Edición 151 - Febrero 2023
2023, Un año desafiante para el sector camaronero ecuatoriano
SSP cumple 5 años liderando la innovación y construyendo un futuro sostenible para la industria acuícola mundial
Aqua Expo Santa Elena 2023 reunió a expertos internacionales y ecuatorianos especializados en larvicultura
Más de 20 heridos y más de medio millón de dólares en pérdidas suma el sector camaronero en lo que va del año
Programas de mejoramiento genético han sido cruciales para potenciar la especie Penaeus Vannamei en los laboratorios de larvas
Identificación de un nuevo solinvivirus con localización nuclear asociado a mortalidades masivas en el cultivo del camarón blanco (Penaeus vannamei)
Incremento en el desempeño del crecimiento y mecanismos de digestión del camarón a través de un potenciador de la digestión con base en lisofosfolípídos bajo diferentes niveles de colesterol en la dieta
Incremento de la supervivencia de camarón blanco (Litopenaeus vannamei) infectado con el virus de la mancha blanca y alimentado con una dieta suplementada con aceite de coco (Cocos nucifera)
Rápida determinación de materia orgánica y carbono orgánico en suelos de camaronera mediante espectroscopia visible e infrarrojo cercano VIS-NIR
Ecosistemas de carbono azul: una estrategia para mitigar el cambio climático global
Presidente Ejecutivo
Ing. José Antonio Camposano
Editora “AquaCultura”

Msc. Shirley Suasnavas ssuasnavas@cna-ecuador.com
Consejo Editorial
MSc. Yahira Piedrahita
PhD. Leonardo Maridueña
Ing. José Antonio Lince Econ. Danny Vélez Ing. Alex de Wind
Exportaciones de camarón y tilapia
Reporte de mercado de China
Reporte de mercado de EE. UU.
Noticias de interés
Noticias empresariales
Diseño y diagramación Ing. Orly Saltos osaltos@cna-ecuador.com
Ing. Roberto Peñafiel rpenafiel@cna-ecuador.com
Corrección de estilo
Daniel Ampuero daniel.ampuero@gmail.com
Comercialización
Gabriela Nivelo gnivelo@cna-ecuador.com
ÍNDICE
INFORMACIÓN DE COYUNTURA ARTÍCULOS TÉCNICOS ESTADÍSTICAS NOTICIAS 8 12 29 16 21 24 38 42 52 57 63 68 72 76 78
Durante el 2022, el sector experimentó una complicada contracción de la demanda, comparable únicamente con la crisis del año 2008 y con el derrumbe de los mercados del primer año de la pandemia en 2020. Conforme se desarrollaba el segundo trimestre del 2022, las suspensiones en China a un considerable número de empacadoras ecuatorianas motivó de inmediato el colapso de la cadena de exportación y el pánico en el mercado, produciendo un derrumbe de precios internacionales. Esta situación trajo como consecuencia la caída del precio a los productores quienes, en su gran mayoría, no podían cubrir siquiera sus costos de producción.
Con el pasar de los meses, en la medida que las suspensiones se empezaron a levantar y la demanda a activarse, el precio empezó un lento proceso de recuperación. A pesar de esta situación, la afectación producida por la devolución de centenas de contenedores desde China era un hecho, y las pérdidas sobrepasaban las decenas de millones de dólares.
Mientras esto sucedía en el mercado más importante para el camarón ecuatoriano, el mercado europeo vio como su moneda, el euro, se desplomaba frente al dólar estadounidense. Automáticamente, nuestro camarón se encareció 20% por efecto de la devaluación de la moneda europea. De igual forma, la ola inflacionaria que golpeó a los Estados Unidos frenó el crecimiento del consumo de camarón en dicho país; es decir, ya no podíamos contar con un mercado para nuestros productos de valor agregado o, por lo menos, ya no a los precios que veníamos recibiendo.
Mientras los mercados internacionales mostraban este sombrío panorama, en el Ecuador los costos de producción se incrementaron de forma permanente. Insumos, alimento balanceado, cartón, plástico y otros materiales experimentaron un acelerado incremento en sus precios, complicando aún más la situación de miles de productores y quitándonos capacidad de competir en los mercados, pues nuestra competencia se beneficiaba de la debilidad de sus monedas para lograr precios más favorables.
Finalmente, al acercarnos al último cuatrimestre del año, los agresivos brotes de COVID-19 en China motivaron masivas
José Antonio Camposano Presidente Ejecutivo

Hay que mirar más allá de las ventas
medidas de confinamiento, lo que asfixió el consumo derribando la demanda en el momento clave del año para venta de camarón a ese mercado. Una vez más, los precios cayeron a niveles inferiores, inclusive, a los de los meses más duros de la pandemia.
La situación anteriormente descrita parece haber pasado desapercibida por las autoridades del Ministerio de Finanzas y su equipo de analistas, pues desde el prisma de la burocracia, se señaló que el sector pasaba por un buen momento, lo que permitía tomar la decisión de eliminar el precio diferenciado del diésel, atestando un golpe más a la actividad productiva en uno de sus peores momentos.
Hoy, los reportes y comunicados del Banco Central, así como los del Ministerio de Comercio Exterior se limitan a resaltar el incremento de las exportaciones de camarón en 2022. No hay una sola mención respecto a que hoy es casi 25% más caro producir una libra de camarón que hace año y medio. No se indica que el precio promedio de la libra exportada en 2022 está fuertemente influenciado porque ahora se pelan casi 4 veces más camarones que en 2020 y, por lo tanto, ese producto se vende a un precio diferente, pero a un costo mucho más alto dada la poca competitividad que tenemos para los procesos de valor agregado en el país.
El escenario experimentado en 2022 nos obliga interpretar con mucha cautela al panorama para este 2023, e insistir con mucha más fuerza en que en el país se ejecuten de inmediato acciones que mejoren la competitividad del sector en lugar de buscar incrementar sus costos. Las posibilidades de continuar mejorando la productividad en la magnitud registrada en la última década no existen, por lo que no se puede seguir apostando, desde lo público, que el sector privado podrá defenderse solo en los mercados internacionales. Es más urgente que nunca que en el Ecuador se tomen decisiones que apunten a bajar costos internos como el de la inseguridad que hoy los privados asumimos, así como otros asociados a la ineficiencia del aparato estatal. Seguiremos exponiendo esta realidad para contrarrestar el “mundo empresarial perfecto” que se vende desde ciertas esferas públicas, y que no es más que una ilusión•
EDITORIAL
PRIMER VICEPRESIDENTE
Ing. Luis Francisco Burgos
DIRECTORIO
PRESIDENTE DEL DIRECTORIO
Ing. Marcelo Vélez
SEGUNDO VICEPRESIDENTE

Ing. Fabricio Vargas
Ing. Ricardo Solá
Dr. Alejandro Aguayo
Ing. Alex Olsen
Ing. Ori Nadan
Ing. Francisco Pons
Ing. José Antonio Lince
Ing. Jorge Redrovan
Ing. Alex de Wind
Ing. Kléber Siguenza
Ing. Rodrigo Vélez
Ing. Iván Rodríguez
Ing. Juan Carlos Vanoni
Ing. Roberto Aguirre
VOCALES
Ing. Alejandro Ruiz-Cámara
Ing. Alvaro Pino
Econ. Heinz Grunauer
Ing. Víctor Ramos
Ing. David Eguiguren
Ing. Humberto Dieguez
Ing. Atilio Solano
Ing. Freddy Arias
Ing. Miguel Uscocovich
Ing. Vinicio Aray Dueñas
Econ. Sandro Coglitore
Ing. Rodrigo Laniado
Blgo. Carlos Sánchez
Ing. Diego Puente
Ing. Johnny Adum
Sra. Verónica Dueñas
Ing. Alex Elghoul
Ing. Bastien Hurtado
Econ. Wolfgang Harten
Econ. Carlos Miranda
Econ. Danny Vélez
Ing. Héctor Marriott
Ing. Oswin Crespo
Ing. Edison Brito
Ing. Santiago León

COYUNTURA
2023, UN AÑO DESAFIANTE PARA EL SECTOR CAMARONERO ECUATORIANO
Después de un 2022 desalentador, el año en curso representa un nuevo desafío a la actividad camaronera.
El cierre del año 2022 dejó como resultado una imagen distorsionada de las complicaciones que viene enfrentando la actividad camaronera y que en algunas ocasiones aparecen en los titulares de prensa. Si bien, las cifras de exportación dan cuenta de un sector que se ha expandido hasta transformarse en el principal generador de divisas para la economía, muy lejos de sus seguidores, los sectores bananero y minero, lo cierto es que, al interior de la cadena de valor del camarón se experimenta un constante proceso de deterioro de su competitividad, que se ha visto contrarrestado exclusivamente por el esfuerzo del sector privado.
No se trata de ocultar las cifras, pues es de conocimiento público que, en la última década, el promedio de crecimiento anual del sector ha sido del 18%, pero lo que no debe confundirse es que esos incrementos en ventas equivalen a incrementos en la rentabilidad de la actividad camaronera, pues lo registrado en los últimos años más bien apunta hacia una sistemática pérdida de competitividad ante la falta de políticas de fomento a la actividad productiva y a la exportación.
Estadísticas no revelan la realidad del sector
Es necesario recordar que los resultados que aparecen en los reportes estadísticos no rescatan la realidad de la actividad, que ha tenido que asumir altos costos y hacer grandes esfuerzos para lograr incrementar su productividad y eficiencia en predios camaroneros, en los que se han implementado nuevas tecnologías como: aireadores, alimentadores automáticos, bombeo eléctrico y sistemas informáticos de control para toma de decisiones.
Por su parte, los establecimientos exportadores ecuatorianos han adaptado sus procesos para diversificar sus productos y satisfacer al consumidor mundial, ya sea con la exportación de camarón entero, preferido en China, o con valor agregado: colas, pelado y desvenado, como lo prefieren en el mercado europeo y en EE.UU. Esta diversificación supone considerables inversiones en equipamiento y contratación de personal, lo que implica desembolsos y agregar costos laborales a la cadena de valor.
En síntesis, el sector ha hecho importantes inversiones y ha visto su estructura de costo abultarse, mientras el precio internacional del camarón y la pérdida de competitividad, agravada por factores exógenos como la inseguridad, los empuja constantemente por debajo de su punto de equilibrio e inclusive a preocupantes pérdidas durante algunos meses del año, incluso siendo este el caso de productores con eficiencias promedio, por lo que la situación de unidades productivas ineficientes es dramática.

“El año 2022 ha sido de los más complicados que hemos tenido que afrontar: mercados deprimidos, dólar fortalecido, costos de producción al alza, costos por ineficientes servicios públicos, inseguridad a niveles nunca antes vistos. Las ventas al exterior no muestran esa realidad” .
José Antonio Camposano Presidente Ejecutivo de la Cámara Nacional de Acuacultura
“Enero arrancó con una leve recuperación de los precios internacionales; sin embargo aún estamos a la expectativa del ‘ánimo’ que muestren los mercados de destino, lo que podría derivar a una nueva caída de precios. Las evidencias que apuntan a una recesión en nuestros mercados aún son evidentes por lo que, al igual que 2022, proyectamos un año 2023 cuesta arriba lo que nos obliga aún más a insistir enérgicamente en que se ejecuten las acciones que alivien la estructura de costos del sector”, precisó José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura, apuntaba a fines del primer mes del año.

COYUNTURA 8
8
Elevación de costos de producción
Actualmente, producir camarón ecuatoriano es 24% más caro en comparación con los costos de hace dos años atrás. Los tres principales factores que han incidido en el aumento del costo de producción se centran en la eliminación el precio diferenciado del diésel, que encareció el rubro en 16 centavos por libra a finales del año pasado, afectando al 82% de la superficie camaronera.

A esto se suma que el costo ya se encontraba visiblemente afectado por el incremento de precio en las materias primas a escala mundial, ya que en comparación con el 2019 el trigo subió el 71%, la pasta de soya el 45%, el aceite de pescado 105% y la harina de pescado 24%; lo que aumentó significativamente el valor de producción del alimento balanceado, uno de los principales rubros para el productor camaronero, que representaba el 55,2% del costo total de producción.
Asimismo, mediante una decisión antitécnica, se incrementó el salario básico una vez más, lo que agregó un centavo de costo adicional por cada libra producida en el país.
Finalmente, el camaronero asume un costo de seguridad que suma más de $80 millones anuales por la implementación de sistemas de video vigilancia, rastreo y comunicación, además del pago de guardias privados en turnos rotativos y las pérdidas por robos que sufren a diario. Se trata de un monto fijo destinado para hacer frente a la delincuencia que golpea sin tregua al sector por vía terrestre y fluvial, dejando al momento, un muerto y 20 personas heridas, según estadísticas de la Dirección de Seguridad de la Cámara Nacional de Acuacultura, hasta febrero 2023.

ECOS DE PRENSA
9
Desventajas ante la competencia
Si bien muchos de estos escenarios afectan a otros países competidores, un elemento que agrava la situación es el hecho que el Ecuador tiene como moneda de uso nacional al dólar de los Estados Unidos de Norteamérica, lo que no le permite, mediante devaluación, bajar costos de producción abaratando sus exportaciones. Por el contrario, el dólar se fortaleció frente a otras monedas durante todo el 2022, lo que hizo más caro producir camarón en Ecuador y, de forma automática, Ecuador se volvió menos atractivo frente a las ofertas de la competencia en los mercados de destino.
Por su parte, India y Vietnam, competidores directos de Ecuador, cuentan con varias ventajas frente a nuestro país. Salarios más bajos, subsidios a insumos y moneda propia. Ambos países registraron depreciaciones de sus respectivas monedas, lo que les da una “ventaja temporal” frente a países como el Ecuador, que están dolarizados y que ven encarecer su oferta frente a la de estos dos competidores.
Ante esta situación, el sector considera que la aplicación del drawback automático aliviaría parcialmente los problemas antes mencionados, a los que se suma la depreciación de las monedas de países competidores o de los mercados de destino como el euro que llegó a la paridad con el dólar en 2022, lo que automáticamente encareció nuestras exportaciones. Este fenómeno se repitió con China, el principal destino de las exportaciones de camarón en 2022.
José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura, aclara que las cifras de cierre del camarón ecuatoriano en el 2022, no reflejan esa dura realidad que ha enfrentado la industria el año pasado. “El año 2022 ha sido de los más complicados que hemos tenido que afrontar: mercados deprimidos, dólar fortalecido, costos de producción al alza, costos por ineficientes servicios públicos, inseguridad a niveles nunca antes vistos. Las ventas al exterior no muestran esa realidad”.
Actualmente, el camarón ecuatoriano es el primer producto de exportación no petrolero
del Ecuador y es un pilar fundamental para la economía del país, generando cerca de 290 mil fuentes de empleo directo e indirecto. Según el boletín Análisis Semanal de Walter Spurrier y Alberto Acosta Burneo: “Por cada 1% que decline la producción camaronera se destruirían 791 empleos; además se reduciría el valor agregado en $19.7 millones y en pago de impuesto a la renta en $1.5 millones”.
La economía de Ecuador sigue siendo vulnerable frente a las conmociones externas (WTO, 2012) por lo que debe enfocarse en fomentar y fortalecer las exportaciones, lo que le permitirá continuar con la generación de divisas de manera continua y mantener una estabilidad macroeconómica.
Los costos relacionados a la producción, procesamiento y comercialización en toda la industria camaronera se han incrementado, mientras que el precio internacional se reduce drásticamente debido a una demanda débil y una competencia agresiva por captar nuevos mercados.

Si no se adoptan medidas compensatorias y de fomento a la competitividad de las exportaciones, según Camposano, se perderá cuota de mercado, empezando por aquellos destinos de precios muy competitivos como los EE. UU. y luego en otros como Europa, cuyas condiciones no son favorables para el consumo de ciertos bienes que se adquieren fuera del hogar en segmentos como la restauración.
Proyecciones
“Estados Unidos es un mercado para el que estamos trabajando con mucho esfuerzo, mediante nuevas inversiones y contratación de personal para ampliar la oferta de valor agregado. A pesar de ello mantenemos
el reto de continuar atendiendo a nuestro principal destino que es China y no descuidar el sitial ganado en Europa tras varias décadas de ser el principal proveedor de camarón con una calidad muy superior a la de nuestra competencia. Finalmente, tenemos la expectativa de que las condiciones sanitarias que exige Corea del Sur sean revisadas para retomar el espacio que hemos perdido en ese mercado. Estamos trabajando con la autoridad sanitaria ecuatoriana, la Secretaría de Calidad e Inocuidad, para alcanzar este objetivo”, advierte Camposano.
En enero de este año, la exportación de camarón alcanzó 209 millones de libras; sin embargo, los factores antes mencionados se mantienen y la falta de políticas de fomento por parte del gobierno ecuatoriano para la industria persiste, por lo que el panorama de la actividad se vuelve aún más incierto. Es evidente que los niveles de eficiencia de muchas unidades productivas han llegado a un tope con la tecnología actual, por lo que no sería extraño un desempeño mucho más conservador que el registrado en años anteriores.
Ante esta situación, desde el sector privado la CNA continuará trabajando con cada eslabón de la cadena de valor de la industria para tratar de sostener los niveles de producción actuales, se continuará insistiendo en la necesidad de atención de parte de las diversas autoridades, pero en especial las del Ministerio de Producción, Comercio Exterior, Inversiones y Pesca, como ente rector de la actividad acuícola y responsable de motivar eficiencias, así como eliminar costos generados por servicios deficientes que entorpecen a la actividad productiva y exportadora•
COYUNTURA 10 10
COYUNTURA

El 12 de marzo de 2018, en el marco de la feria Seafood Expo North America de Boston (EE.UU.), se presentó oficialmente ante el mundo la iniciativa de sostenibilidad del camarón ecuatoriano: Sustainable Shrimp Partnership (SSP), con el objetivo de que nuestro país sea un ejemplo a nivel global en temas de sostenibilidad.


Este importante paso para la acuicultura mundial fue posible gracias a la iniciativa de un grupo de productores de camarón líderes en Ecuador, quienes decidieron unir esfuerzos para trabajar de manera precompetitiva y transformar el futuro de la industria global de la acuicultura del camarón nacional. A través de SSP, los productores se comprometen a cultivar camarón cumpliendo con los más altos estándares, totalmente rastreable, sin antibióticos y de manera sostenible.
SSP desarrolló uno de los protocolos más exigentes para la producción de camarón con el apoyo del Consejo Asesor de SSP, formado por World Wildlife Fund (WWF), The Sustainable Trade Initiative (IDH) y Aquaculture Stewardship Council (ASC); y de la mano del Instituto Colombiano de Normas Técnicas y Certificación - (ICONTEC). Además, se encuentra colaborando con varios organismos internacionales en proyectos que permitan identificar nuevos retos que como industria deben abordar y encontrar alternativas para mejorar aún más las prácticas de producción.


Los miembros de SSP se someten a verificaciones constantes durante cada ciclo de producción para asegurar el uso cero de antibióticos, trazabilidad total e impacto neutro en el agua, demostrando el cumplimiento de las mejores prácticas de producción. Entre los Miembros Fundadores de SSP se incluye a Grupo Almar, Lanec, Omarsa, Promarisco - Grupo Nueva Pescanova y Songa - Sociedad Nacional de Galápagos.
SSP también cuenta con Miembros Asociados, que son empresas relacionadas a la cadena de valor del camarón y que son de gran aporte para la ejecución de actividades y el cumplimiento de los objetivos planteados. Entre los miembros asociados están: BioMar, DSM, INVE Aquaculture, Skretting, Vitapro.
Su visión es impulsar el futuro sostenible de la industria al producir camarones de la más alta calidad y garantizar que sea una actividad sostenible y exitosa para el mundo. Para lograr esa visión, se desarrolló una filosofía basada en cuatro atributos clave que son el marco detrás de todas las acciones:
- FEBRERO 2023
12
cumple 5 años liderando la innovación y construyendo un futuro sostenible para la industria acuícola mundial
Responsabilidad
Asegurar que sus acciones estén impulsadas por el desempeño y los resultados; demostrando el progreso y el compromiso de mejorar continuamente su desempeño ambiental y social.
Transparencia
Brindar información accesible sobre las prácticas detrás del producto, de dónde proviene y el perfil de sostenibilidad para brindarles a los clientes las herramientas que necesitan para tomar decisiones informadas sobre lo que compran.
Inclusión
Al trabajar en colaboración con otras empresas, ONGs y partes interesadas, se incentiva la implementación de las mejores prácticas para elevar a toda la industria al siguiente nivel en prácticas sostenibles.

