Ø.V.


Ø.V.
edizione italiana a cura di carlo


Ø.V.
SJAASTAD O. SAND K. HOVE
Titolo originale: Physiology of Domestic Animals, 2nd edition, by Ø.V. Sjaastad, O. Sand and K. Hove, 2010
Traduzione autorizzata dell’edizione originale in lingua inglese: Sjaastad Ø.V., Sand O. e Hove K., Physiology of Domestic Animals, pubblicata da Scandinavian Veterinary Press, Oslo – prima edizione, 2003; seconda edizione, 2010.
Copyright © Scandinavian Veterinary Press 2010
Copyright © 2013 C.E.A. Casa Editrice Ambrosiana
I diritti di elaborazione in qualsiasi forma o opera, di memorizzazione anche digitale su supporti di qualsiasi tipo (inclusi magnetici e ottici), di riproduzione e di adattamento totale o parziale con qualsiasi mezzo (compresi i microfilm e le copie fotostatiche), i diritti di noleggio, di prestito e di traduzione sono riservati per tutti i paesi. L’ acquisto della presente copia dell’opera non implica il trasferimento dei suddetti diritti né li esaurisce.
Fotocopie per uso personale (cioè privato e individuale) possono essere effettuate, nel limite del 15% di ciascun volume, dietro pagamento alla SIAE del compenso previsto dall’art. 68, commi 4 e 5, della legge 22 aprile 1941, n. 633. Tali fotocopie possono essere effettuate negli esercizi commerciali convenzionati SIAE o con altre modalità indicate da SIAE. Per riproduzioni ad uso non personale (per esempio: professionale, economico o commerciale) l’editore potrà concedere a pagamento l’autorizzazione a riprodurre un numero di pagine non superiore al 15% delle pagine del presente volume. Le richieste per tale tipo di riproduzione vanno inoltrate a: Centro Licenze e Autorizzazioni per le Riproduzioni Editoriali (CLEARedi) Corso di Porta Romana 108 20122 Milano e-mail: autorizzazioni@clearedi.org e sito web: www.clearedi.org
L’editore, per quanto di propria spettanza, considera rare le opere fuori del proprio catalogo editoriale. La riproduzione degli esemplari esistenti nelle biblioteche di tali opere è consentita, non essendo concorrenziale all’opera. Non possono considerarsi rare le opere di cui esiste, nel catalogo dell’editore, una successiva edizione, le opere presenti in cataloghi di altri editori o le opere antologiche.
Revisione: Carlo Tamanini
Traduzione: Paolo Accornero (capp. 5 e 18), Maria Laura Bacci (capp. 13 e 17), Mario Baratta (capp. 7, 20, 21), Barbara Barboni (capp. 6, 16), Giuseppina Basini (capp. 2, 3, 10), Cristiano Boiti (capp. 9, 12), Francesca Grasselli (capp. 4, 8), Mauro Mattioli (cap. 19), Angelo Quaranta (cap. 11), Marcella Spinaci (cap. 15), Antonia Zarrilli (capp. 1, 14)
Realizzazione editoriale: Epitesto, Milano
Impaginazione: Pre&Stampa, Segrate
Disegni (se non diversamente specificato): Anne Langdalen, Kari C. Toverud
Copertina: 46xy, Milano
Immagini di copertina: sezioni sagittali del cranio e dell’occhio di un gatto domestico (Dorling Kindersley/Getty Images): sullo sfondo rappresentazione di cellule nervose (ktsdesign/Shutterstock).
Prima edizione: gennaio 2013
Ristampa 4 3 2 1 0 2013 2014 2015 2016 2017
Realizzare un libro è un’operazione complessa, che richiede numerosi controlli: sul testo, sulle immagini e sulle relazioni che si stabiliscono tra loro. L’esperienza suggerisce che è praticamente impossibile pubblicare un libro privo di errori. Saremo quindi grati ai lettori che vorranno segnalarceli.
Per segnalazioni o suggerimenti relativi a questo libro rivolgersi a:
C.E.A. Casa Editrice Ambrosiana viale Romagna 5, 20089 Rozzano (MI) fax 02 52202260 e-mail: redazione@ceaedizioni.it
Sul sito www.ceaedizioni.it è possibile verificare se sono disponibili errata corrige per questo volume. Accedendo, attraverso il menu sulla sinistra, alla pagina Per l’università, è possibile visualizzare l’elenco dei volumi per i quali è disponibile un errata corrige cliccando sulla voce Errata corrige presente nelle diverse categorie dell’elenco degli strumenti per lo studio. Nel caso, è possibile scaricare il relativo PDF alla sezione Servizi della scheda dedicata al volume, raggiungibile cliccando sul titolo del volume stesso.
Stampato da Pirovano s.r.l. via della Pace 19, San Giuliano M.se (MI) per conto della C.E.A. Casa Editrice Ambrosiana, viale Romagna 5, 20089 Rozzano (MI)
Revisione:
Carlo Tamanini, Dipartimento di Scienze Mediche Veterinarie, Alma Mater Studiorum Università di Bologna
Traduzione di:
Paolo Accornero, Dipartimento di Scienze Veterinarie, Università degli Studi di Torino
Maria Laura Bacci, Dipartimento di Scienze Mediche Veterinarie, Alma Mater Studiorum Università di Bologna
Mario Baratta, Dipartimento di Scienze Veterinarie, Università degli Studi di Torino
Barbara Barboni, Dipartimento di Scienze Biomediche Comparate, Università degli Studi di Teramo
Giuseppina Basini, Dipartimento di Scienze Medico-Veterinarie, Università degli Studi di Parma
Cristiano Boiti, Dipartimento di Scienze Biopatologiche Veterinarie ed Igiene delle Produzioni Animali e Alimentari, Università degli Studi di Perugia
Francesca Grasselli, Dipartimento di Scienze Medico-Veterinarie, Università degli Studi di Parma
Mauro Mattioli, Dipartimento di Scienze Biomediche Comparate, Università degli Studi di Teramo
Angelo Quaranta, Dipartimento di Medicina Veterinaria, Università degli Studi di Bari “Aldo Moro”
Marcella Spinaci, Dipartimento di Scienze Mediche Veterinarie, Alma Mater Studiorum Università di Bologna
Antonia Zarrilli, Dipartimento dell’Emergenza e dei Trapianti di Organi, Università degli Studi di Bari “Aldo Moro”
3 Dalle cellule
4 Il sistema nervoso Aspetti
Conduzione degli impulsi nervosi negli assoni mielinici
Sinapsi
Sinapsi chimiche tra neuroni
Sviluppo e struttura del sistema nervoso centrale
cerebrale degli input provenienti dagli organi dell’equilibrio
Divisione metamerica del midollo spinale e dei nervi spinali
Struttura interna del midollo spinale
Nervi spinali
Riflessi
Encefalo
Tronco dell’encefalo e nervi cranici
Cervelletto
Diencefalo 134
Cervello 135
Sistema nervoso autonomo
Neurotrasmettitori e recettori
Sintesi, secrezione e metabolismo degli ormoni tiroidei
Regolazione della sintesi e della secrezione degli ormoni tiroidei
Azione degli ormoni tiroidei
Ghiandole surrenali
Steroidi
Ormoni
Regolazione ormonale del metabolismo
Aspetti
Composizione e funzioni del tessuto osseo
Formazione di tessuto osseo
Tessuto osseo compatto e trabecolare (spugnoso)
Riassorbimento e rimodellamento osseo
Articolazioni
Regolazione dell’omeostasi del calcio e del fosfato
Vitamina D
Ghiandole paratiroidee
Ormone paratiroideo
Calcitonina
Integrazione della regolazione ormonale del
Tessuti che trasportano Ca2+
Ghiandola del guscio
Meccanismi di difesa innati
Elementi esterni della difesa innata
Elementi interni della difesa innata
Infiammazione
Meccanismi di difesa acquisiti
Immunità mediata da anticorpi
Immunità adattativa cellulare
(difesa da parte delle cellule T)
Trasferimento degli anticorpi materni al feto e al neonato
Gruppi sanguigni
Aspetti principali
Cuore
Struttura
Sistema di conduzione
Elettrocardiografia
Derivazioni bipolari
Derivazioni unipolari
Asse elettrico del cuore
Uso diagnostico
Fasi del ciclo
Gittata cardiaca
Regolazione della frequenza cardiaca
Regolazione della gittata sistolica
Interazione tra la regolazione di VTD e VTS
Lavoro cardiaco
Pressione arteriosa e gittata sistolica
383
383
384
Vasi sanguigni e pressione del sangue 385
Leggi fisiche che regolano il flusso dei fluidi nei vasi
Relazione tra pressione, flusso e resistenza vascolare nella circolazione sistemica
Arterie e pressione arteriosa
Arteriole e regolazione del
385
Reni 460
Struttura del nefrone 461
Flusso ematico e pressione idrostatica nei vasi renali
Funzioni del glomerulo
Velocità di filtrazione glomerulare
Fattori che influenzano la filtrazione
Regolazione della velocità di filtrazione glomerulare
Pressione ematica e VFG
tubulare
Riassorbimento tubulare 473
Relazioni tra morfologia e funzioni delle cellule tubulari 473
Meccanismi di riassorbimento 474
Secrezione 477
Manipolazione tubulare di sostanze importanti
Ormone antidiuretico (ADH) 484
Produzione di urine concentrate o diluite 484
Regolazione del volume e dell’osmolarità dei liquidi corporei
Regolazione del volume del liquido extracellulare
Regolazione dell’osmolarità del liquido extracellulare
Volume urinario
Vie urinarie
Bilancio idrico complessivo
Acquisizione e perdita di acqua
Differenze di specie
Funzioni renali nell’invecchiamento
492
Alterazioni nell’equilibrio acido-base 515
Alterazioni acido-base di tipo respiratorio .
