
DIAGNÓSTICO MOLECULAR DE PRRS: DE LA PCR A TIEMPO REAL A LA SECUENCIACIÓN MASIVA p. 39 Nº7 | Abril 2023
Hazlo fácil
Los protocolos de inseminación

requieren gran cantidad de recursos y esfuerzo y pueden ser una fuente importante de estrés en la granja. La inseminación es crucial, aunque es sólo una parte de tu trabajo. Relájate, ahorra tiempo y trabajo y reduce tu estrés gracias a la simplicidad de un protocolo de inseminación única a tiempo fijo (IAU-TF).
Ahorra tiempo y trabajo Reduce tu estrés Cronograma conocido
Simplemente fácil
FICHA TÉCNICA: OVUGEL 0,1 MG/ML GEL VAGINAL PARA CERDAS PARA LA REPRODUCCIÓN. Composición: Un ml contiene: Sustancia activa: Triptorelina (como acetato de triptorelina) 0,1 mg;

Excipientes: Parahidroxibenzoato de metilo, sal de sodio 0,9 mg, Parahidroxibenzoato de propilo, sal de sodio 0,1 mg. Para la lista completa de excipientes, véase la sección 6.1. de la Ficha Técnica. Indicaciones de uso: Para la sincronización de la ovulación en cerdas destetadas a fin de permitir una única inseminación artificial en un momento determinado. Contraindicaciones: No usar en casos de hipersensibilidad a la sustancia activa o a algún excipiente. No usar durante la gestación ni la lactancia. No usar en cerdas con anomalías evidentes en el aparato reproductor. Precauciones especiales de uso. Advertencias especiales para cada especie de destino. La eficacia de OvuGel no se ha demostrado en cerdas jóvenes (cerdas nulíparas), por lo que no se recomienda el uso del medicamento veterinario en estos animales. La respuesta de las cerdas a los protocolos de sincronización puede verse influida por el estado fisiológico en el momento del tratamiento. Las respuestas al tratamiento no son uniformes ni entre las distintas piaras ni entre los individuos de una misma piara. Precauciones especiales para su uso en animales: El producto no debe utilizarse en cerdas con anomalías en el aparato reproductivo, infertilidad o trastornos médicos en general. Se realizó un estudio de seguridad de la reproducción en cerdas después de la administración de 3 veces la dosis recomendada de OvuGel, que no mostró ningún efecto en el rendimiento de la reproducción ni en los lechones. Sin embargo, no se ha demostrado la seguridad del tratamiento en cerdas en los ciclos reproductivos posteriores. No se pueden excluir los posibles efectos a largo plazo de la aparición de quistes. Precauciones específicas que debe tomar la persona que administre el medicamento veterinario a los animales: El producto puede causar irritación de los ojos. Las personas con hipersensibilidad conocida a los análogos de la GnRH o cualquiera de los excipientes (incluidos los parabenos) deben evitar todo contacto con el medicamento veterinario. Usar un equipo de protección personal consistente en un mono y guantes al manipular el medicamento veterinario. No comer, beber ni fumar al manipular el medicamento veterinario. Evitar el contacto directo con la piel o los ojos, lavarse las manos tras la manipulación del medicamento veterinario. En caso de contacto accidental con los ojos, enjuagar bien y buscar atención médica inmediatamente. En caso de contacto accidental con la piel, lavar las zonas contaminadas con agua y jabón. La triptorelina puede afectar a los ciclos reproductivos de las mujeres y se desconocen los efectos de la exposición accidental en mujeres embarazadas; por consiguiente, se recomienda que las mujeres embarazadas no manipulen el medicamento veterinario y que las mujeres fértiles lo hagan con precaución. Uso durante la gestación, la lactancia o la puesta. Gestación y lactancia: No ha quedado demostrada la seguridad del medicamento veterinario durante la gestación ni la lactancia. No usar durante la gestación ni la lactancia. Tiempo de espera: Carne: Cero días. Titular de la autorización de comercialización Vetoquinol S.A. Número de la autorización de comercialización: EU/2/20/260/001. USO VETERINARIO. MEDICAMENTO SUJETO A PRESCRIPCIÓN VETERINARIA. BAJO CONTROL O SUPERVISIÓN DEL VETERINARIO. Información del medicamento en: https://cimavet.aemps.es/cimavet

GA680-2
“Para quedarte donde estás tienes que correr lo más rápido que puedas. Si quieres ir a otro sitio, deberás correr, por lo menos, dos veces más rápido”.
La Reina Roja en “A través del espejo y lo que Alicia encontró allí”
Lewis Carroll, 1871
El viaje evolutivo de los seres que habitan este planeta es una aventura épica que se reescribe continuamente, una carrera armamentística que requiere adaptarse a los cambios constantes para mantenerse competitivos y evitar la extinción. Es por ello que los organismos biológicos no son estáticos, sino que deben mantenerse en constante “movimiento” para sobrevivir en un entorno aparentemente volátil, caprichoso y, en ocasiones, cruel.
Sin dejar de lado los condicionantes externos relacionados con el entorno, tal y como señalaba el biólogo evolutivo Leigh Van Val en su hipótesis de la Reina Roja, las especies mejoran en un proceso de coevolución con otras especies, ejerciendo una presión selectiva mutua cuyo resultado es el mantenimiento del status quo.
Ante el ojo inexperto, la observación de la historia evolutiva podría llevar al error de dotar a estos fenómenos de cierta intencionalidad hasta tal punto que parecen seguir un “Plan Maestro”…
¡Insectos que fingen ser plantas, plantas que aparentan ser insectos, microorganismos capaces de “manipular” a sus hospedadores para conseguir sus objetivos, mamíferos terrestres que un día “decidieron volver a los océanos” o especies muy alejadas filogenéticamente que convergen en estrategias sospechosamente similares!
¿Cómo no maravillarse ante semejantes obras de “ingeniería biológica”?
Sin embargo, quienes están familiarizados con las teorías evolutivas saben que los organismos biológicos no desarrollan un fenotipo específico con un objetivo final, sino que son las condiciones específicas a lo largo del tiempo las que determinan cuáles son los genotipos y, por ende, los fenotipos predominantes en un momento dado.
En el caso del ganado porcino, a través de la selección y mejora genética, la intervención humana, ya sí con una intencionalidad dirigida, ha sido el principal motor evolutivo que ha culminado en la obtención de las actuales líneas hiperprolíficas y de alto rendimiento productivo.
Trabajar con estas genéticas trae consigo nuevos retos, especialmente en un escenario global en el que el Cambio Climático y el auge de las bacterias multirresistentes son factores determinantes que deben estar presentes en los programas de mejora genética.
LECCIONES APRENDIDAS DE LA BIOLOGÍA EVOLUTIVA
No solo se trata de maximizar la supervivencia de los lechones nacidos de las cerdas hiperprolíficas a través de estrategias de manejo, sino también lograr que estos animales expresen su máximo potencial a lo largo de toda su vida.
Nos encontramos en un punto de inflexión en el que las nuevas genéticas deberán incorporar caracteres enfocados más en la resiliencia que en la productividad, seleccionando a favor de fenotipos que respondan mejor al estrés, a las condiciones ambientales desafiantes y a los agentes infecciosos, pero que también sean capaces de hacer un uso más eficiente de los recursos y minimicen sus emisiones al medio ambiente.
Para asistir en esta encrucijada, la comunidad científica avanza a pasos agigantados, por ejemplo, con las revolucionarias técnicas ómicas y el desarrollo de nuevas tecnologías que tendrán aplicaciones muy interesantes en el ámbito de la ganadería de precisión.
Si bien, la metáfora de la Reina Roja ilustra un interesante principio evolutivo, nos debe hacer reflexionar sobre los mecanismos que impulsan la evolución del sector porcino y la necesidad de adaptar los modelos productivos y empresariales a una realidad siempre cambiante, modelada no solo por condicionantes que vienen impuestos por las propias empresas del sector, sino también por otros que escapan a nuestro control, como las nuevas tendencias de consumo y las demandas de la sociedad.
¡Es necesario que el sector porcino coevolucione con la sociedad y con el ecosistema, y el conformismo no es una opción! Tal y como señalaba la Reina Roja, para ser competitivos no basta con cumplir con los mínimos, requiere mantenerse en continuo movimiento, siempre dos, tres o cuatro pasos por delante, con la mirada puesta en el futuro para adelantarse a los retos que vendrán.
EDITOR
GRUPO DE COMUNICACIÓN AGRINEWS S.L.
DISEÑO GRÁFICO & WEB
Marie Pelletier
Enrique Núñez Ayllón
PUBLICIDAD
Laura Muñoz +34 629 42 25 52 laura@mediatarsis.com
Luis Carrasco +34 605 09 05 13 lc@agrinews.es
REDACCIÓN
Daniela Morales Osmayra Cabrera F.X. Mora
ADMINISTRACIÓN
Mercè Soler
Barcelona
España
Tel: +34 93 115 44 15 info@agrinews.es
www.porcinews.com
www.porcinews.com/revista-porcisapiens/
Precio de suscripción anual:
España 45 €
Extranjero 120 €
ISSN (Revista impresa) 2696-8142
ISSN (Revista digital) 2696-8151
DIRIGIDA A VETERINARIOS DE PORCINO Depósito Legal PorciSapiens B 7620-2021
Revista Cuatrimestral
EDITORIAL
La
dirección de la revista no se hace responsable de las opiniones de los autores. Todos los derechos reservados. Imágenes: Noun Project/Freepik/Dreamstime/BioRender
1
4/14
EL INTESTINO, EL ÓRGANO INMUNITARIO
MÁS GRANDE DEL ORGANISMO
Cândida Azevedo¹ y Henry Cancian²
¹Zootecnista, MsC en Ciencia Animal y DsC en Ciencia Animal y Pasturas
²Zootecnista
15/25
INFLUENCIA DEL PESO CORPORAL SOBRE LA EXPRESIÓN GÉNICA
INTESTINAL DE LOS LECHONES
Sandra Villagómez-Estrada1,2, José F. Pérez1, Diego Melo-Durán1,3 , Francesc Gonzalez-Solè1, Matilde D’Angelo1, Francisco J. Pérez-Cano 4 y David Solà-Oriol1
1Departamento de Ciencia Animal y de los Alimentos, Servicio de Nutrición y Bienestar Animal (SNIBA), Universidad Autónoma de Barcelona

2Carrera de Medicina Veterinaria, Escuela Superior Politécnica de Chimborazo (ESPOCH)
3Facultad de Ciencias Médicas "Eugenio Espejo", Universidad UTE
4Departamento de Bioquímica y Fisiología, Universidad de Barcelona
26/28
SOUNDTALKS®, IA AL SERVICIO DE LA SALUD RESPIRATORIA PORCINA
Maite Bastero
Head of Marketing Livestock, Boehringer Ingelheim Animal Health España
30/38
USO DE NUEVAS TECNOLOGÍAS
DIGITALES PARA LA EVALUACIÓN DEL BIENESTAR EN GRANJA Y MATADERO
Emma Fàbrega1, Marc Bagaria1, Aranzazu Varvaró1, Joaquim Pallisera1, Xavier Averós2 e Inma Estevez2,3
1Programa de Bienestar Animal, IRTA Monells
2Departamento de Producción Animal, NEIKER Instituto Vasco de Investigación y Desarrollo Agrario Alianza Vasca de Investigación y Tecnología (BRTA)
3IKERBASQUE, Fundación Vasca para la Ciencia
39/51
DIAGNÓSTICO MOLECULAR DEL PRRS: DE LA PCR A TIEMPO REAL A LA SECUENCIACIÓN MASIVA
Sofía Lázaro Exopol S.L.
52/58
¿QUÉ MÁS PODEMOS HACER PARA AUMENTAR LA PROTECCIÓN FRENTE
AL PRRS?
Sonia Cárceles, Laura Garza, Salvador Oliver, Carlos Casanovas y David Espigares Servicio Técnico Porcino, Ceva Salud Animal
60/74
USO RACIONAL DE ANTIMICROBIANOS EN PORCINO: ANTIMICROBIANOS INHIBIDORES DE LA SINTESIS DE ARN Y ADN – PARTE II
Sonia Rubio, Mª Dolores San Andrés, Manuel I. San Andrés y Juan Antonio Gilabert
Sección Dptal. de Farmacología y Toxicología, Facultad de Veterinaria, Universidad Complutense de Madrid
Nº 7 | Abril 2023 2
NO 7
SUMARIO
75/89
DETECCIÓN RÁPIDA DE BACTERIAS RESISTENTES TRAS EL TRATAMIENTO CON ENROFLOXACINA
Elena González-Fandos1, Alba Martínez-Laorden1, Ana Abad-Fau2, Eloisa Sevilla2, Rosa Bolea2, María Jesús Serrano2, Olga Mitjana2, Cristina Bonastre2, Alicia Laborda2, María Victoria Falceto2 y Rafael Pagán2
1Departamento de Tecnología de los Alimentos, Centro de Investigación CIVA, Universidad de La Rioja
2Instituto Agroalimentario de Aragón-IA2 (Universidad de Zaragoza-CITA)
90/96
VACUNACIÓN DE CERDAS HIPERPROLÍFICAS. ¿PODEMOS MEJORAR LA INMUNIZACIÓN DE SUS LECHONES FRENTE A E. COLI ?
Agradecemos a nuestros anunciantes por hacer posible la publicación de esta revista: Boehringer Ingelheim, Ceva, MSD Animal Health, S.P. Veterinaria, Tashia y Vetoquinol. WWW.PORCINEWS.COM/


REVISTA-PORCISAPIENS/
Rut Menjón, Marta Jiménez y Marcial Marcos Servicio Técnico MSD Animal Health
EL INTESTINO EL ÓRGANO INMUNITARIO MÁS GRANDE DEL ORGANISMO
Descarga el PDF
LLa integridad intestinal es crucial para la salud y el rendimiento de los cerdos dado que, además de las funciones digestivas del tracto gastrointestinal (TGI), las células inmunitarias y las estructuras linfoides presentes en el TGI constituyen el órgano inmunitario más grande del cuerpo.
Cândida Azevedo¹ y Henry Cancian²

¹Zootecnista, MsC en Ciencia

Animal y DsC en Ciencia Animal y Pasturas
²Zootecnista
O
Nº 7 | Abril 2023 INMUNOLOGÍA El intestino: el órgano inmunitario más grande del organismo 4
LOS PILARES DEL SISTEMA INMUNITARIO INTESTINAL
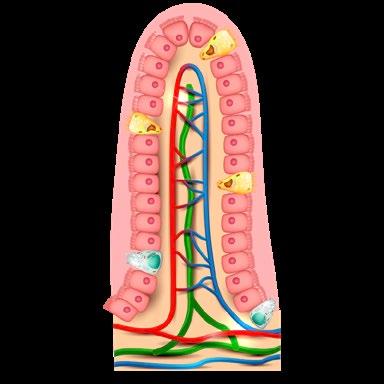
El sistema inmunitario asociado a la mucosa intestinal se sustenta en cinco pilares:
Barrera epitelial
Se trata de una barrera física formada por una capa de células epiteliales (enterocitos y colonocitos), células caliciformes productoras de moco, células de Paneth productoras de sustancias bactericidas (defensinas), células transportadoras especializadas en la captación de antígenos luminales (células M) y linfocitos T intraepiteliales.

Folículos linfoides aislados
Constituyen un punto de inicio de las respuestas inmunitarias adaptativas, siendo estructuras organizadas formadas por poblaciones de células inmunitarias.
Placas de Peyer (folículos linfoides asociados)
Son un punto de inicio de las respuestas inmunitarias adaptativas, siendo estructuras organizadas en forma de varios folículos linfoides con centros germinales.
Tejido linfoide difuso
Está formado por macrófagos, células dendríticas, mastocitos, linfocitos T efectores, linfocitos T reguladores, células plasmáticas secretoras de IgA dispersas en la lámina propia de la mucosa.
Ganglios linfáticos mesentéricos
Forman otro punto de inicio de las respuestas inmunitarias adaptativas, en este caso, frente a los antígenos presentes en el intestino que llegan a través del sistema linfático.
O
5 SALUD INTESTINAL
(Gonçalves et al., 2016)
IMPORTANCIA DE LA INTEGRIDAD DE LA BARRERA INTESTINAL
MUCOSA INTESTINAL
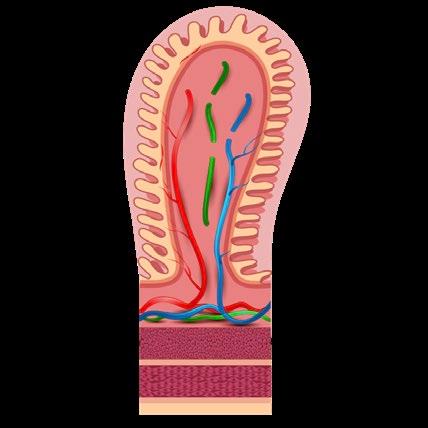
La superficie de la mucosa (Figura 1) está recubierta con un moco formado por mucinas que son secretadas por las células caliciformes, creando una barrera que evita que partículas más grandes, incluyendo la mayoría de las bacterias, entren en contacto directo con la capa de células epiteliales (Turner, 2009).
Mucina
Vellosidad intestinal
Capa mucosa
Epitelio
Lámina própia
Muscular de la mucosa
Plexo submucoso
Vena
Arteria
Capa submucosa
Capa muscular
Capa serosa
FIGURA 1
Vaso linfático
Músculo circular
Plexo mientérico
Músculo longitudinal
MUCINAS MUCINAS
Las mucinas contienen diferentes oligosacáridos, incluyendo las glicoproteínas secretadas y de superficie celular. Las mucinas secretadas (MUC2, MUC5 y MUC6) forman un gel hidratado de 300 a 700 μm de espesor que tiene dos capas:
Capa externa: menos densa y, normalmente, colonizada por bacterias.
Capa interna: más densa, unida al epitelio y libre de bacterias.
Representación esquemática del epitelio de la mucosa intestinal.
Nº 7 | Abril 2023 INMUNOLOGÍA El intestino: el órgano inmunitario más grande del organismo 6
CÉLULAS DE PANETH CÉLULAS DE PANETH
Las células de Paneth, situadas en las criptas del intestino delgado, secretan α-defensina, mientras que las células epiteliales presentes en las criptas intestinales del colon secretan β-defensinas, algunas de forma constitutiva y otras en respuesta a la citoquina proinflamatoria IL-1 (Abbas et al., 2015).
Además, las células de Paneth secretan una lectina de tipo C, la proteína regeneradora de epitelio derivada de islote 3 γ (REGIIIγ), que bloquea la colonización bacteriana en la superficie epitelial.
REGIII γ Y SU HOMÓLOGA REGIII α SE UNEN AL PEPTIDOGLICANO DE LAS BACTERIAS GRAM POSITIVAS
La expresión de REGIIIγ requiere señales de los receptores toll-like (TLR) en respuesta a organismos comensales y su producción aumenta tras la colonización y la infección con patógenos (Abbas et al., 2015).
PATRONES MOLECULARES DE RECONOCIMIENTO
La respuesta inmunitaria innata se inicia por mediación de patrones moleculares de reconocimiento asociados a patógenos (PAMP) y receptores celulares de reconocimiento de patrones (PRR).
En los mamíferos, los TLR (un tipo de PRR) desempeñan un papel esencial en el reconocimiento de componentes microbianos y en el desencadenamiento de la respuesta inmunitaria innata. Estos receptores se clasifican en dos subgrupos, dependiendo de su localización celular y especificidad hacia sus respectivos PAMP:
RECEPTORES TLR-1, TLR-2, TLR-4, TLR-5, TLR-6 Y TLR-10
Se expresan en las superficies celulares y reconocen componentes de la membrana microbiana como lípidos, lipoproteínas y proteínas.
RECEPTORES TLR-3, TLR-7, TLR-8 Y TLR-9
Se expresan exclusivamente en vesículas intracelulares y reconocen el ADN microbiano.
(Cario, 2005)
7 SALUD INTESTINAL
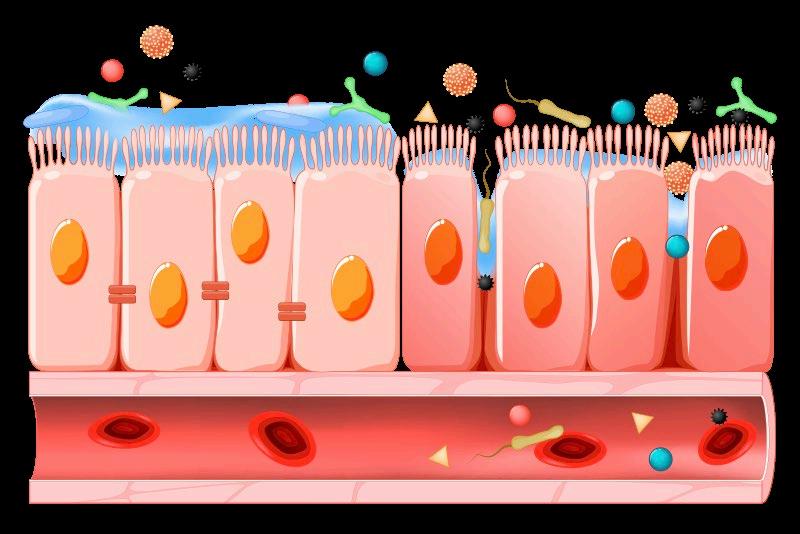
La alteración de la integridad de la barrera epitelial (Figura 2), caracterizada por un aumento de la permeabilidad intestinal, permite la invasión tisular por parte de bacterias patógenas, así como un aumento de la presencia de antígenos intactos procedentes de la dieta en la mucosa intestinal, lo que conduce a un aumento de la síntesis de citoquinas inflamatorias por parte de las células del sistema inmunitario de la mucosa, pudiendo dar lugar a una respuesta inmunitaria exacerbada y patológica que culmina en un proceso de inflamación intestinal (Gonçalves et al., 2016).
En estas condiciones, para estar eficazmente protegido, el organismo del animal debe poseer sistemas de defensa que detecten y eliminen eficazmente a los microorganismos invasores, preferiblemente sin ocasionar daños a los tejidos, estando esta función a cargo del sistema inmunitario adaptativo (Levinson, 2016).
FIGURA 2
La pérdida de la integridad intestinal conlleva una mayor permisividad al paso de microorganismos y antígenos desde el lumen intestinal a la circulación sanguínea, con la consiguiente activación de una respuesta inmunitaria e inflamatoria que puede ocasionar daños a los tejidos circundantes.

