Ecología a mi manera Ariel E. Lugo

Ecología a mi manera
Ariel E. Lugo
Coordinación editorial Pamela Navarro (pamelaapr@gmail.com)
Diseño y diagramación Pascual Estrada (estrada.pascual@gmail.com)
Portada
Fotografía de Jerry Bauer. Bosque elfino en el tope del Pico del Este, Bosque Experimental de Luquillo. El bosque elfino es un humedal de montaña que capta agua y nutrientes de las nubes cuando estas entran al bosque. Las nubes son visibles al fondo de la foto.
Contraportada
La foto antigua es de John C. Gifford y la más reciente de Jerry Bauer. Vista de las Montañas de Luquillo en 1905 y 2008. Las dos fotografías, tomadas desde el pueblo de Río Grande, ilustran la transformación dramática del paisaje de la región, de agrícola a urbano.
ISBN 979-8-9882827-0-9
Ariel E. Lugo
Tamesis #1528, El Paraíso. Río Piedras, Puerto Rico 00926 (aelugo1234@gmail.com)
Río Piedras, Puerto Rico 2023
Cita sugerida
Lugo, A.E. 2023. Ecología a mi manera. HANAEL Asociados Inc. Río Piedras, Puerto Rico. 325 pp.
Ecología a mi manera
Ariel E. Lugo
5 ariel e. lugo | ecología a m i m anera Contenido Prólogo 11 CaPítulo 1 15 Grandes ecólogos 15 CaPítulo 2 29 El ecosistema: unidad de la vida en el planeta 29 CaPítulo 3 45 Estequiometría y alometría 45 Estequiometría 46 Alometría 56 CaPítulo 4 63 l a conservación en el a ntropoceno 63 ¿Qué ha hecho el ser humano? 63 Calentamiento global y el cambio climático 67 Balance de gases: C, N, O3 67 Expansión del agua 69 Cambio en el pH del agua 70 Causas no-antropogénicas vs . antropogénicas del cambio climático: ¿Cuál es la evidencia? 70 Cambio de las condiciones en Puerto Rico 71 Temperatura 71 Lluvia 72 Huracanes 72 Nivel del mar 73 Acidificación del mar 73 Efectos del cambio climático 73 En regiones urbanas 73
6 ariel e. lugo | ecología a m i m anera En regiones agrícolas 74 En regiones rurales 75 Adaptación: la idea de lo novel o novedoso en el Antropoceno 76 Adaptaciones al nivel SETS 77 CaPítulo 5 81 los bosques tropicales 81 Trasfondo histórico 81 Algunos principios fundamentales 85 El sistema de ecorregiones de Bailey 92 ¿Cuáles son las limitaciones ecológicas de clasificaciones basadas en geografía? 94 Sistema de zonas de vida de Holdridge 94 Comparación entre el sistema de Holdridge y el de Bailey 99 CaPítulo 6 103 los bosques de Puerto Rico 103 El estudio y conservación de los bosques de Puerto Rico bajo el Imperio Español 103 El estudio y conservación de los bosques de Puerto Rico bajo el Imperio Norteamericano 105 Clasificación y geografía de los bosques de Puerto Rico 114 Condiciones comunes de todos los bosques de Puerto Rico y sus efectos ecológicos 117 Seis condiciones que ayudan a explicar la biogeografía y diversidad de los bosques de Puerto Rico 120 CaPítulo 7 127 Bosques secos 127 Introducción 127 Bosques secos en general 127 Factores que afectan el crecimiento de los bosques secos 128 Características ecológicas y ecofisiológicas de los bosques secos 129 Efectos antropogénicos 132 Las bases de la resiliencia y sus límites 137 CaPítulo 8 141 Bosques húmedos, muy húmedos y pluviales 141 Introducción 141 Resultados del inventario de bosques 142 Variación edáfica y de biomasa en bosques aparentemente homogéneos 143
7 ariel e. lugo | ecología a m i m anera Disturbios y mortalidad arbórea 144 Uniones de tabonuco 146 Sucesión 148 Dominancia de especies 150 Flujo y almacenaje de masa y nutrientes 151 La catena 156 Bosques de tabonuco remanentes y noveles 157 CaPítulo 9 161 Humedales arbóreos 161 Introducción 161 Definición 161 La energía y los tipos funcionales de humedales 163 El concepto de la huella energética o energy signature 165 La diversidad de humedales en Puerto Rico 165 Modelo del efecto de la actividad humana sobre los humedales 171 CaPítulo 10 173 Manglares 173 Introducción 173 Zonación 174 Relación entre los manglares y las mareas 175 Primer estudio de manglares en cuanto al ecosistema 177 Interpretación de la zonación del mangle 177 Síntesis del manglar y su funcionamiento 180 El manglar y su función en las cadenas alimenticias 183 Cambio histórico en el área de los manglares de Puerto Rico 183 CaPítulo 11 187 El Yunque 187 Introducción 187 Ubicación geográfica 187 Geología e historia natural de Las Montañas de Luquillo 189 Tipos de rocas 189 Cómo eventos geológicos, Ma atrás, afectan el ciclaje de nutrientes de los bosques del presente: simplificando una historia compleja 192 La combinación del levantamiento tectónico, la litología, la erosión química (weathering), insumos atmosféricos y los bosques determinan la disponibilidad de cationes minerales en las Montañas de Luquillo 196
8 ariel e. lugo | ecología a m i m anera Gradientes 197 Parámetros físicos 199 Grupos de organismos 201 Parámetros estructurales de los bosques 204 Procesos ecológicos y tipos de bosques 209 Régimen de disturbios 213 Huracanes 213 Sequías 214 Derrumbes 214 Cambio en cobertura y uso de la tierra 215 Biota y sistemas ecológicos 217 ¿Es o no El Yunque una maravilla natural del planeta? 219 CaPítulo 12 221 Ecología de plantaciones arbóreas tropicales 221 Introducción 221 Los bosques plantados 221 Área de plantaciones 224 Controversias 226 La influencia de las especies en el funcionamiento de la plantación 228 Comparación de plantaciones con bosques nativos de la misma edad 235 Efectos de huracanes sobre la plantación de pino en Cubuy 240 Plantaciones para restaurar biodiversidad, rescatar tierras degradadas y catalizar la sucesión 242 CaPítulo 13 245 El ciclo de carbono de los bosques tropicales 245 El almacenaje y flujo de carbono en un rodal 246 Factores que afectan los almacenajes y flujos de carbono en bosques tropicales 249 El rol de los bosques tropicales en el balance de carbono del mundo 253 El ciclo de carbono global no cuadra con los datos disponibles 256 Factores que afectan el intercambio de carbono entre los bosques tropicales y la atmósfera 258 Sumideros terrestres de carbono atmosférico 258 Estrategias para aumentar los sumideros de carbono atmosférico 258 CaPítulo 14 261 las velocidades de extinción y evolución de las especies 261 Extinciones masivas 261 Estimaciones de tasas de extinción de especies 262
9 ariel e. lugo | ecología a m i m anera Especies y hábitats más vulnerables a la extinción 268 Lecciones de Puerto Rico 268 La homogenización de las comunidades vegetales 269 Recuperando la biodiversidad 270 Especies introducidas, extinciones y aceleración de la evolución 270 CaPítulo 15 275 l a é tica de la tierra 275 Definición 275 Introducción 275 Consultorías 276 Ejemplos 276 Silenciando la ciencia 277 El científico individual 277 Conclusión 278 La ética de la Tierra 278 Epílogo 281 El ambiente del científico 281 El futuro de los bosques tropicales y de la humanidad 282 literatura citada 285 índice de materias 311
ariel e. lugo | ecología a m i m anera
Durante el verano de 2009, mientras compartía con un grupo de participantes del programa AlACiMa (Al ianza para el A prendizaje de Ciencias y Ma temáticas), mi estudiante de maestría Jéssica Fonseca da Silva se quejó de la falta de ofrecimiento de cursos graduados en ecología para el 2010 y el hecho de que se iba a graduar sin haber tomado un curso conmigo. Indicó que a los estudiantes de Biología les interesaría mi curso. El argumento de Jéssica sobre el no haberlo tomado caló hondo y me di cuenta de que lo mismo le pasaría a Oscar Abelleira, otro estudiante de maestría a punto de graduarse. Esto no había ocurrido con los otros ocho estudiantes de maestría y cinco de doctorado que había dirigido en mi carrera hasta este momento. Por esto, esa misma noche pensé en los temas de un curso de ecología, que llamé «Ecología a mi manera», para ver si era del interés de Jéssica. Me dijo que sí, y comencé a desarrollarlo. Siempre he pensado que, si voy a pasar el trabajo de escribir algo, lo mejor es hacerlo para publicarlo. Fue así como decidí compilar los temas de las conferencias en un libro compacto sobre la ecología de Puerto Rico.
¿Por qué «a mi manera»? Principalmente porque tanto la clase como este libro presentan una visión u opinión muy personal de los tópicos ecológicos. Este no es un libro de texto donde se supone que se presente una versión balanceada de los tópicos incluidos. Esta obra está fundamentada en la literatura, mi experiencia como investigador, y representa mi punto de vista; de ahí el que sea «a mi manera».
Durante el curso aprendí mucho de los estudiantes y de las discusiones que tuvimos. En varios temas utilicé la información provista por ellos, asegurándome de darle crédito a sus contribuciones. En la tabla adjunta se encuentra la lista de los estudiantes que participaron en el curso. Para mantener el libro compacto, el material se presenta en forma abreviada, pero la bibliografía es extensa y contiene todos los detalles necesarios para satisfacer al lector interesado en profundizar en uno u otro tema. Las tablas y figuras que acompañan el texto apoyan y expanden el contenido, pero depende del lector interpretar los datos, ya que el libro no contiene las explicaciones que ofrecí a los estudiantes en forma de conferencia durante las clases.
El punto de vista de este trabajo y la selección de los temas es mi responsabilidad. La idea del curso, y por ende de este libro, es presentar a los estudiantes una
ariel e. lugo | ecología a m i m anera Prólogo
11
forma particular de pensar sobre los ecosistemas tropicales, incluidos los de Puerto Rico, y los temas ecológicos del momento. La selección de temas está basada en mi experiencia en el debate científico. En el primer capítulo resumo someramente las contribuciones científicas y escuelas de pensamiento de 15 científicos que influyeron en mí y en la forma como se presenta la información en el libro. Los capítulos dos y tres resumen los aspectos cuantitativos, químicos y energéticos que establecen las condiciones bajo las cuales operan los ecosistemas. El cuarto capítulo define las condiciones noveles que el ser humano ha introducido en el planeta y que ahora rigen el funcionamiento de los ecosistemas y por ende las actividades de conservación durante la Época del Antropoceno. Luego, en el capítulo cinco se definen las condiciones que llevan a la clasificación de los bosques tropicales, amparado en la idea de las zonas de vida de Holdridge. Los próximos siete capítulos resumen la información disponible sobre los bosques de Puerto Rico, principalmente aquellos que he estudiado con algún detalle. Los últimos tres capítulos están dedicados a las grandes controversias que hacen de la ecología una ciencia contenciosa, pero que, por su relevancia al ser humano, envuelven asuntos de vital importancia para la conservación de la biodiversidad. El libro termina con una reflexión sobre los cambios que he observado en el campo de la ecología y algunos pensamientos sobre el futuro de los sistemas tropicales y su biodiversidad.
Dallas, 20 de enero de 2010
La edición final del trabajo se llevó a cabo entre el 2020 y 2021 después de haber ofrecido el curso a otra cohorte de estudiantes en el 2015 (Tabla adjunta). Aproveché para poner al día algunas estadísticas, incluyendo datos sobre los bosques de Puerto Rico e Islas Vírgenes. Para algunos capítulos incluyo una pregunta general sobre el tema del capítulo, al igual que asignaciones al final. A veces doy la contestación a la pregunta y de no hacerlo, el material presentado es suficiente para inferirla. Durante el periodo de clases, Mildred Alayón y Grisel (Lía) Sánchez me ayudaron con la preparación del material para las clases. Mildred Alayón editó todo el manuscrito previo a su publicación y Helen Nunci ayudó con la producción final, incluyendo la traducción al español de todas las figuras. A Ernesto Medina por su lectura y valiosas sugerencias. Josué E. Oquendo Natal, asistido por Melissa Álvarez Xilos, ambos artistas gráficos, prepararon todo el arte gráfico y Olga Ramos González me ayudó con los mapas. La coordinación editorial estuvo a cargo de Pamela Navarro y Pascual Estrada se encargó del diseño, diagramación y presentación de este libro. A
HANAEL Asociados Inc., por el apoyo financiero para la publicación de esta obra. A todos les estoy agradecido.
Río Piedras, 1 de junio de 2023
12 ariel e. lugo | ecología a m i m anera Prólogo
Estudiantes que participaron en los cursos de historia natural de Puerto Rico. El curso se identificó como Biología 5039 en el catálogo del Departamento de Biología de la Universidad de Puerto Rico en Río Piedras.
2010
2015
Raisa Hernández Pacheco Adrián Lugo Bendezú
Jéssica Fonseca da Silva Aixa Ramírez Lluch
Sandra Ayala Cruz Arístides Colón Castillo
Michelle J. Rivera Benjamín Branoff
Gabriel O. Estrada Carlos Germosen Contreras
Julio C. Lazcano Lara Danay D. Macías Reyes
Ruber Rodríguez Barreras Daniela D. Yaffar de la Fuente
Estrella D. Santiago Pérez Diego O. Rosario Díaz
Isis López Q. Gabriela M. Morales Nieves
Marcos González Jardany Díaz Salgado
Gusth Merly Pérez Jorge L. Rivera Velázquez
María Alexandra Addarich Figueroa
Nerybelle Pérez-Rosas
Sara L. Arocho Rivera
Sulpicio Flores Chalco
Wendeline M. Figueroa Hernández
13 ariel e. lugo | ecología a m i m anera Prólogo
CaPítulo 1 Grandes ecólogos
La ecología tiene sus orígenes en los trabajos de historia natural de los siglos XVIII y XIX. Kormondy (1965) acredita los trabajos de Linnaeus y Ferchault de Réaume a mediados del siglo XVIII como los precursores de la ecología moderna. Los historiadores de la ecología están de acuerdo en que la ecología tuvo su desarrollo principal en el siglo XX, especialmente durante la segunda mitad del siglo (Kingsland 2005). Sin embargo, el pensamiento ecológico ha estado presente en todas las épocas, incluyendo aquellas cuando estaban activos los grandes filósofos griegos como Theophrastus o los grandes pensadores de las dinastías chinas o del mundo musulmán. Las bases físicas y químicas de la ecología se desarrollaron durante el siglo XIX, por ejemplo los trabajos de Liebig (1840) y la teoría de la evolución de Darwin sentaron las bases del pensamiento moderno sobre la selección natural (Mayr 1991).
La ecología es una ciencia joven. Conocí a Víctor Shelford, considerado el padre de la ecología animal en los Estados Unidos de América (EE. UU.), en la Universidad de Illinois el 17 de agosto de 1965 durante la reunión anual de la Sociedad Ecológica de Norte América. Durante la primera mitad del siglo XX los ecólogos norteamericanos desarrollaron los conceptos de las comunidades de plantas y animales y la idea de la sucesión ecológica. El concepto de ecosistema surgió en 1935 (Golley 1993) y fue durante las décadas de los años 40 y 50 que surgieron las bases ecológicas para la conservación1 de los sistemas ecológicos. Durante las décadas de los 60 y 70 se establecieron las bases de la ecología cuantitativa, basada en trabajos de Lotka (1956) y de otros a principios del siglo. Coleman (2010) se refiere a este tipo de ciencia como Big Ecology y su libro describe su historia e importancia. Después de la década de 1980, la ecología tomó mucho auge en función de los desarrollos tecnológicos que permitieron estudios a gran escala y de alta sofisticación para incluir aplicaciones de modelaje con el desarrollo de las computadoras. La narrativa de Egerton (2015) sobre la historia de la Sociedad Ecológica de Norte América explica los avances hechos por prominentes ecólogos durante un periodo de 100 años, culminando en el 2015.
ariel e. lugo | ecología a m i m anera
15
1 En mi opinión, la conservación y el manejo son sinónimos (Lugo 1989). Para algunos el manejo tiene connotaciones negativas y apoyan la preservación. El manejo profesional basado en la ciencia es conservación e incluye la preservación.
Para propósitos de este libro, en la lista de científicos que sigue solo enfatizo un grupo muy limitado de grandes ecólogos usando como criterio su influencia sobre los conceptos e ideas que se repiten prominentemente en el manuscrito. Más aún, la lista se limita a los ecólogos más familiares y admirados, sin que esto implique que no haya muchos más ecólogos igualmente importantes, admirados e influyentes en las ideas expuestas en el libro. Finalmente, la información presentada para cada ecólogo no representa la totalidad de la obra de ese científico, ni necesariamente sus contribuciones principales. De nuevo, solo incluyo aquellos asuntos que me llaman la atención. Los lectores interesados en información más sistemática y completa deben consultar la literatura citada.
1. Charles Robert Darwin. Científico británico conocido por ser el proponente de la teoría de la evolución (Mayr 1991). La teoría de evolución de Darwin está basada en un entendimiento fundamental de la ecología y cómo las fuerzas ambientales influyen en la biota. Su pensamiento repercute en todos los aspectos de la ecología, particularmente en el presente, cuando el ser humano está cambiando las condiciones bajo las cuales los organismos evolucionan. Menos conocida es la contribución de Darwin al desarrollo de la ecología de suelos (Ghilarov 1983, Graff 1983, Hayes 1983), y ecología de invertebrados con su libro sobre las lombrices de tierra (Darwin 1881). Se le considera el padre de la ciencia del suelo. También contribuyó al entendimiento de la geología en los trópicos al descifrar el papel de los arrecifes de coral en la formación de las islas del Pacífico.
2. Andreas Franz Wilhelm Schimper. Botánico alemán que inició el uso del concepto de Rain Forest o bosque pluvial y escribió el libro Plant Geography upon a Physiological Basis (Schimper 1903) considerado un clásico que resumió el conocimiento hasta el final del siglo XIX, pero a la vez presentó nueva información y visión fisiológica del estudio de la biota.
3. Víctor Ernest Shelford. Zoólogo norteamericano, padre de la ecología animal norteamericana y reconocido pionero del campo de la ecología (Croker 1991). Junto a Frederic Clements inició el uso del concepto de «Bio-Ecología» o lo que se considera un intento por unir la ecología animal y la de plantas (Clements y Shelford 1939). Estableció la ecología fisiológica para animales, lo que incluye la Ley de Tolerancia de los organismos, la cual reconoce que hay límites ambientales en las adaptaciones de los organismos a su entorno.
4. Frederic Edward Clements. Ecólogo norteamericano. Se le considera el ecólogo más influyente en el desarrollo del concepto de la sucesión ecológica, del
1 16 CAP ariel e. lugo | ecología a m i m anera Grandes ecólogos
pensamiento de la comunidad como un superorganismo y la idea del mono clímax. Con sus muchos libros impulsó una visión dinámica a la ecología norteamericana en contraste con la ecología más estática enfatizada en el continente europeo. Enfatizó la idea de «ecesis» para entender la sucesión de plantas sobre distintos substratos. Su trabajo es fundamental para entender el éxito o fracaso de las restauraciones de áreas degradadas, tan comunes en el presente. Con su esposa Edith, una ecóloga profesional reconocida en su propio mérito, viajaron cientos de miles de millas estudiando comunidades vegetales (E. Clements 1960).
5. Henry Allan Gleason . Ecólogo norteamericano que en 1926 con Melville Thurston Cook, vicedirector de la Estación Experimental Agrícola de Puerto Rico en Río Piedras, publicó Plant Ecology of Porto Rico. Gleason es mejor conocido por su rechazo a las ideas del mono clímax de Clements a favor de comunidades de clímax múltiples que no se comportan como superorganismos (Gleason 1926). Favoreció el concepto de la asociación individualista de plantas. Las asociaciones surgen al azar influenciadas por el ambiente físico y de acuerdo con las características de cada especie.
6. Robert Hardin Whittaker. Ecólogo norteamericano quien documentó que la distribución de especies obedece a la presencia de gradientes ambientales. Para Whittaker, el clímax es un patrón poblacional que obedece al patrón de variables ambientales. Relacionó los conceptos de comunidades a los de ecosistemas y contribuyó al desarrollo de muchas ideas sobre la organización de comunidades, incluso las curvas del Valor de Importancia (Whittaker 1970).
7. Sir Arthur George Tansley. Botánico británico a quien se le encargó resolver la controversia entre las distintas escuelas de pensamiento relativas a la organización y funcionamiento de la comunidad. En el proceso se le acreditó haber presentado el concepto del ecosistema (Tansley 1935). Nunca más publicó trabajos utilizando el concepto.
8. Frank Edwin Egler. Un ecólogo consultor norteamericano con nuevas ideas acerca de la zonificación y funcionamiento de las comunidades de plantas. Fue rechazado por la comunidad académica. Pudo descifrar la zonación de los pantanos salinos de la Florida (Egler 1952) y la función ecológica de las especies introducidas en Hawái décadas antes que sus pares (Egler 1942).
9. Charles Sutherland Elton . Zoólogo inglés que desarrolló la idea de «BioEcología» en Inglaterra y escribió lo que se considera el trabajo clásico relativo
17 1 CAP ariel e. lugo | ecología a m i m anera Grandes ecólogos
Figura
1.1. Genealogía académica de G. Evelyn Hutchinson, profesor de la Universidad de Yale y consejero de notables ecólogos, incluyendo a Howard Thomas Odum. Los nombres incluidos se restringieron a los graduados de doctorados y posdoctorales antes de 1971. Detalles en Egerton (2015).
ProctorV.
1 18 CAP ariel e. lugo | ecología a m i m anera Grandes ecólogos G.E . Hu tc hin so n E.S. Deevey D.A. Livingstone J.L. Richardson M.S. Darling R.L. Kendall R.R. Peterson P.D. Vergl R.E. Hecky P. Kilham M.A. Mantuani H. Ignatius U. Cowgill J. Shapiro R.H. Macarthur P.J. Wangersky E. Mills J. Gilbert R. Doyle F.M. Williams H.O. Werntz I.A. Mclaren S.R. Brown M. Warburg P.H. Klopfer A.J. Kohn R. Benoit H.T. Odum L.B. Slobodkin W. D. Hartman F.E. Smith I.M. Newell D.J. Zinn J.L. Brooks V.T. Bowen J.R. Vallentyne T.F. Goreau E. Leopold J.G. Ogden
Colinvoux G.W. Comita G.C. Anderson S.L. Dodson J.G. Stockner G.J. Lauer C.M. Falter H.L. Sanders A.G. Carey R.E. Ha ner G.A. Bertrand D.C. Gordon H.A. Donaldson E.H. Wheeler S. Mirow G.V. Levin J. Eaton W. Kolman T.M. Allen R.C. Cooke D.D. Adams S.R. Kerr M.J. Bozin C. Kellog A. Covich R.A. Kinzie R.H. Staun E.G. Leigh P.F. Brodie R. Daley G. Gottlieb R. Northen J. Hailman A. Lall R. Hemmes B. Gilbert R. Pulliam D. MacDonald T. Harrington G. Milikan L. Rosenson J. Hatch L. Kepner A.C. Borror C.E. King C. Hall L. DiSalvo M. Canoy J. Ewel R. Kelley F. Little R. Colwell J. Armstrong R. Caldwell M. Rankin C. Lynch R. Lynch S. Saul C. La France H. Dingle C. Istock T. Gri ng D. Futuyma R.T. Paine D. Hall N. Woldow M.F. Cohn D.W. Tappa J. Jacobs J.S. Su ern S.J. Warshaw T.M. Zaret N.R. Andersen T.T. Stonier L.J. Friesen W. Alley B. Carlasa G.N. Oldfield J.TT. Crowe W. Cooper Q. Ross D.M. Johnson L. Leux A. McClary R. Elsenberg A. Wenner D.L. Johnson J.M. Emlen W. Platt D. Arnold G. Gallapin I. Valiela V. Kaczynski J.O’ Brien K.P. Mauzey B.A. Menge C.E. Birkeland P.K. Dayton R.R. Vance T.S. Spight U. Ritte S. Richman T.L. Simpson J.C. Lang J.G. Hechtel R.K. Pang G. Vermeij H.M. Relswig E.M. Hadden Y. Loya S. Nixon R. Beyers R. Wilson T. Hellier P. Roe N.R. Sinclair S.D. Ludlam G. Brunskill L. Pollock D.L. Luchtel R. Strathmann C. Lalli G.E. Beunlands D.C. Grant T.E. Lovejoy M.Blaug D.D. Someoto R. Pocklington K.L. Crowell M. Berril P. Regal E. Maly E.G. Horn W.G. Sprules M.L. Rosenzweig M.L. Cody R. Bernstein E. A. Wood J.F. Hebard R.M. Laurs R.J.Conover W.G. Pearcy G.S. Posner T. Napora C.MacPhee L.W. Huthchins R. Lindeman E. Bonatti J.
A. Frank W.T. Edmonson G.A. Riley S.Hsiao
C.Goulden
P.A.
Armstrong
M.Tsukada
A.
H. Loe er G.A. Longlois E. Cutler W.R. Cobb J.
Ecología a Mi Manera FIG. 1.1
Palmer
Bradbury
a las especies invasoras2 (Elton 1958). A pesar de presentar una situación tétrica sobre especies invasoras, reconoció la inevitabilidad de las invasiones y propuso conceptos ecológicos para apoyar una nueva modalidad de conservación donde las especies colonizadoras ocupan espacios legítimos. Elton fue el primero en observar y explicar las fluctuaciones en el número de individuos en poblaciones animales. Las pirámides de biomasa y número de organismos en comunidades se le atribuyen a Elton.
10. Aldo Leopold. Un dasónomo norteamericano con ideas ecológicas que trabajaba para el Servicio Forestal de los EE. UU. antes de que el concepto de ecólogo fuera popular en la agencia. La enciclopedia Wikipedia lo designa como autor, filósofo, naturalista, científico, ecólogo, dasónomo, conservacionista y ambientalista. Escribió muchos artículos que unidos comprenden nuevos paradigmas prácticos sobre la ética en el uso de la tierra y de su conservación. Un ejemplo es el famoso A Sand County Almanac and Sketches Here and There (Leopold 1949). Integró y expandió las ideas de Pinchot y Muir. Se le acredita el concepto de wilderness dentro del gobierno federal y fundó el Wilderness Society al igual que la nueva ciencia de game management (Meine 1988).
11. George Evelyn Hutchinson. Ecólogo británico asociado con el desarrollo de la ecología moderna. Escribió A Treatise on Limnology, una obra masiva en cuatro volúmenes (Hutchinson 1957, 1967, 1975 y 1993). Visto como la máxima autoridad en ese campo de la ciencia, por lo que es considerado el padre de la limnología en los EE. UU. Contribuyó al desarrollo de la biogeoquímica. Fue profesor y consejero de grandes figuras de la ecología (Fig. 1.1) tales como el norteamericano Raymond Laurel Lindeman (escribió sobre los niveles tróficos en los ecosistemas), el canadiense Robert Helmer MacArthur (escribió sobre ecología de poblaciones), el norteamericano Edward Smith Deevey Jr. (paleolimnólogo que escribió sobre la biogeoquímica de lagos), el norteamericano Lawrence Basil Slobodkin (escribió sobre la ecología de poblaciones) y el norteamericano Howard Thomas Odum (ver más adelante en este capítulo). Hutchinson también desarrolló el concepto del nicho como un hipervolumen y escribió ensayos muy importantes sobre evolución y el número de especies en sistemas acuáticos.
12. Leslie Ransselaer Holdridge. Botánico y climatólogo norteamericano y el primer científico que laboró para el Instituto Internacional de Dasonomía Tropical
2 El uso del término «especies invasoras» ha adquirido connotaciones negativas y normativas en contra de especies introducidas. Para evitar términos normativos, utilizaré «especies colonizadoras».
19 1 CAP ariel e. lugo | ecología a m i m anera Grandes ecólogos
en Puerto Rico. Holdridge estableció la biblioteca y el herbario del Instituto y comenzó la síntesis para describir los árboles de Puerto Rico. Basado en sus experiencias en Puerto Rico y Haití, desarrolló el concepto de zonas de vida (Holdridge 1947, 1967). Se consideraba a sí mismo un cosmólogo.
13. Crawford Stanley Holling. Ecólogo canadiense que desarrolló las ideas de resiliencia (Holling 1973) y de panarquías (Gunderson y Holling 2002). Las panarquías son dinámicas y adaptables al cambio ambiental contrario a las jerarquías. La resiliencia es la capacidad de un sistema para absorber la energía de un disturbio sin cambiar su estado y manteniendo su funcionamiento. Adaptó estas ideas a la conservación de ecosistemas, incluyendo sistemas antropogénicos.
14. Eugene Pleasants Odum. Ecólogo norteamericano, graduado de la Universidad de Chicago, donde se originó el concepto de la ecología norteamericana. Cuando comenzó su carrera en la Universidad de Georgia, su jefe de Departamento le dijo que la ecología no era una ciencia porque no contaba con libros de texto. Eso estimuló a E.P. Odum a escribir el primer libro de texto de ecología (E.P. Odum 1953). Eugene Odum fue un gran comunicador y educador, desarrolló y dirigió el programa de ecología en la Universidad de Georgia. Enfocó su atención en la estructura y funcionamiento del ecosistema, integrando todos los tipos de ecosistemas bajo una serie de principios generales.
15. Howard Thomas Odum (de ahora en adelante Odum). Un innovador ecólogo norteamericano que formó muchos ecólogos gracias a su genio y dotes
1 20 CAP ariel e. lugo | ecología a m i m anera Grandes ecólogos
como educador (Fig. 1.2). Fue responsable de muchas de las nuevas ideas y estudios en el campo de la ecología moderna: economía ecológica, ingeniería ecológica, ecosistemas noveles (Odum 1962), ideas sobre el valor relativo de la energía expresada en equivalentes solares y conocida como EMERGY, el principio de máxima potencia, ecología cuantitativa de ecosistemas en los bosques
21 1 CAP ariel e. lugo | ecología a m i m anera Grandes ecólogos
Figura 1.2. Genealogía y carrera académica de Howard Thomas Odum. Con base en las anotaciones de H.T. Odum.
y mangles de Puerto Rico, arrecifes de coral en el Pacífico, bahías costeras en Texas, estudio clásico en Silver Springs Florida, la geoquímica del fósforo en la Florida, el flujo de energía y productividad de ecosistemas, diagramas de flujo de energía, etc. Fue un pionero en estudios de ecología a gran escala y dentro del enfoque que Coleman (2010) denominó Big Science. Sus aportes al conocimiento ecológico en los trópicos, incluyendo a Puerto Rico, han sido resumidas (Lugo 1995a,b,c, 2003, 2004a).
Otros científicos surgieron durante las discusiones con los estudiantes:
16. Eugene Warming. (Johannes Eugenius Bülow Warming) nacido en 1841 y fallecido en 1924. Era botánico y es considerado uno de los fundadores de la ecología (Goodland 1975). Entre otras ocupaciones, trabajó como profesor de botánica en la Universidad de Copenhague y en el Jardín Botánico de la misma ciudad. Le gustaba la taxonomía, fisiología y morfología de plantas y enseñó la primera clase de ecología en una universidad. Hacía excursiones con los estudiantes para mostrar cómo las plantas se relacionaban y se adaptaban al ambiente. Escribió varios libros en el ámbito universitario y elemental. Hizo diversas excursiones por el mundo, incluyendo países de Europa y de los trópicos, como Brasil y Venezuela. Con el material de sus observaciones por el mundo escribió el primer libro sobre ecología de plantas: Oecology of Plants (Plantesamfund en danés) en 1895 (Coleman 1986), que fue traducido al alemán el año siguiente y al inglés en 1909. El libro fue innovador por no ser apenas una descripción botánica, sino por relacionar la distribución de plantas de acuerdo con condiciones abióticas. Defendía que las características abióticas eran más importantes que los factores bióticos, como la especiación, sobre la distribución y evolución de las especies de plantas. Con su obra influyó en grandes nombres de la ecología, como Arthur Tansley (Tansley 1947), Henry Cowles (Coleman 1986, Cassidy 2007), Andreas Schimper y Christen Raunkiær. Por Jéssica Fonseca da Silva
Naturalista y
alemán que en una expedición realizada entre 1799-1804, coleccionó, junto a otros, 60,000 especímenes de plantas en centro y sur América. Publicó 30 volúmenes en los que describió la flora y fauna de América tropical. Correlacionó los tipos de vegetación con las características ambientales, y acuñó el término asociación. Además, llamó la atención respecto a la necesidad de observaciones sistemáticas de la vegetación, influyendo grandemente en los procedimientos de muestreo de la época. Por Sandra Ayala
1 22 CAP ariel e. lugo | ecología a m i m anera Grandes ecólogos
17. Friedrich Heinrich Alexander von Humboldt (1769-1859).
geógrafo
18. Howard Thomas Odum. Uno de los científicos ecólogos más influyentes del siglo XXI, nacido en Chapel Hill, Carolina del Norte en 1924 y tercer hijo del famoso sociólogo estadounidense Howard W. Odum. Howard Thomas Odum es conocido como un pionero en la ecología de ecosistemas, por sus innovadores trabajos en la teoría general de sistemas. En uno de sus trabajos, titulado Energy, Ecology and Economics, Odum relaciona tres vertientes que otrora serían considerados antagonistas: la energía, la ecología y la economía. Escrito en 1973, este artículo se adelanta a su tiempo y previene sobre los problemas de depender energética y socialmente de los combustibles fósiles. Utilizando su sistema de lenguajes explica cómo la inflación económica mundial es causada por el aumento en el uso de energía para conseguir más energía. Habla sobre el ambiente como un aliado en la conservación energética y cómo la utilización de tecnología solar es ineficiente, ya que no compite con el trabajo solar en la agricultura, en los bosques y en otras formas ancestrales de utilización. Odum también advierte sobre un crecimiento inmediato de la tecnología mundial que se adelantará a los procesos energéticos de la época. Considera que el afán de los gobiernos por promover el crecimiento tecnológico, y por ende económico de un país, será contraproducente y exhorta a regresar a un «estado constante» donde la sociedad encontrará estabilidad y satisfacción individual.
“Worldwide inflation is driven in part by the increasing fraction of our fossil fuels that have to be used in getting more fossil and other fuels.”
Odum 1973, p. 220-227
Aunque no estoy de acuerdo con ciertos puntos de su artículo, tengo en mente que se escribió en la década de los setenta y que muchos de los eventos que predijo ocurrirían en un futuro no tan lejano, como el crecimiento exponencial de la tecnología, ocurrieron en un lapso relativamente corto. También concuerdo en que no se puede depender de una sola fuente de energía, como los combustibles fósiles, sino que (y en esto difiero con Odum) deberíamos buscar otras fuentes de energía como la energía solar, el gas natural o la energía hidráulica. Fuentes energéticas que sean ambientalmente viables y que a la larga provean una economía a la sociedad. Concurro además con la idea de que el medio ambiente no es el enemigo de la energía, sino un gran aliado. El agua, los aceites naturales y hasta el aire nos proveen energía sin necesidad de excavar pozos en búsqueda de petróleo y a su vez nos brindan las herramientas para desarrollar un sistema de energía sustentable. Aunque la idea de permanecer por un tiempo en un estado constante sería de alguna forma beneficiosa para nuestra sociedad, entiendo que no es viable porque hoy en día nos regimos por los adelantos tecnológicos
23 1 CAP ariel e. lugo | ecología a m i m anera Grandes ecólogos
y es casi imposible ponerle un alto a la ciencia y a la tecnología. Por Estrella D. Santiago Pérez.
19. Patrick Holt Leslie (1900-1974). Estudió fisiología en Oxford. En 1935 se unió al Bureau of Animal Population en Oxford, mientras era dirigido por Charles Elton. Trabajó en el mismo hasta su retiro en 1968. Entre sus grandes obras como ecólogo se destacan realizar la primera aplicación de la ecuación de Lotka de la tasa de crecimiento intrínseco de una población animal (Leslie y Ranson 1940) y el desarrollo de los modelos poblacionales basados en matrices. Es por esto por lo que, según Caswell (2001), se considera el científico que inventó el rol del ecólogo matemático contemporáneo. Por Raisa Hernández Pacheco
20. Jean-Baptiste Lamarck (1744-1829) fue un médico, naturalista, biólogo, botánico, zoólogo y en mi opinión uno de los ecólogos más brillantes de todos los tiempos. A pesar de que murió en la miseria, sus trabajos y aportes a la ciencia siguen siendo de admiración y uso en la actualidad. Lamarck nació en Bazantin, Francia, en el seno de una familia militar. Sin embargo, por problemas de salud tuvo que dejar el ejército y así abordó su interés por la ciencia. Entre sus aportes más importantes están: (1) la clasificación de la flora mediante claves dicotómicas que actualmente se usan tanto en la botánica como en la zoología, (2) el término de «Biología», y (3) la primera teoría de la evolución biológica. Además, se encuentran sus importantes contribuciones con la paleontología de invertebrados, la meteorología y la física de la Tierra. Entre sus trabajos, posiblemente el más revolucionario y controversial de la época fue el de la evolución. Este planteaba una adaptación de los organismos a los ambientes cambiantes y por consiguiente la modificación de su estructura conforme el ambiente iba cambiando. Lamarck hace referencia a la evolución de los organismos a través de su vida, y no así de su origen, ya que se encontraba en una época en la que se creía fuertemente en la generación espontánea. Así como Darwin justificó su teoría con la existencia de fósiles de especies intermedias, Lamarck postuló esa posibilidad 50 años antes. A pesar del mínimo reconocimiento que obtuvo en su época, los trabajos de Lamarck, especialmente sobre la teoría de la evolución, fueron base esencial para el desarrollo de la ciencia en general y claramente de la relación del ser vivo y su ambiente, que ahora se desarrolla más profundamente en la ecología. Por Daniela Yaffar.
21. En Puerto Rico, consideramos a Agustín Stahl Stamn como el primer científico local con reputación internacional (Rivero 2008). Aunque nació en Curazao de padres no puertorriqueños, se crió en Aguadilla, vivió y trabajó en Bayamón y se auto consideró puertorriqueño. Fue médico, pero desarrolló la primera flora
1 24 CAP ariel e. lugo | ecología a m i m anera Grandes ecólogos
de Puerto Rico y promovió la historia natural de la Isla cuando nadie pensaba en esas cosas.
22. Ana Roqué de Duprey fue quizás la primera puertorriqueña científica (Foto). Nació en Aguadilla y murió en Río Piedras. Fue estudiante de Agustín Stahl y escribió Botánica antillana, una obra extensa sobre las plantas del Caribe, incluyendo a Puerto Rico. Laboró en esa publicación por casi treinta años (1906 a 1933 cuando murió), pero no logró que se publicara pues los académicos de la época no la apoyaron (lea los detalles en http://periodismoinvestigativo.com/2015/02/ el-tesoro-de-una-cientifica-rebelde/ ).
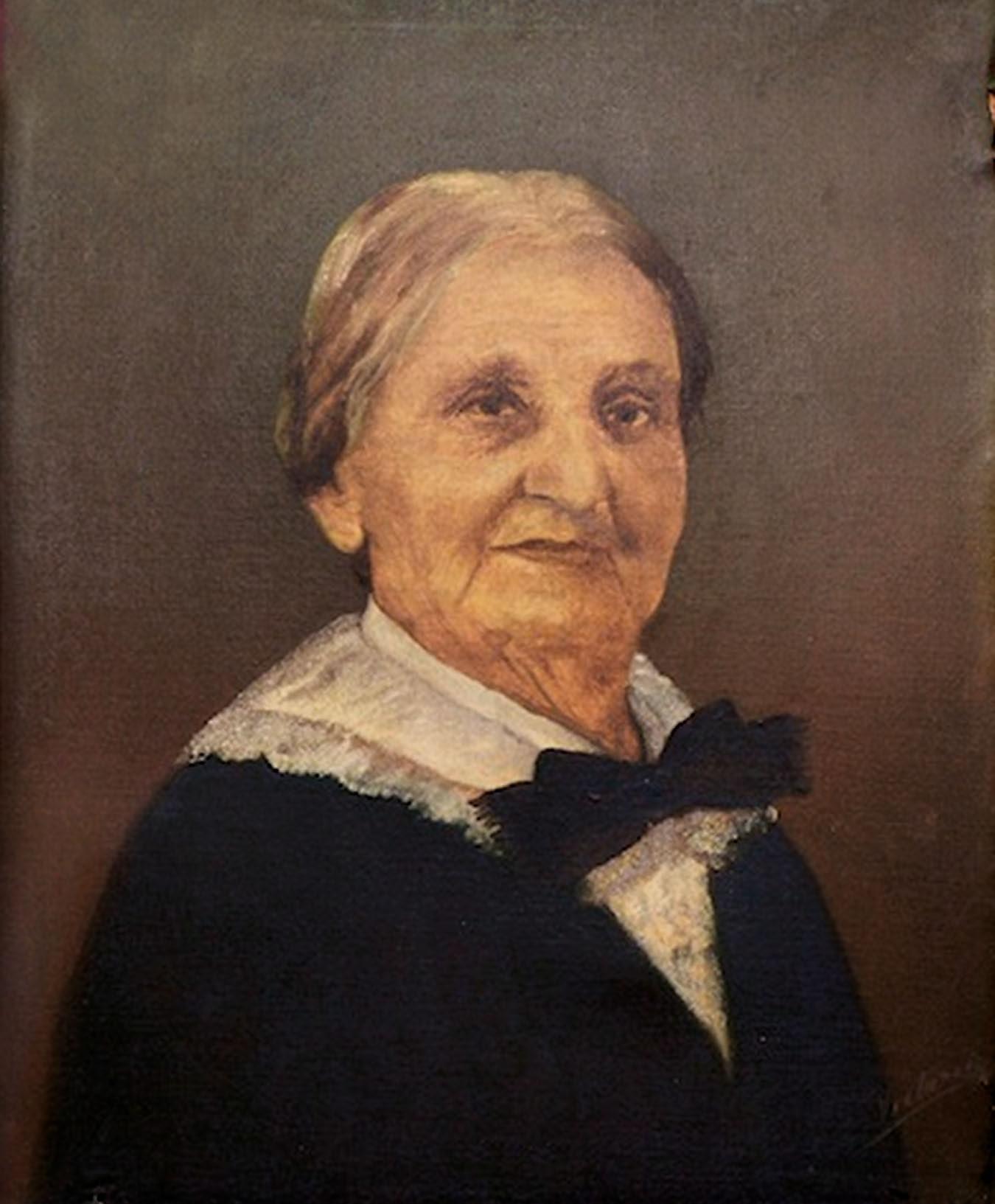
25 1 CAP ariel e. lugo | ecología a m i m anera Grandes ecólogos
Foto por Juan Costa | Centro de Periodismo Investigativo Ana Roqué de Duprey, por el pintor López de Victoria. Retrato de la colección de la Corporación de las Artes Musicales.
Wei Wu
Whendee Silver
Tana Wood
Daniela Cusack
Jennifer Pett-Ridge
Nubes Lluvia H2O et
Olga Mayol
Epifitas y otras plantas
Suelo
Sharon Cantrell Jean Lodge
Elizabeth Britton
Inés Sastre
Barbara Richardson
JoAnne Sharpe
Amelia Merced
Nancy Harris
Sandra Brown
Whendee Silver
Erika Marín Spiotta
Hongos Macroinvertebrados Microorganismos
Grizelle González
Carmen J. Hernández
Animales
Vegetación riparia
Ríos
Kristina Vogt
Karen Beard
Tanya Matlaga
Patricia Burrowes
Migdalia Alvarez
Tamara Heartsill
Elvira Cuevas
Kathy Pringle
Elfie Greathouse
Kathryn Clark
Estuario
Álida Ortiz Sotomayor
Rt
Investigación relacionada:
Heather Erickson
Diana García Montiel
Melanie Lenart
Rebecca Ostertag
Jill Thompson
Sheila Ward
Elvia Meléndez Ackerman
María Uriarte
Martha Scholl
Heather Buss
Jane K. Willenbring
Mei Yu
Molly A. Cavaleri
Sasha C. Reed
Kathleen McGinley
Benedicte Bachelelot
Kelsey Carter
Christine O’Connell
Chris Smith-Martin
Daniela Yaffar
23. Elizabeth Britton quizás fue la primera mujer científica extranjera en Puerto Rico. El Monte Britton en El Yunque se nombró en honor a ella y su esposo Nathaniel Lord Britton, quien dirigió la expedición sobre la Historia Natural de Puerto Rico de la Academia de Ciencias de Nueva York y la Universidad de Puerto Rico. La Dra. Britton era experta en las briófitas. Subsiguientemente, las mujeres científicas, tanto nativas como extranjeras han contribuido significativamente al conocimiento ecológico de El Yunque como se resalta en la figura 1.3.
24. Muchos científicos puertorriqueños han contribuido con el avance de la ciencia y la conservación, sin embargo, no es mi intención mencionarlos a todos. Solo quiero destacar a dos figuras fuera de la ciencia que influenciaron la conservación en Puerto Rico cuando tal actividad no era tan reconocida como lo es
1 26 CAP ariel e. lugo | ecología a m i m anera Grandes ecólogos
Figura 1.3. Ciclo de agua y carbono en los bosques de las Montañas de Luquillo enfatizando la contribución de mujeres científicas que han trabajado es esas montañas y que han culminado sus estudios doctorales. La evapotranspiración es «et », PPG es la productividad primaria bruta y R t es la respiración total del sistema. Grizelle González y Tana Wood contribuyeron con información para esta figura.
Nancy Harris Ashley Van Beusekom Sheila Murphy Nancy Harris
PPG
SOL CO2
CO2
hoy. Clara Livingston, primera aviadora en la Isla y amiga de Amelia Earhart (quien salió de Puerto Rico en su trágico vuelo alrededor del mundo), conservó el bosque de Dorado (Figueroa et al. 1984) a principio del siglo XX, cuando el gobierno en ese momento pretendía desarrollarlo.
25. José Mariano García , pintor impresionista nacido en Utuado, Puerto Rico. Salvó el puente sobre el río Piedras en la Estación Experimental, utilizando sus pinturas para evitar que el gobierno lo demoliese en la década de 1930 (Álvarez 1987-1988). Este puente data de 1853, es el más antiguo en la carretera de San Juan a Ponce y hoy es el símbolo de los estudios social-ecológicos-tecnológicos de San Juan. El cuadro original del pintor está en el lobby del Instituto Internacional de Dasonomía Tropical en el Jardín Botánico de la Universidad de Puerto Rico en Río Piedras (Foto).

27 1 CAP ariel e. lugo | ecología a m i m anera Grandes ecólogos
Foto por Ariel E. Lugo Puente de la Estación Experimental (Norzagaray), óleo de José Mariano García.
CaPítulo 2
El ecosistema: unidad de la vida en el planeta
De acuerdo con Neil A. Campbell (1993), la vida tiene las siguientes características: orden, reproducción, crecimiento y desarrollo, utilización de energía, respuesta al ambiente, homeostasis y adaptación evolutiva. En un momento dado se enseñaba que la unidad de la vida en el planeta era la célula. Pero esta apreciación representa una visión reduccionista. Una célula no puede sobrevivir sin el organismo y el organismo sin el sistema que lo alberga. La unidad de la vida debe buscarse a una escala mayor de funcionamiento ecológico. El ecosistema, según lo voy a definir más adelante en este capítulo, contiene todos los elementos necesarios para sostener la vida indefinidamente siempre y cuando reciba insumos de energía potencial (luz solar en el caso de los sistemas del planeta Tierra). La unidad de la vida en el planeta es el ecosistema.
El concepto de ecosistema es parte de las grandes ideas explicativas de la ciencia de acuerdo con Sterner y Elser (2002). Al ecosistema, ellos le suman la selección natural, la tabla periódica de los elementos, la conservación de materia y energía, retroalimentación positiva y negativa y el dogma central de la biología molecular. Este conjunto de ideas observa los sistemas ecológicos y la vida en el planeta de una forma integrada y regida por leyes fundamentales de la física, química y biología.
El dogma central de biología molecular, o su hipótesis central, no se va a enfatizar en este curso, pero fue propuesto por Francis Crick (1970):
“The central dogma of molecular biology deals with the detailed residue-by-residue transfer of sequential information. It states that information cannot be transferred from protein to either protein or nucleic acid” (p. 561).
Crick presentó su idea en un simposio internacional en 1956 indicando:
“Once information has got into protein, it can’t get out again” (Fig. 1 en Cobb 2017).
Esta idea explica la transferencia de información genética en los sistemas biológicos. Los ácidos nucleicos tienen más información que las proteínas y requieren más
ariel e. lugo | ecología a mi ma nera
29
energía para sintetizarse, por lo que le pueden transferir información a las proteínas, pero el flujo de información no puede operar en la dirección contraria, o sea, de bajo contenido de información a alto contenido de información. Esta hipótesis central reafirma la base energética de la biología, ya que hay una relación directa entre el flujo de información entre las moléculas durante el intercambio de información genética y el flujo de energía (Odum 1983).
Aquí enfatizo las leyes de energía por su relevancia sobre el funcionamiento de los ecosistemas: (1) la ley de conservación de masa y energía, (2) la ley del aumento en la entropía, y (3) la ley de potencia máxima. La ley de conservación de masa y energía establece que la masa y la energía, aunque pueden transformarse, no se destruyen. Dicho de otra forma, la cantidad de masa y energía en el universo es constante. Esta ley permite el balance de reacciones químicas y el estimado de los presupuestos o balances geoquímicos de elementos como el carbono, el agua o los nutrientes. La ley del aumento en la entropía establece que la entropía en el universo tiende a aumentar. Dicho de otra forma, ningún uso de energía es cien por ciento eficiente. Cada vez que se realiza un trabajo y se utiliza energía, una porción contribuye al trabajo útil, mientras que otra se disipa como calor o entropía. Entropía es sinónimo de desorden. El desorden siempre aumenta cuando hay un consumo de energía o cuando la energía deja de fluir. La energía se subdivide en la energía potencial o disponible para realizar un trabajo, la energía cinética (la que permite que se realice el trabajo) y la entropía, la cual no tiene capacidad para realizar un trabajo.
La ley de potencia máxima relaciona las leyes de energía con la evolución y la sucesión ecológica, entre otras aplicaciones. Esta ley está basada en el trabajo de 1924 del biofísico norteamericano Alfred James Lotka el cual se reimprimió en 1956 (Lotka 1956). Él sugirió que esta idea constituía la cuarta ley de termodinámica y dijo que las leyes de termodinámica indican lo que no se puede hacer, pero esta cuarta ley explica cómo se hace. Establece que durante la autoorganización de sistemas, aquellos diseños que se generan y prevalecen son los que maximizan la potencia (el metabolismo o flujo de energía), la transformación de energía y aquellos usos que refuerzan la producción y la eficiencia (Odum 1995, p. 311). Odum y Pinkerton (1955) observaron que en una transformación de energía potencial ( free energy) a energía cinética, la máxima potencia ocurre cuando la eficiencia de uso es el 50 %. A eficiencias más bajas o altas, la potencia que se genera es menor. Bajo condiciones similares, aquellos procesos biológicos y ecológicos con la máxima potencia son los favorecidos por la selección natural.
La energía se define como la capacidad de realizar un trabajo. Se mide en unidades de calorías o gram calorie, aunque en el Sistema Internacional de Unidades se utiliza el «julio» ( joule) (1 gram calorie es igual a 4.2 julios). Un gram calorie se define como la cantidad de energía requerida para aumentar la temperatura de un gramo
2 30 CAP ariel e. lugo | ecología a m i m anera El ecosistema: unidad de la vida en el planeta
o un centímetro cúbico de agua, un grado centígrado cuando el agua está a 15 grados centígrados y bajo una atmósfera de presión.
Tabla 2.1
Equivalencias solares de varios tipos de energía tomado de Odum y Odum 2001.
No todas las calorías tienen la misma capacidad de realizar un trabajo. Algunas, no pueden hacerlo porque ya están disipadas como entropía. Por otro lado, una caloría de madera realiza más trabajo que una caloría solar y una de petróleo más que cualquiera de las anteriores. La diferencia se debe al grado de concentración de energía potencial en el material. Mientras más concentrada la energía, más trabajo útil por caloría puede ocurrir. Sin embargo, más tiempo, trabajo y energía son necesarios para producir unidades con alta concentración calórica. Esto se ilustra en la tabla 2.1 con las equivalencias solares de distintos materiales y procesos. La equivalencia solar indica cuantas calorías solares son necesarias para producir una caloría del material o proceso indicado, o sea, 4,400 calorías solares para producir una de madera en el suelo del bosque. Fíjese que los valores más altos corresponden a la información, incluyendo la información en el material genético. La formación de todos los materiales y la potencia para operar todos los procesos en el planeta, están relacionados con la energía solar.
La constante solar (que no es constante) es aproximadamente 1.951 gram calories por centímetro cuadrado de superficie planetaria por minuto. Todos los sistemas sociales, ecológicos y tecnológicos del planeta Tierra funcionan con esa energía solar, más la energía de calor residual en el centro o núcleo del planeta. En la figura 2.1, se ilustra un perfil del planeta acentuando el acoplamiento entre la atmósfera y la litosfera. Los ciclos de rocas, agua y elementos como el carbono representan los ciclos biogeoquímicos del planeta que a su vez establecen las condiciones bajo las cuales funcionan los sistemas ecológicos. El conjunto de estructuras y funcionamiento en este perfil se conoce como la biosfera, o el espacio que sostiene a los seres y sistemas vivos del planeta.
Homeostasis es el conjunto de fenómenos de autorregulación conducentes al mantenimiento de una relativa constancia en la composición y las propiedades del medio interno de un organismo o del ecosistema. La retroalimentación, ya sea
2 31 CAP ariel e. lugo | ecología a m i m anera El ecosistema: unidad de la vida en el planeta
Tipo
Energía solar 1 Energía eólica 1,500 Materia orgánica (madera, en el suelo) 4,400 Potencial de agua de lluvia elevada 10,000 Energía química de agua de lluvia 18,000 Energía mecánica 20,000 Energía de un río caudaloso 40,000 Energía fósil 50,000 Comida 100,000 Energía eléctrica 170,000 Alimentos de proteínas 1,000,000 Servicios humanos 100,000,000 Información 100,000,000,000 Formación de una especie (evolución) 1,000,000,000,000,000
de caloría Equivalente solar por caloría
FORMACIÓN DE NUBES
Es más abundante sobre la tierra
Lluvia
Ciclo hidrológico
Océano
Erosión
Levantamiento volcánico
Transportación de sedimentos por ríos
Fisura
Desplazamiento
Masa continental compuesta de rocas y sedimentos de menor densidad de rocas profundas
Levantamiento
Rocas muy calientes (lava)
Deposición de sedimentos ?
Leyenda
(modificado de Odum 1971).
positiva o negativa, contribuye a mantener la homeostasis de los sistemas. Una retroalimentación positiva acelera el sistema, y si se prolonga, crea inestabilidad. Por ejemplo, A produce B y B produce A. Este es un sistema inestable, pues se acelera la multiplicación de las «A». La retroalimentación positiva es útil para crecer rápido cuando las condiciones lo permiten. En la retroalimentación negativa, el producto del sistema reduce el insumo. Un ejemplo es el control de temperatura en el cuerpo humano. Cuando aumenta la temperatura, el sistema automáticamente suda y al evaporarse el sudor la temperatura baja. Un termostato tiene retroalimentación negativa. Reacciona a la temperatura en forma contraria y la mantiene estable.
¿Cuántos ecosistemas hay en la figura 2.2? La contestación es que hay tantos ecosistemas como quisiéramos definir, o sea, el número es ilimitado. Los límites del
no son límites estructurales o fisonómicos. El ecosistema es un concepto funcional y la definición que vamos a usar es la de Evans (1956 p. 1127):
2 32 CAP ariel e. lugo | ecología a m i m anera El ecosistema: unidad de la vida en el planeta
ecosistema
«En sus aspectos fundamentales un ecosistema envuelve la circulación, transformación y acumulación de materia y energía a través del medio de los seres vivos y sus actividades».
Figura 2.1. Perfil del planeta Tierra que ilustra sus características geomorfológicas y sus tres grandes ciclos biogeoquímicos: el ciclo de rocas, el ciclo de agua (gaseoso) y el ciclo sedimentario. Las casillas en el fondo del océano representan las placas tectónicas
lateral del suelo oceánico
522
oceánica Fisura en suelo del océano
OcéanoRocas de baja densidad Rocas pesadas y en localidades profundas
Evaporación
SOL
?
Ciclo de rocas
Ciclo de rocas caracterizados por sus rocas de menor densidad.
Esta definición resalta que el concepto de ecosistema es un concepto funcional con límites arbitrarios. El concepto lo que refleja es el funcionamiento del grupo biótico y abiótico que arbitrariamente seleccionemos como ecosistema.
La idea del ecosistema nos permite explorar preguntas que no se plantean en la idea de la evolución. G. E. Hutchinson (1965) observó que en el teatro ecológico se lleva a cabo la trama de la evolución (discutido en el capítulo 4). La ecología y la evolución están fuertemente entrelazadas y cada una influencia a la otra. Pero a pesar de estar estrechamente ligadas, cada una persigue aspectos distintos en la obra del desarrollo y funcionamiento de los sistemas ecológicos. El concepto del ecosistema facilita el entendimiento de la funcionalidad de sistemas a distintas escalas. Le da más énfasis al papel que juega el ambiente físico en el funcionamiento de los sistemas ecológicos. La biología tradicionalmente le presta más atención a los aspectos bióticos y no tanto a los abióticos. La siguiente

