Ethologie
docenten ChristelMoons,WimVanhove
studiegebied biotechniek
bachelor in de agro-enbiotechnologie
campus Roeselare






4.3.1








4.2.1
4.2.2
4.2.3
4.3
4.4

docenten ChristelMoons,WimVanhove
studiegebied biotechniek
bachelor in de agro-enbiotechnologie
campus Roeselare
4.3.1
4.2.1
4.2.2
4.2.3
4.3
4.4
Deze cursus voor het academiejaar 2023-2024 is een gedeeltelijke bewerking van een eerdere versie, met dank aan de personen die daartoe bijdroegen: Prof. Em. Frank Ödberg (Universiteit Gent), meneer Wim Vanhove en Dr. Joke Monteny. Je vindt in deze tekst de te kennen leerstof. Aan het begin van elk hoofdstuk staan ook de leerdoelen vermeld.
Cursiefgedrukte tekst in deze syllabus is geen examenstof. Dit is extra informatie voor de geïnteresseerde student, maar hierover worden dus geen vragen gesteld op het examen. De vetgedrukte woorden doorheen de cursus (titels en subtitels even buiten beschouwing gelaten) zijn begrippen of personen die je dient te kennen. Voor de concepten kan een definitie worden gevraagd of er kan je gevraagd worden het concept toe te passen, bijvoorbeeld in een oefening of casus. Samengevat is dus alle niet-cursief gedrukte tekst examenstof.
In de lessen worden soms nog extra voorbeelden gegeven. Deze voorbeelden hoef je niet van buiten te kennen, maar je dient ze wel te begrijpen. Mocht er een vraag komen die verband houdt met één van die extra voorbeelden, zal je altijd genoeg uitleg over het voorbeeld in de vraagstelling meekrijgen om de vraag over een te kennen concept te beantwoorden.
Voor een aantal items in deze cursus is bepaalde kennis van de fysiologie nuttig. Aangezien Fysiologie I pas in het tweede semester aan bod komt, zal een relevante basis gegeven worden voor het begrijpen van de ethologische leerstof Hierbij is ook een woord van dank op zijn plaats voor mevrouw Lara Simons die toeliet enkele delen uit haar cursus over te nemen. Voor een meer doorgedreven uitleg over de zintuigen, het zenuwstelsel en hormonaal stelsel, wordt echter verwezen naar de volledige cursus Fysiologie I
De structuur van de cursus weerspiegelt de noodzaak om eerst inzicht te hebben in een aantal fundamentele begrippen en gedragsmechanismen, zodat later de soortspecifieke ethologie en welzijnsproblemen beter te kaderen en begrijpen zijn. De eerste hoofstukken van Deel 1 concentreren zich dan ook op het definiëren van de ethologie, het concept gedrag en op de verschillende benaderingen van gedrag via de vier vragen van Niko Tinbergen. Vervolgens gaan we dieper in op de motivatie voor gedrag, leerprocessen, sociale systemen, natuurlijke ritmen, algemene beschouwingen rond domesticatie en gedrag en tenslotte stress en abnormale gedragingen. In Deel 2 komt vervolgens de soortspecifieke ethologie aan bod van hond, kat, paard en rund. In het laatste deel (Deel 3) raken we kort aan het concept dierenwelzijn, wat het is (en wat het niet is), hoe het tot stand kwam, hoe het gemeten kan worden en als laatste geven we ook nog bijzondere aandacht aan het concept pijn.
Tenslotte nog een verduidelijking vooraleer je deze cursus begint te lezen. De evolutietheorie van Charles Darwin (en daarnaast talrijk onderzoek in de vergelijkende anatomie, fysiologie, endocrinologie, …) leerde ons dat de mens ook een dier is. Wanneer we in deze cursus echter spreken over dieren, bedoelen we de niet-menselijke dieren. Zonder enig waardeoordeel te willen geven over de positie van de mens ten opzichte van andere dieren, maken we in deze tekst dus qua terminologie een onderscheid tussen de mens en niet-menselijke dieren, waarbij voor die laatste de term “dier” wordt gehanteerd

Als je dit hoofdstuk verwerkt hebt, beheers je de volgende leerdoelen:
• Je kan de vetgedrukte concepten beschrijven en telkens een voorbeeld geven dat het concept duidelijk maakt.
• Je kan een voorbeeld geven van de observaties die geleid hebben tot het begrip motivatie.
• Je kan de principes van de twee behandelde motivatiemodellen uitleggen en de verschillen en gelijkenissen beschrijven.
• Je kan het verband uitleggen tussen motivatietoestand en de subjectieve ervaring.
• Je kan de onderdelen van doelgericht gedrag opsommen en je kan uitleggen wat ermee wordt bedoeld.