Nº 7 | Abril 2023 INMUNOLOGÍA El intestino: el órgano inmunitario más grande del organismo 8
TEJIDO LINFOIDE ASOCIADO A LAS MUCOSAS
En el tracto gastrointestinal, las respuestas inmunitarias adaptativas se inician en conjuntos organizados de linfocitos y células presentadoras de antígenos estrechamente asociados con el revestimiento epitelial de la mucosa del intestino, así como en los ganglios linfáticos mesentéricos (Abbas et al., 2015).
Los tejidos linfoides organizados no encapsulados formados por células del sistema inmunitario que se encuentran asociadas a las superficies mucosas de los tractos respiratorio, gastrointestinal y urogenital se denominan colectivamente como tejido linfoide asociado a las mucosas (MALT, mucosa-asssociated lymphoid tissue).
PLACAS DE PEYER
Específicamente, en el caso de la mucosa del TGI se encuentra el tejido linfoide asociado al intestino (GALT, gut-asssociated lymphoid tissue), un sistema formado por tejido linfoide denso (Gonçalves et al., 2016).
En el GALT, el tejido linfoide se distribuye a lo largo de estructuras como:
Ganglios linfáticos mesentéricos
Placas de Peyer
Agregados linfoides más difusos (Vega-López et al., 2012)
Además de estas estructuras, el GALT está también constituido por células presentadoras de antígenos, células dendríticas y macrófagos, áreas de linfocitos T y B con centros germinales en la lámina propia y células Natural Killer (NK) (Cunha, 2013).
Entre las estructuras que componen el GALT destacan las placas de Peyer que se encuentran, principalmente, en el íleon distal estructuradas en forma de folículos linfoides con centros germinales que contienen linfocitos B, linfocitos T colaboradores, células dendríticas y macrófagos. Estos centros germinales están rodeados de linfocitos B inmaduros que producen IgM e IgD.
Una región denominada cúpula, situada entre los folículos y el epitelio de revestimiento, contiene linfocitos B y T, células dendríticas y macrófagos (Abbas et al., 2015).
9 SALUD INTESTINAL
CÉLULAS M
La principal vía de distribución de antígenos desde el lumen hasta el GALT se produce a través de células especializadas, las células M (microfold), localizadas en la cúpula o epitelio asociado a folículos (FAE) (Abbas et al., 2015).
Estas células presentan características morfológicas únicas, como la presencia de un glicocálix reducido, un borde en cepillo irregular y microvellosidades pequeñas, estando altamente especializadas en la fagocitosis y transcitosis de macromoléculas del lumen intestinal, antígenos en forma de partículas y microorganismos patógenos o comensales a través del epitelio (Mabbott et al., 2013).
A pesar de que las células M desempeñan un papel primordial en la respuesta inmunitaria frente a microorganismos luminales, algunos microorganismos patógenos han evolucionado para sortear el mecanismo de acción de las células M, utilizándolas como vía de invasión a través de la barrera mucosa.
El ejemplo mejor descrito de este fenómeno es el de Salmonella typhimurium. Las células M expresan lectinas que permiten la unión específica de estas bacterias y su posterior fagocitosis, pero, debido a sus propiedades citotóxicas para las células M, conducen a un aumento de la permeabilidad intestinal y favorecen la proliferación de microorganismos (Abbas et al., 2015).
PROCESAMIENTO Y PRESENTACIÓN DE ANTÍGENOS
Las células dendríticas y los macrófagos son células centinela y procesadoras de antígenos, por lo que el procesamiento del antígeno se suele iniciar de forma simultánea a la eliminación del invasor por parte de las defensas innatas.
Tras la fagocitosis, los microorganismos invasores son procesados intracelularmente y fragmentados en péptidos más pequeños que se unen a moléculas del Complejo Mayor de Histocompatibilidad (CMH).
Estas moléculas están especializadas en la presentación de antígenos, transportándolos a la superficie celular para que sean reconocidos por receptores linfocitarios específicos, desencadenando una respuesta inmunitaria adaptativa cuando sea necesaria (Tizard, 2008).
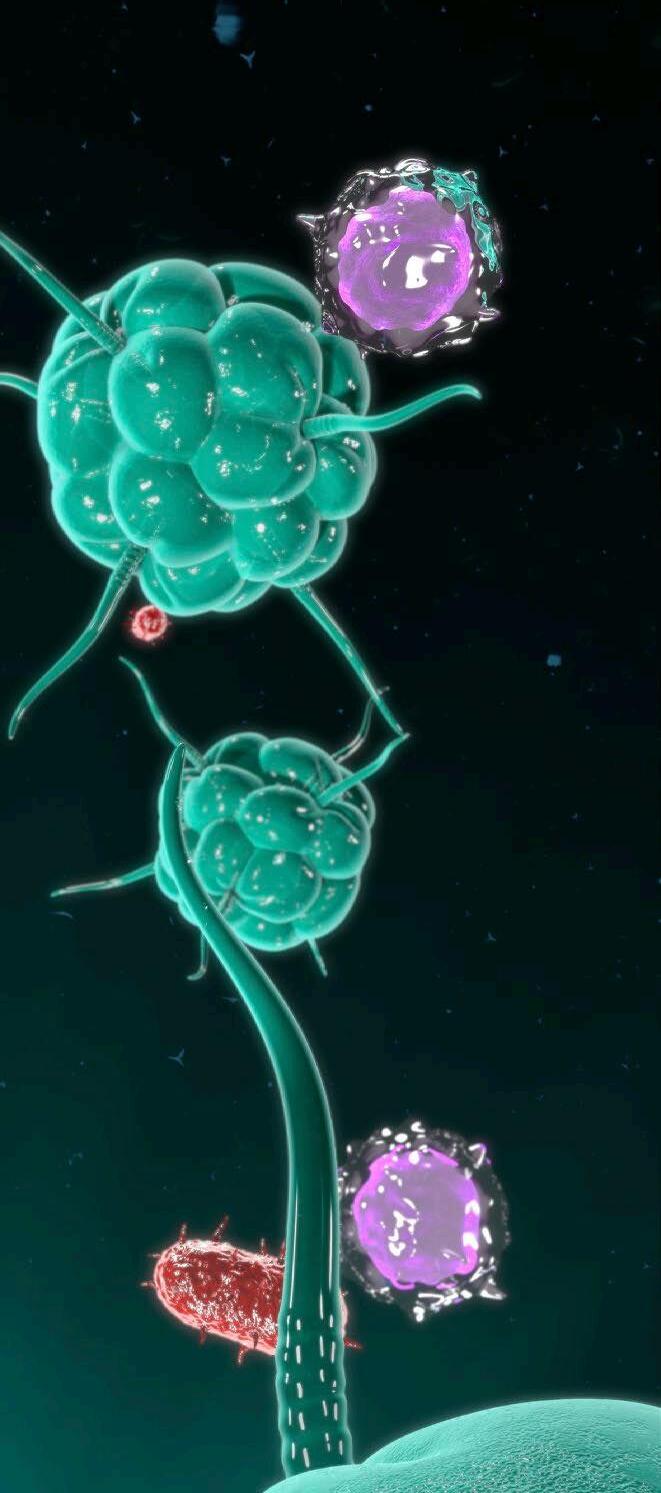
Nº 7 | Abril 2023 INMUNOLOGÍA El intestino: el órgano inmunitario más grande del organismo 10
Fagocitosis y procesamiento de microorganismo
Linfocito T
Tras la fagocitosis de un patógenos por parte de un macrófago, los péptidos resultantes de su fragmentación se presentan a los linfocitos T a través de moléculas CMH.
Las prolongaciones citoplasmáticas de algunas células dendríticas se intercalan entre las células epiteliales, lo que les permite captar, procesar y presentar antígenos directamente desde el lumen a los linfocitos intraepiteliales y a los linfocitos T de los folículos linfoides subyacentes.
LINFOCITOS T
Existen linfocitos T (linfocitos T CD4 colaboradores, linfocitos T CD8 citotóxicos y células plasmáticas secretoras de IgA) previamente activados/diferenciados en el GALT o en los ganglios linfáticos de drenaje (Gonçalves et al., 2016).
Además, ciertas poblaciones de linfocitos T se localizan específicamente encima de la lámina propia y la membrana basal del epitelio intestinal, situándose entre las células epiteliales y presentando características diferentes de otros linfocitos T que se encuentran en la región periférica.
Estos linfocitos son heterogéneos, mayoritariamente (80%) de fenotipo CD8, con abundantes gránulos citoplasmáticos que contienen moléculas citotóxicas, y capaces de producir varias citoquinas (IFN-γ, IL-2, IL-4 o IL-17) y de dividirse en poblaciones celulares que expresan en su superficie receptores de antígenos (TCR) formados por cadenas de tipo αβ o γδ (Gonçalves et al., 2016).
LOS LINFOCITOS T INTRAEPITELIALES PREDOMINANTES
CORRESPONDEN A CD8αβ +/TCR αβ+, QUE PENETRAN EN EL EPITELIO
INTESTINAL GRACIAS A UNA MAYOR EXPRESIÓN DE INTEGRINAS ESPECÍFICAS
Y RECEPTORES DE QUIMIOQUINAS TRAS SU ACTIVACIÓN EN ÓRGANOS
LINFOIDES SECUNDARIOS (GONÇALVES ET AL., 2016)
FIGURA 3
Molécula MHC
SALUD INTESTINAL 11
Presentación de antígeno Macrófago
Por su parte, los linfocitos T intraepiteliales TCRγδ+ desempeñan funciones primordiales en la mucosa intestinal, entre ellas:
Mantenimiento de la función de barrera epitelial (control del crecimiento de las células epiteliales, secreción de TGF-β).
Mantenimiento de la homeostasis tisular.
Protección frente a patógenos intestinales (producción de citoquinas inflamatorias, citotoxicidad).
(Gonçalves et al., 2016)
Además de los linfocitos T intraepiteliales, la mucosa intestinal también contiene otras poblaciones de linfocitos T, localizados dentro de los GALT o de forma difusa a lo largo de la lámina propia del intestino.
Los linfocitos T son esencialmente CD4+ o CD8+ (ambos TCRαβ+), presentando un fenotipo de memoria (CD45RD) y asociándose a la mucosa intestinal a través de la expresión de integrinas específicas (α4β7) y receptores de quimioquinas (CCR9) que dirigen a estos linfocitos tras su activación y diferenciación a células efectoras (Gonçalves et al., 2016).
Para que se produzca una respuesta inmunitaria adaptativa frente a los antígenos entéricos, es necesario que los linfocitos T intravasculares naive pasen al GALT y a los ganglios linfáticos mesentéricos de drenaje, donde se activarán, produciéndose su expansión clonal, polarización/diferenciación en células efectoras Th1 y/o Th17 tras la interacción inicial con la célula presentadora del antígeno.
A continuación, los linfocitos T efectores abandonan el tejido linfoide a través de los vasos linfáticos eferentes, pasando a la circulación sanguínea sistémica y regresando de nuevo al intestino donde colaborarán en la eliminación de un antígeno determinado (Gonçalves et al., 2016).
Al regresar al intestino en un proceso denominado homing, los linfocitos T efectores vuelven a exponerse a los antígenos que desencadenaron la respuesta inmunitaria que, a su vez, ya está amplificada y cuenta con una población más diversa de células presentadoras de antígenos, como:
Macrófagos
Linfocitos B
Células dendríticas
Tras este nuevo contacto con el antígeno (promovido por las células presentadoras de antígeno), los linfocitos T efectores responden más rápida y vigorosamente, secretando citoquinas como IFN-γ, IL-17, TNF-α, linfotoxina-α o IL-2, dependiendo del perfil del linfocito T efector (Th1 o Th17).
Nº 7 | Abril 2023 INMUNOLOGÍA El intestino: el órgano inmunitario más grande del organismo 12
CADA CITOQUINA TIENE UNA FUNCIÓN ESPECÍFICA EN
LA COORDINACIÓN DE LA RESPUESTA INMUNITARIA DESENCADENADA
El IFN-γ estimula a las células presentadoras de antígeno para que produzcan IL-12 y, específicamente en los macrófagos, activa la producción de otras citoquinas inflamatorias como IL-1, IL-6, IL-8 IL-18 y TNF-α, así como especies reactivas de oxígeno y nitrógeno (Gonçalves et al., 2016).
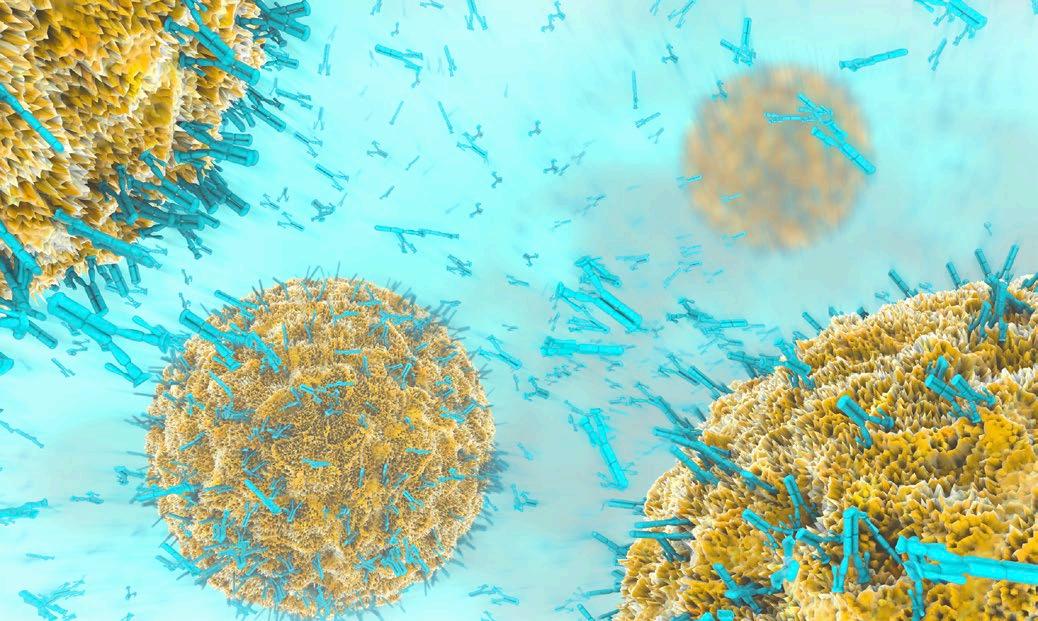
La IL-2 estimula el crecimiento y la proliferación de los linfocitos T y B, así como la producción de otras citoquinas, como el IFN-γ y el TNF-β, lo que resulta en la activación de monocitos, neutrófilos y células NK (Oliveira et al., 2011).
LINFOCITOS B
Los linfocitos B desempeñan un papel fundamental en la respuesta inmunitaria de las mucosas mediante la producción de anticuerpos.
Estas células se activan en los ganglios linfáticos mesentéricos, los folículos linfoides aislados de la mucosa intestinal y en las placas de Peyer, diferenciándose en células productoras de anticuerpos del isotipo IgA en su forma dimérica (IgA/S-IgA secretoras) o multimérica.
Las IgA desempeñan un papel importante en la protección del epitelio de la mucosa frente a patógenos invasores, modulando la composición de la microbiota intestinal y manteniendo la homeostasis frente a antígenos comensales y de origen alimentario (Gonçalves et al., 2016).
SALUD INTESTINAL 13
Las SIgA se unen a bacterias comensales en el lumen intestinal y permiten su transporte a la mucosa mediante la unión del complejo SIgA/ antígeno a receptores específicos expresados en las células M intestinales.
Posteriormente, se estimula a las células dendríticas de fenotipo tolerante de la mucosa para que produzcan IL-10 e induzcan la producción de IgA por parte de los linfocitos B de las placas de Peyer.
Este mecanismo de "mostrar" microorganismos comensales a las células del sistema inmunitario en un microambiente regulador (antiinflamatorio) permite mantener la tolerancia a la microbiota intestinal (Gonçalves et al., 2016).
EL INTESTINO, ADEMÁS DE SER EL ÓRGANO RESPONSABLE DE LOS PROCESOS DE DIGESTIÓN Y ABSORCIÓN DE NUTRIENTES, DESEMPEÑA
UN PAPEL PRIMORDIAL EN LA DEFENSA Y LA RESPUESTA INMUNITARIA
DEL ORGANISMO DE FORMA QUE EL MANTENIMIENTO DE LA SALUD
INTESTINAL SE VE REFLEJADO EN LA SALUD Y EL RENDIMIENTO
PRODUCTIVO DE LOS ANIMALES
BIBLIOGRAFÍA
Abbas, A. K.; Lichtman, A. H.; Pillai, S. Imunidadade Regional: Respostas imunes especializadas em tecidos epiteliais e imunoprivilegiados. Imunologia celular e molecular. 8. ed. Rio de Janeiro: Elsevier, 2015.
Gonçalves, J. L.; Yaochhite, J. N. U.; Queiroz, C. A. A.; Câmara, C. C.; Oriá, R. B. Bases do sistema imunológico associado à mucosa intestinal. Em: Sistema digestório: integração básico-clínica. Editora Edgard Blücher Ltda., ISBN 978-85-8039-189-3, p. 369-377, 2016.
Cario, E. Bacterial interactions with cells of the intestinal mucosa: Toll-like receptors and NOD2. Gut, v. 54, n. 8, p. 1182-1193, 2005.
Cunha, J. A. C. Importância do tecido linfóide associado ao intestino na infecção pelo VIH e suas implicações terapêuticas. Dissertação de mestrado, Universidade do Porto, Porto, p. 14, 2013.
Levinson, W. Microbiologia Médica e Imunologia. 13.ed. Porto Alegre: Amgh, 2016.
Mabbott, N. A.; Donaldson, D. S.; Ohno, H.; Williams, I.R.; Mahajan, A. Microfold (M) cells: important immunosurveillance posts in the intestinal epithelium. Mucosal Immunol., v. 6, n. 4, p. 666-677, 2013.
Nature Video. Immunology in the Gut Mucosa. Disponível em: https://www.youtube.com/watch?v=gnZEge78_78. Acessado em: 26 de junho de 2019. Publicado em 5 de fev de 2013.
Oliveira, C. M. B.; Sakata, R. K.; Issy, A. M.; Gerola, L. R.; Salomão, R. Citocina e dor. Rev Bras Anestesiol., v. 61, n. 2, p. 255-265, 2011.
Rodrigues, F. A. P.; Medeiros, P. H. Q. S.; Prata, M. M.; Lima, A. A. M. Fisiologia da barreira epitelial intestinal. Em: Sistema digestório: integração básico-clínica. Editora Edgard Blücher Ltda., ISBN 978-85-8039-1893, p. 441-475, 2016.
Sang, Y.; Yang, J.; Ross, C. R.; Rowland, R. R.; Blecha, F. Molecular identification and functional expression of porcine Toll-like receptor (TLR) 3 and TLR7. Vet. Immunol. Immunopathol., v.125, n. 1-2, p.162-167, 2008.
Shimosato T.; Tohno, M.; Kitazawa, H.; Katoh, S.; Watanabe, K.; Kawai, Y., et al. Toll-like receptor 9 is expressed on follicle-associated epithelia containing M cells in swine Peyer's patches. Immunol. Lett., v. 98, n. 1, p.83-89, 2005.
Shinkai, H.; Muneta, Y.; Suzuki, K.; Eguchi-Ogawa, T.; Awata, T.; Uenishi, H. Porcine Toll-like receptor 1, 6, and 10 genes: complete sequencing of genomic region and expression analysis. Mol. Immunol., v. 43, n. 9, p. 1474-1480, 2006.
Tizard, I.R. Imunologia Veterinária: Uma Introdução. 8. ed. Rio de Janeiro: Elsevier, 2008.
Tohno, M.; Shimosato, T.; Kitazawa, H.; Katoh, S.; Iliev, I. D.; Kimura, T., et al. Toll-like receptor 2 is expressed on the intestinal M cells in swine. Biochem. Biophys. Res. Commun. v. 330, n. 2, p. 547- 554, 2005.
Tuner, J. R. Intestinal mucosal barrier function in health and disease. Nat Rev Immunol., v. 9, n. 11, p. 799-809, 2009.

Uenishi, H.; Shinkai, H. Porcine Toll-like receptors: the front line of pathogen monitoring and possible implications for disease resistance. Dev. Comp. Immunol., v. 33, n.3, p.353-361, 2009.
Vega-López, M. A.; Cole, M. F.; Bellanti, J. A. The Mucosal Immune System in Health and Disease. Em: Immunology IV Clinical Applications in Health and Disease. 4th ed. Bethesda, p. 255-70, 2012.
Zhang, L.; Liu, J.; Bai, J.; Wang, X.; Li, Y.; Jiang, P. Comparative expression of Toll-like receptors and inflammatory cytokines in pigs infected with different virulent porcine reproductive and respiratory syndrome virus isolates. Virol. J., v.10, p. 135, 2013.
14 Nº 7 | Abril 2023 El intestino: el órgano inmunitario más grande del organismo INMUNOLOGÍA
INFLUENCIA DEL PESO CORPORAL SOBRE LA EXPRESIÓN GÉNICA INTESTINAL DE LOS LECHONES
G
Sandra VillagómezEstrada1,2 *, José F. Pérez1, Diego Melo-Durán1,3 , Francesc Gonzalez-Solè1, Matilde D’Angelo1, Francisco
J. Pérez-Cano 4 y David
Solà-Oriol1
1Departamento de Ciencia Animal y de los Alimentos, Servicio de Nutrición y Bienestar Animal (SNIBA), Universidad Autónoma de Barcelona
2Carrera de Medicina Veterinaria, Escuela Superior Politécnica de Chimborazo (ESPOCH)
3Facultad de Ciencias Médicas
“Eugenio Espejo”, Universidad UTE
4Departamento de Bioquímica y Fisiología, Universidad de Barcelona
*Autor correspondiente: sandra.villagomez@outlook.es
n las actuales condiciones de producción con cerdas hiperprolíficas, los lechones recién nacidos y lactantes de bajo peso corporal tienen desventajas fisiológicas en comparación con sus compañeros de camada de peso medio. Para comprender mejor el impacto que tiene el peso corporal en los lechones, el presente estudio evaluó el grado de debilidad fisiológica de los lechones más pequeños al nacimiento y durante el periodo de lactación (20 días) en comparación con sus compañeros de camada de peso medio a través de su expresión génica yeyunal.
Los lechones recién nacidos de bajo peso presentaron una reducción en la expresión de genes relacionados con la inmunidad, la actividad antioxidante y la actividad digestiva, pero también una tendencia a una mayor expresión de otros genes relacionados con la inflamación y la respuesta al estrés. Al final del periodo de lactación, estas diferencias en la expresión de los genes desaparecieron, apareciendo otras. Los lechones de peso ligero mostraron una menor expresión de genes implicados en la función de barrera y en las respuestas a la inflamación y al estrés, en comparación con sus compañeros de camada de peso medio.
GENÉTICA 15
EEDescarga el PDF
EL RETO DE LA PRODUCCIÓN PORCINA EN LA ERA DE LA CERDA HIPERPROLÍFICA
La intensa selección genética a favor de la hiperprolificidad basada en el rendimiento reproductivo de la cerda (tamaño de la camada; 16-20 lechones nacidos totales) ha conducido a una reducción del peso medio de los lechones al nacimiento, así como a un aumento de la variabilidad de los pesos al nacimiento dentro de la camada (Peltoniemi et al., 2021).
Así, la proporción de lechones pequeños presentes en camadas grandes, así como la frecuencia de aparición de lechones con crecimiento intrauterino restringido (IUGR, por sus siglas en inglés, Intrauterine GrowthRestriction) ha aumentado notablemente (Matheson et al., 2018; Ward et al., 2020).
Las variaciones iniciales de peso al nacimiento dentro de una camada revelan que las cerdas de genéticas hiperprolíficas no son capaces de asegurar una nutrición satisfactoria y equitativa de todos sus lechones, lo que resulta en una intensa competencia pre- y postnatal por unos nutrientes limitados y un alto riesgo de mortalidad para los más débiles (Ward et al., 2020).
La mayoría de los lechones con IUGR y bajo peso al nacimiento también suelen padecer efectos negativos a largo plazo en la estructura de sus órganos, en el crecimiento postnatal y en la eficiencia alimentaria (Ji et al., 2017).
Por ello, es frecuente que en las explotaciones comerciales modernas se tengan que sacrificar lechones IUGR al no existir un apoyo nutricional o de gestión eficaz para garantizar su crecimiento o supervivencia durante los periodos de lactación y postdestete (Kraeling y Webel, 2015). No obstante, los lechones de bajo peso al nacimiento podrían experimentar un crecimiento compensatorio si se les proporciona un manejo y unas condiciones dietéticas óptimas (van Barneveld y Hewitt, 2016; Viott et al., 2018; Farmer y Edwards, 2021).
16 LECHONES Nº 7 | Abril 2023 Influencia del peso corporal sobre la expresión génica intestinal de los lechones
IMPACTO DE LA PROGRAMACIÓN FETAL Y
EL CRECIMIENTO INTRAUTERINO SOBRE LA EXPRESIÓN GÉNICA INTESTINAL
Durante el periodo de gestación se produce una programación fetal que conlleva cambios en la transcripción de los genes, lo que resulta en una alteración de determinadas vías metabólicas y procesos de control homeostático (Burdge et al., 2007).
DEFICIENCIAS EN LA NUTRICIÓN MATERNA DURANTE LA GESTACIÓN
PUEDEN RESULTAR EN ALTERACIONES
PERMANENTES EN LA PROGRAMACIÓN FETAL (Kwon y Kim, 2017)
Con el fin de comprender mejor el impacto que tiene la sobrepoblación fetal intrauterina en los lechones, en el presente estudio se propuso evaluar los cambios en la expresión génica yeyunal como indicador clave de la función y el desarrollo intestinal.
Para ello, se comparó la expresión génica de lechones de bajo peso al nacimiento y al final del periodo de lactación con la de sus compañeros de camada de peso medio, todos ellos nacidos de cerdas hiperprolíficas en condiciones comerciales.
MATERIALES & MÉTODOS
A los 113 días postcubrición (±1,27 días) se registraron las características de 80 camadas (1.542 lechones nacidos en total) de 80 cerdas hiperprolíficas y se realizó un seguimiento de las cerdas y sus camadas hasta el destete (21,6 ± 1,27 días).
Las cerdas recibieron 2,4 kg de pienso/día desde el destete hasta los 35 días de gestación y 2,1 kg de pienso/día desde los 35 días de gestación hasta los 110 días de gestación.
El procesamiento de los lechones se realizó dentro de las 48 h posteriores al nacimiento y se realizaron adopciones cruzadas para estandarizar el tamaño de la camada a 15 lechones/camada.
ANIMALES & ALOJAMIENTO ANIMALES & ALOJAMIENTO
GENÉTICA 17
DIETA EXPERIMENTAL & MEDICIONES MUESTREO DIETA EXPERIMENTAL & MEDICIONES MUESTREO
El peso corporal de las cerdas se registró a los 35 y 110 días de gestación y al destete. Además, durante este periodo, se registró diariamente la ingesta individual de pienso a través de estaciones de alimentación electrónica, mientras que durante la lactación la ingesta se registró manualmente ponderando diariamente la diferencia entre el pienso ofrecido y su desaparición.
Las dietas se formularon para cubrir las necesidades nutricionales (NRC, 2012).
DIETA DE GESTACIÓN
La composición de la dieta de gestación se basó en trigo (35%), cebada (23,5%), salvado de trigo (20%), torta de girasol (10%), maíz (7,70%) y lisina 50 (0,44%).
El resto de ingredientes, aminoácidos y macro y microminerales representaron el 3,36% de la composición total.
El contenido en nutrientes de la dieta de gestación fue de 2.260 kcal/kg de energía neta, 13,0% de proteína bruta y 0,67% de lisina.
DIETA DE LACTACIÓN
La dieta de lactación incluyó como ingredientes principales trigo (37%), maíz (30%), harina de soja (11,5%), torta de girasol (7%), lisina 50 (0,92%) y otros ingredientes (13,58%).
Los valores de energía neta de la dieta de lactación fueron de 2.450 kcal/kg, 15,5% de proteína bruta y 1,08% de lisina.
Del total de 80 cerdas alojadas inicialmente, se eligió un subconjunto de 10 cerdas multíparas (3ª a 5ª paridad) para el muestreo. Se seleccionaron dos lechones de cada camada para tomar muestras de yeyuno durante el parto (antes de la toma de calostro) y durante la lactación (20 días).
El criterio de selección fue el peso corporal de los lechones, clasificado en dos niveles:
Lechones de peso bajo: 600800 g al nacimiento (cuartil inferior) y 2.500-3.800 g a los 20 días de vida.

Lechones de peso medio: 1.200-1.300 g al nacimiento y 4.000-5.100 g al final de la lactación.
Las muestras para el análisis de expresión génica se tomaron según lo descrito en Villagómez-Estrada et al. (2021), obteniéndose a nivel del punto medio del yeyuno con un tamaño aproximado de 1,5 cm, enjuagándose en solución de PBS, congelándose posteriormente en 1 ml de RNAlater (Deltalab, Barcelona, España) y almacenándose a -80 °C hasta su procesamiento.
18 LECHONES Nº 7 | Abril 2023 Influencia del peso corporal sobre la expresión génica intestinal de los lechones
ANÁLISIS DE EXPRESIÓN GÉNICA ANÁLISIS DE EXPRESIÓN GÉNICA
Se analizó la expresión de 56 genes en el tejido yeyunal, todos ellos implicados en múltiples funciones fisiológicas estrechamente relacionadas con la salud intestinal y seleccionados a partir de la bibliografía y agrupados según su función principal de la siguiente manera:
GENES QUE INTERVIENEN EN FUNCIONES DE BARRERA
Miembros de la familia de las claudinas (CLDN)
Mucinas (MUC)
Zonula occludens (ZO)
Factor trefoil (TFF)
Ocludinas (OCLN): CLDN1, CLDN4, CLDN15, MUC2, MUC13, ZO1, TFF3 y OCLN
4
GENES DE ENZIMAS ANTIOXIDANTES
Glutatión peroxidasa (GPX2)
Superóxido dismutasa (SOD2)
GENES DE ENZIMAS DIGESTIVAS Y HORMONAS QUE INTERVIENEN EN LOS PROCESOS DE DIGESTIÓN Y METABOLISMO
Fosfatasa alcalina intestinal (ALPI)
Sucrasa-isomaltasa (SI)
GENES IMPLICADOS EN RESPUESTAS
INMUNITARIAS E INFLAMATORIAS (receptores de reconocimiento de patrones, citoquinas, quimioquinas y proteínas de estrés):
Receptores Toll-Like (TLR2, TLR4)
Interleucinas (IL-1β, IL6, IL8, IL10, IL17A, IL22)
Interferón gamma (IFNγ, IFNγR1)
Factor de necrosis tumoral (TNF)
Factor de crecimiento transformante beta 1 (TGF-β1)
Ligando de quimiocinas (CCL20, CXCL2)
Proteína de choque térmico (HSPB1, HSPA4)
Regenerating family member 3 gamma (REG3G)
Receptor gamma activado por el proliferador de peroxisomas (PPARGC1α)
Dominio hidroxilasa de ácidos grasos (FAXDC2)
Proteína de unión a guanilato (GBP1)
D-aminoácido oxidasa (DAO1)
Histamina N-metiltransferasa (HNMT)
Alanil aminopeptidasa de membrana (ANPEP)
Indoleamina 2,3-dioxigenasa (IDO1)
Glucagón (GCG)
Colecistoquinina (CCK)
Factor de crecimiento similar a la insulina (IGF1R)
1 3 2 GENÉTICA 19
Péptido YY (PYY)
5 6
GENES CODIFICANTES DE TRANSPORTADORES DE NUTRIENTES
Familia de transportadores de solutos (SLC5A1, SLC16A1, SLC7A8, SLC15A1, SLC13A1, SLC11A2, SLC30A1, SLC39A4)
Metalotioneína (MT1A)
GENES DE RESPUESTA AL ESTRÉS
Receptor de la hormona liberadora de corticotropina (CRHR1)
Receptor nuclear (NR3C1)
Hidroxiesteroide (11-beta) deshidrogenasa (HSD11β1)
Para calcular los valores relativos de los datos genéticos se utilizaron cuatro genes de control:
Actina beta (ACTB)
Beta-2-microglobulina (B2M)
Gliceraldehído-3-fosfato deshidrogenasa (GAPDH)
Proteína de unión a TATA (TBP)
El análisis de la expresión génica del ARN se realizó según la metodología descrita en Villagómez-Estrada et al., (2021).
A partir de 50 mg de tejido de yeyuno congelado, se obtuvo el ARN, evaluándose la calidad, cantidad e integridad del mismo.
A continuación, se llevó a cabo la transcripción inversa de aproximadamente 1 μg de ARN total a ADNc monocatenario en un volumen final de 20 μL utilizando un kit de transcripción inversa de ADNc de alta capacidad y cebadores hexaméricos aleatorios, ajustándose ciclo umbral máximo relativo a 26.
20 LECHONES Nº 7 | Abril 2023 Influencia del peso corporal sobre la expresión génica intestinal de los lechones
RESULTADOS
RENDIMIENTO & PESO CORPORAL RENDIMIENTO & PESO CORPORAL
Los resultados del rendimiento de las cerdas y sus camadas se muestran en la Tabla 1.
El peso corporal medio de los lechones de peso bajo y medio muestreados al nacimiento fue de 741 y 1.367 g, mientras que al final del periodo de la lactación fue de 3.125 y 4.733 g, respectivamente.
TABLA 1
Rendimiento reproductivo y crecimiento de las cerdas y los lechones durante el periodo de estudio.
1Los lechones nacidos totales incluyen los nacidos vivos y los nacidos muertos.
2Los datos son medias aritméticas de 10 lechones por categoría y periodo.

PARÁMETRO MEDIA DESVIACIÓN ESTÁNDAR Cerdas (n) 80Peso corporal Cerdas (kg) Gestación (día 35) 248,3 36,97 Gestación (día 110) 270,9 28,80 Destete (día 24) 259,7 33,32 Ingesta media diaria de pienso durante la gestación (kg) 2,33 0,09 Ingesta media diaria de pienso durante la lactación (kg) 7,45 1,22 Parámetros reproductivos Al nacimiento Lechones nacidos totales1 (n) 19,3 4,02 Lechones nacidos vivos (n) 16,1 3,44 Peso al nacimiento de la camada de nacidos vivos (kg) 21,0 3,90 Peso del lechón nacido vivo (kg) 1,3 0,20 Al destete Lechones destetados totales (n) 14,2 1,69 Peso de la camada al destete (kg) 74,7 14,63 Peso medio del lechón al destete (kg) 5,3 0,82 Peso corporal Q1 3,6 0,52 Peso corporal Q2 4,8 0,27 Peso corporal Q3 5,7 0,23 Peso corporal Q4 7,0 0,73 Al muestreo² Camadas (n) 10Peso bajo al nacimiento (kg) 0,7 0,10 Peso medio al nacimiento (kg) 1,3 0,17 Peso bajo al destete (kg) 3,1 0,48 Peso medio al destete (kg) 4,7 0,34 GENÉTICA 21
EXPRESIÓN GÉNICA INTESTINAL EXPRESIÓN GÉNICA INTESTINAL
En la Tabla 2 se muestra el impacto del peso corporal del lechón sobre la expresión intestinal de algunos genes implicados en múltiples funciones fisiológicas.
Al nacimiento, los lechones de bajo peso presentaron una reducción en la expresión de los genes relacionados con la respuesta inmunitaria (FAXDC2, HSPB1, PPARGC1α), así como genes relacionados con enzimas antioxidantes (SOD2m), enzimas digestivas (ANPEP, IDO1, SI) y el transportador de nutrientes (SLC39A4) (P < 0,05).
En cambio, en estos mismos lechones se observó una tendencia a una mayor expresión del ARNm de GBP1 (respuesta inmunitaria/ inflamatoria) y HSD11β1 (enzima relacionada con la respuesta al estrés) en comparación con sus compañeros de camada de mayor peso (P < 0,10). De hecho, la tendencia hacia una mayor expresión génica de HSD11β1 en los lechones de bajo peso también se observó al final del periodo de lactación.
TABLA 2
Expresión génica relativa en lechones de peso bajo y medio al nacimiento y al destete (20 días)1
AL DESTETE, LOS LECHONES DE BAJO PESO PRESENTARON UNA MAYOR EXPRESIÓN DE GENES IMPLICADOS EN LA FUNCIÓN DE BARRERA (CLDN1), LA RESPUESTA PROINFLAMATORIA (CXCL2, IL6), LA DIGESTIÓN (IDO1) Y LA SEÑALIZACIÓN DE LA HORMONA DEL ESTRÉS (HSD11β1)
1Los datos son medias de 10 lechones para cada categoría y corresponden a valores retrotransformados. Los valores de expresión génica se indican como cocientes del valor del ciclo umbral relativo para cada gen normalizado con respecto al de la muestra de referencia. Solo se presentan las diferencias significativas (valor P < 0,05) y de tendencia (valor P < 0,10)
SEM, error estándar de la media; FDR, tasa de falsos descubrimientos.
Genes con expresión disminuida en los lechones de bajo peso Genes con expresión aumentada en los lechones de bajo peso
FUNCIÓN GEN PESO AL NACIMIENTO PESO AL DESTETE Bajo Medio SEM P FDR Bajo Medio SEM P FDR Función de barrera Claudina-1 CLDN1 1,10 0,94 0,071 0,322 0,650 2,41 1,02 0,091 0,46 0,415 Respuesta inmunitaria Dominio hidrolasa de ácidos grasos 2 FAXDC2 1,70 3,12 0,063 0,010 0,138 107,57 129,83 0,152 0,702 0,887 Proteína de unión a guanilato 1 GBP1 0,31 0,06 0,141 0,062 0,319 1,53 1,19 0,093 0,806 0,907 Proteína de choque térmico 1 HSPB1 1,27 1,81 0,029 0,002 0,083 3,47 3,33 0,045 0,970 0,992 Receptor gamma activado por el proliferador de peroxisomas, coactivador 1 alfa PPARGC1α 2,87 4,79 0,062 0,012 0,138 2,73 2,67 0,102 0,391 0,815 Interleucina 6 IL6 1,18 1,44 0,119 0,517 0,820 6,38 1,90 0,097 0,007 0,165 Ligando de quimiocinas 2 CXCL2 1,20 0,90 0,078 0,263 0,650 10,99 4,08 0,101 0,025 0,283 Enzima antioxidante Superóxido dismutasa SOD2m 1,17 1,37 0,027 0,100 0,462 1,40 1,20 0,059 0,673 0,887 Enzimas digestivas Aminopeptidasa-N ANPEP 0,61 0,72 0,027 0,047 0,311 1,68 1,44 0,092 0,615 0,887 Indoleamina 2,3-dioxigenasa IDO1 1,40 4,72 0,120 0,040 0,310 12,90 3,30 0,115 0,024 0,283 Sucrasa-isomaltasa SI 7,27 55,70 0,171 0,006 0,131 130,06 84,69 0,573 0,206 0,815 Transportador de nutrientes Transportador de solutos de la familia 39, miembro 4 (transportador de Zn) SLC39A4 1,31 2,19 0,074 0,028 0,258 1,88 1,64 0,095 0,752 0,887 Enzima relacionada con el estrés Hidroxiesteroide (11-beta) deshidrogenasa 1 HSD11β1 2,88 1,92 0,029 0,062 0,319 3,60 1,73 0,056 0,002 0,079 Hormona relacionada con el estrés Receptor 1 de la hormona liberadora de corticotropina CRHR1 2,33 2,39 0,094 0,982 0,985 7,15 3,63 0,120 0,060 0,451 Nº 7 | Abril 2023 LECHONES Influencia del peso corporal sobre la expresión génica intestinal de los lechones 22
DISCUSIÓN
La mucosa intestinal posee una función compleja, no solo desempeña un papel importante en la barrera epitelial y la digestión de nutrientes, sino que también forma parte de un sistema inmunitario bien organizado (Okumura y Takeda, 2017).
Al igual que el peso corporal, el desarrollo y la funcionalidad del tracto gastrointestinal pueden verse alterados debido a acontecimientos prenatales (Dong et al., 2014; Farmer y Edwards, 2021).
Esto resulta especialmente relevante en el momento del destete, cuando se producen situaciones de estrés y proinflamatorias.
En el presente estudio, la expresión génica intestinal al nacimiento puso de manifiesto la inmadurez del intestino de los lechones de bajo peso relacionada con una menor activación de genes implicados en la digestión (ANPEP, IDO1, SI), transporte de nutrientes (SLC39A4), inmunidad/ inflamación (FAXDC2, GBP1, HSPB1, PPARGC1α) y función antioxidante (SOD2m), acompañada también de una sobreexpresión del gen de la enzima asociada al estrés HSD11β1.
Los genes digestivos con bajo nivel de expresión, como ANPEP, IDO1 y SI, participan principalmente en el proceso digestivo final de proteínas, aminoácidos y carbohidratos, pero también pueden tener una acción antiinflamatoria (Trevisi et al., 2012; Xu et al., 2015).
Además, la baja expresión del gen transportador de nutrientes (SLC39A4), considerado como el principal transportador intracelular de Zn (Martin et al., 2013), puede estar implicada en los cambios observados en otros genes fisiológicos que requieren Zn para su correcta expresión (Suttle, 2010).
Teniendo en cuenta que el intestino delgado desempeña un papel esencial, no solo en la digestión y absorción de nutrientes, sino también en la respuesta inmunitaria, esta reducida expresión génica observada en lechones de bajo peso al nacimiento puede comprometer seriamente la función inmunitaria (Michiels et al., 2013; Qi et al., 2019; Li et al., 2021).
CABE DESTACAR LA SOBREEXPRESIÓN DEL GEN
HSD11β1 EN LOS LECHONES CON MENOR PESO
CORPORAL, TANTO AL NACIMIENTO COMO AL DESTETE
El gen HSD11β1 codifica para la enzima que transforma la cortisona inactiva en cortisol activo, regulando así los niveles tisulares de glucocorticoides y desempeñando un papel fundamental en el metabolismo y la respuesta inflamatoria (Nixon et al., 2012; Huang et al., 2020).
El hecho de que la expresión de este gen se mantuviera elevado hasta el final de la lactación demuestra que el sufrimiento fetal y el estado inflamatorio neonatal no se revierten con la edad, lo que sugiere que este impacto negativo puede durar hasta edades más avanzadas.
Dado que a los lechones recién nacidos no se les permitió consumir calostro, se necesitan más estudios para evaluar la influencia del calostro, fuente primaria de nutrientes e inmunoglobulinas para los lechones, en la mejora de estas funciones fisiológicas de los lechones deterioradas por la sobrepoblación intrauterina.
Exceptuando el gen HSD11β1, todas las diferencias de expresión génica intestinal observadas inicialmente al nacimiento entre lechones de peso bajo y medio desaparecieron al final del periodo de lactación, lo que indica la consolidación de estas funciones a lo largo de la fase de lactación.
Curiosamente, al final de la lactación, se observó un aumento de la expresión del gen CLDN1 (función de barrera) en los lechones de bajo peso, junto con una sobreexpresión de algunos genes proinflamatorios importantes (IL6 y CXCL2, IDO1) y del receptor de la hormona del estrés (CRHR1, HSD11β1) en comparación con los lechones de mayor peso.
GENÉTICA 23
El aumento de la expresión de CLDN1, un componente estructural y funcional esencial de las uniones estrechas (Günzel y Yu, 2013), podría apuntar hacia una mejora de la regulación de la permeabilidad intestinal.
Sin embargo, el aumento concomitante de la expresión de las hormonas del estrés (CRHR1, HSD11β1) y de los genes proinflamatorios (IL6) y sus cofactores (IDO1) también podría reflejar el nivel de estrés que experimentan los lechones de bajo peso. Concretamente, se ha sugerido que los genes HSD11β1 y CRHR1 son mediadores inducidos por el estrés asociado al deterioro de la barrera intestinal y a la hipersecreción en lechones destetados precozmente (Meddings y Swain, 2000; Smith et al., 2010) y en lechones sometidos a un desafío con lipopolisacáridos (Zhu et al., 2016).
Por otro lado, el gen IDO1 codifica la enzima indolamina 2,3-dioxigenasa que cataliza la degradación del L-triptófano, pero también actúa como inmunorregulador cuya actividad parece aumentar en circunstancias proinflamatorias (Frumento et al., 2002).
Tomados estos hallazgos en su conjunto, el retraso del crecimiento postnatal no solo perjudica al rendimiento económico de las explotaciones, sino que también afecta negativamente a varias funciones fisiológicas al aumentar las respuestas proinflamatorias y de estrés.

