de suelo que puede ser explorado por la planta. Se estima que una reducción del 80% de fertilizante rico en P se puede sustituir por la inoculación con hongos MA. Evidentemente, esta reducción del uso de fertilizante tendría un importante impacto económico y ambiental.

Figura 6. Estructuras morfológicas del hongo de la micorriza arbuscular. La raíz ha sido teñida con la técnica del azul tripán

Figura 7. Diversidad funcional de las micorrizas arbusculares (MA) en los ecosistemas terrestres.
Ojo al dato. En el año 2025, habrán desaparecido dos tercios de la tierra cultivable en África, un tercio en Asia y una quinta parte en América del Sur y la superficie cultivable por habitante también se disminuirá a 0.15 ha en 2050. En USA y España, un tercio está en vías de desertificación. ¿No os parece inquietante? La población mundial sigue creciendo, llegaremos a 7000 millones a finales de año y comer es una mala costumbre que nos acompaña cada día. Según la FAO, 925 millones de personas están sufriendo hambre crónica. Inquietante y desesperante. Desde el punto de vista ecológico, una de las líneas de investigación que se lleva a cabo es manipular aislados autóctonos de hongos MA de zonas áridas para la revegetación de zonas degradadas en ecosistemas mediterráneos. Por poner un ejemplo, una cepa de hongo tolerante a la sequía (aislada de una zona afectada por sequía severa) asociada con una bacteria nativa presente también en el suelo, es capaz de reducir en un 42% los requerimientos de agua para la producción de Retama sphaerocarpa. A simple vista, es evidente la mejora que proporciona el hongo sobre la parte aérea y por tanto la biomasa de la planta en condiciones de sequía y salinidad

(Figura 9). ácidas alcanzaron las orillas de los ríos Agrio y Guadiamar, hasta llegar a uno de los mayores emblemas naturales Igual ocurre con el estrés salino, y los cultivos del mundo: el Parque Nacional de Doñana. El agua de olivos en España o los de palma del norte de África. arrastraba oxígeno sin disolver y sólidos en suspensión Son innumerables los estudios que demuestran el papel que provocaron la muerte de toda la vida subacuática: protector de estos pequeños organismos frente a una más de 30 toneladas de peces muertos y 170 de cangrejos gran variedad de condiciones salinas. El mecanismo fueron recogidos en los días siguientes. que utilizan es complejo pero podemos decir que lo ... se considera que consiguen aumentando la captación de agua y nutrientes, las MA desempeñan El accidente minero del el intercambio gaseoso, la un papel crucial en Guadiamar marcó un punto de transpiración y conductancia estomática, el balance iónico y hormonal, y ajustando la supervivencia y desarrollo de las inflexión en la relación entre la ciencia y la conservación de la naturaleza en España. La el balance osmótico y plantas, sobre todo comunidad científica española composición de carbohidratos como la prolina que tiene un papel fundamental. Además en suelos sometidos a condiciones de estrés se volcó en aportar soluciones ante el mayor accidente ambiental de nuestro país. ponen en marcha una serie de Hoy en día, la fitorremediación mecanismos bioquímicos que ( f i t o e x t r a c c i ó n , implican la activación de un pool de enzimas antioxidantes fitoestabilización y rizofiltración) de los suelos y moleculares que abarca la inducción de genes como contaminados por metales pesados es actualmente una aquaporinas, proteínas LEA (Late embriogénesis de las herramientas más efectivas y más “limpia” con el abundant), canales de transporte, etc. medio.
Durante las últimas décadas, se ha acelerado el proceso de contaminación atmosférica por metales pesados provenientes de la minería, hornos de fundición, industria, tratamientos de suelos de cultivo con agroquímicos y sedimentos del suelo etc. Se vierten a los suelos elementos peligrosos como el plomo (Pb), arsénico (As), cadmio (Cd) y mercurio (Hg) con el consiguiente peligro de su conducción a las capas freáticas del suelo. La madrugada del 25 de abril de 1998 una brecha de 60 m de ancho y 30 m de alto quebró la balsa minera de la localidad sevillana de Aznalcóllar (Fig. 10). En pocas horas siete millones de metros cúbicos de lodos tóxicos y aguas Hay plantas que de forma natural hiperacumulan ciertos metales. Pues bien, estas especies vegetales junto con las MA que contengan o aquellas más eficientes a tal fin, se están utilizando como estrategias de fitorremediación…. o de micorrizoremediación.
La inmovilización de metales en la biomasa fúngica constituye un mecanismo mediante el cual estos

Figura 9. (a) Plantas de tomate sometidas a estrés hídrico. Las plantas control no han sido inoculadas y las plantas MA llevan hongo. (b) Plantas de maíz sometidas a estrés por salinidad. Las plantas control no han sido inoculadas y las micorrizadas contienen hongo MA.
microorganismos seleccionados de la rizosfera para minimizar los riesgos tóxicos de los contaminantes y a su vez maximizar el crecimiento y la nutrición. El aislamiento de cepas de hongos MA adaptados al estrés constituye ya y será la clave como herramienta biotecnológica para la inoculación de plantas en ecosistemas alterados.