• Je kan de behandelde methoden om motivatie te meten opnoemen en beschrijven
Motivatie is het geheel van inwendige factoren die tijdelijk en op omkeerbare wijze het gedrag kunnen veranderen (Nelissen 1996). Het is het proces dat bepaalt welk gedrag optreedt en wanneer. De inwendige factoren zijn bijvoorbeeld de prikkels waargenomen in het interne milieu (bloedsuikerspiegel, hormoonspiegel, …), inwendige ritmes waardoor organismen per dag, maand of jaar cyclische veranderingen ondergaan in activiteit (vb. migratie) en het voorafgaand optreden van hetzelfde gedrag en hoe lang dat geleden is. Hoewel vooral bepaald door interne factoren, zullen ook externe factoren het proces van motivatie beïnvloeden en sturen (zo zal de aanblik van smakelijke voeding een dier kunnen aanzetten tot eten ook al is er inwendig in het lichaam nog geen hongersignaal)
Vroeger werd voor motivatie ook de term ‘drang’ (of in het Engels ‘drive’) gebruikt, maar dat is nu niet meer het geval. Het verwarrende aan de term 'drive' is dat het impliceert dat het gaat om één variabele, met één enkele vergelijkingsschaal. Er zijn echter verschillende factoren die een dier 'drive' geven en zo invloed hebben op het al dan niet uitvoeren van een bepaald gedrag.
Enkele observaties uit het verleden die geleid hebben tot het begrip motivatie:
• De Amerikaanse behavioristen bestudeerden de relatie tussen stimulus en reactie of respons bij dieren Ze wilden niet spreken van motivatie, want dat zat in de ‘black box’ volgens hen (“wat we niet kunnen observeren, interesseert ons niet”). Ze kwamen echter in de problemen met die houding toen ze zagen dat vaak eenzelfde stimulus toch een ietwat andere respons kon uitlokken, dat die bijvoorbeeld een andere intensiteit had. Er was duidelijk iets dat de stimulus-respons relatie beïnvloedde. Ze noemden dat dan ‘mediërende variabelen’ of ‘intervening variables’. Later werd dit dan toch motivatie genoemd.
• De observatie dat sommige gedragingen sterker opkomen na een deprivatieperiode, dit is een periode waarin het substraat ontbreekt om een gedrag waarvoor een sterke inwendige motivatie bestaat, te vertonen Eens het substraat weer aanwezig is, zal het dier het gedrag in een hogere frequentie of duur vertonen dan vóór de deprivatieperiode, wat men een reboundeffect noemt. Voorbeeld: hoe langer het geleden is sinds een dier gedronken heeft, hoe harder het naar water zal zoeken en hoe meer het zal drinken eens het weer toegang heeft tot water. Dit gaat echter niet op voor alle motivaties. Agressief gedrag, bijvoorbeeld, komt maar op wanneer er een agressieve stimulus aanwezig is; er wordt niet intensiever gezocht naar een tegenstrever naargelang de deprivatietijd. Een dier kan wel agressiever gemaakt worden (d.w.z. de drempelwaarde verlagen) door genetische selectie (bv. gevechtsstieren en hanen) of door training.
• Sommige gedragingen bleken ogenschijnlijk in afwezigheid van de normaal vereiste externe prikkels toch uitgevoerd te worden (een mooi voorbeeld is de kraai van Lorenz die, volgens de wetenschapper zelf, fictieve vliegen ving). De uitleg is dan dat de motivatie zó sterk is dat het gedragspatroon zuiver door interne prikkels uitgelokt wordt. Deze gedragingen worden “vacuümactiviteiten” of “vacuümgedrag” genoemd. Later werd het bestaan van vacuümgedrag in twijfel getrokken: het zou kunnen dat het dier externe prikkels waarneemt
die wij niet kunnen zien (in het geval van de kraai van Lorenz: misschien kleine stofdeeltjes die reflecteren in het licht). In ieder geval wijst het op een sterke motivatie
• Men nam ook waar dat verschillende gedragspatronen vaak samen voorkomen op om een bepaald doel te vervullen. Bijvoorbeeld bij de hond: tanden ontbloten, pilo-erectie, grommen, uitvallen, bijten, zijn gedragingen die vertoond worden bij de aanval en die gegroepeerd worden onder de benaming "agressief gedrag". De valkuil hierbij was wel dat gedragspatronen ook bij verschillende types van motivatie kunnen gebruikt worden. Zo kan "bijten" ook gebruikt worden in de motivatietoestand "honger" om zich te voeden.
Het psychohydraulisch model werd bedacht door Konrad Lorenz in 1950, om het motivationele proces van Fixed Action Patterns te illustreren Belangrijk is om te onthouden dat het niet bedoeld is als een waarheidsgetrouwe voorstelling van hersenstructuren betrokken bij het proces, maar eerder om een principe duidelijk te maken;
Nemen we het voorbeeld van agressie bij een cichlidensoort (Astatotilapia bloyeti). Adulte mannelijke dieren van deze soort zullen met elkaar vechten volgens een geritualiseerd patroon van opeenvolgende FAP’s, met een toenemende drempelwaarde per gedragselement: (1) het lichaam vertoont een typisch kleurenpatroon, (2) parallel positioneren naast de tegenstander met maximale spreiding van de vinnen, (3) openen van het kieuwdeksel, (4) heen en weer slaan met de staartvin, (5) tegen de tegenstander beuken.