504
506
Funzioni renali nei neonati 506
Funzioni renali negli uccelli 507
14 Regolazione acido-base
Aspetti principali 510
Scala del pH 510
pH dei liquidi corporei
Formazione di H+
511
Anidride carbonica come fonte indiretta di H+ . 511
Acidi non volatili come sorgenti di H+ . .
. 511
Basi provenienti dalla dieta 512
Differenze di specie nella produzione di acidi non volatili e assunzione di basi con la dieta . . . 512
Tamponi 514
Tamponamento di acidi non volatili 515
516
Disturbi acido-base metabolici (non respiratori) 516 Produzione di HCO3– nei reni
Tamponamento di H+ nei tubuli
Regolazione della secrezione renale di H+ . . . . . 522 Correlazione tra K+ ed
Differenze di base nei processi nutritivi e digestivi
Digestione microbica delle proteine 564
Assorbimento a livello dei prestomaci
Metabolismo delle tossine nel rumine
. . 567
. . 569
Omaso 569
Stomaco 570
Funzioni dello stomaco .
570
Anatomia 570
Contrazioni della muscolatura dello stomaco . . 572
Secrezione del succo gastrico
575
Abomaso 579
Pancreas 579
Anatomia
580
Composizione ionica del succo pancreatico 581
Differenze di specie nella secrezione pancreatica 583
Fegato, dotti biliari e cistifellea 584
Anatomia
Aspetti principali del metabolismo energetico . 612
Fase di assorbimento e di postassorbimento 613
Fase di assorbimento 614
Fase di postassorbimento 619
Regolazione del metabolismo dei nutrienti organici 625 Insulina 626 Glucagone .
(adrenalina) e sistema
Aspetti principali
584
Funzioni del fegato 585
Produzione della bile 586
Regolazione della secrezione biliare
. . 587
Digestione nel piccolo intestino 588
Motilità del piccolo intestino 589
Svuotamento del piccolo intestino
590
Secrezione nel piccolo intestino 591
Degradazione e assorbimento dei nutrienti nel piccolo intestino
Aspetti generali dell’assorbimento
591
591
Idrolisi 592
Meccanismi assorbitivi
592
Degradazione e assorbimento dei carboidrati 594
Digestione e assorbimento dei lipidi . . . . . . . . 599
Assorbimento dell’acqua 601
Assorbimento del ferro e del calcio 601
Grosso intestino
603
Anatomia 603
Aspetti generali della digestione nel grosso intestino
604
Digestione nel grosso intestino negli erbivori monogastrici 605
Motilità del grosso intestino
606
Regolazione delle contrazioni del grosso intestino 607
Svuotamento del retto
Digestione negli uccelli
Assunzione dell’alimento e digestione meccanica
608
609
609
Digestione enzimatica e assorbimento dei nutrienti 610
Funzioni della cloaca
610
Regolazione della temperatura corporea 664
Processi regolatori
665
Termoregolazione all’interno della zona di termoneutralità 666
Regolazione della temperatura al di sotto della temperatura critica inferiore 668
Regolazione della temperatura al di sopra della temperatura critica superiore 669
Adattamento al caldo
Febbre . .
Caratteristiche anatomiche importanti per la regolazione termica
Aspetti principali
670
670
671
676
Sistema riproduttivo 676
Maschio
677
Femmina 677
Controllo endocrino della riproduzione 678
Gravidanza
678
Controllo endocrino dell’attività gonadale 678
Sviluppo dei caratteri sessuali
Inattivazione del centro della secrezione fasica nel maschio
679
680
Pubertà 681
Fisiologia della riproduzione maschile
682
Struttura dell’apparato riproduttore maschile 682
Produzione degli spermatozoi 686
Controllo endocrino della riproduzione nel maschio
690
Fisiologia della riproduzione femminile 693
Struttura dell’apparato riproduttore femminile 693
Ciclicità ovarica
696
Maturazione della cellula uovo 700
Ormoni sessuali nella femmina
705
Comportamento sessuale negli animali domestici 708
Comportamento sessuale in animali monogami e poligami 708
Stagionalità riproduttiva
Trasporto degli spermatozoi, capacitazione e fecondazione
711
712
Trasporto degli spermatozoi 712
Capacitazione
713
Fecondazione 713
Gravidanza .
. 714
Invogli fetali 715
Riconoscimento materno di gravidanza e mantenimento del corpo luteo 715
Impianto e sviluppo della placenta 716
Trasferimento di ossigeno, nutrienti, cataboliti e calore attraverso la placenta 718
Produzione di ormoni durante la gravidanza 718
Parto e puerperio 719
Endocrinologia del parto 719 Fasi del parto 720
Involuzione uterina 721
Tecnologie riproduttive negli animali domestici 721
Raccolta e conservazione del seme 721
Trasferimento embrionale e clonazione 722
Riproduzione negli uccelli 722
Organi riproduttivi della femmina .
Organi riproduttivi del maschio
Aspetti principali
della vacca
e crescita della ghiandola mammaria
Sviluppo durante la vita fetale
Sviluppo dalla nascita fino all’età adulta 733
Sviluppo durante la gravidanza e la lattazione 733
Modificazioni del tessuto mammario durante la lattazione 734
Involuzione del tessuto mammario
svezzamento
ormonale della secrezione di latte
Mantenimento della produzione di latte
del latte in diverse specie .
Sintesi dei componenti del latte
epiteliali mammarie
Altri fattori che influenzano lo svuotamento della mammella
Variazioni della produzione di latte
dall’ormone della crescita
Aspetti principali
Assorbimento dei substrati energetici 755
Metabolismo basale 756
Indice metabolico basale 756
Indice metabolico a digiuno
Indice metabolico di mantenimento
Indice metabolico di campo
Relazione tra massa corporea e indice metabolico
756
757
757
Calcolo dell’energia . Necessità energetiche degli animali 758
Misurazione del metabolismo energetico 759
Quoziente respiratorio
Incremento del calore dovuto ai processi digestivi
metabolica aerobica
Utilizzo dell’energia presente nell’alimento
dell’energia nel corpo
Crescita corporea e rigenerazione
Invecchiamento negli animali
di misura
Illustratori e fonti delle figure
Indice analitico
La prima edizione di questo testo di Fisiologia degli animali domestici nella sua versione in lingua inglese è datata 2003; successivamente (2004 e 2007) ci sono state delle ristampe e, finalmente, nel 2010 è uscita la seconda edizione. Il testo ha avuto e ha una vasta diffusione nei Paesi scandinavi, negli Stati Uniti e, in genere, nei Paesi anglosassoni.