definición del ecosistema (p. 8 en Golley 1993):
“But the more fundamental conception is, as it seems to me, the whole system (in the sense of physics), including not only the organism-complex, but the whole complex of physical factors forming what we call the environment of
2 33 CAP ariel e. lugo | ecología a m i m anera El ecosistema: unidad de la vida en el planeta
cita de Tansley (1935, p. 299) captura su
A B C D E F G H J I
Figura 2.2. Ilustración de los límites arbitrarios para delimitar ecosistemas. Se ilustran 10 sistemas (A - I) pero se pueden demarcar muchos más. Cada caja representa una escala distinta del sistema, desde la escala mayor (A) a escalas menores como (I) y (E). Noten que dentro de (G) se delimitan dos sistemas (H) y (J). Se representan sistemas acuáticos (F), terrestres (B) y antropogénicos (I). Basado en Odum (1971).
the biome-the habitat factors in the widest sense. It is the systems so formed which, from the point of view of the ecologist, are the basic units of nature on the face of the earth. These ecosystems, as we may call them, are of the most various kinds and sizes. They form one category of the multitudinous physical systems of the universe, which range from the universe as a whole down to the atom.”
Es útil pensar en términos de jerarquías cuando tratamos de visualizar la organización y funcionamiento de la vida en nuestro planeta. Tal jerarquía aparece en la figura 2.3, pero el ecosistema no aparece, ya que puede incluir cualquier agrupación dentro de la jerarquía. Por lo tanto, el ecosistema no es parte de la jerarquía de la vida en el planeta. La jerarquía ilustra la continuidad funcional y sin impedimento que se observa desde los átomos hasta las estrellas. A esa continuidad funcional, ilustrada por el ecosistema, es a la que se referían Sterner y Elser (2002) al afirmar que podemos unificar el estudio de los sistemas ecológicos utilizando los principios y conceptos que ellos identificaron al principio de este capítulo.
Los procesos de los ecosistemas incluyen la producción, respiración, reciclaje, sucesión, y homeostasis interna por medio de fenología o la simbiosis. La fenología
2 34 CAP ariel e. lugo | ecología a m i m anera El ecosistema: unidad de la vida en el planeta
Figura 2.3. Una jerarquía con continuidad funcional a lo largo de escalas de complejidad y tiempo de reemplazo. La jerarquía consiste en sistemas químicos, biológicos, ecológicos y extraterrestres (Lugo et al. 1999a).
describe los cambios temporales en el sistema, incluso la caída de hojarasca, el crecimiento de nuevas hojas, los eventos de floración y producción de semillas, entre otros. Toda la fenología está sincronizada a cambios en el ambiente físico y a las características genéticas de las especies. La simbiosis describe la relación entre especies, ya sea relaciones de depredación, competencia o mutualismo. Tanto los procesos fenológicos como las relaciones simbióticas ayudan en la integración entre el ambiente físico y el biótico y contribuyen a la homeostasis del ecosistema.
La producción, respiración y reciclaje se ilustran con diagramas en la figura 2.4. La productividad bruta es la conversión de energía radiante a energía química en el cloroplasto de las plantas (Fig. 2.5). La productividad neta es la productividad bruta menos la respiración de las plantas. La productividad neta se mide por el cambio en la biomasa de la planta que incluye la masa de hojas, tallos y raíces. Se mide indirectamente por la caída de hojarasca y el crecimiento de los árboles. La suma de estos dos atributos se conoce como productividad neta aérea, ya que no incluye el crecimiento de las raíces.
2 35 CAP ariel e. lugo | ecología a m i m anera El ecosistema: unidad de la vida en el planeta
Figura 2.4. Ejemplos de sistemas autotróficos cerrados (A), abiertos a la masa (B) y heterotrófico abierto a la masa pero sin fotosíntesis (C). Los almacenajes de nutrientes y materia orgánica son 1 y 2, respectivamente. Fotosíntesis es P y respiración es R.
Figura 2.5. Diagrama que muestra con detalle los principales flujos de energía en un ecosistema con el propósito de aclarar el concepto de productividad. La productividad bruta (Pb) es la conversión total de energía radiante a energía química por las plantas.La productividad neta, sin embargo, puede variar en su valor numérico según los costos respiratorios de las plantas (R) que se tomen en consideración.La velocidad de respiración en las plantas puede acelerarse bajo el efecto de los factores de tensión o estrés. La respiración de los consumidores es Rc. El significado de los flujos enumerado es: 1. Respiración antes de producir azúcares en la hoja. 2. Respiración asociada al almacenaje de las azúcares en la hoja. 3. Respiración asociada al transporte y conversión de las azúcares a estructura vegetal. 4. Respiración de la estructura vegetal. 5. La cosecha de toda la planta. 6. La Cosecha de estructura vegetal particular como los tallos. 7. Cosecha de materia orgánica muerta. 8. Cosecha de consumidores. El diagrama también ilustra el consumo de consumidores, la retroalimentación para maximizar la productividad y el rol de factores climáticos y edáficos (Lugo y Morris 1982).
Químicamente, la respiración es un proceso oxidativo, mientras que la fotosíntesis es un proceso reductor. La respiración es el proceso que permite a las plantas, animales, microbios y células obtener energía de la materia orgánica producida por la fotosíntesis. La fotosíntesis produce materia orgánica y oxígeno por medio de la captura de bióxido de carbono de la atmósfera y la reducción química del agua utilizando la energía solar. La respiración consume oxígeno para producir energía química al oxidar la materia orgánica liberando el bióxido de carbono.
Los procesos de fotosíntesis y respiración están acoplados en los ecosistemas. Sin embargo, solo cuando el ecosistema se coloca en un recipiente cerrado (un microcosmo) o si se considera a todo el planeta como un ecosistema, se puede inferir que el sistema es un sistema cerrado a la materia. Todos los ecosistemas son sistemas abiertos, ya que materiales y organismos pueden cruzar sus límites (Fig. 2.4).
La luz, o energía solar, es el factor que organiza los ecosistemas verticalmente (Fig. 2.6). Recordemos que la energía disponible al sistema está limitada por la
2 36 CAP ariel e. lugo | ecología a m i m anera El ecosistema: unidad de la vida en el planeta
constante solar, y por lo tanto, los organismos fotosintéticos (las plantas) compiten fuertemente por la luz, causando así que los sistemas ecológicos estén estratificados verticalmente, como se ilustra para muchos atributos en la figura 2.6.
La sucesión ecológica describe los cambios a través del tiempo en la estructura, composición de especies y funcionamiento de los ecosistemas. Generalmente, se distinguen dos etapas contrastantes en el desarrollo de estos. Las etapas de sucesión tempranas y las etapas de sucesión maduras. Un sistema joven en sucesión tiene características distintas a uno maduro en las postrimerías de la sucesión. El trabajo clásico de E.P. Odum (1969) contrasta estas dos etapas desde el punto de vista de los atributos de ecosistemas (Tabla 2.2).
En general, las etapas jóvenes de sucesión son etapas transicionales, mientras que las etapas finales se consideran como un estado estable. En el estado estable los insumos de energía son iguales a los usos de energía y el sistema no aumenta sus almacenajes a pesar de tener un flujo alto de energía. La idea del balance de la naturaleza surge del funcionamiento del sistema maduro, donde la tasa de producción es igual a la tasa de respiración. En ese estado, no se acumula materia orgánica y el almacenaje de biomasa es estable. El sistema está balanceado. Estado estable no es un equilibrio. Equilibrio representa la entropía máxima o estado de desorden sin vida. Cuando se utiliza el término equilibrio para referirse al estado estable se comete un error termodinámico. El término alterno y aceptable es el equilibrio dinámico.
La figura 2.7 ilustra distintos tipos de sucesión, los estados estables y el equilibrio. En esa figura también se identifican sistemas autotróficos y sistemas heterotróficos. Los cambios en composición de especies y estructura asociados a distintas
2 37 CAP ariel e. lugo | ecología a m i m anera El ecosistema: unidad de la vida en el planeta SÍMBOLO Ciclo biogeoquímico Zona Fuentes de energía Clorofila (mg/m3) Aire SS Zona de acumulación R > P Zona de regeneración P > R Zona eufótica Reserva de gases Luz P y R g/(m3)(hr) Reactantes en las reacciones de P y R Componentes biológicos P P CO2 CO2 R O2 H2O Minerales y M. Orgánica Energía para Consumidores R Punto de compensación Oxígeno Minerales Plantas Herbívoros Carnívoros Carnívoros secundarios Microconsumidores (g/m3) (g)
Figura 2.6. Relación entre el funcionamiento y la estructuración de ecosistemas. Se ilustra la estratificación vertical de la estructura y el funcionamiento en sistemas ecológicos. La parte superior de la ilustración equivale al tope del ecosistema y la parte inferior al suelo o sedimentos (SS). La fotosíntesis es representada por «P» y la respiración por «R». Modificada de Lugo y Morris (1982) basado en Odum (1971).
Tabla 2.2
Las comparaciones de E.P. Odum (1969) entre sistemas ecológicos jóvenes y maduros. Algunos puntos se han simplificado.
Atributo del ecosistema Etapas en desarrollo Etapas maduras
Energética de la comunidad
Producción primaria bruta/respiración de la comunidad (P/R) Mayor o menor de 1 Se acerca a 1
Producción primaria bruta/biomasa (P/B) Alta Baja
Mantenimiento de biomasa/unidad de flujo de energía (B/E) Baja Alta
Productividad neta (rendimiento) Alta Baja
Cadenas alimentarias Lineares y de pastoreoRedes basadas en detrito Estructura de la comunidad
Materia orgánica total Baja Alta
Nutrientes inorgánicos Extrabióticos Intrabióticos
Diversidad de especies Baja Alta
Diversidad bioquímica Baja Alta
Estratificación y heterogeneidad espacial Pobremente organizado Bien organizado Historia de vida
Especialización del nicho Amplio Estrecho
Tamaño de organismos Pequeños Grandes
Ciclos de vida Cortos y simples Largos y complejos Ciclaje de nutrientes
Ciclos minerales Abiertos Cerrados
Intercambio de nutrientes entre organismos y el ambiente Rápidos Lentos
Función del detrito en la regeneración de nutrientes No es importante Importante Presión de selección
Forma de crecimiento Rápida (selección «r»)
Para retroalimentación (selección «K»)
Producción Cantidad Calidad
Homeostasis
Simbiosis interna Sin desarrollar Desarrollada
Conservación de nutrientes Pobre Buena
Estabilidad ante disturbios Pobre Buena
Entropía Alta Baja
Información Baja Alta
combinaciones de producción y consumo de energía los discutiremos más adelante cuando estudiemos los bosques de Puerto Rico. Normalmente, si el tiempo de sucesión es lo suficientemente largo, el sistema llega a su madurez ecológica con una serie de atributos estructurales, funcionales y de composición de especies, según lo describió E.P. Odum (1969). Pero esa sucesión es una sucesión sin disturbios, ya sean no-antropogénicos o antropogénicos. Ejemplos de disturbios no-antropogénicos son las sequías, las tormentas, los huracanes y las inundaciones. La deforestación, la sobre explotación de poblaciones silvestres y la quema de vegetación son ejemplos de disturbios antropogénicos. Los disturbios afectan la estructura y el funcionamiento
2 38 CAP ariel e. lugo | ecología a m i m anera El
de la vida en el
ecosistema: unidad
planeta
las condiciones iniciales en el eje vertical y las condiciones finales o del estado «clímax» en el eje horizontal. Las condiciones iniciales están determinadas por los almacenajes de materia orgánica (tanque superior) y nutrientes (tanque inferior). Las condiciones finales corresponden al balance entre la productividad y la respiración (sistemas balanceados), más productividad que respiración (sistemas autotróficos) y más respiración que productividad (sistemas heterotróficos). Los sistemas sin producción o insumo de materia orgánica eventualmente llegan al equilibrio.
de los ecosistemas y son determinantes en la composición de especies en los sistemas afectados.
Los disturbios, independientemente de su origen, interrumpen la sucesión y causan oscilaciones en los sistemas ecológicos (Fig. 2.8). Los ecosistemas responden ante los efectos de disturbios, ya sea resistiendo el disturbio o cambiando de estado ante el disturbio. La capacidad para absorber el efecto de un disturbio sin cambiar de estado se conoce como resiliencia, un concepto desarrollado por Holling (1973).
Cuando el disturbio supera la capacidad de resiliencia del sistema ecológico, este cambia de estado a otro tipo de ecosistema, el cual tiene resiliencia ante las nuevas condiciones que causaron el cambio en el sistema anterior. Todos los ecosistemas tienen resiliencia y es un error pensar que la resiliencia se perdió al cambiar de estado.
2 39 CAP ariel e. lugo | ecología a m i m anera El ecosistema:
en
unidad de la vida
el planeta
Figura 2.7. Patrones de productividad primaria (P) y respiración (R) en catorce tipos de sucesión sin interrupciones por disturbios (modificado de Odum 1971). Las gráficas están organizadas en una matriz con
Ciclo de nutrientes abierto Ciclo de nutrientes cerrado
Figura 2.8. ( A ) Patrón de sucesión de un sistema joven a uno maduro de acuerdo con los paradigmas en E.P. Odum (1969) y asumiendo que hay el tiempo suficiente para completar el proceso sin disturbios. (B) Patrón de sucesión en respuesta a múltiples eventos de disturbio. El patrón de la sucesión cambia en respuesta a cada evento y es distinto a la sucesión en ausencia de disturbios. El patrón «B» está basado en los experimentos de Ewel (1971).
La resiliencia es un atributo relativo al factor que causa el disturbio y cuando se excede, ocurre un cambio en el estado del sistema a otro que puede resistir la condición de cambio. Por ejemplo, un bosque caribeño tiene resiliencia hacia los huracanes y siempre se recupera no importa su fuerza. Pero ese mismo bosque no resiste la tala continua de árboles. Ante la tala de árboles, el bosque cambia a pastizal, el cual no se afecta por talas. Si el pastizal se expone a fuegos, los cambios en especies pueden convertir el fuego en un factor que favorece al pastizal. De manera que todos los sistemas tienen resiliencia hacia los factores a los cuales están adaptados, pero no la tienen para los factores que le son desfavorables.
El artículo sobre sucesión de E.P. Odum (1969) fue tan influyente que los conservacionistas y ecólogos concentraron su atención en los sistemas maduros a expensas de los sistemas en etapas jóvenes de sucesión. Eugene P. Odum asoció a los sistemas jóvenes con la producción, el crecimiento y cambio en la cantidad de biomasa. Por el contrario, el sistema maduro lo asoció con protección, estabilidad y calidad. El español Ramón Margalef (1968) también contribuyó a estas comparaciones tan sesgadas con su trabajo sobre la teoría de ecología, la cual enfatizaba los principios de E.P. Odum. Otro factor que contribuyó a la atención desmedida en los sistemas maduros es el hecho de que son más fáciles para modelar, ya que si se asume el estado estable, no hay necesidad de preocuparse por los cambios de biomasa y otros parámetros del ecosistema. Las premisas de ecosistemas en estado estable a su vez causaron que los modelos globales de bosques por décadas dieran lugar a pronósticos erróneos (Lugo y Brown 1986) que discutiremos en el capítulo 13.
2 40 CAP ariel e. lugo | ecología a m i m anera El ecosistema: unidad de la vida en el planeta
Figura 2.9. Los símbolos del lenguaje inventado por H.T. Odum (1971) para diagramar sistemas utilizando las leyes de energía y conservación de masa. El lenguaje sirve para representar procesos comunes a todos los sistemas social-ecológicos-tecnológicos y ha sido explicado y expandido en Odum (1983, 2007). A . Almacenaje pasivo. Este símbolo representa el almacenaje de cualquier substancia en el ecosistema, sin que ocurra una transformación de energía durante el almacenaje. Algunos ejemplos son el almacenaje de hojas en el suelo del bosque, gasolina en el tanque de un automóvil o comida en la alacena. B. Multiplicador. Este símbolo denota la relación multiplicativa entre un flujo energético de baja magnitud (factor controlante) y otro flujo energético de mayor magnitud (insumo). El producto representa un tercer flujo energético y está acompañado por la pérdida de calor. C . Receptor de energía. Este símbolo representa la recepción de energía ondular, como la energía solar, el sonido y el oleaje. La energía ondular activa algún material cíclico en el sistema y este transfiere energía potencial a algún proceso del sistema y retorna a su estado de receptor. Un ejemplo es la recepción de luz por la clorofila. D. Consumidor. Este símbolo representa cualquier población de consumidores en un sistema, por ejemplo el ser humano, sus máquinas, o los animales y microbios del bosque. El sistema consumidor almacena energía potencial activamente (por medio de transformación y pérdida de calor) y utiliza parte del almacenaje para trabajar y obtener más energía potencial. E . Productor. El símbolo de un sistema productor es la combinación de dos símbolos: un receptor de energía y un consumidor, el cual mantiene la maquinaria metabólica y recibe energía potencial del receptor. Ejemplos: un bosque o una planta. F. Fuente de energía. Cualquier fuente de energía de un sistema se representa con el círculo. Ejemplos: el sol, la energía fósil, el agua en una represa o el viento. Para facilitar la comprensión se debe especificar si la energía es transmitida como un flujo continuo, con fuerza continua o con alguna periodicidad específica. G. Presión o tensión. Este símbolo denota la pérdida de energía potencial de un sistema. La pérdida es una función multiplicativa de algún factor de tensión o presión. El producto de la interacción no hace el trabajo, sino que se pierde como calor. Ejemplo: el efecto de un contaminante en un río. H. Multiplicador de dos direcciones. Este símbolo también se denomina símbolo de difusión. Se utiliza para representar procesos que pueden mover materia o energía en dos direcciones. La fuerza motriz del proceso determina la dirección del flujo. Ejemplos: el movimiento de las mareas bajo la acción de la luna, el movimiento de substancias químicas bajo la influencia de mareas, el movimiento vertical del plancton y nutrientes en el mar, o el intercambio de gases entre cuerpos de agua y la atmósfera. I. Interruptor. Este símbolo denota procesos que tienen estados activados y no activados. El flujo no es posible en ausencia de la señal de activación. Cuando esta señal está presente, el interruptor se activa y es posible el flujo de materia o energía. Ejemplos: el interruptor eléctrico o la germinación de una semilla. J. Interruptor activado por un límite. Este símbolo se comporta de la misma manera que el interruptor anterior, pero antes de ser activado, el factor activador tiene que exceder un límite crítico característico del proceso que se está modelando. Por ejemplo, el flujo de agua sobre una represa tiene que exceder el volumen de la represa antes de que ocurra. Del mismo modo, el agua no se escapa del suelo, hasta que no excede su capacidad de retención. Al utilizar este símbolo hay que indicar las condiciones que deben satisfacerse para que funcione el interruptor. K. Transacción económica. Este símbolo se utiliza para representar los flujos de dólares y energía en sistemas económicos. El flujo de dólares se representa con líneas entrecortadas y el de energía con líneas sólidas. Cuando se obtiene un artículo o un servicio se envían dólares en una dirección y se obtiene energía potencial o trabajo en la dirección opuesta. La razón o cociente entre estos dos flujos es el precio del artículo o el servicio. La pérdida de calor de tales convenios es pequeña a menos que aquellos no estén controlados por sistemas de transacción complejos. L . Caja. Este símbolo denota funciones para las cuales no hay símbolo, cuando se representa una acción no especificada o algo de poca importancia para el punto que ilustra el modelo. Si se conoce la función matemática que describe el funcionamiento del compartimiento, esta debe escribirse dentro de la caja. M. Amplificador con ganancia constante. Este símbolo describe sistemas con energía ilimitada, que se utiliza para alimentar el insumo por un factor constante (el factor de amplificación). Esto causa crecimiento exponencial. Una población que tenga una fuente de alimentos y espacio ilimitado puede sostener una razón de reproducción constante y crecimiento exponencial. N. Pérdida de calor y flujos en estado estable. Se utilizan dos símbolos: la línea representa el flujo estable de energía, y la flecha, con el símbolo de tierra que se utiliza en electricidad, representa la pérdida de calor necesario para hacer posible cualquier proceso irreversible (unidireccional).
2 41 CAP ariel e. lugo | ecología a mi man era El ecosistema: unidad de la vida en el planeta
Howard Thomas Odum se dio cuenta de que hay funciones ecológicas que se repiten en el ecosistema y que esas funciones se pueden expresar matemáticamente y simularse con modelos de computadora. Para facilitar el desarrollo de los modelos, Odum inventó una serie de símbolos (Fig. 2.9) que, si se utilizan correctamente, son útiles para simulaciones con computadoras, para desarrollar preguntas de investigación, para cuantificar los flujos de energía y materiales de los sistemas, para identificar las fuerzas que potencian a los sistemas, para describir la interacción entre los componentes de sistemas y para verificar el nivel de entendimiento que uno pueda tener sobre algún sistema de interés (Odum y E.C. Odum 2000).
Asignación
(1) Selecciona cualquier ecosistema de tu interés y construye un diagrama de sus compartimientos principales, la circulación, transformación y acumulación de materia y energía y las externalidades que regulan su funcionamiento. Ven preparado para presentarle tu modelo a la clase.
(2) Utiliza la siguiente información para calcular todos los flujos ilustrados en el siguiente diagrama.
Síntesis de información dispersa
Los físicos reportan que la constante solar promedia 1.951 gram calories por centímetro cuadrado por minuto.
Los demógrafos indican que la población de Puerto Rico es de aproximadamente 3.7 millones de personas.
Los geógrafos calculan que el área de Puerto Rico es de aproximadamente 8,000 kilometros cuadrados.
Dietistas estiman que cada puertorriqueño consume 5,000 kcal por día.
Ecofisiólogos estiman una tasa promedio de fotosíntesis bruta en un bosque tropical de 10 micromoles de CO2 por metro cuadrado de hojas por segundo.
Ecólogos han medido 3 m 2 de superficie foliar/m 2 de superficie terrestre para el área foliar del bosque.
Químicos establecen que un mol de CO2 consumido por la fotosíntesis es el equivalente de 30 g de azúcar producida
Laboratorios estiman que un gramo de azúcar es equivalente a 4 kilocalorías.
L os ecofisiólogos encuentran que el bosque respira el 90 % de lo que fotosintetiza.
Los físicos estiman que 34 % de la energía del sol se refleja al espacio y que solo el 42 % llega al dosel del bosque.
2 42 CAP ariel e. lugo | ecología a m i m anera El ecosistema: unidad de la vida en el planeta
2 43 CAP ariel e. lugo | ecología a m i m anera El ecosistema: unidad de la vida en el planeta kcal/m2 · día 156 16 13.7 140 5,724
CaPítulo 3
Pregunta
¿Cuál de los siguientes dos árboles creció más rápido y cuánto carbono removieron de la atmósfera?
Ambos árboles eran de 10 metros de altura con tallos cilíndricos y la densidad de su madera era de 1 g/cm3
El árbol 1 tenía un diámetro a la altura del pecho (dap) de 10 cm el primero de enero y creció a 11 cm para diciembre 31.
El árbol 2 tenía un diámetro a la altura del pecho de 100 cm el primero de enero y creció a 101 cm para diciembre 31.
Contestación
Estequiometría es la rama de la química que trata sobre la aplicación de las leyes de proporciones definidas y la conservación de masa y energía. Es la relación cuantitativa entre constituyentes en una sustancia química. Se refiere a los patrones y proporciones de elementos en reactantes y productos de reacciones químicas. Stoicheion = elemento; estequiometría: que mide los elementos. Ecología Estequiométrica estudia el balance de múltiples sustancias químicas en interacciones y procesos ecológicos. El estudio del balance de energía y materiales.
Alometría es el estudio del crecimiento relativo de una parte de un organismo en relación con el crecimiento de todo el organismo. Relaciones alométricas describen la
ariel e. lugo | ecología a mi ma nera
alometría
Estequiometría y
Parámetro Árbol 1 Árbol 2 dap inicial 10 100 Crecimiento lineal 1 cm/año 1 cm/ año Crecimiento basal 16.5 cm2/año 158 cm2/año Crecimiento volumétrico 16,500 cm3/año 158,000 cm3/año Crecimiento en masa 16.5 kg/año 158 kg/año Remoción de carbono atmosférico 8.25 kg/año 79 kg/año
45
relación entre dos atributos de organismos vivos y usualmente son relaciones matemáticas y geométricas bien definidas. Alometría se deriva de la palabra griega alloios, que quiere decir diferente.
Las leyes o relaciones de energía, estequiometría, alometría, espacio y adaptabilidad regulan la actividad biótica. Son leyes o relaciones críticas para el entendimiento del funcionamiento de los ecosistemas y son la base de la ecología cuantitativa. En este capítulo enfatizo la estequiometría y alometría, comenzando por la primera. Al final de cada sección, incluyo ejemplos de Puerto Rico.
Estequiometría
Para ilustrar una aplicación de la estequiometría, consideren la siguiente reacción química que en una dirección describe la fotosíntesis y en la otra la respiración.
6 CO2 (gas) + 6H2O = C6H12O6 + 6O2 (gas)
Conociendo los pesos moleculares de los elementos (C = 12, O = 16, H = 1), se puede calcular que 264 g de CO2 se combinan con 108 g de H2O para producir 192 g de oxígeno y 180 g de carbohidratos. La masa envuelta en ambos lados de la reacción es igual (372 g), ya que la masa se conserva. La energía del sol mueve la reacción hacia la derecha y la energía almacenada en el carbohidrato la mueve hacia la izquierda, ambas con pérdidas de calor. Fíjense que 1.07 g de oxígeno gaseoso equivale a uno de carbohidrato y para producir 1 g de carbohidrato se requiere 1.47 g de CO2 y 1.67 g de agua y se emite 1.38 g de oxígeno a la atmósfera. Cuarenta por ciento del carbohidrato es carbono.
Para el ecosistema, E.P. Odum (1953) presentó la siguiente ecuación balanceada y con datos sobre la energía requerida para describir la productividad primaria de un sistema acuático planctónico
1,300,000 kcal de energía radiante + 106 CO2 + 90 H2O + 16 NO3 + 1 PO4 + elementos minerales = 13,000 kcal energía potencial (1 %) en 3,258 g protoplasma (106 C, 180 H, 46 O, 16 N, 1 P, 815 g de cenizas minerales) + 154 O2 + 1,287,000 kcal de calor dispersado (99 %).
Observen que la masa y la energía se conservan a la vez que se genera entropía (el calor dispersado). Los elementos entran en reacciones químicas en proporciones determinadas y los organismos funcionan dentro de los límites impuestos por las proporciones estequiométricas. Como vamos a ver, el funcionamiento de células, organismos y ecosistemas se adapta a estas proporciones para optimizar el flujo de energía y la supervivencia. Justus Liebig (1840) se dio cuenta de estas proporciones en
3 46 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
el movimiento de los elementos y propuso la Ley del Mínimo que postula que entre los materiales químicos requeridos para el funcionamiento de organismos, el que esté en menor cantidad será el factor limitante.
Otra observación sagaz fue la de Alfred Redfield (1934, 1958) quien publicó un trabajo que establece la estequiometría del océano y el plancton: 106 átomos de C: 16 N: 1P, las proporciones utilizadas por E.P. Odum en la ecuación del párrafo anterior. Este descubrimiento implica que los organismos, la vida, influyen la estequiometría del océano y sugiere que las condiciones estequiométricas del planeta son en parte el resultado de la actividad biótica; la vida afectando las condiciones del planeta. Dicho de otra manera, el planeta contiene las condiciones necesarias para sostener la vida, no porque las tuviese originalmente, sino porque emergieron a causa de la evolución de la actividad biótica. La acumulación de oxígeno en la atmósfera como consecuencia de la evolución de la fotosíntesis en las plantas sería otro ejemplo de este fenómeno. Los siguientes ejemplos tomados del libro de Sterner y Elser (2002) ilustran las relaciones estequiométricas en distintos niveles de organización molecular y biótica.
La figura 3.1 destaca la diferenciación entre la composición química del cuerpo humano y la de la corteza terrestre. Además de la importancia del agua (hidrógeno y oxígeno) en ambos, es notable la exclusión de silicio y otros elementos en el cuerpo humano más la concentración en el organismo del carbono, fósforo y nitrógeno. ¿Cuál es la razón por la cual se concentran estos elementos y no otros que están en concentraciones más altas en la corteza terrestre? Estos tres elementos son vitales para la estructura y funcionamiento de células, tejidos, organismos y ecosistemas. Por su importancia para el crecimiento, se consideran nutrientes.
Un nutriente es un elemento necesario para completar el ciclo de vida de organismos. En la ecología los ciclos de carbono, hidrógeno y oxígeno se estudian aparte de los otros elementos esenciales como el nitrógeno, fósforo, azufre, potasio, magnesio y calcio, que se consideran los macronutrientes por su abundancia relativa y efectos limitantes para la productividad de sistemas ecológicos. En contraste, el manganeso,
3 47 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
Figura 3.1. Comparación de la composición química de la corteza de la tierra con la del ser humano (Basado en Lotka 1956 y Sterner y Elser 2002). El ser humano concentra carbono, hidrógeno, nitrógeno y fósforo en comparación con la corteza terrestre, la cual contiene mayores cantidades de potasio, sodio, hierro, silicio y aluminio. La escala para las concentraciones es logarítmica.
Figura 3.2. La relación nitrógeno:fósforo (N:P) de moléculas bióticamente importantes para el funcionamiento de organismos. Noten las moléculas que no contienen fósforo, pero tienen distintas cantidades de nitrógeno. Moléculas de trifosfato de adenosina (ATP, por sus siglas en inglés) contienen la mayor cantidad de fósforo y son moléculas especializadas en la captura, almacenaje y transferencia de energía química en las células de todos los organismos. Los números asociados a las líneas entrecortadas ilustran el cociente de la relación N:P. Modificado de Sterner y Elser (2002).
hierro, cloro, cobre, zinc, boro y molibdeno, se consideran micronutrientes por ser esenciales, pero en cantidades tan bajas que generalmente no son limitantes para la productividad de los sistemas ecológicos. Cada elemento o nutriente, no importa su abundancia, tiene un ciclo biogeoquímico particular y bajo condiciones especiales pueden ejercer su influencia ya sea limitando o estresando los procesos ecológicos.
Dependiendo de su función energética o estructural, diferentes tipos de moléculas exhiben distintas proporciones de nitrógeno y fósforo (Fig. 3.2). La glucosa, que funciona como almacenaje de energía, es rica en carbono, pero no contiene fósforo ni nitrógeno. El carbono provee mejores alternativas de almacenaje de energía y flexibilidad estructural (por su alta capacidad para establecer enlaces entre sí y con otros elementos) que otros elementos en la tabla periódica. Las proteínas son ricas en nitrógeno, pero tienen poco fósforo, mientras que la molécula trifosfato de adenosina (ATP, por sus siglas en inglés) es rica en fósforo con concentraciones relativamente altas de nitrógeno. Los lípidos y carbohidratos son altos en carbono pero bajos en nutrientes.
Figura 3.3. La relación nitrógeno:fósforo de estructuras celulares presentes en bacterias, plantas o animales. Noten la diferencia entre la pared celular de plantas y bacterias. El retículo endoplásmico es «E.R.», por sus siglas en inglés. Los números asociados a las líneas entrecortadas ilustran el cociente de la relación N:P. Modificado de Sterner y Elser (2002).
Cuando las moléculas se combinan para formar organelos o moléculas más complejas, cambia la estequiometría y el cambio depende de la estructura y funcionamiento de las combinaciones resultantes (Fig. 3.3). Por ejemplo, la pared celular
3 48 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
de las plantas no contiene fósforo ni nitrógeno y su función depende de la fortaleza estructural de las moléculas ricas en carbono. No así las paredes de las bacterias que son ricas en nitrógeno y tienen funciones fisiológicas relacionadas con proteínas y enzimas. En contraste, los ribosomas y cromosomas son ricos en fósforo con concentraciones moderadas de nitrógeno y bajas concentraciones de carbono. La molécula de ácido ribonucleico (RNA, por sus siglas en inglés) es responsable de la síntesis de proteínas que a su vez determinan el crecimiento animal, según se infiere de la figura 3.4 para una variedad de artrópodos y microorganismos. Mientras más rápido el crecimiento, más necesario es producir sustancias químicas como las proteínas que aseguren el crecimiento. Esto a su vez depende de la presencia de RNA y de los nutrientes necesarios en las proporciones necesarias para sostener el crecimiento.
positiva entre el crecimiento específico por día en una variedad de microorganismos y artrópodos y la concentración de ácido ribonucleico (RNA, por sus siglas en inglés). El crecimiento específico es la cantidad de masa producida por masa presente durante un intervalo de tiempo, en este caso un día. Modificado de Sterner y Elser (2002).
Las concentraciones1 de nitrógeno y fósforo en plantas de distintos hábitos exhiben poca variación (Fig. 3.5a) y tanto las concentraciones del fósforo como la del nitrógeno, están positivamente relacionadas con las tasas de crecimiento de las plantas (Fig. 3.5b,c). En general, cuando la tasa de crecimiento aumenta, aumentan también las concentraciones de nutrientes. Las bajas tasas de crecimiento ocurren cuando las concentraciones de los nutrientes son bajas. El mantenimiento de las concentraciones de nutrientes dentro de límites específicos a pesar de la alta variación de esas concentraciones en los ambientes de donde proceden es el resultado de procesos de homeostasis en los organismos. En la figura 3.6 se ilustra cómo las bacterias mantienen una estequiometría particular, independientemente de la estequiometría del recurso ambiental de donde devengan su nutrición. En el caso de las plantas, la nutrición proviene del suelo y en la de los animales, de su alimento, ya sea vegetal o
1 La concentración de elementos se reporta en % o mg del elemento por gramo de peso seco; sin embargo, cuando se calculan los cocientes entre elementos, la concentración se estima en mmol/kg que se obtiene dividiendo la concentración en unidades de mg/g, o en % , entre el peso molar del elemento (g/mol).
3 49 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
Figura 3.4. Relación
Figura 3.5. Relación entre alometría (tasa de crecimiento específico) y estequiometría (concentraciones de nitrógeno y fósforo [A , B] y sus cocientes [C ]) en los órganos fotosintéticos de una variedad de plantas terrestres y acuáticas de diferentes tamaños. Aunque la variación en las concentraciones de fósforo y nitrógeno varían poco en las plantas consideradas (A), las de rápido crecimiento tienen concentraciones más altas que las de lento crecimiento (B). El cociente nitrógeno:fósforo (N:P) es más alto en las plantas de lento crecimiento que en las de rápido crecimiento (C) debido a que la concentración de fósforo aumenta más rápidamente en las plantas de rápido crecimiento que la concentración de nitrógeno. Los números asociados con las líneas rectas entrecortadas corresponden a cociente N:P y los números en la línea entrecortada curvada corresponden a la tasa de crecimiento específico. Modificado de Sterner y Elser (2002).
Figura 3.6. Ilustración de la homeostasis de elementos en el consumidor de alimento en relación con la concentración del elemento en el alimento. El organismo puede mantener concentraciones más altas o bajas que en el alimento (A). Los números asociados a las líneas en ‘B ’ representan distintos grados de homeostasis con el 1 representando la ausencia de homeostasis. Modificado de Sterner y Elser (2002).
3 50 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
animal. Los microorganismos descomponedores derivan sus nutrientes del material orgánico muerto o del sustrato donde se encuentren.
Figura
Eficiencia de la fotosíntesis al
La velocidad de fotosíntesis neta en las plantas es también proporcional a la disponibilidad de los nutrientes, como se ilustra para el nitrógeno en la figura 3.7. Altas tasas de fotosíntesis requieren altas concentraciones de nitrógeno disponible. De igual manera, las tasas de descomposición son proporcionales al cociente carbono:nitrógeno en el material en descomposición (Fig. 3.8). Cuando el material en descomposición tiene cocientes altos de carbono:nitrógeno, la descomposición es lenta y se acelera cuando el cociente es bajo, o sea, cuando hay más nitrógeno por unidad de carbono en el material bajo descomposición. Se dice que la calidad nutricional de un
de saturación de luz en relación con la concentración de nitrógeno en
primer producto de carbono en las plantas C3 contiene tres átomos de carbono. Los números asociados a las líneas entrecortadas reflejan la eficiencia de la fotosíntesis en el uso del nitrógeno (mmol CO2/mmol N). Los puntos representan el comportamiento de 21 especies de plantas salvajes con distintos hábitos en zonas áridas de los Estados Unidos de América y Sur África. Modificado de Sterner y Elser (2002).
C 3.
Figura 3.8. Relación inversa entre la constante de descomposición de detrito y los cocientes carbono:nitrógeno (C:N) y carbono: fósforo (C:P) en el detrito bajo descomposición. La descomposición está expresada en la fracción del detrito que se descompone por día, el cual disminuye con el aumento de los cocientes. Observe las escalas logarítmicas. Modificado de Sterner y Elser (2002).
3 51 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
3.7.
punto
hojas
El
tejido aumenta mientras más bajo sea el cociente carbono:nitrógeno. Por otro lado, generalmente las plantas retienen el material nutritivo (por el proceso de retranslocación) antes de desechar hojas o madera muerta por vía de la caída de hojarasca del bosque. Eso le aumenta el cociente carbono:nitrógeno a los materiales disponibles para la descomposición.
Las plantas tienen que concentrar nutrientes del suelo para poder llevar a cabo sus funciones de productividad. Los suelos generalmente tienen concentraciones bajas de nutrientes esenciales comparado con las concentraciones en las plantas, lo que aumenta el costo energético de concentrarlos. En esos casos las plantas deben producir más raíces para aumentar el volumen del suelo disponible para obtener nutrientes. Otra estrategia es la de establecer relaciones simbióticas con hongos llamados micorrizas. La figura 3.9 demuestra que las plantas aumentan su biomasa y la concentración de fósforo en los tallos cuando están asociadas a las micorrizas. A cambio de este servicio, el hongo obtiene de las plantas materia orgánica con energía potencial para su metabolismo. Este es un ejemplo claro de una relación simbiótica donde ambos participantes se benefician mutuamente.
Los hongos y bacterias funcionan como agentes de mineralización de la materia orgánica muerta en el bosque. La mineralización es sinónimo de la descomposición u oxidación o respiración de la materia orgánica muerta en el bosque. La descomposición fragmenta y convierte la materia orgánica en bióxido de carbono y agua, consumiendo oxígeno en el proceso y generando nutrientes (minerales) que se tornan disponibles a las plantas y los microorganismos. Las plantas y los microorganismos compiten por los nutrientes disponibles en el suelo y en la materia orgánica muerta.
En la figura 3.10 se ilustra que la velocidad de mineralización y la producción de hojarasca aumentan con la concentración de nitrógeno en el suelo. Más aún,
3 52 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
Figura 3.9. Ilustración de cómo las micorrizas incrementan la concentración de fósforo en los tallos de plantas y cómo el incremento en fósforo permite mayor biomasa de tallos (plantas más grandes). El efecto de los hongos es mayor en lugares de baja fertilidad y eventualmente desap arece bajo condiciones de alta fertilidad. Modificado de Sterner y Elser (2002).
cuando el cociente de carbono:nitrógeno aumenta en la vegetación, la tasa de mineralización disminuye. Los procesos de mineralización y producción de hojarasca son más rápidos cuando la concentración de nitrógeno en la materia orgánica es alta en relación con la de carbono. Esta tendencia se ilustra también en la figura 3.11. La eficiencia en el retorno de nitrógeno por vía de la caída de hojarasca es alta cuando las tasas de caída son bajas y el contenido de nitrógeno en las hojas que caen es bajo (línea entrecortada con un 20 en la figura 3.11a). Altas tasas de caída de hojarasca con altas cantidades de nitrógeno generalmente exhiben una baja eficiencia en el retorno de nutrientes, como se ilustra con los puntos asociados a la línea entrecortada con un 5 en la figura 3.11a. La eficiencia en el uso de nutrientes durante el proceso de caída de hojarasca se estima dividiendo la cantidad de materia orgánica que cae al suelo entre la cantidad del nutriente en la caída de hojarasca. El resultado se expresa en unidades de masa orgánica retornando al suelo por unidad de masa de nutrientes en la hojarasca que cae al suelo.
Figura 3.10. Mineralización anual (tasa de descomposición) y producción anual (caída) de hojarasca con relación al cociente carbono:nitrógeno (C:N) de gramíneas bajo tres concentraciones de nitrógeno en el suelo luego de tres años de crecimiento. La producción de hojarasca es mayor y la mineralización menor cuando el C:N es alto. La mineralización es más rápida cuando la concentración de nitrógeno en el suelo es alta y el C:N es bajo, pero el efecto de la concentración de nitrógeno en el suelo desaparece cuando el C:N es alto. Modificado de Sterner y Elser (2002).
También, mientras más alta la producción de biomasa, más rápida es la mineralización del nitrógeno (Fig. 3.11b), posiblemente porque los materiales en descomposición están enriquecidos por el nitrógeno cuando la productividad es alta, mientras que cuando la productividad es baja, esos materiales orgánicos contienen más carbón por unidad de nitrógeno. La figura 3.11c ilustra la misma relación entre la concentración de fósforo en el suelo y la caída de hojarasca. Tanto para el fósforo como para el nitrógeno, la eficiencia de crecimiento en relación con el retorno de nutrientes por caída de hojarasca aumentó en suelos infértiles comparado con suelos fértiles para cuatro especies arbóreas de zona templada. Cuando hay alta disponibilidad de nutrientes, la eficiencia en su uso disminuye porque la producción de materia orgánica (energía potencial) aumenta. Si los nutrientes escasean, la tasa de productividad disminuye, pero aumenta la eficiencia en el uso de los nutrientes.
Durante la sucesión ecológica, los bosques acumulan especies, material orgánico (biomasa) y por supuesto, nutrientes como el nitrógeno y fósforo (Fig. 3.12a,b). A
3 53 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
pesar de que las cantidades acumuladas de nutrientes varían en el tiempo y en los distintos componentes vegetales del bosque, la relación estequiométrica entre nitrógeno y fósforo no varía tanto como las cantidades absolutas y la distribución de estos elementos entre los distintos grupos de plantas (Fig. 12c). Este es otro ejemplo de la homeostasis estequiométrica en acción a través del tiempo y en el ecosistema.
El ecofisiólogo venezolano Ernesto Medina (2015) utilizó datos reportados por Sánchez et al. (2015) sobre la química de las hojas de árboles en Puerto Rico para determinar la huella o patrón de concentración de elementos químicos (chemical element signature) de unas 120 especies. El análisis está basado en la noción de que las concentraciones de nutrientes en las hojas de las plantas reflejan las condiciones nutricionales donde crecen dependiendo del comportamiento genético de la
Figura 3.11. Relación entre ( A ) la tasa de caída de hojarasca y la cantidad de nitrógeno en la caída de hojarasca, (B) la productividad de hojarasca, madera y biomasa sobre el terreno y la tasa de mineralización de nitrógeno y (C) la caída de hojarasca con la cantidad de fósforo en el suelo. Los datos corresponden a distintos tipos de bosques de zona templada y demuestran la disminución en la eficiencia del uso de nutrientes cuando la tasa de productividad es alta. Noten que a altas tasas de productividad más unidades de nutrientes fluyen (A y B) o son necesarias (C), lo que reduce la eficiencia (masa producida por masa utilizada o requerida). Los números asociados a las líneas entrecortadas en (A) reflejan la eficiencia en el uso del nitrógeno; números bajos reflejan eficiencias más bajas. Modificado de Sterner y Elser (2002).
3 54 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
Figura 3.12. Acumulación de ( A) nitrógeno (N) y ( B) fósforo (P) y (C) sus respectivos cocientes N:P en distintos tipos de vegetación a lo largo de una sucesión de 20 años después de una tala. Noten la progresión en el aumento de los almacenajes de nitrógeno y fósforo y su distribución entre los tipos de vegetación. A pesar de los cambios en almacenaje y los distintos tipos de vegetación, el cociente N:P no cambió tanto. Modificado de Sterner y Elser (2002).
especie. Por lo tanto es posible comparar la estequiometría de una especie con la disponibilidad de elementos en el suelo. En algunos casos el ambiente determina el tipo de especie que puede crecer en un lugar y en otros, la especie puede acumular ciertos elementos a pesar de que están en bajas concentraciones en el suelo. Medina relacionó la estequiometría de las hojas con tipos de condiciones ambientales y llegó a las siguientes conclusiones: (1) las especies pioneras son ricas en nitrógeno, (2) especies de bosques muy húmedos tienden a tener bajas concentraciones de todos los elementos, (3) las especies del karso se subdividieron en dos grupos, unas acumulando calcio y otras no, (4) las especies de mangle exhibieron concentraciones heterogéneas con relación a la proporción de los elementos, (5) Laguncularia racemosa y Pterocarpus officinalis exhibieron bajas concentraciones de azufre a pesar de las altas concentraciones de azufre en los suelos, (6) Miconia prasina y Dacryodes excelsa exhibieron bajas concentraciones de magnesio, pero Avicennia germinans exhibió alta concentración de magnesio relativo a calcio, y (7) ciertas familias como Melastomataceae, Flacourtiaceae y Phytolaccaceae acumulan aluminio y hierro. En el karso, Medina
3 55 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
et al. (2017) concluyeron que la química del suelo y el régimen de lluvia regulaban la composición elemental en las hojas de los árboles.
Los estudios de estequiometría en Puerto Rico ayudan en la interpretación de los bosques más allá de sus respuestas a las variaciones en clima. Por ejemplo, sabemos que los bosques de Puerto Rico en el presente evidencian tasas de recambio de nutrientes altas como consecuencia del enriquecimiento nutricional de la hojarasca que se acumulan en el suelo de los bosques. El origen nutricional alto en esa hojarasca, reflejado en las concentraciones de fósforo, nitrógeno y la relación carbono:nitrógeno, refleja la composición de especies que dominan estos bosques, por ejemplo Spathodea campanulata (alto fósforo) y Leucaena leucocephala (alto nitrógeno). Uno de los legados de las especies introducidas en los bosques de Puerto Rico es la aceleración de los procesos del ciclaje de fósforo y carbono de los bosques (Erickson et al. 2014, Lugo y Erickson 2017).
Una comparación entre un bosque secundario de Tabebuia heterophylla (roble blanco) y uno primario de Dacryodes excelsa (tabonuco), ambos sobre suelos ácidos en el Bosque Experimental de Luquillo, dio como resultado los siguientes hallazgos (Lugo et al. 2022):
• Cocientes molares de carbono (C:N, C:P, C:S) eran más altos en el bosque secundario y los de nitrógeno (N:Ca, N:K, N:Mg, N:P) eran más altos en el bosque primario.
• El bosque secundario tenía los cocientes molares más altos con fósforo en el denominador (Ca:P, K:P, Mg:P, S:P) que los del bosque primario.
• La estequiometría del suelo separó a los dos bosques en términos de la capacidad de intercambio catiónico, el Ca:Al y la disponibilidad del fosforo; todos más altos en el bosque secundario comparado con el bosque primario.
• La acidez mayor en el bosque primario le causaba más estrés que al bosque secundario y aumentaba el costo de concentrar, almacenar y reciclar nutrientes.
a lome tría
El escocés sir D’Arcy Wentworth Thompson (1860–1948) fue un biólogo, matemático y estudioso de los clásicos. Se considera un pionero de la biología matemática y se recuerda por su libro de 1917, On Growth and Form, considerado por algunos como una joya literaria de la ciencia anglosajona. D’Arcy Thompson pensaba que los biólogos le daban más crédito del necesario a la evolución como responsable por las causas fundamentales que determinan la forma y estructura de los organismos vivos. Para él la responsabilidad recaía sobre las leyes físicas y mecánicas. D’Arcy Thompson
3 56 CAP ariel e. lugo | ecología a mi ma nera Estequiometría y Alometría
sugería un estructuralismo en vez de la supervivencia del más capacitado para explicar la forma de las especies. Escribió sobre el concepto de alometría lo siguiente:
“An organism is so complex a thing, and growth so complex a phenomenon, that for growth to be so uniform and constant in all the parts as to keep the whole shape unchanged would indeed be an unlikely and an unusual circumstance. Rates vary, proportions change, and the whole configuration alters accordingly.”
Thompson 1942, p. 205
Ante la complejidad del tema de la estructura y forma de los organismos vivos, aquí solo consideramos dos aspectos: (1) la relación entre el tamaño y el metabolismo de los organismos, y (2) el uso de la alometría para estimar la biomasa de bosques y construir balances de masa y nutrientes.
La relación entre el tamaño de un organismo (planta o animal) y su metabolismo es una de las relaciones fundamentales de la biología (Fig. 3.13). Mientras más grande es el organismo, menor es el cociente entre el área de superficie y su masa y por lo tanto, la pérdida de calor hacia el ambiente es menor. Esto se debe a la relación matemática entre área (largo al cuadrado) y volumen (largo al cubo). Un mamífero pequeño que mantiene su homeostasis termal variando su metabolismo, va a tener que estar más activo por gramo de peso y consumir más comida por unidad de peso que un mamífero más grande. La relación no es linear, sino geométrica según se aprecia al centro de la figura 3.13. La ley de Kleiber dice que el metabolismo, representado por q 0, es proporcional a la masa elevada a 3/4 (q 0 ~ μ 3/4)) donde μ es la masa del organismo. Esta relación alométrica, aunque controversial, unifica la energética y la estequiometría de los organismos, según se explica a continuación.
En la parte superior de la figura 3.13 se ilustra una cadena alimenticia con los flujos de energía de acuerdo con las leyes energéticas de conservación de energía y aumento en entropía. El sistema está en estado estable porque el insumo de energía es igual a la salida sin cambio de masa o energía almacenada. Si se divide el flujo energético a través de cada eslabón de la cadena alimenticia entre el costo necesario para sostener un gramo de tejido vivo de organismos de distintos tamaños, se obtiene la biomasa por metro cuadrado para las poblaciones de cada nivel trófico ilustradas en los histogramas de la figura. Nótese que se generan dos histogramas distintos con solo variar el tamaño de los organismos, ya que el flujo de energía es el mismo para los dos cálculos. Con la variación en biomasa entre los dos histogramas, cambia también el almacenaje de nutrientes y la estequiometría de las poblaciones.
3 57 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
Figura 3.13. (A) Flujo de energía en una cadena alimenticia con cinco eslabones y en estado estable (el insumo es igual al calor generado). (B) Relación entre el tamaño del organismo y el costo energético necesario para mantenerlo. El costo de mantenimiento de un gramo de organismo disminuye exponencialmente con el aumento en el tamaño. (C) La distribución de biomasa en cadenas alimenticias en dos tipos de ecosistemas: bosque y sistema planctónico. Manteniendo el mismo flujo de energía en « A » y considerando el costo de mantenimiento de organismos de acuerdo con su tamaño (B), se generan dos distribuciones de biomasa distintas con el mismo flujo de energía. Modificado de Odum (1971).
La ecología cuantitativa de sistemas permite estudiar el funcionamiento y papel que juegan los ecosistemas en el paisaje local o global por medio de la simulación de modelos locales o globales y otras técnicas como las de percepción remota. Algunos temas de interés incluyen determinar tasas de secuestro de carbono y la productividad primaria de los bosques, o entender los efectos del funcionamiento del ecosistema en el ciclo del agua, la producción de sedimentos o la utilización de nutrientes. Para practicar esta ecología cuantitativa es necesario conocer y cuantificar los distintos almacenajes de materia y nutrientes en los ecosistemas, al igual que las tasas de los variados procesos ecológicos. Las técnicas para medir procesos como la fotosíntesis y la respiración de plantas han progresado muchísimo, pero el ecólogo siempre tiene el problema de cómo relacionar datos tomados a una escala
3 58 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
Tabla 3.1
Algunas relaciones alométricas en bosques. Relación Cómo se estima y sus unidades Para qué se utiliza*
Índice de área foliar Área de hojas por unidad de área de suelo (m2/m2) Extrapola procesos por unidad de área foliar a unidad de superficie de bosque
Área específica foliar Área foliar por peso seco (cm2/g) Información sobre el espesor de la hoja Para convertir peso a área de hoja
Peso específico foliar Peso seco por área foliar (g/cm2) Densidad de la hoja
Densidad específica de la madera Peso seco por centímetro cúbico de madera (g/cm3)
Densidad aparente del suelo
Peso seco por centímetro cúbico de suelo (g/cm3)
Para convertir peso a área
Para convertir volumen de madera a masa de madera
Para calcular el peso del suelo y su contenido de nutrientes
Área basal de árboles Área transversal de los tallos por hectárea (m2/ha) Espacio que ocupan los árboles
Densidad de árboles Número de árboles por hectárea (número/ha)
Volumen de madera
Volumen de todos los tallos en una hectárea (m3/ha), se deriva de la relación alométrica entre área basal, altura y forma de los tallos
Peso seco por hectárea (Mg/ha)
Para cálculos de volumen y masa
Inventarios de madera
Para estimar la cantidad de la madera
Para estimar su biomasa (Mg/ha) al multiplicar por la densidad de la madera (g/cm3)
Biomasa
Se estima pesando todos los árboles en un área conocida; o por relación alométrica entre la masa de los árboles y su altura, diámetro y densidad de madera multiplicado por la densidad de árboles por hectárea
Cobertura del dosel El porcentaje de la superficie del bosque cubierta por vegetación (%)
Cociente largo:ancho foliar Largo dividido por el ancho de la hoja (cm/cm)
Fundamental para balances de masa y nutrientes y estimados de secuestro de carbono
Para evaluar condiciones de luz para la regeneración
Para evaluar la condición de la hoja El cociente es constante para la especie pero las dimensiones lineales varían con las condiciones; disminuyen con el estrés
Cociente área del dosel:diámetro del tallo Área en m2 y diámetro en cm Área fotosintética relativo al diámetro del tallo
Cociente área del dosel:biomasa Área en m2 y biomasa en Mg Área fotosintética relativo a la masa que respira Índice de superficie de tallos El área acumulativa de superficie de tallos por área de superficie del bosque (m2/m2)
* No incluye todos los posibles usos de estas medidas.
Para estimar la respiración de los tallos
menor de comportamiento (individuos) a escalas mayores (poblaciones y comunidades), por ejemplo, de una hoja a todo el bosque. Para lograrlo se requiere aplicar relaciones alométricas entre los componentes del ecosistema. La tabla 3.1 contiene once relaciones alométricas de uso común en la literatura ecológica. El uso de esas relaciones le permite al ecólogo extrapolar medidas en una escala menor a una escala mayor y viceversa.
El uso de las proporciones alométricas es común en la ecología, especialmente en los estudios de inventarios de bosques o en el estudio de la estructura de rodales arbóreos. El puertorriqueño Gilberto Cintrón Molero escribió un manual para el estudio de manglares (borrador disponible a través de Ariel E. Lugo) donde enfatizaba el uso de las relaciones alométricas. En la publicación de un resumen en 1984 (Cintrón
3 59 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
Tabla 3.2
Volumen sobre la corteza, biomasa y almacenaje aéreo de elementos y cenizas de poblaciones arbóreas de Spathodea campanulata en nueve sitios en el norte central de Puerto Rico (Lugo et al. 2011). Los datos son para árboles con un diámetro a la altura del pecho (dap) ≥ 15.0 cm, correspondiendo a la densidad arbórea en la segunda columna. Altura se refiere a la altura del dosel. El área basal (en paréntesis en los subtítulos) corresponde a la combinación de lugares por substrato (kárstico, aluvial y volcánico). El porcentaje de la densidad total de árboles con dbh > 10 cm correspondiente a Spathodea está en paréntesis en la columna 2. Los datos de volumen aplican al tallo principal y ramas grandes. Los datos fueron redondeados a m3, kg, o Mg más cercano.
y Schaeffer Novelli 1984) se ilustra como la alometría permite estimar la biomasa de los manglares partiendo de medidas simples de diámetro y altura de árboles. Lugo et al. (2011) expandieron el análisis para incluir estimados de las proporciones de elementos en la masa de poblaciones de Spathodea campanulata (Tablas 3.2 y 3.3).
La biomasa, el volumen de madera y los almacenajes de elementos en poblaciones de Spathodea en distintos substratos son altos, particularmente en los suelos aluviales y volcánicos, y proporcionales a la densidad de árboles en las poblaciones (r2 = 0.6 p <.02; Tabla 3.2). Los valores son más bajos en los substratos kársticos. Los valores en la tabla 3.2, basados solamente en las poblaciones de Spathodea, son tan o más altos que valores observados en bosques nativos, particularmente cuando la densidad de Spathodea es alta. Esta generalización es notable, ya que el rango de los diámetros de los árboles considerados es más extenso en los bosques y plantaciones que en las poblaciones de Spathodea (Tabla 3.3). Si se comparan poblaciones de Spathodea con poblaciones de tabonuco (Dacryodes excelsa), palma de sierra (Prestoea montana), pino caribe (Pinus caribaea) y caoba (Swietenia macrophylla) la mayor parte de los valores de masa, volumen y elementos son superiores en Spathodea, con la excepción del almacenaje de sodio, que es bajo en Spathodea
Para consultar los métodos a utilizarse en el estudio cuantitativo e interdisciplinario de los bosques de Puerto Rico, recomiendo a Domínguez Cristóbal (2011), y
3 60 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
Sitio Densidad de árboles (árboles/ha) Altura (m) Volumen (m3/ha) Biomasa (Mg/ha) NPK Ca Mg S Mn Na Al Fe Cenizas (Mg/ha) (kg/ha) Kárstico (54 m2/ha) Juan Nieves 414 (72) 24.4 265 96 309 48 338 480 101 93 0.34 12 422 Pugnado 580 (72) 29.7 503 177 585 84 632 895 190192 0.65 23 834 Ollas y Calderas 536 (73) 24.1 428 152 497 73 539 764 162 156 0.5520 723 Aluvial (84 m2/ha) Cibuco I 814 (83) 22.3 714 252 830 119 899 1272 269266 0.92 32 12 46 Cibuco II 770 (77) 25.3 849 296 988 137 1061 1497 318 343 1.10 38 14 56 Paso del Indio 1388 (83) 24.6 588 221 682 119 766 1098 228 165 0.73 29 10 35 Volcánico (58 m2/ha) Adjuntas 166 (28) 23.1 210 73 245 33 262 36979 85 0.27 9312 Perchas 1061 (68) 23.3 696 251 809 124 885 1257 265 238 0.89 32 12 45 Pozas 344 (44) 16.9 163 60 190 32 209 299 63 57 0.21 8311
Tabla 3.3
Datos estructurales, volumen sobre la corteza, biomasa y almacenaje aéreo de elementos y cenizas en los árboles de varios tipos de bosque en Puerto Rico. Los datos fueron redondeados al m3, kg, o Mg más cercano excepto para algunos almacenajes de fósforo. Área basal es AB y la referencia de los trabajos es Ref. Espacios sin datos significa que ese dato no fue reportado. El valor en paréntesis en la columna 1 es el diámetro a la altura del pecho (cm) correspondiente a los árboles medidos. Todos los rodales, con la excepción del bosque seco, estaban ubicados en el Bosque Experimental de Luquillo. El bosque seco estaba ubicado en Guánica. Todos los bosques son subtropicales sensu Holdridge 1967. Palo colorado es Cyrilla racemiflora, caoba es Swietenia macrophylla y el pino es Pinus caribaea
para calcular los almacenajes y flujos de nutrientes en hojarasca y suelos basadas en medidas de campo y laboratorio a Lugo (1999a).
Asignación
Selecciona 10 árboles y determina si hay alguna relación alométrica entre la circunferencia del árbol y el diámetro de la copa.
Alternativamente, calcula el C:N, C:P y N:P de 10 materiales biológicos como tejidos (plantas o animales), compuestos orgánicos o suelos.
3 61 CAP ariel e. lugo | ecología a m i m anera Estequiometría y Alometría
Sitio Altura (m) AB (m2/ha) Biomasa (Mg/ha) Volumen (m3/ha) NPK Ca Mg Cenizas Ref.* (kg/ha) Bosques Nativos Muy Húmedo Bisley (> 2.5) Cima 20-30 54.2 312 805 44 702 655 153 1 Pendiente 20-30 33.4 186 525 28 446 398 100 1 Valle Alto 20-30 21.3 121 391 21 295 243 97 1 Valle Ripario 20-3020.9 119 370 23 262 246 94 1 Dacryodes excelsa (solo) 20-30 12.6 103 181 10 170 116 21 1 Muy Húmedo El Verde 20-30 284 238 112 917 1,235 335 1,784 2 Muy Húmedo (>4) 20-30 198 350 3 Palmar 17 30.0 174 210 991 66 792 625 187 3, 4 Prestoea montana (solo) 17 18.9 72 541 45 508 195 115 4 Palo colorado 8-20 40.0 81-203 220 3, 5 Secundario Cubuy 19 28.0 90 504 30 423 24 6 Secundario Sabana 25 28.5 68 378 27 404 28 6 Secundario El Verde 24 33.8 78 379 25 336 16 6 Seco 16 98 3 61 981 7, 8 Plantaciones Anthocephalus chinensis 12.5 años 144 348 1,365 9 Pino 18.5 años 33 55.6 166 689 51651 443 90 6 Caoba 17 años 25 29.5 94 145 954 26 337 13 6 Caoba 49 años 27 31.9 121 419 34 388 66 *1.
Scatena et al. 1996, 2. Odum 1970, 3. Weaver y Murphy 1990, 4. Frangi y Lugo 1991, 5. Weaver 1995, 6. Lugo 1992, 7 Murphy y Lugo 1986, 8. Lugo y Murphy 1986, 9. Lugo y Figueroa 1995.
CaPítulo 4
La conservación en el Antropoceno
¿Qué es el Antropoceno? Es una época1 geológica caracterizada por el dominio de la actividad humana sobre el planeta. El proponente del Antropoceno como época geológica fue Paul Jozef Crutzen (2002), holandés y premio nobel de química. Su propuesta desató una polémica entre los geólogos que no pueden ponerse de acuerdo sobre su comienzo. Los geólogos necesitan una señal clara en el récord geológico para identificar las épocas geológicas y algunos argumentaron que el ser humano todavía no ha dejado tal señal en el planeta. Se estableció un comité para estudiar el asunto oficialmente y han surgido numerosas propuestas para identificar el comienzo del Antropoceno (Smith y Zeder 2013). Algunos argumentan que las grandes extinciones de especies por causa de la actividad humana denotan la influencia planetaria del ser humano. Otros apuntan al comienzo de la agricultura o la quema de combustibles fósiles que han transformado el clima. Aún otros señalan al año 1964, cuando el ser humano introdujo a la atmósfera nuevos isótopos radiactivos o en 1610, cuando la concentración de bióxido de carbono llegó a su punto más bajo en el récord atmosférico (Lewis y Maslin 2015).
Las implicaciones del Antropoceno para la ecología y la conservación (para mí, sinónimo del manejo; Lugo 1989) de los sistemas ecológicos son trascendentales, ya que el Antropoceno requiere una nueva forma de pensar al tratar estos asuntos (Lugo 2014). Las condiciones ambientales del Antropoceno exponen los ecosistemas del planeta a nuevos espacios ecológicos que difieren de los espacios ecológicos durante épocas pasadas. Los cambios aparentan ser irreversibles y continuos. El cambio ambiental continuo y direccional del Antropoceno sustituye la noción de los ciclos recurrentes que tipificaron el funcionamiento del planeta durante el Holoceno (la época de los glaciales).
¿Qué ha hecho el ser humano?
La figura 4.1 ilustra el efecto acelerante de la actividad humana en los ciclos biogeoquímicos de los elementos principales para la estequiometría del planeta (Sterner y Elser 2002). La actividad humana mueve más fósforo (400 % más), azufre (113 %
1 De acuerdo con la Enciclopedia Británica, una época geológica es una unidad de tiempo geológico durante el cual se depositan series de rocas; es una subdivisión de los periodos geológicos como el Paleógeno, Neogeno y Cuaternario.
ariel e. lugo | ecología a m i m anera
63
más) y nitrógeno (108 % más) que todos los restantes procesos globales. Más aún, la actividad humana mueve cuantiosas cantidades de otros elementos como el hidrógeno y oxígeno (16 % del flujo no-antropogénico) y carbono (13 % del flujo no-antropogénico). El efecto humano sobre el ciclo del carbono (capítulo 13) tiene como resultado el cambio climático (discutido más adelante en este capítulo). Otras actividades humanas modifican el ciclo del agua, la topografía del planeta, la distribución y abundancia de las especies de plantas y animales y el uso de los productos de la fotosíntesis del planeta (Tabla 4.1). La actividad humana también afecta el funcionamiento de los sistemas acuáticos, la acidez y distribución del calor en los océanos y el nivel del mar.
Tabla 4.1
Ejemplos de la magnitud del efecto humano sobre la topografía, sedimentación, apropiación de los servicios ecológicos y la alteración de ríos. Recopilado de varias fuentes de información.
Moviendo sedimentos y cambiando la topografía
Transporte de 2.3 X 109 Mg/año por los ríos
Reducción del flujo de sedimentos al mar por 1.4 X 109 Mg/año
Represas retienen > 100 billones de Mg de sedimentos
>50 % de superficie terrestre sin hielo ha sido modificada por la gente
Apropiación de los servicios de los ecosistemas
Consumimos entre 1/3 a 1/2 de la productividad del planeta
Los cultivos cubren 40 % de la superficie del planeta
87 % de las pesquerías comerciales del mundo están sobreexplotadas, totalmente consumidas o diezmadas
Alteración de ríos
Más de la mitad de los grandes ríos del planeta están afectados por represas, diques, canalizaciones, alteración de descargas y cambios en el transporte de agua, sedimentos y solutos de las tierras altas
El patrón de descarga de los ríos se ha homogenizado
4 64 CAP ariel e. lugo | ecología a m i m anera La conservación en el Antropoceno
Figura 4.1. Flujos globales de carbono, nitrógeno, fósforo, azufre y oxígeno e hidrógeno atribuidos a la actividad humana (antrópico) y al resto de la biosfera (no-antrópico). El porcentaje de cambio al flujo global atribuible a la actividad humana está en paréntesis. Noten la escala logarítmica. Los datos se obtuvieron de Sterner y Elser (2002).
Intensivo
Figura 4.2. Mapa de los Antromas o biomas antropogénicos de acuerdo con Ellis (2021). El mapa señala las tierras utilizadas por la actividad humana con tierras en distintos niveles de estados no afectados por el ser humano. La intensidad de uso aumenta de izquierda a derecha en la clave de la figura y la intensidad de los colores aumenta con la intensidad de los usos. Las seis grandes categorías de uso son (de izquierda a derecha): tierras salvajes, tierras con usos culturales, pastizales, tierras de cultivo, pueblos y asentamientos densos. Dentro de cada categoría se identifican variantes en la intensidad de los usos. Figura cortesía de E. C. Ellis (2022).
El ser humano ha domesticado especies, modificado el genoma de organismos, introducido luz artificial al ambiente, sintetizado nuevas sustancias químicas, utilizado la energía nuclear y ha interconectado todo el planeta con el transporte y las comunicaciones globales. La actividad humana también ha transformado la cobertura terrestre (Ellis et al. 2010, Ellis 2015, 2021). La cobertura del planeta, que en el siglo pasado se analizaba en términos de biomas (discutidos en el próximo capítulo), ahora se clasifica en Antromas o Anthromes (Fig. 4.2), que son coberturas producto de la actividad humana (Ellis 2021). De acuerdo con este sistema de Antromas, el planeta solo tiene aproximadamente un 23 % de cobertura libre de aparentes efectos de la actividad humana. Muchos sugieren que ya en el planeta no existen áreas «vírgenes» o«silvestres» sin efectos o presencia de la huella humana. La gente utiliza el 40 % de la superficie global y los ecosistemas noveles cubren el 37 %. El ser humano ha podido asumir la dominancia en el planeta gracias al consumo de energía fósil. Todas las actividades de organismos y de procesos ecológicos

4 65 CAP ariel e. lugo | ecología a mi ma nera La conservación en el Antropoceno
Asentamientos densos Tierras de cultivo Pueblos Pastizales
Tierras con usos culturales Tierras salvajes
Urbano Asentamiento mixto
Residencial con riego Residencial de secano Pobladas Remotas
Arroz Irrigado Secano Pastoral
Residenciales Poblados Remotos
Bosques residenciales Bosques poblados Bosques remotos Tierras secas habitadas
Bosques salvajes Tierras secas salvajes Hielo
requieren energía y con el uso de la energía fósil el ser humano incrementó el flujo de energía de todo el planeta como se observa en la figura 4.3. Allí se presentan todas las energías que mueven el planeta Tierra. La energía solar, geotermal y mareal son las fuentes de energía del planeta antes del Antropoceno. Se han mantenido constantes. La energía fósil es de origen reciente, asociada al ser humano y es en órdenes de magnitud más potente que las primeras tres. Mientras dure, la energía fósil facilita el dominio humano sobre el planeta.
Puerto Rico es ejemplo de una unidad geográfica bajo los efectos del Antropoceno (Lugo 2004b). La tabla 4.2 resume algunos de los efectos de la actividad humana en esta isla. Rivera Collazo (2015) estima que la actividad humana comenzó en Puerto Rico hace 4,500 años. He argumentado en el pasado (Lugo 2006) que los procesos sociales y ecológicos en Puerto Rico anticipan el futuro estado de áreas continentales, ya que la isla ha sido sometida a condiciones típicas del Antropoceno por un periodo de tiempo más largo e intensidades superiores a lo que se observa en los continentes. Por sus limitaciones de espacio, las islas están más expuestas a los efectos de la actividad humana que los continentes. Por eso, Puerto Rico no es la única isla expuesta a los efectos de intensa actividad humana. Otras islas caribeñas (Lugo et al 2012c), las de Hawái (Mascaro et al. 2012) o las Seychelles (Kueffer y Kaiser-Bunbury 2013), se
Deforestación masiva resultó en la pérdida de 99 % de los bosques originales y 95 % de la cobertura vegetal
22 % de la cobertura está urbanizada
11 % de la cobertura se ha impermeabilizado
450 personas por km2
Especies introducidas exceden el 25 % de la flora y fauna
Importación por encima del 80 % del alimento y más del 90 % de los combustibles que necesita
3.8 km de carreteras por km2 de superficie
* Algunos indicadores han cambiado pero no significativamente para propósitos ilustrativos.
4 66 CAP ariel e. lugo | ecología a mi ma nera La conservación en el Antropoceno
Tabla 4.2
Cambios al paisaje de Puerto Rico y otros indicadores* de actividades humanas que señalan la presencia del Antropoceno en la Isla (Lugo 2004a y otras fuentes).
Figura 4.3. Las fuentes de energía que potencian todos los sistemas del planeta Tierra. De estas cuatro, solo la energía fósil ha cambiado a través de la historia del planeta. La aceleración de su uso durante el siglo XX ha permitido el desarrollo de la civilización moderna al igual que el cambio en el clima.
comportan similarmente a Puerto Rico. Sin embargo, el caso de Puerto Rico ha sido documentado con más detalle y por más tiempo que en otras islas.
Calentamiento global y el cambio climático
El calentamiento global y el cambio climático se atribuyen a la actividad humana. Estos efectos de la actividad humana fueron una de las principales razones que indujeron a Crutzen a proponer que el Holoceno había terminado. Las leyes de conservación de energía y masa presentadas anteriormente ayudan a explicar el cambio climático. Durante el Holoceno, la entrada de energía del sol al planeta estaba balanceada por su retorno al espacio en forma de calor. El clima se mantenía estable. Sin embargo, con la quema de combustibles fósiles, el ser humano incrementa la generación de calor en el planeta, ya que la energía generada por los combustibles fósiles es muy superior a la energía disponible al ser humano durante el holoceno (Fig. 4.3).
Más aún, la actividad humana altera los ciclos de gases atmosféricos al producir altas cantidades de gases como el bióxido de carbono, metano, ozono, y óxidos de nitrógeno conocidos como gases con efecto de invernadero (de ahora en adelante gases de invernadero). Estas emisiones alteran la química de la atmósfera, ya que los gases de invernadero interaccionan entre sí y con los gases ya presentes en la atmósfera. Además, los gases de invernadero retienen parte del calor que se genera en el planeta y evitan que se escape al espacio, contribuyendo así al calentamiento de la atmósfera. La figura 4.4 compara la capacidad de retención de calor de los gases de invernadero y documenta el efecto neto de los procesos que calientan o enfrían el planeta.
Las altas concentraciones de gases de invernadero en la atmósfera y las aguas de los océanos retienen el calor dentro del planeta en vez de devolverlo al espacio. La retención de energía en la atmósfera y el mar causa que se caliente el planeta. Se espera que en el próximo siglo se observarán las temperaturas más altas en 65 millones de años (Diffenbaugh y Field 2013). Al indicar que el clima cambia, se entiende que el clima describe los promedios a largo plazo (basado en datos de 30 o más años) de las variables que describen el estado del tiempo (temperatura, lluvia, humedad). El estado del tiempo se refiere a los eventos climáticos a corto plazo (días a meses).
Balance de gases: C, N, O3
La composición gaseosa de la atmósfera es dinámica debido a que los procesos como la fotosíntesis y la respiración continuamente modifican la concentración de los gases atmosféricos. Por ejemplo, la fotosíntesis produce oxígeno y remueve bióxido de carbono de la atmósfera, mientras que la respiración consume oxígeno y emite bióxido de carbono a la atmósfera. Dependiendo del balance entre estos dos procesos (fotosíntesis y respiración), la composición atmosférica de los gases envueltos va
4 67 CAP ariel e. lugo | ecología a m i m anera La conservación en el Antropoceno
Figura 4.4.
invernadero en desviar la radiación incidente sobre el planeta hacia la superficie de la tierra (valores positivos en rojo) medido en watts por metro cuadrado. Valores negativos en azul significan que la energía se refleja fuera del planeta. Modificado de Intergovernmental Panel on Climate Change (2007). El efecto neto de todas las actividades (última fila) es significativamente positivo, sobrepasa el efecto de la radiación solar y es el mecanismo por el cual se calienta la atmósfera.
a cambiar con el bióxido de carbono, aumentando cuando predomina la respiración y disminuyendo cuando predomina la fotosíntesis. Algunos organismos remueven nitrógeno de la atmósfera y otros lo añaden. Sin embargo, como la atmósfera tiene altos porcentajes de nitrógeno y oxígeno, la concentración de estos gases no cambia tanto como la del bióxido de carbono (Figs. 4.5 y 4.6). La concentración del bióxido de carbono cambia dramáticamente en la atmósfera, ya que su concentración es baja y los procesos biogeoquímicos pueden alterarla significativamente. El ozono es un gas que se produce y se pierde en la estratosfera donde la luz ultravioleta del sol acelera la fotolisis del oxígeno y la formación de ozono. En el proceso se absorbe la luz ultravioleta que es nociva para la vida en el planeta. El ozono se pierde al combinarse con átomos de oxígeno para formar moléculas de oxígeno. Si la concentración de ozono baja en la estratosfera, aumenta la entrada de luz ultravioleta al planeta, poniendo en riesgo la vida.
4 68 CAP ariel e. lugo | ecología a mi ma nera La conservación en el Antropoceno
Forzamiento radiativo o la influencia de varios gases de
Figura 4.5. Concentración mensual promedio de bióxido de carbono en la atmósfera del Observatorio de Mauna Loa, Hawái y La Parguera, Puerto Rico. La información está disponible en caricoos. org y fue accesada el 6 de septiembre de 2021. CARICOOS es la sigla en inglés del Sistema Caribeño de Observación Costera y Oceánica ubicado en La Parguera, Puerto Rico. El punto rojo indica la concentración de bióxido de carbono en el momento indicado.
Figura 4.6. ( A ) Concentraciones de bióxido de carbono y oxígeno en la atmósfera de varios lugares en el planeta a través de 35 años. Un meg es equivalente a una molécula de oxígeno por una de nitrógeno, en otras palabras el cociente oxígeno:nitrógeno. (B) Las emisiones globales de combustibles fósiles (línea negra) y el porcentaje del C13 en la atmósfera. La escala del isótopo de carbono está invertida para subrayar el hecho de que denota una proporción mayor del C 13 en años recientes. Este récord es del Intergovernmental Panel on Climate Change (2007). La misma organización ha publicado récords más recientes que muestran la continuidad del mismo patrón. Se obtiene en https:// www.ipcc.ch/reports/.
Expansión del agua
Cuando la temperatura del aire o el mar aumentan, tanto los gases atmosféricos como el agua del mar se expanden. Esto se debe a que el calor aumenta el movimiento de las moléculas que componen el aire y el agua de mar, causando que se alejen unas de otras y que se expandan los gases y el agua. En contraste, cuando la temperatura baja, el agua y los gases se contraen debido a que al perder energía, el movimiento molecular disminuye y eso causa que las moléculas se acerquen unas
4 69 CAP ariel e. lugo | ecología a m i m anera La conservación en el Antropoceno
a otras concentrando las masas de agua y aire. Masas de aire y agua fría tienden a hundirse por ser más densas que masas de aire y agua caliente. Como el planeta se está calentando, la tendencia para las masas de agua y aire es la expansión. En el caso de las aguas de los océanos, el calentamiento del agua causa que los niveles del mar aumenten, ya que el agua caliente ocupa más espacio que la fría.
Cambio en el pH del agua
Normalmente las lluvias son levemente ácidas, ya que el bióxido de carbono en la atmósfera se combina con el agua para formar el ácido carbónico. En el mar, altas concentraciones de carbonato neutralizan el ácido de las lluvias. Sin embargo, la actividad humana aumenta la concentración de bióxido de carbono en la atmósfera y por consiguiente la producción de ácido carbónico también aumenta. La quema de basura, la combustión de combustibles fósiles en los vehículos, la manufactura de cemento y los fuegos también producen ácidos, en algunos casos con altos niveles de acidez (incluidos los ácidos sulfúrico y clorhídrico). Estas altas emisiones de ácidos alteran la capacidad de amortiguamiento químico de las aguas marinas, y por consiguiente, acidifican las aguas costeras. Para amortiguar la acidez en el mar, se disuelve el carbonato de calcio que es el material estructural de arrecifes de coral, algunas algas y el plancton marino. Todo organismo marino que dependa del carbonato de calcio para producir su estructura celular está amenazado por la acidificación del océano.
Causas no-antropogénicas vs . antropogénicas del cambio climático:
¿Cuál es la evidencia?
La concentración de bióxido de carbono está aumentando en la atmósfera desde que se comenzó a medir en los años 50 en Mauna Loa, Hawái. En el presente, la concentración de bióxido de carbono se mide continuamente a través de todo el planeta y todas las estaciones de medición reportan aumentos, incluso en Puerto Rico (Fig. 4.5). Como el bióxido de carbono es un gas de invernadero (discutido anteriormente) el aumento en su concentración en la atmósfera es responsable de una fracción importante del calentamiento global. Algunos preguntan ¿cuál es el origen del bióxido de carbono en la atmósfera?
Para contestar esa pregunta se determina la presencia de isótopos de carbono en el bióxido de carbono. Un isótopo es un átomo de un elemento con el mismo número de protones pero distinto número de neutrones que otros átomos del mismo elemento. Esto resulta en una diferencia en la masa de los átomos. A pesar de la diferencia en masa, todos los isótopos de un elemento se comportan igual químicamente. El carbono tiene tres isótopos: uno con una masa de 12 (C12 , el más común, 99 %), otro con masa de 13 (C13) y uno con masa de 14 (C14). Existen metodologías para medir la cantidad de cada uno de estos isótopos en el bióxido de carbono de la atmósfera.
4 70 CAP ariel e. lugo | ecología a m i m anera La conservación en el Antropoceno
Figura 4.7. Anomalías de la temperatura del aire entre 1888 y 1998 medidas en distintas elevaciones en Puerto Rico ( A). Las anomalías aumentan con la elevación. Las líneas entrecortadas indican la tendencia en la anomalía. Estos datos se basan en pocas estaciones con récords largos. (B). Datos de la temperatura diaria (máxima y mínima) promedio del aire en la estación del Aeropuerto Internacional de San Juan (Carolina) no reflejan tendencias de calentamiento a través del tiempo (cortesía de Ferdinand Quiñones Márquez).

Temperaturas máximas y mínimas en el aeropuerto LMM en Carolina, PR 1956-2022
Temperatura máxima y mínima (ºF)
El petróleo se forma de la materia orgánica producida por la fotosíntesis de plantas que vivieron en épocas pasadas. Cuando extraemos ese petróleo, el mismo refleja la proporción atmosférica de los isótopos de carbono disponibles en la atmósfera a las plantas al momento de fabricar la materia orgánica. En épocas pasadas la proporción C13/C12 en el bióxido de carbono era más baja que en el presente. Por lo tanto, el petróleo y sus productos derivados tienen una huella química que los identifica donde quiera que estén. Es entonces posible analizar las proporciones de los isótopos de carbono del bióxido de carbono en la atmósfera presente para determinar su origen y para distinguirlo del bióxido de carbono que emiten las plantas, animales, los fuegos y volcanes de hoy. Al llevar a cabo estas pruebas, se encuentra que la mayor parte del bióxido de carbono en la atmósfera procede de la quema de combustible fósil y no de los sistemas del presente. La tendencia en la atmósfera presente es un paulatino cambio negativo en los valores del porcentaje de C13 en el bióxido de carbono de la atmósfera presente (Fig. 4.6). El patrón es consistente con el aumento en la tasa de emisión de productos de combustión fósil con valores del porcentaje de C13(CO2) más bajos que los de la atmósfera del presente.
Cambio de las condiciones en Puerto Rico
Temperatura. De acuerdo con el Puerto Rico Climate Change Council (2013), la temperatura promedio en Puerto Rico ha aumentado de 0.012 a 0.014 oC por año desde el año 1900 al presente. Hay evidencia de que la temperatura del aire en las zonas montañosas también está aumentando (Fig. 4.7a). Se espera que la temperatura promedio
4 71 CAP ariel e. lugo | ecología a m i m anera La conservación en el Antropoceno
del aire en Puerto Rico aumente unos 4 grados centígrados durante este siglo. La temperatura de las aguas superficiales del mar Caribe aumentó 1.5 oC en los últimos 100 años. Sin embargo, un análisis estadístico del récord de temperaturas diarias en el aeropuerto de San Juan por el hidrólogo puertorriqueño Ferdinand Quiñones Márquez no detecta tendencias de aumento en los pasados 100 años (Fig. 4.7b). Lluvia. Las tendencias en la lluvia de Puerto Rico bajo el efecto del cambio climático no son claras y pueden variar geográficamente a través de la Isla. Se espera un aumento en eventos extremos de lluvia y sequía a medida que progrese este siglo. Larsen (2000) encontró que durante el siglo XX las lluvias en Puerto Rico disminuyeron un milímetro anual, equivalente a una reducción de 2,351 millones de galones de agua por año. Sin embargo, un análisis de tendencia utilizando estaciones meteorológicas con datos de más de 30 años, no encontró tendencia alguna en el patrón de las lluvias en Puerto Rico (Fig. 4.8). La conclusión del estudio fue:
«Sin embargo, y según los datos de lluvia histórica en Puerto Rico, concluyo, sin muchas dudas, que el promedio de lluvia anual en la isla no ha disminuido al menos desde el 1920 hasta el 2020, lo que contrasta con resultados frecuentemente reseñados en la literatura científica y destacados
(Quiñones Márquez 2021, p. 13-14).
Huracanes. No hay evidencia de que la frecuencia o intensidad de los huracanes hayan aumentado en Puerto Rico. Teóricamente, ambas deben aumentar en el futuro debido al calentamiento del mar y la atmósfera.
4 72 CAP ariel e. lugo | ecología a m i m anera La conservación en el Antropoceno
en agencias gubernamentales en general»
Figura 4.8. Precipitación promedio anual en Puerto Rico desde 1900 a 2020 y el promedio corriente (línea verde) basado en 30 años (Quiñones Márquez 2021). No se observa ninguna tendencia en la disminución de la lluvia. Las barras rojas representan años con lluvia promedio anual menor a 60 pulgadas (1524 mm). Para convertir pulgadas a mm multiplique por 25.4.
Nivel del mar. El nivel del mar ha subido continuamente en Puerto Rico desde que se comenzó a medir en 1956 (Fig. 4.9). La tasa de aumento ha sido un promedio de 1.4 mm/año, pero recientemente ha aumentado a 3.79 mm/año, lo que coincide con el patrón global. Globalmente, la velocidad en el aumento del nivel del mar es 4.4 + 0.5 mm/año (Yi et al. 2015). Sin embargo, tanto las tasas globales como las que se miden en Puerto Rico cambian anual y estacionalmente dependiendo de la velocidad de cambio en la temperatura del mar y del derretimiento de los glaciales. Acidificación del mar. Est u d ios en la Parguera indican un aumento en la frecuencia de muestras con valores bajos de pH (Fig. 4.10).
Efectos del cambio climático
En regiones urbanas. Las regiones urbanas por su naturaleza acumulan calor. Durante el día, los materiales que se utilizan para construir la infraestructura gris (la construida) de la ciudad absorben más energía de la que reflejan. De noche, estos materiales irradian calor a la atmósfera. Además, la ciudad tiende a ser seca (poca agua y vegetación) por lo que el enfriamiento por el proceso de evapotranspiración es mínimo. Por ejemplo, los techos y paredes exteriores de edificios y las carreteras se sobrecalientan de día y emiten calor toda la noche, ya
4 73 CAP ariel e. lugo | ecología a m i m anera La conservación en el Antropoceno
Figura 4.9. Anomalías en el nivel promedio mensual del mar en Isla Magueyes (desde 1955) y Bahía de San Juan (desde 1962). Las líneas de tendencia fueron suavizadas. Cortesía de Aurelio Mercado Irizarry. El promedio del nivel del mar es la línea cero.
Figura 4.10. El pH del agua de mar en la vecindad de la Parguera. La información está disponible en caricoos.org y fue accesada el 6 de septiembre de 2021. CARICOOS es la sigla en inglés del Sistema Caribeño de Observación Costera y Oceánica ubicado en La Parguera, Puerto Rico.
que el concreto y la brea absorben pero no reflejan energía. Esto causa la Isla de Calor en zonas urbanas comparado con los alrededores de la ciudad (Murphy 2007).
La Isla de Calor es el aumento en la temperatura en la ciudad comparado con los alrededores no urbanos (Fig. 4.11). En los alrededores de la ciudad donde abunda la vegetación y el agua, la energía del sol se utiliza por la fotosíntesis y evapotranspiración o se refleja durante el día, reduciendo lo que se emite de noche. Como consecuencia, las temperaturas son más bajas y oscilan menos que en la ciudad. La Isla de Calor resulta en una ciudad más caliente de lo normal y cualquier calentamiento adicional a causa del cambio climático empeora la situación. En San Juan se ha estimado que desde el año 1900, la temperatura de la ciudad ha aumentado 0.022 o C por año, más rápido que el resto de la isla.
Altas temperaturas, particularmente cuando ocurren durante varios días consecutivos (olas de calor), afectan la salud humana. Los niños y ancianos son particularmente vulnerables al calor excesivo. Las altas temperaturas también obligan a las personas a utilizar aires acondicionados y abanicos eléctricos, lo que aumenta el gasto energético de la ciudad. Eso a su vez aumenta las emisiones de gases de invernadero para crear un ciclo vicioso. Lo mismo ocurre con el aumento en el uso del agua para mitigar el calor, ya que la misma se bombea a la ciudad con el uso de maquinaria que emite gases de invernadero. La economía se afecta por sequías y altas temperaturas, ya que la población altera su horario de actividades y toma medidas que merman la productividad económica. En regiones agrícolas. Los cultivos son vulnerables al cambio climático, ya que su productividad depende de las condiciones atmosféricas y la ausencia de disturbios. Un aumento en temperatura disminuye la productividad de cultivos y animales. Por ejemplo, durante una sequía o periodos con temperaturas altas extremas, los cultivos no crecen y las vacas producen menos leche. Sequías o inundaciones extremas también disminuyen la productividad agrícola y causa daños a la infraestructura que apoya la agricultura. Para contrarrestar los efectos negativos del cambio climático es necesario incrementar el manejo de los cultivos, lo cual cuesta tiempo,
4 74 CAP ariel e. lugo | ecología a m i m anera La conservación en el Antropoceno
Figura 4.11. La Isla de Calor asociada a la zona metropolitana de San Juan (Murphy 2007).
dinero y combustibles. El aumento en los costos del manejo le resta rentabilidad a la actividad agrícola. La quema de más combustibles para sobreponerse a los efectos negativos del cambio climático genera más gases de invernadero, lo que a la larga empeora la situación. En regiones rurales. Los sistemas ecológicos pueden adaptarse al cambio climático paulatino, pero al hacerlo ocurren cambios en la productividad de los ecosistemas y su composición de especies. Por ejemplo, los cambios en temperatura obligan a los animales a moverse a otros lugares con condiciones más favorables. En Puerto Rico, los lagartijos de la especie Anolis gundlachi, un lagartijo arbóreo de El Yunque se ha movido hacia las tierras bajas donde bosques noveles (discutidos más adelante en este capítulo) contienen condiciones favorables para su crecimiento y allí viven debajo de la copa de los árboles. Aunque cambiaron de clima (de la montaña a las tierras bajas), es posible que el cambio se debió a la reforestación de Puerto Rico y no necesariamente al calentamiento del clima. Es posible que estos lagartijos estaban refugiados en El Yunque cuando se deforestaron las tierras bajas. Estudios en El Yunque, sin embargo, han encontrado movimiento de algunas aves entre distintas elevaciones, posiblemente en respuesta a cambios de temperatura, pero cada especie sigue un patrón de respuesta distinto (Campos Cerqueira et al. 2017).
Tanto los animales como las plantas se han movido en el pasado en respuesta a cambios ambientales y seguirán en movimiento con los cambios climáticos futuros causados por el calentamiento global y otros cambios ambientales. La velocidad del cambio climático sobre el espacio geográfico es más rápida en el presente, y lo será en el futuro, que en el pasado (Diffenbaugh y Field 2013). Se define la velocidad de cambio climático como la distancia por unidad de tiempo que una especie necesita moverse para mantener la temperatura de su hábitat dentro de los límites del entorno presente. Diffenbaugh y Field (2013) estimaron velocidades de varios kilómetros por año en la zona de sus estudios.
Al constituir nuevas comunidades de plantas y animales es posible que cambien algunos servicios ecológicos que recibimos de estos sistemas. Por ejemplo, la calidad de la madera en un bosque novel es menor a la calidad de la madera de bosques nativos2 maduros. Sin embargo, el bosque novel puede sustentar aves introducidas de gran belleza como los guacamayos. Ambos tipos de bosque (noveles y nativos) sostienen la biodiversidad, almacenan carbono, contrarrestan el calentamiento climático y proveen aguas puras.
2 Los bosques noveles, producto de la actividad humana, se contrastan con los bosques nativos, que son bosques precolombinos o bosques históricos compuestos enteramente por especies nativas. En Puerto Rico, los bosques noveles son dominados por especies introducidas. En los continentes, especies nativas forman bosques noveles (Lugo et al. 2020).
4 75 CAP ariel e. lugo | ecología a m i m anera La conservación en el Antropoceno
Los arrecifes de coral se blanquean ante las altas temperaturas del mar. El blanqueamiento se debe a la expulsión de las algas que conviven con los animales que forman el arrecife. Sin el alga, no hay fotosíntesis y los corales mueren por falta de alimento. El aumento en la acidez del agua disuelve el carbonato de calcio en el arrecife, por lo que representa una segunda amenaza a estos sistemas a causa del cambio climático.
Adaptación: la idea de lo novel o novedoso en el Antropoceno
Los cambios asociados al Antropoceno ponen a prueba los mecanismos de adaptación de los sistemas sociales-ecológicos-tecnológicos (SETS, por sus siglas en inglés; definido en McPhearson et al. 2022 y Lugo 2020). Cuando un SETS se enfrenta a un disturbio, ya sea no-antropogénico o antropogénico, se determina su nivel de adaptabilidad al disturbio por el grado de resiliencia que exhibe ante el fenómeno. La resiliencia es la capacidad de un sistema para absorber la fuerza de un disturbio y continuar funcionando. Un sistema con resiliencia resiste y se recupera de un disturbio, mientras que un sistema sin adaptaciones al disturbio tiene poca resiliencia hacia ese tipo de disturbio y como consecuencia no lo resiste y puede ser sustituido por otro sistema mejor adaptado.
Hutchinson (1965) relacionó la evolución de adaptaciones bióticas con la ecología al usar la analogía de que la situación ecológica representaba el teatro donde la
4 76 CAP ariel e. lugo | ecología a m i m anera La conservación en el Antropoceno
Figura 4.12. Diagramas de las ideas de Hutchinson (1965) sobre la situación ecológica (el teatro) y la trama de la evolución dentro del teatro, pero bajo la influencia de fuerzas externas (círculos). (A) La situación durante el Holoceno según la describió Hutchinson. (B) Los ajustes de los colegas de Hutchinson donde los organismos afectan la situación ecológica. (C) Las ideas adaptadas al Antropoceno con una nueva fuerza externa y la aceleración del desarrollo de sistemas noveles.
Figura 4.13. Ilustración del mecanismo de selección que favorece el desarrollo de sistemas noveles en el Antropoceno (Lugo et al. 2018). Bajo condiciones noveles tanto las especies introducidas como las locales (nativas) compiten y pueden formar parte de los sistemas noveles. El movimiento de especies bajo la influencia de la energía fósil que potencia la civilización permite que especies de todas partes del planeta tengan oportunidad de competir por los nuevos ambientes creados por la actividad humana. Sin embargo, para formar parte de los sistemas noveles las especies introducidas deben pasar por varios filtros (se ilustran dos: biogeográfico y local) que determinan su adaptabilidad a nuevas condiciones de vida.
trama era la evolución (Ecological Theater and Evolutionary Play, Fig. 4.12a). Otros científicos añadieron que la ecología y la evolución están interrelacionadas por medio de retroalimentaciones entre las poblaciones participantes en la trama evolutiva y las condiciones ecológicas en el teatro (Fig. 4.12b). En el Antropoceno, la actividad humana subsidiada por energía fósil transforma el teatro, mezcla la diversidad genética en forma impredecible, y participa en la trama evolutiva (Fig. 4.12c). El resultado inmediato es la aceleración de la evolución, la generación de nuevas adaptaciones y el movimiento de la biota hacia la formación de sistemas novedosos o noveles.
El mecanismo evolutivo y de selección natural se aplica a la generación de bosques noveles bajo influencia humana en la figura 4.13. En Puerto Rico, el 75 % de los bosques son bosques noveles (Fig. 4.14) compuestos por nuevas combinaciones de plantas y animales nativos e introducidos (Lugo y Helmer 2004, Lugo et al. 2012b). La Isla se ha enriquecido tanto en la diversidad de condiciones del teatro ecológico, como en el número de especies naturalizadas y capaces de sobrevivir bajo las nuevas condiciones ambientales (Tabla 4.3). Los bosques y otros ecosistemas noveles representan la respuesta de la biota a los cambios ambientales del Antropoceno (Lugo 2013).
Adaptaciones al nivel SETS
Para adaptarnos al cambio climático necesitamos enfoques social-ecológicostecnológicos, en otras palabras, soluciones integradas que tomen en cuenta todos los
4 77 CAP ariel e. lugo | ecología a m i m anera La conservación en el Antropoceno
Bosques nativos
Bosques noveles
Figura 4.14. Distribución geográfica de la dominancia en comunidades arbóreas en el 2003 cuando se llevó a cabo el inventario de bosques de Puerto Rico (Brandeis et al. 2007, 2009). Los colores de la leyenda reflejan el Valor de Importancia en porcentaje de las especies introducidas. Cero % (verde) indica bosques nativos históricos y los valores mayores de 1 al 100 % reflejan el grado de novedad de la comunidad, basado en su composición de especies (Martinuzzi et al. 2013).

