
91 minute read
Exploradores de la biodiversidad hoy: nuevas res puestas a pre guntas antiguas


Colecciones del “Darwin Centre” inaugurado en 2009 en el British Museum de Londres con motivo del bicentenario de Charles Darwin. (Fotografía: Rut Caparrós) El interes ´ por con o c e r los oren la primera ocasión en la que un occidental veía un ornitorrinco, una anguila eléctrica o un quetzal? El asombro y la incredulidad no tendrían límite, y por eso no debeganismos con los que compartimos el planeta y en saber mos extrañar
cuántos son y cómo se distinguen unos de otros es tan
Advertisement
antiguo como el comer: la supervivencia de la comunidad
puede llegar a depender de saber distinguir las plantas
comestibles y medicinales de las venenosas, por ejemplo.
Sin embargo, no fue hasta el siglo XVIII cuando el ansia
por catalogar y describir cuantas especies existieran caló
en el nuevo espíritu científico de la época. Claro que esto
no debe extrañarnos, ya que la humanidad estaba expe
rimentando una curiosidad intensísima por los entresijos

del mundo que le rodeaba; los seres vivos no iban a ser
una excepción. Siempre me ha parecido que ya en los orí
genes de este interminable catálogo, cuando las selvas y
los océanos eran aún vírgenes para los naturalistas (más
aún que hoy, quiero decir), existía una fascinación espe
cial, una expectativa por las formas increíbles que la na
turaleza iba desvelando a la ciencia, y que no tiene com
paración con otras disciplinas. En la actualidad, todos los nos de que hasta épocas relativamente tardías se diera crédito a la existencia de animales míticos como las manticoras o los grifos. El propio Linneo reservó en su Systema Naturae una clase “Paradoxa” donde tenían cabida unicor
clase acabó eliminándose en la sexta edición, pero ¿Quién hubiese apostado que estos animales no existían cuando llegaban algunos más increíbles aún recolectados en los confines de un mundo entonces tan vasto e inabarcable?
en muchas ocasiones una misma especie bajo nombres distintos sin que se pudiese evitar. Este caballo de batalla de la descripción del mundo natural llevaba acompañada una consecuencia, tan ingrata como imprescindible:

la de la metódica revisión de los especímenes guardados
El pez sapo psicodélico (Histiophryne psychedelica), descrito en 2009, es una muestra de cómo las especies que siguen descubriéndose en la actualidad pueden ser tan insólitas como en el pasado. (Fotografía: Wikicommons)
en herbarios y museos para, eventualmente, sintetizar los criterios de los estudiosos anteriores y reunir varias especies en una sola. A menudo se ha destacado la contraposición de estas dos almas de la taxonomía, la de los analíticos (splitters o “fragmentadores”) y la de los sintéticos (lumpers o “amontonadores”). Los críticos sacan a relucir con frecuencia lo difícil y tortuoso que es el camino que lleva a un consenso entre taxónomos (a veces inalcanzable), pero aunque obviamente sobran ejemplos de especialistas cuyos criterios resultaron ser erróneos, ambas actitudes son consecuencias de la envergadura y dificultad del trabajo a realizar. Aunque la labor de los taxónomos fue incesante e ininterrumpida, con el paso del tiempo esta tarea concienzuda perdió reconocimiento frente a otras dedicaciones emergentes que iban más allá de una simple descripción sistemática de la flora y la
fauna, como la ecología, la fisiología o la genética. No faltaron quienes desdeñaban esta empresa y la comparaban al mero coleccionismo de sellos, supongo que con el permiso de los aficionados a la filatelia. entrañar las afinidades de los organismos. Los comienzos de su uso generalizado se enfrentaron a menudo con la desconfianza de los taxónomos “de toda la vida”, pero por suerte podemos empezar a decir que esa dualidad es cosa del pasado. La otra gran aportación que la tecnología ha
Pese a todo, los exploradores y descriptores de dado en tiempos recientes a esta disciplina tricentenaria
nuevos organismos nunca cesaron de realizar su (a mees internet. Internet, de hecho, parece hecho a medida
nudo incomprendido) trabajo, pero la popularización repara el trabajo del taxónomo por su capacidad para hacer
ciente del neologismo “biodiversidad” a finales del siglo accesible una cantidad ingen
XX tuvo como consecuencia una te de información y la facili
revitalización y popularización de dad con la que las imágenes
la taxonomía. De repente, los erpueden difundirse por todo el
mitaños de museo que examinaban hasta el más mínimo detalle mundo de forma inmediata. de las glumas de una gramínea De repente, bases de datos o de las escamas de un pez, eran inmensas, nomenclaturales, estudiosos de la flamante palabra. de especímenes y de sus disOtras disciplinas biológicas o amtribuciones se vuelcan al albientales comenzaron a reconsicance de todos, y el estudio derar la importancia de esta labor. de la biodiversidad específica Conocer la biodiversidad específica se hace inmensamente más era importante en la medida en la sencillo y eficaz. Las publique las especies cumplen roles incaciones sobre especies del sustituibles en el correcto funciootro lado del mundo pueden namiento de los ecosistemas y por consultarse al instante y cada lo tanto en los servicios que éstos vez más instituciones ponen brindan para mantener la habitabia libre disposición imágenes lidad del planeta y la renovación de de los especímenes tipo a alta sus recursos. El segundo gran emresolución. Basta con recurrir, pujón reciente que ha recibido la Espécimen tipo de una saxifragácea, Heuchera woodsiaphila, descrita recientemente en Nuevo por ejemplo, a tropicos.org, exploración de la biodiversidad ha México. Los especímenes tipo depositados en el portal del jardín botánico venido de la mano de la tecnoloinstituciones científicas sirven para mantener la de Missouri, especializado en gía. Por una parte el desarrollo de correcta asignación de los nombres científicos. (Fotografía: Patrick Alexander -Creative Comnomenclatura y bibliografía, la filogenia molecular ha puesto al mons) que alberga más de un millón alcance de los científicos un critede nombres botánicos y rerio poderoso para contribuir a desgistra casi cuatro millones de


El sifaka diademado (Propithecus diadema) y el sifaka de Milne-Edward (P. edwarsii) estaban considerados hasta hace poco pertenecientes a la misma especie. En los últimos años el número de especies reconocidas de lémures casi se ha triplicado. (Fotografía: Rafael Medina) sin saber cuántas especies nos faltan por describir, la ma
especimenes; o FishBase, la referencia en Internet para la ictiología, que pone la información de más de 32.000 especies de peces bajo una licencia Creative Commons. Sinceramente, me gustaría ver qué cara pondría un naturalista del siglo XIX si pudiera verlo. ¿Reconocería a los profesionales de 2011 como colegas de su misma disciplina?
El perfil actual de los taxónomos, lógicamente, yoría de las estimaciones parecen indicar que aún queda
ha cambiado bastante y con razón. En este momento hay un largo camino por recorrer hasta poder responder esta
descritas aproximadamente 1.2 millones de especies [1], pregunta [1]. Pero, ¿dónde y cómo se encuentran en la
frente a los pocos miles de los comienzos de la taxonomía. actualidad las especies nuevas?
cernir cuándo el ejemplar que tenemos enfrente es una novedad científica, el nivel de especialización de los profesionales en grupos de organismos cada vez más concretos se ha ido incrementando con el tiempo. La otra diferencia es que quienes exploran y describen nuevas especies son, cada vez más frecuentemente, equipos multidisciplinares compuestos por varias personas con distintas tareas, un reflejo de cómo la descripción de nuevas especies es cada vez una tarea más polivalente [2]. Lo que desde luego se mantiene totalmente vigente entre los especialistas actuales y sus antecesores es la fascinación y el impulso por desvelar organismos ocultos y desconocidos, por dejarse sorprender y seducir. El mundo ya no es ese planeta inmenso lleno de lagunas en los mapas, con manticoras y unicornios, pero las satisfacciones y sorpresas que aporta la descripción de nuevas especies sigue siendo igual de estimulante para quienes lo consiguen. Aunque seguimos
Para conseguir llegar al nivel de detalle que permite dis
Una primera respuesta, quizá la más intuitiva, es que hay que irse a lugares muy remotos e inexplorados para encontrar especies nuevas, y es parcialmente cierta, porque de hecho siguen existiendo parajes inaccesibles al ser humano. Regiones como las fumarolas volcánicas, a miles de metros de profundidad en los océanos, sólo empezaron a explorarse en los años 70 del siglo pasado, descubriendo ecosistemas capaces de maravillarnos más aún que un ornitorrinco en el gabinete de un naturalista victoriano: gusanos de varios metros de longitud con
anatomías nunca vistas, peces y cangrejos de formas increíbles que serían capaces de mantener su vida normal aunque el mismísimo sol se apagase ¡y apenas se conoce una pequeñísima fracción de los fondos abisales! Otro ejemplo por supuesto, se encuentra en las pluvisilvas tropicales: regularmente podemos leer noticias incluso en la prensa generalista de equipos científicos que visitan estas zonas del planeta y permiten la descripción de docenas o incluso cientos de especies nuevas.
Sin embargo, en realidad no es necesario irse a lugares tan remotos para hacer tales descubrimientos. Muchas de las especies de nueva descripción estaban aquí, delante de nuestras narices, pero era necesario un nuevo enfoque para darnos cuenta. Tomemos por ejemplo el murciélago común (Pipistrellus pipistrellus), que hasta hace unos años no albergaba confusión posible de identificación. Cuando, en lugar de atender sólo al aspecto, se estudiaron los sonidos que emitían estos animales para relacionarse y percibir su entorno, se llegó a la conclusión de que dos linajes distintos compartían el territorio. Cada uno de ellos empleaba para sonidos de distinta frecuencia. Puesto que estos sonidos tienen relevancia a la hora del apareamiento, las distintas frecuencias convierten a estos linajes en dos grupos reproductivamente aislados y que por lo tanto pertenecen a especies distintas: Pipistrellus pipistrellus (con un reclamo a 45 kHz) y P. pygmaeus (que emitiría sonidos de 55kHz). Igualmente, hace sólo unas semanas asistíamos a la descripción de Tursiops australis, una nueva especie de delfín australiano, para cuyo reconocimiento formal se han empleado tanto datos anatómicos como genéticos y biogeográficos [3], poniendo de manifiesto una vez más que la descripción de nuevas especies debe proceder de una sinergia entre distintos campos.
Todos estos ejemplos tienen en común que las nuevas especies, no es que fuesen totalmente desconocidas hasta ahora, sino que habían pasado desapercibidas hasta la fecha por su gran semejanza con sus parientes cercanos, de las que eran virtualmente indistinguibles. Este tipo de especies suelen conocerse como especies crípticas [4]. El papel de las especies crípticas en la estimación de la riqueza de la biosfera es muy controvertido. Existen indicios que nos hacen pensar que este fenómeno puede estar mucho más extendido de lo que pensamos hasta el punto de afectar significativamente las, aún desconocidas, cifras globales [5]. Otra forma de verlo es que la filogenia molecular es la puerta de entrada a un criterio demasiado “fragmentador”. Por ejemplo, se ha llamado la atención de que en apenas unas décadas, las especies reconocidas de lémures de Madagascar han pasado de 36 a un centenar [6]. Esto se debe sin duda a un mejor conocimiento del área y a la aplicación de técnicas de filogenia molecular, pero ¿existe una “inflación taxonómica”? A buen seguro esto ocurre en más de un caso. Un ejemplo flagrante de esta situación corresponde a las orquídeas mediterráneas del género Ophrys, cuya infinidad de atractivas variaciones ha recibido muchísima atención, hasta el punto de que la divergencia de criterios entre taxónomos “fragmentadotes” y “amontonadores” es extrema: entre 32 y 250 especies, según distintos criterios. Estudios
recientes, que combinan tanto datos filogenéticos como de ultraestructura microscópica, sugiere que la cifra más exacta debe estar más cercana al primer valor que al segundo [7]. Muchos autores sostienen que lo que ocurre es que se mira con demasiado detenimiento las diferencias entre individuos de grupos muy idiosincrásicos y vistosos, como los primates o las orquídeas, mientras que otros, mucho menos llamativos (como hongos, gusanos o musgos), no reciben tanta atención [4],[8].
Lo cierto es que aunque los debates entre distintos criterios continúan, los grupos como los mencionados, que son morfológicamente más austeros, no reciben pro-
porcionalmente la atención que merecen y son un campo abonado para la descripción de muchas especies nuevas. De hecho, especializarse en organismos o ambientes poco estudiados es otra de las formas de explorar la biodiversidad específica desconocida con grandes probabilidades de dar con algo nuevo. Los diminutos intersticios entre las partículas de la arena de los fondos marinos es un universo de invertebrados imposibles, algunos de ellos (como los loricíferos o los quinorrincos) parecerían sacados de películas de ciencia ficción si fuesen más grandes, y a buen seguro son sólo una parte de todo lo que queda por conocer. ¿Cómo enfrentarse a esa riqueza oculta que es, si cabe, más escurridiza aún? Es obligado hacer una mención especial a iniciativas como Barcode of Life que aspiran a que la identificación de los organismos se haga de forma rápida y eficiente mediante la secuenciación de fragmentos escogidos de su genoma, que previamente se habrían reconocido como capaces de actuar como un “código de barras” fiable [9], aunque ni siquiera esta aproximación está exenta de críticas [10].
Por último, es fundamental no olvidarse de que uno de los lugares más habituales donde se descubren especies nuevas en la actualidad siguen siendo los museos, herbarios e instituciones científicas. El trabajo de campo que realizaron y realizan los investigadores es sólo una pequeña parte del tiempo invertido en la descripción de la biodiversidad específica, y generalmente nunca es el factor limitante. De hecho, es muy habitual que el legado de un científico incluya materiales recolectados pero nunca examinados por falta de tiempo. Además, y como es lógico, nadie está exento de equivocaciones, por lo que las identificaciones incorrectas son también muy abundantes. La meticulosa revisión de las colecciones históricas de todo tipo de instituciones es una labor muy ardua pero que aporta también recompensas, especialmente cuando es posible aplicar una perspectiva que no existía en el momento de la recolección.
La historia de la descripción de los organismos de nuestro planeta es muy larga, pero aún tremendamente incompleta. Han sido necesarios más de trescientos años para alcanzar el estado actual. Se trata de una tarea titánica para la que son necesarias las carreras científicas completas y la dedicación de miles de especialistas de todas las épocas. Dado que éstos no siempre van a compartir criterios, es normal que los debates entre analíticos y sintéticos hayan copado una parte de esta actividad, dando cierta imagen caótica. Sin embargo, los avances tecnológicos nos ofrece la posibilidad de poder aproximarnos a la tremenda riqueza de la biosfera desde nuevos puntos de vista que permiten llegar a resoluciones mucho más robustas. Por suerte, persiste el entusiasmo por descubrir nuevas especies al igual que en tiempos de Linneo, como persiste también la capacidad de la naturaleza por asombrarnos con formas desconocidas, incluso en los lugares más insospechados.
Referencias
[1] Mora et al. (2011). How Many Species Are There on Earth and in the Ocean?
[2] Joppa et al. (en prensa). The population ecology and social behaviour of taxonomists
[3] Charlton-Robb et al. (2011). A New Dolphin Species, the Burrunan Dolphin Tursiops australis sp. nov., Endemic to Southern Australian Coastal Waters
[4] Bickford et al. (2007). Cryptic species as a window on diversity and conservation [5] Trontelj & Fišer (2009). Cryptic species diversity should not be trivialised
[6] Tattersall (2007). Madagascar’s Lemurs: Cryptic diversity or taxonomic inflation?
[7] Bradshaw et al. (2010). Comparative labellum micromorphology of the sexually deceptive temperate orchid genus Ophrys: diverse epidermal cell types and multiple origins of structural colour
[8] Fernandez et al. (2006). Cryptic species within the cosmopolitan desiccation-tolerant moss Grimmia laevigata [9] Savolainen et al. (2005). Towards writing the encyclopaedia of life: an introduction to DNA barcoding [10] Will & Rubuinoff (2004). Myth of the molecule: DNA barcodes for species cannot replace morphology for identification and classification
Rafael Medina es licenciado en Ciencias Biológicas por la Universidad Autónoma de Madrid, donde está terminando su tesis doctoral sobre taxonomía y filogenia molecular de briófitos epífitos. Observador y fotógrafo aficionado de la naturaleza, de vez en cuando incluye textos divulgativos sobre el estudio y la conservación de la biodiversidad en su blog personal, "Diario de un copépodo".