La nuova edizione è stata sensibilmente migliorata rispetto alla precedente, sia per i contenuti che per la grafica. I diversi capitoli della Fisiologia veterinaria sono trattati in maniera compiuta; i riferimenti ai vari animali domestici sono frequenti e sempre pertinenti. Rispetto alla prima edizione, a margine sono state aggiunte numerose annotazioni che permettono al lettore di focalizzare al meglio l’argomento trattato, nonché una notevole serie di domande finaliz-
zate a una sorta di autovalutazione; siamo sicuri che gli studenti apprezzeranno. Molto belle sono le figure, spesso di una semplicità addirittura sconcertante ma sicuramente utili al fine di facilitare l’apprendimento.
Tutte le caratteristiche di cui sopra ci hanno fatto ritenere opportuna un’edizione italiana. Siamo certi di fornire un valido supporto didattico soprattutto agli studenti di Medicina veterinaria, ma non solo: anche Studiosi e Ricercatori di discipline biologiche nonché professionisti interessati alle Produzioni animali avranno modo di apprezzare la chiarezza con cui vengono descritti i meccanismi fisiologici più complessi.
Prof. Carlo Tamanini
Professore Ordinario di Fisiologia Veterinaria
Alma Mater Studiorum Università di Bologna
La Fisiologia è la scienza che studia le funzioni degli organismi viventi; questo libro ha lo scopo di guidare alla comprensione delle funzioni corporee nei mammiferi e negli uccelli. Sebbene sia stato concepito per e sia rivolto principalmente a studenti dei corsi di Medicina veterinaria e di Scienze animali, l’approccio utilizzato per la sua preparazione lo rende utile anche per i corsi universitari di Fisiologia generale dei mammiferi e degli uccelli.
Lungo tutto il testo abbiamo enfatizzato la spiegazione di aspetti concettuali e abbiamo sottolineato come i meccanismi di regolazione mantengano stabile l’ambiente interno dell’organismo, nonostante l’ambiente esterno si modifichi continuamente (omeostasi). Il volume ha lo scopo di fornire una spiegazione comprensibile del funzionamento dell’organismo e il suo obiettivo finale è di mettere gli studenti nella condizione di acquisire le conoscenze e di sviluppare la capacità di soluzione dei problemi delle quali avranno bisogno nella loro vita lavorativa futura.
Siamo riconoscenti a tutti coloro che ci hanno assistito nel lungo cammino che ha portato alla conclusione di questo lavoro. Un grazie va ad Arne G. Ruud e al National Centre for Ve-
terinary Contract Research and Commercial Service Ltd (VESO) per aver supportato il progetto. Ringraziamo anche i nostri principali disegnatori, Anne Langdalen e Karl C. Toverud e la consulente linguistica della seconda edizione, Lucy Robertson. Per le discussioni oltremodo utili e per il confronto pedagogico e professionale ringraziamo i nostri studenti della Norwegian School of Veterinary Science, Oslo University e Norwegian University of Life Science. Non per ultimi, ringraziamo i Colleghi che hanno contribuito al processo di miglioramento della qualità del testo. Siamo particolarmente grati a Øystein Andresen, Jon-Arne Birkeland, Inge Bjerkås, Peter Bols, David Cotrell, Odd Magne, Egil Haug, Helga Høgåsen, Johan Høgset Jansen, Clarence Kvart, Mette O. Nielsen, B. E. C. Nordin, Kerstin Olsson, Reeta Pøsø, Lucy Robertson, Anne Storset, Rosanne Taylor, Michael Tranulis e Reidun Ursin per i loro utili suggerimenti e le loro critiche costruttive. Un grazie va anche alla Direttrice della Biblioteca della Norwegian School of Veterinary Science, Anne Cathrine Munthe.
Øystein V. Sjastaad Olav Sand Knut Hove
I processi vitali sono complessi e anche nel caso degli organismi unicellulari più semplici i meccanismi fisiologici non sempre sono stati chiariti compiutamente. Pertanto, com’è possibile comprendere il funzionamento dell’organismo dei vertebrati, che può essere costituito da più di cento migliaia di miliardi di cellule, ognuna delle quali costituisce un’unità ben delimitata? Nel loro insieme, le cellule contribuiscono a formare un organismo, con la sua specifica identità. Al fine di permettere questa collaborazione, le cellule si scambiano continuamente informazioni e innumerevoli meccanismi di regolazione mantengono stabile l’ambiente corporeo interno, nonostante continue variazioni dell’ambiente esterno. La Fisiologia animale è pertanto una scienza estremamente complicata e il tentativo di penetrare tale complessità può sembrare uno sforzo inutile. Tuttavia, i principi della regolazione e del controllo della maggior parte dei processi fisiologici sono piuttosto simili e le caratteristiche più importanti della fisiologia animale sono di comprensione relativamente semplice.
Al fine di strutturare al meglio questo campo così complesso, i testi di fisiologia di norma suddividono le funzioni corporee in alcuni capitoli principali come la digestione, la riproduzione o la circolazione del sangue. I titoli dei capitoli di questo volume mettono in luce il sistema di suddivisione da noi scelto. La classificazione della fisiologia in singoli argomenti può portare al concetto che i diversi sistemi operano in maniera indipendente l’uno dall’altro, ma di fatto non è così: l’organismo è un’unica unità funzionale e i processi fisiologici in tutte le cellule e in tutti gli organi sono strettamente collegati fra loro. Noi abbiamo cercato di enfatizzare questa unità mediante continui riscontri incrociati fra i diversi capitoli.
Non è possibile comprendere le funzioni corporee senza delle conoscenze anatomiche di base. Tuttavia, poiché i riferimenti all’anatomia in questo testo sono limitati al minimo indispensabile per la comprensione dei processi fisiologici, si raccomandano ulteriori rivisitazioni dell’anatomia degli animali domestici. Nella fisiologia, l’ingegnosamente semplice e l’incomprensibilmente complesso sono due facce della stessa medaglia. Le nostre personali ricerche in vari ambiti della fisiologia ci hanno dato una sensazione di umile venerazione per l’incredibile sistema di processi e dettagli che costituiscono un organismo vivente. La nostra speranza è che questo testo sia in grado di trasmettere questa sensazione anche ai nostri lettori.
Al fine di avere un’idea della fisiologia sono necessarie conoscenze di base della fisica, della chimica e della biochimica. Pertanto abbiamo previsto un capitolo introduttivo che presenta una panoramica di alcuni dei principali argomenti in questi campi. Non ci sono confini logici fra la biologia cellulare e la fisiologia e la conoscenza della biologia cellulare costituisce una piattaforma per la comprensione della fisiologia animale. Di conseguenza, il secondo capitolo costituisce di fatto un’introduzione alla biologia cellulare. Ogni capitolo inizia con una breve descrizione degli aspetti principali degli argomenti trattati; la parte successiva fornisce le basi per una comprensione più approfondita dei processi fisiologici.
Il testo contiene circa 200 esempi clinici, scelti e selezionati per illustrare le normali funzioni dell’organismo, piuttosto che per entrare nel
Le annotazioni a margine sintetizzano i punti principali del testo e sono utili per il ripasso del capitolo
dettaglio dei processi patologici o del trattamento delle diverse patologie. Un metodo efficace per dimostrare l’importanza di uno specifico meccanismo fisiologico è la descrizione dei sintomi che si manifestano quando questo meccanismo non opera in maniera corretta. Crediamo anche che questi esempi possano motivare lo studente e stimolare la sua curiosità.