Incrementan la tolerancia frente a estreses bióticos
Figura 10. Imagen de las consecuencias del accidente minero de Aznalcóllar.
pesados. Las raíces actúan como una barrera frente al transporte de metales, reduciendo la transferencia y mejorando el ratio de Cd raíz/parte aérea. Una vez más, recurrimos al beneficio del micelio del hongo, que además de aumentar la captación de agua y nutrientes y favorecer la estabilidad del suelo, actúa como medio tamponador de la captación de metales como el cadmio, reduciendo los efectos tóxicos de este en el crecimiento de la planta. Me entran picores de imaginar una cola de pulgones subiéndome por la pierna o una procesionaria recorriéndome la espalda… ¿os imagináis? Pues no creo que a las plantas tampoco le haga ninguna gracia. Yo me
hongos pueden incrementar la tolerancia a metales sacudiría, podría echar a correr si las viera venir y hasta gritar pidiendo ayuda. Ellas no.
Hace ya tiempo que se evidenció la capacidad de las MA para aumentar la resistencia o tolerancia de las plantas frente al ataque de determinados patógenos del suelo. La investigación en España ha demostrado tal protección en plantas de tomate, pimiento, platanera, olivo y otros frutales frente a patógenos tales como Phytophthora, Verticillium, Fusarium, nemátodos , bacterias, etc. (Figura 11)
Este efecto es atribuido a la quitina, compuesto esencial que forma parte de la pared celular del hongo y que posee la capacidad de unir metales. También recientemente, se ha visto que la glomalina anteriormente mencionada puede quelar metales, disminuyendo así la disponibilidad de estos para las plantas. Se propone otro mecanismo consistente en la dilución de la concentración de metales en los tejidos de la planta mediante un mayor crecimiento de la parte aérea.
De cualquier forma, un objetivo de la biotecnología sería usar la inoculación combinada de En esta protección operan mecanismos de distinta índole como la competencia por el nicho y nutrientes, modificaciones en las poblaciones microbianas de la rizosfera, compensación de daños… No se conoce la producción de compuestos antimicrobianos por los hongos MA, lo que descarta un efecto inhibidor directo, o sea, que se traten de “agentes de control biológico” en el sentido estricto del término.
Sin embargo, diversos modelos de estudio han sugerido la posibilidad de una protección a nivel sistémico. Además, se ha apuntado a la posible existencia de

Figura 11. Organismos patógenos de plantas (a) El hongo Fusarium oxysporum crecido en placa. (b) Microfotografía coloreada de un nematodo (Heterodera glycines) parásito de la planta de soja (Glycine max) (c) Pseudomonas aeruginosa al microscopio de barrido, con falso color.