Lorenz stelde voor dat er zich bij dieren, zoals voor het gedragssysteem “agressie” bij de cichlide, inwendige 'nerveuze energie' (of motivationele energie) opbouwt. Hierdoor neemt de motivatie mettertijd toe om gedragspatronen geassocieerd met dat gedragssysteem uit te voeren. Dit wordt voorgesteld door een vat waarin water loopt en waarvan de uitloop afgesloten is door een klep met een veer. Aan de veer hangt een gewicht. Dit gewicht stelt de externe sleutelprikkels voor die het gedragssysteem deblokkeren, in geval van de cichlide is dat een ander mannetje Naarmate de tijd sinds de laatste uitvoering van de gedragspatronen toeneemt, zit er meer water (en dus endogene energie) in het vat, zal de drempelwaarde voor het gedrag verlagen en zijn prikkels van steeds mindere intensiteit of kwaliteit voldoende om de klep van het reservoir te laten openen en een actie te deblokkeren Omgekeerd, als de prikkel heel sterk is (en dus veel doorweegt), zal er minder motivationele energie nodig zijn om de klep te openen. Onder het reservoir bevindt zich een recipiënt waar de gedeblokkeerde energie in terechtkomt. Hoe verder de straal spuit, des te intenser is het gedrag Naarmate de hoeveelheid van de vrijgekomen energie, zal de cichlide meer intense gedragingen in de sequentie gaan vertonen
Voor sommige gedragssystemen kan het vullen van het reservoir zover gaan dat, uiteindelijk, het dier het gedrag uitvoert in afwezigheid van enige externe prikkels, omdat de motivatie zo hoog is (het zogenaamde vacuümgedrag). Zoals we al zagen zal dat bij agressief gedrag niet het geval zijn (zonder een externe sleutelprikkel, in het geval van de cichlide een ander mannetje, is er niets om agressie naar te vertonen). Het is ook mogelijk dat een reservoir zo vol is, dat de energie bij openen van de klep zo ver spuit, dat het niet in het eigen recipiënt maar in het recipiënt van een naburig gedragssysteem terechtkomt. Er wordt dan irrelevant gedrag vertoond. Vandaar de benaming “overspronggedrag”.
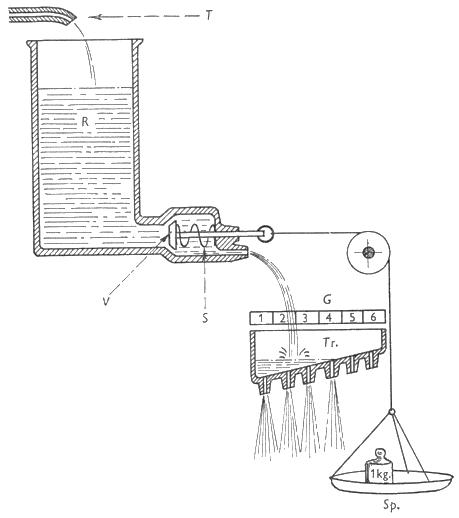
Figuur 7. Psychohydraulisch model van Konrad Lorenz (1950) als model voor het motivationele proces bij Fixed Action Patterns. Een kraan (T) levert continu endogene energie. De vloeistof in reservoir (R) is de energie die steeds ter beschikking is van het individu. Een klep (V) stelt het deblokkerend mechanisme voor en de veer (S) stelt de inhibitorische actie van hogere kernen voor die de uitstroom van de energie verhindert. De weegschaal (Sp) stelt de waarneming van externe sleutelprikkels voor. Het gewicht erop stelt de mate van stimulatie door externe prikkels voor. De interne accumulatie van energie en de externe prikkels werken dus in dezelfde richting, namelijk het wegduwen/-trekken van de klep. Bij weinig energie is er een sterke prikkel nodig om het gedrag te deblokkeren, terwijl wanneer R goed gevuld is, is er minder gewicht (minder sterke prikkel) nodig om het gedrag te deblokkeren. Energie die vrijkomt wordt opgevangen in de trog (Tr) eronder. Naarmate er bij de deblokkering meer energie overloopt van het reservoir naar de trog, zal een FAP met een hoger niveau van excitatie (een hogere drempelwaarde) worden vertoond. (Bron: Lorenz 1950)
Homeostatische modellen zijn actueel de meest gebruikte modellen. Er bestaan verschillende varianten van maar eentje kennen we al, namelijk de cybernetische visie van zelfregulatie uit hoofdstuk 1. Wanneer de actuele toestand van een dier voor een bepaalde parameter te ver afwijkt van de norm (de norm wordt soms ook wel setpoint genoemd), zal het dier een actie vertonen om die afwijking te verkleinen. Eens de situatie hersteld is, stopt de actie via de negatieve feedbacklus Dit model is zeer toepasselijk bij interne toestanden van honger, dorst of stress (cfr. de feedbackmechanismen bij de HPA-as), maar ook op het niveau van gedrag en externe prikkels Bijvoorbeeld: de norm bij de broedende grauwe gans is dat eieren in het nest liggen. Indien in de actuele situatie een ei naast het nest ligt, zal de eirolreactie starten om het ei terug te brengen. Eens het ei in het nest ligt, stopt dit gedrag bij het ouderdier. Homeostatische mechanismen leggen dus even goed de interne als de externe homeostase uit.