A supporto della trattazione dei diversi argomenti sono state previste molte domande, in totale circa 2000, distribuite in tutto il volume, che possono essere utilizzate per verificare se il contenuto dei vari argomenti è stato compreso. Le risposte alle domande possono essere trovate leggendo attentamente il testo principale. Riteniamo che questo approccio sia un metodo molto più efficace per facilitare l’apprendimento
di quanto non sarebbe mettere le risposte a disposizione in una sezione a parte.
A margine del testo sono state inserite inoltre più di 3000 annotazioni che sottolineano i punti principali degli argomenti trattati e che saranno di particolare importanza quando lo studente vorrà ripassare i capitoli. Siamo certi che queste note a margine possano migliorare l’efficacia dell’apprendimento.
Alla fine di ogni capitolo abbiamo incluso un elenco di altri testi raccomandati per ulteriori approfondimenti.
Nell’indice, di facile consultazione, il numero delle pagine in grassetto orienta il lettore verso le principali descrizioni o definizioni, mentre i numeri in corsivo fanno riferimento alle figure.
978-88-08-18293-7
Gli esempi clinici sottolineano l’importanza dei normali processi fisiologici e indirizzano a un ulteriore approfondimento
Le domande si riferiscono ad aspetti fondamentali e sono uno strumento di verifica dell’apprendimento.
Le risposte sono facilmente individuabili nel testo
292Il muscolo
Nell’ipertrofia
di nuclei nelle fibre muscolari aumenta

Nell’atrofia muscolare il numero di nuclei nelle fibre muscolari non varia I movimenti degli arti sono causati da contrazioni sequenziali di muscoli antagonisti
I muscoli propulsivi sono inseriti sulle ossa in modo tale da amplificare movimenti
normale attività. L’atrofia muscolare può svilupparsi rapidamente anche quando l’inattività è la conseguenza di una lesione nervosa. Il tessuto muscolare scheletrico contiene cellule staminali chiamate cellule satellite. L’ipertrofia muscolare è associata all’aumento del numero di nuclei conseguente alla fusione tra cellule satellite e fibre del muscolo in accrescimento. Tuttavia, in una fibra che si sta atrofizzando i nuclei non vengono persi e questo facilita una rapida ricrescita che tende a ripristinare le dimensioni originarie della fibra a patto che il miglioramento delle condizioni consenta una normale attività muscolare. Questa proprietà del muscolo scheletrico è chiamata memoria del muscolo
Interazioni tra muscoli e scheletro
Quando un muscolo si accorcia, i punti di inserzione dei tendini si avvicinano, muovendo così le ossa. In generale, un punto di inserzione rimane quasi fermo durante l’accorciamento del muscolo e viene chiamato quindi origine del muscolo stesso, mentre il punto che si muove di più è chiamato inserzione. Nella maggior parte dei casi l’origine è prossimale all’inserzione. Molti muscoli possono determinare il movimento di ogni articolazione; questi sono dispo-
Confronta
8.19 Muscoli antagonisti. Quando il muscolo semitendinoso si contrae, l’arto posteriore si flette e il muscolo vasto laterale viene stirato. Con la contrazione del muscolo vasto laterale la gamba viene estesa e il muscolo semitendinoso viene stirato.
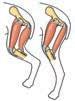
sti in gruppi che causano movimenti opposti a livello articolare (gruppi di muscoli antagonisti fig. 8.19). Per esempio, il muscolo semitendinoso dell’arto posteriore appartiene al gruppo dei muscoli flessori che flettono la gamba, mentre il muscolo vasto laterale fa parte del grande gruppo dei muscoli estensori antagonisti (quadricipite femorale) che la estende. Come già osservato, un muscolo non può stirarsi attivamente, ma la contrazione di un muscolo specifico determina sempre lo stiramento dei suoi antagonisti. La capacità di una forza di far ruotare una leva è determinata dal prodotto della forza stessa per il suo braccio, posto che la forza agisca perpendicolarmente alla leva. Il braccio è la distanza fra il fulcro e la linea di azione della forza. La figura 8.20 illustra la posizione approssimativa delle ossa nell’arto anteriore destro di un gatto che sta correndo quando la zampa si appoggia al suolo. La distanza tra la zampa e il fulcro nell’articolazione della spalla è circa undici volte maggiore della distanza tra il fulcro e l’inserzione tendinea del muscolo grande rotondo sull’omero. Affinché questo muscolo sia in grado di fornire una forza propulsiva dalla zampa al suolo, è necessario che sviluppi una forza undici volte maggiore; in altre parole, il tendine deve esercitare sull’osso una forza molto maggiore della forza esterna. Questo è tipico dei tendini e dei muscoli responsabili di estesi movimenti oscillatori delle estremità. I rapporti anatomici illustrati nella figura 8.20 indicano inoltre che l’ampiezza e la velocità dei movimenti della zampa sono molto maggiori dell’accorciamento del muscolo. Nell’esempio descritto, il muscolo grande rotondo deve accorciarsi solo di 1 cm per muovere la zampa di 11 cm. Tale aumento di ampiezza del movimento a spese della forza ha indubbi vantaggi. Se è necessario un aumento della massima forza contrattile, il muscolo può facilmente aumentare la sua area di sezione trasversa. Poiché i muscoli sono connessi allo scheletro in modo da amplificare i movimenti, le possibili deviazioni delle estremità sono adeguate anche se i muscoli lavorano costantemente nell’ambito della loro limitata lunghezza ottimale. Tendini
Quando la forza contrattile dei muscoli scheletrici viene trasferita allo scheletro, le ossa possono essere sottoposte a forze molto intense; per que
Molti dei muscoli presenti nell’organismo sono in grado di sviluppare una notevole forza contrattile e ciò ovviamente richiede la presenza di tendini forti con robuste inserzioni. Gli sforzi estremi, tuttavia, possono determinare il distacco del tendine dall’osso oppure causare fratture ossee nella
sta ragione i tendini che collegano i muscoli alle ossa devono essere molto forti ed essere inseriti saldamente alle due estremità. I tendini e i legamenti articolari sono costituiti da fibre collagene parallele, densamente ammassate, che conferiscono al tendine una notevole forza elastica. Un tendine con un diametro di 7 mm sopporta senza lacerarsi una forza di stiramento di circa 5000 N. Le fibre collagene nel tendine si estendono direttamente nella rete di fibre collagene che formano il periostio e in quelle che rinforzano il tessuto osseo (fig. 8.1). Ciò crea una connessione organica continua, priva di giunzioni tra l’osso e il tendine. I tendini lunghi localizzati lungo le ossa sono spesso tenuti in situ da bande di tessuto connettivo fissate allo scheletro che, assieme alle ossa adiacenti, delimitano dei canali tendinei. All’interno di questi, i tendini sono circondati da guaine tendinee formate da due strati di membrana sinoviale separati da una piccola quantità di fluido sinoviale (fig. 8.21), che consente ai tendini di muoversi senza esercitare un grande attrito sul tessuto adiacente. Le borse sono sacche di membrana sinoviale ripiene di liquido, frequenti nei punti in cui i muscoli o i tendini decorrono sopra le creste ossee o le articolazioni. Analogamente alle guaine tendinee, le borse riducono l’attrito tra il tendine e il tessuto sottostante.