Liderazgo continuo
Mirando continuamente hacia el futuro e identificando dónde y cómo se puede impulsar el cambio y liderar el progreso para garantizar que el cultivo de camarón sea una práctica sostenible y exitosa para el mundo.
Pionero en tecnología de trazabilidad
Tras 5 años de existencia, entre sus principales logros está ser ejemplo de
transparencia y responsabilidad con sus clientes y consumidores, al convertir el camarón SSP en el pionero en la implementación de tecnología blockchain, que proporciona información sobre su origen y trazabilidad, luego de unirse al ecosistema de IBM Food Trust.
Detrás de cada pieza del camarón premium de SSP hay una gran historia, y esta iniciativa busca que el consumidor conozca los detalles desde su origen, con la tecnología más avanzada, transparente y segura disponible.
Escaneando el código QR que se muestra en la caja, el consumidor podrá conocer el perfil de sostenibilidad en el que se criaron sus camarones, para garantizar que es un producto seguro y saludable.
Su propósito es liderar la lucha contra el fraude alimentario, que se registra en todo nivel y en diferentes cadenas.
Para SSP, la clave está en compartir información que adquiera un valor tangible para los clientes y consumidores, con el propósito de incrementar la capacidad de compra informada.
SSP: ejemplo a escala mundial por el no uso de antibióticos
Sustainable Shrimp Partnership fue escogida
“Hace 5 años, un grupo de productores camaroneros ecuatorianos se juntaron para crear SSP con un mismo objetivo: asegurar que, en las próximas décadas, las actuales y futuras generaciones puedan disfrutar de mariscos saludables. SSP es un claro ejemplo de cómo una industria puede trabajar de manera precompetitiva, en la cual sus miembros comparten objetivos y desarrollan planes claros y ambiciosos para llevar a la industria camaronera global a un siguiente nivel en términos de responsabilidad y sostenibilidad”.
 Pamela Nath Directora Sustainable Shrimp Partnership
Pamela Nath Directora Sustainable Shrimp Partnership
como ejemplo para la elaboración de un documental por la Red Global ReAct, a través de sus representantes de la Escuela de Salud de la Universidad Johns Hopkins de EE. UU.
Este producto audiovisual se enfoca en la resistencia antimicrobiana, uno de los principales problemas para la Organización Mundial de la Salud. En tal sentido, la Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO) y ReAct promueven el uso responsable de los antibióticos mediante campañas, y esta vez con el propósito de abordar el no uso de antibióticos para la cría de proteína animal. La red escogió a Sustainable Shrimp Partnership para mostrar cómo los productores ecuatorianos pertenecientes a la iniciativa SSP proveen las condiciones adecuadas para evitar brotes de enfermedades, y por ende que el animal se enferme y que haya que medicar.
SSP ha podido abordar este tema con sus miembros e incluso poder tener el compromiso por parte de ellos, pues son auditados periódicamente y de forma rigurosa, ya que en cada ciclo de producción deben compartir la evidencia de que no están utilizando antibióticos y así cumplir uno de los criterios fundamentales de este proyecto.
“Es un orgullo para nosotros como equipo ver el nombre y la reputación que SSP y sus miembros han generado a nivel mundial. Hemos sido testigos de cómo organismos internacionales y de referencia para la industria, han reconocido el trabajo de SSP, lo cual es invaluable. Ya sea por el compromiso al cumplimiento de sus criterios, como la voluntad y disposición para asumir los retos que nos llevan hoy a ser el laboratorio de innovación en la industria camaronera. Esto nos llena de compromiso y responsabilidad para seguir trabajando día a día”.
María Fernanda Vilches M. Gerente de Procesos Sustainable Shrimp Partnership
- FEBRERO 2023 COYUNTURA 13
COYUNTURA
El grupo ReACT visitó Ecuador y conoció de cerca el sistema de producción en Ecuador, recorriendo fincas camaroneras, en las que pudieron constatar que los sistemas de producción en Ecuador son naturales, o sea, lo más cercano al hábitat natural del camarón.
Como segundo punto se encuentra el factor humano, en lo que respecta al compromiso, la implementación de las buenas prácticas para preservar las condiciones naturales y hacerlo mucho más eficiente. Se mostró todo el cuidado que se da en cada una de las etapas y lograron evidenciar todo lo que hay detrás de cada camarón.
Identificando nuevos retos que abordar
SSP es miembro del “Aquaculture Working Group on Environmental Footprint”, plataforma que la reconocida Organización IDH – The Sustainable Trade Initiative, creó con la finalidad de mejorar la medición de la huella de carbono de los productos acuícolas y reducir su impacto ambiental.
La plataforma está compuesta por diversas empresas internacionales, que trabajan en conjunto para priorizar problemas, iniciar proyectos, crear métricas y metodologías. Entre los miembros del Aquaculture Working Group on Environmental Footprint están organizaciones como: Global Salmon Initiative, Seafresh Group, Hilton Seafood, Marks & Spencers, Tesco, Wegmans, entre otros.
Implementando nuevos proyectos de mejoras
SSP se encuentra trabajando con miembros de la Corporación de Promoción de Exportaciones de Inversiones (CORPEI) en el diseño de un nuevo proyecto de mejoras que permita a más productores camaroneros ecuatorianos obtener la certificación
Aquaculture Stewardship Council (ASC). Esta certificación es una de la más exigente en temas sociales y ambientales, y permite el acceso a mercados internacionales. Dentro de las actividades que se contemplan están capacitaciones en requisitos ambientales y legales en Ecuador, buenas prácticas de acuicultura y requisitos para la certificación ASC; pre-auditorías para verificar la implementación de las recomendaciones; estudios para verificar el impacto en la biodiversidad y social, entre otras.
De esta forma, Sustainable Shrimp Parnership sigue buscando nuevas alianzas y programas que permitan impulsar el desarrollo de la acuicultura mundial a través de un producto sano, seguro, con sabor inigualable, de alta calidad y totalmente trazable•


- FEBRERO 2023

SANTA ELENA 2023
El evento de mayor trayectoria del sector camaronero AQUA EXPO, realizó su primer encuentro técnico-científico del año especializado en larvicultura, en el Salinas Golf & Tenis Club de la provincia de Santa Elena el 8 y 9 de febrero pasado. Constó de dos partes: congreso técnico y feria comercial.

En el acto inaugural, la mesa directiva estuvo conformada por Andrés Arens, Viceministro de Producción Comercio Exterior Inversiones y Pesca; Yahira Piedrahita, Directora Ejecutiva de la Cámara Nacional de Acuacultura (CNA); Gabriela Nivelo, Directora de Aqua Expo, y Alex Elghoul, Director de la CNA, capítulo Santa Elena.
REUNIÓ A EXPERTOS INTERNACIONALES Y ECUATORIANOS ESPECIALIZADOS EN LARVICULTURA
“Estamos en la cuna del camarón de cultivo: Santa Elena, porque es aquí donde se encuentra la mayor parte de los laboratorios de larvas de camarón, siendo este un importante semillero para el país. Consecuentemente, la Cámara Nacional de Acuacultura, desde el 2010 ha realizado eventos en esta provincia, con el objetivo de capacitar a los larvicultores en diversos temas que permitan enriquecer sus conocimientos para aplicar óptimos protocolos de manejo en las diversas áreas de los laboratorios para una producción más sostenible A esto se suma, que cada evento organizado por la CNA, ha mostrado la innovación de productos y servicios con el propósito de brindar a sus asistentes todas las herramientas que permiten tecnificar al sector. Al momento, son más de 12 años realizando eventos en esta provincia, con más de expositores internacionales y locales, y con empresas participantes de diferentes partes del mundo”.

16
Yahira Piedrahita Directora Ejecutiva de la Cámara Nacional de Acuacultura
“Juntos hemos construido este Aqua Expo. Agradecemos a las autoridades presentes por el apoyo brindado en todos nuestros eventos, a las empresas patrocinadoras por la confianza de todos los años, a las empresas expositoras por ser el corazón de la feria y sobre todo a Uds., ya que este evento es posible gracias a su arduo trabajo, a ser los artífices de que la industria siga creciendo y seamos hoy en día la industria más importante del Ecuador, al ser los productores y exportadores principales del mejor camarón del mundo. damos la bienvenida a su evento, al evento de los larvicultores: AQUAEXPO Santa Elena 2023”.




El congreso contó con la participación de 13 expositores nacionales e internacionales que actualizaron los conocimientos en varios temas de interés e importancia para la producción de larvas y los procesos de maduración. Entre los temas podemos mencionar: identificación de patologías de origen infeccioso, aplicación de nuevas herramientas, actualización del plan de vigilancia epidemiológica, soluciones microbianas en acuacultura, buenas prácticas de producción y manejo de alimentos vivos, presentación de tecnologías de microencapsulación y desarrollo genético. Además, nuevas técnicas para obtener larvas de buena calidad, manejo de altas densidades, sistemas óptimos de producción larvaria y análisis de mercado. Los expositores internacionales fueron: Fabrice De Panthou de Francia, Luke Keeton y Adriana Artiles de Estados Unidos, Marcos Santos y Diogo Filipe Rosso de Brasil, Jef Peeters de Bélgica, Violeta Medina y Diva Aldama de México.
Por su parte, la exposición comercial contó con más de 40 stands que mostraron la innovación de sus productos y servicios para toda la cadena de valor, pero especialmente para los interesados en su primer eslabón: la larvicultura. Se destacaron los productos y servicios basados en inteligencia artificial, alimentadores para larvas y aireadores de última tecnología.


“El Mejor Camarón del Mundo tiene sin duda la mejor larva del mundo. Me queda felicitar a los productores de larvas que se dan cita en este evento y felicitar a la Cámara Nacional de Acuacultura, a través de la mesa directiva y demás representantes que están organizando este evento que significa no solo un intercambio de conocimientos sino también un intercambio comercial, donde se puede evidenciar el crecimiento del sector”.
Andrés Arens Viceministro de Producción, Comercio Exterior, Inversiones y Pesca
"Los visito de México, para ver los avances, el progreso y todas aquellas técnicas que podemos implementar para tener unas mejores producciones".
"Me ha llamado la atención la organización. Yo creo que la Cámara siempre hace un trabajo excepcional, en la logística de los eventos. Me siento muy feliz de haber venido".
"Lo que más me ha llamado la atención aquí en la feria es la forma en que podemos asociarnos con nuevos clientes estratégicos para poder realizar negociaciones futuras".

- FEBRERO 2023 COYUNTURA 17
Gabriela Nivelo Directora de Aqua Expo
José Jesús Reyna Asistente al congreso
Adriana Artiles Expositora en la Feria Comercial
Jennyfer Vera Rivas Expositora en la Feria Comercial
"Actualmente veo que muchísimos sectores están sumando a la Aqua Expo, no solamente alimentación, no solamente alimentación o insumos, sino también empacadoras, empresas de tecnología. Y veo que cada día la Aqua Expo sigue creciendo y sobre todo dando este beneficio con capacitaciones y conferencias para el sector camaronero".




los
Estudiante de la Universidad Estatal de Santa
"Es mi primera vez asistiendo a un congreso de esta índole. Aprendí que la innovación y la utilización de diferentes técnicas para el tratamiento y para el desarrollo productivo nos ayudan a nivel nacional en el sector productivo, en este caso del camarón, en el que se destaca Ecuador".
Estudiante de la Universidad Estatal de Santa Elena
"Me pareció excelente y muy interesante para conocer cómo está actualmente el sector camaronero en proyectos e investigaciones, no solamente de aquí del país, sino a nivel internacional también".
Estefanie Cadena
Visitante de la Feria Comercial
"Destaco la forma cómo expusieron los temas; para las personas que tal vez no conocemos tanto lo hizo bastante didáctico".
Franklin Ramírez
Participante del congreso
"La intervención que más me llamó la atención fue la utilización de bacteriófagos, una tecnología bastante amigable, que llama la atención y que tiene aún mucho potencial por seguir explotando".
La Cámara Nacional de Acuacultura es la entidad que organiza desde hace más de dos décadas este evento, con el objetivo de promover la capacitación permanente del sector e impulsar su innovación para afianzar su desarrollo.
Los próximos encuentros se realizarán en Manabí el 10 y 11 de mayo en el Salinas Golf & Tenis Club de Bahía de Caráquez; en el Oro, del 11 al 13 de julio en el Hotel Oro Verde de Machala y en Guayaquil, del 23 al 26 de octubre en el Centro de Convenciones.


- FEBRERO 2023 COYUNTURA 18
Gabriela Corona Expositora en la Feria Comercial
Elena
Visualiza
videos Aqua Expo Santa Elena


en pérdidas suma el sector camaronero en lo que va del año
Más de 20 eventos delictivos contra camaroneros se han registrado en lo que va del 2023, tanto a nivel fluvial como terrestre en las provincias de Guayas y el Oro. Por vía marítima, las víctimas han sido interceptadas por varias embarcaciones, sus tripulantes los han amedrentado disparando con armas automáticas. Al momento, más de 20 personas han resultado heridas, según estadísticas de la Dirección de Seguridad de la Cámara Nacional de Acuacultura.
En comparación con el año pasado, el robo de camarón se ha incrementado en un 300%, del alimento balanceado un 200% y el número de heridos ha crecido en un 250%.
Ante la grave situación, José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura en su cuenta de Twitter exhortó nuevamente al gobierno a atender con celeridad los problemas de inseguridad, a manos del crimen organizado, que a inicios de este año cobró la vida de un guardia de seguridad vinculado al sector camaronero.
“La inseguridad ha llegado a niveles críticos, nunca antes vistos, en el Ecuador. No existe sector productivo que esté exento de sufrir ataques de grupos criminales. ¡No hay reacción eficaz de las autoridades responsables de la seguridad ciudadana!”, precisó Camposano.

Por su parte, la Cámara Nacional de Acuacultura, a través de su Dirección de Seguridad, coordina mesas de trabajo con representantes de la Policía Nacional en la zona 8-5 y la Armada del Ecuador, con el propósito de diseñar nuevas estrategias de control en corredores fluviales y carreteras. Sin embargo, las autoridades no cuentan con los recursos gubernamentales para efectuar patrullajes en las zonas georreferenciadas como peligrosas.
Nuevamente, el titular de la CNA, solicita al gobierno nacional a tomar acciones concretas
para dotar a sus entidades de control de los insumos necesarios para realizar patrullajes, intensificar labores de inteligencia y aplicar estrategias efectivas que detengan la ola de
hechos delictivos que amenazan la vida de trabajadores, relacionados al sector acuícola.
“Una zona que se ha transformado en una zona roja para las diferentes áreas productivas, pero principalmente para la actividad camaronera. Un área en la que estamos siendo asaltados con violencia, con armamento pesado, prácticamente todas las semanas desde hace cuatro meses atrás. El sector camaronero documentó los hechos violentos que se han dado en esta zona de Taura principalmente, y los expuso a través de un oficio al ministro Juan Zapata, a fines del mes de noviembre, cuando la situación ya era insoportable. A la fecha, ese oficio, esa denuncia por escrito realizada al señor ministro, no ha sido respondida y a pesar de que se han dado las instrucciones para que se realicen los diferentes operativos que permitan desmontar a estas bandas de criminales que están operando con total impunidad”.
José Antonio Camposano Presidente Ejecutivo de la Cámara Nacional de Acuacultura

Más de 20 heridos y más de medio millón de dólares
21
“El último trimestre del año 2022 hemos tenido aproximadamente 12 eventos que cuantifican una pérdida de alrededor de medio millón de dólares, lo que implica que las estructuras que se encuentran involucradas hayan generado una capacidad logística para poder generar muchos y más fuertes delitos alrededor de este sector“.
Christian Correa
“Esperamos que la policía, con la ayuda de la empresa privada, en este caso las camaroneras, por la preocupación que contenemos, obviamente adopten una tendencia más al soporte. Más que una reacción, nosotros pedimos un tema de inteligencia. Este es un tema de bandas organizadas, entonces sí es importante ya generar algún tipo de gestión”.
José Landívar
“Se pide de parte de las autoridades lo que actualmente estamos haciendo a través de la Cámara Nacional de Acuacultura, a través de los organismos respectivos. Que ellos accedan a través de inteligencia policial y puedan hacer un trabajo más efectivo desarmando estas bandas delictivas”.



“Nosotros estamos desplegando el trabajo acá en este sector, vamos a continuar acentuando ciertas estrategias operativas conforme a lo que vamos apreciando. Vamos a continuar la visita en otros puntos específicos, en donde vamos a generar también un mapeo situacional de toda la problemática en ese sector. Lo estamos nosotros analizando y en ese sentido se van a ejecutar otras estrategias que esperemos den buenos resultados en un corto plazo y podamos nosotros generar una situación de mayor seguridad en este sector”.
Crnel. Gustavo Pérez
Miembros de la Policía Nacional, directores de seguridad de las empresas afectadas y representantes de medios de comunicación, recorrieron la parroquia Taura, en la provincia del Guayas, para constatar los daños en este eje vial que facilitan la operación delincuencial.



 Jefe de seguridad de empresa afectada por la delincuencia
Jefe de seguridad de empresa afectada por la delincuencia
Dick Aguilar Jefe de seguridad de empresa afectada por la delincuencia
Dirección General de Seguridad Ciudadana y Orden Público de la Policía Nacional
Jefe de seguridad de empresa afectada por la delincuencia
Jefe de seguridad de empresa afectada por la delincuencia
Dick Aguilar Jefe de seguridad de empresa afectada por la delincuencia
Dirección General de Seguridad Ciudadana y Orden Público de la Policía Nacional
22

Programas de mejoramiento genético han sido cruciales para potenciar la especie Penaeus
Vannamei en los laboratorios de larvas
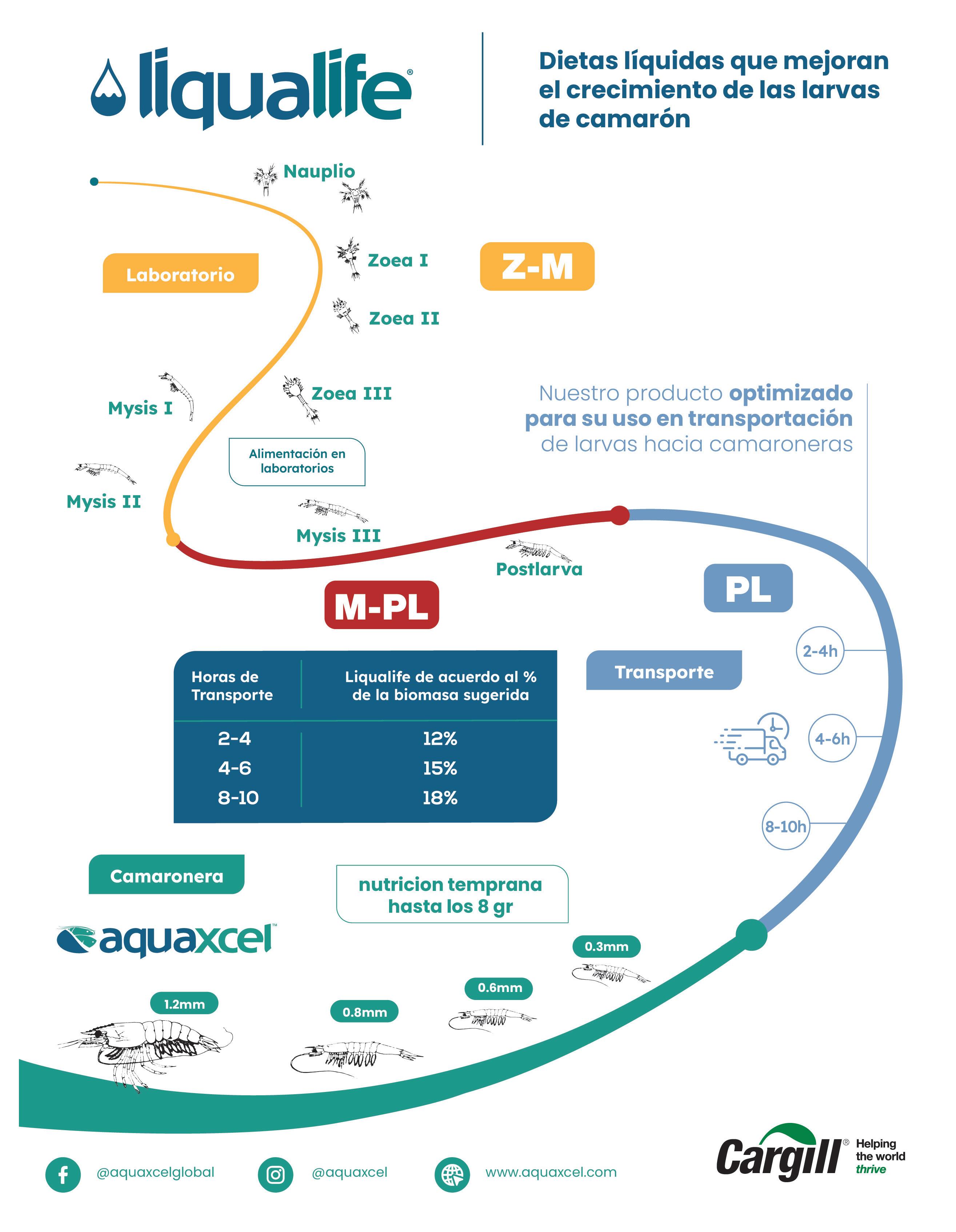
La larvicultura tuvo su origen en la década de los 80. Inicialmente se capturaba larva salvaje de dos especies P. vannamei y P. stylirostris; además se comercializaban reproductores capturados por la “Flota de arrastre de camarón”, pero con el paso del tiempo y con el objetivo de preservar las poblaciones silvestres de camarón y otras especies acompañantes de la captura, el Ministerio de Industrias, Comercio, Integración y Pesca de aquel entonces, resolvió mediante Acuerdo Ministerial la prohibición de por vida de captura de larvas silvestres a en el 2002. Desde entonces, empezó el cultivo de larvas en establecimientos que en su mayoría se encuentran ubicados en la provincia de Santa Elena
Una de estas maternidades de la especie p. vannamei es el laboratorio de Omarsa, ubicado en Mar Bravo. Tiene una extensión de más de 16.000 metros cuadrados. Integra áreas como el departamento de algas, microbiología y biología molecular, investigación, cuarentena, maduración, cría larvaria, artemia y alimento fresco.