In de toegepaste ethologie (vooral in welzijnsonderzoek) is dit model erg belangrijk geworden, dankzij de uitbreiding door de Nederlandse etholoog Piet Wiepkema (1987). Hij stelde dat dieren in
gevangenschap de status van de buitenwereld monitoren en die aftoetsen aan een inwendige voorstelling van de gewenste toestand. Als er een afwijking tussen de twee is, dan zal er actie ondernomen worden om deze te verminderen. Het gaat er dus om of het dier controle heeft om die afwijking te verminderen of niet. Het terugkeren naar de norm (en het hebben van controle daarover) wordt door een dier als aangenaam ervaren, het afwijken van de norm als onaangenaam. Dit laatste is momenteel algemeen aanvaard voor de motivatieprocessen bij alle voelende wezens.
Een interessante vergelijkende analyse werd gemaakt door Jensen en Toates (1993). We geven deze hier mee, omdat je verder in de opleiding beide modellen nog zal tegenkomen en ze dan goed kan plaatsen ten opzichte van elkaar.
Samengevat kunnen we volgende wetmatigheden formuleren voor homeostatische modellen:
• Gedrag wordt gedeblokkeerd (en komt dus tot expressie) wanneer de waarde van een parameter afwijkt van de norm, waarbij de norm een zogenaamd optimaal evenwicht is.
• Het gedrag doet de motivatie afnemen en verdwijnen doordat het de afwijking tussen actuele en gewenste toestand vermindert
• Wanneer er geen afwijking is tussen actuele situatie en norm, dan wordt het gedrag niet gedeblokkeerd
Voor het Lorenziaanse psychohydraulisch model geldt het volgende:
• Een gedrag wordt gedeblokkeerd door een combinatie van interne en externe factoren.
• Wanneer een gedrag wordt uitgevoerd zal, na een initiële periode van positieve feedback, de motivatie gradueel afnemen en het gedrag stoppen, ongeacht de gevolgen van het gedrag.
• Wanneer een bepaald gedrag niet uitgevoerd wordt, zal de motivatie voor die actie gradueel toenemen.
Beide modellen stellen ten eerste dat gedrag gedeblokkeerd wordt als een gevolg van de afwijking van een parameter van een bepaalde waarde. Ten tweede zeggen ze dat het gedrag geïnhibeerd of gestopt wordt wanneer de waarde opnieuw de norm bereikt.
De verschillen zijn echter ook duidelijk. Bij het psychohydraulisch model is de eigenlijke uitvoering van het gedrag het essentiële feedbackmechanisme, terwijl het bij het homeostatisch mechanisme het gevolg van het gedrag is dat de motivatie doet verdwijnen. Bovendien zegt Lorenz dat de motivatie niet gelijk blijft in een constante omgeving, maar dat er een spontane toename van de motivatie is voor een gedrag wanneer dat gedrag niet uitgevoerd wordt (het vat vult zich spontaan met endogene energie). Bij het homeostatisch mechanisme is het niet noodzakelijk zo dat een dier een verandering in interne factoren zou ervaren in functie van de tijd sinds de laatste uitvoering van een specifiek gedrag (wat niet wil zeggen dat de norm niet kan variëren met interne factoren, zoals we ook al zagen in hoofdstuk 1).
Motivatie is het proces dat bepaalt welk gedrag optreedt en wanneer. Het gedrag is het resultaat van de interactie tussen tal van causale factoren, waarbij sommige sterker kunnen zijn dan andere. Bijvoorbeeld een koe in de weide kan zich onder een boom begeven om zich te beschutten tegen de zon, na een bepaalde tijd beginnen grazen en vervolgens gaan liggen en herkauwen.
Misschien is de koe wel hongerig wanneer zij gaat schuilen, maar de zon blaakt te hard zodat de thermoregulatorische homeostase gaat primeren. Nadien is de gewenste lichaamstemperatuur bereikt, ofwel is het hongergevoel zodanig gestegen dat het dier toch gaat grazen. Uiteindelijk bepalen interne prikkels dat het herkauwen moet aanvatten. Stel dat daags ervoor die koe getergd werd door een hond. Daarom gaat zij bijzonder uitkijken naar potentiële gevaren, wat voor gevolg heeft dat het herkauwen ietwat uitgesteld wordt.
Al de gedragingen beschreven in het voorbeeld zijn het resultaat zowel van interne prikkels en normen (homeostatische regeling van de energiebalans, van de temperatuur, vluchtafstand) als van externe prikkels (warmtestralingen, schaduw, gras, af te leggen afstand, gepaste plaats om te liggen) of zelfs de herinnering ervan (de hond). Die herinnering zal ook de algemene waakzaamheid en de graad van excitatie doen stijgen. De drempelwaarde van prikkels kan dus beïnvloed worden door de formatio reticularis (zie hoofdstuk causaliteit). Voor sommige gedragingen zal nog een interne klok een rol spelen (bv. paringsseizoen).
Men spreekt in de literatuur nu meestal van een "motivatietoestand" ("motivational state") in plaats van motivatie om duidelijker te maken dat binnen een bepaalde tijdsspanne verschillende motivaties actief zijn (met hun gemeenschappelijke en eigen neurofysiologische mechanismen). Er kan uiteindelijk maar één gedrag uitgevoerd worden. Een organisme wordt op het ogenblik van de uitvoering van bijvoorbeeld eetgedrag, door méér motivaties beïnvloed dan alleen maar honger.