Figura 12. Algunos productos comercializados por el spin-off Mycovitro con sede en Granada.
fenómenos de priming (potenciación de las defensas), por el que las plantas podrían estar en un “estado de alerta” que les permitiera reaccionar de manera más eficaz a la presencia del patógeno e incluso avisarse entre ellas. Algo así como ¡Ehhh compañera, se acera una procesonaria por el sur! Estos procesos de priming inducidos por la MA podrían ser eficaces frente a patógenos foliares necrótrofos o hemibiotrofos e insectos masticadores, aunque no frente a biotrofos y otros tipos de insectos más especializados.
A pesar de lo que pueda parecer, esta resistencia inducida de carácter sistémico conlleva costes mínimos en la productividad de la planta comparados con una
¿Biotecnología con las micorrizas? El comienzo…
El estudio de las micorrizas como es lógico comenzó por ser de tipo fisiológico (efectos sobre la nutrición vegetal), luego molecular (tratando de dilucidad los aspectos de la protección que confieren) y cómo no. ecológico (presencia de micorrizas en plantas de interés agronómico, pratense y forestal). Posteriormente, se han iniciado aislamientos y caracterización de hongos MA asociados tanto a cultivos de interés como a plantas de la sucesión natural, en las que últimamente se han incluido especies de la flora amenazada y/o endémica, propias de los ecosistemas ibéricos (Sierra Nevada, por poner un ejemplo… y barrer para casa).
Durante 30 años se ha investigado en esto, hasta hongos MA de
el punto de disponer en la actualidad en la Estación Experimental del Zaidín, EEZ (CSIC, Granada) de un Banco de Glomeromicetos (Glomus es el género más famoso de estos hongos) integrado por unos 80 aislados conservados en cultivo puro de los cuales, la mitad están caracterizados molecular y filogenéticamente. Distintas instituciones españolas contribuyen así a incrementar el BEG (Banco Europeo de Glomeromicetos).
He mencionado “cultivo puro”. ¿No os resulta extraño tratándose de un simbionte obligado? En la EEZ se han desarrollado dos patentes, una de las cuales permite obtener micorrizas in vitro mediante cultivo monoxénico de raíces. Esto es, colocar en condiciones totalmente estériles un pequeño fragmento de raíz micorrizada en un compartimento de una placa de Petri
bicompartimentada y dejar que se desarrolle. Las hifas del hongo son capaces de saltar la barrera y seguir creciendo en el otro compartimento, donde encontraremos únicamente al hongo. Precisamente, sobre la base de este procedimiento se ha desarrollado la única colección española y una de las pocas a nivel mundial, de hongos MA conservados in vitro.
Parece ser que el potencial de estos hongos y sus beneficios ecológicos llegan a ser interesantes económicamente. Mycovitro, una empresa de base tecnológica de tipo spin-off surgida de la EEZ, produce y comercializa inoculantes, denominados de forma global como “glomigel”, específicos para cada tipo de cultivo/ ecosistema (Figura 12). No es la única empresa. Mycosim, Agromed, Bio-Triton, Plant Health Care, etc., tienen actividades similares y parece que es un negocio en auge y distribuido por todo el mundo.
En lo referente a ensayos de inoculación en campo hay que destacar la interacción con empresas viverísticas (para la producción de planta micorrizada), de gestión ambiental (restauración de áreas degradadas mediante revegetación con planta micorrizada) y productoras de inoculantes. Concretamente, se ha ensayado la micorrización de Pinus sp., Pseudotsuga sp., Quercus sp., entre otros con hongos ectomicorrícicos. Por su interés económico, merecen especial mención las investigaciones conducentes a implementar la micorrización de encina, orientada a la producción de trufa.
En lo que atañe a ensayos de campo con hongos MA, se han desarrollado experimentos con leguminosas, tomate, calabaza, espárrago y otros hortícolas, pero los aportes más significativos son los referentes a la micorrización de frutales y de plantas de interés en el matorral arbustivo mediterráneo. Como frutales de interés, se ha centrado en olivo, cítrico, vid, frutales de hueso y en cultivos tropicales (aguacate, chirimoyo, piña, platanera, papaya, etc). En cuanto a plantas de interés en ecosistemas naturales (incluyendo aromáticas, medicinales, melíferas etc) cabe destacar los estudios con Retama sphaerocarpa, Genista cinerea, Rosmarinus officinalis, Lavandula latifolia, Genista umbellata, Thymus zigys, Pistacia lentiscos, etc. Teniendo en cuenta lo que hemos visto hasta ahora, parece que las investigaciones encaminadas en la biotecnología se podrían centrar en la aplicación de estos hongos en la agricultura y en la recuperación de ecosistemas degradados y flora amenazada, en interacción con la industria viverista, en gestión ambiental y en programas de control de enfermedades y plagas en cultivos.
Por lo pronto, las macetas de mi casa ven el agua cuando me acuerdo y sí,… pensándolo bien, es una ventaja poderme ir de vacaciones y ¡¡no regarlas!!