Por José Manuel Echevarría
PRÓLOGO 1. HECHOS QUE DEFINEN LA VIDA
La decepción de un peregrino Células, metabolismo y entropía
Aunque la virología ha sido el ámbito de mi ejercicio profesional incluso desde antes de terminar de licenciarme en bioquímica, hace ya treinta y seis años, lo cierto es que siempre he ejercido la profesión muy pegado a los problemas de salud y bastante alejado de las cuestiones más básicas de la materia. No obstante, la pregunta que encabeza estas líneas siempre me provocó, y creo que esa provocación contribuyó a acrecentar mi interés por las cuestiones relativas al origen y la diversificación de la Vida en general.
Hace ya mucho tiempo que, intuitivamente, me decanté por una respuesta afirmativa, pero nunca sentí el impulso de analizar y exponer racionalmente los fundamentos de mi respuesta hasta que no leí –gracias al amable regalo de un buen amigo- el libro titulado The Ancestor’s Tale: a Pilgrimage to the Dawn of Life, publicado en 2004 por el excelente y polémico experto en biología evoluti-excelente y polémico experto en biología evolutiva Richard Dawkins. En él, los seres humanos llegamos a Canterbury –como los peregrinos de Chaucer- acompañados por los seres vivos que se nos han ido sumando en nuestro camino retrógrado hacia el origen de la Vida, desde el resto de los hominoideos actuales hasta la más pequeña de las bacterias. Peregriné con entusiasmo durante días, alegre de conocer a los ancestros que me unen a los seres vivos con los que comparto el planeta, pero cuando llegué allí no encontré, para mi sorpresa, nada más sencillo o primitivo que una célula convencional con un ácido desoxirribonucleico (ADN) dentro. De mis queridos virus no había ni rastro; no me los topé por el camino, ni estaban tampoco, como yo intuía, esperándome en Canterbury. Y entonces caí en la cuenta, no sin sorpresa, de que quizá Dawkins no pensaba que los virus fuesen seres vivos, o que tal vez no deseaba reconocerlo públicamente.
Aunque habrá quien comparta -intuitiva o razonadamente- esa opinión, pienso que habrá también muchos que se sorprendan al saber que la pregunta en cuestión ha desatado en la comunidad científica una intensa polémica que continúa plenamente viva a día de hoy. Decidir si los virus son o no seres vivos requiere escudriñar el concepto de Vida hasta desentrañar sus fundamentos más íntimos, llegando en último extremo a elaborar una definición bien asentada que permita diferenciar con claridad lo que consideramos vivo de lo que es inerte. Desde que el físico alemán Erwin Schröedinger publicase en la década de 1930 su libro titulado ¿Qué es la Vida?, este ejercicio ha devorado, sin embargo, auténticos ríos de tinta sin generar por ahora el consenso necesario para lograrlo. Y en gran medida, los virus son los culpables. Todos los seres que llegaron conmigo al Canterbury de Dawkins compartían una propiedad común. Bien fuera en solitario o en apretado apiñamiento, todos estaban hechos, como yo, de células. Los últimos en sumarse a la peregrinación –los de origen más antiguo- eran todos unicelulares y mostraban su material genético apenas condensado en un rincón del protoplasma, característica distintiva de la célula procariota. Las bacterias y las arqueas son los representantes de esta modalidad celular en el mundo actual. Entre el resto los había unicelulares y pluricelulares, pero en todos los casos encerraban su material genético en una estructura que conocemos como núcleo, que es lo que distingue por encima de cualquier otra cosa a la célula eucariota.
El diseño eucariota es el diseño exclusivo en el mundo que podemos ver con el ojo desnudo, esto es, el de los llamados organismos superiores. El genoma de los seres que responden a ese diseño está siempre constituido por moléculas de ADN de doble cadena estructuradas en esa forma de doble hélice que Watson y Crick describieron en 1953. Al margen de esta diferencia, cualquier célula posee una serie más o menos compleja de estructuras destinadas a gestionar un complicado tráfico de reacciones químicas, meticulosamente acopladas entre sí, que agrupamos bajo el nombre de metabolismo. El entorno en el que tiene lugar el metabolismo celular es un espacio de aspecto granulado que llamamos citoplasma. En la célula procariota, no se distinguen a primera vista grandes estructuras en él. En la eucariota, está ocupado por estructuras membranosas complejas, entre las que destacan unos saquitos cerrados y profusamente compartimentados que hemos dado en llamar mitocondrias (“hilos pequeños”).
La energía que alimenta el metabolismo celular proviene en último extremo del intenso gradiente energético generado por el Sol, que es degradado primero por las plantas y las algas verdes mediante la fotosíntesis y, sucesivamente, por los miembros de una larga cadena trófica que queman con lentitud los compuestos orgánicos que almacenaron en sus enlaces químicos la energía solar capturada. Dicho gradiente, que involucra un alto grado de orden en la energía, es pues degradado por el conjunto de los seres vivos, que contribuyen con ello a generar entropía y a dar cumplimiento a la segunda ley de la termodinámica. Como ya señaló muy bien Schröedinger hace ochenta años, la Vida es, por consiguiente, un proceso termodinámicamente favorecido, y su existencia no sería el fruto de un azar improbable, sino la consecuencia necesaria que
deriva de su enorme capacidad para degradar gradientes de energía, esos gradientes que la naturaleza aborrece y la termodinámica proscribe. Dado que las estrellas generan por doquier tales gradientes, la Vida debiera existir, por imperativo termodinámico, en todos los lugares del Universo en los que las circunstancias lo permitan.
La energía solar llega a los seres vivos no fotosintéticos a través de la alimentación, y su tráfico está sometido a una estricta gestión que se confía a las mitocondrias. Y sucede que, a la postre, esos orgánulos subcelulares gestores de la energía han resultado ser células procariotas que viven en simbiosis con la célula eucariota, algo así como bacterias adaptadas a vivir en el interior de las células de las algas, las plantas, los hongos, los protozoos y los animales. Poseen su propio material genético y una historia evolutiva compartida con la de sus células simbiontes, pero conservan su propia identidad. El esclarecimiento de la naturaleza de las mitocondrias llevó a Lynn Margulis a enunciar su hipótesis de la endosimbiosis como explicación para el nacimiento y el desarrollo de la vida celular. Si consideramos que los cloroplastos de las células vegetales presentan características similares a las de las mitocondrias, la hipótesis de Margulis parece la explicación más plausible para la aparición de la célula eucariota. ¿Son, por consiguiente, las mitocondrias y los cloroplastos seres vivos? Si la respuesta fuese afirmativa, ¿son seres vivos pertenecientes al linaje de las bacterias actuales?; ¿o quizás haya que desplazar el Canterbury de Dawkins hacia atrás en el tiempo hasta llegar a encontrarnos con un ancestro compartido con mitocondrias y cloroplastos?
Interesantes preguntas que habrán de considerarse más adelante.
Por encima de esas nada despreciables diferencias, procariotas y eucariotas comparten otra característica estructural esencial: su citoplasma está repleto de unos diminutos orgánulos que llamamos ribosomas por estar hechos de ácido ribonucleico (ARN). Y esto nos lleva un poco más allá. como expresión génica. En esencia, es un proceso de copia del mensaje en diferentes lenguajes cuyo resultado final es la fabricación de multitud de proteínas que entran a participar en la gestión del metabolismo celular. El resultado de esa gestión es lo que hace que una neurona sea una neurona y trabaje como tal; que un hígado tenga la forma de un hígado y funcione como un hígado; y que un ratón parezca un ratón y se comporte como un ratón. Y entre el ADN y las proteínas, absolutamente todo lo que hay es ARN. La secuencia del ADN se transcribe a una secuencia de ARN, y ese ARN mensajero dirige la traducción del mensaje a una secuencia de aminoácidos específica de la proteína que ese gen codifica. El proceso sucede en la superficie de los ribosomas, que están hechos de ARN ribosomal. Y los aminoácidos que se van insertando en la cadena llegan hasta allí transportados por unas moléculas de ARN llamadas de transferencia. El ADN custodia la información, pero es el ARN quien la ejecuta, y son las proteínas quienes la ponen a trabajar convirtiéndola en formas, funciones, aptitudes y comportamientos.
En las últimas décadas hemos aprendido que la expresión de los genes está, sin embargo, sujeta a una regulación estricta que hace que se expresen sólo en el momento oportuno y en el lugar adecuado. Todas nuestras células poseen un genotipo idéntico, pero su fenotipo es, sin embargo, muy diferente en función de cómo se regule en cada una de ellas la expresión génica. Algunos de los factores que participan en esa regulación se hallan codificados en la propia secuencia del ADN, pero hoy sabemos que otros son ajenos a ella, ya que consisten en sencillas modificaciones químicas del propio ADN o de las proteínas que se asocian íntimamente a él en los cromosomas (las histonas). Esas modificaciones pueden suceder en respuesta a cambios en las condiciones ambientales, y se transmiten a la siguiente generación cuando afectan también a los cromosomas de las células germinales. La joven ciencia que estudia esas modificaciones ha recibido el nombre de Epigenética, y sus descubrimientos han dado en apoyar algunos aspectos de la primera teoría evolutiva que se formuló con seriedad.
La identidad de cualquier especie viva reside en su material genético. Son esas moléculas de ADN las que dicen que un ratón es un ratón, y las que hacen que un ratón y una ratona engendren siempre ratoncitos, nunca otra cosa. ¿Qué cómo lo hacen? Pues bien, en realidad no hacen casi nada. Simplemente se limitan a almacenar un mensaje codificado y a transferir ese mensaje a quienes se encargarán de ejecutar sus instrucciones. El mensaje en sí mismo es lo que conocemos como genotipo y sólo puede conocerse leyendo directamente el ADN. El resultado de la ejecución completa del mensaje es el fenotipo, y buena parte de él salta a la vista: ratón o ratón, hay poco lugar a dudas. Se trata de la Teoría del Medio, que fue desarrollada en el Museo Nacional de CienciasNaturales de París por Jean Baptiste Lamarck a finales del siglo XVIII.
Mutación y diversidad: ¿somos hijos del error?
Si la identidad de una especie reside en su material genético, la conservación de esa identidad a lo largo del tiempo, que es esencial para que una especie pueda considerarse tal, reside en la fidelidad con la que ese material se transfiera a la descendencia. El ADN se copia cada vez que una célula se divide. En un ser unicelular, esa división equivale a la reproducción. El proceso de copia del ADN se conoce como replicación, y sucede merced a la actuación de unas proteínas llamadas polimerasas de ADN dependientes de ADN, que llamaremos por abreviar ADN pol. La replicación del ADN es un proceso complejo, y las ADN pol poseen varias funciones diferentes para llevarlo a cabo. Una
de esas funciones consiste en reparar los errores cometidos. Se lee la copia según se va haciendo, y cuando se detecta un error, se elimina y se corrige. Si la replicación fuese siempre fiel al ciento por cien, todos los descendientes heredarían un genotipo idéntico, que se expresaría como único fenotipo; y si todos sus miembros fuesen siempre idénticos unos a otros, las especies serían inmutables. Pero eso no casa con la realidad, aunque la Historia nos muestre que nos haya costado siglos aceptarlo.
En realidad, la fidelidad de la replicación del ADN mediada por las ADN pol no es nunca del ciento por cien, sino que acumula errores con una frecuencia media de 10- 10, es decir, un error por cada diez mil millones de letras de mensaje copiadas. Estos errores se denominan mutaciones, y el fenotipo resultante de la expresión del genoma mutado se conoce como variante. Aunque parezca muy baja, lo cierto es que esa tasa de error es suficiente para generar, por acumulación, descendencias genéticamente diversas que muestren diferencias individuales en su capacidad de adaptación a los cambios ambientales. Este hecho constituye la base de la evolución de la Vida, al hacer posible que las especies no sean inmutables. entre ellas. Lejos de ser la especiación un fenómeno discontinuo, se trata de un proceso continuo que está permanentemente en marcha en todas las especies, aunque pueda tomar mucho tiempo y sólo a veces culmine en la aparición de especies nuevas. Hasta la llegada de la genética molecular, hace apenas medio siglo, se consideraba imposible observarlo directamente, aunque eso no fuese del todo cierto. Para los microbiólogos actuales, capaces de leer con todo detalle el mensaje contenido en los sencillos genomas de esos diminutos seres de reproducción vertiginosa, observar la Evolución en directo es, por el contrario, un asunto cotidiano.
Llegados hasta aquí, formularé dos proposiciones que tratan de definir lo que yo entiendo que son los seres vivos y las especies:
- Un ser vivo terrestre es un organismo poseedor de un genoma que:
1. Está constituido por un ácido nucleico;
2. Es capaz de mantener su identidad y un linaje evolutivopropio;
Como veremos a continuación, la Vida ha inventado otras formas más eficaces de diversificación genética dentro de las especies, pero la mutación fue antaño, con toda probabilidad, la única causa generadora de diversidad entre los primeros seres vivos terrestres; y es, por consiguiente, la responsable última de la diversidad que la Vida muestra hoy en nuestro planeta. Toda vez que nosotros somos parte integrante de esa diversidad, también somos, en esa misma medida, hijos del error, ya que jamás hubiésemos llegado a existir sin él.
Recombinación genética, sexo y evolución
Pero llegó un día en que sucedió que los seres vivos sencillos que habían evolucionado por mutación y selección hacia genotipos y fenotipos diversos empezaron a hacer algo insólito. De hecho, empezaron a intercambiar entre sí fragmentos de información genética, creando, en un único y breve acto, descendencias enormemente diversas. Ese proceso recibe el nombre de recombinación, y sucede en todos los seres vivos, con independencia de su complejidad. En los seres unicelulares, el acto que dispara el proceso se llama conjugación. En los seres vivos superiores se llama fecundación y tiene que ver, obviamente, con el sexo. Es precisamente el sexo el mecanismo de recombinación que genera la máxima diversidad intraespecífica, y es por eso que la diferenciación en sexos ha tenido un éxito espectacular sobre la Tierra.
La teoría de la evolución por selección natural de Darwin y Wallace se basa en esa diversidad que subyace en las poblaciones de las especies, y explica la aparición de nuevas especies a través de dos fenómenos: la presión selectiva que ejerce el medio ambiente sobre las variantes y el eventual aislamiento reproductor que pueda surgir 3. Evoluciona en interacción permanente con los genomas de otros seres afines cuyos mensajes son semejantes al suyo;
y 4. Participa en el ciclo de la energía contribuyendo a aumentar la entropía del Universo con su expresión.
- El colectivo formado por un conjunto de seres vivos cuyos genomas interaccionan y evolucionan en un linaje evolutivo común definen la identidad genética y fenotípica que llamamos especie.
Me responsabilizo plenamente de estas proposiciones, aunque apunto que hay en ellas mucho de lo que el microbiólogo francés André Lwoff expresó en relación al concepto de la Vida hace más de cuarenta años. La esencia de la Vida no reside, pues, únicamente en la estructura celular y en el metabolismo, sino más bien en el mensaje genético en sí y en su capacidad de proyectarse hacia el futuro en cambio permanente, pero conservando su identidad. Y los virus son, como veremos, mensajes genéticos en estado virtualmente puro que evolucionan, como cualquier otro, sobre la base de la mutación y la recombinación y según las reglas de la selección natural, interaccionando con su medio ambiente y entre sí según los usos y costumbres propios de las especies.
2. LOS VIRUS EN VIVO
Genomas que andan sueltos
He dedicado buena parte de mi tiempo de trabajo durante los últimos años a un interesante virus que está últimamente de moda entre los que nos ganamos la vida con esto, el virus de la hepatitis E. La partícula de este virus –el virión, en la jerga científicaconsta de una molécula
de ARN encerrada en un estuche –la cápside- de simetría icosaédrica formado por el ensamblaje de unidades idénticas de una única proteína. La misión de esa proteína es proteger el genoma de las agresiones del medio ambiente, adherirse a la membrana de la célula en la que el virus va a reproducirse, e interactuar con ella hasta lograr que la cápside llegue al citoplasma. Una vez allí, se desintegra y se libera el genoma viral. A partir de ese momento, ese genoma toma el mando y dirige el metabolismo celular hacia la fabricación masiva de nuevas partículas víricas. Cuando se han acumulado ya miles de ellas en el citoplasma del hepatocito infectado, la célula estalla y las partículas se liberan para infectar nuevas células y propagar así la infección por el hígado del paciente. Por consiguiente, el virus participa decisivamente en los procesos metabólicos de las células infectadas, aunque carezca de una maquinaria metabólica propia.
En esencia, y pese a una muy notable diversidad en los detalles, todos los virus hacen algo parecido a eso cuando infectan a su hospedador, y el fin de esa estrategia no es otro que producir partículas capaces de llevar eficazmente sus genomas al interior de otras células en las que puedan reproducirse, esto es, llevar a cabo su replicación. Aquí no hay pues fabricación de arquitecturas muy complejas, ni metabolismo, ni sexo, ni ningún otro adorno o recurso, tan sólo genomas minúsculos (millones de veces más pequeños que el nuestro) empeñados en la tarea que es propia del genoma de cualquier ser vivo, que no es otra que la de perpetuarse y sobrevivir con la máxima economía de recursos en competencia con otros.
La expresión de los genomas de los virus sucede exactamente igual que la del genoma celular, aunque algunos introducen una necesaria y única modificación. A diferencia del resto de los seres vivos, muchos virus poseen genomas hechos de ARN en lugar de ADN. Las células no poseen ningún enzima que sea capaz de copiar ARN a partir de un molde de ARN (esto no es rigurosamente cierto, pero a los efectos de estas reflexiones lo daremos por bueno), así que todos esos virus fabrican unos enzimas que sí son capaces de hacerlo. Se llaman polimerasas de ARN dependientes de ARN, y les llamaremos ARN pol. Las ARN pol fabrican ARN mensajero copiando el mensaje escrito en el ARN genómico, y a partir de ahí la expresión génica sucede según el mecanismo general. Además, ese mismo enzima participa también en la replicación del genoma vírico.
La replicación de los genomas víricos constituidos por ADN corre siempre a cargo de una ADN pol propia del virus, nunca por una ADN pol celular (de nuevo, soslayemos alguna excepción ya conocida). La fidelidad de copia de estos enzimas es similar a la de sus homólogos celulares, y la tasa de mutación que originan está también en el orden de 10 -10 . No sucede lo mismo, sin embargo, con la replicación de los genomas víricos constituidos por ARN. Las ARN pol no saben detectar y reparar sus errores, de forma que todos los que cometen permanecen en la copia. La tasa de error se dispara así hasta 10 - 6 , esto es, diez mil veces más que la anterior. Esos virus generan, por tanto, variantes a una velocidad de vértigo. Pero no terminan ahí las sorpresas. Algunos virus (no muchos) replican su genoma a través de una copia intermedia que se realiza copiando ARN en ADN. Fabrican para ello unos enzimas que se llaman transcriptasas reversas (RT), ya que hacen exactamente lo contrario de lo que sucede en la transcripción durante la expresión génica. El paso de transcripción reversa mediado por las RT sólo existe en esos virus, carece también de corrección de errores, y la fidelidad final de replicación que proporciona es del orden de 10 -8 , cifra intermedia entre las dos anteriores. El descubrimiento de las RT valió a Howard Temin, David Baltimore y Renato Dulbecco compartir el Premio Nobel de Medicina en 1975.
El ARN, los viroides y el origen de la Vida
Aunque su impacto no fue, ni de lejos, el de El Origen de las Especies de Darwin, la publicación al comienzo de la década de 1960 de la Teoría Evolutiva del Origen de la Vida por Alejandro Oparin merece figurar, en mi opinión, en el registro de los auténticos momentos estelares de la Ciencia. En ella, Oparin se hacía eco de ideas que ya habían expresado antes personajes como John Haldane y François d’Herelle, y postulaba que el primer ser vivo tuvo que poseer un genoma de ARN. En aquéllos tiempos de la Guerra Fría, ser ruso y trabajar en la URSS no era buen asunto para la proyección internacional de un científico, así que esas circunstancias, sumadas a lo revolucionario y aún poco sustentado de su propuesta, privaron a Oparin de un premio Nobel a mi juicio más que merecido.
La oposición a la hipótesis de un mundo vivo primigenio basado en el ARN se sustentaba en dos fuertes pilares. Primero, la inestabilidad del ARN, que se rompe con extrema facilidad por las agresiones del medio ambiente; segundo, y al no poder conformar cadenas muy largas debido a esa inestabilidad, la escasa probabilidad de que hubiese podido formarse y persistir algún ARN lo suficientemente grande como para llegar a fabricar una ARN pol que hiciese posible su replicación. Hubo que esperar casi un siglo para que la genética molecular viniese a afianzar los cimientos de la teoría de Darwin, y él nunca llegó a verlo. La de Oparin sólo necesitó poco más de veinte años para verse respaldada por hechos, aunque por desgracia él no lo vio, ya que murió poco antes de que se realizase el asombroso descubrimiento que explico a continuación.
Los cultivos de árboles frutales se ven afectados por muchas enfermedades infecciosas, y no pocas responden a infecciones víricas. Algunas de estas últimas traían algo desconcertados a los ingenieros agrónomos en la década de 1970, ya que estaban producidas por unos virus ARN que, aparentemente, se reproducían sin originar ninguna proteína propia y sin fabricar viriones. El ARN de esos agentes se encontraba en abundancia en los tejidos
vegetales infectados, pero por más que se buscaba nadie lograba detectar la presencia de una actividad de ARN pol en las células. ¿Quién demonios replicaba el ARN de esos misteriosos “virus”?
La respuesta era tan obvia como difícil de aceptar. Ese ARN se replica a sí mismo, es decir, cataliza su propia replicación. Es, a la vez, un genoma y un enzima. Además, tampoco necesita la ayuda de ninguna proteína que lo proteja de las agresiones del ambiente para pasar de una célula a otra y de una a otra planta, ya que se pliega sobre sí mismo en una forma muy compleja que le hace invulnerable, algo así como el erizo que se hace una bola de espinas para defenderse del zorro.
Estos agentes se conocen como viroides, y sus sorprendentes propiedades se descubrieron y demostraron a principios de la década de 1980, época en la que se acuñaron los términos ribozima y ARN autocatalítico para referirse a esas prodigiosas moléculas. Existen veintiocho especies de viroides conocidas en la actualidad, y se agrupan en dos familias atendiendo a su estructura. El esclarecimiento de la auténtica naturaleza de los viroides granjeó a Thomas Cech y Sidney Altman el Nobel de Química en 1989.
Superados los escollos de la estabilidad y la replicación sin ayuda de polimerasas, el mundo vivo primigenio de ARN que propuso Oparin cobra una fuerza irresistible, y los viroides se convierten en la huella probable de ese mundo ancestral, en los seres que más se parecerían en la actualidad a lo que pudo ser el ancestro común al conjunto de la Vida en el planeta. El Canterbury de Dawkins se desplaza, así, mucho más allá de las bacterias, sin duda mucho más allá de los 2.500 millones de años. Pero para poder llegar hasta allí es necesario admitir que los virus están tan vivos como cualquier otro ser vivo; que la Vida no es sólo arquitectura y metabolismo, sino también, y esencialmente, genomas en permanente evolución que luchan sin cesar por la supervivencia. Llama la atención que sea precisamente Dawkins -que planteó esta idea en su forma más radical en El gen egoísta, y que la ha defendido con vehemencia durante cuarenta años- quien haya sido a la postre incapaz de encontrar virus y viroides en su Canterbury particular.
¿Cuánto importa el tamaño?
Por mucho que los ciudadanos ignoren muchas cosas sobre los virus, yo creo que casi todos saben que son muy, muy pequeños. Su carácter “ultramicroscópico”, que los hace visibles sólo bajo el microscopio electrónico, es quizá lo único que todos los profesores de instituto saben decir a sus alumnos sobre los virus. Los virus se mueven, por tanto, en un umbral físico más cercano al de una molécula orgánica gigante que al de un ser vivo, y este ha sido un argumento habitual entre quienes defienden su pertenencia al mundo de “lo inerte” o lo “no vivo”. Pero, ¿es eso verdad? El microscopio óptico llega a duras penas a un nivel de resolución en torno a los 300 nanómetros (nm) (3x10-7mm). Más allá, la longitud de onda de la luz visible es demasiado larga para resolver la imagen, y hay que iluminar el objeto con radiaciones de longitud de onda más corta. Así, utilizando haces enfocados de electrones, algunos trucos, y una pantalla sensible o una película fotográfica, los virus se nos hacen visibles bajo las lentes magnéticas del microscopio electrónico. Pero, ¿todas las bacterias pueden verse bien al microscopio óptico?; y ¿ningún virus puede verse en absoluto usando luz visible?
Si tomamos la bacteria intestinal Escherichia coli como prototipo bacteriano, nos encontramos con una célula en forma de bastón de unos 700x2000nm, un buen tamaño para el microscopio óptico. Los Mycobacterium (los bacilos de la tuberculosis y la lepra) ya son un poco más difíciles, alrededor de 250x1000nm, y los vemos más bien como hilos que como bastones. Un poco más allá están las Rickettsia (por ejemplo, la del tifus exantemático), en el orden de los 200x600nm, y esto ya empieza a verse como una pulguita con nuestro objetivo más potente. Y por último, los Mycoplasma (los agentes de la neumonía atípica primaria genuina), esferas de unos 500nm de diámetro, ya son puntos que imaginamos más que vemos. Pues bien, exactamente al nivel de una esfera de 500nm de diámetro están las partículas de los Mimivirus y los Phycodnavirus, los virus más grandes de entre los conocidos; y un poco más allá, con forma de bastones de 300x80nm de longitud, están las de los Poxvirus, entre los que se incluye el virus de la viruela. A partir de ahí, sólo el microscopio electrónico nos puede enseñar lo que hay más allá.
Así pues, la frontera que marca la capacidad resolutiva de un haz de luz visible no coincide exactamente con la que separa los tamaños de bacterias y virus. Es más, la realidad es que tal frontera, tal discontinuidad, no existe. Puede argumentarse, en todo caso, que una constricción tan fuerte en el tamaño deja poco lugar a la diversificación estructural en el mundo de los virus, y que ese mundo debe ser pequeño, aburrido y monótono. Nada tan lejos de la realidad.
Los virus, a día de hoy
El catálogo del Comité Internacional de Taxonomía de Virus (ICTV) incluye alrededor de 9500 entradas. Los genotipos caracterizados dentro de cada especie se cuentan como entradas independientes, así que no es fácil saber con exactitud de cuántas especies consta el catálogo, aunque pienso que una estimación en torno a las 5000 debe aproximarse razonablemente a la realidad. La contribución relativa de esas especies a la biomasa marina total ha sido estimada recientemente en un 40%, lo que puede dar una
idea de su importante –y seguro que para muchos sorpre dente- contribución a la biomasa total del planeta. Si un extraterrestre de vista “ultramicroscópica” llegase hoy a la Tierra, nos vería a animales y plantas nadando en un
El catálogo del ICTV incluye virus que infectan bacterias, arqueas, algas, plantas, hongos, protozoos y metazoos pertenecientes a todos los phyla, tanto virus ADN como ARN. También incluye las 28 especies de viroides conocidas. La diversidad de formas en los viriones de todos esos virus es amplísima. Pero no termina ahí la lista, ya que desde hace solo tres años sabemos que existen también ¡virus de virus! Se les ha dado el nombre de virófagos, por semejanza con la denominación genérica bacteriófago que se ha utilizado siempre para los virus que infectan bacterias. Los virófagos parasitan las factorías de viriones que los Mimivirus establecen en el citoplasma de las amebas infectadas, se reproducen en ellas e introducen sus viriones en el interior de los viriones de los Mimivirus mientras se forman (Img. 1). Ocultos en ellos, abandonan la ameba infectada para penetrar junto con ellos en el interior de una nueva ameba y continuar reproduciéndose. Resulta interesante saber que esa parasitación puede causar severas deformidades en las partículas de los Mimivirus, y que incluso puede abortar su formación, así que podría decirse que la “infección” por virófagos causa “malformaciones congénitas” y “aborto espontáneo” en esos virus.
El origen de los virus
Cuando se trata de opinar sobre su origen, es frecuente escuchar la idea de que los virus “proceden de sus hospedadores”. La concreción de esa idea consiste en postular que los virus son fragmentos de material genético celular que han logrado escapar de la célula convertidos en organismos parásitos autónomos. Con mucha frecuencia, los virus muestran una especificidad de especie exquisita en cuanto a su hospedador natural, y es en esa propiedad donde se fundamenta la proposición, por lo demás bastante inconcreta en casi todos sus aspectos. Como esto vendría a sugerir que los virus actuales tienen orígenes diversos, inconexos y probablemente recientes, vale la pena pararse un momento a analizar la cuestión. Si consideramos el rango de hospedadores de las especies agrupadas en las diferentes familias de virus conocidas, y lo referimos a taxones biológicos amplios, encontramos cosas interesantes (Tabla 1). Por un lado, que ninguna familia de virus presenta hospedadores entre los Procariota y los Eucariota a la vez. Por otro, que existen familias de virus que presentan hospedadores incluidos en más de una de las tres grandes divisiones de Eucariota consideradas. Esto no sólo es coherente con la existencia de
fenómenos adaptativos compartidos que han definido e identifican linajes evolutivos independientes (familias), sino que sitúa la divergencia de algunos de esos linajes en momentos muy remotos del pasado. Muchos virus parecen, pues, compartir ancestros comunes que hubieron de infectar a especies ancestrales comunes a sus hospedadores actuales. Y eso se llama filogenia, y es una propiedad característica de la evolución de los seres vivos.
Los virus y la Systemata Naturae de Linneo
La clasificación de los seres vivos continúa basándose en el sistema de agrupamiento jerárquico que ideó Carlos Linneo hace más de dos siglos, en el que las jerarquías principales son Reino, Phylum, Clase, Orden, Familia, Género y Especie. Esto es así porque esas jerarquías reflejan relaciones filogenéticas evolutivas entre sus miembros, aunque el bueno de fray Carlos no lo supiese (acérrimo defensor de las especies inmutables, Linneo llegó a admitir en su vejez que quizá sólo los órdenes eran obra directa del Creador). Por el momento, su aplicación al mundo de los virus por parte del ICTV ha tenido un éxito limitado. Resulta bastante completa en los niveles de género y familia, pero aún bastante pobre en las jerarquías superiores. No obstante, me parece que esas carencias reflejan más la timidez -o la excesiva prudencia- de los virólogos que las dificultades reales. Así, y de la misma forma en que Dawkins propone el Súper-Reino Eukarya para englobar a todos los seres vivos hechos de células eucariotas, no veo gran obstáculo para definir un Súper-Reino o un Reino Viria que englobe a los virus ADN y RT. De igual manera podrían definirse phyla o clases para los virus ADN (Desoxiviria) y los virus RT (Retroviria), y también más órdenes que los definidos hasta la fecha. Por lo demás, pienso que a medida que el ICTV vaya admitiendo mejor la filogenia de genomas para clasificar los virus, las propuestas de esa naturaleza quizá se irán fundamentando mejor. La definición de un taxón superior único para los virus ARN no parece posible a día de hoy.
Imagen 1. Siete partículas de un virófago visibles en el interior del virión de un Mimivirus (Nature, 4 Septiembre, 2008/doi:10.1038/nature07218). En 1993, sólo cuatro años después del Nobel de Cech y Altman, Richard Roberts y Phillip Sharp recibieron el codiciado premio por haber encontrado en la naturaleza otras ribozimas al margen de los viroides. Las hallaron al
Nº de Familias con miembros que infectan a No son pocos los virus actuales de animaespecies incluidas en cada grupo les y vegetales que son capaces de integrar su genoma en el de la célula hospedadoProcariotas 18 ra, el virus del SIDA es quizás el ejemplo más popular. Existen también virus así enAlgas y Plantas 19 tre los que infectan a las bacterias, pero en el mundo bacteriano sucede algo aún más
Eucariotas Hongos Metazoos 7 57 extraordinario, ya que involucra a unas moléculas de ADN de vida absolutamente libre: los plásmidos bacterianos. Se trata de unos elementos genéticos autónomos que se encuentran libres en el citoplasma de las Múltiple (>10 9 7 bacterias, pero que pueden integrarse tamaños) bién en su genoma. En su forma libre están constituidos por un ADN circular, y son caTabla 1. Rango de hospedadores de las familias de virus conocidas en la acpaces de replicarse con independencia del tualidad, referido a taxones biológicos muy amplios. El hecho de que siete genoma bacteriano merced a una ADN pol de ellas presenten un rango múltiple al considerar esos tres taxones de los organismos eucariotas indica un origen común que habría de remontarse más de mil millones de años atrás. propia, diferente de la de las bacterias, que ellos mismos fabrican. Se transfieren muy fácilmente entre bacterias de la misma esestudiar los intrones, que son unas misteriosas secuencias pecie por conjugación, y codifican proteínas de ADN que interrumpen los genes de todas las especies importantes para su supervivencia. Entre ellas se encuende seres vivos superiores conocidas, incluida la nuestra. tran los factores de resistencia a antibióticos, esenciales Están, literalmente, por todas partes, y algunas de ellas han para permitir a las bacterias competir con los hongos en la resultado ser muy parecidas a como son los viroides. Naexplotación de los recursos del entorno. die sabe por qué ni para qué están allí. Cuando la bacteria se divide, las células hijas reciEste hallazgo sorprendente –quizás, incluso, inben copias tanto de los plásmidos citoplasmáticos como de quietante- tenía, si embargo, precedentes cercanos. Diez los que se hallan integrados en el genoma. Al igual que las años antes, Barbara McClintock recibió también un Nobelmitocondrias y los cloroplastos nos hablan de fenómenos por desvelar las propiedades de otros misteriosos elemensimbióticos que pudieron estar en el origen de la célula tos genéticos conocidos como retrotransposones. Estos eucariótica, los plásmidos bacterianos parecen contarnos resultaron ser capaces de fabricar una RT –un enzima que una historia similar que podría estar en el origen de las ninguna célula utiliza para nada en su metabolismo-, y el primeras células realmente vivas, aquéllas que, por poseer objeto de ello no es otro que realizar copias de sí mismos ya un genoma, poseían también una identidad propia y un para propagarse dentro del propio genoma del ser vivo mecanismo para conservarla y para adaptarse al medio. Es en el que residen. Parecerá ciencia-ficción, pero los redifícil, sin embargo, evaluar hasta qué punto haya influitrotransposones suponen un cuarenta por ciento de nuesdo en la evolución de la Vida ese trasiego de información tro genoma, y hasta un setenta por ciento del genoma del asociado a la actividad de los ancestros de todos estos maíz. En los ratones, y bajo ciertas condiciones, la actielementos genéticos autónomos que hoy se nos presentan vación de algunos retrotransposones resulta en la producbajo el aspecto de intrones, retrotransposones y plásmidos ción espontánea de virus, precisamente de esos virus en bacterianos. En cualquier caso, la ubicuidad que exhiben los que se descubrieron las RT por primera vez. En pocas sugiere para ellos un origen muy antiguo, y su impresiopalabras, si sumamos los intrones a los retrotransposones, nante abundancia no mueve a pensar que esa influencia resulta que no sabemos para qué sirve -ni porqué está allí, haya sido pequeña. ni a quién pertenece en realidad- más de la mitad del ADN que contienen nuestros genes. ¿Están vivos los virus?
La interpretación más probable para este hecho es que los genomas de los seres vivos superiores han ido acumulando materiales de procedencias diversas a lo largo de su historia evolutiva, y que esos materiales son fiel reflejo de esa historia. Así, parece que ciertos elementos genéticos de vida autónoma, semejantes a los viroides y a los virus RT actuales, se han ido introduciendo con el tiempo en esos genomas, y que lo han debido hacer desde tiempos ancestrales. Esto parecería una especulación excesiva si no fuese porque sabemos, con muchos detalles, que tal cosa sucede comúnmente hoy día entre las bacterias. Pienso que lo visto hasta aquí justifica afirmar que los virus cumplen con los requisitos necesarios para ser considerados seres vivos, y que sus poblaciones definen, además, especies como lo hacen las de cualquier otro, aunque su extrema diversidad intraespecífica desdibuje con frecuencia el concepto. Su insólita capacidad de diversificación y adaptación en respuesta a las presiones del medio ambiente nos ha ayudado mucho a juzgar lo acertado de los conceptos de diversidad y selección natural postulados hace más de un siglo por Charles Darwin, dando así nuevo respaldo a unas ideas que nos permiten hoy profundizar en
Familia
Organismos celulares
Presente en
Virus Elementos genéticos autónomos
ADN pol*
ADN pol B (tipo)
ADN pol B (RG)*
ADN pol C
ADN pol RT
ARN pol Mitocondrias Cloroplastos Rickettsias
Arqueas Eucariotas
Bacterias Clamidias Micoplasmas Virus ADN
Virus ADN
Virus ADN RT Virus ARN RT
Virus ARN Plásmidos bacterianos
Retrotransposones
Tabla 2. Clasificación de las replicasas y presencia de las diferentes familias en los organismos celulares, los virus y los elementos genéticos autónomos. *: Replicasas que trabajan según el mecanismo conocido como rueda giratoria (RG). Este es, además, el mecanismo que usan las ribozimas (viroides y algunos intrones) en su autorreplicación.
más bien sólo ellos, son los seres vivos que mejor nos pueden hablar sobre la biosfera primigenia, ese mundo basado en el ARN que Alejandro Oparin postuló, a mi entender, con muchos visos de acierto pleno.
No es, por consiguiente, un ejercicio descabellado tratar de buscar la manera de encontrar en el Árbol de la Vida el hueco a rellenar con las ramas que corresponden al mundo de los virus. Nos adentramos con ello en el meollo de una polémica plenamente actual, en un terreno aún poco explorado y por completo abierto a la especulación.
Hemos llegado, por consiguiente, al punto más estimulante de nuestras reflexiones, a ese momento en el que la imaginación humana es ya libre de planear sobre los datos y los hechos para tratar de acercarse a lo que quizá pueda ser la verdad.
3. LOS VIRUS Y EL ÁRBOL DE LA VIDA
El cuento de las replicasas
Para establecer las posiciones relativas de los seres vivos en el Árbol de la Vida resulta conveniente –y cada vez más necesario- comparar sus genomas. Esa estrategia nos ha enseñado, por poner un ejemplo llamativo y reciente, que los hipopótamos no pertenecen como pensábamos al linaje evolutivo de los cerdos y los jabalíes, sino al de
las ballenas y los delfines, encarnando así el retorno a tierra firme de un linaje de mamíferos que halló un día su futuro en el mar sin dejar por ello de respirar con pulmones. Sin embargo, esas comparaciones no se realizan utilizando y que estén presentes en los genomas de todas ellas.
Cuando se pretende comparar los virus con otros seres vivos, la extrema sencillez de sus genomas deja muy poco margen de maniobra, pero no es imposible hacerlo. Los genomas de todos los virus presentan genes que codifican replicasas, que es el nombre genérico que engloba a las ADN pol, las ARN pol y las RT. Por su parte, todas las células, tanto procariotas como eucariotas, poseen también genes que codifican replicasas. Así pues, veamos qué sabemos sobre las replicasas y sobre las relaciones filogenéticas que existen entre ellas.
Según nos cuenta Eugene Koonin -experto en biología evolutiva de los Institutos Nacionales de Salud de los EEUU-, se conocen tres familias de ADN pol que se denominan A, B y C (Tabla 2). Las arqueas, las células eucariotas, los virus ADN y los plásmidos bacterianos presentan ADN pol de la familia B; las bacterias presentan ADN pol C; y las mitocondrias, los cloroplastos y las rickettsias, ADN pol A. Además, están las RT, sólo presentes en los virus RT y en los retrotransposones; y las replicasas ARN pol, que solo poseen los virus ARN. Dentro de la familia de las ADN pol B existen también dos variantes que funcionan con mecanismos algo distintos del que usa el tipo.
Una de ellas se conoce como ADN pol B de rueda giratoria, y se encuentra sólo en algunos virus y en los plásmidos bacterianos. Además, conviene saber que el mecanismo de replicación en rueda giratoria es el que utilizan las ADN pol A, y también las ribozimas en su re
La filogenia predice que las ADN pol A, B y RT evolucionaron independientemente a partir de un ancestro común, en tanto que las ADN pol C lo hicieron a partir de la familia B. Además de esto, el virólogo Luis Villarreal, de la Universidad de California en Irving, ha aportado datos que sustentan la hipótesis de que las ADN pol víricas representan la forma ancestral de la familia ADN pol B, precursora de las ADN pol de arqueas y eucariotas. Por desgracia, no parece que haya muchos datos sobre las ARN pol víricas, aunque Koonin sugiere que su aparición fue posterior a la de A, B y RT. En la cabeza del esquema, el autor coloca una hipotética replicasa ancestral que existió en el contexto de un mundo de ARN. La interpretación de este panorama ofrece aspectos muy interesantes. El primero, que las mitocondrias, los cloroplastos y las rickettsias parecen representar un linaje evolutivo muy antiguo e independiente del de las bacterias modernas. Cabe pensar que los ancestros de vida libre de las mitocondrias y los cloroplastos se extinguieron como organismos autónomos, y que sus linajes específicos sólo se prolongaron hasta hoy día en íntima asociación simbiótica con las células eucariotas. El segundo, que los virus RT y los retrotransposones representan un linaje evolutivo muy anterior al de cualquier célula moderna, quizá con la excepción de las arqueas. El tercero, que los virus ADN pertenecen al mismo linaje evolutivo que las arqueas y las eucariotas y preceden a ambas.
Un camino a Canterbury
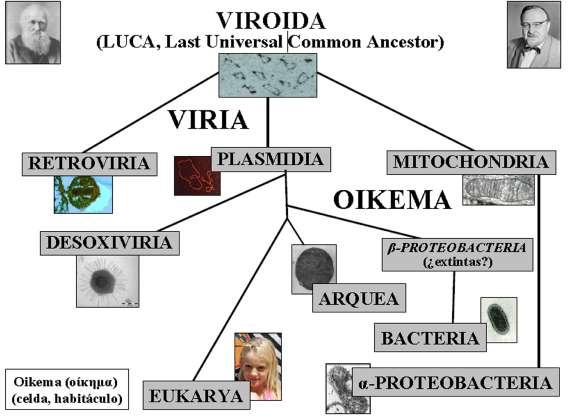
Mi corolario particular al cuento de las replicasas es el esquema de la Figura 1, que pretende representar con cierto detalle la base del Árbol de la Vida. Partiendo de elementos genéticos autónomos semejantes a los viroides actuales, englobados en un único grupo (Viroida), se abren tres ramas que inician dos grandes mundos, el de los virus (Viria) y el de las células (Oikema) (�������οίκημα �: celda, habi- táculo). Una rama entra directamente en este último para generar el linaje de las mitocondrias, los cloroplastos y las α-proteobacterias, el grupo en el que clasificamos a las rickettsias. Las otras dos lo hacen en el primero y originan el linaje de los virus RT y un linaje vírico ancestral dotado de ADN pol B. Este último conduce a los virus ADN actuales, y es también origen de dos linajes adicionales, uno hacia arqueas y eucariotas y otro hacia las bacterias modernas, pasando este último por unas hipotéticas bacterias ancestrales dotadas de ADN pol B(�β-proteobacterias) supuestamente extintas en la actualidad. sa. Se trataría, pues, de un ancestro aún no completamente vírico, de un viroide moderno que ya habría descubierto el ADN y las replicasas, que estaría en la base de los linajes de todos los seres vivos actuales a excepción de las mitocondrias, los cloroplastos, las α-proteobacterias y los virus RT y ARN, y que tendría su descendencia directa en los plásmidos bacterianos, adaptados estos hoy a la simbiosis con las células procariotas como las mitocondrias y los cloroplastos lo están a la vida simbiótica con las eucariotas
Como se puede ver, ninguno de los dos grandes grupos de seres vivos (Viria y Oikema) tiene en este esquema un origen independiente único, sino que hay que remontarse más atrás en él para encontrar ancestros comunes a todos los Viria y todos los Oikema que conocemos hoy. Por otra parte, en tanto que los organismos eucariotas forman un linaje evolutivo único (Eukarya), los procariotas representan tres linajes independientes (α-Proteobacteria, Bacteria y Arquea) que solo encuentran su ancestro común en LUCA (Last Universal Common Ancestor), ya en el corazón mismo de ese nuevo Canterbury.
Por lo demás, los virus ARN permanecen como un enigma a desentrañar, ya que parece que la cuestión de su origen es muy compleja. Es muy probable que presenten orígenes múltiples, y también posible que muchos de ellos sean, realmente, elementos genéticos escapados de células y adaptados a una vida autónoma. Pero también es posible que tal interpretación solo responda a nuestra ignorancia. En todo caso, este esquema ofrece un camino que conduce hasta un Canterbury para mí más real que el Canterbury de Dawkins, toda vez que incluye a esos seres vivos que llamamos virus y termina en otros que se nos presentan como descendientes directos del ancestro común a todos. Dicho ancestro sería un ARN autorreplicante de vida libre y aún no dotado de la habilidad de codificar proteínas.
EPÍLOGO
Estamos hechos de polvo de estrellas
Ignoro hasta qué punto mis especulaciones puedan contener alguna verdad sobre el origen de la Vida, pero pienso que tienen la virtud de integrar las ideas actuales más relevantes al combinar las consideraciones termodinámicas de los seguidores de la endosimbiosis de Margulis con los puntos de vista de Dawkins sobre el protagonismo de la información genética. En cualquier caso, creo que sí son útiles para dar aún más significado a esa hermosa y conocida afirmación del astrofísico Stephen Hawking que encabeza este epílogo.
Pero además, si ubicamos en ese esquema las posiciones que ocupa el mecanismo de replicación en rueda giratoria (ver Tabla 2) sucede algo extraordinario. Los plásmidos bacterianos, dotados de una ADN pol B que trabaja según ese antiguo mecanismo replicativo, vienen a rellenar con exactitud el hueco en el que cabe esperar encontrar un ADN autónomo y libre aún de envolturas protectoras que ha aprendido ya a sintetizar una polimeraLas hipótesis formuladas para explicar los eventos que dieron lugar a la aparición de la Vida en nuestro planeta son diversas. Para unos, las primeras macromoléculas relacionadas con la Vida pudieron formarse en el propio planeta, en ambientes concretos del mar primigenio. La rugosa superficie de las rocas que forman las chimeneas de las fumarolas en las dorsales oceánicas habría proporcionado un microentorno reductor y muy catalítico en el
que los componentes básicos de los azúcares, los ácidos nucleicos y las proteínas habrían podido alcanzar concentraciones suficientemente altas como para permitir la formación de macromoléculas biológicamente activas. Para otros, pudieron hacerlo en otra parte del Universo, llegando repetidas veces hasta aquí con los meteoritos y con los residuos desprendidos de la helada superficie de los cometas, ya que en ambos se han podido detectar compuestos orgánicos de cierta complejidad.
El consenso es, sin embargo, más amplio en postular que los precursores de la vida celular pudieron surgir espontáneamente en el ambiente de las chimeneas submarinas merced a la presencia de esas macromoléculas, fuese cual fuese su procedencia. La razón de su formación habría sido su habilidad para dispersar gradientes energéticos surgidos en ese entorno como consecuencia de la química del azufre. Hasta la aparición de los primeros organismos fotosintéticos, que cambiaron la composición de la atmósfera hasta el punto de iniciar la era del sistema oxígenodióxido de carbono y de la fosforilación oxidativa, la vida terrestre habría sido, por consiguiente, muy diferente de la que conocemos hoy, que nació de la explosión vital del Cámbrico. Cabría, así, fijar el origen de la Vida en el momento en el que surgieron por vez primera esas protoSin embargo, las protocélulas surgidas al calor de esos gradientes de energía química carecían de capacidad para adaptarse a los cambios. Tal capacidad sólo pudieron adquirirla al asociarse con unos ácidos nucleicos, quizá ya expertos en cuestiones de adaptación y supervivencia, que habrían de proporcionarles un genoma y, con él, una identidad. ¿Quién estaba, pues, más “vivo” antes de que tal simbiosis se produjese? Es en este punto en el que yo me decanto más por Dawkins que por Margulis, en tanto que la conservación de la identidad y la capacidad adaptativa me parecen propiedades más “esenciales” de la Vida que las habilidades metabólicas. Pienso, además, que el hecho de que esos seres no celulares hayan llegado hasta nuestros días bajo aspectos tan variados como los que exhiben hoy, y en tal abundancia como la que hoy vemos, es una razón muy sólida para sustentar ese punto de vista. La Figura 2 pretende esquematizar en forma comprensible la participación de todos estos protagonistas en los fenómenos que terminaron por originar la aparición de las formas de vida que observamos hoy sobre el planeta.
“En un principio fue la Palabra..., y la Palabra transfor
Árbol de la vida. Propuesta basada en los datos disponibles sobre la filogenia de las replicasas. Se incluyen en él los virus ADN y RT, los viroides, las mitocondrias, los cloroplastos y los plásmidos bacterianos junto con bacterias, arqueas y organismos eucariotas. Los datos no permiten ubicar a los virus ARN en el esquema.
2. Bamford DH. Do viruses form lineages across different domains of life?. Research in Microbiology 2003; 154:231-236.
Trata muy bien la cuestión de las familias de virus con hospedadores en diferentes taxones básicos de seres vivos. Afirma que los virus han podido preceder a la vida celular, y despliega bien la idea de árboles evolutivos paralelos y quizás entrecruzados.

