bardziej wydajny pod względem odzyskiwania energii w mitochondrialnym utlenianiu substratów. 2.6.5.
Jak nazywa się synteza ATP, która jest możliwa dzięki procesom pokazanym na rycinie 2.6a, b?
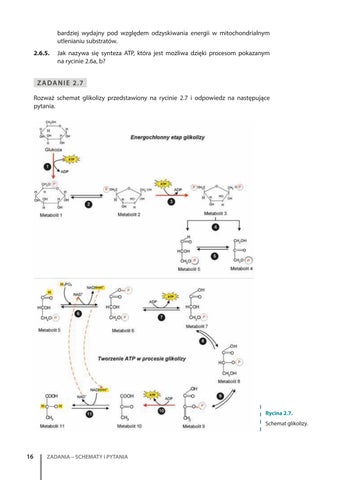
Z A D A N I E 2.7 Rozważ schemat glikolizy przedstawiony na rycinie 2.7 i odpowiedz na następujące pytania.
Rycina 2.7. Schemat glikolizy.
16
ZADANIA – SCHEMATY i PYTANIA