Ecuador First Class Shrimp: la marca creada por la CNA y que lleva el camarón ecuatoriano al escenario mundial
Nuevas autoridades lideran el aparato productivo del Ecuador con enfoque técnico y estratégico
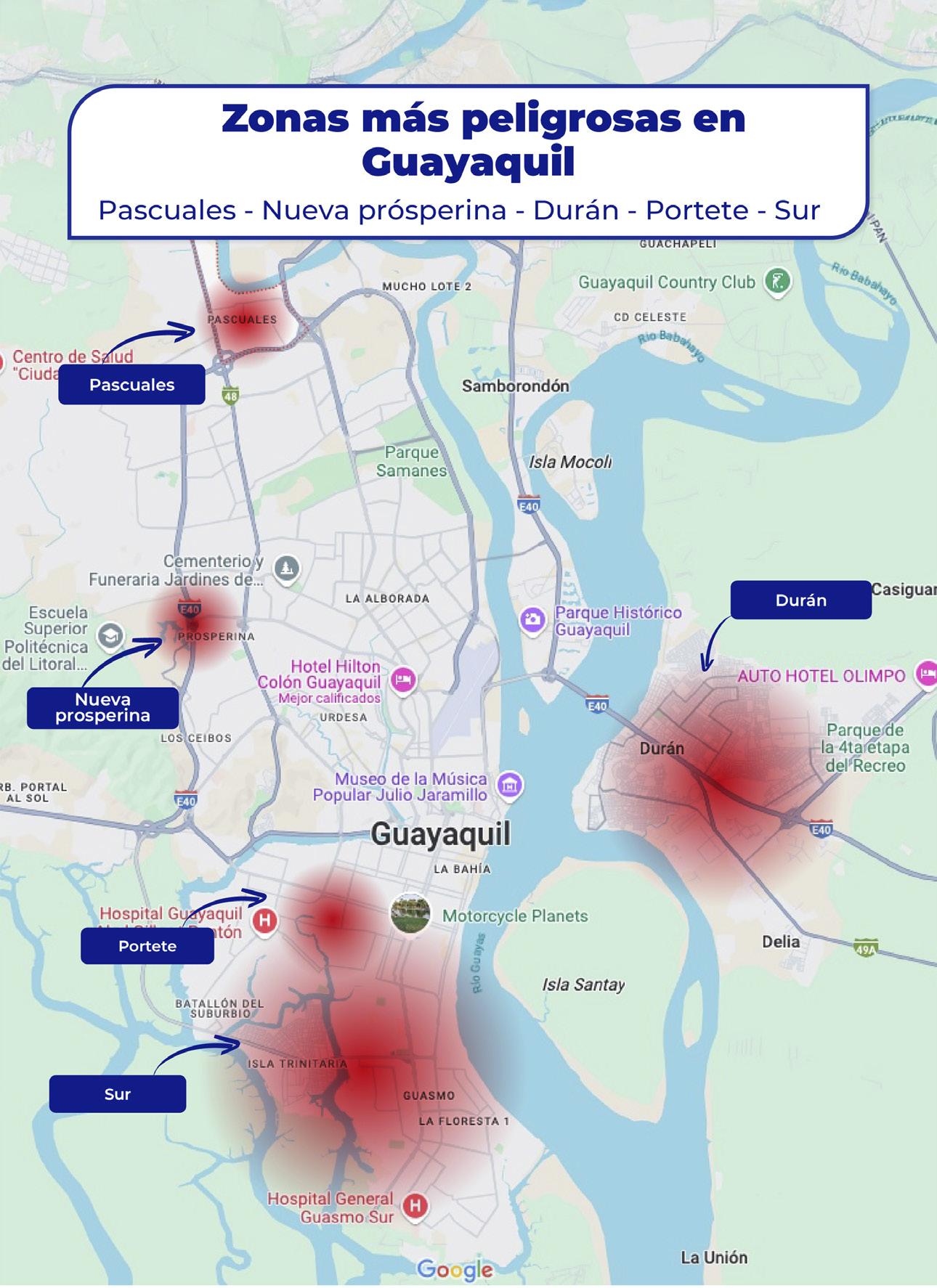
Golpe a la economía criminal: estrategia en marcha
Finanzas que cultivan sostenibilidad
Las pequeñas granjas camaroneras: recomendaciones para ser más eficientes y sostenibles
Aliados invisibles: Cómo los microorganismos transforman la productividad en acuacultura
Evaluación de la diversidad bacteriana como indicador de calidad en suelos de cultivos acuícolas y biorremediación para la mejora productiva
H2 para eficiencia térmica y reducción de emisiones en calderas y MCI
Adición de suplementos promotores de salud en microdietas para postlarvas de camarón blanco (Penaeus vannamei): efectos sobre el crecimiento, la supervivencia y el estado de salud
La trazabilidad total ya es una realidad en el Ecuador
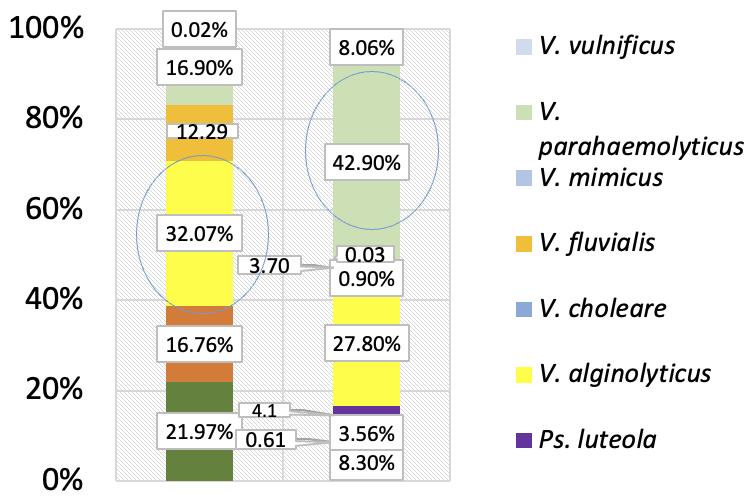
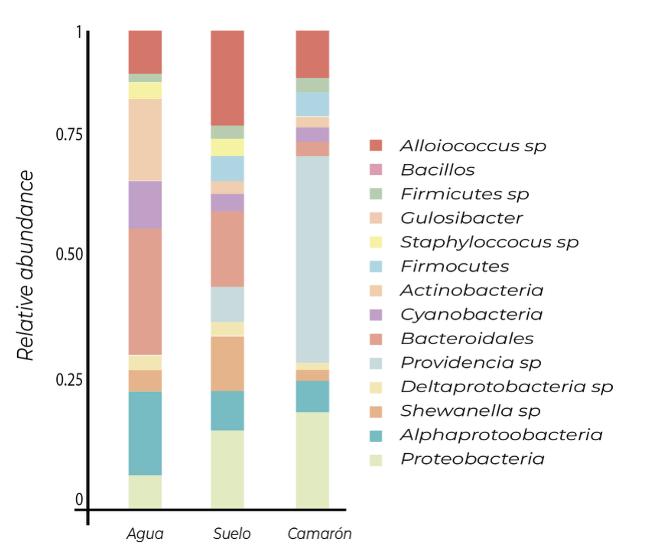
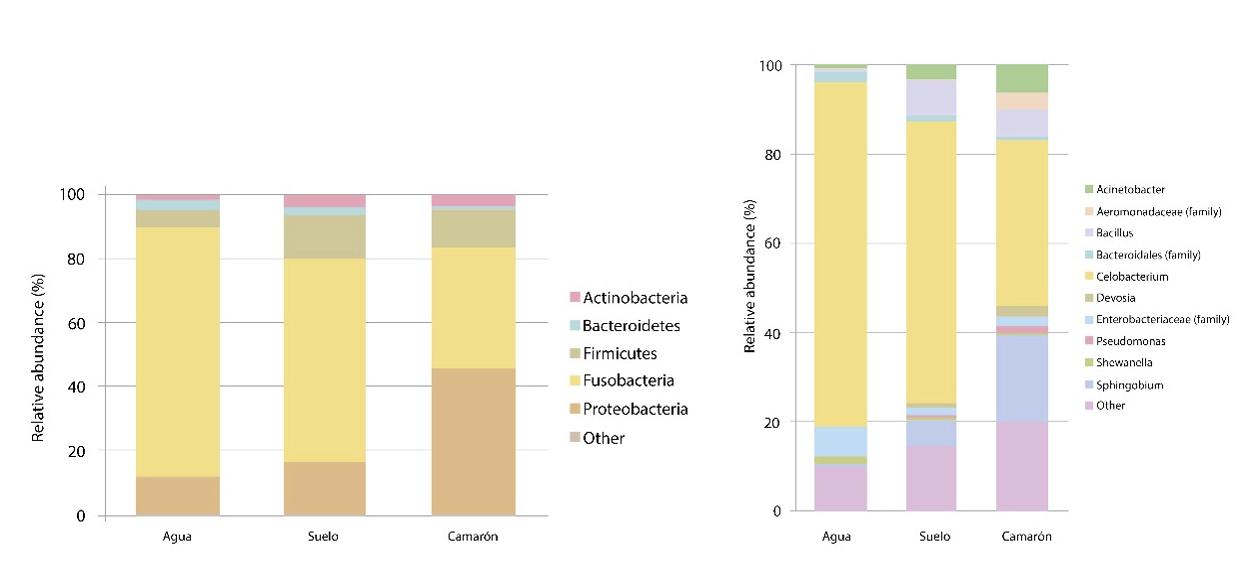
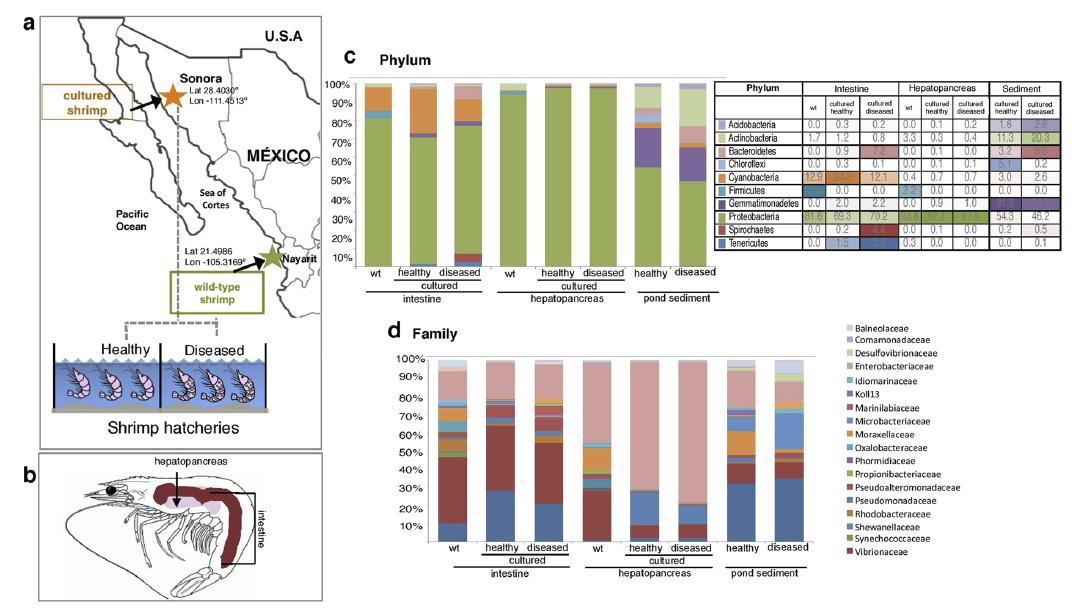
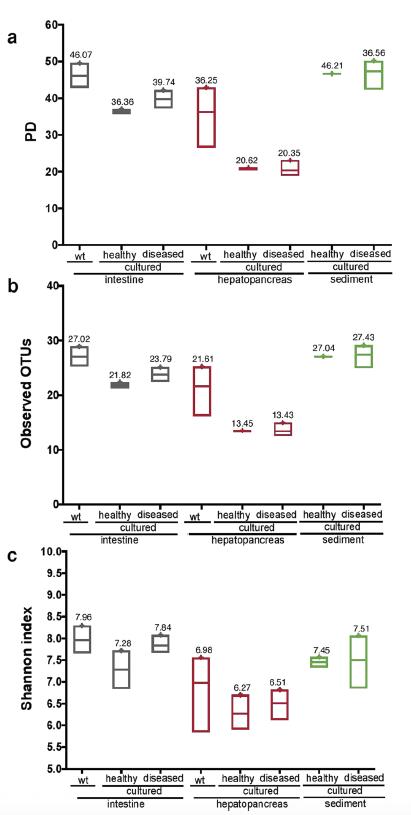
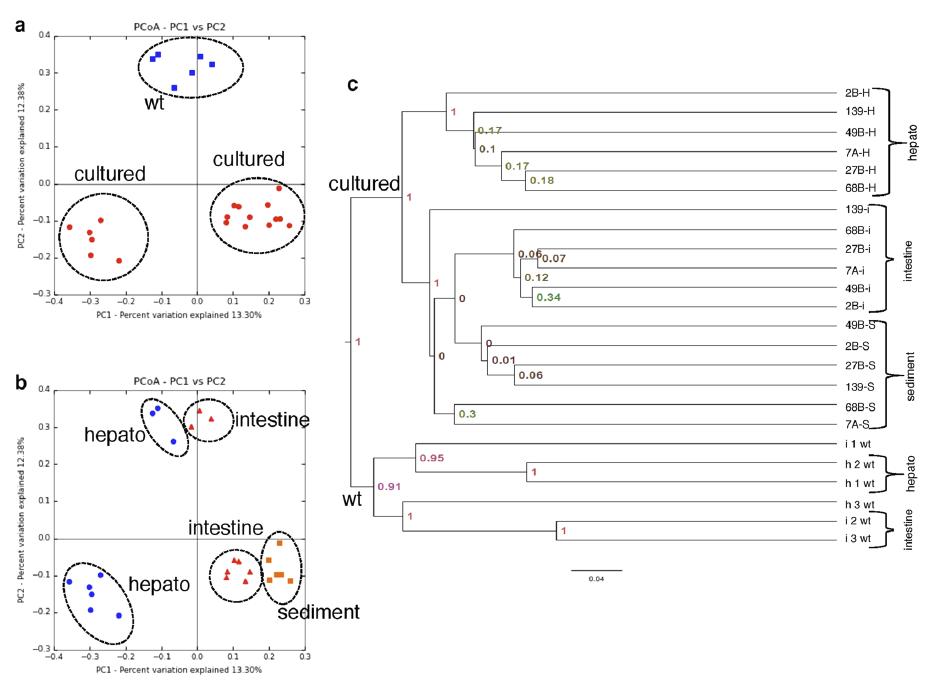
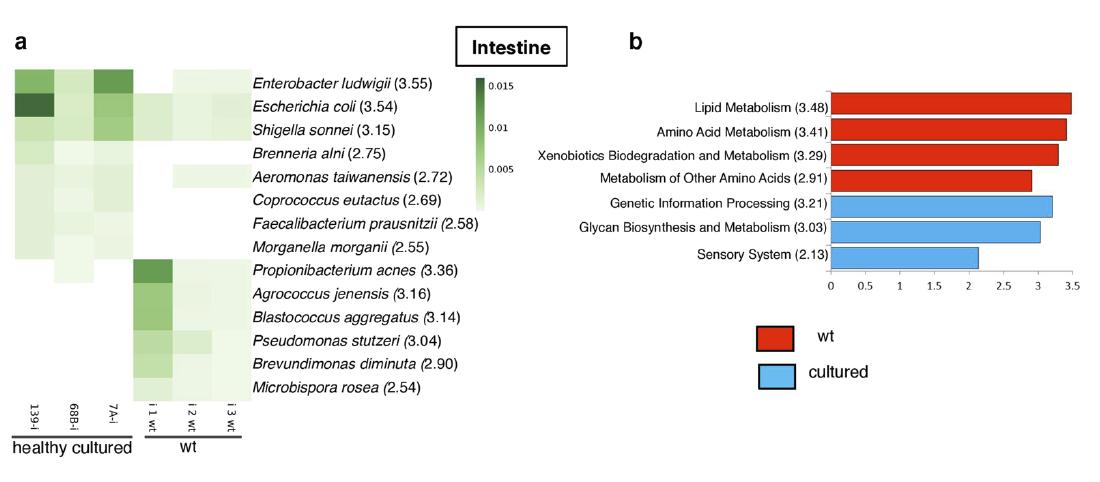
El microbioma del camarón blanco del Pacífico revela una composición diferencial de la comunidad bacteriana entre condiciones naturales, de cultivo y en brotes de AHPND/EMS
Presidente Ejecutivo
Ing. José Antonio Camposano
Editora “AquaCultura”
MSc. Shirley Suasnavas ssuasnavas@cna-ecuador.com
Consejo Editorial
MSc. Yahira Piedrahita
PhD. Leonardo Maridueña
Ing. José Antonio Lince Ing. Alex de Wind
Diseño y diagramación Ing. Orly Saltos osaltos@cna-ecuador.com
Ing. Roberto Peñafiel rpenafiel@cna-ecuador.com
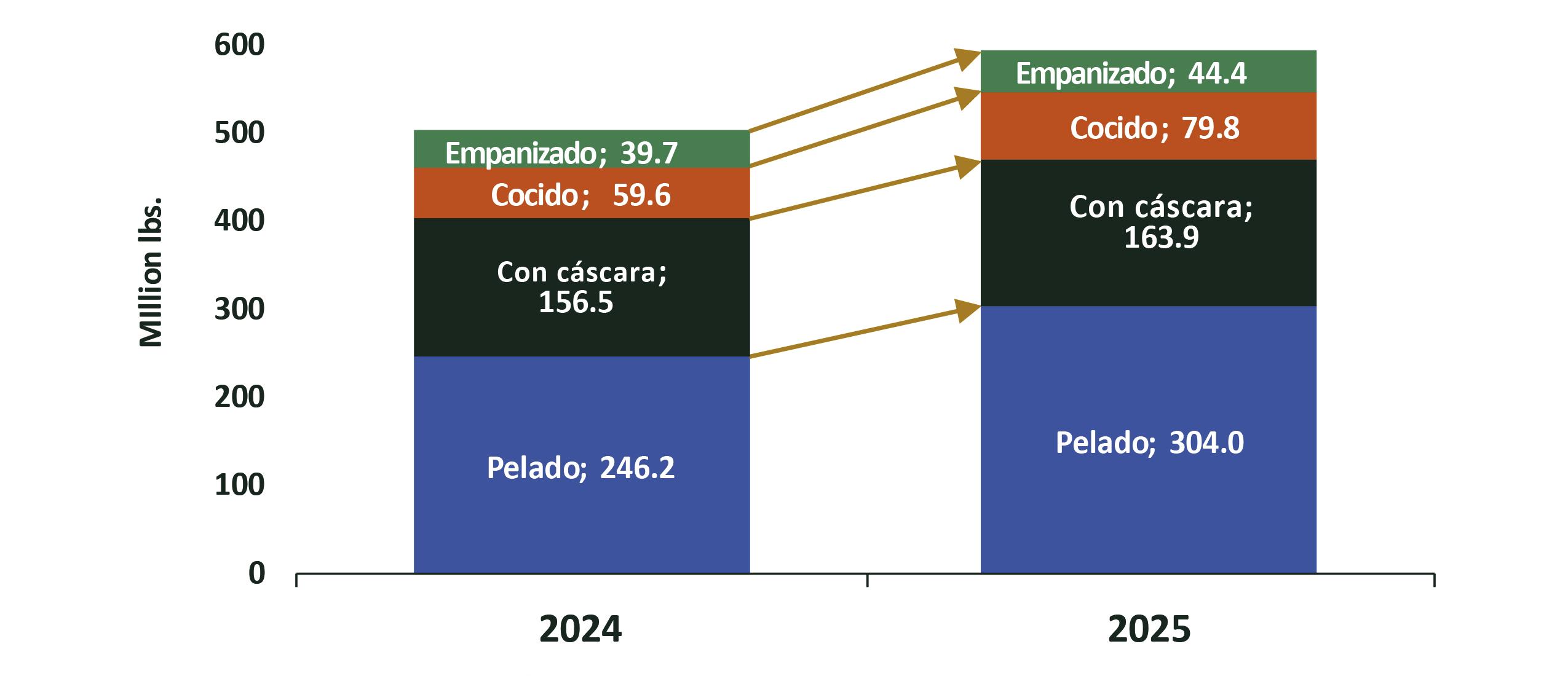
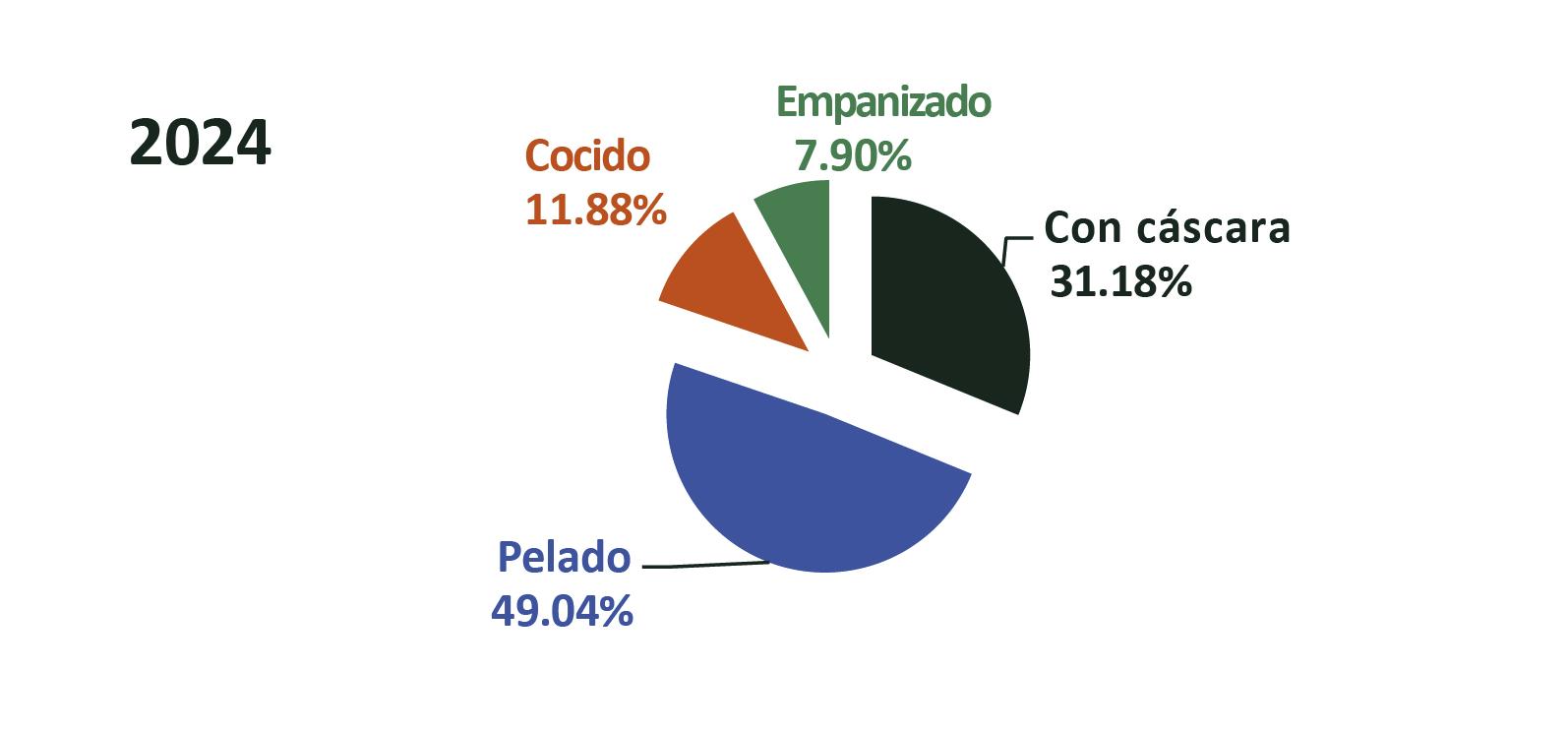
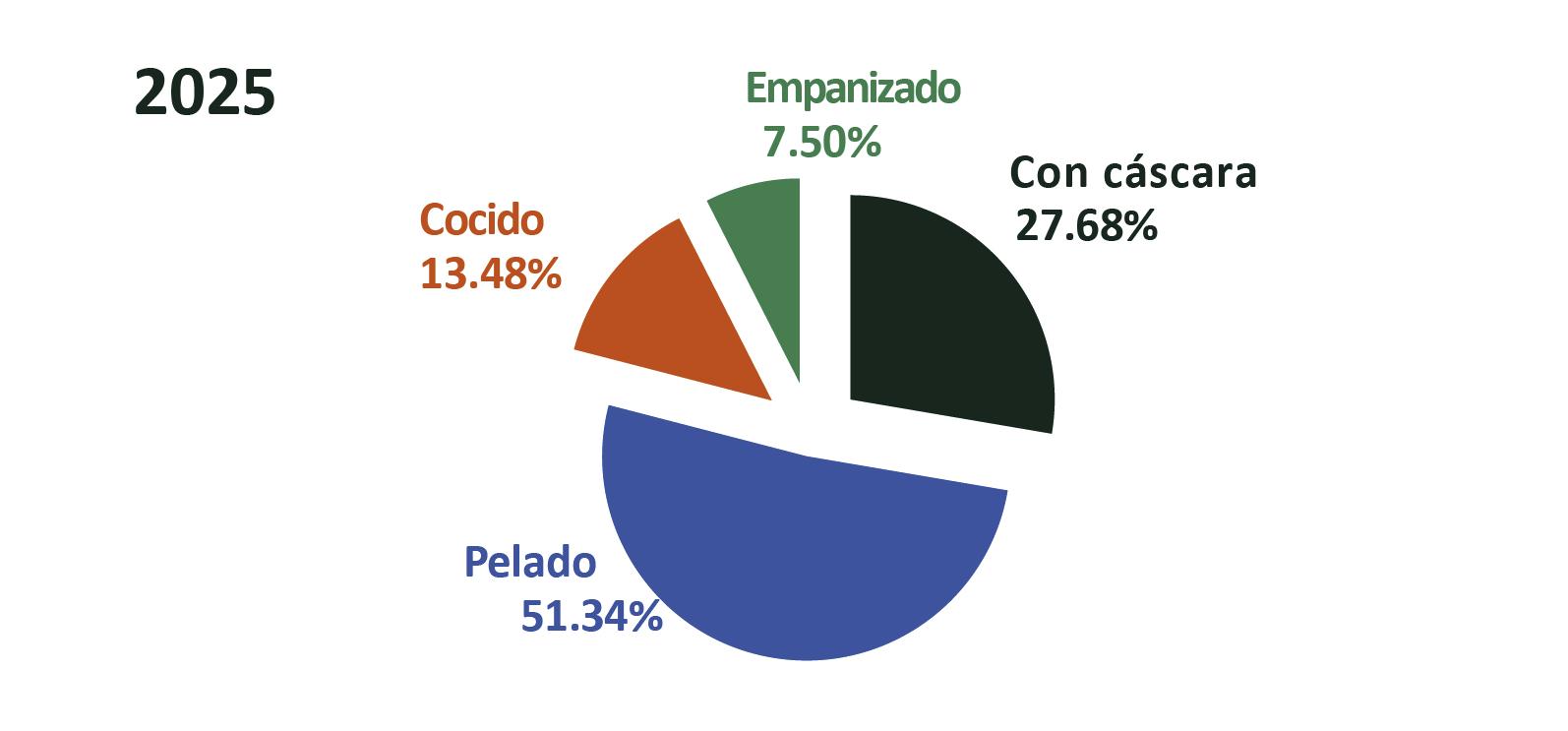
Exportaciones de camarón
Reporte de mercado de EE. UU.
Noticias del sector
Corrección de estilo MSc. Daniel Ampuero daniel.ampuero@gmail.com
Comercialización
MSc. Gabriela Nivelo gnivelo@cna-ecuador.com
EEDITORIAL
José Antonio Camposano Presidente Ejecutivo
Pragmatismo para un comercio sin trabas
Mantener relaciones funcionales y directas con cada destino, sin importar quién gobierne o qué tensiones existan en sus relaciones bilaterales, es una obligación que no podemos descuidar.
n un entorno global cada vez más complejo, la política comercial se ha convertido en un terreno marcado por tensiones geopolíticas, nuevas regulaciones y presiones internas en los países importadores. Para un sector como el nuestro, que ha demostrado su capacidad de competir y liderar con calidad, sostenibilidad y volumen, este escenario exige algo más que excelencia productiva: requiere visión estratégica y relaciones pragmáticas.
Pragmatismo significa anteponer los intereses del país y del sector exportador por sobre simpatías ideológicas o afinidades políticas. Significa entender que, en el comercio internacional, el diálogo constante, el cumplimiento riguroso de estándares y la capacidad de anticipar riesgos son herramientas tan importantes como el producto que ponemos en el mercado.
Las exportaciones ecuatorianas de camarón han crecido gracias a la apertura de mercados clave y al trabajo técnico con nuestros socios comerciales. Pero esa posición no es permanente ni está asegurada. Las decisiones tomadas por gobiernos extranjeros —a veces por motivos económicos, otras por razones políticas— pueden alterar
drásticamente las condiciones de acceso. Por eso, mantener relaciones funcionales y directas con cada destino, sin importar quién gobierne o qué tensiones existan en sus relaciones bilaterales, es una obligación que no podemos descuidar.
Esto implica también fortalecer los espacios de representación del sector privado para fomentar una política exterior que vea en las exportaciones no solo una fuente de divisas, sino un motor de empleo y desarrollo. Ecuador necesita una narrativa común que entienda al comercio como una prioridad real, más allá de cualquier eslogan.
Desde la Cámara Nacional de Acuacultura promovemos un relacionamiento internacional basado en el respeto mutuo, la apertura al diálogo técnico y la defensa de condiciones justas para nuestros exportadores.
Hoy más que nunca, pragmatismo no es indiferencia ni neutralidad. Es inteligencia estratégica para seguir abriendo puertas, incluso en medio de un mundo que parece cada vez más dispuesto a cerrarlas•
DIRECTORIO
PRIMER VICEPRESIDENTE
Ing. Luis Francisco Burgos
Ing. Ricardo Solá
Dr. Alejandro Aguayo
Ing. Chris Olsen
Ing. Francisco Pons
Ing. José Antonio Lince
Ing. Jorge Redrovan
Ing. Alex de Wind
Ing. Kléber Siguenza
Ing. Rodrigo Vélez
Ing. Iván Rodríguez
Ing. Juan Carlos Vanoni
Ing. Alejandro Ruiz-Cámara
PRESIDENTE DEL DIRECTORIO
Ing. Marcelo Vélez
VOCALES
Econ. Heinz Grunauer
Ing. Víctor Ramos
Ing. David Eguiguren
Ing. Humberto Dieguez
Ing. Eduardo Seminario
Ing. Miguel Uscocovich
Ing. Vinicio Aray Dueñas
Econ. Sandro Coglitore
Ing. Rodrigo Laniado
Ing. Roberto Aguirre
Blgo. Carlos Sánchez
Ing. Diego Puente
SEGUNDO VICEPRESIDENTE
Ing. Fabricio Vargas
Ing. Johnny Adum
Sra. Verónica Dueñas
Ing. Alex Elghoul
Ing. Bastien Hurtado
Ing. Luis Burgos
Econ. Wolfgang Harten
Jorge Gonzalez
Andres Rivadulla
Ing. Héctor Marriott
Ing. Edison Brito
Dalian, China
Bruselas, Bélgica
Bruselas, Bélgica
En la década de los 90, el sector camaronero ecuatoriano inició su presencia coordinada en ferias internacionales junto a la Cámara Nacional de Acuacultura. Posteriormente, CORPEI asumió el liderazgo y fortaleció esta estrategia mediante la creación de las primeras oficinas comerciales en el extranjero. A partir de 2011, la Cámara Nacional de Acuacultura retomó este rol, consolidando la promoción del camarón ecuatoriano en los principales mercados del mundo.
- JUNIO 2025
La marca creada por la CNA y
que lleva el camarón ecuatoriano al escenario mundial
La historia de cómo Ecuador llegó a ser reconocido como referente mundial en la industria del camarón no solo se mide en toneladas exportadas o divisas generadas. También se cuenta desde una narrativa de identidad, visión y unidad. Esa historia lleva nombre propio: Ecuador First Class Shrimp (EFCS), la marca creada por la Cámara Nacional de Acuacultura (CNA) ante la necesidad identificada por sus empresas afiliadas, que no solo querían vender camarón ecuatoriano al mundo, sino diferenciarse y representar a su país con orgullo, técnica y profesionalismo en los escenarios más exigentes.
Los orígenes de una representación gremial
La Cámara Nacional de Acuacultura, constituida por el Congreso Nacional el 18 de julio de 1993, nació para acompañar a una industria que, aún en crecimiento, mostraba ya un enorme potencial productivo y exportador. Para la década de los 90, la CNA organizaba la participación de las empresas ecuatorianas en ferias internacionales. Años después, la organización se trasladó a CORPEI, que hasta el 2011 tuvo a cargo esta responsabilidad. La mayoría de las empresas vendía más del 90% de su producción a Estados Unidos, pero el mundo comenzaba a mostrar interés por el camarón ecuatoriano, y era evidente que se necesitaba algo más: identidad.
En 2011, CORPEI entregó nuevamente la organización de ferias internacionales del sector camaronero a la CNA. Sin embargo, no existía una marca o imagen que diferenciara la gestión de la Cámara y la presencia de sus socios. Hasta antes de 2014, las participaciones en ferias internacionales se realizaron bajo la marca país vigente. En ese entonces, la CNA y sus miembros, con el objetivo de diferenciar nuestra participación de los demás, trabajaron en la creación de Ecuador First Class Shrimp.
Esta no fue una estrategia diseñada desde un escritorio en la CNA: ha sido una evolución orgánica, nacida desde las experiencias vividas por empresarios y la industria camaronera ecuatoriana en las ferias más importantes del mundo, desde la primera participación en Boston (2015) hasta la más reciente Seafood Expo Global, donde se evidencia una evolución tangible con pabellos imponentes.
Hoy, el pabellón First Class no solo presenta camarón. Transmite confianza, hospitalidad y profesionalismo. Cada detalle ha sido afinado con los años: el diseño, la iluminación, la atención personalizada, las degustaciones. Todo responde a una misma premisa: Ecuador no solo vende camarón, exporta excelencia.
Esencia
Uno de los pilares fundamentales de Ecuador First Class Shrimp es su carácter colectivo. No pertenece a una sola empresa, ni a una alianza puntual. Es una marca gremial, construida con el aporte voluntario y constante de empresas unidas bajo una misma visión. En un sector donde cada metro cuadrado en feria cuesta, que las compañías apuesten año tras año por el pabellón común es una prueba contundente del valor que le asignan a esta marca y la confianza que depositan en la gestión de la CNA.
El impacto de Ecuador First Class Shrimp no solo se mide por el tamaño del stand o el número de visitantes. También se refleja en la manera en que el país es percibido por compradores, autoridades y organizadores de ferias. Representantes de eventos como Conxemar (España), China Fisheries (Asia) y Diversified Communications (EE. UU.) han reconocido la evolución del pabellón ecuatoriano y su consolidación como uno de los más profesionales y atractivos del circuito internacional.
Construir una marca de esta magnitud no estuvo exento de desafíos. Hubo que alinear agendas empresariales, consensuar presupuestos, desarrollar manuales de uso de marca, asegurar calidad gráfica y audiovisual, y, sobre todo, mantener el estándar a lo largo del tiempo. Pero la recompensa ha sido clara: Ecuador transmite en sus pabellones un mensaje de confianza, excelencia y la visión de largo plazo de su industria.
Actualmente, a diez años de su creación, Ecuador First Class Shrimp no solo es una marca posicionada. Es un símbolo. Un sello que agrupa a quienes decidieron que la excelencia no es un objetivo lejano, sino una práctica diaria. La marca ha sido validada en mercados de alta exigencia, reconocida por compradores internacionales y, lo más importante, sostenida por el compromiso firme de sus actores.
Porque, al final, no se trata solo de exportar un producto. Se trata de contar una historia. Una historia que habla de resiliencia, de innovación, de trabajo conjunto y de orgullo por lo propio. Una historia que se renueva en cada feria, en cada degustación, en cada trato cerrado bajo el sello de Ecuador First Class Shrimp.
Cuando hoy vemos ese pabellón en los grandes eventos del mundo, no vemos solo stands. Vemos el reflejo de un país que aprendió a contarse con voz propia. Vemos una industria que entendió que su mejor carta de presentación no es solo la calidad de su camarón, sino la solidez de sus empresarios y su compromiso colectivo con la excelencia.
Proximos eventos:
Entre agosto y octubre de 2025, la Cámara Nacional de Acuacultura tiene previsto representar al sector camaronero ecuatoriano en cinco
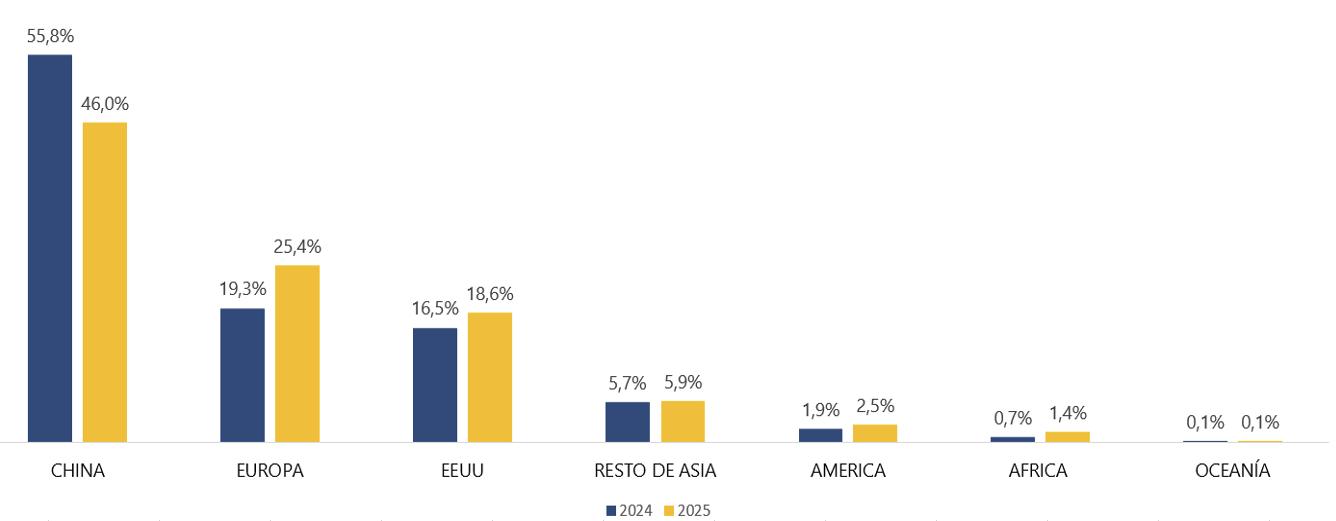
Desde su lanzamiento en 2015, la marca Ecuador First Class Shrimp ha llegado a más de 30 países, consolidando al camarón ecuatoriano como un producto premium en mercados clave de Asia, Europa, América y Medio Oriente, a través de ferias internacionales como Seafood Expo Global, China Fisheries, World Seafood Shanghai y Conxemar.