3. Desjardins C et al. New evolutionary frontiers from unusual virus genomes.
Todos los retos de conocimiento que plantean los “virus gigantes” –Mimivirus y Phycodnavirus- en relación con el origen de los virus y las raíces del árbol de la Vida.
4. Goodier JL, Kazazian HH Jr. Retrotransposons revisited: the restraint and rehabilitation of parasites. Cell 2008; 135:23-35.
Figura 2. Esquema que especula sobre las sucesivas asociaciones simbióticas que, Excelente revisión sobre los retropartiendo de entes metabólicos carentes de mecanismos replicativos y de ácidos transposones. nucleicos autorreplicantes de vida libre, habrían dado en originar seres vivos celu lares dotados de identidad genética y de habilidades adaptativas. 5. Koonin E. Temporal order of evolution of DNA replication systems inferred by comparison of cellular and viral DNA mó el Mar con su mensaje, copiándose sin cesar y para polymerases. Biology Direct 2006; 1:39. siempre.” Mi base para “el cuento de las replicasas”. Con esta hermosa y sugerente frase terminó Matt 6. Villarreal LP. Are viruses alive?. Scientific American, DiRidley su libro titulado Genoma, dedicado a analizar la ciembre 2004. historia evolutiva de nuestra especie a través de los mensajes que están escritos en nuestros genes. Quizás algún día Muy divulgativo, aunque quizá demasiado “mediáti co”, como pasa a veces con lleguemos a conocer la verdad sobre el origen de la Vida y sobre nuestro origen último; quizá no lo logremos nunca. 7. Scientific American. Se puede encontrar traducido en el Pero estoy íntimamente convencido de que si un día lo lonúmero 341 de la revista Investigación y Ciencia, Febrero de 2005 (“¿Tienen vida los virus?”) gramos será porque no habremos apartado nuestra mente de la línea de pensamiento que esa bella frase nos marca. 8. Villarreal LP, de Filippis VR. A hypothesis for DNA viruses as the origin of eukaryotic replication proteins. Journal of ViGracias, lector, por haber continuado peregrinanrology 2000; 74:7079-7084. do conmigo hacia ese otro Canterbury, más lejano, que yo Propuesta razonada de que las ADN pol B son de oriimagino. Tal vez hayamos viajado juntos por más de mil gen vírico, y de que los virus son, por consiguiente, más antimillones de años, y ese es tiempo más que suficiente para guos que la célula eucariota. hacer buenos amigos. 9. Zanotto PM et al.. A reevaluation of the higher taxonomy of viruses based on RNA polymerases. Journal of Virology 1996; MÁS PARA LEER 70:6083-6096.
1. Ruiz-Sáenz J, Rodas JD. Viruses, virophages and their living nature. Acta Virologica 2010; 54:85-90.
Reciente revisión breve sobre las reflexiones que el descubrimiento de los Mimivirus y los virófagos aconsejan introducir en la polémica sobre la pertenencia de los virus al mundo de “lo vivo”. Droga dura, sólo para los muy interesados en tratar de entender los problemas que plantea el origen de los virus ARN. Muestra claramente que los virus ARN y los virus ARNRT no poseen un ancestro común.
Jose Manuel Echevarría Mayo (Madrid, 1953) es licenciado en Ciencias Químicas y doctro en Farmacia por la Universidad Complutense. En la actualidad trabaja como jefe del Área de Virología del Centro Nacional de Microbiología. Ha publicado más de 160 artículos científicos relacionados con la virología, aunque su trabajo como especialista se ha centrado en el problema de las hepatitis víricas. En los últimos años ha impulsado la creación de un grupo de investigación multidisciplinar dedicado al estudio de las enfermedades víricas tropicales. Así mismo es autor de la novela de literatura de ficción Amaroncachi.
Bajo el nombre de rapaces, también llamadas aves de presa, englobamos a un grupo polifilético de aves con una serie de características morfológicas y comportamentales similares que tienen como finalidad la captura de presas vivas, aunque algunas especies se han especializado en alimentarse de carroña. Gozan de una vista muy desarrollada, una capacidad de vuelo magnífica, unas garras potentes y afiladas y un pico igualmente robusto y con gran capacidad para despedazar carne.
Actualmente las aves rapaces se recuperan lentamente de una masacre a manos del ser humano. A pesar de ello, nuevas amenazas asociadas al progreso del hombre aparecieron durante el siglo XX y se ciernen sobre el futuro, comprometiendo en gran medida la supervivencia de poblaciones estables de estas aves.
Un siglo de oscuridad
A pesar de que hoy en día cuentan con una protección legal, no siempre fue así y se las tildó de animales dañinos, ya fuera por su incidencia sobre especies criadas por el hombre o sobre especies cinegéticas, considerándolas alimañas. Las aves rapaces fueron ampliamente perseguidas por ello, llegándose a pagar recompensas por cada ejemplar abatido, lo cual esquilmó las poblaciones hasta el punto de llegar a extinguir algunas especies a ni