Dit alles impliceert dat het organisme een keuze maakt tussen verschillende, soms tegenstrijdige motivaties. Uiteindelijk kan maar één motorisch patroon uitgevoerd worden. Het organisme zou dus op ieder ogenblik het belangrijkste gedrag moeten kunnen uitkiezen voor de overleving. Het is een onderwerp van discussie tussen specialisten, in welke mate het belang op kort termijn in strijd kan zijn met het belang op lange termijn. Het is duidelijk dat wegvluchten voor een predator belangrijker is dan blijven eten. Minder duidelijk is bv. hoe het komt dat sommige dieren toch migreren terwijl zij veel kansen lopen te sterven tijdens die activiteit? Die keuze impliceert op haar beurt dat er een soort gemeenschappelijke “munt” moet bestaan om verschillende motivaties te kunnen vergelijken. Volgens Cabanac zouden dat juist de gevoelens of de subjectieve ervaring kunnen zijn. Of anders gezegd, de mate waarin iets meer of minder aangenaam of onaangenaam is voor een dier
Wat aangenaam en onaangenaam is, is niet alleen individueel verschillend (afhankelijk van smaakvoorkeuren vinden we bepaald voedsel aangenaam om te eten of niet), het hangt ook vaak af van interne en externe factoren. Zo zal bijvoorbeeld voor ons een warme douche aangenaam zijn in de winter, maar niet zo aangenaam aanvoelen op een hete zomerdag
Welke dieren emoties kunnen ervaren en dus voelend zijn, is voer voor heel wat discussies onder wetenschappers. Op basis van vergelijkende studies rond neurale infrastructuur, cognitieve capaciteiten, fysiologische eigenschappen en gedragsmatige indicatoren, is de consensus momenteel dat alle gewervelde dieren en minstens enkele ongewervelden (inktvis en kreeft), voelend zijn.
Wanneer een dier een gedrag uitvoert om een specifiek doel te bekomen, dan spreken we van doelgericht gedrag. Het gedrag zal enkel uitgevoerd worden als er op dat moment een motivatie is om het doel te bekomen (het doel heeft waarde voor het dier), en als er een eerder geleerd verband is tussen het gedrag en het resultaat (het bekomen van het doel).
Hoewel tamelijk arbitrair, is het soms praktisch doelgericht gedrag als volgt in te delen.
• Appetitief gedrag: Het zoeken van het doel, bv. een prooi, een vijver met water, een partner. Vrij flexibel deel; tal van gedragingen kunnen hier ingeschakeld worden.
• Consummatorisch gedrag: De bevrediging zelf, zoals het doden en opeten van de prooi, drinken, baltsen en copulatie.
• Fase van verzadiging: na het bereiken van het doel.
De vraag is in welke mate bij een aantal soorten er een nood bestaat om appetitief gedrag als zodanig te vertonen. In gevangenschap krijgen meestal dieren bijvoorbeeld te eten zonder er iets te moeten voor doen. Op het vlak van de energetische balans is de homeostase bereikt, maar toch vertonen sommigen abnormaal gedrag (zie hoofdstuk stress en abnormaal gedrag) of gedragspatronen die op jacht- of foerageergedrag lijken alvorens de dieren het eten verorberen. In dergelijke gevallen verdwijnen deze gedragingen wanneer huisvesting en management een categorie appetitieve gedragingen toelaten. In andere gevallen zou het eerder gaan om in het algemeen een bepaald tijdspercentage “actief te blijven”, welke het gedrag ook is We komen hier nog op terug in Deel 3 (Dierenwelzijn) van de cursus.
Het meten van motivatie kan interessant zijn om verschillende redenen. Bijvoorbeeld hoe een dier kiest tussen verschillende gedragingen binnen de motivatietoestand en hoe die keuze verandert overheen de tijd (zoals bij leeftijdsgebonden variatie, of diurnale en seizoenale variatie). Ook de motivatie om te kiezen voor een bepaalde reproductieve partner of bepaalde resource kan gemeten worden. Een ander voorbeeld is dat motivatietesten gebruikt kunnen worden om te weten wat er belangrijk is voor dieren die we in gevangenschap houden. Dat laatste zal uitgebreider aan bod komen in Deel 3 wanneer we het over dierenwelzijn hebben.
Hierbij gaat men een dier één of meerdere substraten geven en meten hoe vaak en/of hoe lang het gedrag dat ermee gepaard gaat, wordt vertoond. Men kan ook gaan kijken wat de latentietijd is (de duur vanaf het geven van het substraat totdat het dier het gedrag vertoont) of wat het verbruik van een bepaald substraat is. Een valkuil bij dit type onderzoek is wel dat de metingen sterk kunnen afhangen van de aanwezige stimuli (bijvoorbeeld: als er niet ad libitum voedsel wordt gegeven, dan is voedingsgedrag beperkt en zijn de observaties misschien vertekend).