COYUNTURA
ferias internacionales de alto impacto. El recorrido comenzará en Tokio, Japón, con la Japan International Seafood & Technology Expo del 20 al 22 de agosto; continuará en Shanghái, China, con la World Seafood Shanghai del 27 al 29 del mismo mes; y seguirá en Singapur, con la Asia Seafood Expo Asia del 10 al 12 de septiembre. En octubre, la agenda contempla la participación en Conxemar, del 7 al 9 en Vigo, España, y culminará en Qingdao, China, con la China Fisheries and Seafood Expo del 29 al 31.
Esta intensa agenda internacional refleja el compromiso del sector acuícola ecuatoriano por posicionar al camarón como un producto premium en los mercados más exigentes del mundo. La participación en estas ferias no solo consolida relaciones comerciales estratégicas, sino que permite mostrar los avances en sostenibilidad, trazabilidad e innovación que distinguen al Ecuador como líder en acuicultura responsable. Cada feria es una vitrina para fortalecer la marca EFCS y abrir nuevas oportunidades para los productores que apuestan por la excelencia•
Barcelona, España 2025
Nuevas autoridades lideran el aparato productivo del Ecuador con enfoque técnico y estratégico
En una nueva fase de gestión pública en Ecuador, el presidente Daniel Noboa ratificó a Luis Alberto Jaramillo Arias como ministro de Producción, Comercio Exterior, Inversiones y Pesca. Funcionario con más de 30 años de trayectoria en el sector empresarial e institucional, Jaramillo es ingeniero comercial de formación con estudios de alta dirección en instituciones como el MIT y Kellogg. Ha ocupado cargos gerenciales en empresas privadas y fue director general del Servicio Nacional de Aduana del Ecuador (Senae). Desde su primera gestión en el despacho ministerial en octubre de 2024, ha buscado impulsar una agenda centrada en la atracción de inversiones, el fortalecimiento de la competitividad, la apertura comercial y la articulación con el sector privado.
A su equipo se sumó desde esta primera designación a Ivanova Cereceda Jalil , quien asumió el cargo de viceministra de Acuacultura y Pesca. Economista de profesión con una sólida experiencia en comercio exterior y trazabilidad, Cereceda ha enfocado su gestión en la eficiencia institucional, el combate a la pesca ilegal y la implementación de herramientas tecnológicas como el sistema
Palantir, que fortalecerá el control y seguimiento de productos del mar. Su administración ha priorizado el diálogo con los gremios y la sostenibilidad como ejes del desarrollo acuícola.
Asimismo, fue nombrada como viceministra de Comercio Exterior Alexia Alcívar, especialista en negocios internacionales y con experiencia en promoción comercial en mercados como Estados Unidos, Europa y Medio Oriente. Alcívar ha liderado una activa política exterior que ya muestra resultados concretos: la entrada en vigencia del Tratado de Libre Comercio con China en mayo de 2024, la firma de acuerdos con Costa Rica y la finalización de negociaciones con Canadá y Corea del Sur. Su trabajo se ha enfocado en diversificar mercados, atraer inversión sostenible y consolidar a Ecuador como un socio estratégico confiable.
Con estas designaciones, el Gobierno busca consolidar un liderazgo técnico, eficiente y transparente en una de las carteras más relevantes para la economía nacional. La combinación de experiencia, formación internacional y compromiso público-privado de este nuevo equipo marca una hoja de ruta clara: transformar al Ecuador en un país más competitivo, innovador y sostenible ante los mercados globales.
Con el objetivo de conocer de primera mano las prioridades y líneas de acción del nuevo equipo gubernamental, la revista AQUACULTURA mantuvo entrevistas con las autoridades designadas del Ministerio de Producción, Comercio Exterior, Inversiones y Pesca (MPCEIP). En estos espacios, los funcionarios expusieron su visión, los principales ejes de trabajo y los mecanismos de articulación que plantean para fortalecer la competitividad del sector productivo y exportador, con énfasis en la acuacultura y pesca.
LUIS JARAMILLO: LIDERAZGO Y COOPERACIÓN PÚBLICO – PRIVADA
¿Cuál es su principal objetivo al frente del ministerio, en este contexto económico tan complejo?
Promover la inserción estratégica del Ecuador en el comercio mundial a través del desarrollo productivo, la mejora de la competitividad integral y el impulso de cadenas de valor. Trabajamos con honestidad, de la mano del sector privado y la academia, para impulsar la confianza del mundo y mostrar lo mejor de este país.
¿Cómo se está trabajando para que las exportaciones ecuatorianas sean más competitivas?
Con una política de competitividad de largo plazo, identificando sectores sofisticados, apoyando a las industrias con innovación, formación y sostenibilidad. Reconocemos el esfuerzo de sectores como el acuícola, que han logrado posicionar productos de calidad a nivel global.
¿Cómo se coordina internamente la política productiva sin caer en la tramitología?
Cada viceministerio tiene metas claras. Mantenemos una agenda comercial continua y trabajamos en procesos internos más transparentes, automatizados y ágiles. El objetivo es reintegrar los valores perdidos y mostrar al mundo nuestro potencial.
IVANOVA CERECEDA: EFICIENCIA, TRAZABILIDAD Y DIÁLOGO EN ACUACULTURA Y PESCA
La viceministra Ivanova Cereceda Jalil, conocedora del sector camaronero, aporta una gestión técnica con énfasis en trazabilidad, diálogo abierto y control eficiente.
¿Qué diferencia a esta gestión frente a otras?
No soy partidaria de comparaciones, pero sí puedo afirmar que esta es una administración de puertas abiertas. Buscamos consensos y trabajamos con eficiencia, porque la eficiencia combate la corrupción. El funcionario público debe servir, facilitar y controlar.
¿Cuáles son las prioridades del sector para los próximos seis meses? Salir de la tarjeta amarilla de la Unión Europea es una prioridad. Ya hicimos los deberes: tenemos una Ley de Pesca, un sistema de trazabilidad (SIAP) y ahora implementaremos Palantir, una plataforma inteligente que fortalecerá el control y las operaciones del sector.
¿Qué se está haciendo frente al crimen organizado?
Aunque no es competencia directa del viceministerio, acompañamos al sector como puente con los ministerios de Interior y Defensa.
¿Cómo se construye confianza con productores y mercados internacionales?
Con diálogo franco, transparencia y trazabilidad. Estos sectores son innovadores, creativos y motivo de orgullo nacional.
COYUNTURA
ALEXIA ALCÍVAR: DIPLOMACIA COMERCIAL CON VISIÓN TÉCNICA
Desde el viceministerio de Comercio Exterior, Alexia Alcívar promueve una estrategia moderna de inserción global. Su experiencia en AMCHAM y en misiones comerciales en EE. UU., Europa y Medio Oriente respalda su visión de país como socio estratégico.
¿Qué puertas ha abierto Ecuador recientemente en comercio exterior? Hemos logrado hitos como el TLC con China, vigente desde mayo de 2024, que da acceso preferencial al 99,6 % de nuestras exportaciones. También firmamos con Costa Rica, concluimos negociaciones con Canadá y Corea del Sur, y avanzamos con Emiratos Árabes Unidos. El resultado: un superávit comercial de USD 6.678 millones en 2024, el más alto desde la dolarización.
¿Cómo avanza la relación comercial con EE. UU.?
Trabajamos bajo la hoja de ruta del Consejo de Comercio e Inversiones (TIC), buscando un acuerdo integral que alivie los efectos de los aranceles recíprocos y facilite un comercio bilateral justo, transparente y sostenible.
¿Qué oportunidades ofrece Ecuador en mercados emergentes como Asia o Medio Oriente?
Con China hemos crecido exponencialmente en productos como cacao elaborado, balsa y langosta. En Emiratos Árabes, hay interés en invertir en agroindustria y energía. Además, India y Arabia Saudita muestran alto potencial para productos frescos y de calidad como camarón, banano y rosas.
¿Qué esfuerzos se están haciendo para atraer inversión sostenible? Hemos incorporado principios de sostenibilidad en los acuerdos
El MPCEIP mantiene una agenda de trabajo orientada a la coordinación con el sector privado y la academia para abordar temas relacionados con la eficiencia institucional. Entre sus líneas de acción se encuentran la facilitación de procesos, el fortalecimiento de las cadenas productivas, la trazabilidad en la acuacultura y pesca, y la implementación de mecanismos para la promoción comercial y el acceso a nuevos mercados.
En ese marco, el despacho ministerial ha delineado acciones junto a los viceministerios técnicos. En acuacultura y pesca, la gestión está enfocada en la operatividad del sistema de trazabilidad, la regulación sectorial y la coordinación interinstitucional. En el ámbito del comercio exterior, se han ejecutado procesos de negociación y entrada en vigencia de acuerdos comerciales con países como China y Costa Rica, así como avances con Canadá, Corea del Sur y Emiratos Árabes Unidos. Estas acciones forman parte de una estrategia enfocada en dinamizar las exportaciones, facilitar la inversión y fortalecer el posicionamiento del país en mercados internacionales•
comerciales. Se ofrecen incentivos tributarios y acompañamiento a empresas extranjeras. Apostamos por sectores estratégicos como energía renovable, minería responsable y turismo. Las zonas francas son parte de este esquema integral para fomentar desarrollo, empleo y sostenibilidad.
Ecuador vive una coyuntura compleja en materia de seguridad, donde la violencia criminal ya no se limita a las grandes ciudades ni se restringe a hechos aislados. Hoy, el crimen organizado ha penetrado profundamente en el tejido económico del país, afectando de forma directa a las actividades productivas que sostienen el empleo, la inversión y las exportaciones.
El principal problema ya no es solo el robo o la pérdida de bienes, sino la amenaza latente de incursiones armadas, intimidaciones y extorsiones sistemáticas.
Lo que antes se consideraba excepcional, ahora ocurre con una frecuencia alarmante. Predios sitiados, transportistas interceptados, trabajadores que deben pagar “vacunas” para poder operar, son solo algunas de las postales de una realidad que se expande por todo el mapa productivo del Ecuador.
La cifra, aunque significativa, es solo un reflejo parcial de este problema estructural.
La violencia que hoy enfrenta el sector productivo no solo afecta sus ingresos: altera la planificación, encarece los seguros, paraliza inversiones, disminuye la productividad y pone en riesgo la vida de trabajadores y empresarios.
En sectores como el camaronero o el logístico, las rutas de transporte se han convertido en escenarios de alto riesgo, donde cada cargamento se vuelve un blanco potencial para bandas armadas.
Las extorsiones disfrazadas de “pagos por seguridad” o “autorizaciones” son solo una parte del problema. Muchas veces, los grupos criminales ya no solo buscan dinero, sino control territorial: establecer quién entra, quién sale, cuándo se produce y cuánto se exporta. Esta lógica de dominio pone en peligro la soberanía económica de regiones enteras.
Respuesta del Estado: despliegue operativo y coordinación multisectorial Ante esta realidad, el Gobierno ha desplegado una estrategia combinada entre Policía Nacional y Fuerzas Armadas. Se ha conformado un bloque de seguridad con más de mil uniformados que patrullan zonas productivas por aire, tierra y agua. Se han establecido retenes móviles, puntos de control militar en accesos estratégicos y se ha intensificado el uso de drones y vigilancia remota en coordinación con el ECU911.
En paralelo, desde marzo de 2025, funciona la Mesa Técnica de Seguridad del Sector Productivo, liderada por el Ministerio del Interior, que articula reuniones permanentes con gremios empresariales.
Esta instancia permite coordinar protocolos de respuesta, fortalecer canales de denuncia y compartir inteligencia operativa. La Fiscalía, por su parte, ha habilitado unidades especializadas para acelerar la investigación de casos de extorsión y delitos conexos.
Si bien estas acciones han contenido parcialmente el avance del crimen en ciertas zonas, los gremios coinciden en que la solución requiere una política sostenida,
con inversión en equipamiento, formación policial, tecnología y justicia eficaz. No se trata solo de “reaccionar”, sino de reconstruir la capacidad del Estado para prevenir y controlar el delito en el largo plazo.
En esta nueva etapa de la lucha contra la inseguridad, el objetivo no es solo capturar a los autores materiales, sino debilitar el corazón financiero de las bandas. Por eso, desde 2024, se ha intensificado el trabajo conjunto entre la Policía Nacional y la Unidad de Análisis Financiero y Económico (UAFE), para detectar y desmantelar los flujos de capital que sostienen al crimen organizado.
Se han iniciado procesos de incautación de bienes, congelamiento de cuentas y bloqueo de transacciones sospechosas vinculadas a extorsiones, lavado de activos y financiamiento de grupos delictivos. En el primer trimestre de 2025, la UAFE emitió más de 300 reportes de operaciones inusuales, muchos de ellos con vínculos en zonas productivas clave.
También se ha reactivado el Consejo Nacional contra el Lavado de Activos, con la participación de la Superintendencia de Bancos, la Fiscalía y la Policía Antinarcóticos. Este cuerpo ha logrado la judicialización de redes que operaban bajo fachadas legales, pero financiaban actividades criminales. La inteligencia financiera se convierte así en un pilar fundamental para atacar el crimen de forma estructural.
El rol del sector privado: control, denuncia y cooperación
El sector empresarial no es un actor pasivo en esta lucha. Su participación activa es clave para frenar la expansión del crimen, por lo que las empresas deben fortalecer sus mecanismos internos de trazabilidad, prevención de lavado y verificación de proveedores. Además, deben generar una cultura de denuncia protegida, donde los casos de extorsión y amenazas puedan ser reportados con garantías.
Muchos gremios ya trabajan de la mano con el Estado para diseñar mapas de riesgo, establecer protocolos de evacuación, instalar sistemas de videovigilancia y capacitar al personal en gestión de crisis. Esta alianza público-privada se convierte en un activo
fundamental para blindar la operación de sectores clave para la economía.
La lucha contra la inseguridad en Ecuador ha entrado en una fase estratégica: golpear donde más le duele al crimen organizado, que son sus finanzas. Mientras las bandas buscan controlar territorios y economías locales a través del miedo, el Estado, en alianza con el sector privado, ha comenzado a responder con inteligencia, trazabilidad y cooperación.
Sin un flujo constante de recursos, las estructuras criminales pierden fuerza, capacidad operativa y poder de intimidación. Por ello, cortar el circuito financiero que alimenta la violencia no es solo una acción policial, es una política pública que protege el desarrollo.
Pero ningún esfuerzo será suficiente si no se garantiza la coherencia entre los diferentes poderes del Estado. Cuando las fuerzas del orden arriesgan sus vidas para capturar criminales y la justicia los deja libres por decisiones cuestionables, se desmorona la confianza ciudadana.
Es por eso que urge además una depuración profunda del sistema judicial y un compromiso firme por erradicar la impunidad desde sus cimientos.
Acciones destacadas Golpe a la economía criminal
GUAYAS | Desmantelan red de lavado vinculada a alias “Fito”
En Guayaquil, la Policía Nacional ejecutó el operativo Centinela VI, allanando varios
inmuebles relacionados con alias “Fito” y su organización. Como parte de una acción simultánea en 33 propiedades a escala nacional, se incautaron 47 bienes valorados en más de USD 13 millones y se detuvo a seis familiares del cabecilla.
En paralelo, se capturó a miembros de los Latin Kings, vinculados a extorsión y sicariato. La operación forma parte de una ofensiva coordinada contra estructuras como Tiguerones y Mafia Fénix.
MANABÍ| En Manta caen activos clave de la red financiera ilícita de Los Choneros En Manta, la intervención policial afectó directamente la estructura económica de Los Choneros. Se intervinieron bienes y propiedades utilizadas para el lavado de activos, con apoyo de unidades especializadas que rastrearon flujos financieros y desmantelaron mecanismos de ocultamiento patrimonial usados por la organización.
SANTA ELENA | Fuerzas del orden desmantelan redes criminales vinculadas al narcotráfico
La Policía Nacional y las Fuerzas Armadas desarticularon en Santa Elena una banda que operaba pistas clandestinas en Chanduy, utilizadas para actividades de narcotráfico. Durante la operación, dos sospechosos fueron abatidos y se destruyeron infraestructuras ilegales para aterrizaje de aeronaves. Además, se ejecutaron operativos
simultáneos en Salinas, La Libertad y Centro 5, que dejaron como resultado la captura de sicarios vinculados a Los Choneros, la detención de 10 personas por asociación ilícita y la recuperación de motocicletas y vehículos reportados como robados.
Estas acciones fortalecen la ofensiva contra el crimen organizado en la región costera.
EL ORO| Entre mayo y junio de 2025, las autoridades intensificaron los operativos en la provincia de El Oro para frenar el avance de los grupos de delincuencia organizada vinculados a delitos de extorsión y tráfico de drogas. En cantones como Pasaje y Santa Rosa, la Policía Nacional ejecutó allanamientos que permitieron la detención de presuntos extorsionadores, quienes exigían pagos a cambio de no atentar contra la vida o los bienes de sus víctimas.
Paralelamente, las unidades antidrogas, en coordinación con el Bloque de Seguridad, realizaron intervenciones en Machala y El Guabo, logrando decomisar importantes cantidades de sustancias sujetas a fiscalización. Varios detenidos fueron identificados como integrantes de redes dedicadas al microtráfico, que operan bajo el amparo de organizaciones criminales.
ESMERALDAS|Capturan
a extorsionadores ligados a Los Tiguerones
Cinco presuntos extorsionadores, dos de ellos menores de edad, fueron aprehendidos en el barrio El Coral, Esmeraldas, durante un operativo contra la banda Los Tiguerones. Se incautaron celulares usados para extorsión y motocicletas robadas. La acción permitió desarticular una célula activa y reducir la presión criminal en sectores vulnerables.
La magnitud y complejidad de los desafíos en materia de seguridad requieren no solo el accionar del Estado, sino también el compromiso activo del sector empresarial y de la sociedad en general. La articulación de esfuerzos permite sumar capacidades operativas, tecnológicas, logísticas y humanas para fortalecer la prevención, la vigilancia y la respuesta frente al crimen organizado•
Pamela Nath Directora
Cuando hablamos de sostenibilidad, pocas veces pensamos en un país pequeño que ha conquistado el mundo con un marisco. Y sin embargo, Ecuador lo ha hecho.
En poco más de cinco décadas, la industria camaronera ecuatoriana se ha convertido en el mayor productor y exportador de camarón del mundo, aportando más del 30% de la producción global de esta proteína.
Pero este logro no ha sido producto del azar. Detrás de este crecimiento hay una estrategia integral que ha apostado por la sostenibilidad, la innovación y la visión a largo plazo. Y hoy, ese compromiso puede abrir una nueva puerta para impulsar el crecimiento del sector: el acceso a las finanzas sostenibles.
Cada vez, más bancos e inversionistas están orientando su capital hacia proyectos que generan valor no solo económico, sino también ambiental y social. Según el White Paper del Pacto Mundial de la ONU España, el financiamiento sostenible está en auge. Y esto coincide con lo que los consumidores están exigiendo: el 80% de ellos —especialmente millennials y Generación Z— dice estar dispuesto a pagar más por productos responsables, según la encuesta Global Consumer Insights Pulse Survey de PwC del 2023.
Esto significa que, para las industrias, incluida la del camarón, ser sostenible está dejando de ser un valor agregado: es una condición para competir, crecer y mantenerse relevante.
Aprovechar los instrumentos que las finanzas sostenibles nos ofrecen, pensados para quienes producen con responsabilidad, será clave para seguir avanzando. No solo para sostener lo que ya hemos logrado, sino para fortalecer —y proyectar aún más— la posición de un país pequeño que, con visión y compromiso, ha conquistado el mundo con un marisco… y que hoy tiene todo para liderar también el futuro de la sostenibilidad.
Las finanzas también pueden cultivar sostenibilidad. En Ecuador, este cultivo se da en la industria del camarón, un sector que lidera la exportación global de esta proteína y que hoy busca crecer sin perder de vista el cuidado del ambiente, la innovación y el impacto social.
En los últimos años, el capital financiero global ha comenzado a redirigirse hacia proyectos que cumplen con criterios ASG: ambientales, sociales y de gobernanza. “Las finanzas sostenibles están experimentando una tendencia al alza, principalmente porque inversores y empresas consideran que existen importantes oportunidades de negocio en el ámbito de la sostenibilidad”, afirma el White Paper del Pacto Mundial de la Organización de las Naciones Unidas (ONU) en España
Esta transformación —impulsada por bancos, fondos de inversión y organismos internacionales— reconoce que el desarrollo sostenible no solo es necesario, sino también rentable. Según el Libro Blanco, las finanzas sostenibles son una herramienta para orientar recursos hacia actividades productivas que generen valor económico, social y ambiental a largo plazo.
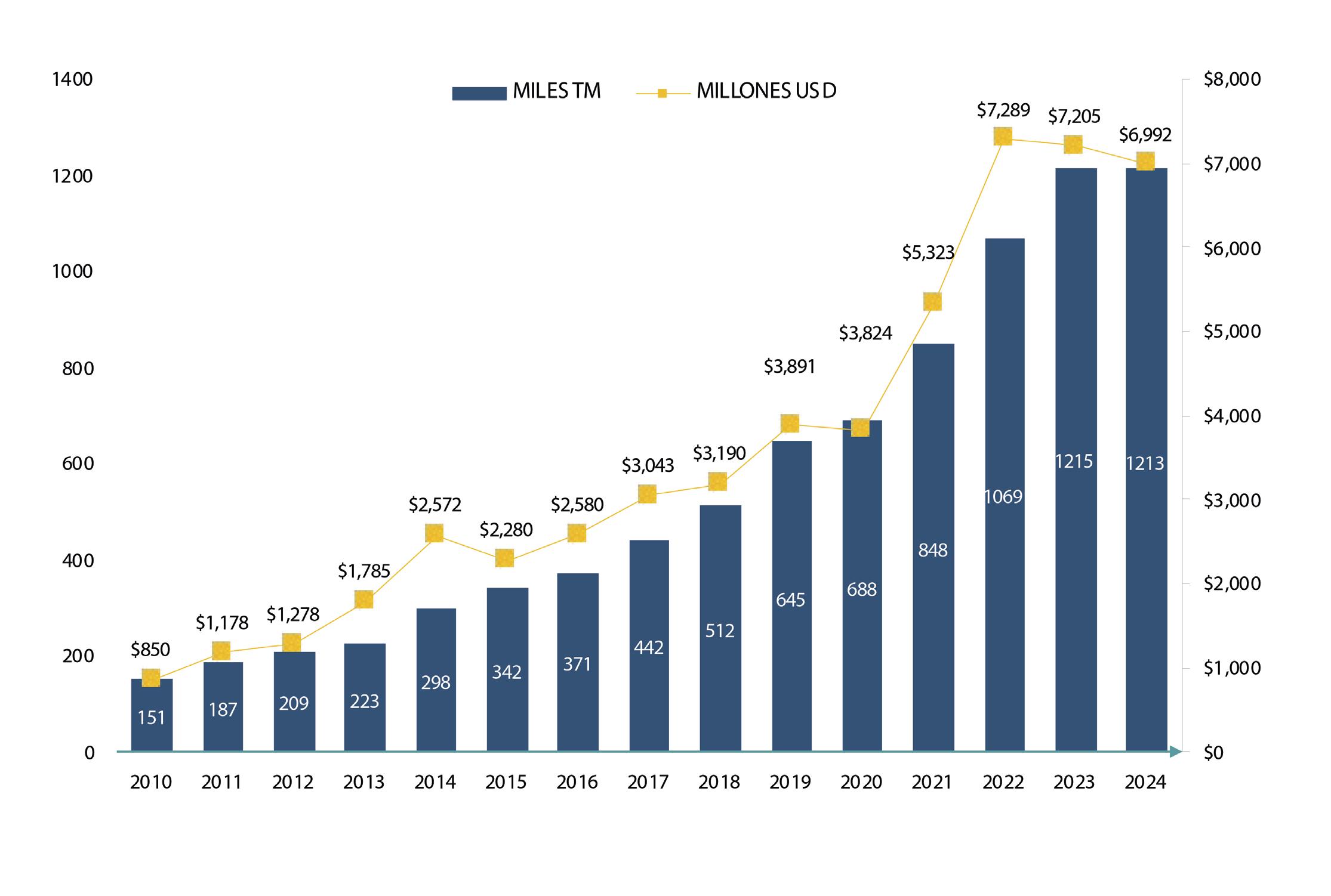
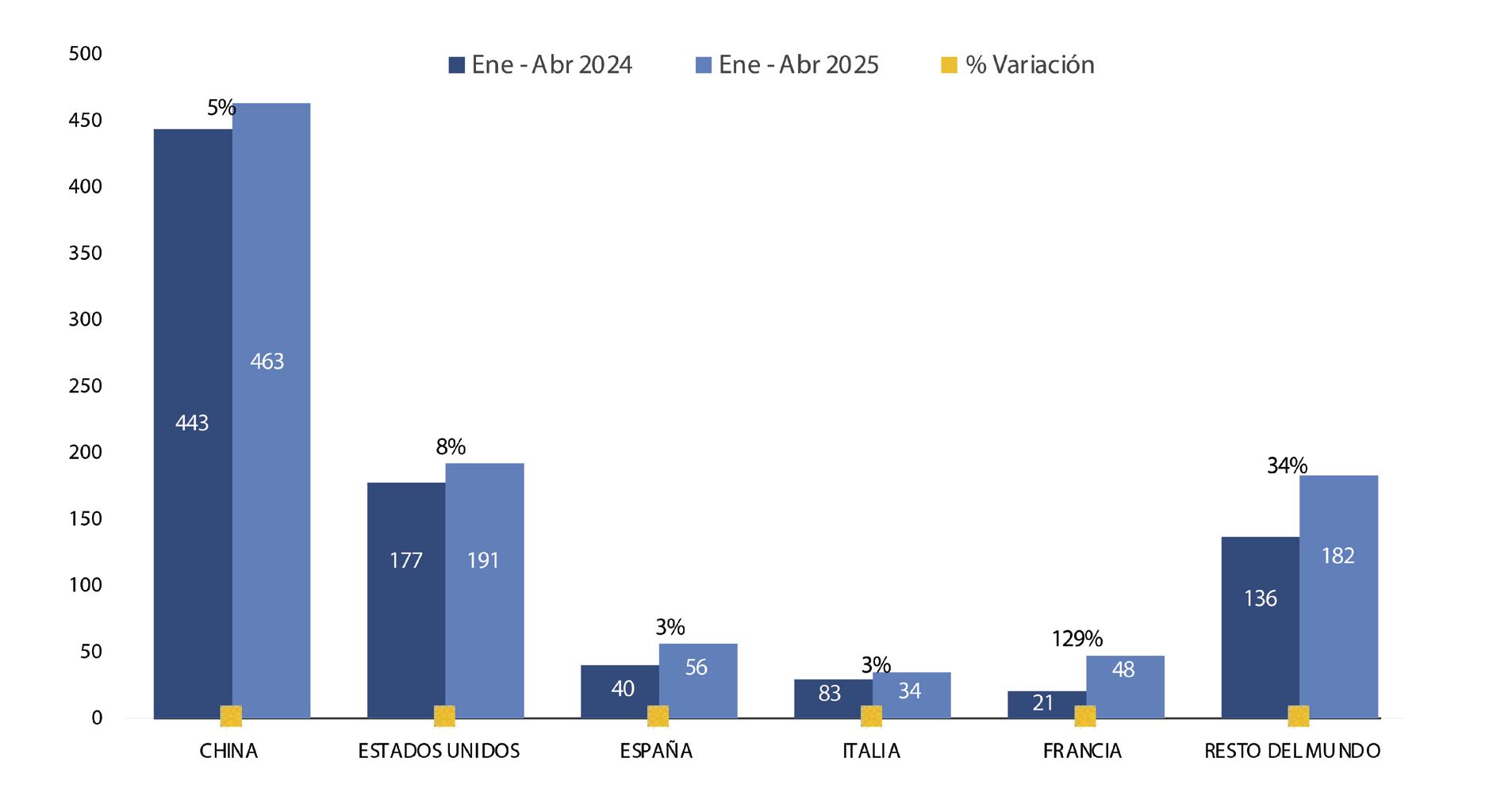
Con este nuevo escenario, el sector camaronero ecuatoriano se encuentra en una coyuntura estratégica para aprovechar este curso. De acuerdo con cifras del Banco Central, en 2024 Ecuador exportó 1,2 millones de toneladas de camarón, generando 6.992 millones de dólares. Ese mismo año, el sector proporcionó más de 290.000 empleos directos e indirectos en toda su cadena de valor. Esta industria, que comercializa su producto en 74 países, ya ha demostrado su capacidad de crecimiento y adaptación. Hoy, la expansión de las finanzas sostenibles representa una oportunidad para acelerar su transición hacia un modelo de desarrollo responsable, competitivo e innovador.
La Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO) respalda esta visión: los sistemas acuáticos ofrecen soluciones clave para la seguridad alimentaria, la reducción de la pobreza y el desarrollo costero, con baja huella ecológica. En 2022, la acuicultura alcanzó un récord de 130,9 millones de toneladas —superando por primera vez a la pesca de captura— y se estima que en 2032 proveerá el 60% de los alimentos acuáticos para consumo humano.
La nueva lógica financiera ofrece a la acuicultura del camarón una oportunidad real de crecer con responsabilidad. Diversas instituciones ya comienzan a impulsar ese cambio. Una de ellas es el Banco de Desarrollo Empresarial (FMO) de Países Bajos. Su oficial de inversiones, Soledad Jirón, comparte cómo desde la banca de desarrollo es posible acompañar al sector camaronero ecuatoriano en este camino.
“FMO tiene un mandato claro: invertir en sectores clave que impulsen la prosperidad local en mercados emergentes”, afirma Jirón. Para el sector acuícola, esto se traduce en tres líneas de acción: financiamiento a largo plazo para inversiones en infraestructura verde — como la electrificación de fincas y plantas de procesamiento eficientes—; asistencia técnica para fortalecer capacidades en sostenibilidad, gobernanza y gestión de riesgos, y la promoción de estándares internacionales como los Principios de Desempeño de la Corporación Financiera Internacional (IFC) y certificaciones como Aquaculture Stewardship Council (ASC), que aumentan la competitividad en mercados internacionales.
La experta señala que para seleccionar proyectos y garantizar que el financiamiento tenga impacto real, FMO prioriza aquellos con impacto ambiental positivo, generación de empleo y reducción de desigualdades, alineados con los Objetivos de Desarrollo Sostenible (ODS) de la ONU, en áreas como trabajo decente, acción climática y seguridad alimentaria. Asimismo, el organismo “espera que las empresas financiadas se comprometan con una mejora continua en sus prácticas ASG”, comparte.
El seguimiento continuo es clave. “No solo evaluamos al inicio, también monitoreamos el desempeño para asegurar el cumplimiento de compromisos e impacto”, señala Jirón. El control incluye indicadores ASG y planes de acción.
En cuanto a su estrategia de financiamiento, FMO utiliza distintos instrumentos: financiamiento directo a empresas líderes, con préstamos de hasta 35 millones de dólares
destinados a la electrificación y expansión de plantas de empaque; financiamiento indirecto a través de instituciones financieras locales, facilitando el acceso a recursos para una mayor diversidad de actores, y la participación en créditos sindicados, ya sea como estructurador o como parte de un consorcio junto a otros bancos de desarrollo, instituciones financieras y fondos, lo que permite movilizar mayores recursos, compartir riesgos y ofrecer condiciones más competitivas para los clientes.
Para acceder a financiamiento sostenible, la oficial de inversiones sugiere a los productores camaroneros seguir algunas recomendaciones clave:
• Adoptar estándares internacionales que demuestren compromiso con la sostenibilidad.
• Fortalecer la gobernanza y la transparencia, incluyendo reportes ASG y trazabilidad.
• Invertir en eficiencia energética y digitalización, que no solo reducen costos, sino que también mejoran el perfil de riesgo ante inversionistas.
• Buscar alianzas con instituciones financieras locales e internacionales enfocadas en el sector.
• Preparar una propuesta clara de
impacto, mostrando cómo el proyecto contribuye a los ODS y a la resiliencia del sector.
Esta visión internacional encuentra un eco creciente en el sistema financiero ecuatoriano. Varios bancos locales ya han empezado a desarrollar productos y estrategias alineadas con los principios ASG, contribuyendo desde dentro a fortalecer la sostenibilidad del sector acuícola ecuatoriano.
Un ejemplo concreto es el caso del Banco Bolivariano, que ha consolidado una propuesta robusta basada en instrumentos financieros especializados.
El gerente de Banca Empresas de la institución, Andrés Crespo, está convencido de que la sostenibilidad no es solo una responsabilidad ética, sino una oportunidad estratégica. “En el caso de la industria camaronera, nuestro compromiso se traduce en facilitar el acceso a financiamiento hacia modelos de producción más responsables y eficientes”, señala.
Crespo indica que, al promover este cambio,
se favorece tanto la reducción de riesgos ambientales y reputacionales como la creación de valor compartido: “Fortalecemos la competitividad del sector y su crecimiento en mercados internacionales, protegemos los recursos naturales y promovemos un crecimiento económico más inclusivo”.
Una de las herramientas clave del Banco Bolivariano son los Créditos Verdes, dirigidos a proyectos con beneficios ambientales comprobables. “Nuestro portafolio de estos créditos creció un 171%, lo que evidencia una demanda sostenida por financiamiento sostenible y un alto potencial para sectores clave como la acuicultura”, destaca.
Los Créditos Verdes se aplican a iniciativas como eficiencia energética, manejo responsable del agua, implementación de tecnologías limpias, energías renovables, conservación de ecosistemas, mejora de las condiciones laborales, entre otras.
Su propósito es impulsar la adopción de tecnologías que optimicen el uso de recursos naturales, protejan ecosistemas y enfrenten los desafíos del cambio climático, en línea con los compromisos de banca responsable.
Además, el banco cuenta con el Bono Azul, que canaliza recursos hacia proyectos que generan impactos positivos en ecosistemas marinos y costeros. “Priorizamos a
productores con certificaciones sostenibles reconocidas como ASC, Marine Stewardship Council (MSC) o Best Aquaculture Practices (BAP), así como a aquellos con compromisos claros en gestión de aguas residuales y economía circular”, detalla Crespo y añade que “los productores camaroneros certificados no solo pueden acceder a condiciones crediticias más favorables, sino que también fortalecen su reputación y su capacidad de posicionarse en mercados internacionales cada vez más exigentes”
Pero estos no son los únicos actores que han incorporado la sostenibilidad como eje estratégico. Banco Guayaquil también ha desarrollado una propuesta específica para apoyar al sector camaronero a través de sus propios créditos verdes, que combinan flexibilidad financiera con impacto ambiental. La entidad ha puesto en marcha productos financieros diseñados para facilitar inversiones en tecnologías limpias, gestión ambiental adecuada y mejoras sociales en comunidades vinculadas al sector acuícola. Uno de estos productos es el Crédito Terra, dirigido a proyectos que buscan mejorar la eficiencia energética, reducir emisiones o manejar residuos de forma responsable.
“Los productores de camarón pueden utilizar este crédito para modernizar sus sistemas de cultivo con tecnologías amigables con el ambiente”, detalla el banco. Las inversiones
elegibles incluyen infraestructura sostenible, innovación tecnológica y programas de capacitación en prácticas responsables.
Los productores interesados en acceder a este financiamiento deben cumplir una serie de criterios, entre ellos el cumplimiento de normativas ambientales y sociales, la implementación de prácticas de manejo responsable, un plan de sostenibilidad que refleje impactos positivos en el ambiente y la comunidad, una capacidad financiera adecuada y gestión transparente.
Más allá del sistema bancario tradicional, también han surgido alianzas que expanden el alcance de las finanzas sostenibles. Un Ejemplo es la alianza estratégica entre Banco Del Austro y Soluciones Ambientales Totales (SAMBITO) para la operativización de los créditos Bonos Azules. para el país.
“Los Bonos y los créditos que financiarán a través de banco del Austro son una herramienta clave para la conservación de nuestros recursos marinos y costeros”, afirma el presidente de Sambito, Gustavo Manrique. Según el experto, la economía azul representa entre el 6 y el 10% del PIB nacional, por lo que su transformación sostenible tiene un alto potencial de impacto.
“Los Créditos Bonos Azules, alinean los intereses de crecimiento económico, conservación ambiental y desarrollo social para financiar proyectos que aseguren la salud de los ecosistemas marinos, la resiliencia de las comunidades costeras y el cumplimiento de normativas internacionales, como las vinculadas al acceso a mercados sostenibles”, explica Manrique.
Los proyectos camaroneros que acceden a este financiamiento deben cumplir con criterios ambientales y sociales, incluyendo reducción de emisiones, eficiencia energética, manejo responsable de efluentes y condiciones laborales dignas. Para asegurar un impacto positivo, Banco del Austro ha desarrollado un proceso de evaluación técnica de monitoreo continuo, con indicadores que, a través, del acompañamiento técnico de SAMBITO garantizan el uso transparente y trazable de los fondos. “Esta alianza ha demostrado que, cuando el sistema financiero y los actores técnicos trabajan juntos, es posible implementar productos innovadores que promueven el desarrollo sostenible y la competitividad del país en los mercados internacionales”, concluye el presidente de la consultora.
Mientras organismos como FMO y la banca local canalizan recursos hacia la sostenibilidad, la electrificación del sector camaronero ilustra cómo estos fondos
pueden generar transformaciones concretas. GPS Group, con más de una década en soluciones energéticas inteligentes, ha electrificado aproximadamente 15.000 hectáreas de camaroneras en Ecuador. En este proceso, ha logrado articular financiamiento sostenible con tecnología aplicada.
Para conocer más, conversamos con el gerente de Viabilidad de la empresa, Alberto Barriga, sobre los desafíos y oportunidades de modernizar las camaroneras del país.
¿En qué consiste exactamente la electrificación de una camaronera?
Se trata de sustituir y/o disminuir el uso de diésel por energía eléctrica, particularmente en dos procesos fundamentales del cultivo de camarón: el bombeo de agua y los sistemas de aireación de las piscinas. Esta transformación requiere acceso a la red eléctrica pública o, alternativamente, sistemas de autogeneración.
¿Pero qué implica esto en términos de desarrollo tecnológico?
Vemos la electrificación como un medio para el acceso a la tecnología, no como un fin. Si bien es cierto que existe un impacto
directo en el costo de producción por el reemplazo o reducción del uso del diésel, cuando un proceso o sistema funciona con energía eléctrica se pueden automatizar procesos, gestionar en tiempo real y manejar información estratégica para la toma de decisiones que permita una acuicultura de camarón más predictible, competitiva y sostenible.
¿Cuál es el impacto ambiental y económico de esta medida?