vel local. En Europa, la Primera y Segunda Guerra Mundial supusieron una tregua para la caza de rapaces y tras ellas, entre los años 50 y 60, comienza a protegerse a este grupo de aves, lo cual se ve reflejado en un ligero aumento de sus poblaciones en la década de los 70. En España, las aves rapaces están bajo protección legal desde 1966.
Esta amenaza directa, si bien se ha reducido mucho, no ha desaparecido del todo ya que año tras año a los CREA (centros de recuperación de especies amenazadas) siguen llegando individuos tiroteados. Ahora bien, aunque es difícil calcular las pérdidas exactas debido a que se trata de una práctica furtiva y por tanto, se trata de ocultar, sí se puede afirmar que la tendencia es descendente y en ningún caso tan alarmante como en épocas anteriores. Un caso excepcional puede considerarse Malta. En esta pequeña isla mediterránea se lleva a cabo una indiscriminada caza ilegal de aves migratorias. Por su posición estratégica, situada en la ruta europea de migración hacia África, miles de pájaros pasan por la isla y lamentablemente los cazadores (Malta tiene la mayor densidad de cazadores por kilómetro cuadrado de Europa) incumplen deliberadamente la normativa europea, hecho que se viene produciendo desde que Malta entró a formar parte de la Comunidad Europea.
Sin hogar
Como ya se ha apuntado, esos niveles de persecución directa parecen haber disminuido pero las poblaciones de aves rapaces se enfrentan actualmente a otras serias amenazas. Una de las más preocupantes es la pérdida y fragmentación de hábitat como consecuencia de incendios forestales, proliferación de pistas, trabajos forestales inadecuados, transformaciones masivas de secanos y terrenos forestales en regadíos intensivos, talas a matarrasa, disminución del alimento, roturación, una urbanización incontrolada, etc.
Las repoblaciones con coníferas de rápido crecimiento privan a estas aves de gran-
des árboles de fronda aislados, viejos y fuertes donde anidar. Debido a las especiales características de sus nidos, las rapaces están entre los grupos de aves cuyo número y éxito de reproducción están limitados con frecuencia por la disponibilidad de lugares para los nidos. A su vez, la desecación de regiones húmedas también produce un empobrecimiento de la naturaleza ya que, si bien no suelen ser zonas donde la gran mayoría de especies de aves rapaces aniden, sí son áreas muy productivas con buena oferta de alimentos.
A todo este conjunto de acciones que degradan el medioambiente hay que sumar las repercusiones del cambio climático, inciertas para muchos ecosistemas y sus pobladores, pero nada alentadoras. Por tanto, se han perdido tantas zonas de hábitat favorable que nunca se restablecerá la densidad inicial de población de tiempos anteriores a los grandes episodios de caza. A lo más que aspiramos es a mantener poblaciones óptimas allí donde el ecosistema aún lo permite.
Traumas
Otra de las principales causas de muerte o lesión que sufren las rapaces son los atropellos, consecuencia directa de la proliferación de pistas, los choques contra las ventanas, contra tendidos eléctricos y aerogeneradores. Generalmente, las colisiones se producen con líneas de alta tensión superiores a 45 kV, que poseen un cable de tierra de menor diámetro situado por encima de los cables de mayor grosor que conducen la corriente eléctrica y, por tanto, es menos visible para los pájaros. Respecto a los aerogeneradores, se suelen ver afectadas las especies planeadoras de mayor envergadura, siendo especialmente vulnerable el buitre leonado (Gyps fulvus). La proliferación de parques eólicos en los últimos años maximiza la
Árboles que matan
Ligado a los tendidos eléctricos, además del problema de las colisiones, encontramos el riesgo de electrocución. Las aves rapaces utilizan los postes como oteaderos y la descarga eléctrica se produce cuando el ave toca dos cables o un cable y la cruceta. Los diseños más peligrosos son aquellos con apoyos con aisladores rígidos, con elementos conductores de corriente por encima de la cruceta o zona de posada, así como los apoyos con transformadores y secuenciadores. Para minimizar el impacto se estudian las zonas o los tendidos donde se detectan mayores índices de mortalidad y, o bien se modifican los postes para hacerlos más seguros o bien se entierran. Ésta última opción es la más adecuada pero su elevado coste hace que no siempre sea posible llevarla a cabo.
Envenenamiento e intoxicación
Otra práctica delictiva cuyo grado de indicidencia es difícil de cuantificar es la persecución directa mediante el uso de cebos envenenados. Especies como el alimoche (Neophron percnopterus) o el milano real (Milvus milvus) son especialmente vulnerables. El uso de estricnina como veneno se expandió por los campos españoles. Actualmente se encuentra muy perseguido aunque siguen apareciendo casos.
La estricnina como veneno.
La estricnina es un alcaloide que se obtiene de la nuez vómica. Actúa como antagonista del aminoácido glicina, que es neurotransmisor de las células de Renshaw.