El laboratorio siembra 300 nauplios por litro y su ciclo de producción es de 18 a 20 días, para despachar postlarvas de 140 a 200 pl./gramo y su capacidad de siembra es de 3.920 toneladas
Aldo Vanoni, destacado empresario
ecuatoriano vinculado desde hace más de cuatro décadas a la acuicultura, contó su experiencia en producción de larvas y manejo de reproductores en el programa CNA VIDEO PODCAST. A continuación, parte de su entrevista.
¿Cómo inició el proceso de selección genética en el Ecuador?
En 1988, tuve la oportunidad de conocer a expertos en genética que estaban trabajando en Panamá, Venezuela y Colombia: Harvey y Amber Persyn; así que tuve esa inmensa alegría de poderlos invitar a que venga al Ecuador a compartir su experiencia con nosotros.
La visita fue clave porque luego de unos años posteriores a esta reunión vino la Mancha Blanca. Es decir, ya habíamos comenzado con nuestro proceso de mejoría genética masal, cuando pasó el problema de la Mancha Blanca que para la industria ecuatoriana fue importante.
Se dieron cuenta de una u otra manera que de los animales que estaban en piscina, prácticamente un cinco o un 10% sobrevivían y decían: bueno, ¿que tienen estos animales que pueden tener la posibilidad de haber estado en un ambiente con tantos problemas y subsistir? Y ahí nos llevó a tener la primera parte de los reproductores resistentes poco a poco a la enfermedad de la Mancha Blanca. No fue inmediato, pero cuatro o
cinco o seis años después comenzamos a ver que las producciones mejoraban y luego lógicamente el proceso fue avanzando tanto que tuvimos que hacer análisis genéticos un poco más avanzados, y eso nos ayudó a evitar el Inbreeding y a evitar la familiaridad más cercana.
También nos permite no tener tantos problemas de enfermedades. Hablo a nivel de país, por que se han hecho muy buenos proyectos y se está trabajando bastante bien en genética.
¿Qué tipo de variables lograron ustedes identificar en aquella época, década de los 80, con este experto para iniciar el mejoramiento?
En aquellos tiempos todavía se trabajaba con larva salvaje, había mucha dificultad para analizar, pero posteriormente a esa época se comenzó a trabajar solamente con larvas que se iniciaban en los laboratorios del Ecuador. Luego se hicieron algunos procesos de investigación en el que se hacían apareamientos dentro del laboratorio de las larvas salvajes. Luego artificialmente se lo hacía también, hasta que finalmente todos entendimos que lo natural es mucho mejor y más viable. Hoy en día, las producciones del manejo larval a nivel interno de los laboratorios, hablando del proceso de reproductores, es mucho más natural que hace décadas.
24
Estableciendo una comparación con aquellos inicios en los 80, ¿qué tipo de variables se deben tomar en cuenta ahora?
Definitivamente el control de las enfermedades, tanto en laboratorio como en las piscinas de producción. Desde que ingresan le hacemos controles de diferentes enfermedades. Tenemos un departamento de microbiología y otro de biología molecular, en los cuales podemos revisar todos los procesos y controles de los animales que están ingresando. Igualmente hacemos los mismos controles cuando ya es producto terminado, cuando es larva o cuando tenemos que hacer los análisis previos de toda la alimentación, para que no tengamos ningún efecto negativo en la producción. Eso nos permite tener una estabilidad, tanto en la parte de la maduración como en la larvicultura. El medio puede venir cargado de patógenos o de vidrios, es decir, lo intentamos controlar con ácidos orgánicos para evitar daño en la producción.
En este sentido, ¿cómo manejan ustedes el afluente y el efluente en su sistema de producción?
Bueno, todos los laboratorios tenemos casi los mismos procedimientos al tener las puntas que le llamamos para la captación del agua y bombas que están al interior, y esa captación natural que se hace, prácticamente forma un control biológico del agua que está ingresando. Hay filtros, pero se forma una película natural prácticamente,
que arma un proceso natural dentro de la captación del agua a través de la punta y hasta que llegue a la bomba, y luego se distribuye internamente a las diferentes áreas de producción. Cuando llega ahí está totalmente analizada, es decir, desde el mar hasta antes de que llegue a la punta y luego de la punta; lo que llega acá son análisis diferentes que se hacen para controlar los procesos.
Una vez que tenemos eso, hacemos el equilibrio del medio para que el animal, en este caso la larva, tenga un agua completamente limpia.

¿Por qué es tan importante analizar la calidad de agua?
Es clave. Todo lo hacemos a través de la microbiología, pero lo que intentamos es controlar todos los procedimientos y evitar inconvenientes en el proceso; es decir, desde el ingreso del agua, el manejo es clave, el cultivo y control de algas también son importantes. Todos los alimentos naturales o alimentos balanceados son controlados a través de microbiología para intentar, a través de los procesos que acabamos de conversar, equilibrar el medio. Que todo esté básicamente controlado.
Aldo Vanoni Destacado larvicultor ecuatoriano

25
“Todos dependemos de la larva, como también dependemos de la producción camaronera para que nuestro país siga liderando en el mundo las producciones camaroneras”.
Es increíble, todo lo que puede cambiar de un día a otro. Entonces muchas veces dice: Bueno, ¿por qué no se produce igual una corrida con otra? Porque los momentos son diferentes.
Hay muchos factores que inciden en eso…
¿Cuál es la importancia del uso de ácidos orgánicos?
Producimos probióticos, hay muchos que están en el mercado y son analizados constantemente antes de que ingresen a producción.
Un aspecto muy importante también, es cómo se devuelve el agua a su cauce, ¿cómo hacen ustedes hacen el tratamiento del efluente?
Todas las aguas, tanto de la larvicultura como de maduración y de algas, pasan por un proceso de control. Así como controlamos el ingreso, también lo hacemos al egresar y usamos los mismos métodos; es decir,
los probióticos, que son necesarios para eliminar los problemas que van saliendo y evitar también que a través de esos efluentes dañemos el ecosistema.
No todos los laboratorios tienen microbiología, pero hay que entender que los procesos se pueden copiar para bajar la incidencia de los problemas que regresan al mar.
Comprendemos que el agua que se toma pierde oxígeno, pero con todos los procesos actuales de tecnología, alimentación y aireación, entendería yo que el agua que regresa al estero esté mejor de calidad que la que ingresa a la producción.
Otro aspecto importante dentro del laboratorio, es realizar un buen proceso de desinfección. ¿Cómo ustedes lo manejan?
Nosotros tenemos ya hace algún tiempo atrás, un grupo especializado solamente en el manejo de la limpieza poscosecha. El control
a través de hisopados del Departamento de Microbiología, en el cual salen resultados y nos indican qué productos, qué problemas tenemos, cómo solucionarlos, y empezamos con un grupo solamente a sacar cada una de las tuberías de agua, cada una de las tuberías de aire en la que tengamos que limpiar, mantener, hasta que venga el nuevo proceso de siembra de cada tanque.


¿Cuánto se toma este tiempo de limpieza y de secado?
Entre ocho y diez dias. Digamos, es un proceso en el que ya has tenido la posibilidad de hacer la limpieza continua de todo lo que va dentro de un tanque y la cantidad de tiempo que has usado para, con productos, eliminar todos los problemas que durante el proceso has captado en esa tubería, tanto de aire y de agua. Luego se vuelve a armar y se pone en funcionamiento.
¿Qué tipos de tecnologías considera usted que se han podido incorporar a su
26
Dr. Leonardo Hidalgo realiza controles en los tanques de maduración.
laboratorio de larvas en los últimos años?


Más que la tecnología, es concientizar a las personas que trabajan en la empresa, y en los diferentes laboratorios, a producir de la mejor manera, en la forma más natural posible. Eso garantiza que el producto tenga una buena calidad y llegue a los productores camaroneros de la mejor manera.
¿Cuáles son los principales desafíos que puede enfrentar la larvicultura en el Ecuador actualmente?
Como la larvicultura, yo no encuentro mayor desafío, sino más bien el crecimiento constante y la necesidad de la cantidad de larva que mes a mes se va realmente aumentando, va in crescendo, porque a partir de que la tecnología es usada en nuestro sistema de producción camaronero, ha aumentado constantemente la producción camaronera, como también debe aumentar la producción de larvas. No caigamos en la desesperación de seguir produciendo más, aumentando densidades que podrían
provocar algún tipo de enfermedades que no las tenemos felizmente, pero que hemos visto estos procesos en otros países y que lamentablemente han tenido nefastas consecuencias para la producción camaronera.
Creo que el Ecuador debe manejarse con límites en cuanto a las siembras, que hasta
ahora siendo extensivos nos ha ido muy bien, y hoy en día creo que estamos en un proceso semiintensivo del que no debemos salir y que debe mantenerse, con relación a la cantidad de larva que se produce•

27
VEA LA ENTREVISTA COMPLETA ESCANÉANDO EL CÓDIGO QR
Departamento de microbiología
ARTÍCULOS TÉCNICOS ÍNDICE
Identificación de un nuevo solinvivirus con localización nuclear asociado a mortalidades masivas en el cultivo del camarón blanco (Penaeus vannamei)

Incremento en el desempeño del crecimiento y mecanismos de digestión del camarón a través de un potenciador de la digestión con base en lisofosfolípídos bajo diferentes niveles de colesterol en la dieta
Incremento de la supervivencia de camarón blanco (Litopenaeus vannamei) infectado con el virus de la mancha blanca y alimentado con una dieta suplementada con aceite de coco (Cocos nucifera)
Rápida determinación de materia orgánica y carbono orgánico en suelos de camaronera mediante espectroscopia visible e infrarrojo cercano VIS-NIR
Ecosistemas de carbono azul: una estrategia para mitigar el cambio climático global
Edición 151
Febrero 2023 29 38 42 52 57
-
Identificación de un nuevo solinvivirus con localización nuclear asociado a mortalidades masivas en el cultivo del camarón blanco (Penaeus vannamei)
Autores:
Roberto Cruz-Flores 1,2,†
Thales P.D. Andrade 2,3,†
Hung N. Mai 2
Rod Russel R. Alenton 2
Arun K. Dhar 2,*
1 Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE), Carretera Ensenada-Tijuana No. 3918, Zona Playitas, Ensenada 22860, Baja California, México
2 Laboratorio de Patología Acuícola, Facultad de Ciencias Biomédicas Comparativas y de Animales, Universidad de Arizona, Tucson, AZ 85721, EE. UU.
3 Laboratorio de Diagnóstico de Enfermedades de Crustáceos, Universidad Estatal de Maranhão, Ciudad Universitaria Paulo VI, 1000 Tirirical, São Luis 65055-970, MA, Brasil

*Correspondencia: adhar@arizona.edu
†Estos autores contribuyeron equitativamente en este trabajo
Publicación original:
https://www.mdpi.com/1999-4915/14/10/2220
https://pubmed.ncbi.nlm.nih.gov/36298775/
Mientras el cultivo de camarón siga siendo uno de los sectores más rentables en la industria de producción de alimento animal, la expansión del cultivo de camarón a nuevas regiones y la implementación de técnicas de cultivo innovadoras continuarán. Sin duda, esto implica el movimiento de líneas genéticas particulares de camarón entre regiones, países y continentes. La aparición y distribución transfronteriza de agentes patógenos asociados con la producción de peneidos está bien documentada [1] y está directamente relacionada con el movimiento de reproductores y subproductos del camarón [2]. La aparición y propagación de enfermedades en la industria acuícola corresponde a prácticas industriales deficientes y ha ocasionado la distribución cosmopolita de todos los patógenos que alguna vez han afectado a la industria camaronera [1,3].
Las enfermedades virales han sido, sin duda, una de las mayores amenazas para la sostenibilidad del cultivo de camarón. Patógenos como el virus de la necrosis hipodérmica y hematopoyética infecciosa (IHHNV), el virus del síndrome de la mancha blanca (WSSV), el virus del síndrome de Taura (TSV), el virus de la cabeza amarilla (YHV) y el virus de la mionecrosis infecciosa (IMNV) han causado importantes panzootias que tuvieron un impacto global en el precio del camarón y en el sustento de muchas personas que dependen directa e indirectamente de la producción de camarón [4–8]. Más recientemente, se ha evidenciado que una cepa particular de IMNV que causa una mortalidad inusual en Brasil podría deberse al movimiento de animales desde Indonesia [9]. Además, estas mismas muestras de Brasil mostraron la presencia de otra secuencia viral única correspondiente a un nuevo virus que inicialmente propusimos que pertenecía a la familia Caliciviridae [10], pero que ahora hemos reclasificado como miembro de los Solinviviridae. La presencia de este nuevo virus en camarón cultivado de Brasil ayuda a explicar las mortalidades inusuales que han estado ocurriendo desde 2016 y muestra el surgimiento de un nuevo patógeno que amerita un monitoreo directo para evitar su propagación a otras regiones de cultivo de camarón.
- FEBRERO 2023 PATOLOGÍA
29
La familia Solinviviridae consta de virus tipo picorna/calici con genomas de ARN lineales de sentido positivo no segmentados de 10–11 kb [11]. A diferencia de otros virus, las proteínas de su cápside están codificadas hacia el extremo 3′ del genoma, donde pueden expresarse de un ARN subgenómico o como una extensión de la replicación [11]. Los miembros de esta familia forman partículas icosaédricas con un diámetro de 26-30 nm y parecen tener proyecciones [12,13]. Actualmente, la familia Solinviviridae incluye dos géneros Invictavirus y Nyfulvavirus, y son parte de un gran grupo diverso de virus que infectan artrópodos dentro del grupo de picornavirus y calicivirus [11]. Actualmente, no existen criterios oficiales de delimitación de género para esta familia.
Los Solinviviridae contienen especies virales que son patógenos conocidos de hormigas e infectan el epitelio del intestino medio [14]. Algunos de estos virus causan infecciones crónicas y baja mortalidad que están restringidas a un tipo de tejido (epitelio del intestino medio), mientras que las especies más virulentas causan infecciones sistémicas y mortalidad aguda [12,14]. Si bien las especies de solinivivirus mejor estudiadas se encuentran en las hormigas (Solenopsis invicta virus 3 y Nylan deria fulva virus 1), estudios recientes sobre la virosfera del ARN de invertebrados han demostrado que secuencias de virus no clasificados pero relacionados a los solinvivirus están presentes en otros insectos y artrópodos [11,15]. En este estudio, reportamos la caracterización genómica de un nuevo solinivivirus en la especie de camarón de cultivo más importante, Penaeus vannamei, y las metodologías de diagnóstico (PCRq e ISH) para su detección. Hemos denominado provisionalmente a este virus “Penaeus vannamei solinvivirus” (PvSV).
Material y Métodos
Origen de la muestra
Los P. vannamei enfermos examinados en este estudio se recolectaron entre 2016 y 2019 de estanques afectados ubicados en la región noreste de Brasil, incluyendo al Estado de Maranhão (Perizes de Baixo), el Estado de Piauí (Mexeriqueira), el Estado de Ceará (Camocim, Jaguaruana, Aracati y Alto Santo) y el Estado de Pará. El peso promedio de los camarones juveniles
enfermos en los estanques afectados fue de aproximadamente 3.0 g. Se recolectaron de cinco a seis camarones de cada granja y se examinaron mediante un análisis RT-PCR en tiempo real, histopatología H&E e ISH. El aislamiento del ARN se realizó utilizando aproximadamente 25 mg de músculo picado y pleópodos de los camarones presumiblemente afectados. Se utilizó el kit de purificación de ARN total Tissue LEV (#AS1220, Promega, Madison, WI, EE.UU.) junto con un sistema automatizado de extracción de ADN/ARN (Maxwell® MDX Promega, EE.UU.) siguiendo las recomendaciones del fabricante. La RT-PCR en tiempo real de un paso se llevó a cabo siguiendo los protocolos descritos por Andrade et al. [16]. Antes del ensayo en tiempo real, el ARN extraído se hirvió a 100°C durante 3 min para desnaturalizar el ARNds, luego se colocó en hielo y finalmente se sometió a un análisis de RT-PCR en tiempo real utilizando el sistema GoTaq® Probe
1-Step RT-qPCR (PROMEGA, Madison, WI, EE. UU.). Cada muestra se analizó por duplicado utilizando un termociclador PCR ViiA7 en tiempo real (Applied Biosystems, Foster city, CA, EE. UU.).
Secuenciación de próxima generación
Se envió una muestra 8-Br al Laboratorio de Patología Acuícola (APL) de la Universidad de Arizona para la detección de patógenos. Esta muestra había dado negativo previamente para todos los patógenos virales, bacterianos y fúngicos conocidos en los camarones cultivados. El ARN de esta muestra se envió para ARN-Seq a OmegaBioservices, Norcross, GA, EE.UU. La preparación de la biblioteca se llevó a cabo utilizando un TruSeq Stranded Total RNA Library Prep (Illumina®, San Diego, CA, EE.UU.). Las muestras se secuenciaron con un sistema Illumina HiSeq 2500 (PE 2X150PE).
Análisis Bioinformático
Se verificó la calidad de las 40,904,762 lecturas pareadas de la muestra 8-Br y se recortaron antes de asignarlas al conjunto de secuenciación del genoma Keihai No.1 RefSeq de P. vannamei (PRJNA508983) para eliminar las lecturas del host utilizando el mapeador Geneious Prime con parámetros predeterminados (biomateria) [17]. Las lecturas duplicadas se eliminaron usando Dedupe en Geneious Prime antes del mapeo.
Las lecturas no asignadas se ensamblaron de nuevo utilizando el ensamblador Geneious Prime con los parámetros predeterminados. Los contigs y frames Open Reading fueron analizados por las herramientas de búsqueda BLASTN y BLASTP [18]. La búsqueda de patrones o “motif” se realizó utilizando Motif Search y Prosite Expasy para identificar dominios conservados en la secuencia viral. Las señales de localización nuclear (NLS) se predijeron utilizando NLStradamus utilizando el modelo dinámico HMM de 2 estados con un límite de predicción de 0.9 [19].
Análisis filogenético
El dominio RdRp del PvSV de Brasil se comparó con otros virus del orden Picornavirales. Las secuencias de las familias Caliciviridae (AHX24377, AAL99277 y AYF53102), Dicistroviridae (NP620562, AAF80998 y AF277675), Iflaviridae (NP_853560, NP_049374, NP_277061 y NP_620559), Marnaviridae (NP_944776), Picornaviridae (NP_740737, BAA31356 y NP_740478), Polycipiviridae (APG774337, ASK12200 y ASK12194), Secoviridae (NP_734447, NP_734463, YP_054443, NP_730557, YP_001039627 y YP_081454) y Solinviridae (AAZ78308, ANQ44728 y ACO37271) se utilizaron en el análisis. Las secuencias se alinearon con Geneious Aligner y el análisis filogenético se realizó con MEGAX [17, 20]. Se construyó un árbol filogenético utilizando el método de unión de vecinos “Neighbor-Joining” [20]. El árbol de consenso “Bootstrap” se infirió a partir de 1000 repeticiones.
Diseño del Primer y Clonación del Fragmento Genómico PvSV
Usando el genoma de PvSV generado a partir del set de data de NGS, se diseñaron un par de primers y una sonda (Tabla 1). Los primers y las sondas se diseñaron con Geneious Prime y se probaron para determinar su especificidad con el PrimerBlast [17,21]. Las regiones de unión del primer y la sonda se seleccionaron mediante el algoritmo de diseño de primers Geneious Prime teniendo en cuenta el rango de tamaño de fragmento óptimo de 100–220 pb y una temperatura de hibridación de 60°C. Los análisis Primer-Blast indicaron que los primers eran específicos para PvSV. La sonda TaqMan se sintetizó y marcó con 6-carboxifluoresceína (FAM) en el extremo 5’
- FEBRERO 2023 30 PATOLOGÍA
y N, N, N’, N’-Tetrametil-6-carboxirrodamina TAMRA en el extremo 3’. Para los ensayos, se utilizó TaqMan Fast Virus 1-Step Master Mix (Applied Biosystems™, Foster city, CA, EE.UU.), la concentración final para cada primer fue de 0.5 µM y 0.1 µM para la sonda TaqMan a un volumen final de 10 µL. El perfil de PCRq fue de 20 s a 95°C seguido de 40 ciclos de 1 s a 95°C y 20 s a 60°C.
El fragmento de ADNc de 133 pb se amplificó a partir de la muestra original 8-Br y se clonó en el vector de clonación pDrive (QIAGEN®, Hilden, Alemania) para usarlo como control positivo. El plásmido con el inserto se denominó PvSV-8-Br. El ADN del plásmido se purificó utilizando el kit QIAprep® Spin Miniprep. La secuencia del fragmento PvSV8-Br se verificó mediante secuenciación en las instalaciones de secuenciación de la Universidad de Arizona, Tucson, AZ, EE. UU.
Histopatología
Todas las muestras de camarón se fijaron con Davidson (330 mL de etanol al 95%, 220 mL de formalina al 100%, 115 mL de ácido acético glacial, 335 mL de agua destilada, pH~3.0–4.0) siguiendo un procedimiento estándar [22]. Después de la fijación, se eliminó el fijador de Davidson y se reemplazó con un volumen igual de etanol al 70%. Para el procesamiento histológico, las muestras se lavaron en una serie de soluciones de alcohol/xileno, se impregnaron en parafina, se seccionaron a 5 µm y se tiñeron con H&E siguiendo un procedimiento estándar [22]. Los portaobjetos histológicos se examinaron usando un microscopio de luz de campo brillante. La gravedad de la infección se clasificó según una escala semicuantitativa que va del grado 0 al grado 4 según una publicación anterior [23].
Hibridación in situ
Todas las secciones se procesaron como se describió anteriormente (histopatología). Sin embargo, para ISH, las secciones se secaron
Nombre del primer/sonda Secuencia del primer (5′ a 3′) Tamaño del producto (nt)
3136 F (Set 1)
3268 R (Set 1)
Probe 3159 (Set 1)
en portaobjetos microscópicos cargados positivamente y el ISH se llevó a cabo siguiendo los protocolos descritos por CruzFlores et al. [24], con una mezcla equitativa de los primers (3136 F y 3268 R) para el PCRq y la sonda (Probe 3159). Estos primers y la sonda se marcaron en el extremo 3’ con digoxigenina-11-dUTP (Sigma-Aldrich™, St. Louis, MO, EE.UU.). Se utilizó como control positivo un bloque de parafina que contenía camarones afectados previamente con PvSV. Como control negativo, se utilizaron bloques de parafina de camarones libres de patógenos específicos. Los portaobjetos se examinaron mediante microscopía óptica para detectar la presencia o ausencia de PvSV que hibridaba con la sonda de ADN, y los portaobjetos que mostraban precipitados de color azul a púrpura se consideraron positivos.
Resultados
Organización del genoma de Penaeus vannamei solinvivirus y análisis de secuencias
El genoma completo de PvSV es de 10.44 kb (excluyendo la cola poli A) y presentó una cobertura promedio de 9853. Identificamos un marco de lectura abierto (ORF) grande (9981 nt) que codifica una poliproteína de 3326 aa con similitud a Picornavirales sp. y Riboviria sp (Tabla 2). La búsqueda de motifs o patrones identificó cuatro dominios conservados que codifican una Helicasa, ARN polimerasa dependiente de ARN (RdRp), proteína cubierta de calicivirus, parche G y proteína de tegumento (Figura 1). La
organización del genoma es similar a la de otros solinvivirus (Nylan deria fulva virus 1) que muestran una gran ORF. El análisis de la señal de localización nuclear (NLS) indica que hay un NLS putativo en la poliproteína alrededor de 4268–4474aa. La secuencia del genoma de PvSV Se depositó en GenBank con el número de acceso OP265432.
El análisis de secuencia de ORF1 mostró una identidad del 93.84% con el virus 8 del camarón Wenzhou (NC_032852.1), un riboviria no clasificado. Además, el análisis de la secuencia de aminoácidos (aa) de los dominios conservados de helicasa y RdRp mostró una similitud del 100% y del 99.22% con la proteína hipotética del virus 8 del camarón Wenzhou sin clasificar (YP_099336733.1), como se ve en la Tabla 2.
Filogenia
Las inferencias filogenéticas utilizando el método de unión de vecinos para RdRp indicaron que los nuevos grupos de virus con la familia Solinviviridae posiblemente representaban un nuevo género dentro de esta familia (Figura 2).
3.3. Distribución del Penaeus vannamei Solinvivirus en Brasil
Los primers diseñados para el PvSV amplifican un fragmento de 133 nt del genoma viral. El fragmento clonado se secuenció utilizando el método de Sanger y mostró una identidad del 99.9% con la secuencia del genoma completo de PvSV. Este plásmido se usó posteriormente como control positivo para los ensayos de detección de PCRq.
Figura 1. Organización del genoma del Penaeus vannamei solinvivirus (PvSV). El genoma consta de un ORF grande con cinco dominios conservados identificados, que incluyen una helicasa de ARN (verde), una señal de localización nuclear (naranja), una ARN polimerasa dependiente de ARN (rosa), una proteína del tegumento (naranja), una proteína cubierta de Calicivirus (azul) y parche G (verde claro).
- FEBRERO 2023 31 PATOLOGÍA
Tabla 1. Primers y sondas diseñadas para la detección específica de Penaeus vannamei solinvivirus (PvSV) por RT-PCR en tiempo real.
TACGCCACGAACGAGAACAA GGACAGCGACAAAGACGAGA [FAM]CGTCGTGACTACTCTCACCG [TAM] 133
El PvSV está ampliamente distribuido en las granjas brasileñas y está presente en los Estados de Maranhão (Perizes de Baixo), Piaui (Mexeriqueira), Ceará (Camocim, Jaguaruana, Aracati y Alto Santo) y Pará. El número total de casos positivos de 2016 a 2019 se registró mediante el ensayo RT-PCR de PvSV en tiempo real. En este estudio, de los 13 casos sospechosos seleccionados, se detectó PvSV en 11 casos (84%) (Tabla 3).
Histopatología e Hibridación In Situ
El PvSV infecta células del tracto gastrointestinal (Figura 3). La señal de las sondas marcadas con DIG se observó en el núcleo de las células epiteliales de los túbulos del hepatopáncreas, las células epiteliales del intestino y el estómago (Figuras 3 y 4F1, F-2). En el órgano linfoide, la señal de la sonda marcada con DIG se observó tanto en el citoplasma como en el núcleo (Figura 4E-1, E-2). De manera similar, la señal se observó en el núcleo de las células del músculo estriado (Figura 4G-1, G-2, H-1, H-2). No se observó ninguna reacción a la sonda específica de PvSV en el tejido del camarón SPF (Figura S1).
Discusión
Desde 2016, se han registrado mortalidades inusuales que progresaron muy rápido y resultaron en una mayor mortalidad acumulada (hasta 80%) en los Estados de Pará, Maranhão, Piauí, Ceará, Río Grande del Norte, Alagoas, Sergipe y Bahía en Brasil [9]. Como mencionamos anteriormente, encontramos que estas mortalidades están asociadas con una nueva cepa de IMNV [9]. Además, durante el análisis del transcriptoma de los camarones infectados con IMNV, identificamos una secuencia viral divergente adicional que corresponde a un nuevo miembro de Solinviviridae que hemos denominado tentativamente PvSV. La identificación de PvSV en camarones brasileños que experimentan mortalidades inusuales en coinfección con IMNV podría sugerir que existe una interacción sinérgica entre los dos virus, contribuyendo a una enfermedad que progresa más rápidamente y a mortalidades más altas observadas en diferentes estados brasileños. El movimiento transfronterizo de reproductores y postlarvas entre las naciones camaroneras sigue siendo una de las razones principales por la cual todos los
vannamei solinvivirus (PvSV) a otros virus de GenBank.
Detección de IMNV y PvSV por RT-PCR en tiempo real en Penaeus vannamei obtenidos de granjas camaroneras que experimentaron brotes de enfermedades. Estas fincas están ubicadas en cuatro Estados diferentes de Brasil. ND = No detectado.