Bibliografía
- Barea JM. (2011). Investigaciones sobre micorrizas en España: pasado, presente y futuro. Fundamentos y aplicaciones agroambientales de las interacciones beneficiosas plantasmicroorganismos.
- De Bary, H.A. (2008) Die Erscheinung der Symbiose “Till death do us part”: coming to terms with symbiotic relationships. Nature Reviews Microbiology 6, 721-724
- Garg N, Chandel S. (2010) Arbuscular mycorrhizal networks: process and functions. A review. Agron. Sustain. Dev. 30:581- 599
- Gianninazzi S, Gollotte A, Binet MN, Van Tuinen D, Redecker D, Wipf D. (2010) Agroecology: the key role of arbuscular mycorrhizas in ecosystem services. Mycorrhiza 20: 519-530
- Harrison MJ (2005). Signaling in the arbuscular mycorrhizal symbiosis. Annu Rev Microbiol. 59: 19–42
- Kirk, P.M., P.F. Cannon, J.C. David & J. Stalpers (2001) Ainsworth and Bisby’s Dictionary of the Fungi. 9th ed. CAB International, Wallingford, UK - Margulis, Lynn (2002). Planeta Simbiótico. Un nuevo punto de vista sobre la evolución.. Victoria Laporta Gonzalo (trad.). Madrid: Editorial Debate
- Wang, B.; Qiu, Y.L. (2006) Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 16 (5): pp. 299–363. - Porcel R, Aroca R, Ruiz-Lozano, JM (2011) Salinity stress alleviation using arbuscular mycorrhizal fungi. A review. Agron. Sustain. Dev. DOI 10.1007/s13593-011-0029-x Published on line.
Rosa Porcel (Granada, 1976) es licenciada en Biología por la Universidad de Granada y doctora en la especialidad de Bioquímica y Biología Molecular (2006). Recibió el Premio Nacional de Investigación en Relaciones Hídricas por su trayectoria científica en 2006 y posteriormente comenzó su etapa postdoctoral con un Juan de la Cierva (2007-2009) y una JAE-doc (2009-2011). Actualmente trabaja en el CSIC en el Departamento de Microbiología del Suelo y Sistemas Simbióticos.
El verdadero sentido de la vida
Por Sergio Ferrer
1962 Francis Crick, James Watson y Maurice Wilkins reciben el Premio Nobel de Fisiología o Medicina por el descubrimiento de la estructura del ADN. La famosa estructura de la doble hélice había sido descubierta unos años atrás, en 1953, gracias en parte a los trabajos de la olvidada (y en ocasiones despreciada por sus compañeros) Rosalind Franklin.
Avery, McCarty, Chargaff… muchos investigadores habían realizado avances importantes antes de la doble hélice, sin embargo, conocer la estructura del ADN abrió las puertas a la tecnología del ADN recombinante y a la genómica. Con el descubrimiento de la Doble Hélice estábamos avanzando, sin saberlo todavía, hacia Dolly, la insulina transgénica, la terapia génica, los chips de ADN… no obstante no nos precipitemos: el estudio de los ácidos nucleicos y del ADN se remonta más allá de 1953.
La historia de la genética está íntimamente ligada a la del ADN, pero podemos decir que la Era del ADN comienza en 1944, con el descubrimiento por parte de Avery, MacLeod y McCarty de que el ADN es el portador de la información genética en las bacterias. En 1950 Chargaff demuestra que la cantidad de residuos de adenina es proporcional a la cantidad de residuos de timina en el ADN de cualquier especie, y lo mismo para los residuos de guanina y citosina. En 1952 tienen lugar dos acontecimientos clave: por un lado el histórico experimento de Hershey y Chase, que confirmaba que era el ADN el portador de la información genética (y no las proteínas como muchos pensaban). Por otro lado se obtiene la famosa Fotografía 51 mediante difracción de Rayos X. Esta imagen del ADN, realizada por Rosalind Franklin, se convirtió en una prueba decisiva a la hora de confirmar la estructura en doble hélice del ADN, en una serie de artículos publicados por Watson y Crick en la revista Nature a lo largo de 1953.
Hace mucho, mucho tiempo, en una charca no muy lejana…
… unas moléculas orgánicas sencillas dieron lugar a una protocélula. Podemos entender “el origen de la vida” como una consecuencia inevitable una vez alcanzada cierta complejidad química; y para que la vida tuviera unas mínimas oportunidades de éxito debían de darse dos condiciones indispensables: el primer paso era la distinción entre “fuera” y “dentro”. El medio interno debía diferenciarse del medio externo. La separación física (mediada por una protomembrana) era el primer paso para que el medio interno fuera separándose cada vez más del externo. Una vez logrado esto, el siguiente paso lógico era la habilidad de perpetuarse. La vida dependía de la capacidad de las células para almacenar, recuperar y expresar las instrucciones genéticas necesarias para la producción y mantenimiento de un organismo vivo. Existen muchas teorías sobre el posible origen de la vida, pero en cualquier caso lo más probable es que fuera el ARN (Ácido Ribonucleico) el “padre” del actual ADN. Algo nada descabellado si tenemos en cuenta que el ARN es el único material genético de ciertos virus.