En dosis mayores de 25 mg puede producir la muerte debido a la contractura de los músculos torácicos. Al inmovilizar el músculo del diafragma el individuo muere por asfixia. Otras veces la intoxicación no viene provocada de una manera directa como la expuesta anteriormente, si no de una indirecta mediante sustancias añadidas al medio generalmente para la erradicación de plagas o la protección de cultivos frente al ataque de insectos y que resultan ser nocivas para las aves. Es el caso de los pesticidas, herbicidas y metales pesados. Este conjunto de biocidas tiene tres propiedades peligrosas, por un lado, son muy estables y persisten en el entorno durante muchos años; después, son solubles y se acumulan en las grasas corporales de los animales, pasándose así de presas a depredadores, con lo cual, las concentraciones de estas sustancias aumentan según se asciende en la cadena alimenticia y por último, dosis pequeñas pueden afectar a la reproducción y reducir el número de crías. Algunos pesticidas muy utilizados fueron los hidrocarburos clorados como el DDT, el dieldrín y el aldrín que son diez veces más tóxicos que el DDT o el BPC (bifenilo policlorado) que es diez veces menos tóxico pero increíblemente estable. Estas sustancias han sido prohibidas y sustituidas por otras cuya magnitud perjudicial es menor, aunque se siguen utilizando en países subdesarrollados. Respecto a los metales pesados, el plomo produce intoxicación por ingesta y el mercurio es letal a dosis muy bajas. Se ha comprobado que en un gavilán (Accipiter nisus) una cantidad de 6 ppm es el límite crítico.
El uso del DDT.
El DDT fue un pesticida usado masivamente durante la Segunda Guerra Mundial contra la malaria. Su descomposición es muy lenta y se acumula en el cuerpo. Es mortal en dosis de 30 ppm (30 mg por kg de tejido cerebral), además incita al hígado a producir enzimas que intervienen en el desequilibrio de las hormonas sexuales y puede tener efecto como hormona sexual. A pesar de ello, no suele ser tan perjudicial como el producto que surge de su descomposición, el DEE, que en muy bajas concentraciones daña la capacidad de producir cáscaras de huevo e interfiere en el intercambio gaseoso entre el embrión y el aire. Una disminución del 15-20% del grosor en la cáscara del huevo es crítica puesto que no soporta el peso de los progenitores al incubar y el huevo se rompe, perdiéndose la puesta. Los descensos poblacionales son más acusados entre las rapaces que se alimentan de otras aves puesto que ellas acumulan estas sustancias en su cuerpo.
La destrucción de nidos
El expolio es el saqueo de nidos y el robo de los huevos. Hoy en día la causa no es tanto el exterminio de las rapaces por ser considerados animales dañinos si no que está más asociado al auge de la cetrería, una modalidad de caza donde se utilizan aves de presa. De esta manera existe todo un mercado negro de huevos y pollos extraídos de nidos en la naturaleza que luego son vendi
Enfermedades
Otro motivo que provocó descensos poblacionales en el siglo pasado fueron las enfermedades sufridas por las presas o por las propias aves rapaces. Entre las primeras, especialmente perjudiciales fueron las relacionadas con el conejo (Oryctolagus cuniculus), especie clave para muchas de las aves rapaces ibéricas, como la mixomatosis, de gran incidencia en los años 60 o la neumonía hemorrágica vírica, más agresiva en los años 80. Recientemente, las encelopatías espongiformes transmisibles, por las que se prohibió el abandono de cadáveres en el campo, han supuesto un duro azote para las poblaciones de rapaces carroñeras que han visto mermada una gran fuente de alimento.
Menos conocidas son las enfermedades propias de las aves rapaces como por ejemplo el virus del Nilo Occidental del cual se vienen registrando brotes en países europeos donde nunca antes había aparecido. Se piensa que esta enfermedad, letal para las rapaces, se está extendiendo debido a los efectos del cambio climático puesto que es transmitido por la picadura del mosquito común de las casas (Culex pipiens) y del mosquito tigre (Aedes albopictus).
Molestias en época de cría
Las aves de presa son especialmente sensibles a las molestias cercanas a sus zonas de cría y reproducción, por ello el aumento de deportes como la escalada o el excursionismo pueden conducir a que muchas puestas sean abandonadas ante la presión que el ser humano realiza sobre el entorno natural. Algunas de estas prácticas se paralizan en épocas críticas de reproducción como la escalada, el paso de aviones sobre zonas protegidas o la interrupción de obras si se descubre algún nido en las inmediaciones.
Una perspectiva futura
El futuro para las aves rapaces lejos de cincelarse con letras diamantinas se aparece incierto. Aunque las poblaciones de la mayoría de las especies se recuperan lentamente, este conjunto de amenazas conlleva que no se pueda bajar la guardia en su conservación. Por fortuna cada vez es mayor el porcentaje de la población concienciada que aprecia y valora el medioambiente y trabaja para que en nuestros cielos nunca dejemos de ver el poderoso y bello vuelo de águilas, buitres, búhos y demás
aves rapaces.
Jesús David Tavira Guerrero es Licenciado en Biología en la especialidad de zoología. Actualmente profundiza en el campo de la ornitología y de evaluación de impacto ambiental. Es posible encontrarlo en los blogs ‘Aves de Guadalajara’ y ‘Rapaces del Mundo’ de los cuales es autor, asi como en su cuenta de twitter @JesusDavidTG
PROFESIÓN VETERINARIA 250 AÑOS