Tabla
Figura
La historia evolutiva de Penaeus vannamei solinvivirus (PvSV) se infirió utilizando el método “NeighborJoining”. La figura muestra que el PvSV (recuadro rojo) se asocia con la familia Solinviviridae y es divergente del resto de los virus de esta familia. Se muestra el árbol óptimo con la suma de longitud de rama = 16.70591706. El porcentaje de árboles replicados en los que los taxones asociados se agruparon en la prueba de arranque o bootstrap (1000 réplicas) se muestran junto a las ramas. El árbol está dibujado a escala, con longitudes de rama en las mismas unidades que las distancias evolutivas utilizadas para inferir el árbol filogenético. Las distancias evolutivas se calcularon usando el método de corrección de Poisson y están en las unidades del número de sustituciones de aminoácidos por sitio. Este análisis involucró 29 secuencias de aminoácidos. Se eliminaron todas las posiciones ambiguas para cada par de secuencias (opción de eliminación por pares). Hubo un total de 610 posiciones en el conjunto de data final.

- FEBRERO 2023 32
PATOLOGÍA
Tabla 2. Similitud de secuencia de aminoácidos de ORF1, Helicasa y ARN polimerasa dependiente de ARN de Penaeus
3.
2.
(PvSV). (A-1-D-1) Inclusiones virales basófilas y eosinofílicas en las células epiteliales de túbulos intestinal, estomacal y hepatopáncreas de P. vannamei con PvSV; estos se muestran con las flechas negras. La señal positiva de las sondas marcadas con DIG se observa en el núcleo de las células epiteliales del intestino y del hepatopáncreas (A-2-D-2). El cuadro con línea continua en el Panel C-2 se muestra en el Panel D-2, y el cuadro con línea punteada se muestra en la esquina derecha en el Panel D-2.

principales patógenos del camarón tienen una distribución cosmopolita [9,25,26]. Una nueva cepa de IMNV identificada hace poco en Brasil posiblemente se originó en Asia y se reintrodujo recientemente en Brasil [9]. El PvSV muestra una gran similitud con el virus 8 del camarón Wenzhou, que se detectó originalmente en Asia en 2016 y podría representar una cepa divergente de este virus [15]. Se ha encontrado que secuencias virales adicionales de Tailandia [27], China [28] y Australia [29] que también presentan
una gran similitud con el virus 8 del camarón Wenzhou y, por lo tanto, también podrían representar virus relacionados con el PvSV. Esto plantea dudas sobre si PvSV se introdujo en Brasil a través del movimiento de animales de las naciones mencionadas que cultivan camarón. Sin embargo, aún se desconoce si una cepa recién emergida de IMNV y PvSV se introdujeron simultáneamente o en diferentes eventos de introducción.
Solinviviridae es una familia de virus que
Referencias
1. Stentiford, G.D.; Neil, D.M.; Peeler, E.J.; Shields, J.D.; Small, H.J.; Flegel, T.W.; Vlak, J.M.; Jones, B.; Morado, F.; Moss, S.; et al. Disease will limit future food supply from the global crustacean fishery and aquaculture sectors. J. Invertebr. Pathol. 2012, 110, 141–157. [CrossRef]
2. Dhar, A.K.; Lakshman, D.K.; Amundsen, K.; Robles-Sikisaka, R.; Kaizer, K.N.; Roy, S.; Hasson, K.W.; Thomas Allnutt, F.C. Characterization of a Taura syndrome virus isolate originating from the 2004 Texas epizootic in cultured shrimp. Arch. Virol. 2010, 155, 315–327. [CrossRef] [PubMed]
3. Lightner, D.V. Global transboundry disease politics: The OIE perspective. J. Invertebr. Pathol. 2012, 110, 184–187. [CrossRef] [PubMed]
4. Shike, H.; Dhar, A.K.; Burns, J.C.; Shimizu, C.; Jousset, F.X.; Klimpel, K.R.; Bergoin, M. Infectious Hypodermal and Hematopoietic Necrosis Virus of Shrimp Is Related to Mosquito Brevidensoviruses. Virology 2000, 277, 167–177. [CrossRef]
5. Van Hulten, M.C.W.; Witteveldt, J.; Peters, S.; Kloosterboer, N.; Tarchini, R.; Fiers, M.; Sandbrink, H.; Lankhorst, R.K.; Vlak, J.M. The white spot syndrome virus DNA genome sequence. Virology 2001, 286, 7–22. [CrossRef] [PubMed]
6. Mari, J.; Poulos, B.T.; Lightner, D.V.; Bonami, J.R. Shrimp Taura syndrome virus: Genomic characterization and similarity with members of the genus Cricket paralysis-like viruses. J. Gen. Virol. 2002, 83, 915–926. [CrossRef] [PubMed]
7. Sittidilokratna, N.; Hodgson, R.; Cowley, J.; Jitrapakdee, S.; Boonsaeng, V.; Panyim, S.; Walker, P. Complete ORF1b-gene sequence indicates yellow head virus is an invertebrate nidovirus. Dis. Aquat. Organ. 2002, 50, 87–93. [CrossRef] [PubMed]
8. Poulos, B.T.; Tang, K.F.J.; Pantoja, C.R.; Bonami, J.R.; Lightner, D.V. Purification and characterization of infectious myonecrosis virus of penaeid shrimp. J. Gen. Virol. 2006, 87, 987–996. [CrossRef] [PubMed]
9. Andrade, T.P.D.; Flores, R.C.; Mai, H.N.; Dhar, A.K. Novel infectious Myonecrosis virus (IMNV) variant is associated with recent disease outbreaks in Penaeus vannamei shrimp in Brazil. Aquaculture 2022, 554, 738159. [CrossRef]
10. Cruz-Flores, R.; Dhar, A.K.; Andrade, T.P.D.; Mai, H.N.; Alenton, R.R.R. Identification of a novel calicivirus with nuclear localization associated with mass mortalities in cultured whiteleg shrimp (Penaeus vannamei). In Aquaculture 2022; Society, W.A., Ed.; San Diego, CA, USA, 2022; p. 448.
11. Brown, K.; Olendraite, I.; Valles, S.M.; Firth, A.E.; Chen, Y.; Guéerin, D.M.A.; Hashimoto, Y.;
- FEBRERO 2023 33 PATOLOGÍA
Figura 3. Secciones teñidas con hematoxilina y eosina e hibridación in situ de secciones paralelas del intestino y hepatopáncreas de Penaeus vannamei infectado con Penaeus vannamei solinvivirus
Herrero, S.; de Miranda, J.R.; Ryabov, E.; et al. ICTV virus taxonomy profile: Solinviviridae. J. Gen. Virol. 2019, 100, 736–737. [CrossRef] [PubMed]
12. Valles, S.M.; Hashimoto, Y. Isolation and characterization of Solenopsis invicta virus 3, a new positive-strand RNA virus infecting the red imported fire ant, Solenopsis invicta. Virology 2009, 388, 354–361. [CrossRef] [PubMed]
13. Valles, S.M.; Oi, D.H.; Becnel, J.J.; Wetterer, J.K.; LaPolla, J.S.; Firth, A.E. Isolation and characterization of Nylanderia fulva virus 1, a positive-sense, single-stranded RNA virus infecting the tawny crazy ant, Nylanderia fulva. Virology 2016, 496, 244–254. [CrossRef]
14. Valles, S.M.; Strong, C.A.; Dang, P.M.; Hunter,W.B.; Pereira, R.M.; Oi, D.H.; Shapiro, A.M.;Williams, D.F. A picorna-like virus from the red imported fire ant, Solenopsis invicta: Initial discovery, genome sequence, and characterization. Virology 2004, 328, 151–157. [CrossRef]
15. Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [CrossRef]
16. Andrade, T.P.D.; Srisuvan, T.; Tang, K.F.J.; Lightner, D.V. Real-time reverse transcription polymerase chain reaction assay using TaqMan probe for detection and quantification of Infectious myonecrosis virus (IMNV). Aquaculture 2007, 264, 9–15. [CrossRef]
17. Kearse, M.; Moir, R.; Wilson, A.; StonesHavas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data.
Bioinformatics 2012, 28, 1647–1649. [CrossRef] [PubMed]
Figura
y
e hibridación in situ de secciones paralelas de tejido de Penaeus vannamei infectado con el Penaeus vannamei solinvivirus (PvSV). (E-1) Órganos linfoides esferoides en camarones enfermos. (E-2) Órganos linfoides esferoides que muestran una reacción citoplásmica y nuclear a la sonda específica de PvSV. (F-2) Epitelio intestinal que muestra gran cantidad de inclusiones intranucleares con reacción positiva a la sonda de PvSV, no se observan anormalidades aparentes en los núcleos infectados (F-1). (G-1-H-2) Bajo y alto aumento del tejido muscular que también muestra una reacción positiva en los núcleos. En la esquina derecha del panel se muestra una vista ampliada del cuadro con línea continua en el Panel F-2.
infecta a las hormigas, pero los virus relacionados infectan a una gran variedad de insectos y otros artrópodos [11–13,30]. Estos virus están relacionados con Caliciviridae y Picornaviridae, y también poseen genomas de ARN lineales, no segmentados y de sentido positivo de ~10–11 Kb [11]. Sin embargo, Solinviviridae difiere de Caliciviridae en términos de tamaño del genoma y organización ORF. Los Caliciviridae tienen genomas más cortos,
como genomas de 6.4-8.5 kb con dos o tres ORFs [31]. Inicialmente habíamos asignado el PvSV a la familia Caliciviridae teniendo en cuenta el patrón o motif de la proteína de la cubierta del calicivirus y un análisis filogenético limitado [10]. Sin embargo, tras un análisis filogenético adicional y la asignación taxonómica del virus 8 del camarón Wenzhou a la familia Solinviviridae por el Comité Internacional de Taxonomía de virus capítulo para Solinviviridae, hemos

18. Altschul, S.F.; Gish,W.; Miller,W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [CrossRef]
19. Nguyen Ba, A.N.; Pogoutse, A.; Provart, N.; Moses, A.M. NLStradamus: A simple Hidden Markov Model for nuclear localization signal prediction. BMC Bioinform. 2009, 10, 202. [CrossRef]
20. Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. B 2018, 33, 1870–1874. [CrossRef] [PubMed]
21. Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3-new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [CrossRef] [PubMed]
22. Bell, T.A.; Lightner, D.V. A Handbook of Normal Penaeid Shrimp Histology, 1st ed.;World Aquaculture Society: Baton Rouge, LA, USA, 1988.
23. Lightner, D.D.V. A Handbook of Shrimp
- FEBRERO 2023 34 PATOLOGÍA
4. Secciones teñidas con hematoxilina
eosina
concluido que el nuevo virus del camarón de Brasil es un miembro de esta familia. 32]. La afiliación taxonómica del Penaeus vannamei picornavirus (PvPV) [28] debe evaluarse más a fondo utilizando la región putativa RdRp para aclarar su ubicación en el orden Picornavirales Solinviviridae es una familia relativamente nueva y la mayoría de los miembros de la familia permanecen sin clasificar. Hasta la fecha, solo se han asignado dos miembros, a saber, el Invictavirus y el Nyfulvavirus [11,13,33]. El PvSV representa un género nuevo dentro de la familia y es el primer solinvivirus conocido que infecta camarones peneidos. El nuevo virus del camarón presenta el tamaño y la organización del genoma típicos de la familia, pero muestra una característica adicional, a saber, la presencia del NLS. No se han descrito NLS para Solinviviridae y esta es la primera evidencia de localización nuclear para este grupo de virus confirmada por hibridación in situ. El tropismo tisular del PvSV (réplicas en el hepatopáncreas) es similar al de otros solinvivirus que infectan el epitelio del intestino medio de los insectos [12,13,30] pero difiere en el hecho de que afecta el tejido muscular. La infección por PvSV es entérica y sistémica como el virus Solenopsis invicta 3 que afecta a todos los tejidos del huésped [12].
La afinidad del PvSV por el hepatopáncreas y su efecto sobre los tejidos hace que este órgano sea más susceptible a las infecciones bacterianas crónicas. El tropismo tisular del PvSV es casi idéntico al virus 8 del camarón Wenzhou detectado en Tailandia [27]. Debido a los signos que muestra el camarón infectado que puede estar asociado con una etiología bacteriana, los productores brasileños generalmente sospechan y analizan otros patógenos que causan enfermedades entéricas como la hepatopancreatitis necrotizante debido a Hepatobacter penaeid, la enfermedad de necrosis hepatopancreática aguda (VpAHPND), microsporidiosis hepatopancreática causada por Enterocytozoon hepatopenaei (EHP) y necrosis hepatopancreática séptica debida a vibriosis (SHPN). Sin embargo, está claro a partir de los resultados de este estudio y de estudios previos nuestros [9] que la coinfección de PvSV e IMNV es una causa probable de las mortalidades inusuales que actualmente afectan al cultivo de camarón
en Brasil. Se requieren más estudios para demostrar el postulado de Rivers para determinar la infectividad de PvSV y evaluar su papel en las mortalidades inusuales que ocurren en las granjas camaroneras en Brasil.
El PvSV está ampliamente distribuido en Brasil, afectando a siete estados de la región noreste y ha sido detectado tanto en juveniles como en postlarvas. Es posible que el PvSV tenga una distribución mucho más amplia en América Latina y se necesita estudios de detección para determinar su prevalencia y sus posibles interacciones con otros patógenos del camarón. Coincidentemente, la distribución de este patógeno abarca un área donde se encuentran concentradas la mayoría de la unidades de producción de camarón en Brasi.
Las áreas nacionales de producción de camarón de Brasil, y esta región actualmente está experimentando un efecto negativo en la producción. Es tentador especular que un efecto acumulativo de una infección dual de IMNV y PvSV está contribuyendo a esta pérdida. Las metodologías de PCRq e ISH descritasLas metodologías de PCRq e ISH descritas aquí permitirán la prueba de reproductores y postlarvas para limitar la propagación del patógeno y ayudarán a mitigar sus efectos negativos en la producción de camarón en Brasil y otras naciones camaroneras. Además, la caracterización genética y la asignación taxonómica del PvSV arroja luz sobre la diversidad genética y el rango de huéspedes de la familia Solinviviridae•
Materiales complementarios: Los siguientes están disponibles en línea en https://www.mdpi.com/article/10.3 390/ v14102220/s1
La data de secuencia de nucleótidos sin procesar reportada en este manuscrito está disponible en la base de datos del archivo de lectura de secuencias (SRA) del NCBI con el ID de BioProject: PRJNA871275, SubmissionID (SUB11953069) (https:// www.ncbi.nlm.nih.gov/bioproject/871275 (consultado el 20 de septiembre de 2022)).
La traducción de este artículo al español estuvo a cargo de la Revista Aquacultura de Ecuador.
Pathology and Diagnostic Procedures for Diseases of Cultured Penaeid Shrimp, 1st ed.;World Aquaculture Society: Baton Rouge, LA, USA, 1996.
24. Cruz-Flores, R.; Mai, H.N.; Noble, B.L.; Schofield, P.J.; Dhar, A.K. Detection of Enterocytozoon hepatopenaei using an invasive but non-lethal sampling method in shrimp (Penaeus vannamei). J. Microbiol. Methods 2019, 162, 38–41. [CrossRef] [PubMed]
25. Senapin, S.; Phewsaiya, K.; Briggs, M.; Flegel, T.W. Outbreaks of infectious myonecrosis virus (IMNV) in Indonesia confirmed by genome sequencing and use of an alternative RT-PCR detection method. Aquaculture 2007, 266, 32–38. [CrossRef]
26. Longshaw, M.; Bateman, K.S.; Stebbing, P.; Stentiford, G.D.; Hockley, F.A. Disease risks associated with the importation and release of non-native crayfish species into mainland Britain. Aquat. Biol. 2012, 16, 1–15. [CrossRef]
27. Srisala, J.; Thaiue, D.; Saguanrut, P. Wenzhou shrimp virus 8 (WZV8) diagnosis by unique inclusions in shrimp hepatopancreatic E-cells and by RT-PCR. bioRxiv 2022, 8, 1–20.
28. Liu, S.; Xu, T.;Wang, C.; Jia, T.; Zhang, Q. A Novel Picornavirus Discovered in White Leg Shrimp Penaeus vannamei. Viruses 2021, 13, 2381. [CrossRef]
29. Huerlimann, R.;Wade, N.M.; Gordon, L.; Montenegro, J.D.; Goodall, J.; McWilliam, S.; Tinning, M.; Siemering, K.; Giardina, E.; Donovan, D.; et al. De novo assembly, characterization, functional annotation and expression patterns of the black tiger shrimp (Penaeus monodon) transcriptome. Sci. Rep. 2018, 8, 13553. [CrossRef]
30. Yang, Y.T.; Nai, Y.S.; Lee, S.J.; Lee, M.R.; Kim, S.; Kim, J.S. A novel picorna-like virus, Riptortus pedestris virus-1 (RiPV-1), found in the bean bug, R. pedestris, after fungal infection. J. Invertebr. Pathol. 2016, 141, 57–65. [CrossRef]
31. Vinjé, J.; Estes, M.K.; Esteves, P.; Green, K.Y.; Katayama, K.; Knowles, N.J.; L’Homme, Y.; Martella, V.; Vennema, H.; White, P.A. Ictv Report Consortium ICTV Virus Taxonomy Profile: Caliciviridae. J. Gen. Virol. 2019, 100, 1469–1470. [CrossRef]
32. Brown, K.; Olendraite, I.; Valles, S.M.; Firth, A.E.; Chen, Y.; Guérin, D.M.A.; Hashimoto, Y.; Herrero, S.; de Miranda, J.R.; Ryabov, E. Solinviviridae. Available online: https://ictv.global/report/chapter/ solinviviridae/solinviviridae (accessed on 20 September 2022).
33. Valles, S.M.; Porter, S.D.; Firth, A.E. Solenopsis invicta virus 3: Pathogenesis and stage specificity in red imported fire ants. Virology 2014, 461, 66–71. [CrossRef]
- FEBRERO 2023 35 PATOLOGÍA