Entre los cambios fisiopatológicos asociados a las citoquinas proinflamatorias y las moléculas de estrés cabe destacar la redistribución de nutrientes, como la energía y los aminoácidos, que inicialmente estarían destinados al crecimiento (Huntley et al., 2018).
Se podría especular que las provisiones de nutrientes e inmunidad durante el periodo de lactación no son suficientes para satisfacer las necesidades de los lechones poco desarrollados que deben competir con sus compañeros de camada por la provisión de leche.
De hecho, los lechones de bajo peso han demostrado tener un desarrollo postnatal y postdestete desfavorable (Bérard et al., 2008; Ji et al., 2017; Rodrigues et al., 2020), con órganos y canales con características deficientes (Rehfeldt y Kuhn, 2006; Bérard et al., 2008) en comparación con los lechones de mayor peso.
Nº 7 | Abril 2023 LECHONES Influencia del peso corporal sobre la expresión génica intestinal de los lechones 24
Artículo
El deterioro de las funciones fisiológicas de los lechones de bajo peso observado en el presente estudio apoya el argumento de que el suministro insatisfactorio de nutrientes al feto/neonato es un factor importante que influye en su rendimiento fisiológico postnatal (Larriestra et al., 2006; Farmer y Edwards, 2021).
En conclusión, los lechones de bajo peso al nacimiento y al final de la lactación parecen tener un desarrollo intestinal y una absorción de nutrientes deficientes, así como mayor tendencia a desarrollar respuestas proinflamatorias en comparación con sus compañeros de camada de peso medio.
Se necesitan más estudios para determinar las consecuencias a corto y largo plazo de la inmadurez intestinal de los lechones de bajo peso al nacimiento procedentes de camadas grandes, así como sobre su eficiencia alimentaria y su capacidad para hacer frente a los retos posteriores al destete.
of Animal Science, Volume 100, Issue 6, June 2022, skac161, https://doi.org/10.1093/jas/skac161 (CC BY 4.0).
BIBLIOGRAFÍA
van Barneveld, R. J., and R. J. E. Hewitt. 2016. Reducing variation in pork production systems through maternal and pre- and post-weaning nutrition strategies. Anim. Prod. Sci. 56:1248. doi:10.1071/AN15396.
Bérard, J., M. Kreuzer, and G. Bee. 2008. Effect of litter size and birth weight on growth, carcass and pork quality, and their relationship to postmortem proteolysis. J. Anim. Sci. 86:2357–2368. doi:10.2527/jas.2008-0893.
Burdge, G. C., M. A. Hanson, J. L. Slater-Jefferies, and K. A. Lillycrop. 2007. Epigenetic regulation of transcription: a mechanism for inducing variations in phenotype (fetal programming) by differences in nutrition during early life? Br. J. Nutr. 97:1036–1046. doi:10.1017/ S0007114507682920.
Dong, L., X. Zhong, H. Ahmad, W. Li, Y. Wang, L. Zhang, and T. Wang. 2014. Intrauterine growth restriction impairs small intestinal mucosal immunity in neonatal Piglets. J. Histochem. Cytochem. 62:510–518. doi:10.1369/0022155414532655.
Farmer, C., and S. A. Edwards. 2021. Review: improving the performance of neonatal piglets. Animal 100350. doi:10.1016/j.animal. 2021.100350.
Frumento, G., R. Rotondo, M. Tonetti, G. Damonte, U. Benatti, and G. B. Ferrara. 2002. Tryptophan-derived catabolites are responsible for inhibition of T and natural killer cell proliferation induced by indoleamine 2,3 dioxygenase. J. Exp. Med. 196:459–468. doi:10.1084/ jem.20020121.
Günzel, D., and A. S. L. Yu. 2013. Claudins and the modulation of tight junction permeability. Physiol. Rev. 93:525 569. doi:10.1152/physrev.00019.2012.
Huang, P., Y. Li, C. Xu, G. Melino, C. Shao, and Y. Shi. 2020. HSD11B1 is upregulated synergistically by IFNγ and TNFα and mediates TSG-6 expression in human UC-MSCs. Cell Death Discov. 6:24. doi:10.1038/s41420020-0262-7.
Ji, Y., Z. Wu, Z. Dai, X. Wang, J. Li, B. Wang, and G. Wu. 2017. Fetal and neonatal programming of postnatal growth and feed efficiency in swine. J. Anim. Sci. Biotechnol. 8:1–15. doi:10.1186/s40104- 017-0173-5.
Kraeling, R. R., and S. K. Webel. 2015. Current strategies for reproductive management of gilts and sows in North America. J. Anim. Sci. Biotechnol. 6:3. doi:10.1186/2049-1891-6-3.
Kwon, E. J., and Y. J. Kim. 2017. What is fetal programming?: a lifetime health is under the control of in utero health. Obstet. Gynecol. Sci. 60:506. doi:10.5468/ ogs.2017.60.6.506.
Larriestra, A. J., S. Wattanaphansak, E. J. Neumann, J. Bradford, R. B. Morrison, and J. Deen. 2006. Pig characteristics associated with mortality and light exit weight for the nursery phase. Can. Vet. J. 47(6):560–566. PMID: 16808228; PMCID: PMC1461411.
Li, T., S. Huang, L. Lei, S. Tao, Y. Xiong, G. Wu, J. Hu, X. Yuan, S. Zhao, B. Zuo, et al. 2021. Intrauterine growth restriction alters nutrient metabolism in the intestine of porcine offspring. J. Anim. Sci. Biotechnol. 12:15. doi:10.1186/s40104-020-00538-y.
Matheson, S. M., G. A. Walling, and S. A. Edwards. 2018. Genetic selection against intrauterine growth retardation in piglets: a problem at the piglet level with a solution at the sow level. Genet. Sel. Evol. 50:1–11. doi:10.1186/s12711-018-0417-7.
Meddings, J. B., and M. G. Swain. 2000. Environmental stress–induced gastrointestinal permeability is mediated by endogenous glucocorticoids in the rat. Gastroenterology 119:1019–1028. doi:10.1053/gast.2000.18152.
Michiels, J., M. De Vos, J. Missotten, A. Ovyn, S. De Smet, and C. Van Ginneken. 2013. Maturation of digestive function is retarded and plasma antioxidant capacity lowered in fully weaned low birth weight piglets. Br. J. Nutr. 109:65–75. doi:10.1017/S0007114512000670.

Nixon, M., D. J. Wake, D. E. Livingstone, R. H. Stimson, C. L. Esteves, J. R. Seckl, K. E. Chapman, R. Andrew, and B. R. Walker. 2012. Salicylate downregulates 11β-HSD1 expression in adipose tissue in obese mice and in humans, mediating insulin sensitization. Diabetes 61:790–796. doi:10.2337/db11-0931.
Okumura, R., and K. Takeda. 2017. Roles of intestinal epithelial cells in the maintenance of gut homeostasis. Exp. Mol. Med. 49:e338–e338. doi:10.1038/ emm.2017.20.
Peltoniemi, O., J. Yun, S. Björkman, and T. Han. 2021. Coping with large litters: the management of neonatal piglets and sow reproduction. J. Anim. Sci. Technol. 63:1–15. doi:10.5187/jast.2021.e3.
Rehfeldt, C., and G. Kuhn. 2006. Consequences of birth weight for postnatal growth performance and carcass quality in pigs as related to myogenesis. J. Anim. Sci. 84:E113–E123. doi:10.2527/2006.8413_supplE113x.
Rodrigues, L. A., M. O. Wellington, J. M. Sands, L. P. Weber, T. D. Olver, D. P. Ferguson, and D. A. Columbus. 2020. Characterization of a swine model of birth weight and neonatal nutrient restriction. Curr. Dev. Nutr. 4(8):nzaa116:1–8. doi:10.1093/cdn/nzaa116.
Smith, F., J. E. Clark, B. L. Overman, C. C. Tozel, J. H. Huang, J. E. F. Rivier, A. T. Blisklager, and A. J. Moeser. 2010. Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol. Liver Physiol. 298:G352–G363. doi:10.1152/ ajpgi.00081.2009.
Suttle, N. F. 2010. Mineral Nutrition of Livestock, 4th ed. Oxon, United Kingdom.
Trevisi, P., E. Corrent, S. Messori, S. Formica, D. Priori, and P. Bosi. 2012. Supplementary tryptophan downregulates the expression of genes induced by the gut microbiota in healthy weaned pigs susceptible to enterotoxigenic Escherichia coli F4. Livest. Sci. 147:96–103. doi:10.1016/j.livsci.2012.04.008.
Villagómez-Estrada, S., J. F. Pérez, S. van Kuijk, D. Melo-Durán, A. Forouzandeh, F. Gonzalez-Solè, M. D., ’ Angelo, F. J. Pérez-Cano, and D. Solà-Oriol. 2021. Strategies of inorganic and organic trace mineral supplementation in gestating hyperprolific sow diets: effects on the offspring performance and fetal programming. J. Anim. Sci. 99:1–14. doi:10.1093/jas/skab178.
Viott, R. C., T. A. Menezes, A. P. G. Mellagi, M. L. Bernardi, I. Wentz, and F. P. Bortolozzo. 2018. Performance of low birth-weight piglets upon protein-energy and/or colostrum supplementation. Arq. Bras. Med. Veterinária e Zootec. 70:1293–1300. doi:10.1590/1678-4162-9798.
Ward, S. A., R. N. Kirkwood, and K. J. Plush. 2020. Are larger litters a concern for piglet survival or an effectively manageable trait? Animals 10:309. doi:10.3390/ ani10020309.
Xu, S., D. Wang, P. Zhang, Y. Lin, Z. Fang, L. Che, and D. Wu. 2015. Oral administration of Lactococcus lactis-expressed recombinant porcine epidermal growth factor stimulates the development and promotes the health of small intestines in early-weaned piglets. J. Appl. Microbiol. 119:225–235. doi:10.1111/jam.12833.
traducido y adaptado de: Sandra Villagómez-Estrada, José F. Pérez, Diego Melo-Durán, Francesc Gonzalez-Solè, Matilde D’Angelo, Francisco .J Pérez-Cano, David Solà-Oriol, Body weight of newborn and suckling piglets affects their intestinal gene expression, Journal
25 GENÉTICA
SOUNDTALKS® IA AL SERVICIO DE LA SALUD RESPIRATORIA PORCINA
ENTREVISTA CON MAITE BASTERO, HEAD OF MARKETING LIVESTOCK DE BOEHRINGER INGELHEIM

ANIMAL HEALTH ESPAÑA
M M
Descarga el PDF
aite Bastero, Head of Marketing Livestock de Boehringer Ingelheim Animal Health España, analiza todos los detalles de SoundTalks®, un sistema que pone la inteligencia artificial al servicio de los veterinarios y productores de porcino.
Aportar valor a través de la innovación es uno de los valores fundacionales de Boehringer Ingelheim Animal Health. Un valor perfectamente alineado con la misión de trabajar cada día para mejorar la salud y el bienestar de los animales.
SoundTalks® es el reflejo perfecto de esta vocación. Se trata de una tecnología aplicada a la producción porcina que monitoriza de forma continua el estado de salud respiratoria de los cerdos para facilitar la predicción de enfermedades.
ENTREVISTA Nº 7 | Abril 2023 26 SoundTalks®, IA al servicio de la salud respiratoria porcina - Entrevista con Maite Bastero,
Boehringer Ingelheim Animal Health España
¿EN QUÉ CONSISTE SOUNDTALKS ® ?
SoundTalks® es una herramienta que analiza el sonido de la granja y que está diseñada para la vigilancia de la salud respiratoria en explotaciones de cerdos.
Está basada en Inteligencia Artificial y es capaz de predecir la aparición de un proceso respiratorio hasta con 5 días de antelación.
¿CUÁLES SON SUS PRESTACIONES?
El sistema de SoundTalks® se compone de unas campanas o monitores en cuyo interior hay 3 sensores: sonido, temperatura y humedad.
A través de los micrófonos, SoundTalks® es capaz de registrar y distinguir toses, estornudos y otros sonidos asociados a la respiración de los cerdos, enviar la información a la nube y, gracias a un algoritmo matemático, predecir la aparición de un problema respiratorio en los siguientes 5 días.
¿EN QUÉ PAÍSES ESTÁ IMPLEMENTADO SOUNDTALKS ® Y EN QUÉ PUNTO DE IMPLANTACIÓN NOS ENCONTRAMOS EN ESPAÑA?
SoundTalks® está implementado en cerca de una veintena de países como EEUU, China, Japón, Brasil y gran parte de los países de Europa.
En España empezamos a finales de 2021 con algún proyecto piloto y es ahora, en 2023, cuando estamos en pleno lanzamiento.
Además, próximamente estará disponible en Argentina, Australia y Canadá.
Cuando el sistema detecta la aparición de un brote respiratorio envía una alerta al móvil del usuario y queda registrada toda la información en la web, donde también hay un sistema de alertas que llega al profesional en forma de mensaje.
¿A QUIÉN VA DIRIGIDO SOUNDTALKS ® ?
Además de esto, es importante detallar que los monitores colgados en el techo de la granja funcionan con un código de luces como un semáforo.
Cuando todo es correcto, las luces están de color verde
Si el sistema detecta un posible brote, se pone de color amarillo (alerta).
Si el proceso respiratorio ya está instaurado aparece el color rojo.
SoundTalks® va dirigido a productores de porcino y a veterinarios que quieran tener un mejor control en sus granjas. Este sistema funciona 24/7, por lo que permite tener controlados a los animales en todo momento.
Gracias a esta herramienta podemos gestionar mejor nuestro tiempo y centrarnos en aquellos lotes que están en color amarillo o rojo.
SANIDAD 27
¿CUÁL ES LA APORTACIÓN DE ESTA TECNOLOGÍA A LA PREVENCIÓN DE ENFERMEDADES?
SoundTalks® no previene la aparición de una enfermedad, sino que predice su aparición (en este caso, el Complejo Respiratorio Porcino - CRP).
Al tener la información por adelantado, el productor puede tomar decisiones y aplicar determinadas medidas o protocolos de actuación que harán que el proceso dure menos tiempo.
En este sentido, está demostrado que hay una fuerte correlación entre días en verde y mejores resultados técnico-económicos.
¿QUÉ EFECTO TIENE EL USO DE SOUNDTALKS ®
SOBRE LA PRODUCTIVIDAD DE UNA EXPLOTACIÓN?
Las pérdidas de productividad se producen a medida que la enfermedad progresa en una granja. Cuanto mayor es la duración de los procesos patológicos, mayores son las pérdidas, ya que hay un mayor número de animales infectados, generalmente el CRP se agrava y, además, aparecen infecciones secundarias.
Con la implementación de SoundTalks®, lo que conseguimos es adelantarnos a la aparición de un posible brote respiratorio. Si somos capaces de actuar a tiempo, antes que las alarmas de la herramienta y aplicando el protocolo consensuado con el veterinario, entonces conseguiremos varias cosas:
Reducir el uso de antibióticos.
Reducir la duración del proceso clínico y, en definitiva, la mortalidad.
LA DETECCIÓN TEMPRANA
EFECTIVA PERMITE
MEJORAR LOS PARÁMETROS
PRODUCTIVOS, COMO LA
GANANCIA MEDIA DIARIA Y, POR SUPUESTO, EL BIENESTAR ANIMAL
¿QUÉ IMPORTANCIA TIENE LA PREVENCIÓN EN LA FILOSOFÍA Y VALORES DE BOEHRINGER INGELHEIM?
Uno de los mayores hitos para Boehringer Ingelheim Animal Health ha sido ser capaces de entender que teníamos que cambiar el paradigma y pasar de un modelo pensado en tratar enfermedades a otro centrado en la prevención.
Dicho esto, en salud porcina la prevención siempre ha sido uno de nuestros grandes objetivos y ha sido el eje central de nuestro lema “Prevention Works”
Ahora, hemos dado un paso más y ya no solo hablamos de prevención, si no de predicción.
La detección temprana de enfermedades es clave para abordar la salud porcina del futuro. Esto queda reflejado en nuestro nuevo lema: “Moving from prevention to prediction”.
Si hablamos de SoundTalks®, la detección temprana de una enfermedad respiratoria puede conducir a mejores protocolos preventivos y tratamientos más tempranos y precisos, lo que puede resultar en una mayor productividad y reducción de costes.
ENTREVISTA Nº 7 | Abril 2023 28 SoundTalks®, IA al servicio de la salud respiratoria
- Entrevista
Bastero,
Health
porcina
con Maite
Boehringer Ingelheim Animal
España
Escucha tu granja
Detecta la tos de tus cerdos de forma temprana y responde antes y desde cualquier lugar con SoundTalks®
+info
USO DE NUEVAS TECNOLOGÍAS
DIGITALES PARA LA EVALUACIÓN DEL BIENESTAR EN GRANJA Y MATADERO
T E
El Bienestar Animal es un aspecto que ha ganado importancia en la producción animal, tanto para la sociedad en general como para los ganaderos y sus intereses económicos. Se puede definir como “el estadodeunindividuoconrespectoasusintentos deadaptarsealmedioenelquevive”(Broom,1988).
Emma Fàbrega1, Marc Bagaria¹, Aranzazu Varvaró1, Joaquim Pallisera1, Xavier Averós2 e Inma Estevez2,3.
1Programa de Bienestar Animal, IRTA Monells, Girona



2Departamento de Producción
Animal, NEIKER Instituto Vasco de Investigación y Desarrollo Agrario

Alianza Vasca de Investigación y Tecnología (BRTA), Arkaute
3IKERBASQUE, Fundación Vasca para la Ciencia, Bilbao
Cuantificar este “estado” es complejo, por lo que resulta necesaria la evaluación de múltiples parámetros objetivos para poder obtener una imagen completa y fiable del bienestar de un individuo. Sin embargo, utilizar sistemas de evaluación con dichos criterios requiere demasiado tiempo, por lo que resulta crucial que la industria ganadera tenga acceso a protocolos de evaluación y a herramientas prácticas que faciliten esta tarea.



GANADERÍA DE PRECISIÓN Nº 7 | Abril 2023 Uso de nuevas tecnologías digitales para la evaluación del bienestar en granja y matadero 30
Descarga el PDF
En este artículo se presentan dos herramientas basadas en nuevas tecnologías con la finalidad de ser utilizadas por ganaderos y otros profesionales del sector (veterinarios, auditores, etc.).
1 2
La primera de ellas es una App para evaluar el bienestar de los cerdos a la llegada al matadero.
La segunda es una herramienta de formación y autoevaluación sobre los factores de riesgo de aparición de caudofagia en granja.
APP PARA LA EVALUACIÓN DEL BIENESTAR DE LOS CERDOS EN EL MATADERO
Los equipos de IRTA y NEIKER participaron en el desarrollo de los protocolos de evaluación de bienestar Welfare Quality® (WQ) y AWIN® (Marchewka et al., 2013). Estos protocolos incluyen: Medidas basadas en el animal que nos informan sobre su estado individual de bienestar. Medidas basadas en las instalaciones y el ambiente de cría que permiten identificar factores de riesgo que puedan tener un impacto negativo sobre el bienestar.

Estos protocolos han sido desarrollados tanto para granja como para matadero, siendo fácilmente aplicables a condiciones comerciales con mínimas molestias para los animales.
Sin embargo, pese a su éxito, aplicar estos protocolos requiere de un buen entrenamiento específico y experiencia para poder llevar a cabo las evaluaciones de manera adecuada, además de unos conocimientos mínimos para poder procesar los datos y entender e interpretar los resultados.
En el marco del proyecto TECHWEL , se ha trabajado con el objetivo principal de desarrollar herramientas digitales para la evaluación del Bienestar Animal, centrándonos en su utilidad y accesibilidad para el usuario final. Estas herramientas consisten en dos aplicaciones para dispositivos móviles, una para la evaluación en granjas de pollos de engorde y otra para el sacrificio de cerdos en matadero.
LA TRANSFORMACIÓN DE LOS PROTOCOLOS DE EVALUACIÓN EN APLICACIONES
DIGITALES
PRETENDE
SIMPLIFICAR LA APLICACIÓN PRÁCTICA DE LOS PROTOCOLOS Y HACERLOS ACCESIBLES Y FÁCILES DE IMPLEMENTAR
T
BIENESTAR ANIMAL 31
Las aplicaciones se han diseñado para ser intuitivas y guiar al usuario paso a paso durante la evaluación, simplificando así el procedimiento de evaluación. Las aplicaciones WellPig App y Broiler App : Facilitan la recopilación de datos de salud y bienestar de forma más eficiente.
Facilitan la detección de factores de riesgo para el Bienestar Animal de manera estandarizada.
Permiten la optimización de las prácticas de manejo.
Los usuarios potenciales de estas Apps incluyen a agencias certificadoras, veterinarios y técnicos que deseen una herramienta de apoyo en sus rutinas diarias de toma de datos, así como ganaderos para que puedan llevar a cabo sus propias autoevaluaciones.
Las autoevaluaciones permiten a los ganaderos determinar cuantitativamente el estado de sus animales de forma que puedan tomar decisiones de manejo de acuerdo con sus resultados, mejorando tanto el bienestar de los animales, en caso de detectar deficiencias, como su eficiencia productiva.
La obtención de resultados a tiempo real puede ayudar a los ganaderos a estar mejor preparados para superar un proceso de auditoría o comparar sus resultados con evaluaciones previas. Además, estas aplicaciones funcionan offline, lo que contribuye a un uso extensivo de protocolos de evaluación del Bienestar Animal sin limitaciones geográficas.

GANADERÍA DE PRECISIÓN Nº 7 | Abril 2023 Uso de nuevas tecnologías digitales para la evaluación del bienestar en granja y matadero 32
INDICADORES DE BIENESTAR
El primer paso en el desarrollo de WellPig App fue definir las variables a evaluar y el número de mataderos de cerdos a incluir en el estudio, trabajando en base al protocolo Welfare Quality® con algunas adaptaciones para potenciar su aplicabilidad en formato digital.
La App permite la evaluación del bienestar de los cerdos a su llegada al matadero en base a indicadores de bienestar que incluyen:

Información general del matadero.
Evaluación de las instalaciones.
Evaluación de los animales antes del sacrificio.
Evaluación de los animales después de su sacrificio.
EVALUACIÓN ANTES DEL SACRIFICIO
Antes del sacrificio, se evalúa: La descarga de los animales con parámetros como los resbalones o los intentos de retroceso (Figura 1)
Las condiciones de estabulación, como las vocalizaciones cuando son conducidos al aturdimiento.
El correcto aturdimiento.
EVALUACIÓN DESPUÉS DEL SACRIFICIO
Después del sacrificio se evalúa: Las canales. La formación de los trabajadores.
La formación de los transportistas.
FIGURA 1
Portada de la aplicación WellPig App y una de las pantallas para la evaluación de resbalones y caídas en el momento de la descarga.
BIENESTAR ANIMAL 33
Al final de la evaluación se realiza un checklist de cumplimiento de la legislación.
DISEÑO DE LA HERRAMIENTA DIGITAL
A continuación, se procedió al diseño y arquitectura de la herramienta digital (aplicación móvil), obteniendo una versión beta de la App.
Esta versión beta fue testada en matadero por personal técnico formado en la aplicación de los protocolos y en el uso de la app con el objetivo de optimizar su funcionamiento y detectar posibles errores.
Basándonos en los resultados del testaje y, trabajando de manera conjunta con los desarrolladores de software, se refinó la app para mejorar los resultados y la experiencia del usuario final. Este proceso se repitió hasta conseguir la versión final de la aplicación que en la actualidad permite llevar a cabo una evaluación de bienestar completa en matadero y presentar los resultados obtenidos de forma clara y visual.
LA APLICACIÓN ES FÁCIL DE USAR, PERO REQUIERE DE UN ENTRENAMIENTO BÁSICO EN LAS METODOLOGÍAS
ESPECÍFICAS UTILIZADAS
EN LOS PROTOCOLOS Y EN LOS CRITERIOS DE PUNTUACIÓN DE LOS INDICADORES DE BIENESTAR
El entrenamiento es esencial para conseguir una homogeneidad en las evaluaciones que sea independiente del evaluador, y obtener, por tanto, resultados fiables. Su uso puede mejorar el Bienestar Animal, pero también la eficiencia y sostenibilidad a largo plazo de los sistemas productivos.
Todo ello puede redundar en una mayor aceptación de los productos de origen animal por parte de los consumidores, y en una mayor responsabilidad social de los sistemas ganaderos modernos.
ESTE PROYECTO
HA DADO UN PASO
ADELANTE EN LA IMPLEMENTACIÓN
DEL USO DE NUEVAS
TECNOLOGÍAS EN EL CAMPO DE LA EVALUACIÓN DEL BIENESTAR ANIMAL
La aplicación Wellpig App, desarrollada a partir del protocolo Welfare Quality, estará disponible de manera gratuita en Google Play Store a partir de septiembre de 2023, permitiendo su uso libre y extensivo.

GANADERÍA DE PRECISIÓN Nº 7 | Abril 2023 34
“PÍLDORAS” DE CONOCIMIENTO Y EVALUACIÓN DE RIESGOS DE CAUDOFAGIA
La caudofagia es un problema de origen multifactorial que tiene consecuencias adversas importantes, tanto para el Bienestar Animal como por sus importantes repercusiones económicas.
El raboteo ha sido tradicionalmente la estrategia preventiva más utilizada para evitar las mordeduras de cola. Sin embargo, se trata de una práctica que no soluciona totalmente el problema y que, además, de acuerdo con la legislación europea, no está permitida de forma sistemático.
El último informe de Bienestar Animal en porcino de la EFSA (Agencia de Seguridad Alimentaria Europea, 2022), señala como uno de los grandes retos de futuro en bienestar porcino el abandono del raboteo.
En los sistemas de producción actuales el control de todos los factores de riesgo resulta muy complejo.
Impedir que se produzcan brotes de caudofagia importantes y que pueda ser viable plantearse el abandono progresivo del corte de colas depende, en gran medida, de una buena estrategia de detección precoz y prevención.
A nivel europeo, en países como Alemania, Francia, Irlanda o Reino Unido se está trabajando en sistemas de evaluación de los riesgos de caudofagia para su implementación a través de plataformas virtuales o aplicaciones móviles.
Algunos ejemplos se pueden encontrar en propuestas en desarrollo como:
WebHat de AHDB en Reino Unido.
SchwIP, desarrollado inicialmente en Alemania (Grüter et al., 2019) y en fase de prueba en proyectos como PigNoDock de Irlanda en Teagasc.
En países como Finlandia, donde el raboteo lleva más de 10 años totalmente prohibido, también se dispone de programas de evaluación de la incidencia de caudofagia y herramientas como SAPARO, una App gratuita para dar apoyo a los ganaderos en la prevención y control de la caudofagia mediante una monitorización continua de la situación en la granja y una ayuda para realizar los planes de acción para prevenir las mordeduras (Imagen 2).
PIGLOW App que se ha desarrollado para sistemas de producción ecológica y tiene una cobertura de evaluación de bienestar más amplia, no solo caudofagia.
A partir de estas iniciativas ya en desarrollo en otros países europeos, la Generalitat de Catalunya ha trabajado con el IRTA y una empresa de desarrollo de soluciones tecnológicas y formativas en un proyecto conocido como “Píldoras de Conocimiento”.
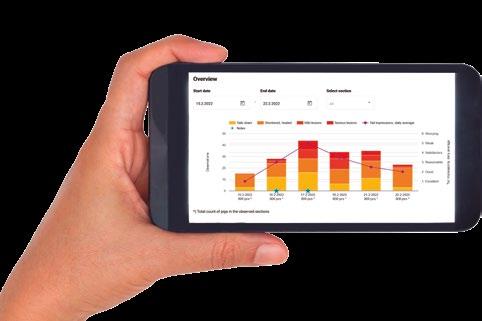 ACCEDER A WebHat
ACCEDER A PigNoDock
ACCEDER a SAPARO
ACCEDER a PIGLOW App
IMAGEN 2
Ejemplo de una gráfica de SAPARO de monitorización caudofagia en una explotación porcina
ACCEDER A WebHat
ACCEDER A PigNoDock
ACCEDER a SAPARO
ACCEDER a PIGLOW App
IMAGEN 2
Ejemplo de una gráfica de SAPARO de monitorización caudofagia en una explotación porcina
BIENESTAR ANIMAL 35
OBJETIVO 1 –APORTAR INFORMACIÓN
El objetivo de estas píldoras es, en primer lugar, generar módulos formativos muy interactivos con el fin de aportar información a los ganaderos sobre cómo prevenir la caudofagia.
En una primera fase, estos módulos formativos fueron elaborados por parte del IRTA y consensuados con el Departament d’Acció Climàtica, Alimentació i Agenda Rural y se trabajó con la empresa de desarrollo sobre qué métodos pedagógicos podían ser más adecuados.
De este modo, en las primeras versiones de la herramienta se incluyeron vídeos interactivos o motivacionales, protagonizados por ganaderos, actividades de drag-and-drop, preguntas de solución múltiple, etc.
En la Imagen 3 se muestran ejemplos de algunas de las pantallas con actividades disponibles.
Como herramienta formativa, estos materiales en la actualidad están disponibles en el portal Ruralcat , aunque ya no es posible realizar el curso con un tutor, a la espera que se consiga el objetivo 2, pudiendo utilizarse como herramienta de autoevaluación.
 ACCEDER a Ruralcat
IMAGEN 3
ACCEDER a Ruralcat
IMAGEN 3
GANADERÍA DE PRECISIÓN Nº 7 | Abril 2023 Uso de nuevas tecnologías digitales para la evaluación del bienestar en granja y matadero 36
Ejemplos de pantallas interactivas de las “Píldoras de Conocimiento” disponibles en Ruralcat (autores de los materiales didácticos: TCP group).
OBJETIVO 2 –HERRAMIENTA DE AUTOEVALUACIÓN
El segundo objetivo de estas píldoras formativas es que sirvan como herramienta de autoevaluación de los factores de riesgo.
La formación sobre los factores de riesgo de mordeduras se ha diseñado para que pueda realizarse recopilando datos de la propia granja que serán almacenados online y contrastados con datos de referencia para, de este modo, convertirse en la evaluación de riesgos de caudofagia que los ganaderos deben realizar según la legislación.
ESTA COMPARACIÓN CON DATOS ESPERADOS U ÓPTIMOS TIENE
COMO OBJETIVO PODER AYUDAR EN LA TOMA DE DECISIONES Y LA ELABORACIÓN DE PLANES DE ACCIÓN
PARA PREVENIR LA CAUDOFAGIA

La herramienta está en fase de mejora mediante un proceso conocido como living-lab, un proceso de co-creación en el que 10 ganaderos ya han participado utilizando las versiones beta para aportar sus ideas de mejora que se están incorporando para obtener la versión final optimizada.
Al igual que las Apps desarrolladas en el proyecto TECHWEL, estas versiones finales estarán disponibles en modo abierto, para trabajar online, en móviles u otros dispositivos digitales.
El objetivo de la herramienta de evaluación de caudofagia es que permanezca abierta durante un año con la finalidad de que los ganaderos puedan acceder para realizar su evaluación de riesgos y planes de acción. Para facilitar su uso, se podría seguir un vídeo tutorial con los diferentes pasos a seguir.
BIENESTAR ANIMAL 37
CONCLUSIONES
La monitorización del Bienestar
Animal puede resultar compleja para el ganadero porque requiere de una inversión de tiempo considerable si se incorporan todos los parámetros y factores a controlar necesarios para obtener una buena evaluación.
Las nuevas tecnologías digitales pueden ayudar a que el ganadero u otros profesionales realicen esta monitorización de una forma más óptima porque pueden ser sistemas que no sirvan únicamente para almacenar información, sino que, mediante modelos de contraste con datos de referencia, permitan aportar avisos y apoyar las decisiones sobre estrategias preventivas o de control de problemas como la caudofagia o las autoevaluaciones de bienestar en granja.
Agradecimientos
El estudio TECHWEL recibió financiación por parte del Ministerio de Ciencia e Innovación, Agencia Estatal de Investigación, cofinanciación Fondos FEDER (MINECO, RTI2018096034-R-C21), y las Píldoras de Conocimiento fueron financiadas por la Generalitat de Catalunya, Departament d’Acció Climàtica, Alimentació i Agenda Rural.