Figura 1: Rosalind Franklin, una de las científicas más brillantes e influyentes junto a su famosa Fotografía 51
Cabría preguntarnos, dicho esto, por qué no se estableció el ARN como material genético. La explicación la encontramos en su estructura: el ARN no puede plegarse como el ADN (algo imprescindible para que la información genética ocupe el menor espacio posible) ni tampoco tiene una vida muy larga (minutos o como mucho días).
El secreto de la vida
Dicen que el día de su descubrimiento más importante Crick, que por entonces era un joven de 35 años, dijo: “Hemos descubierto el secreto de la vida”. No estaba muy desencaminado, ya que entender la estructura del

ADN era imprescindible para comprender plenamente su función. Conocer su estructura resolvió inmediatamente el problema de cómo puede copiarse (replicarse) el ADN, y aportó indicios de cómo una molécula de ADN puede codificar la información para la síntesis de proteínas. Pero, ¿qué es realmente el ADN?
El ADN (Ácido Desoxirribonucleico) es una molécula larga (macromolécula), que recuerda a una escalera, y que forma una doble hélice. Cada cadena de esta hélice es una molécula lineal formada por subunidades llamadas nucleótidos.
Una molécula de ADN está formada por dos largas cadenas de polinucleótidos compuestas por cuatro tipos de subunidades nucleotídicas. Estas cadenas (o hebras) permanecen unidas entre sí por enlaces de hidrógeno que se forman entre las bases de los nucleótidos. Estos nucleótidos están a su vez formados por un azúcar de cinco carbonos (pentosa) que tiene unido uno o más grupos fosfato y una base nitrogenada. En el ADN el azúcar es una desoxirribosa (de ahí el nombre de ácido desoxirribonucleico) unida a un único grupo fosfato, y la base puede ser Adenina (A), Guanina (G), Timina (T) y Citosina (C). Los nucleótidos se unen covalentemente entre sí a través de los azúcares y los fosfatos, formando una cadena que constituye el “esqueleto” de azúcar-fosfatoazúcar-fosfato. Podemos simplificar diciendo que la diferencia se encuentra en las bases nitrogenadas, por lo que cada cadena de polinucleótidos del ADN es un collar (el esqueleto de azúcar-fosfato) formado por cuatro tipos de cuentas (las cuatro bases nitrogenadas). Puesto que la única diferencia está en las bases, se utilizan los símbolos A,G,T,C para identificar a los cuatro nucleótidos. Dicho de otra forma: la unidad fundamental del ADN es el nucleótido, compuesto por la unión covalente de una pentosa, un
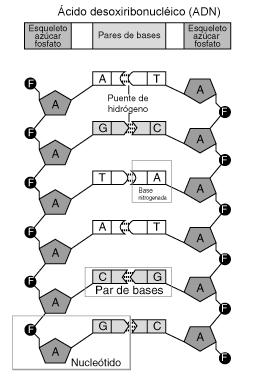
Figura 3: Podemos apreciar que los nucleótidos se diferencian únicamente en la base nitrogenada
grupo fosfato y una base nitrogenada, siendo esta última la fuente de la variación.
Por lo tanto, son estas cuatro bases nitrogenadas las que constituyen el código genético, que con sus combinaciones

Figura 4: Modelo en 3 dimensiones de una molécula de ADN
determina la secuencia de aminoácidos de las proteínas. Si la parrafada le ha resultado abrumadora, imagine que esas cuatro letras son el alfabeto genético. El secreto de la vida, que decía Crick. Los ordenadores pueden hacer maravillas con unos y ceros, y los seres vivos no quedamos atrás con nuestras cuatro letras.
Las subunidades nucleotídicas se unen entre sí
formando la cadena de ADN. Estas subunidades presentan una polaridad química que transmiten a la molécula final, de manera que los dos extremos de la cadena serán fácilmente diferenciables: por un lado encontraremos un fosfato (extremo 5’) y por el otro un hidroxilo (extremo 3’). Watson y Crick establecieron con su modelo que en la molécula de ADN, ambas cadenas son complementarias entre sí, ya que en cada peldaño de la escalera encontramos los pares A-T o G-C (pares de bases). Esta relación complementaria es fundamental tanto para la replicación como para la expresión génica. En realidad, la estructura tridimensional del ADN no es más que una consecuencia de las propiedades químicas y estructurales que tienen las cadenas de polinucleótidos. Las dos cadenas se mantienen unidas entre sí gracias a los enlaces de hidrógeno que se forman entre las bases de las diferentes cadenas. Por lo tanto, las bases se encuentran en el interior de la doble hélice y el esqueleto de azúcar-fosfato en la periferia. Además, los miembros de cada par de bases sólo pueden unirse si las dos cadenas son antiparalelas (la polaridad de una cadena es opuesta a la de la otra). to de la doble hélice contestó a dos preguntas que atormentaban a los científicos desde hace tiempo: cómo se puede almacenar químicamente la información necesaria para formar un organismo vivo, y cómo se puede replicar de manera fiel esta información.
Ya hemos jugado con las similitudes entre el sistema de almacenamiento de información de un ordenador y el de un organismo, comentando que cada una de las cuatro bases (A,T,G,C) pueden considerarse como letras de un alfabeto biológico con el que escribir mensajes en el ADN, pero ¿cómo funciona esto exactamente?