Incremento en el desempeño del crecimiento y mecanismos de digestión del camarón a través de un potenciador de la digestión con base en lisofosfolípídos bajo diferentes niveles de colesterol en la dieta
Autores:
Yu-Hung Lin
Waldo G. Nuez-Ortín Gilberto Hernández-González
Professor Yu-Hung Lin is with the Department of Aquaculture, National Pingtung University of Science and Technology, Taiwan.
Waldo G. Nuez-Ortín, PhD is Lead Scientist Aquaculture, Adisseo, Belgium.
Gilberto Hernández-González, MSc is Regional Manager Aquaculture, Adisseo, Latam.
gilberto.hernandez@adisseo.com
Durante muchos años, la industria de la acuicultura ha buscado mejorar la sostenibilidad mediante la reducción del uso de harina de pescado en los alimentos acuícolas. La harina de soya es una alternativa ampliamente utilizada a la harina de pescado en la alimentación de camarones debido a su composición nutricional bien equilibrada y su fácil disponibilidad. Sin embargo, el uso de harina de soya en la alimentación de camarones debe considerarse cuidadosamente debido a su deficiencia de algunos nutrientes, especialmente colesterol. El colesterol es un requerimiento dietético crítico para los camarones que carecen de la capacidad de sintetizar este nutriente de novo (NRC, 2011). Se informa que los requerimientos de colesterol para el camarón blanco son del
0,11 al 0,14 % (NRC, 2011), mientras que los niveles de colesterol en las formulaciones estándar de alimentos para camarones actualmente oscilan entre el 0,05 % y el 0,1 %. En un estudio por Lin et al. 2017, los niveles de inclusión de harina de soya disminuyeron las concentraciones de colesterol en el hepatopáncreas y la hemolinfa de los camarones. Tal hipocolesterolemia se atribuye tanto a la deficiencia de colesterol en la harina de soya como a la mala utilización del colesterol por parte de los camarones.
Los camarones carecen de un sistema de jugos biliares, por lo que el proceso de digestión de los lípidos no es tan eficiente como en los peces. Los potenciadores de la digestibilidad basados en emulsionantes naturales han demostrado ser capaces de

38 - FEBRERO 2023 NUTRICIÓN
complementar el proceso de emulsificación y optimizar la digestión y utilización de importantes nutrientes lipídicos como el colesterol.
En la actualidad, factores como el incremento en los costos de energéticos, materias primas y problemas con las cadenas de suministro han ejercido una fuerte presión sobre los costos de producción en los piensos para camarón. Por eso, encontrar ingredientes alternativos con capacidades funcionales se ha vuelto una prioridad en la estrategia de los productores de piensos. Uno de estos ingredientes funcionales es un potenciador digestivo (Adisseo) a base de lisofosfolípidos producido por la hidrólisis controlada de fosfolípidos en lecitina de soya. La lecitina facilita la digestión y absorción de los lípidos y es ampliamente usada en los alimentos para camarón. Sin embargo, los lisofosfolípidos tienen propiedades emulsionantes superiores en relación con la lecitina, por lo que una aplicación cada vez más común hoy en día en los alimentos para camarones es el reemplazo de la lecitina para reducir el costo de la formulación y mantener el rendimiento del crecimiento.
Se realizó una prueba de crecimiento para investigar los beneficios de lisofosfolípidos (Adisseo) en las formulaciones actuales de alimentos para camarones que contienen
concentraciones altas y bajas de colesterol hoy en día. Los niveles dietéticos de harina de pescado y harina de soya se ajustaron para proporcionar concentraciones de colesterol altas (0,12%) y bajas (0,05%) en los alimentos experimentales.
Diseño experimental
Se formularon y se produjeron en peletizadora cuatro alimentos experimentales isoproteicas e isolipídicos, mismos que se presentan en la Tabla 1. Se diseñaron dos alimentos de control que contenían 1% de lecitina, con concentraciones altas o bajas de colesterol: dieta alta en colesterol (HIGH CHOL, 0.12%) y dieta baja en colesterol (LOW CHOL, 0,05%). Los lisofosfolípidos (AQL) se suplementaron al 0,1% en ambas formulaciones: HIGH CHOL+0,1 % AQL y LOW CHOL+0,1 % AQL.
Los cuatro alimentos experimentales se asignaron aleatoriamente a 12 tanques (300 l, 3 repeticiones por tratamiento) en un sistema de recirculación cerrado. Se sembraron veinte camarones en cada tanque. El sistema consta de un filtro común, biofiltro, filtro (skimmer) de proteínas y luz ultravioleta para mantener la calidad del agua. La temperatura del agua del sistema de cultivo se controló a 28 ± 1°C. Los
camarones fueron alimentados al 6% de su peso húmedo 4 veces al día a las 07:00, 12:00, 17:00 y 22:00. Los camarones se pesaron una vez cada 2 semanas y se realizó un recambio de agua del 50% al mismo tiempo. Los camarones fueron alimentados con las dietas experimentales durante 8 semanas.
Al final de la prueba de alimentación, los camarones se pesaron a granel para calcular el rendimiento del crecimiento. Después de esto, se recolectaron al azar hepatopáncreas de dos camarones en cada tanque. Se midió la expresión génica de las enzimas digestivas, incluidas la quimotripsina, la tripsina, la amilasa y la lipasa. El diseño de cebadores (primers) y la cuantificación relativa se efectuó siguiendo la descripción de Castro-Ruiz et al. (2021) y Livak y Schmittgen (2001).
Se evaluó la normalidad y la homogeneidad de la varianza de los datos mediante la prueba de Kolmogorov-Smirnov y la prueba de Bartlett, respectivamente. Los resultados se analizaron mediante un análisis de varianza de una vía (ANOVA). Cuando el ANOVA identificó diferencias entre los grupos, se realizaron comparaciones múltiples

39 - FEBRERO 2023 NUTRICIÓN
entre las medias utilizando la prueba de rango múltiple de Duncan. La significación estadística se determinó ajustando el error de tipo I agregado a p<0,05.
La suplementación refuerza el desempeño en crecimiento
La suplementación del potenciador digestivo a base de lisofosfolípidos impulsó la ingesta de alimento y el rendimiento del crecimiento de los camarones alimentados con alimentos altos y bajos en colesterol (Figura 1). El consumo de alimento mejoró significativamente en un 13 y un 21 % con la suplementación en los alimentos altos y bajos en colesterol, respectivamente. Los mejores efectos de crecimiento se encontraron en el alimento alto en colesterol suplementado con lisofosfolípidos, mostrando mejoras significativas del 16% y 11% en la ganancia de peso y la tasa de crecimiento específica (SGR), respectivamente. Se observó un patrón similar en el alimento bajo en colesterol; con la suplementación las mejoras numéricas fueron del 10% y 8% en ganancia de peso y SGR, respectivamente. Más interesante aún, la suplementación en el alimento bajo en colesterol igualó el rendimiento del alimento alto en colesterol, demostrando la eficacia de este potenciador digestivo a base de lisofosfolípidos para reducir el costo de la formulación y mantener el rendimiento de un alimento de mejor calidad. La suplementación con aditivos no afectó significativamente la eficiencia de conversión alimenticia de las dietas que contenían niveles altos y bajos de colesterol. Ni los niveles de colesterol ni la suplementación aditiva afectaron la supervivencia, que promedió el 76% en todos los tratamientos.
Hemos demostrado previamente que 0,1% de lisofosfolípidos puede reemplazar con éxito la lecitina al 0,75-1% de un alimento de control que contiene lecitina al 2% (Lin et al. 2021). En el presente estudio, se agregó una cantidad promedio de lecitina de soya al 1% a todas las dietas para satisfacer el requerimiento mínimo de fosfolípidos para los camarones (NRC, 2011). Bajo tal estrategia de formulación, el rendimiento de crecimiento similar entre el alimento rico en colesterol y el alimento bajo en colesterol suplementado con lisofosfolípidos se puede atribuir al efecto digestivo aditivo para mejorar la emulsificación y promover
Tabla 1. Formulación y composición de las dietas experimentales.
*Incluye 15% de pasta fermentada de soya, 8% almidón de maíz, 10% alpha-almidón, 7% harina de hígado de calamar, 1.5% cloruro de colina, 1% premezcla vitaminas y 2% premezcla mineral.
(AQL) sobre el consumo de alimento (A), la ganancia de peso (B) y la tasa de crecimiento específica (C) de camarones blancos alimentados con dietas con colesterol alto (HIGH CHOL) o colesterol bajo (LOW CHOL). Las barras con letras diferentes indican diferencias significativas (p<0,05).


Figura 2. Efectos de los lisofosfolípidos de la dieta (Aqualyso®, AQL) sobre la expresión génica relativa de quimotripsina (A), tripsina (B), amilasa (C) y lipasa (D) de camarones blancos alimentados con dietas con colesterol alto (HIGH CHOL) o colesterol bajo (CHOL BAJA). Las barras con letras diferentes indican que son significativamente diferentes (p<0,05) entre sí.

- FEBRERO 2023 40 NUTRICIÓN Ingredientes (%) Harina de pescado 20 20 7 7 Harina de Soya 25 25 43.5 43.5 Aceite de pescado 3 3 1 1 Aceite de soya (refinado) 0 0 3 3 Lecitina de soya 1 1 1 1 Aqualyso® 0 0.1 0 0.1 Alpha-celulosa 6.5 6.0 3.0 2.5 Otros* 29.9 29.9 29.9 29.9 Composición (%) Humedad 9.32 11.04 9.96 11.01 Ceniza 7.62 6.40 7.57 6.72 Proteína cruda 37.15 36.68 37.15 36.85 Grasa cruda 7.64 7.90 7.74 8.12 Cholesterol 0.118 0.052 0.122 0.053 HIGH CHOL LOW CHOL HIGH CHOL+ 0.1% AQL LOW CHOL+ 0.1% AQL
Figura 1. Efectos de los lisofosfolípidos de la dieta
una absorción y utilización más eficiente del colesterol en alimentos con 1% de lecitina.

Estudios previos en especies de peces, como el rodaballo y el bagre de canal, también han reportado un efecto positivo de la suplementación con lisofosfolípidos que se cree que está relacionado con una mejor absorción y utilización de los nutrientes lipídicos esenciales (Li et al., 2019; Liu et al., 2019). En salmón, datos no publicados más recientes muestran que lisofosfolipidos aceleran la absorción y el transporte de nutrientes en el intestino, así como el procesamiento de nutrientes en el hígado.
Influencia en la actividad enzimática digestiva
El presente estudio también mostró que los lisofosfolípidos pueden influir potencialmente en la actividad enzimática digestiva. El análisis de la expresión génica de las proteasas, como la quimotripsina y la tripsina, aumentó tanto en los alimentos con colesterol alto como bajo (Figura 2). Sin embargo, solo la suplementación en el alimento alto en
colesterol demostró ser significativa. Se observó un patrón de regulación ascendente similar pero no significativo para la amilasa y la lipasa (Figura 2). Específicamente, la suplementación con lisofosfolípidos indujo numéricamente un cambio de 1,5 veces en la expresión génica de la lipasa en el alimento bajo en colesterol, lo que sugiere una optimización de la actividad de la lipasa debido a la emulsificación mejorada. El patrón regulatorio en la expresión de proteasas, amilasa y lipasa estuvo en línea con la ingesta y un mejor rendimiento observado en los grupos suplementados con lisofosfolípidos, lo que indica que la mejora del rendimiento se puede atribuir a una mayor ingesta de alimento respaldada por los mecanismos digestivos que se mantienen al día con el aumento de las demandas digestivas.
En resumen, demostramos en este estudio que 0,1% de lisofosfolípidos mejora el rendimiento del crecimiento y la expresión génica de enzimas digestivas en camarones alimentados con diferentes niveles de
colesterol. Dada la necesidad de reducir aún más la inclusión de harina de pescado en los alimentos para camarones, junto con el costo creciente de la lecitina, la suplementación con aditivo a base de lisofosfolípidos bajo las estrategias de reemplazo de lecitina parece una estrategia efectiva para promover la sostenibilidad y optimizar el costo y el rendimiento de los alimentos para camarones.
El área de investigación y desarrollo continúa combinando la investigación específica de cada especie sobre las estrategias de aplicación de sus lisofosfolípidos y la experiencia en formulación, al mismo tiempo que brinda servicios relacionados con la formulación y el procesamiento de alimentos•
Para más información sobre este artículo escriba a: gilberto.hernandez@adisseo.com
Bibliografía
Las referencias están disponibles bajo petición.
- FEBRERO 2023 NUTRICIÓN
Incremento de la supervivencia de camarón blanco (Litopenaeus
vannamei) infectado con
el
virus de la mancha blanca y alimentado
dieta suplementada con aceite de coco (Cocos nucifera)
Autores:
Hernández-Sandoval Pedro3ID
Timaná-Morales María1,2ID
Robles-Ravelero Maricruz2ID
Peraza-Gómez Viridiana1,2*ID
Ascencio Felipe4ID
Jiménez-Ruíz Edgar1,5ID
1Programa de Maestría en Ciencias Biológicas Agropecuarias, Universidad Autónoma de Nayarit, Xalisco, Nayarit, México.
2Escuela Nacional de Ingeniería Pesquera, Universidad Autónoma de Nayarit, San Blas, Nayarit, México.
3Unidad Regional Los Mochis de la Universidad Autónoma de Occidente, Departamento de Ciencias Naturales y Exactas, Los Mochis, Sinaloa, México.
4Programa de Acuicultura, Centro de Investigaciones Biológicas del Noroeste, S.C. La Paz, BCS, México.
5Unidad de Tecnología de Alimentos, Secretaría de Investigación y Posgrado, Universidad Autónoma de Nayarit. Tepic, Nayarit, México.
*Autor responsable y para correspondencia: Viridiana Peraza-Gómez, Laboratorio de Biotecnología Molecular Experimental, Escuela Nacional de Ingeniería Pesquera, Universidad Autónoma de Nayarit. Bahía Matanchén, Carretera Los Cocos Km