Dergelijke studies kunnen spontaan gedrag bestuderen (bijvoorbeeld het bitter maken van het water om te kijken hoe groot motivatie om te drinken is), maar soms kan er ook eerst een leerperiode aan vooraf gaan. Men geeft bijvoorbeeld een kip toegang tot een nestplaats dat ze kan bereiken door over een metalen rooster te wandelen. Eens men zeker is dat de kip dit gedrag uitvoert, gaat men lichte elektrische schokken van verschillende intensiteit geven via het rooster om te kijken of de kip nog naar de nestplaats gaat en vanaf welke schokintensiteit ze dit niet meer doet (op dat moment is de motivatie om de schokken te vermijden groter dan de motivatie om het nest te bereiken)
Hierbij werkt men met een onderzoeksopzet gelijk aan de principes van de Skinnerdoos (zie hoofdstuk leerprocessen). Men leert een dier eerst om een bepaalde taak uit te voeren in ruil voor een resource (vb. nestmateriaal). Daarna gaat men de prijs verhogen door het dier meerdere keren de taak te laten uitvoeren alvorens er toegang tot de resource is. Hoe vaker het dier blijft duwen, hoe belangrijker het doel moet zijn. Er wordt dan nagegaan wat de maximumprijs is die het dier bereid is te ‘betalen’.
Ben-Ami Bartal I, Decety J, Mason P. 2011. Empathy and pro-social behavior in rats. Science, 334(6061):1427-1430.
Blakemore C, Cooper GF. 1970. Development of the brain depends on the visual environment. Nature: 228:477-478.
Braastadt BO, Mcbride A, Newberry RC. 2022. The cat: Behaviour and Welfare. CABI, Wallingford, 209 pp.
Broom D. 1986. Indicators of poor welfare. British Veterinary Journal, 142:524-526.
Broom D, Johnson KG. 2019. Stress and animal welfare (2e ed.). Springer Nature Switzerland, Cham. 223 pp.
Chu B, Marwaha K, Sanvictores T, Ayers D. 2022. Physiology, Stress Reaction. National Library of Medicine. Physiology, Stress Reaction - StatPearls - NCBI Bookshelf (nih.gov)
Cooper RM, Zubek JP. 1958. Effects of enriched and restricted early environments on the learning ability of bright and dull rats. Canadian Journal of Psychology 12, 159-164.
Cooper JJ, Nicol CJ. 1994. Neighbour effects on the development of locomotor stereotypies in bank voles (Clethrionomys glareolus). Animal Behaviour, 47, 222-224.
D’Eath RB, Arnott G, Turner SP, Jensen T, Lahrmann HP, Busch ME, Niemi JK, Lawrence AB, Sandøe. 2014. Injurious tail biting in pigs: how can it be controlled in existing systems without tail docking? Animal: 8(9):1479-1497.
de Waal FBM, Leimgruber K, Greenberg AR. 2008. Giving is self-rewarding for monkeys. Proceedings of the National Academy of Sciences (PNAS), 105(36):13685-13689
Dias BG,n Ressler KJ. 2014. Parental olfactory experience influences behavior and neural structure in subsequent generations. Nature Neuroscience: 17(1):89-96.
Domjan M. 1998. The principles of learning and behavior (4e ed.) Thomson Brooks/Cole Publishing Co. Pacific Grove. 435pp.
Dugatkin LA. 2018. The silver fox domestication experiment. Evolution: Education and Outreach: 11, artikelnummer 16
Duncan IJH en Petherick JC. 1991. The implications of cognitive processes for animal welfare. Journal of animal Science, 69:5017-5022.
Epstein R, Kirschnit CE, Lanza RP, Rubin LC. 1984. ‘Insight’ in the pigeon: antecedents and determinants of an intelligent performance. Nature, 308:61-62.
FAWC. 1979. Press release 5 December 1979. Klik hier voor het persartikel. Laatst geraadpleegd op 7 september 2023.
FAWC. 1993. Second report on priorities for animal welfare research and development. FAWC, Surrey. 26 pp.
Francis DD, Dorio J, Plotsky PM, Meaney MJ. 2002. Environmental enrichment reverses the effects of maternal separation on stress reactivity. Journal of Neuroscience, 22(18)7840-7843.
Gelez H, Fabre-Nys C. 2004. The “male effect” in sheep and goats: a review of the respective roles of the two olfactory systems. Hormones and Behaviour, 46(3):257-271.
Grammer K, Fink B, Neave N. 2005. Human pheromones and sexual attraction. European Journal of Obstetrics & Gynecology and Reproductive Biology, 118(2):135-142.
Jensen P, Toates FM. 1993. Who needs ‘behavioural needs’? Motivational aspects of the needs of animals. Applied Animal Behaviour Science, 37:37:161-181.
Leon M, Moltz H. 1972. The development of the pheromonal bond in the albino rat. Physiology and Behaviour, 8(4):683-686.
Liu D, Dioro J, Tannenbaum B, Francis D, Freedman A, Sharma S, Pearson D, Plotsky PM, Meaney MJ. 1997. Maternal care, hippocampal glucocorticoid receptors and hypothalamic-pituitary-adrenal responses to stress. Science, 277(5332):1659-1662.
Lorenz K. 1950. The comparative method in studying innate behavior patterns. Symposia of the Society for Experimental Biology, 47:221-254. 1950-InnateBehavior (klha.at)
Lu A, Feder JA, Snyder-Mackler N, Bergman TJ, Beehner JC. 2021 Male-mediated maturation in wild Geladas. Current Biology, 31(1):214-219.