Las camaroneras, al dejar de usar diésel, reducen significativamente sus emisiones de gases de efecto invernadero y el manejo de desechos peligrosos, como lubricantes y filtros. En cifras, dependiendo de cada camaronera, una hectárea electrificada puede reducir entre 2.000 y 4.000 galones de diésel al año, lo que se traduce en una disminución de entre 10 y 14 toneladas de CO2 por hectárea anualmente.
¿Cuáles son las barreras que enfrenta este cambio y cuál el rol del financiamiento sostenible?
Al momento de incorporar sistemas eléctricos en las camaroneras, existen diversas barreras. Primero, hay que considerar la visión del dueño de la camaronera; luego, el acceso a la energía, que muchas veces se ve limitado por la falta de inversión en la expansión de las redes públicas. También hay un desconocimiento sobre cómo
implementar un proyecto de infraestructura de este tipo y, por supuesto, el acceso al capital.
¿El acceso a financiamiento sostenible puede ser un impulsor clave? Definitivamente, los productos financieros están bien diseñados pero existen oportunidades para mejorar el entendimiento mutuo: por un lado, la banca no siempre está familiarizada con los aspectos técnicos del sector camaronero, y por otro, los productores enfrentan desafíos al manejar el lenguaje financiero. Por ello, se sugiere el uso de asesoría técnica y soporte técnico para tomar decisiones acertadas.
La electrificación de camaroneras, con sus
repercusiones positivas, es una muestra concreta del impacto que pueden tener las finanzas sostenibles cuando se articulan con soluciones tecnológicas y visión de largo plazo. Pero este es solo uno de muchos caminos posibles. La transformación del sector requiere una estrategia integral que conecte innovación, financiamiento y compromiso en toda la cadena de valor.
“Lo que ocurre en Ecuador es una muestra de cómo una industria puede transformarse con propósito. La colaboración entre productores, banca, expertos y alianzas como SSP está impulsando una acuicultura más competitiva y sostenible. Las herramientas ya existen. Ahora se trata de usarlas estratégicamente para generar valor y bienestar. Es una
oportunidad real para liderar un cambio global desde lo local”, concluye Pamela Nath, directora de Sustainable Shrimp Partnership (SSP).
Así, las finanzas pueden cultivar sostenibilidad cuando respaldan decisiones que combinan crecimiento y responsabilidad económica, social y ambiental de forma integrada•
Para más información sobre este artículo, contactar a: pnath@sustainableshrimp.org
ARTÍCULOS TÉCNICOS ÍNDICE
Edición 165 - Junio 2025 26 31 36
Las pequeñas granjas camaroneras: recomendaciones para ser más eficientes y sostenibles
Aliados invisibles: Cómo los microorganismos transforman la productividad en acuacultura
Evaluación de la diversidad bacteriana como indicador de calidad en suelos de cultivos acuícolas y biorremediación para la mejora productiva
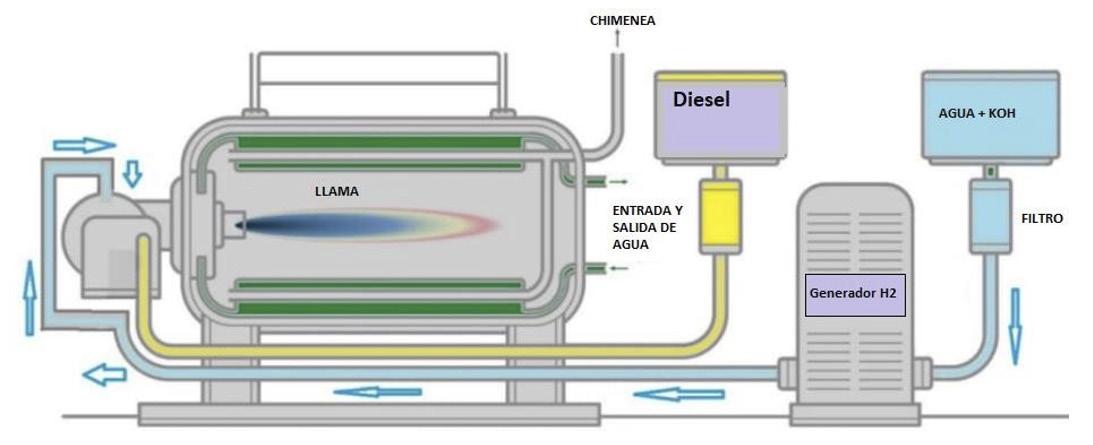
H2 para eficiencia térmica y reducción de emisiones en calderas y MCI
Adición de suplementos promotores de salud en microdietas para postlarvas de camarón blanco (Penaeus vannamei): efectos sobre el crecimiento, la supervivencia y el estado de salud
La trazabilidad total ya es una realidad en el Ecuador
El microbioma del camarón blanco del Pacífico revela una composición diferencial de la comunidad bacteriana entre condiciones naturales, de cultivo y en brotes de AHPND/EMS
Las pequeñas granjas camaroneras: recomendaciones para ser más eficientes y sostenibles
Autores:
Leonardo S. Maridueña
Simone Maridueña L.
Dirección de Ambiente
Cámara Nacional de Acuacultura
lmariduena@cna-ecuador.com
La acuicultura es el sistema de producción que más rápido ha crecido en el mundo, sobrepasando a la pesquería y la producción de carne. Si ponemos en contexto la cantidad de producción, tiempo y todo el esfuerzo involucrado, los acuicultores han aprendido muy rápido en convertirse en más eficientes y producir más con pocos cambios y efectos. Sin embargo, la situación económica no es igual entre los productores y existen los grandes, medianos y pequeños; estos últimos, por la economía de escala, son quienes más necesitan orientación, para que, con una dirección adecuada, puedan ser también resistentes a los cambios que inesperadamente genera la naturaleza y a los efectos antropocéntricos que se suscitan en su entorno.
El presente artículo tiene por objetivo dar una orientación técnica a pequeños productores, con la finalidad de hacer eficientes y sustentables las pequeñas granjas camaroneras (menores a 100 hectáreas), aplicando una estrategia integral enfocada en bioseguridad, tecnología, manejo ambiental y economía circular, sin perder de vista la rentabilidad.
Los pasos a seguir se describen a continuación:
Diagnóstico inicial
El primer paso fundamental para encaminar una pequeña granja camaronera hacia la eficiencia y la sostenibilidad consiste en realizar un diagnóstico integral del sistema de producción. Este análisis debe considerar no solo las condiciones biofísicas del entorno, sino también la infraestructura, el historial productivo y los principales cuellos de botella operativos.
En cuanto a los aspectos ambientales, se requieren estudios detallado de los suelos y el agua que ingresa al sistema. Estos deben realizarse al menos dos veces al año —una en la estación lluviosa y otra en la seca— para captar las variaciones estacionales.
Los parámetros que deben evaluarse incluyen pH, sólidos suspendidos, demanda bioquímica de oxígeno (DBO), demanda química de oxígeno (DQO), nitrógeno total, fósforo, aceites y grasas, así como un análisis bacteriológico completo. Esta información permite identificar riesgos potenciales
para la salud del cultivo y orientar acciones correctivas basadas en evidencia.
La revisión de la infraestructura constituye otro componente esencial del diagnóstico. Es necesario inspeccionar todas las instalaciones físicas, incluyendo estanques, compuertas, canales de ingreso y drenaje, así como equipos como aireadores y bombas. La identificación de estructuras obsoletas o deterioradas puede evitar fallas operativas durante el ciclo productivo.
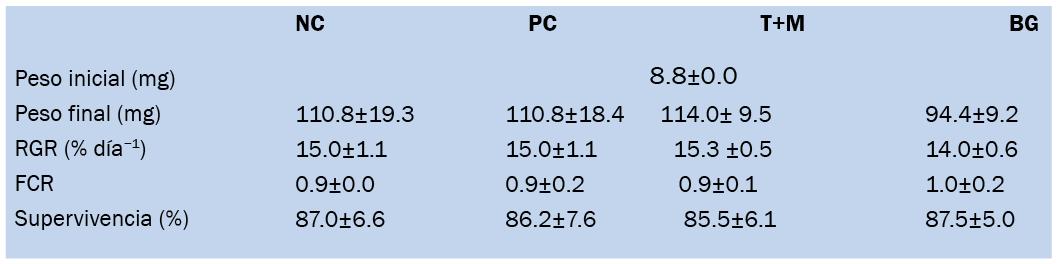
Asimismo, se debe analizar el historial productivo de la granja, recopilando datos de ciclos anteriores como fechas de siembra y cosecha, crecimiento biométrico, factores de conversión alimenticia (FCR), eventos de mortalidad y aparición de enfermedades. Estos datos permiten establecer patrones de desempeño y sirven como línea base para las mejoras propuestas.
Finalmente, es crucial identificar problemas críticos que afecten la rentabilidad del sistema, tales como niveles bajos de oxígeno disuelto, episodios frecuentes de mortalidad, tasas de crecimiento por debajo del promedio y sobrealimentación. La correcta identificación de estos puntos vulnerables permite diseñar un plan de manejo específico y adaptado a la realidad de cada unidad productiva.
Producción sustentable
La implementación de prácticas de producción sustentable permite aumentar la eficiencia del cultivo sin comprometer la salud ambiental ni la viabilidad económica del sistema. Esta estrategia debe centrarse en tres pilares: calidad del agua, manejo de densidades y eficiencia alimenticia.
a) Mejora de la calidad del agua sin uso de químicos
Una adecuada gestión del agua es esencial para prevenir enfermedades, mejorar el crecimiento de los camarones y reducir la dependencia de productos farmacológicos. Se recomienda implementar sistemas de recirculación parcial que promuevan la estabilidad de los parámetros fisicoquímicos. Estos también contribuyen a mitigar el impacto ambiental de los vertidos. En lugar del uso de antibióticos, que puede inducir resistencia bacteriana y dañar la
microbiota del ecosistema, se promueve la aplicación de probióticos y biorremediadores. Estos productos biológicos ayudan a mantener una comunidad microbiana equilibrada y a degradar materia orgánica en exceso, mejorando la calidad del agua de forma natural.
El monitoreo constante de parámetros clave como oxígeno disuelto, pH, amonio, nitritos y temperatura es indispensable. Este control no solo permite tomar decisiones rápidas ante cambios críticos, sino que también facilita la evaluación del desempeño de las estrategias implementadas.
b) Control de densidades y siembras
Una densidad de siembra adecuada es un factor determinante en la salud del cultivo. En pequeñas granjas, se recomienda mantener densidades moderadas —entre 10 y 30 individuos por metro cuadrado— para reducir el estrés, disminuir la incidencia de enfermedades y facilitar el manejo técnico del sistema.
El tipo de cultivo más apropiado dependerá de la infraestructura disponible. Los sistemas semiintensivo o intensivo pueden ser viables siempre que exista una aireación eficiente. Es fundamental asegurar que la capacidad de oxigenación del sistema sea proporcional a la biomasa cultivada, lo cual requiere una planificación técnica previa.
El uso de postlarvas certificadas, libres de patógenos como el virus del síndrome de la Mancha Blanca (WSSV) o el síndrome de Mortalidad Temprana (EMS), es una práctica esencial para reducir riesgos sanitarios. Estos organismos deben provenir de laboratorios que garanticen trazabilidad, sanidad y buena genética.
c) Alimentación eficiente
El manejo alimenticio es uno de los componentes con mayor impacto económico y ecológico dentro de la acuicultura. Por ello, es necesario establecer programas de alimentación basados en tablas de crecimiento, consumo real y observaciones in situ
La sobrealimentación no solo implica un desperdicio económico, sino que también deteriora la calidad del agua al aumentar la
carga orgánica. Para evitarlo, se recomienda el uso de bandejas de alimentación (trays) y, cuando sea posible, sensores de movimiento para ajustar las raciones según el comportamiento de los animales.
Asimismo, la implementación de comederos automáticos, ya sean simples o semimanuales con temporizador, permite una dosificación más precisa del alimento, reduciendo pérdidas y mejorando el FCR. Esta estrategia también disminuye la necesidad de alimentación nocturna manual, optimizando el uso del recurso humano.
Tecnología apropiada y accesible
El acceso a tecnologías simples pero efectivas puede marcar la diferencia en la productividad de las pequeñas granjas. La adopción de soluciones adaptadas al contexto local permite modernizar la producción sin incurrir en altos costos.
Una opción viable es la incorporación de energía solar para alimentar equipos como aireadores o sensores, especialmente en zonas donde el suministro eléctrico es limitado y/o costoso. Los paneles solares, aunque requieren una inversión inicial, pueden reducir significativamente los gastos energéticos a mediano plazo.
En cuanto a la aireación, se recomienda sustituir equipos obsoletos o ineficientes por sistemas más modernos como los paddle wheels (ruedas de paleta) o difusores de burbuja fina, que ofrecen una mayor eficiencia en la transferencia de oxígeno por unidad de energía consumida.
Por otro lado, el registro de datos productivos —como biomasa, consumo de alimento, mortalidad y parámetros del agua— puede realizarse mediante aplicaciones móviles o plantillas en Excel. Estas herramientas, de bajo costo y fácil uso, simplifican el análisis técnico y permiten una mejor toma de decisiones basada en evidencia.
Enfoque ambiental
La sostenibilidad ambiental es un componente clave en la modernización de las pequeñas granjas camaroneras. Una estrategia de manejo ambiental adecuada no solo minimiza el impacto ecológico, sino que también permite aprovechar los recursos de
manera más eficiente y crear valor agregado. El tratamiento del agua de descarga debe ser una prioridad. La construcción de estanques de sedimentación o humedales artificiales permite retener sólidos y nutrientes antes de que el efluente llegue a cuerpos de agua receptores naturales.
Cuando la calidad lo permite, el agua utilizada durante la cosecha puede ser filtrada y reutilizada para el prellenado de los estanques en el siguiente ciclo, reduciendo así el consumo de líquido y los costos operativos.
El manejo de los lodos es otro aspecto fundamental. En lugar de desechar estos residuos, se propone su conversión en compost mediante la integración con residuos vegetales de fincas cercanas. El resultado es un fertilizante orgánico que puede ser utilizado localmente o comercializado, generando ingresos adicionales para la granja. Esta práctica se alinea con los principios de la economía circular y contribuye al cumplimiento de normas ambientales más estrictas.
Otro factor importante es la reforestación, ya sea natural o a través de la siembra del manglar. Se debe mantener un amortiguamiento con estas especies, ya que protegen de la erosión costera y por lo tanto contribuyen al cuidado de la infraestructura camaronera; además, se destaca su capacidad de absorber entre 102 y 376 toneladas de CO2 por hectárea durante su vida útil, lo que representa más del doble que otros ecosistemas naturales.
Capacitación y organización del personal El capital humano constituye un elemento fundamental para el éxito de las pequeñas granjas camaroneras. La capacitación continua del personal permite mejorar la eficiencia operativa, reducir errores y fomentar una cultura de manejo técnico y responsable.
Es esencial que el personal de campo reciba formación básica en temas clave como bioseguridad, técnicas de alimentación, monitoreo de parámetros de agua, identificación de enfermedades y buenas prácticas de manejo. Estas capacitaciones pueden ser brindadas por técnicos del
sector público, asociaciones de productores, universidades o a través de cursos en línea accesibles.
Una estrategia efectiva es asignar roles claros y responsabilidades específicas a cada trabajador. La creación de manuales operativos simples, con instrucciones para cada etapa del ciclo productivo, facilita el cumplimiento de los procedimientos y estandariza el trabajo.
La motivación del personal también juega un rol clave. El reconocimiento de logros, incentivos por resultados y un ambiente laboral positivo contribuyen a mejorar el compromiso del equipo con los objetivos productivos y ambientales de la granja.
Indicadores de eficiencia
Para evaluar el desempeño de una granja camaronera y orientar procesos de mejora continua, es necesario establecer indicadores de eficiencia técnica y económica. Algunos de los más relevantes incluyen:
• Tasa de supervivencia (%): mide el porcentaje de camarones sobrevivientes respecto al número sembrado. Valores superiores al 70% son considerados aceptables en sistemas bien manejados.
• FCR (factor de conversión alimenticia): indica la cantidad de alimento requerida para producir un kilogramo de biomasa. Un FCR eficiente se encuentra entre 1.4 y 1.8 en cultivos bien controlados.
• Ganancia diaria de peso (g/día): refleja el crecimiento promedio diario de los animales. Dependerá de la genética, la calidad del alimento y el ambiente, pero valores mayores a 0.15 g/día son deseables.
• Rendimiento por hectárea (kg/ha/ ciclo): representa la productividad del sistema. En granjas semiintensivas bien manejadas puede superar los 3.000 kg/ha/ciclo.
• Costo de producción por kilogramo: es un indicador económico clave. Debe incluir costos directos e indirectos para evaluar la rentabilidad real del sistema.
El análisis de estos indicadores permite identificar puntos críticos, comparar resultados entre ciclos y establecer metas de mejora. Su registro debe ser sistemático, utilizando herramientas sencillas como hojas
de cálculo o software especializado cuando esté disponible.
Innovación y mejora continua
La sostenibilidad de una granja camaronera a largo plazo depende de su capacidad de adaptarse, aprender y mejorar constantemente. La innovación no necesariamente implica grandes inversiones, sino la incorporación gradual de cambios que aumenten la eficiencia o reduzcan impactos negativos.
Se recomienda fomentar una cultura de mejora continua basada en el análisis de datos, la identificación de problemas recurrentes y la experimentación con soluciones prácticas. Por ejemplo, probar nuevas estrategias de alimentación, ajustar densidades o modificar técnicas de cosecha pueden arrojar resultados valiosos.
La colaboración con otras granjas, asociaciones o centros de investigación también puede generar sinergias positivas. El intercambio de experiencias, la participación en capacitaciones colectivas y el acceso a información técnica actualizada son factores que fortalecen la resiliencia del pequeño productor.
Caso práctico
Una granja ubicada en el archipiélago de Jambelí constituye un ejemplo representativo de una unidad productiva que ha logrado mejorar sus indicadores técnicos mediante la implementación de prácticas sostenibles. Esta finca, con una extensión de 10 hectáreas, operaba inicialmente con sistemas tradicionales de bajo rendimiento y presentaba problemas recurrentes de mortalidad y sobrealimentación.
Tras realizar un diagnóstico técnico en 2023, se identificaron deficiencias en el sistema de aireación, un mal manejo del alimento y falta de control sobre los parámetros del agua.
Con base en estos hallazgos, se diseñó un plan de mejora que incluyó:
• Instalación de dos aireadores de paleta alimentados con energía solar.
• Implementación de bandejas de alimentación y uso de sensores para evitar el exceso de comida.
• Capacitación del personal en
bioseguridad y manejo eficiente del alimento.
• Monitoreo semanal de oxígeno, pH y amonio, con registros en programa de Excel.
• Siembra con postlarvas certificadas de alta supervivencia.
Al finalizar el primer ciclo posterior a la intervención, la tasa de supervivencia aumentó del 55 al 78%, el FCR mejoró de 2.1 a 1.6, y el rendimiento por hectárea se incrementó en un 35%. Además, se redujo el uso de antibióticos y productos químicos, disminuyendo el costo operativo y el impacto ambiental.
Este caso demuestra que, con planificación técnica y decisiones estratégicas, las pequeñas granjas camaroneras pueden lograr avances significativos en eficiencia y sostenibilidad, incluso con recursos limitados.
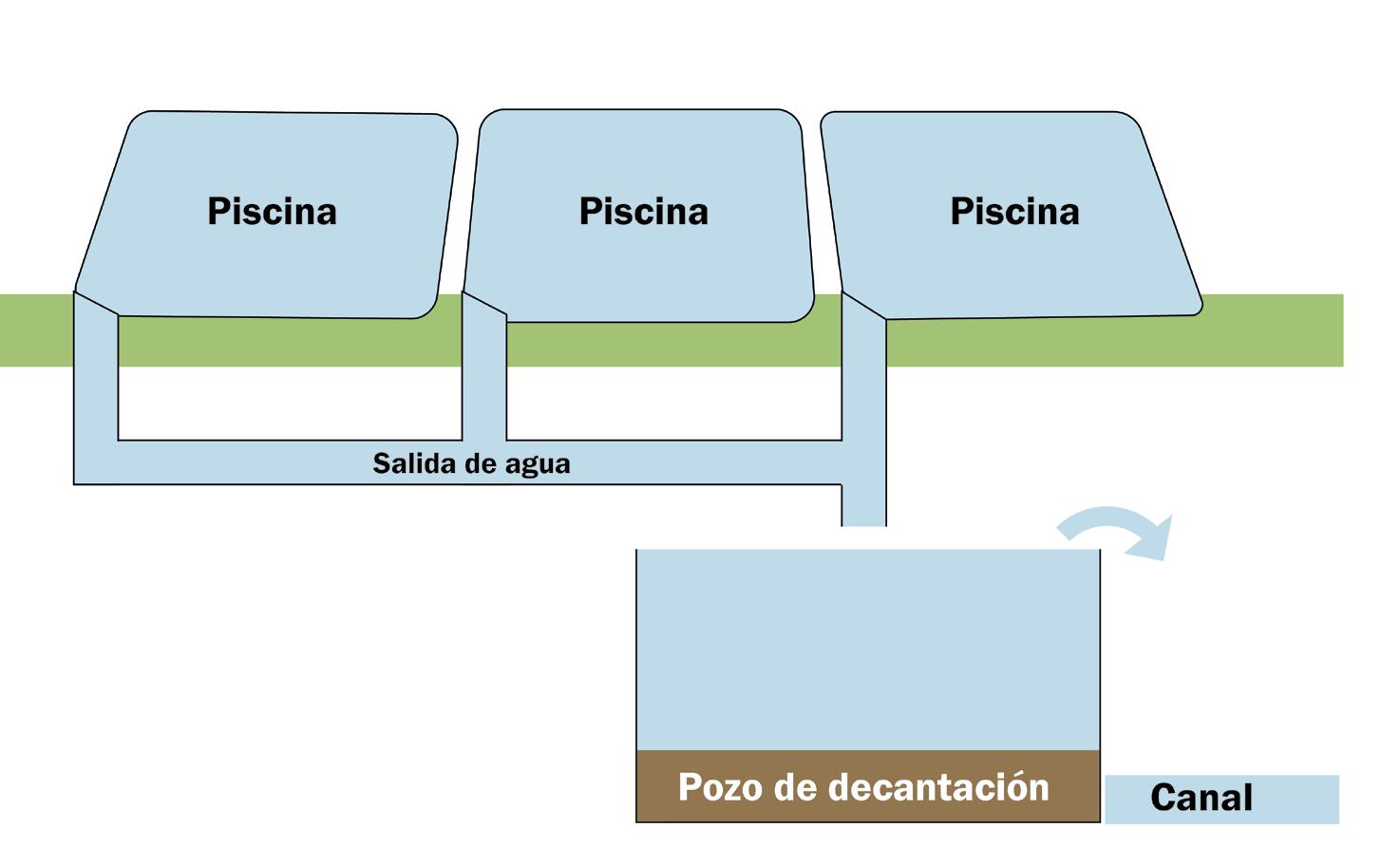
Importancia de los pozos de decantación en el manejo ambiental de granjas camaroneras Para cualquier tamaño de finca, uno de los elementos técnicos clave es la incorporación de pozos de decantación con batería de piscinas (Fig. 1), las cuales pueden ser construidas para dar servicio a una piscina en particular o para un grupo de estas. Estas estructuras, sencillas pero eficaces, cumplen una función crítica en el tratamiento primario de las aguas residuales generadas por el cultivo de camarón. Su implementación responde tanto a la necesidad de mejorar la calidad del efluente como a los objetivos de reutilización de recursos y reducción de la carga contaminante hacia el cuerpo receptor.
Los pozos de decantación se construyen en puntos estratégicos del sistema de drenaje, específicamente donde convergen las salidas de varias piscinas (usualmente tres a cuatro). El diseño básico incluye una fosa de aproximadamente 1.5 a 2 metros de profundidad, revestida con geomembrana, ladrillo, cemento, o incluso apisonado compacto si se trata de una solución temporal. Su función es permitir que el agua fluya de forma lenta a través de la estructura, favoreciendo la sedimentación de sólidos suspendidos como restos de alimento, heces y partículas orgánicas, antes de que el
efluente sea descargado o reutilizado.
Desde el punto de vista operativo, los pozos de decantación ofrecen múltiples ventajas. En primer lugar, centralizan el manejo de lodos, lo que reduce significativamente los costos y el esfuerzo necesario para su extracción. Al concentrar los residuos en un solo punto, se puede realizar su remoción periódica mediante bombas de lodos o retroexcavadoras, optimizando recursos. Además, estos lodos pueden ser aprovechados para la producción de compost, especialmente si se combinan con residuos vegetales provenientes de fincas agrícolas vecinas, generando un fertilizante útil para cultivos locales o incluso comercializable.
Desde el enfoque ambiental, su eficacia es notable: se estima que los pozos de decantación pueden retener entre un 80 y 90% de los sólidos suspendidos, lo que representa una reducción sustancial en la carga orgánica vertida al medio. Esto no solo mejora la calidad del agua en los canales y cuerpos receptores, sino que también contribuye a disminuir la eutrofización y el impacto negativo sobre los ecosistemas acuáticos adyacentes. Asimismo, al disponer de agua parcialmente clarificada, se facilita su reutilización en procesos como el prellenado de estanques para el siguiente ciclo productivo, favoreciendo el ahorro hídrico.
En el contexto de pequeños y medianos productores, los pozos de decantación representan una solución accesible, de bajo costo y alto impacto, que mejora la sostenibilidad del cultivo sin requerir grandes inversiones. Su adopción forma parte de un enfoque integral de bioseguridad y manejo ambiental responsable, y puede ser la base para obtener certificaciones de buenas prácticas o producción sustentable en el mediano plazo.
Conclusión
Las pequeñas granjas camaroneras enfrentan múltiples desafíos en su camino hacia una producción rentable y ambientalmente responsable. Sin embargo, mediante la aplicación de prácticas apropiadas, el uso eficiente de los recursos, la capacitación del personal y una cultura de mejora continua es posible transformar estos sistemas en unidades sostenibles y resistentes•
Para más información sobre este artículo, contactar a: lmariduena@cna-ecuador.com
Bibliografía Consultada
C. Boyd y A.McNevin. 2015. Aquaculture, Resource, and the Environment Ed. Wiley Blackwell
Fig. 1: Pozos de decantación con batería de piscinas
Aliados invisibles: Cómo los microorganismos transforman la productividad en acuacultura
Autores:
Dr. Dariano Krummenauer1
Dr. Otávio Augusto Lacerda Ferreira Pimentel2
Dra. Bianca de Oliveira Ramiro1
Dr. Wilson Wasielesky Jr.1
1 Instituto de Oceanografía, Universidad Federal de Río Grande - FURG, Brasil
2 Departamento de Pesca y Acuicultura, Universidad Federal Rural de Pernambuco, Brasil
darianok@gmail.com
Consultor Alltech
La producción de camarones se ha expandido significativamente en las últimas décadas, convirtiéndose en una actividad económica fundamental para diversas regiones costeras. Sin embargo, este crecimiento también ha traído consigo desafíos ambientales, sociales y económicos. Para garantizar su viabilidad a largo plazo, es esencial adoptar criterios holísticos que integren la rentabilidad económica con la sostenibilidad ambiental y social.
El modelo tradicional de cultivo de camarones, basado en alto consumo de agua, grandes extensiones de tierra y el uso intensivo de antibióticos, no es sostenible. Se necesitan nuevas estrategias de cultivo para satisfacer la demanda de sistemas más bioseguros con menor producción de residuos ricos en materia orgánica, que pueden afectar los ambientes acuáticos naturales. El uso de recirculación de agua y el manejo microbiológico del sistema son alternativas que promueven una mayor eficiencia en el uso del agua y el control de sus condiciones de calidad, además de ser ambientalmente responsables.
Los microorganismos juegan un papel fundamental en los procesos biogeoquímicos y en la colonización del tracto intestinal de los animales. Por lo tanto, su gestión debe recibir mayor atención para lograr el éxito en el cultivo del camarón. Por ello, este artículo destaca los beneficios del uso de microorganismos, bioflocs y biorremediación para promover una mayor eficiencia productiva, control de enfermedades y reducción de impactos ambientales.
El
papel de los microorganismos en la calidad del agua y la productividad
La gestión eficiente del agua, el control de la calidad del suelo y la reducción de contaminantes mediante tecnologías limpias son acciones esenciales para optimizar el uso de los recursos naturales. La implementación de sistemas de recirculación de agua (RAS), el uso de biofiltros, bioflocs y el tratamiento adecuado de los efluentes son ejemplos que ayudan a minimizar el impacto ambiental al mismo tiempo que aumentan la productividad.
El uso de estrategias RAS y bioflocs estimula
PRODUCCIÓN
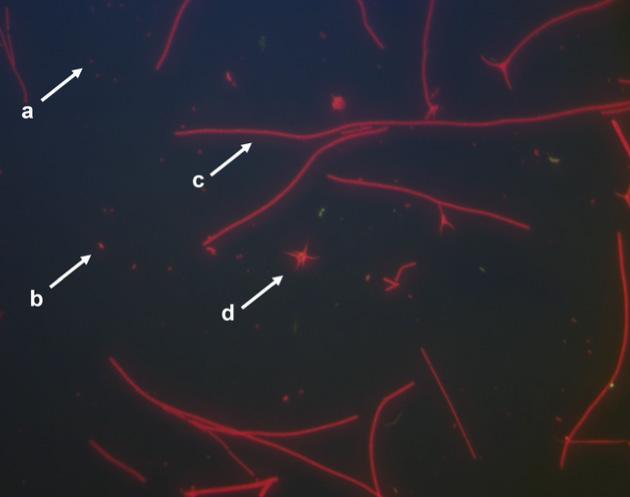
la actividad de las bacterias que actúan para eliminar los compuestos nitrogenados tóxicos que quedan disponibles en el sistema de cultivo a través de los residuos de alimento no consumidos y del proceso de excreción del camarón. El principal compuesto nitrogenado disuelto en el medio de cultivo es el amoníaco, el cual, dependiendo de variables como la temperatura, la salinidad y el pH del agua, ve alterado su nivel de toxicidad. En el sistema de bioflocs (Figura 1), el control del amoníaco se realiza a través de la comunidad de bacterias heterotróficas, que utilizan este compuesto para producir biomasa, creando agregados microbianos. Además de bacterias como cocoides (Figura 2a), Bacillus (Figura 2b) y bacterias filamentosas (Figura 2c), estos bioflocs también están compuestos de fitoplancton, zooplancton, hongos y materia orgánica. En este sistema y en el RAS, el control de los compuestos nitrogenados también se realiza a través del proceso de nitrificación. Este proceso lo llevan a cabo grupos de bacterias oxidantes del amoníaco que actúan para convertir este compuesto en nitrito, así como bacterias oxidantes de nitrito que convierten el nitrito en nitrato, que es la forma de nitrógeno menos tóxica para el camarón.
Además del fascinante impacto que las bacterias tienen en la calidad del agua, también pueden afectar la productividad del sistema de cultivo. Los bioflocs representan una fuente nutritiva de alimento suplementario para los camarones y están disponibles en todo momento suspendidos en la columna de agua. Por tanto, los bioflocs tienen el potencial de reducir las tasas de conversión alimenticia de los animales, aumentando la productividad de los sistemas de cultivo y mejorando el estado de salud de los camarones.
Biofilm y ciclo microbiano
El uso de biofilm en el cultivo de camarón es una estrategia que combina el control de la calidad del agua y la mejora del crecimiento del camarón. Los biofilms están compuestos por comunidades microbianas que se forman en superficies sumergidas. Generalmente, el crecimiento de biofilm es estimulado por la presencia de sustratos artificiales, los cuales tienen la función de aumentar el área disponible para la adhesión y crecimiento de bacterias nitrificantes. Estas bacterias tienen
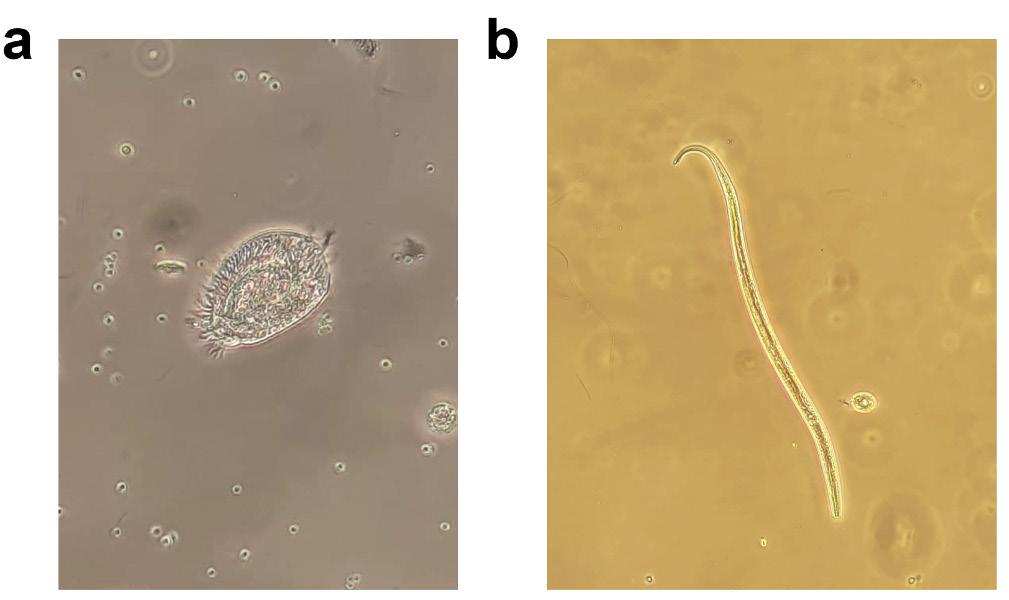
Figura 2. Microbioma de un cultivo de camarón con sistema biofloc. a: bacteria cocoide; b: Bacillus; c: bacterias filamentosas; d: ameba. Fuente: Otávio Augusto L. F. Pimentel.
Figura 1. Precriadero de camarón Penaeus vannamei con sistema biofloc. Fuente: Egidia Andrade (FURG).
preferencia por crecer en sustratos. En el cultivo intensivo de camarón con bioflocs, se recomienda utilizar sustratos artificiales en una proporción que varía entre el 200 y el 400% del área lateral del tanque de cultivo. Esta estrategia mejora el control de los compuestos nitrogenados, reduce la densidad relativa de almacenamiento y mejora las tasas de producción de cultivos.
Un aspecto importante en el uso de biofilm en el cultivo de camarón es el manejo de sustratos artificiales entre ciclos de cultivo. Es importante conocer la mejor estrategia de empaque de estos dispositivos para mantener la comunidad microbiana viable, de modo que no haya pérdidas en el control de los compuestos nitrogenados durante los sucesivos ciclos de cultivo. Un estudio reciente desarrollado por FURG abordó precisamente este tema. Se mantuvieron varios sustratos artificiales durante 30 días bajo diferentes condiciones (en sistema de biofloc con camarones [tratamiento control], sumergido en agua, sumergido en agua con aireación y expuesto al aire). Después de este período, los sustratos fueron insertados en el cultivo intensivo de camarón. Los resultados indicaron que mantener los sustratos sumergidos en agua no limitó el proceso de nitrificación entre ciclos de cultivo. Mantener los sustratos expuestos al aire tampoco afectó el proceso de nitrificación, provocando una recuperación de la comunidad bacteriana con el crecimiento de varios grupos de bacterias. Estas estrategias son prácticas y representan un ahorro energético al mantener los sustratos sumergidos en agua aireada.
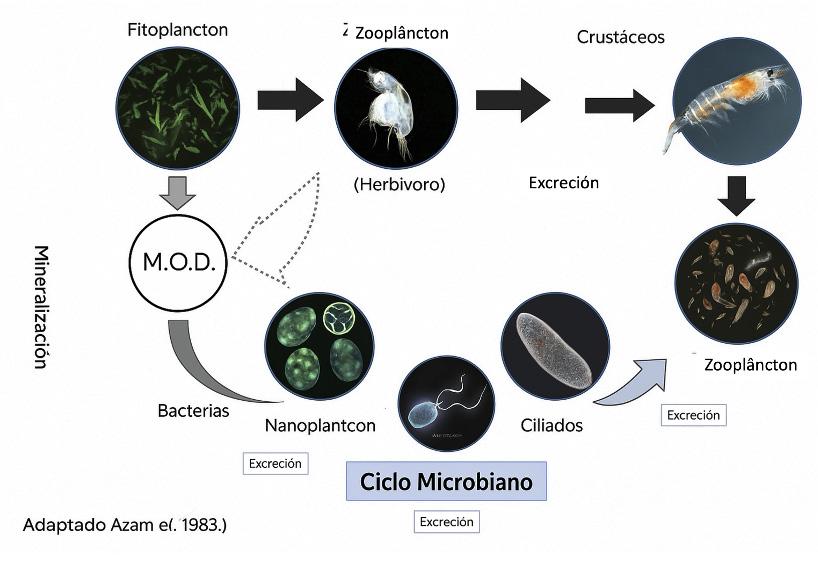
Tanto en los biofilms como en los bioflocs, un proceso clave para el correcto funcionamiento del sistema es el llamado “microbial loop” (Figura 3). Se trata de una vía trófica caracterizada por el reciclaje del carbono orgánico disuelto y su reintroducción en la red trófica. Este ciclo comienza con su asimilación por la comunidad bacteriana, que lo transforma en biomasa. Estas bacterias son consumidas por protozoos como amebas (Figura 2d), ciliados (Figura 4a) y flagelados. Estos microorganismos, a su vez, son consumidos por organismos más grandes, como rotíferos, nematodos (Figura 4b) y copépodos. Este efecto transfiere energía y materia orgánica disuelta a niveles
Figura 3. Concepto de “microbial loop” donde una serie de microorganismos surgen de la materia orgánica generada por el fitoplancton y otros organismos muertos. Fuente: Julio Zemor (FURG).
4. Protozoo ciliado (a) y nematodo (b) encontrados en cultivos de camarón utilizando el sistema biofloc. Fuente: Otávio Augusto L. F. Pimentel.
superiores de la cadena alimentaria. Este proceso microbiano ha sido observado en varios estudios con sistemas de biofloc desarrollados por FURG y es intrínseco al sistema de biofloc, contribuyendo a la reducción del uso de agua y alimento en el cultivo de camarón.
Reutilización del agua y reducción de impactos
Al estimular el crecimiento microbiano en el medio de cultivo, el uso del sistema biofloc
permite una reducción drástica en el uso de agua en comparación con los sistemas tradicionales. Estudios realizados en FURG han demostrado que la reutilización de agua (hasta el 100%) en sistemas biofloc no compromete el rendimiento zootécnico, siempre que el medio de cultivo esté adecuadamente madurado. Se debe mantener una relación carbono:nitrógeno entre 10 y 20:1 para que el sistema funcione correctamente. El ajuste de esta relación debe realizarse con una fuente de carbono
Figura
orgánico simple, como la melaza y la dextrosa.