Dal muscolo alla carne Nei mammiferi e negli uccelli il tessuto muscolare contiene circa il 20% di proteine, l’1-5% di grassi e il 75% di acqua. La composizione aminoacidica delle proteine muscolari rende la carne una fonte proteica adatta agli animali non erbivori, come l’uomo. Poiché contiene una quantità elevata dell’aminoacido essenziale lisi-
na, la carne è un buon supplemento alle diete a base di cereali, che ne sono carenti. Quando con la macellazione l’animale viene dissanguato, il metabolismo aerobico nelle cellule muscolari si interrompe, ma la produzione di ATP continua per alcune ore grazie alla degradazione anaerobica del glicogeno ad acido lattico. Fino a quando è presente una quantità di ATP sufficiente a staccare i ponti trasversali tra actina e miosina, il muscolo può essere stirato e la carne dell’animale appena macellato risulta tenera. Dopo 8-15 ore, tuttavia, insorge il rigor mortis e la carne si indurisce. a1 a
Tessuto connettivo Guaina tendinea Tendine
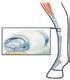
Lume con liquido sinoviale
Periostio Osso

Traduzione di Antonia Zarrilli
2 Materia, massa, forza
2 Lavoro ed energia
3 Atomi e particelle elementari
5 Elettricità
6 Legami chimici
9 Mole e concentrazione
9 Diffusione
11 Reazioni chimiche
13 Acqua
17 Acidi, basi e scala del pH
18 Chimica organica
20 Molecole organiche nell’organismo
32 ATP ed energia - Metabolismo cellulare
Gli organismi viventi contengono un grande numero di composti chimici. Tra questi, molti sono piccole e semplici molecole, come la componente predominante di tutti gli organismi viventi, l’acqua. Altre molecole sono grandi e complesse e possono aggregarsi a formare strutture ancora più complesse, per sostenere le quali è necessario un continuo rifornimento di energia; inoltre, gli organismi crescono e si riproducono, con ulteriore necessità di energia. La più semplice cellula vivente è più complessa di ogni sistema inanimato presente in natura. Tuttavia, le reazioni che avvengono in un organismo vivente seguono le stesse leggi fisiche e chimiche che si applicano alla parte inanimata; pertanto, per comprendere il funzionamento degli organismi è necessario conoscere le basi della chimica e della fisica. Questo capitolo offre una breve rassegna di aspetti rilevanti e basilari di chimica e fisica.
La materia ha massa e occupa spazio
La forza è la capacità di modificare la forma o la velocità di una massa
Le forze si presentano come coppie di forza azione-reazione
Le forze sono o di contatto o senza contatto
Il lavoro è una forza moltiplicata per la distanza
La potenza è lavoro per unità di tempo
L’energia è la capacità di svolgere un lavoro
L’energia può essere cinetica o potenziale
È definita materia qualsiasi cosa abbia massa e occupi spazio; l’unità di misura della massa è il chilogrammo (kg). In natura la maggior parte delle sostanze è costituita da composti chimici, che possono essere scissi in sostanze più semplici. I più semplici prodotti che possono essere ottenuti dalla degradazione di un composto chimico sono gli atomi; tutti gli atomi che hanno lo stesso numero di protoni costituiscono un elemento. La Terra contiene 92 elementi naturali.
Viene definita forza la capacità di modificare la forma di una massa o di cambiare la sua velocità, cioè di accelerarla. Un oggetto è accelerato in proporzione alla forza netta che agisce su di esso e in proporzione inversa alla sua massa:
Forza = massa · accelerazione
Se la velocità della massa diminuisce, l’accelerazione è negativa (decelerazione). L’unità di misura della forza è il newton (N). 1 N è la forza che determina su una massa di 1 kg un’accelerazione di 1 m/s2; la forza ha sia una grandezza che una direzione e quindi è una grandezza vettoriale.
Le forze sono distinte in forze di contatto e forze senza contatto. Le prime sono esercitate quando due oggetti entrano in contatto diretto l’uno con l’altro; le seconde possono agire a distanza, anche attraverso il vuoto senza materia dello spazio e includono la gravitazione e le forze elettriche e magnetiche.
Atomi e molecole attuano un moto termico
Il calore è l’energia cinetica interna di una massa
L’energia potenziale esprime sempre una differenza
Quando la forza viene usata per accelerare una massa su una data distanza, si compie un lavoro Questo lavoro è definito come la forza (misurata in newton) moltiplicata per la distanza (misurata in metri) e l’unità di misura è quindi il newton · metro (Nm), detto anche joule (J). La potenza è un’espressione di lavoro per unità di tempo; l’unità di misura della potenza è quindi J/s, normalmente chiamato watt (W).
L’energia di una massa è definita come la capacità di compiere un lavoro e l’unità di misura dell’energia è quindi identica a quella del lavoro (J). Un tempo l’unità comune dell’energia era la caloria (cal), laddove 1 cal corrisponde alla quantità di energia necessaria per aumentare di
1 °C la temperatura di 1 g di acqua (1 J = 0,230 cal).
Le due forme principali di energia sono: • energia cinetica, • energia potenziale.
Ogni massa in movimento possiede energia cinetica. Si prenda, per esempio, un martello che batte contro un chiodo: l’energia cinetica permette al martello di compiere un lavoro sul chiodo, che può essere forzato in un pezzo di legno quando la sua testa è colpita dal martello.
Nella materia gli atomi e le molecole sono continuamente in movimento, casuale e disorganizzato, che aumenta all’aumentare della temperatura. L’energia cinetica interna di una massa è chiamata energia termica e il movimento delle singole particelle è detto moto termico Quando una massa viene riscaldata, gli atomi e le molecole si muovono più rapidamente; ne consegue un aumento dell’energia cinetica interna, cioè dell’energia termica della massa.
L’energia potenziale è l’energia che una massa possiede come risultato della sua posizione. Essa esprime sempre una differenza, cioè la differenza di energia di una massa tra due diverse posizioni. Per esempio, quando una forza viene utilizzata per sollevare un vaso su una mensola, il vaso possiede energia potenziale a causa della sua posizione rispetto al pavimento; se il vaso cade dalla mensola, la forza gravitazionale converte l’energia potenziale in energia cinetica. Quando il vaso colpisce il pavimento, l’energia cinetica permette al vaso di compiere il lavoro richiesto per andare in frantumi.
Esistono molte forme di energia potenziale (immagazzinata). L’energia chimica è energia potenziale conseguente alle posizioni relative di atomi legati tra loro; l’energia contenuta nei nutrienti è energia potenziale di questo tipo (p.
1 Definisci materia, massa, elemento e composto chimico.
2 Qual è la relazione tra forza, massa e accelerazione?
3 Definisci lavoro, energia e potenza.
4 Quali sono le due principali forme di energia?
5 Fai un esempio di tipi diversi di energia per ognuna delle due principali forme.
32). L’energia elettrica è energia potenziale conseguente alla separazione di cariche elettriche opposte; infatti l’energia viene rilasciata allorquando le cariche elettriche opposte tornano insieme (p. 5).
Il campo della fisica che descrive le trasformazioni e le conversioni dell’energia è chiamato termodinamica. È utile considerare l’energia entro un sistema definito che può essere sia aperto che chiuso. Tutto ciò che si trova all’esterno del sistema osservato è definito ambiente. Un sistema aperto può scambiare energia con l’ambiente, mentre ciò non è possibile per un sistema chiuso; tuttavia, possono esserci scambi di energia all’interno di un sistema chiuso. In relazione alle condizioni studiate, possono esistere sistemi di energia di grandezze differenti: l’organismo animale è un esempio di sistema aperto che scambia energia con l’ambiente. Un altro esempio è la Terra, che riceve energia dall’ambiente, in particolare dal sole, e la trasmette all’ambiente (spazio) sotto forma di radiazioni elettromagnetiche (luce e radiazione termica). L’universo è il più vasto sistema chiuso che possiamo immaginare. Poiché non esiste alcun ambiente esterno all’universo, ogni possibile scambio di energia deve avvenire all’interno del sistema. La prima legge della termodinamica afferma che la quantità totale di energia in un sistema chiuso è costante: ciò significa che l’energia non può essere creata né distrutta ma può solo essere trasferita da un posto a un altro o convertita da una forma a un’altra.