Manning A, Dawkins M S 2012 An Introduction to Animal Behaviour. Cambridge University Press, Cambridge, 458 pp.
Marshall-Pescini S, Dale R, Quervel-Chaumette M, Range F. 2016. Critical issues in experimental studies of prosociality in non-human species. Animal Cognition, 19:679-705.
McBride G. 1964. A general theory of social organization and behaviour. University of Queensland Papers, 1(2): 110 pp.
McClearn GE. 1962. The inheritance of behavior. In: Psychology in the making, Postman LJ (ed ), Alfred A. Knopf, New York City, p 144-252.
Meaney MJ, Aitken D, Van Berkel C, Bhatnagar S, Sapolsky RM. 1988. Effect of neonatal handling on age-related impairments associated with the hippocampus. Science, 239(4841:766-768).
Meaney MJ, Diorio J, Francis D, Weaver S, Yau J, Chapman K, Seckl JR. 2000. Postnatal handling increases the expression of cAMPinducible transcription factors in the rat hippocampus: the effects of thyroid hormones and serotonin. Journal of Neuroscience, 20:3926–3935.
Mellor D. 2016. Moving beyond the “Five Freedoms” by Updating the “Five Provisions” and introducing Aligned “Animal Welfare Aims”. Animals, 6:ani6100059.
Mellor D, Beausoleil NJ, Littlewood KE, McLean AN, McGreevy PD, Jones B, Wilkins C. 2020. The 2020 Five Domains Model: Including Human-Animal Interactions in Assessments of Animal Welfare.
Monclus R, Rödel HG, Von Holst D, De Miguel J. 2005. Behavioural and physiological responses of naïve European rabbits to predator odour. Animal Behaviour: 70(4):753-761.
Mostafa T, El Khouly G, Hassan A. 2012. Pheromones in sex and reproduction: Do they have a role in humans? Journal of Advanced Research, 3(1):1-9.
Nelissen M. 1996. Lexicon van de gedragsbiologie. Galant, Leuven, 112 pp
Owczarczak-Garstecka SC en Burman OHP. 2016. Can sleep and resting behaviours be used as indicators of welfare in shelter dogs (Canis lupus familiaris)? PLoS One, 11(10): e0163620.
Purves WK, Orians GH, Heller HC, Sadava D. 1998. Live – The science of biology (5e ed.). Sinauer Associates, Massachussetts, 1243 pp
’t Hart M. 1978. De stekelbaars. Het spectrum, Utrecht, 112 pp
Rault JL. 2019 Be kind to others: Prosocial behaviours and their implications for animal welfare. Applied Animal Behaviour Science, 210:113-123.
Sankarganesh D, Kirkwood R, Nagnan-Le Meillour P, Angayarkanni J, Achiraman S, Archunan G. 2022. Pheromones, binding proteins, and olfactory systems in the pig (Sus scrofa): a review. Frontiers in Veterinary Science, 9: artikelnummer 989409.
Schleidt WM. 1974. How “fixed” is the fixed action pattern? Zeitschrift zur Tierpsychologie, 36:184211. Klik hier om naar de paper te gaan.
Silva Moreira P, Almeida PR, Leite-Almeida H, Sousa N, Costa P. 2016. Impact of chronic stress protocols in learning and momory in rodents: systematic review and meta-analysis. PLoS One, 11(9):e0163245.
Skolnick NJ, Ackerman SH, Hofer MA, Weiner H. 1980. Vertical transmission of acquired ulcer susceptibility in the rat. Science: 208(4448),1161-1163.
Sokolowski K en Corbin JG. 2012. Wired for behaviors: from development to function of innate limbic system circuitry. Frontiers in Molecular Neuroscience, 5:55
Stamp Dawkins M en Guilford T 1991. The corruption of honest signalling. Animal Behaviour, 41:865873.
Tinbergen N. 1951. The Study of Instinct. Oxford University Press, Londen, 228 pp
Tinbergen N. 1978. In ’t vrije veld. Het spectrum, Utrecht, 279 pp
Van Oortmerssen GA. 1981. Gedrag en erfelijkheid. In: Ethologie – De biologie van gedrag. Baerends GP (ed.), Centrum voor Landbouwpublikaties en Landbouwdocumentatie, Wageningen, p 115135.
Webster J. 1994. Assessment of animal welfare: The Five Freedoms. In: Animal Welfare: A Cool Eye Towards Eden; Blackwell Science, Oxford, UK, pp10-14.
Weiss JM. 1972. Psychological factors in stress and disease. Scientific American, 226(6):104-113.
Wiedenmayer C. 1997. Causation of the ontogenetic development of stereotypic digging in gerbils. Animal Behaviour, 53(3):461-470.
Wiepkema PR. 1987. Behavioural aspects of stress. In: Biology of Stress in farm Animals: An Integrative Approach. Wiepkema PR, van Adrichem PWM (eds.). Martinus Nijhoff, Dordrecht, pp. 113-133.
Wispé LG. 1972. Positive forms of social behavior: an overview. Journal of Social Issues, 28:1-19.