Esta reutilización del agua puede realizarse a través del volumen de líquido según el tanque de cultivo o mediante la inoculación de bioflocs en el agua a una determinada concentración de sólidos suspendidos totales. Un estudio reciente realizado por FURG, que comparó el sistema biofloc con el sistema simbiótico fertilizado con diferentes salvados vegetales, utilizó un inóculo de bioflocs maduros a una concentración inicial de sólidos suspendidos totales de 5 mg/L. Los autores encontraron un buen control de los compuestos de nitrógeno en todo el cultivo con un pico reducido de amoníaco y nitrito, y un control rápido del amoníaco. Además, no hubo daño al crecimiento y supervivencia del camarón.
Esta reducción en el uso de agua es un factor que contribuye significativamente a reducir los impactos que la producción de camarón ocasiona sobre los ambientes naturales. La reducción de los intercambios y la posibilidad de reutilización del agua está en línea con los objetivos de desarrollo sostenible de consumo y producción responsables, garantizando la gestión y el uso eficiente de los recursos naturales finitos.
Probióticos e inmunidad
Otro aspecto esencial para la industria camaronera es la salud de los animales. El monitoreo constante de los parámetros fisicoquímicos del agua, la selección genética de especies más resistentes y el uso de buenas prácticas de manejo sanitario reducen la necesidad de antibióticos y el riesgo de enfermedades, lo que resulta en menores pérdidas y una mayor rentabilidad. Se estima que brotes de enfermedades virales o bacterianas pueden causar pérdidas de hasta un 40% en la producción si no se implementan controles adecuados.
Los camarones no tienen inmunidad adaptativa. Por lo tanto, la inoculación de microorganismos probióticos fortalece la inmunidad innata, mejora el microbiota intestinal y reduce la presencia de patógenos como Vibrio spp., aumentando la supervivencia de los lotes. Bacterias como las del género Bacillus, Lactobacillus y Pediococcus realizan exclusión competitiva,
compitiendo por espacio y nutrientes con microorganismos patógenos. De esta forma estos microorganismos ganan espacio y limitan el desarrollo de la expresión de patógenos en el medio de cultivo. Además, desempeñan un papel importante en la descomposición de la materia orgánica, y estabilización compuestos nitrogenados, como el amoníaco y el nitrito, lo que reduce significativamente la necesidad de renovación frecuente del agua.
Los microorganismos probióticos, además de su extraordinario papel como biorremediadores, son consumidos por los camarones y colonizan su tracto intestinal. De esta forma, ayudan a mejorar la digestibilidad de los alimentos, reduciendo el factor de conversión alimenticia y mejorando la respuesta inmune de los animales. El uso de estos microorganismos en la industria camaronera representa una revolución que aporta ganancias en todos los aspectos, mejorando el desarrollo sostenible de la acuicultura.
Consideraciones finales
El uso de estrategias de cultivo innovadoras como los bioflocs, biofilms y la inoculación de microorganismos biorremediadores en la acuacultura moderna permite alcanzar altos niveles de productividad con menor impacto ambiental, siendo herramientas clave para un modelo de cultivo más eficiente, resiliente y sostenible. Sin embargo, se debe considerar también el bienestar de las comunidades locales involucradas en la cadena productiva. Es importante garantizar condiciones laborales dignas, ofrecer capacitación técnica y promover una distribución más justa de los beneficios económicos. Integrar aspectos ambientales, sociales y económicos en la gestión de la actividad es esencial para construir un modelo más equilibrado, competitivo y resiliente frente a los desafíos del futuro•
Referencias
Abakari G, Luo G, Kombat EO. Dynamics of nitrogenous compounds and their control in biofloc technology (BFT) systems: A review. Aquaculture and Fisheries. 2021; 6: 441447. https://doi.org/10.1016/j.aaf.2020.05.005
Ballester ELC, Wasielesky W, Cavalli RO, Santos MRS, Abreu PC. Influência do biofilme no crescimento do camarãorosa Farfantepenaeus paulensis em sistemas de berçário. Atlantida. 2003; 25 (2): 117 - 122.
Decamp O, Cody J, Conquest L, Delanoy G, Tacon AGJ. Effect of Salinity on Natural Community and Production of Litopenaeus vannamei (Boone), within Experimental Zerowater Exchange Culture Systems. Aquaculture Research 2003; 34: 345-355. https://doi.org/10.1046/j.13652109.2003.00842.x
Decamp O, Warren A, Sánchez R. The role of ciliated protozoa in subsurface flow wetlands and their potential as bioindicators. Water Sci. Technol. 1999; 40: 91 – 98. https:// doi.org/10.1016/S0273-1223(99)00444-8
Hargreaves JA. Biofloc Production Systems for Aquaculture. Southern regional aquaculture center. 2013; 4503. Hlordzi V, Kuebutornye FKA, Afriyie G, Abarike ED, Lu Y, Chi S, Anokyewaa MA. The use of Bacillus species in maintenance of water quality in aquaculture: a review. Aquac. Rep. 2020; 18: 100503. https://doi.org/10.1016/j.aqrep.2020.100503
Krummenauer D, Cavalli RO, Ballester ELC, Wasielesky W. Feasibility of pacific white shrimp Litopenaeus vannamei culture in southern Brazil: effects of stocking density and a single or a double CROP management strategy in earthen ponds. Aquaculture Research. 2010; 41: 240-248. https:// doi.org/10.1111/j.1365-2109.2009.02326.x
Krummenauer D, Samocha T, Poersch L, Lara G, Wasielesky W. The reuse of water on the culture of pacifc white shrimp, Litopenaeus vannamei, in BFT system. J World Aquac Soc 2014; 45(1): 3–14. https://doi.org/10.1111/jwas.12093 Lara GR, Poersch LH, Wasielesky W. The quantity of artificial substrates influences the nitrogen cycle in the biofloc culture system of Litopenaeus vannamei. Aquacultural Engineering. 2021; 94: 102171. https://doi.org/10.1016/j. aquaeng.2021.102171
Pimentel OALF, Wasielesky W, da Silva NP, Borges LV, Krummenauer D. Fertilizing synbiotic system with different vegetable brans: effects on nitrification, plankton composition, and growth of Penaeus vannamei in the nursery phase. Aquaculture International. 2024; 32: 6407–6429 https://doi.org/10.1007/s10499-024-01471-x
Ramiro BO, Wasielesky W, Pimentel OALF, San Martin NP, Borges LV, Krummenauer D. Different management strategies for artificial substrates on nitrification, microbial composition, and growth of Penaeus vannamei in a superintensive biofloc system. Aquaculture. 2025; 596: 741853. https://doi.org/10.1016/j.aquaculture.2024.741853
Robles-Porchas GR, Gollas-Galván T, Martínez-Porchas M, Martínez-Cordova LR, Miranda-Baeza A, Vargas-Albores F. The nitrification process for nitrogen removal in biofloc system aquaculture. Rev. Aquacult. 2020; 12: 2228-2249. https://doi.org/10.1111/raq.12431
Samocha TM. Sustainable biofloc systems for marine shrimp – The Texas A&M AgriLife Research Experience. Baton Rouge: The World Aquaculture Society. 463p. 2019. Sanders, R.W. Protists: Flagellates and Amoebae. Encyclopedia of Inland Waters (2nd Edition). 2022; 630-638. https://doi.org/10.1016/B978-0-12-819166-8.00079-7
Para más información sobre este artículo, contactar a: darianok@gmail.com
Silva KR, Wasielesky W, Abreu PC. Nitrogen and phosphorus in the biofloc production of the pacific white shrimp, Litopenaeus vannamei. Journal of the World Aquaculture Society. 2013; 44 (1): 30 - 41. https://doi.org/10.1111/ jwas.12009
Silveira LGP, Rosas VT, Krummenauer D, Poersch LH, Wasielesky W. Comparison between horizontal and vertical substrate in shrimp super-intensive culture in bioflocs system. Aquacultural engineering. 2022; 96: 102218. https://doi.org/10.1016/j.aquaeng.2021.102218
Evaluación de la diversidad bacteriana como indicador de calidad en suelos de cultivos acuícolas y biorremediación para la mejora productiva
Autores:
Sonnya Mendoza L. Ph.D.
Laboratorio de Análisis NOVAGESTIÓN
Universidad Estatal Península de Santa Elena sonnya.mendoza@gmail.com
Ecuador, con más de 220 mil hectáreas dedicadas a la producción del camarón, se convirtió en el primer exportador mundial en el 2021, según información estadística de la Cámara Nacional de Acuacultura (CNA). Esta masificación e intensificación ha provocado la degradación de los suelos, y se ha determinado que la estrecha relación de eventos patológicos bacterianos o virales en la acuacultura del camarón están asociados a la calidad del suelo (Shafi et al., 2021), siendo la estructura de las comunidades bacterianas, del tracto digestivo del camarón y la composición de la microbiota del sedimento similares (Hou et al., 2018).
Es importante saber que la tendencia de incrementar las densidades para lograr una mayor recuperación en reducción de tiempos y biomasa cosechada, ha conllevado a la acumulación de alta carga orgánica, lo que se ha convertido en un problema que deteriora las interacciones bacterianas benéficas. Esto indiscutiblemente resulta en una calidad incontrolable de la relación agua-suelo, específicamente en sus procesos químicos como nitrito, amoníaco, sulfuros, fosfatos, etc.; esto, a su vez, impacta de manera negativa en la estabilidad de la microbiota de los ambientes acuáticos y dulce acuícolas, realizando una variación bacteriana tanto del agua, suelo y bacterias intestinales en el camarón, potencializando los patógenos causantes de enfermedades, reduciendo las posibles especies benéficas que ayudan a los ecosistemas, y ocasionando un estrés y posteriormente la muerte en los animales cultivados (Zhang et al., 2014).
Mientras tanto, la mayor abundancia de patógenos en el ambiente puede ingresar al intestino de un camarón estresado a través del sistema digestivo y luego alterar significativamente la microbiota intestinal; esto se relaciona de manera directa al suelo, vinculando con las tendencias del control de altas cargas de algas en las columnas de agua, lo que ha llevado a la práctica rutinaria de emplear floculantes, en su mayoría calcáreos, que precipitan todo lo suspendido en la columna hacia los suelos haciendo que reciban todas las descargas o desperdicios, y convirtiéndolo en un caldo de cultivo para protozoos, bacterias, virus y una verdadera “capa tóxica”, que incrementa el consumo
de oxígeno biológico y químico, nitrógeno amoniacal, nitrito y otras sustancias tóxicas, excediendo seriamente la cantidad estándar (Fazio et al., 2012; Fiorino et al., 2018). Si la capa tóxica no se ha eliminado a tiempo, agravará la carga de bacterias patógenas del hepatopáncreas del camarón, ocasionando una descamación de la mucosa intestinal e incrementando la inapetencia, lo que enferma al camarón y provoca su muerte.
En los sistemas intensivos y semiintensivos de cultivo de Penaeus vannamei, la microbiota presente en el agua, el suelo del estanque y el intestino del camarón cumple funciones críticas para el equilibrio ecológico, el metabolismo del hospedador y la prevención de enfermedades infecciosas. Esta comunidad microbiana incluye bacterias, arqueas y otros microorganismos beneficiosos, cuya estructura y dinámica pueden evaluarse mediante el uso de Unidades Taxonómicas Operativas (OTUs, por sus siglas en inglés), especialmente a través de herramientas de secuenciación masiva (Zhou et al., 2023).
Se ha demostrado que la diversidad y abundancia de estas OTUs microbianas varía significativamente dependiendo de factores ambientales como el tipo de manejo del sistema, la calidad del agua, el tipo de alimento y, sobre todo, la acumulación de materia orgánica (Xiong et al., 2019). El exceso de materia orgánica proveniente de restos de alimento no consumido, heces y material vegetal en descomposición genera condiciones anóxicas y un aumento de compuestos tóxicos como el amoníaco, lo que propicia el crecimiento de bacterias patógenas como Vibrio spp. y Aeromonas spp., reduciendo la riqueza microbiana y afectando la salud del camarón (Rungrassamee et al., 2014; Liu et al., 2022). El intestino del camarón, que alberga una microbiota diversa que actúa como una extensión de su sistema inmunológico, también se ve afectado por el desequilibrio en el ecosistema acuático. Estudios recientes mostraron que las disbiosis intestinales están directamente relacionadas con la presencia de condiciones ambientales adversas, especialmente con altas cargas de materia orgánica en el agua y sedimento, lo que favorece la colonización por OTUs patógenas (Gómez-Gil et al., 2020).
Ante este panorama, la aplicación de probióticos y bacterias biorremediadoras en el agua de cultivo y en la dieta del camarón ha emergido como una herramienta eficaz para restablecer el equilibrio microbiano. Cepas del género Bacillus, Lactobacillus y Pseudomonas entre otras han mostrado ser capaces de degradar materia orgánica, mejorar la calidad del agua, ser antagónicas con bacterias patógenas y estimular la respuesta inmune del hospedador (Zhang et al., 2021; Sharma et al., 2023). Estas bacterias probióticas pueden incrementar la abundancia y diversidad de OTUs beneficiosas y reducir significativamente las OTUs patógenas, mejorando los índices de supervivencia y crecimiento del camarón. Por tanto, un manejo adecuado del sistema acuícola debe contemplar no solo el control físico-químico del agua, sino también una gestión microbiológica activa a través del uso de probióticos, con el objetivo de mantener una microbiota saludable que contribuya al éxito del cultivo, biorremediando de manera constante los suelos donde mayor tiempo se alberga el camarón.
En los sistemas de acuicultura intensiva y semiintensiva, la microbiota asociada al suelo, agua e intestino de Penaeus vannamei desempeña un papel clave en la regulación del ecosistema, la digestión, la inmunomodulación y la prevención de enfermedades infecciosas. Esta microbiota se compone de una amplia gama de bacterias, cuya diversidad puede describirse mediante Unidades Taxonómicas Operativas (OTUs) a través de tecnologías de secuenciación de alto rendimiento (Zhou, Liu, & Wang, 2023). Sin embargo, factores como la acumulación excesiva de materia orgánica, derivada de restos de alimento y heces, pueden alterar profundamente estas comunidades microbianas, favoreciendo la proliferación de bacterias patógenas como Vibrio spp., Aeromonas spp. y Photobacterium spp. (Liu et al., 2022).
El desequilibrio microbiano en el intestino del camarón, generalmente inducido por estrés ambiental o contaminación orgánica, puede comprometer la integridad intestinal y aumentar la susceptibilidad a infecciones bacterianas (Rungrassamee et al., 2014; Gómez-Gil, Roque, & Turnbull, 2020). Frente a este escenario, la aplicación de probióticos
y bacterias biorremediadoras ha demostrado ser una estrategia eficaz para restaurar la composición de la microbiota, reducir los niveles de materia orgánica y limitar el crecimiento de bacterias oportunistas (Zhang et al., 2021).
Entre las bacterias probióticas más comúnmente utilizadas se encuentran diversas cepas del género Bacillus, incluyendo B. subtilis, B. licheniformis, B. megaterium, B. polymyxa y B. coagulans, las cuales han demostrado capacidad para degradar compuestos nitrogenados y competir eficazmente con patógenos intestinales (Chatterjee & Dey, 2023; Zhang et al., 2021). En aplicaciones prácticas, se ha reportado que B. subtilis, incorporado en el alimento a una dosis de 10⁶ UFC/g y suministrado al 5% del peso corporal por 56 días, mejora significativamente la ganancia de peso y la supervivencia de los camarones, al mismo tiempo que reduce la presencia de OTUs asociadas a patógenos (Chatterjee & Dey, 2023).
Otras bacterias benéficas identificadas en el suelo e intestino de camarones incluyen Lactobacillus plantarum, que ha sido utilizado como probiótico incorporado al alimento a una concentración de 10⁶ UFC/ mL en una proporción de 100 mL/kg de alimento, logrando mejorar la microbiota intestinal, reducir la carga de Vibrio spp. y aumentar la actividad enzimática digestiva (Sukenda et al., 2020). Asimismo, cepas de Pseudomonas balearica y P. stutzeri, aisladas de sistemas biofloc, han mostrado potencial como biorremediadores por su capacidad de oxidar materia orgánica y producir sustancias antimicrobianas contra bacterias entéricas (Utomo et al., 2023).
Exiguobacterium profundum ha emergido recientemente como un probiótico prometedor en acuicultura, al ser detectado en ambientes de cultivo biofloc y demostrar efectos positivos sobre la salud intestinal del camarón cuando se administra a 1 × 10⁶ UFC/g en el alimento durante 60 días (Utomo et al., 2023). Por su parte, Rhodovulum sulfidophilum, una bacteria fototrófica púrpura no azufrada, ha sido usada en el agua de cultivo a concentraciones de 10³ UFC/mL, reduciendo compuestos tóxicos y mejorando el crecimiento, al mismo tiempo
MANEJO
que suprime floraciones nocivas de algas y bacterias patógenas (Sharma et al., 2023).
Estos probióticos, al establecerse en el intestino y el sistema acuático, no solo desplazan físicamente a los patógenos a través de exclusión competitiva, sino que también modulan la expresión de genes inmunes del camarón, aumentando la producción de proteínas antimicrobianas y enzimas digestivas (Gómez-Gil et al., 2020). Además, su aplicación mejora la estabilidad de los parámetros físico-químicos del sistema, como la reducción del amonio, nitrito y compuestos orgánicos volátiles, que están directamente relacionados con la salud de la microbiota (Liu et al., 2022; Zhang et al., 2021).
En conjunto, la integración de cepas probióticas y bacterias biorremediadoras en el manejo microbiológico del cultivo de camarón constituye una herramienta poderosa para optimizar la producción, reducir el uso de antibióticos y mitigar los efectos negativos del exceso de materia orgánica. La selección racional de cepas, combinada con estrategias de dosificación y monitoreo basadas en datos microbiológicos (OTUs), ofrece un enfoque sostenible para el control de enfermedades en acuicultura intensiva.
Adaptaciones bacterianas en diferentes ambientes de cultivo han demostrado que la salinidad es un factor importante para la selectividad de estas a su presencia (Zhang et al., 2016), debido a que se piensa que la salinidad puede actuar de manera selectiva para un cierto grupo de bacterias; sin embargo, lo reportado en condiciones de monitoreo microbiológico y de secuenciación reportan una adaptabilidad no selectiva de ciertos géneros bacterianos sin importar este parámetro.
Este trabajo se fundamentó en la realización de análisis para conocer la abundancia bacteriana o microbioma en cultivos de agua dulce y salada; conocer la relación de estas bacterias en suelo, agua y camarón; medir las cargas bacterianas empleando productos biorremediadores y determinar factores de controles químicos en los suelos.
Materiales y Métodos
Muestreos microbiológicos
En este estudio se emplearon 10 estanques para la caracterización molecular de muestras de agua, suelo e intestinos, con una densidad de 200 mil animales por ha. durante 80 días de cultivo. Se tomaron muestras de agua, sedimento e intestinos de camarones en piscinas de agua dulce y agua marina, rotuladas como sedimento dulce, sedimento marino e intestino marino. Las placas fueron cultivadas a 32 °C por 24 horas y el conteo de colonias se realizó posterior a las 24 horas, tomando en cuenta las colonias que se encontraban distantes para evitar contaminación. Se seleccionaron las colonias más representativas de cada placa y se realizaron purificaciones en agar TSA con el rayado de estrías, y los recuentos bacterianos fueron cuantificados en UFC/g (Muthukrishnan et al., 2019)
Los suelos fueron enriquecidos y analizados empleando la metodología de siembra por vertido,10 gramos de suelos en 90 ml de medio enriquecido, con agares selectivos para Vibrios sp y Pseudomonas sp, las diluciones de siembra a 10-1, 102, para su posterior cuantificación según normativas estándares (BAM, 2024). Las muestras de intestino recolectadas fueron lavadas con agua destilada estéril y se realizó la extracción de 10 organismos. Se tomaron tubos Eppendorf colocando la muestra del hepatopáncreas hasta llegar al volumen de 1 gramo, y en la cámara de flujo laminar se realizó la siembra en disoluciones seriadas de 1:10, 1:100 y 1:1000. Las muestras recolectadas fueron sembradas por el método de vertido en agar.
Los suelos fueron sembrados en agar TCBS, Cromoagar vibrio, agar Cetrimide, y las cepas bacterianas aisladas, caracterizadas y criopreservadas para su posterior extracción de ADN.
Extracción de ADN
El kit de aislamiento de ADN fue el PowerSoil (MO BIO, EE.UU.), la técnica de secuenciación Ilumina de amplicones del gen 16S rRNA. Antes de la secuenciación, se realizaron PCR con el conjunto de cebadores bacterianos utilizando el conjunto de cebadores
universales 8F como cebador directo: AGAGTTTGATCCTGGCTCAG y 1492R como cebadores inversos: GGTTACCTTGTTACGACT (Kang et al., 2022), que amplifica V4- V5 . Regiones del gen 16S rRNA. El ADN genómico extraído se analizó mediante electroforesis en gel de agarosa al 1% y se determinó la concentración y pureza del ADN con un espectrofotómetro UV-vis Nano-Drop 2000 (Thermo Scientific, Wilmington, NC, EE.UU.). La secuenciación se hizo por el método de Sanger y el producto de PCR fue purificado y secuenciado en Macrogen Company - Seúl, Corea (dna.macrogen.com) por el método de Sanger. (Yen et al., 2021). De la misma manera se trabajó en la obtención de las unidades taxonómicas operativas según las instrucciones del fabricante y cuantificado con el fluorómetro Quantus™ (Promega, Madison, WI, EE.UU.) para la secuenciación de alto rendimiento (OTU) Illumina MiSeq, mediante el método Macrogen CompanySeúl, Corea (dna.macrogen.com) (Yen et al., 2021).
Análisis de materia orgánica
En paralelo se tomaron submuestras en 36 piscinas como seguimiento en paralelo en la misma granja donde se trabajaron las partes de extracción molecular, recuperando muestras compuestas de 454 gr. Estas piscinas tuvieron una aplicación de bacterias biorremediadoras y enzimas. El tiempo de análisis fue al inicio de la preparación de la siembra, considerada como antes del tratamiento, y muestras al término de las corridas, consideradas como después de los tratamientos. Análisis de carbón orgánico, materia orgánica, pH, así como otros parámetros químicos fueron realizados. La materia orgánica se determinó por el método de Walkley-Black AASHTO (2013).
Medidas de diversidad bacteriana
La diversidad fue estimada con los índices Shannon, Simpson y Chio, emulando el estimador de riqueza ACE. La prueba de Kruskall-Wallis fue aplicada para evaluar la diferencia de riqueza y uniformidad de las comunidades bacterianas entre las muestras. Adicionalmente, se aplicaron pruebas de abundancia diferencial (DA) significativamente. Los análisis estadísticos fueron realizados en R, con los paquetes: MicrobiomeAnalyst (Dhariwal et al., 2017) y Microbiome Marker (Yang, 2020).
Aplicación de productos probióticos y biorremediadores
Se emplearon en simultáneo tanto productos probióticos como enzimáticos en la preparación de los suelos de las piscinas camaroneras. Los probióticos biorremediadores fueron bioaumentados en tanques de 1 tonelada, usando 1000 gramos de bacteria con una composición de Bacillus subtilis, Lactobacillus Lactis, Nitrosomonas sp. y Nitrobacter sp. con exponente a la 10 ufc/g. por tonelada de agua; además, una aplicación de melaza al 20% en relación al volumen de agua empleada para su levante, que fue desinfectada con peróxido de hidrógeno en una dosis promedio de 1500 a 2000 ppm. También se aplicaron productos vitamínicos del complejo B para su crecimiento exponencial. La determinación de su cinética bacteriana se obtuvo entre las 16 a 18 horas con la concentración en ufc/ ml indicada por el proveedor para uso directo. La aplicación fue de 100 litros por hectárea aplicados 2 veces por semana por los 80 días, empleando mangueras inyectando directo al suelo, además de aplicación a la columna de agua.
Las enzimas fueron aplicadas también durante la corrida en los suelos, debidamente homogenizadas con agua para asegurar su distribución en dosis de 500 ml, 250 ml, 250ml, 250 ml por ha, distribuidas de manera equitativa en la preparación de los suelos humedecidos en la siembra, a los 15, 30 y 45 días al fondo, respectivamente.
Resultados
Los aislamientos bacterianos realizados a muestras de intestino, de agua y suelo mostraron una similitud en los géneros y especies de agua salada mediante
Figura 1. Porcentajes bacterianos identificados en intestinos de camarón y sedimento Nota: Caracterizaciones bacterianas identificadas bioquimicamente en suelo e intestino respectivamente.
aislamiento e identificaciones bioquímicas por API 20En, los cuales fueron similares a los géneros representativos de los intestinos y suelos analizados. El intestino tiene una mayor diversidad de especies y presencia balanceada de varias especies de Vibrio, especialmente V. alginolyticus y V. fluvialis. En el suelo, destacó claramente V. parahaemolyticus como la especie dominante, con menor diversidad en general. Esto sugiere que el ambiente intestinal favorece a ciertos Vibrios como V. fluvialis y V. vulnificus, mientras que el suelo puede ser más favorable para V. parahaemolyticus, razón de la vinculación con enfermedades bacterianas (Figura 1).
Los géneros que representaron una mayor abundancia de las diferentes muestras realizadas fueron de la especie Providencia; estas bacterias están relacionadas a géneros que pertenecen a la familia Enterobacteriaceae, y tienen importancia médica, ambiental, acuícola y suelos contaminados. Es una bacteria oportunista en larvas o juveniles inmunosuprimidos o bajo estrés; se aisló de intestinos y suelos como una posible infección secundaria, relacionada con suelos contaminados, con una vinculación claramente con el intestino, siendo responsables con procesos de degradación orgánica y transformación de nitrógeno.
De la misma manera, las Proteobacterias se encontraron mayormente en camarón, aunque también en suelo y agua, siendo estas relacionadas con las Pseudomonadota como
Pseudomonas aeroginosas, Psychrobacter spp. vibrionales como el Vibrio cholera, V. parahaemolyticus, V. vulnificus, Pasteurellas consideradas como oportunistas, pueden estar relacionadas también en procesos de nitrificación degradación en ambientes extremos (Figura 2).
Deltaproteobacterium fue encontrada tanto en agua, suelo y camarón. Entre sus principales géneros podemos citar al Delsufobibrio sp. Desulfobacter spp. Desulfobulbus spp. Geobacter spp, Myxococcus spp. intervienen en la reducción del sulfato, producen H2S, reportadas en ambientes marinos anóxicos, pantanos y aguas residuales, encargadas de reducir hierro y ciertos metales pesados, en variaciones ambientales que seguramente relacionan baja calidad de las producciones con poca diversidad bacteriana (Figura 2).
2. Porcentajes bacterianos cuantificados en suelo e intestino de camarón
Figura
Probiótico bioaumentado en el muro de las piscinas
Es importante determinar que aunque se trata de ambientes diferentes, cada hábitat está limitado por presencia de especies menos tolerantes, reflejando su papel como un ecosistema de transición con alto valor ecológico (Figura 2).
En un análisis comparativo de la abundancia relativa de bacterias en tres ambientes –agua, suelo y tracto intestinal de camarones– se observó un predominio claro de Fusobacteria, particularmente en agua (80%) y suelo (70%), mientras que su presencia en camarones fue relativamente menor (30%). Este grupo, asociado con ambientes anóxicos, sugiere un papel relevante en condiciones de baja oxigenación, especialmente en cuerpos de agua con alta carga orgánica (Kostic et al., 2013) (Figura 3).
En contraste, Proteobacteria fue el grupo más abundante en el tracto intestinal de los camarones (40%), superando su representación en suelo (15%) y agua (10%). Esta clase es reconocida por su versatilidad metabólica y su rol fundamental en la digestión y la respuesta inmunológica de organismos acuáticos (Gómez-Gil et al., 2014; Wang et al., 2020). Firmicutes mostró una presencia moderada, con una representación más notable en el suelo (10%), seguido por camarones (8%) y agua (5%), lo cual podría estar relacionado con su capacidad para formar esporas y tolerar condiciones adversas (Zhou et al., 2017).
Finalmente, Actinobacteria, aunque con baja representación (<2% en todas las muestras), desempeñan funciones relevantes en la degradación de compuestos recalcitrantes y producción de antibióticos naturales, lo que podría influir en la estabilidad microbiana del entorno (Barka et al., 2016) (Figura 3).
Estos resultados evidencian patrones de especialización ecológica y funcionalidad bacteriana, donde Fusobacteria dominan los entornos acuáticos y sedimentarios, mientras que Proteobacteria prevalecen en el microbioma del camarón. Esta información resulta crucial para entender las interacciones microbiota hospedero y sus implicaciones en sistemas acuáticos, especialmente en acuicultura intensiva (Figura 3).
Asimismo, se detectó la presencia
Porcentajes bacterianos cuantificados en suelo e intestino de camarón
Nota: Abundacia relativa de bacterias analizadas en agua, suelo e intestino de camarón.
de Sphingobium y Acinetobacter, con abundancias moderadas en suelo y camarón. Sphingobium, conocido por su capacidad para degradar compuestos xenobióticos y aromáticos (Mendes et al., 2015), ha sido reportado en ambientes contaminados y suelos agrícolas tratados con pesticidas (Yabuuchi & Kosako, 2005), lo que podría reflejar la historia de uso agrícola de las zonas aledañas. Por su parte, Acinetobacter, aunque con baja abundancia, ha sido identificado tanto en suelos como en intestinos de crustáceos, donde participa en procesos de fijación de nitrógeno y tolerancia a condiciones adversas (Chen et al., 2020; Li et al., 2017) (Figura 3).
Estos hallazgos sugieren una posible transferencia de bacterias edáficas y acuáticas hacia el microbioma intestinal del camarón, mediada por la interacción trófica con el sustrato, lo que resalta la importancia del manejo microbiológico en ambientes de cultivo multitrófico.
La microbiota intestinal está asociada a composiciones bacteriana del sedimento. a) La abundancia relativa de la microbiota intestinal, del sedimento y agua. b) El sedimento y el intestino comparten 2100 OTUs. El agua y el intestino comparten 290 OTUs, lo que confirma una relación directa entre los suelos y el contenido microbiano en los intestinos, lo que hace que la biorremediación sea considerada como la principal herramienta de la salud y éxito en una piscina camaronera.
El análisis del índice de riqueza estimada Chao evidenció diferencias notables en la diversidad bacteriana entre los tipos de sedimento analizados. El sedimento dulce presentó el valor más alto (Chao1 = 1400), indicando una mayor riqueza de especies bacterianas en comparación con las otras muestras. Esta alta diversidad puede atribuirse a la heterogeneidad ambiental característica de las zonas de transición entre agua dulce y marina, donde la variación en
Figura 3.
Figura 4. Interrelaciones bacterianas del agua, sedimentos e intestinos analizados..
parámetros fisicoquímicos como salinidad, oxígeno disuelto y contenido de materia orgánica generaron una amplia variedad de nichos ecológicos microbianos.(Cornejo et al.,2017).
En contraste, los análisis de los intestinos de camarones marinos registró una riqueza significativamente menor (Chao1 = 400), lo cual sugiere un ambiente más restrictivo o especializado, posiblemente asociado a condiciones físicas más estables y homogéneas, lo que limita la diversidad bacteriana presente. Esta baja riqueza puede estar relacionada con una menor heterogeneidad del hábitat o con la predominancia de ciertos grupos microbianos altamente adaptados al ambiente marino.
Por su parte, el sedimento marino submareal mostró un valor intermedio (Chao1 = 1100), lo que sugiere una riqueza bacteriana moderada, posiblemente influenciada por factores como la proximidad a zonas costeras, el tipo de sustrato o el flujo de nutrientes en la columna de agua. Este valor intermedio podría reflejar condiciones ambientales relativamente estables pero con cierto grado de productividad o influencia continental que favorece la presencia de comunidades bacterianas más diversas que en el intestino, aunque sin alcanzar la complejidad del sedimento dulce (Zhou et al., 2017).
El índice de Shannon, que combina tanto la riqueza de especies como la forma en que están distribuidas, proporcionó una visión más equilibrada de la diversidad bacteriana en los ecosistemas evaluados. En este estudio, se observaron valores de Shannon de 4.0 para el sedimento dulce, 4.0 para el intestino marino y un valor ligeramente más alto de 5.0 en el sedimento marino.
Estos resultados sugieren que, en general, los tres ambientes albergan comunidades bacterianas diversas y relativamente equilibradas. Sin embargo, el sedimento marino destaca por presentar una diversidad más alta, lo que puede atribuirse a la confluencia de condiciones variables propias de la zona de transición entre ambientes acuícolas dulces y marinos. En estos hábitats mixtos, la diversidad de nichos ecológicos y la fluctuación de factores como salinidad,
Figura 4.
Índices aplicados al nivel de OTUS. La diversidad microbiana se reduce ligeramente en agua de mar en comparación con el agua dulce, e independiente del ambiente la diversidad es mayor en los sedimentos. c) Índice Simpson de nivel de OTU Figura 5.
pH y disponibilidad de nutrientes crean un entorno propicio para una comunidad microbiana más compleja y diversa.
El análisis de la diversidad bacteriana mediante el índice de Simpson reveló diferencias notables entre los ambientes evaluados. Los sedimentos de agua dulce presentaron el menor valor (D = 0.015), lo que indica una alta diversidad y baja dominancia de grupos bacterianos. En contraste, los intestinos de camarones marinos mostraron un valor intermedio (D = 0.035), mientras que los sedimentos marinos exhibieron el valor más alto (D = 0.075), lo que sugiere una menor diversidad y mayor dominancia de ciertos OTUs en estos ambientes. Estos resultados evidencian una mayor heterogeneidad microbiana en ambientes dulceacuícolas en comparación con los sistemas marinos (Figura 5).
Los análisis realizados en 36 piscinas acuícolas evidenció una reducción significativa de la carga orgánica acumulada en el sedimento, con valores de carbono orgánico que fluctuaban entre 5% y 2% antes de la aplicación del tratamiento. Luego de la implementación de un protocolo regular basado en biorremediación bacteriana y aplicación enzimática, se observó una disminución promedio superior al 25% en los niveles de materia orgánica en todos los sistemas evaluados. Además, el proceso permitió una estabilización del pH del sedimento, con una reducción aproximada del 10% en la variabilidad de este parámetro, favoreciendo así la recuperación de la biodiversidad bacteriana autóctona.
Estos resultados refuerzan la efectividad de los tratamientos biológicos para la regeneración de suelos acuáticos y el mantenimiento de condiciones óptimas en sistemas de producción intensiva, alineándose con lo reportado por Vázquez-Sánchez et al. (2020) y Luo et al. (2022), quienes demostraron que el uso de microorganismos degradadores y enzimas específicas mejoraron la calidad del sedimento favoreciendo las comunidades microbianas más estables y funcionales. La sinergia entre frecuencia de aplicación y tiempo de contacto ha sido clave en el éxito de la estrategia empleada, permitiendo optimizar el proceso de biodegradación de residuos orgánicos acumulados en ambientes de cultivo acuático (Zhang et al., 2014; Gao et al., 2019),asegurando que la salud del camarón no se vea disminuida.
Discusión
La intensificación de la acuicultura, especialmente del cultivo de Penaeus vannamei, ha incrementado la densidad de organismos por unidad de área, lo que resulta en una acumulación excesiva de materia orgánica en los sedimentos de los estanques. Esta situación favorece la generación de ambientes anóxicos que alteran la dinámica microbiana y causan estrés fisiológico en los camarones, aumentando su susceptibilidad a enfermedades bacterianas como las causadas por Vibrio spp., Pseudomonas spp., Streptococcus spp. y miembros de la familia Enterobacteriaceae (Zhou et al., 2021). Según el estudio de He et al. (2021), los ambientes eutróficos con bajo contenido de oxígeno disuelto estimulan la proliferación de bacterias patógenas facultativas, lo que
compromete gravemente la supervivencia del cultivo.
En este contexto, el uso de probióticos y enzimas ha cobrado gran relevancia como estrategia de biorremediación y control preventivo en sistemas intensivos (Novriadi et al. 2023) demostraron que la combinación de probióticos con enzimas digestivas mejora significativamente la calidad del agua al reducir los niveles de amoníaco y nitrito, a la vez que aumenta la supervivencia y el crecimiento de los camarones. Esta sinergia entre microorganismos y enzimas facilita el análisis eficiente de la materia orgánica acumulada en los sedimentos, reduciendo la formación de zonas anóxicas y promoviendo un ambiente más oxigenado y saludable para el cultivo.
Además de su impacto directo en la calidad del agua, estas estrategias microbianas influyen indirectamente en la microbiota intestinal del camarón. Estudios recientes han confirmado la estrecha relación entre el microbioma del sedimento y la flora intestinal del camarón. La investigación de Wang et al. (2022) identificó que más del 70% de las unidades taxonómicas operativas (UTO) presentes en el intestino de P. vannamei coincidían con las del sedimento, lo que indica que una adecuada modulación del microbioma del suelo puede tener efectos positivos en la salud intestinal y la respuesta inmunitaria del organismo cultivado. En este sentido, la aplicación de cepas probióticas como Bacillus spp., Pediococcus spp., Nitrosomonas spp. y Nitrosospira spp. ha demostrado ser eficaz no solo en la
remineralización de nutrientes, sino también en el control de bacterias patógenas como Vibrio spp. (Sutthi et al., 2023).
La microbiota del suelo en cultivos de camarón presenta una abundancia bacteriana dominada principalmente por géneros como Bacillus, Vibrio, Pseudomonas, y otros pertenecientes a las clases Gammaproteobacteria, Alphaproteobacteria, Flavobacteriia, Bacilli y Actinobacteria (Mendoza, 2022; Mendoza, 2024). Estudios recientes indican que el género Bacillus fue predominante en suelos de lagunas camaroneras, junto con otros como Exiguobacterium, Acinetobacter, Arthrobacter y Planococcus (Mendoza, 2022). Además, se ha observado que en sistemas de cultivo intensivo, la comunidad
bacteriana incluye una mayor proporción de Vibrio, lo cual puede estar asociado a riesgos patógenos para los camarones (Robi, 2022). Otro aspecto clave es el control de la materia orgánica en los sedimentos. Reducir este componente por debajo del 5% se ha identificado como un parámetro esencial para mantener la estabilidad del sistema. Gao et al. (2022) concluyen que el uso de bacterias nitrificantes y probióticas. así como enzimas bacterianas. aceleran la oxidación de compuestos nitrogenados, mejorando las concentraciones de oxígeno disuelto y reduciendo significativamente las toxinas derivadas de la descomposición anaeróbica. Esto se traduce en un entorno más estable, resistente y productivo.
Finalmente, la implementación estratégica de aplicaciones tanto al suelo como a la columna de agua ha demostrado ser eficaz. Protocolos de campo recientes sugieren que dosis de 1 L/ha cada 15 días aplicadas al suelo y 1,5 L dos veces
por semana al agua, utilizando mezclas de bacterias biorremediadoras y enzimas, no solo mejoran los parámetros físico-químicos del sistema, sino que también ejercieron un efecto profiláctico contra patógenos. Este manejo integrado contribuyó a mejorar la conversión alimenticia, aumentar las tasas de supervivencia y, en consecuencia, a una producción más eficiente y sostenible (Tran et al., 2023). En nuestro estudio, la aplicación combinada de probióticos y enzimas constituyó una de las mejores estrategias para lograr mantener mejores resultados productivos.