Tabla 4.3
Número de especies introducidas y naturalizadas en Puerto Rico (animales) y el Caribe (plantas) (Lugo et al. (2012a,c).
aspectos que influencian un SETS. Un ejemplo es la conversión de la ciudad de un lugar vulnerable al calor, a un sitio fresco y agradable para la actividad humana con menos dependencia de la energía fósil. Ese cambio requiere incorporar elementos verdes (vegetación), azules (agua) y púrpura (humedales) a la ciudad que normalmente es gris por su infraestructura construida. Espacios verdes abiertos y agua en movimiento contrarrestan la Isla de Calor, enfrían la ciudad, reducen la necesidad del uso de acondicionadores de aire y la hacen más atractiva para vivir. Tales cambios requieren diálogo entre personas de todos los sectores y aceptación social para
4 78 CAP ariel e. lugo | ecología a m i m anera La conservación en el Antropoceno
Grupo Número de especies Introducidas (%) Oligochaetes 29 39.3 Formicidae 77 23.4 Anfibios 24 25.0 Reptiles 55 7.3 Aves 354 12.7 Mamíferos de cuatro patas 13 100.0 Todos los mamíferos 62 16.1 Familias de plantas 231 13 Géneros de plantas 1,953 26 Especies de plantas 12,847 15 Leyenda 0 % 1 a 19 % 20 a 39 % 40 a 59 % 60 a 79 % 80 a 100 % Tierras no forestadas Sin datos
que funcione. La sociedad también puede decidir sustituir infraestructura verde por infraestructura gris cuando tal sustitución es conveniente. Por ejemplo, alejar estructuras de áreas inundables y sustituirlas por zonas riparias con vegetación da como resultado múltiples beneficios para combatir el cambio climático. Si ocurren eventos de lluvia extremos, estas zonas riparias acomodan la inundación y protegen las estructuras contra daños. Si la temperatura aumenta, estas zonas ayudan a moderar la temperatura. Si ocurre una sequía, estas zonas ayudan a conservar el agua. Si construimos detrás de las dunas de arena, evitamos problemas con el aumento en el nivel del mar y con las marejadas, ya que las dunas absorberían la energía de esos eventos en lugar de las estructuras donde vive la gente. Mejor aún, la infraestructura verde es mucho más barata que la infraestructura gris y cuesta menos mantenerla.
4 79 CAP ariel e. lugo | ecología a m i m anera La conservación en el Antropoceno
CaPítulo 5 Los bosques tropicales
Pregunta
¿Qué es un bosque tropical y cuántos tipos de bosques tropicales existen?
Para contestar esta pregunta hay que entender un poco sobre la clasificación de la vegetación y la biogeografía, la cual se inició con los grandes exploradores del Nuevo Mundo. Científicos europeos y de EE. UU. estudiaban regiones climáticamente homogéneas y por eso inicialmente no le dieron tanta importancia a los factores climáticos. Enfatizaron los factores locales tales como el suelo, la exposición, o la topografía. El campo de la geografía es responsable por conceptos como el bioma y las formaciones vegetales y por los primeros intentos de entender y clasificar la vegetación del mundo. Las ideas basadas en geografía dominan aún la clasificación de la vegetación, particularmente con el trabajo de Robert G. Bailey, un geógrafo norteamericano del Servicio Forestal de los EE. UU. Sin embargo, ecológicamente, la más objetiva y efectiva forma de clasificar y entender la vegetación global y el funcionamiento de los ecosistemas es el sistema de zonas de vida de Holdridge, el cual enfatizaremos en este capítulo.
Trasfondo histórico
De acuerdo con E.P. Odum (1953), el bioma es la unidad comunitaria más grande que se puede reconocer en términos de cobertura geográfica. Se define por el clima, los suelos y la vegetación clímax. «Formaciones vegetales» es un término que es sinónimo con bioma. En 1807, el alemán Alexander von Humboldt, reconocido como el padre de la geografía moderna universal, relacionó el clima con la geografía de la vegetación cuando escribió sobre el volcán Chimborazo en Ecuador (Fig. 5.1). El volcán está inactivo y tiene la distinción de que su cima es el punto más distante del centro del planeta.
A finales del siglo XIX, el alemán Andreas Franz Wilhem Schimper y el danés Eugenius Warming resumieron el conocimiento ecológico disponible y formalizaron la geografía de la vegetación desde un punto de vista fisiológico (Schimper) y
ariel e. lugo | ecología a mi ma nera
81
ecológico (Warming). Andrew John Herbertson (1905), geógrafo escocés, definió la geografía como «la ciencia de las distribuciones» y elaboró un mapa de las regiones naturales del planeta en función del clima y la vegetación (Fig. 5.2). Herbertson además sugirió que las actividades humanas eran influenciadas por las condiciones ambientales en las regiones donde ocurrían.
En 1894 el norteamericano Clinton Hart Merriam desarrolló el concepto de zonas de vida definidas por la temperatura, pero no corrigió por elevación y su aplicación fue principalmente para el oeste de los EE. UU. El sistema se utiliza para exponer la distribución de animales, pero no la vegetación. El Departamento de

5 82 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Figura 5.1. Portada de la traducción del ensayo sobre la geografía de las plantas de Alexander von Humboldt y Aimé Bonpland (1807), versión de tapa blanda de 2013 (Jackson y Romanowski 2009). La portada fue diseñada por Maia Wright de la Biblioteca Central del Museo Nacional de Historia Natural de París. La ilustración original de Humboldt del Monte Chimborazo en Ecuador se considera un ícono de la ciencia e ilustración científica.
Figura 5.2. Las principales seis regiones naturales del planeta Tierra según Herbertson (1905). La clave es: 1. Regiones polares: tierras bajas (a) y tierras altas (b). 2. Regiones con temperaturas frías: margen occidental (a), margen oriental (b), tierras bajas interiores, (c) áreas interiores montañosas (d). 3. Regiones con temperaturas cálidas: margen occidental con lluvias en invierno (a), margen oriental con lluvias en verano (b), tierras bajas interiores (c), mesetas (d). 4a desiertos tropicales del oeste. 4b Tierras tropicales del este. 4c Mesetas intertropicales. 5 Altas montañas tropicales o subtropicales. 6 Tierras bajas ecuatoriales.
Agricultura de los EE. UU. publica un mapa con la distribución geográfica de las temperaturas para guiar a los agricultores sobre las zonas de crecimiento vegetal en la nación (Fig. 5.3). Varios ecólogos norteamericanos discreparon de Merriam en el uso de las zonas de vida y dos de ellos, John Weaver y Frederick Clements, desarrollaron un mapa para la vegetación de Norteamérica que en 1932, Víctor Shelford modificó (Fig. 5.4).
El concepto de zonas de vida de Holdridge (1947) es distinto al de Merriam y está diseñado para clasificar todas las condiciones climáticas (no solamente la temperatura) que determinan la distribución de las zonas de vida. El sistema de Holdridge tiene más precisión que el de Merriam y es el sistema de clasificación más utilizado en los trópicos del Nuevo Mundo, aunque se ha aplicado globalmente. Las agencias federales de los EE. UU. utilizan el sistema de Bailey (1987, 1996) para clasificar los ecosistemas y ese sistema se ha aplicado a todo el mundo terrestre y oceánico. El énfasis del trabajo de Bailey es proveer una herramienta útil para la conservación de los ecosistemas. En el Caribe, el sistema de Beard (1944, 1949, 1955) es el más conocido y efectivo. Este sistema está basado en la fisonomía de la vegetación (Tabla 5.1).
En este capítulo vamos a comparar los sistemas de Bailey y Holdridge para ilustrar dos enfoques contrastantes al estudio y conservación de la vegetación. Antes de
5 83 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Figura
América
las zonas de temperatura mínima promedio distribuido por el Departamento de Agricultura como guía de las condiciones de siembra. El mapa es la versión moderna de las zonas de vida utilizadas por el norteamericano Clinton Hart Merriam para estudiar la distribución de los animales.

Tundra
Bosque boreal Bosque caducifolio
Bosque costero
Bosques montano y subalpino Estepa de artemisa
Matorral del desierto Pradera Clímax desconocido
5 84 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales Temperatura (ºC) -45.6 y más baja -42.8 a -45.5 -40.0 a -42.7 -37.3 a -40.0 -34.5 a -37.2 -31.7 a -34.4 -28.9 a -31.6 -26.2 a -28.8 -23.4 a -26.1 -20.6 a -23.3 -17.8 a -20.5 -15.0 a -17.7 -12.3 a -15.0 -9.5 a -12.2 -6.7 a -9.4 -3.9 a -6.6 -1.2 a -3.8 1.6 a -1.1 4.4 a 1.7 4.5 y más alta Temperatura (ºF) más baja de -50 -45 a -50 -40 a-45 -35 a -40 -30 a -35 -25 a -30 -20 a -25 -15 a -20 -l0 a -15 -5 a -10 0 a -5 5 a 0 10 a 5 15 a 10 20 a 15 25 a 20 30 a 25 35 a 30 40 a 35 40 y más alta Zona 1 2a 2b 3a 3b 4a 4b 5a 5b 6a 6b 7a 7b 8a 8b 9a 9b 10a 10b 11 Promedio de la temperatura mínima anual
5.3. Mapa de Estados Unidos de
con
Figura 5.4. Mapa de los biomas de Norteamérica de Shelford (1932) basado en el trabajo de Weaver y Clements. Más tarde, Shelford actualizó este mapa (Shelford 1963).
describir los dos sistemas debemos entender algunos conceptos básicos sobre nuestro planeta, los factores físicos que controlan la vegetación y el contraste entre un enfoque geográfico vs. uno ecológico. Es importante entender la diferencia entre estos dos puntos de vista, pues gran parte de la literatura ecológica refleja la visión geográfica pretendiendo hacerla pasar por la ecológica. Al entender la diferencia entre Bailey y Holdridge, se entiende la diferencia entre un enfoque ecológico y funcional (Holdridge) y uno geográfico (Bailey).
Algunos principios fundamentales
La incidencia de la energía solar sobre el planeta no es homogénea como se aprecia en la figura 5.5. En el ecuador del planeta la energía que llega es más concentrada que en las latitudes más altas, ya que en el ecuador la incidencia es directa, mientras que en altas latitudes la energía solar entra sesgada y tiene que viajar más distancia antes de llegar a la superficie del planeta. Los rayos de luz pierden más calor en su trayectoria a las latitudes altas y están más dispersos sobre la superficie planetaria. Esas diferencias en el insumo de energía solar sobre distintas latitudes controlan el clima de todo el planeta, incluyendo los vientos, las lluvias, la temperatura del aire y las corrientes marinas. Cuando las masas de aire sobre el ecuador se calientan, se expanden y elevan en la atmósfera, crean zonas de baja presión atmosférica. Esas masas de aire caliente se transportan hacia latitudes más altas donde se enfrían, se contraen y descienden nuevamente creando zonas de alta presión
Tabla 5.1
Sistema de clasificación de asociaciones vegetales de Beard (1944, 1955) traducido en Lugo (2005a). El orden de las categorías corresponde a Beard (1955). Los números corresponden a series y el resto de las categorías son formaciones. La duración de hojas en los árboles se clasifica de la siguiente manera: caducifolio, > 66 % de las especies deciduas; semisiempreverde, 33 a 66 % de las especies deciduas; siempreverde, < 33 % de las especies deciduas.
1. Bosque pluvial tropical
2. Formaciones estacionales
Bosque siempreverde estacional
Bosque semisiempreverde estacional
Bosque caducifolio estacional
Arboleda espinosa
Matorral de cactus
Desierto
3. Formaciones montanas
Bosque pluvial tropical montano bajo
Bosque montano pluvial tropical
Palmares en laderas
Arboleda elfina
Arboleda con heladas
Pinares montanos
Bosque de bambú en laderas
Páramo
Tundra
4. Formaciones siempreverdes secas
Bosque pluvial seco
Bosque siempreverde seco
Arboleda siempreverde seca y arboleda del litoral
Matorral (thicket) seco siempreverde y matorral del litoral
Matorral (bushland ) siempreverde y cobertura (hedge) litoral
Vegetación sobre pavimento rocoso
6. Formaciones de ciénagas o pantanos estacionales
Selva en lodazal estacional
Arboleda en lodazal estacional
Matorral pantanoso estacional
Pantano herbáceo (sabana)
5. Formaciones pantanosas
Bosque pantanoso y manglares
Arboleda pantanosa
Matorral pantanoso
Pantano herbáceo
5 85 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Figura 5.5. La luz solar que llega al ecuador es mayor a la que llega a latitudes más altas porque comparada con la que incide en altas latitudes, los rayos del sol llegan directos con más energía concentrada por unidad de área. Además, los rayos del sol que llegan al ecuador atraviesan un volumen menor de la atmósfera, la cual absorbe parte de su energía en proporción al volumen (Modificado de Bailey 2009).
Figura 5.6. La distribución de la energía solar sobre el planeta crea regiones de alta y baja presión atmosférica que regula los vientos que se mueven de lugares con alta presión a lugares de baja presión. En el ecuador, la alta energía solar calienta el aire lo que causa que baje en densidad y se eleve creando zonas de baja presión. Al elevarse, el aire se enfría, aumenta en densidad y tiende a bajar creando zonas de alta presión atmosférica. El ciclo del movimiento del aire entre zonas de alta y baja presión crea las celdas de convección ilustradas en la figura. Las zonas de alta presión son zonas secas y las de baja presión son zonas húmedas. La ubicación de las zonas de alta y baja presión es dinámica geográficamente dependiendo de la rotación e inclinación del planeta respecto al sol (Modificado de Bailey 2009).

atmosférica (Fig. 5.6). Las zonas de alta presión crean climas áridos, mientras que las de baja presión crean climas húmedos (Fig. 5.7).
Tanto el suelo como la biota son el resultado del clima y la topografía. O sea, el efecto del insumo de energía solar al planeta termina controlando el clima, la topografía, los suelos y eventualmente la biota. En la figura 5.8 se ilustra que el límite altitudinal de crecimiento vegetal (plantas vasculares y árboles) ocurre a elevaciones más altas cerca del ecuador que en latitudes altas, donde el crecimiento de plantas vasculares casi desaparece. Las condiciones climáticas e insumo de energía son más favorables cerca del ecuador que en altas latitudes.
Como el insumo de energía varía latitudinalmente, se puede organizar el planeta en zonas de temperatura que afectan el comportamiento de la biota (Fig. 5.9).



5 86 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
las isotermas de -3 °C para el mes promedio más frío en el hemisferio norte y sur, y la de 22 °C para el mes promedio más caliente en el hemisferio norte y sur. Las isotermas se desvían cuando atraviesan los límites entre tierra y océano.

Para los organismos conquistar las zonas frías del planeta necesitan adaptaciones fisiológicas extremas que no son necesarias en las zonas cálidas. Desarrollar y mantener estas adaptaciones tiene un costo energético, lo que ayuda a explicar por qué hay más biodiversidad cerca del ecuador que cerca de los polos.
Anteriormente, habíamos examinado la organización vertical de los ecosistemas desde el punto de vista estructural y funcional (Fig. 2.6). Ese perfil vertical cambia


5 87 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Figura 5.7. Subdivisión de los trópicos basado en la estacionalidad de la lluvia (Bailey 2009). Las líneas sólidas representan
Figura 5.8. Distribución de los límites del crecimiento vegetal y de los árboles con la elevación en distintas latitudes (Modificado de Bailey 2009).
en cuanto a su extensión vertical y funcionamiento, incluyendo su estequiometría, dependiendo del clima en el cual se encuentre (Fig. 5.10). Mientras más húmedo el clima, más desarrollo estructural se observa en el perfil vertical de esos ecosistemas.
Tanto el desarrollo del suelo como el de la vegetación exhiben mayor complejidad con la disponibilidad del agua, ya que la temperatura no varía en ese transecto. La nomenclatura de los suelos y sus características químicas también cambia en este gradiente de humedad. En lugares áridos predominan los suelos con altas concentraciones de carbonatos y altos pH. En contraste, en los climas húmedos predominan los suelos ácidos (bajo pH) con altas concentraciones de óxidos de aluminio y hierro. El estudio de Lugo et al. (2022), resumido en el capítulo 3, encontró que los suelos muy ácidos (pH < 4.0) afectan la estequiometría limitando la disponibilidad del fósforo y calcio, lo que aumenta el costo metabólico del reciclaje y potencialmente afecta la productividad.
Para describir la vegetación y los suelos del planeta, los geógrafos y luego los ecólogos, desarrollaron clasificaciones de los climas del planeta para que, con base en los climas y la topografía, se pudiese derivar los tipos de vegetación y suelos. Uno de los sistemas de clasificación de climas más utilizados es el del ruso-alemán Wladimir Peter Köppen el cual han modificado varios científicos y de acuerdo con Bailey tiene 13 climas básicos para todo el planeta (Tabla 5.2). El geógrafo norteamericano Glen Thomas Trewartha usó y modificó los climas de Köppen para desarrollar un mapa de las zonas ecoclimáticas del planeta (Fig. 5.11). En el mapa de Trewartha las zonas montañosas constituyen un grupo ecoclimático independientemente de la latitud donde se encuentren, lo cual reconoce la complejidad climática, edáfica y biótica de las montañas, pero es silencioso relativo a esa complejidad.
5 88 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Figura 5.9. Mapa global de las zonas termales que delimitan el crecimiento de especies arbóreas de distintas zonas biogeográficas (Modificado de Bailey 2009).
Figura 5.10. Ilustración de la relación entre clima, vegetación y suelos en un transecto desde el suroeste hasta la región de los grandes lagos de los Estados Unidos de América. La ilustración está basada en Bailey 2009. Los distintos horizontes en los suelos están demarcados por letras mayúsculas (A, B, etc.). El carbonato de calcio (CaCO3) ocurre a profundidades menores en las zonas áridas y a profundidades mayores con el aumento en la lluvia y la acidez del suelo.
Tabla 5.2
Trece climas básicos del planeta Tierra basados en el sistema de Köppen modificado por Trewartha y reportado por Bailey (2009).
Símbolo Definición
Límites
Ar Tropical muy húmedo Todos los meses >18 ºC y sin periodo de sequía
Aw Tropical muy húmedo-seco Igual a Ar pero dos meses secos en el invierno
BSh Tropical/subtropical semiárido La evaporación potencial > precipitación y todos los meses >0 oC
BWh Tropical/subtropical árido La mitad de la precipitación en BSh y todos los meses >0 oC
BSk Templado semiárido Igual a BSh pero por lo menos un mes <0 oC
BWk Templado árido Igual a BWh pero por lo menos un mes <0 oC
Cs Subtropical verano seco (Mediterráneo)Ocho meses 10 ºC, el mes más frío bajo 18 ºC y el verano seco
Cf Subtropical húmedo Igual a Cs sin periodo de sequía
Do Templado oceánico Cuatro a siete meses >10 ºC, el mes más frío > 0 oC
Dc Templado continental Igual a Do pero el mes más frío < 0 oC
E Boreal o subártico Hasta tres meses sobre 10 ºC
Ft Tundra Todos los meses bajo 10 ºC
Fi Capa de hielo polar Todos los meses bajo 0 oC
5 89 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Para cada tipo de clima en el sistema de Köppen se identifican formaciones vegetativas como las siguientes: Clima A Tropical lluvioso- Bosque Pluvial Tropical; Clima B o secos- Desierto Tropical; Clima C templado cálido- Bosque Templado Deciduo; Clima D con bosques bajo efectos de nevadas- Bosque Boreal, y Clima E opolar- la Tundra. Además, se reconoce en cada tipo de clima la presencia del suelo y la vegetación zonal (Tabla 5.3) que representan el máximo desarrollo de estos componentes para cada tipo de clima. Sin embargo, tanto la vegetación como el suelo zonal no representan los únicos componentes en un clima dado. Hay otros tipos de vegetación y suelos en cada clima dependiendo de las variaciones climáticas o edáficas, los disturbios y la actividad humana.
De los patrones globales de clima, vegetación y suelo, surgieron patrones de productividad (Tabla 5.4) y descomposición (Tabla 5.5) de la materia orgánica. Es evidente que las tasas más altas de estos procesos están asociadas a la humedad y alta temperatura. El contraste en la velocidad de procesos entre las zonas tropicales y las polares, por ejemplo, es notable.
Los climadiagramas de Walter (1971) representan una forma muy efectiva para describir el clima de una localidad a la vez que se comunica importante información

5 90 CAP ariel e. lugo | ecología a mi ma nera Los bosques tropicales
BSk
BWk
Cs
Cf
Do
Dc
E Boreal Ft Tundra
Fi
H Tierras
Ar Tropical muy húmedo
Aw
Tropical muy húmedo – seco
BSh
Tropical/subtropical semiárido
BWh
Tropical/subtropical árido
Templado semiárido
Templado árido
Subtropical verano seco
Subtropical húmedo
Templado oceánico
Templado continental
polar
Capa de hielo polar
altas
Figura 5. 11. Las zonas eco climáticas de Trewartha según la interpretación de Bailey 2009. Todas las zonas montañosas se clasifican como tierras altas.
Tabla 5.3
Relación entre el ecoclima y la vegetación y los suelos zonales de acuerdo con el trabajo de Bailey (2009). Los nombres en paréntesis corresponden a los órdenes de suelo del Departamento de Agricultura de los Estados Unidos de América. Las subzonas ecoclimáticas corresponden a los climas de Köppen modificado por Trewartha en la tabla 5.2. En Fi no hay vegetación zonal.
Subzona ecoclimática Suelo zonal Vegetación zonal
Ar Latisoles (Oxisoles) Pluvial siempreverde tropical (selva)
Aw Latisoles (Oxisoles) Bosque deciduo o sabana tropical
BS Chestnut, suelos marrones y serozem (Mollisoles, Aridisoles) Hierbas cortas
BW Desierto (Aridisoles) Arbustos o hierbas dispersas
Cs Tierras marrones mediterráneas Arbolada esclerófila
Cf Suelos rojos y amarillos podzólicos (Ultisoles) Bosques de coníferas o bosque mixto de coníferas y deciduas
Do Bosque marrón y gris-marrón podzólico (Alfisoles) Bosque de coníferas
Dc Gris-marrón podzólico (Alfisoles) Bosques de coníferas o bosque mixto de coníferas y deciduas
E Podzólicos (Spodosoles e Histosoles asociados) Bosque boreal de coníferas (taiga)
Ft Tundra humus con solifluxión (Entisoles, Inceptisoles e Histosoles asociados) Vegetación de tundra (sin árboles)
Fi
Tabla 5.4
Distribución de la productividad primaria anual del planeta Tierra basado en Bailey (1996).
Tasas de descomposición de hojas anchas y agujas de pino en varios climas o ecozonas. La tasa de descomposición (k) se calculó dividiendo la producción anual de hojarasca entre la acumulación de hojarasca. El tiempo requerido para descomponer el 95 % del material se obtuvo dividiendo tres entre k. Basado en Bailey (2009).
5 91 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Zona climática Productividad promedio (Mg/ha·año) Productividad total (Mg/año) X 109 Polar 1.6 1.33 Boreal 6.5 15.2 Templada Húmeda 12.6 9.34 Semiárida 8.2 6.44 Árida 2.8 1.99 Subtropical Húmeda 25.5 15.9 Semiárida 13.8 11.5 Árida 7.3 7.14 Tropical Húmeda 29.2 77.3 Semiárida 14.1 22.6 Árida 2.0 2.62
Tabla 5.5
Ecozona Descomposición anual Tiempo (años) requerido para descomponer el 95 % del materiales Polar/subpolar; tundra 0.03 100 Boreal 0.21 14 Latitudes templadas medias 0.77 4 Latitudes medias secas (estepas) 1.5 2 Trópicos estacionales 3.2 1 Trópicos húmedos 6.0 0.5
Figura 5.12. Ejemplos de los diagramas del clima de Walter (1971). Se presentan cuatro tipos de clima contrastantes: el clima muy húmedo en las tierras bajas tropicales de Douala en Camerún; el templado húmedo de Hohenheim, Alemania; el clima montano, frío y seco con un periodo de sequía fuerte de Ankara, Turquía; y el clima seco y costero de Odessa en el suroeste de Ucrania en la costa norte del mar Negro. La explicación del diagrama y la información que contiene se obtiene del significado de las letras en itálicas en distintas partes del diagrama: a, nombre de la estación de donde provienen los datos; b, elevación sobre el nivel del mar; c, número de años de datos (temperatura primero y luego la precipitación); d, temperatura anual promedio (oC); e, precipitación anual promedio en mm; f, temperatura diaria mínima promedio durante el mes más frio; g, temperatura mínima más baja en el récord; h, temperatura diaria máxima promedio durante el mes más caluroso; i, temperatura más alta en el récord; j, rango de temperatura diaria promedio; k, curva de la temperatura mensual promedio en incrementos de 10 oC en la escala; l, curva de la precipitación mensual promedio con la escala de 10 oC = 20 mm; m, periodo de sequía (punteada); n, periodo húmedo (patrón de escotilla vertical); o, precipitación mensual promedio en exceso a 200 mm (se reduce la escala a 1 oC = 10 mm), área negra; p, curva de precipitación ajustada a 10 oC = 30 mm (patrón de escotilla horizontal superior refleja periodo seco); q, meses con temperatura diaria promedio menor a 0 oC (negro); r, meses con el mínimo absoluto menor a 0 oC (escotilla diagonal); s, duración promedio de temperaturas diarias sobre 0 oC (sin heladas) en días y en itálicas.
con relevancia ecofisiológica (Fig. 5.12). Además de información geográfica del lugar y su elevación, las escalas de lluvia y temperatura (10 o C = 20 mm de lluvia) permiten evaluar periodos de sequía, humedad y exceso de agua.
El sistema de ecorregiones de Bailey
Bailey basó su sistema de ecorregiones en el trabajo de Köppen, según modificado por Trewartha (Tabla 5.2). Como nos interesan los trópicos, es útil entender cómo se define esta región por este sistema de clasificación.
En el clima tropical de Köppen los 12 meses promedian 18 °C/64.4 oF o más). Los climas «A» son climas sin invierno y libres de heladas. Trewartha modificó el sistema de Köppen para incluir estaciones secas en los trópicos. Climas con no más de dos meses secos (menos de 60 mm en promedio) se clasifican Ar y Aw, si la estación seca ocurre durante periodos de días cortos debido al bajo ángulo del sol. Se clasifica As, si la época seca ocurre cuando los días son largos por lo alto del sol.
5 92 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Climas con monzón caracterizados por estaciones secas de más de tres meses son Am. Un clima monzónico se caracteriza por cambios en la dirección prevalente de los vientos, de manera que causan sequías o épocas de lluvia, dependiendo de la dirección del viento.
En el sistema de Bailey, el tamaño del área geográfica es determinante al identificar sus categorías jerárquicas de ecosistemas (Fig. 5.13). Su sistema identifica sitios, paisajes y ecorregiones. Cuando el Servicio Forestal de los EE. UU. decidió desarrollar mapas de las ecorregiones de Bailey, introdujo subdivisiones geográficas para incrementar su resolución (Fig. 5.14). Por ejemplo, las ecorregiones se subdividieron en Dominios, Divisiones y Provincias, que para los trópicos incluyen dos Dominios (seco y húmedo) y ocho Divisiones y regímenes montañosos (cuatro en cada Dominio).
5 93 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Figura 5.13. Jerarquía geográfica utilizada por Bailey en su sistema de clasificación de eco regiones. Contrario a la jerarquía funcional en la figura 2.3, esta jerarquía está basada en el área de la región bajo estudio (modificado de Bailey 2009).
Figura 5.14. Comparación entre las jerarquías utilizadas por el Servicio Forestal de los Estados Unidos de Norteamérica (Ecomapa) y las propuestas por Bailey. El Servicio Forestal aumenta la resolución de las categorías, lo que baja la variabilidad de los datos coleccionados para caracterizar sistemas ecológicos (Modificado de Bailey 2009).
¿Cuáles son las limitaciones ecológicas de clasificaciones basadas en geografía?
La limitación principal es que las unidades ecológicas están predeterminadas por su área geográfica y, por lo tanto, no constituyen unidades ecológicas que puedan atribuirse a una jerarquía funcional como la presentada en la figura 2.3 del capítulo 2 sobre ecosistemas. Es difícil relacionar una unidad geográfica a una ecológica porque se pierde mucha resolución ecológica al agrupar distintos tipos de ecosistemas bajo una sola categoría geográfica. Más aún, la determinación de los sistemas geográficos no es reproducible cuando distintas personas utilizan la clasificación, ya que cada persona que aplique los criterios de clasificación llegará a una solución distinta. Los criterios de clasificación no son objetivos ni empíricos. El sistema de zonas de vida de Holdridge trata de resolver asuntos funcionales ignorados por otros sistemas, resultando en una clasificación eminentemente ecológica y con un enfoque en el funcionamiento de ecosistemas.
Sistema de zonas de vida de Holdridge
El sistema de zonas de vida de Holdridge se basa en data empírica y utiliza reglas objetivas para asegurar que un lugar se clasifique consistentemente, no importa quién aplica las reglas de clasificación. El sistema está limitado por la disponibilidad de datos. El sistema de Holdridge también es jerárquico (Fig. 5.15). Incluye regiones latitudinales, bandas altitudinales, provincias de humedad, zonas de vida, asociaciones y etapas de sucesión. En el sistema de Holdridge se delimitan zonas de vida en las montañas en vez de agregar las montañas como una unidad de clasificación. Como Holdridge trabajó en Puerto Rico y el Caribe, aprovechó su experiencia con las montañas caribeñas para desarrollar sus ideas de zonas de vida, que también se basan en las ideas de Merriam, aunque con más fundamentos climáticos y fisiológicos.
La asociación (de plantas, animales, sustrato geológico y suelos) es la unidad de estudio. Las asociaciones en cada zona de vida son únicas para esa zona de vida. La fisonomía y no las especies, definen la asociación. Una zona de vida se delimita por su clima, pero representa un grupo de asociaciones relacionadas por el efecto del calor, la humedad y la precipitación. Tales efectos sobre la asociación ocurren en todas las asociaciones (no-antropogénicas, maduras, la actividad animal, el crecimiento secundario y las actividades del ser humano).
Los factores universales que definen las asociaciones y zonas de vida son el calor, la precipitación y la humedad. Se descartan criterios taxonómicos, edáficos, disturbios, geológicos, topográficos y muchos factores climáticos. Aunque sean importantes y relevantes a la clasificación, el problema es que no son prácticos ni lo suficientemente universales.
Las bases del sistema de Holdridge incluyen el calor que se representa por el concepto de biotemperatura y se utilizan escalas logarítmicas para reflejar el efecto
5 94 CAP ariel e. lugo | ecología a mi ma nera Los bosques tropicales
la región latitudinal están calculados al nivel del mar utilizando una tasa de lapso promedio de 6 oC por 1,000 m. La línea entrecortada originándose en la biotemperatura aproximada de 18 oC, delimita los sistemas tropicales y subtropicales en las tierras bajas y premontanas.
del calor y la precipitación sobre la vegetación. La biotemperatura es una medida del calor que es efectivo sobre el crecimiento de la vegetación. Se determina asignándole valor de cero a las temperaturas diarias o medidas cada hora que den valores menores a 0 o C o mayores a 30 o C. Las escalas logarítmicas están basadas en el concepto del factor limitante, donde para adquirir un incremento en la productividad igual hay que aumentar la cantidad que limita al proceso en una progresión logarítmica.
El diagrama de zonas de vida de Holdridge (Fig. 5.16) describe 112 zonas, de las cuales 52 son boscosas (Tabla 5.6). Noten que en los trópicos, Holdridge reconoce más zonas de vida que en el resto del mundo combinado. Para designar una zona de vida es necesario saber la biotemperatura promedio anual, la lluvia promedio anual, la elevación y latitud del lugar. Si el lugar está en las montañas, es necesario saber la tasa de lapso adiabático (lapse rate o gradiente vertical de temperatura que indica el cambio en la temperatura con la elevación). Tal información permite calcular la biotemperatura al nivel del mar correspondiente a la biotemperatura en la montaña. La biotemperatura al nivel del mar determina la región latitudinal del sitio y si es tropical o subtropical en el caso de Puerto Rico.
5 95 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Figura 5.15. Regiones latitudinales y pisos altitudinales definidas por la biotemperatura promedio anual en el sistema de zonas de vida del mundo de Holdridge (1967). La biotemperatura, calculada con base en temperaturas diarias donde valores sobre 30 oC y bajo 0 oC se consideran como cero en el estimado del promedio. Los valores en esta ilustración que determinan
La nomenclatura de las zonas de vida es de acuerdo con Ewel y Madriz (1968). La región latitudinal «templada subtropical» la deno -
La extensión en área de las zonas de vida no es rígida. Varían espacial y temporalmente. Un ejemplo son los manglares de Jobos, en el sur de Puerto Rico. Están ubicados entre dos zonas de vida cuyas condiciones se mueven temporal y espacialmente por ciclos en décadas. A veces dan la impresión de ser manglares áridos y en otras ocasiones aparentan ser manglares de zonas húmedas. Eso se debe a que el clima oscila entre uno con más evapotranspiración potencial (PET) que precipitación (P; PET/P > 1) y uno con menos evapotranspiración potencial que precipitación (PET/P < 1).
5 96 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Figura 5.16. Diagrama utilizado para clasificar las zonas de vida o formaciones de plantas del mundo de acuerdo con el sistema de zonas de vida del mundo de Holdridge (1967). La línea entrecortada identificada como «línea de escarcha o línea de temperatura crítica» separa los trópicos y subtrópicos de las otras regiones latitudinales.
minó Holdridge (1967) como « templada cálida» que es la traducción preferible.
Categorías Número de zonas de vida Tropicales Templada Boreal Total Seca 33 16 3 52 Húmeda 11 7 2 20 Muy húmeda 22 14 4 40 Total 66 37 9 112 Bosques 33 16 3 52
Tabla 5.6 Número de zonas de vida en el sistema de Holdridge (Lugo y Brown 1991).
En el sistema de Holdridge hay cuatro tipos de asociaciones: climáticas (consideradas zonales si están sobre el suelo zonal), edáficas, atmosféricas e hídricas (humedales). La condensación de agua sobre la vegetación se considera en las asociaciones atmosféricas. Una misma especie puede predominar en más de una zona de vida, pero su fisonomía cambia. La fisonomía es crítica en la identificación de una asociación.
Holdridge propone dos herramientas ecológicas para estudiar las asociaciones. Una es el Índice de Complejidad (CI) y la otra, el perfil idealizado de la vegetación. El perfil idealizado (Fig. 5.17) permite visualizar la complejidad vertical
5 97 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Figura 5.17. Ejemplos de dos perfiles idealizados de dos bosques maduros en distintas zonas de vida (Holdridge 1967). El perfil del tope es una asociación climática en la zona de vida bosque tropical premontano pluvial en valle Escondido, Costa Rica. El perfil de abajo es una asociación monzón en la zona de vida bosque tropical húmedo, Potrero Grande, Costa Rica. Noten la persona en la parte inferior izquierda de cada perfil. En el original se identifican las especies en el diagrama.
de la asociación. La complejidad se calcula empíricamente de acuerdo con la siguiente ecuación:
CI = 10-3 (altura en m)*(área basal en m2)*(densidad de árboles)*(número de especies)
Para un área de muestreo de 10 m x 100 m y árboles con un diámetro a la altura del pecho (dap) ≥10 cm. La figura 5.18 ilustra que el CI aumenta hacia los trópicos superhúmedos de tierras bajas.
Al igual que Herbertson, Holdridge propuso que las condiciones climáticas de las zonas de vida indican la importancia del clima para la actividad humana. La actividad humana se beneficia de climas en las zonas de vida húmeda y secas en lugares donde PET/P no se aleja mucho de 1. Hacia los extremos de humedad PET/P << 1 y aridez (PET/P >> 1) el costo de supervivencia aumenta y la actividad humana disminuye o se imposibilita sin subsidios energéticos. Además del clima, por supuesto, otros factores a considerar al evaluar la actividad humana son la disponibilidad de suelos fértiles y la topografía favorable. Con el gasto de energía fósil, el ser humano puede sobreponerse a las limitantes climáticas, edáficas y topográficas del planeta.
5 98 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Figura 5.18. El Índice de Complejidad de Holdridge para bosques en Costa Rica de acuerdo con la zona de vida donde se obtuvo el resultado. Solo se consideraron bosques maduros sin fuertes restricciones de crecimiento. Todos los valores corresponden a la latitud tropical y disminuyen con la elevación y la disponibilidad del agua con el valor más alto en la zona de vida bosque tropical pluvial en tierras bajas.
Bosque pluvial boreal
Zonas de vida de Holdridge
Tundra muy húmeda alpino templada fría
Tundra pluvial alpino templada fría
Maleza seca subalpino templada fría
Bosque húmedo subalpino templada frío
Bosque muy húmedo subalpino templado frío
Bosque pluvial subalpino templado frío
Desierto templado frío
Maleza desértica templada fría
Estepa templada fría
Bosque húmedo templado frío
Bosque muy húmedo templado frío
Bosque pluvial templado frío
Nival templada cálida
Tundra muy húmeda alpino templada cálida
Tundra muy húmeda templada cálida

Tundra pluvial alpina templada cálida
Maleza desértica subalpina templada cálida
Bosque húmedo subalpino templado cálido
Bosque muy húmedo subalpino templado cálido
Bosque pluvial subalpino templado cálido
Desierto montano templado cálido
Maleza desértica montana templada cálida
Estepa montana templada cálida
Bosque húmedo montano templado cálido
Bosque muy húmedo montano templado cálido
Bosque pluvial montano templado cálido
Desierto templado cálido
Maleza desértica templada cálida
Estepa espinosa templada cálida
Bosque seco templado cálido
Bosque húmedo templado cálido
Bosque muy húmedo templado cálido
Maleza desértica subtropical
Monte espinoso subtropical
Bosque seco subtropical
Bosque húmedo subtropical
Bosque seco tropical
Comparación entre el sistema de Holdridge y el de Bailey
Con el mapa de las zonas de vida de Holdridge para los EE. UU. (Fig. 5.19) es posible representar el país por sus zonas altitudinales, latitudinales y de humedad (Fig. 5.20) y comparar los mapas con los de Bailey y otros mapas similares. Latitudinalmente, EE. UU. es un país bastante homogéneo, pero en términos de zonas altitudinales tiene mucha diversidad, particularmente en el oeste. En términos de humedad, la porción oriental es homogénea y la región occidental exhibe múltiples gradientes que influencian la diversidad ecológica en esa región del país.
En la figura 5.21 se comparan cinco mapas para los EE. UU., agregando las unidades de acuerdo con categorías similares. Se puede apreciar el nivel de detalle que emerge de los distintos esquemas de clasificación. El esquema de Loveland es una clasificación basada en una imagen de satélite y por eso la categoría agrícola resalta.
5 99 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Figura 5.19. Las zonas de vida de los Estados Unidos de América excluyendo a Alaska y Hawái (Lugo et al. 1999b). La zona montañosa del oeste contiene la mayor complejidad ecológica y diversidad de zonas de vida comparada con el este de la nación.
0 500 1000 km
Zonas latitudinales (celdas)
Boreal (9)
Templado frío (172,378)
Templado (311,847)
Subtropical (648)
Tropical (288)
Provincias de humedad (celdas)
Superhúmedo (17,278)
Perhúmedo (42,705)
Húmedo (219,080)
Subhúmedo (139,615)
Semiárido (51,828)
Árido (11,008)
Perárido (3,841)
Superárido (153)
Zonas altitudinales (celdas)
Nival (51)
Alpina (5,221)
Subalpina (36,401)
Montano (85,925)

Finalmente, la figura 5.22 compara las zonas de vida de Holdridge con las ecorregiones de Bailey de acuerdo con su rango de cobertura espacial. Queda muy claro que el sistema de Holdridge reconoce más unidades que el de Bailey. Las diferencias que se ven distinguen claramente la visión geográfica vs. la ecológica. De acuerdo con Holdridge, hay más unidades ecológicas en los EE. UU. que las que reconoce el sistema de ecorregiones.
La contestación a la pregunta al inicio de este capítulo es que un bosque tropical es uno que no está sujeto a heladas y el número de tipos de bosques tropicales es variable, pues depende del nivel de resolución ecológica que se utilice para numerarlos.
Asignación
Selecciona cualquier atributo estructural o funcional de un bosque tropical y compáralo a lo largo de un gradiente de humedad o de temperatura.
5 100 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Figura 5.20. El clima de los Estados Unidos de América excluyendo a Alaska y Hawái segregado por las zonas latitudinales y altitudinales y las provincias de humedad de acuerdo con el sistema de zonas de vida de Holdridge (Lugo et al. 1999b). Los números en paréntesis corresponden al número de celdas incluidas.
Zonas de vida de Holdridge
Ecorregiones de Bailey
Vegetación potencial de Kuchler
Zona de vida
Ecorregiones
Figura 5.21. Comparación de cinco sistemas de clasificación de vegetación o cobertura utilizando la misma nomenclatura para las coberturas (Lugo et al. 1999b). La cobertura de la tierra de Loveland se determinó con un radiómetro avanzado de muy alta resolución.
Figura 5.22. Comparación del sistema de Bailey con el de Holdridge en términos de los rangos basados en cobertura aérea de las unidades de clasificación: provincias ecológicas de Bailey y zonas de vida de Holdridge (Lugo et al. 1999b). El sistema de Holdridge provee más resolución ecológica que el sistema de Bailey porque delimita las unidades por el clima, en vez de utilizar criterios geográficos preseleccionados por el observador.

5 101 CAP ariel e. lugo | ecología a m i m anera Los bosques tropicales
Pastizal Arbustal
Bosque
Sin vegetación Cultivos
Bioma
Cobertura de tierra de Loveland
CaPítulo 6 Los bosques de Puerto Rico
Pregunta
¿Cuántos tipos de bosque hay en Puerto Rico?
El estudio y conservación de los bosques de Puerto Rico bajo el Imperio Español
El cialeño Carlos Manuel Domínguez Cristóbal (2000) escribió la obra más completa sobre la historia forestal de Puerto Rico y con base en ese trabajo resumo a continuación algunos temas relacionados con el estudio y conservación de los bosques durante la época española. De allí se desprende la importancia de la flora y fauna y los bosques de Puerto Rico durante todo el periodo colonial español de la isla hasta la llegada del gobierno de EE. UU. Cuando los españoles llegaron, Puerto Rico estaba 100 % cubierto de bosques con una gran riqueza natural. Desde el principio, árboles como el guayacán (Guaiacum officinale), tabonuco (Dacryodes excelsa), cedro (Cedrela odorata), uva de playa (Coccoloba uvifera), úcar (Bucida buceras), la ceiba (Ceiba pentandra) y muchos otros eran reconocidos por sus valores madereros o por sus otros productos no maderables como las medicinas o la resina del tabonuco.
La deforestación de los bosques comenzó durante el siglo XVII para darle paso a la agricultura y pastizales. Las causas de la deforestación en Puerto Rico fueron las mismas que se observaron en otros países tropicales: la necesidad de tierras agrícolas y combustible (leña y carbón derivados de la madera de los árboles) y el desarrollo de infraestructura. La deforestación comenzó en las zonas llanas costeras y poco a poco fue extendiéndose a laderas y acelerándose durante el siglo XVIII, y llegó a su máxima intensidad durante el siglo XIX. La aceleración coincidió con el aumento poblacional y una expansión agrícola.
En 1797 el francés André Pierre Ledru (1797) visitó y vivió un año en Puerto Rico. Encontró manglares, helechos gigantes, plantas trepadoras y bosques húmedos en el sector de San Patricio de San Juan, un lugar que le tomó días llegar (cruzando la Bahía de San Juan) y explorar por vía del cauce del río Piedras. Ledru le atribuyó la
ariel e. lugo | ecología a m i m anera
103
fecundidad de las tierras de Puerto Rico a las lluvias que inundan la isla durante el invierno, el crecido número de ríos con agua todo el año, el abundante rocío de la noche y los despojos de los árboles que bajo la influencia del clima se descomponen y forman tierra vegetal. Lo que se desprende de los escritos sobre Puerto Rico hasta finales del siglo XIX es el verdor del paisaje tropical, la exuberancia de la vegetación y la abundancia de especies en los bosques de la isla. Los bosques se describen como densos y extensos en lugares remotos en relación con los poblados de donde procedían los escritores de esos tiempos.
La Circular 493 de 1824 fue el primer documento oficial en el desarrollo de la política forestal española en Puerto Rico. La Circular estableció zonas de protección contra la deforestación en las cabeceras y cauces de ríos y quebradas y contenía medidas para asegurar el cumplimiento de las disposiciones de protección de las aguas, los suelos y los árboles.
En 1867 se publicó el primer inventario de los bosques de Puerto Rico, municipio por municipio, basado en los reportes de cada uno de ellos. Se encontró una cobertura forestal de 106,864 ha (12 % de la isla), de las cuales 18 % eran bosques públicos. En las páginas 259-260 de Domínguez Cristóbal (2000) se reportan los resultados por municipio. Utuado, Salinas y Mayagüez fueron los municipios con mayor cobertura forestal. En 1870 se llevó a cabo un inventario de los montes (bosques) públicos (18,682 ha). El inventario reportó la distribución de las especies forestales más prominentes y ayuda a establecer la distribución de especies arbóreas hace más de cien años. El inventario no incluyó manglares porque estaban bajo la jurisdicción de la Marina. En 1899, el Departamento de Guerra de los EE. UU. reportó 316,016 cuerdas (124,207 ha) de bosques (46 % bosques altos y 54 % bosques bajos) en Puerto Rico. Además, reportaron 477,987 cuerdas (187,868 ha) cultivadas.
A mediados de la década de 1870, el gobierno español estableció la Inspección de Montes de Puerto Rico hasta 1889, cuando fue suprimida, esta organización tuvo logros significativos en la conservación de los bosques de Puerto Rico. En el año 1876 el rey Alfonso XII emitió una ordenanza para establecer la política forestal del gobierno español en Puerto Rico, Cuba y otras colonias en el Caribe. Se definió un monte como todos los terrenos destinados particularmente a la producción de madera y leña y las tierras de pastos no cultivados. La ordenanza incluyó la designación de 10,000 cuerdas (3,930 ha) de El Yunque como una reserva pública, convirtiéndose en una de las primeras áreas protegidas en el Caribe. Los ingleses habían designado reservas forestales en sus colonias caribeñas antes de la acción de los españoles.
La Inspección de Montes mantuvo récord de la utilización de los bosques, estableció colindancias y reguló la explotación y mercado de la madera en la isla. Debe aclararse que la proporción de los bosques bajo manejo profesional era muy
6 104 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
baja comparada con la explotación y contrabando de madera fuera del control de la Inspección de Montes. El manejo de los bosques para productos madereros dio como resultado las primeras estimaciones del volumen de madera disponible en los bosques, al igual que un récord del nivel de las cosechas. Para 1887, el montero Lope Marinero Cardenal reportó la disponibilidad de madera por especie en dos bosques públicos (Monte Santa de Cayey y Monte El Roble de Aibonito), especificando por especie la longitud, ancho y cantidad de madera disponible.
A finales del siglo XIX, antes de la llegada de los norteamericanos, conocidos naturalistas e historiadores visitaron Puerto Rico y escribieron sobre su flora y fauna. Además, varios puertorriqueños, incluyendo a Román Baldorioty de Castro, catedrático de botánica en el Seminario Conciliar de San Juan y Agustín Stahl, también estudiaron la biota de la isla. Estos estudiosos entraron a los bosques para coleccionar plantas y animales, pero su atención estaba centrada en la taxonomía, y las propiedades y usos de las plantas y animales; no en el estudio de los bosques en sí.
El estudio y conservación de los bosques de Puerto Rico bajo el Imperio Norteamericano
La descripción sistemática de los bosques de Puerto Rico comenzó a finales del siglo XIX y principios del siglo XX y continúa en el presente con mucho vigor. Inicialmente, un geólogo y dos dasónomos visitaron la isla para estudiar y reportar a las oficinas del Departamento de Agricultura en Washington DC la condición de sus bosques. En 1913, el director del Jardín Botánico de Nueva York, el norteamericano Nathaniel Lord Britton, organizó y llevó a cabo una expedición científica sin precedentes en el Neotrópico para estudiar la historia natural de Puerto Rico y las Islas Vírgenes. Ese amplio estudio publicado en 19 volúmenes, culminó en la década de 1940 después de la muerte del Dr. Britton en 1934. Los gobiernos de Puerto Rico e Islas Vírgenes ayudaron a financiar el proyecto (Lugo 2021). Algunos aspectos de este trabajo se revisaron en el 80.º aniversario de su comienzo (Figueroa Colón 1996).
Entre la década de 1940 y el presente, el estudio de los bosques consistió en:
(1) estudios dasonómicos del Servicio Forestal de los EE. UU. (de ahora en adelante Servicio Forestal) en los bosques de toda la isla, pero especialmente los de Luquillo, (2) estudios ecológicos en la Sierra de Luquillo por la Universidad de Puerto Rico, otras universidades y el Servicio Forestal, (3) estudios ecológicos en el bosque seco de Guánica por el Departamento de Recursos Naturales (hoy conocido como Departamento de Recursos Naturales y Ambientales y de aquí en adelante DRNA), el Servicio Forestal y universidades locales y del extranjero, y (4) estudios de manglares y otros bosques dispersos por toda la isla conducidos por una gran variedad de organizaciones científicas locales y del extranjero, incluyendo las ya mencionadas y otras más.
6 105 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Tabla 6.1
Algunas expediciones y programas de investigación ecológica sobre los bosques de Puerto Rico. Programas activos se identifican con un asterisco (*) y el periodo de duración está basado en el 2020.
Año Científico Propósito Auspiciador Duración1
1899 R. T. Hill Evaluar las condiciones de los bosques Servicio Forestal de los Estados Unidos1 mes
1905 J. C. Gifford Formular un plan de manejo para El Yunque Servicio Forestal de los Estados Unidos ¿?
1912 L. Murphy Evaluar la situación forestal de Puerto Rico Servicio Forestal de los Estados Unidos 6 meses
1926 H. A. Gleason y M. T. CookEcología de Puerto RicoAcademia de Ciencias de Nueva York 6 meses
1939 Varios Dasonomía y ecología tropicalServicio Forestal de los Estados Unidos 75 años*
1962 H.T. Odum Efectos de radiación ionizante sobre el bosque Comisión de Energía Atómica 6 años
1965 R. Howard Estudio del bosque elfino Fundación Nacional de Ciencias 4 años
1972 Varios Estudio de bosques secos, manglares y otros recursos naturales Departamento de Recursos Naturales y Ambientales 43 años*
1982 Varios Inventarios periódicos de los bosquesServicio Forestal de los Estados Unidos 38 años*
1988 F. N. Scatena Estudios de ecohidrología en El Yunque Servicio Forestal de los Estados Unidos 27 años*
1988 Varios Estudios a largo plazo en El Yunque Fundación Nacional de Ciencias 27 años*
1994 Varios Estudios de los sistemas del karso Ciudadanos del Karso y colaboradores 21 años 1Hasta el 2020.
A continuación describo brevemente la naturaleza de las investigaciones ecológicas en Puerto Rico de acuerdo con la agrupación de estudios en la tabla 6.1 y termino el capítulo con algunas generalizaciones que aplican a todos los bosques de Puerto Rico, al igual que algunas particularidades que se manifiestan en algunos pero no todos los bosques de la isla. Utilizo como fuente de información varios resúmenes que he publicado anteriormente (Lugo 1983, 1996, 2005a). En los próximos capítulos profundizaré sobre las peculiaridades de los distintos tipos de bosque de Puerto Rico.
El geólogo norteamericano Robert Thomas Hill (1899) sugirió la importancia de los suelos para explicar las variaciones que observó en los bosques de Puerto Rico. También observó discontinuidades en el número de especies presentes entre los suelos arcillosos y los calcáreos. Reportó, además, que los bosques costeros habían desaparecido para darle paso a la agricultura. El norteamericano John Clayton Gifford (1905) reportó altas tasas de erosión en Puerto Rico y notó la capa de raíces en los bosques de El Yunque, las cuales hacían difícil caminar por el bosque. Gifford no le atribuyó valor alguno al bosque de palma de sierra y de hecho sugirió exterminar el palmar usando lechones salvajes (él favorecía la introducción de animales al bosque para fomentar la caza). Sin embargo, reconoció el valor de El Yunque desde el punto de vista escénico. Fue el primer dasónomo desde la época española en reportar medidas cuantitativas de la estructura de los bosques de El Yunque.
6 106 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Figura 6.1. Análisis de la situación forestal de Puerto Rico a principios del siglo XX (Murphy 1916). El tamaño del círculo representa la disponi bilidad de madera en Puerto Rico para el uso, mayormente combustible (carbón). El círculo en el centro representa la productividad de madera en un año, por lo que tomaría más de 40 años reemplazar la cantidad disponible al momento del estudio. Las porciones triangulares representan solamente el uso doméstico de la madera en un año. Por lo tanto, tomaría 12 años en agotar toda la madera disponible al principio del estudio y la isla se quedaría sin madera para producir carbón para cocinar.
El norteamericano Louis Sutliffe Murphy (1916) fue el primero que clasificó los bosques de Puerto Rico usando las siguientes formaciones arbóreas: arboladas del litoral (manglares y arboladas mareales secas), bosques húmedos deciduos, bosques tropicales pluviales, bosques secos deciduos, crecimiento en pastizales, bosques culturales (los bosques más conspicuos en la isla: palmares de coco, cafetales, plantaciones de cacao). Solo el 2 % de la cobertura de Puerto Rico tenía bosques libres de disturbios. Murphy hizo dos observaciones muy sagaces sobre Puerto Rico. Primero, observó que el centro geográfico de la isla coincidía con el centro de la distribución poblacional, o sea, la distribución geográfica de la población en la isla era proporcional con poca aglomeración en ciudades. Segundo, presentó un cálculo sobre la sustentabilidad del uso de la madera en el país. Demostró que al paso que iba la isla, en 12 años consumiría toda la producción maderera de sus bosques (Fig. 6.1). Como en 1909, Puerto Rico consumía más energía solar que energía fósil (Tabla 6.2), la observación de Murphy indicaba que el ritmo de la actividad humana en Puerto Rico era insostenible y requería alguna acción inmediata. Murphy recomendó la creación de un servicio forestal insular para conservar los bosques, sugerencia que avaló la legislatura de Puerto Rico al establecer el Servicio Forestal de Puerto Rico en 1918.
Consumo de energía solar y fósil en Puerto Rico en 1909. (Detalles en Lugo 1983). El cálculo no incluye la agricultura, lo cual multiplicaría aún más el uso de la energía solar.
6 107 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Tabla 6.2
Fuente energética Equivalentes de calor Equivalentes fósiles kcal x 109 Consumo fósil 287 287 Consumo solar 1,818 909 Total de consumo 19,105 1,196 Solar/Fósil 6.3 3.2
Tabla 6.3
La vegetación de Puerto Rico de acuerdo con Gleason y Cook (1926).
Planicie costera del norte (20 asociaciones) La región montañosa central Mésico Bosque montano de baja elevación
Bosque mesofítico
Matorral de crecimiento secundario
Arcilla margosa y roja en las planicies
Bosque en tierras bajas sobre arenas blancas
Bosque pluvial
Bosque de palma de sierra
Bosque musgoso
Matorrales secundarios sobre arenas blancas Región costanera del sur y cerros adyacentes (17 asociaciones)
Árido
Vegetación playera
Matorral playero
Dunas de Coccoloba uvifera
Matorrales de sotavento
Bosque sobre dunas
Vegetación secundaria sobre dunas
Salino
Asociación de manglares
Asociación de Batis - Sesuvium
Salinas
Bosque clímax
Hídrico
Lagunas
Hídrico Pantanos herbáceos
Ríos y valles inundables Árido
Lagunas de agua dulce
Vegetación en lodazales
Vegetación sobre arena
Matorrales playeros
Salino
Asociación de manglares
Asociación de Acrostichum
Asociación de Pterocarpus
Vegetación clímax
Playas y matorrales costeros
Bosque xerofítico
Vegetación de montes de esquisto
Vegetación sobre suelos ultramáficos
Vegetación semimesofítica
El análisis ecológico más completo de la vegetación de Puerto Rico hasta el momento fue el de Gleason y Cook (1926). El estudio fue inicialmente concebido por Cook, quien era patólogo de plantas de la Estación Experimental Agrícola de Río Piedras. Gleason y Cook consideraban la asociación de plantas como la unidad de la vegetación. Para ellos la asociación es el producto de la migración de especies y la selección natural por el ambiente. Su duración depende de la estabilidad del ambiente, larga cuando el ambiente es estable y corta cuando el ambiente cambia con más rapidez. La tabla 6.3 contiene la clasificación de la vegetación de Puerto Rico de acuerdo con el estudio de Gleason y Cook. El poco detalle para las montañas se debe a que no las visitaron tanto como a las comunidades costeras, quizás por dificultad de acceso. Además, los datos de lluvia que tenían eran limitados (Fig. 6.2a).
El trabajo de Gleason y Cook (1926) se llevó a cabo desde el punto de vista de la sucesión (Fig. 6.3) y una serie de principios que todavía son relevantes:
6 108 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Precipitación anual: promedio para el periodo 1963 - 1995

6 109 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Figura 6.2. El récord de lluvia disponible a (A) Gleason y Cook (1926), (B) el utilizado por el Departamento de Recursos Naturales (1973) durante la década de los 70, y (C) el que cualquier ciudadano puede disponer con acceso a internet en el presente. Los datos en «C» corresponden al promedio anual entre 1963-1995. Los datos en «A» y «B» están en mm/año y los datos en C en pulgadas/año Una pulgada es equivalente a 25.4 mm.
Precipitación (pulgadas) 50 km 28 - 45 45 - 65 65 - 85 85 - 100 100 - 150 150 - 181
• El ambiente afecta y a veces controla la estructura y comportamiento de las plantas. Las diferencias genéticas entre las especies evitan que la respuesta al ambiente sea homogénea en una comunidad, pero bajo ciertas condiciones algunas peculiaridades estructurales son comunes a muchas de las plantas.
• Ninguna especie puede sobrevivir en todos los ambientes, ya que cada una tiene un rango de condiciones a los cuales puede adaptarse. Por lo tanto, cuando se repiten las mismas condiciones en distintos lugares, se repite la presencia de las mismas especies. Estas modifican las condiciones ambientales favoreciendo el desarrollo de asociaciones de especies.
• El ambiente afecta y controla la distribución de cada especie en todo el mundo. Todos los individuos de la misma especie tienen, en general, las mismas demandas del ambiente y las especies solo pueden crecer en aquellas partes del mundo donde las condiciones necesarias ocurren. Muchas especies de plantas en el presente no ocupan todas las condiciones disponibles para las cuales están adaptadas.
• El desarrollo y mantenimiento de una asociación de plantas es el producto de migración de las especies y el ambiente.
• Las plantas tienden a controlar y cambiar el ambiente, por lo que pueden influenciar el éxito de la migración y el ambiente.
• Algunas especies, por el tamaño o abundancia de sus individuos, tienen más influencia sobre el ambiente que otras.
• El ambiente de las plantas está constantemente cambiando, por lo que la duración de las asociaciones es limitada y lleva a la sucesión.
• La velocidad del cambio ambiental regula la velocidad de la sucesión.
El Instituto Internacional de Dasonomía Tropical (Instituto), fundado en el 1939 bajo otro nombre y adscrito al Servicio Forestal, ha auspiciado o colaborado en la mayor cantidad de estudios ecológicos de los bosques de Puerto Rico (Lugo 2021). Los científicos de esta institución, empezando por Holdridge, han estudiado y contribuido al conocimiento de todos los bosques del país. Los estudios incluyen los bosques nativos, los noveles, los urbanos, los primarios, los secundarios, los humedales de agua dulce, los manglares y las plantaciones desde diversos puntos de vista como la silvicultura, hidrología, limnología, ornitología, ecofisiología, geoquímica, mensuración, dendrología, geografía, etc. El trabajo de Frank H. Wadsworth (1949) sobresale por ser el pionero en las medidas de crecimiento de los bosques en la isla y por establecer parcelas de investigación a largo plazo al principio de la década de 1940. Estas parcelas de estudio son las más antiguas en el Neotrópico y permiten analizar el comportamiento de los bosques en respuesta a múltiples
6 110 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
y el clima
y
y
(1926) durante el estudio de la vegetación (A) del sur de Puerto Rico (bosques secos) y la de la región del karso norteño (B). Las sucesiones incluyen series xerofíticas, hídricas de agua fresca, halárquicas de agua salada (en el sur de Puerto Rico) y las mismas series más la serie mesófila en el norte de Puerto Rico. El esquema propone que todas las series en cada región llegan a un bosque clímax.
eventos como los huracanes, la tala, eventos climatológicos con poca frecuencia, o el cambio climático.
En la década de 1960 se llevaron a cabo dos proyectos de investigación complementarios a los trabajos del Servicio Forestal en las Montañas de Luquillo encabezados por dos prominentes científicos norteamericanos. Richard A. Howard,
6 111 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Figura 6.3. Esquema de sucesión y relación entre la vegetación
en tierras bajas
costeras utilizado por Gleason
Cook
director del Arnold Arboretum de la Universidad de Harvard en Boston, dirigió un grupo de científicos norteamericanos en el estudio de los bosques elfinos (elfin forest sensu Beard 1955) del Pico del Oeste en El Yunque (Howard 1968). Esos estudios contribuyeron al entendimiento taxonómico, ecológico y fisiológico de los bosques elfinos, que algunos catalogan erróneamente como bosques enanos. También se utiliza bosque musgoso (mossy forest) al referirse a ellos por la abundancia de musgos y epifitas que tienen. Este crecimiento abundante de epifitas responde a que estos bosques tienen contacto con las nubes de donde obtienen agua y nutrientes, y por consiguiente, también se denominan bosques nublados o nubosos (cloud forests).
Los estudios dirigidos por Odum (Odum y Pigeon 1970) se llevaron a cabo en el sector El Verde de El Yunque, donde se sometió un predio del bosque de tabonuco a tres meses de radiación ionizante para estudiar su resiliencia ante la radiación gamma. Este estudio tenía importancia nacional e internacional, ya que en esa época surgía el uso de energía atómica con propósitos militares y no militares y tales usos ponían en peligro todos los ecosistemas del planeta. Era necesario entender cuáles serían sus efectos, pero el conocimiento ecológico no existía y era necesario adquirirlo. Por lo tanto, el estudio en El Verde fue el primero a gran escala de la ecología funcional de los bosques tropicales y sirvió para adelantar la ecología de ecosistemas tropicales que hasta entonces no había enfocado su atención al nivel funcional del ecosistema.
Cuando se estableció el DRNA en 1972, se incluyó una secretaría auxiliar de investigaciones científicas que a lo largo de los años hizo muchos trabajos ecológicos sobre los bosques de Puerto Rico. Aunque la secretaría auxiliar de investigaciones científicas fue eliminada posteriormente, el Departamento ha continuado promoviendo la investigación en sus bosques. Los trabajos más significativos fueron en los manglares (por ejemplo, Cintrón et al. 1978) y en el bosque seco de Guánica donde cientos de estudios se han llevado a cabo (ver número especial de Acta Científica 4:1990). El bosque seco de Guánica es segundo a El Yunque en términos de la intensidad de investigaciones ecológicas, lo que permite contrastar la resiliencia de dos bosques ubicados en polos opuestos en cuanto a la disponibilidad de agua para su funcionamiento (Lugo et al. 2006).
En 1988 el Instituto estableció las instalaciones para estudiar la ecohidrología del bosque de tabonuco en el sector Bisley de El Yunque. El estudio fue dirigido por el norteamericano Frederick N. Scatena (1989) y ha generado cientos de trabajos científicos sobre el comportamiento ecohidrológico de cuencas hidrográficas en las zonas escarpadas de los trópicos. El proyecto continúa activo y es el estudio de cuencas tropicales con mayor continuidad temporal en el Neotrópico. Uno de los logros notables del estudio fue la documentación ecohidrológica del efecto de un huracán categoría 3 (huracán Hugo) sobre la cuenca y su subsiguiente recuperación (Brokaw et
6 112 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
al. 2012a). Cuando la sucesión pos-Hugo llevaba 27 años, los huracanes Irma y María la interrumpieron y el proceso de sucesión tomó otro rumbo.
Cuando Odum terminó su estudio de irradiación, la Universidad de Puerto Rico, en colaboración con el Servicio Forestal, mantuvo estudios ecológicos en El Verde. En 1988, EE. UU. a través de la Fundación Nacional para las Ciencias (NSF por sus siglas en inglés), formalizó el estudio en el Verde con la designación del Bosque Experimental de Luquillo como un lugar para estudios ecológicos a largo plazo (Lugo 2021). La NSF también auspicia un estudio multidisciplinario con enfoque en lo que llaman la zona crítica. La zona crítica es el volumen de espacio desde el tope del dosel del bosque hasta la litosfera. Este proyecto tiene la participación de docenas de científicos de Puerto Rico y los EE. UU., incluyendo biogeoquímicos, hidrólogos, geólogos y químicos de la atmósfera (Anderson et al. 2016). Si se combinan estos estudios al estudio en el sector Sabana de El Yunque donde experimentalmente se le eleva la temperatura a una porción de un bosque novel (auspiciado por el Servicio Forestal y el Departamento de Energía de los EE. UU.), se devela un panorama de estudios en progreso con un alto nivel científico y con relevancia en la búsqueda del entendimiento del funcionamiento de los bosques tropicales en el contexto del Antropoceno.
En la región del karso, que cubre el 27 % de Puerto Rico (Lugo et al. 2001), se estableció una estación de campo en la reserva El Tallonal, administrada por los Ciudadanos del Karso, un grupo no gubernamental. Allí se llevan a cabo estudios en murciélagos, reptiles y anfibios, la flora del karso y la ecología de los bosques noveles. Recientemente, el puertorriqueño Omar Gutiérrez del Arroyo Santiago (2014) completó un estudio de la respiración edáfica y produjo el récord más largo de medidas continuas de la respiración de los suelos tropicales.
En las Islas Vírgenes, el Instituto estableció programas a largo plazo en el Bosque Experimental Estate Thomas (Weaver 2006a) y en los bosques de la cuenca de Cinnamon Bay (Weaver 2006b) en las islas de Santa Cruz y San Juan, respectivamente. El Bosque Experimental se estableció en 1964 y tiene representación de la zona de vida subtropical seca. Allí se ejecutaron numerosos trabajos dasonómicos y experimentos con el establecimiento y rendimiento de plantaciones de caoba (Swietenia macrophylla, S. mahagoni y un híbrido entre estas especies), teca (Tectona grandis) y cedro (Cedrela odorata) entre otras especies. Actualmente, las investigaciones en el Bosque Experimental incluyen además aspectos relacionados con bosque urbanos. En la cuenca de Cinnamon Bay, se estudiaron bosques secos, riparios y húmedos a través de 20 años que cubrieron sequías y el paso de dos huracanes.
Finalmente, a principios de 1982 comenzó el programa de inventarios periódicos de los bosques de Puerto Rico y las Islas Vírgenes (Birdsey y Weaver 1982). Este programa se lleva a cabo bajo el auspicio del Servicio Forestal y en colaboración con el DRNA de Puerto Rico y el Departamento de Agricultura de las Islas Vírgenes.
6 113 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Tabla 6.4
Algunos experimentos llevados a cabo en los bosques de Puerto Rico.
Experimento
Remoción de área basal para estimular el crecimiento de árboles
Siembra de distintas especies en pastizales abandonados para restaurar el bosque después de la deforestación
El cilindro gigante para medir el metabolismo de todo el bosque
Aplicación de radiación ionizante para estudiar la resiliencia de bosques tropicales
Aplicación de herbicidas para estudiar la efectividad de los químicos en relación con la apertura del dosel
Tala del bosque y aplicación de herbicidas para medir resiliencia
Tala del bosque para medir velocidad de sucesión y acumulación de biomasa
Tala del bosque para medir la eficiencia de retención de nutrientes
Fertilización del bosque para ver qué nutrientes son limitantes al crecimiento
Fertilización con nitrógeno a largo plazo en dos tipos de bosque con distintas elevaciones
Simulación de sequías para estudiar sus efectos geoquímicos
Remoción del dosel para estudiar los procesos del bosque después de un huracán
C alentar el sotobosque para estudiar la respuesta al calentamiento global
E xperimentos comparativos y recíprocos de descomposición de materiales diversos y de distintas localidades y latitudes para entender los factores que regulan el reciclaje de nutrientes
Referencia
Wadsworth (1949)
Marrero (1950)
Odum y Jordan (1970)
Odum y Pigeon (1970)
Dowler y Tschirley (1970)
Ewel (1971)
Murphy y Lugo (1986b)
Silver (1992)
Zimmerman et al. (1995)
Cusack et al. 2009
Wood y Silver (2012)
Shiels y González (2014)
Cavaleri et al. (2015)
González y Lodge (2017)
La tabla 6.4 contiene una lista parcial de experimentos de campo hechos en los bosques de Puerto Rico. La mayor parte de estos se llevaron a cabo en El Yunque y el bosque seco de Guánica. Los experimentos representan el estado más avanzado en los estudios de los bosques, ya que por medio de la replicación y reducción de la variabilidad ecológica, permiten hacer inferencias más precisas sobre los controles de la actividad biótica y los procesos ecológicos de los ecosistemas. Wood et al. (2019) contiene un resumen de los experimentos más recientes y su significado.
Clasificación y geografía de los bosques de Puerto Rico
En el capítulo anterior discutí el sistema de Beard para clasificar la vegetación del Caribe. Aquí, introduzco la clasificación geoclimática de los bosques de Puerto Rico que desarrolló originalmente el puertorriqueño Julio Figueroa Colón. El mapa que utilizo es la versión cartografiada por el Servicio Geológico de los EE. UU. en colaboración con el Servicio Forestal. Este sistema combina seis zonas de vida de Holdridge en Puerto Rico (Ewel y Whitmore 1973) con seis formaciones geológicas para delimitar las condiciones geoclimáticas bajo las cuales crecen los bosques en Puerto Rico (Fig. 6.4). De las 36 combinaciones de clima y geología potenciales, 28 ocurren en Puerto Rico (las otras 8 no). Agrupé las 28 combinaciones en 10 tipos de bosque para resumir el conocimiento disponible hasta el momento sobre la estructura
6 114 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Bosque pluvial subtropical
Bosque pluvial montano bajo
Bosque muy húmedo montano bajo
Bosque muy húmedo subtropical
Bosque húmedo tropical Bosque seco tropical
Terreno aluvial y otros sedimentos no consolidados Roca caliza y otras rocas de carbonato
Rocas sedimentarias sin carbonato
Rocas de efusivo, volcánicas y que sufren alteraciones hidrotermales Rocas intrusivas
Rocas ígneas ultramáficas
y composición de especies de los bosques de Puerto Rico (Lugo 2005a). Con el uso de sistemas de información geográfica, imágenes de satélite de alta resolución y los inventarios periódicos de los bosques llevados a cabo por el Servicio Forestal, es posible analizar la distribución geográfica de los bosques desde distintos puntos de vista. Por ejemplo, en términos de su cobertura (Fig. 6.5), su edad (Fig. 6.6), o la composición de especies (Fig. 4.14). Estos estudios dan como resultado estadísticas notables como por ejemplo:

• Los bosques cubren el 60 % del país, el valor más alto en los últimos dos siglos (Fig. 6.7).
• Los bosques urbanos cubren igual área que los bosques públicos protegidos.
• Los bosques de Puerto Rico son mayormente bosques jóvenes (menos de 64 años).
• Los bosques de Puerto Rico son mayormente bosques noveles con especies introducidas.
• Solamente el 25 % de los bosques de Puerto Rico son bosques nativos.
• Aproximadamente un 1 % de los bosques de Puerto Rico son primarios.
6 115 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Figure 6.4. Mapa de las zonas geoclimáticas de Puerto Rico. Este mapa lo desarrolló Julio Figueroa Colón mientras trabajaba en el Servicio Forestal de los Estados Unidos de Norteamérica.
Tipos de bosque
Urbano
Seco/AVS
Seco/calizo
Seco, Húmedo/ultramáfico
Húmedo/AVS
Húmedo, Muy húmedo/calizo
Muy húmedo AVS
Muy húmedo/LM/ultramáfico
Montano alto
Palma abandonada
Humedales boscosos
Tierras no forestadas
Descripción
Bosques en áreas urbanas
Bosques secos en sustratos aluviales, volcánicos y sedimentarios
Bosques secos en sustratos calizos
Bosques secos y húmedos en sustratos ultramáficos (serpentina)
Bosques húmedos en sustratos aluviales, volcánicos y sedimentarios
Bosques húmedos y muy húmedos en sustratos calizos
Bosques muy húmedos en sustratos aluviales, volcánicos y sedimentarios
Bosques muy húmedos montano bajo en sustratos ultramáficos
Bosques maduros sobre 600 m de elevación
Plantaciones de palmas de coco abandonadas
Bosques de mangle y Pterocarpus
Figura 6.5. Mapa con la distribución de los bosques de Puerto Rico y el área que ocupaban (hectáreas) según Gould et al. (2008) y Martinuzzi et al. (2013).

Todos los bosques
Figura 6.6. Mapa de los bosques de Puerto Rico clasificados por su edad (Kennaway y Helmer (2007). La escala está en kilómetros y las categorías incluyen áreas sin bosque, desarrollos urbanos y las distintas clases de edad de los bosques.