Zhang L, Hastings MH, Green EW, Tauber E, Sladek M, Webster SG, Kyriacou CP, Wilcockson DC. 2013. Dissociation of Circadian and Circatidal Timekeeping in the Marine Crustacean Eurydice pulchra. Current Biology, 23(19):1863-1873.
Hieronder volgen nog enkele publicaties die een overzicht geven van de verschillende aspecten van de ethologie of van aanverwante gedragswetenschappen.
Als inleiding tot de ethologie:
Jensen P. (Editor). 2017. The ethology of domestic animals (3rd edition). CABI Publishing, Oxfordshire, UK. ISBN 9781786391650.
Manning, A. and Dawkins, M. S. (2012): An Introduction to Animal Behaviour. Cambridge University Press, Cambridge. (6e uitgave) ISBN 0521165148
Nordell, S.E. and Valone, T.J. (2017): Animal Behaviour. Second International Edition. Oxford University Press, Oxford. ISBN: 978-0-19-027678-2.
Een mooi voorbeeld van de neuroethologie:
Sillar, K.T., Picton, L.D. and Heitler, W.J. (2016): The Neuroethology of Predation and Escape. WileyBlackwell, Hoboken, New Jersey. ISBN: 978-0-470-97224-3
Ethologie van de menselijke soort:
Campan, R. et Scapini, F. (2002): Ethologie. Approche Systémique du Comportement. De Boeck Université, Bruxelles. Chapitre 10: Ethologie Humaine (pp. 603-668) ISBN 2804137651
de Lannoy, J.-D. et Feyereisen, P. (1987): L'Ethologie Humaine. Presses Universitaires de France, Paris. (Que Sais-Je? No. 2339) ISBN 2130398456
Nelissen, M. (2000): De Bril van Darwin. Lannoo, Tielt. ISBN 902094116X
Ethologische visie van de psychiatrie:
Demaret, A. (1979): Ethologie et Psychiatrie. Mardaga, Bruxelles. ISBN 2870091133
Stevens, A. And Price, J. (2000): Evolutionary Psychiatry. Routledge, London. ISBN 0415219795
Panksepp, J. (Ed.) (2004): Textbook of Biological Psychiatry. Wiley, New York. ISBN 0471434787
Ethologische observatietechnieken:
Rees, P.A. (2015): Studying Captive Animals. Wiley Blackwell, Hoboken, New Jersey, ISBN 1118629361
Lehner, P.N. (2002): Handbook of Ethological Methods. Cambridge University Press, Cambridge.
ISBN 0521637503
Martin, P. and Bateson, P. (2021): Measuring Behaviour. 4th Edition, Cambridge University Press, Cambridge.
ISBN 9781108745727
Genetica en gedrag
Moleculaire en populatiegenetica:
Anholt, R.R. and Mackay, T.F.C. (2010): Principles of Behavioral Genetics. Elsevier, Amsterdam.
ISBN 978 0 12 372575 2
Populatiegenetica:
Grandin, T. (Ed.)(2014): Genetics and Behavior of Domestic Animals. 2nd Edition, Academic Press, San Diego.
ISBN 9780123945860
Hahn, M.E. et al. (Eds) (1990): Developmental Behavior Genetics. Oxford University Press, New York & Oxford. ISBN 0195054466
Plomin, R., DeFries, J.C. and McClearn, G.E. (1980): Behavioral Genetics. A Primer. Freeman and Co., San Francisco. ISBN 0716728249
Leertheorieën en behavioristische visie van het gedrag:
Daffin L. 2021. Principles of learning and behavior. 2nd edition. Washington State University. Principles of Learning and Behavior – Principles of Learning and Behavior (wsu.edu) Het boek is gratis raadpleegbaar online en ook te downloaden als PDF via de link bovenaan de pagina
Domjan M. 2013. Principles of Learning and Behavior. 7th edition. Cengage Learning, Inc Publishing.
ISBN 1285088565
Pierce J.M. 2008. Animal Learning and Cogntion. 3rd Edition. Psychology Press. 432p. ISBN 1841696552
Schwartz, B., Wasserman, E.A. and Robbins S.J. (2002): Psychology of Learning and Behavior. Norton & Co, New York & London. (5e uitgave). ISBN 9780393975918
Stress, psychosomatiek, neurofysiologie, neurobiochemie:
Buckingham, J.C., Gillies, G.E. and Cowell, A.-M. (1997): Stress, Hormones and the Immune System. J. Wiley & Sons, Chichester.
Moberg, G.P. and Mench, J.A. (Eds)(2000): The Biology of Animal Stress. Basic Principles and Implications for Animal Welfare. CABI, Wallingford. ISBN 0851993591
Volgende auteurs presenteren daarenboven een heel mooie interdisciplinaire synthetische visie van de werking van hogere organismen. Het zijn oudere boeken maar niet verouderd.
Henry, J.P. and Stephens, P.M. (1977): Stress, Health, and the Social Environment. A Sociobiological Approach to Medicine. Springer Verlag, New York, Heidelberg, Berlin. ISBN 0387902937
Karli, P. (1987): L'Homme Agressif. Editions Odile Jacob, Paris. ISBN 202009553X
Laborit, H. (1986): L'Inhibition de l'Action. Biologie Comportementale et Physio-Pathologie. Masson, Paris. ISBN 2225806411