Nuestros genes contienen instrucciones para la producción de proteínas, por lo que los “mensajes” de nuestro ADN deben codificar proteínas de alguna forma. Es necesario comprender la naturaleza química de las proteínas para entender cómo: las propiedades de una proteína (directamente responsables de su función), están determinadas por su estructura tridimensional, la cual es
Figura 6: Podemos apreciar los sucesivos pasos de enrrollamiento del ADN, desde la doble hélice hasta el cromosoma final.
consecuencia directa de la cadena de aminoácidos que la

Las funciones del ADN: la clave de la herencia genética

Figura 8. Podemos apreciar las diferencias en la replicación de la hebra líder y de la hebra retrasada. Obsérvese cómo se sintetiza cada fragmento de Okazaki independientemente y luego se unen.
forman. Por lo tanto, la secuencia de nucleótidos de un gen debe describir la secuencia de aminoácidos de una proteína. El alfabeto del ADN (de 4 letras), se traduce de esta forma en el alfabeto de los aminoácidos (de 20 letras o aminoácidos) mediante distintas combinaciones. Esta correspondencia no se deduce directamente de la estructura del ADN, y llevó tiempo dilucidarla por completo. A este mecanismo por el cual la secuencia de nucleótidos de un gen se traduce a la secuencia de aminoácidos de una proteína se lo conoce como expresión génica.
Los mecanismos por los cuales el ADN codifica proteínas y por el cual se replica serán tratados más adelante.
La mayor parte de nuestro ADN es “basura”
Hemos dicho que el objetivo último del ADN es permitir a la célula sintetizar todas las proteínas que necesita, pero no hemos mencionado no todo el ADN codifica proteínas (es decir, compone genes). Distinguimos entre el ADN codificante (codifica proteínas) y el no codificante (o basura, que no codifica proteínas). Durante muchos años los científicos pensaron que este ADN no tenía utilidad, de ahí el nombre que recibió. No obstante, no parecía lógico que en el ser humano (por poner un ejemplo) el 90% del ADN no codificara nada. Esto contradecía la idea, poco ortodoxa aunque a menudo acertada, de que en la naturaleza las cosas están por algo. tructural: ADN centromérico y telomérico. Por otra parte, algunos genes no codifican proteínas, aunque sí que se transcriben en ARN.
El ADN de una de tus células es más alto que tú.
Llamamos genoma al conjunto completo de la información almacenada en el ADN de un organismo y que contiene la información de todas sus proteínas (también se utiliza genoma para referirse al ADN que contiene esta información). La cantidad total de esta información es tan enorme que cada célula humana contiene 2 metros de ADN, que la célula es capaz de recoger en un espacio de unas 6 micras. No sólo eso, sino que si escribiéramos toda la secuencia de ADN del genoma humano llenaríamos más de 1.000 libros (todos más gruesos que el más grueso de los libros de George R.R. Martin). Siguiendo con datos asombrosos sobre nuestro genoma, el genoma humano contiene la información necesaria para producir unas 30.000 proteínas. Por cierto, que toda esta información fue secuenciada gracias al proyecto científico internacional Proyecto Genoma Humano, en el que participó el propio Watson, junto a Francis Collins (y Craig Venter en el otro lado del cuadrilátero, véase la bibliografía recomendada para más información).
Toda esta ingente cantidad de información es copiada con una fidelidad asombrosa (ver siguiente apartado) y empaquetada en un conjunto de cromosomas (en nuestro caso 46). Cada cromosoma lo forma una molécula de ADN aso
ARN: la otra cara de la moneda
Aunque hasta ahora casi sólo hemos hablado del Ácido Desoxirribonucleico, lo cierto es que este no es el único ácido nucleico que nos podemos encontrar. No podemos dejar de mencionar al Ácido Ribonucleico (ARN), que ya fue presentado como el “padre” del ADN.
El ARN es una macromolécula relativamente similar al ADN, con singulares diferencias: la primera es que la pentosa presente en sus nucleótidos es ribosa (en vez de desoxirribosa) y que en vez de Timina encontramos Uracilo (las otras tres bases son iguales que en el ADN). Además suele formar cadenas más cortas, y suele ser de cadena simple (monocatenario).
No obstante, la diferencia más notable la encontramos en lo relativo a su función: todos los tipos de ARN que se sintetizan en la célula son una copia complementaria de una secuencia (más o menos larga) de desoxirribonucleótidos del ADN. Su papel principal es actuar de intermediario entre el contenedor de la información (el ADN) y el fin último de esa información (normalmente sintetizar alguna proteína). A pesar de esto, sería un enorme error decir que la única función del ARN es la de colaborar en el proceso de síntesis proteica, ya que muchos tipos de ARN regulan la expresión génica, al ser complementarios de determinadas secuencias de ADN o ARNm. Y no sólo eso, determinadas clases de ARN tienen una función catalítica, actuando como biocatalizadores.
Por último, y como ya comentamos en un apartado anterior, el ARN puede almacenar la información genética de algunos organismos, como es el caso de ciertos virus y viroides.