BIBLIOGRAFÍA

A. Grümpel, A., J. Krieter, J., Dippel., S. (2019) Reducing estimated tail biting risk in German weaner pigs using a management tool. The Veterinary Journal, 254, 105406 //doi.org/10.1016/j.tvjl.2019.105406

Broom, D. (1988). The scientific assessment of animal Welfare. Applied Animal Behaviour Science, 20, 5-19. //doi.org/10.1016/0168-1591(88)90122-0.
Marchewka, J., Watanabe, T. T. N., Ferrante, V., and Estevez, I. (2013). Welfare assessment in broiler farms: transect walks versus individual scoring. Poult. Sci. 92, 2588–2599. doi: 10.3382/ ps.2013-03229
Welfare Quality (2009). Welfare quality assessment protocol for pigs (sows, piglets an growing pigs). Welfare Quality consortium, Lelystad, Netherlands. http://www.welfarequalitynetwork.net/ media/1018/pig_protocol.pdf
GANADERÍA DE PRECISIÓN Nº 7 | Abril 2023 38 Uso de nuevas tecnologías digitales para la evaluación del bienestar en granja y matadero
DIAGNÓSTICO MOLECULAR DE PRRS: DE LA PCR A TIEMPO REAL A LA SECUENCIACIÓN MASIVA

EEl Síndrome Reproductivo y Respiratorio Porcino (PRRS) es una de las enfermedades más importantes en la industria porcina a nivel mundial ya que ocasiona elevadas pérdidas económicas1,2. Este impacto económico está asociado principalmente al aumento de la mortalidad en lechones, la necesidad del uso de vacunas y la implementación de medidas de bioseguridad3.
La enfermedad afecta a cerdas reproductoras, lechones y cerdos jóvenes en crecimiento. En cerdas se observan problemas reproductivos como abortos, repeticiones, mayor incidencia de mortinatos, fetos momificados y lechones nacidos débiles. Además, puede ocasionar disminución en el crecimiento y aumento de la tasa de mortalidad en lechones por problemas respiratorios y susceptibilidad a otros agentes patógenos 4 .
Sofía Lázaro Exopol S.L.
SANIDAD 39
Descarga el PDF
EL VIRUS PRRS
El agente etiológico de este síndrome, el virus PRRS (PRRSV), es un virus de genoma ARN de la familia Arteviridae. Genéticamente está clasificado en dos genotipos:
Genotipo I: presente principalmente en Europa.
Genotipo II: predominante en América del Norte y Asia.
Entre ambos genotipos existen grandes diferencias a nivel de genoma, ya que solo comparten entre un 50-70% de identidad nucleotídica.
Dentro de un mismo genotipo existe gran variabilidad entre cepas, pudiendo llegar a diferir en hasta el 15% en el genoma6,7
El tamaño del genoma del PRRSV es de unos 15.000 pares de bases, correspondiendo tres cuartas partes al ORF1, que codifica para proteínas no estructurales. El resto del genoma comprende el resto de ORF (ORF2a, ORF2b, ORF3, ORF4, ORF5, ORF6 y ORF7) que codifican para las proteínas estructurales del virus (Figura 1)8
FIGURA 1
Esquema del genoma ARN de PRRS indicando los Open Reading Frames (ORFs) y el tamaño de ORF5 y ORF2-7 (Adaptado de Kappes & Faaberg, 20155, creado con BioRender.com).

LA GRAN VARIABILIDAD GENÉTICA DEL PRRSV ES FRUTO DE SU ELEVADA TASA DE MUTACIÓN Y DE LOS FENÓMENOS DE RECOMBINACIÓN
En un animal infectado, lo habitual es que se encuentre un conjunto de cuasiespecies virales sobre las que actúa la presión selectiva derivada de la inmunidad del hospedador9,10. Todo ello ha conducido al extenso abanico de cepas circulantes PRRS en todo el mundo11
Nº 7 | Abril 2023 DIAGNÓSTICO Diagnóstico molecular del PRRS: de la PCR a tiempo real a la secuenciación masiva 40
DIAGNÓSTICO DE PRRS
El grave impacto productivo derivado del PRRS ha hecho que, desde su descubrimiento a finales de los años ochenta, el virus siga siendo objeto de numerosas investigaciones con el objetivo de entender los mecanismos que intervienen en su patogénesis y epidemiología, así como desarrollar estrategias para el control efectivo de la enfermedad.
Para esto último, las medidas de bioseguridad, el manejo, la inmunización y el diagnóstico temprano son aspectos claves12,13
Existen diferentes herramientas de diagnóstico disponibles actualmente. A continuación, repasaremos las principales técnicas empleadas actualmente, incidiendo en su utilidad práctica.
TÉCNICAS INMUNOLÓGICAS Y CULTIVO VIRAL
El uso de técnicas serológicas es común para la detección de anticuerpos frente al virus, aunque no permiten diferenciar anticuerpos de infecciones activas de los producidos como respuesta a la vacunación.
Además, los resultados negativos no pueden descartar una infección, ya que en procesos agudos puede no haberse producido la seroconversión en el momento del análisis y la respuesta inmunitaria puede ser muy variable.
Por otro lado, a pesar de la complejidad y el largo tiempo de análisis, el aislamiento y cultivo viral también se han empleado como método de diagnóstico. Además, técnicas como la inmunohistoquímica y la inmunofluorescencia permiten detectar el antígeno viral pero su sensibilidad es limitada14
TÉCNICAS MOLECULARES
Las técnicas moleculares basadas en la detección del genoma viral, como la reacción en cadena de la polimerasa (PCR), confieren gran sensibilidad y especificidad15
Requieren de un menor tiempo de análisis, lo que resulta clave en ciertas aplicaciones como descartar la presencia del virus en dosis seminales de forma previa a la inseminación6
LA PCR ES LA HERRAMIENTA MÁS UTILIZADA
EN LA ACTUALIDAD Y PERMITE ANALIZAR
VARIOS TIPOS DE MUESTRAS DIFERENTES
SANIDAD 41
La elección del tipo de muestra dependerá del objetivo del análisis:
Confirmación de una sospecha clínica
Fetos o placentas
Suero de hembras en caso de abortos
Sangre de lechones
débiles
Pulmones
Lavados o raspados broncoalveolares de animales con sintomatología
Monitorización del virus para conocer el estado epidemiológico de la explotación
Fluidos orales
Lenguas
Semen
Fluidos del procesado de testículos o de colas*
*El uso de estas últimas cada vez está más restringido por la legislación vigente17-20
La PCR a tiempo real o cuantitativa (qPCR) es actualmente la técnica más extendida entre los laboratorios veterinarios para realizar el diagnóstico y monitorización de patógenos, incluido el PRRSV.
Aunque la PCR convencional con retrotranscripción o transcripción inversa (RT-PCR) ya se utilizaba para el diagnóstico de PRRSV21, la qPCR, llevando a cabo el paso previo necesario de retrotranscripción del genoma ARN del virus (RT-qPCR), ha terminado por reemplazarla, ya que presenta importantes ventajas:
Reducción del tiempo de análisis y de la probabilidad de obtener falsos positivos (la PCR convencional requiere de un análisis postPCR del fragmento amplificado de ADN, lo que aumenta el riesgo de contaminación de otras muestras).
Aumento de la sensibilidad y especificidad, y obtención de información cuantitativa21
Al igual que la PCR convencional, la qPCR consiste en la amplificación de un fragmento del genoma utilizando un par de oligonucleótidos, llamados cebadores, que son complementarios a dicha región. En la qPCR, este fragmento se detecta por fluorescencia tras cada ciclo (en “tiempo real”).
Para ello, se pueden emplear agentes que se intercalan en el ADN de doble cadena y emiten fluorescencia o mediante un tercer oligonucleótido (sonda) que está marcado con una molécula fluorescente que emite luz cuando la secuencia diana está presente (Figura 2)22
PCR A TIEMPO REAL PCR A TIEMPO REAL Nº 7 | Abril 2023 DIAGNÓSTICO Diagnóstico molecular del PRRS: de la PCR a tiempo real a la secuenciación masiva 42
1Retrotranscripción
Síntesis del ADN complementario por par te de la retrotranscriptasa utilizando el cebador reverso
ADN complementario
Cebador reverso
2 Desnaturalización
Aumento de la temperatura a 95ºC para separar las hebras de ADN
Fragmento de ARN diana
3 Unión de cebadores y amplificación
Disminución de la temperatura a 60ºC, unión de los cebadores a las regiones complementarias
La ADN polimerasa sintetiza la hebra complementaria
La ADN polimerasa tiene la capacidad de degradar moléculas de ADN que encuentra a su paso lo que lleva a la rotura de la sonda Esto provoca una señal fluorescente que es captada por el equipo
Las copias del fragmento diana se duplican
4 Repetición de ciclos (40x)
La repetición sucesiva de los pasos 2 y 3 lleva al aumento exponencial del producto amplificado y, en consecuencia, de moléculas que emiten fluorescencia
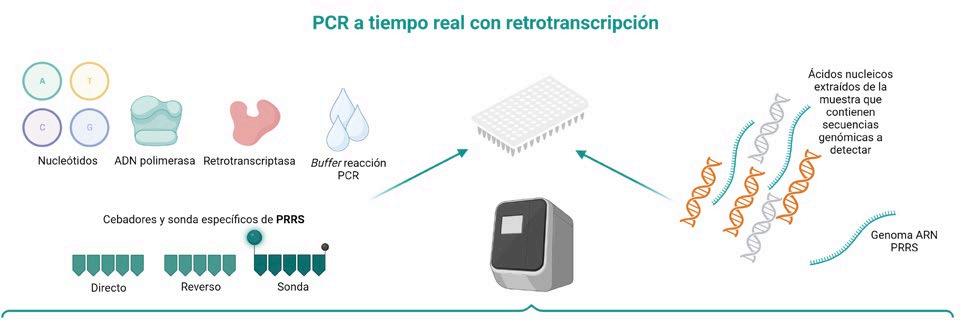
Aumento de fluorescencia al amplificarse el fragmento diana
Cebador directo
Sonda
FIGURA 2
Cebador reverso
Detección de fluorescencia
Esquema de la reacción de PCR a tiempo real con retrotranscripción para detección de PRRS utilizando sondas marcadas con moléculas fluorescentes (creado con BioRender.com).
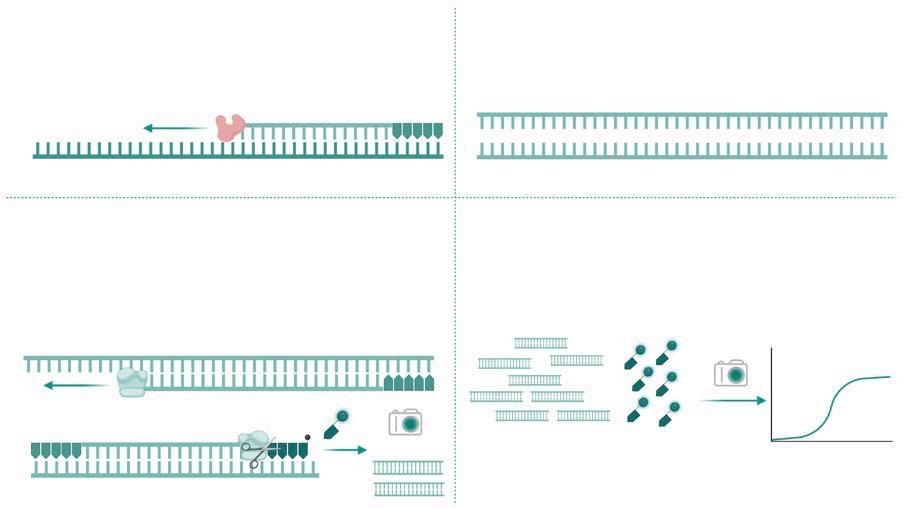
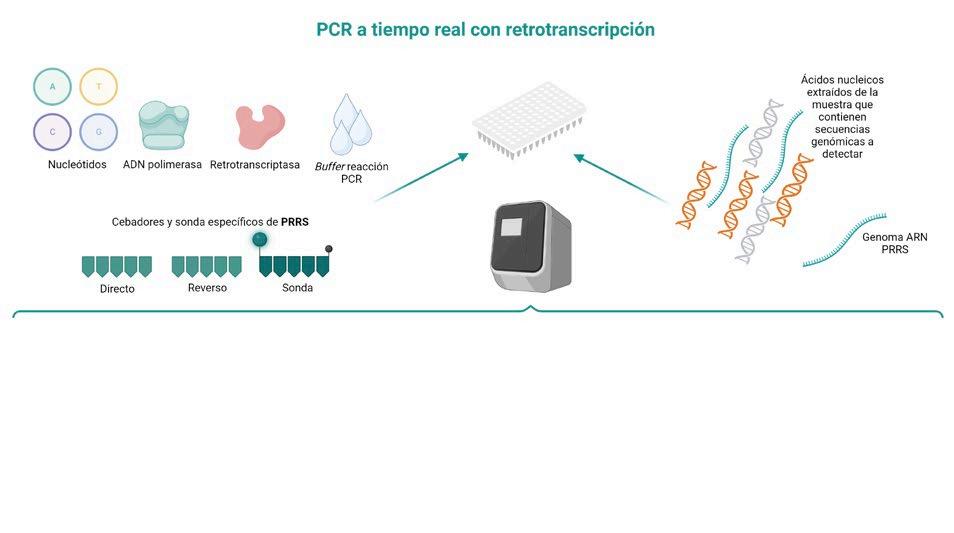
SANIDAD 43
La gran diferencia a nivel genómico entre los genotipos I y II posibilita su detección específica en una única reacción de RT-qPCR utilizando diferentes parejas de cebadores y sondas marcadas con moléculas fluorescentes distintas. Del mismo modo puede incorporarse la detección adicional de controles internos

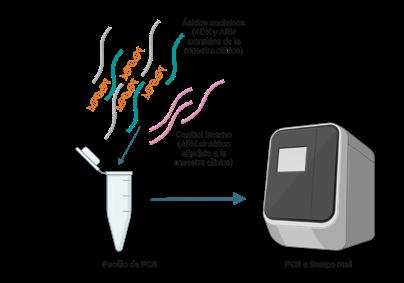
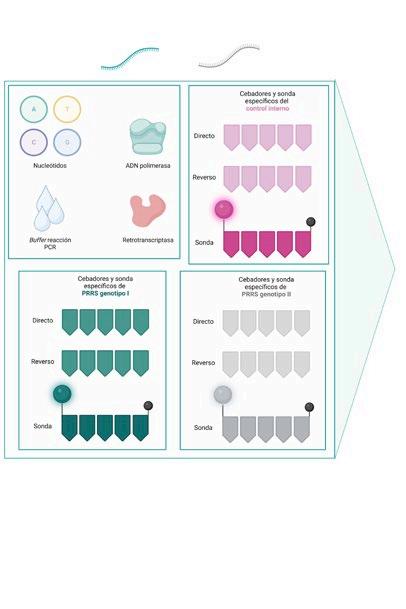
El uso de estos controles consiste en la adición de un fragmento de ARN o ADN a la muestra clínica de forma previa a la extracción, y que luego es detectado en la qPCR. Permite verificar el proceso de extracción, la reacción de amplificación y la ausencia de inhibidores, evitando así resultados falso-negativos (Figura 3)23
Dentro de cada genotipo, el diseño de los cebadores y sondas está dirigido a regiones genómicas más conservadas entre las distintas cepas, como el ORF723. Sin embargo, el diagnóstico por RT-qPCR supone un reto constante dada la elevada tasa de mutación del virus PRRS. Se requiere un trabajo continuo de revisión y actualización de estos ensayos para asegurar la detección del mayor número de cepas circulantes.
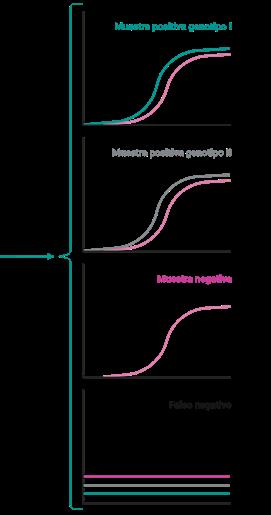 PRRS genotipo I PRRS genotipo II
FIGURA 3
Esquema de los componentes de una reacción multiplex para la detección simultánea y diferenciación en un único pocillo de los genotipos I, II y un control interno, así como los posibles resultados que se pueden obtener (creado con BioRender.com).
PRRS genotipo I PRRS genotipo II
FIGURA 3
Esquema de los componentes de una reacción multiplex para la detección simultánea y diferenciación en un único pocillo de los genotipos I, II y un control interno, así como los posibles resultados que se pueden obtener (creado con BioRender.com).
Directo
Cebadores y sonda específicos de PRRS genotipo
Cebadores y sonda específicos de PRRS genotipo II
Directo Directo Nucleótidos ADN polimerasa Retrotranscriptasa Buffer reacción PCR Reverso Reverso Reverso Sonda Sonda Sonda Nº 7 | Abril 2023 DIAGNÓSTICO Diagnóstico molecular del PRRS: de la PCR a tiempo real a la secuenciación masiva 44
Cebadores y sonda específicos de control interno
La capacidad de esta técnica para identificar regiones genómicas concretas ha permitido el diseño de ensayos específicos para detectar cepas de alta patogenicidad o virulencia.
Por ejemplo, se han diseñado ensayos de RT-qPCR para detectar algunas cepas de este tipo descritas en China y caracterizadas por carecer de determinados segmentos (lo que se conoce como deleciones) en el ORF1a23,24.
Considerando que las estrategias de prevención se basan en gran parte en la vacunación con cepas vivas atenuadas25 y que los kits comerciales para la detección de PRRSV no diferencian entre cepas vacunales y cepas campo, es importante conocer si un resultado qPCR positivo se debe a la presencia de una cepa vacunal para poder tomar las medidas de manejo apropiadas.
En este sentido, hay trabajos en los que se han desarrollado ensayos de qPCR para la detección de cepas vacunales26 y ya existen kits comerciales para este fin.
SECUENCIACIÓN DE SANGER SECUENCIACIÓN DE SANGER
La secuenciación de Sanger es una metodología que consiste en la determinación de los nucleótidos que conforman una región de ADN Permite obtener secuencias de hasta 900-1000 pares de bases, lo que resulta muy útil para secuenciar genes o secuencias cortas.
La secuenciación de Sanger del virus PRRS tiene aplicación directa en estudios epidemiológicos y en la mejora y actualización continua de los kits de qPCR de diagnóstico para no perder sensibilidad frente a nuevas cepas emergentes.
ORF5
El ORF5, de una longitud de 606 bp, contiene regiones inmunogénicas y regiones hipervariables que posibilitan una discriminación entre las diferentes cepas27
Para obtener su secuencia, se realiza una RT-PCR utilizando parejas de cebadores que se unen a las zonas flanqueantes del ORF, amplificando esa región y consiguiendo un número elevado de copias para el posterior paso de secuenciación. No obstante, no todas las muestras positivas a PRRS por PCR a tiempo real son susceptibles de ser secuenciadas, ya que muestras con títulos bajos (Cq superiores a 30) disminuyen las probabilidades de éxito en la obtención de una secuencia de buena calidad.
Una vez obtenida la secuencia final, es posible compararla con las secuencias ORF5 de las cepas vacunales o con otras cepas secuenciadas previamente en la misma explotación o pirámide productiva (Figura 4A y B).
LA
SECUENCIACIÓN DE SANGER DEL VIRUS PRRS, EN CONCRETO DEL ORF5, ES UN ANÁLISIS DE RUTINA EN LA MAYORÍA DE LOS
LABORATORIOS VETERINARIOS
Si bien, se trata de un valor arbitrario, está generalmente aceptado un cut-off de 97% de similitud en la secuencia de nucleótidos para el ORF5 para determinar que con alta probabilidad se trate de la misma cepa. No obstante, para llegar a esta conclusión, es importante considerar:
La ubicación geográfica de estas dos cepas.
El periodo de tiempo en el que se han detectado.
SANIDAD 45
¿Qué nos dicen los resultados de la secuenciación de Sanger sobre la bioseguridad de la granja?
Problema de bioseguridad interna
Si se determina que se trata de la misma cepa que la detectada y secuenciada en un momento anterior, se sospechará de un problema de bioseguridad interna.
Problema de bioseguridad externa
Cuando no exista una elevada identidad nucleotídica con cepas anteriores, podrá atribuirse a un fallo en los sistemas de bioseguridad externa.
De esta forma se podrán tomar las acciones correctivas más apropiadas para disminuir las probabilidades de transmisión del virus.
ANÁLISIS FILOGENÉTICOS
Si la secuenciación se realiza de forma rutinaria y se dispone de secuencias a lo largo del tiempo y de una región geográfica, el análisis de las relaciones filogenéticas entre ellas da una imagen muy útil de la situación epidemiológica del virus en un área y su progresión y transmisión en un periodo de tiempo determinado (Figura 4C).
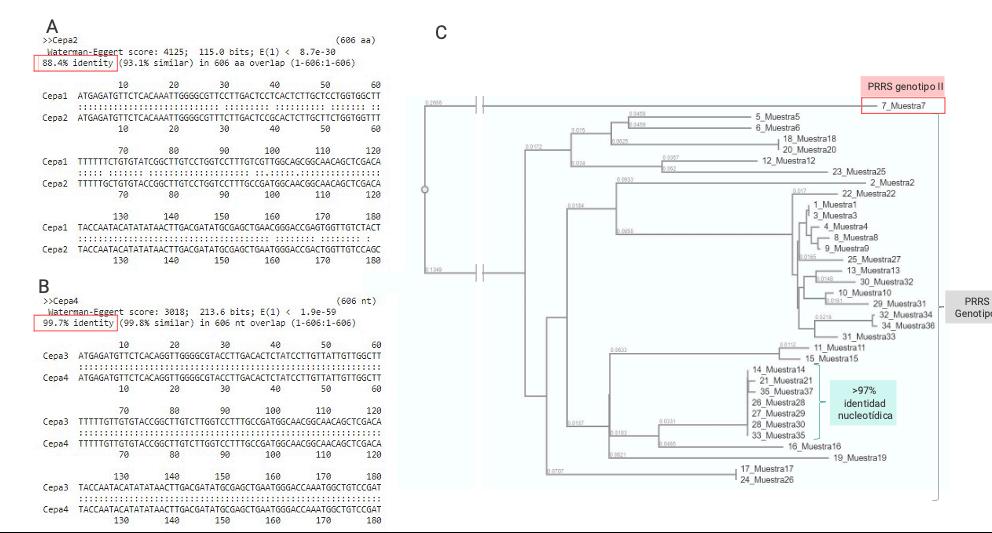
PRRS genotipo II
7_Muestra7
Análisis de secuencias Sanger. Visualización de un alineamiento de la parte inicial del ORF5 entre dos secuencias con una similitud inferior a 97% (A) y superior al 97% (B). (C) Árbol filogenético que incluye secuencias ORF5 de 35 muestras diferentes, una de ellas del genotipo II, que queda clasificada fuera del resto (genotipo I). Se observa un clúster de muestras donde la comparación dos a dos entre ellas arroja un resultado superior a 97%.
>97% identidad nucleotídica
PRRS genotipo I
FIGURA 4
Nº 7 | Abril 2023 DIAGNÓSTICO Diagnóstico molecular del PRRS: de la PCR a tiempo real a la secuenciación masiva 46
Es cierto que la secuenciación de ORF5 proporciona una valiosa información epidemiológica y tiene aplicación directa en campo. Sin embargo, no se puede pasar por alto que esta región representa un 5% del genoma de PRRS por lo que no permite captar toda la variabilidad genética de una cepa (Figura 1)7 .
En este punto, cabe destacar los fenómenos de recombinación que se han descrito para ambos genotipos de PRRS y que pueden ocurrir a lo largo de diversos puntos en todo el genoma, incluso dentro de un mismo ORF.
Estos eventos de recombinación generan cepas quimera que contienen secuencias de diferentes cepas parentales Ocurren cuando dos o más cepas de PRRSV infectan una misma célula de forma simultánea5. Por ejemplo, se han descrito cepas de campo que contienen el ORF5 procedente de cepas vacunales vivas atenuadas, por lo que se podrían clasificar cepas como vacunales cuando en realidad no lo son28
Además, ante una infección por una cepa recombinante se puede realizar una asunción errónea de una recirculación si esta comparte una alta similitud en ORF5 con otras cepas detectadas previamente.
LAS DISCREPANCIAS ENTRE EL DIAGNÓSTICO Y LOS SIGNOS CLÍNICOS
Comúnmente denominada NGS (del inglés Next Generation Sequencing), la secuenciación masiva permite la obtención miles de secuencias cortas, unos 200300 pares de bases, de forma paralela.
En estos casos, mediante secuenciación Sanger se pueden amplificar varios fragmentos del genoma que cubran una región más amplia (ORF2-5 u ORF2-7) o simplemente de otro ORF no adyacente al ORF5, por ejemplo, el ORF1a u ORF2, permitiendo identificar este tipo de cepas30.
Finalmente, es importante conocer las limitaciones de la secuenciación Sanger:
El resultado de esta técnica es una única secuencia consenso, lo que significa que, con alta probabilidad, se obtiene la variante mayoritaria en la muestra e impide la observación del resto de mutantes presentes o, dentro de un mismo individuo, de las cuasiespecies, dificultando el estudio de la evolución del virus.
En muestras que contengan dos cepas diferentes puede no llegar a obtenerse una secuencia consenso final.
Estos pequeños fragmentos se someten a un complejo análisis con herramientas bioinformáticas para armar un puzle llegando a obtener secuencias más largas o genomas completos en poco tiempo.
En comparación a la secuenciación de Sanger:
El proceso es menos laborioso. El coste por nucleótido secuenciado es menor.
No es necesario un conocimiento previo del genoma del agente.
Recientemente, se han desarrollado nuevas tecnologías que ofrecen secuenciación de fragmentos mucho más largos (>10.000 pares de bases).
En su aplicación al estudio del PRRSV, la secuenciación masiva ha aumentado el número de genomas completos disponibles. Gracias a ello, se ha determinado que, aparte del ORF5, existen otras regiones con más variabilidad como el principio del ORF1a31
OBSERVADOS
CEPA RECOMBINANTE
MASIVA
SANIDAD 47
PUEDEN SER UNA PISTA PARA SOSPECHAR DE UNA
29 SECUENCIACIÓN
SECUENCIACIÓN MASIVA
CONOCER LA TOTALIDAD DEL GENOMA IMPLICA
TENER UNA CEPA COMPLETAMENTE CARACTERIZADA Y ESTABLECER DE FORMA MUCHO MÁS PRECISA LAS
RELACIONES FILOGENÉTICAS ENTRE CEPAS
Además, impide una malinterpretación epidemiológica debido a los eventos de recombinación mencionados anteriormente y se pueden identificar marcadores epidemiológicos y/o detectar nuevas cepas emergentes24
Con la secuenciación masiva no se obtiene una única secuencia como en la secuenciación de Sanger, sino que todo el material genético en una muestra, donde pueden encontrarse distintas variantes o mutantes de PRRSV, se secuencia (Figura 5A)
Por ello, si se realizara de forma rutinaria, se podría observar cómo se comporta o cómo varía la presencia de posibles variantes en una explotación en respuesta a condiciones concretas (por ejemplo, tras una campaña de vacunación)32. Por supuesto, también permitiría identificar muestras con una coinfección por dos cepas diferentes.
El ADN de la muestra se fragmenta en trozos pequeños
Cada fragmento se ampli ca y se secuencia cientos de veces
Mediante herramientas bioinformáticas las secuencias obtenidas se ensamblan consiguiendo secuencias de mayor tamaño o incluso el genoma completo
FIGURA 5
El resultado nal son miles de secuencias diferentes y se pueden observar las distintas variantes, cuasiespecies o cepas Ácido nuc e co de a muestra
(A) Esquema del proceso de secuenciación masiva de fragmentos cortos (200-300 bp) y obtención de diferentes secuencias consenso dentro de una misma muestra. (B) Esquema del sistema de secuenciación por nanoporos donde la molécula de ADN o ARN atraviesa un sistema que detecta señales de corriente eléctrica, las cuales varían entre los diferentes nucleótidos.
A
-8 kb 0 kb Genoma ARN PRRS Tiempo Secuencia en t empo real S e ñ a l A G C T C
B
A A G G T Nº 7 | Abril 2023 DIAGNÓSTICO Diagnóstico molecular del PRRS: de la PCR a tiempo real a la secuenciación masiva 48
A nivel de investigación, con la secuenciación masiva se puede ahondar, entre otros, en:
La diversidad genética y la evolución del virus.
Las variaciones de las cepas circulantes a lo largo del tiempo.
Los mecanismos de transmisión y evasión de la respuesta inmunitaria
El conjunto de cuasiespecies virales en un individuo y su distribución en los diferentes tejidos.
Todo este conocimiento constituye la base para continuar mejorando el diagnóstico, las estrategias de control de la enfermedad y el diseño de nuevas vacunas que confieran mayor protección frente a las cepas circulantes32,33
El coste de la secuenciación masiva y el tiempo y dificultad de análisis son todavía una limitación en su uso como herramienta diagnóstica para el veterinario de campo. Además, la carga de PRRSV y el tipo de muestra pueden afectar al éxito de la secuenciación34
La variabilidad observada en los datos de secuenciación masiva es únicamente a nivel cualitativo por lo que no es posible obtener información cuantitativa, especialmente de aquellas variantes en menor proporción, que incluso podrían llegar a no detectarse32.
OTRAS HERRAMIENTAS PARA EL DIAGNÓSTICO MOLECULAR DE PRRS
OTRAS HERRAMIENTAS PARA EL DIAGNÓSTICO MOLECULAR DE PRRS