Nelle piante verdi, per esempio, l’energia luminosa (una forma di energia cinetica) è convertita in energia chimica (energia potenziale) immagazzinata nelle molecole organiche. L’energia chimica che noi otteniamo dal cibo può essere trasferita a nuove molecole sintetizzate nel corpo o può essere trasformata in altre forme di energia. Nei muscoli scheletrici, per esempio, l’energia chimica viene convertita in energia cinetica quando viene compiuto un lavoro esterno. Come si è visto, la quantità totale di energia, prima e dopo i vari trasferimenti della stessa, rimane sempre uguale, ma durante ogni trasferimento o conversione almeno una parte viene degradata sotto forma di energia termica. Quando un chiodo viene conficcato con un martello, tutta l’energia cinetica del martello è alla fine
convertita in energia termica; un grosso chiodo conficcato in un pezzo di legno duro può diventare così caldo da determinare dolore al tatto. Le varie forme di energia non sono ugualmente adatte a compiere un lavoro: l’energia termica può essere utilizzata per compiere un lavoro solo quando viene trasferita da un posto a un altro e questo richiede una differenza di temperatura. Nei sistemi che mantengono temperature costanti, come il corpo, l’energia termica non può essere utilizzata per compiere lavoro. La frazione massima di energia che può essere utilizzata per ottenere lavoro in condizioni ottimali è detta energia libera. La seconda legge della termodinamica afferma che in un sistema chiuso tutte le conversioni di energia portano a una riduzione della quantità di energia libera. In pratica, questo significa che una sempre crescente proporzione dell’energia totale esisterà come energia termica e che in un sistema chiuso le differenze di temperatura tenderanno in modo graduale ad annullarsi.
In un organismo la quantità di energia libera può essere mantenuta costante o anche incrementata nonostante conversioni di energia continue; tuttavia questo non è in contrasto con la seconda legge della termodinamica. Come abbiamo già visto, un organismo è un sistema aperto in comunicazione con l’ambiente; ogni incremento dell’energia libera dell’organismo è sempre contemporaneo a una diminuzione dell’energia libera dell’ambiente.
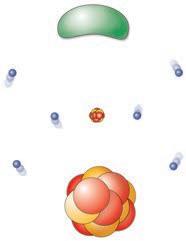
Gli atomi consistono di varie particelle elementari; di queste le più importanti sono (fig. 1.1):
• i protoni, che hanno carica elettrica positiva;
• i neutroni, che non hanno carica elettrica;
• gli elettroni, che hanno carica elettrica negativa.
Le cariche elettriche opposte di un elettrone e di un protone hanno eguale grandezza e ognuna viene definita carica elementare. I protoni e i neutroni sono contenuti nel nucleo, mentre gli elettroni si muovono a elevata velocità in uno spazio limitato intorno al nucleo. Le particelle con cariche opposte si attraggono, mentre particelle con lo stesso tipo di carica si respingono
In un sistema chiuso la quantità di energia è costante
Nelle conversioni di energia si perde sempre una quota di energia libera
6 Definisci un sistema di energia aperto e uno chiuso.
7 Qual è la prima legge della termodinamica?
8 Qual è la seconda legge della termodinamica?
9 Fai un esempio di conversione dell’energia in una cellula.
Gli atomi sono costituiti da protoni, neutroni ed elettroni
Protoni ed elettroni hanno cariche elettriche opposte di eguale grandezza Cariche elettriche opposte si attraggono; cariche elettriche simili si respingono
Figura 1.1 Le più importanti particelle elementari presenti in un atomo. Gli elettroni hanno una carica elettrica negativa, i protoni hanno una carica positiva e i neutroni non hanno carica. Gli elettroni si muovono intorno al nucleo dell’atomo, che contiene le altre particelle elementari.
Materia Atomo
Elettrone
Nucleo atomico
Figura 1.2 Forze elettriche tra particelle cariche elettricamente. Cariche opposte si attraggono, mentre cariche simili si respingono.
Atomi con lo stesso numero di protoni costituiscono un elemento
Neutrone
(fig. 1.2); le forze di attrazione tra gli elettroni negativi e i protoni positivi evitano che gli elettroni possano fuggire dall’atomo.
La massa di un protone è pressoché identica alla massa di un neutrone ed è circa 2000 volte più grande della massa di un elettrone. Quindi, l’intera massa di un atomo è concentrata nel nucleo e la massa di un elettrone è spesso trascurabile nel calcolo della massa atomica. Poiché anche le masse dei protoni e dei neutroni sono molto piccole, solo 1,7 × 10–27 kg, l’utilizzazione del chilogrammo come unità per confrontare le masse di atomi e molecole non è pratica. Pertanto è stata introdotta un’unità di misura definita unità di massa atomica, detta anche dalton: una massa di 1 dalton equivale alla massa di un protone, che ha la stessa massa di un neutrone. Il numero totale di protoni e neutroni presenti nel nucleo di un atomo è detto numero di massa
Nelle reazioni chimiche tra atomi sono solo gli elettroni a interagire (p. 6). In un atomo elettricamente neutro il numero dei protoni è identico al numero degli elettroni; atomi con lo stesso numero di protoni, quindi, hanno le stesse proprietà chimiche e sono definiti come appartenenti allo stesso elemento
La Terra contiene 92 elementi naturali, ognuno dei quali è indicato con un simbolo accettato in ambito internazionale; tra questi, 25 sono essenziali per la vita. I quattro elementi ossigeno,
Tabella 1.1 Elementi nell’organismo ElementoSimbolo
Iodio I53
Rame Cu29
Zinco Zn30
ManganeseMn25
CobaltoCo27
Cromo Cr24
Selenio Se34
MolibdenoMo42
Fluoro F9
Stagno Sn50
Silicio Si14
VanadioV23
* Ogni oligoelemento costituisce meno dello 0,01% della massa corporea.
carbonio, idrogeno e azoto rappresentano più del 96% del peso corporeo di un organismo (tab. 1.1).
Tra gli elementi essenziali per la vita ci sono anche molti metalli: per esempio, nei vertebrati il ferro è necessario per permettere al sangue di trasportare ossigeno (p. 306). Elementi essenziali per la vita che costituiscono meno dello 0,01% della massa corporea sono detti oligoelementi La maggior parte degli elementi esiste in più di una variante, poiché il numero di neutroni
Traduzione di Barbara Barboni
612 Aspetti principali del metabolismo energetico
612 Fa se di assorbimento e di postassorbimento
625 Regolazione del metabolismo dei nutrienti organici
Le cellule possono essere considerate come centrali che producono energia tramite l’ossidazione continua di nutrienti organici; l’energia prodotta viene utilizzata dalle cellule per la crescita, la proliferazione e le loro specifiche funzioni. L’energia complessiva che ciascun organismo utilizza proviene, in definitiva, dai nutrienti organici introdotti con la dieta. I carboidrati e i lipidi sono utilizzati principalmente come substrati energetici, mentre gli aminoacidi, assorbiti dopo la digestione delle proteine alimentari, possono servire sia come precursori per la sintesi di nuove proteine, strutturali o enzimatiche, sia come substrati energetici. Tuttavia, il supporto energetico negli animali superiori può essere garantito anche in assenza di un’assunzione continua di nutrienti, in quanto essi hanno sviluppato la capacità di immagazzinare molecole a elevato contenuto energetico durante i periodi di abbondante disponibilità di alimento con la possibilità poi di attingervi quando le condizioni metaboliche interne e l’assenza di nuovo alimento lo rendono necessario.