Por Fernando Adam
Nada más y nada menos que 250 años cumple la profesión veterinaria y no podríamos encontrar un mejor año para reivindicar el papel de estos profesionales. Precisamente por eso las celebraciones para conmemorar tal magno acontecimiento tenían que estar a la altura de las circunstancias, razón por la cual se han llevado a cabo conmemoraciones a nivel mundial. No debemos dejar pasar la oportunidad de comunicación que nos brinda la revista Journal of Feelsynapsis para recordar la versatilidad de esta profesión. La historia de la Medicina Veterinaria en el mundo se remonta a las primeras civilizaciones establecidas en la tierra. Nace la Medicina Veterinaria asociada a la cría de animales con fines de proveer alimentos de origen animal, vestido, calzado, y trabajo, luego para la guerra.
Nace con una cultura mitad práctica, mitad esotérica o religiosa. Evolucionando en cada etapa histórica hasta ser fundada hace 250 años. Varias son las pruebas en papiros, murales, vasijas y esculturas antiguas donde se demuestra pastores atendiendo a los animales y sus prácticas de manejo. Si bien, la enseñanza reglada de la profesión data su origen en esos 250 años, la primera referencia escrita a los veterinarios como profesión aparece recogida en el mesopotámico Código de Hammurabi, datado entre los siglos XVII y XVIIi a. de C. En él se incluyen dos artículos, el 224 y el 225, que regulan la actividad de los veterinarios, encargados de cuidar a los bueyes y a los asnos en aquella época. digo de Hammurabi, que podríamos considerar el “primer tratado de Medicina Veterinaria” de la Historia, el papiro de Kahun. Aunque en él no se habla de la figura del veterinario como tal, pues parece que esta figura no existía en el antiguo Egipto y las labores de curación de los animales las realizaban los sacerdotes.
Flinders y Petrie, dos arqueólogos ingleses, encontraron en la ciudad de Kahun esta serie de papiros originarios de la época del faraón Amenemhaït III, que reinó en el imperio del Nilo, durante el siglo XX a. de C.
Como en una enciclopedia, los papiros recogen el saber egipcio sobre matemáticas, medicina (fundamentalmente ginecología y obstetricia) o veterinaria. En ellos se describen enfermedades del ganado y su tratamiento, así como patologías de perros y de gatos. Extensos fragmentos de estos documentos han llegado muy mal conservados hasta nuestros días, y en algunos casos no es posible traducirlos.
La historia de la Medicina Veterinaria en el mundo se remonta a las primeras civilizaciones establecidas en la tierra.