La rapidez en el diagnóstico para aplicar medidas correctivas en el menor tiempo posible es clave ante un brote de una enfermedad y el PRRS no es una excepción.
Existen técnicas en desarrollo para hacer diagnósticos in-situ sin la necesidad de equipos especializados. Entre otros sistemas, la PCR isoterma y la amplificación por recombinasa se han descrito como posibles herramientas para detectar el virus PRRSV con buena sensibilidad y especificidad.
Son técnicas similares a una PCR, pero se realizan a temperatura constante, pudiendo observarse el resultado de forma visual -por ejemplo, con un cambio de color en el tubo de reacción o una tira de flujo lateral (similar a un test rápido)- y no requieren de equipamiento complejo ni personal altamente cualificado.
No obstante, para poder explotar su potencial todavía se debe optimizar su sensibilidad y su robustez para que su resultado llegue a ser comparable a la qPCR o poder usar muestras clínicas directas sin extracción previa de ácidos nucleicos35,36
SANIDAD 49
Las últimas tecnologías de secuenciación desarrolladas (tercera generación) han conseguido ampliar la longitud de las secuencias obtenidas, así como poder secuenciar directamente moléculas ARN.
Una de estas tecnologías se basa en el uso de nanoporos (tecnología MiniON Oxford Nanopore) y se realiza con un pequeño dispositivo portable conectado a un ordenador (Figura 5B).
En estudios de PRRSV con este equipo se se ha demostrado que es posible secuenciar directamente la molécula ARN de su genoma en pocas horas y, aplicando un análisis bioinformático más sencillo, pueden obtenerse lecturas de secuencias más largas. Con este sistema también fue posible identificar coinfección de dos cepas en la misma muestra37
Cada día, en los laboratorios veterinarios se realizan cientos de análisis moleculares de muchos patógenos, generando multitud de informes individuales. El análisis de la información diagnóstica desde una perspectiva global de una enfermedad como el PRRSV, incluso en combinación con otras enfermedades, proporciona información muy valiosa
Con el fin de exprimir toda esta información, existen actualmente en algunos países bases de datos que agregan resultados de laboratorios privados junto con información pública para obtener información en nuevos patógenos emergentes y en su diseminación38
CONCLUSIONES
La RT-qPCR es una herramienta indispensable para el diagnóstico de PRRS de forma sensible y rápida ante un brote o como forma de monitorización. Sin embargo, debe ir de la mano de la secuenciación para proporcionar la información genética de la cepa detectada, indispensable para la toma de decisiones a nivel de granja.
ES IMPORTANTE QUE LA INTERPRETACIÓN DEL RESULTADO DE SECUENCIACIÓN SE REALICE JUNTO
CON LA OBSERVACIÓN CLÍNICA Y LA SITUACIÓN
EPIDEMIOLÓGICA DE LA EXPLOTACIÓN O LA REGIÓN
El seguimiento epidemiológico basado en la secuenciación de ORF5 resulta de gran utilidad, pero la secuenciación de otras regiones más amplias u ORFs no contiguos puede ampliar la información obtenida y confirmar presencia de cepas recombinantes.
La secuenciación masiva podrá en el futuro reemplazar a la secuenciación de ORFs individuales, brindando mucha más información y proporcionando una caracterización genética completa.
Asimismo, el desarrollo y la optimización de las nuevas tecnologías surgidas podrán ofrecer un diagnóstico más rápido y a pie de granja, algunas de ellas aportando información genética en tiempo real.
Todo ello en conjunto y recopilando de forma adecuada la información en las bases de datos permitirá abrir nuevas vías para seguir avanzando en el control y conocimiento del PRRS.
HAY QUE DESTACAR LA IMPORTANCIA DE REGISTRAR, ANALIZAR Y USAR TODA LA INFORMACIÓN QUE SE RECOPILA DE FORMA GLOBAL
Nº 7 | Abril 2023 DIAGNÓSTICO Diagnóstico molecular del PRRS: de la PCR a tiempo real a la secuenciación masiva 50
BIBLIOGRAFÍA
1. Neumann, E. J. et al. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J Am Vet Med Assoc (2005) doi:10.2460/javma.2005.227.385.
2. Holtkamp, D. J. et al. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. Journal of Swine Health and Production Preprint at (2013).
3. Nathues, H. et al. Cost of porcine reproductive and respiratory syndrome virus at individual farm level – An economic disease model. Prev Vet Med (2017) doi:10.1016/j.prevetmed.2017.04.006.
4. Albina, E. [Porcine reproductive and respiratory syndrome: ten years of experience (1986-1996) with this undesirable viral infection]. Vet Res (1997).
5. Kappes, M. A. & Faaberg, K. S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology Preprint at https://doi. org/10.1016/j.virol.2015.02.012 (2015).
6. Forsberg, R. Divergence time of porcine reproductive and respiratory syndrome virus subtypes. Mol Biol Evol (2005) doi:10.1093/molbev/msi208.
7. Murtaugh, M. P., Stadejek, T., Abrahante, J. E., Lam, T. T. Y. & Leung, F. C. C. The ever-expanding diversity of porcine reproductive and respiratory syndrome virus. Virus Research Preprint at https://doi.org/10.1016/j. virusres.2010.08.015 (2010).
8. Dokland, T. The structural biology of PRRSV. Virus Research Preprint at https://doi.org/10.1016/j. virusres.2010.07.029 (2010).
9. Domingo, E. & Schuster, P. What is a quasispecies? Historical origins and current scope. Curr Top Microbiol Immunol (2016) doi:10.1007/82_2015_453.
10. Lu, Z. H. et al. Quasispecies evolution of the prototypical genotype 1 porcine reproductive and respiratory syndrome virus early during in vivo infection is rapid and tissue specific. Arch Virol (2017) doi:10.1007/ s00705-017-3342-0.
11.Shi, M. et al. Molecular epidemiology of PRRSV: A phylogenetic perspective. Virus Research Preprint at https://doi.org/10.1016/j.virusres.2010.08.014 (2010).
12. Pileri, E. & Mateu, E. Review on the transmission porcine reproductive and respiratory syndrome virus between pigs and farms and impact on vaccination. Veterinary Research Preprint at https://doi.org/10.1186/ s13567-016-0391-4 (2016).
13. Drew, T. W. The emergence and evolution of swine viral diseases: To what extent have husbandry systems and global trade contributed to their distribution and diversity? OIE Revue Scientifique et Technique (2011) doi:10.20506/rst.30.1.2020.
14. Yaeger, M. J. The diagnostic sensitivity of immunohistochemistry for the detection of porcine reproductive and respiratory syndrome virus in the lung of vaccinated and unvaccinated swine. Journal of Veterinary Diagnostic Investigation (2002) doi:10.1177/104063870201400104.
15. Belák, S. Molecular diagnosis of viral diseases, present trends and future aspects. A view from the OIE Collaborating Centre for the Application of Polymerase Chain Reaction Methods for Diagnosis of Viral Diseases in Veterinary Medicine. Vaccine (2007) doi:10.1016/j. vaccine.2006.11.068.
16. Christopher-Hennings, J. et al. Detection of porcine reproductive and respiratory syndrome virus in boar semen by PCR. J Clin Microbiol (1995) doi:10.1128/ jcm.33.7.1730-1734.1995.
17. Kittawornrat, A. et al. Porcine reproductive and respiratory syndrome virus (PRRSV) in serum and oral fluid samples from individual boars: Will oral fluid replace serum for PRRSV surveillance? Virus Research Preprint at https://doi.org/10.1016/j.virusres.2010.07.025 (2010).
18. Rotolo, M. L. et al. Sampling guidelines for oral fluidbased surveys of group-housed animals. Vet Microbiol (2017) doi:10.1016/j.vetmic.2017.02.004.
19. Lopez, W. A., Angulo, J., Zimmerman, J. J. & Linhares, D. C. L. Porcine reproductive and respiratory syndrome monitoring in breeding herds using processing fluids. Journal of Swine Health and Production Preprint at (2018).
20. Baliellas, J., Novell, E., Enric-Tarancón, V., Vilalta, C. & Fraile, L. Porcine reproductive and respiratory syndrome surveillance in breeding herds and nurseries using tongue tips from dead animals. Vet Sci (2021) doi:10.3390/vetsci8110259.
21. Kleiboeker, S. B., Lehman, J. R. & Fangman, T. J. Concurrent use of reverse transcription-polymerase chain reaction testing of oropharyngeal scrapings and paired serological testing for detection of porcine reproductive and respiratory syndrome virus infection in sows. Journal of Swine Health and Production (2002).
22.Lurchachaiwong, W. et al. Rapid detection and strain identification of porcine reproductive and respiratory syndrome virus (PRRSV) by real-time RT-PCR. Lett Appl Microbiol (2008) doi:10.1111/j.1472-765X.2007.02259.x.
23.Wernike, K. et al. Detection and typing of highly pathogenic porcine reproductive and respiratory syndrome virus by multiplex real-time RT-PCR. PLoS One (2012) doi:10.1371/journal.pone.0038251.
24. Liu, Y. et al. Emergence of Different Recombinant Porcine Reproductive and Respiratory Syndrome Viruses, China. Sci Rep (2018) doi:10.1038/s41598-018-22494-4.
25. Linhares, D. C. L. et al. Effect of modified-live porcine reproductive and respiratory syndrome virus (PRRSv) vaccine on the shedding of wild-type virus from an infected population of growing pigs. Vaccine (2012) doi:10.1016/j.vaccine.2011.10.075.
26. Rawal, G. et al. Development and validation of a reverse transcription real-time PCR assay for specific detection of PRRSGard vaccine-like virus. Transbound Emerg Dis 69, 1212–1226 (2022).
27. Díaz, I. et al. In silico prediction and ex vivo evaluation of potential T-cell epitopes in glycoproteins 4 and 5 and nucleocapsid protein of genotype-I (European) of porcine reproductive and respiratory syndrome virus. Vaccine (2009) doi:10.1016/j.vaccine.2009.07.029.
28. Nan, Y. et al. Improved vaccine against PRRSV: Current Progress and future perspective. Frontiers in Microbiology Preprint at https://doi.org/10.3389/ fmicb.2017.01635 (2017).
29. Risser, J. et al. Porcine reproductive and respiratory syndrome virus genetic variability a management and diagnostic dilemma. Virology Journal Preprint at https:// doi.org/10.1186/s12985-021-01675-0 (2021).
30. Vandenbussche, F., Mathijs, E., Tignon, M., Vandersmissen, T. & Cay, A. B. Wgs versus orf5 based typing of prrsv: A belgian case study. Viruses (2021) doi:10.3390/v13122419.
31. Lu, Z. H., Archibald, A. L. & Ait-Ali, T. Beyond the whole genome consensus: Unravelling of PRRSV phylogenomics using next generation sequencing technologies. Virus Res (2014) doi:10.1016/j. virusres.2014.10.004.
32. Cortey, M., Díaz, I., Martín-Valls, G. E. & Mateu, E. Next-generation sequencing as a tool for the study of Porcine reproductive and respiratory syndrome virus (PRRSV) macro- and micro- molecular epidemiology. Veterinary Microbiology Preprint at https://doi. org/10.1016/j.vetmic.2017.02.002 (2017).
33. Dortmans, J. C. F. M., Buter, G. J., Dijkman, R., Houben, M. & Duinhof, T. F. Molecular characterization of type 1 porcine reproductive and respiratory syndrome viruses (PRRSV) isolated in the Netherlands from 2014 to 2016. PLoS One (2018) doi:10.1371/journal.pone.0218481.
34. Gagnon, C. A., Lalonde, C. & Provost, C. Porcine reproductive and respiratory syndrome virus wholegenome sequencing efficacy with field clinical samples using a poly(A)-tail viral genome purification method. Journal of Veterinary Diagnostic Investigation (2021) doi:10.1177/1040638720952411.
35. Park, J. Y. et al. Reverse-transcription loop-mediated isothermal amplification (RT-LAMP) assay for the visual detection of European and North American porcine reproductive and respiratory syndrome viruses. J Virol Methods (2016) doi:10.1016/j.jviromet.2016.08.008.
36. Tian, X. X. et al. Rapid visual detection of porcine reproductive and respiratory syndrome virus via recombinase polymerase amplification combined with a lateral flow dipstick. Arch Virol (2022) doi:10.1007/ s00705-021-05349-8.
37. Tan, S., Dvorak, C. M. T. & Murtaugh, M. P. Rapid, unbiased PRRSV strain detection using MinION direct RNA sequencing and bioinformatics tools. Viruses (2019) doi:10.3390/v11121132.
38. Anderson, T. K. et al. The United States Swine Pathogen Database: integrating veterinary diagnostic laboratory sequence data to monitor emerging pathogens of swine. Database (2021) doi:10.1093/database/baab078.
51 ONE HEALTH
PODEMOS HACER PARA AUMENTAR LA PROTECCIÓN FRENTE AL PRRS?
P ¿QUÉ MÁS
Descarga el PDF
EEl Síndrome Respiratorio y Reproductivo Porcino (PRRS), cuyo agente etiológico es un virus (PRRSv), es una de las enfermedades del ganado porcino que mayores pérdidas económicas ocasiona, siendo de muy difícil control en los sistemas de producción actuales.

Hoy en día, en algunos países no solo se está trabajando en el control de la enfermedad a nivel de granja, sino que se está apostando por el control regional del virus e incluso se han llegado a implementar planes de erradicación de la enfermedad a nivel nacional que incluyen la combinación de estrategias de bioseguridad, manejo (Sistema McRebel) y protocolos de vacunación.
 Sonia Cárceles, Laura Garza, Salvador Oliver, Carlos Casanovas y David Espigares
Sonia Cárceles, Laura Garza, Salvador Oliver, Carlos Casanovas y David Espigares
Nº 7 | Abril 2023 VACUNACIÓN ¿Qué más podemos hacer para aumentar la protección frente al PRRS? 52
Servicio Técnico Porcino, Ceva Salud Animal
LA VACUNACIÓN REDUCE LA CIRCULACIÓN DEL PRRSV EN GRANJAS INFECTADAS Y DISMINUYE LA CLÍNICA ASOCIADA A LA ENFERMEDAD
En los últimos años, para el control y erradicación del PRRSv se han utilizado diversas estrategias, entre ellas, los protocolos de vacunación mixtos.
Estos protocolos, que están basados en la combinación de una vacuna viva atenuada (MLV) y una vacuna inactivada (KV), han demostrado ser una alternativa eficaz para controlar la circulación del PRRSv en las granjas, especialmente cuando la vacuna inactivada se aplica a las cerdas al final del periodo de gestación.
EL USO DE VACUNAS INACTIVADAS
PROPORCIONA MAYOR SEGURIDAD A PRODUCTORES Y VETERINARIOS AL NO INTRODUCIR MÁS CEPAS VIVAS DEL VIRUS EN LA GRANJA CON POSIBILIDAD DE RECOMBINAR CON LAS CEPAS PRESENTES EN LA MISMA

En este artículo se exponen los resultados de dos nuevos estudios de campo publicados en 2022.
El primer estudio (Martin-Valls y cols.) valora los resultados reproductivos y la inmunidad frente a PRRSv en lechones descendientes de cerdas vacunadas con dos protocolos vacunales distintos que incluyen o no una KV (Progressis®)
1 2 INMUNIDAD
P
El segundo trabajo expuesto (Pertich y cols.) describe las acciones llevadas a cabo en una granja y los resultados de estas para la erradicación de PRRSv, iniciando el proceso utilizando una KV (Progressis®) 53
EXPERIENCIA DE CAMPO Nº 1 RESULTADOS REPRODUCTIVOS
E INMUNIDAD
DISEÑO EXPERIMENTAL
Este estudio se llevó a cabo en dos granjas, ambas en una situación de inestabilidad frente al PRRS, con producción de lechón de 6-7 kg:
GRANJA 1
300 reproductoras y manejo en bandas cada tres semanas.
GRANJA 2
1.700 reproductoras y manejo en bandas semanales.
Al inicio del estudio, se introdujeron cerdas de reposición libres de PRRSv que fueron vacunadas con dos dosis de MLV separadas cuatro semanas. Además, todas las reproductoras fueron vacunadas en sábana con una dosis de MLV.
Durante seis bandas consecutivas, se establecieron dos grupos de cerdas en cada banda en función del protocolo vacunal a aplicar (Tabla 1), Grupo Control (GC) y Grupo Progressis (GP).
Ambos grupos fueron vacunados con una MLV en la 8ª semana de gestación.
A las cerdas del Grupo Progressis (GP) se les administró una dosis de vacuna inactivada (Progressis®, Ceva) cuatro semanas antes de la fecha prevista de parto.
En la Granja 2, las nulíparas en cuarentena del Grupo Progressis recibieron una dosis de KV antes de la 1ª inseminación, mientras que en la Granja 1 solo se les aplicó a las nulíparas del 6º lote del Grupo GP.
Los lechones descendientes de las cerdas del Grupo Progressis y del Grupo Control fueron destetados y alojados en salas de destete separadas e independientes.
PROTOCOLO VACUNAL Nulíparas Reproductoras en SÁBANA Reproductoras en CICLO

GRUPO C (CONTROL)
2 dosis MLV (separadas 4 semanas)
2 dosis MLV (separadas 4 semanas)
GRANJA 1
GRUPO PG (KV-Progressis)
GRUPO C (CONTROL)
1 dosis KV (4 semanas después)
2ª dosis MLV (4 semanas antes de la cubrición), solo en el 6º lote
2 dosis MLV (separadas 4 semanas)
GRANJA 2
TABLA 1
GRUPO PG (KV-Progressis)
2 dosis MLV (separadas 4 semanas)
1 dosis KV (4 semanas después)
2ª dosis MLV (4 semanas antes de la cubrición)
Protocolos vacunales frente a PRRSv aplicado en las cerdas.
3 veces/año
3 veces/año
MLV: 8 semanas de gestación
MLV: 8 semanas de gestación
KV: 4 semanas antes del parto
3 veces/año
MLV: 8 semanas de gestación
MLV: 8 semanas de gestación
3 veces/año
KV: 4 semanas antes del parto
Nº 7 | Abril 2023 VACUNACIÓN ¿Qué más podemos hacer para aumentar la protección frente al PRRS? 54
En ambos grupos se seleccionan aleatoriamente lechones procedentes de diferentes camadas (Tabla 2) para evaluar los niveles de anticuerpos mediante ELISA y seroneutralización, para determinar la presencia de anticuerpos neutralizantes (AN).
GRANJA GRUPO Nº CAMADAS Nº LECHONES MUESTREO (Nacimiento: cordones umbilicales; al destete y transición: sangre)
6 cordones umbilicales /camada
GRANJA 1 (155 camadas)
GRUPO C (CONTROL) 78 480
GRUPO PG (KVProgressis) 77 473
≥3 lechones al destete/cerda (45 muestras/lote)
6 y 9 semanas (30 muestras/lote)
6 cordones umbilicales/camada
≥3 lechones al destete/cerda (45 muestras/lote)
6 y 9 semanas (30 muestras/lote)
6 cordones umbilicales/camada
GRANJA 2 (148 camadas)
TABLA 2
GRUPO C (CONTROL) 71 418
GRUPO PG (KVProgressis) 77 448
Número de animales por grupo y granja y tipo de muestreo.
RESULTADOS
≥3 lechones al destete/cerda (45 muestras/lote)
6 y 9 semanas (30 muestras/lote)
6 cordones umbilicales/camada
≥3 lechones al destete/cerda (45 muestras/lote)
6 y 9 semanas (30 muestras/lote)
Los resultados obtenidos (Tabla 3) muestran una mayor homogeneidad en los valores de anticuerpos de los lechones descendientes de cerdas del GP, presentando unos títulos de anticuerpos neutralizantes más elevados, con una mayor proporción de animales ELISA positivos, lo que sugiere que este protocolo potencia la inmunidad humoral neutralizante frente al virus.
TÉCNICA/ EDAD GRANJA 1 GRANJA 2 (2 primeros lotes)
TÉCNICA/ EDAD GRANJA 1 GRANJA 2 (2 primeros lotes)
ELISA POSITIVO AL DESTETE GRUPO PG GRUPO C GRUPO PG GRUPO C TEST GRUPO PG GRUPO C GRUPO PG GRUPO C 94,1% lechones 86,5% Media 55 INMUNIDAD
En este trabajo no se observaron diferencias en la productividad al parto o en el número de camadas positivas a PRRSv al nacimiento. Sin embargo, los valores obtenidos de Ct >30 (Tabla 4) muy probablemente pueden estar relacionados con una contaminación ambiental de las instalaciones, secreciones de cerdas o lechones de la camada infectados.
TÉCNICA/ EDADMUESTRA GRANJA 1 GRANJA 2
RT-QPCR GRUPO PG GRUPO C GRUPO PG GRUPO C
CORDÓN UMBILICAL
12,7% camadas positivas Ct: 34,5 ± 3,4
6 SEMANAS DE VIDA
TABLA 4
14,8% incidencia (p < 0,05) Ct: 32,6 ± 4,8 (p = 0,0503)
10,9% camadas positivas Ct: 36,5 ± 3,1
25,5% incidencia (Ct) 29,7 ± 5,8
Resultados del análisis de RT-qPCR.
14,3% camadas positivas Ct: 37,9
2 primeros lotes: 11,3% incidencia (p < 0,05)
Ct 34,3 ± 2,9
8.4% camadas positivas Ct: 37,5
2 primeros lotes: 22,4% incidencia Ct: 32., ± 3,2
En la mayoría de los lotes, la incidencia de PRRSv a las 4 o 6 semanas de vida fue menor en los lechones del GP, lo que puede estar relacionado con una mayor proporción de animales seropositivos al destete, sugiriendo un efecto beneficioso de la vacunación preparto.
Se observó que los descendientes de cerdas nulíparas del Grupo GP tendieron a tener un título de AN más alto, aunque en la Granja 2 esto se detectó en todas las paridades.
No se apreciaron diferencias a nivel reproductivo. Sin embargo, los resultados sugieren que la vacunación tiene un efecto significativo sobre la reducción de la mortalidad de los lechones en lactación.
No se observaron diferencias significativas en la mortalidad durante la etapa de transición. No obstante, la transmisión del virus fue menos eficiente en el Grupo GP, lo que podría estar relacionado con una mayor protección después del nacimiento.
DIFERENCIAS OBSERVADAS
ESTOS RESULTADOS REFLEJAN EL IMPACTO DE LA VACUNACIÓN CON UNA SOLA DOSIS DE VACUNA INACTIVADA ANTES DEL PARTO, CON LO QUE CABRÍA DEDUCIR QUE, A TRAVÉS DE SUCESIVAS VACUNACIONES EN LOS SIGUIENTES CICLOS, SE LOGRARÍAN AUMENTAR LAS
Nº 7 | Abril 2023 VACUNACIÓN ¿Qué más podemos hacer para aumentar la protección frente al PRRS? 56
EXPERIENCIA DE CAMPO Nº 2 ERRADICACIÓN DE PRRSV
DISEÑO EXPERIMENTAL
Esta experiencia se realizó en una granja de ciclo cerrado de Hungría con 1.250 reproductoras y 225 cerdas de reposición (autoreposición) infectada por PRRSv en 2008.
Al mes de la infección, se inició un protocolo de inmunización frente a PRRS que se mantuvo hasta 2014 y que incluyó la aplicación de una vacuna inactivada (Progressis®, Ceva).
A partir de ese momento, comenzó un programa nacional de erradicación del virus, con lo que se intensificó el desecho de cerdas, produciéndose lechones de 70-80 días de vida, además de continuar con la producción cerdas de reposición.
Las cerdas de reposición se vacunaban con 5 dosis de vacuna inactivada:
Las dos primeras dosis se aplicaban a los 60 y 90-100 días de vida, respectivamente.
La tercera dosis se administraba un mes antes de la inseminación.
La cuarta vacunación se realizaba a los 60-70 días de gestación.
La quinta dosis se aplicaba en el momento del destete.
LA
VACUNACIÓN
SOSTENIDA EN EL TIEMPO CON ESTE PROTOCOLO PERMITE PRODUCIR ANIMALES NEGATIVOS A PRRSV MEDIANTE LA TÉCNICA RT-PCR A LOS 90 DÍAS DE VIDA


En una siguiente fase, las cerdas de reposición se transportaban desde la granja de reproductoras (Granja A) a dos naves vacías (Granjas B y C) que estaban alejadas 6-20 km de la granja A y de otras granjas endémicas a PRRSv.
Las cerditas de reposición producidas en la Granja A se llevaban negativas a PRRSv (RT-PCR) con 7 semanas de vida a la Granja B, permaneciendo allí hasta las 14 semanas de vida, y siendo trasladadas posteriormente a la Granja C tras confirmar que eran serológicamente negativas a PRRSv.
Las futuras reproductoras eran muestreadas a los 60 días de su entrada en la Granja C y aquellas serológicamente negativas eran inseminadas al llegar a la edad reproductiva.
Al mismo tiempo, en la Granja A se dejó de inseminar y todas las cerdas fueron sacrificadas después del último parto, realizándose una limpieza y desinfección de esta granja y, aproximadamente, a los 3 meses una repoblación con nulíparas gestantes libres de PRRSv, procedentes de la Granja C
INMUNIDAD 57
1 2 VACUNACIÓN
RESULTADOS
La clasificación de la Granja A como libre de PRRSv se estableció testando serológicamente a todas las cerdas dos veces con un intervalo de seis meses y fue declarada libre de PRRSv en 2019, manteniendo ese estado libre del virus (17 de febrero de 2022).
CONCLUSIONES
De los trabajos expuestos en este artículo se puede concluir lo siguiente:
Reducir la transmisión de PRRSv en lactación y transición es crucial para controlar la enfermedad.
Este objetivo está apoyado en protocolos de vacunación que incluyen una vacuna inactivada.
Estos programas vacunales producen una inmunidad maternal específica de PRRSv más elevada y homogénea, reduciendo la infección por PRRSv a edades tempranas.
BIBLIOGRAFÍA
Berton P, Normand V, Martineau GP, Bouchet F, Lebret A, WaretSzkuta A. Evaluation of porcine reproductive and respiratory syndrome stabilization protocols in 23 French Farrow-to-finish farms located in a high-density swine area. Porcine Health Manag. 2017;3:11.
Boivent et al, 2016. 24th IPVS/8th ESPHM

Defoort et al, 2014. Proceedings 23rd IPVS, p:565
Dekens et al, 2013. Proceedings 5th ESPHM, p: 190
Díaz I, Gimeno M, Callén A, Pujols J, López S, Charreyre C, et al. Comparison of different vaccination schedules for sustaining the immune response against porcine reproductive and respiratory syndrome virus. Vet J. 2013;197:438–44.
Fablet C, Renson P, Eono F, Mahé S, Eveno E, Le Dimna M, et al. Maternally-derived antibodies (MDAs) impair piglets’ humoral and cellular immune responses to vaccination against porcine reproductive and respiratory syndrome (PRRS). Vet Microbiol. 2016;192:175–80
Holtkamp DJ, Kliebenstein JB, Neumann EJ, Zimmerman JJ, Rotto HF, Yoder TK, et al. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J Swine Health Prod. 2013;21:72–84
Jung-Ah L, Nak-Hyung L, Joong-Bok L, Seung-Yong P, Chang-Seon S, In-Soo C, et al. Augmented immune responses in pigs immunized with an inactivated porcine reproductive and respiratory syndrome virus containing the deglycosylated glycoprotein 5 under feld conditions. Clin Exp Vaccine Res. 2016;5:70–4.
Knockaert et al, 2015. 7th ESPHM
Lopez OJ, Oliveira MF, Garcia EA, Kwon BJ, Doster A, Osorio FA. Protection against porcine reproductive and respiratory syndrome virus (PRRSV) infection through passive transfer of PRRSV-neutralizing antibodies is dose dependent. Clin Vaccine Immunol. 2007;14:269–75
Martín-Valls GE, Mortensen P, Clilvert H, Li Y, Cortey M, Sno M,Barna T, Terré M, Guerra N, Mareu E. The use of a whole inactivated PRRS virus vaccine administered in sows and impact on maternally derived immunity and timing of PRRS virus infection in piglets.
Vet Rec Open. 2022;9:e34
Martín-Valls GE, Hidalgo M, Cano E, Mateu E. Testing of umbilical cords by real time PCR is suitable for assessing vertical transmission of porcine reproductive and respiratory syndrome virus under field conditions.
Vet J. 2018;4:27–9
Nilubol et al, 2007. 5th ISERPD, p:146
Pertich A, Barna Z, Makai O, Farkas J, Molnár T, Bálin A, Szabó I, Albert M. Elimination of porcine reproductive and respiratory syndrome virus infection using an inactivated vaccine in combination with a roll-over method in a Hungarian large-scale pig herd. Acta Veterinaria Scandinavica. 2022; 64:12
Schindler et al, 2019. Proceedings 11th ESPHM, IMMPP-46, p:312
Schindler et al, 2019. Proceedings 11th ESPHM, VVDPP-52, p:417
Spaans et al, 2016. 24th IPVS/8th ESPHM
Willems et al, 2015. Proceedings 7th ESPHM p:165
Yoon IJ, Joo HS, Goyal SM, Molitor TW. A modified serum neutralization test for the detection of antibody to porcine reproductive and respiratory syndrome virus in swine sera. J Vet Diagn Invest. 1994;6: 289–92.
Zimmerman et al, 2012. 10th edition Diseases of Swine
La combinación de estrictas medidas de bioseguridad y manejo (realizar la fase de engorde fuera de la granja de reproductoras, aumento del desecho de cerdas, adecuada gestión de la reposición, etc.), junto a la realización de una vacunación regular con una vacuna inactivada puede proporcionar animales negativos a PRRSv y ha demostrado ser clave en el éxito de un programa de erradicación de PRRS. 58 Nº 7 | Abril 2023 ¿Qué más podemos hacer para aumentar la protección frente al PRRS?
Progressis® Emulsión inyectable para cerdos Composición cualitativa y cuantitativa. Cada dosis de 2 ml contiene: Virus del Síndrome Respiratorio y Reproductivo Porcino (PRRS) inactivado Cepa EU P120: 102,5 unidades IF Gentamicina como tazas Excipiente oleoso Indicaciones En cerdas adultas y futuras reproductoras: reducción de los trastornos de la reporducción causados por el Virus del Síndrome Respiratorio y Reproductivo Porcino (Cepa europea) en medio contaminado La vacunación reduce el número de partos prematuros y el número de lechones mal nacidos muertos Posología y modo de administración: Respetar las condiciones habituales de asepsia. Una dosis de 2 ml por vía intramuscular profunda, en los músculos del cuello, detrás de la oreja, de acuerdo con el protocolo de vacunación siguiente: Primovacunación: nulíparas: 2 inyecciones, con 3-4 semanas de intervalo al menos 3 semanas antes de la cubrición; cerdas adultas: 2 inyecciones, con 3-4 semanas de intervalo, (se recomienda vacunar a todas las cerdas de la explotación en un corto período de tiempo). Revacunación: 1 inyección a los 60-70 días de gestación, a partir de la primera gestación siguiente a la primovacunación. Precauciones particulares de uso: Vacunar solamente animales sanos Respetar las condiciones habituales de manipulación de los animales Utilizar la vacuna inmediatamente después de la apertura del frasco En caso de auto-inyección accidental de la vacuna, consúltese inmediatamente con un médico mostrándole el prospecto o la etiqueta. Precauciones particulares para la la especie de destino: En las explotaciones contaminadas por PRRS, la infección vírica es heterogénea y variable a lo largo del tiempo En este contexto la instauración de un programa de vacunación es una herramienta para mejorar los parámetros reproductivos y puede contribuir al control de la enfermedad en combinación con medidas sanitarias Utilización durante la gestación o la lactación No se ha observado ningún efecto indeseable después de la administración de la vacuna a las hembras gestantes o en lactación. Efectos secundarios: La vacunación puede inducir un edema ransitorio (máximo 3 cm) que generalmente no persiste más de una semana y una reacción local de pequeña magnitud (granuloma) que no tiene ninguna consecuencia en la salud ni en los rendimientos reproductivos de los animales A veces se han observado reacciones más extensas (máximo 7cm) después de vacunaciones repetidas frecuentemente En raros excepto los mencionados en el epígrafe “Efectos secundarios Contradicciones Ninguna conocida. Interacciones: No se ha observado ningún efectoindeseable en la respuesta serológica después de la administración simultánea, pero en un punto de inyección diferente, Incompatibilidades: No mezclar con otros medicamentos Presentación: Viales de 5 dosis (10ml) y 25 dosis (50ml). Tíempo de espera: Cero días Precauciones especiales de almacenamiento Consérvese en el refrigerador, entre 2ºC-8ºC, al abrigo de la luz. Precauciones especiales para la eliminación del producto no utilizado o material de desecho: Los frascos y cualquier resto de producto deben ser eliminados de acuerdo con los requirimientos de la legislación nacional vigente sobre residuos



Salud Animal, S.A. Avda. Diagonal 609 – 615 08028 Barcelona. Sí al cambio, sí al progreso
r e f o r z a n d o la inmunidad Ceva Salud Animal, Avenida Diagonal, 609 - 615, 9ª planta 08028 Barcelona Tel. (+34) 902 367 218 - www.ceva.es Vacuna
PRRS Consulte con su veterinario
Uso veterinario: Dispensación: “Con prescripción veterinaria” Nº de registro: 1359. Esp Titular de la autorización de comercialización: Ceva
PR GRES SI S
inactivada frente
A USO RACIONAL DE ANTIMICROBIANOS
EN PORCINO: ANTIMICROBIANOS
INHIBIDORES DE LA SÍNTESIS DE ARN Y ADN

– PARTE II
Descarga el PDF
EEn la primera parte de este artículo abordamos los antibióticos que ejercen su mecanismo de acción a través de la inhibición de los ácidos nucleicos ARN y ADN, centrándonos en las fluoroquinolonas.