6 116 CAP ariel e. lugo | ecología a mi ma nera Los bosques de Puerto Rico
Hectáreas 27,311 21,592 19,822 4,365 143,543 67,445 125,408 3,364 31,432 491 7946 452,718 418,042 27 % 1 % 0.1 % 2 % 6 % 5 % 4 % 1 % 32 % 15 %
67º0'0''O 66º30'0''O 66º0'0''O 67º0'0''O 66º30'0''O 66º0'0''O 18º0'0'' N 18º30'0'' N 18º0'0'' N 18º30'0'' N Clase de edad de los bosques No forestados Desarrollos urbanos 1-9 años 10-22 años 23-49 años 50-> 64 años 0 5 10 20 km
Condiciones comunes de todos los bosques de Puerto Rico y sus efectos ecológicos
La figura 6.8 es un modelo de las fuerzas externas en la Isla que afectan la estructura, funcionamiento y composición de especies de todos sus ecosistemas. Los efectos antropogénicos se introdujeron en el cuarto capítulo y se discutirán en otros capítulos al presentar tipos de bosques, incluyendo los bosques noveles. En este capítulo enfatizo tres de las siete fuerzas no-antropogénicas que convergen en la Isla.
• La temperatura. La temperatura regula la velocidad del metabolismo de los organismos, particularmente la respiración y la fotosíntesis. Por su latitud, Puerto Rico tiene un régimen de temperatura que lo ubica en la banda subtropical del sistema de zonas de vida de Holdridge. La figura 6.9 demuestra que las temperaturas del aire en la base de El Yunque y en Guánica son similares a pesar de la discrepancia en la lluvia; son respectivamente regiones húmedas y secas.
• Los vientos alisios. Estos vientos son determinantes en la regulación del clima de Puerto Rico, ya que su presencia es confiable, moderan las temperaturas y traen masas de aire húmedo. Además de su función en la regulación del clima, los vientos alisios ventilan los bosques y estimulan el intercambio gaseoso necesario para mantener las tasas de producción y respiración de las plantas. Las masas de aire nutren las plantas epifitas con los nutrientes que cargan las nubes. Los vientos alisios y los vientos con fuerza de huracán también determinan la fisonomía de los bosques (discutido abajo).
6 117 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Figura 6.7. Cambios a través del tiempo en la cobertura (área) de los bosques en Puerto Rico, islas adyacentes y las Islas Vírgenes de los Estados Unidos de Norteamérica (EE. UU.). Los datos están basados en los reportes de los inventarios de bosques del Servicio Forestal de los EE. UU. (Birdsey y Weaver 1982,1987; Franco et al. 1997; Brandeis y Oswalt 2007; Brandeis et al. 2007; Brandeis y Turner 2013a,b; Marcano Vega 2019). Noten la escala logarítmica.
• Los disturbios. Los bosques de Puerto Rico están sujetos a sequías periódicas, pero el disturbio principal es el huracán. Un huracán, sin embargo, está compuesto de varios tipos de disturbios como lo son el viento, las lluvias, escorrentías altas, inundaciones, saturación de los suelos y derrumbes. El potencial de los huracanes como agentes evolutivos en el Caribe
6 118 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Figura 6.8. Factores biofísicos que inciden sobre Puerto Rico y contribuyen a la organización, estructura, composición de especies y funcionamiento de los sistemas ecológicos isleños (tope) y las principales masas de aire que convergen en la isla (abajo) (Lugo y Scatena 1995).
se puede apreciar al estimar que la flora de Puerto Rico ha sido expuesta a por lo menos 4.5 millones de eventos huracanados (asumiendo 0.45 huracanes por año) en los últimos 10 millones de años.
Estos tres factores afectan todos los bosques isleños y contribuyen a la determinación de su fisonomía, metabolismo, estructura y la composición de especies. Como los factores mencionados son comunes en la región insular del Caribe, no debe sorprender que se observen similitudes tanto entre los bosques de Puerto Rico como en los bosques caribeños en general.
Los huracanes y otros disturbios que ocasionan condiciones de rápido crecimiento vegetal son responsables por la presencia de cohortes arbóreos de la misma edad en los bosques de Puerto Rico. Después del disturbio, las condiciones en los bosques favorecen el establecimiento de poblaciones arbóreas que aprovechan la apertura del dosel para crecer juntos. La densidad de especies (número de especies por unidad de área) es otro parámetro que varía poco, no importa el régimen de humedad. El bosque seco y el pluvial, por ejemplo, tienen la misma densidad de especies arbóreas (aproximadamente 60 especies por hectárea). La incidencia de energía solar, la temperatura, los vientos alisios y el paso de huracanes aparentan ser las causas principales de este patrón de similitud en los bosques de Puerto Rico.
Otra característica común en todos los bosques caribeños son los doseles parejos (Fig. 6.10) sin árboles emergentes. Esto se debe a la poda continua de los vientos alisios y los efectos agudos de los vientos huracanados. El viento continuo favorece los doseles parejos y actúa como una fuerza selectiva contra los árboles emergentes. Los bosques de Puerto Rico también están en transición debido a la recurrencia y
6 119 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Figura 6.9. Temperatura máxima, mínima y media promedio en dos estaciones en las tierras bajas pero en costas opuestas, central Guánica en la costa sur y río Blanco en la costa norte de Puerto Rico (Lugo 2005a).
los efectos de los huracanes. Además, exhiben alta dominancia por parte de pocas especies arbóreas y baja diversidad de especies. Estas dos características las he atribuido como una respuesta a la frecuencia de los huracanes (Lugo 1991).
Seis condiciones que ayudan a explicar la biogeografía y diversidad de los bosques de Puerto Rico
Puerto Rico tiene 28 zonas geoclimáticas dentro de las cuales ocurren los bosques (Fig. 6.4). Sin embargo, la riqueza de asociaciones vegetales dentro de esas zonas geoclimáticas es alta debido a por lo menos seis factores no-antropogénicos adicionales que diversifican el paisaje. El efecto antropogénico y las variaciones de sucesión de cada asociación aumentan aún más el número de asociaciones vegetales en la isla. Termino este capítulo con una descripción breve de cómo estos seis factores no-antropogénicos contribuyen a la diversidad de bosques en Puerto Rico.
1. La topografía y geomorfología. La topografía y geomorfología de Puerto Rico son diversas y proveen múltiples oportunidades para diversificar el paisaje y sus bosques. Topografía escarpada y cambios geomorfológicos entre zonas como la del karso, las aluviales o volcánicas en un área geográfica tan limitada acentúa los contrastes en los bosques que, aunque exhiban la misma
6 120 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Figura 6.10. Contraste en la fisonomía de bosques tropicales en el Caribe (A) bajo la influencia de vientos alisios y huracanes y (B) bosques ecuatoriales sin esos efectos (Odum 1970).
fisonomía, difieren en el tipo de especie que albergan, en su productividad primaria o la velocidad del reciclaje. En los humedales, el tipo de vegetación cambia con variaciones sutiles (cm) en la topografía del terreno. Cambios topográficos afectan el hidroperiodo (discutido abajo) y la saturación del suelo.
2. Los suelos. Puerto Rico tiene casi todos los órdenes de suelo existentes en el mundo (menos dos) y 213 series de suelos (Fig. 6.11, Tabla 6.5). Los dos órdenes de suelo que no ocurren en Puerto Rico son Gelisoles, debido al clima, pues son suelos con permafrost, y los Andisoles, que requieren la presencia de volcanes activos. Los órdenes con mayor cobertura son los Inceptisoles y Ultisoles. La mayor diversidad de series ocurre en los suelos Inceptisoles y Molisoles (Muñoz et al. 2018). Los Inceptisoles ocurren en superficies geomórficas jóvenes en las montañas interiores y los valles aluviales costeros. Los suelos Molisoles son los más fértiles y ocurren en dos zonas de Puerto Rico: en las zonas calizas del Terciario y en zonas inundables aluviales en los valles costeros

6 121 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
67º0'0''O 66º40'0''O 66º20'0''O 66º0'0''O 65º40'0''O 67º53'30''O 67º53'30''O 00.751.5 3 4.5 6 millas 18º4'30''N Alfisoles Aridisoles Entisoles Histosoles Inceptisoles Leyenda Miscelaneos Molisoles Oxisoles Spodosoles Ultisoles Vertisoles 18º4'30''N 67º20'0''O 67º0'0''O 66º40'0''O 66º20'0''O 66º0'0''O 65º40'0''O 18º40'0''N 18º20'0'' N 18º0'0''N 17º40'0'' N 18º40'0''N 18º20'0'' N 18º0'0''N 17º40'0'' N 65º33'0''O 65º22'30''O 65º12'0''O 65º33'0''O 65º22'30''O 65º12'0''O 0 3 6 12 18 24 millas 18º19'30''N 18º19'30''N 18º9'0''N 18º9'0''N 0 5 10 20 30 40 millas N
Figura 6.11. Distribución de los órdenes de suelos en Puerto Rico e islas adyacentes (Muñoz et al. 2018).
Tabla 6.5
Los órdenes de suelos, el número de series en cada orden y la cobertura del suelo en Puerto Rico (Muñoz et al. 2018). Orden
de seriesCobertura
de la isla)
e interiores, particularmente en el sur de Puerto Rico. Los Ultisoles son suelos en etapas avanzadas de meteorización con pH bajo y altas concentraciones de aluminio. Son comunes en la región oeste de Puerto Rico y en las Montañas de Luquillo. La diversidad de suelos contribuye a la diversidad de bosques en Puerto Rico, ya que cada tipo de suelo presenta distintos problemas y oportunidades a los bosques en términos de su capacidad para suplir agua, nutrientes y anclaje a las plantas. Estas diferencias a su vez se manifiestan en ajustes ecofisiológicos de las plantas y en efectos acumulativos en la biogeoquímica de los bosques.
3. La elevación. Los cambios en elevación introducen múltiples gradientes en los factores ambientales que regulan el funcionamiento de los bosques. En general, la temperatura disminuye, la lluvia aumenta, la radiación solar disminuye, la velocidad del viento aumenta, el poder evaporativo de la atmósfera disminuye y la humedad relativa aumenta con la elevación. Los sistemas bióticos responden a estos gradientes en formas complejas, no todas conocidas (González et al. 2013). Por lo tanto, aunque algunas respuestas de los bosques a la elevación son fáciles de observar, como la disminución en la altura de los bosques (Brown et al. 1983), hay otras que son difíciles de observar como la reducción en la productividad primaria. La maximización de la complejidad de los bosques (sensu Holdridge 1967; Brown et al. 1983) y la potencia máxima del flujo de energía (Harris et al. 2013) en El Yunque, ocurren entre 300 y 700 m de elevación, o sea, no son proporcionales a la elevación. Estudios de la vegetación a lo largo de un transecto entre 250 y 1,000 m indican que cuando se llega al nivel de condensación de las nubes (600 m de elevación) ocurren cambios fundamentales en la composición y funcionamiento de los bosques, a pesar de que las especies se distribuyen en forma individual de acuerdo con sus adaptaciones a lo largo del transecto (Fig. 6.12).
4. La catena. El gradiente topográfico que incluye una cima o tope, ladera y valle, conocido como catena, exhibe condiciones que afectan el comportamiento
6 122 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
(%
Inceptisoles 45 29.2 Ultisoles 29 19.7 Molisoles 36 15.1 Oxisoles 24 7.7 Alfisoles 17 4.6 Vertisoles 19 4.3 Entisoles 17 2.3 Aridisoles 15 1.5 Histosoles 8 0.5 Spodosoles 3 0.2 Total 213 85.1
Número
Prestoea montana
Psychotria berteriana
Dacryodes excelsa
Manilkara bidentata
Rheedia portoricensis
Magnolia splendens
Cyathea bryophila
Lobelia portoricensis
Guarea glabra
Heliconia caribaea
Hirtella rugosa
Miconia sintenisii
Miconia tetrandra
Micropholis garciniifolia
Ocotea leucoxylon
Ocotea spathulata
Psychotria maleolens
Rheedia portoricensis
Sloanea berteriana
Tabebuia rigida
Daphnopsis philippiana
Figura 6.12. Variación en la densidad de árboles de 21 especies arbóreas dominantes a lo largo de un transecto entre <300 m y 1,000 m en las laderas de la cuenca del río Sonadora en el Bosque Experimental de Luquillo (Heartsill Scalley 2012). de las especies, las condiciones edáficas y el funcionamiento del bosque (Fig. 6.13). Los bosques en el tope de una catena tienen estructura, funcionamiento y composición de especies distintas a los que crecen en los valles o las laderas (Scatena y Lugo 1995). Más aún, las características de las catenas varían con la elevación, el aspecto, y las condiciones climáticas donde ocurren (Weaver 1995).
5. El aspecto o exposición al viento y el sol. Exposiciones o aspectos hacia el norte de Puerto Rico reciben más lluvia que los aspectos al sur (Fig. 6.14). Aspectos hacia el viento (barlovento) reciben vientos más fuertes y frecuentes que exposiciones o aspectos de sotavento (protegidos de los vientos prevalecientes). El efecto del aspecto también se refleja en la producción de agua de las cuencas hidrográficas (Murphy y Stallard 2012). Por supuesto, la trayectoria de un huracán puede exponer cualquier aspecto a vientos fuertes. En el karso del norte de Puerto Rico, exposiciones hacia el oeste son más secas que aspectos hacia el

6 123 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
6 124 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Figura 6.13. Cambios estructurales y geoquímicos a lo largo de catenas en el bosque de tabonuco del sector Bisley, Bosque Experimental de Luquillo (Scatena y Lugo 1995).
este (Aukema
Los
acumulativos del aspecto de los rodales se manifiestan en la estructura y composición de especies de esos rodales.
6. El hidroperiodo, o tiempo de inundación o saturación de suelos. Los suelos se saturan con mucha lluvia o si tienen poca capacidad de drenaje, como ocurre con suelos arcillosos. Generalmente, un suelo saturado pierde oxígeno y se torna anaeróbico, una condición típica de humedales. En Puerto Rico ocurren humedales en las laderas de las montañas (Frangi 1983) debido a la alta precipitación y condiciones edáficas. Los humedales de las montañas tienen suelos con bajo oxígeno, lo que causa que la vegetación tenga menos diversidad y adaptaciones para airear los tallos y raíces. Las comunidades microbianas y los procesos de descomposición cambian radicalmente bajo condiciones anaeróbicas y es común que emitan el gas metano (Silver et al. 1999).


Asignación
Selecciona cualquier tipo de bosque en Puerto Rico y lee cualquier trabajo sobre ese bosque. Para la clase, determina en qué zona de vida estuvo ubicado el estudio y qué aprendiste de la lectura sobre ese bosque.
6 125 CAP ariel e. lugo | ecología a m i m anera Los bosques de Puerto Rico
Figura 6.14. Perfil transversal de Puerto Rico entre el océano Atlántico y el mar Caribe que ilustra la variación en la precipitación anual y el aspecto y altura de las nubes (Lugo 2005a).
et al. 2007).
efectos
CaPítulo 7 Bosques secos
Pregunta
¿Por qué el bosque seco de Guánica es tan seco?
Introducción
Los bosques secos son los bosques tropicales con mayor extensión geográfica (el 42 % de los bosques tropicales son bosques secos) y los más intensivamente utilizados por el ser humano (Murphy y Lugo 1986a). El uso intensivo crea problemas de conservación por sus altas tasas de deforestación y degradación. A pesar de estar limitados por el agua y a veces por nutrientes, los bosques secos antillanos tienen alta resiliencia (Lugo et al. 2002, 2006). En Puerto Rico y las Islas Vírgenes ocurren bosques secos sobre substratos calizos, volcánicos, ultramáficos (igual a serpentina) y aluviales (Lugo 2005a, Weaver y Gould 2013). Además, el clima sobre las costas de Puerto Rico tiende a ser seco, por lo que se encuentran bosques secos (bosques litorales dominados por uva de playa [Coccoloba uvifera] y estudiados por Gleason y Cook 1926) en lugares donde los mapas indican costas húmedas. En este capítulo enfatizo los bosques secos sobre substratos calizos y volcánicos por ser los más abundantes en Puerto Rico y porque contrastan en su desarrollo estructural.
Bosques secos en general
En el sistema de Holdridge (1967) un bosque seco crece en zonas de vida donde el cociente PET/P (capítulo 5) es mayor a uno, indicando más evapotranspiración potencial que precipitación (lluvia) o sea, un déficit de agua. En Guánica, el cociente PET/P es de 1.4. En general, los bosques crecen cuando la precipitación anual es > 500 mm. Los bosques secos ocurren entre 500 y 1,000 mm de precipitación anual. La lluvia anual promedio en Guánica es de 880 mm, pero dependiendo del año puede variar entre 300 y 1300 mm.
La Tabla 7.1 resume algunas características y coberturas de los bosques secos en Puerto Rico y las Islas Vírgenes. Las diferencias estructurales entre los bosques secos en ambas islas son menores, mayormente en la densidad de árboles, la biomasa
ariel e. lugo | ecología a mi ma nera
127
Tabla 7.1
Algunos parámetros geográficos y ecológicos de los bosques secos de Puerto Rico (Brandeis y Turner 2013) e Islas Vírgenes de los Estados Unidos de América (Brandeis y Oswalt 2007). El diámetro a la altura del pecho de los árboles es dap. Los datos de Puerto Rico corresponden al 2009 y los de las Islas Vírgenes al 2004. Las unidades están en paréntesis. El área total de bosques en Puerto Rico era de 472,791 ha y en las Islas Vírgenes 21,237 ha.
subterránea y el almacenaje total de carbono, todas favoreciendo a los bosques secos en las Islas Vírgenes. En 2009, los bosques secos representaban el 17 % de los bosques de Puerto Rico. En el año 2004, los bosques secos en las Islas Vírgenes representaban el 59 % de los bosques en esas islas. Al interpretar los datos en la tabla 7.1 y compararlos con estudios puntuales, es importante entender el tipo de muestreo de los estudios y los métodos de estimación de parámetros ecológicos. Los datos en la tabla 7.1 están basados en inventarios en el paisaje con muestreos aleatorios. Los estudios puntuales en general son menos representativos que los inventarios forestales debido al área reducida que cubren.
Factores que afectan el crecimiento de los bosques secos
El balance hídrico, generalmente definido por la lluvia, es el factor que define lo que es un bosque seco, y sus gradientes estructurales reflejan la lluvia anual que reciben. Sin embargo, por su efecto fisiológico sobre las plantas, el número de días y meses secos pueden ser tan importantes como la lluvia. Los bosques secos experimentan más días y meses secos que cualquier otro tipo de bosque tropical (Medina 1995).
La profundidad del suelo es crítica para almacenar agua y nutrientes. Dependiendo de la profundidad y el tipo de suelo, se observan variaciones en la densidad y altura de la vegetación de bosques secos. Las cactáceas crecen sobre rocas donde la presencia del suelo se limita a hendiduras entre las rocas calizas típicas de Guánica e Isla de Mona. Dependiendo del tipo de suelo y su profundidad, se desarrollan también
7 128 CAP ariel e. lugo | ecología a m i m anera Bosques secos
Parámetro Puerto Rico^
Vírgenes Área (ha) 80,937 12,533 Árboles con dbh entre 12.7 y < 27.9 cm (%) 93 41* Biomasa aérea (Mg/ha) 40.4 39 Biomasa subterránea (Mg/ha) 4.5 10 Carbono total (Mg/ha) 22.4 24.5 Crecimiento en volumen (m3/ha.año) 1.19 Mortalidad (m3/ha·año) 0.84 Densidad (tallos/ha) 3,088** 4,000** Área basal (m2/ha) 9.37** 10** Volumen (m3/ha) 16.8** 12** ^ dap ≥ 5.0 cm * 10 a 30 cm dap ** dap ≥ 2.5 cm
Islas
gradientes en la disponibilidad de nutrientes. Más aún, el carbonato de calcio de los suelos calizos puede inmovilizar el fósforo, lo que causa limitaciones nutricionales a las plantas. No así, en los suelos volcánicos que son más profundos que los suelos kársticos en Puerto Rico.
La comunidad arbórea con el máximo desarrollo estructural en Guánica ocurre en los valles de las catenas y a lo largo de los cañones, formados por quebradas intermitentes que normalmente no tienen agua, pero que durante eventos de lluvia intensa descargan enormes cantidades de agua. Los árboles más grandes en Guánica se encuentran en estos cañones (Castilleja 1991).
En bosques secos costeros hay gradientes de viento y salitre que afectan la vegetación, lo que ocasiona que los árboles se achaparren en lugares con condiciones extremas de viento y salitre. Medina et al. (2012) describieron un bosque enano costero expuesto a fuertes vientos y salitre, lo que causó que la altura del bosque fuese entre 0.4 y 2.3 m en un transecto entre el mar y 150 m de este. La poca estatura y baja diversidad florística del bosque contrasta con la aparente madurez, que se considera bosque primario. Allí, Medina et al. (2012) encontraron a Tabebuia heterophylla, una especie de bosque húmedo, empequeñecida y creciendo con especies del bosque seco que también tomaban forma enana.
Las brisas marinas crean condiciones atmosféricas nocturnas cuando la humedad del aire se condensa de sobre la vegetación. En Guánica, Gleason y Cook (1926) observaron un frente de epifitas perpendiculares a las brisas marinas y a una elevación particular donde coincidían con la condensación atmosférica nocturna. Murphy y Lugo (1986b) encontraron que como consecuencia de este fenómeno, el bosque seco de Guánica tenía más biomasa de epifitas que los bosques húmedos amazónicos de Venezuela.
Características ecológicas y ecofisiológicas de los bosques secos
La alta capacidad de evaporación de la atmósfera y las altas temperaturas diurnas en los bosques secos afectan el funcionamiento ecofisiológico de las plantas. Las plantas de los bosques secos tienen adaptaciones foliares que optimizan la temperatura de las hojas, el intercambio gaseoso con la atmósfera y la conservación de nutrientes limitantes. Algunos ejemplos incluyen: (1) El movimiento foliar donde las hojas pueden estar perpendiculares o verticales con relación a la incidencia de los rayos de luz. La posición de las hojas depende de la intensidad en la incidencia de energía y la disponibilidad de agua para evaporar. Bajo condiciones extremas, las hojas se enrollan antes de caer. (2) Predominan hojas pequeñas en términos del área foliar y con área específica intermedia (92 cm 2 /g), lo cual ayuda en la conservación de agua y nutrientes. (3) Algunas plantas del bosque seco son suculentas lo que les ayuda a lidiar con la sequía, o almacenan agua en los tallos
7 129 CAP ariel e. lugo | ecología a m i m anera Bosques secos
Tabla 7.2
Comparación entre bosques secos y muy húmedos con base en varios parámetros ecológicos (Murphy y Lugo 1995).
como el almácigo (Bursera simaruba). (4) Las plantas suculentas como el cactus pueden invertir el patrón diurno de intercambio de gases abriendo las estomas de noche y así evitan perder agua de día. El bióxido de carbono que fijan estas plantas de noche en forma de ácido málico, lo procesan de día con los estomas cerrados. El proceso se conoce como el metabolismo ácido de crasuláceas (CAM, por sus siglas en inglés). (5) Altas tasas de reabsorción o retranslocación de nutrientes limitantes como el fósforo en hojas senescentes antes de que caigan.
Los bosques secos tienen bajos índices estructurales en comparación con los bosques húmedos (Tabla 7.2). Por ejemplo, bajos índices de área foliar, baja altura del dosel y baja biomasa. Sin embargo, la densidad de tallos puede ser superior a la de los bosques húmedos debido a que los árboles son de menor diámetro y ocupan menos espacio. La distribución de la biomasa es también distinta a la de los bosques húmedos. Los bosques secos tienen una alta proporción de biomasa subterránea en contraste con bosques húmedos que tienen una fracción más alta de biomasa aérea (Murphy y Lugo 1986a). En Guánica, la mitad de la biomasa del bosque siempreverde (sensu Beard) es subterránea (Murphy y Lugo 1986b). Este bosque tiene una biomasa de 89.9 Mg/ha, un valor menor al bosque seco en Ceiba sobre substrato volcánico (163 Mg/ha; Weaver y Gould 2013). La tabla 7.3 sugiere que la distribución de carbono y nitrógeno en los bosques secos antillanos es diferente a la de los bosques secos continentales. Bosques secos antillanos tienen 2.9 y 15.3 veces más carbono y nitrógeno (respectivamente) bajo la superficie comparado con la cantidad aérea que los bosques secos continentales con 1.4 y 7.2, respectivamente. La causa de estas diferencias pude ser la presencia de huracanes en el Caribe y no en Centro América, ya que la masa subterránea está protegida del viento y sirve para impulsar el nuevo crecimiento después del evento. Los almacenajes subterráneos de carbono y nitrógeno contribuyen a la resiliencia ante los huracanes y otros eventos como la tala, según demostró el norteamericano John Jeffrey Ewel en experimentos con el bosque seco de Guánica (Ewel 1971).
7 130 CAP ariel e. lugo | ecología a m i m anera Bosques secos
Parámetro Rango de valores a nivel global Seco Muy húmedo Especies arbóreas en 1-3 ha 33-90 50-200 Altura del dosel (m) 10-40 20-84 Área basal (m2/ha) 17-40 20-75 Biomasa total (Mg/ha) 98-320 269-1186 Biomasa de raíces (por ciento del total) 8-50 5-33 Productividad primaria aérea neta (Mg/ha año) 6-16 10-22
Tabla 7.3
Almacenaje de carbono y nitrógeno sobre y debajo de la superficie de dos bosques secos; uno insular antillano (Guánica) el otro continental en México (Lugo et al. 2006).
Funcionalmente, los bosques secos tienen la misma capacidad productiva por unidad de área foliar que otros tipos de bosques en ambientes más favorables para el crecimiento (Lugo et al. 1978). Sin embargo, el tiempo disponible para maximizar su capacidad productiva es más corto en el bosque seco que en un bosque húmedo. Por ejemplo, los bosques secos no pueden mantener altas tasas fotosintéticas durante épocas de sequía y su índice de área foliar es menor al de los bosques húmedos. Por lo tanto, al expresar la productividad anual por unidad de área suelo, los bosques secos exhiben menos productividad que los húmedos. Sin embargo, la eficiencia en el reciclaje de nutrientes por unidad de materia orgánica en circulación es más alta en los bosques secos que en los bosques húmedos (Lugo y Murphy 1986).
Las comunidades vegetales de los bosques secos exhiben alta dominancia, según se aprecia en sus curvas de Importancia Relativa (Lugo 1991; Fig. 7.1). Estas curvas tienden a ser más verticales y cortas en los bosques secos comparados con los bosques húmedos (Lugo 2005a,b). La dominancia es mayor en los árboles grandes
7 131 CAP ariel e. lugo | ecología a m i m anera Bosques secos
Parámetro Continental Antillano Carbono (Mg/ha) Aéreo 58.3 37.8 Subterráneo Raíces a 60 cm 6.7 22.5* Suelo a 60 cm 76.2 86.5* Total 141.1 146.8 Nitrógeno (kg/ha) Aéreo 940 631 Subterráneo Raíces a 60 cm 106 546* Suelo a 60 cm 6,659 9,100* Total 7,704 10,277
de profundidad
* a 85 cm
Figura 7.1. Rango de las especies arbóreas de los bosques secos de Puerto Rico y las Islas Vírgenes de los Estados Unidos de América (EE. UU.) basado en sus Valores de Importancia. Los datos son del inventario de bosques de 2009 (Brandeis y Turner 2013a,b).
Tabla 7.4
Índices en porcentaje relacionados a la importancia de las especies nativas y naturalizadas en los bosques secos de Puerto Rico y las Islas Vírgenes de los Estados Unidos de Norteamérica. El área muestreada fue de 3.95 ha y 4.96 ha para árboles en Puerto Rico con menos de 12.7 cm y más de 12.7 cm de diámetro a la altura del pecho (dap, respectivamente) y 1.68 ha para los datos de las Islas Vírgenes. Los datos corresponden al 2009 para Puerto Rico (Brandeis y Turner 2013) y 2004 para las Islas Vírgenes (Brandeis y Oswalt 2007).
Grupo
* < 12.7 cm dap
** > 12.7 cm dap
comparados con los más pequeños, pero la riqueza de especies es mayor en los árboles pequeños comparado con los árboles grandes (Fig. 7.1). En Puerto Rico, se encontraron más especies en los bosques secos que en las Islas Vírgenes. Los bosques secos en estas islas tienen baja diversidad florística comparada con bosques húmedos y generalmente están dominados por especies nativas (Tabla 7.4), ya que pocas especies introducidas tienen adaptaciones a las condiciones extremas de sequía o de baja disponibilidad de nutrientes que son comunes en bosques secos. Leucaena leucocephala (zarcilla o tantan), Prosopis juliflora (bayahonda o mesquite) y Swietenia mahagoni (caoba), todas introducidas a Puerto Rico, son excepciones. En el bosque seco de las Islas Vírgenes, las especies introducidas con los Valores de Importancia (en paréntesis) más altos y diámetros ≥ 12.7 cm son Melicoccus bijugatus (9.37), Swietenia mahagoni (5.29) y Acacia farnesiana (1.4).
Efectos antropogénicos
Ewel (1971) y Molina Colón y Lugo (2006) estudiaron la resiliencia de los bosques secos a disturbios antropogénicos. Ewel, la respuesta a la tala y los herbicidas. La puertorriqueña Sandra Molina Colón estudió la respuesta del bosque seco de Guánica a los cambios en la cobertura, incluyendo vivienda, agricultura, parque de pelota y carboneras (Fig. 7.2). A pesar de las limitaciones climáticas y nutricionales, los bosques secos de Guánica respondieron a esas perturbaciones con rapidez (Figs. 7.3 y 7.4).
Los patrones de cambio en la densidad de especies durante la sucesión después de un disturbio en el bosque seco son similares a los patrones observados
7 132 CAP ariel e. lugo | ecología a m i m anera Bosques secos
de especies Número Densidad relativa Dominancia relativa Frecuencia relativa Valor de importancia Puerto Rico Nativas* 72 77.9 73.4 86.9 79.4 Naturalizadas* 6 22.1 26.6 13.1 20.6 Nativas** 35 84.1 82.9 84.7 83.9 Naturalizadas** 8 15.9 17.1 15.3 16.1 Islas Vírgenes Nativas* 20 81.9 75.4 82.1 79.8 Naturalizadas* 7 18.1 24.6 17.9 20.2
tras el abandono del uso: carboneras, agricultura, vivienda, juego de pelota y bosque. Noten la
7 133 CAP ariel e. lugo | ecología a m i m anera Bosques secos Bosque estatal de Guánica El Maniel Bosquemaduro
N
m Tierra agrícola Bosque cerrado Bosque abierto Suelo desnudo Carretera 334 Sendero Camino Carbonera Casas
Parquedepelota
0200600
Figura 7.2. Mapa de la región del Bosque Estatal de Guánica donde Sandra Molina Colón (1998) condujo su estudio de sucesión luego del abandono de la actividad agrícola. Se ilustra la ubicación de distintos usos de la tierra donde se evaluaron los bosques allí presentes
distribución de las carboneras.
en otros bosques de Puerto Rico (Fig. 7.5). Los bosques secos son bosques con alta resiliencia (Lugo et al. 2002, 2006), ya que tienen características que los pre adaptan a disturbios no-antropogénicos y antropogénicos (Fig.7.6). Una de las adaptaciones más importantes para contrarrestar talas y huracanes es el alto almacenaje de materia orgánica y nutrientes en el suelo y las raíces (Fig. 7.7, Tablas 7.2 y 7.3).
La resiliencia de los bosques secos a la fragmentación también se observa al nivel o escala del paisaje (Ramjohn et al 2012). Ellos observaron una alta densidad de especies en rodales aislados o fragmentados en la costa sur de Puerto Rico. Encontraron que con poca área de bosque, incluyendo fragmentos aislados, se conservaban fracciones importantes de las especies, incluyendo especies endémicas, que caracterizan los bosques maduros de la región (Fig.7.8). Específicamente, observaron entre 1 y 86 % de las especies de bosque seco maduro en fragmentos aislados.
7 134 CAP ariel e. lugo | ecología a m i m anera Bosques secos
Figure 7.3. Parámetros estructurales (A , B), funcionales (C) y de composición de especies (A) en el bosque seco en Guánica luego de 42 años de sucesión en tres tipos de tierras abandonadas: vivienda, agricultura y carboneras. Los valores reportados están expresados en porcentaje relativo a los respectivos valores en un bosque maduro cercano (Molina Colón 1998). El índice de cobertura del dosel es CAI, por sus siglas en inglés.
Figura 7.4. Parámetros estructurales (biomasa, altura, densidad de tallos, porcentaje de cobertura, índice de área foliar) y riqueza de especies en el bosque seco en Guánica luego de una tala (tiempo cero). Los puntos representan datos de campo proporcionados entre ellos pero la línea entrecortada es hipotética y representa el patrón esperado en respuesta a la tala (Dunevitz 1985).
Figura 7.5. Patrones de cambio en el número de especies arbóreas luego de disturbios en bosques muy húmedos (huracán de 1932 sobre el bosque de tabonuco de El Verde en el Bosque Experimental de Luquillo) y secos (disturbios antropogénicos sobre el bosque seco de Guánica) en Puerto Rico. Como los datos provienen de distintas áreas de muestreo, esta figura solamente ilustra el patrón común en la respuesta al disturbio (Lugo et al. 2002).
7 135 CAP ariel e. lugo | ecología a m i m anera Bosques secos
7 136 CAP ariel e. lugo | ecología a m i m anera Bosques secos
Figura 7.7. Distribución con la profundidad del suelo de la biomasa de raíces en el bosque seco de Guánica (Murphy y Lugo 1986b).
Figura 7.6. Combinación de factores que contribuyen a la alta resiliencia de los bosques secos antillanos (Modificado de Lugo et al. 2006). La información en negritas es de Dickinson et al. 2001).
Las bases de la resiliencia y sus límites (Fig. 7.6)
Los bosques secos adaptados a condiciones ambientales extremas (humedad, temperatura y nutrientes) exhiben resiliencia no solo a los extremos en las perturbaciones no-antropogénicas tales como sequía, eventos de calor o huracanes, sino que también presentan resiliencia ante disturbios antropogénicos como la tala, deforestación e incluso herbicidas. Las bases de esta resiliencia son las siguientes: diversidad de formas de vida (Tabla 7.5; Medina 1995), resistencia al viento (Van Bloem et al. 2005), alta proporción de raíces (Murphy y Lugo 1986b), alto almacenaje subterráneo de nutrientes y materia orgánica (Tabla 7.3; Lugo y Murphy 1986), alta eficiencia en el reciclaje de nutrientes (Lugo y Murphy 1986), alta dominancia de especies (Fig. 7.1; Lugo 1992, 2005a) y alta capacidad para formar rebrotes en respuesta a la tala o la deforestación (Molina Colón y Lugo 2006, Van Bloem et al. 2003).
Fundamentalmente, la resiliencia de los bosques secos se basa en altas acumulaciones de materia orgánica y nutrientes en compartimientos protegidos por los efectos mecánicos del viento o la deforestación. Estos compartimientos se conocen como variables lentas (slow variables) que proveen estabilidad y amortiguamiento a los disturbios periódicos. La función estabilizadora de estas variables está complementada por procesos que aseguran la utilización óptima de los recursos disponibles al bosque. Por ejemplo, los rebrotes, la eficiencia del reciclaje, o la diversidad de formas de vida. Según se presenta en la tabla 7.5, las diversas formas de vida representan diversas funciones que contribuyen a la utilización óptima de los recursos de luz, agua y nutrientes.
Sin embargo, los bosques nativos de Puerto Rico, incluyendo los bosques secos, carecen de adaptaciones para sobreponerse a los efectos de los incendios. Esto
7 137 CAP ariel e. lugo | ecología a mi ma nera Bosques secos
Figura 7. 8. Relación entre el área del fragmento de bosque seco y el porcentaje de especies del bosque maduro presentes en el fragmento (Ramjohn et al. 2012). Los fragmentos se estudiaron en la costa sur de Puerto Rico. Noten la escala logarítmica.
Tabla 7.5
Diversidad de formas de vida en bosques secos y sus atributos funcionales (Medina 1995). El metabolismo ácido de la crasuláceas es CAM, por sus siglas en inglés.
Forma de vida y características morfológicas Atributos funcionales
Árboles y arbustos
Mesófilos Deciduos
Esclerófilos
Siempreverdes
Suculentos C 3 y Fotosíntesis CAM
Herbáceas
Yerbas Anuales, C 3 y C4 Fotosíntesis
Dicotiledóneas
Perennes
Suculentas C 3 y Fotosíntesis CAM
Bejucos
Herbáceos
Leñosos
Mesófilos
Deciduos
Esclerófilos Siempreverdes
Suculentos C 3 y Fotosíntesis CAM
Epifitas
Mesófitas
Esclerófilas
Suculentas
Parásitas
Deciduas
Siempreverdes
C 3 y Fotosíntesis CAM
Conectadas al xilema de árboles
quiere decir que hay disturbios antropogénicos que retardan o redirigen la sucesión, causando cambios fundamentales en la composición de especies de los bosques secos. Otros disturbios antropogénicos para los cuales los bosques secos carecen de suficiente resiliencia para contrarrestarlos son: apertura desmedida del dosel, erosión del suelo y su materia orgánica o remoción de raíces. Cuando estos disturbios extremos ocurren, las especies nativas tienen problemas para restablecer el bosque original, la sucesión pierde velocidad, entran especies naturalizadas o introducidas y se regeneran bosques noveles, quizás con menor estatura o complejidad dependiendo de la intensidad y duración de los disturbios antropogénicos. Los bosques noveles secos han sido estudiados por Molina Colón y Lugo (2006), Molina Colón et al. (2011) y Ramjohn et al. (2012).
En el capítulo 3 se mencionaron varios trabajos sobre la estequiometría de bosques secos en Puerto Rico. Por ejemplo, Medina et al. (2017) encontraron que hojas maduras de árboles en bosques secos exhiben altos valores del cociente nitrógeno:fósforo, sugiriendo limitaciones de fósforo y posiblemente hierro en suelos calizos con altos pH. Bajo esas condiciones, las plantas acumulan calcio y aluminio. Altos valores del cociente C13:C14 en el bosque seco sugiere altos niveles de eficiencia en el uso
7 138 CAP ariel e. lugo | ecología a m i m anera Bosques secos
del agua. Comparando la estequiometría de la hojarasca en el suelo de bosques secos noveles con la hojarasca promedio de todos los bosques de Puerto Rico, Lugo y Erickson (2017) encontraron que los bosques noveles tenían menores concentraciones de manganeso y sodio y cocientes más bajos de carbono:nitrógeno y carbono:fósforo. Además, comparados con bosques secos nativos, los bosques noveles tenían anomalías de carbono más altas, anomalías de concentración de calcio y sodio más bajas, y anomalías más bajas de carbono:nitrógeno. Estas características tienen influencia en la estructura y funcionamiento de estos bosques.
Asignación
Prepara una lista de adaptaciones de los bosques secos a: sequías, poco fósforo, huracanes, fuego, el ser humano.
7 139 CAP ariel e. lugo | ecología a m i m anera Bosques secos
CaPítulo 8 Bosques húmedos, muy húmedos y pluviales
Pregunta
¿Cuál es la zona de vida más extensa de Puerto Rico y qué implicaciones tiene para la ecología de bosques?
Introducción
Los bosques de la zona de vida subtropical húmeda fueron casi todos modificados dramáticamente por la actividad agrícola (Lugo 2005a). La especie Dacryodes excelsa otabonuco en un momento se extendía por tres zonas de vida, desde la subtropical húmeda hasta la subtropical muy húmeda montano bajo (Lugo y Wadsworth 1990). Los mejores ejemplos de estos bosques se conservan en la zona de vida subtropical muy húmeda, aunque aún hoy hay rodales de tabonuco en la zona de vida subtropical húmeda (Álvarez Ruiz y Lugo 2012). Los bosques de tabonuco son los bosques nativos con el mayor desarrollo estructural y las mejores posibilidades de producir madera para aserrar en abundancia. Una gran proporción de los estudios ecológicos sobre los bosques de la Isla se han llevado a cabo en la zona de vida subtropical muy húmeda, específicamente sobre los bosques de tabonuco (Odum y Pigeon 1970, Basnet et al 1993, Drew et al. 2009).
En este capítulo centro la atención en los atributos estructurales y funcionales de los bosques de tabonuco en las Montañas de Luquillo (Lugo y Scatena 1995) al final de la década de 1980, cuando estos llegaron a su máxima madurez antes del paso del huracán Hugo. Luego, analizo algunos efectos del huracán sobre estos bosques y cómo este evento cambió la visión incompleta que tenía sobre la dinámica de estos sistemas. Finalmente, comparo los bosques noveles de tabonuco a lo largo de un gradiente climático, desde la zona de vida subtropical húmeda, hasta la zona de vida subtropical muy húmeda montano bajo (Álvarez Ruiz y Lugo 2012). Antes de comenzar, resumo someramente algunos resultados del inventario de bosques de Puerto Rico y las Islas Vírgenes donde se presentan datos para bosques subtropicales húmedos, muy húmedos, pluviales, y montano bajo muy húmedos y pluviales. En los capítulos 8 y 11 presento más datos ecológicos para otros bosques de las
ariel e. lugo | ecología a mi ma nera
141
Tabla 8.1
Algunos parámetros geográficos y ecológicos de los bosques subtropicales húmedos, muy húmedos y pluviales de Puerto Rico (Brandeis y Turner 2013a) e Islas Vírgenes de los Estados Unidos de América (Brandeis y Turner 2013b). Es diámetro de los árboles a la altura del pecho es dap. Los datos corresponden al 2009. Las unidades están en paréntesis. El área total de bosques en Puerto Rico era de 472,791 ha y en las Islas Vírgenes 18,277 ha.
montañas de Puerto Rico (subtropical muy húmedo, pluvial y montano bajo muy húmedo y pluvial).
Resultados del inventario de bosques
Entre todos los tipos de bosques existentes en Puerto Rico, los subtropicales húmedos cubren el área más extensa (Tabla 8.1). En las Islas Vírgenes, los bosques subtropicales húmedos cubren una proporción pequeña del área total de bosques. Los bosques pluviales cubren poca área en Puerto Rico, mientras que los bosques muy húmedos les siguen a los húmedos en abundancia. Los datos estructurales en la tabla 8.1 reflejan los efectos antropogénicos sobre los bosques de Puerto Rico, y en particular sobre los bosques húmedos. Los bosques montano bajo muy húmedos y pluviales, reflejan más estructura que los húmedos porque son bosques nativos, maduros y sin efectos antropogénicos visibles. Los datos de los bosques húmedos reflejan rodales jóvenes en sucesión y por eso aparentan ser menos desarrollados estructuralmente. La tabla 8.1 también ilustra la diferencia entre la estructura de bosques húmedos en Puerto Rico vs. los bosques de las Islas Vírgenes, cuyos bosques exhiben menos biomasa y volumen y crecimiento de árboles más lentos que en los de Puerto Rico.
8 142 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
Parámetro Puerto Rico^ Islas Vírgenes Húmedo Muy húmedo y pluvial Montano bajo muy húmedo y pluvial Húmedo Área (ha) 234,233 161,124 8,770 6,582 Biomasa aérea (Mg/ha) 85 94 134 77.4* Biomasa subterránea (Mg/ha) 20 21 25 15.9* Carbono total (Mg/ha) 50 58 80 45.8* Crecimiento en volumen (m3/ha·año) 4.2 4.6 3.7 2.0^ Mortalidad (m3/ha·año) 1.7 2.5 1.4 0.4^ Densidad (tallos/ha) 403 474 719 4,908* Área basal (m2/ha) 10.8 17.8 23.7 17.5* Volumen (m3/ha) 73.2 103.4 99.2 32.3^ ^ dap > 5.0 cm * dap > 2.5 cm
Variación edáfica y de biomasa en bosques aparentemente
homogéneos
La figura 8.1 ilustra la variación espacial en la biomasa del bosque de tabonuco en seis rodales ubicados dentro de un área relativamente pequeña en el sector El Verde del Bosque Experimental de Luquillo. Noten que la biomasa varía por un factor de cuatro. Lo mismo pasa con la acumulación de hojarasca en el suelo, que puede variar por un factor de casi 30 veces y con obvias diferencias dependiendo de la posición topográfica (Fig. 8.2). Las concentraciones de nitrógeno (Fig. 8.3), calcio (Fig. 8.4) y densidad aparente del suelo (Fig. 8.5) también varían considerablemente en estos bosques de tabonuco. El significado de estos resultados no es solamente su relevancia a la variación estructural y edáfica dentro de un bosque homogéneo de tabonuco. La variación estructural y edáfica es intrínseca en todos los sistemas ecológicos y se revela aquí por las medidas cuantitativas de campo como parte de investigaciones científicas.
8 143 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
Figura 8.1. Variación espacial en la cantidad y distribución de biomasa en ocho rodales de bosques de tabonuco en El Verde, Bosque Experimental de Luquillo. Los datos originales son de Ovington y Olson (1970) y la síntesis de Lugo y Scatena (1995).
Figura 8.2. Frecuencia de la distribución de la masa de hojarasca en el bosque de tabonuco en Bisley, Bosque Experimental de Luquillo (Lugo y Scatena 1995).
Figura 8.3. Frecuencia de la distribución de la concentración de nitrógeno en los suelos del bosque de tabonuco en Bisley, Bosque Experimental de Luquillo. Los datos se tomaron al azar y representan la capa superior del suelo (0-10 cm). Más detalles en Lugo y Scatena 1995.
Disturbios y mortalidad arbórea
Figura 8.4. Frecuencia de la distribución de la concentración de calcio en los suelos del bosque de tabonuco en Bisley, Bosque Experimental de Luquillo. Los datos se tomaron al azar y representan la capa superior del suelo (0-10 cm). Más detalles en Lugo y Scatena 1995.
La figura 8.6 ilustra la intensidad de lluvia y la frecuencia de recurrencia de eventos de lluvia asociados a distintos tipos de disturbios. Para las distintas intensidades de eventos se ilustran algunas respuestas en el paisaje. En 100 años, los huracanes han afectado el 100 % del área del bosque en las Montañas de Luquillo.
8 144 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
Figura 8.5. Frecuencia de la distribución de la densidad aparente de los suelos del bosque de tabonuco en Bisley, Bosque Experimental de Luquillo. Los datos se tomaron al azar y representan la capa superior del suelo (0-10 cm). Más detalles en Lugo y Scatena 1995.
Figura 8.6. El patrón de retorno de lluvias de distintas intensidades en Bisley, Bosque Experimental de Luquillo. Las intensidades están asociadas a los eventos identificados con los años y las condiciones meteorológicas en el margen derecho. En el margen izquierdo, se ilustran las respuestas a las lluvias de parte del paisaje en término de la respuesta de árboles y procesos geológicos e hidrológicos (Lugo y Scatena 1995).
Los derrumbes afectaron el 0.08 % por centuria a elevaciones entre 530 y 850 m en zonas volcanoclásticas y 0.20 % por centuria a las mismas elevaciones pero en geología intrusiva. Estos datos ayudan a explicar el efecto de los disturbios sobre la geomorfología del paisaje, el cual depende de la frecuencia, intensidad del disturbio y la topografía del área afectada. Eventos intensos, pero con poca extensión geográfica, ej., los derrumbes, tienen efectos notables en pequeñas escalas dependiendo de
8 145 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
la geología donde ocurren. Otros, como los huracanes, pueden ser intensos y afectar extensas regiones.
La mortalidad de árboles es un proceso que responde a los disturbios y, dependiendo de la longevidad de las especies, a la edad de los árboles. La mortalidad base (background mortality) varía entre 0.5 y 2.6 % por año, lo que implica que en 100 años afecta entre 50 y 260 % de la población. El porcentaje de mortalidad asociado a disturbios es sensible al área bajo consideración, y la intensidad y duración del disturbio. Mortalidades anuales sobre el 5 % reflejan los efectos de disturbios. En un estudio en el sector Bisley del Bosque Experimental de Luquillo, el 30 % de los árboles murieron durante las cuatro horas que los vientos del huracán Hugo afectaron al sector. Cuando se conoce la causa y se mide el efecto en intervalos de tiempo corto se denomina el proceso como mortalidades instantáneas. Sin embargo, es posible que un árbol se considere muerto en un momento y aparezca vivo meses o años después. Igualmente, hay árboles que sobreviven un disturbio y mueren meses o años después. En esos casos se describe la mortalidad retrasada (delayed mortality). Es posible que la mortalidad retrasada asociada a un evento sea mayor a la mortalidad instantánea después del evento. ¿Puedes explicar por qué?
Surgen dos paradojas de la información disponible sobre eventos extremos y la respuesta de los bosques de tabonuco, de hecho, de todos los bosques de Puerto Rico. Primero, ¿cómo se explica la presencia de árboles centenarios (sobre 500 años) en estos bosques si los huracanes y la mortalidad natural afectan sobre el 100 % del área de bosque cada 100 años? Segundo, ¿cómo se explica la resurrección de árboles considerados muertos después de un disturbio? La explicación de la primera paradoja está en la distribución geográfica de la intensidad de un evento extremo. La misma no es uniforme, lo que ofrece a los organismos la posibilidad de encontrar refugios. Además, hay regiones topográficas que son menos vulnerables a los efectos de los disturbios, al igual que biológicamente hay especies más resistentes a los disturbios que otras. La segunda paradoja tiene una explicación biológica. Muchas especies arbóreas en el Caribe y Puerto Rico se han adaptado al efecto de los vientos y otros disturbios por medio de su capacidad de rebrote y el almacenaje de nutrientes y materia orgánica en raíces subterráneas que sobreviven los vientos y sostienen los rebrotes. Los tabonucos en particular aparentan desafiar el viento dominando la parte alta (cima o tope) de la catena donde los vientos son más fuertes. Es allí donde crecen tabonucos centenarios a pesar de altas frecuencias de vientos huracanados. La solución a la paradoja se discute a continuación.
Uniones de tabonuco
Los árboles de tabonuco forman uniones (Basnet et al. 1993) interconectando sus raíces, lo que les permite intercambiar materia orgánica y nutrientes entre los
8 146 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
miembros de la unión (Fig. 8.7). La unión incluye árboles de todos los tamaños y su extensión geográfica cubre todo el tope de la catena. Los tabonucos forman uniones porque tienen la capacidad de injertar raíces entre individuos. Estas uniones de tabonuco no son clones de árboles ya que los individuos se reproducen independientemente y se interconectan posteriormente. El funcionamiento de las uniones de tabonuco se resume en el modelo de la figura 8.8. La unión concentra energía y nutrientes, y mantiene la dominancia de los tabonucos con poca oportunidad para que otras especies entren al lugar donde está la unión. La estructura y funcionamiento de la unión protege los tabonucos contra los vientos de los huracanes y es un componente importante de la resiliencia a huracanes de estos bosques (Lugo et al. 2002). Por ejemplo, tocones de árboles cortados en la década de 1940 y marcados muertos en numerosos inventarios del rodal, produjeron nuevas ramas y hojas después que abrió el dosel ante los vientos del huracán Hugo. La transferencia de materia orgánica y energía de los árboles vivos en la unión claramente fueron responsable de este comportamiento.
8 147 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
Figura 8.7. Mapa de un rodal o unión de tabonuco en Bisley, Bosque Experimental de Luquillo, enfatizando las conexiones entre los árboles por medio de los injertos entre las raíces (Basnet et al. 1993).
Sucesión
La figura 8.9, resume la respuesta a un huracán de la estructura y diversidad de especies arbóreas de un bosque de tabonuco. Durante el intervalo de tiempo señalado, el huracán Hugo pasó al noreste de la ubicación del rodal, el cual estaba protegido por la topografía de las montañas, por lo que sus efectos fueron mínimos en comparación al efecto sobre rodales expuestos a los vientos (Lugo et al. 2017). En el 1932 el huracán San Cipriano había afectado los bosques de esta región, por lo que los datos en la figura 8.9 ilustran la sucesión ininterrumpida de un bosque de tabonuco después del huracán antes mencionado. El efecto del huracán Hugo se refleja en un aumento en densidad de árboles en el 1989 (Fig. 8.9a). En ausencia de huracanes, los rodales tienden a reducir la densidad de los árboles a medida que estos crecen y ocupan más espacio. La figura 8.9b refleja el crecimiento en el área basal de los árboles entre huracanes, con una pequeña interrupción en 1989 por el huracán Hugo. Los cambios en densidad y área basal dieron como resultado un aumento en la biomasa aérea del rodal sin aparente efecto del huracán Hugo. La diversidad y equitabilidad (evenness) de las especies disminuyó con el tiempo, pero ambas aumentaron ligeramente cuando el huracán Hugo proporcionó la entrada de especies al rodal. Los huracanes aumentan la riqueza de especies en rodales
8 148 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
Figura 8.8. Modelo del funcionamiento de una unión de tabonuco propuesto por Lugo y Scatena (1995). Se enfatiza la función de concentración de recursos y la protección ante eventos climáticos extremos.
arbóreos al permitir la entrada de especies de crecimiento rápido o pioneras en la sucesión del bosque.
En las Montañas de Luquillo el número de etapas de sucesión entre la etapa con vegetación pionera y la etapa con vegetación madura disminuye con la elevación (Fig. 8.10). A elevaciones altas, la sucesión se torna en autosucesión, donde las mismas especies ocurren en las etapas pioneras y maduras según ocurren en los manglares (Lugo 1980). A medida que las condiciones ambientales se tornan más exigentes, como ocurre con el aumento en elevación en las montañas, menos especies tienen la capacidad de sobrevivir y se autorreemplazan después de un disturbio. No así con los bosques de tabonuco donde hay presencia de especies pioneras como el yagrumo (Cecropia schreberiana) o el roble blanco (Tabebuia heterophylla), especializadas para aumentar en abundancia luego del disturbio. La palma de sierra (Prestoea
) se comporta como el mangle rojo (Rhizophora mangle) en términos de su comportamiento como pionera y especie de bosque maduro. Estas dos especies forman humedales arbóreos en la montaña y la costa, respectivamente.
8 149 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
montana
Figura 8.9. Sucesión ininterrumpida en el bosque de tabonuco tras el paso de un huracán en 1932. Este rodal, ubicado en El Verde, Bosque Experimental de Luquillo, fue levemente afectado por el huracán Hugo en 1989 por su exposición de sotavento hacia el huracán. El cambio en el patrón de los índices de diversidad y uniformidad de especies reflejan el paso de Hugo. Los datos son del Instituto Internacional de Dasonomía Tropical y la síntesis de Harris y Lugo (2012).
El huracán Hugo causó un desplazamiento de las comunidades arbóreas en el espacio ecológico, como se observa en la figura 8.11. Quince años después del huracán, comunidades con combinaciones de abundancia de especies similares a las presentes antes del huracán, comenzaron a aparecer, pero es obvio que rodales con combinación de especies típicas de 1988, no retornan, a pesar de que hubo poco cambio en el número y tipo de especies en los rodales. Lo que cambia es la proporción de las especies medida por sus respectivos Valores de Importancia. Cuando los huracanes Irma y María azotaron el bosque 27 años después de Hugo, los rodales ya se habían alejado bastante de su condición inicial en 1988.
Dominancia de especies
La dominancia de especies, medida en términos del Valor de Importancia de la especie más importante de un rodal, es alta en los bosques caribeños en comparación con los bosques continentales. Le he atribuido esa diferencia al efecto de los huracanes en el Caribe (Lugo 1991). Mi razonamiento es que los huracanes ejercen una fuerte selección sobre las especies y pocas logran adaptarse a sus frecuentes disturbios. Aquellas que logran hacerlo ejercen dominancia sobre otras especies en
8 150 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
Figura 8.10. Modelos de sucesión a lo largo de un gradiente de elevación en el Bosque Experimental de Luquillo (Lugo y Scatena 1995). El número de comunidades serales disminuye con la elevación. Las flechas indican la dirección de la sucesión. Las líneas verticales entrecortadas representan cambios significativos en la estructura y composición de especies en las comunidades. La palma de sierra funciona como especie de sucesión y como especie de comunidad estable. El diagrama está basado en varios estudios citados en Lugo y Scatena 1995.
proporción a la efectividad de sus adaptaciones y otras características ecofisiológicas. La dominancia de especies y la forma de las curvas de Valor de Importancia cambian con la elevación en El Yunque (Fig. 8.12). La dominancia aumenta y el número de especies disminuye con la elevación. Los bosques de tabonuco ubicados en elevaciones bajas tienen menos dominancia y más especies de árboles comparados con los bosques elfinos en la cima de las montañas.
No importa la forma de vida de las plantas del bosque de tabonuco, todos los grupos de especies exhiben alta dominancia (Fig. 8.13). La diversidad de formas de vida y la abundancia de especies en estos grupos en la figura 8.13 es notable y apunta a una alta diversidad de funciones ecológicas en estos bosques.
Flujo y almacenaje de masa y nutrientes
El flujo de masa y nutrientes en bosques varía con el tipo de bosque, su edad y estado de sucesión, incluyendo los efectos de disturbios. Como los nutrientes generalmente fluyen en proporción a la masa, particularmente durante la caída de hojarasca, podemos ver estas relaciones en los datos de caída de hojarasca en tres tipos de bosques, todos en la zona de vida subtropical muy húmeda en el Bosque Experimental de Luquillo (Fig. 8.14). Los datos corresponden a un periodo de tiempo sin la influencia de un huracán y reflejan las tasas altas en las plantaciones,
8 151 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
Figura 8.11. Trayectoria de la sucesión en el bosque de tabonuco en Bisley, Bosque Experimental de Luquillo, entre 1988 y 2018 pos huracán María. La trayectoria de la sucesión está basada en inventarios periódicos del bosque por el Instituto Internacional de Dasonomía Tropical y un análisis de ordenación de la composición de especies por la Dra. Tamara Heartsill Scalley utilizando el método escalado multidimensional no métrico (NMS, por sus siglas en inglés). El análisis utilizó datos de ocho inventarios para tallos con diámetro a la altura del pecho > 2.5 cm en 85 parcelas que cubrían 0.7 ha.
8 152 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
Figura 8.12. Rango de las especies arbóreas en tres tipos de bosque en el Bosque Experimental de Luquillo basado en el Valor de Importancia de la especie (Lugo y Scatena 1995).
Figura 8.13. Rango de las especies pertenecientes a distintas formas de vida en el bosque de tabonuco en El Verde, Bosque Experimental de Luquillo, basado en el Valor de Importancia de la especie (Lugo y Scatena 1995).
seguida por los bosques secundarios y finalmente el bosque maduro de tabonuco. Solamente cuando las plantaciones o bosques secundarios son muy jóvenes su caída de hojarasca es menor al bosque maduro. Mientras las plantaciones y bosques secundarios varían anualmente en la caída de hojarasca, el bosque maduro mantiene tasas muy constantes reflejando un alto grado de homeostasis resultado de su madurez.
El movimiento de la biomasa y nutrientes en la caída de hojarasca es uno de muchos flujos que sostienen el funcionamiento de los bosques. La figura 8.15 compara flujos de fósforo y nitrógeno bióticos y abióticos para el bosque de tabonuco en las montañas de Luquillo. La lluvia que cae sobre el bosque es un insumo de estos elementos esenciales para el funcionamiento del sistema y se observa que si el volcán en Montserrat está activo, la lluvia en Puerto Rico transporta más nitrógeno y fósforo al bosque de tabonuco. Más aún, la lluvia se enriquece de nutrientes al pasar por el dosel (transcolación) y caer al suelo y las aguas de escorrentía cargan cantidades más altas que el insumo de estos elementos en la lluvia. En el bosque de tabonuco la salida de nutrientes por vía acuática es superior a la entrada de estos en la lluvia, reflejando la erosión de las rocas volcanoclásticas. En la zona de rocas intrusivas la discrepancia entre la entrada y salida de nutrientes es mayor. Así, los bosques contribuyen a los ciclos de elementos de ríos, quebradas y estuarios aguas abajo. Los datos de retranslocación significan el reciclaje de nutrientes
8 153 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
Figura 8.14. Contraste en las tasas de caída de hojarasca a través del tiempo en bosques secundarios, bosque de tabonuco maduro y plantaciones de distintas edades en el Bosque Experimental de Luquillo. Las fuentes de datos están contenidas en Lugo y Scatena (1995) donde apareció esta figura.
dentro de los árboles antes de que las hojas caigan con la caída de hojarasca. Es un flujo de conservación de nutrientes. Los flujos asociados a los coquíes son particularmente notables por su magnitud, en comparación con el flujo de otros componentes del bosque.
La biomasa de distintos componentes del bosque de tabonuco (excluyendo los árboles) reafirma la importancia de las epifitas y las lombrices de tierra (Fig. 8.16). Sin embargo, a pesar de su menor biomasa en comparación con las epifitas, los hongos y bacterias almacenan más nitrógeno y fósforo que las epifitas (Fig. 8.17). Estos
8 154 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
Figura 8.15. Flujos de nitrógeno y fósforo asociados a distintos procesos ecológicos en el Bosque Experimental de Luquillo. La producción primaria neta aérea es NPP. Referencias de los trabajos utilizados están en Lugo et al (2012d), donde apareció esta figura.
Figura 8.16. Biomasa de una variedad de organismos no arbóreos en el Bosque Experimental de Luquillo. Referencias de los trabajos utilizados están en Lugo y Harris (2012), donde apareció esta figura. La biomasa de lombrices de tierra no está incluida en la barra de «todos los animales» pues proceden de estudios distintos. Las lombrices de tierra contienen la biomasa animal más alta en el bosque.
Figura 8.17. Almacenaje de nitrógeno y fósforo en grupos de animales, hongos y tipos de plantas no arbóreas en el bosque de tabonuco y un palmar en valle inundable (las briofitas) en el Bosque Experimental de Luquillo. Las fuentes de información están en Lugo et al. (2012d) donde apareció esta figura.
8 155 CAP ariel e.
lugo | ecología a m i m anera
Bosques húmedos, muy húmedos y pluviales
datos reflejan las distintas funciones de los distintos organismos del bosque, ya sea almacenando o circulando masa y nutrientes.
La catena
El diagrama de la catena asociada a los bosques de tabonuco (Fig. 8.18), pero presente en todos los bosques de Puerto Rico, resalta los gradientes en estas catenas discutidas anteriormente. La posición dominante de las uniones de tabonuco y la interacción con las nubes son particularmente notables. La alta intercepción de lluvia del bosque de tabonuco (puede ser > 50 %) también se ilustra con la recirculación de materia orgánica y nutrientes en el dosel. En los doseles de los bosques muy húmedos y pluviales de Puerto Rico se acumulan suelos orgánicos producto de la actividad fotosintética de las plantas del dosel, donde se sostiene una alta diversidad de organismos, tanto plantas como animales y microbios. Los flujos contra la gravedad de los ríos y quebradas a la zona riparia que se ilustran incluyen la absorción de nutrientes por las plantas y la actividad de los animales anfibios o terrestres que mueven materiales del río o quebrada al sistema terrestre.
8 156 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
Figura 8.18. Esquema del flujo de materiales en una catena con una unión de tabonucos en la cima en el Bosque Experimental de Luquillo (Lugo y Scatena 1995). Las líneas y flechas indican la dirección y ruta de los intercambios de materiales, enfatizando las áreas de interfaz con la atmósfera, el suelo y los sistemas acuáticos. Los lugares con mayor reciclaje ocurren en el dosel del bosque, la hojarasca y la vegetación.
Áreas de estudio
Elevación entre 200-900 m y
20 km
de
de
subtropical húmeda. Distribución histórica (basado en la literatura) Municipios
Bosques de tabonuco remanentes y noveles
La distribución original de los bosques de tabonuco cubría casi toda la cordillera central de Puerto Rico (Fig. 8.19). La puertorriqueña Migdalia Álvarez Ruiz (2002) estudió rodales remanentes y encontró que se separaban a lo largo de gradientes de precipitación y elevación de acuerdo con la composición de especies (Fig. 8.20). Otros rodales quedaban fuera del gradiente de precipitación y elevación. Al estudiar la composición de especies de los rodales de tabonuco, se notaron gradientes en la presencia de especies como la pomarrosa (Syzygium jambos), cacao motillo (Sloanea berteriana) y el ausubo (Manilkara bidentata). El ausubo y cacao motillo eran más abundantes en los rodales de tabonuco en áreas protegidas con mucha presencia del tabonuco, pero no se encontraban en áreas bajo la influencia agrícola, particularmente el tabaco. En las áreas afectadas por la agricultura el tabonuco se encontraba en densidades menores y la pomarrosa dominaba muchos rodales. Estas distribuciones de especies reflejaban la naturaleza novel de los bosques remanentes de tabonuco. Álvarez Ruiz y Lugo (2012) formularon un modelo de la transformación y sucesión de estos bosques (Fig. 8.21) partiendo de bosques nativos sin perturbaciones antropogénicas. El paisaje donde se encontraban estos

8 157 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
Figura 8.19. Mapa con la distribución del tabonuco de acuerdo con la literatura y de la zona de vida y elevación que favorecen el crecimiento del tabonuco. Los puntos representan los lugares de estudio donde Álvarez Ruiz y Lugo (2012) analizaron la composición de especies y la estructura de los rodales de tabonuco.
zona
vida
Holdridge
rodales y los usos del ser humano a estos paisajes, fueron determinantes en la composición de especies de los bosques de tabonuco que sobrevivieron la deforestación. Subsiguientemente, la puertorriqueña Sandra Ayala, de la Universidad del Turabo, encontró evidencia en el Cerro la Tiza de Comerío, de la restauración por sucesión del tabonuco en áreas dominadas anteriormente por la pomarrosa. Este estudio es el primero que documenta la restauración de un bosque nativo donde había prevalecido un bosque novel.
8 158 CAP ariel e. lugo | ecología a m i m anera Bosques húmedos, muy húmedos y pluviales
Figura 8.20. Ordenación de las especies arbóreas en rodales de tabonuco estudiados por Álvarez Ruiz y Lugo (2012). La mayor parte de las especies se ordenaron en función de la elevación y precipitación, pero los rodales del Barrio Sumidero y el Cerro Peña, por los efectos de usos agrícolas, resultaron ser bosques noveles por la abundancia de especies no nativas como Syzygium jambos (pomarrosa).
Figura 8.21. Diagrama que ilustra los efectos de la actividad agrícola, la deforestación y otros disturbios sobre los bosques de tabonuco en Puerto Rico. En ausencia de la actividad humana, los rodales de tabonuco en regiones boscosas se mantienen como tabonuco por medio de una sucesión cíclica dominada por especies nativas. La tala selectiva también sostiene los rodales de tabonuco y permite que desarrollen su aspecto maduro. No así con las intervenciones agrícolas como el café y el tabaco, que permiten la colonización de especies que normalmente no son parte del bosque de tabonuco maduro. El resultado es bosques noveles con tabonuco los cuales, con pocas excepciones, no revierten (retorno en la figura) a las condiciones originales (modificado de Álvarez Ruiz y Lugo 2012).
Asignación
Compara un atributo estructural y uno funcional de los bosques de tabonuco con los mismos atributos de otros bosques que no sean bosques secos o humedales.
8 159 CAP ariel e. lugo | ecología a m i m
y pluviales
anera Bosques húmedos, muy húmedos
CaPítulo 9
Humedales arbóreos
Pregunta
¿Qué es un humedal?
Introducción
El estudio de humedales ha evolucionado a lo largo del tiempo y en 1984 lo resumí en las siguientes etapas (Lugo 1984): 1) las primeras observaciones entre 325 a. de C. y 136 d. de C., 2) primeras descripciones (123 –1731), 3) formalización de las descripciones comenzando en 1753, 4) estudios ecológicos durante el siglo XX, y 5)estudios de los atributos funcionales comenzando en la década de 1930. En la década de 1970 se iniciaron los estudios comparativos y experimentales en la Florida bajo la dirección de Odum (Ewel y Odum 1984). En el ámbito global, los estudios de los manglares, humedales estuarinos y de agua salada, generalmente estaban más avanzados, aproximadamente por una década, que los estudios en los humedales de agua dulce. Lo mismo ocurrió en Puerto Rico, donde los manglares llamaron la atención de los ecólogos antes que los humedales de agua dulce.
Definición
Un humedal es un sistema ecológico inundado o con suelos saturados por aguas superficiales o subterráneas a tal frecuencia y duración que organismos adaptados a la pobre aeración crecen y se reproducen en esos lugares.
La definición, modificada de Lugo (1990), se basa en dos criterios: sustrato saturado o con pobre aeración y la respuesta biótica a estas condiciones (Tabla 9.1). Es posible definir el humedal, ya sea por el hidroperiodo si es obvio, como en los manglares o los pantanos donde la inundación es fácil de observar, o por la composición de especies de la vegetación o los animales cuando la presencia de la inundación es invisible a la vista. Por ejemplo, a veces caminando por un bosque terrestre, de pronto uno se encuentra con un sotobosque desprovisto de
ariel e. lugo | ecología a mi ma nera
161
Tabla 9.1
Características comunes en los humedales (Lugo 1990).
Característica
Presencia de palmeras
Esclerofilia*
Estructuras para intercambio gaseoso: lenticelas, neumatóforos, raíces aéreas, hinchamiento de la base de los tallos
Zonación de vegetación
Doseles parejos
Raíces tabulares o de zanco
Raíces superficiales o aéreas
En ambientes húmedos: baja en la riqueza de especies relativas a sistemas terrestres cercanos
En sistemas áridos: más riqueza, abundancia y área basal de especies relativas a sistemas terrestres cercanos
Explicación
Tienen efectivos sistemas de aeración por ser monocotiledóneas
Por limitación de nutrientes o agua
Transportan oxígeno y bióxido de carbono de la atmósfera a las raíces y viceversa
Reflejan gradientes ambientales como el hidroperiodo o la salinidad
Reflejan edad pareja de las plantas, dominancia de especies
Proveen estabilidad en pendientes o suelos inestables
Relacionadas con pobre aeración y limitación de nutrientes
Hidroperiodo es más largo
Aumento en la humedad del suelo
*Desarrollo excepcional de esclerénquima en las hojas, dando como resultado un follaje endurecido y espeso resistente a la pérdida de agua.
vegetación y quizás aglomeraciones de especies de humedales, creciendo donde no se esperaban. Al caminar sobre el lugar, se puede sentir que el suelo está saturado. Cuando el suelo es aplastado, cede ante el paso de una persona y el agua se desplaza ante la presión.
El régimen hidrológico o hidroperiodo es el factor principal que determina la presencia de un humedal. El hidroperiodo se mide por la profundidad del agua de inundación o el grado de saturación del suelo según ambos varían a través del tiempo. La frecuencia y la duración del periodo de inundación o saturación representan el factor tiempo dentro de la definición de un humedal. La frecuencia del periodo de inundación o saturación varían entre horas y milenios, como ocurre en humedales en los desiertos. La duración del periodo de inundación o saturación es igualmente variable.
Tanto la definición como la clasificación de humedales son conflictivas, ya que distintas clasificaciones utilizan criterios distintos para identificar y clasificar los humedales. La clasificación de Cowardin et al. (1979) es la clasificación de humedales prevaleciente en los EE. UU. y la utilizada por las agencias del gobierno federal, incluyendo Puerto Rico. La estructura de la clasificación es compleja (Tabla 9.2). Esta clasificación fue desarrollada por un grupo de científicos norteamericanos especializados en humedales que formaron un grupo de trabajo y llevaron a cabo talleres con el propósito único de desarrollar una clasificación uniforme para los EE. UU.
El geocientífico australiano Bruce Graham Thom, sugirió que la distribución, composición y estructura de los humedales (en su caso refiriéndose a los manglares)
9 162 CAP ariel e. lugo | ecología a m i m anera Humedales arbóreos
Tabla 9.2
La clasificación de humedales de Cowardin et al. (1979). Esta clasificación está basada en forma de vida, tipo de suelo, régimen de agua y calidad de agua.
Seis niveles principales
Diez provincias regionales
Cinco sistemas ecológicos: marinos, estuarinos, ribereños, lacustrinos, palustrinos
Diez subsistemas ecológicos
De 8 a 18 clases basadas en la vegetación (arbórea, arbustal, sumergida, etc.)
De 12 a 17 subclases
De 8 a 25 órdenes
se entendían mejor en el contexto de las condiciones de sedimentación y la geomorfología de las costas (Thom 1975). Independientemente, Lugo y Snedaker (1974) clasificaron los tipos de manglares al nivel de rodales utilizando las condiciones fisiográficas (relieve y relación con el movimiento de las aguas) en las cuales se encuentran (próximo capítulo). Ambos enfoques, el geomorfológico a gran escala y el fisiográfico a escalas menores, los combinó el norteamericano Patrick C. Kangas (1990) utilizando el concepto de la huella energética de Odum (próximo tópico).
La energía y los tipos funcionales de humedales
Kangas se basó en la noción de que el tipo y potencia de la energía incidente en un sistema determinan las características de los ecosistemas. Ilustró cómo la dirección, concentración y potencia hidrológica forman distintos tipos de humedales (Figs. 9.1 y 9.2). La figura 9.1 identifica cuatro tipos de distribución espacial de la energía:
9 163 CAP ariel e. lugo | ecología a mi ma nera Humedales arbóreos
Figura 9.1. Representación esquemática de cuatro tipos básicos de la distribución espacial de la energía cinética sobre el terreno. La flecha indica la dirección hacia donde se dirige la energía potencial (modificado de Kangas 1990).
Figura 9.2. (A) Forma, orientación y tamaño relativo de las formas de relieve. (B) Patrones de la forma de ecosistemas cuando se combina el efecto de la distribución espacial de la energía (Fig. 9.1) con las formas, orientación y tamaño relativo de las formas de relieve (A) (modificado de Kangas 1990). Los patrones emergentes en (B) son estructuras (humedales) que se adaptan a, interactúan con y amplifican la energía espacial en sus entornos.
9 164 CAP ariel e. lugo | ecología a m i m anera Humedales arbóreos
el manto, el punto, el frente y la línea. El manto y frentes son energías diluidas espacialmente, mientras que el punto y la línea son energías concentradas. El manto y el punto inciden perpendicularmente al sistema y el frente y línea inciden paralelamente al sistema. El frente es análogo al frente meteorológico que se desplaza paralelamente con el sistema y puede estar estacionario (un gradiente). Ejemplos son escorrentías, mareas, y masas de aire. Un río es un ejemplo de una línea. Un manantial artesiano es un ejemplo de un punto, otro ejemplo de un punto sería un rayo. La luz del sol y la lluvia son ejemplos de mantos, como lo es una formación rocosa uniforme emergiendo por fuerzas tectónicas. La figura 9.2 contiene ejemplos de formas de ecosistemas en respuesta a la energía que converge sobre el sistema con el ejemplo del movimiento y tipo de energía.
El concepto de la huella energética o energy signature (Fig. 9.3)
la suma de todos los tipos de energía que inciden en un ecosistema y su patrón de entrada se conoce como la huella energética del sistema. Este concepto lo desarrolló Odum (1983). La idea ayuda a entender a qué fuerzas le responde el ecosistema, como se ve en la figura 9.3 en el caso de un pantano costero que recibe efluentes calientes. Al corregir por la calidad de la energía, la importancia de las mareas y las olas aumenta relativo a su valor expresado en calorías.
La diversidad de humedales en Puerto Rico
Los humedales en Puerto Rico son diversos (Lugo y Brown 1988) e incluyen humedales marinos, estuarinos, terrestres, arbóreos, herbáceos, sin vegetación emergente, en tierras bajas, en las montañas, micro humedales y en la zona de chapoteo de cataratas (lacrimitic wetlands). De los trabajos tradicionales de la vegetación del Caribe, Lugo y Brown (1988) encontraron sobre 50 asociaciones vegetales identificadas como humedales por los autores.
El puertorriqueño Manuel del Llano (1988) preparó el primer mapa de los humedales de Puerto Rico (Fig. 9.4) utilizando el sistema de Cowardin et al. (1979). La abundancia de humedales en el país sorprendió a muchos que no habían considerado que la saturación de los suelos en las montañas creaba condiciones de humedal con humedales arbóreos capaces de funcionar en suelos saturados pobres en oxígeno. El argentino Jorge Luis Frangi (1983) fue el primero en reconocer como humedales a los bosques sobre laderas y valles de las montañas expuestas a altas precipitaciones y con suelos arcillosos que retienen agua (Fig. 9.5). El estudio de Silver et al. (1999) confirmó la propuesta de Frangi, ya que al medir la concentración de oxígeno en los suelos en distintos tipos de bosque, encontraron un gradiente en la concentración del oxígeno en los suelos de El Yunque (Fig. 9.6). El bosque de tabonuco tenía concentraciones similares a las concentraciones atmosféricas, pero sobre 600 m de
9 165 CAP ariel e. lugo | ecología a m i m anera Humedales arbóreos
Figura
energética de un humedal costero (Odum 1983). ( A ) Diagrama de los flujos de energía al humedal recibiendo aguas calientes de una generadora nuclear en la Florida. ( B ) Cálculo de los insumos energéticos al humedal en equivalentes de calor. (C) Los mismos insumos de energía, corregidos por su calidad energética y expresados en equivalentes de carbón. Al corregir por la calidad de la energía la huella energética revela la importancia de la marea y las olas (basado en Kemp 1977).

Sistemas pantanosos de Puerto Rico e islas limítrofes
9 166 CAP ariel e. lugo | ecología a m i m anera Humedales arbóreos
9.3. Ilustración del concepto de huella o firma
Figura 9.4. Mapa de los humedales de Puerto Rico de Manuel del Llano (1988).