Conclusiones
El estudio del microbioma en suelos de camaroneras revela que una alta biodiversidad microbiana es clave para el mantenimiento de un ecosistema funcional y resistente a enfermedades. La implementación de estrategias de manejo que incluyan la aplicación regular y continua de probióticos, enzimas y biorremediadores
favorece el equilibrio bacteriano, disminuye la acumulación de materia orgánica y mejora significativamente la calidad del agua y del suelo.
Asimismo, el monitoreo mediante índices de diversidad microbiana permite evaluar la salud del sistema y ajustar las prácticas productivas de manera más precisa y preventiva. Estas acciones, orientadas al fortalecimiento de la diversidad y riqueza bacteriana, no solo promueven el bienestar del camarón, sino que también contribuyen a una producción acuícola más sostenible, reduciendo la dependencia de tratamientos químicos y el riesgo de enfermedades. Por lo tanto, el manejo integral de bacterias benéficas debe considerarse un pilar esencial en la bioseguridad y eficiencia de los sistemas de cultivo de camarón, asegurando la inocuidad del producto final exportable•
Para más información sobre este artículo, contactar a: sonnya.mendoza@gmail.com
H2 para eficiencia térmica y reducción de emisiones en calderas y MCI
Autor:
Marco Ledesma eng.marco.ledesma@gmail.com
Stuttgart – Alemania
Coautor:
David Valdez dvaldez@estud.usfq.edu.ec
Quito – Ecuador
MANEJO
¿Qué se conoce? La industria térmica, con sus calderas de vapor, hornos de proceso y motores de combustión interna estacionarios, consume alrededor del 40% del gas natural y diésel industrial mundial, emitiendo 3,3 Gt CO2 año-¹. Múltiples gobiernos han fijado metas de reducción de gases de efecto invernadero (GEI) ≥ 50% al 2035; sin embargo, la sustitución total de activos por tecnología eléctrica o 100% H2 es económicamente inviable a corto plazo.
¿Qué problema persiste? Las mezclas bajas de hidrógeno (≤ 10% vol) solo recortan 3 al 7% de CO2 y se limitan al parque de gas natural; por encima del 30% H2, surgen desafíos de seguridad (velocidad de llama, retroceso) y materiales (fragilización). Es necesario un consenso cuantitativo sobre el punto de equilibrio entre impacto climático y viabilidad operativa.
Hipótesis y objetivo. Se plantea que la sustitución del 20% energético equivalente a la quinta parte de la demanda de BTU logra reducciones proporcionales de consumo de combustible y emisiones de CO2 sin comprometer la integridad de los sistemas. El objetivo es cuantificar dicha reducción para calderas y motores de combustión interna, contrastar los resultados con balances conceptuales y exponer barreras técnicas y económicas para su adopción.
Marco teórico
El hidrógeno posee un poder calorífico inferior (PCI) de 120 MJ kg-¹, casi el triple comparado con el diésel (42 MJ kg-¹). Su velocidad de llama (≈ 265 cm s-¹ a 1 atm, 25 °C) supera siete veces a la del metano, favoreciendo una combustión rápida y completa (Haraldsen & Meyer, 2022). En calderas, esto se traduce en un frente de llama más corto y temperaturas pico más altas; en motores, permite mezclas más homogéneas y una disminución del retardo de ignición (Li et al., 2024).
El balance estequiométrico muestra una relación lineal entre la fracción energética de H2 y la reducción de CO2 siempre que la eficiencia térmica permanezca constante (Kersting, 2014). No obstante, la termodinámica de radiación de llama puede variar en calderas, y el índice adiabático de mezcla altera el Rendimiento Térmico de Combustión (RTC).
Ledesma, M. (2025). Esquema de caldera diésel con inyección dual de H2 [Diagrama]
En MCI, la adición de H2 eleva el Índice de Mezcla Equivalente (Φ) y favorece la combustión en mezclas pobres, una estrategia clave para abatir los GEI y aumentar la eficiencia (Zhang et al., 2023). Sin embargo, la temperatura pico mayor incrementa la formación de NO2; de allí la importancia de recurrir a recirculación de gases de escape (EGR) o postratamiento catalítico (Patel & Ravi, 2023).
Metodología
Diseño
• Tipo de estudio: revisión sistemática y meta análisis de efectos aleatorios.
• Fuentes: Scopus, Web of Science, ScienceDirect, repositorios de congresos SAE e IEA (enero 2015-diciembre 2024).
• Valores cuantitativos pre y posmezcla de consumo fósil, CO2 y eficiencia.
• Simulaciones validadas con medición experimental (Δ < 5%).
Selección y extracción
De 1222 registros iniciales, 34 cumplieron los criterios (20 calderas, 14 motores). Los datos se normalizaron a 100% de carga nominal. Se empleó el método DerSimonian Laird; la heterogeneidad se midió con I².
Análisis complementario
Se construyó un balance teórico usando ecuaciones de combustión estequiométrica (Guerra, 2021) para verificar la proporción CO2 energía. Adicionalmente, se estimó la sensibilidad de los resultados a variaciones de eficiencia ± 3 p.p.
Resultados
Ahorro de combustible y eficiencia
n= Número de estudios o ensayos / I² Estadístico I Cuadrado / p.p Puntos Porcentuales
Reducción de CO2
Las aplicaciones reportan reducciones entre 19 y 26%, consistentes con la fracción energética. La dispersión en motores se explica por el aumento de eficiencia y las condiciones de carga parcial.
Contaminantes secundarios
• NO2: aumento promedio de 12% en calderas sin EGR; en motores diésel
con EGR > 35%, se observa -8%.
• CO y HC: caídas de 30 al 60% en motores.
• PM (diésel): -25% en servicio pesado.
Discusión
Los hallazgos confirman la proporcionalidad teórica 1:1 entre energía fósil desplazada y recorte de CO2, validando la hipótesis central.
La heterogeneidad moderada (I² ≈ 25%) sugiere robustez transversal a tecnologías, combustibles y potencias.
Motores diésel. La eficiencia gana 5 p.p. porque la ignición del diésel piloto inicia una propagación de llama acelerada por el H2, disminuyendo la duración de combustión y pérdidas por calor. Estos resultados concuerdan con pruebas en flotas norteamericanas (Guan et al., 2024), que informan reducciones de 24% de diésel a HEF ≈ 22%.
Calderas. La leve pérdida de eficiencia (< 2 p.p.) se atribuye a la menor radiación infrarroja del H2 frente al CH2 o diésel; no obstante, estudios de optimización de quemador mayor turbulencia y staging han recuperado totalmente el RTC (Stevens, 2025).
Limitaciones. Más del 60% de la bibliografía emplea H2 de laboratorio; los costos industriales de H2 verde influyen en la adopción.
Proyección económica. Con precios de electricidad renovable < 35 USD MWh-¹, la electrólisis alcalina integrada en caldera GN logra TIR > 14% (Torres et al., 2021). En transporte pesado, los sistemas on board de hidrólisis muestran ROI < 2 años para recorridos > 120 000 km año-¹.
Conclusiones
Sustituir el 20% del poder calorífico por H2 gaseoso reduce entre 19 y 21% de CO2 y combustible en calderas industriales sin rediseños mayores.
Motores diésel dual-fuel obtienen reducciones de 22 al 25% y mejoras de eficiencia de 5 p.p., ampliando el beneficio climático.
n= Número de estudios o ensayos / I² Estadístico I Cuadrado / p.p Puntos Porcentuales
Riesgos operativos (retroceso, NO2,
Fig.1
Sistema
fragilización) son manejables mediante EGR, válvulas antirretroceso y selección de aceros austeníticos.
El costo extra de producción y almacenamiento de H2 verde puede recuperarse en 3-8 años según precio de energía y factor de carga.
Se requiere normativa específica que establezca límites de mezcla energéticos (no volumétricos) y protocolos de seguridad en la cadena de suministro.
Recomendaciones
• Fase piloto. Desarrollar estudios primarios de factibilidad, estudios de balances de energía y emisiones con base en el consumo y características de los equipos.
• Motores de combustión Interna > 200 kW. Implementar sistemas dual-fuel con H2 en admisión y diésel piloto; EGR ≥ 35%.
• Desarrollo normativo. Definir estándares materiales y pruebas de hermeticidad para líneas de H2 a presión media (≤ 30 bar).
• I+D cooperativa. Evaluar blends de H2-biogás y H2-biodiésel para maximizar descarbonización neta•
Bibliografía
Chen, M., Albuquerque, F., Hirsch, J., & Tarr, R. W. (2024). Hydrogen co-firing in large- scale utility boilers. Fuel, 346, 128943.
Guan, Y., Zhang, X., & Li, H. (2024). Performance of a dual-fuel diesel engine at 20% hydrogen energy fraction. International Journal of Hydrogen Energy, 49(6), 3401- 3412.
Haraldsen, J., & Meyer, T. (2022). Flame speed of hydrogen–air mixtures under elevated pressure. Combustion and Flame, 240, 112137.
Kersting, L. (2014). Characterizing dual-fuel diesel–hydrogen combustion near the knock limit (Doctoral dissertation). North Dakota State University.
Li, C., Wang, J., & Zhao, H. (2024). Influence of hydrogen addition on cycle-to-cycle variations in a turbocharged gasoline engine. Energy Conversion and Management, 296, 118945.
Patel, S., & Ravi, M. (2023). NO2 mitigation strategies for hydrogen-assisted combustion. Energy, 263, 125598.
Stevens, R. (2025). Operational limits for 20% hydrogen blends in industrial steam generators. Journal of Cleaner Production, 441, 141873.
Torres, F., Gómez, P., & Martínez-Ruiz, A. (2021). Comparative life-cycle assessment of hydrogen pathways for process heat. Renewable & Sustainable Energy Reviews, 151, 111612.
Zhang, K., Liu, H., & Huang, Y. (2023). Combustion characteristics of hydrogen-enriched natural gas in spark-ignition engines. Applied Energy, 344, 120214.
(Se emplea formato APA 7. Todas las fuentes cumplen la regla de no mencionar marcas comerciales). Declaración de autoría
Marco Ledesma concibió la hipótesis, diseñó la metodología y lideró la discusión; David Valdés Ledesma realizó la búsqueda bibliográfica, el desarrollo de la programación para las interacciones de los resultados, y el modelo financiero y la redacción de resultados. Ambos autores revisaron y aprobaron la versión final.
Adición de suplementos promotores de salud en microdietas para postlarvas de camarón blanco (Penaeus vannamei): efectos sobre el crecimiento, la supervivencia y el estado de salud
Autores:
André Barreto 1,2,3,*
Diogo Peixoto 2,3
Carlos Fajardo 2,4
Wilson Pinto 5
Rui J. M. Rocha 1
Luis E. C. Conceição 5
Benjamín Costas 2,3
1 Riasearch Lda, Muelle del río Pardelhas, n◦ 21, 3870-168 Murtosa, Portugal
2 Centro Interdisciplinario de Investigaciones Marinas y Ambientales (CIIMAR), Terminal de Cruceros del Puerto de Leixões, Av.
3 Instituto de Ciencias Biomédicas Abel Salazar (ICBAS-UP), Universidad de Oporto, Rua de Jorge Viterbo
Ferreira n◦ 228, 4050-313 Oporto, Portugal
4 Departamento de Biología, Facultad de Ciencias del Mar y Ambientales, Instituto Universitario de Investigación Marina (INMAR), Campus de Excelencia Internacional del Mar (CEI·MAR), Universidad de Cádiz, 11510 Puerto Real, España
5 Sparos Lda, Marim Business Area, Lote C, 8700-221 Olhão, Portugal
andrebarreto@riasearch.pt
Publicación original https://www.mdpi.com/journal/animals
El camarón blanco del Pacífico (Penaeus vannamei) es actualmente la especie animal más representativa en la acuicultura, representando en 2020 una participación del 4.7% en la producción mundial [1]. Para satisfacer la demanda del mercado, la producción de larvas y postlarvas (PL) de camarón blanco en los laboratorios ha aumentado considerablemente. Sin embargo, los problemas en la larvicultura pueden tener un enorme impacto en el rendimiento del camarón a largo plazo, afectando la producción posterior de juveniles y adultos de alta calidad. Las etapas iniciales del desarrollo se asocian frecuentemente con un crecimiento subóptimo, una alta dispersión de tallas y una baja supervivencia debido al canibalismo y a la menor resistencia a patógenos oportunistas. Esto último se debe a una alta dependencia de condiciones zootécnicas y nutrición óptimas, ya que el camarón carece de una respuesta inmunitaria adaptativa y depende exclusivamente de su sistema inmunitario innato para mantener un buen estado de salud y evitar brotes de patógenos que pueden tener consecuencias desastrosas para la producción y pérdidas económicas significativas [2–4]. Además de que el camarón no puede vacunarse debido a la falta de un sistema inmunitario adaptativo, el uso de antibióticos en la acuicultura es limitado debido a preocupaciones inherentes a la seguridad alimentaria, problemas ambientales y el aumento de la resistencia a los antimicrobianos [5].
En consecuencia, el uso de aditivos funcionales en dietas para estimular el sistema inmunitario del camarón se ha estudiado como una alternativa profiláctica y se considera una estrategia fundamental para superar las limitaciones del cultivo intensivo de camarón. Los ácidos orgánicos, los extractos de plantas y algas, los nucleótidos, los aminoácidos funcionales, las vitaminas y los compuestos inmunoestimulantes naturales, como los β-glucanos, se han estudiado exhaustivamente en dietas para peces y crustáceos, ya que pueden mejorar el crecimiento, la supervivencia, el estrés y la resistencia a enfermedades, como fue recientemente revisado por Dawood et al. [6]. Su potencial para ser incluidos en dietas para camarón blanco juvenil y/o adulto también ha sido demostrado en cierta medida por
ALIMENTACIÓN
varios autores [7–18]. Sin embargo, existen muchos menos estudios disponibles sobre el uso de estos suplementos en dietas para las etapas iniciales del desarrollo del camarón blanco. Existe evidencia de que la vitamina C suplementada en dietas para PL puede ser un antioxidante eficaz a nivel tisular [19]. Recientemente, se han verificado aumentos en el rendimiento del crecimiento, la actividad de las enzimas digestivas y la mejora del estado inmunitario al incorporar mezclas comerciales de prebióticos y probióticos en dietas para PL [20-22].
Por lo tanto, las soluciones nutricionales innovadoras que mejoran el desarrollo y la resistencia al estrés y a los factores patógenos durante estas etapas críticas, y, por consiguiente, mejoran la calidad del camarón en las fases posteriores de producción, tienen un enorme potencial para impulsar el éxito del cultivo. Por lo tanto, este estudio tuvo como objetivo evaluar los efectos de varios nutrientes/aditivos beneficiosos para la salud (p. ej.: vitaminas C y E, β-glucanos, taurina y metionina) suplementados en microdietas inertes sobre el rendimiento del crecimiento y el estado de salud de las postlarvas de camarón blanco.
Materiales y Métodos
Tratamientos Dietéticos
Se evaluaron cuatro microdietas experimentales por triplicado. Se formuló una dieta como control positivo (PC) para satisfacer los requerimientos nutricionales de las postlarvas de camarón blanco, conteniendo 515 g kg−1 de la mezcla patentada de proteína marina SPAROS, 160 g kg−1 de la mezcla patentada de proteína vegetal SPAROS, 103 g kg−1 de hidrolizado de proteína de pescado, 19 g kg−1 de aceite de pescado, 28 g kg−1 de fosfolípidos marinos y 57 g kg−1 de la premezcla patentada de vitaminas y minerales SPAROS. En los tratamientos restantes, se utilizaron tres variantes experimentales basadas en el PC, difiriendo únicamente en la formulación de los ingredientes como se detalla a continuación: (1) disminuyendo los niveles de inclusión de la premezcla de vitaminas y minerales en 7 g kg−1 para reducir el contenido de vitaminas C y E en la dieta control negativo (NC); (2) suplementando 5 g kg−1 de taurina y 10 g kg−1 de metionina para aumentar los niveles de ambas moléculas en
la dieta T + M; y (3) suplementando la dieta PC con 1 g kg−1 de β-(1, 3)/(1, 6)-glucanos (BG) de Saccharomyces cerevisiae. La formulación de las dietas experimentales se puede consultar en la Tabla 1, y su composición proximal fue analizada por Eurofins Food Testing Portugal siguiendo sus procedimientos estándar (Tabla 2).
Todas las dietas se elaboraron en las instalaciones de Sparos Lda (Olhão, Portugal), utilizando la extrusión a baja temperatura como principal proceso de producción, de la siguiente manera: mezcla de los ingredientes en polvo según la formulación deseada mediante un mezclador de doble hélice; molienda en un molino de martillos micropulverizador (SH1, Hosokawa-Alpine, Augsburgo, Alemania); adición de la fracción de aceite; humidificación y aglomeración mediante extrusión a baja temperatura (Dominioni Group, Lurate Caccivio, Italia); secado de pellets resultantes en un horno de convección (OP 750-UF, LTE Scientifics, Oldham, Reino Unido) durante 4 h a 60 °C; desmenuzado (Neuero Farm, Melle, Alemania), y tamizado hasta alcanzar los rangos de tamaño deseados.
Cultivo y muestreo de camarón
Las postlarvas de camarón blanco (PL16), procedentes de Blue Genetics (La Paz, México), se cultivaron durante 18 días en las instalaciones de Riasearch Lda (Murtosa, Portugal). Los camarones se distribuyeron aleatoriamente en 12 tanques de aproximadamente 50L, conectados a un sistema de recirculación de agua. Cada tanque contaba con 200 individuos con un promedio de 9 mg de peso húmedo. Estos se mantuvieron bajo un fotoperiodo de 12h de luz y 12h de oscuridad, y se alimentaron ad libitum con comederos automatizados que suministraban ocho comidas al día. Los comederos se limpiaron a diario y se cargaron con cantidades de alimento ajustadas según la observación de los tanques y la presencia o ausencia de restos del día anterior. El tamaño del alimento fue de 400 a 600 µm durante la primera semana y de 600 a 800 µm durante el resto del período de alimentación. La temperatura del agua se mantuvo a 28.8 ± 0.3 °C, la concentración de oxígeno disuelto a 7.5 ± 0.4 mg L−1, la salinidad a 20.3 ± 1.2, el pH a 7.96 ± 0.1, el NH3 a 0.0 ± 0.0 mg L−1 y el NO2 a 0.36 ± 0.3 mg L−1
Al inicio del experimento, se seleccionó aleatoriamente un total de 60 camarones del stock inicial y se pesaron por grupo para la determinación inicial del peso húmedo. Al final del experimento, todos los camarones se pesaron en grupos de 20 individuos para la determinación final del peso húmedo de cada tanque. Además, se seleccionaron aleatoriamente 40 camarones de cada tanque para el análisis de estrés oxidativo y parámetros inmunitarios, y 10 camarones para el análisis de expresión génica. Los camarones se mantuvieron en ayunas durante 12 h antes de los muestreos para asegurar que sus intestinos estuvieran vacíos al momento de la recolección. Los camarones muestreados para estrés oxidativo y parámetros inmunitarios se almacenaron a -80 °C para su posterior análisis. Los camarones muestreados para análisis de biología molecular se mantuvieron en RNAlater (Sigma, St. Louis, MO, EE. UU.) en una proporción de volumen >1:5, a 4 °C, durante 24h antes de ser almacenados a -20 °C. Al final del experimento se evaluaron la tasa de crecimiento relativo (RGR), la tasa de conversión alimenticia (FCR) y la supervivencia de cada tratamiento.
Estrés oxidativo y biomarcadores relacionados con la inmunidad
Preparación de la muestra
Al final del ensayo se pesaron y homogeneizaron 40 postlarvas enteras de camarón blanco de cada tanque muestreado en grupos cuádruples de 10 individuos para el análisis de estrés oxidativo y parámetros inmunitarios. Se añadió tampón de fosfato de potasio (0.1 M) a cada grupo en una proporción de 1/10 (p/v) seguido de ua homogeneización utilizando un instrumento de dispersión de alto rendimiento (SilentCrusher M, Heidolph Instruments, Schwabach, Alemania). Se reservó una alícuota para la peroxidación lipídica (LPO) con hidroxitolueno butilado antes de la centrifugación. Después de la centrifugación (5500 rpm durante 20 min), se recogió el sobrenadante de la muestra y se distribuyó en alícuotas separadas para los parámetros de estrés oxidativo y los parámetros inmunes. Los 10 camarones restantes muestreados para el análisis de biología molecular se homogeneizaron en NZYol (Nzytech, proporción p/v según las instrucciones del fabricante) utilizando un homogeneizador
de tejidos Precellys 24 (Bertin instruments, Montigny-le-Bretonneux, Francia).
Determinación de biomarcadores de estrés oxidativo
Se determinaron las actividades de catalasa (CAT), peroxidación lipídica (LPO) y glutatión total (tGSH), así como el contenido de proteínas totales, en las muestras homogeneizadas. Las proteínas totales se midieron utilizando el kit de ensayo de proteínas Pierce™ BCA, según lo descrito por Costas et al. [23]. Las muestras se diluyeron en tampón de fosfato potásico (01 M; pH 7.4) y se utilizó albúmina sérica bovina (BSA, 2 mg mL−1) como estándar. Posteriormente, se sembraron por triplicado 25 µL de cada muestra diluida y estándares y se leyeron a 562 nm en un lector de microplacas Synergy HT. Los resultados se calcularon mediante una curva estándar y se expresaron en mg mL−1
Los niveles de actividad de CAT se determinaron midiendo la disminución de la concentración de peróxido de hidrógeno (H2O2 30%, Sigma), según lo descrito por Clairborne [24]. La mezcla de reacción se compuso de tampón de fosfato potásico (0.05 M, pH 7.0) y H2O2 (30%) como sustrato. Se añadieron 10 µL de homogeneizado a la mezcla de reacción, alcanzando un volumen total de 300 µL. La absorbancia se leyó a 240 nm en microplacas UV durante 2 min (1 lectura cada 15 s) en un lector de microplacas Synergy HT. Los resultados se expresaron como unidades de enzima por miligramo de proteína total (U mg−1 de proteína). Una unidad de enzima es la cantidad de enzima necesaria para catalizar un micromol de sustrato por minuto.
La LPO endógena se evaluó midiendo las sustancias reactivas al ácido tiobarbitúrico (TBARS), previniendo la oxidación lipídica artificial mediante la adición de butilhidroxitolueno (4%; Sigma) [25]. Las muestras de homogeneizado se incubaron durante 60 min a 100 °C con 100 µL de solución de ácido tricloroacético al 100% y 1 mL de ácido 2-tiobarbitúrico al 0.73% (Sigma), clorhidrato de trizma (Sigma) y ácido dietilentriaminopentaacético (Fluka) en microtubos de poliestireno. Posteriormente, se centrifugaron durante 5 min a 11 500 rpm y se añadió el sobrenadante (200 µL) a los
Tabla 1. Formulación dietética de las dietas experimentales utilizadas para el cultivo de PL de camarón blanco (P. vannamei) durante 18 días.
Ingredientes (g kg-1)
Mezcla de proteínas marinas1
Hidrolizado de proteína de pescado2
Mezcla de proteínas vegetales3
Celulosa4
Aceite de pescado5
Fosfolípidos marinos6
Lecitina7
Vitaminas y minerales8
Colesterol9
Antioxidante10
Fosfato monoamónico11
β-(1,3)/(1,6)-glucanos12
DL-metionina13
Taurina14
1Producto patentado para camarones: 37% de proteína cruda, 5% de grasa cruda — SPAROS, Portugal
2 Sopropêche, Francia
3 Producto patentado para camarones: 13 % de proteína cruda, 1 % de grasa cruda — SPAROS, Portugal
4 Disproquimica, Portugal
5 Sopropêche, Francia
6 Triple nine, Dinamarca
7 Lecico, Alemania
8 Premezclas/productos patentados para camarones — SPAROS, Portugal
9 Carbogen, Países Bajos
10 Kemin Italia
11 Timab Ibérica, España
12 MacroGard—Orffa, Países Bajos
13 Premix—Especialidades Agrícolas e Pecuárias Lda, Portugal
14 Producto patentado para peces marinos y camarones—SPAROS, Portugal
pocillos de la microplaca. La absorbancia se leyó a 535 nm y los resultados se expresaron como nmol g wt−1
El contenido total de glutatión en las muestras de homogeneizado de postlarvas se midió con base en la oxidación del glutatión por el ácido 5,5’t-ditiobis-(2-nitrobenzoico) (DTNB; Sigma), según lo descrito por Rodrigues et al. [26]. Las muestras se diluyeron en tampón de fosfato potásico (0.1 M, pH 7.4) para obtener 0.7 mg mL−1 de proteína. Posteriormente, se añadieron 50 µL de cada muestra diluida a los pocillos de la microplaca, seguidos de la adición de 250 µL de una solución de reacción compuesta por DTNB, tampón de fosfato potásico (0.1 M, pH 7.4), NADPH (sal tetrasódica reducida de ß-nicotanimida adenina dinucleótido 2t-fosfato; Alpha Aesar) y glutatión reductasa (Sigma). La absorbancia se leyó a 412 nm durante 3 min (una lectura cada 20 s) en un lector de microplacas Synergy HT y los resultados se expresaron como nmol mg de proteína−1
Análisis de parámetros inmunitarios
Se determinaron las actividades de
lisozima, profenoloxidasa y bactericida en las muestras homogeneizadas. La actividad de la lisozima se midió mediante un ensayo turbidimétrico, según lo descrito por Costas et al. [27]. Brevemente, se preparó una solución de Micrococcus lysodeikticus (0.25 mg mL−1, tampón de fosfato de sodio 0.05 M, pH 6.2) y se añadieron 40 µL de las muestras homogeneizadas y 130 µL de esta suspensión a una microplaca, alcanzando un volumen final de 170 µL. La reacción se llevó a cabo a 25 °C y se midió la absorbancia (450 nm) después de 0.5 y 30 minutos en un lector de microplacas Synergy HT. La lisozima liofilizada de clara de huevo de gallina (Sigma) se diluyó en tampón de fosfato de sodio (0.05 M, pH 6.2) y se utilizó para desarrollar una curva estándar. La cantidad de lisozima en la muestra se calculó mediante una curva estándar. La lisozima se expresó como µg mg proteína−1
La actividad de la profenoloxidasa se midió espectrofotométricamente utilizando L-DOPA (L-3,4-dihidroxifenilalanina) como sustrato y tripsina (Sigma) como activador, siguiendo el método descrito por Ji et al. [28] con
ALIMENTACIÓN
modificaciones. Se diluyeron muestras de homogeneizado de 50 µL en 100 µL de solución de tripsina (0.1% en solución de cacodilato) en una en solución de cacodilato). La absorbancia se midió cada minuto durante 5 min a 490 nm utilizando un lector de microplacas Synergy HT. Los resultados se calcularon mediante la ley de Beer-Lambert y el coeficiente de extinción molar de la L-DOPA (3700). Los resultados se expresaron como unidades de profenoloxidasa mL−1 de muestra.
Se utilizó Vibrio harveyi en el ensayo de actividad bactericida. Las bacterias con crecimiento exponencial se resuspendieron en HBSS estéril y se ajustaron a 3.1 × 109 cfu mL−1. La siembra de diluciones seriadas de las suspensiones en placas TSA-2 y el recuento del número de cfu tras la incubación a 22 °C confirmaron la concentración bacteriana del inóculo. La actividad bactericida de las muestras homogeneizadas se determinó posteriormente siguiendo el método descrito por Machado et al. [29]. Brevemente, se añadieron 60 µL de muestras homogeneizadas a una placa de 96 pocillos en forma de U. Se añadió HBSS a algunos pocillos en lugar de muestras homogeneizadas, lo que sirvió como control positivo. A cada pocillo se añadieron 20 µL de V. harveyi (3.1 × 109 cfu mL−1) y la placa se incubó durante 2.5 h a 25 °C. A cada pocillo se añadieron 25 µL de bromuro de 3-(4,5 dimetil-2-il)-2,5-difenil tetrazolio (1 mg mL−1; Sigma) y se incubó durante 10 min a 25 °C para permitir la formación de formazán. A continuación, las placas se centrifugaron a 2000 × g durante 10 min y el precipitado se disolvió en 200 µL de dimetilsulfóxido (Sigma). La absorbancia del formazán disuelto se midió a 560 nm. La actividad bactericida se expresó como un porcentaje calculado a partir de la diferencia entre las bacterias supervivientes y el número de bacterias de los controles positivos (100%).
Análisis de la expresión génica
La extracción de ARN se realizó con el kit de aislamiento de ARN total NZY (NZYTech, Lisboa, Portugal), siguiendo las instrucciones del fabricante. La concentración y pureza del ARN se analizó mediante espectrofotometría con DeNovix DS-11 FX (Wilmington, NC, EE. UU.). La concentración del ARN varió de 123.9 a 2180.7 ng µL−1 y la proporción
Tabla 2. Composición proximal de las dietas experimentales utilizadas para el cultivo de PL de camarón blanco (P. vannamei) durante 18 días.
Materia seca (MS, %)
Proteína bruta (% MS)
Grasa bruta (% MS)
Fibra (% MS)
Cenizas (% MS)
Fósforo (% MS)
Energía (MJ/kg MS)
Vitamina C (mg/kg MS)
Vitamina E (mg/kg MS)
Taurina (g/100 g MS)
Metionina (g/100 g MS)
260:280 de 1.99 a 2.17, respectivamente. La integridad de las muestras de ARN se verificó mediante un gel de agarosa al 2 %. El ADNc se obtuvo con el kit de síntesis de ADNc de primera cadena NZY (NZYTech). Este paso se utilizó para estandarizar la concentración de las muestras. La transcripción inversa se llevó a cabo en un termociclador Veriti DX de 96 pocillos (Applied Biosystems, Waltham, MA, EE.UU.), utilizando 4.4 µL de ADNc diluido (20 ng µL−1) mezclado con 5 µL de NZYSpeedy qPCR Green Master Mix® (NZYTech) y 0.3 µL (10 µM) de cada cebador específico en un volumen final de 10 µL.
Se realizó PCR cuantitativa en tiempo real, por duplicado para cada muestra, utilizando un sistema de detección de PCR en tiempo real CFX384 Touch (Biorad, Hercules, CA, EE.UU.). Se seleccionaron y analizaron nueve genes debido a su papel en la respuesta inmunitaria. Se evaluó la eficiencia del primer para cada gen (Tabla 3). Las condiciones de ciclado fueron las mismas entre los diferentes genes, consistentes en un ciclo de 95 °C durante 10 min, seguido de 40 ciclos de 2 pasos de 95 °C durante 15 s y 62 °C durante 1 min, con un ciclo final a 95 °C durante 1 min, seguido de 35s a 62 °C y finalizando a 95 °C durante 0.5 s. Se utilizó el método de Pfaffl [30] para realizar análisis de expresión génica, y los genes diana se normalizaron utilizando bactn y rpl-8 como mantenimiento.
Análisis de data
La tasa de crecimiento relativo (RGR, % peso día−1) se calculó de la siguiente manera: RGR = (eg − 1) × 100, donde e = exponencial y g = (lnWf − lnWi) × t−1. Wf y Wi corresponden al peso final e inicial, respectivamente. El índice de conversión alimenticia (FCR) se calculó de
la siguiente manera: FCR = (Fi/Wg), donde Fi corresponde al alimento administrado (g) y Wg a la ganancia de peso media (g). La supervivencia se expresó como porcentaje y se calculó de la siguiente manera: S = (Lf/ Li) × 100, donde Li y Lf corresponden al número inicial y final de postlarvas en los tanques, respectivamente. Las diferencias en el rendimiento del crecimiento, el FCR, la supervivencia, el estrés oxidativo, el estado inmunitario y la expresión génica entre los tratamientos dietéticos se evaluaron mediante ANOVA de una vía, seguido de pruebas de comparación múltiple de Tukey. Se utilizaron pruebas de análisis de varianza unidireccional de Kruskal-Wallis, seguidas de pruebas de comparación por pares de Wilcoxon, cuando los datos no cumplían con los supuestos del ANOVA unidireccional. Los resultados se expresaron como media ± desviación estándar (SD). En los resultados expresados como porcentaje, se realizó una transformación de arcoseno antes de cualquier prueba estadística: T = ASIN (SQRT (valor/100)). El nivel de significancia considerado fue p < 0.05 para todas las pruebas realizadas.
Todas las actividades se llevaron a cabo dentro de los estrictos límites de los marcos legales nacionales y de la UE, bajo la dirección de científicos/técnicos cualificados y de acuerdo con las directrices europeas sobre la protección de los animales utilizados con fines científicos (Directiva 2010/63/ UE del Parlamento Europeo y del Consejo de la Unión Europea), y bajo la estricta supervisión y control de la DGAV (Direção Geral de Alimentação e Veterinária), División de Bienestar Animal, autoridad competente responsable de la aplicación de la legislación sobre la protección de los animales utilizados con fines científicos.
Tabla 3. Genes seleccionados y primers específicos utilizados para evaluar el estado inmunitario de las PL de camarón blanco (P. vannamei) al final del período experimental.
Actina 4 de tipo citoplasmático
Proteína ribosomal L8
PvHm117 crustina P
Penaeidina-3a
Hemocianina
Lisozima tipo C
Lectina tipo C tipo 2
Tiorredoxina 1
Glutatión transferasa
Glutatión
Caspasa 3
Resultados
Rendimiento del crecimiento
No se observaron diferencias significativas en el rendimiento del crecimiento ni en la supervivencia entre los tratamientos dietéticos. El peso húmedo final promedió alrededor de 100 mg, los valores de RGR fueron del 15 % día−1, el FCR fue cercano a 1 y la supervivencia osciló entre el 86% y el 88% para todos los tratamientos (Tabla 4).
Biomarcadores relacionados con el estrés oxidativo y el estado inmunitario
En cuanto a los parámetros de estrés oxidativo medidos, los niveles de CAT fueron similares, sin detectarse diferencias significativas entre los tratamientos; los niveles de LPO fueron significativamente menores en los camarones PL alimentados con el tratamiento dietético BG que en los alimentados con la dieta PC, sin diferencias significativas entre los tratamientos restantes; los niveles de tGSH fueron significativamente mayores en los camarones PL alimentados con el tratamiento BG que en sus contrapartes alimentadas con la dieta NC, sin diferencias significativas entre los tratamientos restantes. En cuanto al estado inmunitario, no se observaron diferencias
significativas entre los tratamientos en cuanto a los parámetros medidos (Tabla 5).
Análisis de la expresión génica
La expresión relativa normalizada del ARNm del gen PvHm117 crustina P disminuyó significativamente en los camarones PL alimentados con la dieta NC en comparación con aquellos alimentados con los tratamientos dietéticos T + M y BG. De igual manera, el nivel de expresión del ARNm de penaeidina-3a disminuyó significativamente en los camarones PL alimentados con la dieta NC en comparación con sus contrapartes alimentadas con los tratamientos dietéticos PC y BG. Los transcritos de hemocianina aumentaron significativamente en los camarones PL alimentados con la dieta NC en comparación con los alimentados con el tratamiento dietético T + M. En cuanto a la expresión relativa normalizada del ARNm de los genes restantes, no se observaron diferencias significativas entre los tratamientos (Tabla 6).
Discusión
Este estudio tuvo como objetivo evaluar la inclusión de varios suplementos dietéticos en microdietas inertes para PL de camarón
F:CACGAGACCACCTACAACTCCATC
R: TCCTGCTTGCTGATCCACATCTG
F:AGCCAAGCAAGATGGGTCG
R: TGTAACGATAAGGGTCACGGAAG
F:GAAACCACCACCAACACCTACTCC
R: TCTGTGCGGCCTCTTTACGG
F:ATACCCAGGCCACCACCCTT
R: TGACAGCAACGCCCTAACC
F:GTCTTAGTGGTTCTTGGGCTTGTC
R: GGTCTCCGTCCTGAATGTCTCC
F:CGGGAAAGGCTATTCTGCCT
R: CCAGCACTCTGCCATGTACT
F:GCTTCTGTTGGTGCTGTTGGC
R: GTTCCCTTCCCGTATGTGGC
F:TTAACGAGGCTGGAAACA
R: AACGACATCGCTCATAGA
F: AAGATAACGCAGAGCAAGG
R: TCGTAGGTGACGGTAAAGA
F: AGGGACTTCCACCAGATG
R: CAACAACTCCCCTTCGGTA
F: ACATTTCTGGGCGGAACACC
R: GTGACACCCGTGCTTGTACA
blanco y estudiar los posibles efectos beneficiosos para su salud. Se seleccionaron vitaminas C y E, metionina, taurina y β-glucanos para este propósito, ya que su potencial para mejorar el estado de salud del camarón blanco en las etapas iniciales de desarrollo es prometedor, pero aún no se ha validado experimentalmente. Se utilizó una dieta control formulada para satisfacer los requerimientos nutricionales de las PL de camarón blanco como control positivo, y las dietas experimentales restantes se basaron en ella, difiriendo únicamente en la reducción o adición de los nutrientes mencionados previamente.
En general, los cambios en la formulación de las dietas no comprometieron su idoneidad, ya que se obtuvieron buenos rendimientos de crecimiento y resultados de supervivencia en todos los tratamientos experimentales, lo que también reveló que se mantuvieron buenas condiciones zootécnicas durante el ensayo. Los resultados de crecimiento fueron similares a los reportados por Wang et al. al utilizar niveles graduados de harina de Schizochytrium [31] y como reemplazo del aceite de pescado [32] en dietas prácticas para PL de camarón
ALIMENTACIÓN
blanco, pero los resultados de supervivencia fueron considerablemente inferiores en esos estudios (40.3–44.5% y 42.7–45.6%, respectivamente) que en el ensayo actual.
El aumento de los niveles de suplementación de vitaminas C y E en el tratamiento dietético PC no produjo cambios en el rendimiento del crecimiento ni en la supervivencia en comparación con la dieta NC, lo que sugiere que los niveles de estas vitaminas presentes en la dieta NC permitieron que las postlarvas de camarón mantuvieran un desarrollo adecuado. Al igual que otras vitaminas, las vitaminas C y E son nutrientes esenciales, ya que los animales no pueden sintetizar cantidades suficientes para satisfacer sus necesidades fisiológicas, y un aporte deficiente en la dieta suele provocar un crecimiento deficiente, lo que puede derivar en graves problemas de salud e incluso comprometer la supervivencia [6,33]. Además, no se verificaron cambios en los niveles de actividad de la mayoría de los parámetros inmunitarios y antioxidantes medidos en este estudio al aumentar los niveles de estas vitaminas en las dietas.