12. C.P. 63740, San Blas, Nayarit, México.
pedro.hernandez@uadeo.mx mafe280901@gmail.com maricruz_ro@hotmail.com
viridiana.peraza@uan.edu.mx ascencio@cibnor.mx edgar.jimenez@uan.edu.mx
Publicación original: https://abanicoacademico.mx/revistasabanico-version-nueva/index.php/abanico-veterinario).
El cultivo de camarón en los países en desarrollo contribuye a su crecimiento económico y proporciona fuentes de empleo (Patil et al., 2021); dentro de esta actividad, la producción de camarón L. vannamei a nivel mundial (4,966241 t) supera a la de otros camarones peneidos (154,615 t) (FAO, 2018); sin embargo, las enfermedades infecciosas y las diferentes condiciones ambientales han sido un continuo desafío en el cultivo de cualquier especie (FAO, 2018) generando grandes pérdidas económicas e inestabilidad en la industria acuícola (Varela et al., 2017). Las enfermedades virales son las más difíciles de controlar debido a su gran poder de diseminación, sus múltiples vías de infección y el amplio espectro de huéspedes tanto silvestres como los cultivados (Wang et al., 1997). El virus del síndrome de la mancha blanca (WSSV) es uno de los virus del camarón más patógenos que infectan a una amplia gama de crustáceos decápodos en todo el mundo; algunos de estos tienen el papel de vectores o reservorios (Flegel & Alday, 1998; OIE, 2019). Este virus también puede causar mortalidades masivas llegando a casos de hasta el 100% durante un período de tres a diez días después de los primeros signos clínicos de la enfermedad (Marks, 2005).
A pesar de los tratamientos para el manejo y control de la enfermedad y mortalidad causada por la presencia del WSSV, la única alternativa efectiva hasta el momento en la producción de camarón es la implementación de prácticas de bioseguridad o exclusión como la filtración y desinfección (EsparzaLeal et al., 2009). Sin embargo, la investigación científica actualmente está enfocada en mejorar las prácticas de manejo y reducir las condiciones de estrés a través de la evaluación y desarrollo de aditivos inmunoestimulantes, los cuales se han convertido en una estrategia novedosa y prometedora para establecer las bases de la resistencia y medidas profilácticas de L. vannamei contra patógenos (Barracco et al., 2014).
Las sustancias inmunoestimulantes se obtienen a través de fuentes naturales y también por síntesis química en base a la estructura molecular de productos naturales, estas sustancias tienen la capacidad de
- FEBRERO 2023
con una
42
regular o modificar la respuesta inmune; por lo que también se conocen como inmunomoduladores o inmunopotenciadores, pudiendo definirse como un componente natural que regula el sistema inmunológico y aumenta la resistencia del huésped frente a enfermedades causadas por patógenos (Bricknell & Dalmo, 2005).
Niu et al. (2018) mencionan que utilizando la macroalga Porphyra haitanensis (2.51 y 3.14%) como suplemento en alimentos acuícolas, mejora la respuesta inmune de L. vannamei aumentando la resistencia contra virus y bacterias, generando una mejora en la función intestinal y una resistencia al estrés oxidativo.
DebMandal & Mandal (2011) revisan los compuestos bioactivos de C. nucifera en la medicina tradicional india, en términos de promoción de la salud y prevención de enfermedades. Entre estos compuestos bioactivos del coco se encuentran principalmente los ácidos grasos de cadena media, como el ácido láurico, el mirístico y el palmítico, los compuestos fenólicos y la vitamina E (Montoya, 2021). Dado el impacto negativo del síndrome de la mancha blanca en la acuicultura, se destaca que el coco puede presentar actividad antiviral, antibacteriana, antiparasitaria y también tiene un efecto inmunoestimulador en varios modelos animales, incluyendo un potencial efecto antiviral contra el SARS-CoV-2 (DebMandal & Mandal, 2011; Elsbaey et al., 2021; Ángeles-Agdeppa et al., 2021).
En este trabajo se evaluó el aceite de coco (C. nucifera), suplementado en la dieta con potencial antiviral para reducir las mortalidades causadas por el virus de la mancha blanca en cultivo de camarón blanco
L. vannamei
Material y métodos
Consideraciones éticas. Este estudio cumple con la Norma Oficial Mexicana NOM-062ZOO-1999, especificaciones técnicas para la producción, cuidado y uso de animales de laboratorio. L. vannamei no se considera una especie protegida o en peligro de extinción.
Aclimatación del camarón a condiciones de cultivo. Para los bioensayos se colectaron camarones presuntivamente sanos de granjas acuícolas de San Blas, Nayarit, y se
transportaron inmediatamente al Laboratorio Experimental de Biotecnología Molecular (LABME) de la Escuela Nacional de Ingeniería Pesquera (ENIP), en recipientes plásticos de 100 L, con agua de los estanques de cultivo y aireación (= 5.4 ± 0.12 mg/L). La aclimatación del camarón a las condiciones de cultivo se realizó durante 48 h, ajustando la salinidad 2‰/h hasta llegar a 35‰. Se utilizó un tanque plástico con capacidad de 1000 L que contenía 800 L de agua de mar filtrada (20 µm); los organismos se mantuvieron con aireación continua y a su vez, fueron alimentados con una ración diaria correspondiente al 5% de su biomasa total dos veces al día 08:00 y 17:00 h con alimento comercial (AC, 35% proteína).
Elaboración de las dietas experimentales con aceite de coco (C. nucifera): El EVCO utilizado para la elaboración de las dietas experimentales fue sometido a un análisis de perfil de ácidos grasos, según el método propuesto por la AOAC (2012). Para la inclusión de aceite de coco extra virgen (prensado en frío), se pulverizó AC en un procesador de alimentos (Oster® modelo FPSTFP1355); el AC se mezcló con aceite de coco (EVCO) al 2.5% y 5% por 0.5 kg-1 de alimento. Una vez realizada la mezcla, se añadió una solución de grenetina (4%) en 180 mL de agua destilada hasta formar una pasta. A partir de la pasta resultante se elaboró el pellet en una picadora de carne (Torrey modelo 22), equipada con una matriz de 3 mm (3/32”). Los pellets formados se colocaron en bandejas para su deshidratación y se secaron a temperatura ambiente durante 24 h; Luego, el alimento se almacenó a 4°C.
Análisis proximal de dietas experimentales. Se realizó un análisis proximal de AC y AC con 2.5 % EVCO y AC con 5% EVCO. Los análisis realizados fueron humedad con método NMX-F-083-1986, cenizas con método NMX-F-607-NORMEX-2013, lípidos (extracto etéreo) con método NOM-086SSA1-1994 (anexo C normativo, numeral 1), proteína (NX6. 25) con método NMX-F-608NORMEX-2011, fibra cruda con método NMXF-613-NORMEX-2017 y carbohidratos totales con método NOM-051-SCFI/SSA1-2010 (por diferencia de análisis proximal).
Inóculo viral de WSSV y su detección
molecular por PCR. El inóculo viral de WSSV (251018) fue donado por el Centro Interdisciplinario para el Desarrollo Integral Regional (CIIDIR IPN Unidad Sinaloa), y previo a su uso fue analizado mediante PCR simple (Kimura et al., 1996). La detección del WSSV en el inóculo y los organismos se realizó mediante PCR de punto final en dos fases; primero las muestras se analizaron con PCR simple, y las que resultaron negativas se analizaron con PCR anidada con los oligonucleótidos propuestos por Kimura et al. (1996).
Análisis de prevalencia del WSSV. La prevalencia se determinó según el porcentaje de organismos enfermos y/o infectados con WSSV entre el número de organismos analizados según Margolis et al. (1982).
Extracción de hemolinfa y recuento total de hemocitos (THC). La extracción de hemolinfa se realizó antes de la primera ración de alimento (7:00-8:00 a. m.), para evitar diferencias en el ciclo circadiano; la hemolinfa se obtuvo con una jeringa estéril (1 mL) en la arteria abdominal ventral (Fisher et al., 1995). La jeringa fue precargada con 200 µL de solución anticoagulante isotónica SIC-EDTA (450 mM NaCl, 10 mM KCl, 10 mM EDTA-Na2, 10 mM HEPES, pH 7.3 y 850 mOsm kg-1) pre-enfriada (4°C) para camarón (Vargas-Albores et al., 1993), depositados en tubos estériles de 1.5 mL en hielo. Se tomaron 50 µL de la dilución 2:1 de SIC-EDTAhemolinfa y se mezclaron con 150 µL (1:3 v/v) de una solución de formaldehído al 4% pre-enfriada para fijar los hemocitos. A partir de esta dilución se realizó el conteo individual utilizando una cámara Neubauer con retícula de 0.01 mm bajo un microscopio binocular (Leica DM300). La respuesta inmune de los camarones se evaluó mediante conteo de hemocitos totales según Cabrera-Pérez et al. (2019):
donde: Hc es el número total de hemocitos contados; D es el factor de dilución de hemolinfa y C es el factor de conversión x 0.1 mm3 x mL (1000).
Bioensayo I. Evaluación de la concentración de aceite de C. nucifera como inmunoestimulante. La duración de
43 - FEBRERO 2023 NUTRICIÓN
este bioensayo fue de 20 días; se realizó en tanques plásticos de 200 L de capacidad, conteniendo 180 L de agua de mar (35 ‰) filtrada (20 µm) y aireación constante. El bioensayo constó de tres tratamientos cada uno por triplicado (30 camarones por tanque con un peso inicial promedio de 5.46 ± 0.19 g): I) control AC; II) AC + 2.5 % EVCO; III) AC + 5% EVCO. Los organismos experimentales fueron alimentados dos veces al día (8:00 a.m. y 17:00 horas). Durante el bioensayo se monitorearon variables fisicoquímicas dos veces al día: se midió pH con un potenciómetro Hanna Hi98130, salinidad con un refractómetro ATAGO modelo 2491, temperatura y oxígeno disuelto con un multiparamétrico YSI pro20i.
Se determinó amonio, nitrito y nitrato cada 10 días con un fotómetro YSI 9500. Al final de cada bioensayo, la supervivencia se determinó mediante la ecuación:
triplicado (10 camarones por acuario con un peso promedio de 7.7 ± 0.25 g): I) CF + WSSV; II) CF + 2.5% EVCO + WSSV; III) Control CF; IV) Control CF + 2.5% EVCO. Los organismos experimentales fueron alimentados dos veces al día (8:00 a.m. y 17:00 p.m.). Durante el bioensayo, las variables fisicoquímicas se monitorearon dos veces al día de la misma manera que en la prueba bienal I.
Al final de cada bioensayo se determinó la supervivencia según Escobar-Gil et al. (2017) y EGR según Ziaei-Nejad et al. (2006). Antes de la infección, se realizó un análisis inicial de prevalencia del WSSV en los camarones de la granja, que fue del 100 % (determinado por PCR anidada). El día 23, los camarones en los tratamientos I y II fueron re infectados con 10 µL de inóculo de camarones previamente infectados con WSSV, el cual se inyectó en la parte dorsal del segundo segmento abdominal con una jeringa de insulina de 0.3 Ml; además, los organismos de estos tratamientos (I y II) fueron alimentados con pasta de camarones infectados con WSSV.
(StatSoft 2003). Los valores de p < 0.05 se consideraron significativamente diferentes. La data de supervivencia y prevalencia se transformaron mediante una función arcoseno según Font et al. (2007).
Resultados
Perfil de ácidos grasos del aceite de coco (C. nucifera). Se identificó un total de 36 ácidos grasos, de los cuales 12 presentaron valores porcentuales (g/100 g grasa) (Tabla 1). El resto de los ácidos grasos presentaron valores < 0.01 por debajo del límite mínimo de calibración detectable. Dentro de esta determinación el 94.52% corresponde a grasas saturadas, el 4.90% a grasas monoinsaturadas y el 0.83% a grasas poliinsaturadas.
donde N° ind f es el número de individuos vivos al final del bioensayo y N° ind i es el número de individuos vivos al inicio del bioensayo (Escobar-Gil et al., 2017). La tasa de crecimiento específica (SGR) se determinó mediante la ecuación:
donde: W2 es el peso final, W1 es el peso inicial y t es el número de días de cultivo (Ziaei-Nejad et al., 2006).
La respuesta inmune de los camarones se evaluó mediante el conteo total de hemocitos según lo descrito por CabreraPérez et al. (2019). Se analizaron organismos individuales (27) en el THC, 9 organismos por tratamiento representando el 30% de la población total de cada tratamiento.
Bioensayo II. Evaluación de la actividad antiviral del aceite de coco suplementado en la dieta de L. vannamei infectados con WSSV. La duración del bioensayo fue de 27 días, donde el día 23 los camarones fueron infectados con el inóculo de WSSV. Se llevó a cabo en acuarios con capacidad de 80 L, los cuales se llenaron con 50 L de agua de mar (35‰) filtrada (20 µm). El bioensayo constó de cuatro tratamientos, cada uno por
La prevalencia del WSSV en camarones se cuantificó como se describió anteriormente (Margolis et al., 1982). Para determinar la prevalencia del WSSV, los camarones infectados se mantuvieron bajo observación hasta que mostraron signos evidentes de la enfermedad de la mancha blanca. Treinta organismos por tratamiento fueron testeados individualmente; además, se recolectaron y sacrificaron organismos moribundos para verificar la presencia del WSSV con PCR simple y anidada; se almacenaron a -80°C. La respuesta inmune de los camarones también se evaluó mediante el recuento total de hemocitos (Cabrera-Pérez et al., 2019). Se analizaron individualmente tres organismos por réplica para THC, 9 organismos por tratamiento que representan el 30% de la población total de cada tratamiento.
Análisis estadístico. Se realizaron pruebas de normalidad (Kolmogorov Smirnov) y homocedasticidad (Bartlett), análisis de varianza unidireccional (ANOVA) y prueba de comparaciones múltiples de Tukey (HSD) para identificar la naturaleza de estas diferencias en supervivencia, tasa de crecimiento específico, recuento de hemocitos totales y prevalencia del WSSV; utilizando el programa STATISTICA Versión 6
Composición químico-proximal de las dietas experimentales. Los resultados del análisis proximal del tratamiento I (CF), tratamiento II (CF + 2.5% EVCO) y tratamiento III (CF + 5% EVCO) se presentan en la Tabla 2. El porcentaje de humedad fue menor en el tratamiento III con un valor de 10.91%, seguido del tratamiento II con un valor de 12.30% y el tratamiento I con 12.37%. El porcentaje de cenizas fue de 8.12%, 8.34% y 8.52% para el tratamiento I, III y II, respectivamente. En cuanto a los lípidos, se obtuvo un porcentaje de 5.26% para el tratamiento I, seguido de 7.64% para el tratamiento II y 10.09% para el tratamiento III. El porcentaje de proteína fue de 32.89%, 34.53% y 35.28%, correspondientes al tratamiento III, II y I, respectivamente. El porcentaje de fibra cruda fue de 4.53% para el tratamiento II, 6.86% para el tratamiento III y 9.67% para el tratamiento I; mientras que para carbohidratos totales el porcentaje más bajo fue para el tratamiento II con 37.01%, seguido del tratamiento III con 37.77% y el tratamiento I con 38.97%. Los porcentajes de proteína, lípidos y carbohidratos con respecto a la dieta total en cada uno de los tratamientos estuvieron dentro de los requerimientos nutricionales recomendados para L. vannamei según Martínez-Córdova et al. (2014)
Bioensayo I.
Parámetros fisicoquímicos. Los parámetros fisicoquímicos evaluados se mantuvieron dentro de los rangos óptimos para el cultivo de L. vannamei durante los 20 días del
- FEBRERO 2023 44 NUTRICIÓN
Tabla 1. Perfil de ácidos grasos del aceite de coco puro
Nombre Común Tipo de Grasa Concentración de Grasa
Ácido lignocérico
Ácido araquídico
Ácido caproico
Ácido esteárico
Ácido cáprico
Ácido palmítico
Ácido caprílico
Ácido mirístico
Ácido laurico
Ácido butírico, ácido pentadecanoico, ácido pentadecanoico, ácido heptadecanoico, ácido heneicosanoico, ácido behénico, ácido tricosanoico
cis-11- Ácido eicosenoico
Ácido oleico (grupo omega 9)
Ácido miristoleico, ácido cis-10pentadecenoico, ácido palmitoleico, ácido cis-10-heptadecenoico, ácido nervónico (grupo Omega 9), ácido erúcico (grupo Omega 9)
Ácido linoleico (Grupo Omega 6)
g- ácido linolénico (grupo Omega 6), ácido linolénico (grupo Omega 3), cis4,7,10,13,16,19- ácido docosahexaenoico (grupo Omega 3), cis- 5,8,11,14, 17- ácido eicosapentaenoico (Grupo Omega 3), cis-11, 14- ácido eicosadienoico, cis-8,11,14- ácido eicosatrienoico (Grupo Omega 6), ácido cis11-14-17-’eicosatrienoico (Grupo Omega 3), ácido araquidónico (Omega Grupo 6), ácido cis-13,16- docosadienoico (Omega Grupo 6) ácido elaídico (TFA C18:1), ácido linolelaídico (TFA C18:2), TFA C18:3
*Por debajo del nivel de calibración más
bioensayo, según Brock & Main (1994).
Calidad del agua (amonio, nitrito y nitrato). Durante el cultivo, las concentraciones de compuestos nitrogenados estuvieron dentro del rango óptimo para el cultivo de camarón blanco, según Boyd (2001) y SENASICA (2003).
Supervivencia y tasa de crecimiento específica (SGR). No se encontraron diferencias significativas en la supervivencia
entre tratamientos (p = 0.256); sin embargo, la supervivencia (Fig. 1) en el tratamiento I fue de 100%, mientras que en los tratamientos II y III con 2.5% y 5% de EVCO se obtuvieron valores de 93.33% y 98.89%, respectivamente.
El análisis estadístico no mostró diferencias significativas en EGR (% día-1), entre tratamientos (p = 0.668); sin embargo, el EGR fue mayor en el tratamiento I con 1.44 ± 0.32 (%día-1), con respecto al tratamiento
Bibliografía
AFTABUDDIN S, Siddique MAM, Romkey SS, Shelton WL. 2017. Antibacterial function of herbal extracts on growth, survival and immunoprotection in the black tiger shrimp Penaeus monodon. Fish and Shellfish Immunology. 65: 52-58. ISSN:10504648. https://doi.org/10.1016/j.fsi.2017.03.050
AKINNUGA AM, Jeje SO, Bamidele O, Amaku EE, Otogo FO, Sunday VE. 2014. Virgin Coconut Oil: Remedial Effects on Renal Dysfunction in Diabetic Rats. Physiology Journal. 1-5. ISSN: 14697793. https://doi.org/10.1155/2014/495926
ANGELES-AGDEPPA I, Nacis JS, Capanzana MV, Dayrit FM, Tanda KV. 2021. Virgin Coconut oil is effective in lowering C-reactive protein levels among suspect and probable cases of COVID-19. Journal of functional foods. (83). 104557. ISSN: 17564646. https://doi.org/10.1016/j.jff.2021.104557
APRAKU A, Liu L, Leng X, Rupia EJ, Ayisi CL. 2017. Evaluation of blended virgin Coconut oil and fish oil on growth performance and resistance to Streptococcus iniae challenge of Nile tilapia (Oreochromis niloticus). Egyptian Journal of Basic and Applied Sciences. 4 (3) 175-184. ISSN:2313808X. https://doi.org/10.1016/j.ejbas.2017.06.002
BARRACCO MA, Perazzolo LM, Rosa RD. 2014. Avances en la Inmunología del Camarón. In: Morales V, Cuéllar-Anjel J. (Eds.). Guía Técnica Patología e Inmunología de Camarones Penaeidos. OIRSA, Panamá, Rep. de Panamá. Pp. 237306. ISBN: 978-9962-661-02-3. https://utm. edu.ec/fcv/acuicultura/images/acuicultura/ pdf_revistas/Guia_Tecnica_Patologia_ Inmunologia_de_Camarones_Penaeidos.pdf BERGER C. 2019. Aportes de la bio-tecnología a la alimentación y a la inmunoestimulación de camarones peneidos. Avances En Nutrición Acuícola. Pp. 19-22. https://nutricionacuicola. uanl.mx/index.php/acu/article/view/268/266 BOYD CE. 2001. Consideraciones Sobre La Calidad Del Agua y Suelo en Cultivos de Camarón En Haws, MC, Boy CE (eds). Métodos para mejorar la camaronicultura en Centroamérica. Editorial-Imprenta UAC, Managua, Nicaragua. 1ra Edición. Pp 1-30. BRICKNELL I, Dalmo RA. 2005. The use of immunostimulants in fish larval aquaculture. F_i_s_h_ _a_n_d_ _s_h_e_l_l_f_i_s_h_ _i_m_m_u_n_o_l_o_g_y_. 19(5): 457-472. ISSN: 1050-4648. https://doi.org/10.1016/j.fsi.2005.03.008 BROCK JA, Main, KA. 1994. Guide to the common problems and diseases of cultured P_e_n_a_e_u_s_ _v_a_n_n_a_m_e_i_. The Oceanic Institute, Honolulu, EUA. Pp 242. ISBN: 1886608008. CABRERA E, Marcelo Z, Reyes W, Azañero C. 2019. Efecto de dietas con alta concentración de S_a_c_c_h_a_r_o_m_y_c_e_s_ _c_e_r_e_v_i_s_i_a_e_ _sobre la proliferación de hemocitos en camarones C_r_y_p_h_i_o_p_s_ _c_a_e_m_e_n_t_a_r_i_u_s_ _machos. R_e_v_i_s_t_a_ _d_e_ _I_n_v_e_s_t_i_g_a_c_i_o_n_e_s_ _V_e_t_e_r_i_n_a_r_i_a_s_ _d_e_l_ _P_e_r_ú. 30 (3): 1057-1067. ISSN: 1609-9117. http:// dx.doi.org/10.15381/rivep.v30i3.16733 CAHU C, Guillaume JC, Stéphan G, Chim L. 1994. Influence of phospholipid and highly unsaturated fatty acids on spawning rate and egg and tissue composition in P_e_n_a_e_u_s_ _v_a_n_n_a_m_e_i_ _fed semi-purified diets. A_q_u_a_c_u_l_t_u_r_e_. 126(1-2): 159-170. ISSN: 0044-8486. https:// doi.org/10.1016/0044-8486(94)90257-7 DAYRIT CS. 2000. Coconut oil in health and disease: its and monolaurin's potential as cure for HIV/ AIDS. 31(4): 19-24. https://doi.org/0.1.1.554.2770 DEBMANDAL M, Mandal S. 2011. Coconut (C_o_c_o_s_ _n_u_c_i_f_e_r_a_ _L.: Arecaceae): in health promotion and disease prevention.
A_s_i_a_n_ _P_a_c_i_f_i_c_ _j_o_u_r_n_a_l_ _o_f_ _t_r_o_p_i_c_a_l_ _m_e_d_i_c_i_n_e_._ _4(3):241-247. ISSN:1995-7645. https:// doi.org/10.1016/S1995-7645(11)60078-3 ELSBAEY M, Ibrahim MA, Bar FA, Elgazar AA. 2021. Chemical constituents from coconut waste and their i_n_ _s_i_l_i_c_o_ _evaluation as potential antiviral agents against SARS-CoV-2. S_o_u_t_h_ _A_f_r_i_c_a_n_ _J_o_u_r_n_a_l_ _o_f_
_B_o_t_a_n_y_. (141): 278-289. ISSN: 0254-6299. https://doi.org/10.1016/j.sajb.2021.05.018 ESCOBAR GC, Pachamoro LM, Reyes AW. 2017. Supervivencia y crecimiento de machos adultos del camarón de río
45 - FEBRERO 2023 NUTRICIÓN
Saturado Saturado Saturado Saturado Saturado Saturado Saturado Saturado Saturado Saturado Monoinsaturado Monoinsaturado Monoinsaturado Poliinsaturado Poliinsaturado Trans 0.03 0.08 0.77 2.92 6.38 8.03 8.60 17.97 49.47 <0.01* 0.03 4.87 <0.01* 0.83 <0.01* <0.01*
crítico
(g/100g de grasa g/100g
(%)
de grasa)
II con 1.41 ± 0.13 (%día-1) y el tratamiento III con 1.28 ± 0.11 (%día-1) (fig. 2).

Recuento total de hemocitos (THC). El valor promedio del recuento total de hemocitos por mililitro de hemolinfa en el tratamiento I fue de 3.83 x 106 ± 0.52 x 106 células/mL. En los tratamientos II y III fue de 4.11 x 106 ± 0.36 x 106 cel/mL y 3.92 x 106 ± 0.50 x 106 cel/ mL, respectivamente. El análisis de varianza (ANOVA) no mostró diferencias significativas (p = 0.449) entre tratamientos (Fig. 3).
Con base en los resultados obtenidos, aunque no se encontraron diferencias significativas entre los tratamientos EVCO, se seleccionó el tratamiento II (2.5% EVCO) para el siguiente bioensayo, por ser el que menos cantidad de EVCO se utilizó y también los resultados de EGR y THC en este tratamiento fueron más alto que en los otros tratamientos.
Bioensayo II.
Parámetros fisicoquímicos. Los parámetros fisicoquímicos evaluados se mantuvieron dentro de los rangos óptimos para el cultivo de L. vannamei durante los 27 días del bioensayo, según Brock & Main (1994).
Calidad del agua (amonio, nitrito y nitrato). Durante el cultivo, la concentración de compuestos nitrogenados estuvo dentro del rango óptimo para el cultivo de camarón blanco, según Boyd (2001) y SENASICA (2003).
Tasa de crecimiento específica (SGR). La SGR (%día-1) no mostró diferencias significativas (p = 0.644) entre tratamientos (Fig. 4). El tratamiento I presentó 0.60 ± 0.10% día-1, el tratamiento II presentó 0.58 ± 0.08% día-1; mientras que el tratamiento III presentó la mayor tasa de crecimiento específico por día con 0.73 ± 0.29 (%día-1), y el tratamiento IV presentó 0.69 ± 0.10 %día1. Estos resultados indican que EVCO no tuvo efecto sobre la SRG (%día-1).
Supervivencia y prevalencia del WSSV. La supervivencia (Fig. 5) mostró diferencias significativas (p = 0.0030) entre el tratamiento I y el resto de los tratamientos. En el tratamiento I (control-infectado con WSSV) la supervivencia fue menor con 83.3%, seguido del tratamiento II (infectado con WSSV) con 93.3%. En los tratamientos
Composición proximal (% en peso)
Lípidos (extracto etéreo)
Proteínas (N-6.25)
Carbohidratos totales
Tratamientos:
III y IV la supervivencia fue del 100%. Los resultados muestran que a pesar de estar infectados con WSSV, los organismos en el tratamiento II con 2.5% de EVCO mostraron una mayor supervivencia que en el tratamiento I también infectado con WSSV, por lo que podría considerarse un efecto beneficioso de EVCO contra WSSV.
La prevalencia inicial de WSSV de los camarones de la granja fue del 100% (determinada por PCR anidada), y con reinfección solo en los tratamientos I y II, la prevalencia fue del 100% (determinada por PCR simple). Sin embargo, al final del bioensayo se analizó la prevalencia del WSSV de todos los tratamientos y no se encontraron diferencias significativas (p = 0.2192).
La prevalencia final del WSSV en camarones del tratamiento I fue 100% (determinada por PCR simple), en camarones del tratamiento II fue 96.67% (de los cuales 86.21% fueron determinados por PCR simple y 13.79% determinados por PCR anidada), donde
93.33% se infectaron vivos, 3.33% se infectaron muertos y el 3.33% restante no se infectaron. En los camarones del tratamiento III la prevalencia fue del 100% (determinada por PCR anidada) y en los camarones del tratamiento IV fue del 93.33%, (de los cuales el 100% se determinó por PCR anidada), donde el 93.33% estaban infectados vivos y el 6.66% estaban vivos, pero sin la presencia de virus.
Se observó que en el tratamiento II no solo se redujo la prevalencia a 3.33%, sino también la carga viral (determinada por PCR anidada); mientras que para el tratamiento IV la prevalencia disminuyó 6.67% (determinada por PCR anidada). Los resultados muestran que los tratamientos con 2.5% EVCO tuvo un efecto en la disminución de la prevalencia y la carga viral del WSSV (Fig. 5)
Recuento
total de hemocitos (THC).
El análisis estadístico del recuento de hemocitos totales no mostró diferencias significativas (p = 0.413) entre tratamientos.
- FEBRERO 2023 46 NUTRICIÓN
Tratamientos Humedad Ceniza
Tabla 2. Composición químico-proximal de las dietas experimentales
Fibra cruda
12.37 12.30 10.91 8.12 8.52 8.34 5.26 7.64 10.09 35.28 34.53 32.89 9.67 4.53 6.86 38.97 37.01 37.77 I II III
I) CF (35% proteína); II) CF + 2.5% EVCO; III) CF + 5% EVCO
Figura 1. Supervivencia de L. vannamei. Tratamientos: I) Control CF (35% de proteína); II) CF + 2.5% EVCO; III) CF + 5% EVCO.
Figura 2. Tasa de crecimiento específico de L. vannamei. Tratamientos: (I) Control CF (35% proteína); (II) CF + 2.5% EVCO; (III) CF + 5% EVCO. Las barras indican el valor promedio ± desviación estándar y (P) análisis de varianza ANOVA.