En la segunda parte de este artículo, nos ocuparemos de las sulfamidas e inhibidores de la dihidrofolato reductasa, rifamicinas, novobiocina y nitrofuranos y nitroimidazoles. No obstante, estos dos últimos no están autorizados en animales productores de alimentos, al no poder establecer sus LMR (Límite Máximo de Residuos) (Anexo 2 del Reglamento 37/2010).
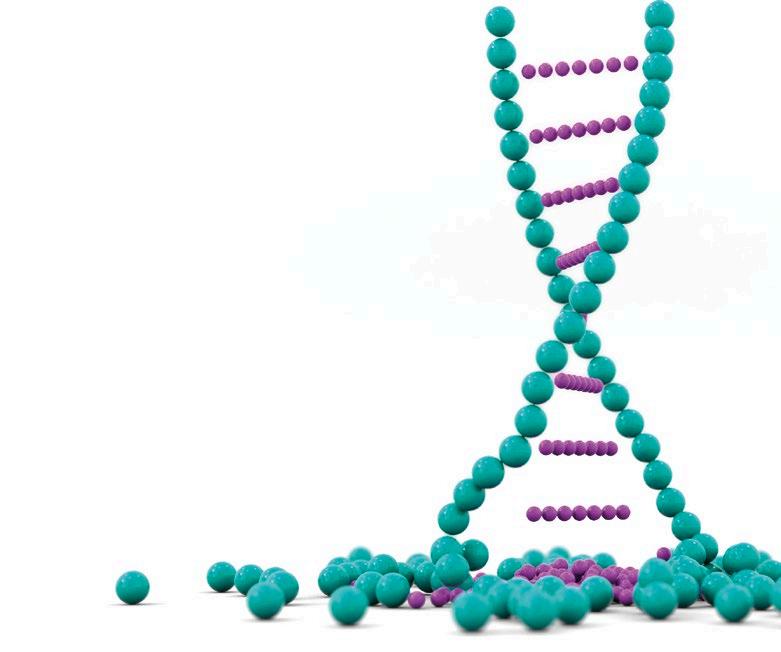
Sonia Rubio, Mª Dolores San Andrés, Manuel I. San Andrés y Juan Antonio Gilabert

Sección Dptal. de Farmacología y Toxicología, Facultad de Veterinaria, Universidad Complutense de Madrid

Nº 7 | Abril 2023 ANTIMICROBIANOS Uso racional de antimicrobianos en porcino: antimicrobianos inhibidores de la síntesis de ARN y ADN – Parte II 60
SULFAMIDAS (SULFONAMIDAS) Y TRIMETOPRIMA ESTRUCTURA QUÍMICA

Las sulfamidas fueron el primer gran grupo de medicamentos utilizados para el tratamiento de infecciones bacterianas en el ser humano, especialmente las causadas por estreptococos.
Fueron descubiertas en la década de 1930 a partir del Prontosil, un colorante rojo con actividad antiinfecciosa, pero que era ineficaz in vitro. Más tarde, se descubrió que en el interior del organismo se metabolizaba a sulfanilamida, un compuesto con capacidad antibacteriana y a partir de la cual se sintetizaron otras sulfamidas con menor toxicidad y amplio espectro.
Las sulfamidas comparten cierta semejanza estructural con el ácido p-aminobenzoico (para-aminobenzoico o PABA), con posibilidad de añadir diferentes radicales, siendo un grupo muy amplio de compuestos sintéticos.
SULFAMIDAS
Las sulfamidas o “sulfas” son un grupo de compuestos sintéticos derivados amida del ácido sulfónico benceno (sulfonamidas) con un amplio espectro de acción antibacteriana bacteriostática (Gram positivas, Gram negativas y protozoos como los coccidios), pero ineficaces, en gran medida, frente a anaerobios obligados.

TRIMETOPRIMA
La trimetoprima (TMP) es un fármaco diaminopirimidina sintético activo frente a bacilos Gram negativos aerobios y algunos microorganismos Gram positivos.
Las sulfamidas y la trimetoprima se suelen usar en asociación, ya que inhiben pasos enzimáticos secuenciales en la ruta de síntesis del ácido fólico en las bacterias que no pueden obtenerlo del medio (Figura 1), como ocurre en los mamíferos.
El ácido fólico es imprescindible para la síntesis de precursores de ácidos nucleicos (purinas y timidina), pero las células de los mamíferos requieren ácido fólico preformado, ya que no pueden sintetizarlo y, por tanto, no se ven afectadas por la acción de las sulfamidas.
A
DE
DE ACTIVIDAD
ESPECTRO
ACTIVIDAD ESTRUCTURA QUÍMICA ESPECTRO
ONE HEALTH 61
MECANISMO DE ACCIÓN MECANISMO DE ACCIÓN
SULFAMIDAS
Debido a su analogía estructural con el PABA, las sulfamidas se unen de forma competitiva a la enzima dihidropteroato sintetasa que se encarga de sintetizar dihidropteroato (ácido dihidropteroico) a partir de PABA y pteridina (dihidropteroato difosfato), y al cual se unirá seguidamente ácido glutámico para formar ácido dihidrofólico (ácido fólico).
TRIMETOPRIMA
La TMP impide la síntesis de ácido tetrahidrofólico (o ácido folínico) a partir de dihidrofolato al unirse a la enzima dihidrofolato reductasa (DHFR) con una afinidad 10.000 veces mayor por la DHFR bacteriana que por la de los mamíferos, por lo que representa una diana selectiva.
Las diaminopirimidinas, como la trimetoprima, compiten por la enzima dihidrofolato reductasa que cataliza la conversión de ácido dihidrofólico en ácido tetrahidrofólico, existiendo otros derivados sintéticos de las diaminopirimidinas como la pirimetamina (uso en aves ornamentales junto a sulfaquinoxalina).

TRIMETOPRIMA
Ácido dihidrofólico
SULFAMIDA
Dihidropteroato sintetasa (DHPS)
Ácido dihidropteroico
Dihidropteroato reductasa (DHFR)
Ácido tetrahidrofólico
FIGURA 1
Ruta de síntesis del ácido tetrahidrofólico y puntos de disrupción de la misma debido a sulfonamidas y trimetoprima. La sulfonamida, debido a su similitud estructural con el PABA, se une de forma competitiva a la dihidropteroato sintetasa (DHPS), impidiendo la formación de ácido dihidropteroico. En un paso posterior, la trimetoprima se une a la dihidrofolato reductasa (DHFR), impidiendo la formación de ácido tetrahidrofólico (creado con BioRender.com).
Nº 7 | Abril 2023 ANTIMICROBIANOS Uso racional de antimicrobianos en porcino: antimicrobianos inhibidores de la síntesis de ARN y ADN – Parte II 62
CARACTERÍSTICAS FARMACOCINÉTICASCARACTERÍSTICAS FARMACOCINÉTICAS
SULFAMIDAS
Las sulfamidas son fármacos poco solubles, más solubles a pH alcalino, por lo que cuando aumenta la diferencia entre su pK y el pH del medio (ej. orina) precipitan.
Son ácidos débiles y, dado su pKa entre 5 y 10,5, suelen encontrarse en el organismo en su forma neutra. Por ello, atraviesan las barreras biológicas con facilidad y tienen buena distribución tisular, salvo excepciones.
LA MAYORÍA DE LAS SULFAMIDAS SE ABSORBEN
BIEN EN EL TRACTO GASTROINTESTINAL
Y TIENEN UNA AMPLIA DISTRIBUCIÓN ALCANZANDO INCLUSO LOS LÍQUIDOS SINOVIAL Y CEFALORRAQUÍDEO
Al ser un grupo de compuestos muy amplio, las propiedades de las sulfamidas pueden ser muy variables. Por ejemplo, sus tasas de unión a proteínas plasmáticas suelen ser altas y sus semividas muy variables.
El metabolismo de las sulfamidas es principalmente hepático y se inactivan por acetilación, glucuronoconjugación e hidroxilación, excretándose el fármaco y sus metabolitos principalmente a través del riñón.
No obstante, se debe tener en cuenta que los derivados acetilados son menos solubles en la orina y pueden precipitar en el riñón (túbulos renales) y causar cristaluria, por lo que se recomienda una buena ingesta de agua durante el tratamiento y, si es necesario, alcalinizar la orina.

TRIMETOPRIMA
Las diaminopirimidinas (trimetoprima) son bases débiles y muy liposolubles, con una absorción oral muy rápida. Tienen una amplia distribución en el organismo y persisten unidas a los tejidos, pudiendo quedar retenidas en próstata y leche debido a que su pH es menor que el del plasma.
Su metabolismo es hepático, parcialmente, y su eliminación renal y, en menor porcentaje, digestiva.
LAS DIAMINOPIRIMIDINAS SE USAN
NORMALMENTE COMO POTENCIADORAS DE LAS
SULFAMIDAS EN UNA RELACIÓN 1:5
El efecto de la trimetoprima es bacteriostático, pero cuando se combina con las sulfonamidas es bactericida.
De esta forma, las sulfonamidas combinadas con trimetoprima (también denominadas sulfonamidas potenciadas) provocan un bloqueo sucesivo de dos enzimas que desempeñan un importante papel en el metabolismo de las bacterias y los protozoos. Su efecto combinado es, por tanto, sinérgico y de más amplio espectro.
ONE HEALTH 63
MECANISMOS DE RESISTENCIA MECANISMOS DE RESISTENCIA
SULFAMIDAS
La resistencia a las sulfamidas es frecuente y puede producirse por distintos mecanismos. Dado que son compuestos muy similares estructuralmente, la resistencia cruzada entre las diferentes sulfamidas es completa y aquellas cepas bacterianas resistentes a una sulfamida serán resistentes a muchas otras, por lo que no se debe cambiar de una a otra si el tratamiento no es satisfactorio.

La resistencia más relevante clínicamente suele estar mediada por plásmidos que provocan cambios en la afinidad por la enzima dihidropteroato sintetasa o bien de origen cromosómico produciendo una disminución de la entrada de fármacos o un exceso de PABA.
Su eficacia también se ve disminuida por un aumento en la concentración de PABA en el medio bacteriano o por lesiones purulentas (con alta cantidad de purinas y timidina).
TRIMETOPRIMA
La resistencia a la TMP suele estar mediada por cambios en la enzima DHFR que reduce su afinidad por el fármaco.
Debido a la probable variabilidad (temporal, geográfica) en la sensibilidad bacteriana a las sulfamidas potenciadas, la aparición de las resistencias bacterianas puede diferir de un área geográfica o región a otra e incluso de una explotación a otra, por lo que se recomienda efectuar un muestreo bacteriológico y ensayos de sensibilidad.
EL USO DE ESTOS ANTIMICROBIANOS DEBE BASARSE EN LA SENSIBILIDAD DE LOS MICROORGANISMOS EN CULTIVO AISLADOS
DE ANIMALES ENFERMOS DE LA PROPIA
EXPLOTACIÓN O EN LA EXPERIENCIA PREVIA
RECIENTE DE ÉSTA
Hoy en día, en algunas patologías como la colibacilosis hay un aumento en el uso de alternativas como la vacunación y mejoras en la nutrición, medidas que deben ir acompañadas de una focalización en la mejora de las condiciones de manejo, de higiene y de bioseguridad.
Nº 7 | Abril 2023 ANTIMICROBIANOS Uso racional de antimicrobianos en porcino: antimicrobianos inhibidores de la síntesis de ARN y ADN – Parte II 64
INTERACCIONES, EFECTOS SECUNDARIOS Y TOXICIDAD INTERACCIONES, EFECTOS SECUNDARIOS Y TOXICIDAD
Las sulfamidas presentan antagonismo con el ácido p-aminobenzoico (PABA) y sus derivados (procaína -penicilina-, benzocaína, tetracaína, etc.) por lo que no deben administrarse simultáneamente con estos compuestos ni con sustancias que aporten o liberen PABA y/o ácido fólico. Tampoco se debería administrar con anticoagulantes orales o acidificantes urinarios. Los efectos secundarios principales de las sulfamidas, aunque se producen en muy raras ocasiones (1 cada 10.000 animales) son:
Inflamación en el punto de inyección
Cristaluria, hematuria y obstrucción de vías renales
Rigidez muscular
Shock anafiláctico
Shock respiratorio
Shock circulatorio
Trastornos hematopoyéticos (trombocitopenia y leucopenia) o discrasias
Déficit de vitamina K o hipoprotrombinemia
Emesis, irritación gastrointestinal y diarrea

El uso de estos medicamentos implica que se excreten en mayor o menor medida al medio ambiente a través de la orina o de las heces.
Los antibióticos mal metabolizados también se pueden acumular en el suelo, lo que puede afectar las comunidades y funciones microbianas del mismo o pueden bioacumularse también en el medio acuático si no están expuestos a la luz solar.
Por otro lado, se puedan encontrar en niveles bajos, pero detectables, en los tejidos comestibles de animales productores de carne tratados.
65 ONE HEALTH ONE HEALTH
SOLAS
SULFADIMIDINA (Categoría D)
Existen autorizadas 2 especialidades de sulfadimidina sódica en polvo para la administración en agua de bebida.
Posee un amplio espectro de acción frente a un gran número de patógenos entre los que caben destacar bacterias Gram negativas (Salmonella spp., E. coli, Klebsiella spp., Haemophilus spp., Pasteurella spp., Bordetella bronchiseptica, Fusobacterium necrophorum, etc.), algunas bacterias Gram positivas (Streptoccoccus spp.) y coccidios.


Están indicadas en porcino para el tratamiento de infecciones respiratorias y enteritis.
Tras la administración por vía oral, la sulfadimidina se absorbe rápidamente y se distribuye ampliamente por el organismo, alcanzando la concentración plasmática máxima entre 1 y 2 horas.
La posología es de una dosis de choque de 110 mg de sulfadimidina/ kg peso vivo (p.v.) el primer día y 55 mg de sulfadimidina/kg p.v./día durante 4 días consecutivos.
No se debe usar en casos de hipersensibilidad conocida a las sulfonamidas y/o a algún excipiente, en animales con insuficiencia hepática o renal o con discrasias sanguíneas.
Se debe manipular el medicamento con cuidado para evitar el contacto durante la incorporación al agua, así como durante la administración a los animales, tomando precauciones específicas como la de llevar equipo de protección individual (mascarilla antipolvo, guantes, mono de trabajo y gafas de seguridad aprobadas).
El tiempo de espera en carne es de 28 o 40 días (según la especialidad).

USO TERAPÉUTICOUSO TERAPÉUTICO
Nº 7 | Abril 2023 ANTIMICROBIANOS Uso racional de antimicrobianos en porcino: antimicrobianos inhibidores de la síntesis de ARN y ADN – Parte II 66
COMBINACIONES
SULFADIAZINA CON TRIMETOPRIMA (SDZ+TMP) (COTRIMACINA) (Categoría D)
La asociación de sulfadiazina con trimetropima se presenta en varias especialidades y formas. Esta combinación está indicada en el tratamiento del síndrome mastitis-metritis-agalaxia, infecciones respiratorias causadas por Bordetella bronchiseptica (ej. rinitis atrófica), Actinobacillus pleuropneumoniae, Pasteurella multocida, Haemophilus parasuis o Streptococcus suis y, así como diarreas causadas por cepas de E. coli y Salmonella spp. sensibles a la sulfadiazina y trimetoprima.

La sulfadiazina se considera una sulfonamida semiretardada con una persistencia de sus niveles plasmáticos bastante larga, con una buena distribución y alta unión a proteínas plasmáticas. Ambos ingredientes activos se metabolizan parcialmente en el hígado. Su excreción es esencialmente renal.
Existen autorizadas 5 especialidades como premezcla medicamentosa para mezclar con el alimento con 250 mg de sulfadiazina y 50 mg de trimetoprima (5:1).
La dosis para administrar es de 25 mg de sulfadiazina y 5 mg de trimetoprima/kg de p.v./día durante 5 días, constituyendo este pienso medicado la única ración durante este período.
Se debe evitar el contacto con la piel y los ojos y, en caso de contacto con la piel, lavar el área expuesta con agua.
Los tiempos de espera para animales productores de carne son:

Vía oral: 5 días
Vía intramuscular (IM): 7 días
Vía intravascular (IV): 15 o 20 días
Existen 2 especialidades como polvo para administración en agua de bebida.
La dosis es de 25 mg de sulfadiazina + 5 mg de trimetoprima/kg de p.v./día durante 5 días consecutivos.
El tiempo de espera para animales productores de carne es 4 o 19 días (según especialidad).
Existen 2 especialidades como solución para la administración en agua de bebida o en leche.
La dosis es de 25 mg de sulfadiazina y 5 mg de trimetoprima por kg de p.v. por día, durante 4 a 7 días consecutivos, por vía oral, diluidos en agua potable o leche. El agua medicada será la única fuente de bebida y deberá ser renovada cada 24 horas.
El tiempo de espera es de 6 o 12 días (según especialidad).
ONE HEALTH 67
SULFAMETOXAZOL CON TRIMETOPRIMA (COTRIMOXAZOL) (Categoría D)
El sulfametoxazol es una sulfamida de acción corta a intermedia. La asociación sulfametoxazol con trimetoprima (SMX+TMP) también se conoce con el nombre de cotrimoxazol.
La dosis recomendada es de 20 mg de trimetoprima y 100 mg de sulfametoxazol por ml o 1:5, lo que en el plasma se transforma en 1:20, dado que trimetoprima tiene un volumen de distribución más amplio que sulfametoxazol.

La asociación que se obtiene es una sinergia y al actuar sobre dos etapas de la síntesis de ácido folínico puede llegar a tener un efecto bactericida.
Existen autorizadas tres especialidades que se presentan en la forma de solución para administración en agua de bebida.
La dosis recomendada en cerdos de engorde es de 5 mg de trimetoprima y 25 mg de sulfametoxazol/kg de p.v./día durante 4 a 7 días.
Tras la administración oral, los dos principios activos se absorben rápidamente a nivel intestinal y se observan concentraciones elevadas de trimetoprima en los riñones, el hígado y los pulmones. Salvo en el caso de los riñones, las concentraciones de sulfametoxazol en los tejidos son significativamente menores que en el plasma. La unión a proteínas de la trimetoprima y el sulfametoxazol no es muy alta.
En porcino, la semivida de eliminación de las dos sustancias es de aproximadamente 2,5 horas. En las 48 horas posteriores a la administración de la última dosis, la trimetoprima, el sulfametoxazol y sus metabolitos ya no son detectables en la orina ni en las heces.
Están indicadas para el tratamiento (cuando la enfermedad ha sido detectada en la piara) de distintos procesos causados por microorganismos sensibles a TMP+SMX como la diarrea postdestete causada por cepas de Escherichia coli beta-hemolítica K88-positivas, K99 positivas o 987P o infecciones bacterianas secundarias provocadas por cepas de Pasteurella multocida, Actinobacillus pleuropneumoniae, Streptococcus spp. y Haemophilus parasuis.
No se debe utilizar en animales que sufran enfermedad renal o hepática grave, oliguria o anuria, ni en animales con alteración del sistema hematopoyético y/o en caso de hipersensibilidad conocida a las sulfamidas o a la trimetoprima.
El excipiente N-metilpirrolidona presente en estas especialidades de SMX+TMP puede dañar al feto tras el contacto con la piel. Por tanto, las mujeres en edad fértil o embarazadas deben actuar con mucho cuidado para evitar la exposición por contacto a estos medicamentos veterinarios. Además, puede causar irritación de la piel y las vías respiratorias, así como daño a los ojos por lo que se deben manipular con guantes impermeables (p. ej., de goma o látex) y gafas protectoras.
Los tiempos de espera en carne son de 5 u 8 días (según la especialidad).
Nº 7 | Abril 2023 ANTIMICROBIANOS Uso racional de antimicrobianos en porcino: antimicrobianos inhibidores de la síntesis de ARN y ADN – Parte II 68
SULFADOXINA CON TRIMETOPRIMA (Categoría D)
La asociación de sulfadoxina con trimetoprima se presenta en una sola especialidad en forma de solución inyectable para su administración por vía intramuscular en cerdos.
Está indicada para el tratamiento de infecciones respiratorias causadas por Actinobacillus pleuropneumoniae, Bordetella bronchiseptica, Pasteurella multocida y/o Mannheimia haemolytica, así como otras infecciones causadas por Haemophilus y/o Streptococcus suis o en diarreas causadas por Escherichia coli.
La dosis recomendada es de 12,5 mg de sulfadoxina + 2,5 mg de trimetoprima/kg de p.v. en dosis única.
En la mayoría de los casos, una única dosis es suficiente, pero si no se alcanza un efecto terapéutico puede repetirse el tratamiento a intervalos de 24 horas durante un máximo de 3 días. No debe de inyectarse un volumen superior a 10 ml en un mismo punto.
Ambas sustancias se absorben individualmente tras la administración parenteral, alcanzándose concentraciones plasmáticas entre las 2 y las 8 horas postadministración. La semivida de eliminación es de 5 a 14 horas para la sulfadoxina y de 0,5 a 4 horas para la trimetoprima, distribuyéndose a todos los tejidos.
El tiempo de espera en carne es de 10 días.
Fármaco Vía de administración
SULFADIMIDINA Oral (PO)
Forma farmacéutica Indicaciones Tiempo de espera en
Polvo para administración en agua de bebida
Premezcla medicamentosa
Polvo para administración en agua de bebida
SULFADIAZINA + TRIMETOPRIMA
Oral (PO)
Intramuscular (IM)
Intravascular (IV)
SULFAMETOXAZOL + TRIMETOPRIMA Oral (PO)
Solución para administración en agua de bebida/leche
Inyectable
Solución para administración en agua de bebida
SULFADOXINA + TRIMETOPRIMA
Intramuscular (IM)
Inyectable
Coccidiosis, Enteritis infecciosa, Infecciones respiratorias

Actinobacilosis, Bordetelosis, Colibacilosis, Complejo respiratorio porcino, Diarrea por E. coli, Pasteurelosis, Rinitis atrófica, Salmonelosis, Síndrome metritis-mastitis-agalaxia, Estreptococosis
Actinobacilosis, Enfermedad de Glässer, Infecciones estreptocócicas, Infecciones respiratorias, Pasteurelosis, Pleuroneumonía
Actinobacilosis, Bordetelosis, Diarrea por E. coli, Infecciones respiratorias, Pasteurelosis
28 o 40 días (según la especialidad)
5 días
4 o 19 días (según la especialidad)
6 (lechones) o 12 días (según la especialidad)
7 días (IM), 15 o 20 días (IV) (según la especialidad)
5 u 8 días (según la especialidad)
10 días
TABLA 1
ONE HEALTH 69
Diferentes especialidades de sulfamidas y trimetoprima de uso terapéutico en porcino.
NOVOBIOCINA
ESTRUCTURA QUÍMICA
ESTRUCTURA QUÍMICA
La novobiocina, también conocida como albamicina o catomicina, es un antibiótico aminocumarina (contiene un núcleo cumarínico) producido por la bacteria Streptomyces niveus.
ESPECTRO DE ACTIVIDAD
ESPECTRO DE ACTIVIDAD
El espectro de la novobiocina es reducido, limitándose a bacterias Gram positivas (Staphycolococcus aureus y Streptococcus) y Gram negativas Brucella spp. y Haemophilus spp
EN VETERINARIA, LA NOVOBIOCINA SE USA
CASI EXCLUSIVAMENTE EN EL TRATAMIENTO DE MASTITIS
MECANISMO DE ACCIÓN
MECANISMO DE ACCIÓN
La novobiocina actúa sobre la topoisomerasa II, concretamente sobre la subunidad GyrB de la enzima involucrada en la transducción de energía.
Actúa como un inhibidor competitivo de la reacción de la enzima ATP sintasa catalizada por GyrB para impedir el superenrollamiento del ADN.
CARACTERÍSTICAS FARMACOCINÉTICAS
CARACTERÍSTICAS FARMACOCINÉTICAS
La novobiocina tiene buena absorción oral (60-70%), pero su volumen de distribución es escaso. Una vez metabolizado, se elimina a través de la bilis y un poco por orina, sufriendo circulación enterohepática.
MECANISMOS DE RESISTENCIA
MECANISMOS DE RESISTENCIA
En el caso de la novobiocina, es frecuente observar resistencias de tipo cromosómico.
INTERACCIONES, EFECTOS SECUNDARIOS Y TOXICIDAD
INTERACCIONES, EFECTOS SECUNDARIOS Y TOXICIDAD
La novobiocina puede provocar reacciones alérgicas, náuseas, vómitos y alteraciones hemáticas como eosinofilia, trombocitopenia y leucopenia.
Nº 7 | Abril 2023 ANTIMICROBIANOS Uso racional de antimicrobianos en porcino: antimicrobianos inhibidores de la síntesis de ARN y ADN – Parte II 70
RIFAMICINAS
ESTRUCTURA QUÍMICA
ESTRUCTURA QUÍMICA
Las rifamicinas son moléculas complejas derivadas del Streptomyces Amycolatopsis mediterranei.
Poseen estructura naftoquinónica, con una larga cadena alifática. Pertenecen al grupo de las ansamicinas.
LA RIFAMPICINA ES LA RIFAMICINA MÁS
UTILIZADA EN VETERINARIA
ESPECTRO DE ACTIVIDAD
ESPECTRO DE ACTIVIDAD
Las rifamicinas afectan a microrganismos extra e intracelulares, incluidos los que se encuentran dentro de macrófagos y neutrófilos, sin afectar la fagocitosis, entre ellos a bacterias Gram positivas, micobacterias, Chlamydia, Rickettsiae, Haemophilus, Brucella, protozoos, hongos y poxvirus.
MECANISMO DE ACCIÓN
MECANISMO DE ACCIÓN
El mecanismo de acción de las rifamicinas consiste en bloquear la subunidad β ARN-polimerasa, dependiente del ADN, impidiendo la síntesis del ARNm. A dosis terapéuticas no afecta a la enzima polimerasa de los mamíferos.
CARACTERÍSTICAS FARMACOCINÉTICAS
CARACTERÍSTICAS FARMACOCINÉTICAS
La rifampicina tiene una absorción oral del 40-68% (según especies), no debiendo administrarse con alimentos porque interfieren su absorción. Se absorben bien vía parenteral y tienen una amplia distribución, alcanzando concentraciones tisulares superiores a las plasmáticas, excepto en líquido cefalorraquídeo (LCR) donde su acceso es muy limitado.
Se metaboliza en hígado (25-40%) y se excreta a través de la bilis y poco por orina, existiendo circulación enterohepática.
LA RIFAMPICINA SE TRANSFORMA EN 25-DESACETILRIFAMPICINA, UN POTENTE INDUCTOR ENZIMÁTICO DEL P-450
MECANISMOS DE RESISTENCIA
MECANISMOS DE RESISTENCIA
Las resistencias a las rifamicinas son de tipo cromosómico por mutaciones de la ARN polimerasa que impide la unión del antibiótico.
INTERACCIONES, EFECTOS SECUNDARIOS Y TOXICIDAD
INTERACCIONES, EFECTOS SECUNDARIOS Y TOXICIDAD
En el caso de la administración de rifamicinas, se pueden observar fenómenos de hepatotoxicidad y reacciones alérgicas (desde leves hasta trombocitopenia anemia hemolítica, shock e insuficiencia renal).
ONE HEALTH 71
NITROFURANOS
ESTRUCTURA QUÍMICA
ESTRUCTURA QUÍMICA
Los nitrofuranos son compuestos sintéticos de amplio espectro basados en el 5-nitrofuraldehido.
EL POTENCIAL EFECTO CARCINÓGENO DE LOS NITROFURANOS HA HECHO QUE EN LA UE ESTÉ
PROHIBIDO SU USO EN ANIMALES PRODUCTORES DE ALIMENTOS
ESPECTRO DE ACTIVIDAD
ESPECTRO DE ACTIVIDAD
Los nitrofuranos tienen un amplio espectro de acción frente a bacterias Gram positivas (Staphylococcus aureus, Streptococcus spp. y Corynebacterium) y Gram negativas (E. coli, Salmonella y Klebsiella), micoplasmas, protozoos (coccidios, tripanosomas) y algunos hongos, mejorando su eficacia a pH 5,5 y reduciéndose ésta en medio alcalino.
Las bacterias anaerobias en condiciones de anaerobiosis son algo sensibles, mientras que las bacterias aerobias son sensibles tanto en condiciones de aerobiosis como de anaerobiosis.
En países donde no está prohibido, se utilizan en cerdos frente a diarrea hemorrágica, diarrea infecciosa, tifosis y paratifosis.
MECANISMO DE ACCIÓN
MECANISMO DE ACCIÓN
El mecanismo de acción de los nitrofuranos se basa en la actividad de las nitrorreductasas bacterianas que los degradan, convirtiéndolos en productos de reducción poco definidos y cuya composición varía de acuerdo con el nitrofurano del que proceden.
Estos compuestos de reducción ocasionan la rotura de las hebras de ADN bacteriano, así como el bloqueo de la traducción del ARNm y de varios procesos metabólicos bacterianos como la producción de acetil coenzima A, a partir de piruvato, alterando la producción de energía por parte de la bacteria.
CARACTERÍSTICAS FARMACOCINÉTICAS
CARACTERÍSTICAS FARMACOCINÉTICAS
La nitrofurantoina se absorbe bien vía oral, tiene una semivida corta y un volumen de distribución bajo, excretándose por orina tras filtración renal, dando un color amarillo. En cambio, la nitrofurazona y furazolidona no se absorben vía oral, utilizándose para procesos intestinales asociados a Salmonella, Treponema o Giardia.
MECANISMOS DE RESISTENCIA
MECANISMOS DE RESISTENCIA
Los nitrofuranos presentan escasas resistencias por su variedad de mecanismos de acción y, cuando aparecen, son de tipo cromosómico, manifestándose con una menor capacidad de penetración del antibiótico o disminución de síntesis de reductasas bacterianas.
INTERACCIONES, EFECTOS SECUNDARIOS Y TOXICIDAD
INTERACCIONES, EFECTOS SECUNDARIOS Y TOXICIDAD
Los nitrofuranos pueden inducir fenómenos de mutagénesis y son procancerígenos, ocasionando también trastornos a nivel del sistema nervioso central (SNC), diátesis hemorrágica, trombocitopenia, anemia, leucocitopenia, aumento del tiempo de hemorragia, anorexia y vómitos.
NITROIMIDAZOLES
ESTRUCTURA QUÍMICA
ESTRUCTURA QUÍMICA
Los nitroimidazoles son compuestos heterocíclicos con un núcleo básico que es un anillo pentagonal similar al nitrofurano (grupos 5-nitro).
En este grupo se incluyen el metronidazol (categoría D, según clasificación UE AMEG), el dimetridazol, ronidazol, tinidazol, ipronodazol y cornidazol.
Nº 7 | Abril 2023 ANTIMICROBIANOS Uso racional de antimicrobianos en porcino: antimicrobianos inhibidores de la síntesis de ARN y ADN – Parte II 72
DÍAS IM 7 Porcino DÍAS IM 3Ovino
DÍAS IM 5Caprino DÍAS IM 6Bovino
Ovino y caprino 2,5DÍASLeche