En consecuencia, otros estudios han reportado umbrales en los niveles de inclusión de estas vitaminas en dietas para camarón blanco juvenil en los cuales sus efectos beneficiosos no aumentaron después de un cierto porcentaje de incorporación [7,14]. La única disimilitud significativa detectada fue la menor expresión relativa de pen-3 en los camarones alimentados con la dieta NC en comparación con los alimentados con la dieta PC, lo que sugiere que niveles más bajos de estas vitaminas podrían afectar el estado inmunitario del camarón. Las penaidinas, un grupo clave de péptidos antimicrobianos en los camarones peneidos, poseen actividad antibacteriana y antifúngica, particularmente eficaz contra bacterias Gram+ y hongos filamentosos [34,35]. Estos hallazgos indican que la suplementación con niveles más altos de vitaminas C y E en microdietas para PL de camarón blanco no mejoró directamente el crecimiento ni la supervivencia en el presente estudio, pero podría haber mejorado su robustez. Aunque no se confirmó en este experimento, esto podría indicar que los camarones podrían tener una mayor capacidad de supervivencia a largo plazo,
Tabla 4. Peso inicial y final, tasa de crecimiento relativo (RGR), tasa de conversión alimenticia (FCR) y supervivencia de las PL de camarón blanco (P. vannamei) durante el período experimental.
Resultados expresados como media ± desviación estándar. Para el peso inicial, n = 60 unidades observacionales; para peso final, FCR, RGR y supervivencia, n = 3 unidades experimentales.
Tabla 5. Niveles de catalasa (CAT), peroxidación lipídica (LPO), glutatión total (tGSH), lisozima, profenoloxidasa y actividad bactericida en PL de camarón blanco (P. vannamei) alimentadas con dietas experimentales durante 18 días.
CAT (U mg−1 proteína)
LPO (nmol g peso−1)
tGSH (nmol mg proteína−1)
Lisozima (µg mg proteína−1)
Profenoloxidasa
(×10−3 U mL−1)
Actividad bactericida (%)
Resultados expresados como media ± desviación estándar (n = 3 unidades experimentales). También se representan los valores p para un ANOVA de una vía. Las diferentes letras en superíndice indican diferencias estadísticas (p < 0.05) entre tratamientos en una prueba de comparación múltiple post hoc de Tukey.
especialmente en una situación de cultivo potencialmente desafiante. Por lo tanto, las dietas inertes con niveles adecuados de vitaminas C y E pueden ser vitales durante etapas críticas de la producción, en particular en granjas donde se emplea un sistema de pre-criadero (etapa intermedia entre la etapa temprana de PL y la fase de engorde), donde las PL se mantienen a densidades de población extremadamente altas que pueden inducir estrés y vulnerabilidad a patógenos oportunistas [36].
La suplementación con metionina para equilibrar los perfiles nutricionales de alimentos acuícolas ricos en proteínas vegetales se ha convertido en una práctica común [16,37]. Tradicionalmente, el suministro de metionina se aseguraba con harina de pescado, pero se están realizando esfuerzos continuos para reducir la dependencia de la industria de este ingrediente y reemplazarlo con proteínas vegetales, donde la metionina y
la lisina generalmente presentan un perfil de aminoácidos bajo. Varios autores han demostrado que, cuando no se satisfacen los requerimientos dietéticos, generalmente, al usar dietas bajas en harina de pescado simultáneamente, el crecimiento y la supervivencia del camarón blanco pueden verse afectados [13,16,38–40]. Además, la metionina tiene un papel reconocido en el sistema inmunitario y recientemente se ha utilizado para mejorar la capacidad antioxidante, la respuesta inmunitaria innata y/o la resistencia a enfermedades de camarón blanco juvenil [16,18,41].
De igual forma, la metionina también es un precursor de la taurina. Se recomienda la suplementación de este nutriente en dietas para camarón blanco, ya que puede tener efectos beneficiosos en su crecimiento y respuesta inmunitaria [12,17]. Sin embargo, aún es necesario evaluar la suplementación de ambas moléculas en dietas para las etapas iniciales del desarrollo del camarón. En este
Tabla 6. Expresión relativa al mantenimiento (bactn y rpl-8) de genes inmunorrelacionados diana en PL de camarón blanco (P. vannamei) alimentada con dietas experimentales durante 18 días.
Expresión relativa
Gen Acrónimo
PvHm117 crustina P
Penaeidin-3a
Hemocianina
Lisozima tipo C
Lectina tipo C tipo 2
Tiorredoxina 1
Glutatión transferasa
Glutatión peroxidasa
Caspasa 3
Resultados expresados como media ± desviación estándar (n = 3 unidades experimentales). También se representan los valores p para un ANOVA de una vía. Las diferentes letras superíndices indican diferencias estadísticas (p < 0.05) entre tratamientos en una prueba de comparación múltiple de Tukey post hoc.
estudio, las PL alimentadas con la dieta T + M mostraron rendimientos de crecimiento, supervivencia, estado oxidativo y condición inmune similares a los alimentados con el tratamiento dietético PC, lo que sugiere que la formulación de los ingredientes de la dieta control fue capaz de cubrir los requerimientos de taurina y metionina de las PL, y no se obtuvieron beneficios adicionales a través de la suplementación de estos aminoácidos. Estos resultados probablemente se pueden explicar por el hecho de que la PC era una dieta de alta calidad con niveles considerables de proteína de origen marino. Sin embargo, las PL alimentadas con la dieta T + M mostraron expresiones relativas significativamente más altas de crus y transcripciones de hmc significativamente más bajas en comparación con las alimentadas con el tratamiento dietético NC.
Tanto la crustina P de PvHm117 como la hemocianina están asociadas con importantes péptidos antimicrobianos de amplio espectro involucrados en la primera línea de la defensa del camarón [42–44]. Estos resultados podrían considerarse contradictorios, ya que se esperaría que las variaciones en las expresiones de ambos genes siguieran la misma tendencia. Aun así, es importante tener en cuenta que la hemocianina es una proteína multifuncional
involucrada en varios procesos fisiológicos más allá de la inmunidad innata, como el transporte de oxígeno, el almacenamiento de proteínas, el ciclo de muda, la formación del exoesqueleto y la osmorregulación [45,46]. Al mismo tiempo, la taurina es uno de los principales osmolitos orgánicos en la osmorregulación de los decápodos [47,48], y se ha demostrado que el aumento de la inclusión de taurina en la dieta incrementa el contenido molecular en diferentes tejidos y hemolinfa del camarón blanco [17]. Por lo tanto, los niveles de hemocianina necesarios para mantener la osmolalidad fueron probablemente menores en los camarones alimentados con la dieta T + M, lo que podría haber causado una regulación negativa del gen hmc. Sin embargo, cabe destacar que el análisis solo se centró en la medición de las transcripciones del ARNm de hmc y no en los niveles de la molécula de taurina. Por lo tanto, para comprender mejor estas interacciones y aclarar si la suplementación de taurina y metionina en dietas para PL de camarón blanco es beneficiosa cuando se utilizan niveles más bajos de inclusión de vitaminas C y E, se deben realizar más estudios.
La inclusión de β-glucanos en las dietas no afectó significativamente el crecimiento y la supervivencia de las PL de camarón.
Aun así, las PL alimentadas con la dieta BG tendieron a crecer menos y alcanzaron pesos finales alrededor de un 15% menores que los alimentados con PC, aunque esto no fue respaldado por el análisis estadístico. No obstante, los niveles de peroxidación lipídica disminuyeron significativamente en las PL alimentadas con el tratamiento dietético BG en comparación con los alimentados con la dieta PC, lo que sugiere que los β-glucanos mejoraron los mecanismos antioxidantes de los animales. De hecho, se han reportado las propiedades inmunoestimulantes y antioxidantes de los β-glucanos como aditivos para alimentos acuícolas para varias especies, como lo revisaron recientemente Pogue et al. [49]. Estos pueden ser tremendamente valiosos en las etapas larvarias/PL, donde los camarones experimentan un desarrollo extremadamente rápido, ya que es probable que el crecimiento acelerado produzca un exceso de especies reactivas de oxígeno que pueden resultar en estrés oxidativo, dañando estructuras fisiológicas clave [50].
Aunque existen reportes de la mejora de la resistencia a enfermedades del camarón blanco a través de la suplementación dietética de β-glucanos [51–53], en este estudio no se observaron mejoras significativas en la condición inmune en las PL alimentadas con
ALIMENTACIÓN
la dieta BG, en comparación con aquellas alimentadas con la dieta PC. Bai et al. [54] sugirieron que la alimentación discontinua, cambiando entre una dieta basal y una con la inclusión de β-glucanos, es la estrategia más adecuada para mejorar la inmunidad del camarón blanco, ya que la alimentación continua por largos períodos de tiempo con la dieta suplementada puede causar fatiga inmune, mitigando los efectos beneficiosos proporcionados a corto plazo. Considerando esta hipótesis, el potencial de la dieta BG para mejorar la inmunidad de PL del camarón blanco podría haber sido más claro si se hubiera empleado una estrategia de alimentación diferente. Cuando se comparó con la dieta NC, las PL alimentadas con BG mostraron aumentos significativos en los niveles de tGSH así como en la expresión relativa de crus y pen-3. Esto sugiere que la suplementación con β-glucanos, junto con mayores niveles de vitamina C y E, puede potenciar la capacidad antioxidante y el estado inmunitario de PL del camarón blanco.
De hecho, Wu et al. [14] propusieron que existe una interacción entre los β-glucanos y la vitamina C capaz de aumentar la respuesta inmunitaria inespecífica del camarón blanco. Los resultados obtenidos en dicho estudio corroboran esta hipótesis, ya que la adición de β-glucanos a la dieta PC amplificó las diferencias en el estado inmunitario y la capacidad antioxidante del camarón en comparación con la dieta NC.
Conclusión
En resumen, los resultados obtenidos en este estudio sugieren que, si bien no se observaron mejoras en el crecimiento ni en la supervivencia al final del período experimental, todos los aditivos dietéticos probados tienen el potencial de añadir valor a las microdietas inertes para PL de camarón blanco. Los beneficios en la capacidad antioxidante y la robustez de las PL fueron más evidentes cuando los niveles de vitamina C y E fueron superiores a los utilizados en la dieta NC, similares a los utilizados en la dieta PC. Sin embargo, la dieta control puede
considerarse una opción premium, y es de esperar que los efectos positivos de estos suplementos se incrementen al incorporarse a alternativas más económicas. Entre los aditivos probados, la inclusión de β-glucanos en las dietas parece ser la más prometedora, ya que redujo la peroxidación lipídica en los camarones, incluso en comparación con una dieta control de alta calidad. En comparación con la NC, la interacción entre la suplementación de β-glucanos y mayores niveles de vitaminas C y E también parece beneficiosa para la capacidad antioxidante de las PL de camarón blanco•
La trazabilidad total ya es una realidad en el Ecuador
La industria camaronera ecuatoriana avanza con firmeza hacia una trazabilidad total en toda su cadena de valor. Actualmente, las etapas de cultivo y procesamiento cuentan con controles establecidos que garantizan altos estándares de calidad. Uno de los siguientes pasos estratégicos es fortalecer aún más el tramo de transporte entre las piscinas camaroneras y las plantas procesadoras. Este segmento es clave para asegurar que el producto mantenga su calidad desde el origen hasta su destino final. Existen grandes oportunidades para implementar soluciones tecnológicas que permitan monitorear en tiempo real variables críticas como la temperatura, tiempos de traslado y la integridad de los contenedores. Con un sistema robusto de trazabilidad en esta fase, los exportadores podrán contar con evidencia verificable para respaldar la cadena de custodia ante clientes, autoridades o aseguradoras, elevando así la confianza y reputación del camarón ecuatoriano en los mercados internacionales.
La hipótesis es que el uso de protocolos de comunicación inálambrica IoT combinados con una red LoRaWAN puede brindar una solución efectiva para lograr trazabilidad total durante el transporte acuícola. La meta de este estudio es demostrar la viabilidad de implementar sensores especializados que recojan datos clave —como ubicación, temperatura, apertura de tapa y nivel— y los transmitan en tiempo real a plataformas de monitoreo, asegurando que toda la ruta logística del camarón esté documentada, controlada y optimizada.
Metodología
Este estudio es de tipo aplicado y descriptivo, orientado a validar el funcionamiento de una red LoRaWAN para lograr trazabilidad completa en el transporte acuícola. Las pruebas iniciales se realizaron utilizando cinco Smartbins equipados con sensores de ubicación, temperatura y apertura, en recorridos dentro de la ciudad de Guayaquil. Estas pruebas permitieron verificar la operatividad del sistema, evaluar la estabilidad de la red y confirmar que los datos eran transmitidos y recibidos correctamente a lo largo del trayecto monitoreado.
Actualmente, la red LoRaWAN ya está
desplegada a lo largo de toda la ruta logística del camarón, cubriendo desde Huaquillas hasta Posorja, garantizando una trazabilidad completa durante el transporte del producto. Una característica destacada de esta red es su alta escalabilidad, ya que puede expandirse fácilmente hacia nuevas áreas según las necesidades de cobertura y monitoreo que vayan surgiendo en la industria. Esto permite anticipar futuros crecimientos hacia otras zonas o sectores donde se requiera control detallado y documentación en tiempo real de los procesos logísticos.
Los materiales utilizados incluyeron sensores IoT validados previamente, gateways LoRaWAN integrados a la red y una plataforma de monitoreo en línea que muestra los datos captados en campo. La medición de resultados basada en la recolección automática de datos generados por los bins durante las pruebas asegura que la trazabilidad se mantiene de forma continua, confiable y verificable a lo largo de toda la cadena de transporte.
¿Por qué LoRaWAN para la ruta del camarón?
LoRaWAN es un protocolo de comunicación de red de área amplia y baja potencia (LPWAN) que funciona sobre la tecnología Low Range (LoRa), que permite la transmisión de señales de radio a larga distancia con bajo consumo de energía. Es un estándar internacional para la conectividad IoT, especialmente para dispositivos de bajo consumo que necesitan enviar pequeñas cantidades de datos a largas distancias.
La elección de esta tecnología de comunicación es uno de los pilares fundamentales para garantizar una trazabilidad completa en la cadena logística camaronera. No todas las soluciones disponibles en el mercado son adecuadas para cubrir las extensas rutas que atraviesan zonas rurales, áreas costeras y corredores logísticos de difícil acceso. Por eso, antes de definir la arquitectura final del sistema, se realizó un análisis comparativo entre diferentes tecnologías IoT disponibles: 5G, LTE Cat 1, LTE-M, NB-IoT, y LoRaWAN. Este análisis consideró aspectos como el costo por módulo, el consumo energético, la capacidad de cobertura en áreas remotas y la escalabilidad de la solución. Como
resultado, LoRaWAN fue seleccionada por su equilibrio entre bajo costo, bajo consumo de batería y su capacidad de operar en zonas sin infraestructura celular, garantizando la continuidad del monitoreo en tiempo real.
Como se observa en la Figura 1, LoRaWAN ocupa una posición privilegiada frente a otras alternativas tecnológicas, ofreciendo costos por módulo mucho más bajos (alrededor de €2) y un consumo energético significativamente reducido, lo que permite a los dispositivos IoT operar durante años con una sola batería. Aunque otras tecnologías como tecnología sw comunicación inalámbrica de comunicación LTE-M o NB-IoT ofrecen mayores velocidades de transmisión, estas características no son prioritarias en aplicaciones como la trazabilidad acuícola, donde los datos transmitidos son pequeños pero críticos (posiciones GPS, valores de temperatura, alertas de apertura de tapa).
La cobertura extendida de LoRaWAN permite garantizar conectividad incluso en zonas donde la red celular no está disponible, asegurando que la trazabilidad no se pierda en ningún punto de la ruta. Además, la naturaleza escalable de esta red significa que puede crecer fácilmente hacia nuevas zonas de operación conforme surjan nuevas necesidades, convirtiéndola en la mejor opción para cubrir la ruta del camarón ecuatoriano desde Huaquillas hasta Posorja y más allá. Esta elección tecnológica no solo responde a las demandas actuales, sino que prepara a la industria para afrontar los
desafíos del futuro en términos de monitoreo, control y optimización logística.
Topología utilizada en el sistema de monitoreo
La arquitectura de red implementada para garantizar la trazabilidad completa en la ruta del camarón sigue una topología simple pero robusta, optimizada para aplicaciones IoT de bajo consumo y largo alcance. El diseño contempla tres componentes principales: los dispositivos SmartBins, la red LoRaWAN y la plataforma de gestión accesible por los usuarios finales.
Los SmartBins, equipados con sensores de posición, temperatura y apertura, transmiten datos periódicos a través de la red LoRaWAN, que cuenta con gateways distribuidos estratégicamente. Estos recolectan la información de los contenedores y la envían hacia un servidor central ubicado en la nube, donde los datos son procesados, almacenados y visualizados en tiempo real. Los usuarios, ya sean transportistas, procesadores o exportadores, pueden acceder a esta información a través de plataformas web o aplicaciones móviles, permitiendo un monitoreo continuo y una gestión eficiente de la logística acuícola.
Este esquema permite una comunicación eficiente y escalable, evitando la dependencia de redes celulares tradicionales y asegurando que los datos captados en campo lleguen de manera confiable al sistema central. Además, la flexibilidad de
Figura 1. Comparación de tecnologías IoT según costo, velocidad de transmisión y consumo energético.
TECNOLOGÍA
la arquitectura permite incorporar nuevos dispositivos o sensores según evolucionen las necesidades del sector, manteniendo la infraestructura preparada para futuros desafíos tecnológicos.
Parámetros de monitoreo y recolección de datos
El sistema de monitoreo implementado en la ruta del camarón está diseñado para capturar parámetros críticos en tiempo real, garantizando una trazabilidad completa y, sobre todo, aumentando la seguridad y el control sobre la carga durante todo el trayecto logístico.
El cuerpo del bin permite registrar constantemente la ubicación GPS, permitiendo visualizar en todo momento la posición exacta del contenedor y la temperatura interna, asegurando que se mantiene la cadena de frío necesaria para preservar la calidad del producto.
En la tapa del bin, los sensores se enfocan en aspectos clave para la seguridad:
• Apertura de tapa, detectada a partir de la inclinación, lo que permite saber si se ha manipulado el contenedor durante el trayecto.
• Temperatura en la tapa, para monitorear cambios externos que podrían impactar las condiciones internas.
• Distancia de la tapa al líquido, un parámetro que permite estimar el nivel de agua dentro del bin, útil para validar que se mantiene la cantidad adecuada de líquido durante el transporte. Estos parámetros no solo permiten un monitoreo técnico preciso, sino que también ofrecen un respaldo verificable ante eventos no autorizados o inesperados. La combinación de estos datos refuerza la seguridad del transporte acuícola, brindando a los productores y exportadores la tranquilidad de que su carga está protegida y que cualquier irregularidad será detectada y registrada automáticamente.
Resultados
Durante las pruebas de campo realizadas con los cinco SmartBins se recolectaron miles de mediciones de temperatura, apertura de tapa y nivel de líquido, transmitidas a través de la red LoRaWAN. Uno de los indicadores clave analizados fue la confiabilidad de la
red, definida como el porcentaje de datos válidos recibidos sobre el total esperado, tras eliminar registros vacíos y outliers extremos.
Estos resultados confirman que la red de medición operó con una confiabilidad muy alta, alcanzando valores superiores al 98% en los bins principales (001, 002, 003) y manteniéndose por encima del 92% incluso en los contenedores que presentaron mayores desafíos operativos (004 y 005).
Las diferencias observadas se atribuyen principalmente a la menor cantidad de registros originales en estos últimos bins, lo que amplifica proporcionalmente el impacto de cualquier pérdida puntual de datos, así como a variaciones locales en la calidad de la señal o las condiciones de operación.
En resumen, los resultados respaldan la hipótesis central del estudio: la implementación de una red LoRaWAN con dispositivos IoT permite lograr una trazabilidad completa y confiable en la ruta del camarón, ofreciendo a la industria una herramienta robusta para garantizar la calidad, seguridad y control logístico del
producto.
Discusión
Los resultados obtenidos en este estudio respaldan los objetivos planteados originalmente, evaluar la viabilidad técnica de implementar una red LoRaWAN con dispositivos IoT para lograr trazabilidad total en la ruta logística del camarón en Ecuador. Las pruebas de campo mostraron una confiabilidad de red superior al 92% en todos los contenedores monitoreados, confirmando que la solución propuesta permite recolectar, transmitir y gestionar datos críticos como ubicación, temperatura, apertura de tapa y nivel de líquido de forma continua y robusta.
En comparación con desarrollos previos de trazabilidad en cadenas logísticas acuícolas y agrícolas, los resultados aquí presentados destacan por su enfoque en el transporte, etapa difícil de controlar. Mientras que otros esfuerzos se han centrado principalmente en trazabilidad en plantas de procesamiento o en fases de cultivo, este trabajo demuestra que es técnicamente viable extender el
Los resultados tienen una base común de 8.800 registros por bin y se resumen en la siguiente tabla:
Figura 2. Topología general del sistema de monitoreo LoRaWAN aplicado a los SmartBins en la ruta del camarón.
monitoreo al trayecto mismo, utilizando una arquitectura de red ligera y eficiente como LoRaWAN.
Entre las principales fortalezas del estudio se encuentran la implementación de pruebas en condiciones de operación, el volumen amplio de datos recolectados (más de 30.000 registros totales) y la validación cruzada de los sensores, que permitió asegurar la calidad y precisión de las mediciones. Además, la escalabilidad de la red implementada abre la posibilidad de ampliar su cobertura a otras rutas y sectores, posicionando esta solución como una herramienta flexible para la industria acuícola.
No obstante, el estudio presenta limitaciones. Las pruebas se realizaron con una muestra piloto de cinco SmartBins, lo cual, si bien representativo, no cubre la totalidad de los escenarios logísticos posibles (por ejemplo, rutas más largas, diferentes tipos de transporte o variaciones climáticas extremas). Además, aunque se alcanzaron porcentajes de confiabilidad altos, algunos contenedores mostraron niveles levemente inferiores, lo que sugiere que aún es necesario optimizar factores como la densidad de gateways y la gestión de interferencias locales.
A partir de estos resultados, exploraremos futuras investigaciones que aborden la integración de nuevos sensores (como calidad del agua, vibración o impactos), o también explorar la operabilidad con otros sistemas de monitoreo (como blockchain para certificación) y evaluar modelos predictivos basados en inteligencia artificial para anticipar fallas o eventos críticos en la cadena logística.
En última instancia, este estudio demuestra que es posible cerrar la brecha en la trazabilidad del transporte camaronero, ofreciendo una solución tecnológica concreta que puede transformar las prácticas operativas del sector. Los hallazgos aquí reportados no solo aportan evidencia empírica sobre la viabilidad técnica de la red LoRaWAN para aplicaciones logísticas, sino que también sientan las bases para futuras mejoras que fortalezcan la sostenibilidad, transparencia y competitividad de la industria
acuícola ecuatoriana en los mercados globales.
Conclusión
La implementación de redes LoRaWAN para trazabilidad total en el transporte acuícola representa un avance significativo no solo para la industria, sino también para el campo de las aplicaciones científicas del IoT en logística. Este estudio demuestra que es posible cerrar una brecha histórica en la cadena de valor del camarón, integrando tecnologías de bajo consumo y alto alcance para generar datos confiables y continuos, incluso en condiciones operativas complejas.
Desde una perspectiva más amplia, estos resultados subrayan la importancia de considerar soluciones tecnológicas adaptadas al contexto local, priorizando la eficiencia y la escalabilidad por encima de la simple adopción de tecnologías de última generación. Además, refuerzan la noción de que la digitalización de procesos logísticos no debe centrarse únicamente en fases visibles como producción o exportación, sino también en los tramos intermedios que tradicionalmente han sido menos desarrollados.
Como consideración final, este trabajo
invitamos a la comunidad científica a reflexionar sobre el potencial de las redes LoRaWAN que funcionan en conjunto con IoTs no solo como herramientas técnicas, sino como elementos transformadores de modelos productivos, abriendo nuevas líneas de investigación en interoperabilidad, resiliencia de redes y análisis predictivo aplicado a cadenas de suministro sensibles. La experiencia generada aquí sienta una base sólida para futuras investigaciones orientadas a ampliar el alcance de la trazabilidad digital y a fortalecer la Para más información sobre este artículo, contactar a: alopez@plasticosrival.com
El microbioma del camarón blanco del Pacífico revela una composición diferencial de la comunidad bacteriana entre condiciones naturales,
Autores:
Fernanda Cornejo-Granados¹
Alonso A. López-Zavala²
Luigui Gallardo-Becerra¹
Alfredo Mendoza-Vargas³
Filiberto Sánchez¹
Rodrigo Vichido4
Luis G. Brieba5, Maria Teresa Viana6
Rogerio R. Sotelo-Mundo7
Adrián Ochoa-Leyva¹
1Departamento de Microbiología Molecular, Instituto de Biotecnología (IBT), Universidad Nacional Autónoma de México (UNAM), Av. Universidad #2001, Col. Chamilpa, Cuernavaca, Morelos, 62210, México.
2Departamento de Ciencias Químico-Biológicas, Universidad de Sonora (UNISON). Blvd., Rosales y Luis Encinas, Hermosillo, Sonora, 83000, México.
3Instituto Nacional de Medicina Genómica, Secretaría de Salud (INMEGEN), Periférico Sur No. 4809, México, D.F., 14610, México.
4Centro Nacional de Servicios de Constatación en Salud Animal (CENAPA), Carr. Fed. Cuernavaca-Cuautla No. 8534 Jiutepec, Morelos, 8534, México.
5Laboratorio Nacional de Genómica para la Biodiversidad (LANGEBIO), Centro de Investigación y Estudios Avanzados (CINVESTAV Unidad Irapuato) Km 9.
6Libramiento Norte Carretera Irapuato-León, Apartado Postal 629, Irapuato, Guanajuato, 36500, México. 6Instituto de Investigaciones Oceanológicas, Universidad Autónoma de Baja California (UABC), Ensenada, B.C., México.
7Laboratorio de Estructura Biomolecular, Centro de Investigación en Alimentación y Desarrollo, A.C. (CIAD), Carretera a Ejido La Victoria Km 0.6, Apartado Postal 1735, Hermosillo, Sonora, 83304, México. Fernanda Cornejo- Granados and Alonso A. Lopez-Zavala contributed equally to this work. Correspondence and requests for materials should be addressed to R.R.S.-M.
Los crustáceos conforman el segundo subphylum más grande de la Tierra, que incluye a Penaeus vannamei (camarón blanco del Pacífico), uno de los camarones más cultivados a nivel mundial. A pesar de los esfuerzos por estudiar la microbiota de la especie, se conoce poco sobre ella en el caso del camarón de mar abierto y sobre el papel que desempeña la acuicultura en la remodelación de la microbiota. En este estudio, se caracterizó la microbiota del hepatopáncreas y el intestino de camarones silvestres (wt) y de cultivo, así como de sedimentos de estanques de laboratorios, mediante la secuenciación de siete regiones hipervariables del gen ARNr 16S. También se incluyeron camarones de cultivo con síntomas de la enfermedad AHPND/EMS. Se observó que (i) la microbiota y sus funciones metagenómicas predichas difirieron entre camarones silvestres y de cultivo; (ii) independientemente del origen del camarón, la microbiota del hepatopáncreas y el intestino fue diferente; (iii) la diversidad microbiana entre el sedimento y los intestinos de los camarones de cultivo fue similar; y (iv) asociado con un desarrollo temprano de la enfermedad AHPND/EMS, se observaron cambios en el microbioma y la aparición de bacterias específicas de la enfermedad. En particular, en condiciones de cultivo, identificamos taxones bacterianos enriquecidos en camarones sanos, como Faecalibacterium prausnitzii y Pantoea agglomerans, y comunidades enriquecidas en camarones enfermos, como Aeromonas taiwanensis, Simiduia agarivorans y Photobacterium angustum.
En los organismos terrestres, la colonización bacteriana inicial se produce a partir de la microbiota materna, mientras que en los sistemas acuáticos proviene del agua y los sedimentos circundantes1. La microbiota desempeña un papel fundamental en el desarrollo y la fisiología de su huésped, previniendo el crecimiento de bacterias patógenas, modulando la respuesta inmunitaria, afectando la absorción de nutrientes, regulando los procesos metabólicos y sintetizando vitaminas2. La estructura (abundancia y composición) y la función de la microbiota se ven influenciadas por la ingesta de alimento, los probióticos, los prebióticos, la secreción hormonal, el estrés, los antibióticos, la etapa de desarrollo, las
condiciones ambientales y fisiológicas2, el metabolismo del huésped y la respuesta inmunitaria3. Los crustáceos pertenecen al grupo de los artrópodos, que representa el segundo subphylum más grande de la Tierra e incluye cangrejos, langostas, cangrejos de río, krill y camarones. La producción artificial de crustáceos ha impulsado una creciente industria acuícola que ha evitado la captura de este recurso, aunque su cultivo podría tener un impacto negativo en las marismas y las zonas costeras de manglar4. Al igual que en muchas áreas de la nutrición animal, ahora se pone un gran énfasis en comprender los roles de la microbiota en la salud, el crecimiento y la supervivencia de los organismos cultivados5 En estudios recientes, se utilizaron técnicas convencionales dependientes del cultivo para caracterizar las comunidades bacterianas en crustáceos, seguido de enfoques moleculares tradicionales, como la electroforesis en gel de gradiente desnaturalizante (DGGE) y las bibliotecas de clones6–10. Más recientemente, la disponibilidad y el bajo costo de la secuenciación de alto rendimiento del gen de la subunidad ribosomal pequeña 16S (ARNr 16S) han facilitado un análisis más profundo de la microbiota del camarón en condiciones de cultivo. Se descubrió una gran diversidad de microbiota que no se podía identificar con los métodos tradicionales dependientes del cultivo. Se han descrito cambios en la microbiota de especies de camarón, como Penaeus monodon (camarón tigre negro)11, Fenneropenaeus chinensis (camarón chino)10, Penaeus penicillatus12 , Penaeus merguiensis (camarón banana)13 y Penaeus vannamei14 (camarón blanco del Pacífico), en diversas condiciones de crecimiento y calidad del agua. Sin embargo, la microbiota de ambientes naturales solo se ha estudiado en el intestino de P. monodon11. Hasta donde sabemos, se desconoce la microbiota intestinal y del hepatopáncreas, así como sus posibles funciones, en P. vannamei silvestre (wt) capturado en el Océano Pacífico. Por lo tanto, es necesaria la caracterización de la microbiota natural de P. vannamei, para utilizarla como referencia en la comparación con camarones de cultivo sanos y enfermos en laboratorio. De esta forma, la modificación de la microbiota podría utilizarse como un nuevo enfoque contra las enfermedades del camarón15.
Existe poca información sobre la microbiota en órganos del camarón distintos del intestino, dejando de lado la importante conexión que existe entre la función de este órgano y el hepatopáncreas debido a que el tracto digestivo de los crustáceos es continuo16. Este último es un órgano esencial para la digestión y absorción de nutrientes, y desempeña un papel en la inmunidad innata en invertebrados17. Hasta la fecha, solo ha habido un estudio que examina la microbiota del hepatopáncreas de N. denticulata16 . Además, el hepatopáncreas puede verse drásticamente afectado por patógenos como V. parahemolyticus, que causa el devastador Síndrome de Mortalidad Temprana (EMS)18,19, también llamado enfermedad de Necrosis Hepatopancreática Aguda (AHPND), en camarones20. La AHPND/EMS está obstaculizando la producción mundial del camarón, y se ha realizado un gran esfuerzo para comprender la base molecular de la enfermedad, incluyendo estudios genómicos de V. parahaemolyticus21. Un estudio seminal demostró que las toxinas Pir A/B están asociadas con la presencia de AHPND/EMS, atacando el hepatopáncreas, y su presencia explica muchos de los aspectos fenotípicos de la enfermedad18. Hasta donde sabemos, no se han realizado estudios de los efectos de las toxinas bacterianas en la microbiota, aunque algunos autores han demostrado que las toxinas químicas de hecho tienen efectos en las comunidades bacterianas de animales22. Pocos estudios han cuantificado cómo la microbiota se ve afectada por la aparición de enfermedades en condiciones de cultivo23,24. Los patógenos pueden interactuar con la microbiota del huésped por competencia por recursos, liberación de compuestos antimicrobianos o antagonismo. Por lo tanto, un patógeno puede causar disbiosis de la microbiota. Debido a la interacción constante entre el sistema digestivo del camarón y su entorno circundante (agua y sedimento), la microbiota del intestino y el hepatopáncreas debería participar en los mecanismos de resistencia a las enfermedades. Sin embargo, no hay investigaciones publicadas que describan los cambios en la microbiota del camarón durante un brote de AHPND/EMS.
P. vannamei, comúnmente conocido como camarón blanco del Pacífico, es una de las especies de camarón más cultivadas (>70%) a
nivel mundial y contribuye significativamente al desarrollo económico de las zonas costeras y humedales25. En este estudio, presentamos la estructura y función de la microbiota intestinal y del hepatopáncreas de muestras de camarón blanco del Pacífico provenientes de su ambiente natural y de laboratorios de producción, incluyendo sedimentos de estanques (Fig. 1). Nuestro estudio incorpora la secuenciación de siete regiones hipervariables del gen ribosomal 16S. Además, incluimos el análisis de la microbiota de camarones con síntomas de AHPND/EMS. Nuestros principales objetivos fueron determinar: (i) la estructura y función de la microbiota entre camarones de ambientes naturales y de cultivo; (ii) el impacto de la enfermedad de AHPND/EMS en la estructura y función de la microbiota del hepatopáncreas y el intestino de camarones de cultivo; y (iii) el análisis de la composición de la microbiota entre sedimentos de estanques de laboratorio que contiene camarones sanos y enfermos. Hasta donde sabemos, este estudio fue el primer estudio de secuenciación de alto rendimiento de la microbiota de P. vannamei utilizando intestinos y hepatopáncreas individuales de entornos naturales y de producción, incluyendo la microbiota asociada con el desarrollo temprano de la enfermedad APHND/EMS.
Resultados
Clasificación de lecturas por región hipervariable del ARNr 16S. El kit ION 16STM Metagenomics incluye seis primers patentados y diseñados para dirigirse a las regiones V2, V4 y V8 en una reacción de PCR simultánea y a las regiones V3, V6-7 y V9 en una segunda reacción. El porcentaje de lecturas fue uniforme para todas las regiones entre tres réplicas biológicas de cada grupo (Fig. Suplementaria S1). Se observó que las regiones V2, V4, V8 y V9 no mostraron sesgo en la abundancia de lecturas hacia ningún grupo de muestra en particular. Sin embargo, las lecturas en la región V3 se enriquecieron significativamente para el hepatopáncreas de wt (p < 0.05). En contraste, las lecturas en V6-V7 se enriquecieron significativamente en los intestinos y hepatopáncreas cultivados (Fig. Suplementaria S2). Curiosamente, este sesgo dependió del origen del camarón, lo que sugiere que existe una mayor diferencia en la abundancia de taxones microbianos
específicos (con una amplificación sesgada en las regiones V3, V6-7) entre las muestras wt y las cultivadas. Cabe destacar que las unidades taxonómicas operativas (OTUs) asignadas fueron diferentes entre las siete regiones secuenciadas (Fig. Suplementaria S3).
Características generales del microbioma mediante siete regiones hipervariables. Tras la eliminación de singletons, se obtuvieron 1,006 OTUs, que se asignaron a 36 phyla, 81 clases, 152 órdenes, 234 familias, 489 géneros y 298 especies con nombres taxonómicos. Aproximadamente el 47% de las lecturas no coincidieron con ninguna OTU conocida reportada en la base de datos GreenGenes (Fig. Suplementaria S4a). Curiosamente, las muestras de sedimento de estanque presentaron el mayor número de lecturas desconocidas (~60%), seguidas de las muestras de camarón wt (Fig. Suplementaria S4b). La mayor parte de la abundancia de OTU fue variable: el phylum Proteobacteria varió del 28.2 al 98.6%, Cyanobacteria del 0.0 al 62.5%, Actinobacteria del 0.1 al 50.5%, Gemmatimonadetes del 0.0 al 44.7%, Bacteroidetes del 0.0 al 16.7% y Firmicutes del 0.0 al 10.9% (Fig. 1c y Tabla Suplementaria S1). Vibrio, Pseudomonas, Photobacterium y Acinetobacter fueron los géneros más abundantes en el hepatopáncreas y el intestino, mientras que Acinetobacter, Vibrio, KSA1 y Pseudomonas fueron los más abundantes en el sedimento (Tabla Suplementaria S2). Enterobacteriaceae, Vibrionaceae, Pseudoalteromonadaceae, Moraxellaceae, Pseudomonadaceae y Rhodobacteraceae fueron las familias más abundantes en todas las muestras (Fig. 1d). Las comunidades bacterianas entre los triplicados biológicos fueron similares en las OTUs, con abundancias superiores al 0.1 % (correlaciones de Spearman en la Tabla Suplementaria S3).
Las métricas de diversidad alfa se calcularon a partir de las curvas de rarefacción a nivel de OTUs para cada grupo (Fig. 2). Para aumentar la profundidad de secuenciación, descartamos de este análisis dos muestras con baja profundidad de secuenciación (68B-S y 139-I). Por lo tanto, las métricas de diversidad alfa se calcularon a una profundidad de secuencia de 2,078 lecturas.
Figura 1. Recolección de muestras de camarón y características generales de la microbiota. (a) Puntos de recolección de especímenes de Penaeus vannamei. Los camarones sanos se recolectaron en la zona del Pacífico mexicano, a 5 km de la costa estatal (estrella verde), mientras que los camarones de cultivo se recolectaron en estanques de laboratorios (estrella naranja) ubicados en el estado de Sonora, México. Los camarones sanos y enfermos con AHPND/EMS se recolectaron en estanques adyacentes. El mapa se creó con la versión en línea de SmartDraw https://www.smartdraw.com/ . (b) Ubicación anatómica del hepatopáncreas y el intestino disecados. Diversidad taxonómica y abundancia de las 24 muestras secuenciadas a nivel de: (c) phylum; y (d) familia.
Las curvas de rarefacción mostraron que los índices de Shannon fueron relativamente estables a la profundidad de muestreo de >2,000 lecturas (Fig. Suplementaria S5a). Todos los valores de Chao1 fueron mayores que las OTUs observadas, lo que indica que aún podría esperarse encontrar más OTUs en todas las muestras (Tabla Suplementaria S4). En consecuencia, la cobertura de Good reveló que obtuvimos, en promedio, ~80% de las OTUs totales (Tabla Suplementaria S5), lo que indica que es necesaria una profundidad de secuenciación significativa para descubrir las OTUs totales en los microbiomas analizados. La cobertura de Good fue uniforme entre los grupos: 83% en promedio para intestinos, 90% para hepatopáncreas y 81% para sedimentos (Tabla Suplementaria S5). La diversidad filogenética (PD), el índice de Shannon y las OTUs observadas mostraron que los sedimentos y los intestinos tenían más diversidad bacteriana que los hepatopáncreas (Fig. 2). Las curvas de rarefacción se obtuvieron utilizando solo aquellas secuencias asignadas a un género como una estimación de la diversidad en ese nivel taxonómico. Las curvas de rarefacción no alcanzaron la saturación
(Fig. Suplementaria S5b), incluso cuando las secuencias de las tres muestras de cada ambiente se agruparon para obtener una mayor profundidad de muestreo de secuencia. Este hallazgo indicó que los géneros observados para cada ambiente parecen no saturados incluso después de 16,000 lecturas (Fig. Suplementaria S5c), sin embargo, se necesitará una profundidad de secuenciación significativamente mayor para descubrir géneros adicionales. La mayor parte de los índices de diversidad para todas las muestras secuenciadas se describe en la Tabla Suplementaria S4.