C_r_y_p_h_i_o_p_s_ _c_a_e_m_e_n_t_a_r_i_u_s_ _Molina, 1782 (Crustacea, Palaemonidae) expuestos a salinidades. E_c_o_l_o_g_ía_ _A_p_l_i_c_a_d_a_.16 (2): 75-85. ISSN:17262216. http://dx.doi.org/10.21704/rea.v16i2.1010
ESPARZA LHM, Escobedo BCM, Casillas HR, Álvarez RP, Portillo CG, Valerio GR. C, Magallón BFJ. 2009. Detection of white spot syndrome virus in filtered shrimp-farm water fractions and experimental evaluation of its infectivity in Penaeus (Litopenaeus) vannamei. Aquaculture. 292:16-22. ISSN: 0044-8486. https://doi.org/10.1016/j. aquaculture.2009.03.021
ESQUENAZI D, Wigg MD, Miranda MMFS, Rodrigues HM, Tostes JBF, Rozental S, Alviano CS. 2002. Antimicrobial and antiviral activities of polyphenolics from Cocos nucifera Linn. (Palmae) husk fiber extract. Research in Microbiology. 153 (10): 647-652. ISSN: 0944-501. https:// doi.org/10.1016/S0923-2508(02)01377-3 FAO (Organización de las naciones unidad para la alimentación y la agricultura). 2018. El estado mundial de la pesca y la acuicultura. Cumplir los objetivos de desarrollo sostenible. Roma: Organización de las Naciones Unidas. ISBN 978-92-5-1306888. http://www.fao.org/3/i9540es/i9540es.pdf
FIFE B. (2013). El milagro del aceite de coco. Agro Food Industry Hi-Tec. 24(3): 7-10. ISNB: 9788478087136.
FISHER W, Krupp F, Schneider W, Sommer C, Carpenter KE, Niem VH. 1995. Guía FAO para la identificación de especies para los fines de la pesca: Pacífico Centro-Oriental. Roma, Italia. (3). ISBN 92-5-303409-2. https:// decapoda.nhm.org/pdfs/29676/29676.pdf
FLEGEL TW, Alday SV. 1998. The crisis in Asian shrimp aquaculture: current status and future needs. Jornal of Applied Ichthyology. 14 (3-4): 269-273. ISSN:1439-0426. https://doi. org/10.1111/j.1439-0426.1998.tb00654.x
FONT H, Torres V, Herrera M, Rodríguez R. 2007. Cumplimiento de la normalidad y homogeneidad de la varianza en frecuencias de medición acumulada de la variable producción de huevos en gallinas White Leghorn. Revista Cubana de Ciencia Agrícola. 41(3). ISSN: 0034-7485. https:// www.redalyc.org/articulo.oa?id=193017693002 http://www.cesasin.com. mx/CentroAmerica/10%20 Practicas%20de%20manejo.pdf
Figura 3. Recuento total de hemocitos (hemocitos/mL) en L. vannamei. Tratamientos: (I) Control CF (35% proteína); (II) CF + 2.5% EVCO; (III) CF + 5% EVCO. Las barras indican el valor promedio ± desviación estándar y (P) Análisis de varianza ANOVA.

El conteo total de hemocitos por mililitro de hemolinfa fue mayor en el tratamiento III con un promedio de 4.37 x 106 ± 0.52 x 106 cel/ mL, seguido del tratamiento IV con 4.27 x 106 ± 0.46 x 106 cel/mL, tratamiento II con un promedio de 4.10 x 106 ± 0.46 x 106 cel/ mL y tratamiento I con un promedio de 3.99 x 106 ± 0.57 x 106 células/mL (Figura 6).
Discusión
Debido al impacto negativo que generan las enfermedades causadas por diferentes agentes etiológicos como virus y bacterias, así como su resistencia a los antibióticos utilizados para el tratamiento y control de enfermedades infecciosas (Lieberman et al., 2006), surge la necesidad de implementar nuevas estrategias para mejorar la inmunidad de especies de importancia
acuícola y aumentar la resistencia contra virus y bacterias, promoviendo también la resistencia al estrés mediante la suplementación de alimentos acuícolas con aditivos inmunoestimulantes naturales (Berger, 2000; Rendón & Balcázar, 2003).
Según Turchini et al. (2009) y Turchini et al. (2011), una de las principales alternativas que se está aplicando como aditivo dietético son los aceites vegetales, ya que ofrecen una amplia gama de ácidos grasos de cadena media (AGCM) de bajo peso molecular. Según Kapilan & Reddy (2008) y Akinnuga et al. (2014), el aceite de coco se compone principalmente de MCFA; además de una amplia variedad de compuestos fenólicos, que según Nevin & Rajamohan (2009) potencian la actividad de las enzimas
HUYNH TG, Yeh ST, Lin YC, Shyu JF, Chen LL, Chen JC. 2011. White shrimp Litopenaeus vannamei immersed in seawater containing Sargassum hemiphyllum var. chinense powder and its extract showed increased immunity and resistance against Vibrio alginolyticus and white spot syndrome virus. Fish and Shellfish Immunology. 31(2): 286-293. ISSN: 1050-4648. https://doi.org/10.1016/j.fsi.2011.05.014
JIANG YS, Zhan WB, Wang SB, Xing J. 2006. Development of primary shrimp hemocyte cultures of Penaeus chinensis to study white spot syndrome virus (WSSV) infection. Aquaculture. 253: 114-119. ISSN: 0044-8486. https://doi.org/10.1016/j. aquaculture.2005.07.045
KAPILAN N, Reddy RP. 2007. Evaluation of Methyl Esters of Mahua Oil (Madhuca Indica) as Diesel Fuel. Journal of the American Oil Chemists’ Society. 85 (2): 185-188. ISSN: 1558-9331. https://doi.org/10.1007/s11746-007-1179-5
KIMURA T, Yamano K, Nakano H, Momoyama K, Hiraoka M, Inouye K. 1996. Detection of penaeid rod-shaped DNA virus (PRDV) by PCR. Fish Pathology. 31(2): 93-98. ISSN: 18817335. https://doi.org/10.3147/jsfp.31.93
LIEBERMAN S, Enig MG, Preuss HG. 2006. A Review of Monolaurin and Lauric Acid: Natural Virucidal and Bactericidal Agents. Alternative and Complementary Therapies. 12(6): 310-314. ISSN: 1557-9085. https://doi.org/10.1089/act.2006.12.310
LÓPEZ MSG. 2020. Efecto del aceite vegetal (canola) y animal (sardina y salmón) en el alimento sobre la expresión transcripcional del camarón blanco Penaeus vannamei. Tesis de maestría. Centro de Investigaciones Biológicas del Noroeste. La Paz, Baja California Sur, México. Pp 97. https://cibnor.repositorioinstitucional. mx/jspui/handle/1001/1865
LU Y, Jin M, Yuan Y, Xiong J, Ma H, Zhou Q. 2018. Effects of different lipid sources on growth performance, body composition, the serum biochemical indices, fatty acids composition and antioxidant capacity in juvenile yellow catfish
47 - FEBRERO 2023 NUTRICIÓN
antioxidantes y eliminan el exceso de radicales libres en las células. Por otro lado, Lu et al. (2018) señalan que, en el caso del ácido láurico, este estimula el crecimiento y la salud de algunas especies acuáticas como Pelteobagrus fulvidraco. En el análisis del perfil de ácidos grasos de EVCO utilizado en este trabajo, el ácido láurico presentó una concentración de 49.47% (g/100g de grasa), donde la alta concentración sugiere que este aceite podría ser utilizado como tratamiento alternativo al problema actual contra patógenos en el cultivo de camarón, ya que se puede agregar al alimento para combatir el WSSV o disminuir su carga viral.
Según Niu et al. (2018), las sustancias aplicadas en los sistemas de cultivo o alimentación no deben alterar el valor nutricional ni afectar negativamente el desempeño productivo de los organismos. Esto coincide con los resultados obtenidos sobre la composición químico-proximal, donde se demuestra que la adición de EVCO no afectó los valores nutricionales de las dietas experimentales y se encontraron dentro de los valores óptimos para el desarrollo de L. vannamei reportados por Martínez-Córdova et al. Alabama. (2014). Con respecto a EGR, en el Bioensayo 1 y 2 no se observó afectación en camarones cuando se utilizó EVCO en diferentes concentraciones.
En el Bioensayo 1 se observó que a medida que aumentaba la concentración de lípidos (% en peso), la EGR mostraba una tendencia a disminuir. Según Cahu (1994), los niveles óptimos de lípidos recomendados para la alimentación comercial de camarón varían de 6 a 7.5%, sin exceder el 10%; sin embargo, en los resultados de esta investigación se encontraron niveles entre 5.26% (FC), 7.64% (2.5% EVCO) y 10.09% (5% EVCO), a pesar de estar dentro del nivel óptimo de lípidos totales (5- 10%) en una dieta para camarón. Según Martínez-Córdova et al. (2014) y López-Marcos (2020), se podría considerar que los EGR más bajos se deben a un exceso en los niveles de lípidos (Cahu, 1994).
La literatura científica sobre el efecto del aceite de coco en L. vannamei es escasa; sin embargo, los resultados de EGR de este estudio son consistentes con los reportados por Apraku et al. (2017), quienes evaluaron la mezcla de aceite de coco virgen y aceite de
de crecimiento específico de L. vannamei. Tratamientos: TI) CF + WSSV; TII) CF + 2.5% EVCO + WSSV; TIII) Control CF; TIV) Control CF + 2.5% EVCO. Las barras indican el valor promedio ± desviación estándar y (P) Análisis de varianza ANOVA.



Figura 5. Supervivencia (%), prevalencia (%), infectados vivos (%), infectados muertos (%) de L. vannamei. Tratamientos: I) CF + WSSV; II) CF + 2.5% EVCO + WSSV; III) Control CF; IV) Control CF + 2.5% EVCO. Las barras indican el valor promedio ± error estándar y (P) análisis de varianza ANOVA. Las letras indican diferencias significativas entre tratamientos según la prueba de rango múltiple de Tukey.
Figura
Conteo total de hemocitos (hemocitos/mL) en L. vannamei. Tratamientos: (I) CF + WSSV; (II) CF + 2.5% EVCO + WSSV; (III) Control CF; (IV) Control CF + 2.5% EVCO. Las barras indican el valor promedio ± desviación estándar y (P) Análisis de varianza ANOVA.
- FEBRERO 2023 48 NUTRICIÓN
Figura 4. Tasa
6
pescado sobre el crecimiento y la resistencia a Streptococcus iniae en la tilapia del Nilo (Oreochromis niloticus), encontrando que el crecimiento no se vio afectado al sustituir parcial y completamente el aceite de coco virgen por aceite de pescado. Además, estos resultados concuerdan con otros trabajos, donde se han utilizado aceites vegetales, como el de Mozanzadeh et al. (2015); quienes indican que el rendimiento de crecimiento de juveniles de Sparidentex, incluso alimentados con dietas alternativas de aceite vegetal (aceite de canola y girasol), no se vio comprometido al reemplazar parcial o totalmente la dieta de aceite de pescado.
En cuanto a la supervivencia, en el Bioensayo 1 no se encontraron diferencias significativas entre tratamientos; mientras que en el Bioensayo 2, el 100% de los organismos provino de estanques de camaroneras infectados con WSSV (carga viral baja), pero adicionalmente se realizó una reinfección en los tratamientos I y II, resultando con una carga viral alta al final del bioensayo y el menor supervivencia (83.3%) observada en el tratamiento I (CF con WSSV), en comparación con el resto de los tratamientos II (EVCO con WSSV), III (CF) y IV (EVCO).
Estos resultados concuerdan con Raa (1996), quien menciona que el propósito del uso de inmunoestimulantes en el sector acuícola es mejorar la supervivencia e inmunidad de los organismos cultivados. Esto es consistente con el hecho de que cuando se agregó EVCO al alimento para camarón, las tasas de supervivencia de los camarones testeados con carga viral alta y baja (WSSV) fueron significativamente más altas. Esto podría deberse a Fife (2013), quien señala que, al utilizar ácido láurico a través de la ingesta diaria de aceite de coco, se puede proteger y prevenir infecciones provocadas por diferentes virus, como el virus de la inmunodeficiencia humana (VIH), sarampión, Herpesviridae, virus de la influenza, entre otras enfermedades.
Los hemocitos son responsables de la coagulación, el endurecimiento del exoesqueleto y la eliminación de materiales extraños (AftabUddin et al., 2017). En el bioensayo I de este estudio se obtuvo un número mayor en el conteo total de hemocitos en los tratamientos (II y III), que contenían
el aditivo EVCO con respecto al tratamiento I (CF); lo que indica que hubo un aumento en la respuesta inmune de los camarones alimentados con este aditivo. Al respecto, Sequeira et al. (1996) mencionan que la proliferación de hemocitos puede aumentar hasta tres veces cuando los camarones son alimentados con un inmunoestimulante; sin embargo, aunque en el bioensayo II no se encontraron diferencias significativas entre tratamientos, se observó una disminución de THC del tratamiento I y II (infectados con WSSV) con respecto a los tratamientos III y IV. Estos resultados concuerdan con lo informado por Niu et al. (2018), quienes alimentaron camarones con diferentes valores dietéticos inmunoestimulantes de macroalgas, y tras el bioensayo con WSSV, la cantidad de hemocitos se redujo significativamente. La disminución en el recuento total de hemocitos puede atribuirse a la migración de hemocitos al sitio de la infección (Söderhäll et al., 2003) o donde hay daño tisular (Niu et al., 2018).
La enfermedad de la mancha blanca ha causado severas mortalidades en los cultivos de camarón a nivel mundial (Jiang et al., 2006), siendo responsable de grandes pérdidas económicas en esta industria (Sahul et al., 2006), por lo que se hace necesario prevenir o controlar la enfermedad de la mancha blanca (Huynh et al., 2011).
En el desarrollo del bioensayo II se observó que la prevalencia inicial fue de 100% WSSV (determinada por PCR anidada) con baja carga viral, lo que significó que todos los organismos eran de organismos infectados de la granja. Cabe señalar que los tratamientos I y II fueron re infectados con inóculo viral para realizar el experimento y al finalizar el bioensayo se observó que en el tratamiento I la prevalencia final fue del 100% (determinada por PCR simple) con carga viral alta, en el tratamiento III la prevalencia fue del 100% (determinada por PCR anidada); mientras que en los tratamientos EVCO, tratamiento II (2.5% EVCO + WSSV) y tratamiento IV (2.5% EVCO). La prevalencia disminuyó a 96.67% y 93.33%, respectivamente; sin embargo, se observó que en el tratamiento II no solo disminuyó la prevalencia del virus (96,67 %), sino también la carga viral; de los cuales el 86.21% se determinó por PCR
(Pelteobagrus fulvidraco). Journal of Fisheries of China. 42 (7): 1094-1110. ISSN:1000-0615. https://doi.org/10.11964/jfc.20170310739
MARGOLIS L, Esch GW, Holmes JC, Kuris AM, Schad GA 1982. The use of ecological terms in parasitology (report of an ad hoc committee of the American Society of Parasitologists). Journal of Parasitology. 68 (1): 131-133 ISSN: 0022-3395. https://doi.org/10.2307/3281335
MARKS H. 2005. Genomics and transcriptomics of white spot syndrome virus. Wageningen University. Promotor(en): Just Vlak; R.W. Goldbach, co-promotor (en): M.C.W. van Hulten. Wageningen. Pp 160. ISBN: 90-8504-318-2. https://library.wur.nl/WebQuery/ wurpubs/fulltext/121730
MARTÍNEZ CL, Martínez PM, López EJA, Enríquez OLF. 2014. Uso de microorganismos en el cultivo de crustáceos. Biotecnia.16 (3): 50-55. ISSN: 1665-1456. https://doi.org/10.18633/bt.v16i3.141
MONTOYA Ballesteros LC. 2021. Compuestos bioactivos en coco (Cocos nucifera L.). Efecto del cultivar y región de cultivo. Biotecnia. 23(2):22-29. ISSN:1665-1456. https:// doi.org/10.18633/biotecnia.v23i2.1336
MOZANZADEH MT, Marammazi JG, Yavari V, Agh N, Mohammadian T, Gisbert E. 2015. Dietary n3 LC-PUFA requirements in silveryblack porgy juveniles (Sparidentex hasta). Aquaculture. ISSN: 0044-8486 448: 151-161. https://doi.org/10.1016/j. aquaculture.2015.06.007
MURRAY M. 1994. Natural alternative to Overthe Counter and prescription Drugs. Nueva York: William Morrow. Pp 357. ISBN: 0-688-12358-9. NEVIN KG, Rajamohan T. 2009. Wet and dry extraction of coconut oil: impact on lipid metabolic and antioxidant status in cholesterol coadministered rats. Canadian journal of physiology and pharmacology. 87 (8): 610-616. ISSN: 1205-7541.
https://doi.org/10.1139/Y09-045
NIU J, Xie SW, Fang HH, Xie JJ, Guo TY, Zhang YM, Liu YJ. 2018. Dietary values of macroalgae Porphyra haitanensis in Litopenaeus vannamei under normal rearing and WSSV challenge conditions: Effect on growth, immune response and intestinal microbiota. Fish and Shellfish Immunology. 81: 135-149. ISSN: 1050-4648. https://doi.org/10.1016/j.fsi.2018.06.010
NOM (Norma Oficial Mexicana NOM-051-SCFI/ SSA1-2010), Especificaciones generales de etiquetado para alimentos y bebidas no alcohólicas preenvasados-Información comercial y sanitaria. https://www.dof.gob.mx/normasOficiales/4010/ seeco11_C/seeco11_C.htm
NOM (Norma Oficial Mexicana, NOM-062ZOO-1999) de especificaciones técnicas para la producción, cuidado y uso de los animales de laboratorio. México. http://dof.gob.mx/nota_ detalle.php?codigo=764738&fecha=18/06/2001
NOM (Norma Oficial Mexicana, NOM-086SSA1-1994). Bienes y Servicios. Alimentos y bebi-das no alcohólicas con modificaciones en su composición. Especificaciones Nutrimentales. Secretaría de Salud. México. https://www.dof.gob.mx/nota_detalle. php?codigo=4890075&fecha=26/06/1996 OIE (Organización mundial de sanidad animal). 2019. Manual de las Pruebas de Diagnóstico para los Animales Acuáticos. En capítulo 2.2.8. Infección por el virus del síndrome de las manchas blancas. Pp 1-16. https://www. oie.int/es/que-hacemos/normas/codigos-ymanuales/acceso-en-linea-al-manual-acuatico/ OYI AR, Onaolapo JA, Obi RC. 2010. Formulation and antimicrobial studies of coconut (Cocos nucifera Linne) oil. Research Journal of Applied Sciences, Engineering and Technology. 2 (2): 133-137. ISSN: 2040-7467. https://www.researchgate. net/publication/285568592
PATIL PK, Geetha R, Ravisankar T, Avunje S, Solanki HG, Abraham TJ, Vijayan KK. 2020. Economic loss due to diseases in Indian shrimp farming with special reference to Enterocytozoon hepatopenaei (EHP) and white spot syndrome virus (WSSV). Aquaculture. (533): 736231. ISSN: 0044-8486. https://doi.org/10.1016/j. aquaculture.2020.736231
RAA J. 1996. The use of immunostimulatory substances in fish and shellfish farming. Reviews in Fisheries Science. 4(3): 229-288. ISSN: 1547-6553. https://doi.org/10.1080/10641269609388587
RENDON L, Balcazar JL. 2003. Inmunología de
49 - FEBRERO 2023 NUTRICIÓN
simple y el 13.79% por PCR anidada. La disminución de la prevalencia y carga viral en los tratamientos con EVCO sugiere que su aplicación como suplemento dietético ejerce actividad antiviral contra WSSV y reduce la carga viral de manera similar a lo reportado por Lieberman et al. (2006) y Fife (2013), señalando que EVCO está compuesto por AGCM tales como: ácido láurico con cerca de 48-50% en su composición, seguido del ácido mirístico con aproximadamente 15% (Fife, 2013; Ruiz et al., 2016), ácido caprílico (8%) (Fife, 2013; Akinnuga et al., 2014) y ácido cáprico (7%) (Fife, 2013).