Nei monogastrici i periodi di accumulo di energia si alternano a fasi di mobilizzazione di substrati energetici
Nei ruminanti la digestione è un processo quasi continuo L’eccesso di energia viene immagazzinato sotto forma di lipidi e di glicogeno
Il fegato svolge un ruolo chiave nel metabolismo e nella conversione dei nutrienti
Gli aminoacidi possono essere notevolmente modificati a livello epatico dopo l’assorbimento
Gli aminoacidi essenziali non possono essere sintetizzati dall’organismo
Nei ruminanti la composizione aminoacidica della dieta è ininfluente
1 Come viene immagazzinata nell’organismo l’energia in eccesso?
2 Qual è organo più importante per la conversione delle molecole organiche?
3 Spiega l’importanza della composizione aminoacidica delle proteine alimentari nei ruminanti e nei monogastrici.
L’energia viene utilizzata continuamente dalle cellule per le loro funzioni; al contrario, essa viene introdotta nell’organismo sotto forma di molecole organiche unicamente dopo la digestione e l’assorbimento dei nutrienti assunti durante il pasto. In un carnivoro selvatico tra un pasto e l’altro possono intercorrere molte ore o addirittura giorni. Un erbivoro, invece, trascorre normalmente la maggior parte della sua giornata assumendo cibo. Per esempio, i cavalli al pascolo occupano mediamente 10-12 ore della loro giornata ad alimentarsi, in sessioni di 0,5-3 ore ciascuna. Gli onnivori, come i maiali, mangiano 3-5 volte al giorno, con sessioni alimentari della durata di 10-15 minuti ciascuna. Naturalmente il comportamento alimentare della maggior parte degli animali è stato profondamente modificato dall’uomo con l’addomesticamento. Nei carnivori e negli onnivori la digestione e l’assorbimento di un pasto richiedono, in genere, circa quattro ore. Le cellule di questi animali, quindi, ricevono dall’intestino i nutrienti solo per circa 12 ore, se l’animale riesce ad alimentarsi tre volte al giorno; durante le restanti 12 ore, lo stomaco e l’intestino tenue sono essenzialmente vuoti e inattivi. Per questo, per mantenere l’omeostasi energetica, l’organismo deve essere in grado di immagazzinare, durante la fase digestiva, substrati energetici che successivamente potranno essere mobilizzati nelle fasi di mancata assunzione di alimento. Alternando periodi di stoccaggio a fasi di mobilizzazione dei nutrienti vengono evitate le fluttuazioni ematiche del glucosio e di altri composti energetici. Nel presente capitolo sono trattati in dettaglio i diversi quadri metabolici che si osservano nei carnivori e negli onnivori durante la fase di assorbimento dei nutrienti, così come durante il periodo di postassorbimento. In particolare, verrà trattata la regolazione dei flussi delle sostanze nutritive durante queste due fasi metaboliche e il ruolo di controllo svolto dal sistema endocrino e nervoso. Negli erbivori, e in particolare nei ruminanti, la digestione, invece, è un processo continuo. In questi animali non si può quindi fare una chiara distinzione fra fase assorbitiva e fase postassorbitiva.
Durante la digestione grandi molecole organiche vengono degradate a piccole molecole che possono essere assorbite dall’intestino; quando
queste molecole entrano nelle cellule, una loro frazione variabile viene ulteriormente degradata e, infine, ossidata a CO2 e H2O (p. 37). L’energia liberata durante i processi ossidativi viene trasferita all’ATP, che viene poi utilizzato dalle stesse cellule (p. 32). L’energia presente nell’ATP può essere utilizzata, per esempio, per far contrarre le cellule muscolari, per il trasporto attivo di sostanze attraverso la membrane cellulari o per la sintesi di molecole complesse. Le cellule, inoltre, possono utilizzare alcune delle molecole organiche assorbite a livello intestinale come elementi base per la sintesi di loro componenti stutturali; fra queste componenti molecolari si ricordano gli enzimi, gli ormoni, gli anticorpi, così come molte altre molecole con funzioni specializzate.
Quando le sostanze nutritive assorbite dall’intestino non sono totalmente utilizzate, l’eccedenza può essere stoccata sotto forma di molecole di deposito. La maggior parte delle riserve energetiche dell’organismo è costituita dai trigliceridi immagazzinati nel tessuto adiposo, che contiene una grande quantità di energia se espressa per unità di massa.
Ogni tipologia di nutriente può essere generalmente convertita in un’altra attraverso specifiche vie metaboliche. Per questa ragione la quantità di carboidrati, proteine e grassi che assumiamo con la dieta può variare anche considerevolmente senza conseguenze. Per esempio, gli aminoacidi possono essere convertiti in glucosio e in acidi grassi; alcuni aminoacidi, quelli non essenziali, possono essere utilizzati per trasferire gruppi aminici alle molecole dei carboidrati. Le cellule epatiche svolgono un ruolo chiave nella regolazione del metabolismo dell’organismo e nella conversione dei nutrienti; tuttavia, alcuni tipi di aminoacidi sono necessari all’organismo in quanto le cellule epatiche non sono in grado di produrli ex novo. Questi sono noti come aminoacidi essenziali e vengono introdotti nell’organismo esclusivamente tramite la dieta; pertanto, la composizione aminoacidica delle proteine presenti nell’alimento è di vitale importanza per molte specie di mammifero. Nei ruminanti, invece, la maggior parte degli aminoacidi assorbiti a livello intestinale proviene dalla digestione delle proteine microbiche sintetizzate nei prestomaci (p. 566). La composizione di queste proteine è poco influenzata dalla composizione aminoacidica delle proteine presenti nella dieta, ma dipende quasi totalmente dall’attività delle cellule batteriche; pertanto, nei ruminanti la
Stoccaggio e sintesi di proteine
Tratto gastrointestinale
Aminoacidi Glucosio
Aminoacidi
UreaChetoacidi
Fegato
Proteine
Glucosio
EnergiaAcidi grassi Glicogeno
Muscolo scheletrico
TrigliceridiLipoproteine
Trigliceridi
Tessuto adiposo
Tutti i tessuti
GlucosioEnergia
Metabolismo ossidativo
composizione aminoacidica della dieta è sostanzialmente ininfluente.
Oltre alle proteine, l’alimento deve contenere una quantità sufficiente di vitamine, minerali e acidi grassi essenziali. Per informazioni sulle funzioni di questi nutrienti, si rimanda il lettore a testi specifici di nutrizione animale.
Nei carnivori, negli onnivori e, in misura minore, negli erbivori il metabolismo dell’organismo alterna due diverse fasi:
1. di assorbimento, 2. di postassorbimento.
Durante la fase di assorbimento l’organismo attinge l’energia dall’ossidazione dei nutrienti che entrano nel flusso ematico dopo la digestione e l’assorbimento intestinale.
Durante la fase postassorbitiva l’organismo attinge la propria energia mobilizzando dai tessuti di deposito del corpo i substrati energetici.
In caso di mancanza di cibo per tempi prolungati, si assiste a una graduale transizione dalla fase di postassorbimento alla fase di digiuno.
Figura 16.1 Principali vie metaboliche nella fase di assorbimento. Aminoacidi, glucosio, trigliceridi e acidi grassi volatili (AGV) vengono trasportati dal tratto gastrointestinale ai vari tessuti, dove le molecole vengono elaborate. Oltre al glucosio, piccole quantità di acidi grassi liberi (FFA) sono ossidati per finalità energetiche (non mostrato).
I carboidrati sono principalmente assorbiti sotto forma di glucosio I monosaccaridi e gli aminoacidi sono trasportati direttamente dal tratto digestivo al fegato
Il sangue proveniente dal tratto digestivo passa attraverso due reti capillari
I lipidi vengono rimossi dal tratto digestivo tramite il circolo linfatico
Nei ruminanti la maggior parte dei carboidrati ingeriti viene convertita in acidi grassi volatili
Il glucosio è l’elemento base delle molecole complesse di amido, glicogeno e cellulosa
Il metabolismo dei carboidrati nei non erbivori è costituito principalmente dal metabolismo del glucosio
Il fegato processa la maggior parte del glucosio che riceve dall’intestino
Nel fegato il glucosio viene stoccato come glicogeno o convertito in lipidi
di assorbimento
Nella fase di assorbimento gli aminoacidi, i monosaccaridi e i trigliceridi assunti con la dieta vengono trasferiti, dopo l’assorbimento dal tratto digestivo, ai vari tessuti (fig. 16.1) attraverso il circolo ematico. La fase di assorbimento è un periodo anabolico durante il quale i nutrienti vengono messi a disposizione dell’organismo e utilizzati per finalità energetiche, per sintetizzare nuove molecole e per stoccare substrati energetici sotto forma di molecole di deposito. Nei carnivori e negli onnivori i carboidrati sono principalmente assorbiti sotto forma di glucosio, mentre le proteine, in tutti i mammiferi, vengono assorbite sotto forma di aminoacidi. Il glucosio e gli aminoacidi vengono trasportati dal lume intestinale, in primo luogo, ai capillari intestinali e poi, tramite la vena porta, raggiungono i capillari epatici; il sangue del tratto gastrointestinale passa quindi attraverso due reti capillari (fig. 15.43). La circolazione dall’intestino ai capillari epatici consente al fegato di elaborare tutti i nutrienti assorbiti e di regolarne la concentrazione e la composizione prima che il sangue raggiunga il circolo sistemico. A differenza del glucosio e degli aminoacidi, i lipidi vengono rimossi dall’intestino tramite la linfa; i dotti linfatici sfociano in una grande vena posta nella parte craniale del torace in prossimità del cuore (vena cava craniale). I capillari polmonari costituiscono pertanto la prima rete capillare attraverso cui passano i lipidi alimentari; ne consegue che il fegato non ha la possibilità di elaborare i lipidi assorbiti prima che questi vengano distribuiti a tutto il corpo.