Figura 9: Esquema sencillo de la transcripción. Obsérvese que sólo una de las dos hebras de ADN se emplea como molde para la síntesis del ARN
Replicación, transcripción y traducción.
Ya sabemos que el ADN es capaz de crear copias idénticas de sí mismo, y que la información contenida en su secuencia de nucleótidos es capaz de traducirse en cadenas de aminoácidos que finalmente dan lugar a proteínas. Igualmente, debe ser capaz de repararse, ya que numerosos factores pueden producir alteraciones como productos químicos, radiaciones, accidentes térmicos o moléculas reactivas. Veamos brevemente cómo se producen todos estos procesos. Cada uno de ellos podría dar por sí sólo un artículo, de forma que simplificaremos todo lo que podamos.

El apareamiento de las bases (recordemos que las hebras de una molécula de ADN son complementarias) constituye el fundamento de su reparación y replicación. Antes de comenzar, resulta importante recordar que todos estos procesos no se producen necesariamente
Figura 10: Esquema del proceso de traducción. El ribosoma lee las instrucciones del ARNm mientras el ARNt va incorporando los aminoácidos necesarios.
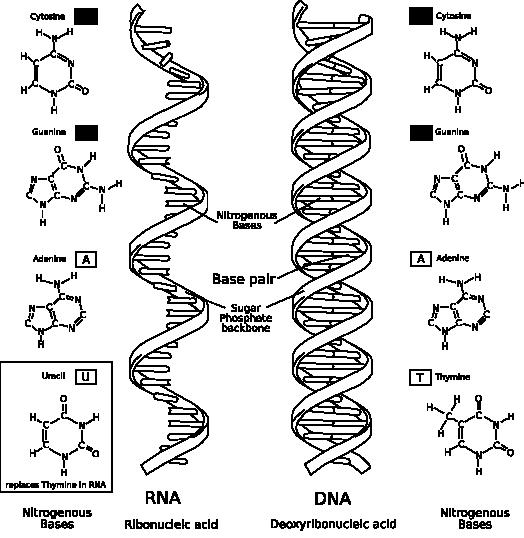
Figura 11: Comparativa entre la estructura del ADN y la del ARN
de manera simultánea. No hay que imaginarlos como una cadena de producción donde se comienza con la replicación y se termina con la traducción. La replicación tiene lugar una sola vez en la vida de la célula: cuando ésta inicia su división (mitosis). Por otra parte, transcripción y traducción se producen de manera constante: continuamente se necesitan nuevas proteínas. A pesar de esto, ambas se encuentran separadas en el espacio (y por lo tanto en el tiempo) en los eucariotas, ya que la transcripción (que utiliza el ADN como molde) se realiza en el núcleo mientras que la traducción se produce en el citoplasma. Ahora sí, pónganse el cinturón que vamos. diante el apareamiento de bases complementarias (A con T, y G con C), produciendo una secuencia complementaria de ADN. Cada una de las hebras existentes actúa como patrón de la nueva cadena: nuevos nucleótidos complementarios se van uniendo a las cadenas patrón, las cuales deben estar necesariamente separadas para que esto se produzca. La enzima responsable de este proceso, descubierta en 1957, es la ADN polimerasa. La región de replicación, que se va desplazando por la doble hélice, recibe el nombre de horquilla de replicación. Debido a que la ADN polimerasa sintetiza en dirección 5’ a 3’, el problema se soluciona de la siguiente forma: en una de las dos hebras patrón (líder) la replicación se produce de forma continua, mientras que en la otra (retrasada. Sí, se llama así) se produce una replicación discontinua (a trozos). Es
el momento de contestar a la pregunta oculta en el título del artículo: el verdadero sentido de la vida es pues, 5’-3’ (y no 42 como erróneamente se piensa). El ADN de la célula no pasa directamente a formar proteínas, sino que se utiliza el ARN como intermediario (ver apartado sobre el ARN para más información). Cuando una célula necesita una proteína determinada, la secuencia de nucleótidos del ADN necesaria (una secuencia determinada en un océano de información) se copia primero a ARN, y a ese proceso lo conocemos como transcripción. En este caso, la enzima responsable de copiar las secuencias de ADN a secuencias de ARN es la ARN polimerasa. El ARN resultante, que transmite la información desde el ADN hasta el ribosoma (donde se sintetizará la proteína) recibe el nombre ARN mensajero (ARNm).