Pero a pesar de todo lo anterior, la enseñanza reglada de la veterinaria no aparece hasta que Luis XV, conocido como el Rey Sol, siguió los buenos consejos de su “Caballerizo Mayor Real”, Claude Bourgelat y dispuso la creación de la primera escuela de Veterinaria que se instaló en Lyon el 4 de Agosto de 1761. Se constituye así la primera escuela veterinaria del mundo, tiempo más tar

de se abrieron tres escuelas más en Francia, y a partir de
esas instancias de la historia, se fue replicando por todo el mundo la iniciativa, hasta nuestros días. Al crear los primeros establecimientos de formación, Bourgelat inventaba al mismo tiempo el oficio de veterinario. Pero el genio de Bourgelat fue más lejos. Efectivamente, gracias a su fructífera colaboración con los cirujanos lioneses, también fue el primer científico que se atrevió a decir que al estudiar la biología y la patología del animal, se podrían entender mejor las del hombre. El año 2011 es pues también el 250 aniversario del concepto de “patología comparada”, sin el cual la medicina moderna no habría nacido nunca. Si realizamos un salto gigantesco de esa época a la actual, vemos que la situación ha cambiado en cuanto a las responsabilidades del veterinario. Tal vez al ciudadano común no le interese mucho este aspecto, pero sí y en gran medida, en lo que significa “el veterinario” como garantía de su salud y de la salud de toda la población. La veterinaria a lo largo de su historia ha acumulado una impresionante colección de conocimientos, a la vez que explorando nuevos campos de actividad. La utilidad de los animales de compañía en la sociedad ya no necesita ser demostrada, como tampoco el interés de la función de los veterinarios como médicos de estos fieles compañeros del hombre y el principal defensor de su bienestar.

Es de lamentar, sin embargo, que la profesión veterinaria no siempre sepa transmitir el mensaje de que sus actividades representan un auténtico bien público mundial. Hoy el veterinario vela, como siempre, por la sanidad animal y la producción animal. Pero a su vez se erige como principal instrumento para garantizar la seguridad de los alimentos, la salud pública, la protección de la biodiversidad y la sostenibilidad de los ecosistemas del planeta.
No debemos por tanto conformarnos ante la reducción de nuestra actividad a la de médicos de animales que una buena parte de la sociedad hace de la labor vete
rinaria, por muy importante y reconocida que sea esta. ...la enseñanza reglada de la veterinaria no aparece hasta que Luis
XV ... siguió los buenos consejos de su “Caballerizo Mayor Real”, Claude
Bourgelat ... en. .. Agosto de 1761...
Muchas veces oímos decir que somos lo que comemos, pero se tiende a ignorar que en la misma medida estamos sanos porque comemos alimentos sanos y saludables, y eso se debe en gran parte a la implacable vigilancia que nuestra profesión ejerce sobre la calidad y la inocuidad de los alimentos.
El veterinario también respalda y protege uno de los componentes más importantes de la economía como lo son las producciones agropecuarias que permiten, al mejorar las economías, mejores posiciones sociales. Hoy en día, en una sociedad que vive de espaldas al campo este papel es fundamental. Todos debemos luchar y esforzarnos para dotar de visibilidad al campo y favorecer la conservación de medios y modos de vida tradicionales.

...la primera referencia escrita a los veterinarios como profesion aparece recogida en el mesopotamico Codigo de Hammurabi...
Pero a pesar de todos los avances, la esencia de la profesión veterinaria continua siendo lo misma: una elevada curiosidad científica y grandes dosis de respeto y amor hacia los animales y el medio ambiente. Esto es la auténtica vocación de veterinario, entender de ladridos, aullidos, mugidos, relinchos y cacareos. Es sentirse feliz cuando estamos junto a nuestros amigos de pelos y plu-pelos y plumas. Por lo anterior y por muchas cosas más los 250 años de la profesión veterinaria deben ser motivo de consideración y valoración por la sociedad en general y no sólo por los profesionales veterinarios.
¡Muchas Felicidades a la profesión veterinaria y a la nueva revista que nace con este número!

Fernando Adam Fresno, es Licenciado en Veterinaria por la Universidad Complutense de Madrid y posee un Máster Universitario en Investigación en Ciencias Veterinarias. Desde el 2006 desarrolla su trabajo en el Centro Nacional de Microbiología, simultaneando su trabajo como veterinario clínico de equipo de atención primaria e impartiendo cursos de formación en el ámbito veterinario y de la cooperación internacional.