Bovino DÍAS IV2Leche


















la vía intravenosa. Dosis: En todas las especies: 20 mg de sulfadiazina + 4 mg de trimetoprima / kg p.v./ día (equivalente a 0.1 ml de medicamento/ kg p.v./ día). En infecciones graves la dosis inicial puede ir seguida de la mitad de la dosis recomendada cada 12 horas. El tratamiento debe continuarse hasta 2-3 días después de la desaparición de los síntomas. No prolongar el tratamiento más de 7 días consecutivos. Si no se observa mejoría en tres días, se suspenderá el tratamiento y se revisará el diagnóstico. Debe determinarse el peso de los animales con la mayor exactitud posible para evitar una dosificación insuficiente. En la administración intramuscular, prever una separación suficiente entre los puntos de inyección cuando sean necesarios varios lugares de administración. TIEMPO DE ESPERA: Carne: Bovino
Bovino (únicamente por via intravenosa): 2 días. Ovino y Caprino: 2,5 días (5 ordeños).. PRESENTACIÓN:









ULTRADIAZINE
SOLUCIÓN INYECTABLE
Siempre protegidos COMPOSICIÓN: Sulfadiazina (sódica): 20 g (equivalente a 21,758 g de sulfadiazina sódica). Trimetoprim: 4 g. Excipiente c.s.p.: 100 ml ESPECIES DE DESTINO: Bovino porcino, ovino y caprino. INDICACIONES: Bovino, ovino y caprino: Tratamiento de infecciones respiratorias, gastrointestinales y de metritis causadas por microorganismos sensibles a la asociación sulfadiazina/trimetoprima. Porcino: Tratamiento de infecciones respiratorias, gastrointestinales y del síndrome de disgalaxia postparto – SDP (Síndrome metritis-mastitis-agalaxia) causadas por microorganismos sensibles a la asociación sulfadiazina/trimetoprima. POSOLOGÍA Y MODO DE ADMINISTRACIÓN: Agitar bien antes de usar. Vía intramuscular (IM) o intravenosa (IV) lenta. La administración en vacas lecheras deberá realizarse únicamente mediante
días / IV 15 días, Ovino IM 3 días / IV 15 días y Caprino IM 5
/ IV
250
ESP.
·
·
IM 6 días / IV 15 días, Porcino IM 7
días
15 días. Leche:
Envases de 100 y
ml. Nº de registro: 857
ESPECTRO DE ACTIVIDAD
ESPECTRO DE ACTIVIDAD
Los nitroimidazoles pueden actuar sobre bacterias aerobias, pero en condiciones de anaerobiosis, teniendo actividad bactericida sobre casi todas las bacterias Gram negativas y sobre muchas Gram positivas anaerobias y protozoos.
Se utilizan en veterinaria frente a bacterias anaerobias.
EL USO DE LOS NITROIMIDAZOLES ESTÁ
PROHIBIDO EN ANIMALES PRODUCTORES DE ALIMENTOS POR SU POTENCIAL CARCINOGÉNICO
En los países donde esta aceptado su uso se emplean frente a disentería porcina e infecciones por Trichomonas foetus.
MECANISMO DE ACCIÓN
MECANISMO DE ACCIÓN
Cuando entran en la bacteria los nitroimidazoles sufren una nitrorreducción en condiciones de anaerobiosis que, a diferencia de los nitrofuranos, no es controlada enzimáticamente.
Estos elementos son inestables y responsables de la actividad antibacteriana porque provocan roturas de las hebras de ADN y la inhibición de la enzima reparadora ADNasa I.
BIBLIOGRAFÍA
AEMPS. (2021) Agencia Española de Medicamentos y Productos Sanitarios. Guía de Prescripción Terapéutica: Información de medicamentos autorizados en España. http://www.aemps.gob.es.
AEMPS. Base de datos CIMAVET. Disponible en línea: https://cimavet.aemps.es/cimavet/publico/home.html
Botana, L.M. Farmacología Veterinaria. (2022). Fundamentos y aplicaciones terapéuticas. 2ª ed. Editorial Médica Panamericana.
Botana, L.M. Farmacología Veterinaria. Fundamentos y aplicaciones terapéuticas. 2ª ed. Editorial Médica Panamericana. 2022
Brunton,L.; Hilal-Dandan, R.; Knollmann, B.C.; Goodman, L.S. y Goodman Gilman, A. (2018). The pharmacologial basis of therapeutics.
Bush, N.; Diez-Santos, I.; Abbott, L.R. y Maxwell, A. (2020). Quinolones: Mechanism, Lethality and Their Contributions to Antibiotic Resistance.. Molecules, 25(23), 5662.
Calvo, J.; Martínez-Martínez, L. (2009). Mecanismos de acción de los antimicrobianos. Enfermedades Infecciosas y Microbiología Clínica, 27(1): 44–52.
EMA (European Medicines Agency) and ESVAC (European Surveillance of Veterinary Antimicrobial Consumption) (2018). Sales of veterinary antimicrobial agents.
European Commission, ECDC (European Center for Disease prevention and control) (2017). EU Guideline for the prudent use of antimicrobial in human health.
https://ec.europa.eu/health/amr/sites/amr/files/ amr_guidelines_prudent_use_en.pdf
https://www.ema.europa.eu/en/documents/report/ sales-veterinary-antimicrobial-agents-30-european-countries-2016-trends-2010-2016-eighth-esvac_en.pdf
in 30 European countries in 2016. (EMA/275982/2018).
Millanao, A.; Mora, A.; Villagra, N.; Bucarey, S. e Hidalgo, A. (2021). Biological Effects of Quinolones: A Family of Broad-Spectrum Antimicrobial Agents. Molecules. Nov 25;26(23):7153.
Ovung, A.; Bhattacharyya, J. (2021). Sulfonamide drugs: structure, antibacterial property, toxicity, and biophysical interactions. Biophysical Reviews, 13: 259-272.
Perez-Trallero, D.V. Tetraciclinas, sulfamidas y metronidazol. Enferm Infecc Microbiol Clin 2010; 28:122-30. 10.1016
CARACTERÍSTICAS FARMACOCINÉTICAS
CARACTERÍSTICAS FARMACOCINÉTICAS
Los nitroimidazoles son liposolubles y se absorben bien por vía oral y parenteral, con un amplio volumen de distribución y penetración tisular, incluyendo LCR.
Se oxidan en el hígado y se conjugan dando formas menos activas, excretándose 2/3 partes a través de la orina, la mayor parte de forma activa.
MECANISMOS DE RESISTENCIA
MECANISMOS DE RESISTENCIA
Los nitroimidazoles apenas presentan resistencias.
INTERACCIONES, EFECTOS SECUNDARIOS Y TOXICIDAD
INTERACCIONES, EFECTOS SECUNDARIOS Y TOXICIDAD
Debido a su potencial efecto carcinogénico en animales de laboratorio, no se deben usar los nitroimidazoles al principio de gestación. También se han observado neuropatías periféricas, ataques epileptiformes, orina oscura, náuseas y exantemas.
Riviere, J.E.; Papich, M.G. (2017) Journal of Veterinary Pharmacology and Therapeutics. 10th ed. Wiley- Blackwell.
Scott, H.M.; Acuff, G.; Bergeron, G.; Bourassa, M.W.; Gill, J.; Graham, D.W.; (2019) Critically important antibiotics: criteria and approaches for measuring and reducing their use in food animal agriculture. Annals of the New York Academy of Sciences, 2019, 1441: 8-16.
Silva J. M. B.; Hollembach, C.B. (2010) Fluoroquinolonas X resistência bacteriana na medicina veterinária. Arquivos do Instituto Biológico de São Paulo, 77(2):363-369.
Stahl, J.; Zessel, K.; Schulz, J.; Finke, J.H., Müller-Goymann, C.C.; Kietzmann M. (2016). The effect of miscellaneous oral dosage forms on the environmental pollution of sulfonamides in pig holdings. BMC Veterinary Research 12: 68.
Vicente, D.; Pérez-Trallero, E. (2010). Tetraciclinas, sulfamidas y metronidazol. Enfermedades Infecciosas y Microbiología Clínica, 28(2): 122-130.
Yang, F.; Liu, Y.; Li, Z.; Wang, Y.; Liu M.B.;, Zhao, Z.; Zhou, B. y Wang, G. (2017) Tissue distribution of marbofloxacin in pigs after a single intramuscular injection. J Vet Scim, Jun 30;18(2):169-173.
74 ANTIMICROBIANOS Nº 7 | Abril 2023 Uso racional de antimicrobianos en porcino: antimicrobianos inhibidores de la síntesis de ARN y ADN – Parte II
DETECCIÓN RÁPIDA DE BACTERIAS RESISTENTES TRAS EL TRATAMIENTO CON ENROFLOXACINA

La aparición de antibiorresistencias entre los microorganismos, tanto patógenos como comensales, es un problema que genera una creciente preocupación en todo el mundo, ya que este fenómeno es responsable de la pérdida de eficacia de los antibióticos frente a enfermedades infecciosas que habitualmente tenían cura.


Es por ello que llevar a cabo un control adecuado de la existencia de estos microorganismos, conociendo cuáles son e incluso qué sensibilidad presentan frente a los antibióticos, resulta de gran importancia de cara a frenar su expansión y gestionar de una forma más eficiente los eventuales brotes que puedan producirse en las explotaciones porcinas.



1Departamento de Tecnología de los Alimentos, Centro de Investigación CIVA, Universidad de La Rioja
2Instituto Agroalimentario de Aragón-IA2 (Universidad de Zaragoza-CITA)
*Contacto: mjserran@unizar.es





E
y Rafael Pagán2
75 ONE HEALTH
Descarga el PDF
La creciente preocupación sobre la generación de resistencias antibióticas existente en la actualidad, junto con las pautas de la Agencia Europea del Medicamento hacia una reducción en la administración de antibióticos, una mejor selección de la terapia antibiótica efectiva mediante la realización de antibiogramas y la categorización de los antibióticos para su administración en medicina humana/veterinaria (EMA, 2020), han sido responsables de una reducción ostensible de su consumo en ganadería (EFSA, 2021). Sin embargo, las antibiorresistencias todavía son responsables de un elevado número de decesos anualmente (O´Neill, 2014; Wall et al., 2016; WHO, 2019; WHO, 2021).
Teniendo en cuenta que la administración de antibióticos en producción animal es en muchos casos necesaria para mantener un adecuado estatus sanitario y un buen nivel productivo, encontrar un equilibrio entre ambos aspectos puede resultar difícil.
Un conocimiento más profundo de los efectos que los tratamientos terapéuticos con antibióticos tienen sobre poblaciones bacterianas, como la microbiota intestinal y genital de individuos tratados, podría resultar de gran ayuda en el momento de tomar la decisión de seleccionar el tratamiento más adecuado y gestionar de forma más eficiente las implicaciones posteriores del mismo.
Puesto que las resistencias a los antibióticos generadas en poblaciones bacterianas procedentes de animales de producción pueden transmitirse a humanos (Bergšpica et al., 2020; Fournier et al., 2021), microorganismos como Enterococcus spp. y Enterobacteriaceae (Dandachi et al., 2018; Unal et al., 2020) resultan de especial importancia, ya que la microbiota fecal es una de las vías de transmisión más común desde los animales a los humanos.
Entre estas bacterias, los enterococos resistentes a la vancomicina (VRE) y las enterobacterias productoras de betalactamasas de espectro extendido (ESBL) son consideradas una amenaza para la salud pública (Fournier et al., 2012; Adler et al., 2016; Madec et al., 2017; EFSA, 2021)

Nº 7 | Abril 2023 Detección rápida de bacterias resistentes tras el tratamiento con enrofloxacina 76 ANTIMICROBIANOS
ENTEROCOCOS RESISTENTES A LA VANCOMICINA (VRE)
Los enterococos forman parte de la microbiota de animales y humanos, siendo bacterias ubicuas que se pueden encontrar en agua, suelos y plantas (Novais et al., 2013), pudiendo diseminarse a través de heces, alimentos contaminados o por contacto directo con animales (Novais et al., 2013; Nilsson et al., 2012)
De este grupo, Enterococcus faecalis y Enterococcus faecium figuran entre las especies con un mayor impacto sobre la salud humana, y las resistencias bacterianas adquiridas por estos microorganismos son una de las primeras causas relacionadas con su morbilidad (Gastmeier et al., 2014), aparte de actuar como reservorio de genes de resistencia transferibles a otras bacterias.
Puesto que la vancomicina es uno de los antibióticos de elección para el tratamiento de infecciones causadas por enterococos multirresistentes en medicina humana, la presencia de VRE en animales es especialmente preocupante (Hammerum, 2012) y su uso en Medicina Veterinaria está prohibido de acuerdo a la categorización de la EMA (EMA, 2020).
ENTEROBACTERIAS PRODUCTORAS DE BETALACTAMASAS DE ESPECTRO EXTENDIDO (ESBL)
Las enterobacterias forman parte de la microbiota habitual del tracto gastrointestinal de animales y humanos, y uno de sus principales mecanismos de resistencia es a través de la producción de β-lactamasas (Doi et al., 2017).
Es por ello que los ESBL son de especial importancia, ya que son capaces de contrarrestar el efecto de la mayor parte de los β-lactámicos, a excepción de los carbapenems y cefamicinas (Tamta et al., 2020).
Los ESBL se han encontrado frecuentemente en animales de granja, entre ellos el porcino (Stefani et al., 2014), desde donde pueden pasar a humanos.
Además, los genes de resistencia suelen localizarse en plásmidos (Cantas et al., 2015), por lo que puede producirse transferencia horizontal de este mecanismo de resistencia.
Puesto que el grado de resistencias a antibióticos detectado en animales de producción se ha relacionado con la administración de antibióticos, se puede hablar de la existencia de una presión positiva hacia la selección de bacterias resistentes (Tan et al., 2018; Aasmäe et al., 2019).
77 ONE HEALTH
La enrofloxacina es una fluoroquinolona utilizada para el tratamiento de un amplio rango de enfermedades en Medicina Veterinaria, pasando por infecciones respiratorias y gastrointestinales en cerdos (De Smet et al., 2020) y, puesto que podría afectar de forma directa a la microbiota comensal, podría ser responsable de la selección de bacterias resistentes (De Smet et al., 2020)
Es por ello que el objetivo de este trabajo fue evaluar el impacto del tratamiento con enrofloxacina vía intramuscular en cerdos sobre la aparición de VRE y ESBL en mucosa genital y rectal.
NUEVOS AVANCES EN IDENTIFICACIÓN MICROBIANA
Actualmente, existe una gran variedad de métodos para la identificación microbiana, desde las tradicionales galerías de identificación bioquímica hasta los más novedosos métodos de identificación cromatográfica.
Entre este abanico de métodos, los sistemas automatizados de identificación microbiana*, destacan por su versatilidad y la posibilidad de llevar a cabo no solo la identificación sino también el estudio de la susceptibilidad antibiótica de un amplio abanico de microorganismos de una forma precisa, barata y rápida.

La rapidez en el diagnóstico del agente causal de una infección no solo reduce la morbilidad y mortalidad, sino que puede ser clave a la hora de frenar la propagación de la infección.
En este sentido, este tipo de equipos son capaces de ofrecer resultados en un periodo de 2-7 horas (dependiendo del microorganismo y la prestación, es decir, identificación o estudio de la sensibilidad antibiótica) frente a las 15-24 horas requeridas por los métodos tradicionales.
Tanto para llevar a cabo la identificación como la evaluación de la sensibilidad antibiótica, estos equipos realizan diversas pruebas bioquímicas reunidas en tarjetas de identificación que se inoculan de forma sencilla y estandarizada con un cultivo procedente de la muestra a identificar.
LAS TARJETAS DE IDENTIFICACIÓN SON
ECONÓMICAS, FÁCILES DE USAR Y GENERAN POCOS RESIDUOS EN COMPARACIÓN CON OTROS
MÉTODOS, ADEMÁS DE MINIMIZAR SITUACIONES DE CONTAMINACIÓN CRUZADA CON AEROSOLES, SALPICADURAS, VERTIDOS O CONTACTO
Lyon, Francia)
*Vitek®2 Compact (BioMérieux, Marcy-l'Étoile,
Nº 7 | Abril 2023 Detección rápida de bacterias resistentes tras el tratamiento con enrofloxacina 78 ANTIMICROBIANOS
Una vez obtenidos los resultados, el equipo conecta de forma automática con las bases de datos que tiene asociadas para dar el resultado requerido, que se recibe de forma inmediata tras finalizar la identificación.
Se trata de un equipo automatizado, eficiente e innovador, de fácil manejo y de gran utilidad en su campo, ya que, además de todas estas características, mejora el flujo de trabajo al reducir los tiempos de espera para la identificación microbiana o la evaluación de la resistencia antibiótica de los microorganismos deseados.

Puesto que es capaz de identificar una amplia gama de microorganismos (bacterias gram-positivas, gram-negativas, levaduras y otros), ofreciendo un alto grado de discriminación entre especies y bajas tasas de resultados erróneos o no concluyentes, se trata de un sistema muy adecuado para su propósito. De hecho, está siento ampliamente utilizado en:
Laboratorios de diagnóstico clínico, del sector agroalimentario, ambientales y otros para la identificación microbiana (Lim et al., 2015; Matajira et al., 2020; Morka et al., 2018)
La evaluación de la sensibilidad antibiótica (Founou et al., 2019, Gundran et al., 2020)
La gestión de infecciones bacterianas (Taipa et al., 2008)
La gestión de resistencias antimicrobianas en diversos ámbitos (Gundran et al., 2020; Mittal et al., 2018)
La detección de resistencias específicas (Dégi et al., 2019; Hwang et al., 2009)
El estudio de infecciones fúngicas (Chun et al., 2019)
IMAGEN 1
Sistema automatizado de identificación microbiana (Vitek®2 Compact, BioMérieux, Marcy-l'Étoile, Lyon, Francia), situado en los laboratorios del grupo de investigación Zoonosis Bacterianas: Brucelosis, Salmonelosis y Estreptocócicas (ZooBac).
79 ONE HEALTH
EL ESTUDIO PERMITIÓ PERFILAR LA IMPLICACIÓN
DEL TRATAMIENTO
INTRAMUSCULAR CON ENROFLOXACINA EN LA SELECCIÓN DE VRE Y
ESBL
IMAGEN 2
Tarjetas para la identificación bioquímica de microorganismos (Fuente: Biomérieux).

EVALUACIÓN DEL IMPACTO DE LA ENROFLOXACINA SOBRE LA MICROBIOTA PORCINA
A pesar de que son muchas las aplicaciones que los sistemas automatizados de identificación microbiana pueden tener, puesto que existe una preocupación creciente y generalizada por el incremento en las poblaciones de bacterias resistentes en animales de producción y su potencial transmisión a humanos, en el Instituto Agroalimentario de Aragón (IA2) en colaboración con la Universidad de la Rioja se planteó un estudio en el que se evaluase el impacto del tratamiento con enrofloxacina sobre la generación de resistencias, tanto en la microbiota intestinal como en la de la mucosa genital de un grupo de cerdos.

En este sentido, en muchas ocasiones, la generación de este tipo de bacterias viene ligada a la presión selectiva ejercida por los tratamientos antibióticos y conocer su impacto podría ser de gran ayuda a la hora gestionar de forma más adecuada su uso.
Para ello, se planteó un estudio en el que 12 cerdos recibieron una pauta completa de enrofloxacina vía intramuscular y, tras finalizar el tratamiento, se fueron tomando muestras de la mucosa rectal y genital a intervalos predeterminados dentro del periodo de supresión para llevar a cabo la identificación de microorganismos comúnmente utilizados en el estudio del efecto de los tratamientos antibióticos sobre la generación de antibiorresistencias en producción porcina:
Enterococos resistentes a la vancomicina (VRE)
Poblaciones productoras de betalactamasas de espectro extendido (ESBL)
Nº 7 | Abril 2023 Detección rápida de bacterias resistentes tras el tratamiento con enrofloxacina 80 ANTIMICROBIANOS
DISEÑO EXPERIMENTAL
En conjunto, 12 cerdos se sometieron a una pauta completa de tratamiento con enrofloxacina vía intramuscular y 4 cerdos se mantuvieron sin tratar y en condiciones libres de antibióticos como controles negativos
Una vez finalizado el tratamiento, se tomaron muestras de la mucosa genital y del recto, utilizando un hisopo estéril, a lo largo de todo el periodo de supresión.
En total, se recogieron 42 muestras de mucosa genital y 42 de mucosa rectal procedentes de cerdos tratados, así como muestras de cada mucosa procedentes de los animales control.
La Tabla 1 muestra los días en los que se tomaron muestras tras la finalización del tratamiento.
*Los códigos de muestras que comienzan por C proceden de cerdos control, no tratados y mantenidos en condiciones libres de antibióticos. Los códigos que comienzan por E proceden de animales tratados con enrofloxacina.
TABLA 1
Tipo de tratamiento Día Post-tratamiento Código muestra* Control 0 C0 14 C14 Enrofloxacina 0 E0 3 E3 4 E4 5 E5 6 E6 7 E7
Número de animales muestreados cada día, tanto control como tratados con enrofloxacina y muestreados a lo largo del periodo de supresión.
81 ONE HEALTH
AISLAMIENTO E IDENTIFICACIÓN BACTERIANA
De cara al aislamiento bacteriano, se utilizaron CHROMID® VRE y CHROMID® ESBL (BioMérieux, Marcy l’Etoile, France), que se sembraron e incubaron a 37°C durante 24h en condiciones de aerobiosis para la selección de E. faecium y E. faecalis resistentes a vancomicina y enterobacterias productoras de ß-lactamasas, respectivamente.
Las colonias sospechosas se confirmaron mediante el sistema automatizado de identificación microbiana. Todos estos métodos son fiables para la identificación de E. faecalis, E. faecium, E. coli y K. pneumoniae (Fang et al., 2012; Bilecen et al., 2015)
PARA CALCULAR EL PORCENTAJE DE ANIMALES CON MICROBIOTA RESISTENTE, SE CONSIDERARON POSITIVOS LOS ANIMALES
QUE ALBERGABAN AL MENOS UN TIPO DE MICROORGANISMO RESISTENTE
RESULTADOS
ENTEROCOCOS RESISTENTES A LA VANCOMICINA (VRE)
ENTEROCOCOS RESISTENTES A LA VANCOMICINA (VRE)
La Gráfica 1 muestra el porcentaje de animales con aislados VRE en muestras de mucosa rectal.
GRUPO CONTROL
En el grupo control, que no recibió ningún tratamiento antibiótico y se crio en condiciones libres de antibióticos, se detectó E. faecium en el 75% de los animales en los días 0 y 14, y E. faecalis en el 50% y 25%, respectivamente (C0, C14).
GRUPO EXPERIMENTAL
En los cerdos tratados con enrofloxacina, E. faecium resistente a la vancomicina (VRE) se detectó en el 83% de los animales en los días 0 y 7 y en el 100% en el día 4, mientras que E. faecalis se detectó únicamente en el 25% de los animales en el día 0.
Nº 7 | Abril 2023 Detección rápida de bacterias resistentes tras el tratamiento con enrofloxacina 82 ANTIMICROBIANOS
GRÁFICA 1
Efecto del tratamiento con enrofloxacina sobre enterococos resistentes a la vancomicina (VRE) en muestras de mucosa rectal. Para encontrar la descripción de los grupos, ir a TABLA 1: C0, C14, E0, E3, E4, E5, E6, E7.
La Gráfica 2 muestra el porcentaje de animales en los que se detectaron VRE en muestras de mucosa genital de animales tratados intramuscularmente con enrofloxacina.
E. faecium se detectó en el 17-50% de los animales en función del día de muestreo.
Únicamente se detectó E. faecalis en el día 3 en el 17% de los animales.
GRÁFICA 2
Efecto del tratamiento con enrofloxacina sobre enterococos resistentes a la vancomicina (VRE) en muestras de mucosa genital. Para encontrar la descripción de los grupos, ir a TABLA 1: C0, C14, E0, E3, E4, E5, E6, E7.

100% 90% 80% 70% 60% 50% 40% 30% 20% 10% 0% 100% 90% 80% 70% 60% 50% 40% 30% 20% 10% 0%
E. faecalis
E. faecalis
faecium Porcentaje de animales positivos Porcentaje de animales positivos C0 C0 C14 C14 E0 E0 E3 E3 E4 E4 E5 E5 E6 E6 E7 E7 83 ONE HEALTH
E. faecium
E.
POBLACIONES PRODUCTORAS DE BETALACTAMASAS DE ESPECTRO EXTENDIDO (ESBL)
POBLACIONES PRODUCTORAS DE BETALACTAMASAS DE ESPECTRO EXTENDIDO (ESBL)
La Gráfica 3 muestra el porcentaje de animales en que se detectaron E. coli y K. pneumoniae productoras de ß-lactamasas (ESBL) en mucosa rectal.
E. coli ESBL
Mientras que en los animales no tratados no se detectó E. coli ESBL, sí se detectó en el 25% de los tratados en el día 0, en el 17% en los días 3 y 4 y en el 33 y 50% en los días 5 y 7, respectivamente.
K. pneumoniae ESBL
K. pneumoniae ESBL se detectó en los animales control no tratados, en los días 0 y 14, en un 25 y 50% de los animales, y en un 17% de los animales tratados en los días 4, 6 y 7, respectivamente.
GRÁFICA 3
Efecto del tratamiento con enrofloxacina sobre poblaciones de E. coli y K. pneumoniae de mucosa rectal de cerdo. Para la descripción de los grupos, ver TABLA 1: C0, C14, E0, E3, E4, E5, E6, E7.
La Gráfica 4 muestra el porcentaje de animales en los que se aislaron E. coli y K. pneumoniae ESBL de mucosa genital.
E. coli ESBL

E. coli ESBL no se detectó en el grupo control, mientras que sí se encontró en el 17% y 33% de la mucosa genital de los cerdos tratados en los días 5 y 6, respectivamente.
K. pneumoniae ESBL
K. pneumoniae ESBL únicamente se detectó en mucosa genital del grupo control el día 14.
100% 90% 80% 70% 60% 50% 40% 30% 20% 10% 0% E. coli Klebsiella pneumoniae Porcentaje de animales positivos C0 C14 E0 E3 E4 E5 E6 E7 Nº 7 | Abril 2023 Detección rápida de bacterias resistentes tras el tratamiento con enrofloxacina 84 ANTIMICROBIANOS
GRÁFICA 4
Efecto del tratamiento con enrofloxacina sobre poblaciones de E. coli y K. pneumoniae de mucosa genital de cerdo. Para la descripción de los grupos, ver TABLA 1: C0, C14, E0, E3, E4, E5, E6, E7.
DISCUSIÓN
Algunos estudios llevados a cabo en cerdos muestran que los aislados de Enterococcus spp. son sensibles a la vancomicina (Tan et al., 2018), mientras que otros autores han demostrado la presencia de VRE en cerdos.
Por ejemplo, Aarestrup et al. (2000) observaron que el 17% de los aislados de E. faecium en porcino de Dinamarca eran VRE.
Este hecho se ha asociado al ya prohibido uso de avoparcina como promotor del crecimiento (Lauderdale et al., 2007), puesto que es un análogo a la vancomicina capaz de generar resistencias cruzadas.
100% 90% 80% 70% 60% 50% 40% 30% 20% 10% 0%
E. coli
Porcentaje de animales positivos
Klebsiella pneumoniae
85 ONE HEALTH
C0 C14 E0 E3 E4 E5 E6 E7
ENTEROCOCOS RESISTENTES A LA VANCOMICINA (VRE)
En el presente estudio, se detectaron enterococos resistentes a la vancomicina, tanto en cerdos no tratados como tratados.
En conjunto, E. faecium fue más prevalente que E. faecalis, lo que podría deberse a una mayor prevalencia general de E. faecium en cerdos (Aasmäe et al., 2019), siendo común encontrar E. faecium VRE (Wist et al., 2020).
Se observó un menor porcentaje de VRE en muestras de mucosa genital, lo que podría explicarse por el hecho de que los enterococos forman parte de la microbiota normal de los cerdos (Hammerum, 2012)
LOS DATOS
ENTEROBACTERIAS PRODUCTORAS DE BETALACTAMASAS DE ESPECTRO EXTENDIDO (ESBL)
Las enterobacterias productoras de ß-lactamasas (ESBL), específicamente E. coli ESBL, son comúnmente detectadas en porcino en todo el mundo (Escudero et al., 2012; Ewers et al., 2012; Bergšpica et al., 2020; Fournier et al., 2021) en porcentajes mayores en granjas en las que se administran antibióticos de forma más frecuente (De Koster et al., 2021; Benavides et al., 2021), lo que sugiere un fenómeno de presión selectiva.
No obstante, cabe señalar que incluso en granjas con un bajo uso de antibióticos, se detecta E. coli ESBL en gran parte de los animales (Fournier et al., 2021), pudiendo estas poblaciones presentar resistencia cruzada frente a otros antibióticos, incluida enrofloxacina (Picozzi et al., 2014) y multirresistencias (Liu et al., 2018)
Puesto que E. coli es una bacteria típicamente intestinal, se encontraron más animales positivos en mucosa rectal en comparación con genital (Dandachi et al., 2018)
Hay pocos estudios sobre K. pneumoniae ESBL, y es que sus recuentos son mucho menores en comparación con E. coli (De Koster et al., 2021).
Sin embargo, se ha detectado en carne (Ojer-Usoz, 2013) y cerdo (Leangapichart et al., 2021)
INTRAMUSCULAR
A pesar de que la enrofloxacina se excreta principalmente por orina, también se han detectado elevadas concentraciones en heces de cerdos tras su administración intramuscular (Zhou et al., 2008) ya que también se metaboliza a nivel hepático (Giguère et al., 2013) y puede ser excretada a nivel intestinal (Álvarez et al., 2008)
Esto afecta a la microbiota intestinal, especialmente a microorganismos como E. coli, frente al que la enrofloxacina es muy efectiva (Edlund & Nord, 1999):
Ejerciendo presión hacia la selección de bacterias resistentes (Erwin et al., 2020)
Inactivando cepas salvajes a nivel intestinal (Béraud et al., 2008; De Smet et al., 2020)
PRESENTADOS DEMUESTRAN LA PRESENCIA DE CEPAS RESISTENTES EN LOS ANIMALES, ESPECIALMENTE EN MUCOSA RECTAL, TRAS EL TRATAMIENTO CON ENROFLOXACINA VÍA
Nº 7 | Abril 2023 Detección rápida de bacterias resistentes tras el tratamiento con enrofloxacina 86 ANTIMICROBIANOS
La microbiota de animales no tratados también podría verse afectada debido a la transferencia ambiental de estas cepas.
Es por ello que se debe tener especial cuidado durante fases críticas del proceso productivo, tales como el faenado tras el sacrificio, para evitar la contaminación de las canales por poblaciones resistentes de microorganismos de la familia Enterobacteriaceae (enterobacterias) y Enterococcaceae (enterococos) que, por otra parte, suelen ser bacterias multirresistentes (Doi et al., 2017).
La contaminación cruzada de la carne con poblaciones resistentes de origen fecal supone un importante riesgo sanitario (Wiuff et al, 2002; Ramos et al., 2012), ya que contribuye a agravar el ya preocupante fenómeno de resistencias antibióticas.
CONCLUSIONES
Este estudio demuestra que la administración intramuscular de enrofloxacina puede seleccionar E. coli productoras de ß-lactamasas (ESBL) en el tracto intestinal y, en menor medida, a nivel genital, así como las poblaciones de E. faecium resistentes a la vancomicina (VRE), por lo que se debe tener especial cuidado para evitar la contaminación de las canales tras el sacrificio.
El sistema automatizado de identificación microbiana** ha mostrado ser un método de detección fiable, rápido y efectivo para identificar bacterias resistentes a antibióticos, aunque sus posibilidades son mucho amplias.
**El VITEK®2 utilizado en esta investigación es propiedad del Instituto Agroalimentario de Aragón IA2 – Universidad de Zaragoza y forma parte del “Servicio de Identificación y Evaluación de la Sensibilidad Antibiótica” que ofrece dentro de su cartera de servicios a entidades, empresas y centros de investigación públicos y privados.