9 167 CAP ariel e. lugo | ecología a m i m anera Humedales arbóreos
elevación la concentración de oxígeno disminuía con la elevación. Los suelos saturados creaban condiciones de humedal en las montañas, o sea, condiciones de bajas concentraciones de oxígeno e incluso condiciones anaeróbicas, y esas condiciones afectan el metabolismo de microorganismos y árboles (Fig. 9.7). El potencial de oxidación-reducción es sensible al oxígeno y cambia dramáticamente con el grado de oxigenación o condiciones anaeróbicas, afectando el estado químico del nitrógeno, hierro, sulfato y el bióxido de carbono (Fig. 9.8). Los suelos anaeróbicos emiten metano en vez de bióxido de carbono exacerbando la producción de gases de invernadero.
Bosques de palo colorado (Cyrilla racemiflora) creciendo sobre geología granodiorita a elevaciones sobre 600 m en El Yunque, los palmares de palma de sierra (Prestoea montana) sobre laderas con aspectos hacia el norte en todas las montañas altas de Puerto Rico, y los bosques elfinos, incorrectamente llamados bosques enanos, en las cima de las montañas, todos resultaron ser humedales cuando se estudiaron con
9 168 CAP ariel e. lugo | ecología a m i m anera Humedales arbóreos
Figura 9.5. Los ecosistemas pantanosos de las Montañas de Luquillo (modificado de Frangi 1983). Las flechas indican el flujo del agua por los humedales. La evapotranspiración no está incluida.
sep92oct 92nov 92dec 92jan93feb 93mar 93apr93may93jun93jul93aug93sep93oct 93nov 93dec 93jan94feb 94mar 94
. 1999).
detenimiento. Comunidades del musgo Sphagnum portoricense son humedales no arbóreos que se encuentran en lugares planos en las cimas de las montañas. La planta que domina los humedales en los Everglades de la Florida, Cladium jamaicense, es una planta común en los humedales no arbóreos en las cimas de las Montañas de Luquillo.
Además, los palmares de palma de sierra también forman humedales en los valles inundables de ríos y quebradas a elevaciones altas, mientras que árboles de palo de pollo (Pterocarpus officinalis) forman humedales en los valles inundables a
9 169 CAP ariel e. lugo | ecología a m i m anera Humedales arbóreos
21 A B Bosque de tabonuco Oxígeno en el suelo (%) Bosque de palo colorado Bosque elfino
18 15 12 9 6 3 0 Fecha Bosque de tabonuco Bosque de palo colorado Bosque elfino 21 18 15 12 9 6 3 0
Figura 9.6. Concentración de oxígeno en el suelo a través del tiempo a 10 cm (A) y 35 cm (B) de profundidad en tres tipos de bosque en el Bosque Experimental de Luquillo. La elevación aumenta entre el tabonuco y el bosque elfino. Los datos representan el promedio + 1 error estándar (modificado de Silver et al
Figura 9.7. Factores que afectan la tolerancia a la inundación de cinco especies arbóreas en humedales de zona templada. Se ilustran adaptaciones ecofisiológicas y morfológicas de cada una de las especies en relación con la inundación (modificado de Hook y Scholtens 1978).
elevaciones bajas y en zonas costeras del norte de Puerto Rico detrás de los manglares. En la zona del karso, ocurren micro humedales en los valles de la región y donde ocurren afloramientos de agua a través del karso. Micro humedales con palma de sierra y palo colorado también son comunes dentro de los bosques de tabonuco en depresiones del terreno con suelos arcillosos.
9 170 CAP ariel e. lugo | ecología a m i m anera Humedales arbóreos
Modelo del efecto de la actividad humana sobre los humedales
La figura 9.9 ilustra cinco efectos de actividad humana sobre los humedales. Las acciones humanas están categorizadas como disturbios o estresores del humedal. El funcionamiento del ecosistema sugiere que la vulnerabilidad del humedal a estas acciones disminuye de izquierda a derecha en el diagrama. En otras palabras, el humedal es más vulnerable a las acciones uno y dos ilustradas en el diagrama y menos vulnerable a las acciones cuatro y cinco. Eso se debe a que las canalizaciones, embalses y otras reducciones de las fuentes energéticas primarias del ecosistema afectan todo el sistema, incluyendo su productividad, mientras que la aplicación de sustancias tóxicas o el corte parcial de árboles afecta solamente una porción del ecosistema sin afectar significativamente su productividad o funcionamiento. Al analizar los efectos de un disturbio sobre un ecosistema se debe siempre considerar que sector del sistema es afectado por la actividad humana (Lugo 1978, 2020).
9 171 CAP ariel e. lugo | ecología a m i m anera Humedales arbóreos
Figura 9.8. El cambio en el potencial de REDOX con la profundidad de los sedimentos en humedales y el resultado químico en términos del estado redox de cinco elementos (oxígeno, nitrógeno, hiero, azufre y carbono) como resultado de los procesos microbianos relacionados (modificado de Lugo et al. 1990a).
1990a). Cada tipo de estresor (numerados del 1 al 5 y con ejemplos) tiene efectos distintos sobre el humedal y se postula que la severidad del estresor a todo el sistema disminuye del 1 al 5. Todos los heterótrofos del sistema están representados por el símbolo denominado «respiración».
Asignación
Selecciona cualquier humedal de Puerto Rico. Indica su ubicación y clasificación y discute aquellos factores ambientales que (1) promueven su productividad y (2) le causan estrés.
9 172 CAP ariel e. lugo | ecología a m i m anera Humedales arbóreos
Figura 9.9. Diagrama que ilustra el punto donde un estresor o disturbio interactúa con un componente de un humedal (modificado de Lugo et al.
CaPítulo 10 Manglares
Pregunta
¿Cuántos tipos de manglares hay en Puerto Rico?
Introducción
Los manglares son humedales de agua salada. También son sistemas estuarinos sujetos a las mareas y dominados por especies arbóreas que crecen en latitudes tropicales, subtropicales y templada cálida (warm temperate). El manglar tolera heladas de baja intensidad y duración, por lo que sobrevive en la zona templada cálida (Lugo y Patterson Zucca 1977). En los humedales costeros de agua salada de zonas templadas frías, contrario a las zonas tropicales y templada cálidas, dominan especies herbáceas de varios géneros, incluyendo Spartina. Mangle se refiere a las especies del manglar, siendo manglar la designación del ecosistema.
El estudio de los manglares está dominado por ideas de zonación, ya que las distintas especies de mangle tienden a formar zonas monoespecíficas que en muchas ocasiones se confunden con progresiones de sucesión (Lugo 1980). Al estudiar las zonificaciones se descubren múltiples gradientes ambientales tales como gradientes de salinidad, oxígeno, profundidad del agua, frecuencia y duración de inundación (hidroperiodo), concentración de sulfatos; REDOX; además de gradientes topográficos y edáficos. Algunos de estos factores se presentaron en el capítulo 9 y otros se desarrollan en el próximo capítulo. Las zonaciones no solo reflejan la distribución de las especies y los gradientes ambientales, sino que también reflejan cambios fisonómicos en la vegetación.
Los manglares de Puerto Rico pertenecen a uno de dos grandes grupos: (1) los que crecen en contacto directo con el mar, y (2) los que crecen tierra adentro protegidos del mar por dunas de arena, en las márgenes de ríos y quebradas donde son sensibles las mareas o en las márgenes de lagunas estuarinas. Los manglares crecen en contacto directo con el mar en el sur de Puerto Rico, generalmente en las orillas del mar Caribe. La costa norte está bajo la influencia del océano Atlántico y sus grandes oleajes. Sin embargo, los manglares no toleran la alta energía de los oleajes
ariel e. lugo | ecología a mi ma nera
173
y por eso requieren refugio detrás de las dunas de arena o tierra adentro hasta donde llegue la influencia de la marea. El funcionamiento y la estructura del manglar varía alrededor de Puerto Rico dependiendo del clima local y de la escorrentía terrestre de agua fresca, sedimentos y nutrientes que lleguen. Por ejemplo, algunos manglares reciben altas descargas de agua, sedimentos y nutrientes como los de Vacía Talega y Piñones, otros reciben solo descargas de agua subterránea como los manglares de Camuy e Isla de Mona, mientras que otros, reciben muy pocas descargas de las cuencas terrestres a las que pertenecen como los manglares de Guánica o la Parguera. Los manglares en los cayos e islas adyacentes a la isla grande de Puerto Rico no reciben escorrentía terrestre y están solamente conectados al mar y la atmósfera por medio de la lluvia. Por la complejidad de condiciones bajo las cuales se encuentran los manglares, cada sector de costa requiere estrategias distintas para la conservación de sus manglares (Lugo y Cintrón 1975). Durante la década de 1970, el DRNA condujo inventarios de los manglares visitando sistemáticamente todos los rodales alrededor de la Isla (Martínez et al. 1979). Cada sector de la zona costanera se caracteriza por tener manglares creciendo bajo distintas condiciones que influencian sus características.
En Puerto Rico se han llevado a cabo estudios pioneros sobre el funcionamiento y productividad de manglares (Golley et al. 1962) al igual que en la descripción sistemática de los manglares de costas áridas y su sucesión ecológica en respuesta a la salinidad del suelo (Cintrón et al. 1978). El área y los cambios en área de los manglares en Puerto Rico a través del tiempo reflejan fielmente aspectos socioeconómicos de la isla, al igual que la efectividad de las medidas de protección (más adelante en este capítulo).
La literatura sobre los manglares es masiva (más de 5,000 trabajos disponibles en la década de 1970, cuando publiqué la revisión sobre la ecología de los manglares con el ecólogo norteamericano Samuel C. Snedaker) pero recientemente ha aumentado dramáticamente, pues se ha acelerado el interés por su estudio. Inicialmente, los manglares se veían como curiosidades botánicas, fisiológicas y ecológicas debido a la presencia de árboles creciendo en agua salada de mar donde la mayor parte de las plantas no pueden sobrevivir. Recientemente, se ha evidenciado el valor de los manglares como protectores de costas ante el aumento en el nivel del mar y como sumideros de carbono que mitigan el calentamiento global. Los trabajos claves en la evolución de los estudios de manglares los resumo a mi manera a continuación.
Zonación
Cualquier estudiante de los manglares notará de inmediato la zonación de las especies perpendicular a la marea. En la clasificación de Kangas en el capítulo anterior, la marea es un manto energético que incide ante el manglar. La secuencia clásica en los manglares del nuevo mundo consiste en un borde frente al mar
10 174 CAP ariel e. lugo | ecología a m i m anera Manglares
dominado por el mangle rojo (Rhizophora mangle), seguido por una zona de mangle negro ( Avicennia germinans), una de mangle blanco (Laguncularia racemosa) y finalmente en la parte exterior del manglar, una zona de Conocarpus erectus, el mangle botón. El norteamericano John Henry Davis (1938) fue uno de los pioneros en los estudios de los manglares, no solo estudiando la zonación, sino también la acumulación de suelo orgánico (turba o peat en inglés) bajo los manglares y otros humedales de la Florida (Davis 1946). Estas acumulaciones de turba sugerían la capacidad de los manglares para crear tierra y avanzar sobre el mar. Hoy sabemos que la acumulación de turba es una respuesta al aumento en el nivel del mar. Durante los últimos 4,000 a 5,000 años, los manglares del Caribe han ajustado sus pisos ante la subida del mar durante todo ese tiempo, como se demostró en el manglar enano de Ceiba (Medina et al. 2010).
La síntesis del neozelandés Valentine Jackson Chapman (1976) para los manglares del mundo culmina los trabajos clásicos de zonación e historia natural de manglares, ya que Chapman resumió todo el material disponible hasta el momento de su publicación. El libro de Chapman coincidió con el momento histórico cuando el estudio de manglares se tornó más ecológico, funcional y a escalas de paisaje (Lugo y Snedaker 1974, Thom 1975). El pase del batón ocurrió en Hawái, del 8 al 11 de octubre de 1974, cuando se reunieron expertos en manglares de todo el mundo, con la presencia de Chapman y muchos otros ecólogos de renombre. Las actas de la reunión están resumidas en dos volúmenes editados por Walsh et al. (1975). La publicación fue dedicada a William Macnae, zoólogo escocés que trabajó en las costas de África del Sur. Entre sus diversos trabajos en zoología y ecología se distinguió la síntesis sobre los manglares de la región Indopacífico occidental (Macnae 1968).
Relación entre los manglares y las mareas
James Gilbert Watson, dasónomo inglés, era el segundo del Conservador de Bosques de los Estados Federados de Malasia. Su publicación sobre los manglares resume todos los trabajos de conservación, uso y silvicultura de manglares en la península de Malasia a principios del siglo XX (Watson 1928). Se distinguió por establecer una relación cuantitativa entre la frecuencia y profundidad de las mareas y la presencia de las especies de mangle en Malasia. Como se aprecia en la figura 10.1, las especies de mangle establecen zonas de dominancia en función a la frecuencia de las mareas. Watson estableció cinco clases de inundación (Tabla 10.1) para explicar la zonificación de las especies y guiar su uso y conservación. Este trabajo de Watson, mejorando un esfuerzo anterior de su colega A.B.S. Boswell, fue de los primeros en establecer una relación cuantitativa entre la zonación de los manglares con las mareas. Permitió un mejor manejo de las especies de acuerdo con las condiciones mareales. Ese trabajo también sirve de base para evaluar los cambios recientes en
10 175 CAP ariel e. lugo | ecología a m i m anera Manglares
Sección
de pr ima ver norma infer ior m uer to od os
Niv e le s equinocciale s
Rhizophoramucronata Niv le equinocciale de pr ima ver a norma infer ior m uer to od os
Rhizophora conjugata
Avicennia intermedia
Bruguiera caryophylloides
Bruguiera caryophylloides
Bruguiera parviflora
Avicennia alba
Bruguiera eriopetala
Rhizophora conjugata
Rhizophora mucronata
Sonneratia gri thii
Avicennia alba
Bruguiera parviflora
Rhizophora conjugata
Avicennia intermedia
Rhizophora mucronata
Rhizophora conjugata
Bruguiera parviflora
Bruguiera caryophylloides
Bruguiera caryophylloides
Bruguiera eriopetala
Bruguiera gymnorhiza
Bruguiera eriopetala
Rhizophora conjugata
BruguieraRhizophoraconjugata parviflora
Bruguiera parviflora Carapaobovata
Rhizophoraconjugata
Bruguiera gymnorhiza
Rhizophoramucronata
Sonneratia gri thii
Rhizophora mucronata
Sección A-B
Referencia
Carapa obovata
Carapa moluccensis
Intsia retusa
Figura 10.1. Diagrama para ilustrar la distribución típica de las especies de mangle más importantes en la península de Malasia (Watson 1928). El marco del mapa de zonación contiene tres transectos marcados en el diagrama (A-B, C-D y E-F) que ilustran cortes verticales de la vegetación y la topografía, incluyendo líneas horizontales con las clases de inundación por las mareas (Tabla 10.1). Watson aclaró que las zonaciones diagramadas son típicas pero no inevitables.
Rhizophora conjugata
Ceriops candolleana
Sonneratia gri thii
Avicennia intermedia
Bruguiera gymnorhiza
Carapa moluccensis
Intsia retusa
Oncosperma, Ficus y formas semi-interior
Intsiaretusa Bruguieragymnorhiza Avicennia alba Bruguiera parviflora Carapa obovata 12345 54321
Avicennia alba
Acrostichum aureum
10 176 CAP ariel e. lugo | ecología a m i m anera Manglares
C-D
E-F 1 2 3 4 5 1 2 3 4 5 Aguas d el estuario MA R ABI ER TO A
Sección
Carapamoluccensis Ceriops candolleana Rhizophora mucronata C F B D E Avicennia intermedia Carapa obovata Rhizophora conjugata Bruguiera gymnorhiza Carapa obovata Sonneratiaalba Sonneratia alba Carapa obovata Bruguiera gymnorhiza Carapa obovata Rhizophora conjugata Bruguiera parviflora Rhizophora conjugata Rhizophora mucronata Bruguiera parviflora Rhizophora conjugata Sonneratia gri thii Avicennia alba Rhizophora conjugata Rhizophora mucronata Rhizophoramucronata Ceriops candolleana
Rhizophora conjugata Sonneratia gri thii
Tabla 10.1
Esquema propuesto por Watson (1928) para clasificar la distribución de especies de mangle en la península de Malasia de acuerdo con la clase de inundación, la cual es determinada por el número de inundaciones por las mareas. Para convertir de pies a centímetros, multiplique por 30.48. Las alturas de las mareas varían con la ubicación geográfica del manglar.
Clase de inundación Inundado por: Altura sobre el datum del almirantazgo (pies) Número
marea inunda mensualmente
veces que
el nivel del mar y sus efectos sobre los manglares. Watson entendía desde entonces que los manglares tenían limitaciones en cuanto a su capacidad de robarle terreno al mar como creían, y aún creen, muchos.
Primer estudio de manglares en cuanto al ecosistema
A final de la década de 1950, Odum y el norteamericano Frank Benjamín Golley colaboraron en estudios pioneros del metabolismo de bosques tropicales, incluyendo el bosque de tabonuco en el sector Bisley de El Yunque (Odum et al. 1970) y los manglares de la Parguera en el sur de Puerto Rico (Golley et al. 1962). Esa colaboración dio eventualmente como resultado el estudio de radiación en El Verde (capítulos 6 y 8) y en el primer trabajo sobre el metabolismo de los manglares. En ese estudio el manglar fue subdividido por compartimientos y por medio de medidas de intercambio gaseoso y características estructurales. Golley et al. estimaron la productividad bruta, la productividad neta y la respiración del manglar durante el mes de mayo (Fig. 10.2).
Interpretación de la zonación del mangle
Egler (1952) fue uno de los primeros en reconocer que la zonación en los manglares de los Everglades de la Florida estaba asociada a gradientes de salinidad en vez de ser la recapitulación de la sucesión a través del tiempo. Egler también enfatizó la importancia del pasaje de huracanes en el reajuste de las distintas capas (belts) o zonas de vegetación, ya que las lluvias asociadas a los huracanes pueden modificar la salinidad del paisaje. En 1980, publiqué un resumen sobre la influencia de gradientes ambientales que afectaban la dirección de la sucesión en los manglares (Fig. 10.3). El diagrama solo consideró las gradientes no antropogénicos bajo la influencia de no menos de 19 fuerzas externas al manglar. En general enfaticé sucesiones asociadas a la profundidad del hidroperiodo, disminución en la salinidad del suelo o en los nutrientes y aumentos en salinidad o disminución en la temperatura. Tales
10 177 CAP ariel e. lugo | ecología a m i m anera Manglares
de
MínimoMáximoMínimoMáximo 1 Todas las mareas altas < 8 56 62 2 Mareas altas medianas 8 11 45 59 3 Mareas altas normales 11 13 20 45 4 Mareas altas de primavera 13 15 2 20 5 Mareas anormales o equinocciales > 15 2
la
Figura 10.2. Balance de carbono de un manglar en la Parguera durante el mes de mayo. Se ilustran las tasas de fotosíntesis (P), respiración (R) y exportación de carbono. El sistema respira y exporta más de lo que fotosintetiza debido a la exportación de materia orgánica. Sin embargo, la fotosíntesis es superior a la respiración (8.23 vs. 7.79 gramos de carbono por metro cuadrado por día), lo que indica que es un sistema autotrófico que exporta una porción de su productividad (modificado de Golley et al. 1962).
condiciones generaron sobre 15 trayectos de sucesión, todos potencialmente reversibles. Tales sucesiones no incluyeron sucesiones dirigidas por la actividad humana como la conversión en el uso del terreno, el calentamiento global o cambios en el hidroperiodo. Claramente, la dinámica de los manglares a través del tiempo puede ser bastante diversa y compleja.
Dado el dinamismo ambiental en los sistemas de manglar, la vegetación madura que se observa en estos lugares representa un estado estable temporero compuesto por especies de mangle con la capacidad de ser pioneras o primarias, exhibiendo el fenómeno de autosucesión cuando las condiciones así lo exigen.
10 178 CAP ariel e. lugo | ecología a m i m anera Manglares
AV-L Bos
Huracán nive de mar más alto, hundimiento de ierra, tormentas, descarga de a marea en aumento
Ecosistema terre stre
Fuego
Agua más profunda, mayo energía de olas o mareas
Agua fresca en aumento salinidad de suelo en descenso
RH-AV Franjas, cuenca o bosque en isla rebasado por la marea
Levantamiento de a tierra nive de ma bajo, salinidad de suelo más alta menos acción de marea o de olas
Franjas de RH Cuenca, ribereño o bosques en isla rebasado por la marea Menos energía de marea o de olas, levantamiento costero, descenso en e nivel de mar
Substrato poco profundo bajos nutrientes
Cambio geomorfológico o ambiental dramático
Ecosistemas marinos
Temperatura mínima más baja o amplitud de temperatura más alta
AV Bosque de cuenca
Huracanes, tormentas aumento de lluvia, baja salinidad en e suelo rebasado de mar
AV enano Matorrales
Huracanes, tormentas nivel de mar más alto nive de a tierra más bajo
AV enano Plantas de pantanos salados
Salinidad en el suelo
> 100&
Baja salinidad en el suelo
Huracanes, tormentas nivel de la tierra más bajo nivel de ma más alto
RH enanos Matorrales
Pérdida de suelo
RH enanos Plantas de agua fresca Salinidad de suelo en aumento
Profundidad de suelo en aumento
RH Bosque de hamaca Agua dulce
Ecosistema de marismas herbáceas
Ecosistema terre stre
Dism nución de frecuencia de heladas y amplitud de temperatura
Ecosistema hipersalino
Figura 10.3. Diagrama de posibles sucesiones en manglares y los factores responsables por ellas. Las cajas representan las etapas serales (RH es Rhizophora, L es Laguncularia y AV es Avicennia). Las flechas grandes (símbolo multiplicador en el sistema de Odum [Fig. 2.9b]) indican la dirección de la sucesión y están conectadas con los factores físicos responsables (círculos) por esa línea de sucesión. Todas las sucesiones son reversibles. Cajas con paredes dobles son sistemas ecológicos que no son manglares. Modificado de Lugo (1980), donde hay más detalles.
En Puerto Rico, Cintrón et al. (1978) hicieron un trabajo pionero en los manglares de las costas áridas de la isla, enfatizando la importancia de la salinidad del suelo en el funcionamiento y estructura de los manglares en zonas áridas (Fig. 10.4). En ese estudio se estableció la concordancia entre distintos niveles de salinidad en los suelos con las especies de mangle y su estructura. Tal como se sugiere en la figura 10.3, aumentos paulatinos en salinidad afectan la altura y composición de especies de mangle y al llegar a valores que exceden 100 partes por mil o más del doble de la salinidad
10 179 CAP ariel e. lugo | ecología a m i m anera Manglares
que de cuenca L Bosque de cuenca Acumulación de sedimentos, nivel freático más profundo, salinidad del suelo en descenso Ecosistema de agua fresca
del mar (35 partes por mil), el mangle da paso a una salina sin vegetación arbórea. Esa sucesión se revierte con la lluvia extrema de sistemas atmosféricos estacionarios o con la lluvia de huracanes, que lavan las sales del suelo, permitiendo el crecimiento arbóreo. Esta sucesión cíclica es típica en las costas áridas pobladas por manglares.
Síntesis del manglar y su funcionamiento
Lugo y Snedaker (1974) reorientaron el estudio de los manglares hacia un foco funcional, reconociendo que la geomorfología e hidrología determinaban tipos funcionales de manglares (Fig. 10.5). Estos tipos de manglar eran los de borde, los de cuenca, los ribereños, los de islotes lavados por las mareas y los enanos. Esta clasificación coincide con la clasificación de Kangas basada en la energía de las fuerzas hidrológicas que potencian los humedales y con las ideas geomorfológicas de Thom (1975). La clasificación permite analizar la estructura y el funcionamiento del manglar en un contexto de sistemas, sustituyendo el análisis basado en las especies. Sin embargo, la composición de especies es importante para entender las adaptaciones ecofisiológicas de los manglares ante los retos ambientales que enfrentan. Las
10 180 CAP ariel e. lugo | ecología a m i m anera Manglares
Figura 10.4. Perfiles de islotes de mangle en la costa sur de Puerto Rico. Se ilustra el nivel de las aguas y del terreno y su relación con la altura de la vegetación y la salinidad del suelo y del agua superficial (modificado de Cintrón et al. 1978).
10.5. Seis tipos de manglares propuestos por Lugo y Snedaker (1974). La geomorfología del terreno con relación al hidroperiodo y la dirección y energía de las aguas mareales y ribereñas determinan el tipo de manglar. En el caso de los manglares de hamacas y los enanos interceden factores históricos y nutricionales.
10 181 CAP ariel e. lugo | ecología a m i m anera Manglares
Figura
especies se segregan de acuerdo con sus adaptaciones a las condiciones prevalecientes en distintos puntos del ecosistema.
Los avances más recientes en el estudio de los manglares se basan en estudios detallados sobre la ecofisiología de las especies y estudios globales sobre el funcionamiento de los manglares en contextos regionales y globales. O sea, por un lado, se estudian los mecanismos internos de un rodal a escalas ecológicas menores a la del rodal, incluyendo el nivel ecofisiológico, y por otro, se estudia la posición del rodal a escalas ecológicas mayores, incluyendo la escala social-ecológica-tecnológica (ver revisiones bibliográficas en Medina et al. 2008, Lugo y Medina 2014, Lugo et al. 2014).
La visualización de las jerarquías de estudio que aplican a los manglares la presentaron Twilley et al. (1996) y está resumida en la figura 10.6. Partiendo de la distribución global de los manglares, mayormente regulada por la temperatura, el segundo eslabón de la jerarquía son los cinco entornos geomorfológicos que propuso Thom y que Twilley et al. (1996) resumen en cuatro. Dentro de cada entorno, y de hecho en cualquier paisaje dominado por manglares, se ubican rodales cuyo funcionamiento es
10 182 CAP ariel e. lugo | ecología a m i m anera Manglares
Figura 10.6. Clasificación jerárquica utilizada para describir los diversos patrones de la estructura y funcionamiento de los mangles basado en los factores globales (temperatura), regionales (geomorfológicos) y locales (ecológicos) que controlan las concentraciones de recursos (nutrientes y agua) y niveles de estresores en los suelos de los manglares (modificado de Rivera Monroy et al. 2017). Todos son sistemas estuarinos.
conforme a los tipos de manglar propuestos por Lugo y Snedaker (1974). Esos rodales constituyen el tercer eslabón en la jerarquía funcional de los manglares. El funcionamiento de un manglar responde no solo a los criterios geomorfológicos e hidrológicos considerados en el sistema de Lugo y Snedaker, sino que también responde a los recursos nutricionales disponibles y las fuerzas de estrés y disturbios en su entorno. Las escalas de tiempo y espacio inherentes en la figura 10.6 se amplifican entrando a los varios componentes del rodal según se ilustra en la figura 10.7. Allí se indica que los estudios de manglares pueden cubrir escalas de tiempo y espacio, desde horas o menos y centímetros cuadrados o menos de los estudios ecofisiológico de las hojas de mangle hasta más de miles de hectáreas y miles de años de los estudios geomorfológicos.
El manglar y su función en las cadenas alimenticias
Los trabajos doctorales del norteamericano Erick Heald (1971) y del hijo de E.P. Odum, William Odum (1971), en los manglares de la Florida, demostraron la importancia del detrito de los manglares en el sustento de las cadenas alimenticias de pesquerías comerciales en aguas oceánicas (Fig. 10.8). Esos estudios fueron inicialmente controversiales, pero sus hallazgos originales se han sostenido con estudios utilizando isótopos. No hay duda de que las hojas de los manglares, cuando caen y son procesadas por animales y microorganismos que las fragmentan y aumentan la calidad nutricional de los fragmentos foliares, alimentan los peces en los estuarios y en el mar. Las pescas comerciales costeras dependen de la productividad de los manglares.
Cambio histórico en el área de los manglares de Puerto Rico
Los manglares de Puerto Rico han sufrido enorme presión antropogénica. En un momento dado, el gobierno auspiciaba la tala y relleno de los manglares por causas de salud pública, temiendo erróneamente que los manglares eran fuente de enfermedades transmitidas por mosquitos. Los manglares de Ocean Park, el Condado, Puerto Nuevo y Hato Rey se rellenaron para dar paso al desarrollo urbano en más de un 20 % del municipio de San Juan. Hoy, con el alza en el nivel del mar y el deterioro de la infraestructura pluvial en San Juan, todas estas áreas rellenadas y construidas se inundan frecuentemente, causando extensas pérdidas al municipio, los comercios y los residentes. Los manglares también fueron deforestados para dar paso a la agricultura, desarrollos urbanos, y para depositar basura. Más de la mitad del área original de manglares en la Isla desapareció. El cambio histórico en las áreas de mangle en Puerto Rico fue analizado por Martinuzzi et al. (2011), quienes demostraron la pérdida y recuperación de estos sistemas (Fig. 10.9). Los cambios en el área de manglares en Puerto Rico reflejan cambios sociales, culturales y económicos en la isla. En el presente, los manglares están protegidos en Puerto Rico y son parte de la zona marítimo-terrestre que por ley les pertenecen a todos por igual y a nadie en particular.
10 183 CAP ariel e. lugo | ecología a m i m anera Manglares
10 184 CAP ariel e. lugo | ecología a m i m anera Manglares
Figura 10.7. La jerarquía en tiempo y espacio de los estudios de manglares desde el nivel de la hoja al entorno ambiental geomorfológico y de horas a milenios (modificado de Twilley et al. 1988).
Asignación
¿Por qué es que en zonas templadas frías los pantanos costeros están dominados por herbáceas y en los trópicos por árboles?
10 185 CAP ariel e. lugo | ecología a m i m anera Manglares
Figura 10.8. Procesos ecológicos costeros y flujos de energía asociados que convergen en los consumidores al final de la cadena alimenticia que beneficia al ser humano por medio de la pesca comercial (modificado de Odum et al. 1982). Los organismos incluidos dependen de varios ecosistemas, incluyendo los manglares y de varios procesos de intercambio energético. Las líneas gruesas resaltan la contribución del manglar a estas cadenas alimenticias. El manglar contribuye con todas las fuentes de alimentación que llegan a los consumidores mayores y por ende a la pesca comercial.
Figura 10.9. Cambios a través de tiempo en la cobertura de los manglares en Puerto Rico, modificado de Martinuzzi et al. (2009). Se sugieren eventos históricos asociados con los cambios en cobertura.
CaPítulo 11 El Yunque
Pregunta
¿Es o no es El Yunque una maravilla natural del mundo y cómo defiende usted su opinión, o sea, ¿por qué?
Introducción
Sobre los bosques de El Yunque se han escrito miles de artículos y libros, mucho más de lo que se ha escrito sobre el resto de los bosques de Puerto Rico combinados. En este capítulo paso por alto los detalles y le doy énfasis a asuntos generales como la estructura y funcionamiento de los sistemas geológicos y ecológicos de El Yunque. El lector interesado debe consultar a Brokaw et al. (2012b) para un resumen reciente sobre la ecología de los sistemas ecológicos de El Yunque. Otros resúmenes disponibles son los de Wadsworth (1949), Odum y Pigeon (1970), Brown et al. (1983), Weaver (1994, 2012), Robinson (1997), Harris et al. (2012), González et al. (2013), Robinson et al. 2014 y Quiñones et al. (2018).
Ubicación geográfica
El Yunque es parte de las Montañas de Luquillo y es a estas montañas a las que me refiero cuando lo menciono. El Yunque es también el nombre de uno de los picos de las Montañas de Luquillo y es el nombre que se le ha conferido al Bosque Nacional. Los puertorriqueños en general se refieren al Bosque Nacional cuando mencionan El Yunque. Para los taínos, las Montañas de Luquillo eran sagradas por ser la residencia del espíritu benefactor Yucahú o Yucayú. La imagen del cemí o ídolo de tres puntas con cabeza y pies humanoides, cargando la isla en la espalda, quizás evolucionó de la contemplación de las montañas. Mabuya y Juracán eran los espíritus temibles asechando el entorno.
Las Montañas de Luquillo están ubicadas en el noreste de Puerto Rico y emergen abruptamente a sobre 1,000 metros sobre el nivel del mar. La distancia lineal entre el pico El Yunque y una playa es de 8 km. Las Montañas de Luquillo interceptan los vientos alisios y ocupan la esquina noreste de la placa tectónica del Caribe, formando
ariel e. lugo | ecología a m i m anera
187
Figura 11.1. Ubicación de la base y el borde de la plataforma del Oligoceno-Mioceno de Puerto Rico (A) y cortes transversales de los dos transectos (B y C) al centro (A-A’) y al este (B-B’) de Puerto Rico. La base de la plataforma y la formación Oligoceno-Mioceno están sobrepuestas sobre la forma actual de la Isla (modificado de Brocard et al. 2015). La formación Oligoceno-Mioceno corresponde al karso del presente y las rocas plutónicas a las montañas de Puerto Rico. Las arenas de manta son producto de la erosión en las montañas y su transporte fluvial a las tierras bajas. Las Montañas de Luquillo están ubicadas en el noreste y la Isla era más angosta debido al nivel del mar que coincidía con la base de la plataforma Oligoceno-Mioceno. Puerto Rico y las Islas Vírgenes compartían el mismo bloque tectónico.
11 188 CAP ariel e. lugo | ecología a m i m anera El Yunque
un bloque tectónico con las Islas Vírgenes (Fig. 11.1). Estas Montañas son jóvenes habiendo emergido en los últimos 4 millones de años (Ma) y aún no han llegado a un equilibro tectónico dinámico (Brocard et al. 2015).
Caliga del Oligoceno - Mioceno
Capas de Rocas del Eoceno
Capas de rocas de Cretáceo Alto
Capas de rocas del Cretáceo Bajo Serpentinita, perdernal, meta-basitas
Geología e historia natural de Las Montañas de Luquillo
Bajo
Tipos de rocas. Rocas ígneas (procedentes del fuego) se forman del magma (roca líquida) dentro de la corteza terrestre. Si el magma se enfría y solidifica dentro de la corteza terrestre, se forman rocas intrusivas denominadas plutónicas, aludiendo al dios romano Plutón que reinaba en el bajo mundo. El magma que sale por un volcán se conoce como lava. Cuando la lava se enfría y solidifica sobre la corteza de la tierra, forma rocas extrusoras denominadas volcánicas (dios romano Vulcano, dedicado al trabajo con metales). Ejemplos de rocas plutónicas son los batolitos (rocas profundas), el granito, granodioritas ( granodiorite) y dioritas de cuarzo (quartz diorite). El basalto es un ejemplo de rocas volcánicas. Cada tipo de roca tiene una mineralogía particular.
Las rocas plutónicas forman la base de los continentes y parte de Puerto Rico. El batolito de San Lorenzo, con un diámetro de 20 km y edad de entre 66 y 78 Ma, es un ejemplo (Joyce 1992). La corteza terrestre bajo Puerto Rico tiene un espesor de 25 km. Las rocas volcánicas, como el basalto, forman la base de los océanos y en Puerto Rico, parte de las montañas. Las Montañas de Luquillo están en la provincia geológica ígnea del noreste, delimitada al sur por la falla geológica del Cerro Mula (Fig. 11.2). Los picos volcánicos de El Yunque corresponden al cretáceo superior (97 a 113 Ma; ver Fig. 11.3 para la escala geológica del tiempo) y los de dioritas de cuarzo tienen una edad de 47 Ma (Porder et al. 2015).
11 189 CAP ariel e. lugo | ecología a m i m anera El Yunque
Rocas intrusivas
Terciario
Rocas intrusivas del Cretáceo 10 11 12 13 14 6 7 5 89 km Fall a geo ló gic a C err o Mu la 3020100 4 3 2 1 Provincia ígnea del noreste Provincia ígne a central
del
Provincia ígnea del suroeste
Figura 11.2. Mapa de tres provincias de rocas ígneas y las calizas del Oligoceno- Mioceno de Puerto Rico con las edades de las distintas rocas. Se ilustra también la ubicación de las dos fallas geológicas principales en Puerto Rico (Joyce 1992). Los números corresponden a paradas en una excursión guiada por el Dr. James Joyce.
Etapa
PL
ALTO
BAJO NEÓGENO PALEÓGENO CRETÁ CE O CUATER NA RI O
DANIENSE
MA AS T RI CHT IENSE
CAMPANIENSE
S AN TONIENSE
CONIACIENSE
TURONIENSE
CENO MANIENSE
ALBIENSE
APT ENSE
BA RR EMENSE
HAUTERIVIENSE
VALANGIENSE
¹ Hace millones de años.
KIMMERIDG ENSE
OXFOR DIENSE
CALLOVIENSE
BAT HO NIENSE
BAJOCIENSE
AALENIENSE
TOARCIENSE
PLIENSBAQ UENSE
SINEMURIENSE
HETTANGIENSE
R HA ETIENSE
NO RIENSE
KAR NIENSE
LADINIENSE
ANISIENSE
OLENEKIENSE
ENSE CHA NGH SINGIENSE WUCHIAPINGIENSE CAPITANENSE WOR DIENSE ROADIENSE KUNGURIENSE ARTINSKIENSE SAK MA RIENSE ASSELIENSE G ZHELLANIENSE KASIMOVIENSE MOSCOVIENSE BA SHKIRIENSE SERPUKHOVIENSE VISIENSE TOUR NA ISIENSE Etapa / Edad Series /Época Sistema / Pe riodo Eratema / Era E on ostema / Eón m /año -145.0 152.1 + 0.9 157.3 + 1.0 163.5 + 1.0 166.1 + 1.2 168.3 + 1.3 170.3 + 1.4 174.1 + 1.0 182.7 + 0.7 190.8 + 1.0 199.3 + 0.3 201.3 + 0.2 -2 08 .5 -227.0 -237.0 -242.0 247.2 251.2 251.902 + 0.024 254 .14 + 0.7 259.1 + 0. 5 265.1 + 0.4 268.8 + 0. 5 272.95 + 0.11 283.5 + 0.6 290.1 + 0.26 2 95 .0 + 0.18 29 8.9 + 0.15 303.7 + 0.1 307.0 + 0.1 315.2 + 0.2 323.2 + 0.4 330.9 + 0.2 346.7 + 0.4 358.9 + 0.4 ALTO ALTO ALTO ALTO MEDIO MEDIO MEDIO MEDIO BAJO BAJO LOPINGIANO GUADALUPI AN A CISURALIANO BAJO BAJO MESOZ OI CA TRIÁSICO JU RÁSICO PÉ RMICO CA RBO NÍ F ER O PE NNSILVIANO 2 MISSIS SI PP IANO 2 PALEOZ OI CA FANEROZ OI CO E on ostema / Eón FANEROZ OI CO
² Las unidades de tiempo Mississippiano y Pennsilviano están formalmente designados como subperiodos del Periodo Carbonífero.
El eón 5 La é y la
confir
11 190 CAP ariel e. lugo | ecología a m i m anera El Yunque P re sente Era Mesozoica 1,000 1 2,000 1 3, 00 0 1 4, 00 0 1 4, 60 0 1 Era C eno zoica Era Paleozoica Tiempo Precámbrico ALTO Hol oceno 1950 CE 0.0117 0.126 0.781 1.80 2.58 3.60 0 5. 33 3 7.246 11.63 13. 82 15 .97 20.44 23.03 27.82 33.9 37.8 41.2 47.8 56.0 59.2 61.6 66.0 72.1 = 0.2 83.6 = 0.2 8 6. 3 = 0.5 89.8 = 0.3 93.9 10 0. 5 -113 -125.0 -129.4 -132.9 -139.8 -145.0 Antropoceno 5 MEDIO CALABRIENSE GELA SIENSE PIACENZIENE ZANCLAY ENSE MES SINIENSE TORTO NIENSE SERR AVALIENSE LA NG HIENSE BURDIGALIENSE AQUITA NIENSE CHATIENSE RUPELIENSE PRIA BO NIENSE BARTONIENSE LU T ECIENSE YPRESIENSE
HA
T
NETIENSE SELA NDENSE
BERRÁSIENSE EISTOCENO PIOCENO
NOZ OI CA
OI CA FANEROZ OI CO
MIOCENO OIGOCENO EOCENO PALEOCENO
CE
MESOZ
TI TONIENSE
INDU
/ Edad Series / Época Sistema / Pe riodo Eratema / Era E on ostema / Eón m /año
³ Varios límites de la unidad de edad C ambriana son informales en espera de ratificación.
Carbonífero.
MEDIO
Escala de tiempo geológica
BAJO
ALTO
O NIENSE
R HUDDA NIENSE
HIR NA NT IENSE
KATIENSE
SANDBIENSE
DA RR IWILL ENSE
MEDIO BAJO SERIE
DAPINGIENSE
FLOIENSE
TREMA DOCIENSE
ETAPA 10
JIA NGSHA NIENSE
PALBIA NIENSE
GUZHA NG L ENSE
DRUR NIENSE
IENSE
El eón Hadeano es un intervalo de tiempo informal del tiempo geológico.
5 La épo ca del Antropo ceno fue declarada un intervalo de tiempo formal por el grupo de trabajo del Antropo ceno (AWG) y la Comisión Internacional de Estratigrafía (ICS), en agosto de 2018. La formalización del intervalo aún espera confir mación del ICS y de la Unión Internacional de Ciencas Geológicas (IUGS).
Figura 11.3. Escala de tiempo geológico de acuerdo con la Enciclopedia Británica (https:// cdn.britannica. com/67/73167-050B9A74092/chart. jpg). El Triásico ya no se reconoce como una Época y se ha subdividido en dos: Paleógeno y Neógeno. Elegí esta escala porque reconoce la Época del Antropoceno.
11 191 CAP ariel e. lugo | ecología a m i m anera El Yunque 0.9 1.0 1.2 1.3 1.4 0.7 1.0 0.3 0.2 0.024 0.7 5 0.4 5 0.11 0.6 0.26 0.18 0.15 0.1 0.2 0.4 0.2 0.4 0.4
Pe riodo Eratema / Era E on ostema / Eón m
358.9 + 0.4 372.2 + 1.6 382.7 + 1.6 387.7 + 0.8 393.3 + 1.2 407.6 + 2.6 410.8 + 2.8 419.2 + 3.2 423.0 + 2. 3 425.6 + 0.9 427.4 + 0. 5 43 0. 5 + 0.7 43 3. 4 + 0.8 438.5 + 1.1 440.8 + 1.2 44 3. 8 + 1.5 445.2 + 1.4 453.0 + 0.7 458.4 + 0.9 467.3 + 1.1 470.0 + 1.4 477.7 + 1.4 48 5. 4 + 1.9 -4 89.5 - 49 4.0 -497.0 -50 0. 5 -504.5 -509.0 - 514.0 -521.0 -529.0 541.0 = 1.0 ALTO FAMENNIENSE FRA SNIENSE GIVETIENSE EIFELIENSE EM SIENSE PRAGUIENSE LO CHKOVIENSE PRIDO LI L UDLOWIENSE WENLOKIENSE LLANDOVERY L UDFOR DIENSE GORSTIENSE HO MERIENSE SHEINWOODIENSE TELYCHIENSE A ER
Etapa / Edad Series /Época Sistema /
/año
ETAPA
ETAPA
ETAPA
ETAPA
SERIE 2 TE
ENEUV
3
5
4
2 FORT UNIENSE
3
RR
GIANO P AL EOZ OI CA DE NO N IA NO SILÚ RI CO ORD OVÍCICO CÁMB RI CO 3 FANEROZ OI CO Sistema / Pe rio do Eratema / Éra E on otema / Eón m /año 541.0 = 1.0 -635 -720 1,00 0 1,200 1,400 1,60 0 1,800 2,050 2,300 2, 500 2,800 3,200 3,60 0 4,00 0 -4,60 0 HA DEICA 4 EOARCAICA PALEOARCAICA MESOARCAICA NEOARCAICA
FURON
PALEO PROT EROZOICA
MESO PROT EROZOICA
RIÁSICO SIDÉRI CO S TATHERIANO ORO SÍRICO ECTASIANO CALIMIANO TONIA N CRIOGÉNICO EDIACARÁN STENIANO ARCAICO PRO T ERO ZOICO PRECÁMBR IC O
NEO PROT EROZOICA
Cuando las rocas ígneas se exponen a la atmósfera o las condiciones dentro de la corteza terrestre, sufren cambios químicos, análogos a la descomposición de la materia orgánica. El producto de la descomposición química de las rocas ígneas es la saprolita. La erosión generalmente se refiere al efecto de fuerzas físicas sobre el terreno, mientras que la denudación se refiere a la erosión más los efectos químicos sobre el terreno. Ambos procesos bajan o reducen la elevación del paisaje. Cuando la erosión es más rápida que la disolución de minerales, se remueven minerales primarios antes de que se disuelvan, dejando rocas expuestas y no erosionadas en pendientes sin suelos. Cuando la erosión es más lenta que la disolución de minerales primarios, se desarrollan suelos y saprolita altamente desgastados (weathered) y la erosión química se limita por la disponibilidad de minerales recientemente expuestos. Para rocas graníticas, las tasas de denudación máximas ocurren a 0.1 mm/año. En el valle del río Icacos, con rocas dioritas de cuarzo, las tasas de denudación son ≈0.05 – 0.1 mm/año y en la cuenca volcanoclástica (rocas volcánicas fragmentadas) de Bisley la denudación es ≈0.4 mm/año (Porder et al. 2015).
Cómo eventos geológicos, Ma atrás, afectan el ciclaje de nutrientes de los bosques del presente: simplificando una historia compleja1. Los ríos transportan al mar los sedimentos y minerales, producto de la denudación de sus laderas, reflejando las fuerzas erosivas que reducen la elevación de las montañas. Las fuerzas tectónicas elevan las montañas, depositando rocas ígneas intrusivas y extrusoras. A largo plazo se balancea el aumento en elevación y la denudación de la masa montañosa. El transporte de sedimentos por los ríos se afecta por procesos cuyos efectos cubren distintos intervalos de tiempo. Por ejemplo, alteraciones antropogénicas, derrumbes de tierra y terremotos son procesos distintos que afectan el transporte de sedimentos por los ríos por meses hasta décadas. Fluctuaciones climáticas afectan el transporte de sedimentos por siglos y milenios. El levantamiento tectónico desenlaza procesos fluviales que duran millones de años, o sea, tiempo geológico. Les toma a los ríos entre cientos de miles y Ma responder a los levantamientos tectónicos, como los que ocurrieron en las Montañas de Luquillo hace 4 a 5 Ma.
La historia es la siguiente: sobre 30 Ma atrás, los picos de las Montañas de Luquillo eran una (Fig. 11.4a) de varias paleo-islas (Fig. 11.4b) en Puerto Rico. En ese momento, existía un alto nivel del mar y por eso los picos de las montañas aparecían como islas rodeadas del mar (Fig. 11.4c). El efecto de los cauces de los ríos sobre las rocas ígneas reflejó un balance entre las tasas erosivas y las tectónicas que denudaban y levantaban (respectivamente) las Montañas de Luquillo. Esos mares permitieron también el desarrollo de toda la región del
11 192 CAP ariel e. lugo | ecología a m i m anera El Yunque
1 Esta sección está basada en Brocard et al. (2015).
karso de Puerto Rico, donde se depositaron rocas calizas de origen biogénico (arrecifes de coral).
Cuando se retiró el mar, y debido a un pulso de levantamiento tectónico de las Montañas de Luquillo hace 4 a 5 Ma, ocurrió en la región un desbalance entre los procesos erosivos y los tectónicos. Se favoreció el levantamiento de las montañas y se expusieron rocas ígneas a la erosión. Debido al desbalance entre levantamiento tectónico y erosión (a favor del levantamiento) se crearon regiones en desequilibrio generalizado y se creó un paisaje compuesto de dominios con distintas tasas de erosión separados por puntos de inflexión (knickpoints en la literatura inglesa) (Fig. 11.5).
El punto de inflexión quiere decir recodo, hendidura o repliegue en el cauce de un río, es un cambio abrupto en la pendiente del río debido a una variedad de factores geomorfológicos tales como la presencia de una falla geológica que puede causar una catarata de agua. Aquí estamos refiriéndonos a los cambios en pendiente observados a 600 m de elevación en El Yunque (Fig. 11.5). El cambio en pendiente se genera por diferencias en la erosión de la roca madre (bedrock), cambios en las tasas de levantamiento del substrato, o tamaño del grano del sedimento en el lecho del río (bedload). También se forman puntos de inflexión temporales al frente de una onda de aceleración de erosión, propagándose río arriba en respuesta a cambios temporales por causas climáticas o tectónicas.
Los ríos reajustaron sus cauces formando puntos de inflexión en la zona de encuentro (alrededor de los 600 m de elevación) entre las rocas geológicamente más recientes y las más antiguas. Las partes altas de las montañas son más viejas y se erosionan con mayor lentitud que las partes bajas más jóvenes (Fig. 11.6). El aumento de la tasa de erosión genera suelos con mayores concentraciones de minerales primarios. Los puntos de inflexión separan suelos con baja concentración de minerales primarios (localizados a mayor elevación) de suelos con alta concentración de minerales primarios (localizados a baja elevación). Esos minerales son la fuente de nutrientes que necesitan las plantas para su productividad, ej., calcio, magnesio, hierro, sodio y potasio. Como las rocas más jóvenes tienen más nutrientes que las más geológicamente antiguas, los bosques formados en asociación a los puntos de inflexión tienen disponibles más nutrientes que los que se forman sobre suelos geológicamente antiguos.
La historia, resumida en los párrafos anteriores, es un área de investigación activa en El Yunque bajo el programa Critical Zone Observatory de la Fundación Nacional de Ciencias de los EE. UU. (https://czo-archive.criticalzone.org/national/). Los científicos utilizan isótopos (10Be con una media vida de 1.39 × 106 años se utiliza para inferir la tasa de erosión de rocas; estroncio [87Sr/86Sr] se utiliza para estudiar el intercambio de calcio en los suelos), cientos de calicatas, perforaciones profundas (30 m), geoquímica, hidrología, microbiología y otros tipos de investigaciones.
11 193 CAP ariel e. lugo | ecología a m i m anera El Yunque
El Toro (1074)Pico del Este (1051)
Figura 11.4. (A) Ubicación de paleo-islas con elevaciones mayores a los 900 m en Puerto Rico incluyendo la paleo-isla El Yunque en las Montañas de Luquillo. La cordillera Central es CC y LM corresponde a las Montañas de Luquillo. La zona de la gran falla del norte es GNFZ por sus siglas en inglés. (B) Detalle de la paleo-isla El Yunque que indica la elevación donde los puntos de inflexión en los ríos Icacos y Sabana reflejan el nivel del mar. Los números identifican la proyección de las pendientes de los picos, por ejemplo el número dos une a El Yunque con Pico del Este y el 3 a El Toro. Noten que la base caliza del Mioceno tenía una elevación superior a la de hoy y similar al de las Montañas de Luquillo. (C) Delimitación de la orilla del mar alrededor de la paleo-isla El Yunque durante el Paleoceno coincidente con los puntos de inflexión en los ríos Sabana (SAB), Icacos (IC) y Cubuy (CUY). El río Mameyes es MAM, el Espíritu Santo es ES y el río Prieto es PRI. El punto inicial del estudio de modelaje es IP por sus siglas en inglés. Modificado de Brocard et al. (2015).
11 194 CAP ariel e. lugo | ecología a m i m anera El Yunque Isla de el Yunque El Toro (1074)Pico del Este (1051) Río Icacos Río Sabana Windgap Punto de inflexión del río 0 1100 1000 900 800 700 600 500 400 300 200 100 0 2 4 6 8 10 12 14 Cresta SE El Yunque (1080) 3 1 4 3 2 2
Plataforma ondulada NW B Distancia (m) Elevación (m) Falla Paleo-islas 700 600 100 200 300 400 500 200 CC San Juan A Elevación de la superficie del envolvente Barranquitas-Cayaguas A LM GNFZ A’ 30 km 900 m 0 m 0 0 C
BasecalizadelMioceno
Isla de el Yunque
Río Icacos Río Sabana Windgap Punto de inflexión del río 0 1100 1000 900 800 700 600 500 400 300 200 100 0 2 4 6 8 10 12 14 Cresta SE El Yunque (1080) 3 1 4 3 2 2
Plataforma ondulada NW B Distancia (m) Elevación (m) Falla Paleo-islas 700 600 100 200 300 400 500 200 CC San Juan A Elevación de la superficie del envolvente Barranquitas-Cayaguas A LM GNFZ A’ 30 km 900 m 0 m 0 0 C
BasecalizadelMioceno
4.5 Ma. En ese punto se desarrolla un punto de inflexión y hoy se observan distintas tasas de erosión: baja tasa de erosión de capas gruesas de saprolita sobre los 600 m y altas tasas de erosión de capas finas de saprolita debajo de los 600 m de elevación. Los lugares de estudio se ubican con estrellas y con varios códigos indicando la cobertura como «palma, CO o TAB». Modificado de Brocard et al. (2015).
11 195 CAP ariel e. lugo | ecología a m i m anera El Yunque 14 Distancia (m) LM A’ 30 km Perfil de tope Perfil de río Diorita de cuarzo Remanente de la plataforma El Y unque El Toro Pico del Este 2 5 MAM 3 3 4 2 CUY La Mina SAB PRI IP IC E5 E7 900 900 700 600 900 700 400 500 Costa - Paleo 3km 0 1 ES C Cresta en el tope Punto de inflexión del río
Figura 11.5. Perfiles de los ríos Sabana (SAB) y Cubuy (CUY) con cambios en sus pendientes coincidentes con la elevación de 600 m donde ubicaba el nivel del mar hace
Río abajo de los puntos de inflexión
TC+VC+SD+QE+T
TC+VC+SD+QE
TC+VC+SD
1995
Figura 11.6. Tasas de erosión aguas arriba y aguas debajo de los puntos de inflexión de los siguientes ríos: Espíritu Santo (ES), Icacos (IC), Prieto (PRI), Sabana (SAB) y Cubuy (CUY). Cada punto representa condiciones distintas en las características del suelo o cobertura. Por ejemplo: suelos finos son «f» y suelos gruesos son «c» (por su sigla en inglés). Los símbolos, explicados en la caja superior izquierda, representan distintas combinaciones de cobertura, incluyendo suelos expuestos. Para detalles del estudio y las condiciones en estos resultados, consulte a Brocard et al. (2015) de donde se obtuvo y se modificó esta ilustración.
La combinación del levantamiento tectónico, la litología, la erosión química (weathering), insumos atmosféricos y los bosques determinan la disponibilidad de cationes minerales en las Montañas de Luquillo. Esta combinación de factores y procesos fue estudiada por Porder et al. (2015) quienes buscaban entender los controles de la disponibilidad de minerales primarios en los suelos de los bosques de El Yunque. Para lograr sus objetivos excavaron 216 calicatas o perfiles de suelos de 0 a 80 cm de profundidad, en dos litologías (volcanoclásticas y dioritas de cuarzo), a lo largo de catenas (valle, ladera y tope) en tres tipos de bosque: palo colorado, tabonuco y palmares. La región con rocas dioritas de cuarzo se dividió en dos subregiones: elevaciones sobre los puntos de inflexión erosionándose más lentamente y elevaciones menores a la de los
11 196 CAP ariel e. lugo | ecología a m i m anera El Yunque
300 250 200 150 100 50 0 Paisaje relicto río arriba C C C C C C C C f f f f f f f f f f f f f f f C C C Sitios de muestreo ESICPRISAB SAB CUY CUY IC Tasa de erosión (m · año)
TC+VC TC NC B,
puntos de inflexión, que respondiendo al levantamiento tectónico hace 4 a 5 Ma, están erosionándose más rápidamente. Los suelos derivados de dioritas de cuarzo bajo los puntos de inflexión han perdido el 80, 65 y 57 % del calcio, magnesio y potasio relativo a la roca madre, mientras que los suelos derivados del mismo material sobre los puntos de inflexión han perdido el 95, 81 y 83 %, respectivamente. En el paisaje volcanoclástico no hay puntos de inflexión y sus suelos han perdido más del 90 % de los cationes en la roca madre.
Bosques de tabonuco tienen más intercambio catiónico que bosques de palo colorado. El tipo de bosque explica el 30 % de la variabilidad en el intercambio de magnesio y potasio y el 10 % de la variabilidad en el intercambio de calcio. Los suelos bajo el punto de inflexión derivados de dioritas de cuarzo tienen el doble del calcio intercambiable que los suelos sobre el punto de inflexión y se duplica de nuevo cerca del punto de inflexión. Ese calcio se deriva de minerales primarios de la erosión de la roca madre, mientras que sobre el punto de inflexión los minerales proceden mayormente de la lluvia y las nubes (de la atmósfera).
Gradientes. La combinación de condiciones climáticas, edáficas, geológicas e hidrológicas de las Montañas de Luquillo da como resultado innumerables gradientes ecológicos que se pueden analizar sobre el gradiente geográfico de la elevación. O sea, se puede proyectar el espacio ecológico sobre el espacio geográfico para visualizar la complejidad estructural y funcional de los ecosistemas de las Montañas de Luquillo. La elevación siempre se ha utilizado como la ordenada al describir los gradientes en las montañas porque es una variable útil e imposible de ignorar. Sin embargo, la relación entre las variables ambientales que afectan la estructura y funcionamiento de los ecosistemas montanos no todas son lineales ni siguen el mismo patrón con la elevación. Por ejemplo, mientras aumenta la lluvia en forma no lineal con la elevación, la temperatura disminuye linealmente (Fig. 11.7), cada uno afectando los procesos ecológicos en direcciones opuestas. Un aumento en la lluvia aumenta la productividad hasta cierto límite, y la reducción en temperatura reduce la tasa de respiración potencialmente limitando la generación de energía de los organismos. Los organismos y los ecosistemas ajustan sus respuestas a los múltiples gradientes altitudinales a los que se enfrentan, optimizando sus respuestas de acuerdo con sus adaptaciones. El resultado es que al organizar características bióticas y ecológicas de acuerdo con la elevación, se observan distintos tipos de funciones como se ilustra en la figura 11.8.
González et al. (2013) resumieron el conocimiento sobre gradientes de elevación en las Montañas de Luquillo. De ese trabajo y de otros anteriores, seleccioné los gradientes principales de acuerdo con parámetros físicos, grupos de organismos, parámetros estructurales de los bosques, procesos ecológicos y tipos de bosques.
11 197 CAP ariel e. lugo | ecología a m i m anera El Yunque
Figura 11.7. Relación entre la precipitación anual y la temperatura promedio anual del aire con la elevación en las Montañas de Luquillo (García Martinó et al. 1996). La identificación de las estaciones de medición es la siguiente: RR = Roosevelt Roads, Faj = Fajardo, Can = Canóvanas, BLT = torre baja en Bisley, EV = El Verde, LM = Montañas de Luquillo, EY = El Yunque y EP = Pico del Este (con nubes). La lluvia anual promedio en mm/año = 1037 (metros sobre el nivel del mar) 0.207; (R 2 = 0.92). La temperatura diaria promedio del aire = 26.51 – (6.69e -3) (metros sobre el nivel del mar); (R 2 = 0.92).
Figura 11.8. Patrones de cambio en las características ambientales con aumento en la elevación en las montañas. Distintos gradientes ambientales explican los cambios observados. (A) patrón estocástico. (B) patrones monotónicos donde la tasa de cambio disminuye, es constante o aumenta con la elevación. (C) patrón asintótico aumentando. (D) patrón de función de paso con cambios abruptos con cambios leves en elevación. (E). patrón de función de paso con amplios ecotonos en elevación. (F) patrones con una moda o una fosa a elevaciones intermedias. Las barras grises reflejan la amplitud del ecotono o interfaz y la línea entrecortada horizontal es el nivel de la asíntota (González et al. 2013).
11 198 CAP ariel e. lugo | ecología a m i m anera El Yunque
y= 3.2903x + 1667.8, R2 = 0.77
Bosques de nubes de las montañas de Luquillo
y= 0.8459x + 1460.3, R2 = 0.2657
Figura 11.9. Precipitación promedio anual en las Montañas de Luquillo y en otras montañas en la cordillera Central de Puerto Rico (González et al. 2013). La relación es más empinada para las Montañas de Luquillo indicando que a una elevación dada, las Montañas de Luquillo reciben más lluvia que las montañas de la cordillera Central.
Parámetros físicos. Anteriormente (Fig. 11.7), había indicado que la lluvia aumenta y la temperatura disminuye con la elevación en las Montañas de Luquillo. El aumento en la lluvia con la elevación en El Yunque es más alto que en las otras montañas de Puerto Rico (Fig. 11.9). Con la elevación disminuye la incidencia de luz solar sobre el dosel de los bosques, en parte por el aumento en la cobertura de nubes. La conductividad de las lluvias disminuye con la elevación (Fig. 11.10), reflejando menos concentración de solutos en el agua de lluvia que cae en las cimas de las montañas. La cobertura del bosque afecta la temperatura del aire y del suelo, que disminuyen con la elevación (Fig. 11.11). Sin embargo, la humedad del suelo aumenta con la elevación. Asociado a ese aumento en la saturación del suelo, disminuye la concentración de oxígeno en los suelos (Fig. 9.6).
11 199 CAP ariel e. lugo | ecología a m i m anera El Yunque
7000 6000 5000 4000 3000 2000 1000 0 0 400 600 800 1000 1200 1400 200 Elevación (m) Promedio de precipitaición anual (mm)
Figura 11.10. Conductividad promedio en la lluvia a lo largo de un transecto con elevación en las Montañas de Luquillo (Medina et al. 2013). Las estaciones están en orden de elevación y se agregaron de acuerdo con los siguientes grupos (por sus siglas en inglés): tierras bajas costaneras (LC), tierras bajas interiores (LI), montano bajo (LM), nivel de formación de nubes (CFL) y nivel alto de formación de nubes (UCFL). Las barras son del error estándar.
Figura 11.11. Comparación en la temperatura y humedad del suelo y la temperatura del aire con el aumento en la elevación con y sin cobertura de bosque (Waide et al. 2013). Las barras son del error estándar.
11 200 CAP ariel e. lugo | ecología a m i m anera El Yunque
Crecropia schreberiana
Dacryodes excelsa
Prestoea montana
Micropholis garciniifolia
Grupos de organismos. La masa y contenido de nitrógeno por metro cuadrado de hojas en El Yunque aumenta con la elevación (Fig. 11.12), pero la concentración de nitrógeno y clorofila en las hojas de las plantas disminuye con la elevación. Las comunidades de gastrópodos (clase de moluscos con conchas) disminuyen en su riqueza y abundancia de especies y aumentan levemente en dominancia y el componente de uniformidad de la diversidad de especies con el aumento en elevación (Fig. 11.13).
El número, abundancia por especies y número de especies de organismos en los tanques de las bromelias disminuyen con la elevación. Sin embargo, la biomasa de
11 201 CAP ariel e. lugo | ecología a m i m anera El Yunque
Figura 11.12. Patrones en las propiedades estructurales de hojas de dosel de cinco especies arbóreas en distintas elevaciones del Bosque Experimental de Luquillo (Harris y Medina 2013). (A) concentración de nitrógeno, (B) concentración de clorofila, (C) contenido de nitrógeno, (D) masa.
Manilkara bidentata
Figura 11.13. Relación entre la elevación y el promedio de cinco características de comunidades de gasterópodos y su abundancia. Los datos se tomaron en dos transectos (bosque de palmeras, símbolos abiertos y bosques mixtos, símbolos sólidos) en el Bosque Experimental de Luquillo (Willig et al. 2013). (A) riqueza de especies, (B) diversidad de Shannon, (C) índice de igualdad de Camargo, (D) dominancia de Berger-Parker, (E) rareza y (F) abundancia. Datos del bosque de tabonuco son los triángulos negros, bosque de palo colorado son los círculos negros y el bosque elfino son los cuadrados negros. Las barras verticales son el error estándar para el promedio por tipo de bosque de los componentes de la biodiversidad. El punto está ubicado a la elevación promedio de los valores para cada bosque.
animales en las bromelias está asociada a la acumulación de materia orgánica en estas plantas y al número de plantas. Estos parámetros aumentan con la elevación (Tablas 11.1 y 11.2). Por lo tanto, la biomasa de animales aumenta con la elevación en los tanques de bromelias. Lo mismo sucede con los invertebrados en la hojarasca de los bosques. La acumulación de nutrientes en las bromelias aumenta con la elevación (Tabla 11.3).
11 202 CAP ariel e. lugo | ecología a m i m anera El Yunque
Abundancia, tamaño y biomasa de animales en bromelias relativo al material orgánico muerto en tres tipos de bosques en el Bosque Experimental de Luquillo. La elevación aumenta entre el bosque de tabonuco, el bosque de palo colorado y el bosque elfino. Modificado de Richardson y Richardson (2013).
Parámetros físicos y biomasa de bromelias en tres tipos de bosque en el Bosque Experimental de Luquillo. La elevación aumenta del bosque de tabonuco al bosque de palo colorado y el bosque elfino. Modificado de Richardson y Richardson (2013).
Almacenaje de nutrientes (kg/ha) en las bromelias de tres bosques del Bosque Experimental de Luquillo. La elevación aumenta desde el bosque de tabonuco al bosque de palo colorado y el bosque elfino. Modificado de Richardson y Richardson (2013). El número en paréntesis representa el porcentaje almacenado en el agua. El trabajo original reporta almacenajes por animales, materia orgánica muerta, plantas y agua. Los porcentajes han sido redondeados.
La diversidad de hongos y su cobertura disminuye con la elevación y es particularmente alta en las tierras bajas (Fig. 11.14). La diversidad de estas comunidades se estima químicamente con distintos marcadores que detectan distintos patrones de abundancia y funcionamiento. Tales técnicas de investigación detectaron bacterias especializadas para funcionar bajo condiciones anaeróbicas tanto en las tierras bajas como en las altas.
11 203 CAP ariel e. lugo | ecología a m i m anera El Yunque
Tabla 11.1
Tabonuco Palo colorado Elfino Organismos/planta 166 141 58 Peso seco animal/planta (mg) 75 113 15 Peso seco del material orgánico muerto/planta (g) 28.1 39.3 4.6 Biomasa animal (g/ha) 3.4 350 480 Masa orgánica muerta total de bromelias (kg/ha) 1.3 122 146 Biomasa animal en por ciento de masa orgánica muerta 0.26 0.29 0.33 Peso promedio de los animales (mg) 0.45 0.80 0.26
Tabla 11.2
Tabonuco Palo colorado Elfino Área/planta (m2) 0.49 0.57 0.08 Hojas verdes/planta 36 29 22 Hojas muertas/planta 8 6 19 Agua (ml/planta) 168 134 103 Agua (Mg/ha) 0.007 0.41 3.3 Densidad de plantas (por ha) 45 3,100 32,000 Peso seco/planta (g) 27.1 35.5 17.2 Materia muerta (g/planta) 28.1 39.3 4.6 Biomasa de bromelias (kg/ha) 1.2 111 550 Biomasa más materia muerta (kg/ha) 2.5 233 696
Tabla 11.3
C N P K Ca Mg S Tabonuco 1.61 (27) 0.05 (45) 0.004 (80) 0.15 (80) 0.04 (70) 0.02 (60) 0.02 (33) Palo colorado 118 (7) 2.95 (12) 0.15 (50) 6.03 (48)2.29 (49) 1.15 (74) 1.28 (35) Elfino 380 (7) 8.35 (24) 0.24 (41) 18.6 (53) 2.91 (65) 2.97 (73) 4.34 (57)
Parámetros estructurales de los bosques. Odum reconoció el cambio en la distribución de raíces y altura de los árboles con la elevación en las Montañas de Luquillo (Fig. 11.15). Asoció esos cambios a la saturación y estructura del suelo y a cambios en el déficit de saturación de la atmósfera. El déficit de saturación atmosférico es la diferencia entre la cantidad de agua en una masa de aire en un momento dado y su capacidad máxima para almacenar agua. Si la masa de aire está saturada al 100 %, su déficit de saturación es nulo, ya que no puede almacenar más agua y, por lo tanto, la evapotranspiración, que es proporcional al déficit de saturación, es igualmente nula. Pero si el déficit es alto, o sea la masa de aire tiene capacidad para absorber agua, las tasas de evapotranspiración son altas siempre y cuando haya agua disponible para evaporar y energía solar para evaporarla. La capacidad de la atmósfera para absorber agua depende de su humedad relativa y la temperatura de la masa de aire. La capacidad de almacenaje aumenta con la temperatura. De acuerdo con Odum, los árboles (bosques) son más altos en las tierras bajas con suelos oxigenados y altos déficits de saturación en la atmósfera y pierden altura cuando el déficit de saturación disminuye. La transpiración de los árboles ayuda al movimiento de minerales del suelo a las hojas,
11 204 CAP ariel e. lugo | ecología a m i m anera El Yunque
Figura 11.14. (A) El cociente hongo:bacteria, (B) la riqueza de especies de hongos mucosos, (C ) la diversidad de Shannon para ácidos grasos microbianos en el suelo y (D) la cobertura de mantos de hongos basidiomicetos a lo largo de la elevación en el Bosque Experimental de Luquillo (Cantrell et al. 2013). Las líneas verticales son el error estándar, letras diferentes indican que los promedios son distintos.
11 205 CAP ariel e. lugo | ecología a m i m anera El Yunque
Figura 11.15. Cambios con la elevación (la elevación aumenta de «A» en las tierras bajas a «C» en las cimas de las montañas) de la estructura del suelo, la ubicación de las raíces grandes y la altura de la vegetación en el Bosque Experimental de Luquillo (Odum 1970).
y la disponibilidad de nutrientes en las hojas (al igual que su enfriamiento) aumenta proporcionalmente con la tasa de transpiración. Por otro lado, suelos saturados tienen menos oxígeno, limitan la respiración de las raíces y afectan su función de extracción de nutrientes. Las raíces en estos ambientes crecen sobre la superficie del terreno en elevaciones altas comparado con su crecimiento profundo en el suelo en elevaciones bajas, donde la concentración del oxígeno es mayor.
Los gradientes estructurales de la vegetación a lo largo de la elevación en las Montañas de Luquillo se ilustran en la figura 11.16. La densidad de árboles aumenta
11 206 CAP ariel e. lugo | ecología a m i m anera El Yunque
Figura 11.16. Cambios con la elevación en parámetros estructurales en los bosques de las Montañas de Luquillo (Brown et al. 1983). El índice de complejidad es el de Holdridge.
con la elevación, mientras que la altura y el diámetro promedio de los árboles disminuyen. El índice de complejidad de Holdridge y el número de especies son más altos a elevaciones intermedias. En capítulos anteriores discutimos que las especies se mueven independientemente de acuerdo con sus límites de tolerancia hacia los factores ambientales. El número de especies y la biomasa disminuyen con la elevación y responden también a las condiciones en las catenas con un mayor número de especies y biomasa en los topes que en los valles (Fig. 11.17 y 11.18). La densidad de tallos en la catena aumenta con la elevación, pero el efecto de la catena es similar, o sea, más densidad en los topes que en los valles (Fig. 11.19).
Tanto la hojarasca sobre el suelo como el almacenaje de carbono y nitrógeno en el perfil del suelo son parámetros que integran los efectos de múltiples factores como la edad del perfil, el clima, la productividad del bosque y la velocidad de descomposición. La cantidad de hojarasca sobre el suelo varía con el gradiente de elevación. El máximo ocurre a elevaciones intermedias en el bosque de tabonuco (Fig. 11.20). El patrón del almacenaje de materia orgánica y el cociente C:N en el perfil del suelo con la elevación es complejo (Fig. 11.21) pero se observan varias tendencias: (1) Con pocas excepciones la capa superior del suelo almacena más
11 207 CAP ariel e. lugo | ecología a m i m anera El Yunque
Figura 11.17. Disminución en la densidad de especies en los bosques con el aumento en elevación en el Bosque Experimental de Luquillo (Weaver y Gould 2013). A cada elevación los bosques en la catena tienen más especies en las cimas (topes) seguido por la pendiente y finalmente el valle (barranco). Las correlaciones son significativas.
Figura 11.18. Disminución en la biomasa de los bosques con el aumento en elevación en el Bosque Experimental de Luquillo (Weaver y Gould 2013). A cada elevación los bosques en la catena tienen más biomasa en las cimas (topes) seguido por la pendiente y finalmente el valle (barranco). Solo la correlación de los barrancos no es significativa.
Figura 11.19. Aumento en la densidad de tallos en los bosques con el aumento en elevación en el Bosque Experimental de Luquillo (Weaver y Gould 2013). A cada elevación los bosques en la catena tienen más densidad de árboles en las cimas (topes) seguido por la pendiente y finalmente el valle (barranco). Los asteriscos (*) indican que la correlación es significativa.
11 208 CAP ariel e. lugo | ecología a m i m anera El Yunque
carbono que las capas más profundas evaluadas individualmente. (2) Los humedales costeros (manglares y bosques de Pterocarpus) son la excepción, pues almacenan carbono en altas cantidades a lo largo de todo el perfil. (3) Cada tipo de bosque en el gradiente altitudinal exhibe variaciones entre distintos rodales del mismo tipo de bosque. (4) Los bosques de tierras bajas (el bosque seco y el bosque húmedo) almacenan menos carbono en el suelo que los bosques a elevaciones intermedias ( bosque de tabonuco) o en elevaciones altas (palmares, bosque de palo colorado y bosque elfino). (5) El bosque de tabonuco y el de palo colorado almacenan cantidades similares de carbono en sus suelos. (6) El almacenaje de carbono en los suelos es mayor en los humedales costeros (manglares y bosques de Pterocarpus) seguido por los humedales en el tope de la montaña (palmares y bosques elfinos). (7) El orden de los bosques en el almacenaje de nitrógeno es el opuesto al orden en el almacenaje de carbono. De mayor a menor almacenaje: bosques secos y húmedos en las tierras bajas, bosques de palo colorado y tabonuco, bosques elfinos y los palmares y bosques de Pterocarpus y manglares.
Procesos ecológicos y tipos de bosques. La tabla 11.4 resume la velocidad de procesos ecológicos en un gradiente de elevación con atención a la contribución de las comunidades de las bromelias. La productividad primaria neta disminuye con la elevación, al igual que la caída de hojarasca. En contraste, la productividad de las bromelias aumenta con la elevación y su importancia relativa a los procesos del bosque aumenta también, llegando al 13 % en el bosque elfino.
11 209 CAP ariel e. lugo | ecología a m i m anera El Yunque
Figura 11.20. Variación en la biomasa de hojarasca reconocible como material vegetal y hojarasca en estado parcial o avanzado de descomposición (duff ) en varios bosques ubicados en distintas elevaciones en las Montañas de Luquillo (González y Luce 2013). Los asteriscos indican que las cantidades de los dos tipos de hojarasca son estadísticamente distintas en magnitud. «NS» significa que no es significativo.
11 210 CAP ariel e. lugo | ecología a m i m anera El Yunque
en distintas elevaciones en las Montañas de Luquillo (Ping et al. 2013). Los datos incluyen el tipo de suelo, la cantidad de carbono orgánico en distintas profundidades, el almacenaje total de carbono orgánico y el cociente carbono:nitrógeno integrado para los suelos del bosque.
La importancia de las bromelias en relación con la velocidad de los procesos ecológicos en tres bosques del Bosque Experimental de Luquillo. La elevación aumenta del bosque de tabonuco al bosque de palo colorado y al bosque elfino. Modificado de Richardson y Richardson (2013).
La respiración del bosque disminuye con la elevación (Fig. 11.22a) y también la productividad primaria bruta (Fig. 11.22b). La proporción de la productividad primaria bruta atribuible a la productividad neta aumenta con la elevación (Fig. 11.22c),
11 211 CAP ariel e. lugo | ecología a m i m anera El Yunque
Figura 11.21. Perfiles de suelo en ocho tipos de bosques
Tabla 11.4
Tabonuco Palo colorado Elfino Productividad primaria neta (Mg/ha·año) 10.5 7.6 3.7 Caída de hojarasca (Mg/ha·año) 8.6 6.8 3.1 Porcentaje de la caída atribuible a las bromelias 0.015 1.79 4.71 Hojarasca en el suelo (Mg/ha) 6 8.76 4.34 Materia muerta de bromelias (g/m2 de bromelias) 57.6 68.9 57.5 Porcentaje de la hojarasca atribuible a las bromelias 0.02 1.4 3.4 Productividad de las bromelias (kg/ha·año) 0.42 38.8 474.1 Porcentaje de la productividad del bosque atribuible a las bromelias 0.004 0.51 12.8
Figura 11.22. Variación con la elevación de (A) la tasa de respiración del suelo, (B) la productividad primaria bruta (GPP), (C) la productividad primaria neta (NPP) y (D) el porcentaje de la productividad bruta atribuida a la productividad neta en el Bosque Experimental de Luquillo (Harris et al. 2013). Los datos están basados en medidas in situ de intercambio gaseoso.
11 212 CAP ariel e. lugo | ecología a m i m anera El Yunque
y ese flujo neto se optimiza a elevaciones intermedias (Fig. 11.22d). Este patrón de comportamiento en el flujo de energía de los bosques en El Yunque está de acuerdo con la ley de máxima potencia propuesta por Odum y Pinkerton (1955) y discutida en el capítulo 2.
Régimen de disturbios
Un disturbio es una interrupción en los procesos ecológicos por una fuerza externa al ecosistema bajo consideración. Los disturbios pueden ser crónicos o agudos de acuerdo con su duración. Los disturbios agudos son eventos de corta duración (horas a meses). Los disturbios crónicos tienen duraciones mayores (años a décadas omás). En décadas pasadas, los disturbios se conocían como estresores (Lugo 1978). En el presente se enfatizan los disturbios extremos debido al efecto sobre los sistemas social-ecológicos-tecnológicos (Lugo 2020). En este capítulo voy a mencionar disturbios meteorológicos (huracanes y sequías), geológicos (derrumbes), hidrológicos (descarga de ríos) y antropogénicos (uso de la tierra). El cambio climático se discutió en el capítulo 4 y arriba mencioné los disturbios tectónicos. El conjunto de disturbios y sus características constituyen el régimen de disturbios de un lugar, en este caso de El Yunque.
Al analizar un disturbio es importante conocer su intensidad, frecuencia de recurrencia, duración y extensión espacial. Además, algunos disturbios como los huracanes están compuestos de varias fuerzas que afectan los ecosistemas individualmente y en sinergia, o sea que la interacción de dos fuerzas es más que la sumatoria de su magnitud, puede ser multiplicativa. Finalmente, es importante identificar para cada disturbio y para cada una de sus fuerzas asociadas, la parte del ecosistema más afectada por la fuerza externa. Por ejemplo, el viento afecta principalmente el dosel del bosque, mientras que los derrumbes afectan los suelos.
En esta sección no discuto en detalle la respuesta de los sistemas ecológicos a los disturbios, ya que estas se discutirán en los capítulos donde se presentan tipos de bosques específicos o en discusiones de cómo los bosques de Puerto Rico reflejan el régimen de disturbio histórico que han experimentado. Brokaw et al. (2012a) contiene un listado de los disturbios que afectan los bosques de El Yunque y la respuesta de organismos y sistemas ecológicos a esos disturbios.
Huracanes. El huracán es el disturbio principal de los bosques de El Yunque, de Puerto Rico y el Caribe. La tabla 11.5 contiene los eventos climáticos e hidrológicos asociados a los huracanes y su relativa intensidad y recurrencia durante el huracán Hugo. Es evidente que la intensidad de las lluvias durante el huracán, relativo a las lluvias durante periodos de ondas tropicales, fue baja, mientras que la intensidad de los vientos fue alta. Cada elemento del huracán afectó un sector distinto del bosque,
11 213 CAP ariel e. lugo | ecología a mi manera El Yunque
Tabla 11.5
Distintas medidas para evaluar las características de un huracán utilizando datos del huracán Hugo (Scatena y Larsen 1991, Lugo 2008). El huracán pasó sobre las Montañas de Luquillo como un huracán de categoría 3, una recurrencia de 60 años y duró cuatro horas generando 285 derrumbes. Las unidades en paréntesis corresponden a la columna de intensidad. Celdas en blanco indican falta de información.
por lo que los efectos de los huracanes en los bosques tienen que analizarse desde el punto de vista de la complejidad de sus interacciones en el tiempo y espacio, además de su intensidad y duración.
Sequías. Puerto Rico está sujeto a sequías periódicas, las cuales se intensifican durante los eventos fuertes de El Niño en el Pacífico. La figura 11.23 ilustra el efecto de una sequía en la humedad de los suelos de una catena en el sector El Verde de El Yunque. La pérdida de humedad a medida que progresaba la sequía revela dos asuntos: (1) La capa superior del suelo mantuvo más humedad que las otras capas del suelo y no sufrió el descenso rápido observado en las capas inferiores. (2) A pesar de la sequía, se mantuvo el gradiente de humedad entre el tope, pendiente y valle de la catena. La sequía tuvo mucha influencia en el funcionamiento de los bosques, en este caso, el funcionamiento de los microorganismos responsables por el reciclaje de nutrientes y la respiración edáfica. Paradójicamente, la sequía aumentó el oxígeno disponible, ya que al reducir la humedad, aumenta la capacidad del suelo para retener el oxígeno. El resultado es que con más oxígeno en el suelo aumentan las tasas de respiración y reciclaje de los suelos. Durante la sequía de 1994, un estudio de campo realizado por Ernesto Medina sobre la condición hídrica de los árboles encontró que los efectos eran menores, sugiriendo adaptaciones a las sequías de parte de los árboles en El Yunque.
Derrumbes. La susceptibilidad a derrumbes se debe a la geología y a la intensidad de las lluvias. La región con rocas plutónicas y suelos diorita en altas elevaciones es la más propensa a derrumbes (Guariguata 1990, Guariguata y Larsen 1990). Los derrumbes ocurren cuando la intensidad de las lluvias excede un umbral, luego de que los suelos se han saturado. Los derrumbes ocurren sobre todas las geologías de las Montañas de Luquillo, pero son más abundantes en la geología de rocas dioritas de cuarzo y a elevaciones superiores a los 600 m (Fig. 11.24).
11 214 CAP ariel e. lugo | ecología a m i m anera El Yunque
Componente Intensidad Frecuencia (años) Duración (h) Escala espacial (km2) Punto de interacción con el bosque Lluvia (mm) 339 5 <4 Cientos Dosel y suelo Viento (km/h) 148 100 4 Cientos Dosel Descarga pico de ríos (m3/s) 70-666 10-31 12-24 Décimas Cause del río Presión barométrica (mbs) 946.2 <4 Cientos Hojas
Cambio en cobertura y uso de la tierra. En el paisaje, la mayor actividad antropogénica en el noreste de Puerto Rico ocurre fuera de las Montañas de Luquillo, particularmente a elevaciones menores a 500 m (Fig. 11.25). Estudios en la región de El Verde descubrieron que tras de la deforestación, uso y abandono de los terrenos, la composición de especies del bosque cambió y luego de más de 70 años no ha retornado a la composición original (Thompson et al. 2002). Imágenes térmicas, demuestran que los bosques operan a temperaturas más bajas que las regiones urbanizadas en las tierras bajas donde la actividad humana es mayor (Fig. 11.26). La distribución de temperatura entre estos dos sectores (Fig. 11.27) ilustra el calentamiento en las tierras bajas, el cual afecta la formación de nubes en las laderas de las montañas. El aire caliente en las tierras bajas necesita más elevación para enfriarse y por eso las nubes se forman a elevaciones más altas de lo normal, potencialmente disminuyendo la humedad que reciben los bosques.
11 215 CAP ariel e. lugo | ecología a m i m anera El Yunque
Figura 11.23. Contenido volumétrico de agua en los suelos del bosque de tabonuco en El Verde, Bosque Experimental de Luquillo (Cortesía de Whendee Silver). La saturación es 1.0 y los datos se tomaron en una catena desde el tope al valle con los números identificando medidas en estaciones entre el tope y el valle. El récord entre noviembre 14, 2014 hasta septiembre 7, 2015 refleja un evento de sequía.
con la elevación en las Montañas de Luquillo de los deslizamientos de terreno causados por tres eventos de lluvia entre el 2003 y 2004 (Shiels y Walker 2013). El 80 % del área de las Montañas de Luquillo ocurre entre 200 y 800 m de elevación. Es difícil segregar el efecto de los suelos y de la elevación, ya que la mayor parte de los deslices de terreno ocurren en los suelos dioríticos y estos ocurren entre 600 y 900 m de elevación.
11.24.
11.25. Variación con la elevación de las Montañas de Luquillo del área de cada banda de elevación y el porcentaje de cobertura de pastizales, áreas desarrolladas, arboladas y matorrales y bosques cerrados (Weaver y Gould 2013).
11 216 CAP ariel e. lugo | ecología a m i m anera El Yunque
Figura
Distribución
Figura
Biota y sistemas ecológicos. El inventario biótico y ecológico de las Montañas de Luquillo es impresionante y se puede resumir de la siguiente manera: 225 especies de árboles, 16 especies de anfibios, 18 especies de reptiles, 16 especies de mamíferos, 150 especies de aves residentes y migratorias, más de 350 especies de hongos, 98 especies raras y en peligro de extinción y 23 especies de plantas endémicas. Los múltiples gradientes bióticos y abióticos en estas montañas permiten que muchas especies y taxones encuentren condiciones favorables de crecimiento y, como resultado,