Ciencia Policial

FENÓMENOS CADAVÉRICOS Y ENTOMOLOGÍA FORENSE PARA EL ESTABLECIMIENTO DE LA DATA DE LA MUERTE.
por Xavi P.
De las cámaras fotográficas de madera a la tecnología digital; de los retratos a mano alzada a la informática más precisa; de las armas rudimentarias a los avances punteros. Esa es la evolución de la Policía Científica que, con el paso de la historia, se ha ido adecuando a la modernidad para poner a disposición de los ciudadanos los mejores avances de cara a su tranquilidad. Lejos quedan las lupas y las intuiciones. Ahora, el rigor científico y la única voz de las pruebas son las armas con las que la Policía Científica lucha contra el crimen.
La rama “forense” de la ciencia se podría definir como el conjunto de disciplinas cuyo objeto común es el de la materialización de la prueba a efectos judiciales mediante una metodología científica. Cualquier ciencia se convierte en forense en el momento que sirve al procedimiento judicial. La primera disciplina precursora de la criminalística fue lo que en la actualidad se conoce como Dactiloscopia; ciencia que estudia las huellas dactilares. La criminalística tal como la entendemos nace de la mano de la medicina forense, en torno al siglo XVII, cuando los médicos toman parte en los procedimientos judiciales. A lo largo de la historia de la Policía Científica ha habido grandes nombres que han marcado un antes y un después por la incorporación de técnicas que supusieron una auténtica revolución y avance, poniendo a la Ciencia al servicio de la Criminología. Algunos de estos nombres son Alphonse Bertillon (1879), Juan Vucetich (1892) y Francis Galton, dentro de lo que podríamos considerar como la Época Contemporánea, pero existen evidencias de que ya mucho antes, algunas disciplinas científicas ya se aplicaban.
Si alguien creó un punto de inflexión en la evolución de esta disciplina fue Alphonse Bertillon , quien creó en Pa- creó en Pa-creó en París el Servicio de Identificación Judicial en 1882. Este mé-1882. Este método antropométrico se basaba en el registro de las diferentes características óseas métricas y cromáticas de las personas mayores de 21 años en 11 partes diferentes del cuerpo. En esa época Bertillon publicó una tesis sobre el retrato hablado. Desde 1884, Bertillon tomó fotografías de los lugares de los hechos con todos sus indicios. Fue en 1886, cuando Alan Pinkerton puso en práctica la fotografía criminal para reconocer a los delincuentes. Por su parte, Sir Francis Galton en el Londres de 1885 instaló los fundamentos para la solución del problema que representaba hacer una clasificación de las impresiones dactilares. En 1905 modificará su sistema citado en “Fingerprint Directories”.
de Buenos Aires (en la ciudad de La Plata), Argentina, dejara de utilizar el método antropométrico de Bertillon y redujo a cuatro los tipos fundamentales de Dactiloscopia, determinados por la presencia o ausencia de los deltas (figura triangular que surge como resultado de la confluencia de las diferentes zonas en las que se divide la impresión dactilar), sistema que se aplica en la actualidad para la clasificación de las huellas dactilares.
En la actualidad cabe destacar al biólogo y criminalista alemán Mark Benecke (1970), especialista en entomología fo-(1970), especialista en entomología forense, disciplina clave para la determinación de la data de la muerte, motivo de este artículo.
El establecer el momento de la muerte constituye una de las cuestiones más complicadas desde el punto de vista médico-forense, actividad que se lleva a cabo mediante el llamado cronotanatodiagnóstico. Cuando se produce el hallazgo de un cadáver, se realiza una inspección general del lugar y del cadáver, tratando de determinar las circunstancias relacionadas con el momento de la muerte así como la forma y causa en que se produjo. Si bien, aunque sea competencia de los profesionales de la medicina el establecer la data de la muerte, existen una serie de indicadores que nos pueden orientar para establecerla de un modo aproximado y que conviene tener en cuenta ante la presencia de un cadáver: los fenómenos cadavéricos.
FENÓMENOS CADAVÉRICOS
Los fenómenos cadavéricos son los cambios que se suceden en el cuerpo sin vida a partir del momento en que se extinguen los procesos bioquímicos vitales, sufriendo pasivamente la acción de las influencias ambientales. Existe una gran variabilidad de estos fenómenos, de forma que en la práctica no existen reglas fijas, si bien, la teoría supone una orientación. Veamos en qué consisten los fenómenos cadavéricos más importantes:
FENÓMENOS CADAVÉRICOS
TEMPRANOS
Acidificación tisular Enfriamiento* Deshidratación* Livideces* Rigidez*
Espasmo cadavérico
DESTRUCTORES
Autolisis* Putrefacción*
TARDÍOS
Antropofagia cadavérica*
CONSERVADORES
Momificación Adipocira 1 Corificación
El enfriamiento cadavérico o algor mortis es el elemento más útil para establecer la data de la muerte. Se debe al cese de la actividad metabólica. Tras la muerte, el cadáver pierde calor progresivamente hasta igualar su temperatura con la del medio ambiente (aproximadamente 1ºC por hora; entre 0-12h desciende de 0.8 a 1,1 ºC/h y entre 12-24 h desciende de 0,4 a 0,5 ºC/h, es la llamada curva de dispersión térmica). Sin embargo, hay que considerar determinadas circunstancias que pueden modificar el enfriamiento, haciéndolo más rápido o por el contrario retardándolo.
1-2 horas 4-5 horas 6-8 horas 10-12 horas 24 horas ENFRIAMIENTO CADAVÉRICO
Aparece en manos, pies y cara. Aparece en miembros, pecho y dorso. En vientre, cuello y axilas. En vísceras abdominales. Enfriamiento completo al tacto. La temperatura del cadáver se iguala a la del medio ambiente.
La deshidratación cadavérica se debe a la pérdida de líquidos por evaporación y lleva consigo una pérdida de peso corporal (que es más manifiesta en cadáveres jóvenes); una apergaminamiento cutáneo y una desecación de las mucosas. La pérdida de peso corporal es más manifiesta en cadáveres jóvenes, a razón de 8 gramos por kilogramo de peso y día. Sus principales manifestaciones se observan en los ojos y constituyen los llamados signos de Stenon-Louis y de Sommer-Larcher (Figura 1).
DESHIDRATACIÓN CADAVÉRICA
Pérdida de peso corporal (es más manifiesta en cadáveres jóvenes) Apergaminamiento cutáneo Desecación de las mucosas
Triángulo oscuro con base de la córnea -a los 45 minutos: en ojo con párpados abiertos - a las 24 h: en ojo con párpados cerrados -varía de rosa pálido a azul oscuro -alcanza máximo tono a las 12-15 h
1 Adipocira: (Del latín, adeps, grasa, y cera, cera). Sinónimo: GRASA DE CADÁVER. Sustancia gris blanquecina, blanda y untuosa al tacto, que procede de la saponificación de la materia animal en los cadáveres que han permanecido largo tiempo en el agua o enterrados en sitios húmedos.
*Desarrollados posteriormente.
El signo de Sommer-Larcher, también llamado mancha negra esclerótica, consiste en un triángulo oscuro con la base en la córnea, y otras veces, en una línea oscura que sigue el ecuador del ojo (Fig.ura 1d) El signo de Stenon-Louis consiste en el hundimiento del globo ocular, pérdida de la transparencia de la córnea y depósito de polvo de aspecto arenoso (Figura 1e). Existe la creencia de que el cabello y las uñas crecen tras la muerte, si bien esto no es cierto. Lo que sucede es que, durante el proceso de deshidratación, la piel se arruga y se contrae dando lugar a que queden a la vista las raíces de las uñas y del cabello, apareciendo así unos milímetros de pelo y de uña que anteriormente no se veían. Esto, añadido a la reducción de volumen que experimenta el cuerpo, puede provocar la ilusión visual de que han crecido las uñas y el pelo.
3. Livideces cadavéricas o hipostasias post mortem (livor mortis): cuando la dinámica circulatoria se suspende, el plasma y los glóbulos rojos de la sangre se acumulan y sedimentan en las partes más declives del cadáver. Esto obedece a las leyes físicas, es decir, influenciados por la gravedad se dirigen a los niveles más bajos del sistema vascular y se asientan en las zonas más bajas del cuerpo,
Fig. 1. Fenómenos cadavéricos
formando zonas de coloración variable. Las livideces tienen su importancia porque orientan sobre la posición que pudo haber tenido el cadáver (manchas de posición), teniendo en cuenta que las manchas no van a aparecer en aquellas zonas en las que haya habido presión, es decir, los puntos de apoyo del cuerpo en la superficie (Figura 1b).
4. La rigidez cadavérica (rigor mortis) consiste en el endurecimiento y retracción de los músculos del cadáver. Entre la muerte y este fenómeno hay una etapa de relajación muscular. El fenómeno desaparece en forma inversa y su desaparición coincide con el inicio de la putrefacción, porque entonces la proteína muscular se desnaturaliza y no puede mantener su contracción. Entre las 3 y 6 horas comienza en el corazón y diafragma, mandíbula y miembros inferiores, siendo completa entre las 8 y 12 horas, alcanzando su máxima intensidad a las 24 horas (Figura 1a). Comienza a desaparecer de forma inversa hacia las 36 horas. Curiosos fenómenos asociados a esta contracción post mortem son aquellos que pueden dar lugar a la eyaculacion, expulsión del feto o erección capilar.
Dentro de los fenómenos tardíos destructores, la autolisis es el más precoz de los procesos transformativos cadavéricos, consiste en la disolución de los tejidos por la acción de enzimas o fermentos propios de las células, es decir, anatómicamente estaríamos ante una necrosis celular.
Otro de los fenómenos tardíos destructores es la putrefacción cadavérica. Es el signo inequívoco de la muerte, ya que no se trata del cese de una actividad vital, sino de la modificación de los tejidos, modificación que sólo ocurre en el cadáver. Consiste en la descomposición de la materia orgánica del cadáver por acción de las propias bacterias que habitan los intestinos, que tras la muerte, se propagan por la sangre, dando lugar a un estadio del cadáver conocido como periodo cromático-enfisematoso (Figura 1c). En su desarrollo actúan primero las bacterias aerobias, luego las aeróbicas facultativas y finalmente, en ausencia de oxígeno, intervienen las bacterias anaeróbicas.
El último de los fenómenos cadavéricos tardíos destructores es la antropofagia cadavérica, un proceso que consiste en la destrucción del cadáver por la acción principalmente por los insectos. La evolución natural del cadáver finaliza con la destrucción del mismo. Sin embargo, si se modifican las condiciones del ambiente, puede detenerse la descomposición y virar hacia un fenómeno conservador. Así se producen los fenómenos tardíos conservadores como la momificación.
ENTOMOLOGÍA FORENSE
Una de las cuestiones de mayor importancia a ser resuelta para los investigadores frente a un cadáver es averiguar el tiempo transcurrido desde que ocurrió la muerte, pudiendo llegar a ser vital en la investigación de un homicidio. Generalmente, el tiempo transcurrido desde la muerte es determinado mediante otras disciplinas forenses (como el anteriormente citado cronotanatodiagnóstico, por ejemplo), pero cuando el cuerpo se encuentra en un estado de putrefacción avanzado, o el cadáver presenta distintas fases de descomposición a la vez, la validez de estos métodos es limitada, y es en estos casos donde la Entomología Forense puede llegar a ser más exacta, siendo con frecuencia el único método fiable para determinar el intervalo postmortem.
La “Entomología Médico Criminal” o Entomología Forense es una disciplina incluida en las Ciencias Forenses que utiliza los datos que los artrópodos pueden aportar en la investigación médico-legal de la muerte proporcionando una fuerte evidencia científica que puede ser defendida con éxito. El primer documento escrito de un caso resuelto por la entomología forense se remonta al siglo XIII en un manual de Medicina Legal chino referente a un caso de homicidio en el que apareció un labrador degollado por una hoz. Para resolver el caso hicieron que todos los labradores de la zona que podían encontrarse relacionados con el muerto, depositasen sus hoces en el suelo, al aire libre, observando que tan solo a una de ellas acudían las moscas y se posaban sobre su hoja, lo que llevó a la conclusión de que el dueño de dicha hoz debía ser el asesino, pues las moscas eran atraídas por los restos de sangre que habían quedado adheridos al ‘arma’ del crimen.
decible, esto es, existiendo una sucesión que se repite siempre en el mismo orden, ya que los distintos grupos van apareciendo por oleadas a medida que progresa el proceso de descomposición y el cadáver va pasando por distintas condiciones que resultan atractivas a distintos grupos de insectos.
El proceso de descomposición cadavérica se divide en cinco fases (según nomenclatura recomendada por los entomólogos forenses internacionales):
Fresco (también llamado período cromático o colorativo, por la coloración verdosa o negruzca del cadáver, durante la primera semana)
Hinchado (o periodo enfisematoso o de desarrollo gaseoso, durante las siguientes dos semanas) Descomposición activa
Descomposición avanzada
Esqueletización
Fase de cadáver “fresco”
A los pocos minutos de producirse la muerte, los dípteros comienzan la ovoposición sobre orificios naturales, fundamentalmente ojos, fosas nasales, en y alrededor de las orejas y, si están expuestos, ano y pliegues inguinales. Este estado se caracteriza por la abundancia de dípteros de la familia Calliphoridae (sobre todo Lucilia sericata, considerado el primer colonizador, incluso cuando aún no se detecta olor a putrefacción), Muscidae y Sarcophagidae (considerados colonizadores secundarios generalmente). También es posible encontrar himenópteros de la familia Formicidae, sobre todo cuando los cadáveres están expuestos a la intemperie. Los individuos de la especie Chrysomya albiceps (Fam. Calliphoridae), son necrófagos y depredadores, aunque son considerados colonizadores secundarios, si bien cuando aparecen lo hacen de forma mucho más abundante (Figura 2).
Figura 2. Dípteros

Fase de cadáver “hinchado”
Se produce la eclosión de las puestas, observándose el desarrollo de las larvas con actividad muy marcada, las cuales se alimentan de las vísceras. Los dípteros que se aprecian siguen siendo los de las familias anteriormente citadas, mientras que aparecen nuevos insectos, adultos de coleópteros necrófagos de la familia Silphidae y depredadores de las familias Staphylinidae e Histeridae (Figura 3).
Fase de “descomposición activa”
En esta fase aumenta la actividad larvaria, apareciendo grandes masas larvarias descomponiendo los tejidos blandos del cadáver. Siguen alimentándose dípteros de las familias Calliphoridae, Muscidae y Sarcophagidae, aunque ya no se localizan puestas ni larvas de edades tempranas. Los coleópteros son los mismos de la fase anterior, fundamentalmente de la familia Silphidae, que aparecen en fases tempranas para realizar puestas, ya que sus larvas se alimentarán de
Fase de “descomposición avanzada”
Las larvas de dípteros necrófagos de la familia Calliphoridae empiezan su actividad migratoria, pupando en zonas ubicadas debajo del cadáver y en los alrededores del mismo, en un radio de acción de unos dos metros de distancia. Se produce la mayor pérdida de

biomasa y se observan puntos de “preesqueletización” marcada en la cabeza.
Figura 3. Coleópteros En esta fase se encuentran, además de los coleópteros de hábitos depredadores antes citados, otras familias con hábitos necrófagos, pero con preferencia por tejidos secos, tales como las familias Cleridae (Necrobia rufipes), Histeridae, Staphylinidae y Nitidulidae (Figura 3). Hay que tener en cuenta que los adultos de las familias Histeri
dae y Staphylinidae se sienten atraídos al cadáver debido a la presencia de las larvas de dípteros, las cuales forman parte de su dieta, haciendo que su presencia en el cadáver no esté asociada a una fase concreta de la descomposición.
Fase de “esqueletización”
La duración de este estadio es la más larga. Los cadáveres son colonizados por grupos dermatófagos y queratófagos, fundamentalmente especímenes de la familia Dermestidae (también coleópteros).
Para establecer la data de la muerte en estadios avanzados de descomposición, las especies de interés forense serán aquellas que son atraídas por elementos químicos del proceso degradativo de la putrefacción y que son activas desde el momento de la colonización (margen mínimo) hasta la fecha de abandono del cadáver (margen máximo). De esta manera, son los estadios larvarios de las familias Cleridae, Silphidae, Dermestidae y Nitidulidae, quienes marcan los umbrales mínimos y máximos de tiempo, ya que los adultos de estas familias pueden aparecer durante todo el proceso, careciendo de valor en la estimación del intervalo post-mortem (Figura 4).
Fundamentalmente, el estudio de la sucesión de insectos sobre el cadáver se utiliza para realizar la estimación del intervalo postmortem en estados de descomposición avanzados. La estimación se basa en la comparación de los insectos que se recolectan en el caso real sobre el cadáver con los datos existentes en la bibliografía para la misma zona geográfica y estación del año. Hay que tener en cuenta que existen una serie de variables que afectan a la aparición y sucesión de unas especies u otras sobre el cadáver. Dentro de ellas están las inherentes al individuo (constitución física,

Figura 4. Larvas edad, salud, toxicología, existencia de traumatismos ante-mortem, etc.) y las geográfico-ambientales (temperatura, humedad, entorno rural o urbano, exposición del cuerpo, etc.)
REFERENCIAS
1. Data de la muerte, fenómenos cadavéricos. Sandra Muñoz Carrasco. Policia, 240; 18-27.
2. Determinación del intervalo postmortem mediante el estudio de la sucesión de insectos en dos cadáveres hallados en el interior de una finca rústica en Madrid. A.M. García-Rojo1, L. Honorato1, M. González1 y A. Téllez1. Cuad Med Forense 2009; 15(56):137-1.
3. La entomología forense. Mariano González González. Ciencia Policial, 102; 87-104.
4. La entomología forense y la práctica policial en España: estimación del intervalo post-mortem en un cadáver hallado en el interior de una arqueta en la comunidad de Madrid. Ana García-Rojo y Lourdes Honorato. Ciencia Forense, 8/2006: 57-62.
5. La entomología forense y su aplicación a la medicina legal. Data de la muerte. Concha Magaña. Aracnet 7 -Bol. S.E.A., nº 28 (2001): 49—57.
6. http://cienciacriminalistica.blogspot.com/2010/09/rigidez-cadaverica-instalada.html
7. http://criminaldescubierto.blogspot.com/2011/03/signos-de-muerte-tanatologia.html
8. http://nicoleb.org/mnblog/2011/08/flesh-fly-sarcophagidae/ wishis.html http://plantpath.wisc.edu/~young/hbhomage/
9. http://www.entomofauna.es.tl/Sucesion-cadaverica.htm
10. http://www.flycontrol.novartis.com/species/flsstablefly/es/index.shtml
11. http://www.insectimages.org
12. http://www.schoepfung.eu/leben-in-wasser-und-luft/in-der-luft/schmeissfliege.htm
13. http://www.umb.edu.co/cidfe/revista/articulos/panorama/n02_2006.html
14. http://www.wikipedia.org