El análisis de componentes principales (PCoA) utilizando distancias UniFrac ponderadas y no ponderadas, mostró que las muestras wt se agruparon por separado de las muestras cultivadas (Fig. 3a y Fig. Suplementaria S6a). Además, el PCoA con muestras marcadas por órganos y sedimentos de estanque formó diferentes grupos (Fig. 3b). Las comunidades bacterianas en el hepatopáncreas fueron distintas a las de los intestinos, ya sea de muestras wt o cultivadas, y cada órgano se agrupó por separado según su origen (Fig. 3b). Cabe destacar que las muestras de
intestino de camarón de cultivo se agruparon junto con el sedimento de estanque en el PCoA ponderado (Fig. Suplementaria S6b). También se observaron grupos similares en el árbol UPGMA de distancias UniFrac no ponderadas (Fig. 3c), en el que todas las muestras wt formaron un grupo, y las muestras cultivadas formaron otro, ambos con un 100% de Soporte Jackknife (JS) (Fig. 3c). Además, el hepatopáncreas se agrupó por separado del intestino en las muestras wt y cultivadas, mientras que el intestino y el sedimento se agruparon con un 100% de JS en las muestras cultivadas (Fig. 3c). Este hallazgo concordó con la proximidad entre los grupos de intestino y sedimento observados en el PCoA no ponderado (Fig. 3b) y con un solo grupo observado que contenía todas las muestras de intestino y sedimento en el PCoA ponderado (Fig. Suplementaria S6b). Por lo tanto, la diversidad microbiana de los sedimentos del estanque y los intestinos del camarón fue similar en condiciones de cultivo de camarón. El árbol ponderado UPGMA (Fig. Suplementaria S7) mostró resultados similares a los observados en el árbol UPGMA no ponderado (Fig. 3c).
Microbioma intestinal. Todos los índices de diversidad (OTUs observados, PD y Shannon) mostraron una biodiversidad significativamente mayor (p < 0.05) para los intestinos wt en comparación con los intestinos cultivados sanos (Fig. 2 y Tabla Suplementaria S4). También encontramos que tanto los intestinos wt como los cultivados tenían significativamente (p < 0.05) más diversidad bacteriana que los hepatopáncreas wt y cultivados. Proteobacteria y Cyanobacteria fueron los filos más abundantes en la mayoría de las muestras (Fig. 1c). Vibrionaceae y Enterobacteriaceae fueron las familias más abundantes, constituyendo ~50% de las lecturas totales (Fig. 1d). A nivel de género, las comunidades fueron ricas en Vibrio, Photobacterium y Paracoccus (Tabla Suplementaria S2). El análisis discriminante lineal (LDA) utilizando LEfSe26, que utiliza las pruebas no paramétricas de KruskalWallis y de Wilcoxon-Mann-Whitney, se aplicó para identificar biomarcadores (taxones y funciones metagenómicas) que explican las mayores diferencias entre nuestros diferentes grupos (n = 3). Debido a que LEfSe requiere que todas las comparaciones por pares
rechacen la hipótesis nula para detectar un biomarcador, no se necesitan correcciones de pruebas múltiples26. Después del análisis LEfSe, los phyla Actinobacteria y Nitrospirae fueron significativamente más abundantes en las muestras wt (n = 3), mientras que Bacteroidetes, Gemmatimonadetes, Fusobacteria y Spirochaetes fueron más abundantes en las muestras cultivadas (n = 3) (Tabla Suplementaria S6). Las clases, órdenes, familias, géneros y especies que fueron diferencialmente abundantes entre las muestras wt y cultivadas se muestran en la Fig. 4a y la Tabla Suplementaria S6. Las comunidades bacterianas con una abundancia superior al 0.1 % (OTUs > 0.1 %) presentaron una marcada diferencia entre los camarones de cultivo wt y los sanos (prueba de Spearman, r = 0.026, p = 0.7144), en concordancia con los hallazgos observados en los árboles PCoA y UPGMA.
Utilizamos PICRUSt27 para predecir la funcionalidad del metagenoma del camarón a partir de la data del gen ARNr 16S. Se predijeron un total de 377 vías KEGG utilizando todas las muestras secuenciadas (Tabla Suplementaria S7). En el nivel dos de la subcategoría funcional, los genes implicados en cuatro vías (metabolismo de lípidos y aminoácidos, biodegradación de xenobióticos y metabolismo de otros aminoácidos) se enriquecieron significativamente en los camarones wt, mientras que el procesamiento de la información genética, los sistemas sensoriales, la biosíntesis de glicanos y las vías metabólicas predichas se enriquecieron en las muestras de cultivo sanas (Fig. 4b). Curiosamente, las comunidades bacterianas de muestras de intestino cultivado tuvieron una abundancia significativamente mayor de proteínas de replicación, recombinación y reparación, y vías de transducción de señales (Fig. Suplementaria S8a). En el nivel tres de la subcategoría funcional, la degradación de valina, leucina e isoleucina, el metabolismo del propanoato, el metabolismo de los ácidos grasos, la degradación del benzoato y el metabolismo del triptófano se enriquecieron en la microbiota derivada del camarón wt (Fig. Suplementaria S8a). Curiosamente, encontramos genes predichos involucrados en la biodegradación de xenobióticos, incluidos estireno, caprolactama, aminobenzoato, naftaleno y bisfenol (Fig.
Figura 2. Índices de diversidad alfa. (a) Diversidad filogenética (PD). (b) Número de OTUs observadas. (c) Índice de Shannon. Todos los grupos analizados mostraron diferencias significativas tras una prueba de Mann-Whitney con un nivel de confianza del 99% (p ≤ 0.01) en cada índice. El valor promedio de cada grupo de muestras (n = 3) se muestra junto a cada diagrama de caja, y la media se indica en rojo.
Suplementaria S8a). Además, para medir la confiabilidad de las predicciones funcionales para cada muestra, calculamos el índice de taxón secuenciado más cercano (NSTI) para las predicciones de PICRUSt. Esta puntuación estima qué tan estrechamente relacionados están los microbios en una muestra dada con los microbios representativos con genomas secuenciados. Los NSTI para intestinos wt y sanos fueron 0.081 ± 0.035 y 0.112 ± 0.072, respectivamente (Tabla Suplementaria S8).
Microbioma del hepatopáncreas. Los índices de diversidad mostraron que el hepatopáncreas wt y sano tuvo significativamente (p < 0.05) menos diversidad bacteriana que los intestinos wt
Figura 3. Análisis de diversidad beta. Análisis de coordenadas principales (PCoA) no ponderado de las distancias UniFrac: (a) muestras marcadas como wt y cultivadas; y (b) muestras marcadas por órgano o sedimento de camarón. Solo se muestran los dos primeros componentes. (c) Árbol UPGMA de distancias UniFrac no ponderadas. El valor del nodo representa el soporte jackknife, utilizando 1,000 réplicas.
Figura 4. Abundancia relativa y resultados de LEfSe de la microbiota enriquecida y funciones microbianas predichas entre muestras wt (n = 3) e intestino cultivado (n = 3). (a) Abundancia relativa de especies significativamente diferentes. La puntuación del análisis discriminante lineal (LDA) se muestra entre paréntesis. (b) LDA de las vías KEEG con abundancia diferencial. Las diferencias fueron significativas (p < 0.05) entre clases (prueba de Kruskal-Wallis) y subclases (prueba de Wilcoxon). El umbral de la puntuación logarítmica del LDA fue de 2.0.
Microbiota compartida entre camarones
y sanos (Fig. 2 y Tabla Suplementaria S4). El hepatopáncreas del camarón wt tuvo significativamente (p < 0.05) más diversidad bacteriana que el del camarón cultivado sano (Fig. 2). Proteobacteria fue el phylum más abundante (Fig. 1c y Tabla Suplementaria S1), y la familia Enterobacteriaceae representó el 44 y el 70% del total de secuencias derivadas wt y cultivadas, respectivamente. Las familias Vibrionaceae, Moraxellaceae y Pseudomonadaceae representaron el 27, el 12 y el 1%, respectivamente, del total de lecturas en muestras wt, mientras que estas familias representaron el 7, el 0.5 y el 19% del total de lecturas en muestras cultivadas. A nivel de género, Photobacterium (16%), Acinetobacter (12%) y Vibrio (8%) fueron los más abundantes en muestras wt, mientras que Pseudomonas (18%), Vibrio (3%) y Escherichia (3%) fueron los más abundantes en muestras cultivadas (Tabla Suplementaria S2). El LDA mostró que los phyla Actinobacteria, Firmicutes y TM7 fueron significativamente más abundantes en muestras wt, mientras que Proteobacteria, Gemmatimonadetes y Spirochaetes fueron más abundantes en muestras cultivadas (Tabla Suplementaria S9). Las clases, órdenes, familias, géneros y especies que fueron diferencialmente abundantes entre muestras wt y cultivadas se describen en la Fig. 5a y la Tabla Suplementaria S9. Las comunidades bacterianas con abundancia mayor a 0.1% (OTUs > 0.1%) fueron muy diferentes entre camarones wt y camarones cultivados sanos (prueba de correlación de Spearman, r = 0.2036, p = 0.0022). Este hallazgo coincidió con los observados en el análisis de árboles de PCoA y UPGMA. La comparación de LDA entre las vías predichas de la subcategoría de primer nivel mostró que los genes involucrados en los sistemas de los organismos y el metabolismo se enriquecieron significativamente en el camarón wt, mientras que el procesamiento de la información ambiental y genética se enriqueció significativamente en el camarón de cultivo. En el nivel dos, se observaron varias vías enriquecidas, incluyendo siete vías metabólicas (metabolismo de terpenoides y policétidos, metabolismo de nucleótidos, metabolismo de aminoácidos y lípidos, metabolismo de otros aminoácidos, metabolismo de cofactores y vitaminas, y metabolismo energético) y vías del sistema endocrino (Fig. 5b) en las muestras wt. Por el contrario, cinco vías predichas, incluyendo tres involucradas en el procesamiento de la información ambiental (transducción de señales, transporte de membrana y moléculas de señalización e interacción), se enriquecieron en las muestras de cultivo (Fig. 5b). El análisis de vías de mayor resolución (nivel tres) de los metagenomas predichos se describe en la Fig. Suplementaria S8b. El NSTI para hepatopáncreas wt y sano fue de 0.046 ± 0.005 y 0.052 ± 0.003, respectivamente (Tabla Suplementaria S8).
wt y sanos de cultivo. Para determinar la comunidad microbiana compartida entre las muestras de camarones wt y de cultivo, analizamos la presencia/ausencia de OTUs con una frecuencia >0.005. Este umbral de frecuencia se seleccionó para evitar la presencia de microorganismos transitorios en las muestras28. Se consideró que una OTU era compartida cuando aparecía en al menos una de las tres muestras de cada grupo. En este sentido, se encontró que 255 OTUs se compartían entre muestras de intestino (Fig. Suplementaria S9a), mientras que 158 OTUs se compartían entre muestras de hepatopáncreas (Fig. Suplementaria S9b). Además, se encontraron OTUs únicas que no se compartían entre las muestras wt y cultivadas (Fig. Suplementaria S9). Estas OTUs podrían representar poblaciones bacterianas transitorias, ya que podían aparecer solo en una de las tres muestras. Por lo tanto, para análisis posteriores, solo se seleccionaron como OTUs únicas las que presentaban lecturas en todas las réplicas biológicas de un órgano, pero no en ningún otro.
En este sentido, en la Tabla Suplementaria S10 se muestran las OTUs únicas con clasificaciones taxonómicas a nivel de género y especie en el intestino y el hepatopáncreas. Curiosamente, varios géneros, como Faecalibacterium, Bacteroides y Bifidobacterium, y especies, como Coprococcus eutactus y Faecalibacterium prausnitzii, fueron exclusivos del intestino a partir de muestras cultivadas (Tabla Suplementaria S10), mientras que los géneros Bacteroides y KSA1 y las especies Zobellella taiwanensis y Enterobacter cowanii fueron exclusivos del hepatopáncreas a partir de muestras cultivadas (Tabla Suplementaria S10).
Finalmente, analizamos las OTUs compartidas entre los intestinos, hepatopáncreas y sedimentos de muestras cultivadas y observamos un núcleo compartido compuesto por 187 OTUs. La mayoría de las OTUs se compartieron entre los sedimentos y los intestinos (286 OTUs), seguidas de 221 OTUs compartidas entre los intestinos y el hepatopáncreas, y 200 OTUs compartidas entre los sedimentos y el hepatopáncreas (Fig. Suplementaria S9c). El alto número de OTUs compartidas entre los sedimentos y los
Figura 5. Resultados de abundancia relativa y LEfSe de la microbiota enriquecida y funciones microbianas predichas entre muestras de hepatopáncreas wt (n = 3) y cultivadas (n = 3). (a) Abundancia relativa de especies con abundancia significativamente diferencial. La puntuación del análisis discriminante lineal (LDA) se muestra entre paréntesis. (b) LDA de las vías KEEG con abundancia diferencial. Las diferencias fueron significativas (p < 0.05) entre clases (prueba de Kruskal-Wallis) y subclases (prueba de Wilcoxon). El umbral de la puntuación logarítmica del LDA fue de 2.0.
intestinos sugirió una fuerte transferencia de bacterias entre estos dos órganos.
Impacto del desarrollo del brote de la AHPND/EMS en la estructura y función del microbioma del camarón. La microbiota intestinal desempeña un papel importante en la salud del huésped, contribuyendo al mantenimiento del equilibrio contra patógenos oportunistas. Para investigar el impacto de la enfermedad de la AHPND/ EMS en la microbiota del intestino y el hepatopáncreas, analizamos muestras de camarones de cultivo que presentaron síntomas de la AHPND/EMS e incluimos sedimento de estanque de donde se muestrearon camarones enfermos y sanos (Fig. 1a). Según las OTUs observadas, la PD y los índices de Shannon, los intestinos de los camarones enfermos tuvieron una diversidad bacteriana significativamente mayor (p < 0.001) que los de los camarones sanos (Fig. 2). Por el contrario, el hepatopáncreas enfermo tuvo una diversidad bacteriana significativamente menor (p < 0.001) (medida por las OTU observadas y la PD) que el hepatopáncreas de los camarones sanos (Fig. 2a y b). Solo el índice de Shannon mostró una mayor diversidad bacteriana en el hepatopáncreas enfermo en comparación con el de los camarones sanos (Fig. 2c). Los sedimentos de los estanques con camarones enfermos presentaron una diversidad bacteriana significativamente menor (p < 0.05) que los de los estanques con camarones sanos (Fig. Suplementaria S2).
El PCoA de las distancias UNIFRAC ponderadas y no ponderadas, y los árboles UPGMA con muestras etiquetadas como sanas o enfermas, no mostraron patrones de agrupamiento según los síntomas de la AHPND/EMS (Figs. Suplementarias S10 y S11). Se encontró un enriquecimiento diferencial de varias bacterias entre los intestinos enfermos y sanos (Tabla Suplementaria S11) y los hepatopáncreas (Tabla Suplementaria S12). Las espiroquetas y los verrucomicrobios se enriquecieron en los sedimentos de las muestras enfermas (Tabla Suplementaria S13). Vibrio shilonii y Faecalibacterium prausnitzii se enriquecieron en muestras de intestino sano, mientras que Aeromonas taiwanensis, Microbispora rosea y Simiduia agarivorans se enriquecieron en muestras de intestino enfermo (Fig. 6a). Pantoea agglomerans se enriqueció en hepatopáncreas sano, mientras que Photobacterium angustum se enriqueció en hepatopáncreas enfermo (Fig. 6b). Las especies diferencialmente enriquecidas observadas en los sedimentos se muestran en la Fig. 6c.
La mayoría de las OTUs (198) se compartieron entre el intestino, el hepatopáncreas y el sedimento del camarón enfermo (Fig. Suplementaria S12), el sedimento y el intestino compartieron 315 OTUs, seguidos por el sedimento y el hepatopáncreas, que compartieron 229 OTUs, y finalmente el intestino y el hepatopáncreas, que compartieron 220 OTUs. Crenothrix solo
se encontró en el intestino de organismos sanos, mientras que Microbispora, Shinella y Teredinibacter solo se encontraron en el intestino de muestras enfermas (Tabla Suplementaria S14). Escherichia solo se encontró en el hepatopáncreas de muestras enfermas (Tabla Suplementaria S14). Pseudomonas y Pseudomonas veronii solo se encontraron en el sedimento de muestras sanas, mientras que Inquilinus, Desulfobacter, Salinibacter, Flavobacterium, Halothiobacillus, Acidaminobacter, Alishewanella y Salinivibrio solo se encontraron en el sedimento de muestras enfermas (Tabla Suplementaria S14).
Se observaron diferencias significativas en los metagenomas predichos entre camarones de cultivo sanos y enfermos. En la subcategoría de primer nivel, los genes involucrados en el procesamiento de la información genética se enriquecieron significativamente en el hepatopáncreas de camarones enfermos. En la subcategoría de nivel dos, el metabolismo de nucleótidos se enriqueció, mientras que en la subcategoría de tercer nivel, los genes involucrados en la replicación y reparación, metabolismo de purinas y biosíntesis de valina, leucina e isoleucina, entre otros, se enriquecieron en el hepatopáncreas de camarones enfermos (Fig. Suplementaria S13a). En los intestinos de camarones enfermos, las vías de metabolismo de aminoácidos, lípidos y carbohidratos se enriquecieron significativamente en el nivel secundario, mientras que el ciclo del citrato (ciclo TCA) y la degradación de valina, leucina e isoleucina se enriquecieron en el nivel tres (Fig. Suplementaria S13b). Por el contrario, los intestinos de muestras sanas se enriquecieron en mecanismos de transducción de señales y vías de secreción pancreática en la subcategoría de nivel tres (Fig. Suplementaria S13b).
Finalmente, la comparación metagenómica entre los sedimentos de camarones sanos y enfermos solo resultó en diferencias significativas en seis vías en la subcategoría de nivel tres (Fig. Suplementaria S13c). El NSTI para intestinos sanos, hepatopáncreas y sedimentos fue de 0.112 ± 0.072, 0.052 ± 0.003 y 0.142 ± 0.027, respectivamente (Tabla Suplementaria S8). El NSTI para intestinos enfermos, hepatopáncreas y sedimentos fue de 0.1 ± 0.007, 0.050 ±
Figura 6. Abundancia relativa y resultados de LEfSe de las especies con abundancia diferencial entre camarones de cultivo sanos (n = 3) y enfermos (n = 3). (a) Intestino, (b) hepatopáncreas y (c) muestras de sedimento. La puntuación LDA se muestra entre paréntesis. Las diferencias fueron significativas (p < 0.05) entre clases (prueba de Kruskal-Wallis) y subclases (prueba de Wilcoxon). El umbral de la puntuación LDA logarítmica fue de 2.0.
0.003 y 0.132 ± 0.029, respectivamente (Tabla Suplementaria S8).
Los resultados del método AP3 para la amplificación del gen ToxA y la detección de la AHPND/EMS fueron negativos para todos los órganos intestinales y hepatopáncreas de las muestras wt (Tabla Suplementaria S15). Por lo tanto, el camarón wt no presentó el plásmido asociado a la AHPND/EMS. Todos los hepatopáncreas de las muestras cultivadas dieron positivo para la presencia del plásmido. Por el contrario, dos intestinos de muestras sanas cultivadas y un intestino de muestras enfermas cultivadas arrojaron resultados negativos, mientras que un intestino de muestras sanas cultivadas y dos intestinos de muestras enfermas cultivadas
arrojaron resultados positivos. Este hallazgo es previsible en las primeras etapas de la AHPND/EMS, ya que los tejidos con mayor densidad bacteriana durante la enfermedad son el hepatopáncreas y el estómago29
Discusión
El camarón blanco del Pacífico, Penaeus vannamei, es la especie de camarón de mayor importancia económica a nivel mundial. En este trabajo, caracterizamos la microbiota y predijimos sus funciones metagenómicas en muestras de hepatopáncreas e intestino de ambiente natural y camarones P. vannamei de cultivo sanos y enfermos. Dado que el suelo es un componente fundamental del microbioma del camarón, también incluimos sedimento de estanques de los laboratorios
de camarón. La secuenciación de siete regiones hipervariables del gen ARNr 16S, utilizando el Ion 16STM Metagenomics Kit, se ha reportado previamente para otros modelos30,31, lo que sugiere que se necesita más de una región hipervariable para proporcionar inferencias confiables de diversidad30. Estudios previos en la microbiota del camarón solo se enfocaron en dos regiones hipervariables (típicamente V1-V3 o V3-V4)6,8,11,14,16. En este trabajo, reportamos la primera caracterización de la microbiota del camarón que incorpora la secuenciación de siete regiones hipervariables mediante el kit de metagenómica Ion 16STM, evitando al mismo tiempo el sesgo de diversidad inherente a la selección de solo una o dos regiones hipervariables del gen2,31 Sin embargo, es importante destacar que también encontramos varias OTUs que fueron clasificadas utilizando solo una de las regiones hipervariables (Fig. Suplementaria S3), lo que sugiere: (i) que diferentes regiones hipervariables del mismo gen ribosomal podrían asignarse a diferentes taxonomías; o (ii) que existen genes ribosomales que solo pueden clasificarse utilizando una región hipervariable. Se ha observado un comportamiento similar al utilizar varias regiones hipervariables para clasificar la microbiota en otros estudios31,32
Para comprender las consecuencias de estos sesgos en el análisis de la microbiota, es necesario realizar más pruebas con el kit de metagenómica Ion 16STM utilizando muestras control bien caracterizadas que contengan cepas bacterianas conocidas en proporciones bien cuantificadas. Considerando que comparamos amplicones no superpuestos de siete regiones hipervariables (V2, V3, V4, V6-V7, V8 y V9) del gen ARNr 16S y que esta comparación no permitió la identificación de nuevos organismos en un proceso de selección de OTUs de novo, solo pudimos detectar los microorganismos reportados en la base de datos GreenGenes. Por lo tanto, todavía existe la oportunidad de investigar más a fondo el papel que juega la nueva diversidad bacteriana en la composición de la microbiota de este modelo. También se han observado diferencias en la abundancia de lecturas para algunas regiones hipervariables utilizando este kit30,31, y también se observaron en nuestro estudio (Fig. Suplementaria S2). Curiosamente,
las diferencias se asociaron con el origen de la muestra (es decir, wt o cultivada) y el órgano analizado (es decir, hepatopáncreas o intestino). Estas diferencias podrían sugerir abundancias distintas de microorganismos específicos, pero podrían normalizarse utilizando las secuencias de las siete regiones hipervariables. El uso de diferentes regiones hipervariables o errores de secuenciación, como los que surgen de la pirosecuenciación y los secuenciadores de moléculas individuales (Roche 454 y PacBio), puede llevar a una inflación artificial de las estimaciones de diversidad33,34. Es bien sabido que la data de Ion Torrent exhibe una mayor tasa de errores de secuenciación, en comparación con la data de la plataforma Illumina7–9,35–37. Sin embargo, un estudio reciente que utilizó Illumina MiSeq e Ion Torrent para el perfilado de la comunidad bacteriana mediante la secuenciación de amplicones de ARNr 16S mostró que la diferencia absoluta en las tasas de error entre las dos plataformas no fue significativa35
Nuestras curvas de rarefacción no alcanzaron la saturación incluso después de 16,000 lecturas (Fig. Suplementaria S5b). Si bien la cobertura de Good reveló que obtuvimos, en promedio, aproximadamente el 80% del total de OTUs con una profundidad de secuenciación de 2,078 lecturas (Tabla Suplementaria S5), se requiere una profundidad de secuenciación significativamente mayor para descubrir el total de OTUs en los microbiomas analizados. Esta data sugiere que se requieren más investigaciones con muestras control bien caracterizadas para cuantificar el impacto real que la combinación de diferentes regiones hipervariables podría tener en la sobreestimación de la diversidad.
En general, encontramos una diversidad significativamente menor en el hepatopáncreas que en el intestino, independientemente del origen del camarón (wt o de cultivo), a diferencia de lo estimado previamente en N. denticulata16, donde los autores reportaron que el hepatopáncreas presenta mayor diversidad bacteriana que el intestino. Las condiciones controladas de laboratorio utilizadas en el estudio de N. denticulata podrían explicar estas diferencias, ya que, en este trabajo, las muestras wt se recolectaron en mar
abierto y los especímenes de cultivo se obtuvieron en condiciones de laboratorio. Las proteobacterias representaron aproximadamente el ~76, ~95 y ~50% de todas las lecturas de muestras de intestino, hepatopáncreas y sedimento, respectivamente. El predominio de proteobacterias en camarones se había observado previamente, pero solo en camarones cultivados en condiciones de laboratorio6,8,11,14,16. La similitud encontrada entre la microbiota intestinal y la del sedimento de camarones de cultivo fue consistente con el comportamiento de excavación, bien descrito, en el que los camarones pasan largos períodos de tiempo enterrados en el sedimento. En este sentido, Xing et al.38 sugirieron varias vías que explican cómo el metagenoma gastrointestinal del lenguado adulto de cultivo se origina a partir del sedimento38. Por lo tanto, la gestión basada en el conocimiento del sedimento podría conducir a una modificación de la microbiota del camarón y a estrategias ecológicamente sólidas para promover el estado sanitario del cultivo de camarón.
Las OTUs compartidas entre el camarón wt y el camarón de cultivo podrían ser bacterias seleccionadas por la fisiología y la genética del huésped, mientras que las OTUs únicas en las muestras wt o cultivadas podrían estar influenciadas por el entorno (natural vs. cultivado) y por las condiciones de alimentación (detritos naturales y microalgas vs. dietas formuladas). Los análisis de PCoA y UPGMA mostraron que la microbiota wt es diferente de la microbiota cultivada y que la microbiota del mismo órgano es más similar entre camarones del mismo origen (wt o cultivado) que la microbiota de diferentes órganos del mismo camarón, en consonancia con las observaciones en animales superiores, como el ser humano, el koala y otros39,40.
Estos resultados también indicaron que el entorno tiene una mayor influencia en la diversidad bacteriana que el órgano del camarón. Sin embargo, en el árbol UPGMA de las muestras wt, un hepatopáncreas se agrupó con muestras de intestino, y se observó el mismo fenómeno para otro intestino que se agrupó con muestras de hepatopáncreas (Fig. 3c). Los perfiles bacterianos de las OTUs superiores al 0.1
% (OTUs > 0.1 %) mostraron correlaciones bajas de Spearman para estas dos muestras entre sus réplicas (Tabla Suplementaria S3). Adicionalmente, las correlaciones de OTU (OTUs > 0.1%) entre los triplicados wt fueron bajas y variaron de 0.38 a 0.87, mientras que las correlaciones entre los triplicados cultivados fueron altas y estables, variando de 0.77 a 0.94 (Tabla Suplementaria S3).
El hecho de que las muestras wt tuvieran correlaciones más bajas entre réplicas podría sugerir que la heterogeneidad del alimento en condiciones naturales podría tener un impacto en la composición de la microbiota, en comparación con las correlaciones estables observadas en camarones cultivados que viven en condiciones de alimento controladas. Existe la noción general de que los animales en sus entornos naturales tienen una microbiota más rica41.
En este sentido, el intestino del camarón tigre wt P. monodon tiene más diversidad bacteriana que el de los camarones cultivados11. De acuerdo con esta hipótesis general, nuestros camarones domesticados cultivados en condiciones controladas con dietas formuladas y sistemas de manejo de agua tienen una diversidad bacteriana menor que los organismos wt. Estas condiciones controladas también se reflejan en las correlaciones de OTUs más altas y similares entre nuestros triplicados de cultivo, a diferencia de las observadas en las muestras wt (Tabla Suplementaria S3), lo que también sugiere la homogeneidad de la microbiota del camarón en condiciones de cultivo.
Recientemente, Xiong et al.24 reportaron que el desarrollo del camarón desde nauplios hasta organismos adultos puede explicar solo el 9.7 % de la variación de las comunidades bacterianas intestinales en P. vannamei. Además, identificaron 28 taxones discriminatorios por edad para el camarón. Encontramos 7 de estos 28 taxones en nuestra data de microbiota (Materiales y métodos). Debido a que los camarones muestreados para este trabajo tenían diferentes tamaños (~44 g en peso y ~16 g en cultivo) y, por lo tanto, probablemente diferentes edades adultas, realizamos un análisis de β-diversidad entre todas las muestras, eliminando estos 7 taxones discriminatorios por edad (Tabla Suplementaria S16). Observamos que la
agrupación de la comunidad era muy similar a la observada incluyendo estos taxones asociados con la edad del camarón (Figura Suplementaria S14), lo que sugiere que una diferencia en la edad del camarón no agrupó la microbiota en nuestras muestras.
Junto con la caracterización de la microbiota de todas las muestras de camarón, utilizamos la data de secuenciación del gen 16S para predecir las funciones del metagenoma. La precisión de estas predicciones con PICRUSt depende de la estrecha relación entre los microorganismos de la muestra estudiada y los microorganismos con genomas ya secuenciados. Esta precisión se mide mediante el valor de NSTI, donde los valores más bajos de NSTI indican una relación más estrecha con los genomas secuenciados. Como era de esperar, los valores de NSTI fueron mayores para los sedimentos filogenéticamente más diversos (0.137 ± 0.026), intermedios para los intestinos (0.098 ± 0.043) y menores para el hepatopáncreas (0.049 ± 0.004). A modo de comparación, en el estudio de Langille et al.27, se encontró que los intestinos de mamíferos tenían una NSTI = 0.14, las muestras de suelo tenían una NSTI = 0.17, mientras que las muestras de microbioma humano bien cubiertas tenían una NSTI = 0.03. Por lo tanto, nuestras muestras de camarones y sedimentos coinciden con la NSTI descrita en otros microbiomas27,42,43
El hepatopáncreas es la principal glándula digestiva de los invertebrados e integra numerosas funciones digestivas, metabólicas e inmunitarias17. El sistema de depuración de patógenos y procesamiento de antígenos44 contiene moléculas relacionadas con la inmunidad innata, como lectinas, hemocianina, ferritina, péptidos y proteínas antivirales y antibacterianos, enzimas proteolíticas y óxido nítrico17. La menor diversidad observada en el hepatopáncreas sugiere que la inmunidad innata restringe la microbiota natural a bacterias que no activan los mecanismos de protección del huésped contra los patógenos. Curiosamente, esta microbiota parece conservarse entre especies, ya que se ha reportado que dichos mecanismos inmunitarios ocurren en el intestino del ratón45. Además, se sabe que los ácidos grasos de cadena corta (SCFAs) y los metabolitos del triptófano desempeñan
un papel importante en la homeostasis de las células inmunitarias adaptativas en varios organismos46. De forma consistente, encontramos genes predichos que se enriquecieron para el metabolismo del butirato y el propionato, y para la biosíntesis de triptófano, en la microbiota del hepatopáncreas wt, lo que podría modular la diversidad bacteriana y desempeñar un papel protector. También encontramos varias vías, como el metabolismo de aminoácidos, cofactores, vitaminas, nucleótidos, lípidos, terpenoides, energía y el sistema endocrino, además de moléculas de señalización, que se enriquecieron en el hepatopáncreas wt.
Las vías para la degradación de aminoácidos ramificados (valina, leucina e isoleucina) se enriquecieron significativamente en el hepatopáncreas y el intestino del camarón wt. Estos aminoácidos son componentes importantes de la hemocianina, la proteína respiratoria en crustáceos y moluscos47, que también tiene una función en la respuesta inmune48. Todas las funciones derivadas de la microbiota mencionadas anteriormente no se encontraron en camarones de cultivo sanos.
Los linajes actuales de camarones utilizados en laboratorios comerciales han sido seleccionados mediante métodos tradicionales de selección genética durante décadas para obtener mejores tasas de conversión alimenticia, crecimiento y resistencia a enfermedades, lo que también podría haber cambiado la microbiota natural49. Estos cambios han sido mucho menos estudiados y comprendidos en el camarón debido a que existen diferencias fisiológicas críticas entre los peces vertebrados y los camarones invertebrados. Además, la pérdida de diversidad bacteriana podría favorecer la aparición de enfermedades oportunistas15. De manera similar, se observó en primates no humanos mantenidos en cautiverio que existía una asociación entre la pérdida de la microbiota intestinal natural y la convergencia hacia la microbiota humana41
La composición de la dieta modula la microbiota49. Los carbohidratos complejos son críticos debido a su función como sustrato prebiótico para el crecimiento bacteriano y en la señalización. Los invertebrados
poseen mecanismos generales para detectar estructuras específicas, como los betaglucanos, y para desencadenar respuestas inmunitarias innatas. Los glicanos suelen estar presentes en los alimentos comerciales para camarón que se utilizan en los laboratorios de acuicultura, y son un factor importante que configura la composición de la microbiota de muchas especies, incluidos los humanos50. Curiosamente, los genes previstos para la biosíntesis y el metabolismo de los glicanos se enriquecieron en el intestino y el hepatopáncreas de camarones de cultivo sanos (Fig. Suplementaria S8), lo que sugiere que la microbiota del intestino y el hepatopáncreas podría responder a los cambios dietéticos de glicanos, lo que permitiría manipular las comunidades bacterianas para promover la salud del huésped añadiendo prebióticos de glicanos naturales a las dietas tradicionales para camarones. Además, el aumento del metabolismo de xenobióticos observado en la microbiota intestinal del camarón wt apunta a la necesidad de gestionar la contaminación ambiental marina.
En cuanto a la enfermedad, no existe una línea clara que establezca las etapas iniciales de la AHPND/EMS. El diagnóstico se basa en un ensayo de PCR que detecta el plásmido que codifica las toxinas Pir A/B. La producción de las toxinas dependiente de la detección de quórum (quorum sensing)29,, y hasta la fecha, no existen métodos para detectar directamente la presencia de las proteínas Pir A/B. En todas las muestras wt, los resultados de la PCR-AP3 fueron negativos, lo que indica la ausencia del plásmido en estas muestras.
Los resultados de la PCR-AP3 para camarones de cultivo fueron controvertidos debido a que la PCR-AP3 fue positiva en 4 de 6 muestras sanas y en 5 de 6 muestras enfermas (Tabla Suplementaria S15). Sin embargo, una cepa no patógena de V. parahaemolyticus también ha arrojado previamente resultados positivos para el diagnóstico de AP329. Sin embargo, ninguno de los camarones cultivados sanos mostró síntomas de la AHPND/ EMS (Fig. Suplementaria S15). Además, los organismos de los estanques enfermos presentaron los síntomas y la mortalidad de la enfermedad, incluyendo letargo, intestino vacío y hepatopáncreas acuoso pálido y
blanco (Fig. Suplementaria S15).
Estos resultados sugirieron que la presencia de las toxinas Pir A/B que codifican plásmidos requiere factores desencadenantes adicionales para conducir a la enfermedad AHPND/EMS. Como se mencionó, se sabe que el quórum sensing es un factor desencadenante para AHPND/ EMS ya que se ha reportado que la densidad infectiva umbral es de 104 CFU ml−1, y menor que esa densidad bacteriana, no ocurrió mortalidad29. También observamos una diferencia no significativa en la agrupación de la comunidad bacteriana entre muestras sanas y enfermas (Fig. 3). Curiosamente, este hallazgo concordó con un estudio reciente en el que no se observaron cambios significativos en la agrupación ni la diversidad de la comunidad bacteriana en las primeras etapas de una enfermedad en P. vannamei24 .
En general, y en todas las especies, la pérdida de diversidad se asocia con enfermedades, como se ha demostrado en humanos2 Observamos que la AHPND/EMS se asoció con una pérdida significativa de la diversidad de la microbiota del hepatopáncreas, en comparación con los camarones sanos (p < 0.01), medida mediante PD y el número de OTUs (Fig. 2), aunque la AHPND/EMS se asoció con una toxina bacteriana y no con una infección bacteriana per se18. Se observó un efecto opuesto en el intestino enfermo, donde la diversidad de la microbiota aumentó significativamente (p < 0.01) como consecuencia de la enfermedad AHPND/EMS (Fig. 2). Cabe destacar que una limitación de nuestro estudio fue el pequeño tamaño de la muestra (n = 3) y la baja profundidad de secuencia, lo que hace necesario estudiar más muestras en futuros estudios para confirmar estos cambios en la diversidad bacteriana. Sin embargo, nuestro análisis lleva a la noción de que algunas bacterias se pierden durante la domesticación del camarón (wt vs. cultivado), lo que podría resultar en la necesidad de producir cepas probióticas para ser administradas en organismos cultivados para promover la salud del camarón.
Con respecto a esto, encontramos algunas bacterias enriquecidas tanto en muestras wt como sanas, lo que sugiere su uso como probióticos para P. vannamei
Primero, algunos miembros de los géneros Janthinobacterium, Dietzia, Streptomyces y Bacillus, que produjeron antibióticos y moléculas antifúngicas51,52 solo se encontraron en los intestinos de camarones wt. Adicionalmente, solo en estas muestras estuvo presente Pseudomonas stutzeri, que ha sido reportada como un probiótico que protege contra Vibrio patógeno en Artemia53 y mejora el crecimiento, supervivencia e inmunidad de P. monodon54. Además, se sabe que varias especies del género Micrococcus poseen potencial probiótico al inhibir la colonización intestinal por patógenos, como se observó en larvas juveniles de L. vannamei55 y Macrobrachium rosenbergii56 Curiosamente, el género Microccocus solo estaba presente en el hepatopáncreas de camarones wt.
Por el contrario, las bacterias de los géneros Faecalibacterium y Bifidobacterium, que se utilizan típicamente como probióticos en humanos57,58, se enriquecieron en los intestinos de camarones de cultivo sanos. Específicamente, Faecalibacterium prausnitzii solo estaba presente y se enriqueció significativamente en los intestinos de camarones de cultivo sanos, en comparación con los mismos órganos en camarones enfermos. Esta bacteria productora de butirato es la más abundante en la microbiota intestinal de adultos sanos, en la que los cambios en su abundancia se han relacionado con varios trastornos humanos59,60
En consecuencia, nuestros resultados sugirieron que esta bacteria podría tener efectos beneficiosos en la salud de los camarones y podría usarse como probiótico para P. vannamei afectado por la AHPND/ EMS. Además, Pantoea agglomerans se enriqueció en el hepatopáncreas de camarones de cultivo sanos versus enfermos. Se ha reportado que la administración oral de lipopolisacárido (LPS) derivado de esta bacteria mejora la resistencia a enfermedades contra la viremia aguda de peneidos en la hemolinfa del camarón kuruma, Penaeus japonicus61. Además, el LPS de otras bacterias se ha utilizado previamente como un “inmunoestimulante” en P. vannamei, pero principalmente contra infecciones virales62. Nuestros resultados sugieren la idea de que P. agglomerans
podría utilizarse como un posible probiótico contra la enfermedad AHPND/EMS en P. vannamei.