Glucosio
4 Definisci le fasi di assorbimento e di postassorbimento.
5 In quale forma i carboidrati della dieta vengono assorbiti dal tratto digerente?
6 In quale forma vengono assorbite dal tratto digerente le proteine?
La dieta di solito contiene sia carboidrati di piccole dimensioni, come il glucosio, il lattosio e il saccarosio, che molecole complesse (polisaccaridi), quali l’amido, il glicogeno e la cellulosa. Negli animali non erbivori i polisaccaridi α-glicosidici come l’amido e il glicogeno vengono degradati, al termine della digestione enzimatica, a monosaccaridi (p. 595). Il glucosio è il principale monosaccaride assorbito dall’intestino; esso è elemento costitutivo dei due più importanti carboidrati alimentari, l’amido e il glicogeno. Tuttavia, a livello intestinale possono essere assorbite anche piccole quantità di galattosio e fruttosio (p. 596). Nel fegato il galattosio e la maggior parte del fruttosio vengono
convertiti in glucosio. Negli erbivori il metabolismo cellulare dei carboidrati non coinvolge, al contrario, in via prioritaria il metabolismo del glucosio: negli erbivori, infatti, la cellulosa è il principale carboidrato alimentare. Nei ruminanti la maggior parte dei carboidrati, fra cui la cellulosa, viene convertita in acidi grassi volatili (AGV) dagli enzimi microbici che operano nei prestomaci (p. 559). Negli erbivori monogastrici una simile degradazione della cellulosa ha luogo nell’intestino crasso, mentre la digestione dell’amido e dei piccoli carboidrati è sovrapponibile a quella degli animali non erbivori.
Il fegato elabora la maggior parte del glucosio che riceve attraverso la vena porta; nel fegato il glucosio viene in parte immagazzinato come glicogeno e in parte convertito in lipidi (vedi oltre). Il glucosio viene, inoltre, trasportato nelle cellule muscolari, dove viene stoccato sotto forma di glicogeno. Di conseguenza, anche se negli animali non erbivori durante la fase di assorbimento vengono assorbite notevoli quantità di glucosio, la sua concentrazione non aumenta mai in modo consistente nel circolo sistemico. Per questo motivo la concentrazione plasmatica di glucosio nell’uomo aumenta solo del 20% durante la fase di assorbimento raggiungendo livelli da 4 a 5 mmol/L dopo un pasto ricco di carboidrati. La concentrazione di glucosio riscontrata nel circolo portale è, invece, considerevolmente più elevata. Un altro momento cruciale dell’omeostasi del glucosio è legato alla sintesi e alla demolizione del glicogeno; il glucosio viene inoltre assorbito dalle cellule del tessu-
Le malattie da accumulo di glicogeno sono caratterizzate da un deposito patologico della molecola glucidica a causa di carenze ereditarie degli enzimi deputati al normale metabolismo del glicogeno. Oltre al fegato, possono essere coinvolti anche altri tessuti, quali il sistema nervoso centrale, i reni, il cuore e il muscolo scheletrico. Nell’uomo sono stati riportati ben otto diversi tipi di malattie ereditarie da alterato accumulo di glicogeno, alcuni dei quali sono stati anche riscontrati negli animali. Il tipo I, che è stato diagnosticato nel cane, riconosce un deficit epatico di glucosio-6-fosfatasi; in mancanza di questo enzima, il fegato perde la capacità di formare glucosio libero con conseguente quadro di ipoglicemia (fig. 16.11).
Intestino
Sintesi di lipoproteine (VLDL+HDL)
Trigliceridi
Fosfolipidi
Aminoacidi
Colesterolo
Fegato
Degradazione
Endocitosi mediata da recettori
LPL (lipoproteina lipasi)
Chilomicroni VLDL
Capillari sanguigni HDL
Chilomicroni residui
VLDL (IDL) residui
HDL ricche di colesterolo
LDL
Eccesso di colesterolo nella bile
Cistifellea
Tutte le cellule
Colesterolo
Recettori LDL
Adipociti

FFA
Sintesi di trigliceridi
FFA (acidi grassi liberi)
Ossidazione a CO2 + H2O FFA
Cellule muscolari

Figura 16.2 Presentazione semplificata del metabolismo delle lipoproteine durante la fase di assorbimento. I chilomicroni assorbiti dall’intestino e le lipoproteine a densità molto bassa (VLDL) prodotte nel fegato forniscono acidi grassi liberi (FFA) depositati come trigliceridi nel tessuto adiposo e, in certa misura, ossidati a livello cellulare. La lipoproteina lipasi (LPL) presente nella parete dei capillari catalizza il rilascio di FFA dai trigliceridi. Le apolipoproteine sintetizzate nel fegato sono trasferite alle VLDL e ai chilomicroni dalle lipoproteine ad alta densità (HDL). Questo trasferimento di apolipoproteine (non mostrato) facilita l’assorbimento dei lipidi attivando la lipoproteina lipasi. I chilomicroni e alcune VLDL residue vengono assunti dalle cellule epatiche e ulteriormente metabolizzati; le lipoproteine a bassa densità (LDL) sono formate a partire dalle VLDL. Il colesterolo è trasportato a diverse tipologie cellulari dalle LDL.
to adiposo, dove viene convertito in trigliceridi. Il glucosio, oltre a essere convertito in lipidi e conservato sotto forma di glicogeno, è la fonte energetica principale nella fase di assorbimento dei monogastrici (fig. 16.1).
La captazione del glucosio da parte delle cellule epatiche aumenta in modo proporzionale alla sua concentrazione nel circolo portale; di conseguenza, dopo un pasto ricco di carboidrati, la captazione epatica di glucosio aumenta notevolmente. Come è noto, gli epatociti formano il glicogeno che, analogamente all’amido, è un polisaccaride composto da molecole di glucosio le-
gate fra loro da legami α-glicosidici (fig. 15.22a).
La sintesi di glicogeno epatico continua finché la molecola stoccata non raggiunge livelli pari a circa il 5% della massa dell’organo. Se l’apporto alimentare di glucosio eccede la capacità del fegato di accumulare glicogeno, il glucosio viene convertito in trigliceridi. Nei ruminanti i livelli di glucosio rielaborati dal fegato sono minori. Parte dei trigliceridi sintetizzati nel fegato a partire dal glucosio viene stoccata all’interno dell’organo stesso (fig. 16.1); la maggior parte, tuttavia, viene riversata nel sangue come aggregati di trigliceridi, fosfolipidi, molecole di
Il glucosio viene captato dalle cellule del tessuto adiposo e convertito in trigliceridi
Anche il muscolo accumula il glucosio sotto forma di glicogeno
Dopo un pasto ricco di carboidrati, la formazione di glicogeno nel fegato continua fino a un accumulo pari a circa il 5% della sua massa