11 217 CAP ariel e. lugo | ecología a m i m anera El Yunque
Figura 11.26. Imagen termal del sensor ATLAS de la región noreste de Puerto Rico con 10 metros de resolución (Waide et al. 2013). La imagen se adquirió como parte de una misión el 13 de febrero de 2004. Mientras más cálidos los colores, mayor es la temperatura de la superficie en grados centígrados.
ras en las superficies de los bosques en El Verde y de las áreas urbanizadas alrededor de las Montañas de Luquillo (Waide et al. 2013). Los valores promedio para la cobertura de bosque son de 26 oC y llegaron hasta 42.7 oC en asfalto y sobre 53.5 oC en los techos.
Tabla
Comparación de la distribución de las especies arbóreas de las Montañas de Luquillo (Little 1970). Basado en 207 especies arbóreas nativas en 133 géneros y 55 familias en el Bosque Experimental de Luquillo. Hay además 50 especies introducidas.
82 especies comunes con los continentes
Puerto Rico a Suramérica y América Central (incluye México y Florida): 53 especies
Puerto Rico a Suramérica incluso Panamá: 23 especies
Puerto Rico y Centro América o México o Florida: 6 especies
Indias Occidentales pero no en continentes: 64 especies
Antillas Mayores y Antillas Menores: 31 especies
Antillas Mayores solamente: 27 especies
Puerto Rico y Antillas Menores solamente: 6 especies
Endémicas de Puerto Rico o Islas Vírgenes: 61 especies
Puerto Rico e Islas Vírgenes solamente: 3 especies
Puerto Rico solamente: 32 especies
Montañas de Luquillo solamente: 26 especies
Montañas de Luquillo vs. La Española (República Dominicana y Haití): 122 especies
Montañas de Luquillo vs. Cuba: 105 especies
Montañas de Luquillo vs. Suramérica: 76 especies
Montañas de Luquillo vs. Venezuela: 65 especies
Montañas de Luquillo vs. América Central: 59 especies
Montañas de Luquillo vs. México: 44 especies
Montañas de Luquillo vs. Florida: 7 especies
Montañas de Luquillo vs. el Viejo Mundo: 4 especies
11 218 CAP ariel e. lugo | ecología a m i m anera El Yunque
11.6
Figura 11.27. Distribución de temperatu
muchos de estos grupos de organismos exhiben sus máximas expresiones aquí. La tabla 11.6 compara la riqueza taxonómica y distribución de los árboles con otros lugares en Puerto Rico, el Caribe, Centro y Suramérica. Los datos comparativos reflejan la afinidad de los árboles en las Montañas de Luquillo con otras regiones del Neotrópico, particularmente con otras Antillas Mayores y Suramérica. Cuatro especies del viejo mundo tropical ocurren en las Montañas de Luquillo. El análisis se puede repetir con otros grupos de organismos con resultados similares.
¿Es o no El Yunque una maravilla natural del planeta?
La respuesta es sí y me explico (Lugo 2005b). Los estudios modernos de ecología no dejan duda de que cada hectárea del planeta es única en el mismo sentido en que cada individuo de una población es único porque tiene su propia genética, historial de desarrollo y comportamiento. Las Montañas de Luquillo, uno de los lugares más húmedos en la región, representan el Caribe insular lluvioso expuesto periódicamente a huracanes. Ante estas condiciones que incluyen 4,000 años de presencia humana y más de 40 Ma de desarrollo tectónico, la biodiversidad en las Montañas de Luquillo adquiere exuberancia. Lo maravilloso se desprende del conocimiento que tenemos de esta expresión de la vida, los múltiples gradientes en el tiempo y espacio, la resiliencia a los disturbios y el historial de cambio a través de los milenios. El Yunque ha sido y seguirá siendo un sitio especial para las muchas generaciones humanas que lo han observado, vivido, atravesado, estudiado, utilizado, respetado y adorado por milenios y quizás para siempre.
Asignación
Selecciona cualquier zona de vida dentro de los límites geográficos de El Yunque y preséntale a la clase cinco atributos de los sistemas ecológicos en esa zona de vida.
11 219 CAP ariel e. lugo | ecología a m i m anera El Yunque
CaPítulo 12 Ecología de plantaciones arbóreas tropicales
Pregunta
¿Cómo comparas una plantación de caoba con un bosque de caoba?
Introducción
En este capítulo presento una introducción global de las plantaciones arbóreas y luego menciono las tropicales. Como parte de esta introducción, resumo algunas definiciones importantes y presento la historia de vida de una plantación. Luego, examino aspectos ecológicos de las plantaciones tropicales basados en estudios en Puerto Rico y termino con una discusión sobre su uso para rescatar tierras degradadas.
Los bosques plantados
Es un reto determinar qué es una plantación y cómo diferenciarla de otro tipo de bosque bajo manejo intensivo. Para Evans (1992), lo que delimita la plantación es la regeneración artificial. Él definió la plantación como un cultivo forestal o rodal arbóreo cultivado artificialmente, ya sea utilizando semillas o la siembra de plántulas o esquejes. Dentro de esta definición de plantaciones Evans incluye la aforestación o la siembra de árboles donde no han existido por 50 años y la reforestación o la siembra de árboles donde han existido durante los 50 años previos. La reforestación incluye siembra de árboles que reemplazan vegetación distinta a la sembrada o siembra de árboles que reemplazan árboles similares a los que se siembran. Un bosque que se regenera sin ayuda humana, con o sin manejo intensivo post-regeneración, no cuenta como plantación.
Las estadísticas globales sobre bosques y plantaciones están centralizadas en la Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO, por sus siglas en inglés). La FAO utiliza una nomenclatura estándar para recopilar y reportar los datos globales o por países. La categorización de los bosques plantados usada por la FAO (2007) utiliza alguna terminología (ej. naturales, seminaturales, etc.) que he descartado por implicar que la actividad humana no es parte de la naturaleza.
ariel e. lugo | ecología a m i m anera
221
Tabla 12.1
Bosques plantados en el conjunto de características forestales (basado en la FAO 2007). Bosques regenerados naturalmente Bosques plantados Árboles fuera del bosque Primarios Naturales modificados
Bosques de especies indígenas regeneradas de manera natural en donde existen indicaciones evidentes de actividad humana
Prácticas silvícolas parte de una ordenación intensiva:
• Deshierbe
• Fertilizantes
• Aclareo
• Corte selectivo
Bosques de especies indígenas establecidos mediante la plantación o siembra y de ordenación intensiva
Bosques de especies introducidas o indígenas establecidos mediante la plantación o siembra principalmente para producir madera o productos forestales no madereros
Seminaturales Plantaciones Regeneración natural asistida Componente plantado Para la producción Para la protección Bosques de especies indígenas en donde no existe una indicación claramente visible de actividades humanas y en donde los procesos ecológicos no son alterados de manera significativa
Bosques de especies introducidas o indígenas establecidos mediante la plantación o siembra principalmente para suministrar servicios
Formaciones inferiores a 0.5 ha; cubierta forestal de los terrenos agrícolas (sistemas agroforestales, huertos familiares, huertas); árboles en ambiente urbano; y a lo largo de las carreteras y dispersos en el paisaje
Sin embargo, por el valor que tiene la estandarización en las estadísticas globales, reproduzco la tabla utilizada por esta organización (Tabla 12.1). La FAO considera como bosques regenerados sin ayuda humana a los bosques primarios, los secundarios que han sido modificados por la actividad humana, y aquellos que reciben cierto tipo de subsidio humano, pero mantienen su capacidad de regeneración sin asistencia humana.
Este último tipo de bosque puede tener un componente de regeneración asistido por el ser humano, por lo que la FAO lo considera dentro del grupo de las plantaciones. Las plantaciones se subdividen en las que se utilizan para la producción de productos forestales o las que se utilizan para la protección de los suelos, la biodiversidad, u otros servicios ecológicos. La selección de especies puede ser de especies nativas o no nativas (introducidas). Los árboles fuera del bosque tienen menor área de cobertura en comparación a los anteriores y pueden ocurrir a lo largo de caminos, en áreas agrícolas, o dispersos en el paisaje. En Puerto Rico contamos con muchos fragmentos de tierra con cobertura de bosque que serían clasificados por la FAO como árboles fuera del bosque. Estas arboladas son importantes para la isla por ser pequeña, pero son insignificantes a nivel global.
La historia de vida de una plantación refleja que el proceso para establecerla, cultivarla y cosecharla es complejo y costoso y presenta numerosos retos técnicos (Fig. 12.1). Además, es un proceso que dura décadas y, por lo tanto, requiere planificación, organización, alta capacidad técnica y perseverancia. La columna que identifica las preguntas principales que se deben contestar en cada etapa de la vida de una plantación establece claramente que múltiples especialidades técnicas
12 222 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
Figura 12.1. Diagrama de flujo que describe la secuencia de las operaciones silviculturales, decisiones principales y el tiempo asociado a las operaciones durante las etapas de siembra, cultivo, cosecha y resiembra de plantaciones arbóreas. Este diagrama lo adaptó Ruiz (2002) de Evans (1992). Las referencias a capítulos se refieren al manual editado por Ruiz (2002).
12 223 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
adquieren inherencia en el proceso de establecimiento y manejo. Someramente, se pueden identificar las siguientes: taxonomía, fisiología, ecofisiología, botánica, zoología, ecología, genética, edafología, ingeniería, química, geoquímica, climatología, dasonomía, silvicultura y muchas más. Por todo lo anterior, el establecimiento, manejo, cosecha y regeneración de plantaciones arbóreas es una actividad que requiere conocimiento y atención profesional, que correctamente aplicado puede garantizar la sostenibilidad de la operación sin necesariamente afectar negativamente la productividad de los suelos.
Área de plantaciones
En el 2005, el área global, excluyendo el componente plantado de los bosques seminaturales en la tabla 12.1, era de aproximadamente 225,000 millones de hectáreas, a los que se puede sumar un 47 % por el componente plantado de los bosques seminaturales. Asia y Europa son los continentes con mayor área de plantaciones y los 10 países con la mayor área son (en orden): China, India, EE. UU., Rusia, Japón, Suecia, Polonia, Sudán, Brasil y Finlandia. Las plantaciones para producir madera son más comunes que para la protección y suministro de servicios ecológicos. Los pinos representan el grupo de árboles más comúnmente utilizados para plantaciones en el mundo (Fig. 12.2). Es notable que la mayor parte de las plantaciones arbóreas estén compuestas por un grupo reducido de especies. Esto debido a que se utilizan especies cuyas características y técnicas de producción son conocidas y, por lo tanto, ameritan la inversión de tiempo, energía y dinero requerido para su cultivo.
Evans (1992) estimó que para el 2000 las plantaciones tropicales cubrirían más de 60 millones de hectáreas y hoy se estiman en 100 millones de hectáreas, muy por debajo del área en otras regiones del planeta. Esto se debe a la falta de conocimiento técnico y las dificultades de controlar el crecimiento de los árboles en los trópicos.
Las primeras plantaciones tropicales se remontan al siglo XIX, pero no fue hasta la década de 1960 que el área de plantaciones tropicales comenzó a ascender exponencialmente (Evans 1992). En Puerto Rico se establecieron plantaciones en la década de 1930 y durante mediados del siglo XX se llevaron a cabo investigaciones para identificar las especies y las técnicas de silvicultura necesarias para establecerlas bajo las condiciones tropicales de Puerto Rico. Las investigaciones sobre bosques neotropicales comenzaron en Puerto Rico de manera temprana (Lugo 2021), lo que permitió experimentar con plantaciones antes que otros países de la región. Esto convirtió a Puerto Rico en un líder mundial en las investigaciones de plantaciones tropicales (Francis 1995). Más de 300 especies nativas e introducidas han sido plantadas y evaluadas en la isla (Wadsworth 1995). La mayor parte de estas fracasaron, pero se identificaron unas 30 especies útiles para plantar bajo distintos climas y suelos en Puerto Rico.
12 224 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
porcentajes están basados en 132,095,000 ha de plantaciones para la producción y 49,597,000 ha de plantaciones para la protección para un total de 181,693,000 ha de plantaciones distribuidas entre 10 países (China, India, Estados Unidos de América, Federación de Rusia, Japón, Suecia, Polonia, Sudán, Brasil y Finlandia; en orden de cobertura de plantaciones arbóreas).
12 225 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
Figura 12.2. Especies utilizadas en plantaciones con propósitos de producción de madera (A) y protección ambiental (B) y el porcentaje de la superficie cubierta por género a nivel global en el 2005 (FAO 2007). Los
Se debe reconocer la contribución del utuadeño José Marrero Torrado, agrónomo, silvicultor y empleado del Instituto de Dasonomía Tropical, quien el Dr. Frank H. Wadsworth consideró el regalo de Puerto Rico a los árboles (Wadsworth 2007). José Marrero estableció más plantaciones en Puerto Rico que ninguna otra persona u organización. Su esfuerzo se concentró en rehabilitar tierras degradadas en la isla (Marrero 1950).
Controversias
Las plantaciones han sido controversiales debido a una variedad de razones sociales, económicas y ecológicas, pero fundamentalmente por una falta de confianza y comunicación entre ecólogos, dasónomos y conservacionistas. El establecimiento de monocultivos de especies introducidas, el alto costo de establecimiento y su mantenimiento, el área de cobertura dedicado a las plantaciones, el consumo de agua por algunas de las especies utilizadas y los efectos que tienen las plantaciones sobre la biodiversidad son algunas de las objeciones levantadas en contra del establecimiento de plantaciones arbóreas en los trópicos.
Algunas de las controversias no están basadas en información confiable o se basan en prejuicios en contra de monocultivos o especies introducidas. Cuando es necesario intensificar o maximizar la producción primaria neta, ya sea para producir alimentos o madera, el monocultivo, generalmente de especies introducidas, es el mecanismo más costo-efectivo. De igual manera, cuando se maximiza la fotosíntesis en las plantas y se obtiene la máxima productividad, el consumo de agua, bióxido de carbono y nutrientes también se acelera. Ambas generalizaciones responden a las leyes físicas y químicas que regulan el funcionamiento de los ecosistemas. Sin embargo, es posible que algunas plantaciones utilicen menos agua, sean más eficientes en su uso y produzcan más materia orgánica que los bosques con especies nativas (Tabla 12.2). El ser humano puede manejar la situación de acuerdo con las circunstancias locales para optimizar el uso de recursos y el funcionamiento de los sistemas ecológicos, evitando así la degradación y asegurando cierto grado de sustentabilidad.
Un ejemplo particularmente notorio es la discusión internacional sobre el comercio de una de las tres especies designadas como «caoba». Estas especies pertenecen al género neotropical Swietenia (S. macrophylla, S. mahagoni y S. humilis). La controversia reciente fue con la caoba de hoja grande (S. macrophylla) oriunda de Centro y Suramérica. Organizaciones internacionales propusieron incluir esta caoba en el apéndice 1 de la Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres (CITES, por sus siglas en inglés). La designación asumía que la caoba es una especie en peligro de extinción, y como tal, amerita la mejor protección disponible. Tal designación hubiese paralizado el comercio internacional de caoba.
12 226 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
Tabla 12.2
Uso de agua promedio, fijación de carbono y eficiencia en el uso del agua en bosques secundarios y plantaciones de teca (Tectona grandis). El error estándar está en paréntesis (n = 6). Los valores en negritas son estadísticamente distintos a p = .0039. Basado en Abelleira Martínez (2015).
La caoba se caracteriza por su madera de enorme valor económico que la hace una de las principales especies tropicales en el comercio maderero internacional. Su remoción del mercado afectaba la economía de naciones tropicales. Cuando analicé la situación en detalle (Lugo 1999b) me di cuenta de que la acción propuesta estaba basada en información incompleta o errónea sobre la caoba, una especie que no estaba en peligro de extinción. Más aún, la caoba crece bien en plantaciones y es posible reducir la presión en las poblaciones silvestres produciendo madera. La caoba fue finalmente incluida en el apéndice 2 de CITES, el cual regula el comercio requiriendo certificaciones y análisis de las poblaciones de caoba de donde procede la madera antes de autorizar el comercio internacional. Las investigaciones en Puerto Rico, resumidas en un libro (Lugo et al. 2003), fueron fundamentales en clarificar la ciencia que apoya la conservación de la caoba.
Otro ejemplo en el ámbito local fue el estudio que llevé a cabo (Lugo 1992) en respuesta a alegatos de la Asociación Médica de Puerto Rico en contra de las plantaciones de pino. Alegaron que las plantaciones de pino acidificaban el suelo, reducían la biodiversidad y evitaban la regeneración de especies nativas. Los alegatos, basados en la ecología de plantaciones de pino en zonas templadas, resultaron ser infundados. Encontré que en Puerto Rico, las plantaciones de pino mejoran la condición del suelo incrementando la materia orgánica y la concentración y cantidad de nutrientes. La acumulación de hojarasca ayuda a reducir la erosión del suelo y crea condiciones donde las lombrices de tierra y otros organismos del suelo abundan. Más aún, las plantaciones de pino (Cruz 1988) y las de caoba (Cruz 1987) atraen aves y otros animales que ayudan con la dispersión de las plantas que germinan y crecen en el sotobosque. La riqueza de especies nativas en plantaciones de pino aumenta con la edad de la plantación, razón por la cual las plantaciones de pino desatendidas dan paso a bosques secundarios dominados por especies nativas.
El establecimiento de plantaciones es un uso legítimo de los suelos y, cuando se manejan adecuadamente, contribuyen positivamente a la economía y la conservación, siendo la forma más efectiva disponible para producir productos madereros de
12 227 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
Parámetro y unidad Bosque secundario Plantación de teca Uso de agua durante temporada de lluvia (kg/ha·día) 15,488 (3450) 9,183 (1351) Uso anual de agua (Mg/ha·año) 4,400 (978) 2,514 (370) Fijación de carbono (kg/ha·año) 2,281 (745) 4,007 (948) Eficiencia en el uso del agua (kg C/Mg H2O 0.495 (0.062) 1.700 (0.340)
calidad conocida y a tasas altas de productividad. Múltiples sectores económicos se benefician de plantaciones, lo que incluye las industrias de madera, papel, celulosa, muebles y energía.
La influencia de las especies en el funcionamiento de la plantación
En el Arboretum de Ciénaga Alta en el Bosque Experimental de Luquillo se establecieron plantaciones de múltiples especies en bloques monoespecíficos (Fig. 12.3). La presencia de plantaciones maduras de las especies comúnmente utilizadas en los trópicos brindó la oportunidad única de estudiar los contrastes y las similitudes en el comportamiento ecológico y ecofisiológico de las especies cuando están expuestas a las mismas condiciones de suelo y clima. A continuación, analizo algunos aspectos ecofisiológicos que, en conjunto, aportan información sobre los mecanismos que regulan la dinámica del carbono y los nutrientes en bosques tropicales
Lugo et al. (1990b) encontraron que la acumulación de hojarasca en esas plantaciones variaba con la especie (Fig. 12.4). Las de pino acumulaban mucha más hojarasca que las plantaciones de especies nativas como Hernandia sonora. Sin embargo, debido a diferencias en la concentración de nutrientes en la hojarasca de las distintas especies, la acumulación de nutrientes no seguía el mismo patrón que la acumulación de biomasa y variaba con el nutriente (Fig. 12.5). La masa de hojarasca acumulada es inversa o negativamente relacionada con la concentración de los nutrientes en la hojarasca (Fig. 12.6). Lo mismo ocurre con la masa de madera en el suelo. Más aún, la concentración de nutrientes varía con el estado de la hojarasca durante el proceso de descomposición (recién caídas, fragmentadas o viejas; Fig. 12.7). Con el tiempo de descomposición, las concentraciones de nutrientes tienden a aumentar, siendo el potasio una excepción porque está sujeto a lixiviación o lavado por las aguas de lluvia.
Los resultados sobre el almacenaje de hojarasca indican que las plantaciones tienden a acumular masa y nutrientes en la hojarasca en contraste con bosques primarios y secundarios que acumulan menos. La tendencia de acumulación de materia orgánica y nutrientes en las plantaciones contribuye a rehabilitar suelos degradados, una función en que son efectivas. La acumulación de hojarasca en proporción a la concentración de nutrientes resalta dos aspectos del funcionamiento de estos sistemas. (1) Hojarasca con baja concentración de nutrientes sugiere alta eficiencia en el uso de los nutrientes, ya que presumiblemente los nutrientes en las hojas vivas se reciclaron en el árbol antes de la caída (proceso de retranslocación). (2) Hojarasca con baja concentración de nutrientes es menos apetecible a los microorganismos que conducen la descomposición en el bosque, y como consecuencia la velocidad de descomposición es inicialmente más lenta, lo que causa la acumulación de hojarasca. Sin embargo, a medida que la hojarasca se descompone
12 228 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
et al. 1990b: 1 Pinus caribaea, 2 Hibiscus elatus, 3 Pinus elliottii, 5 Eucalyptus x patentinervis, 9 Khaya nyasica, 12 Hernandia sonora, 16 Anthocephalus chinensis , 17 Eucalyptus saligna




(fragmentándose y envejeciendo en su apariencia), la calidad nutricional aumenta, según se observa en la figura 12.7. Esto beneficia a la cadena alimenticia del suelo. En un perfil de hojarasca, las hojas en la superficie son las más recientes en caer y tienen una menor calidad nutricional, las hojas en el fondo del perfil, llevan más