El género Aeromonas es un importante patógeno causante de enfermedades en peces y es responsable de complicaciones infecciosas tanto en humanos inmunocompetentes como inmunocomprometidos63. Aquí, reportamos del enriquecimiento de Aeromonas taiwanensis en el intestino de camarones enfermos. Esta bacteria se ha reportado en ambientes acuáticos en todo el mundo y causa enfermedades humanas64, pero no se ha reportado en camarones. Simiduia agarivorans es una bacteria marina capaz de degradar diversos polisacáridos complejos, como la quitina65, un biopolímero abundante en hábitats acuáticos (p. ej., en los exoesqueletos de camarones). Además, se ha demostrado que V. parahemolyticus puede adsorberse y multiplicarse en la quitina y sus derivados solubles66. Por lo tanto, el enriquecimiento de S. agarivorans en los intestinos de camarones enfermos podría estar relacionado con la presencia de V. parahemolyticus en ellos.
Otras vías metabólicas interesantes fueron la biosíntesis y degradación de aminoácidos ramificados, que se sabe que aumentan durante los procesos de infección en otros organismos, incluidos los humanos67
Curiosamente, estas vías se enriquecieron en los microbiomas del hepatopáncreas e intestino enfermos, lo que podría estar asociado con el proceso patológico de la APHND/EMS. Las especies de Photobacterium son ubicuas en el agua de mar, organismos marinos, sedimentos marinos y lagos salinos68 P. damselae se considera un patógeno bacteriano de humanos y muchos organismos acuáticos, incluyendo peces, moluscos y crustáceos68. Recientemente, se han reportado varias cepas de P. damselae que causan mortalidad en camarones Exopalaemon carnicauda, P. monodon y P. vannamei69,70. Curiosamente, los síntomas de los camarones afectados por estas bacterias fueron muy similares a los observados en el hepatopáncreas de camarones con la AHPND/EMS causado por V. parahaemolyticus69. En nuestro estudio, P. angustum se enriqueció en el
hepatopáncreas de camarones enfermos (Fig. 6b), y aunque no se ha reportado que esta bacteria sea un patógeno para camarones, nuestros resultados enfatizan que estudios adicionales aclararán su papel en el desarrollo de la enfermedad AHPND/ EMS.Finalmente, encontramos algunas bacterias enriquecidas en el sedimento del estanque. Primero, Acinetobacter johnsonii, que reduce el fósforo en el agua en un 10-30% y libera fosfato inorgánico en condiciones anaeróbicas, se enriqueció en el sedimento de camarones sanos. También se enriqueció en estas muestras Pseudomonas veronii, que se ha utilizado para la biorremediación de suelos contaminados con compuestos orgánicos aromáticos71. Estas dos bacterias podrían usarse potencialmente para tratar la enfermedad AHPND/EMS. Por el contrario, Pseudoalteromonas ruthenica, una bacteria de biopelícula productora de exopolímeros, se enriqueció en sedimentos enfermos. Esta bacteria se ha aislado en el agua de mar de la entrada de una estación de bombeo en zona tropical72. Sin embargo, se necesitarán más investigaciones para analizar los roles de estas especies enriquecidas en camarones sanos y enfermos y sus sedimentos para comprender completamente sus roles en la enfermedad AHPND/EMS.
Materiales y métodos
Declaración ética. No se requirió una declaración ética para este trabajo ni permisos específicos para el estudio descrito. El lugar de recolección de especímenes no es de propiedad privada ni está protegido de ninguna manera, y los estudios de campo no involucraron especies en peligro de extinción o protegidas. Los animales fueron sacrificados según los protocolos universitarios para evitar su sufrimiento.
Recolección de muestras. Todos los especímenes se identificaron como Penaeus vannamei mediante claves morfológicas73. Los camarones (peso, n = 3, peso promedio = 43.8 ± 2.1 g) (Tabla Suplementaria S17) se obtuvieron del Pacífico mexicano a 5 km de la costa del estado de Nayarit, a 21.4986° de latitud y −105.3169° de longitud (Fig. 1a), utilizando un sistema tradicional de pesca de arrastre de camarón a 15 m de profundidad (salinidad = 32 ppm y temperatura = 24°C). Los camarones de cultivo se recolectaron de un laboratorio de
camarones ubicado a 1000 km del sitio de la recolección de camarones wt (estado de Sonora, 28.4030° de latitud y −111.4513° de longitud, Fig. 1a). Los camarones de cultivo enfermos (n = 3, peso promedio = 15.3 ± 0.4 g) (Tabla Suplementaria S17) se obtuvieron de un estanque en el que los camarones presentaron mortalidad y tenían los síntomas clínicos y patológicos habituales de la enfermedad, como letargo, intestino vacío y hepatopáncreas acuoso pálido y blanco (Fig. Suplementaria S15).
Los tres camarones seleccionados mostraron síntomas claros de la enfermedad (Fig. Suplementaria S15). Los camarones de cultivo sanos (n = 3, peso promedio = 17.1 ± 1.2 g) (Tabla Suplementaria S17) se obtuvieron de un estanque sin la sintomatología de la enfermedad AHPND/ EMS. Después de una prueba ANOVA de una vía, no hubo tendencias entre los camarones de diferente peso dentro del mismo grupo (Tabla Suplementaria S17). Los camarones de cultivo fueron alimentados tres veces al día con alimento comercial (~35% de proteína) durante tres meses (salinidad del agua ~40 ppm y temperatura ~29°C). Los hepatopáncreas e intestinos de los camarones se diseccionaron asépticamente in situ.
Los tejidos se mantuvieron en solución RNAlater durante 24 h (~4°C) y se almacenaron a -80°C hasta su uso. Además, se recolectaron tres muestras de sedimento (50 g) de cada estanque utilizando un núcleo de sedimento (3.8 cm de diámetro por 10 cm de profundidad). Las muestras se homogeneizaron y se almacenaron inmediatamente en nitrógeno líquido. Todas las muestras se analizaron individualmente mediante el método de PCR diagnóstica para la AHPND/EMS74. Las muestras de camarón de cultivo que dieron positivo en la prueba AP3 se etiquetaron como EMS+, y las muestras negativas, como EMS- (Tabla Suplementaria S15).
Extracción de ADN de muestra, preparación de la biblioteca y secuenciación. El ADN total se extrajo de todas las muestras utilizando el ZR Soil Microbe DNA MicroPrep (Zymo Research, Irvine, CA, EE.UU.). La concentración de ADN se determinó utilizando un fluorómetro Qubit (Life Technologies,
Carlsbad, CA, EE.UU.) y la integridad del ADN se confirmó mediante electroforesis en gel de agarosa. Los amplicones se generaron como se describe en la guía del usuario del Ion 16STM Metagenomics Kit (Thermo Fisher, Waltham, MA, EE.UU.) con algunas modificaciones. Brevemente, los amplicones se prepararon utilizando 200 ng de ADN total de muestras de intestino y hepatopáncreas y 300 ng de ADN total de muestras de sedimento.
Se realizaron dos reacciones para cada muestra, una para el conjunto de primers V2-4-8 (mezcla V2) y otra para el conjunto de primers V3-6-7-9 (mezcla V3). Todas las reacciones se ajustaron a un volumen final de 20 µl y se amplificaron durante 35 ciclos siguiendo las condiciones del termociclador del fabricante. Tras la amplificación, cada reacción se realizó en un gel de agarosa de electroforesis, observándose en todos los casos bandas de 200-300 pb.
Todos los amplicones se purificaron directamente del gel de agarosa utilizando el kit de extracción de gel QIAquick (Qiagen, Venlo, Países Bajos) y se cuantificaron con el fluorómetro Qubit. A continuación, con base en las cuantificaciones, se mezclaron cantidades equimolares de V2 y V3 en cada muestra, y la mezcla se utilizó para preparar las bibliotecas de secuenciación Ion Torrent, como se describe en la Guía del usuario del kit de metagenómica Ion 16STM La cantidad y la distribución del tamaño de cada biblioteca se evaluaron utilizando el fluorómetro Qubit y el bioanalizador Agilent 2100 (Agilent Technologies, Santa Clara, CA, EE. UU.). Finalmente, todas las bibliotecas se secuenciaron utilizando la tecnología Ion PGMTM Sequencing 400 en dos chips Ion 316™ v2 (Thermo Fisher, Waltham, MA, EE.UU.).
Bioinformática y análisis estadístico. Las lecturas de secuenciación se generaron utilizando la química de longitud de lectura de 400 bases del sistema Ion PGM. Tras el filtrado de calidad (≥Q25) y la eliminación de bases ambiguas, se obtuvieron 1,060,822 lecturas. Tras el recorte de lecturas (≥120 pb), se conservaron 305,209 lecturas y se analizaron con QIIME75 (versión 1.8). Estas secuencias se agruparon mediante UCLUST en unidades taxonómicas operativas (OTUs),
basándose en una similitud de secuencia del 97% mediante el comando pick_closed_ reference_otus.py con la colección de secuencias de referencia de Green Genes (versión 13_5).
Se habilitó la opción de coincidencia de cadena inversa y cualquier lectura que no coincidiera con una secuencia en la colección de secuencias de referencia se excluyó de los análisis posteriores. Para asignar taxonomía a las OTUs, proporcionamos asignaciones taxonómicas para la base de datos de referencia Green Genes. Seleccionamos el comando de selección de OTUs de referencia cerrada porque estábamos comparando amplicones que no se superponían.
Este método de selección de OTUs fue un enfoque basado en la referencia. Por lo tanto, no fue necesaria la eliminación de quimeras. Se recuperó un total de 153,361 lecturas filtradas de alta calidad con una longitud de lectura media de 215 pb para las 24 muestras con números de secuencia que oscilaban entre 1,032 y 15,814 (Tabla Suplementaria S4).
Las OTUs representadas por una sola lectura (singleton) se descartaron de los análisis posteriores, lo que ayudó a mantener las estimaciones de la diversidad α realistas y a evitar la pérdida de información. Posteriormente, se generaron resúmenes de taxonomía con datos de abundancia relativa mediante el comando summary_taxa_ through_plots.py.
La secuencia más abundante dentro de una OTU se seleccionó como representativa de la misma, y su secuencia se obtuvo mediante el comando pick_rep_set.py. Las secuencias representativas se alinearon con Green Genes mediante el comando align_seqs.py y PyNAST con una identidad de secuencia mínima del 75%. La alineación se filtró mediante filer_alignment.py y se construyó un árbol filogenético mediante el comando make_phylogeny.py y el método FastTree.
Las métricas de diversidad alfa y beta de la tabla OTU final sin singletons se obtuvieron utilizando los comandos QIIME75 alpha_ rarefaction.py y beta_diversity_through_ plots.py, respectivamente. Las curvas de rarefacción hasta la biblioteca más profunda
(15,000 lecturas) se muestran en la Fig. S5b. Para aumentar la profundidad de la secuencia, descartamos dos muestras con baja profundidad de secuenciación (68B-S y 139-I). Por lo tanto, las métricas de diversidad alfa se calcularon a una profundidad de secuencia de 2,078 lecturas por muestra con 10,000 iteraciones y luego se promediaron.
La profundidad de muestreo máxima seleccionada correspondió al número mínimo de lecturas obtenidas de cualquiera de las muestras secuenciadas restantes. La diversidad beta se estimó calculando a partir del árbol filogenético las distancias UniFrac ponderadas y no ponderadas entre las muestras a una profundidad de secuencia de 1,032 lecturas por muestra, y las matrices de distancia UniFrac se visualizaron utilizando el análisis PCoA.
El umbral de profundidad de secuencia de 1,032 lecturas se eligió para incluir todas las muestras para el análisis de diversidad beta. La robustez del árbol UPGMA se estimó con jackknife, con base en 1,000 réplicas y utilizando los comandos beta_diversity. py, multiple_rarefactions_even_depth.py, upgma_cluster.py y tree_compare.py. Para determinar las clasificaciones taxonómicas que fueron significativamente más abundantes en las muestras cultivadas y wt, aplicamos una prueba de suma de rangos no paramétrica de Wilcoxon, seguida de LDA utilizando el programa LEfSe26
Las comparaciones de diversidad alfa se evaluaron mediante la prueba de MannWhitney (prueba t no paramétrica) con un nivel de confianza del 95% (p < 0.05). La tabla final de OTU también se utilizó como dato de entrada para la predicción metagenómica funcional mediante PICRUSt27
El contenido de la vía KEGG obtenido por PICRUSt se normalizó y se utilizó para obtener las predicciones funcionales metagenómicas en diferentes niveles jerárquicos de KEGG (1, 2 y 3). Además, se utilizó la opción -a en el comando predict_metagenomes.py de PICRUSt para calcular el valor NSTI de las predicciones funcionales en cada muestra.
Las OTUs y las vías predichas se sometieron a un análisis de abundancia diferencial
con LEfSe para obtener las OTUs y las vías que presentaron diferencias significativas entre las muestras. Se utilizó un nivel de significación (alfa) de 0.05 y el umbral LDA > 2 para determinar las OTUs y las vías significativamente abundantes. Posteriormente, aplicamos una prueba de tasa de falsos descubrimientos (FDR) para priorizar los resultados de enriquecimiento más significativos de LEfSe, sin embargo, no se detectaron diferencias significativas entre todos los grupos, probablemente debido al pequeño número de réplicas de cada grupo (n = 3)76,77
Xiong et al.24 reportaron los 28 taxones con capacidad de discriminación por edad en un análisis de novo. Por lo tanto, para compararlo con nuestro estudio, que utilizó un análisis cerrado, los taxones se compararon con la base de datos GreenGenes (versión 13_5) para identificar los genes ribosómicos correspondientes en nuestra base de datos.
Posteriormente, solo encontramos 21 OTUs con una identidad de secuencia ≥97%,
de las cuales solo 7 OTUs presentaron abundancia de lecturas en nuestra data (Tabla Suplementaria S16). Por lo tanto, se realizó el análisis de β-diversidad entre todas las muestras, eliminando estas OTUs con capacidad de discriminación por edad de nuestra tabla biom original.
Separación de lecturas en diferentes regiones hipervariables. El kit de metagenómica 16S de Ion Torrent contiene seis pares de primers patentados diseñados para amplificar siete regiones hipervariables diferentes (V2, V4 y V8 en una reacción de PCR y V3, V6-7 y V9 en una segunda reacción de PCR). Dado que la empresa no publicó las secuencias de primers, requeríamos una referencia de genes ribosomales para la alineación. Para ello, dividimos las secuencias de ARNr 16S alineadas de longitud completa de la referencia de GreenGenes (versión 13_5), utilizando los puntos de corte informados previamente para las regiones V2, V3, V4, V6-V7, V8 y V978
Estas subregiones se dividieron en 6 archivos que se utilizaron como referencia de ARNr 16S para cada región hipervariable. Las 153,361 lecturas previas con taxonomías asignadas se agruparon mediante UCLUST en OTUs, basándose en una similitud de secuencia del 97%, utilizando el comando pick_closed_reference_otus.py para cada uno de los seis archivos de regiones hipervariables.
Se habilitó la opción de coincidencia de cadena inversa y el proceso se ejecutó seis veces para obtener tablas de OTU para cada región hipervariable. La taxonomía se asignó mediante el comando assignment_taxonomy. py en QIIME, y se obtuvieron las abundancias relativas para cada región hipervariable y para cada muestra secuenciada•
Para más información sobre este artículo, contactar a: rrs@ciad.mx
ESTADÍSTICAS ÍNDICE
Edición 165 - Junio 2025
77 80 Exportaciones de camarón Reporte de mercado de EE. UU.
CAMARÓN
Estadísticas de Comercio Exterior
Elaborado por: María Andrea Dicindio - Subgerente de Comercio Exterior de la Cámara Nacional de Acuacultura
EVOLUCIÓN DE EXPORTACIONES 2010 - 2024
La gráfica presentada muestra un crecimiento constante en el volumen y valor de las exportaciones de camarón, con un incremento notable hasta 2022, cuando se alcanzó el pico máximo de ingresos. Aunque a partir de 2023 el volumen exportado se mantiene en niveles récord, el 2024 muestra una ligera reducción en el valor de las exportaciones que se da, principalmente, por ajustes en los mercados internacionales. Este desempeño resalta el fortalecimiento del sector camaronero, evidenciando su capacidad productiva y consolidación en el mercado global, así como la necesidad de estrategias para mantener la rentabilidad ante fluctuaciones comerciales con los principales mercados de destino.
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
ESTADÍSTICAS
MILLONES DE LIBRAS EXPORTADAS: COMPARATIVO MENSUAL 2021 AL 2025
Las exportaciones de camarón han mostrado un incremento constante desde 2021 hasta 2025 en casi todos los meses, con algunos meses alcanzando picos considerables.
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
En mayo de 2024, Ecuador alcanzó un récord histórico al exportar 275 millones de libras de camarón, un hito impulsado por la optimización de su producción para obtener camarones de tallas más grandes. Este logro refleja la capacidad del sector para adaptarse a las tendencias del mercado internacional, maximizando tanto la calidad como el valor del producto exportado. No obstante, para el mes de diciembre del 2024 se produjo una caída importante debido, principalmente, a una disminución de la demanda en el mercado chino. Con respecto al año 2025, se observa una tendencia creciente, especialmente marcada entre los meses de febrero y abril, sugiriendo un crecimiento sostenido del sector camaronero ecuatoriano.
PARTICIPACIÓN POR MERCADOS DESTINO: ABRIL 2024 VS ABRIL 2025
El siguiente gráfico muestra la participación porcentual de las exportaciones de camarón por destino en libras durante abril del 2024 y abril del 2025. Como se puede observar, aunque China sigue siendo el principal mercado, se observa una ligera disminución en su participación de 10 puntos. Por el contrario, Europa presenta un aumento en su participación de 6.1 puntos porcentuales.
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
PRINCIPALES PAÍSES DESTINOS (MILLONES DE LIBRAS): ENERO - MARZO (2024 VS 2025)
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
La tendencia general muestra crecimiento en todos los países/regiones, destacando principalmente a Francia y España, presentando un crecimiento del 129% y 40% en el volumen exportado respectivamente. Por otro lado, se observa a Estados Unidos con un aumento del 8%, pasando de 177 a 191 millones de libras entre el período Enero - Abril del 2025 versus mismo período del 2024.
EVOLUCIÓN DE EXPORTACIONES (MILLONES DE LIBRAS):
CHINA Y EE. UU. ENERO - ABRIL 2025
El gráfico presenta la evolución mensual de las exportaciones de camarón hacia China y Estados Unidos durante los meses de enero y abril del 2025. Se observa una marcada tendencia decreciente en los envíos hacia China durante el primer trimestre con un repunte en abril, pasando de 104 millones de libras en marzo a 117 millones en abril, lo que representa un crecimiento del 13%. En contraste, el comportamiento de las exportaciones hacia Estados Unidos muestra una tendencia positiva de enero a marzo, con un decrecimiento en abril del 19% en cuanto a las libras exportadas.
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
Importación de camarón de Estados Unidos
Autores: Jim Kenny jkenny@urnerbarry.com
Ángel Rubio angel.rubio@expanamarkets.com
Urner Barry
Importaciones de todo tipo, por tipo
Según los datos comerciales de abril de 2025, las importaciones de camarones de EE.UU. ascendieron a 154,2 millones de libras, lo que supone un ligero descenso del 0,1% con respecto a marzo, pero un aumento del 38,0% en comparación con abril de 2024. Las cifras acumuladas hasta la fecha muestran un fuerte impulso, con 606,5 millones de libras importadas, lo que supone un aumento del 18,1% con respecto al mismo período de 2024. El desglose por categorías de productos pone de relieve la fortaleza continuada en todos los segmentos, con un aumento de las importaciones con cáscara del 4,7% en lo que va del año, mientras que las variedades peladas y cocidas registraron un crecimiento excepcional del 23,5% y el 33,8%, respectivamente.
La dinámica del mercado se ha vuelto cada vez más compleja a medida que los participantes del sector se preparan para posibles modificaciones arancelarias tras la fecha límite del 9 de julio. La proximidad de esta fecha crítica ha creado un mercado bifurcado en el que el sólido ciclo de cosecha de la India y la proximidad logística de Ecuador proporcionan un posicionamiento competitivo en medio de una incertidumbre generalizada. Muchos importadores han adoptado la postura cautelosa de esperar y ver qué pasa, sin saber si la actual estructura arancelaria uniforme del 10% volverá a las tasas diferenciadas por país (India 26%, Ecuador 10%, Vietnam 46%, Indonesia 34%). A pesar de esta indecisión, el mercado estadounidense parece tener un suministro abundante, ya que la aceleración de las compras a Ecuador compensa cualquier preocupación sobre el suministro, mientras que los compradores reevalúan sus estrategias de abastecimiento a largo plazo para India, Vietnam e Indonesia.
Los patrones de suministro de los principales países de origen revelan notables disparidades en el rendimiento. India entregó 58,7 millones de libras en abril, lo que supone un considerable avance del 25,7% con respecto a abril de 2024, mientras que los envíos de Ecuador aumentaron un 101,9%, hasta alcanzar los 44,0 millones de libras. Indonesia registró un impresionante
aumento interanual del 34,0%, hasta alcanzar los 30,7 millones de libras, y Vietnam registró 8,5 millones de libras, con una modesta contracción interanual del 4,2%. Tailandia contribuyó con un notable impulso de crecimiento, avanzando un 13,3% con respecto al año anterior.
Los costos de sustitución de las importaciones alcanzaron una media de 3,84 dólares por libra en abril, lo que ilustra la complejidad del entorno del mercado, en el que algunos países de origen mantienen la flexibilidad de precios, mientras que otros ejercen disciplina en medio del aumento de los gastos de producción y la incertidumbre normativa.
Ciclos mensuales de importación por país (todos los tipos)
•India: Los envíos de abril desde India ascendieron a un total de 58,7 millones de libras, lo que supone un sólido aumento del 25,7% con respecto a abril de 2024 y un avance del 6,7% en relación a los volúmenes de marzo. Las importaciones acumuladas han alcanzado los 236,7 millones de libras en lo que va del año, lo que representa una fuerte expansión del 21,6% con respecto a 2024; esto subraya la importancia estratégica de India a pesar de las inminentes preocupaciones arancelarias. Fuentes del sector indican una intensificación de la actividad de adquisición para aprovechar los períodos de máxima producción antes de la fecha límite arancelaria del 9 de julio.
•Ecuador: Los volúmenes de abril registraron 44,0 millones de libras, lo que supone un excepcional crecimiento del 101,9% con respecto a abril de 2024, aunque supone un descenso del 10,5% con respecto a los niveles de marzo. Los envíos acumulados en lo que va del año ascienden a 166,1 millones de libras, lo que refleja una saludable mejora del 13,8% con respecto al mismo período de 2024. La ventaja geográfica de Ecuador en cuanto a la duración del tránsito ha atraído una mayor atención de los compradores, ya que las empresas buscan reducir su exposición a los aranceles, y los sólidos resultados de abril podrían indicar este cambio estratégico en los patrones de abastecimiento.
•Indonesia: Las entregas de abril alcanzaron
Importaciones YTD de todos los tipos de camarón por año de EE.UU. y promedio Importación $/lb.
Fuente: USDOC. Urner Barry
los 30,7 millones de libras, lo que supone un notable crecimiento interanual del 34,0% y un aumento del 10,8% con respecto a marzo. Los volúmenes acumulados se sitúan en 106,9 millones de libras, superando los niveles de 2024 en un 17,4%. Ante la posibilidad de que se apliquen aranceles del 34% a partir del 9 de julio, los equipos de compras informan de patrones de compra más selectivos, mientras que el impulso de abril sugiere una acumulación acelerada de existencias para eludir los posibles efectos de la fecha límite.
•Vietnam: Los envíos vietnamitas ascendieron a 8,5 millones de libras en abril, lo que supone un descenso interanual del 4,2%, pero un aumento del 8,6% con respecto a marzo. Las importaciones acumuladas en lo que va del año, de 35,2 millones de libras, se mantienen un 6,1% por encima de las de 2024. Las categorías especializadas mantienen características de rendimiento diferenciadas, y los productos cocinados siguen registrando un sólido avance en lo que va del año. Dados los posibles aranceles del 46%, los más elevados entre los principales proveedores, los participantes en el mercado informan de una mayor planificación estratégica y de plazos de compra más ajustados.
Importaciones de camarones con cáscara,
cíclicas y por tamaño
Las importaciones de camarones con cáscara en abril de 2025 ascendieron a 40,6 millones de libras, lo que supone un aumento sobresaliente del 54,4% con respecto a abril de 2024 y contribuye a un incremento del 4,7% en lo que va del año, hasta alcanzar los 163,9 millones de libras.
El resurgimiento de esta categoría coincide con una aceleración más generalizada del mercado, lo que indica una adquisición estratégica antes de los cambios normativos, a pesar de la abundante oferta del mercado. El análisis del tamaño revela ganancias generalizadas, con un aumento interanual del 52,4% en la categoría U/15 en abril, mientras que el tamaño 16-20 subió un 58,7%. El segmento 21-25 mostró un impulso excepcional con una expansión interanual del 93,6% en abril. Los tamaños medianos de 26-30 registraron un sólido avance del 43,6%. Esta fortaleza generalizada en todas las especificaciones de tamaño refleja una intensificación de la actividad de compra, ya que los participantes posicionan sus existencias antes de la posible aplicación de aranceles, aunque los niveles de oferta actuales parecen más que adecuados.
Ecuador mantuvo su posición dominante como principal proveedor de camarón con cáscara, mientras que Indonesia e India realizaron importantes aportaciones. La combinación de orígenes sigue evolucionando
a medida que los compradores evalúan la exposición a los aranceles, y la eficiencia de los envíos de Ecuador ofrece ventajas tácticas para las necesidades inmediatas.
Precios del camarón con cáscara + Índice HSLO
Los costos de sustitución del camarón con cáscara en abril alcanzaron una media de 3,58 dólares/lb, manteniendo unos precios moderados a pesar de las compras selectivas motivadas por los aranceles. Las variaciones de precios entre los distintos tamaños reflejan tanto las restricciones de suministro como el comportamiento estratégico de compra, con los tamaños más grandes U/15 alcanzando valoraciones superiores, mientras que los tamaños medianos muestran una relativa firmeza.
Persiste la diferencia entre los costos de sustitución de las importaciones y las cotizaciones mayoristas nacionales, con el índice Urner Barry White Shrimp alcanzando los 4,24 dólares/lb hasta junio de 2025, por encima de las lecturas de abril. Esta diferencia de precios abarca factores estacionales y la acumulación estratégica de existencias antes de posibles ajustes arancelarios, incluso aunque la disponibilidad general de la oferta parezca sólida.
Importaciones
de camarones pelados con valor agregado
El producto pelado mantuvo un impulso de crecimiento excepcional en abril, con volúmenes que alcanzaron los 78,7 millones de libras, un aumento sustancial del 38,0% con respecto a abril de 2024. Los totales acumulados en lo que va del año han ascendido a 304,0 millones de libras, lo que supone un destacado aumento del 23,5% con respecto a 2024. Los costes de importación promediaron 3,84 dólares/lb en abril, lo que demuestra una relativa estabilidad de los precios con respecto a marzo.
Los compradores siguen navegando por las diversas condiciones del mercado en todas las categorías, mostrando enfoques más específicos que hacen hincapié en tamaños y parámetros de calidad concretos. India lideró este segmento con 42,3 millones de libras en abril, lo que supone un crecimiento del 22,9% con respecto a abril de 2024, al tiempo que mantiene una fuerte expansión del 17,1% en lo que va del año. Ecuador mostró un rendimiento especialmente sólido, con envíos en abril de 19,0 millones de libras, lo que supone un aumento del 126,9% con respecto a abril de 2024 y un crecimiento del 40,5% en lo que va del año. Indonesia mantuvo un fuerte impulso con un volumen de 10,5 millones de libras en abril, lo que supone un aumento del 29,8% con respecto a abril de 2024. Vietnam entregó 2,2 millones de libras, experimentando un notable descenso interanual del 24,2% en
Detalle de importaciones de camarón YTD por tipo
abril, mientras que registró una moderada disminución del 19,5% en lo que va del año.
Sustitución de producto pelado frente a venta al por mayor en EE.UU.
Los costos de sustitución de las importaciones peladas en abril alcanzaron una media de 3,84 dólares/lb, manteniendo las diferencias de precios específicas de cada país. El producto indio mantuvo su posición competitiva, mientras que el material vietnamita alcanzó primas que reflejaban sus atributos de calidad y sus ventajas de procesamiento. El índice de valor añadido de Urner Barry avanzó hasta los 5,16 dólares/ lb en junio, lo que indica un fortalecimiento de los fundamentos del mercado junto con un posicionamiento estratégico a medida que se acerca la fecha límite del 9 de julio.
Este impulso de los precios abarca múltiples elementos más allá de la mecánica tradicional de la oferta y la demanda, ya que la incertidumbre arancelaria genera urgencia en las compras a pesar de la buena situación del mercado en cuanto a suministro. Los plazos de entrega de origen asiático de entre 6 y 9 semanas (a veces más) han creado una urgencia específica para las compras de India, Vietnam e Indonesia, ya que el material enviado después de principios de mayo podría enfrentarse a una elevada exposición arancelaria si no se resuelven las negociaciones comerciales en curso.
Importaciones de camarones cocidos, empanados y otros
Las importaciones de camarones cocidos en agua caliente alcanzaron los 18,9 millones de libras en abril de 2025, lo que supone un impresionante crecimiento del 53,7% con respecto a abril de 2024. Los resultados acumulados del año revelan un avance sustancial del 33,8% en comparación con 2024, con un total de 79,8 millones de libras. India sigue liderando esta categoría con 10,2 millones de libras en abril, lo que supone un importante crecimiento del 42,1% con respecto a abril de 2024 y mantiene una sólida expansión del 14,4% en lo que va de año. Indonesia alcanzó los 5,0 millones de libras, con un crecimiento interanual del 33,9%, mientras que Vietnam contribuyó con 2,7 millones de libras, lo que supone un avance interanual del 7,3%.
Los productos empanizados alcanzaron un total de 11,4 millones de libras en abril de 2025, lo que supone un crecimiento del 15,5% con respecto a abril de 2024. El volumen acumulado alcanzó los 44,4 millones de libras, superando en un 11,9% el dato de 2024. Indonesia mantuvo el liderazgo con 5,0 millones de libras en abril, registrando un crecimiento del 10,9% con respecto a abril de 2024 y contribuyendo a un avance del 8,0% en lo que va de año. Este segmento sigue demostrando consistencia en medio de la volatilidad general del mercado, y Indonesia mantiene su posición dominante a pesar de la incertidumbre arancelaria.
Fuente: USDOC. Urner Barry
Venta al por menor (retail)
La actividad promocional minorista en mayo de 2025 registró 47.900 oportunidades de compra (anuncios x número de tiendas), lo que supone un descenso con respecto al nivel de 52.000 de abril, en concordancia con los patrones estacionales habituales. Esta moderación sigue las tendencias establecidas tras la Semana Santa, al tiempo que se mantiene una infraestructura promocional estable. El precio medio anunciado en mayo se situó en 7,91 dólares/lb, lo que supone un aumento de 0,30 dólares/lb con respecto a marzo, y también muestra una continuidad con los niveles promocionales recientes. Las cifras acumuladas hasta mayo indican que el volumen promocional alcanzó los 265.000 anuncios, manteniendo un rendimiento estable en relación con períodos comparables, aunque por debajo de los niveles de referencia previos a la pandemia, que se situaron entre 242.000 y 248.000 en 2020-2022.
El precio promocional medio en lo que va de año muestra una notable estabilidad, lo que refleja la consistencia de los precios a pesar de las incertidumbres en el suministro y la próxima aplicación de aranceles. Esta estabilidad promocional y de precios sugiere que los minoristas mantienen estrategias de comercialización de camarones coherentes mientras navegan por el complejo entorno de suministro con la inminente fecha límite de julio.
Camarones del Golfo
La producción nacional en febrero de 2025 ascendió a 1,33 millones de libras, lo que supone un descenso del 11,6% con respecto a los 1,51 millones de libras de febrero de 2024. Aunque aún no se dispone de las estadísticas actualizadas de desembarques de marzo a mayo, los precios se han fortalecido considerablemente a lo largo del segundo trimestre en todas las categorías, y las variedades marrones nacionales han mostrado una firmeza excepcional en medio de una disponibilidad muy limitada. Los valores de mercado han avanzado sustancialmente, y el precio de las variedades marrones nacionales de 21-25 unidades ha superado el umbral de 6,50 dólares/lb en transacciones recientes. Dada la limitada disponibilidad de reposición y las entregas de buques por debajo de los
objetivos, los proveedores han implementado enfoques estratégicos de gestión de inventarios. Los informes del sector indican que las existencias son escasas a finales de la primavera, lo que respalda las expectativas de que los precios se mantengan firmes hasta mediados de 2025, a la espera de nuevas estadísticas de producción. Las actuales limitaciones de suministro han creado un entorno favorable para el producto nacional en medio de las difíciles condiciones del mercado de importación y los próximos cambios normativos.
Exportaciones de Ecuador
Las exportaciones de camarones ecuatorianos en abril de 2025 ascendieron a aproximadamente 244,0 millones de libras, manteniendo un sólido rendimiento exportador. Los volúmenes acumulados alcanzaron los 963,2 millones de libras, lo que representa un crecimiento aproximado del 20% con respecto a 2024. Los patrones de distribución mostraron una evolución continua, con destinos asiáticos que recibieron el 52,0% de las exportaciones de abril, los mercados europeos que representaron el 25,5% y los envíos a
Estados Unidos que representaron el 18,8%.
Esta distribución geográfica refleja las continuas adaptaciones a los patrones de demanda mundial y las consideraciones arancelarias de Estados Unidos. Los envíos acumulados con destino a Estados Unidos ascendieron a 190,8 millones de libras, aumentando a medida que los importadores estadounidenses aprovechan las ventajas de tránsito de Ecuador para minimizar la exposición arancelaria a medida que se acerca la fecha límite del 9 de julio.
Las configuraciones actuales de las exportaciones sugieren que los productores ecuatorianos están diversificando estratégicamente los mercados de destino en medio de la incertidumbre comercial mundial, posicionándose para lograr flexibilidad operativa independientemente de los resultados de las negociaciones arancelarias de EE.UU. Con la fecha límite ahora inminente y el mercado americano aparentemente bien abastecido, esta estrategia de diversificación proporciona una importante mitigación del riesgo•
CNA y Gobierno alinean esfuerzos para potenciar exportaciones de camarón en mercados estratégicos
ESTADOS UNIDOS: En el marco de las negociaciones para la restitución del arancel cero con Estados Unidos, el ministro de Producción, Comercio Exterior Inversiones y Pesca (MPECIP), Luis Jaramillo, planteó la creación de un Cuarto Adjunto, organización en la que el sector exportador, a través de la CNA, puede aportar técnicamente al proceso negociador y asegurar una representación directa en la defensa de sus intereses comerciales del sector camaronero. El 29 de mayo de 2025, el Tribunal de Comercio Internacional de EE.UU. declaró inconstitucionales la mayoría de los aranceles globales aplicados por Donald Trump, al considerar que excedían sus facultades ejecutivas. La medida prohíbe su aplicación permanente y establece un plazo de 10 días para su eliminación administrativa. La administración actual anunció que apelará la decisión.
UNIÓN EUROPEA:
La viceministra de Acuacultura y Pesca, Ivanova Cereceda, se reunió con los equipos ejecutivo y técnico de la Cámara Nacional de Acuacultura. En el encuentro se analizó el flujo comercial entre mercados europeos como Francia, que han presentado observaciones al camarón ecuatoriano. En este contexto, el Gobierno inició gestiones diplomáticas y técnicas para aclarar las inconsistencias detectadas en los procedimientos de inspección europeos. CNA, por su parte, gestionó reuniones bilaterales entre autoridades sanitarias de Ecuador, Francia y España durante el evento de la Organización Mundial de Sanidad Animal (OMSA) el diálogo permitió al país explicar sus controles y coordinar medidas correctivas que garanticen la continuidad del comercio y el cumplimiento de los requisitos sanitarios europeos.
CHINA: La CNA organizó un encuentro estratégico entre representantes del Banco de China y actores clave del sector acuícola ecuatoriano, con el objetivo de identificar oportunidades de cooperación tecnológica, financiamiento e inversión.
En este contexto, funcionarios del Bank of China presentaron líneas de crédito y financiamiento destinadas a impulsar la modernización tecnológica del sector camaronero. El objetivo es mejorar la eficiencia operativa, reducir costos y fortalecer la sostenibilidad y trazabilidad, elevando la competitividad del sector. El Banco de China tiene presencia operativa en 58 países y regiones, además de oficinas de representación en seis naciones. Su red internacional abarca 28 regiones del Asia-Pacífico (incluido Medio Oriente), 19 países europeos, 9 en América y 8 en África.
Sector camaronero ecuatoriano presente en Asamblea Mundial de la OMSA
CNA integra la red de formadores que capacitarán a aspirantes diplomáticos en materia de comercio exterior
La directora ejecutiva de la Cámara Nacional de Acuacultura (CNA), Yahira Piedrahita, integró la misión ecuatoriana que participó en la 92° Asamblea Mundial de la Organización Mundial de Sanidad Animal (OMSA), en particular, en la sesión de la Comisión de Animales Acuáticos. La delegación nacional sostuvo reunionestécnicas con representantes de Francia y España para abordar algunas observaciones relacionadas con las exportaciones a dichos mercados.
Durante el encuentro, Ecuador impulsó la creación de una comisión ad hoc para revisar la normativa del Código Acuático y Manual Acuático en lo referente al IHHNV, con el objetivo final de excluirlo de la lista de enfermedades de notificación obligatoria. La misión brindó soporte técnico al punto focal de Agrocalidad y fortaleció alianzas regionales en una agenda que incluyó a autoridades de alto nivel como la Dra. Alicia Gallardo, presidenta de la Comisión Acuática de la OMSA, y la presidenta del organismo, Dra. Marina Pombo.
SSP y CNA capacitan sobre innovación nutricional
Con el objetivo de fortalecer la eficiencia y sostenibilidad del sector, Sustainable Shrimp Partnership (SSP) y la CNA realizaron la quinta edición del programa educativo SustainED, enfocada en la innovación nutricional como herramienta clave para optimizar el cultivo de camarón.
El encuentro, titulado "Innovación nutricional: cómo aprovechar al máximo el alimento en cada etapa del cultivo", tuvo lugar el jueves 22 de mayo en el Hotel Hilton Colón de Guayaquil y reunió a cerca de 150 expertos, académicos y representantes del sector productivo,
La CNA, a través de la Corporación de Gremios Exportadores del Ecuador (CORDEX) y la Academia Diplomática, desarrollarán un plan conjunto para capacitar a 25 aspirantes a diplomáticos en estrategias de negociación, diplomacia comercial e inserción en mercados internacionales.
El programa incluirá formación técnica y visitas a sectores productivos para fortalecer su conocimiento del comercio exterior ecuatoriano.
consolidándose como un espacio técnico para elevar los estándares de producción responsable.
El siguiente SustainED está previsto para el 14 de agosto, en el que se abordará las nuevas tecnologías en el sector camaronero.