El estudio ha sido cofinanciado al 65% por el Fondo Europeo de Desarrollo Regional (FEDER) a través del Programa Interreg V-A España-FranciaAndorra (POCTEFA 2014-2020).
Para más información, consultar la siguiente referencia: GonzálezFandos, E., Martínez-Laorden, A., Abad-Fau, A., Sevilla, E., Bolea, R., Serrano, M. J., & Pagán, R. (2022). Effect of intramuscularly administered oxytetracycline or enrofloxacin on vancomycinresistant enterococci, extended spectrum beta-lactamase-and carbapenemase-producing Enterobacteriaceae in pigs. Animals, 12(5), 622.
87 ONE HEALTH
REFERENCIAS
Aarestrup, F.M.; Agerso. Y.; Gerner-Smidt. P.; Madsen. M.; Jensen, L.B. Comparison of antimicrobial resistance phenotypes and resistance genes in Enterococcus faecalis and Enterococcus faecium from humans in the community, broilers, and pigs in Denmark. Diagn. Microbiol. Infect. Dis. 2000, 37, 127-137.
Aasmäe, B.; Häkkinen, L.; Kaartt, T.; Kalmus, P. Antimicrobial resistance of Escherichia coli and Enterococcus spp. isolated from Estonian cattle and swine from 2010 to 2015. Acta Vet. Scand. 2019, 61,5.
Adler, A.; Katz, D.E.; Marchaim, D. The continuing plague of extended-spectrum beta-lactamase-producing Enterobacteriaceae infections. Infect. Dis. Clin. North Am. 2016, 30, 347-375.
Álvarez A.; Pérez M.; Prieto J.; Molina A.; Real R, Merino, G. Fluoroquinolone efflux mediated by ABC transporters. J Pharm Sci. 2008, 97, 3483–3493.
Benavides, J.A.; Streicker, D.G.; Gonzales, M.S.; Rojas-Paniagua, E.; Shiva, C. Knowledge and use of antibiotics among low-income small-scale farmers of Peru. Prev. Vet. Med. 2021b, 189, 105287.
Béraud, R.; Huneault, L.; Bernier, D.; Beaudry, F.; Letellier, A.; del Castillo, J.R.E. Comparison of the selection of antimicrobial resistance in fecal Escherichia coli during enrofloxacin administration with a local drug delivery system or with intramuscular injections in a swine model. Can. J. Vet. Res., 2008, 72, 311-319.
Bergšpica, I.; Kaprou, G.; Alexa, E.A.; Prieto, M.; Alvarez-Ordóñez, A. Extended spectrum β-lactamase (ESBL) Escherichia coli in pigs and pork meat in European Union. Antibiotics 2020, 9, 678.
Bilecen, K.; Yaman, I.G.; Ciftci,U.; Laleli, Y.R. Performances and reliability of Bruker Microflex LT and VITEK MS MALDI-TOF mass spectrometry systems for the identification of clinical microorganisms BioMed Res. Int., 2015, 516410.
Cantas, L.; Suer, K.; Guler, E.; Imir, T. High emergence of ESBL producing E. coli cystitis: time to get smarter in Cyprus. Front. Microbiol. 2015, 6, 1446.
Chun, L. Y., Dolle-Molle, L., Bethel, C., Dimitroyannis, R. C., Williams, B. L., Schechet, S. A., ... & Skondra, D. (2019). Rapid pathogen identification and antimicrobial susceptibility testing in in vitro endophthalmitis with matrix assisted laser desorption-ionization Time-of-Flight Mass Spectrometry and VITEK 2 without prior culture. PLoS One, 14(12), e0227071.
Dandachi, I.; Chabou, S.; Daoud, Z.; Rolain, J.M. Prevalence and emergence of extended-spectrum cephalosporin, carbapenem and colistin-resistant Gram negative bacteria of animal origin in the Mediterranean basin. Front. Microbiol. 2018, 9, 2299.
De Koster, S.; Ringenier, M.; Lammens, C.; Stegeman, A.; Tobias, T.; Velkers, F.; Vernooij, H.; Kluytmans-Van Den Bergh, M.; Kluytmans, J.; Dewulf, J.; Goossens, H. ESBL-producing, carbapenem and ciprofloxacin-resistant Escherichia coli in belgian and dutch broiler and pig farms: A cross-sectional and cross-border study. Antibiotics, 2021, 10. 945.
De Smet, J.; Boyen, F.; Croubels, S.; Rasschaert, G.; Haesebrouck, F.; Temmerman, R.; Rutjens, S.; De Backer, P.; Devreese, M. The impact of therapeutic-dose induced intestinal enrofloxacin concentrations in healthy pigs on fecal Escherichia coli populations. BMC Vet. Res. 2020, 16, 382.
Dégi, J., Cristina, R. T., Dégi, D. M., Muselin, F., Dumitrescu, C., & Iancu, I. (2019). Research of methicillin resistance staphylococci in a pig’s farm. Lucrări Ştiinţifice, 27.
Doi, Y.; Iovleva, A.; Bonomo, R.A. The ecology of extended-spectrum B-lactamases (ESBLs) in the developed world. J. Travel Med. 2017, 24, S44-S51.
Edlund, C.; Nord, C.E. Effect of quinolones on intestinal ecology. Drugs 1999, 58, 65-70.
EFSA, 2021. European Centre for Disease Prevention and Control (ECDC), European Food Safety Authority (EFSA), & European Medicines Agency (EMA). (2021). Third joint inter-agency report on integrated analysis of consumption of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from humans and food-producing animals in the EU/EEA: JIACRA III 2016-2018. EFSA Journal, 19(6), e06712.
EMA (European Medicine Agency). Categorisation of antibiotics for use in animals for prudent and responsible use. 2020 Available online: https://www.ema.europa.eu/en/ documents/report/infographic-categorisation-antibiotics-use-animals-prudent-responsible-use_en.pdf (accesed 27 April 2021)
Erwin, S.; Foster D.M.; Jacob, M.E.; Papich, M.G.; Lanzas, C. The effect of enrofloxacin on enteric Escherichia coli: Fitting a mathematical model to in vivo data. PLoS ONE 2020, 15 e0228138.
Escudero, E.; Vinue, L.; Teshager, T.; Torres, C.; Moreno, M.A. Resistance mechanisms and farm-level distribution of fecal Escherichia coli isolates resistant to extended-spectrum cephalosporins in pigs in Spain. Res. Vet. Sci. 2010, 88, 83-87.
European Food Safety Authority and European Centre for Disease Prevention and Control).
The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2018/2019. EFSA J. 2021, 19, e06490.
Ewers, C.; Bethe, A.; Semmler, T.; Guenther, S.; Wieler, L. H.. Extended spectrum beta-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: a global perspective. Clin. Microbiol. Infect. 2012, 18, 646-655.
Fang, H.; Ohlsson, A.K.; Ullberg, M.; Özenci, V. Evaluation of species-specific PCR, Bruker MS, VITEK MS and the VITEK 2 system for the identification of clinical Enterococcus isolates Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 3073-3077.
Founou, L. L., Founou, R. C., Ntshobeni, N., Govinden, U., Bester, L. A., Chenia, H. Y., ... & Essack, S. Y. (2019). Emergence and spread of extended spectrum β-lactamase producing Enterobacteriaceae (ESBL-PE) in pigs and exposed workers: a Multicentre Comparative Study between Cameroon and South Africa. Pathogens, 8(1), 10.
Fournier, C.; Nordmann, P.; Pittet, O.; Poirel, L. Does an antibiotic stewardship applied in a pig farm lead to low ESBL prevalence? Antibiotics, 2021, 10, 574.
Fournier, S.; Brun-Buisson, C.; Jarlier, V. Twenty years of antimicrobial resistance control programme in a regional multi hospital institution, with focus on emerging bacteria (VRE and CPE). Antimicrob. Resist. Infect. Control 2012, 1, 9.
Gastmeier, P.; Schröder, C.; Behnke, M.; Meyer, E.; Geffers, C. Dramatic increase in vancomycin-resistant enterococci in Germany. J. Antimicrob. Chemother. 2014, 69, 1660-1664.
Giguère S.; Prescott J.; Dowling, P. Antimicrobial therapy in veterinary medicine, 5th ed; John Wiley & Sons, New Jersey, USA, 2013.
Gundran, R. S., Cardenio, P. A., Salvador, R. T., Sison, F. B., Benigno, C. C., Kreausukon, K., ... & Punyapornwithaya, V. (2020). Prevalence, Antibiogram, and Resistance Profile of Extended-Spectrum β-Lactamase-Producing Escherichia coli Isolates from Pig Farms in Luzon, Philippines. Microbial Drug Resistance, 26(2), 160-168.
88 ANTIMICROBIANOS Nº 7 | Abril 2023 Detección rápida de bacterias resistentes a enrofloxacina en cerdos
Hammerum, A.M. Enterococci of animal origin and their significance for public health. Clin. Microbiol. Infect. 2012, 18, 619-625.
Hwang, I. Y., Ku, H. O., Lim, S. K., Park, C. K., Jung, G. S., Jung, S. C., & Nam, H. M. (2009). Species distribution and resistance patterns to growth-promoting antimicrobials of enterococci isolated from pigs and chickens in Korea. Journal of Veterinary Diagnostic Investigation, 21(6), 858-862.
Lauderdale, T.L.; Shiau, Y.R.; Wang, H.Y.; Lai, J.F.; Huang, I.W.; Chen, P.C.; Chen, H.Y.; Lai, S.S.; Liu, Y.F.; Ho., M. Effect of banning vancomycin analogue avoparcin on vancomycin-resistant enterococci in chicken farms in Taiwan. Environ. Microbiol. 2007, 9, 819-823.
Leangapichart, T.; Lunha, K.; Jiwakanon, J.; Angkititrakul, S.; Järhult, J.D.; Magnusson, U.; Sunde, M. Characterization of Klebsiella pneumoniae complex isolates from pigs and humans in farms in Thailand: Population genomic structure, antibiotic resistance and virulence genes. J. Antimicrob. Chemoth. 2021, 76, 2012-2016.
Lim, J. S., Han, D. W., Lee, S. R., Hwang, O. H., Kwag, J. H., & Cho, S. B. (2015). A Bacterial Strain Identified as Bacillus licheniformis using Vitek 2 Effectively Reduced NH 3 Emission from Swine Manure. Journal of Animal Environmental Science, 21(3), 83-92.
Liu, X.; Liu, H.; Wang, L.; Peng, Q.; Li, Y.; Zhou, H.; Li, Q. Molecular characterization of extended-spectrum β-lactamase-producing multidrug resistant Escherichia coli from swine in Northwest China. Front. Microbiol. 2018, 9, 1756
Madec, J.Y.; Haenni, M.; Nordmann, P.; Poirel, L. Extended-spectrum beta-lactamase / AmpC- and carbapenemase-producing Enterobacteriaceae in animals: a threat for humans? Clin. Microbiol. Infect. 2017, 23, 826-833.
Matajira, C. E. C., Poor, A. P., Moreno, L. Z., Monteiro, M. S., Dalmutt, A. C., Gomes, V. T. M., ... & Moreno, A. M. (2020). Vagococcus sp. a porcine pathogen: molecular and phenotypic characterization of strains isolated from diseased pigs in Brazil. The Journal of Infection in Developing Countries, 14(11), 1314-1319.
Mittal, D., Grakh, K., Prakash, A., Moudgil, P., Devi, B., & Jadhav, V. (2018). Isolation, Identification and Characterization of Enteric Bacteria from Post Weaning Diarrheic Pigs and their Resistance to Multiple Antibiotics. International Journal of Current and Applied Microbiology, 7, 2377-2384.
Morka, K., Bystroń, J., Bania, J., Korzeniowska-Kowal, A., Korzekwa, K., Guz-Regner, K., & Bugla-Płoskońska, G. (2018). Identification of Yersinia enterocolitica isolates from humans, pigs and wild boars by MALDI TOF MS. BMC Microbiology, 18(1), 1-10.
Nilsson, O. Vancomycin resistant enterococci in farm animals. Occurrence and importance. Infect. Ecology Epidemiol. 2012, 2, 16959.
Novais, C.; Freitas, A.R.; Silveira, E.; Antunes, P.; Silva, R.; Coque, T.M.; Peixe, L. Spread of multidrug-resistant Enterococcus to animals and humans: an underestimated role for the pig farm environment. J. Antimicrob. Chemother. 2013, 68, 2746-2754.
O’Neill, J. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations. London: Review on Antimicrobial Resistance; 2014. Available online: https://amr-review. org/sites/default/files/AMR%20Review%20 Paper%20-%20Tacklng%20a%20crisis%20 for%20the%20health%20and%20wealth%20 of%20nations_1.pdf (accessed on 2 November 2021).
Ojer-Usoz, E.; Gonzalez, D.; Vitas, A.I.; Leiva, J.; Garcia-Jalon, I.; Febles-Casquero, A.; Escolano, M.S. Prevalence of extended-spectrum beta-lactamase-producing Enterobacteriaceae in meat products sold in Navarra, Spain. Meat Sci. 2013, 93, 316-321.
Picozzi, S.C.M.; Casellato, S.; Rossini, M.; Paola, G.; Tejada, M.; Costa, E.; Carmignani L. Extended-spectrum beta-lactamase-positive Escherichia coli causing complicated upper urinary tract infection: Urologist should act in time. Urol. Annals. 2014, 6, 107-112.
Ramos, S.; Igrejas, G.; Capelo-Martinez, J.L.; Poeta, P. Antibiotic resistance and mechanisms implicated in fecal enterococci recovered from pigs, cattle and sheep in a Portuguese slaughterhouse. Ann. Microbiol. 2012, 62, 1485-1494.
Stefani, S.; Giovanelli, I.; Anacarso, I.; Condo, C.; Messi, P.; de Niederhausern, S.; Bondi, M.; Iseppi, R.; Sabia, C. Prevalence and characterization of extended-spectrum beta-lactamase-producing Enterobacteriaceae in food-producing animals in Northern Italy. New Microbiol. 2014, 37, 551-555.
Taipa, R., Lopes, V., & Magalhães, M. (2008). Streptococcus suis meningitis: first case report from Portugal. Journal of Infection, 56(6), 482-483.
Tamta, S.; Vonodh-Kumar, O.R.; Pruthvishree, B.S.; Karthikeyan, R.; Ramkumar, R.R.; Chethan, G.E.; Dubal, Z.B.; Sinha, D.K.; Singh, B.R. Faecal carriage of extended spectrum beta-lactamase (ESBL) and New Delhi metallobeta-lactamase(NDM) producing Escherichia coli between piglets and pig farmworkers. Comparative Immunol. Microbiol. Infect. Dis. 2020, 73, 101564.
Tan, S.C.; Chong, C.W.; Teh, C.S.J.; Ooi, P.T.; Thong, K.L. Occurrence of virulent multidrug resistant Enterococcus faecalis and Enterococcus faecium in the pigs, farmers and farm environments in Malaysia. Peer J. 2018, 6, e5353.
Unal, N.; Bal, E.; Kapagoz, A.; ALtun, B.; Kozag, N. Detection of vancomycin-resistant enterococci in samples from broiler flocks and houses. Acta Vet. Hungarica 2020, 68,117-122.
Wall, B.A.; Mateus, A.; Marshall, L.; Pfeiffer, D.; Lubroth, J.; Ormel, H.J.; Otto, P.; Patriarchi, A. Food and Agriculture Organization of the United Nations. Drivers, dynamics and epidemiology of antimicrobial resistance in animal production. Food and Agriculture Organization (FAO): Rome, Italy, 2016.
Wist, V.; Morach, M.; Schneeberger, M.; Cernela, N.; Stevens, M.J.A.; Zurfluh, K.; Stephan, R.; Nüesch-Inderbinen, M. Phenotypic and genotypic traits of vancomycin-resistant enterococci from healthy food-producing animals. Microorganisms 2020, 8, 261.
Wiuff, C.; Lykkesfeldt, J.; Aarestrup, F.M.; Svendsen, O. Distribution of enrofloxacin in intestinal tissue and contents of healthy pigs after oral and intramuscular administrations. J. Vet. Pharmacol. Therap. 2002, 25, 335-342.
World Health Organization. Antibacterial agents in clinical development: an analysis of the antibacterial clinical development pipeline; World Health Organization, Geneva, Switzerland, 2019. Available online: http://www.who.int (accessed on 18 Nov 2021).
World Health Organization. Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report. World Health Organization, Geneva, Switzerland, 2021. Available online: http:// www.who.int (accessed on 18 Nov 2021).
Zhou, X.J.; Chen, C.X.; Yue, L.; Sun Y.X.; Ding, H.Z.; Liu, Y.H. Excretion of enrofloxacin in pigs and its effect on ecological environment. Environ. Toxicol. Phar. 2008, 26, 272–277.
89 ONE HEALTH
VACUNACIÓN DE CERDAS
HIPERPROLÍFICAS: ¿PODEMOS MEJORAR LA INMUNIZACIÓN DE SUS LECHONES
V
FRENTE A E. COLI?
Descarga el PDF
LLa diarrea neonatal en lechones se considera un problema mundial que conlleva problemas de bienestar animal y pérdidas para los productores. E. coli es un importante patógeno implicado en la aparición de esta enfermedad durante los períodos de lactación y el destete temprano.
Rut Menjón, Marta Jiménez y Marcial Marcos
Servicio Técnico
MSD Animal Health
Nº 7 | Abril 2023 VACUNACIÓN Vacunación de cerdas hiperprolíficas: ¿Podemos mejorar la inmunización de sus lechones frente a E. coli? 90
Durante el primer mes de vida del lechón, las enterobacterias son las principales colonizadoras del intestino delgado, por lo que será normal que siempre que tengamos estos procesos diarreicos se aísle algún Escherichia coli (E. coli).
Esto no quiere decir que sea el causante primario, ya que para ello debe tratarse de un E. coli enteropatógeno (EPEC) o enterotoxigénico (ETEC), es decir, debe poseer al menos un factor de adhesión y producir al menos una enterotoxina.
La diarrea neonatal que aparece normalmente a los 0-4 días de vida, entre otros patógenos, puede estar causada por E. coli enterotoxigénico (ETEC) que produce:
Tres tipos de enterotoxinas:
Las toxinas termoestables STa y STb
La toxina termolábil (LT)
Una o más de las fimbrias de adhesión:
F4 (K88)
F5 (K99)
F6 (987P)
F41
UNA CEPA DE E. COLI ETEC DEBERÁ SER CAPAZ DE ADHERIRSE
Y COLONIZAR LA MUCOSA INTESTINAL PARA LIBERAR NIVELES ADECUADOS DE ESTAS
ENTEROTOXINAS Y PRODUCIR EL PROCESO DIARREICO
E. coli con la fimbria K88 es el más común, existiendo tres variantes antigénicas:
K88ac, muy común, con una distribución mundial.
K88ab, menos frecuente, limitada a Europa.
K88ad, del que hay muy pocos aislamientos reportados.
La toxina LT está asociada principalmente a las cepas de E. coli productoras de la adhesina F4 (K88) y contribuye más a la virulencia que la ST.
IMPORTANCIA DE LA INMUNIDAD MATERNAL FRENTE A E. COLI
La adecuada inmunización de las cerdas es fundamental para proveer a los lechones de una protección efectiva a través del calostro. La mejor forma de conseguirlo es mediante la vacunación de las cerdas al final de la fase de gestación.
Para ello es importante elegir una vacuna que proporcione la máxima protección frente a esta patología, tomando como referencia el nivel de anticuerpos específicos producidos frente a los principales factores de virulencia.
V
INMUNIDAD 91
LA INMUNIDAD PASIVA TRANSFERIDA A TRAVÉS
DEL CALOSTRO A LOS LECHONES NEONATOS ES CLAVE PARA CONSEGUIR
QUE LOS ANTICUERPOS
ACTÚEN CONTROLANDO
Las IgG son capaces de bloquear la adherencia de E. coli a la mucosa intestinal y neutralizar su capacidad de producción de enterotoxinas, de forma que la patología no llegue a desarrollarse.
La ingesta del calostro de madres inmunizadas protege a los lechones del desarrollo de la diarrea, ya que, debido a la placenta epitelocorial de la cerda, la inmunidad inicial del lechón depende casi exclusivamente de los anticuerpos obtenidos de la madre vía calostro.
LAS CLAVES DE LA EFICACIA VACUNAL FRENTE A E. COLI
El éxito de una vacuna depende de dos factores:
La capacidad para inducir la producción del máximo nivel posible de anticuerpos frente a los factores de adhesión de E. coli (F) en la cerda, con el fin de que éstos impidan su unión al intestino del lechón y, por tanto, su multiplicación.
La capacidad de inducir la producción de anticuerpos frente a las enterotoxinas con el fin de prevenir la diarrea y la deshidratación.
Para conocer la inmunidad que se puede proporcionar a los lechones a través de la vacunación, se pueden realizar estudios y valorar la cantidad de inmunoglobulinas que se producen específicamente frente a cada toxina, y frente a las fimbrias o factores de adhesión.
Estas valoraciones son importantes para un eficaz control de esta patología, puesto que la cantidad de calostro producido por una cerda es especifica del individuo, e independiente del tamaño de camada, por lo que interesa partir del mayor nivel de inmunoglobulinas posible, de forma que la calidad del calostro sea la máxima para que todos los lechones estén por encima de los niveles de protección.
El nivel de anticuerpos obtenidos vía calostro depende de:
La cantidad de calostro que produce la hembra
La calidad del calostro producido
La cantidad del calostro ingerido
EL CALOSTRO ES FUNDAMENTAL PARA
ADQUIRIR UNA BUENA INMUNIDAD, PERO NO
TODO EL CALOSTRO ES IGUAL
Los lechones nacidos en las primeras horas pueden llegar a tomar un calostro un 50% más concentrado que los nacidos en último lugar. En el caso de cerdas hiperprolíficas esta diferencia puede verse incrementada debido a la mayor duración del parto. Por ello, conseguir un calostro con mayor cantidad de inmunoglobulinas permitirá que los lechones gocen de mayor protección.
1 2 VACUNACIÓN Vacunación de cerdas hiperprolíficas: ¿Podemos mejorar la inmunización de sus lechones frente a E. coli? 92
LA COLIBACILOSIS Nº 7 | Abril 2023
VALORACIÓN DEL NIVEL DE ANTICUERPOS
ALCANZADO EN CERDAS HIPERPROLÍFICAS CON DIFERENTES PROGRAMAS DE VACUNACIÓN FRENTE A LA DIARREA NEONATAL POR ETEC
Poster publicado en el ESPHM 2022
J. Grandia Anso1, M. Jiménez2, M. Marcos2 y R. Menjón2
1Piensos Costa
2MSD Animal Health
INTRODUCCIÓN
La diarrea neonatal asociada a E. coli ETEC es una de las patologías más comunes en las salas de parto, ocasionando importantes pérdidas económicas debido a su sintomatología y su elevada tasa de morbilidad.
LA ESTRATEGIA MÁS EFECTIVA PARA LA PREVENCIÓN ES LA INMUNIZACIÓN PASIVA CALOSTRAL
El objetivo de este estudio fue diferenciar entre distintos programas vacunales en reproductoras hiperprolíficas primíparas y de segundo parto, valorando los anticuerpos producidos para finalmente determinar el programa de prevención más adecuado.
MATERIALES & MÉTODOS
El estudio se llevó a cabo en una granja comercial del noreste de España, con genética hiperprolífica en la que se estudiaron dos vacunas comerciales (vacuna A, Porcilis® ColiClos y vacuna B) para determinar la concentración de anticuerpos presentes frente a los factores de virulencia de E. coli más prevalente en las diarreas neonatales (987P, K88ac, K88ab, K99 y LT), mediante un test ELISA ECO de MSD AH.
Se seleccionaron 75 cerdas (1er y 2o parto) que se repartieron en 5 grupos con dos programas de vacunación diferentes:
VACUNA A
Grupo 1 (cerdas de 1er parto)
Grupo 3 (cerdas de 2o parto)
Dos dosis: vacunación a las 6 y 4 semanas preparto
VACUNA A
Grupo 4 (cerdas de 2o parto)
Una dosis: Vacunación a las 4 semanas preparto
VACUNA B
Grupo 2 (cerdas de 1er parto)
Grupo 5 (cerdas de 2o parto)
Dos dosis: Vacunación a las 6 y 4 semanas preparto
INMUNIDAD 93
Se tomaron muestras de sangre a las 24h postparto, a los 7-10 días y al destete en los lechones (n=209), y en todas las cerdas en las semanas 6, 4 y 2 preparto, realizándose el estudio estadístico con el programa IBM SPSS Statistics 22.0.
*Se implementó la revacunación de 2 dosis para permitir la comparación en este estudio.
del programa vacunal en cada uno de los grupos de estudio.
RESULTADOS
En los lechones nacidos de cerdas de los grupos 1 y 3 (vacuna A) se detectó un mayor nivel de anticuerpos que en los lechones nacidos de cerdas de los grupos 2 y 5 (vacuna B) (p<0,001) en las mediciones al 1er día y a los 7-10 días de vida.
Asimismo, los títulos de anticuerpos en las cerdas a las 2 semanas preparto fueron mayores en los programas de dos dosis en su segundo parto (p< 0,05), siendo además más elevados estos títulos en todos los animales de los grupos de la vacuna A con respecto a la vacuna B (p<0,05).
Títulos de anticuerpos en primíparas para cada protocolo vacunal.
TABLA 1
Distribución
GRÁFICA 1
GRUPO PARIDAD VACUNA MOMENTO DE VACUNACIÓN (semanas preparto) 1ª dosis 2ª dosis 1 1ª Vacuna A (Porcilis® ColiClos) 6 4 2 1ª Vacuna B 6 4
2ª Vacuna A (Porcilis® ColiClos) 6 4 4 2ª Vacuna A (Porcilis® ColiClos) 45 2ª Vacuna B 6 4 12 10 8 6 4 2 0 F4ab Vacuna A (Porcilis® ColiClos) primíparas Vacuna B primíparas F4ac F5 F6 LT 1 er muestreo 2 o muestreo 3 er muestreo 1 er muestreo 2 o muestreo 3 er muestreo 1 er muestreo 2 o muestreo 3 er muestreo 1 er muestreo 2 o muestreo 3 er muestreo 1 er muestreo 2 o muestreo 3 er muestreo Nº 7 | Abril 2023 VACUNACIÓN Vacunación de cerdas hiperprolíficas: ¿Podemos mejorar la inmunización de sus lechones frente a E. coli? 94
3
a ,bDiferentes superíndices dentro de la misma fila representan diferencias estadísticas en los títulos de anticuerpos entre los diferentes programas de vacunación (p<0,001).
Títulos medios de anticuerpos en cerdas de 2o parto para cada protocolo vacunal.
 TABLA 2 Título medio de anticuerpos en primíparas.
TABLA 2 Título medio de anticuerpos en primíparas.
F4ab 1º 10,9a 7,33b 2º 9,35a 6,37b 3º 8.26 6,29 F4ac 1º 10,95a 8.97b 2º 9,26 8,49 3º 8,5 7,72 F5 1º 10,14a 8,86b 2º 7,95 7,88 3º 7,38 6,7 F6 1º 10,45a 8,4b 2º 8,61 7,2 3º 7,35 5,7 LT 1º 9,34a 8,54b 2º 7,44 7,55 3º 6,2 6,7
Factor virulencia Muestreo Vacuna A (Porcilis® ColiClos) Vacuna B
GRÁFICA 2
1 er muestreo 2 o muestreo 3 er muestreo 1 er muestreo 2 o muestreo 3 er muestreo 1 er muestreo 2 o muestreo 3 er muestreo 1 er muestreo 2 o muestreo 3 er muestreo 1 er muestreo 2 o muestreo 3 er muestreo 12 10 8 6 4 2 0 F4ab Vacuna A
2o parto 2 dosis Vacuna
2º
1
Vacuna B 2º
2 dosis F4ac F5 F6 LT INMUNIDAD 95
(Porcilis® ColiClos)
A (Porcilis® ColiClos)
parto
dosis
parto
a ,b, cDiferentes superíndices dentro de la misma fila representan diferencias estadísticas en los títulos de anticuerpos entre los diferentes programas de vacunación. (p<0,001)
DISCUSIÓN Y CONCLUSIONES
Disponer de un calostro con una elevada concentración de anticuerpos se hace indispensable hoy en día ante la gran cantidad de lechones en camadas hiperprolíficas.
Los resultados ponen de manifiesto la existencia de diferencias entre los programas vacunales utilizados, incrementándose el nivel de anticuerpos con programas de dos dosis, pudiendo esto mejorar el control de la diarrea neonatal en genéticas hiperprolíficas. Además, hubo mayor concentración de anticuerpos en los lechones que utilizan el programa de Porcilis® ColiClos.

Factores virulencia Muestreo Vacuna A (Porcilis® ColiClos) 2 dosis Vacuna A (Porcilis® ColiClos) 1 dosis Vacuna B 2 dosis F4ab 1º 10,83a 8,52b 9,21c 2º 9,76a 8,7b 7,9c 3º 8,8 7,79 7,68 F4ac 1º 11,9a 9,56b 10,2b 2º 10,6a 9,6ab 8,9b 3º 9,47 8,6 8,46 F5 1º 11,52a 8,45b 9,6b 2º 10,55 8,72 8,88 3º 8,57 8,01 6,9 F6 1º 10,37a 7,35b 7,23b 2º 9,15a 7,51a 6,24b 3º 7,4 6,7 6,44 LT 1º 8,7a 7,62b 7,3b 2º 7,9 7,5 6,3 3º 6,3 6,57 6,68
TABLA 3
Títulos medios de anticuerpos en cerdas de 2o parto.
Nº 7 | Abril 2023 VACUNACIÓN Vacunación de cerdas hiperprolíficas: ¿Podemos mejorar la inmunización de sus lechones frente a E. coli? 96
Prevención de la diarrea neonatal
AYUDANDO A LA DESCENDENCIA A SUPERAR LOS DESAFÍOS

Prevención de la diarrea neonatal
SUSPENSIÓN INYECTABLE PARA CERDOS.
COMPOSICIÓN POR DOSIS: Sustancias activas: Componentes de Escherichia coli: adhesina fímbrica F4ab ≥9,7 log2 título AC1, adhesina fímbrica F4ac ≥8,1 log2 título AC1, adhesina fímbrica F5 ≥8,4 log2 título AC1 adhesina fímbrica F6 ≥7,8 log2 título AC1, toxoide LT ≥10,9 log2 título AC1; componentes de Clostridium perfringens: tipo C (cepa 578) toxoide beta ≥20 UI2 Adyuvante: Acetato de dl-a-tocoferilo 150 mg. 1Título medio de anticuerpos (AC) obtenido tras la vacunación de ratones con 1/20 o 1/40 de la dosis para cerda. 2Unidades internacionales de antitoxina b de acuerdo a Farm. Eur. INDICACIONES Y ESPECIES DE DESTINO: Cerdas (adultas y nulíparas). Para la inmunización pasiva de la progenie mediante la inmunización activa de cerdas adultas y nulíparas, para reducir la mortalidad y síntomas clínicos durante los primeros días de vida, causados por cepas de E. coli, que expresan las adhesinas F4ab (K88ab), F4ac (K88ac), F5 (K99) o F6 (987P) y causados por C. perfringens tipo C. CONTRAINDICACIONES: Ninguna. PRECAUCIONES: Precauciones especiales para su uso en animales: Vacunar solamente animales sanos. La protección de los lechones se consigue mediante la ingesta del calostro. Por este motivo, deberán adoptarse precauciones para garantizar que cada lechón ingiera una cantidad de calostro suficiente. Precauciones específicas que debe tomar la persona que administre el medicamento a los animales: En caso de autoinyección accidental, consulte con un médico inmediatamente y muéstrele el prospecto o la etiqueta. Puede utilizarse durante la gestación. Conservar en nevera (entre 2 °C y 8 °C). No congelar. Proteger de la luz. Período de validez después de abierto el envase primario: 10 horas. TIEMPO DE ESPERA: Cero días. Uso veterinario – medicamento sujeto a prescripción veterinaria. Instrucciones completas en el prospecto. Mantener fuera de la vista y el alcance de los niños. Reg. Nº: EU/2/12/141/001-009. Merck Sharp & Dohme Animal Health, S.L. Ficha técnica actualizada a 10 de julio de 2017.

SUSCRÍBASE
El medio técnico-científico, innovador y exclusivo para veterinarios de porcino porcinews.com/revista-porcisapiens porciSapiens® agriNews®
GRATIS