12 229 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
Figura 12.3. Mapa del arboreto del Instituto Internacional de Dasonomía Tropical ubicado en el sector de Ciénaga Alta en el Bosque Experimental de Luquillo. Cada cuadrito representa un rodal de una especie arbórea. Los cuadritos punteados representan la ubicación de las siguientes especies estudiadas por Lugo
, 19 Swietenia macrophylla, 53 Terminalia ivorensis
Figura 12.4. Masa total en la hojarasca de 10 plantaciones en el arboreto de Ciénaga Alta en el Bosque Experimental de Luquillo (Lugo et al. 1990b). Las especies están en orden de mayor a menor masa acumulada en el suelo y las abreviaciones corresponden a: Pinus caribaea (Pc), Hibiscus elatus (He), Pinus elliottii (Pe), Eucalyptus x patentinervis (Ep), Khaya nyasica (Kn) Hernandia sonora (Hs), Anthocephalus chinensis (Ac), Eucalyptus saligna (Es), Swietenia macrophylla (Sm) y Terminalia ivorensis (Ti). Los números bajo las especies significan el rango de productividad de hojarasca entre las especies del estudio y los asteriscos identifican las tres especies más productivas. Anthocephalus chinensis fue la menos productiva. Las barras verticales representan el error estándar del promedio basado en 10 réplicas.
tiempo descomponiéndose (perdiendo carbono) y tienen una calidad nutricional mayor. Generalmente, las raíces las invaden para reciclar los nutrientes al árbol. El mecanismo del enriquecimiento de esas hojas es similar a lo que ocurre con el detrito de los manglares discutido en el capítulo 10. Los cuerpos de los microorganismos descomponiendo las hojas fragmentadas enriquecen nutricionalmente a los fragmentos de hojas en descomposición en el fondo de la hojarasca.
En el mismo lugar y con las mismas especies, Cuevas y Lugo (1998) estudiaron la caída de hojarasca. El orden de las especies en la producción de hojarasca no fue el mismo al orden de las especies en el almacenaje de hojarasca (comparen la figura 12.4 con la figura 12.8). Hubo tres resultados notables en la comparación entre caída y almacenaje de materia orgánica en estas plantaciones. (1) Anthocephalus chinensis mostró la menor tasa de caída a pesar de estar cerca del promedio en el almacenaje
12 230 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
corresponden a: Pinus caribaea (Pc), Hibiscus elatus (He), Pinus elliottii (Pe), Eucalyptus x patentinervis (Ep), Khaya nyasica (Kn) Hernandia sonora (Hs), Anthocephalus chinensis (Ac), Eucalyptus saligna (Es), Swietenia macrophylla (Sm) y Terminalia ivorensis (Ti). Las barras verticales representan el error estándar del promedio basado en 10 réplicas.
12 231 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
Figura 12.5. Almacenaje de cinco macronutrientes en la hojarasca de diez plantaciones en el arboreto de Ciénaga Alta en el Bosque Experimental de Luquillo (Lugo et al. 1990b). Las especies están en orden de mayor a menor masa acumulada en el suelo y las abreviaciones
Figura 12.6. Relación entre la masa de hojarasca en el suelo de 10 plantaciones y la concentración en la hojarasca de tres macronutrientes: nitrógeno (A), potasio (B) y magnesio (C; Lugo et al. 1990b). Las plantaciones estaban ubicadas en el arboreto de Ciénega Alta en el Bosque Experimental de Luquillo. Los promedios están basados en 10 réplicas y las barras verticales representan el error estándar. La escala de concentración de nutrientes varía con el nutriente.
12 232 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
Figura 12.7. Concentración de cinco macronutrientes en la hojarasca de 10 plantaciones en el arboreto de Ciénaga Alta, Bosque Experimental de Luquillo: A - nitrógeno, B - fósforo, C- potasio, D - calcio y E - magnesio (Lugo et al. 1990b). Se presentan resultados de hojas recién caídas, hojas viejas y hojas fragmentadas. Las especies están en orden de mayor a menor masa acumulada en el suelo y las abreviaciones corresponden
a: Pinus caribaea (Pc), Hibiscus elatus (He), Pinus elliottii (Pe), Eucalyptus x patentinervis (Ep), Khaya nyasica (Kn) Hernandia sonora (Hs), Anthocephalus chinensis (Ac), Eucalyptus saligna (Es), Swietenia macrophylla (Sm) y Terminalia ivorensis (Ti). Los puntos están conectados para facilitar la interpretación. La escala de concentración de nutrientes varía con el nutriente.
12 233 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
12.8.
total que está subdivida en cinco componentes (hojas de la especie plantada, hojas de otras especies, madera de diámetro pequeño, partes reproductivas, y miscelánea). Barras con la misma letra no son significativamente distintas entre sí. Las abreviaciones de las especies corresponden a: Pinus caribaea (Pc), Hibiscus elatus (He), Pinus elliottii (Pe), Eucalyptus x patentinervis (Ep), Khaya nyasica (Kn) Hernandia sonora (Hs), Anthocephalus chinensis (Ac), Eucalyptus saligna (Es), Swietenia macrophylla (Sm) y Terminalia ivorensis (Ti).
de la
de hojarasca. (2) Hibiscus elatus y Eucalyptus saligna junto a Pinus caribaea exhibieron las tasas de caída más rápidas. (3) La especie nativa, Hernandia sonora con una tasa de caída baja similar a Anthocephalus chinensis, Swietenia macrophylla y Terminalia ivorensis. Estas diferencias destacan el hecho de que están en juego dos procesos distintos (producción y descomposición de hojarasca) y las especies exhiben distintas estrategias o patrones de comportamiento para cada uno. Igualmente, las especies exhibieron distintas fenologías en la caída de hojarasca a pesar de encontrarse en las mismas condiciones de clima. La fenología, un proceso interno de cada especie, es otro ejemplo de las adaptaciones de las especies a distintos regímenes ambientales y, por lo tanto, se observa la aceleración (aumento en caída) o la reducción en la caída de hojarasca en distintos meses del año (Fig. 12.9).
Con el aumento de la velocidad de caída de la hojarasca, la concentración de nutrientes en las hojas que caen varía (Fig. 12.10). Cuando la velocidad de caída es alta, la concentración de nutrientes disminuye. La concentración de nutrientes es más alta en las hojas que caen durante los periodos con tasas de caída bajas. Estos cambios se deben a distintas tasas de retranslocación en los árboles en distintos
12 234 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
Figura
Caída de hojarasca en 10 plantaciones en el arboreto de Ciénaga Alta, Bosque Experimental de Luquillo (Cuevas y Lugo 1998). Las especies están ordenadas por la magnitud
producción de hojarasca
momentos; o sea, una fenología química distinta en cada especie. La eficiencia en el movimiento de carbono por la caída de hojarasca varía con la especie y dependiendo si se mide como eficiencia en el uso del fósforo o del nitrógeno (Fig. 12.11).
Es notable la alta eficiencia para ambos nutrientes en Pinus caribaea y, en contraste, la baja eficiencia en Hernandia sonora. Estas diferencias sugieren tasas más rápidas de recambio de nutrientes en Hernandia sonora y la tendencia del Pinus caribaea a retener y almacenar nutrientes. Pinus elliottii, un pino de zona templada, exhibe la eficiencia más alta en el uso del nitrógeno y una eficiencia notablemente más baja para el uso del fósforo.
Comparación de plantaciones con bosques nativos de la misma edad
En la sección anterior se señalaron diferencias en el comportamiento ecológico entre la especie nativa Hernandia sonora y otras especies introducidas comúnmente utilizadas en plantaciones arbóreas. Esas diferencias se confirmaron al comparar plantaciones con bosques secundarios nativos en los sectores de Guzmán, Cubuy, Sabana, y El Verde en el Bosque Experimental de Luquillo (Lugo 1992). En cada
12 235 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
Figura 12.9. Patrón y caída mensual de hojas en 10 plantaciones creciendo en el arboreto de Ciénaga Alta, Bosque Experimental de Luquillo (Cuevas y Lugo 1998). Las abreviaciones corresponden a: Pinus caribaea (Pc), Hibiscus elatus (He), Pinus elliottii (Pe), Eucalyptus x patentinervis (Ep), Khaya nyasica (Kn) Hernandia sonora (Hs), Anthocephalus chinensis (Ac), Eucalyptus saligna (Es), Swietenia macrophylla (Sm) y Terminalia ivorensis (Ti).
Figura 12.10. Contraste entre especies y dentro de cada especie del patrón y magnitud mensual de caída de hojas y la concentración de nitrógeno en las hojas que caen (Cuevas y Lugo 1998). En Eucalyptus saligna cuando aumenta la caída de hojas, baja la concentración de nitrógeno en las hojas y la concentración de nitrógeno es alta cuando la caída de hojas es baja. Swietenia macrophylla tiene un pico marcado en la caída de hojas cuando la concentración de nitrógeno llega a su mínimo. El patrón de Hernandia sonora contrasta con las dos anteriores, ya que la concentración de nitrógeno varía independientemente de la tasa de caída de hojarasca. Noten la variación en todas las escalas.
12 236 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
en el uso de nitrógeno y fósforo (masa de caída:masa del nutriente en la caída) en la caída de hojarasca de 10 plantaciones en el arboreto de Ciénaga Alta, Bosque Experimental de Luquillo (Cuevas y Lugo 1998). Noten la diferencia en escalas. Barras verticales representan el error estándar del promedio. Las especies están en el orden de caída de hojarasca (mayor a menor) y las abreviaciones corresponden a: Pinus caribaea (Pc), Hibiscus elatus (He), Pinus elliottii (Pe), Eucalyptus x patentinervis (Ep), Khaya nyasica (Kn) Hernandia sonora (Hs), Anthocephalus chinensis (Ac), Eucalyptus saligna (Es), Swietenia macrophylla (Sm) y Terminalia ivorensis (Ti). Los pinos exhiben altas eficiencias en el uso del nitrógeno y el fósforo y Hernandia sonora las más bajas. lugar había un rodal secundario y una plantación. Las comparaciones, hechas simultáneamente, están basadas en rodales de similar edad creciendo en condiciones comparables (Lugo 1992). Los bosques nativos invierten más energía en raíces que las plantaciones (Fig. 12.12). En el caso de los pinos de Cubuy, la biomasa de raíces es similar a la del bosque secundario, excepto la distribución de las raíces por diámetro. La gran mayoría de las raíces de la plantación eran raíces gruesas, por lo que la diversidad de tamaños de raíces era superior en el bosque secundario nativo. La productividad primaria neta de las plantaciones generalmente es superior a la de los bosques secundarios nativos (Tablas 12.2 y 12.3). A pesar de eso, el intercambio de nutrientes es más rápido en los bosques nativos que en las plantaciones (Fig. 12.13) donde ya habíamos anticipado que la tendencia es acumular nutrientes. La pendiente de la relación entre flujo y almacenaje (o sea la tasa de intercambio) es más alta en los bosques secundarios que en las plantaciones (Fig. 12.13b,c,d) a pesar de que la pendiente en la relación análoga para la masa (Fig. 12.13a) es similar para ambos tipos de bosque. Por lo tanto, mientras el flujo de
12 237 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
Figura 12.11. Eficiencia
Figura
la
raíces
cm en plantaciones de Pinus caribaea de 4 y 18.5 años en Guzmán y Cubuy, respectivamente, y de caoba (Swietenia macrophylla ) de 17 y 49 años en Sabana y El Verde, respectivamente (círculos superiores) y de bosques secundarios pareados con la plantación por edad y ubicación (círculos inferiores), Bosque Experimental de Luquillo. La biomasa total a un metro de profundidad está debajo de cada círculo (Lugo 1992).
su
12 238 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
12.12. Distribución de
biomasa (kg/ha) de
por
diámetro en
Figura 12.13.
Relación entre la masa (A), nitrógeno (B), fósforo (C) y potasio (D) en la caída de hojarasca (cantidad por hectárea por año) y el almacenaje de la masa y elementos (cantidad por hectárea) en la hojarasca del suelo de plantaciones (puntos sólidos) y de los bosques secundarios pareados (puntos abiertos; Lugo 1992). La relación o recambio de la masa (pendiente de la línea) aparenta ser similar en plantaciones y bosques secundarios. Para el nitrógeno y el fósforo la relación es más rápida en los bosques secundarios comparados con las plantaciones. El potasio no exhibe diferencias o tendencias excepto que hay más retorno en los bosques secundarios (D). Las plantaciones operan con masas de hojarasca más altas que los bosques secundarios (A).
Tabla 12.3
Productividad primaria neta aérea en Mg/ha · año para bosques nativos y plantaciones en el bosque experimental de Luquillo (Lugo 1992). En Guzmán y Cubuy las plantaciones son de Pinus caribaea y en Sabana y El Verde de Swietenia macrophylla
12 239 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
. Sitio Plantación Bosque nativo Guzmán 11.4 7.9 Cubuy 15.1 10.0 Sabana 16.6 8.5 El Verde 19.1 12.3
biomasa es similar en plantaciones y bosques secundarios, nutrientes como nitrógeno, fósforo y potasio circulan más rápido en los bosques secundarios nativos que en las plantaciones.
Efectos de huracanes sobre la plantación de pino en Cubuy
En el año 1982, los pinos dominaban las clases de diámetro más grandes en su plantación (Fig. 12.14). La distribución de diámetros cambió en 2014 después de que dos huracanes casi eliminaron los árboles grandes; entonces, predominaron los árboles pequeños. Después del huracán, el crecimiento en área basal en la plantación se acercó más al observado en el bosque secundario nativo, predominando crecimientos bajos y casi desapareciendo las tasas de crecimientos altas (Fig. 12.15). Se observó que el crecimiento de árboles de distintas especies siguió patrones distintos para diferentes especies (Lugo et al. 2017). Una de las sorpresas en las plantaciones de pino después de los huracanes fue que estas aumentaron en la riqueza (número) de especies (Fig. 12.16). Este aumento estuvo acompañado por una reducción en la dominancia y el Valor de Importancia del pino (Fig. 12.17), el cual casi desapareció de la plantación de Cubuy. Como consecuencia de estos resultados, no se puede recomendar el pino para producir madera para aserrar en
12 240 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
Figura 12.14. Distribución del diámetro de los árboles en una plantación de Pinus caribaea en el 1982 antes de los huracanes Hugo y Georges y después de los huracanes en el 2014. Los árboles de pino con diámetros mayores de 15 cm desaparecieron y la clase dominante de diámetros (5 a 10 cm) aumentó en frecuencia (Lugo et al. 2017).
Distribución de las tasas anuales de crecimiento arbóreo en área basal en una plantación de Pinus caribaea y un bosque secundario pareado por edad y lugar de crecimiento (Cubuy, Bosque Experimental de Luquillo) antes (1982-1990) y después (2007-2014) del paso de huracanes (Lugo et al. 2017). La tasa prevaleciente fue la de 0 a 4 cm2/año. Las tasas altas de crecimiento (> 11 cm2/año) del pino antes de los huracanes se redujeron después de los huracanes y entonces prevalecieron tasas de crecimiento más bajas.
El número de especies arbóreas aumento en dos plantaciones de Pinus caribaea después del huracán Hugo en el 1989 (Lugo et al. 2017). El área de muestreo fue de 50 metros cuadrados por plantación.
12 241 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
Figura 12.15.
Figura 12.16.
lugares expuestos periódicamente a huracanes. El pino se puede recomendar para la producción de papel o para la rehabilitación de suelos altamente erosionados, ya que para estos usos no es necesario cultivar árboles grandes.
Plantaciones para restaurar biodiversidad, rescatar tierras degradadas y catalizar la sucesión
Los resultados de investigaciones y observaciones de plantaciones con bajo nivel de mantenimiento en Puerto Rico dieron paso al uso de plantaciones con propósitos de rehabilitación de tierras degradadas, restauración de biodiversidad y la aceleración de la sucesión (Parrotta y Turnbull 1997). Lugo et al. (1993) habían encontrado que una plantación abandonada puede acumular una nueva especie arbórea por año, un valor similar a lo observado en sucesiones de bosques nativos. Silver et al. (2000, 2004) demostraron que los bosques maduros regenerados artificialmente en pastizales degradados (reforestados por José Marrero) se caracterizan por una alta diversidad de árboles (principalmente especies nativas) y el secuestro de carbono en suelos y vegetación.
Varios procesos en la plantación contribuyen a su función de rehabilitación y restauración de suelos, bosques y biodiversidad. Por ejemplo:
12 242 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
Figura 12.17. El Valor de Importancia de Pinus caribaea se redujo en dos plantaciones después del huracán Hugo, pero la reducción fue mayor en Cubuy donde la plantación era 14.5 años más vieja que en Guzmán (Lugo et al. 2017).
L as plantaciones pueden establecerse en tierras degradadas donde la sucesión de especies nativas es muy lenta o arrestada.
El desarrollo de un sotobosque en una plantación no afecta su productividad maderera, pero sirve para atraer vida silvestre, incluso dispersadores de semillas.
El cierre del dosel de una plantación cambia el microclima (luz, temperatura y humedad) lo que facilita la regeneración de especies nativas. Especies madereras de alto rendimiento no crecen bajo su propia sombra, lo que indica que les dan paso a otras especies.
L a plantación concentra nutrientes y materia orgánica y reduce la compactación del suelo, para así fomentar la restauración de la fertilidad del mismo.
El retorno de materiales con altas cantidades (procedentes del dosel) y altas concentraciones (procedentes del sotobosque) de nutrientes acelera el reciclaje de nutrientes y el crecimiento de especies nativas.
Asignación
Selecciona cualquier argumento controversial a favor o en contra de las plantaciones madereras y preséntale a la clase una posición balanceada basada en el conocimiento científico.
12 243 CAP ariel e. lugo | ecología a m i m anera Ecología de plantaciones arbóreas tropicales
CaPítulo 13
El ciclo de carbono de los bosques tropicales
Los procesos que transportan el carbono y los lugares de almacenamiento del carbono en los bosques tropicales son conocidos en términos generales, pero la disponibilidad de datos no es suficiente para establecer cómo se cuadra el ciclo de carbono partiendo de los insumos de la actividad humana a la atmósfera. Los datos disponibles cubren un área limitada de bosques comparada con el área global cubierta por los bosques tropicales. Estos datos apoyan estimaciones sobre el papel de los bosques tropicales en el ciclo de carbono del planeta, pero la incertidumbre es alta, lo que crea controversias dentro de la comunidad científica. La falta de datos lleva a los científicos a emitir opiniones o hacer estimados que dependen de la experiencia y creencias del investigador o al uso de datos regionales extrapolados a escala global. Como consecuencia, el asunto sobre el papel que juegan los bosques tropicales en el ciclo de carbono global lleva más de 50 años de discusión intensa sin que aún haya un consenso entre los diversos grupos de participantes en estas discusiones. Investigaciones en Puerto Rico han sido fundamentales para aclarar el papel de los bosques tropicales en el ciclo de carbono global, según la narrativa de Brown y Lugo (2017) que resumo en este capítulo.
El tema de este capítulo es de importancia práctica porque el ciclo de carbono de los bosques tropicales afecta la concentración de bióxido de carbono en la atmósfera y esta a su vez, afecta el clima por su efecto en la retención de calor en la atmósfera. Más aún, para mitigar el calentamiento global, la comunidad internacional recompensa a los países que manejan sus bosques para optimizar el secuestro del carbono. Muchos países han adoptado la estrategia de secuestrar carbono en sus bosques como uno de los objetivos principales de la conservación.
El bosque mejor estudiado en Puerto Rico desde la perspectiva del ciclo de carbono es el bosque de tabonuco en el sector El Verde del Bosque Experimental de Luquillo (Odum 1970). En el sector de Bisley en el mismo tipo de bosque, el análisis del ciclo de carbono ha incluido la distribución del carbono de acuerdo con la topografía (Scatena y Lugo 1995) y en respuesta al huracán Hugo (Scatena et al. 1996). También se ha estudiado la distribución, almacenaje y flujos del carbono en varios otros bosques de Puerto Rico tales como el manglar de la Parguera (Golley et al. 1962), el bosque seco de Guánica (Lugo et al. 1978, Murphy y Lugo 1986b, Lugo y Murphy 1986),
ariel e. lugo | ecología a m i m anera
245
los palmares de Prestoea montana en el Bosque Experimental de Luquillo (Frangi y Lugo 1985, 1998), los bosques noveles de Spathodea campanulata y Castilla elastica (Lugo et al. 2008, 2011, 2012b; Fonseca da Silva 2011, Gutiérrez del Arroyo Santiago 2014) y una variedad de plantaciones arbóreas y bosques secundarios (Lugo y Figueroa 1985, Lugo 1992, Silver et al. 2004).
En este capítulo primero resumo los resultados de trabajos sobre el ciclo de carbono en los bosques de Puerto Rico para ilustrar la variabilidad en los almacenajes y distribución de carbono y de paso resumir algunas lecciones aprendidas al examinar la dinámica del carbono en estos bosques isleños. Luego cambio de escala en el análisis para concentrar la atención en los factores que hacen que los flujos y almacenajes de carbono varíen entre un tipo de bosque y otro. Con ese entendimiento es posible inferir el efecto agregado de los bosques tropicales sobre el ciclo de carbono del mundo, el cual depende del área de los bosques, su edad y la velocidad de sus procesos. Luego menciono los factores que están operando para aumentar o disminuir los flujos de carbono entre los bosques tropicales y la atmósfera a nivel global. Termino con una discusión sobre el balance del ciclo de carbono en el planeta.
Antes de presentar datos es recomendable entender las equivalencias entre datos expresados en carbono, materia orgánica (o masa) y bióxido de carbono. Las proporciones están basadas en la estequiometría de las ecuaciones de fotosíntesis y respiración discutidas en el capítulo 3. Los estudios de biomasa se basan en el peso seco de la masa del bosque y esos valores se convierten a carbono dividiendo la masa por dos. Aunque el carbono representa el 40 % del azúcar producto de la fotosíntesis, cuando los azúcares se transforman en tallos, hojas y raíces, la proporción de carbono aumenta y varía según la parte de la planta y la especie. Por eso, se utiliza el 50 % como factor de conversión de masa a carbono. En estudios más precisos se puede utilizar la concentración de carbono en los distintos compartimientos de la biomasa vegetal bajo estudio. Para convertir la masa de carbono a bióxido de carbono, se multiplica por 3.67.
El almacenaje y flujo de carbono en un rodal
Los principales procesos que controlan el flujo de carbono en un bosque son cuatro (Fig. 13.1). En orden de magnitud del flujo de carbono: (1) la fotosíntesis de las plantas; (2) la respiración de las plantas, la vida silvestre, los microorganismos y animales del suelo; (3) el reciclaje o movimiento del carbono dentro y fuera del bosque (incluyendo la caída de hojarasca, mortalidad de tallos y ramas), y (4) la cadena alimenticia la cual en la figura 13.1 solo ilustra el papel de los microorganismos. Estos cuatro flujos interconectan los almacenajes de carbono para que el rodal funcione como un sistema. Los almacenajes de carbono se pueden categorizar entre los
13 246 CAP ariel e. lugo | ecología a mi ma nera El ciclo de carbono de los bosques tropicales
almacenajes sobre el terreno y los subterráneos. La biomasa viva (hojas, tallos, ramas y la vida silvestre es un almacenaje sobre el terreno. Las raíces (vivas y muertas), los microbios, y los animales del suelo, como las lombrices de tierra, constituyen los almacenajes subterráneos. El suelo almacena agua, nutrientes y una cantidad significativa de materia orgánica. La materia orgánica del suelo es producto de la actividad biótica del rodal y tiene dos estados: materiales orgánicos recientes y lábiles y materiales orgánicos más antiguos y recalcitrantes. No todos estos almacenajes están ilustrados en la figura 13.1.
La figura 13.2 ilustra los almacenajes y flujos principales de carbono en un rodal generalizado y presenta ejemplos para varios tipos de bosque en Puerto Rico. La plantación de caoba y el palmar tienen la productividad neta más alta, seguida por el bosque maduro de tabonuco, el bosque secundario de tabonuco y finalmente el bosque seco. El bosque seco tiene tanta biomasa en raíces como biomasa aérea, mientras que los otros bosques tienen más biomasa aérea que biomasa en raíces. La plantación de caoba tiene la biomasa de raíces menor entre los bosques (Fig. 13.2). El palmar tiene una alta acumulación de material orgánico en el suelo y alta biomasa en las plantas, seguido por el bosque de tabonuco maduro. La plantación, establecida en suelos erosionados, tiene bajos almacenajes de materia orgánica en el suelo.
Los flujos de carbono en cuatro bosques en Puerto Rico basados en medidas directas de intercambio gaseoso están resumidos en la figura 13.3. El orden en la
247 13 CAP ariel e. lugo | ecología a m i m anera El ciclo de carbono de los bosques tropicales
Figura 13.1. Diagrama simplificado del balance de carbono en un rodal arbóreo con énfasis en las plantas y los microbios.
Figura 13.2. Diagramas comparativos de los almacenajes (Mg/ha) y flujos (Mg/ha · año) principales de carbono en cuatro rodales arbóreos en Puerto Rico. Primera fila izquierda: bosque secundario de tabonuco (Dacryodes excelsa) y plantación de caoba (Swietenia macrophylla) creciendo en El Verde, Bosque Experimental de Luquillo (Lugo 1992); segunda fila izquierda: palmar de Prestoea montana en el Bosque Experimental de Luquillo (Frangi y Lugo 1985); segunda fila derecha: bosque deciduo en el Bosque Estatal de Guánica (Lugo y Murphy 1986). Los diagramas comparan la biomasa de plantas sobre y debajo del terreno y el almacenaje de materia orgánica en el suelo, incluyendo la hojarasca. Los flujos principales son la productividad primaria neta sobre el terreno (izquierda) y la caída de hojarasca (derecha). La profundidad de muestreo para la materia orgánica del suelo es de 1 metro, excepto para el bosque seco en Guánica (85 cm). Los valores de carbono se convierten a materia orgánica multiplicando por dos.
productividad primaria bruta de los sistemas, comenzando por el más productivo, es: (1) bosque maduro de tabonuco (Fig. 13.3a), (2) bosque novel de Castilla elastica (Fig. 13.3b), (3) el manglar (Fig. 13.3c), y (4) el bosque seco (Fig. 13.3d). Si se ordenan con base en la productividad primaria neta comenzando por el más productivo, el orden es: (1) bosque novel de Castilla elastica, (2) el manglar, (3) el bosque seco, y
13 248 CAP ariel e. lugo | ecología a m i m anera El ciclo de carbono de los bosques tropicales
(4) el bosque maduro de tabonuco. La complejidad estructural del bosque maduro de tabonuco requiere una proporción alta de la productividad bruta del bosque para sostenerse por medio de la respiración. El bosque novel es joven y en etapa rápida de crecimiento. El bosque seco se midió durante la época del año cuando la productividad neta exhibe sus tasas más altas debido a la disponibilidad de agua. El manglar es generalmente un sistema con alta productividad neta y por el efecto de las mareas, exporta carbono a las aguas circundantes. La mayor respiración en el bosque novel es en el suelo, la cual representa los valores más altos entre los cuatro bosques. En el bosque seco la variación en la respiración del suelo es alta y es segunda entre los cuatro bosques. En el bosque maduro de tabonuco la respiración edáfica es menos variable que en el bosque seco y su magnitud es similar a la del bosque seco. En el manglar, la exportación de materia orgánica reduce la disponibilidad de combustible para la respiración edáfica y combinado con las limitaciones de oxígeno durante el periodo de inundación da como resultado las tasas más bajas de respiración del suelo.
Factores que afectan los almacenajes y flujos de carbono en bosques tropicales
Brown y Lugo (1982) encontraron que la disponibilidad del agua, medida como el cociente T:P (temperatura anual promedio:precipitación anual promedio), correlaciona significativamente con la cantidad de biomasa que se acumula en los bosques maduros tropicales (Fig. 13.4). Aquellos bosques en zonas de vida seca (menos lluvia que evapotranspiración) acumulan menos biomasa en la vegetación que bosques húmedos donde la precipitación es equivalente a la evapotranspiración. Bosques en zonas de vida extremadamente húmedas (exceso de lluvia sobre evapotranspiración) acumulan más biomasa que los de zonas de vida secas, pero menos que los bosques en zonas de vida húmedas. La acumulación de carbono en el suelo es mayor en las zonas de vida extremadamente húmedas y disminuye rápidamente hacia las zonas de vida secas. El almacenaje total (vegetación más suelo) disminuye linealmente con la disminución en la disponibilidad de agua.
La caída de hojarasca es un indicador del reciclaje de carbono dentro de los bosques y sigue el mismo patrón que la biomasa aérea (Fig. 13.5). El crecimiento de los árboles (tasa de acumulación de carbono en la vegetación), no varía con la disponibilidad del agua y es relativamente constante desde los trópicos a la zona templada (Jordan 1971). Por lo tanto, la caída de hojarasca en la figura 13.5 presenta un indicador de la variación en la productividad neta en los bosques tropicales. Los valores máximos ocurren en los bosques húmedos y disminuyen hacia los muy húmedos o hacia los secos. La explicación es que los bosques húmedos, expuestos a un balance hídrico, no experimentan sequías, como ocurre en los bosques secos, ni saturación de suelos o de la atmósfera, como ocurren en los bosques muy húmedos, incluyendo
249 13 CAP ariel e. lugo | ecología a m i m anera El ciclo de carbono de los bosques tropicales
Figura
y
(g/m2) principales de carbono (uno en unidades de materia orgánica) en cuatro rodales estudiados con técnicas de intercambio gaseoso. Los rodales son (A) el bosque muy húmedo y maduro de tabonuco (Dacryodes excelsa; Odum 1970), reportado en unidades de materia orgánica) (B) un bosque maduro de Castilla elastica en la zona del karso húmedo (Lugo et al. 2008), (C) un manglar en La Parguera (Golley et al. 1962) y (D) un bosque deciduo en el Bosque Estatal de Guánica (Lugo et al. 1978). Los datos para el diagrama del manglar están en la figura 10.2. La caída de hojarasca en el bosque de Castilla es de Gutiérrez del Arroyo y Wood (2019). El círculo en los diagramas representa la fuente de energía solar que potencia la fotosíntesis bruta, el primer número que aparece a la izquierda del símbolo de productores (ver figura 2.9 para la descripción de los símbolos utilizados). Los datos de respiración están asociados al símbolo de pérdida
13 250 CAP ariel e. lugo | ecología a m i m anera El ciclo de carbono de los bosques tropicales
13.3. Diagramas simplificados de los flujos (g/m2 · día)
almacenajes
de calor. En el diagrama del bosque de Castilla, se presentan dos valores para cada flujo. El valor sobre la línea, o como numerador, es para rodales dominados por Castilla elastica (una especie introducida) y el valor debajo de la línea, o como denominador, es para rodales dominados por especies nativas. Se presentan dos flujos respiratorios para las plantas. El primero es la respiración diurna, que al restársele a la fotosíntesis bruta resulta en la fotosíntesis neta. El segundo flujo respiratorio ocurre a expensas de la fotosíntesis neta. Los valores para B, C , y D están reportados en unidades de carbono y los de A en unidades de materia orgánica. Para A , el estudio original reportó unidades de energía y se convirtieron a materia orgánica dividiendo por 4 kcal/gramo. Para convertir los datos de carbono en materia orgánica hay que multiplicarlos por dos.
251 13 CAP ariel e. lugo | ecología a m i m anera El ciclo de carbono de los bosques tropicales
13 252 CAP ariel e. lugo | ecología a m i m anera El ciclo de carbono de los bosques tropicales
Figura 13.4. Relación entre el almacenaje de materia orgánica en vegetación, suelos y el total de materia orgánica de bosques tropicales y el cociente temperatura promedio anual:precipitación promedio anual (Brown y Lugo 1982).
Figura 13.5. Relación entre la caída total de hojarasca en bosques tropicales y el cociente temperatura promedio anual:precipitación promedio anual (Brown y Lugo 1982).
Tabla 13.1
Estimado del almacenaje promedio de biomasa en zonas de vida tropicales y subtropicales a nivel global (Brown y Lugo 1982). El error estándar está en paréntesis.
TB = tierras bajas tropicales
S/T = tierras bajas subtropicales y zonas de vida a elevaciones sobre las tierras bajas y todas las zonas de vida tropicales altitudinales.
los pluviales. Como consecuencia, los bosques húmedos generalmente exhiben las tasas de productividad primaria neta más altas entre los bosques tropicales. Tanto las sequías como la saturación hídrica interrumpen o disminuyen (dependiendo de la intensidad) la fijación de carbono por medio de la fotosíntesis y pueden acelerar la respiración debido al costo metabólico para sobreponerse a las condiciones limitantes. El resultado es una reducción en la tasa de secuestro de carbono, o sea, una menor tasa de productividad primaria neta.
El rol de los bosques tropicales en el balance de carbono del mundo
Para establecer este rol, se necesitan datos del área de los bosques y del almacenaje de carbono por unidad de área para cada tipo de bosque. Como el almacenaje varía con las zonas de vida (Fig. 13.4), es necesario conocer el área de bosques por tipo de zona de vida (Tabla 13.1).
Los estimados de la tabla 13.1 están basados en datos tomados en los años antes del 1980 por ecólogos y sirven como estimados aplicables a bosques primarios (Brown y Lugo 1982). Los mismos autores encontraron que datos recolectados por dasónomos resultaban en estimados de biomasa más bajos (Brown y Lugo 1984). La razón era que los inventarios forestales cubren más terreno que los estudios ecológicos y están diseñados para la recolección al azar de los datos. Esto da como resultado una densidad de carbono más baja y es consistente con la realidad de su variación en el paisaje. Brown y Lugo (1990) luego estimaron la densidad de carbono en bosques secundarios, que son el tipo de bosque que prevalece por todo el trópico. Estos resultados resaltan la dificultad de desarrollar un estimado confiable de la densidad de biomasa en un área tan grande y compleja como la cobertura de los bosques tropicales.
253 13 CAP ariel e. lugo | ecología a m i m anera El ciclo de carbono de los bosques tropicales
Zona de Vida Área Vegetación Suelo Total 109 ha 109 Mg 109 Mg 109 Mg (%) TB muy húmedo y pluvial 209.7 72.1 (6.5) 48.2 (13.4) 121.615 TB húmedo 347.0 128.0 (9.4)59.0 (4.2) 189.1 24 TB seco 177.5 51.8 (3.7) 25.2 (4.3) 78.1 10 S/T muy húmedo y pluvial 255.8 82.4 (5.1) 89.5 (15.4) 173.9 22 S/T húmedo 259.7 75.6 (7.5) 50.9 (8.3) 127.8 16 S/T seco 588.6 47.1 (13.0) 45.9 (12.4) 96.5 12 Total 1838.3 457.0 (45.2) 318.7 (58.0) 787 100
Tabla 13.2
Secuestro de carbono en los sedimentos (burial) de la vegetación costera (carbono azul) en comparación con bosques terrestres en varias latitudes. Los datos son de Mcleod et al. (2011) donde se pueden encontrar los detalles y aspectos estadísticos.
Los estimados de área de bosque tropical utilizados en la tabla 13.1 se obtuvieron antes de que se tuviesen disponible los datos de cobertura basados en imágenes de satélite y antes de estar disponibles los estimados de biomasa e intercambio de gases entre el bosque y la atmósfera utilizando nuevas tecnologías (Saatchi et al. 2011). Sin embargo, a pesar de los avances tecnológicos, los rangos en los resultados presentados no varían mucho con los valores utilizados en el presente.
Lo que sí ha cambiado es la atención dada a tipos de vegetación que anteriormente no se consideraban en los cálculos globales por su baja cobertura de terreno. Actualmente, se habla del carbono azul (blue carbon) para reconocer la importancia de la vegetación costera en el secuestro de carbono. Los ecosistemas costeros que contribuyen al carbono azul son los manglares, praderas de gramíneas marinas y los pantanos salinos de herbáceas (Mcleod et al. 2011).
Para ilustrar estos avances, consideremos la importancia global de la vegetación costera, incluidos los manglares (Tabla 13.2). La vegetación costera tiene una cobertura muy inferior a la de los bosques terrestres. Sin embargo, la intensidad del proceso de sedimentación de carbono por unidad de ecosistema costero es muy superior a la de los bosques terrestres. Por lo tanto, a pesar de que cubren poca superficie, los ecosistemas costeros remueven igual o mayor cantidad de carbono que los bosques terrestres. Claramente, el carbono azul representa un flujo planetario de importancia al ciclo del carbono.
Otra forma de visualizar la importancia de los bosques tropicales en el balance global del carbono es al comparar los datos de almacenaje en la tabla 13.1 con valores para otros tipos de vegetación (Fig. 13.6). Los datos indican que los bosques tropicales comprenden el 9.9 % del área total de vegetación en el planeta. El almacenaje de carbono en los suelos de los bosques tropicales es proporcional a su área, pero no así el almacenaje de carbono en la biomasa aérea, ya que los bosques tropicales almacenan casi el 45 % del total.
13 254 CAP ariel e. lugo | ecología a m i m anera El ciclo de carbono de los bosques tropicales
Ecosistema Sedimentación (g C/m2 · año) Área (km2) Sedimentación global (Tg C/año) Pantano salino de herbáceas 218 22,000 a 400,000 4.8 a 87.2 Manglares 226 137,760 a 152,361 31.1 a 34.4 Praderas marinas 138 177,000 a 600,000 48 a 112 Bosques templados 5.1 10,400,000 53.0 Bosques tropicales 4.0 19,622,846 78.5 Bosques boreales 4.6 13,700,000 49.3
refieren a los siguientes totales: un área de 149.3 millones de kilómetros cuadrados, un almacenaje total de 559.8 X 1015 gramos (peta gramos) de carbono almacenado en vegetación y un almacenaje de 1,635.6 X 1015 gramos de carbono en el suelo hasta un metro de profundidad. Los bosques tropicales con el 9.9 % de la cobertura terrestre, almacenan el 43.5 % del carbón de la vegetación global, el 7.5 % del almacenaje en los suelos y el 16.7 % del almacenaje de carbono global terrestre.
255 13 CAP ariel e. lugo | ecología a m i m anera El ciclo de carbono de los bosques tropicales
Figura 13.6. Distribución del área de cobertura (A) y el almacenaje de carbono en la vegetación (B), el suelo (C) y la suma de los dos (total; D) en distintos biomas terrestres del mundo (Ajtay et al. 1979). Los porcentajes se
El ciclo de carbono global no cuadra con los datos disponibles
El consenso en el año 2000 sobre el ciclo de carbono en el planeta se resume en la figura 13.7. El consenso cambia porque la quema de combustibles fósiles continúa en ascenso, al igual que el almacenaje de carbono en la atmósfera (Fig. 4.6). Como la quema de combustible sigue cambiando al igual que la concentración del carbono en la atmósfera, es necesario balancear el ciclo del carbono con sumideros en los océanos y la vegetación terrestre. Identificar la magnitud de esos sumideros de carbono es el reto principal de los que estudian los ciclos globales, pues estos ciclos deben funcionar dentro de los parámetros de la ley de conservación de masa. En el caso del carbono, sabemos que el ciclo está en transición, ya que la concentración del carbono en la atmósfera y los océanos aumenta continuamente.
13 256 CAP ariel e. lugo | ecología a m i m anera El ciclo de carbono de los bosques tropicales
Figura 13.7. Flujos y almacenajes globales de carbono que ilustra la importancia relativa de los almacenajes en rocas y sedimentos, petróleo, océanos, plantas y suelos y la atmósfera (Houghton 2000).
Tabla 13.3
Flujos y balance global del carbono basado en el análisis de Pan et al. (2011). Las unidades son Pg/año.
El almacenaje principal de carbono en el planeta se encuentra en los sedimentos y las rocas sedimentarias seguidas por el océano. La atmósfera es el reservorio de carbono más pequeño y el más dinámico. En un sistema balanceado, la respiración del planeta iguala a su fotosíntesis y la concentración de carbono en la atmósfera sería constante. El desbalance en el flujo de carbono entre la atmósfera y los varios reservorios de carbono lo crea la suma de tres flujos antropogénicos: la producción de cemento (que voy a ignorar por ser un flujo estable y de baja magnitud, i.e., <10 % del total antropogénico), la quema de combustible fósil y la conversión de bosques a otros usos. Todos generan un flujo neto de carbono en la atmósfera. Sin embargo, los números no cuadran, ya que el flujo conocido hacia los sumideros de carbono atmosférico es menor que las fuentes conocidas de carbono atmosférico. El resultado es que hay un sumidero de carbono desconocido o los estimados de los sumideros contienen un error de estimación. Eso se debe a incertidumbres con el papel que juega la vegetación terrestre, particularmente en los trópicos.
A mi entender, el problema está en cómo conceptualizamos el papel que juegan los bosques tropicales en el ciclo del carbono. Por un lado, se asume que un área significativa de vegetación madura es neutral (en estado estable) con relación al balance de carbono. O sea, se asume que su captura de carbono atmosférico a través de la fotosíntesis iguala su respiración. Sin embargo, normalmente la vegetación se comporta como un sumidero de carbono atmosférico, según resumen de Lugo y Brown (1986). Por otro lado, se exagera el flujo de carbono a la atmósfera cuando se altera la cobertura vegetal, ya que se asume que todo el carbono almacenado en la vegetación se convierte en bióxido de carbono y se subestima el sumidero debido al crecimiento posterior de la vegetación.
Pan et al. (2011) cuadraron el ciclo de carbono del planeta basado en la presencia de sumideros de carbono atmosférico en los bosques (Tabla 13.3). Noten el aumento
257 13 CAP ariel e. lugo | ecología a m i m anera El ciclo de carbono de los bosques tropicales
Emisiones y sumideros 1990-1999 2000-2007 Emisiones Combustibles fósiles y cemento 6.5 + 0.4 7.6 + 0.4 Cambio en uso de terreno 1.5 + 0.7 1.1 + 0.7 Total 8.0 + 0.8 8.7 + 0.8 Sumideros Atmósfera 3.2 + 0.1 4.1 + 0.1 Océanos 2.2 + 0.4 2.3 + 0.4 Vegetación terrestre 2.5 + 0.4 2.3 + 0.5 Total 7.9 + 0.6 8.7 + 0.7 Residuo global 0.1 + 1.0 0.0 + 1.0
reciente en los valores y el residuo en el intervalo 1990-1999. Estos cálculos contienen mucha incertidumbre, particularmente en los trópicos donde aún se discuten las magnitudes asociadas a los sumideros de carbono atmosférico en los bosques tropicales, al igual que las emisiones de carbono a la atmósfera por deforestación y urbanización (cambio en uso de terrenos en la Tabla 13.3).
Factores que afectan el intercambio de carbono entre los bosques tropicales y la atmósfera
Cuando la fotosíntesis de un bosque es mayor a lo que respira, acumula biomasa y remueve más carbono de la atmósfera de lo que añade. Si, por el contrario, el bosque tiene más respiración que fotosíntesis, entonces añade más carbono a la atmósfera de lo que remueve. En este caso su biomasa debe disminuir. Como las medidas de fotosíntesis y respiración de bosques tropicales son muy escasas, se utilizan otros criterios para establecer el balance neto de intercambio gaseoso entre el bosque y la atmósfera. Si un bosque se quema, se deforesta o degrada, se considera una fuente de carbono a la atmósfera, ya que reduce su almacenaje de biomasa al oxidarse y convertirse en bióxido de carbono. Por el contrario, un bosque secundario en rápido crecimiento acumula materia orgánica y se considera un sumidero de carbono atmosférico (Brown y Lugo 1990). Estas determinaciones se hacen utilizando fotos aéreas o imágenes de satélite, en distintos momentos para determinar si hubo deforestación o crecimiento en el área de los bosques. Se utilizan Sistemas de Información Geográfica (GIS, por sus siglas en inglés) y modelos matemáticos para estimar los balances de carbono. El problema con estos esfuerzos es que no hay datos de campo suficientes para apoyarlos y como consecuencia tienen que depender de premisas con altos márgenes de error.
Sumideros terrestres de carbono atmosférico
Recientemente, se ha utilizado la variación en la concentración de bióxido de carbono en la atmósfera para estimar el balance neto de carbono de grandes extensiones de terreno y a largo plazo. Por ejemplo, para zonas templadas, Crevoisier et al. (2010) encontraron sumideros regionales en los EE. UU. entre el 2004 y 2006 con una magnitud de 0.5 + 0.4 Pg/año. Estudios similares sobre bosques tropicales llegan a la misma conclusión: encuentran sumideros netos de carbono.
Estrategias para aumentar los sumideros de carbono atmosférico
La vegetación en general y los bosques en particular son sumideros de carbono atmosférico. Ryan et al. (2010) proponen las siguientes estrategias de manejo de bosques para incrementar los sumideros de carbono atmosférico:
(1) Prevenir la deforestación.
(2) Promover la aforestación o el establecimiento de bosques en tierras que no han
13 258 CAP ariel e. lugo | ecología a m i m anera El ciclo de carbono de los bosques tropicales
tenido bosques por algún tiempo o que nunca los han tenido. (3) Aumentar el intervalo de tiempo para el corte de madera. (4) Acelerar el crecimiento de los árboles. (5) Realizar raleos para minimizar los fuegos. (6) Promover los bosques urbanos. (7) Usar biomasa en vez de energía fósil como fuente energética. (8) Almacenar carbono a largo plazo en productos forestales como muebles y sustituir el acero por la madera. Además, promover y conservar árboles grandes (> 70 cm dap) en los bosques incrementa la magnitud del sumidero de carbono, ya que un árbol grande por hectárea almacena más de la mitad del carbono en la hectárea (Brown y Lugo 1992).
259 13 CAP ariel e. lugo | ecología a m i m anera El ciclo de carbono de los bosques tropicales
CaPítulo 14
Las velocidades de extinción y evolución de las especies
Pregunta
¿Estamos en medio del evento de extinción de especies más grande en la historia del planeta Tierra? De acuerdo con Barnosky et al. (2011) la respuesta es sí. Estiman que dentro de varios siglos, si continúan las cosas como van, el planeta tendrá su sexta extinción masiva, esta vez causada por el ser humano.
¿Cuán confiable es tal aseveración?
Extinciones masivas
Los paleontólogos definen un evento de extinción masiva cuando la tierra pierde más de tres cuartas partes de las especies en un intervalo geológico relativamente corto (Barnosky et al. 2011). Las extinciones masivas han ocurrido cinco veces en los últimos 540 Ma (Tabla 14.1).
Se estima que de los 4 billones de especies de organismos que han evolucionado en el planeta, aproximadamente el 99 % se han extinguido. En general, la evolución de las especies contrarresta las extinciones, aunque representan procesos distintos que necesariamente no están relacionados. Durante extinciones masivas las tasas de evolución no igualan a las de extinción. Durante otros intervalos, la especiación por evolución supera las extinciones. Se estima que las especies tardan cientos de miles de años en evolucionar (aunque hay excepciones discutidas más adelante en este capítulo), lo que causa que se requieran millones de años en recobrar el número de las especies después de una extinción masiva. Erwin (1998) reconstruyó las altas y bajas en el número de familias marinas durante seis extinciones masivas en un periodo de tiempo de 600 Ma (Fig. 14.1). La biodiversidad total aumentó durante el intervalo de tiempo a pesar de las extinciones.
ariel e. lugo | ecología a mi ma nera
261
Tabla 14.1
Extinciones masivas del pasado y sus probables causas (Barnosky et al 2011).
Evento de extinción
Evento del Ordovícico, terminó aproximadamente hace 443 Ma, entre 3.3 y 1.9 Ma se perdieron 57 % de los géneros y aproximadamente 86 % de las especies
Evento del Devónico, terminó aproximadamente 359 Ma, entre 29 y 2 Ma se perdieron aproximadamente el 35 % de los géneros y 75 % de las especies
Evento del Perminan terminó aproximadamente
hace 251 Ma, entre 2.8 Ma y 160 mil años se perdieron aproximadamente el 56 % de los géneros y 96 % de las especies
Evento del Triásico terminó aproximadamente hace 200 Ma, entre 8.3 Ma y 600 mil años se perdieron aproximadamente el 47 % de los géneros y el 80 % de las especies
Evento del Cretácico terminó aproximadamente hace 65 Ma, entre 2.5 Ma a menos de un día se perdieron el 40 % de los géneros y el 76 % de las especies
Causas propuestas
Alternación de periodos glaciales e interglaciares, repetidas transgresiones del mar, levante y erosión de las montañas Apalaches lo que afecta la química del océano y atmósfera. Secuestro del CO2
Enfriamiento global seguido de calentamiento global relacionado con la diversificación de las plantas con desgaste de rocas, pedogénesis y disminución del CO2 Evidencia de anoxia en las aguas profundas del mar y su esparcimiento por transgresiones del mar
Se debate la ocurrencia de impactos de cometas
Volcanismo en Siberia
Calentamiento global. Expansión de aguas marinas anóxicas y profundas Altas concentraciones de H2S y CO2 en los entornos marinos y terrestres
Acidificación del océano
Se debate la ocurrencia de impactos de cometas
Actividad en la provincia magmática en el centro Atlántico (CAMP), se cree elevó los niveles de CO2 en la atmósfera, los cuales elevaron las temperaturas y llevaron a una crisis de calcificación en los océanos
Impacto de cometa en Yucatán llevó a un cataclismo global y rápido enfriamiento Anteriormente, la biota estaba en descenso por otras causas Volcanismo con calentamiento global, levantamiento tectónico alteró la biogeografía y aceleró la erosión, potencialmente contribuyendo a la eutroficación de los océanos y a episodios anóxicos Pico en las concentraciones de CO2 antes de las extinciones y descenso durante el evento de extinción
Estimaciones de tasas de extinción de especies
Para estimar la magnitud de la extinción de especies se necesita información sobre el número de especies en el planeta y la velocidad de su extinción. No se conoce el número de especies en el planeta y la información disponible está basada en estimaciones que varían entre 2 y más de 50 millones de especies (Scheffers et al. (2012). La tabla 14.2 contiene estimaciones del número de especies para distintos grupos de organismos. El número de especies descrito, catalogado y estimado no concuerda y luego de 200 años de trabajo taxonómico se estima que costaría 263 billones de dólares del 2010 y varios siglos más para describir las especies de animales ahora desconocidas. Como no sabemos cuántas especies hay en el planeta ni la velocidad a la cual se extinguen o evolucionan (ambos parámetros discutidos más adelante en este capítulo), se calcula la tasa de extinción de especies basada en la relación entre área de hábitat y la acumulación de especies por área de hábitat (Fig. 14.2).
S = CA z
Donde S es el número de especies, A es el área y Z y C son constantes. La pendiente de la relación especie-área es Z que refleja la densidad de especies (especies por área). La idea es estimar las extinciones invirtiendo la curva de acumulación de
14 262 CAP. ariel e. lugo | ecología a m i m anera Las velocidades de extinción y evolución de las especies
de
importancia relativo a las extinciones masivas (Erwin 1998). Utilizando la figura 11.3 para identificar los periodos geológicos: Proterozoico es Ven, Cambriano es Camb, Ordovícico es Ord, Siluriano es Sil, Devoniano es Dev, Carbonífero es Carb, Pérmico es Per, Triásico es Tri, Jurásico es Jur, Cretáceo es Cret y Ter (de Terciario) corresponde ahora al Paleógeno, Neógeno y el Cuaternario.
especies. Se estima la tasa de extinción si se sabe la tasa de pérdida del hábitat, por ejemplo, la tasa de deforestación.
Distintas personas y organizaciones definen lo que es deforestación, cada uno a su manera, pero hay una organización que organiza y publica datos para todo el planeta utilizando criterios objetivos. La FAO define deforestación como un cambio en uso de terrenos con la pérdida de la cobertura del dosel arbóreo a menos del 10 %. La degradación del bosque no se incluye en la tasa de deforestación.
Se asume que las extinciones ocurren en alguna proporción con la tasa de deforestación (Lugo y Brown 1996). Algunos autores asumen que la relación entre pérdida de hábitat y extinción de especies es lineal, otros que la relación es exponencial y aún otros que la relación aumenta súbitamente luego de exceder un umbral en la pérdida del hábitat (Fig. 14.2). La magnitud del estimado de extinción varía dependiendo de qué valor de «Z» se utilice.
Wright y Muller Landau (2006) examinaron críticamente las premisas utilizadas en los cálculos de la extinción de especies. En su análisis utilizaron la relación de
263 14 CAP ariel e. lugo | ecología a m i m anera Las velocidades de extinción y evolución de las especies
Figura 14.1. Número de familias de organismos marinos a través 600 millones de años de tiempo geológico. Las flechas con números indican extinciones conocidas. El asterisco señala una crisis
menor
Tabla 14.2
Estimaciones del número de especies de hongos, plantas y animales catalogadas y descritas. Modificado de una tabla publicada por Scheffers et al (2012). Muchos valores son estimados y para conocer las fuentes de información y salvedades consulte la tabla original. Millones es M.
Reino Filo/División Dentro del Filo División principal Catalogadas Descritas
14 264 CAP. ariel e.
| ecología a m i m anera Las
lugo
velocidades de extinción y evolución de las especies
Estimadas Hongos 98,998 45,173-300,000 1.5 M 611,000 9.9 M 3.5-5.1 M 1.62 M Plantas 310,129 390,800 215,644 298,000 Vasculares Magnoliophyta 268,600 223,300-315,903 352,000 Monocots y algunos no-monocots 352,000 Gimnospermos 1,021 846-1,021 1,050 Helechos y aliados 12,000 10,000- 15,000 15,000 Briofitas 16,236 13,370- 23,000 22,750 Algas 12,272 12,205- 12,272 Animales Terrestres 1,233,500 8,740,000 Marinos 193,756 2,210,000 Porifera 6,000 5,500- 10,000 18,000 Cnidaria 9,795 9,000- 11,000 Mollusca 85,000 200,000 Annelida 16,763 30,000 Anthropoda Tropicales 3.611.4 M Arachnida 102,248 60,000- 102,248 600,000 Myriapoda 16,072 8,160- 17,923 90,000 Insecta 1 M720,000- > 1M5 M Coleoptera 360,000- 400,000 1.1 M Dipetera 152,956 240,000 Hemiptera 80,000- 88,000 Hymenoptera 115,000 >300,000 Lepidoptera 174,250 300,000- 500,000 Crustacea 47,000 25,000- 68,171 150,000 20,000 20,000- 25,000 80,000 Plathyhelminthes <25,000 12,000- 80,000 500,000 Nematoda >1 M 7,003 6,100- 7,003 14,000 Echinodermata 12,673 20,000 Otros invertebrados 64,788 80,500 Chordata Mamíferos 5,487 4,300- 5,487 5,500 Aves 9,990 9,000- 9,990 >10,000 10,052 Reptiles 8,734 6,300- 8,734 10,000 Anfibios 6,515 4,950- 6,515 15,000 Peces 31,269 25,000- 31,269 40,000
especie y área, pero tomando en consideración estimaciones más realistas de crecimiento poblacional, tasas de deforestación y crecimiento de bosques secundarios. El resultado fue que para los trópicos de Asia y África, respectivamente, un 21-24 y 16-36 % de las especies estarían amenazadas (mi énfasis) por la extinción para el 2030. No pudieron hacer cálculos confiables para el Neotrópico y por esto, sus datos están muy por debajo de los valores promulgados anteriormente por diferentes autores. El norteamericano William Laurance (2006), quien suele enfocarse en visiones apocalípticas sobre el futuro de los bosques tropicales, criticó fuertemente el trabajo de Wright y Muller Landau (2006) por las bajas tasas de potenciales extinciones que pronostican. En mi opinión, y por las razones que explico más adelante en este capítulo, las estimaciones de Wright y Muller Landau todavía parecen ser muy altas. El valor de ese análisis fue que por primera vez los autores explícitamente presentan
265 14 CAP ariel e. lugo | ecología a m i m anera Las velocidades de extinción y evolución de las especies
Figura 14.2. Variación en la curva entre el porcentaje de especies perdidas por el porcentaje de área deforestada. Cada curva tiene un valor de z distinto en la ecuación S = CA z donde S es el número de especies, A es el área, C es una constante y z es la pendiente de la curva. Cuando z = 1, la curva es recta (Lugo 1988b).
Tabla 14.3
Estimaciones de las tasas de extinción de especies tropicales y las bases utilizadas para sustentar el estimado. Basado en Lugo (1988a).
Estimado Bases del estimado Autor(es)
Una especie por día a una especie por hora entre el 1970 y 2000
30-50 % de todas las especies entre 1970 y 2000
Un millón o más de especies para el final de siglo XX
Tan alto como el 20 % de las especies
50 % de las especies para fines del siglo XX o principios del siglo XXI
Varios cientos de miles de especies en solo unas pocas décadas
25-30 % de todas las especies o de 500,000 a varios millones de especies para el fin del siglo XX
500,000-600,000 especies para el fin del siglo XX
0.75 millones de especies para el fin del siglo XX
33 % o más de todas las especies en el siglo XXI
20-25 % de las especies durante el próximo cuarto de siglo
15 % de las especies y 2 % de las familias de plantas para el fin del siglo XX
Desconocido
Una curva cóncava en la relación entre deforestación y pérdida de especies
De seguir las tendenciasde deforestación observadas
Desconocido
Distintas premisas incluyendo una relación exponencial entre pérdida de hábitat y pérdida de especies
Desconocido
Desconocido
Desconocido
Los bosques tropicales desaparecerán y la mitad de las especies se extinguirán
De seguir las tendencias de deforestación observadas
De seguir las tendencias de deforestación observadas
La pérdida de bosques procede según pronóstico hasta el 2000 y luego se detiene completamente
Myers (1979)
Lovejoy (1980)
National Research Council (1980)
Lovejoy (1981)
Ehrlich y Ehrlich (1981)
Myers (1982)
Myers (1983)
Oldfield (1984)
Peter Raven comunicación personal al World Resources Institute y IIED (1986)
Simberloff (1983)
Norton (1986)
Simberloff (1986)
las premisas de sus análisis de suerte que puedan reproducirse las estimaciones. Las altas estimaciones de Myers, Ehrlich y Ehrlich, Lovejoy y otros reportados en la tabla 14.3 no presentaban las premisas de sus cálculos, por lo que no se pueden verificar y con solo mirarlos es obvio que ninguno se dio, y, por el contrario, los pronósticos fueron exagerados.
Más aún, se desprende del trabajo de He y Hubbell (2011) que matemáticamente los estimados de extinciones basados en la relación de especie-área siempre (mi énfasis) sobreestiman las extinciones. Ellos encontraron que el área requerida para remover una especie es mucho más alta que la requerida para encontrar el primer espécimen de la especie, que es lo que refleja la relación especie-área (Fig. 14.3). La sobreestimación puede ser más del 160 %. Por eso, como vamos a ver a continuación, las estimaciones de extinción de especies siempre son más altas que las observaciones empíricas de extinción. Independientemente de cómo se estime la extinción de especies, Fisher y Bloomber (2010) observaron lo difícil que es detectar la extinción de una especie, inclusive en los grupos mejor conocidos como los
14 266 CAP. ariel e. lugo | ecología a m i m anera Las velocidades de extinción y evolución de las especies
SAR
el
donde individuos de la especie aparecen por primera vez y la curva identificada como EAR indica el área, generalmente mayor, cuando el último individuo de la especie desaparece. Esta situación genera una tercera relación (línea entrecortada) que sugiere que se requiere más área de pérdida de hábitat para reducir el número de especies que las anticipadas por la curva SAR (He y Hubbell (2011).
mamíferos. Muchas especies consideradas extintas reaparecen, particularmente cuando la causa ofrecida para la de extinción es la pérdida del hábitat.
¿Cuándo se considera extinta una especie? En la época moderna, una especie se considera extinta cuando no se observan especímenes de la especie después de búsquedas adecuadas. Sin embargo, a veces especies que se consideraban extintas aparecen, como sucedió con el Peltophryne lemur o sapo concho en Guánica. Igualmente, el camarón Palaemon pandaliformis se encontró en el río Piedras, que corre por el centro del área Metropolitana, donde había escapado a la detección por más de cien años. Además, hay especies que son extirpadas o eliminadas de
267 14 CAP ariel e. lugo | ecología a m i m anera Las velocidades de extinción y evolución de las especies
Figura 14.3. Relación entre el número de especies y el área donde habitan (SAR, por sus siglas en inglés). La curva identificada como
refleja
área
una región sin que se consideren extintas, pues ocurren individuos de la especie en otros lugares. Extinciones en el récord fósil requieren que el taxón desaparezca del récord fósil.
Especies y hábitats más vulnerables a la extinción
Se estima que un 40 % de los anfibios del mundo están en el umbral de sufrir extinción (Wake 2012). Los factores agravantes incluyen el cambio climático (temperatura y humedad), efectos de pesticidas, la acidificación del agua, y los efectos de hongos patógenos (Chytridomycosis). La piel húmeda y delicada de los anfibios los hace vulnerables a cambios en la atmósfera. En Puerto Rico, Rafael Joglar (1998) estima que tres especies de anfibios posiblemente se han extinguido en las últimas décadas. Estas son: Coquí Palmeado (Eleutherodactylus karlschmidti), Coquí Dorado (E. jasperi) y Coquí de Eneida (E. eneidae). Organismos grandes, particularmente animales grandes, son vulnerables a la extinción por causa antrópica y sufrieron altas tasas de extinción ante la depredación de parte de poblaciones humanas primitivas. Especies en hábitats donde los organismos están confinados en el espacio son más vulnerables a la extinción que organismos con oportunidades de escape y dispersión. Por ejemplo, organismos en las islas pequeñas, aisladas y con poco relieve topográfico como las islas del Pacífico u organismos en lagos o canales de agua son particularmente vulnerables a la extinción.
Lecciones de Puerto Rico
Varias características de Puerto Rico hacen del lugar un buen caso de estudio sobre la extinción de especies. En primer lugar, Puerto Rico es una isla y las especies en las islas son más susceptibles a extinción. En segundo lugar, Puerto Rico tiene un largo historial de presencia humana y sufrió un evento de deforestación que casi eliminó la cobertura arbórea de la isla (Fig. 6.7). Teóricamente, Puerto Rico debió sufrir extinciones masivas, pero en realidad no ha perdido especies en la proporción esperada (Lugo 1988a). Un ejemplo son las aves, de las cuales solo se ha documentado una tasa de extinción del 11.6 % (Brash 1987) y en el presente la isla tiene más especies de aves que las que tenía antes del contacto humano debido a la introducción y naturalización de especies. Además, muchas de las extinciones ocurrieron antes del evento de deforestación que culminó en el siglo XX. ¿Cómo se explica el caso de Puerto Rico relativo a lo esperado en términos de la extinción de especies?
El comportamiento de la biota de Puerto Rico después de la deforestación masiva que culminó en el siglo XX ilustra el principio de persistencia que discutieron Botkin et al. (2007) al observar que las extinciones globales a causa del cambio climático no eran de las magnitudes esperadas. Se entiende que hay resiliencia en la biota y que hay mecanismos que contribuyen a la persistencia
14 268 CAP. ariel e. lugo | ecología a m i m anera Las velocidades de extinción y evolución de las especies
de las especies ante los efectos de los cambios ambientales. Algunos de estos mecanismos que contribuyeron a la persistencia de las especies en Puerto Rico después de la deforestación son: (1) La presencia de refugios para las especies, como los cafetales o fragmentos aislados de bosques. (2) La supervivencia de especies en zonas deforestadas o degradadas. (3) La dispersión de especies de los bosques que sobrevivieron a la deforestación a zonas adyacentes. (4) La sucesión ecológica, ya que algunas zonas deforestadas revierten a cobertura boscosa con el abandono de la actividad humana. (5) La alta densidad de especies de los bosques de Puerto Rico, lo que facilita que muchas especies sobrevivan en pequeñas áreas forestadas. (6) La adaptación de la biota puertorriqueña a disturbios no-antropogénicos basada en los ciclos de vida preadaptados a condiciones de disturbio (Lugo y Zimmerman 2002). Por ejemplo, la rápida floración y los ciclos de vida acelerados permiten que las especies sobrevivan después de disturbios y se adapten a sobrevivir en ambientes cambiantes.
La homogenización de las comunidades vegetales
Una de las consecuencias de la extinción de especies es el proceso de la homogenización de la biota a nivel global. El argumento de la homogenización de la biota del planeta se basa en la extinción preferencial de especies endémicas a causa de la actividad humana y su sustitución por especies comunes (McKinney y Lockwood 1999, Lockwood y McKinney 2001). El resultado de tal tendencia sería que eventualmente la biota estaría despojada de sus especies más únicas para ser sustituidas por especies muy comunes. La idea de la homogenización ha sido ilustrada con la biota de las ciudades, donde abundan especies cosmopolitas y donde presumiblemente causan que la biota de las ciudades sea similar en todas partes.
El argumento de la homogenización de especies tiene una lógica muy atractiva y es innegable que de extinguirse todas las especies endémicas, la pérdida genética sería muy significativa para la biodiversidad del planeta. Los conceptos de homogenización de especies deben alertarnos a la importancia de proteger la flora y fauna endémica y de estar continuamente vigilantes a los efectos de la actividad humana sobre las especies del planeta.
Sin embargo, la demostración de que comúnmente ocurre la homogenización de especies no ha sido tan contundente como la idea del concepto mismo. Aunque es cierto que las ciudades europeas comparten similares listados de especies, no es cierto que esas especies estén formando las mismas comunidades en esas ciudades ni que la biota de todas las ciudades del mundo sea la misma. Obviamente, las ciudades tropicales como San Juan tienen una biota muy distinta a la de las ciudades templadas de Europa o las boreales de altas latitudes. Más aún, los estudios que apoyan la homogenización se basan en simples comparaciones de especies
269 14 CAP ariel e. lugo | ecología a m i m anera Las velocidades de extinción y evolución de las especies
(usando índices de similitud) e ignoran la realidad ecológica que las mismas especies, presentes en abundancias relativas disimilares, pueden formar comunidades distintas. Para demostrar que en distintas ciudades se forman comunidades distintas con las mismas especies, se requiere datos sobre la abundancia relativa de las especies y el uso de un índice de similitud que considere estos datos (Índice de Sørenson modificado; Mueller-Dombois y Ellenberg 1974). Tales análisis no se llevan a cabo por falta de datos de abundancia de especies, por lo que se crea la impresión de que todas las ciudades son iguales bióticamente.
En la naturaleza las especies se homogenizan cuando el hábitat es homogéneo. Por ejemplo, humedales monoespecíficos surgen cuando la topografía, clima e hidrología son homogéneos. El ser humano tiende a formar hábitats homogéneos donde se observa homogenización de especies, por ejemplo los campos agrícolas. Muchas ciudades comparten hábitats similares a través de largas distancias y uno debe esperar que compartan la misma biota. Sin embargo, la ciudad es heterogénea, ya que contiene una alta diversidad de hábitats que sin duda albergan distintas combinaciones o comunidades de especies. Es obvio que uno puede concentrar la atención en lo que se percibe cómo homogéneo o en lo heterogéneo.
Recuperando la biodiversidad
Las especies que componen la biota están en continuo movimiento en tiempo y espacio, por lo que siempre existe la posibilidad de que procesos tan intensivos como la deforestación, puedan revertirse. Esto permite la recuperación de la biodiversidad (Lugo et al. 1993, Lugo 1997). La biodiversidad se recupera por medio de procesos de dispersión, establecimiento, crecimiento y sucesión de especies. Los procesos de dispersión son acelerados por agentes abióticos como el viento y el agua y agentes bióticos como la actividad de las aves, los murciélagos, la gente y los insectos. El puertorriqueño Tomás Carlo y sus colegas han demostrado que el pitirre (Tyrannus dominicensis) es un ave cuya actividad incesante mueve semillas de bosques a pastizales, por lo que acelera la restauración de la biodiversidad boscosa en los pastizales de Puerto Rico (Carlo y Morales 2016). En plantaciones monoespecíficas arbóreas es posible observar el establecimiento de sotobosques diversos de especies nativas. Como resultado del uso de la plantación por las aves (Cruz 1987, 1988) y murciélagos (Gannon et al. 2005). La tasa de establecimiento de especies es de 1 por año. Lo mismo ocurre durante la sucesión en bosques secundarios (Brown y Lugo 1990).
Especies introducidas, extinciones y aceleración de la evolución
Es casi axiomático pensar que la segunda causa más importante en la extinción de las especies (después de la pérdida de hábitats) es las especies introducidas. Sin embargo, la demostración de esta causa y efecto no es trivial y existe una literatura
14 270 CAP. ariel e. lugo | ecología a m i m anera Las velocidades de extinción y evolución de las especies
creciente que sugiere que las especies introducidas no causan extinciones de especies en la mayoría de los casos examinados críticamente (Gurevitch y Padilla 2004).
Los cambios en las tasas de procesos ecológicos causados por la introducción de especies son reales, pero no deben confundirse como las causas de la extinción de las especies.
Vellend et al. (2007), señalan que la introducción de especies crea condiciones ideales para promover la diversificación, evolucionaría inclusive el establecimiento de poblaciones alopátricas bajo nuevas condiciones ambientales, creando condiciones ecológicas alteradas para las especies nativas, y creando nuevas condiciones para la hibridación de grupos previamente alopátricos. Otros procesos en juego en esta diversificación evolutiva son: la deriva genética o genetic drift, selección natural direccional, o la selección natural disruptiva (causando valores más bajos de aptitud genética o fitness). Abbott (1992) resaltó la formación de nuevas especies por medio del proceso de hibridación entre una especie nativa y otra introducida. Tales hibridaciones fueron confirmadas con técnicas moleculares y de enzimas.
En Arecibo Puerto Rico, herpetólogos como la norteamericana Kristin Winchell, han estudiado como lagartijos del género Anolis progresivamente se aclimatan y adaptan a los ambientes urbanos. Compararon lagartijos de la especie Anolis cristatellus, una especie urbanófila, con la especie Anolis stratulus, una especie que tolera, pero evita los ambientes urbanos (Winchell et al. 2016). Lagartijos urbanos de A. cristatellus superaba a A. stratulus en experimentos de laboratorio donde individuos competían con base en su capacidad de locomoción sobre distintas superficies urbanas. Los lagartijos urbanos A. cristatellus exhibían cambios en su morfología, incluyendo el tamaño del cuerpo, largo de las extremidades, y modificaciones en los dedos que le permitían mejor adhesión a superficies planas. Tenían además más disposición para explorar nuevos hábitats y podían llevar a cabo todas sus funciones en el ambiente urbano sin necesidad de retornar al ambiente rural. Tales cambios morfológicos y de comportamiento representan los primeros pasos en la evolución de poblaciones urbanas desconectadas del hábitat rural inicial y distintas a las poblaciones que le dieron su origen.
En el capítulo 4 presenté el argumento de Hutchinson (1965) sobre la trama evolutiva en el teatro ecológico (Fig. 4.12). Modifiqué la metáfora de Hutchinson para concluir que en el Antropoceno la actividad del ser humano alteraba el teatro ecológico y la dispersión de especies, dando como resultado la aceleración de la evolución. Thomas (2015) formalizó este argumento y observó que en Europa un mayor número de nuevas especies han surgido en los últimos 300 años que las que se han documentado como habiéndose extinguido durante el mismo periodo de tiempo. Él espera que estas tasas de evolución se acelerarán el futuro cercano (Fig. 14.4), aumentando la biodiversidad en el Antropoceno (Thomas 2013).
271 14 CAP ariel e. lugo | ecología a m i m anera Las velocidades de extinción y evolución de las especies
Durante el Antropoceno, la velocidad de evolución de especies se acelera. GB se refiere a especies en Inglaterra. La unidad «msy» es millones de especies-años, por su sigla en inglés.
¿Estamos en la sexta extinción masiva del planeta?: Barnosky et al. (2011) concluyeron que sí, pero tal respuesta conlleva las siguientes salvedades o caveats: (1) Es dificilísimo comparar tasas de extinción en el presente con extinciones en el récord fósil. (2) Las extinciones causadas por el ser humano hasta el momento son insignificantes comparadas con las extinciones en la tabla 14.1, esto a pesar de que no sabemos cuántas especies se han perdido sin darnos cuenta. (3) Si se extinguen todas las especies hoy consideradas como críticamente en peligro de extinción, observaríamos tasas de extinción que solo se han visto cinco veces en los últimos 540 Ma. (4) De extinguirse las especies clasificadas como en peligro y vulnerables a extinción, entonces en los próximos 200 años llevaríamos al planeta a su sexta extinción masiva. Sin embargo, cuando es necesario predecir un evento biótico tanto tiempo hacia el futuro, y cuando el resultado depende de la actividad humana, es necesario cuestionar si para el ser humano existen futuros alternativos con diferentes resultados.
14 272 CAP. ariel e. lugo | ecología a m i m anera Las velocidades de extinción y evolución de las especies
Figura 14.4. Velocidad de la evolución de especies en Europa a través del tiempo y en el Antropoceno (Thomas 2013). Las barras horizontales indican el rango observable para distintos grupos.
Briggs (2014) examinó data empírica sobre la extinción de especies y encontró que la documentación de extinciones, aun cuando se extrapolen a otros grupos, son mucho más bajas de lo esperado. Argumentó que la biodiversidad aumentaría a consecuencia de la aceleración evolutiva del ser humano. Estimó que el asunto a considerar relativo a la biodiversidad no es la extinción de especies, sino la acumulación de especies que en un momento eran más comunes, pero ahora tienen poblaciones pequeñas y vulnerables a los cambios ambientales.
Pregunta
Considerando los puntos y contrapuntos presentados sobre la extinción y evolución de especies, ¿cómo crees que se deben dirigir los limitados recursos disponibles para la conservación de la biodiversidad?
273 14 CAP ariel e. lugo | ecología a m i m anera Las
de
de
velocidades
extinción y evolución
las especies
CaPítulo 15
La ética de la Tierra
Pregunta
¿Qué importancia tiene la ética en el quehacer científico? ¿Cómo se aplica la ética a la conservación, definida como la relación armónica entre el ser humano y la Tierra?
Definición
La ética se define como un conjunto de comportamientos morales o moralidades. La ética también puede referirse a las teorías o principios que rigen la moral. Dicho en otras palabras:
«para una persona o grupo… el [conjunto] de creencias fundamentales sobre lo que se constituye bueno o malo en las relaciones entre personas (y entre sociedades, seres humanos y otros seres vivientes)»
(traducido de Gasper 2004, p. 18).
Introducción
Un alto sentido de ética es imprescindible para sustentar una carrera científica. El proceso científico requiere que los participantes sean éticos y cuando no lo son, la ciencia sufre. Por ejemplo, nadie supervisa la recolección de datos de un científico; se asume que son recolectados éticamente. Se asume que un científico publica datos una sola vez y que son verídicos. El sistema de publicación de artículos asume que los participantes, lo que incluye a los revisores de los trabajos, operan sin conflicto de interés y sin prejuicios a favor o en contra de ideas o participantes. Algunas profesiones se desempeñan bajo códigos de ética, pero otras profesiones no tienen tales códigos. La biología no tiene códigos de ética para el comportamiento de sus participantes, aunque hay códigos que regulan el uso de animales en los laboratorios de investigación. En este capítulo mi intención es compartir experiencias que he tenido cuando los principios éticos fueron necesarios para guiar la toma de decisiones. Termino el capítulo relacionando la ética con la conservación, específicamente
ariel e. lugo | ecología a m i m anera
275
la conservación de la tierra, según fue concebida por Aldo Leopold en su famoso escrito (Leopold 1949) y la colección de las citas de Leopold por Meine y Knight (1999).
Consultorías
La consultoría representa una opción importante de trabajo para biólogos y ecólogos. Sin embargo, trabajos de consultoría pueden ocultar trampas éticas que pueden afectar reputaciones. Mi primera experiencia fue cuando acepté un trabajo de consultoría en el sur de la Florida, para ese entonces era profesor de la Universidad de la Florida y las reglas universitarias permitían los trabajos de consultoría. En algunas universidades los profesores aprovechan las facilidades que ofrece la universidad con todos sus beneficios (biblioteca, internet gratis, oficinas e incluso estudiantes, entre otras) para trabajar privadamente, lo cual les permite incrementar sus ganancias personales. No siempre estas prácticas son éticas o legales; dependen de las reglas de las universidades. En mi experiencia, los trabajos de consultoría eran muy buenos desde el punto de vista económico. Sin embargo, me enseñaron mi primera lección de ética. Como siempre llevé a cabo mi trabajo científico en el marco público, no sabía que cuando uno firma un contrato de consultoría el trabajo resultante le pertenece al que pagó por el servicio y no al que lo ejecutó. De hecho, el que paga decide el contenido del informe y qué excluye. Después de esta experiencia, nunca más presté mis servicios para causas privadas con ganancia financiera. He dedicado todo mi trabajo al servicio público.
Ejemplos
Conozco personas prominentes en la academia o en la sociedad civil que en el desempeño de sus carreras profesionales han perdido su credibilidad. ¿Qué han hecho profesionalmente para afectar su reputación? Han rendido informes de estudios incompletos, equivocados, tendenciosos, o sesgados a favor de los proyectos de los proponentes que pagaron por los estudios. Específicamente han propuesto que existen manglares de agua dulce que no son sujetos a las mareas y, por ende, pueden ser sustituidos por desarrollos urbanos. Han sugerido que las aguas de la laguna Piñones reflejan condiciones anormales por tener más respiración que fotosíntesis. Por lo tanto, no ameritan la protección que recibirían si tuviesen tasas de fotosíntesis más altas que las tasas de respiración. Todos los argumentos discutidos anteriormente se debatieron en una corte federal donde prevaleció el conocimiento ecológico que sostiene que todos los manglares, sin excepción, están expuestos a la acción de la marea y a la salinidad, aunque estas sean influencias débiles bajo ciertas condiciones. De igual manera, un ecosistema no tiene que ser autotrófico para ameritar conservación. Las cuevas son sistemas heterotróficos con un alto valor de conservación igual que la laguna Piñones.
15 276 CAP ariel e. lugo | ecología a m i m anera La ética de la Tierra
El punto geográfico más alto de Puerto Rico es el cerro Puntas. Una visita al cerro demuestra que está ocupado por antenas y los edificios adyacentes a estas. La forma del cerro está severamente modificada por los cortes hechos para instalar las antenas. El cerro es parte del Bosque Estatal de Toro Negro. ¿Quién tiene la responsabilidad ética? ¿El consultor que justificó la eliminación de esa zona del cerro Puntas, o los oficiales que aprobaron el proyecto?
Silenciando la ciencia
En varias ocasiones, colegas científicos estadounidenses y puertorriqueños han argumentado que hay cierta información científica que no debe divulgarse al público porque tal información ayuda a los que quieren enriquecerse con el desarrollo de los recursos naturales. Además, para llamar la atención del público sobre la conservación es necesario asustarlos un poco, ya que, si piensan que los problemas no son graves, no le van a prestar atención. Ejemplos de información que no debe divulgarse incluyen: (1) la baja tasa de deforestación en los trópicos, (2) bajas tasas de extinción de especies, (3) la resiliencia de los bosques tropicales, (4) la necesidad de deforestar terrenos en Puerto Rico para favorecer la agricultura, (5) la autosuficiencia alimentaria del país, y (6) la resiliencia del clima de Puerto Rico ante el cambio climático. ¿Son éticas estas posiciones? ¿Se deben esconder los resultados de estudios científicos por temor a cómo puedan usar el conocimiento adquirido por grupos o personas que creemos no tienen los mejores intereses públicos en mente?
Siempre he resistido esas sugerencias con base en la noción de que el objetivo de la ciencia es buscar la verdad y es la obligación del científico publicar los resultados de sus investigaciones. El estado del conocimiento ecológico se utiliza generalmente para desarrollar políticas de conservación y tales políticas se benefician de la mejor información disponible. Un científico no puede a priori decidir qué información hace pública y cuál no con base en sus valores personales. De hecho, es responsabilidad del científico minimizar la influencia de valores normativos en sus estudios y expresiones científicas.
El científico individual
La complejidad de la ciencia moderna requiere que los trabajos se lleven a cabo en equipo y que se publiquen con múltiples autores. En el pasado, la mayoría de los científicos trabajaban y publicaban solos, por lo que los problemas de autoría no eran tan comunes como los de hoy. Hay grupos de trabajo que incluyen al director del grupo en todos los escritos, sin importar si contribuyó o no al estudio y al escrito. Piensa que el mero hecho de utilizar su equipo y las facilidades que le otorga su institución (lo que no es cierto, pues generalmente son instalaciones públicas) le da el derecho de ser coautor de los resultados. Tal política no es ética, ya que todo autor de
277 15 CAP ariel e. lugo | ecología a m i m anera La ética de la Tierra
un trabajo científico debe haber contribuido sustancialmente al mismo y debe poder explicar todo lo que se hizo y sus implicaciones. El hecho de ser jefe, supervisor, director de un comité graduado, o dueño de equipo, no es motivo para recibir el crédito de coautoría de forma automática en los trabajos que se realicen bajo su liderazgo.
Conclusión
Todos nos apoyamos sobre los hombros de los que nos precedieron y nos ayudaron a llegar donde estamos. Es importante entender estas circunstancias y darle crédito a quien lo merece, siempre reconocer la ayuda de otros. Los principios éticos y morales nos deben servir de guía al navegar por el complejo mundo de la ciencia moderna.
La ética de la Tierra
Aldo Leopold sostiene que una ética de la tierra cambia el papel del Homo sapiens de conquistador de la comunidad de la tierra a uno de miembro ordinario y ciudadano. Significa respeto tanto para su prójimo como para la comunidad como tal. Añadía Leopold: «La cúspide de la ignorancia es el hombre que dice de un animal o planta: ¿Para qué sirve?» Tal forma de pensar llevó a Leopold a definir la conservación como: «… una situación de armonía entre él [ser humano] y la tierra». Más aún, de acuerdo con Leopold, «[la] motivación fundamental [de la conservación] no es la economía, sino la ética». En el Puerto Rico de hoy y a pesar de lo que dice la constitución del país, la economía tiene prioridad sobre la conservación. Este criterio ignora el hecho de que sin conservación no hay progreso económico, pero, aun así, los conservacionistas invierten mucho esfuerzo en la justificación económica de la conservación, ignorando su valor intrínseco y legítimo a la economía de cualquier país.
Leopold relacionó la conservación con la ética. De acuerdo con el filósofo J. Baird Callicott (1999) la filosofía ambiental comenzó en 1970 y es holística y no-antropogénica.
Es holística porque ve la ética a través del lente de la ecología, considerando la integridad, estabilidad y belleza de la comunidad biótica. La comunidad biótica trasciende los derechos de sus componentes (plantas, animales, suelo y agua). Sin embargo, Callicott, reconoció que existe tensión entre lo holístico no-antropogénico y el individualismo antropogénico, la cual requiere aún explicación.
La ética de la Tierra no es solo la responsabilidad del gobierno o de los que proponen proyectos sin tomar en consideración los aspectos de conservación. Es responsabilidad de todos los seres humanos. Como dijo Leopold en 1938: «La verdadera sustancia de la conservación yace, no en los proyectos físicos de gobierno, sino en los procesos mentales de la ciudadanía». En 1949, señaló que «… el peor obstáculo ante la evolución de una ética de la tierra… es que nuestros sistemas educativo y económico, en vez de acercarnos hacia una conciencia intensa de la tierra,
15 278 CAP ariel e. lugo | ecología a m i m anera La ética de la Tierra
nos alejan». Esta idea es la actual realidad de Puerto Rico y de su sistema educativo y económico. Necesitamos que los diferentes sectores social-ecológico-tecnológicos integren sus visiones y acciones en torno a las ideas de conservación de Leopold en busca de mejores opciones futuras para el bienestar de los puertorriqueños.
El criterio de lo que es correcto o incorrecto para Leopold se centralizaba en nuestra relación con la biota, según se desprende de esta cita: «Algo está en lo correcto cuando tiende a preservar la integridad, estabilidad y belleza de la comunidad biótica. Está equivocado cuando tiende a lo contrario». El mundo, la ciencia y la biota han cambiado desde que Leopold expresó sus ideas sobre la conservación. La comunidad biótica ya no es íntegra, ni estable; continúa siendo bella, pero es inestable, cambiante y novedosa. La conservación se enfrenta a un problema ético al favorecer unas especies sobre otras y perseguir especies no nativas y estados noveles de la comunidad biótica. Por lo tanto, una ética de la tierra reconoce el valor intrínseco de todas las especies y todos los estados de la naturaleza. Tal enfoque es consistente con los principios que esbozó Leopold. Por ejemplo, en 1927 escribió: «La conservación… deberá prevalecer finalmente en todas las tierras, públicas y privadas, o pasar a la historia como un ensueño agradable pero estéril».
Para mí es evidente que debemos:
Desarrollar un nuevo enfoque de la ciencia hacia lo social-ecológico-tecnológico para mejorar como esta informa a la filosofía y la economía y las dirige hacia la formulación de una ética de la Tierra.
Re dirigir la educación, el gobierno y la ciudadanía hacia la aplicación de una ética de la Tierra.
A plicar la conservación a todas las actividades humanas y todos los estados de la comunidad biótica, asegurando omitir prejuicios infundados contra la biota.
279 15 CAP ariel e. lugo | ecología a m i m anera La ética de la Tierra
El ambiente del científico
Cuando conocí a Víctor Shelford en 1965 durante la reunión anual de la Sociedad Ecológica de Norte América (ESA, por sus siglas en inglés), el ambiente científico era muy distinto al del presente. La ESA tenía menos de 3,000 miembros y era posible reunir la sociedad en un campus universitario junto con otras sociedades biológicas. Aprovechaba la ocasión para escuchar charlas de los autores de los libros de texto del momento, por ejemplo a Katherine Esau, la autora del libro de texto de morfología de plantas. Actualmente, la ESA tiene unos 10,000 miembros desparramados por todo el mundo. Cuando se reúnen en gigantescos centros de convenciones, lo hacen solos y a pesar de eso es imposible encontrar a las personas y mucho menos conocer a todos los líderes intelectuales de la ecología moderna que se ha fragmentado en múltiples especialidades. Si el presupuesto de viajes es limitado, hay que pensar si es más conveniente reunirse con los limnólogos, o con los que estudian humedales, ola sociedad de restauración ecológica, o los que se enfocan en sistemas, o mejor con las sociedades que enfatizan lo tropical. Es un ambiente muy complicado y difícil de navegar.
Lo mismo ha ocurrido con la literatura ecológica. Al principio bastaba con leer unas pocas revistas para mantenerse al día de los avances en la ecología. Las bibliotecas eran el lugar esencial para estar actualizado con los avances de la ciencia. Sin embargo, el aumento en el número de personas que se dedican a la investigación ha provocado un aumento acelerado en las publicaciones, lo que a su vez ha incrementado el número de revistas y libros disponibles. Justo cuando las bibliotecas comienzan a perder la capacidad de mantener inventarios actualizados de sus libros y revistas. Tradicionalmente, las revistas las publicaban las sociedades profesionales como la ESA. Hoy es muy difícil para las sociedades profesionales mantener las publicaciones y se ha comercializado la producción de estas. La necesidad de publicar para avanzar en la academia y obtener prestigio ha afectado las tasas de publicación. Lo que anteriormente constituía un estudio y una publicación, ahora se fragmenta en varias publicaciones. Las coautorías se han multiplicado a tal extremo que he visto artículos donde la lista de coautores ocupa más de una página
ariel e. lugo | ecología a mi ma nera Epílogo
281
de la revista. He escrito artículos en revistas donde limitan las citas bibliográficas y, peor aún, donde le dictan al autor la proporción a utilizar en función de la fecha de publicación de los artículos citados. Algunos editores prefieren citas recientes. La consecuencia es que se reinventa la rueda una y otra vez. Los medios de comunicación masiva, apoyados por instituciones que promueven sus publicaciones, continuamente anuncian «descubrimientos» que no lo son y muchas veces ni siquiera representan un avance en el conocimiento. En este ambiente intelectual toman importancia los trabajos de síntesis.
A pesar de lo dicho anteriormente, la tecnología ha revolucionado la ciencia y el científico moderno tiene a su alcance una serie de herramientas que facilitan el quehacer científico y hacen de la profesión un deleite. Para empezar, los equipos de campo que utilicé para mi tesis de maestría y doctoral ocupaban tanto espacio que tuve que agregarle un remolque a mi Volkswagen sedán de 1966, incluyendo un generador de gasolina con una capacidad de 6.5 kilowatts. Hoy en día, una persona puede transportar un equipo portátil que pesa no más de 20 libras para hacer las mismas mediciones y en cuestión de minutos tiene los resultados analizados que a mí me tomaba horas hacer empleando una calculadora Monroe de última generación porque tenía la capacidad de almacenar una constante, lo que aceleraba los cálculos. El científico de hoy tiene mucha movilidad y acceso rápido a cualquier parte de Puerto Rico. Podemos trabajar en el campo todo el día y regresar a la comodidad del hogar por la noche. Si uno tiene el respaldo de una agencia, es posible tener acceso a todo el conocimiento existente gratuitamente por medio de la computadora y las licencias necesarias para accesar a la literatura y programas de computación sin costo alguno. Sin el respaldo de una agencia, las computadoras personales y la internet permiten acceso a cualquier tipo de información que uno necesite. Las comunicaciones permiten establecer contacto con cualquier científico en el mundo y, de hecho, el científico de hoy está conectado a una red donde incluso ciudadanos interesados pueden contactarlo. La riqueza de información disponible, contactos con pares y personas en otros quehaceres no científicos, y oportunidades de ver cualquier lugar electrónicamente facilitan y enriquecen la actividad científica a niveles no imaginados al principio de mi carrera.
El futuro de los bosques tropicales y de la humanidad
La capacidad demostrada de la biota caribeña (incluyendo la de Puerto Rico) para formar comunidades noveles ante la intensa presión de uso antropogénico me da optimismo hacia el futuro. A veces pensamos que la naturaleza necesita de nuestra protección, sin entender que la naturaleza es la que nos protege con su enorme
282 ariel e. lugo | ecología a m i m anera Epílogo
capacidad de adaptación a los cambios ambientales. Los principios de sistemas que regulan el comportamiento de todos los ecosistemas garantizan la supervivencia y la adaptabilidad ante la presión antropogénica (Von Bertalanffy 1968, Odum 1983). Lo que no queda claro es el papel que jugaremos en el futuro. ¿Cuánto tiempo durará la época del Antropoceno? A menos que se descubra una fuente de energía más potente que la energía fósil, el Antropoceno durará mientras duren las reservas de petróleo, o sea no mucho tiempo. Odum y Odum (2001) anticiparon un futuro donde nos enfrentaremos a una reducción significativa en la generación de energía para mantener las ciudades que ahora prevalecen en el planeta. Recomiendan una estrategia de descenso próspero hacia una nueva relación con los sistemas no-antropogénicos. Tal estrategia está basada en una relación más directa con los sistemas ecológicos y el uso de la energía solar, cuya potencia en la generación de energía es menor que la del petróleo. El ser humano mantendrá su posición en la Tierra, pero solo si reduce su huella energética.
283 ariel e. lugo | ecología a m i m anera Epílogo
Abbott, R. J. 1992. Plant invasions, interspecific hybridization and the evolution of new plant taxa. Trends in Ecology & Evolution 7:401-405.
Abelleira Martínez, O. J. 2015. Water use of reforested lands in dry tropical Costa Rica: a functional traits and ecosystem services approach. Disertación. University of Idaho, Moscow, ID.
Ajtay, G. L., P. Ketner, and P. Duvigneaud. 1979. Terrestrial primary production and phytomass. Pages 129-181 in B. Bolin, E. T. Degens, S. Kempe, and P. Ketner, editors. The global carbon cycle. John Wiley & Sons, Chichester, England.
Álvarez, E. 1987-1988. La pintura impresionista de José Mariano García (en su centenario, 1887-1987). Revista de Estudios Generales 2:9-55.
Álvarez Ruiz, M. 2002. Effects of human activities on stand structure and composition, and genetic diversity of Dacryodes excelsa Vahl (tabonuco) forests in Puerto Rico. Disertación. University of Puerto Rico, Río Piedras, PR.
Álvarez Ruiz, M. y A. E. Lugo. 2012. Landscape effects on structure and species composition of tabonuco forests in Puerto Rico: Implications for conservation. Forest Ecology and Management 266:138-147.