Al último de los procesos, en el que ya no interviene el ADN sino el ARN, se lo conoce como traducción. La traducción del ARNm consiste en la polimerización de aminoácidos en cadenas polipeptídicas. La “mesa de trabajo” para este proceso es el ribosoma. En el proceso entra en juego un nuevo tipo de ARN: el ARN de transferencia (ARNt), que como su nombre indica se encarga de transportar los aminoácidos a los ribosomas para que estos se vayan ensamblando en el orden correcto que indica
Figura 12: La PCR, un gran invento
el ARNm.

Figura 14. Si no se pone esta imagen de Watson y Crick en un artículo sobre el ADN es como que falta algo importante

Y todo esto, ¿a mí de qué me sirve?
Resulta imposible encontrar una rama de la biología que no sea tocada por el largo tentáculo (tentáculo de doble hélice dextrógira, todo hay que decirlo) de la genética. Si cogemos un periódico y consultamos cualquier noticia sobre algún tipo de avance médico, veremos que casi seguro que se utilizaron PCRs durante esos ensayos. Por lo tanto podríamos decir que la respuesta a la pregunta que nos hacemos es: para todo. En este apartado intentaremos convenceros de la enorme cantidad de técnicas y avances relacionados con los ácidos nucleicos.
Para empezar, el conocimiento de la estructura del ADN permitió desarrollar toda una serie de herramientas tecnológicas que aprovechan sus características
físico-químicas: la PCR (Reacción en Cadena de la Polimerasa), la Secuenciación del ADN, los Southern Blots o los Chips de ADN son el pan de cada día en muchos laboratorios de todo el mundo, con fines de lo más diversos. Conocer la estructura del ADN permitió desarrollar la tecnología del ADN recombinante (ADN artificial desarrollado in vitro) y gracias a eso nació la ingeniería genética, que nos permite transferir ADN entre organismos, corregir defectos genéticos y sintetizar numerosos compuestos. Las aplicaciones de la ingeniería genética ocuparían medio artículo, pero entre ellas encontramos la creación artificial de compuestos como la insulina, el desarrollo de vacunas, mejora de especies de interés humano, usos en terapia génica…
Por otro lado, la secuenciación del ADN consiste en conocer la secuencia de nucleótidos de un fragmento o fragmentos de ADN, y permite no sólo realizar estudios filogenéticos, sino que tiene también una aplicación bastante más directa en medicina forense.
Otro ejemplo muy interesante sobre las aplicaciones de los ácidos nucleicos son los chips de ADN. Un chip de ADN (o microarray, biochip...) es una estructura sólida a la cual se une una colección de fragmentos de ADN. Sus aplicaciones incluyen el estudio de genes, la detección de mutaciones o la clasificación molecular de enfermedades (asociar ciertas enfermedades a mutaciones presentes en determinados genes). Epigenética, microARNs, bioinformática… la genética cada vez tiene un mayor peso en el día a día del Homo sapiens. Su potencial para mejorar nuestra calidad de vida y la de los que nos rodean es infinito: vacunas, medicinas, alimentación… y todo empieza con el descubrimiento, en 1952, de el secreto de la vida.
Fuentes:
-Conceptos de Genética. 8º edición (Klug, Cummings & Spencer).
-Biología molecular de la célula, 4º edición (Alberts et al)
Lecturas recomendadas:
-La doble hélice (James D. Watson): el propio Watson nos narra en primera persona el mayor descubrimiento del siglo XX. Imprescindible tanto por su parte científica como histórica y social (atención a sus comentarios sobre Franklin).
- Qué loco propósito (Francis Crick): lo mismo pero contado desde la perspectiva de Crick, un peculiar personaje, defensor de la extravagante teoría de la Panspermia Dirigida, cuenta con algunas anécdotas de lo más jugosas.
- La conquista del genoma humano (Kevin Davies): Crónica de la carrera por la obtención del mapa genético del ser humano. Davis, director de la revista Nature Genetics nos narra el espectacular relato de uno de los logros más impresionantes del s. XX. Amaréis u odiaréis a Venter al terminarlo (o ambas cosas).
Sergio Ferrer es (micro)biólogo, apasionado de la ciencia y de la divulgación. Podéis encontarlo de vez en cuando en su blog ‘La Muerte de un Ácaro’, y demasiado a menudo en su Twitter @SergioEfe.