Anderson, S., S. Brantley, L. Derry, W. Dietrich, G. Grant, S. Hart, P. Kumar, K. Lohse, W. McDowell, J. McIntosh, N. Moloch, T. Papanicolaou, J. Richardson, D. Richter, C. Riebe, T. Russo, M. Seyfried, S. Thompson y T. White. 2016. A strategy for advancing critical zone science - 3-5 February 2016. CZO Strategy Meeting, Boulder, CO.
Aukema, J. E., T. A. Carlo y J. A. Collazo. 2007. Landscape assessment of tree communities in the northern karst region of Puerto Rico. Plant Ecology 189:101-115.
Bailey, R. G. 1987. Ecoregion-based design for sustainability. Springer, New York, NY.
Bailey, R. G. 1996. Ecosystem geography. Springer, New York, NY.
Bailey, R. G. 2009. Ecosystem geography. From ecoregions to sites. Second edition. Springer, New York, NY.
ariel e. lugo | ecología a m i m anera Literatura
citada
285
Barnosky, A. D, N. Matzke, S. Tomiya, G. O. U. Woga, B. Swartz, T. B. Quental, C. Marchall, J. L. McGuire, E. M. Lindsey, K. C. Maguire, B. Mersey y E. Ferrer. 2011. Has the Earth’s sixth mass extinction already arrived? Nature 471:51-57.
Basnet, K., F. N. Scatena, G. E. Likens y A. E. Lugo. 1993. Ecological consequences of root grafting in tabonuco (Dacryodes excelsa) trees in the Luquillo Experimental Forest, Puerto Rico. Biotropica 25:28-35.
Beard, J. S. 1944. Climax vegetation in tropical America. Ecology 25:127-158.
Beard, J. S. 1949. The natural vegetation of the windward and leeward islands. Oxford Forestry Memoirs 21. The Clarendon Press, Oxford, England.
Beard, J. 1955. The classification of Tropical American vegetation types. Ecology 36:89-100.
Birdsey, R. A. y P. L. Weaver. 1982. The forest resources of Puerto Rico. Resource Bulletin SO-85, USDA Forest Service Southern Forest Experiment Station, New Orleans, Louisiana.
Birdsey, R. A. y P. L. Weaver. 1987. Forest area trends in Puerto Rico. United States Department of Agriculture Forest Service Southern Forest Experiment Station Research Note SO-331, New Orleans, LA.
Botkin, D. B. H. Saxe, M. B. Araujo, R. Betts, R. H. W. Bradshaw, T. Cedhagen, P. Chesson, T. R. Dawson, J. R. Etterson, D. P. Faith, S. Ferrier, S. Guisan, A. S. Hansen, D. W. Hilbert, C. Loehle, C. Margules, M. New, M. J. Sobel y D. R. B. Stockwell. 2007. Forecasting the effects of global warming on biodiversity. BioScience 57:227-236.
Brandeis, T. J., E. Helmer, H. Marcano Vega y A. E. Lugo. 2009. Climate shapes the novel plant communities that form after deforestation in Puerto Rico and the U.S. Virgin Islands. Forest Ecology and Management 258:1704-1718.
Brandeis, T. J., E. H. Helmer y S. N. Oswalt. 2007. The status of Puerto Rico’s forests, 2003. USDA Forest Service, Southern Research Station Resource Bulletin SRS-119, Asheville, NC.
Brandeis, T. J. y J. A. Turner. 2013a. Puerto Rico’s forests, 2009. Southern Research Station, USDA Forest Service Resource Bulletin SRS-191, Ashville, NC. 85 p.
Brandeis, T. J. y J. A. Turner. 2013b. U.S. Virgin Islands’ forests, 2009. Southern Research Station, USDA Forest Service Resource Bulletin SRS-191, Ashville, NC.
Brandeis T. J. y S. N. Oswalt. 2007. The status of U.S. Virgin Islands’ forests, 2004. USDA Forest Service, Southern Forest Experiment Station Resources Bulletin SRS-122, Ashville, NC. 61 p.
Brash, A. R. 1987. The history of avian extinction and forest conversion on Puerto Rico. Biological Conservation 39:97-111.
286 ariel e. lugo | ecología a m i m anera Literatura citada
Briggs, J. C. 2014. Global biodiversity gain is concurrent with declining population sizes. Biodiversity Journal 5:447-452.
Brocard, G. Y., J. K. Willenbring, F. N. Scatena y A. H. Johnson. 2015. Effects of tectonically-triggered wave of incision on riverine exports and soil mineralogy in the Luquillo Mountains of Puerto Rico. Applied Geochemistry 63:586-598.
Brokaw, N., J. K. Zimmerman, M. R. Willig, G. R. Camilo, A. P. Covich, T. A. Crowl, N. Fetcher, B. L. Haines, D. J. Lodge, A. E. Lugo, R. W. Myster, C. M. Pringle, J. M. Sharpe, F. N. Scatena, T. W. Schowalter, W. L. Silver, J. Thompson, D. J. Vogt, K. A. Vogt, R. B. Waide, L. R. Walker, L. L. Woolbright, J. M. Wunderle Jr. y X. Zou. 2012a. Response to disturbance. Páginas 201-271 en N. Brokaw, T. A. Crowl, A. E. Lugo, W. H. McDowell, F. N. Scatena, R. B. Waide y M. R. Willig, editores., A Caribbean forest tapestry: the multidimensional nature of disturbance and response. Oxford University Press, New York, NY.
Brokaw, N., T. A. Crowl, A. E. Lugo, W. H. McDowell, F. N. Scatena, R. B. Waide y M. R. Willig, editores. 2012b. A Caribbean forest tapestry: the multidimensional nature of disturbance and response. Oxford University Press, New York, NY.
Brown, S. y A. E. Lugo. 1982. The storage and production of organic matter in tropical forests and their role in the global carbon cycle. Biotropica 14:161-187.
Brown, S. y A. E. Lugo. 1984. Biomass of tropical forests: a new estimate based on forest volumes. Science 223:1290-1293.
Brown, S. y A. E. Lugo. 1990. Tropical secondary forests. Journal of Tropical Ecology 6:1-32.
Brown, S. y A. E. Lugo. 1992. Above ground biomass estimates for tropical moist forests of the Brazilian Amazon. Interciencia 17:8-18.
Brown, S. y A. E. Lugo. 2017. Trailblazing the carbon cycle of tropical forests from Puerto Rico. Forests 8, 101 doi:10.3390/f8060204.
Brown, S., A. E. Lugo, S. Silander y L. Liegel. 1983. Research history and opportunities in the Luquillo Experimental Forest. USDA Forest Service, Southern Forest Experiment Station, General Technical Report SO-44, New Orleans, LA.
Callicott, J. B. 1999. Land ethics: into terra incognita. Páginas 296-313 en C. Meine y R. L. Knight, editores. The essential Aldo Leopold quotations and commentaries. The University of Wisconsin Press, Madison, WI, USA.
Campbell, N. A. 1993. Biology, Third edition. The Benjamin/Cummings Publishing Company Inc., Redwood City, CA.
Campos Cerqueira, M., W. J. Arendt y J. M. Wunderle Jr. 2017. Have bird distributions shifted along an elevational gradient on a tropical mountain? Ecology and Evolution 2017:1-11.
287 ariel e. lugo | ecología a m i m anera Literatura citada
Cantrell, S. A., D. J. Lodge, C. A. Cruz, L. M. García, J. R. Pérez Jiménez y M. Molina. 2013. Differential abundance of microbial functional groups along the elevation gradient from the coast to the Luquillo Mountains. Ecological Bulletins 54:87-100.
Carlo, T. A. y J. M. Morales. 2016. Generalist birds promote tropical forest regeneration and increase plant diversity via rare‐biased seed dispersal. Ecology 97:1819-1831.
Cassidy, V. M. 2007. Henry Chandler Cowles: pioneer ecologist. Sigel Press, Chicago, IL.
Castilleja, G. 1991. Seed germination and early establishment in a subtropical dry forest. Disertación. Yale University, New Haven, CN.
Caswell, H. 2001. Matrix population models: construction, analysis, and interpretation. 2nd Ed. Sinauer Associates, Inc.
Cavaleri, M. A., S. C. Reed, W. Kolby Smith y T. E. Wood. 2015. Urgent need for warming experiments in tropical forests. Global Change Biology 21:2111-2121.
Chapman, V. J. 1976. Mangrove vegetation. J. Cramer, Leutershausen, Germany.
Cintrón, G. y Y. Schaeffer Novelli. 1984. Methods for studying mangrove structure. Páginas 91-113 en S. C. Snedaker and J. G. Snedaker, editores. The mangrove ecosystem: research methods. United Nations Educational, scientific and cultural organization, Paris, France.
Cintrón, G., A. E. Lugo, D. J. Pool y G. Morris. 1978. Mangroves of arid environments in Puerto Rico and adjacent islands. Biotropica 10:110-121.
Clements, E. S. 1960. Adventures in ecology: half a million miles...from mud to macadam. Pageant Press, Inc., New York, NY.
Clements, F. E y V. E. Shelford. 1939. Bio-ecology. John Wiley & Sons, Inc., New York, NY.
Cobb, M. 2017. 60 years ago, Francis Crick changed the logic of biology. PLoS biology 15:e2003243.
Coleman, D. C. 2010. Big ecology. The emergence of ecosystem science. University of California Press, Berkeley, CA.
Coleman, W. 1986. Evolution into ecology? The strategy of Warming’s ecological plant geography. Journal of the History of Biology 19(2):181-196.
Cowardin, L. M., V. Carter, F. C. Golet y E. T. La Roe. 1979. Classification of wetlands and deep-water habitats of the United States. U.S. Fish and Wildlife Service (70/31), Washington, DC.
Crevoisier C., C. Sweeney, M. Gloor, J. L. Sarmiento y P. Tans. 2010. Regional US carbon sinks from three-dimensional atmospheric CO2 sampling. PNAS 107:18348-18353.
288 ariel e. lugo | ecología a m i m anera Literatura citada
Crick, F. 1970. Central dogma of molecular biology. Nature 227:561-563.
Croker, R. A. 1991. Pioneer ecologist: the life and work of Victor Ernest Shelford 1877-1968. Smithsonian Institute Press, Washington, DC.
Crutzen, P. J. 2002. Geology of mankind. Nature 415:23.
Cruz, A. 1987. Avian community organization in a mahogany plantation in a neotropical island. Caribbean Journal of Science 23:286-296.
Cruz, A. 1988. Avian resource in a Caribbean pine plantation. Journal of Wildlife Management 52:274-279.
Cuevas, E. y A. E. Lugo. 1998. Dynamics of organic matter and nutrient return from litterfall in stands of ten tropical tree plantation species. Forest Ecology and Management 112:263-279.
Cusack, D. F., W. Silver y W. H. McDowell. 2009. Biological oxygen fixation in two tropical forests: ecosystem-level patterns and effects of nitrogen fertilization. Ecosystems 12:1299-1315.
Darwin, C. 1881. The formation of vegetable mould through the action of worms with observation on their habits. Murray, London, England.
Davis Jr, J. H. 1938. Mangroves- makers of land. Nature 31:551-553.
Davis, J. H. 1946. The peat deposits of Florida. Their occurence, development, and uses. Geological Bulletin 30 Florida Geological Survey, Tallahassee FL. 246 p.
del Llano, M. 1988. Inventario de terrenos anegadizos y hábitats de aguas profundas de Puerto Rico. Acta Científica 2: contraportada.
Departamento de Recursos Naturales. 1973. Uso de terreno en Puerto Rico. División de Inventario Científico, San Juan, PR.
Dickinson, M. B., S. M. Hermann, and D. F. Whigham. 2001. Low rates of background canopy-gap disturbance in a seasonally dry forest in the Yucatan Peninsula with a history of fires and hurricanes. Journal of Tropical Ecology 17:895-902.
Diffenbaugh, N. S. y C. B. Field. 2013. Changes in ecologically critical terrestrial climate conditions. Science 341:486-492.
Domínguez Cristóbal, C. M. 2000. Panorama histórico forestal de Puerto Rico. Editorial de la Universidad de Puerto Rico, Río Piedras, PR.
Domínguez Cristóbal, C. M. 2011. Manual interdisciplinario de investigación forestal. USDA Forest Service International Institute of Tropical Forestry, Río Piedras, P.R., 146 p.
Dowler, C. C. y F. H. Tschirley. 1970. Effects of herbicides on a Puerto Rican rain forest. Páginas B-315- B-323 en Odum HT, Pigeon RF, editores. A tropical rain forest. National Technical Information Service, Springfield, VA.
Drew, A. P, J. D. Boley, Y. Zhao, M. H. Johnston y F. H. Wadsworth. 2009. Sixty-two years of change in subtropical wet forest structure and composition at El Verde, Puerto Rico. Interciencia 34:34-40.
289 ariel e. lugo | ecología a m i m anera Literatura citada
Dunevitz, V. L. 1985. Regrowth of clearcut subtropical dry forest: mechanisms of recovery and quantification of resilience. Tesis. Michigan State University, East Lansing, MI.
Egerton, F. N. 2015. A centennial history of the Ecological Society of America. CRC Press, Taylor & Francis Group, Boca Raton, FL.
Egler, F. E. 1942. Indigene versus alien in the development of arid Hawaiian vegetation. Ecology 23:14-23.
Egler, F. E. 1952. Southeast saline Everglades vegetation, Florida, and its management. Vegetatio 3:213-265.
Ehrlich, P. and A. Ehrlich. 1981. Extinctions. The causes of the disappearance of species. Random House, New York, NY.
Ellis, E. C. 2015. Ecology in an anthropogenic biosphere. Ecological Monographs 85:287-331.
Ellis, E. C. 2021. Land Use and Ecological Change: a 12,000-Year History. Annual Review of Environment and Resources 46:1-33.
Ellis E. C., K. K. Goldewijk, S. Siebert, D. Lightman y N. Ramankutty. 2010. Anthropogenic transformation of the biomes, 1700 to 2000. Global Ecology and Biogeography 19: 589-606.
Elton, C. S. 1958. The ecology of invasions by animals and plants. Chapman and Hall, London, England.
Erickson, H. E., E. H. Helmer, T. J. Brandeis y A. E. Lugo. 2014. Controls of fallen leaf chemistry and forest floor element masses in native and novel forests across a tropical island. Ecosphere 5(4):48.
Erwin, D. H. 1998. The end and the beginning: recoveries from mass extinctions. Trends in Ecology & Evolution 13:344-349.
Evans, F. 1956. Ecosystem as the basic unit in ecology. Science 123:1127-1128.
Evans, J. 1992. Plantation forestry in the tropics, Second edition. Clarendon Press, Oxford, England.
Ewel, J. J. 1971. Experiment in arresting succession with cutting and herbicides in five tropical environments. Disertación. University of North Carolina, Chapel Hill, NC.
Ewel, J. J., y A. Madriz. 1968. Zonas de vida de Venezuela. Memoria explicativa sobre el mapa ecológico. Ministerio de Agricultura y Cria, Dirección de Investigación, República de Venezuela, Caracas, Venezuela.
Ewel, J. J. y J. L. Whitmore. 1973. The ecological life zones of Puerto Rico and the U.S. Virgin Islands. Forest Service Research Paper ITF-18, Institute of Tropical Forestry, USDA Forest Service, Río Piedras, PR.
Ewel, K. C. y H. T. Odum, editores. 1984. Cypress swamps. University Presses of Florida, Gainesville, FL.
290 ariel e. lugo | ecología a m i m anera Literatura citada
FAO. 2007. Situación de los bosques del mundo 2007. The Food and Agriculture Organization of the United Nations, Rome, Italy.
Figueroa Colón, J. C., editor. 1996. The scientific survey of Puerto Rico and the Virgin Islands: an eighty-year reassessment of the island’s natural history. Annals of the New York Academy of Sciences 776:1-273.
Figueroa, J. C., C. L. Totti, A. E. Lugo y R. O. Woodbury. 1984. Structure and composition of moist coastal forests in Dorado, Puerto Rico. Página 11. USDA Forest Service, Southern Forest Experiment Station Research Paper SO-202, New Orleans, LA.
Fisher, D. O. y S. P. Blomberg. 2010. Correlates of rediscovery and the detectability of extinction in mammals. Proceedings of the Royal Society 10.1098/rspb.2010.1579.
Fonseca da Silva, J. 2011. Ecophysiology and productivity of Castilla elastica, an introduced tropical tree species. University of Puerto Rico, Rio Piedras, Puerto Rico.
Francis, J. K. 1995. Forest plantations in Puerto Rico. Páginas 210-223 en A. E. Lugo y C. A. Lowe, editores. Tropical forests: management and ecology. Springer-Verlag, New York, NY.
Franco, P. A., P. L. Weaver y Eggen McIntosh. 1997. Forest resources of Puerto Rico, 1990. Resource Bulletin SRS-22, USDA Forest Service, Southern Research Station, Ashville, North Carolina.
Frangi, J. L. 1983. Las tierras pantanosas de la montaña Puertorriqueña. Páginas 233-247 en A. E. Lugo, editor. Los bosques de Puerto Rico. USDA Forest Service Institute of Tropical Forestry and Puerto Rico Department of Natural Resources, San Juan, PR.
Frangi, J. L. y A. E. Lugo. 1985. Ecosystem dynamics of a subtropical floodplain forest. Ecological Monographs 55:351-369.
Frangi, J. L. y A. E. Lugo. 1998. A flood plain palm forest in the Luquillo Mountains of Puerto Rico five years after hurricane Hugo. Biotropica 30:339-348.
Gannon, M. R., A. Kurta, A. Rodriguez Durán y M. R. Willig. 2005. Bats of Puerto Rico. An island focus and a Caribbean perspective. Texas Tech University Press, Lubbock, TX.
García Martinó, A. R., G. S. Warner, F. N. Scatena y D. L. Civico. 1996. Rainfall and elevation relationships in the Luquillo Mountains of Puerto Rico. Caribbean Journal of Science 32:413-424.
Gasper, D. 2004. The ethics of development. Edinburgh University Press, Edinburgh, England.
Ghilarov, M. S. 1983. Darwin’s formation of vegetable mould-its philosophical basis. Páginas 1-4 en J. E. Satchell, editor. Earthworm ecology: from Darwin to vermiculture. Chapman and Hall, London, England.
291 ariel e. lugo | ecología a m i m anera Literatura citada
Gifford, J. C. 1905. The Luquillo Forest Reserve, Porto Rico. U.S. Department of Agriculture, Division of Forestry, Bulletin 54, Washington, DC.
Gleason, H. A. 1926. The individualistic concept of the plant association. Bulletin of the Torrey Botanical Club 53:7-26.
Gleason, H. A. y M. T. Cook. 1926. Plant ecology of Porto Rico. Páginas 1-173 en Scientific Survey of Porto Rico and the Virgin Islands. New York Academy of Sciences, New York, NY.
Golley, F. B. 1993. A history of the ecosystem concept in ecology: more than the sum of the parts. Yale University Press, New Haven, CN.
Golley, F. B., H. T. Odum y R. F. Wilson. 1962. The structure and metabolism of a Puerto Rican red mangrove forest in May. Ecology 43:9-19.
González, G. y D. J. Lodge. 2017. Soil biology research across latitude, elevation, and disturbance gradients: a review of forest studies from Puerto Rico during the past 25 years. Forests 8, 178, doi:10.3390/ f8060178.
González, G. G. y M. M. Luce. 2013. Woody debris characterization along an elevation gradient in northeastern Puerto Rico. Ecological Bulletins 54:181-193.
González, G. G., M. R. Willig y R. B. Waide. 2013. Ecological gradient analyses in a tropical landscape. Ecological Bulletin 54:7-250.
Goodland, R. J. 1975. The tropical origin of ecology: Eugen Warming’s jubilee. Oikos 26:240-245.
Gould, W. A., C. Alarcón, B. Fevold, M. E. Jiménez, S. Martinuzzi, G. Potts, M. Quiñones, M. Solórzano y E. Ventosa. 2008. The Puerto Rico gap analysis project. Volumen 1: land cover, vertebrate species distributions, and land stewardship. General Technical Report IITFGTR-39, USDA Forest Service International Institute of Tropical Forestry, Río Piedras, PR.
Graff, O. 1983. Darwin on earthworms-the contemporary background and what the critics thought. Páginas 5-18 en J. E. Satchell, editor. Earthworm ecology: from Darwin to vermiculture. Chapman and Hall, London, England.
Guariguata, M. R. 1990. Landslide disturbance and forest regeneration in the upper Luquillo Mountains of Puerto Rico. Journal of Ecology 78:814-832.
Guariguata, M. R. y M. C. Larsen. 1990. Preliminary map showing locations of landslides in El Yunque quadrangle, Puerto Rico. United States Department of Interior, United States Geological Survey, Open File Report 39-257.
292 ariel e. lugo | ecología a m i m anera Literatura citada
Gunderson, L. H. y C. S. Holling, editores. 2002. Panarchy. Understanding transformations in human and natural systems. Island Press, Washington, DC.
Gurevitch, J. y D. K. Padilla. 2004. Are invasive species a major cause of extinctions? TRENDS in Ecology and Evolution 19:470-474.
Gutiérrez del Arroyo Santiago, O. 2014. Soil respiration of a novel subtropical moist forest: from diel to seasonal patterns. MS Thesis. University of Puerto Rico, Río Piedras, PR.
Gutiérrez del Arroyo, O. y T. A. Wood. 2019. Large seasonal variation of soil respiration in a secondary tropical moist forest in Puerto Rico. Ecology and Evolution 2020;00:1-10.
Harris, N. L. y A. E. Lugo. 2012. Disturbances. Páginas 21-49 en N. L. Harris, A. E. Lugo, S. Brown y T. Heartsill Scalley, editores. Luquillo Experimental Forest: research history and opportunities. U.S. Department of Agriculture Forest Service. EFR-1, Washington, DC.
Harris, N. L. y E. Medina. 2013. Changes in leaf properties across an elevation gradient in the Luquillo Mountains, Puerto Rico. Ecological Bulletins 54:169-179.
Harris, N. L., A. E. Lugo, S. Brown y T. Heartsill Scalley, editores. 2012. Luquillo Experimental Forest: research history and opportunities. U.S. Department of Agriculture, EFR-1, Washington, DC.
Harris, N. L, C. A. S. Hall y A. E. Lugo. 2013. Gradients, primary productivity and maximum power in Puerto Rico’s Luquillo Mountains. Ecological Bulletin 54:233-243.
Hayes, M. H. B. 1983. Darwin’s ‘vegetable mould’ and some modern concepts of humus structure and soil aggregation. Páginas 19-33 en J. E. Satchell, editor. Earthworm ecology: from Darwin to vermiculture. Chapman and Hall, London, England.
He, F. y S. P. Hubbell. 2011. Species-area relationships always overestimate extinction rates from H loss. Nature 473:368-371.
Heald, E. J. 1971. The production of organic detritus in a south Florida estuary. Miami Sea Grant Technical Bulletin 6:1-110.
Heartsill Scalley, T. 2012. Vegetation. Páginas 64-72 en N. L. Harris, A. E. Lugo, S. Brown y T. Heartsill Scalley, editores. Luquillo Experimental Forest: research history and opportunities. U.S. Department of Agriculture, EFR-1, Washington, DC.
Herbertson, A. J. 1905. The major natural regions: an essay in systematic geography. Geography Journal 25:300-312.
Hill, R. T. 1899. Notes on the forest conditions of Porto Rico. U.S. Department of Agriculture, Division of Forestry, Bulletin 25, Washington, DC.
Holdridge, L. R. 1947. Determination of the world plant formations from simple climatic data. Science 105:367-368.
293 ariel e. lugo | ecología a m i m anera Literatura citada
Holdridge, L. R. 1967. Life zone ecology. Tropical Science Center, San José, Costa Rica.
Holling, C. S. 1973. Resiliency and stability of ecological systems. Annual Review of Ecology and Systematics 4:1-23.
Hook, D. D. y J. R. Scholtens. 1978. Adaptations and flood tolerance of tree species. Páginas 299-331 en D. D. Hook and R. M. M. Crawford, editores. Plant life in anaerobic environments. Ann Arbor Science, Ann Arbor, MI.
Houghton, R. 2000. A new estimate of global sources and sinks of carbon from land-use change. EOS. 81. S281.
Howard, R. A. 1968. The ecology of an elfin forest in Puerto Rico, 1. Introduction and composition studies. Journal of the Arnold Arboretum 49:381-418.
Humboldt, A., von, and A. Bonpland. 1807. Essai sur la géographie des plantes; Accompagné d’un tableau physique des régions equinoxiales. Levrault, Schoell et Compagnie, Paris, France.
Hutchinson, G. E. 1957. A treatise on limnology. Geography, physics, and chemistry. John Wiley & Sons, Inc., New York, NY, USA.
Hutchinson, G. E. 1965. The ecological theater and the evolutionary play. Yale University Press, New Haven, CN.
Hutchinson, G. E. 1967. A treatise on limnology. Introduction to lake biology and the limnoplankton. John Wiley & Sons Inc., New York, NY, USA.
Hutchinson, G. E. 1975. A treatise on limnology. Limnological botany. Wiley Interscience Publications, New York, NY, USA.
Hutchinson, G. E. (edited by Y. H. Edmondson) 1993. A treatise on limnology. The zoobenthos. John Wiley & Sons Inc., New York, NY, USA
Intergovernmental Panel on Climate Change. 2007. Climate change 2007: the physical science basis. Contribution of Working Group I to the fourth assessment report of the Intergovernmental Panel on Climate Change [Solomon, S., D. Qin, M. Manning, Z. Chen, M. Marquis, K. B. Averyt, M. Tignor y H. L. Miller (editores)]. Cambridge University Press, Cambridge, United Kingdom.
Jackson, S. T., and S. Romanowski, editors. 2009. Essay on the geography of plants. The University of Chicago Press, Chicago, IL, USA.
Joglar, R. L. 1998. Los coqíes de Puerto Rico. Su historia natural y conservación. Editorial de la Universidad de Puerto Rico, Río Piedras, PR.
Jordan, C. F. 1971. Productivity of a tropical forest and its relation to a world pattern of energy storage. Journal of Ecology 59:127-142.
Joyce, J. 1992. Geology of the east coast of Puerto Rico field guide. Department of Geology, University of Puerto Rico Mayagüez, Puerto Rico.
294 ariel e. lugo | ecología a m i m anera Literatura citada
Kangas, P. C. 1990. An energy theory of landscape for classifying wetlands. Páginas 15-23 en A. E. Lugo, M. M. Brinson y S. Brown, editores. Forested wetlands. Elsevier, Amsterdam.
Kemp, W. M. 1977. Energy analysis and ecological evaluation of a coastal power plant. Disertación, University of Florida, Gainesville, FL.
Kennaway, T. y E. H. Helmer. 2007. The forest types and ages cleared for land development in Puerto Rico. GIScience & Remote Sensing 44:356-382.
Kingsland, S. E. 2005. The evolution of American ecology, 1890-2000. The Johns Hopkins University Press, Baltimore.
Kormondy, E. J. editor. 1965. Readings in ecology. Prentice-Hall International, London, England.
Kueffer, C. y C. N. Kaiser-Bunbury. 2013. Reconciling conflicting perspectives for biodiversity conservation in the Anthropocene. Frontiers in Ecology and the Environment 12(2):131-137.
Larsen, M. C. 2000. Analysis of 20th century rainfall and streamflow to characterize drought and water resources in Puerto Rico. Physical Geography 21:494-521.
Laurance, W. F. 2006. Have we overstated the tropical biodiversity crisis? TRENDS in Ecology & Evolution 22:65-70.
Ledru, A. P. 1797. Viaje a la isla de Puerto Rico. Editado por Manuel A. Domenech Ball, Oficina Historiador Oficial de Puerto Rico, San Juan, PR.
Leopold, A. B. 1933. Game management. Charles Scribner’s Sons, New York, NY.
Leopold, A. 1949. A sand county almanac and sketches here and there. Oxford University Press, New York.
Leslie, P. H. y R. M. Ranson. 1940. The mortality, fertility and rate of natural increase of the vole (Microtus agrestis) as observed in the laboratory. Animal Ecology 9:27-52.
Lewis, S. L y M. A. Maslin. 2015. Defining the Anthropocene. Nature 519:171-180.
Liebig, J. 1840. Chemistry in its applications to agriculture and physiology. 4th edition. Taylor and Walton, London.
Little, E. L. 1970. Relationship of trees of the Luquillo Experimental Forest. Páginas B-47-B-58 en H. T. Odum y R. F. Pigeon, editores. A tropical rain forest. National Technical Information Service, Springfield, VA.
Lockwood, J. L. y M. L. McKinney, editores. 2001. Biotic homogenization. Kluer Academic/Plenum Publishers, New York, NY.
Lotka, A. J. 1956. Elements of mathematical biology. Dover Publications, New York, NY.
295 ariel e. lugo | ecología a m i m anera Literatura citada
Lovejoy, T.E. 1980. A projection of species extinctions. Páginas 328331, volumen 2 en G.O. Barney (study director). The global 2000 report to the President. Entering the twenty-first century. Council on Environmental Quality, Washington, D.C
Lovejoy, T. E. 1981. Prepared statement in Tropical deforestation, an overview, the role of international organizations, the role of multinational corporations. Pages 175-178 Subcommittee on International Organizations of the Committee on Foreign Affairs. US Printing Office, Washington, D.C.
Lugo, A. E. 1978. Stress and ecosystems. Páginas 62-101 en Thorp JH, Gibbons JW, eds. Energy and environmental stress in aquatic systems. National Technical Information Services, Springfield, VA.
Lugo, A. E. 1980. Mangrove ecosystems: Successional or steady state? Biotropica 12 (supplement 2):65-72.
Lugo, A. E., editor. 1983. Los bosques de Puerto Rico. USDA Forest Service and Department of Natural Resources, Río Piedras, PR.
Lugo, A. E. 1984. A review of early forested wetland literature in the United States. Páginas 7-15 en K. C. Ewel y H. T. Odum, editores. Cypress swamps. University Presses of Florida, Gainesville, FL.
Lugo, A. E. 1988a. Estimating reductions in the diversity of tropical forest species. Páginas 58-70 en E. O. Wilson y F. M. Peter, editores. Biodiversity. National Academy Press, Washington, D.C.
Lugo, A. E. 1988b. Diversity of tropical species: questions that elude answers. Biology International Special Issue 19:1-37.
Lugo, A. E. 1989. Biosphere reserves in the tropics: An opportunity for integrating wise use and preservation of biotic resources. Páginas 53-67 en J. W. P. Gregg, S. L. Krugman y J. D. Wood, editores. Proceedings of the symposium on biosphere reserves. Fourth World Wilderness Congress. U.S. Department of Interior, National Park Service, Atlanta, GA.
Lugo, A. E. 1990. Introduction. Páginas 1-14 en A. E. Lugo, M. M. Brinson y S. Brown, editores. Forested wetlands. Elsevier, New York, NY.
Lugo, A. E. 1991. Dominancia y diversidad de plantas en Isla de Mona. Acta Científica 5:65-71.
Lugo, A. E. 1992. Comparison of tropical tree plantations with secondary forests of similar age. Ecological Monographs 62:1-41.
Lugo, A. E. 1995a. A review of Dr. Odum’s early publications: from bird migration studies to Scott Nixon’s turtle grass model. Páginas 3-10 en C. A. S. Hall, editor. Maximum power: the ideas and applications of H.T. Odum. University Press of Colorado, Niwot, CO.
Lugo, A. E. 1995b. Contributions of H.T. Odum to tropical ecology. Páginas 23-24 en C. A. S. Hall, editor. Maximum power: the ideas and applications of H.T. Odum. University Press of Colorado, Niwot, CO.
296 ariel e. lugo | ecología a m i m anera Literatura citada
Lugo, A. E. 1995c. Reminiscences of H.T. Odum with a focus on his educational techniques. Página 359 en C. A. S. Hall, editor. Maximum power: the ideas and applications of H.T. Odum. University Press of Colorado, Niwot, CO.
Lugo, A. E. 1996. Ninety years of plant ecology research in Puerto Rico. Annals of the New York Academy of Sciences 776:73-88.
Lugo, A. E. 1997. The apparent paradox of re-establishing species richness on degraded lands with tree monocultures. Forest Ecology and Management 99:9-19.
Lugo, A. E. 1999a. Metodología para estimar el almacenaje y flujo de nutrientes en la hojarasca y suelo de bosques secundarios. Acta Científica 13:11-26.
Lugo, A. E. 1999b. Point-counterpoints on the conservation of big-leaf mahogany. USDA Forest Service General Technical Report WO-64, Washington, D.C.
Lugo, A. E. 2003. Obituary: Professor H.T. Odum (1924-2002). Tropical Ecology 44:267-268.
Lugo, A. E. 2004a. H.T. Odum and the Luquillo Experimental Forest. Ecological Modelling 178:65-74.
Lugo, A. E. 2004b. The homogeocene in Puerto Rico. Páginas 366-375 en D. J. Zarin, J. R. R. Alavalapati, F. E. Putz y M. Schmink, editors. Working forests in the Neotropics: conservation through sustainable management? Colombia University Press, New York, NY.
Lugo, A. E. 2005a. Los bosques. Páginas 395-548 en R. L. Joglar, editor. Biodiversidad de Puerto Rico. Vertebrados terrestres y ecosistemas. Editorial del Instituto de Cultura Puertorriqueña, San Juan, PR.
Lugo, A. E. 2005b. The ecosystems of the Luquillo Mountains. Acta Científica 19:3-5.
Lugo, A. E. 2006. Lecciones ecológicas de una isla que lo ha visto todo. Ecotropicos 19:1-13.
Lugo, A. E. 2008. Visible and invisible effects of hurricanes on forest ecosystems: an international review. Austral Ecology 33:368-398.
Lugo, A. E. 2013. Novel tropical forests: nature’s response to global change. Tropical Conservation Science 6:325-337.
Lugo, A. E. 2014. Evolving conservation paradigms for the Anthropocene. Páginas 47-59. USDA Forest Service RMRS-P-71, Fort Collins, CO.
Lugo, A. E. 2020. Effects of extreme disturbance events: from ecesis to social-ecological-technological systems. Ecosystems 23:1726-1747.
297 ariel e. lugo | ecología a m i m anera Literatura citada
Lugo, A. E. 2021. The Luquillo Experimental Forest: A Neotropical example of the interaction between forest conservation and long-term ecological research Páginas 119-149 en R. B. Waide y S. E. Kingsland, editores. The challenges of long-term ecological research: A historical analysis. Springer International Publishing, Switzerland AG.
Lugo, A. E. y S. C. Snedaker. 1974. The ecology of mangroves. Annual Review Ecology and Systematics 5:39-64.
Lugo, A. E. y G. Cintrón. 1975. The mangrove forests of Puerto Rico and their management. Páginas 825-846 en Proceedings of international symposium on biology and management of mangroves. Institute of Food and Agricultural Sciences, University of Florida, Gainesville, FL. 846 p., Honolulu, HI.
Lugo, A. E. y C. Patterson Zucca. 1977. The impact of low temperature stress on mangrove structure and growth. Tropical Ecology 18:149-161.
Lugo, A. E. y G. L. Morris. 1982. Los sistemas ecológicos y la humanidad. Secretaría General de la Organización de los Estados Americanos Programa Regional de Desarrollo Científico y Tecnológico, Washington, DC.
Lugo, A. E. y J. Figueroa 1985. Performance of Anthocephalus chinensis in Puerto Rico. Canadian Journal of Forest Research 15:577-585.
Lugo, A. E. y P. G. Murphy. 1986. Nutrient dynamics of a Puerto Rican subtropical dry forest. Journal of Tropical Ecology 2:55-76.
Lugo, A. E. y S. Brown. 1986. Steady state ecosystems and the global carbon cycle. Vegetatio 68:83-90.
Lugo, A. E. y P. G. Murphy. 1986. Nutrient dynamics of a Puerto Rican subtropical dry forest. Journal of Tropical Ecology 2:55-76.
Lugo, A. E. y S. Brown. 1986. Steady state ecosystems and the global carbon cycle. Vegetatio 68:83-90.
Lugo, A. E. y S. Brown. 1988. The wetlands of the Caribbean. Acta Científica 2:48-61.
Lugo, A. E. y F. H. Wadsworth. 1990. Dacryodes excelsa Vahl. Tabonuco. Páginas 284-287 en R. M. Burns y B. H. Honkala, editores. Silvics of North America. Agriculture Handbook No. 654. USDA Forest Service, Washington, D.C.
Lugo, A. E. y S. Brown. 1991. Comparing tropical and temperate forests. Páginas 319-330 en J. Cole, G. Lovett y S. Findlay, editores. Comparative analysis of ecosystems: patterns, mechanisms, and theories. Springer-Verlag, New York, NY, USA.
Lugo, A. E. y F. N. Scatena. 1995. Ecosystem-level properties of the Luquillo Experimental Forest with emphasis on the tabonuco forest. Páginas 59-108 en A. E. Lugo y C. Lowe, editores. Tropical forests: management and ecology. Springer Verlag, New York, NY, USA.
298 ariel e. lugo | ecología a m i m anera Literatura citada
Lugo, A. E. y S. Brown. 1996. Management of land and species richness in the tropics. Páginas 280-295 en R. C. Szaro y D. W. Johnston, editores. Biodiversity in managed landscapes. Theory and practice. Oxford University Press, New York, NY.
Lugo, A. E. y J. K. Zimmerman. 2002. Ecological life histories. Páginas 191-213 en J. A. Vozzo, editor. Tropical tree seed manual. Agriculture Handbook 721. USDA Forest Service, Washington, DC., USA.
Lugo, A. E. y E. Helmer. 2004. Emerging forests on abandoned land: Puerto Rico’s new forests. Forest Ecology and Management 190:145-161.
Lugo, A. E. y N. L. Harris. 2012. Ecosystem processes. Páginas 98-106 en N. L. Harris, A. E. Lugo, S. Brown y T. Heartsill Scalley, editores. Luquillo Experimental Forest: research history and opportunities. U.S. Department of Agriculture Forest Service, EFR-1, Washington, D.C.
Lugo, A. E. y E. Medina. 2014. Mangrove forests. Páginas 343-352 en Encyclopedia of Natural Resources: Land. Taylor and Francis, New York, NY.
Lugo, A. E. y H. E. Erickson. 2017. Novelty and its ecological implications to dry forest functioning and conservation. Forests 8:161
Lugo, A. E., J. A. González Liboy, B. Cintrón y K. Dugger. 1978. Structure, productivity, and transpiration of a subtropical dry forest in Puerto Rico. Biotropica 10(4):278-291.
Lugo, A. E., S. Brown y M. M. Brinson. 1990a. Concepts in wetland ecology. Páginas 53-85 en A. E. Lugo, M. M. Brinson y S. Brown, editores. Forested wetlands. Elsevier, New York, NY.
Lugo, A. E, E. Cuevas y M. J. Sánchez. 1990b. Nutrients and mass in litter and top soil of ten tropical tree plantations. Plant and Soil 125:263-280.
Lugo, A. E., J. A. Parrotta y S. Brown. 1993. Loss in species caused by tropical deforestation and their recovery through management. Ambio 22:106-209.
Lugo, A. E., J. S. Baron, T. P. Frost, T. W. Cundy y P. Dittberner. 1999a. Ecosystem processes and functioning. Páginas 219-254 en R. C. Szaro, N. C. Johnson, W. T. Sexton, A. y J. Malk, editores. Ecological stewardship: A common reference for ecosystem management. Elsevier Science, Oxford, UK.
Lugo, A. E., S. L. Brown, R. Dodson, T. S. Smith y H. H. Shugart. 1999b. The Holdridge life zones of the conterminous United States in relation to ecosystem mapping. Journal of Biogeography 26:1025-1038.
Lugo, A. E, L. Miranda Castro, A. Vale, T. dM. López, E. Hernández Prieto, A. García Martinó, A. R. Puente Rolón, A. G. Tossas, D. A. McFarlane, T. Miller, A. Rodríguez, J. Lundberg, J. Thomlinson, J. Colón, J. H. Schellekens, O. Ramos y E. Helmer. 2001. Puerto Rican karst – A. vital resource. USDA Forest Service, General Technical Report WO-65, Washington, DC.
299 ariel e. lugo | ecología a m i m anera Literatura citada
Lugo, A.E., F.N. Scatena, W. Silver, S. Molina Colón y P.G. Murphy 2002. Resilience of tropical wet and dry forests in Puerto Rico. Páginas 195225 en L. H. Gunderson, L. Pritchard Jr., editores. Resilience and the behavior of large-scale systems. Island Press, Washington, DC. 287 p.
Lugo, A. E., J. C. Figueroa Colón y M. Alayón, editores. 2003. Big-leaf mahogany: genetics, ecology, and management. Springer Verlag, New York, NY.
Lugo, A. E., E. Medina, J. C. Trejo Torres y E. Helmer. 2006. Botanical and ecological basis for the resilience of Antillean dry forests. Páginas 359381 en R. T. Pennington, G.P. Lewis y A. Ratter, editores. Neotropical savannas and seasonally dry forests. Taylor & Francis, Boca Raton, FL. USA.
Lugo, A. E., J. Fonseca da Silva y A. Sáez Uribe. 2008. Balance de carbono en un bosque de Castilla elastica: resultados preliminares. Acta Científica 22:13-28.
Lugo, A. E., O. J. Abelleira, A. Collado, C. A. Viera, C. Santiago, D. O. Vélez, E. Soto, G. Amaro, G. Charón, H. Colón-Jr., J. Santana, J. I. Morales, K. Rivera, L. Ortíz, L. Rivera, M. Maldonado, N. Rivera y N. J. Vázquez. 2011. Allometry, biomass, and chemical content of novel African tulip tree (Spathodea campanulata) forests in Puerto Rico. New Forests 42:267-283.
Lugo A. E., T. A. Carlo y J. A. Wunderle, Jr. 2012a. Natural mixing of species: novel plant-animal communities on Caribbean Islands. Animal Conservation 15:233-241.
Lugo, A. E., O. Abelleira Martínez y J. Fonseca da Silva. 2012b. Aboveground biomass, wood volume, nutrient stocks and leaf litter in novel forests compared to native forests and tree plantations in Puerto Rico. Bois et Forêts Tropiques 314:7-16.
Lugo, A. E., E. H. Helmer y E. Santiago Valentín. 2012c. Caribbean landscapes and their biodiversity. Interciencia 37:705-710.
Lugo, A. E., N. L. Harris y W. L. Silver. 2012d. Nutrient cycling in the Luquillo Experimental Forest. Páginas 107-118 en N. L. Harris, A. E. Lugo, S. Brown y T. Heartsill Scalley, editores. Luquillo Experimental Forest: research history and opportunities. U.S. Department of Agriculture Forest Service, EFR-1, Washington, D.C.
Lugo, A. E., E. Medina y K. McGinley. 2014. Issues and challenges of mangrove conservation in the Anthropocene. Madera y Bosques 20:11-38.
Lugo, A. E., C. D. Rodríguez Pedraza, I. Fremont y I. Vicéns. 2017. Response to hurricanes of Pinus caribaea var hondurensis plantations in Puerto Rico. Caribbean Naturalist 43:1-16.
Lugo, A. E., K. M. Winchell y T. A. Carlo. 2018. Novelty in ecosystems. Páginas 259-271 en D. A. DellaSala y M. I. Goldstein, editores. The Encyclopedia of the Anthropocene. Elsevier, Oxford.
300 ariel e. lugo | ecología a m i m anera Literatura citada
Lugo, A. E., O. J. Abelleira Martínez, E. Medina, G. Aymard, and T. Heartsill-Scalley. 2020. Novelty in the tropical forests of the 21st century. Advances in Ecological Research 62:53-116.
Lugo, A. E., M. J. Sánchez, L. S. Betancourt, O. M. Ramos González, y I. Vicens. 2022. A stoichiometric comparison of primary and secondary forest stands on acid soils Caribbean Naturalist 89:1-32.
Macnae, W. 1968. A general account of the fauna and flora of mangrove swamps and forests in the Indo-West-Pacific region. Advances in Marine Biology 6:73-270.
Marcano Vega, H. 2019. Los bosques de Puerto Rico, 2014. Resource Bulletin SRS-224, USDA Forest Service, Southern Research Station, Asheville, NC.
Margalef, R. 1968. Perspectives in ecological theory. University of Chicago Press, Chicago, IL.
Marrero, J. 1950. La reforestación de tierras degradadas de Puerto Rico. Caribbean Forester 11:16-24.
Martínez, R. G., G. Cintrón y L. A. Encarnación. 1979. Mangroves in Puerto Rico: a structural inventory. Department of Natural Resources, Commonwealth of Puerto Rico, San Juan, PR.
Martinuzzi, S., W. A. Gould, A. E. Lugo y E. Medina. 2011. Conversion and recovery of Puerto Rican mangroves: 200 years of change. Forest Ecology and Management 257: 75-84.
Martinuzzi, S., A. E. Lugo, T. J. Brandeis y E. H. Helmer. 2013. Geographic distribution and level of novelty of Puerto Rican forests. Páginas 8187 en R. J. Hobbs, E. S. Higgs y C. Hall, editores. Novel ecosystems: Intervening in the new ecological world order? Wiley, Oxford.
Mascaro, J., F. R. Hughes y S. A. Schnitzer. 2012. Novel forests maintain ecosystem processes after the decline of native tree species. Ecological Monographs 82: 21-238.
Mayr, E. 1991. One long argument: Charles Darwin and the genesis of modern evolutionary thought. Harvard University Press, Cambridge, Massachusetts.
McKinney, M. y J. L. Lockwood. 1999. Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends in Ecology and Evolution 14:450-452.
Mcleod, E., G. L. Chmura, S. Bouillon, R. Salm, M. Björk, C. M. Duarte, C. E. Lovelock, W. H. Schlesinger y B. R. Silliman. 2011. A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2 . Frontiers in Ecology and the Environment 9:552-560.
301 ariel e. lugo | ecología a m i m anera Literatura citada
McPhearson, P. T., E. M. Cook, M. Berbés-Blázquez, C. Cheng, N. B. Grimm, E. Andersson, O. Barbosa, D. G. Chandler, H. Chang, M. V. Chester, D. L. Childers, S. R. Elser, N. Frantzeskaki, Z. J. Grabowski, P. M. Groffman, R. L. Hale, D. M. Iwaniec, N. Kabisch, C. Kennedy, S. A. Markolf, M. Matsler, L. McPhillips, T. R. Miller, T. A. Munoz-Erickson, E. J. Rosi y T. G. Troxler. 2022. A social-ecological-technological systems framework for urban ecosystem services. One Earth 5:505-518.
Medina, E. 1995. Diversity of life forms of higher plants in neotropical dry forests. Páginas 221-242 en S. H. Bullock, H. A. Mooney y E. Medina, editores. Seasonally dry tropical forests. Cambridge University Press, Cambridge, Great Britain.
Medina, E. 2015. Elemental composition of species from contrasting environments in Puerto Rico. Páginas 61-84 en M. J. Sánchez, E. Lopez y A. E. Lugo, editores. Chemical and physical analyses of selected plants and soils from Puerto Rico (1981-2000). USDA Forest Service International Institute of Tropical Forestry, General Technical Report IITF-GTR-45, Río Piedras, PR.
Medina, E., M. Francisco y A. Quilice. 2008. Isotopic signatures and nutrient relations of plants inhabiting brackish wetlands in the northeastern coastal plain of Venezuela. Wetlands Ecology and Management 16:51-64.
Medina, E., E. Cuevas y A. E. Lugo. 2010. Nutrient relations of dwarf Rhizophora mangle L. mangroves on peat in eastern Puerto Rico. Plant Ecology 207:13-24.
Medina, E., E. Cuevas, S. Molina, A. E. Lugo y O. Ramos. 2012. Structural variability and species diversity of a dwarf Caribbean dry forest. Caribbean Journal of Science 4:1-13.
Medina, E., G. González y M. M. Rivera. 2013. Spatial and temporal heterogeneity of rainfall inorganic ion composition in northeastern Puerto Rico. Ecological Bulletins 54:157-167.
Medina, E., E. Cuevas y A. E. Lugo. 2017. Substrate chemistry and rainfall regime regulate elemental composition of tree leaves. Forests 8, 339, doi:10.3390/f8060182.
Meine, C. 1988. Aldo Leopold. His life and work. The University of Wisconsin Press, Madison, Wisconsin.
Meine, C. y R. L. Knight, editores. 1999. The essential Aldo Leopold. Quotations and commentaries. The University of Wisconsin Press, Madison, WI.
Molina Colón, S. 1998. Long-term recovery of a Caribbean dry forest after abandonment of different land uses in Guánica, Puerto Rico. Disertación. University of Puerto Rico, Río Piedras, PR.
Molina Colón, S. y A. E. Lugo. 2006. Recovery of a subtropical dry forest after abandonment of different land uses. Biotropica 38(3):354-364.
302 ariel e. lugo | ecología a m i m anera Literatura citada
Molina Colón, S., A. E. Lugo y O. Ramos. 2011. Novel dry forests in southwestern Puerto Rico. Forest Ecology and Management 262:170-177.
Mueller-Dombois, D. y H. Ellenberg. 1974. Aims and methods of vegetation ecology. John Wiley and Sons, New York, NY.
Muñoz, M. A., W. I. Lugo, C. Santiago, M. Matos, S. Ríos y J. Lugo. 2018. Taxonomic classification of the soils of Puerto Rico, 2017. College of Agricultural Sciences and Agricultural Experiment Station Bulletin 313, Mayagüez, PR.
Murphy, D. A. J. 2007. The relation between land-cover and urban heat island in northeastern Puerto Rico. Tésis. State University of New York, Syracuse, NY.
Murphy, L. S. 1916. Forests of Puerto Rico: past, present, and future and their physical and economic environment. US Department of Agriculture Bulletin 354, Washington, DC.
Murphy, P. G. y A. E. Lugo. 1986a. Ecology of tropical dry forest. Annual Review of Ecology and Systematics 17:67-88.
Murphy P. G. y A. E. Lugo. 1986b. Structure and biomass of a subtropical dry forest in Puerto Rico. Biotropica 18(2):89-96.
Murphy, P. G. y A. E. Lugo. 1995. Dry forests of Central America and the Caribbean. Páginas 9-34 en S. H. Bullock, H. A. Mooney, y E. Medina, editores. Seasonally dry tropical forests. Cambridge University Press, Cambridge, Great Britain.
Murphy, S. F. y R. F. Stallard. 2012. Hydrology and climate of four watersheds in eastern Puerto Rico. Páginas 43-83 en S. F. Murphy y R. F. Stallard, editores. Water quality and landscape processes of four watersheds in eastern Puerto Rico. Professional Paper 1789 US Department of the Interior, Geological Survey. Reston, VA.
National Research Council. 1980. Research priorities in tropical biology. National Academy of Sciences, Washington, D.C. 116 p.
Norton, B.J., editor. 1986. The preservation of species. Princeton University Press, Princeton, N.J. 305 p.
Myers, N. 1979. The sinking ark. A new look at the problem of disappearing species. Pergamon Press, New York, N.Y.
Myers, N. 1982. Forest refuges and conservation in Africa with some appraisal of survival prospects for tropical moist forests throughout the biome. Pages 658-672 in G. T. Prance, editor. Biological diversification in the tropics. Columbia University Press, New York, N.Y.
Myers, N. 1983. Conservation of rain forests for scientific research, for wildlife conservation, and for recreation tourism. Pages 325-334 in F. B. Golley, editor. Tropical rain forest ecosystems. Structure and function. Elsevier, Amsterdam.
303 ariel e. lugo | ecología a m i m anera Literatura citada
Odum, E. P. 1953. Fundamentals of ecology, first edition. W.B. Saunders Co., Philadelphia, PA.
Odum, E. P. 1969. The strategy of ecosystem development. Science 164:262-270.
Odum, H. T. 1962. Man and the ecosystem. Páginas 57-75 en P. E. Waggoner y J. D. Ovington, editores. Lockwood conference on the suburban forest and ecology. The Connecticut Agricultural Experiment Station, New Haven, CN.
Odum, H. T. 1970. Summary: an emerging view of the ecological systems at El Verde. Páginas I191-I289 en H. T. Odum y R. F. Pigeon, editores. A tropical rain forest. National Technical Information Service, Springfield, VA.
Odum, H. T. 1971. Environment power and society. Wiley Interscience, New York, NY.
Odum, H. T. 1973. Energy, ecology, and economics. Ambio. 2(6):220-227.
Odum, H. T. 1983. Systems ecology. Wiley, New York, NY.
Odum, H. T. 1995. Self-organization and maximum empower. Páginas 311-330 en C. A. S. Hall, editor. Maximum Power: The ideas and applications of H.T. Odum. University Press of Colorado, Niwot, CO.
Odum, H. T. 2007. Environment, power, and society for the twenty-first century. The hierarchy of energy. Columbia University Press, New York, NY, USA.
Odum, H. T. y R. C. Pinkerton. 1955. Time’s speed regulator: the optimum efficiency for maximum power output in physical and biological systems. American Scientist 43:321-343.
Odum H. T y R. F. Pigeon, editores. 1970. A tropical rain forest. National Technical Information Service, Springfield, VA.
Odum H. T. y C. F. Jordan. 1970. Metabolism and evapotranspiration of the lower forest in a giant plastic cylinder. Páginas I-165 - I-189 en H. T. Odum y R. F. Pigeon, editores. A tropical rain forest: a study of irradiation and ecology at El Verde, Puerto Rico. USAEC Division of Technical Information Extension, Oak Ridge, TN.
Odum, H. T., W. Abbott, R. K. Selander, F. B. Golley y R. F. Wilson. 1970. Estimates of chlorophyll and biomass of the tabonuco forest of Puerto Rico. Páginas I3-I19 en H. T. Odum y R. F. Pigeon, editores. A tropical rain forest. National Technical Information Service, Springfield, VA.
Odum, H. T. y E. C. Odum. 2000. Modelling for all scales: an introduction for system simulation. Academic Press, San Diego, CA.
Odum H. T. y E. C. Odum. 2001. A prosperous way down: principles and policies. University Press of Colorado, Boulder, CO.
Odum, W. E. 1971. Pathways of energy flow in a south Florida estuary. Sea Grant Technical Bulletin 7, University of Miami, Miami, FL.
304 ariel e. lugo | ecología a m i m anera Literatura citada
Odum, W. E., C. C. McIvor, y T. J. Smith III. 1982. The ecology of the mangroves of South Florida: a community profile. U.S. Fish and Wildlife Service, Office of Biological Services FWS/OBS-81/24, Washington DC.
Oldfield, M.I. 1984. The value of conserving genetic resources. US Department of Interior National Park Service, Washington D.C. 360 p.
Ovington, J. D. y J. S. Olson. 1970. Biomass and chemical content of El Verde lower montane rain forest plants. Páginas H53-H77 en H. T. Odum y R. F. Pigeon, editores. A tropical rain forest. A study of irradiation and ecology at El Verde, Puerto Rico. National Technical Information Service, Springfield, Virginia.
Pan Y, R. A. Birdsey, J. Fang, R. Houghton, P. E. Kauppi, W.A. Kurz, O. L. Phillips, A. Shvidenko, S. L. Lewis, J. G. Canadell, P. Ciais, R. B. Jackson, S. W. Pacala, A. D. McGuire, S. Piao, A. Rautiainen, S. Sitch y D. Hayes. 2011. A large and persistent carbon sink in the world’s forests. Science 333:988-993.
Parrotta, J. A. y J. W. Turnbull. 1997. Catalyzing native forest regeneration on degraded tropical lands. Forest Ecology and Management 99:1-290.
Ping, C. L., G. J. Michaelson, C. A. Stiles y G. González. 2013. Soil characteristics, carbon stores, and nutrient distribution in eight forest types along an elevation gradient, eastern Puerto Rico. Ecological Bulletins 34:67-86.
Porder, S., A. H. Johnson, H. X. Xing, G. Brocard, S. Goldsmith y J. Pett Ridge. 2015. Linking geomorphology, weathering and cation availability in the Luquillo Mountains of Puerto Rico. Geoderma 249-250:100-110.
Puerto Rico Climate Change Council (PRCCC). 2013. Puerto Rico’s state of the climate 2010-2013: assessing Puerto Rico’s social-ecological vulnerabilities in a changing climate. Puerto Rico Coastal Zone Management Program. Department of Natural and Environmental Resources. NOAA Office of Ocean and Coastal Resource Management. San Juan, PR.
Quiñones Márquez, F. 2021. Análisis estadísticos de la lluvia promedio diaria en Puerto Rico entre 1920-2020 no reflejan disminuciones en la precipitación anual en la mayor parte de la isla. Acta Científica 32:5-25.
Quiñones, M., I. K. Parés, W. A. Gould, G. González, K. McGinley y P. Ríos. 2018. Atlas del Bosque Nacional El Yunque y el Bosque Experimental de Luquillo. General Technical Report (IITF-GTR-47), USDA Forest Service, Río Piedras, PR.
Ramjohn, I. A., P. G. Murphy, T. M. Burton y A. E. Lugo. 2012. Survival and rebound of Antillean dry forests: role of forest fragments. Forest Ecology and Management 284:124-132.
Redfield, A. C. 1934. On the proportions of organic derivatives in sea water and their relation to the composition of plankton. Páginas 176-192 en R. J. Daniel, editor. James Johnstone memorial volume. Liverpool University Press, Liverpool, U.K.
305 ariel e. lugo | ecología a m i m anera Literatura citada
Redfield, A. C. 1958. The biological control of chemical factors in the environment. American Scientist 46:205-221.
Richardson, B. A. y M. J. Richardson. 2013. Litter-based invertebrate communities in forest floor and bromeliad microcosms along an elevational gradient in Puerto Rico. Ecological Bulletins 54:101-115.
Rivera Collazo, I. C. 2015. Por el camino verde: Long-term tropical socioecosystem dynamics and the Anthropocene as seen from Puerto Rico. The Holocene 25:1604-1611.
Rivera Monroy, V. H., M. J. Osland, J. W. Day, S. Ray, A. Rovai, R. H. Day y J. Mukherjee. 2017. Advancing mangrove macroecology. Páginas 347381 en V. H. Rivera Monroy, S. Y. Lee, E. Kristensen y R. R. Twilley, editores. Mangrove ecosystems: a global biogeographic perspective. Structure function, and services. Springer, Cham, Switzerland.
Rivero, J. A. 2008. Agustín Stahl: el precursor. Páginas 27-75 en Joglar R. L., editor. Biodiversidad de Puerto Rico. Agustín Stahl, flora, hongos. La Editorial Universidad de Puerto Rico, Río Piedras, PR.
Robinson, K. 1997. Where dwarfs reign: a tropical rain forest in Puerto Rico. University of Puerto Rico Press, Río Piedras, PR.
Robinson, K., A. E. Lugo y J. Bauer, editores. 2014. Passing the baton from the Taínos to tomorrow: forest conservation in Puerto Rico. USDA Forest Service International Institute of Tropical Forestry, San Juan, PR FS-862, 196 p.
Ruiz, B. I., editor. 2002. Manual de reforestación para América tropical. General Technical Report IITF-GTR-18, International Institute of Tropical Forestry, Río Piedras, PR.
Ryan, M.G., M. E. Harmon, R. A. Birdsey, C. P. Giardina, L. S. Heath, R. A. Houghton, R. B. Jackson, D. C. McKinley, J. F. Morrison, B. C. Murray, D. E. Pataki y E. Skog. 2010. A synthesis of the science on forests and carbon for U.S. forests. Issues in Ecology 13:1-16.
Saatchi, SS, N. L. Harris, S. Brown, M. Lefsky, E.T.A. Matched, W. Salas, B. R. Zeta, W. Buhrmann, S. L. Lewis, S. Hagen, S. Petrova, L. White, M. Silman y A. Morel. 2011. Benchmark map of forest carbon stocks in tropical regions across three continents. PNAS 108:9899-9904.
Sánchez, M. J., E. Lopez y A. E. Lugo. 2015. Chemical and physical analyses of selected plants and soils from Puerto Rico (1981-2000). USDA Forest Service, General Technical Report IITF-GTR-45, Río Piedras, PR.
Scatena, F. N. 1989. An introduction to the physiography and history of the Bisley Experimental Watersheds in the Luquillo Mountains of Puerto Rico. General Technical Report SO-72, USDA Forest Service, Southern Forest Experiment Station, New Orleans, LA.
306 ariel e. lugo | ecología a m i m anera Literatura citada
Scatena, F. N. y A. E. Lugo. 1995. Geomorphology, disturbance, and the soil and vegetation of two subtropical wet steepland watersheds of Puerto Rico. Geomorphology 13:199-213.
Scatena, F. N. y M. C. Larsen. 1991. Physical aspects of hurricane Hugo in Puerto Rico. Biotropica 23:317-323.
Scatena, F. N., S. Moya, C. Estrada y J. D. Chinea. 1996. The first five years in the reorganization of aboveground biomass and nutrient use following hurricane Hugo in the Bisley Experimental Watersheds, Luquillo Experimental Forest, Puerto Rico. Biotropica 28:424-440.
Scheffers, B. R., L. N. Joppa, S. L. Pimm y W.F. Laurance. 2012. What we know and don’t know about Earth’s missing biodiversity. Trends in Ecology & Evolution 1553:1-10.
Schimper, A. F. W. 1903. Plant-geography upon a physiological basis. Clarendon Press, Oxford, England.
Shelford, V. E. 1932. Life zones, modern ecology, and the failure of temperature summing. The Wilson Bulletin September: 144-157.
Shelford, V. E. 1963. The ecology of North America. University of Illinois Press, Urbana, IL.
Shiels, A. B. y G. G. González. 2014. Tropical forest responses to largescale experimental hurricane effects. Forest Ecology and Management 332:1-135.
Shiels, A. B. y L. R. Walker. 2013. Landslides cause spatial and temporal gradients at multiple scales in the Luquillo Mountains of Puerto Rico. Ecological Bulletins 54:211-221.
Simberloff, D. 1983. Are on the verge of mass extinction in tropical rain forest? Unpublished monograph, July.
Simberloff, D. 1986. Are we on the verge of a mass extinction in tropical rain forest? Pages 165-180 in D. K. Elliott, editor. Dynamics of extinctions. John Willey and Sons, New York, NY.
Silver, W. L. 1992. Effects of small-scale and catastrophic disturbance on carbon and nutrient cycling in a lower montane subtropical wet forest in Puerto Rico. Disertación. Yale, New Haven, CT, USA.
Silver, W. L, A. E. Lugo y M. Keller. 1999. Soil oxygen availability and biogeochemistry along rainfall and topographic gradients in upland wet tropical forest soils. Biogeochemistry 44:301-328.
Silver, W. L., R. Ostertag y A. E. Lugo. 2000. The potential for carbon sequestration through reforestation of abandoned tropical agricultural and pasture lands. Restoration Ecology 8:394-407.
Silver, W. L., L. M. Kueppers, A. E. Lugo, R. Ostertag y V. Matzek. 2004. Carbon sequestration and plant community dynamics following reforestation of tropical pasture. Ecological Applications 14:1115-1127.
307 ariel e. lugo | ecología a m i m anera Literatura citada
Smith, B. D. y M. A. Zeder. 2013. The onset of the Anthropocene. Anthropocene 4:8-13.
Sterner, R. W. y J. J. Elser. 2002. Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton University Press, Princeton, NJ.
Tansley, A. G. 1935. The use and abuse of vegetational concepts and terms. Ecology 16:284-307.
Tansley, A. G. 1947. The early history of modern plant ecology in Britain. Journal of Ecology, 35:130-137.
Thom, B. G. 1975. Mangrove ecology from a geomorphic viewpoint. Páginas 469-481 en G. Walsh, S. Snedaker y H. Teas, editores. Proceedings of the international symposium on biology and management of mangroves. Institute of Food and Agricultural Sciences, University of Florida, Gainesville, FL.
Thomas, C. D. 2013. The Anthropocene could raise biological diversity. Nature 502:7.
Thomas, C. D. 2015. Rapid acceleration of plant speciation during the Anthropocene. Trends in Ecology & Evolution 30:448-455.
Thompson, D. W. 1942. On growth and form. The complete revised edition. Dover Publications, Inc., New York, NY, USA.
Thompson, J., N. Brokaw, J. K. Zimmerman, R. B. Waide, E. M. Everham III, D. J. Lodge, C. M. Taylor, D. García Montiel y M. Fluet. 2002. Land use history, environment, and tree composition in a tropical forest. Ecological Applications 12:1344-1363.
Twilley, R. R., V. H. Rivera Monroy, R. Chen y L. Botero. 1988. Adapting an ecological mangrove model to simulate trajectories in restoration ecology. Marine Pollution Bulletin 37:404-419.
Twilley, R. R., S. C. Snedaker, A. Yáñez-Arancibia, y E. Medina. 1996. Biodiversity and ecosystem processes in tropical estuaries: perspectives of mangrove ecosystems. Páginas 327-370 en H. A. Mooney, J. H. Cushman, E. Medina, O. E. Sala, y E. D. Schulze, editores. Functional role of biodiversity: a global perspective. John Wiley & Sons, New York, NY.
Van Bloem, S. J., P. G. Murphy y A. E. Lugo. 2003. Subtropical dry forest trees with no apparent damage sprout following a hurricane. Tropical Ecology 44:1-9.
Van Bloem, S. J., P. G. Murphy, A. E. Lugo, R. Ostertag, M. Rivera Costa, I. Ruiz Bernard, S. Molina Colón y M. Canals Mora. 2005. The influence of hurricane winds on Caribbean dry forest structure and nutrient pools. Biotropica 37:571-583.
Vellend, M., L. J. Harmon, J. L. Lockwood, M. M. Mayfield, A. R. Hughes, J. P. Wares y D. F. Sax. 2007. Effects of exotic species on evolutionary diversification. TRENDS in Ecology and Evolution 22:481-488.
308 ariel e. lugo | ecología a m i m anera Literatura citada
Von Bertalanffy, L. 1968. General system theory. Foundations, development applications. George Braziller, New York, N.Y.
Wadsworth, F.H. 1949. The development of the forest land resources of the Luquillo Mountains of Puerto Rico. Disertación. University of Michigan, Ann Arbor, MI.
Wadsworth, F. H. 1995. A forest research institution in the West Indies: the first 50 years. Páginas 33-56 en A. E. Lugo and C. Lowe, editores. Tropical forests: management and ecology. Springer Verlag, New York, NY, USA.
Wadsworth, F. H. 2007. José Marrero Torrado, regalo puertorriqueño a los árboles. Acta Científica 21:29-35.
Waide, R. B., D.E. Comarazamy, J.R. González, C.A.S. Hall, A.E. Lugo, J.C. Luvall, D.J. Murphy, J.R. Ortiz Zayas, N.D. Ramírez Beltran, F.N. Scatena y W.L. Silver. 2013. Climate variability at multiple spatial and temporal scales in the Luquillo Mountains, Puerto Rico. Ecological Bulletins 54:21-41.
Wake, D. B. 2012. Facing extinction in real time. Science 335:1052-1053.
Walsh, G., S. Snedaker y H. Teas, editores. 1975. Proceedings of international symposium on biology and management of mangroves. Institute of Food and Agricultural Sciences, University of Florida, Gainesville, FL., Honolulu, HI. 846 p.
Walter, H. 1971. Ecology of Tropical and Subtropical Vegetation. Van Nostrand Reinhold Company, New York, NY.
Watson, J. G. 1928. Mangrove forests of the Malay Peninsula. Malayan Forest Records 6:1-274.
Weaver, P. L. 1994. Baño de Oro natural area Luquillo Mountains, Puerto Rico. USDA Forest Service, Southern Forest Experiment Station General Technical Report SO-111, New Orleans, LA.
Weaver, P. L. 1995. The colorado and dwarf forests of Puerto Rico’s Luquillo Mountains. Páginas 109-141 en A. E. Lugo y C. Lowe, editores. Tropical forests: management and ecology. Springer Verlag, New York, NY.
Weaver, P. L. 2006a. Estate Thomas Experimental Forest, St. Croix, U.S. Virgin Islands: research history and potential. International Institute of Tropical Forestry, Río Piedras, General Technical Report IITF-30.
Weaver, P. L. 2006b. A summary of 20 years of forest monitoring in Cinnamon Bay Watershed, St. John, U.S. Virgin Islands. International Institute of Tropical Forestry, General Technical Report IITF-34, Río Piedras, PR.
Weaver, P. L. 2012. The Luquillo Mountains: forest resources and their history. International Institute of Tropical Forestry General Technical Report IITF-44, Río Piedras, PR.
Weaver, P. L. y W. A. Gould. 2013. Forest vegetation along environmental gradients in northeastern. Ecological Bulletins 54:43-65.
309 ariel e. lugo | ecología a m i m anera Literatura citada
Whittaker, R. H. 1970. Communities and ecosystems. The Macmillan Company, Toronto, Ontario.
Willig, M. R., S. J. Presley, C. P. Bloch y J. Alvarez. 2013. Population, community, and metacommunity dynamics of terrestrial gastropods in the Luquillo Mountains: a gradient perspective. Ecological Bulletins 54:117-140.
Winchell, K. M., R. G. Reynolds, S. R. Prado-Irwin, A. R. Puente-Rolón y L. J. Revell. 2016. Phenotypic shifts in urban areas in the tropical lizard Anolis cristatellus. Evolution 70:1009-1022.
Wood, T. E., G. González, W. L. Silver, S. C. Reed y M. A. Cavaleri. 2019. On the shoulders of giants: continuing the legacy of large-scale ecosystem manipulation experiments in Puerto Rico. Forests 10, 210: doi:10.3390/f10030210.
Wood, T. E. y W. L. Silver. 2012. Strong spatial variability in trace gas dynamics following experimental drought in a humid tropical forest. Global Biogeochemical Cycles 26:GB3005.
Wright, S. J. y H. C. Muller Landau. 2006. The future of tropical forest species. Biotropica 38:287-301.
Yi, S., W. Sun, K. Heki y A. Quian. 2015. An increase in the rate of global mean sea level rise since 2010. Geophysical Research Letters 42:3998-4006.
Zimmerman, J. K., W. M. Pulliam, D. J. Lodge, V. Quiñones Orfila, N. Fetcher, S. Guzmán Grajales, J. A. Parrotta, C. E. Asbury, L. Walker y R. B. Waide. 1995. Nitrogen immobilization by decomposing woody debris and the recovery of tropical wet forest from hurricane damage. Oikos 72:314-322.
310 ariel e. lugo | ecología a m i m anera Literatura citada
A
Acidificación del agua 268
Ácido ribonucleico (RNA) 49
Ácidos nucleicos 48
Actividad agrícola 75, 133, 141, 159
Aforestación 221, 258
Agentes abióticos 270
Agentes bióticos 270
Alfonso XII 104
Almacenaje aéreo de elementos 60, 61
Almacenaje de carbono 207, 209, 246, 253 -256
Almacenaje de cinco macronutrientes en la hojarasca 231
Almacenaje promedio de biomasa en zonas de vida tropicales y subtropicales 253
Almacenaje sobre el terreno 247
Almacenajes subterráneos 130, 247
Alometría 45, 46, 56
Alometría, concepto 57
Altas montañas tropicales 83
Aluviales 60
Álvarez Ruiz, Migdalia 157
Anaeróbico 125
Anfibios 113, 156, 217, 268
Animales
, 264, 268, 275, 278
Antillas Mayores 218, 219
Índice de materias
Antropoceno 12, 63, 66, 76, 77, 113, 191, 271, 272, 283
Antropoceno, época del 283
Antropogénica 183, 278, 283
No-antropogénica 278
Antropogénica, actividad 215
Antropogénicos 20, 33, 38, 65, 117, 132, 134, 135, 137, 138, 142, 177, 213, 257
No-antropogénicos 38, 120, 134, 269, 283
Árboles centenarios 146
Árboles fuera del bosque 222
Arbóreos 59, 119, 149, 155, 161, 165, 169, 248
Arboretum de Ciénaga Alta 228
Área basal de árboles 59
Área de plantaciones 224
Área específica 129
Área específica foliar 59
Área foliar 42, 59, 129, 130, 131, 135
Arrecifes de coral 16, 22, 70, 76, 193
Artrópodos 49
Asociación Médica de Puerto Rico 227
Aspecto o exposición al viento y el sol 123
Atlántico 125, 173, 262
ATP. Véase Moléculas de trifosfato de adenosina (ATP)
Atributos funcionales 138, 161
Autosucesión 149, 178
Ayala, Sandra 22, 158
Azufre 47, 55, 63, 64, 171
311 ariel e. lugo | ecología a m i m anera Índice de materias
15
16, 19
36
41, 48, 49, 61, 64, 71, 74 -78, 82, 84, 94, 105, 106, 155, 156, 161, 183, 202, 203, 227, 246, 247, 262
,
,
,
B
Bacterias 48, 49, 52, 154, 203
Bahía de San Juan 73, 103
Baird Callicott, J. 278
Baldorioty de Castro, Román 105
312 ariel e. lugo | ecología a m i m anera Índice de materias
12, 75, 87, 202, 219, 222, 226, 227, 242, 261, 269-271, 273
restauración de 242 Biomasa 19, 35, 37, 38, 40, 52-54, 57- 61, 114, 127-130, 135, 136, 142, 143, 148, 153 -155, 201-203, 207-209, 237, 238, 240, 246 -249, 253, 254, 258, 259
acumulación de 114, 228 Biomasa aérea 142, 249 Biomasa de raíces 130, 136, 237, 238, 247 Biomasa subterránea 127, 130, 142 Biomasa viva 247 Biota 16, 77, 86, 105, 217, 262, 268-270, 279, 282 Biota, adaptación de la 269 Biota, homogenización de la 269 Bióxido de carbono 36, 52, 63, 67-71, 130, 162, 168, 226, 245 -258
de carbono, concentración en la atmósfera de 70, 245, 258 Bisley 61, 112, 124, 143 -147, 151, 177, 192, 198, 245 Boro 48 Bosque bajo manejo intensivo 221 Bosque de palo colorado 168, 197, 202, 203, 209, 211 Bosque de tabonuco 112, 124, 135, 141, 143 -145, 148, 149, 151-159, 165, 170, 177, 202, 203, 207, 209, 211, 215, 245, 247 Bosque elfino 106, 112, 151, 168, 169, 202, 203, 209, 211 Bosque Estatal de Toro Negro 277 Bosque Experimental de Luquillo 56, 61, 113, 123, 124, 135, 143 -147, 149-156, 169, 201-205, 207, 208, 211, 212, 215, 218, 228 -235, 237, 238, 241, 245, 246, 248
Experimental
113 Bosque húmedo 103, 107, 116, 129, 130, 131, 132, 141, 142, 209, 249, 253 Bosque maduro 97, 98, 134, 137, 149, 153, 242, 247, 248 -250 Bosque montano bajo muy húmedos y pluviales 141, 142 Bosque Nacional 187 Bosque nativo 60, 75, 78, 110, 115, 137, 141, 142, 157, 158, 235, 237, 239, 242 Bosque novel 75, 77, 113, 115, 117, 138, 139, 141, 158, 159, 246, 248, 249 Bosque primario 56, 129, 222, 228, 253 Bosques cerrados 216 Bosques de tierras bajas 209 Bosque secundario 56, 222, 227, 235, 237, 238 -241, 247, 248, 258 Bosque secundario nativo 240 Bosques muy húmedos 55, 135, 142, 156, 249 Bosques secos 61, 105 -107, 111-114, 119, 127-132, 134 -139, 159, 209, 245, 247-249 Bosques secos antillanos 127, 130, 136 Bosques secos continentales 130 Bosques secos costeros 129 Bosques secundarios 153, 246, 253, 265, 270 Bosques subtropicales húmedos 141, 142 Bosques terrestres 254 Bosques tropicales 12, 81, 100, 107, 112-114, 120, 177, 228, 245, 246, 249, 252-255, 257, 258, 265, 266, 277, 282 Britton, Elizabeth 26 Bromelias 201-203, 209, 211 C Cadena alimenticia 57, 58, 185, 229, 246 Calcio 47, 55, 70, 76, 88, 89, 129, 138, 139, 143, 144, 193, 197, 233 Calcio, concentración de 139, 144 Calculadora Monroe 282
Basalto 189 Biodiversidad
Biodiversidad,
Biomasa,
Bióxido
Bosque
Estate Thomas
Cationes minerales 196
Cemí 187
Centroamérica 219, 226
Cerro la Tiza de Comerío 158
Cerro Puntas 277
Chapman, Valentine Jackson 175
Ciclo de nutrientes abierto 40
Ciclo de nutrientes cerrado 40
Ciclos de carbono, hidrógeno y oxígeno 47
Ciénaga Alta 228 -235, 237
Cinnamon Bay 113
Cintrón Molero, Gilberto 59
Circular 493 de 1824 104
CITES 226, 227
Ciudades europeas 269
Ciudades tropicales 269
Clasificación geoclimática 114
Clements, Frederic Edward 16
Clímax 39, 81, 108, 111
Cloro 48
Clorofila 48
C:N Detrito 51
C:N Planta 53
Cobertura
Cobertura de los bosques en Puerto Rico 117
Cobertura, cambio en 215
Cobertura del dosel 59
Cobertura, porcentaje de 135, 216
Cobre 48
Cociente área del dosel:biomasa 59
Cociente área del dosel:diámetro del tallo 59
Cociente C13:C14 138
Cociente C:N 207
Cociente hongo:bacteria 204
Cociente largo:ancho foliar 59
Cociente PET/P 127
Cocientes N:P 55
Cociente temperatura promedio anual:precipitación promedio anual 252
Cociente T:P 249
Competencia 35
313 ariel e. lugo | ecología a m i m anera Índice de materias Calentamiento global 67, 70, 75, 114, 174, 178, 245, 262 Caliza
Calizos 116
127, 129, 138 Cambio climático 64, 67, 70, 72-77, 79, 111
213
268
277
263 Canalizaciones 64, 171 Caoba 60, 61, 113, 132, 221, 226, 227, 238, 247 Carbonífero 263 Carbono 26, 30, 31, 36, 45 - 49, 51-53, 56, 58, 59, 63, 64, 67-71, 75, 128, 131, 139, 162, 168, 171, 174, 178, 207, 209, 211, 226 -228, 230, 235, 242, 245, 246 -251, 253 -259 Carbono atmosférico 45, 257, 258 Carbono azul (blue carbon) 254 Carbono, ciclo del 64, 245, 246, 254, 256, 257 Carbono:fósforo 51, 139 Carbono:nitrógeno 51-53, 56, 139, 211 Carbono, reciclaje o movimiento del 246 Carbono, secuestro de 58, 59, 242, 253, 254 Carbono total 128, 142 Cardenal, Lope Marinero 105 Caribe 25, 72, 78, 83, 94, 104, 114, 118 -120, 125, 130, 146, 150, 165, 173, 175, 187, 213, 219 Catena 122, 123, 146, 147, 156, 207, 208, 214, 215
123, 124, 129, 196
115, 194
,
,
,
,
Cambriano
Catenas
65, 66, 81, 85, 100, 101, 104, 107, 115, 117, 121, 122, 132, 134, 135, 185, 195, 196, 199, 200, 203, 204, 215, 216, 218, 222, 225, 226, 253-255, 257, 263, 268, 269
Composición química de la corteza de la tierra 47
Composición química del ser humano 47
Comunidades microbianas 125
Condado 183
Condiciones anaeróbicas 125, 168, 203
Conductividad de las lluvias 199
Conservación 15, 19, 20, 23, 26, 29, 30, 41, 45, 57, 63, 67, 83, 103 -105, 127, 129, 154, 174, 175, 227, 245, 256, 273, 275 -279
Conservador de Bosques de los Estados Federados de Malasia 175
Constante de descomposición 51
Constante solar 31, 37, 42
Consumidores 36, 41, 185
Consumo 30, 36, 38, 65, 107, 226
Contribución de mujeres 26
Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres (CITES) 226
Coquíes 154
Corteza de la tierra 47, 189
Cosecha 36, 223, 224
C:P Detrito 51
Crecimiento arbóreo 180, 241
Crecimiento de nuevas hojas 35
Crecimiento específico 49, 50
Cretáceo 263
Critical Zone Observatory 193
Cuaternario 63, 263
Cubuy 61, 194 -196, 235, 237-242
Cultivo 65, 221, 223, 224
Cultivo forestal 221
Curvas de Importancia Relativa 131 D
Darwin, Charles Robert 16 Deforestación
Deforestación, tasa de 263, 277
Deforestación y urbanización 258
Degradación 127, 226, 263
del Llano, Manuel 165, 166
Densidad aparente de los suelos 145
Densidad aparente del suelo 59
Densidad de árboles 59, 60, 98, 123, 127, 148, 206, 208
Densidad de especies 119, 132, 134, 207, 262, 269
Densidad de tallos 130, 135, 207, 208
Densidad específica de la madera 59
Depredación 35, 268
Deriva genética (genetic drift) 271
Derrumbes 118, 145, 192, 213, 214
Descomposición 51, 52, 53, 90, 91,
Descomposición, proceso de 228
Desiertos tropicales 83
Deslizamientos de terreno 216
38, 51, 183, 230
Disturbios agudos 213
Disturbios crónicos 213
Disturbios meteorológicos 213
Disturbios tectónicos 213
de hábitats 270
de hongos 203
Dominancia de especies 137, 150, 151, 162
Domínguez Cristóbal, Carlos Manuel 103
Dunas 79, 108, 173, 174
314 ariel e. lugo | ecología a m i m anera Índice de materias
38, 103, 104, 114, 127, 137, 158, 159, 215, 258, 263, 265, 266, 268 -270, 277
114
125
192
207
209
228
230
,
,
,
,
,
,
, 234
Devoniano
Dioritas 189, 192, 196, 197, 214 Disturbio 20, 39, 40, 76, 118, 119, 132, 135, 145, 146, 149, 171, 172, 213, 269 Disturbios 38 - 40, 74, 90, 94, 107, 118, 119, 132, 134, 135, 137, 138, 144 -146, 150, 151, 159, 171, 183, 213, 219, 269
Detrito
263
Diversidad
Dominancia
78
120
131
147
151
Diversidad
65,
,
,
,
,
, 175, 201, 202, 240
E Earhart, Amelia 27
Ecología cuantitativa de sistemas 58 Ecología estequiométrica 45
Epifitas 112, 117, 129, 154
Equilibrio 39
Equilibrio dinámico 37
Equivalencias solares 31
Erosión 106, 138, 153, 188, 192, 193, 195 -197, 227, 262
Erosión de las rocas volcanoclásticas 153
Ecosistema, definición de Evans 32
Ecosistema, definición de Tansley 33
Ecosistemas tropicales 112
ESA. Véase Sociedad Ecológica de Norte América (ESA)
Esau, Katherine 281
Escala de tiempo geológica 191
Especiación por evolución 261
Especie en peligro de extinción 226
Especies, abundancia de 104, 150, 151, 158, 201, 270
Especies del karso 55
Especies de mangle 55, 173, 175 -179
Frank Edwin 17
Especies, dispersión de 269, 271
Especies
Especies, persistencia de las 268, 269
Especies pioneras 55, 149
Especies, refugios para las 269
Especies, supervivencia de 269
Especie urbanófila 271
Estado
Estequiometría 45, 46
Estrés 36, 56, 59, 172, 183
Estrés, fuerzas de 183
Estresor 172
Estresores 171, 182, 213
Estroncio 193
, 178, 257
Estuarinos 161, 163, 165, 173, 182
Ética 19, 275, 277-279
Europa 22, 224, 269, 271, 272
315 ariel e. lugo | ecología a m i m anera Índice de materias
15, 17, 20, 29, 31- 42, 46, 54, 58, 59, 112, 165, 171, 173, 177, 182, 213, 254
Ecosistema
, 276
Ecotono
Eficiencia 30, 51, 53, 54, 114, 131, 137, 138, 227
Eficiencia
Eficiencia
Eficiencia
Egler,
Elevación 71, 82, 92, 95, 98, 108, 116, 122, 123, 129, 149-151, 157, 158, 168, 169, 192-195, 197, 198 -209, 211, 212, 215, 216 Elton,
Sutherland 17 El Toro 194 El Verde 61, 112, 113, 135, 143, 149, 152, 177, 198, 214, 215, 218, 235, 238, 239, 245, 248 El Yunque 26, 75, 104, 106, 112-114, 117, 122, 151, 165, 168, 177, 187, 189, 193, 194, 196, 198, 199, 201, 213, 214, 219 Embalses 171 Energía 30, 46 Energía, flujos de 22, 30, 36, 38, 42, 46, 57, 58, 66, 122, 166, 185, 213 Energía hidráulica 23 Energía potencial 29, 30, 31, 41, 46, 52, 53, 163 Energía química 35, 36, 48 Energía radiante 35, 36
46
solar 23, 31, 36, 41, 66, 85, 86, 107, 119, 204, 250, 283
30, 31, 37, 46, 57
198
, 228, 235
en el retorno de nitrógeno 53
en el uso de nitrógeno y fósforo 237
en el uso de nutrientes 53
Charles
,
Energía
Entropía
Especies endémicas 134, 269
56
78
218
222
226
235
75
132
138
159
222
224
226 -228, 242, 243, 251, 270
271
Especies introducidas 17, 19,
, 75, 77,
, 115, 132,
,
,
,
, 270, 271 Especies nativas
,
,
,
,
,
,
,
no nativas 158, 279
40
41
57
58
estable 37,
,
,
,
Evapotranspiración 26, 73, 74, 96, 127, 168, 204, 249
Everglades de la Florida 169, 177
Evolución, aceleración de la 77, 270, 271
Evolución de las especies 22, 261
Evolución, trama de la 33, 76
Ewel, John Jeffrey 130
Explotación 38, 104, 105
Extinción 217, 226, 227, 261-263, 265 -273, 277
Extinciones masivas del pasado 262
Extinción masiva, evento de 261
Extinción, tasa de 262, 263, 268
F
Falla geológica del Cerro Mula 189
FAO. Véase Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO)
Fenología 34, 35, 234
Fenología química 235
Figueroa Colón, Julio 114, 115
Fijación de carbono 227, 253
Filosofía ambiental 278
Filósofos griegos 15
Floración 35, 269
Flujos de nitrógeno y fósforo 154
Flujo y almacenaje de masa y nutrientes 151
Formación Oligoceno-Mioceno 188
Fosfoalginina 48
Fosfocreatina 48
Fosfolípidos 48
Fósforo
Frangi, Jorge Luis 165
Fuerzas tectónicas 165, 192
Fundación Nacional de Ciencias de los EE. UU. 193
Fundación Nacional para las Ciencias (NSF) 113
G
García, José Mariano 27
Gases de invernadero 67, 68, 74, 75, 168
Gasterópodos 202
Gastrópodos 201
Geológicos 7, 94, 192, 213, 263
Geomorfología 120, 145, 163, 180, 181
Gifford, John Clayton 106
Gleason, Henry Allan 17
Glucosa 48
Golley, Frank Benjamín 177
Gradientes ecológicos 197
Gradientes topográficos y edáficos 173
Granito 189
Granodioritas 189
Guánica
,
Guánica, Bosque Estatal de 133, 250
Gutiérrez del Arroyo Santiago, Omar 113
Hato Rey 183
Hawái 17, 66, 69, 70, 99, 100, 175
Herbáceos 108, 165
Heterótrofos 172
Hibridación 271
Hibridación de grupos 271
Hidrógeno 47, 64
Hidrológicos 145, 183, 213
Fotosíntesis neta 51
Fragmentación 134
Hidroperiodo 121, 125, 161, 162, 173, 177, 178, 181
Hierro 47, 48, 55, 88, 138, 168, 193
Hill, Robert Thomas 106
316 ariel e. lugo | ecología a m i m anera Índice de materias
22, 47-56, 61, 63, 64, 88, 129, 130, 138, 139, 153 -155, 233, 235, 237, 239, 240
35 -37, 42, 46, 47, 58, 64, 67, 68, 71, 74, 76, 117, 138, 178, 226, 246, 250, 251, 253, 257, 258, 276
Fotosíntesis
112
114
117
119
127
129
132
134
136
174
245
248
267
61, 105,
,
,
,
,
,
-
,
-
,
,
,
H
Historia de vida de una plantación 221,
Huracán 38, 40, 72, 111-114, 117-120, 123, 130, 134, 135, 137, 139, 141, 144, 146 -151, 177, 180, 213, 214, 219, 240 -242, 245
Huracán Hugo 141, 146, 147, 150, 241, 242, 245
Hutchinson, George Evelyn 19
Icacos 192, 194, 196
Hojarasca, caída total de 252
Hojarasca, concentración de cinco macronutrientes en la 233
Hojarasca en el suelo 211
Hojarasca, invertebrados en la 202
Hojarasca, masa total en la 230
Hojarasca, producción de 53
Hojarasca sobre el suelo 207
Hojarasca, tasa de caída de 54, 236
Hojas fragmentadas 230, 233
Holdridge, Leslie Ransselaer 19 Holística 278 Holling, Crawford Stanley 20
Inceptisoles 91, 121, 122
Índice de área foliar 59, 130, 131, 135
Índice de complejidad 206
Índice de complejidad de Holdridge 98, 207
Índice de similitud 270
Índice de superficie de tallos 59
Inspección de Montes de Puerto Rico 104
Instituto de Dasonomía Tropical 226
Instituto Internacional de Dasonomía
Tropical 19, 27, 110, 149, 151, 229
Intercambio catiónico 56, 197
Intercambio energético 185
Inundación 38, 74, 79, 118, 125, 161, 162, 170, 173, 175 -177, 249
Inventario biótico y ecológico 217
Irradiación, estudio de 113
Hongos mucosos 204
Hongos patógenos 268
Howard, Richard A. 111
Huella energética 163, 165, 166, 283
von Humboldt, Friedrich Heinrich Alexander 22
Humedal
,
,
Humedales, características comunes en los 162
Humedales, clasificación de 162, 163
Humedales costeros 173, 209
Isla de Mona 128, 174
Islas de Santa Cruz y San Juan 113
Islas Vírgenes 105, 113, 117, 127, 128, 131, 132, 141, 142, 188, 218
Isótopos 63, 70, 71, 183, 193
Jerarquías de estudio que aplican a los manglares 182
Juracán 187
Jurásico 263
317 ariel e. lugo | ecología a m i m anera Índice de materias
222
91
139
143
151
154
156
239
246
248
227
228
caída
35, 52-54, 151, 153, 154, 209, 211, 230, 234 -237, 239, 246, 248
Hojarasca 53, 56, 61,
,
,
,
,
,
, 202, 207, 211, 230 -237,
,
,
-250, 252 Hojarasca, acumulación de 91, 143,
,
Hojarasca,
de
-250
Homeostasis 29
31, 32, 34, 35, 38, 49, 50, 54, 57, 153 Hongos 52
154
155, 203, 204, 217, 264, 268
,
,
,
Hongos basidiomicetos 204
97
110
121
125
149
159
161
166
168
173
175
180
209
270
78 ,
,
,
,
,
-
,
-
,
,
,
,
, 281
I
J
K
Kangas, Patrick C. 163
Karso 55, 106, 111, 113, 120, 123, 170, 188, 193, 250
Karso, ciudadanos del 106, 113
L
Laderas 85, 103, 123, 125, 165, 168, 192, 215
Laguna Piñones 276
Lamarck, Jean-Baptiste 24
Latitudes 85 - 87, 114, 173, 254, 269
Laurance, William 265
Ledru, André Pierre 103
Leopold, Aldo 19, 276, 278
Leslie, Patrick Holt 24
Levantamiento tectónico 192, 193, 196, 197, 262
Ley de conservación de masa y energía 30, 45
Ley de Kleiber 57
Ley del aumento en la entropía 30
Ley del mínimo 47
Ley de máxima potencia 30, 213
Liebig, Justus 46
Lípidos neutrales 48
Litología 196
Livingston, Clara 27
Lixiviación 228
Lombrices de tierra 16, 154, 155, 227, 247
Los símbolos del lenguaje inventado por H.T. Odum (1971) 41
M
Mabuya 187
Macnae, William 175
Macronutrientes 47, 232, 233
Magnesio 47, 55, 193, 197, 232, 233
Manejo 63, 74, 75, 104 -106, 175, 221, 224, 258
Manganeso 47, 139
Manglar 59, 60, 85, 96, 103 -108, 110, 112, 149, 161-163, 170, 173 -175, 177-185, 209, 230, 245, 248 -250, 254, 276
Manglares de agua dulce 276
Mangle 55, 116, 149, 173, 175 -180, 183
Mangle blanco 175
Mangle botón 175
Mangle negro 175
Mangle rojo 149, 175
Maravilla natural del mundo 187
Maravilla natural del planeta 219
Mar Caribe 72, 125, 173
Mareas 41, 165, 173, 175, 176, 177, 180, 249,
Marrero Torrado, José 226
Materia
-252, 258
Materia orgánica muerta 36, 203
Matorrales 108, 216
Medina, Ernesto 54, 214
Mesetas 83
Mesetas intertropicales 83
Metabolismo ácido de crasuláceas (CAM) 130, 138
Metano 67, 125, 168
Meteorológicos 213
Micorrizas 52
Micorrízico 52
No-micorrízico 52
Micro humedales 165, 170
Micronutrientes 48
,
Microorganismos 49, 51, 52, 168, 183, 214, 228, 230, 246
Microorganismos descomponedores 51
Minerales primarios 192, 193, 196, 197
Mineralización 52, 53, 54
Mineralización de nitrógeno 54
Mioceno 188, 189, 194
318 ariel e. lugo | ecología a m i m anera Índice de materias
276
39
52
53
71
90
131
134
137
138
146
147
156
178
192
207
226
228
230
243
orgánica 35 -37,
,
,
,
,
,
,
,
,
,
,
,
,
,
, 202, 203,
,
-
,
,
246
Modelos de computadora 42
Moléculas de trifosfato de adenosina (ATP) 48
Molibdeno 48
Molina Colón, Sandra 132, 133
Molisoles 121, 122
clímax 17
275
Mortalidad base 146 Mortalidad de árboles 146 Mortalidad retrasada 146
Movimiento foliar 129
Murphy, Louis Sutliffe 107 Mutualismo 35
191, 263
49, 54,
, 232-234
Nutrientes, disponibilidad de 53, 129, 132, 206
Oligoceno 188, 189
Ordovícico 262, 263
Organelos 48
Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO) 221
Oxígeno 36, 46, 47, 52, 64, 67- 69, 125, 162, 165, 168, 171, 173, 199, 206, 249
Oxígeno, concentración de 165, 168, 199, 206
Oxígeno en el suelo 169, 214
P
Paleoceno 194
Paleógeno 63, 191, 263
Paleo-islas 192, 194
Palmares 107, 168, 169, 196, 209, 246
Palo colorado 168, 170, 196, 197, 202, 203, 209, 211
Parámetros estructurales 134, 135, 204
Parguera 69, 73, 174, 177, 178, 245, 250
Pastizales 65, 103, 107, 114, 216, 242, 270
Patrón asintótico 198
Patrón de función de paso 198
Patrón de sucesión 40
Patrones con una moda 198
Patrones de cambio 135, 198
Patrones de productividad primaria 39
Patrones monotónicos 198
Patrón estocástico 198
Peptidoglicano 48
Perfil del planeta Tierra 32
Perfiles de islotes de mangle 180
Perfiles de suelo 211
Periodos geológicos 63, 263
Pérmico 263
Océano Atlántico 125, 173
Ocean Park 183
Odum, Eugene Pleasants 20
Odum, Howard Thomas 18 -21, 23, 42
Peso específico foliar 59
Pesticidas 268
Pico del Este 194, 198
Pico del Oeste 112
319 ariel e. lugo | ecología a m i m anera Índice de materias
Montañas de Luquillo 26
111, 122, 141, 144, 149, 168, 169, 187-189, 192-194, 196 -200, 204, 206, 209, 211, 214 -219
Mono
,
Montserrat 153 Moral
N Neógeno
Neotrópico 105, 110, 112, 219, 265 Nitrógeno 47-56, 64, 67- 69, 114, 130, 131, 138, 139, 143, 144, 153 -155, 168, 171, 201, 207, 209, 211, 232, 233, 235 -237, 239, 240 Nitrógeno, concentración de 50 -53, 143, 144, 201, 236 Nitrógeno, masa y contenido de 201 Nutriente 47
48, 53, 228, 232, 233, 237 Nutrientes, acumulación de 202
228 Nutrientes, concentración de
,
,
228
O
Pino 60, 61, 91, 227, 228, 235,
Placa tectónica 187
Productividad bruta 35, 36, 177, 212, 249 Productividad maderera 243
Productividad neta 35, 36, 177, 211, 212, 247, 249
Plantación, definición de Evans 221
Plantaciones
Productividad primaria bruta 26, 211, 212, 248
Productividad primaria neta 211, 239
Productividad promedio 91
Productividad total 91
Programas de investigación ecológica sobre los bosques de Puerto Rico 106
Propiedades estructurales de hojas de dosel 201
Protección ambiental 225
Proteína 29, 30, 31, 48, 49 Proterozoico 263 Puerto Nuevo 183
320 ariel e. lugo | ecología a m i m anera Índice de materias
240
242
-
Plantación 221, 222, 227, 228, 237, 238, 240 -243, 247, 248, 270
Plantación de teca 227
arbóreas
, 223, 224, 225,
madereras
Plantaciones tropicales 221, 224 Plantas 15 -17, 22, 25, 26, 35 -37, 47-52, 54, 58, 61, 64, 71, 75, 77, 82, 86, 94, 96, 103, 105, 108, 110, 117, 122, 128 -130, 138, 151, 155, 156, 162, 174, 193, 201-203, 217, 226, 227, 246 -248, 251, 256, 262, 264, 266, 278, 281
221
226, 235, 246 Plantaciones
243
188
214 Pluviales 107
141, 142, 156, 253
alopátricas
47
193
197
228, 232, 233, 239, 240 Potencial de oxidación-reducción 168 Precipitación 89, 92, 94 -96, 125, 127, 157, 158, 198, 249, 252 Precipitación anual 92, 125, 127, 198, 249 Primer inventario de los bosques de Puerto Rico 104 Principio de persistencia 268 Principios de sistemas 283 Proceso oxidativo 36 Proceso reductor 36 Procesos geológicos 145 Producción 12, 30, 34, 35, 37- 40, 52, 53, 58, 70, 91, 104, 107, 117, 123, 154, 168, 222, 224, 225, 226, 230, 234, 242, 257, 281 Producción de semillas
Productividad 22, 26, 35, 36, 38, 39, 46 - 48, 52-54, 58, 64, 74, 75, 88, 90, 91, 95, 107, 121, 122, 130, 131, 171-174, 177, 178, 183, 193, 197, 207, 209, 211, 212, 224, 226, 228, 230, 237, 239, 243, 247-249, 253
Plataforma del Oligoceno-Mioceno 188 Plutónicas
,
,
Poblaciones
271 Potasio
,
,
,
35
Productividad primaria 26, 39, 46, 58, 91, 121, 122, 209, 211, 212, 237, 248, 253
194
196
197 Q Quema 38, 63, 67, 70, 71, 75, 256, 257, 258 Quema de combustible 71, 256, 257 Quitina 48 R Radiación 177 Raíces 35, 52, 106, 125, 130, 134, 136 -138, 146, 147, 162, 204 -206, 230, 237, 238, 246, 247
grandes
Raíces subterráneas
Reciclaje 34, 35, 88, 114, 121, 131, 137, 153, 156, 214, 243
246
249 Reciclaje de nutrientes 114, 131, 137, 153, 214
Reforestación
221
Puntos de inflexión (knickpoints) 193,
,
,
Raíces
205
146
,
,
, 243 Redfield, Alfred 47
75,
Regeneración asistido 222
Régimen de disturbios 213
Rehabilitación de tierras degradadas 242
Rehabilitación y restauración de suelos 242
Relación entre alometría y estequiometría 50
Relación entre área de hábitat y la acumulación de especies por área de hábitat 262
Relaciones alométricas en bosques 59
Relación especie-área 262, 266
Relación nitrógeno:fósforo 48
Reserva El Tallonal 113
283
Respiración del bosque 211
Respiración del suelo, tasa de 212 Respiración edáfica 113, 214, 249
Resurrección de árboles 146
Retranslocación 52, 130, 153, 228, 234
Retroalimentación negativa 32
Retroalimentación positiva 29, 32
Río Espíritu Santo 194, 196
Río Piedras 27, 103, 267
Río Prieto 194, 196
Río Sabana 194, 195, 196
RNA. Véase Ácido ribonucleico (RNA)
Roca madre (bedrock) 193, 197
Rocas calizas 128, 193
Rocas extrusoras 189
Rocas ígneas 115, 189, 192, 193
Rocas intrusivas 153, 189
Rocas plutónicas 188, 189, 214
Rocas volcánicas 189, 192
Rocas volcanoclásticas 153
Roqué de Duprey, Ana 25
S
Sabana 61, 113, 235, 238, 239
Salinidad 162, 173, 174, 177, 179, 180, 276
Salinidad en los suelos 179
San Juan 27, 71- 74, 103, 105, 113, 183, 269
San Patricio 103
Sapo concho en Guánica 267
Saprolita 192, 195
Saturación 51, 118, 121, 125, 162, 165, 199, 204, 215, 249, 253
Saturación hídrica 253
Scatena, Frederick N. 112
Sectores social-ecológico-tecnológicos 279
Secuestro de carbono en suelos y vegetación 242
Selección natural direccional 271
Selección natural disruptiva 271
Sequías 38, 74, 93, 113, 114, 118, 139, 213, 214, 249, 253
Servicio Forestal de Puerto Rico 107
Shelford, Víctor Ernest 15, 16, 83, 281
Siembra 84, 221, 222, 223
Siluriano 263
Simbiosis 34, 35
Síntesis de información dispersa 42
Sistema autotrófico 35, 37, 39, 178
Sistema de Holdridge 83, 94, 96, 97, 99, 100, 101, 127
Sistema ecológico, estratificación vertical de la estructura y el funcionamiento 37
Sistema ecológico joven 38
Sistema ecológico maduro 38, 40
Sistemas autotróficos abiertos 35
Sistemas autotróficos cerrados 35
Sistemas balanceados 39
321 ariel e. lugo | ecología a m i m anera Índice de materias
76, 112, 114, 127, 130, 132, 134, 136 -138, 147, 219, 268, 277 Respiración 26, 34 -39, 46, 52, 58, 59, 67, 68, 113, 117, 172, 177, 178, 197, 206, 211, 212, 214, 246, 249-251, 253, 257, 258, 276
Reservas de petróleo
Resiembra 223 Resiliencia 20, 39, 40,
Sistemas de Información Geográfica
(GIS) 258
Sistemas ecológicos
Sumidero de carbono atmosférico 174,
, 257, 258
218, 219, 226
Sistemas geológicos 187
Sistemas heterotróficos 37, 39, 276
Sistemas heterotróficos abiertos 35 Sistemas social-ecológicos-tecnológicos
41, 213
Sociedad Ecológica de Norte América
(ESA) 15, 281
Sodio 47, 60, 139, 193
Stahl Stamn, Agustín 24, 25, 105
Substratos 17, 60, 127
Substrato volcánico 130
Tansley, Sir Arthur George 17 Tasas de denudación 192
Tasas de erosión aguas arriba y aguas debajo 196 Teatro ecológico 33, 77
15, 16, 30, 37, 53, 174, 269
Sucesiones en manglares 179 Sucesión madura 37
Sucesión, sistemas en etapas jóvenes 40 Sucesión temprana 37 Sucesión,
de 37, 39 Suculentas 129, 130 Suelo, Órdenes de 91, 121
Temperatura promedio anual 198, 252
Templada cálida 96, 99, 173
Terciario 121, 263
Terrestres 33, 50, 156, 162, 165, 174, 254, 255, 258, 262
Thom, Bruce Graham 162
Thompson, D’Arcy Wentworth 56
Tierras bajas ecuatoriales 83
Tierras bajas interiores 83, 200 Tierras tropicales 83
Tipos funcionales de manglares 180
Topografía 64, 81, 86, 88, 98, 120, 121, 145, 148, 176, 245, 270
Tormentas 38
Trama evolutiva en el teatro ecológico 271
Transcolación 153
Transpiración 204, 206
322 ariel e. lugo | ecología a m i m anera Índice de materias
15
29
31
33
34
37
39
48, 63
75, 93, 118, 143, 163, 179
226
283
,
,
,
,
,
-
, 47,
,
, 187, 213, 217, 219,
,
61, 83, 95, 141, 142, 173, 253 Sucesión 15 -17, 30, 34, 37- 40, 53, 55, 94, 108, 110, 111, 113, 114, 120, 132-134, 138, 142, 148 -151, 157-159, 173, 174, 177-180, 242, 243, 269, 270
Sucesión
Subtropicales
Sucesión, aceleración de la 242
ecológica
Suelos 16, 52, 53, 55, 56, 60, 61, 81, 86, 88, 89-91, 94, 98, 104, 106, 108, 113, 118, 121, 122, 125, 129, 138, 144, 145, 156, 161, 162, 165, 168, 170, 179, 182, 192, 193, 196, 197, 199, 204, 206, 209, 211, 213 -216, 222, 224, 227, 228, 242, 247, 249, 252, 254 -256
arcillosos
165, 170
tipos
Suelos ácidos 56 Suelos anaeróbicos 168 Suelos
106, 125,
Suelos oxigenados 204
256
Suramérica
Sustratos aluviales 116
127 T Tabonuco 56, 60, 103, 112, 124, 135, 141, 143 -149, 151-159, 165, 169, 170, 177, 196, 197, 202, 203, 207, 209, 211, 215, 245, 247-250 Taínos 187 Tala 40, 55, 111, 130, 132, 135, 137, 159, 183
,
Tanques de bromelias 201, 202
Teca
227 Temperatura 30, 32, 67, 69, 71-75, 79, 82- 86, 88, 90, 92, 95, 96, 100, 113, 117, 119, 122, 129, 137, 177, 182, 197-200, 204, 215, 217, 243, 249, 252, 268
, 271
113,
Trayectoria de la sucesión 151
Trayectos de sucesión 178
Triásico 191, 262, 263
Tropicales 81, 90, 92, 95, 100, 103, 107,
W
Wadsworth, Frank H. 110, 226
Warming, Eugene 22
Watson, James Gilbert 175
Whittaker, Robert Hardin 17
Wilhelm Schimper, Andreas Franz 16
Winchell, Kristin 271
U
Ultisoles 91, 121, 122
Ultramáficos 108, 116, 127
Unión de tabonucos 156
Uso de agua promedio 227
Uso del agua 74, 138, 227
Uso de la tierra 19, 213, 215
V
Valles 108, 121, 123, 129, 165, 169, 170, 207
Valles aluviales costeros 121
Valor de Importancia 17, 78, 150 -152, 240, 242
Variables lentas 137
Variación espacial 143
Vegetación costera 254
Vida de una plantación 221, 222
Vientos alisios 117, 119, 120, 187
Volcán 81, 153, 189
Volcánicos 60, 116, 127, 129, 189
Volcanoclásticas 145, 153, 196
Volkswagen sedán de 1966 282
Volumen de madera 59
Volumen sobre la corteza, biomasa y almacenaje aéreo 60
Yucahú (Yucayú) 187
Zinc 48
Zonación 17, 173, 176
Zona de chapoteo de cataratas 165
Zona de vida subtropical húmeda 141
Zonas aluviales 120
Zonas geoclimáticas 115, 120
Zonas volcanoclásticas 145
Zonificaciones 173
323 ariel e. lugo | ecología a m i m anera Índice de materias
227
258
112, 114, 120, 173, 177, 213, 221, 224,
, 228, 246, 249, 252, 253, 255, 257,
, 266, 269, 277, 282
Y
Z
Acacia farnesiana 132
Acrostichum 108, 111
Acrostichum aureum 176
Agave 111
Amyris 111
Anolis cristatellus 271
Anolis gundlachi 75
Anolis stratulus 271
Anthocephalus chinensis 229 -231, 233 -235, 237
Avicennia 179
Avicennia alba 176
Avicennia germinans 55, 175
Avicennia intermedia 176
Batis 108, 111
Borrichia 111
Bruguiera caryophylloides 176
Bruguiera eriopetala 176
Bruguiera gymnorhiza 176
Bruguiera parviflora 176
Bucida 111
Bucida buceras 103
Bursera simaruba 130
Cacti 111
Carapa moluccensis 176
Carapa obovata 176
Castalia 111
Castilla elastica 246, 248, 250, 251
Cecropia schreberiana 149
Cedrela odorata 103, 113
Ceiba pentandra 103
Índice de especies
Ceratophyllum 111
Ceriops candolleana 176
Cladium jamaicense 169
Coccoloba uvifera 103, 108, 127
Coccolobis 111
Crecropia schreberiana 201
Cyathea bryophila 123
Cyrilla racemiflora 61, 168
Dacryodes excelsa 55, 56, 60, 61, 103, 123, 141, 201, 248, 250
Daphnopsis philippiana 123
Eleutherodactylus eneidae 268
Eleutherodactylus jasperi 268
Eleutherodactylus karlschmidti 268
Eucalyptus saligna 229 -231, 233 -237
Eucalyptus x patentinervis 229 -231, 233 -237
Fimbristylis 111
Fraxinus pennsylvanica 170
Guaiacum officinale 103
Guarea glabra 123
Guazuma 111
Heliconia caribaea 123
Hernandia sonora 228 -231, 233, 234 -237
Hibiscus elatus 229 -231, 233 -235, 237
Hirtella rugosa 123
Intsia retusa 176
Khaya nyasica 229 -231, 233 -235, 237
Laguncularia 179
Laguncularia racemosa 55, 175
Leucaena leucocephala 56, 132
324 ariel e. lugo | ecología a m i m anera Índice de especies
Liquidambar styraciflua 170
Liriodendron tulipifera 170
Lobelia portoricensis 123
Magnolia splendens 123
Manilkara bidentata 123, 157, 201
Mariscus 111
Melicoccus bijugatus 132
Miconia prasina 55
Micropholis garciniifolia 201
Miconia sintenisii 123
Miconia tetrandra 123
Micropholis garciniifolia 123, 201
Nyssa aquatica 170
Ocotea leucoxylon 123
Ocotea spathulata 123
Palaemon pandaliformis 267
Parkisonia 111
Peltophryne lemur 267
Philoxerus 111
Pinus caribaea 60, 61, 229 -235, 237-242
Pinus elliottii 229 -231, 233 -235, 237
Pistia 111
Pithecellobium 111
Platanus occidentalis 170
Potamogeton 111
Psychotria berteriana 123
Psychotria maleolens 123
Prestoea montana 60, 61, 123, 149, 168, 201, 246, 248
Prosopis juliflora 132
Pterocarpus 108, 111, 116, 209
Pterocarpus officinalis 55, 169
Rheedia portoricensis 123
Rhizophora 179
Rhizophora conjugata 176
Rhizophora mangle 149, 175
Rhizophora mucronata 176
Sesuvium 108, 111
Sloanea berteriana 123, 157
Sonneratia griffithii 176
Spathodea 60
Spathodea campanulata 56, 60, 246
Sphagnum portoricense 169
Sporobolus 111
Swietenia 226
Swietenia humilis 226
Swietenia macrophylla 60, 61, 113, 226, 229 -231, 233 -239, 248
Swietenia mahagoni 113, 132, 226
Syzygium jambos 157, 158
Tabebuia heterophylla 56, 129, 149
Tabebuia rigida 123
Tectona grandis 113, 227
Terminalia ivorensis 229 -231, 233 -235, 237
Typha 111
Tyrannus dominicensis 270
325 ariel e. lugo | ecología a m i m anera Índice de especies
El libro se basa en un curso de la Universidad de Puerto Rico sobre la historia natural de Puerto Rico. El énfasis es en los bosques de Puerto Rico y cubre los tópicos más relevantes sobre la conservación de la biodiversidad tropical. El contenido es rico en material gráfico que ilustra las bases científicas de las ideas discutidas en el texto.
En Puerto Rico no existe ningún texto similar que resuma de forma sucinta el conocimiento actual sobre la estructura y funcionamiento de los bosques de la isla. Más aún, contiene un examen crítico de las grandes controversias de la conservación moderna, incluyendo la extinción de especies, las especies no nativas que algunos llaman invasivas, el rol de los bosques tropicales en el ciclo del carbono, el cambio climático, la importancia de plantaciones arbóreas, el efecto del Antropoceno sobre los bosques de Puerto Rico, la ética en la ecología, entre otras.