Estos compuestos han demostrado ser efectivos contra bacterias, hongos y virus (Esquenazi et al., 2002). En el caso de los virus, Oyi et al. (2010) mencionaron que un componente del aceite de coco llamado monocaprina destruyó el VIH y el virus del herpes simplex (VHS) en un minuto. De manera similar, Fife (2013) reportó que los MCFA presentes en el aceite de coco pueden inactivar virus como el VIH, el coronavirus asociado con el síndrome respiratorio agudo, el virus del sarampión, el virus de la rubéola, Herpesviridae, el virus del sarcoma, el virus respiratorio sincitial, el virus linfotrópico humano (tipo 1), virus de la estomatitis vesicular, Visna virus, citomegalovirus humano (CMV), virus de Epstein-Barr (EBV), virus de la influenza, virus de la hepatitis C (HCV) y virus Coxsackie B4. Al respecto, Murray (1994) y Dayrit (2000) registraron una disminución de la carga viral del VIH a
niveles indetectables en personas infectadas con este mismo virus y una marcada mejoría en su salud luego de consumir coco. Por lo tanto, se sugiere que el EVCO compuesto por MCFA como el ácido láurico ofrece la posibilidad de disminuir la carga viral, lo que a su vez puede ser utilizado como una forma de terapia inmunoterapéutica y preventiva en situaciones de infecciones por WSSV.
Conclusiones
De acuerdo con los resultados obtenidos, la adición de EVCO 2.5% en el alimento promovió una disminución en la prevalencia y carga viral de WSSV en juveniles de L. vannamei; no mostró efectos negativos sobre la supervivencia y el THC de L. vannamei Adicionalmente, no hubo inmunosupresión ni reducción de la respuesta inmune, ya que no hubo diferencias significativas en el THC entre los tratamientos de ambos bioensayos. Este trabajo de investigación es el primer reporte sobre el efecto de Cocos nucifera EVCO en la supervivencia del camarón blanco (Litopenaeus vannamei) testeado con WSSV. Por lo que se recomienda realizar estudios sobre el uso de EVCO en alimentos bajo diferentes esquemas de formulación y tiempos de aplicación en la alimentación del camarón. Además, es esencial estudiar más a fondo las respuestas inmunonutricionales de EVCO cuando se incluye en la formulación del alimento•
Para más información sobre este artículo escriba a: pedro.hernandez@uadeo.mx
camarones: Conceptos básicos y recientes avances. Revista AquaTIC. 19:27-33. ISSN: 1578-4541.
http://www.revistaaquatic.com/ojs/ index.php/aquatic/article/view/256/244
RUIZ NB, Dijck BDAJ, Muskiet FAJ. 2016. The relation of saturated fatty acids with low-grade inflammation and cardiovascular disease. The Journal of Nutritional Biochemistry. 36: 1-20. ISSN: 0955-2863. https://doi.org/10.1016/j.jnutbio.2015.12.007
SAHUL HAS, Sarathi M, Sudhakaran R, Balasubramanian G, Syed S. 2006. Quantitative assessment of apoptotic hemocytes in white spot syndrome virus (WSSV)-infected penaeid shrimp, Penaeus monodon and Penaeus indicus, by flow cytometric analysis. Aquaculture. 256(1-4): 111-120. ISSN: 0044-8486. https://doi.org/10.1016/j. aquaculture.2006.02.054
SENASICA (Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria. 2003. Manual de Buenas Prácticas de Producción de Acuícola de Camarón para la Inocuidad Alimentaria. Pp 95. ISBN: 9685384-04-5. https://cesasin.mx/wp-content/ uploads/2017/12/Cam-Manual-de-buenaspracticas-de-produccio%CC%81n-acuicola-decamaro%CC%81n-para-la-inocuidad-alimentaria.pdf
SEQUEIRA T, Tavares D, Arala Chaves M. 1996. Evidence for circulating hemocyte proliferation in the shrimp Penaeus japonicus. Developmental and Comparative Immunology. 20(2): 97-104. ISSN: 0145-305X. https://doi.org/10.1016/0145-305X(96)00001-8
SÖDERHÄLL I, Bangyeekhun E, Mayo S, Söderhäll K. 2003. Hemocyte production and maturation in an invertebrate animal; proliferation and gene expression in hematopoietic stem cells of Pacifastacus leniusculus. Developmental and Comparative Immunology. 27(8): 661-672. ISSN: 0145-305X. https://doi.org/10.1016/S0145-305X(03)00039-9 STATSOFT INC. 2003. Statistica (Data Analysis Software System), versión 06. www.statsoft.com
TURCHINI GM, Francis DS, Senadheera SPSD, Thanuthong T, De Silva, SS. 2011. Fish oil replacement with different vegetable oils in Murray cod: evidence of an “Omega-3 sparing effect” by other dietary fatty acids. Aquaculture. 315 (3-4): 250-259. ISSN: 0044-8486. https:// doi.org/10.1016/j.aquaculture.2011.02.016
TURCHINI GM, Torstensen BE, Ng WK. 2009. Fish oil replacement in finfish nutrition. Reviews in Aquaculture. 1(1): 10-57. ISSN: 1753-5131. https:// doi.org/10.1111/j.1753-5131.2008.01001.x
VARELA MA, Peña NN, Aranguren CLF. 2017. Necrosis aguda del hepatopáncreas: una revisión de la enfermedad en Penaeus vannamei. Agronomía Mesoamericana. 28 (3): 735-745. ISSN: 22153608. http://doi.org/10.15517/ma.v28i3.27788
VARGAS AF, Guzmán MA, Ochoa JL. 1993. A lipopolysaccharide-binding agglutinin isolated from brown shrimp (Penaeus californiensis Holmes) haemolymph. Comparative Biochemistry and Physiology Part B: Comparative Biochemistry. 104(2): 407-413. ISSN: 0305-049. https:// doi.org/10.1016/0305-0491(93)90387-K WANG CS, Tang KFJ, Kou GH, Chen SN. 1997. Light and electron microscopic evidence of white spot disease in the giant tiger shrimp, Penaeus monodon (Fabricius), and the kuruma shrimp, Penaeus japonicus (Bate), cultured in Taiwan. Journal of Fish Diseases. 20(5): 323-331. ISSN: 0140-7775. https:// doi.org/10.1046/j.1365-2761.1997.00301.x
ZIAEI NS, Rezaei MH, Takami GA, Lovett DL, Mirvaghefi AR, Shakouri M. 2006.The effect of Bacillus spp. bacteria used as probiotics on digestive enzyme activity, survival and growth in the Indian white shrimp Fenneropenaeus indicus. Aquaculture. 252(2-4): 516-524. ISSN: 0044-8486. https:// doi.org/10.1016/j.aquaculture.2005.07.021
La traducción de este artículo al español estuvo a cargo de la Revista Aquacultura de Ecuador.
- FEBRERO 2023 50 NUTRICIÓN

El desarrollo tecnológico en sistemas de posicionamiento, sensado y control ha abierto una nueva era, en la que se dejan atrás las prácticas acuícolas tradicionales. La acuacultura de precisión es un término utilizado para describir el manejo de la variabilidad dentro de un campo, aplicando insumos acuícolas en el lugar, en el momento y en la cantidad correcta para mejorar la eficiencia económica y disminuir el impacto ambiental adverso de la producción (Earl et al., 2000). El análisis y la comprensión de la variabilidad espacial del suelo es muy importante, ya que la variabilidad del suelo hace que el rendimiento de las piscinas se distribuya de manera desigual en una camaronera (Liu et al., 1999).
Entre los factores que deben tomarse en consideración para obtener mejores resultados en el cultivo de camarón están el conocimiento y manejo de los suelos de los fondos de los estanques. La importancia que juegan los suelos a nivel de la productividad en los estanques encavados en el medio natural es un hecho conocido hace bastante tiempo (Gordin et al. 1984, Shilo 1984).
Debido a factores productivos, en algunos casos poco beneficiosos se incrementa la materia orgánica presente en el suelo, que se agudiza en la época de lluvia, siendo responsable del incremento de nivel de amonio, reducción del oxígeno y deterioro de la calidad del agua.

Por lo tanto, el suelo es un factor crítico para la producción; sin embargo, la atención brindada hacia este ha estado limitada por el tiempo requerido para su análisis. El aumento de materia orgánica producida por las excretas de los camarones, debido a la alimentación excesiva y por otros insumos añadidos en los estanques de cultivo, ha sido una preocupación desde los inicios de la actividad (Sandifer y Hopkings, 1995).
Las propiedades del suelo, generalmente, son determinadas mediante métodos de laboratorio, por química húmeda o seca, los cuales en la mayoría de los casos son laboriosos, tardados y costosos (Ge et al., 2011); además de generar residuos químicos que, de no manejarse en forma adecuada, pueden causar contaminación ambiental (Zornoza et al., 2008). Aunado a lo anterior, algunas características químicas del suelo tienen un ciclo muy dinámico y una gran
- FEBRERO 2023
Autores:
Silvia Medranda
César Molina-Poveda*
Skretting Aquaculture Innovation *cesar.molina@skretting.com
52
Rápida determinación de materia orgánica y carbono orgánico en suelos de camaronera mediante espectroscopia visible e infrarrojo cercano VIS-NIR
variabilidad espacial. Esto dificulta y encarece la obtención de información confiable, lo cual hace indispensable el análisis de grandes cantidades de muestras para lograr un buen conocimiento del comportamiento de dichas propiedades (Plant, 2001).


Espectroscopia visible e infrarrojo cercano (VIS-NIR)
La espectroscopia VIS-NIR es una técnica analítica rápida y no destructiva que correlaciona la radiación infrarroja cercana difusamente reflejada con las propiedades químicas y físicas de los materiales (Chang y Laird, 2002) y se ha utilizado para evaluar las cualidades del grano y del suelo (Morra et al., 1991; Ben-Dor y Banin, 1995; Delwiche y Hruschka, 2000) demostrando ser conveniente, simple, preciso y capaz de analizar muchos constituyentes al mismo tiempo. Además de las ventajas antes mencionadas frente a los análisis convencionales, la tecnología de reflectancia VIS-NIR es eficaz, de bajo costo, requiere tiempo mínimo de análisis por muestra, es sencilla y puede ser un complemento ideal, o incluso sustituir a los métodos clásicos, una vez que se desarrollan calibraciones robustas (Terhoeven et al., 2008; Xie et al., 2012); además posee la capacidad de predecir diversas propiedades a partir de un solo espectro.

Actualmente, la técnica de espectroscopia de reflectancia VIS-NIR se ha propuesto como una alternativa para reemplazar los métodos convencionales en la determinación de las propiedades del suelo. Los métodos analíticos utilizados demandan largas horas de recolección de datos para obtener los resultados y requieren insumos químicos que pueden generar impactos ambientales (Nanni y Demattê, 2006).

Se debe tomar en cuenta que la materia orgánica (MO) en el suelo tiene una química muy complicada y su fuerte absorción puede enmascarar algunas características asociadas con los minerales del suelo que tienen una profunda influencia en las propiedades físicas, químicas y biológicas de los suelos.
Se propone un enfoque alternativo para la determinación del contenido de MO en el suelo, utilizando espectroscopia de VIS-
- FEBRERO 2023 53 TECNOLOGÍA
Figura 2. Diferencia en la determinación de materia orgánica en suelo de camaronera por el método de calcinación y espectroscopia VIS-NIR.
Figura 1. Equipo NIR analizando Materia Orgánica en muestras suelo cuya ilustración de como analiza, se acompaña.
Figura 3. Diferencia en la determinación de materia orgánica en suelo de camaronera por el método Walkley-Black modificado y espectroscopia VIS-NIR.
NIR y regresión de mínimos cuadrados parciales para el modelado de datos. Este método se aplicó a diferentes tipos de suelo de camaroneras, desde arcillosos hasta arenosos, en diferentes zonas geográficas donde están ubicadas las mismas. Se obtuvieron cien muestras de suelo, recolectadas con diferentes contenidos de MO de 1 a 6%.

Para la determinación de la MO se siguió el método de calcinación propuesto por Schulte y Hopkins (1996) que se esquematiza en la figura 2. El método de calcinación también se comparó con el método de Walkley-Black modificado para las mismas 100 muestras de suelo; los datos de referencia se obtuvieron por medición del exceso de cromo (VI) con solución estándar de ácido sulfúrico, después de la digestión de la muestra con dicromato de potasio en un medio ácido en tubos de digestión, después de dejar en reposo y filtrar se realizó la lectura en el espectrofotómetro. Los valores obtenidos se multiplicaron por el factor de Van Benmelen para determinar la MO, no encontrándose diferencias en los resultados para la determinación de la MO.
Los contenidos de MO determinados por el método del tubo de digestión se correlacionaron con los obtenidos por espectroscopia de VIS-NIR, utilizando regresión de mínimos cuadrados parciales.

Como se puede estimar a través de las figuras 2 y 3, una vez secas y molidas las muestras del suelo, la determinación de materia orgánica por método ignición toma 7 horas y 30 minutos mientras que por el método de Walkley-Black dura más de 12h, tiempos de análisis superiores a los requeridos por espectroscopia VIS-NIR que están entre 6 y 7 minutos.
El objetivo de este estudio fue evaluar la capacidad de la espectroscopia de reflectancia VIS-NIR para predecir la composición de la MO en los suelos de camaroneras.

El suelo recolectado se dividió en dos porciones: A y B. La porción A se colocó en bolsas debidamente cerradas y se llevó al laboratorio acreditado de análisis del suelo, donde se analizó el contenido de carbono orgánico (CO) y MO. El grupo B,
ser analizado mediante espectroscopia NIR.
La espectroscopia VIS-NIR es básicamente un método empírico, cuyo procedimiento completo consta de dos etapas: la 1ra de calibración, donde se desarrolla la ecuación de predicción, y la 2da etapa de validación, donde se valida la etapa anterior. En la etapa de calibración se utilizaron 60 muestras
de suelo. Las ecuaciones de calibración se desarrollaron utilizando técnicas de regresión de mínimos cuadrados parciales modificados que permitieron relacionar los espectros de reflectancia del infrarrojo cercano con los valores medidos de CO y MO en el suelo. El análisis de datos y desarrollo de la curva empleó el software WinISI 4. Para el desarrollo de la calibración se usó la primera derivada y la corrección SNVDetrend para el procesamiento de datos
- FEBRERO 2023 54 TECNOLOGÍA
una vez
Figura 4. Muestras luego del tratamiento en la región visible (VIS) e Infrarrojo cercano (NIR).
Figura 5. Relación entre los porcentajes de materia orgánica del suelo por el método químico y VISNIR
Figura 6. Porcentajes de Materia y Carbono Orgánico reportados en muestreos aleatorios en varias camaroneras ubicadas en los principales sectores de la provincia del Guayas secado en estufa, fue molido y tamizado a <2 mm, para posteriormente
reduciendo las diferencias iniciales leves. Estos tratamientos matemáticos previos se utilizan para corregir efectos por tamaño de partícula y desplazamiento de la línea base y pendiente (Fig 4).
Resultados
La espectroscopia de infrarrojo cercano predijo bien. El conjunto de predicción que muestra que NIR puede predecir CO y MO en el suelo está de acuerdo con los hallazgos de Ben-Dor y Banin (1995), Fidencio et al. (2002), Sudduth y Hummel (1991), Chang y Laird (2002) y Martín y Malley (2003). En la Fig. 3 se muestra estadísticamente para CO y MO pronosticados. El coeficiente de correlación entre los valores medidos y pronosticados de CO y MO de 0,92 y 0,93 (Fig. 5), respectivamente, mostró que el método NIR tiene la robustez para predecir con precisión estos constituyentes en tipos de suelos de camaronera expuestos a diferentes salinidades. A pesar de que la composición porcentual de MO no estaba por encima del 5% más que una muestra, se encontró una correlación ligeramente mejor en la predicción de este constituyente, aunque la diferencia no fue muy significativa. El R=0,93 obtenido en la predicción de MO estuvo dentro del rango propuesto por Martin y Malley (2003) en su investigación sobre el mismo constituyente (0,81-0,97) en suelos agrícolas y considerado como una buena aproximación. En el caso de CO, el R=0,92 obtenido también mostró una buena precisión.
Para corroborar los resultados, se hizo una serie de muestreos a camaroneras de las provincias de Guayas y El Oro durante el 2022. Es necesario realizar análisis para medir la MO y CO, para lo cual se tomaron muestras al menos en 5 puntos por piscina en una profundidad de 10 cm del suelo (según metodología de Estándar Métodos).
A continuación, se muestran resultados de MO analizada usando la metodología por VIS-NIR en los laboratorios Skretting, resultados obtenidos en principales zonas de producción camaronera de Guayas y El Oro.
La figura 6 muestra los resultados de MO en diferentes zonas de Guayas, con valores que oscilan entre 2,16% y 5,30%. Siendo la zona de Taura, Balo y Chongon las que arrojaron

los porcentajes más altos en relación con los demás sectores, sin que estos superen a los valores esperados en la curva del NIR.
Figura 7 muestra los resultados de MO en diferentes zonas de El Oro, con valores que oscilan entre 1,32% como mínimo y 5,01% como máximo. Siendo las zonas de Tendales y Jambelí, las que arrojaron valores altos en relación con los demás sectores. En general los valores estaban de 2 a 5 según método por pérdida de ignición (Davies, 1974).
Conclusión
Los resultados demostraron una fuerte relación entre las predicciones VIS-NIR y los valores medidos por los análisis de laboratorio habituales. Realizado el análisis de varios modelos matemáticos y a partir de los resultados obtenidos, se observa que la técnica de infrarrojo cercano es una gran alternativa para el rápido análisis de suelos, siendo un análisis medioambientalmente responsable, confiable y sencillo para la determinación de MO y CO en muestras de suelo de camaroneras, mientras que los métodos convencionales son laboriosos, con alto costo y generan gran cantidad de residuos químicos.
Los resultados obtenidos mediante los modelos de predicción generados por VISNIR correlacionan bien con los datos de MO y CO de las muestras de suelos analizados por el método base utilizado para crear la curva patrón en el VIS-NIR con base en los modelos de calibración (datos no mostrados)•
Para más información sobre este artículo escriba a: cesar.molina@skretting.com
Bibliografía
Ben-Dor E, Banin A. 1995 Near-infrared analysis as a rapid method to simultaneously evaluate several soil properties. Soil Sci Soc Am J. 59:364–372
Caicedo, N. 2018. Análisis de oferta y demanda del camarón en la provincia de El Oro y Ecuador en los Últimos Ocho Años. In Universidad Técnica de Machala.
Chang CW, Laird DA. 2002Near-infrared reflectance spectroscopy analysis of soil C and N. Soil Science. 167(2):110–116.
Delwiche S, Hruschka WR. 2000 Protein content of bulk wheat from near-infrared reflectance of individual Kernel. Cereal Chem. 77(1):86–88
Ge, Y., C. L. S. Morgan, S. Grunwald, D. J. Brown y D.C. Sarkhot. 2011. Comparison of soil reflectance spectra and calibration models obtained using multiple spectrometers. Geoderma 161:202-211 Morra MJ, Hall MH, Freeborn LL. 1991 Carbon and Nitrogen analysis of soil fractions using nearinfrared reflectance spectroscopy. Soil Sci Soc Am J. ;55:288–291.
Nanni, M.R.& Demattê, J.A.M. 2006. Spectral reflectance methodology in comparison to tradicional soil analysis. Soil Science Society of America Journal, 70, 393-407.
Pérez N., J. C., J. Soler A., G. Arango P., E. Meneses O. y O. S. Ruiz V. 2014. NIR Spectroscopy as Quick Exploratory Technique for Detection of Chrysanthemum Leaf Yellowing (Dendranthema grandiflora var. Zembla). Revista Facultad Nacional de Agronomía Medellín 67(1): 7163-7168.
Plant, R. 2001. Site-specific management: The application of information technology to crop production. Computers and Electronics in Agriculture 30:9-29.
Schulte, EE & BG Hopkins. 1996. Estimation of organic matter by weight loss-on-ignition. In: FR Magdoff et al. (ed) Soli organic matter: Analysis and interpretation. SSSA Spec. Publ. 46. SSSA, Madison, WI. P 21-31.
Terhoeven U., T., H. Schmidt, R. G. Joergensen y B. Ludwig. 2008. Usefulness of near-infrared spectroscopy to determine biological and chemical soil properties: importance of sample pre-treatment. Soil Biology and Biochemistry 40:1178-1188.
Viscarra R., R. A., D. J. Walvoort, A. B. McBratney, L. J. Janik y J. O. Skjemstad. 2006. Visible, near infrared, mid infrared or combined diffuse reflectance spectroscopy for simultaneous assessment of various soil properties. Geoderma 131: 59-75.
Zornoza, R., C. Guerrero, J. Mataix-Solera, K. M. Scow, V. Arcenegui y J. Mataix-Beneyto. 2008. Near infrared spectroscopy for determination of various physical, chemical and biochemical properties in Mediterranean soils. Soil Biology and Biochemistry 40:1923- 1930.
- FEBRERO 2023 55 TECNOLOGÍA
Figura 7. Porcentajes de Materia y Carbono Orgánico reportados en muestreos aleatorios en varias camaroneras ubicadas en los principales sectores de la provincia de El Oro

Ecosistemas de carbono azul: una estrategia para mitigar el cambio climático global

Autores:
Sahina Akter
Suman Nama
División de Manejo de Cosecha y Post Cosecha de Recursos Pesqueros
ICAR-Instituto Central de Educación Pesquera, Pinch Marg, Off Yari Road, Versova, Mumbai, India.
sumancau2017@gmail.com
Publicación orginal: http://aquainternational.in/i/jan-2023.pdf
El calentamiento de la Tierra en las últimas décadas ha provocado un abrumador consenso entre científicos expertos en clima debido a que las actividades antropogénicas han aumentado la cantidad de gases de efecto invernadero (GEI) en la atmósfera. Para mitigar el severo impacto del cambio climático necesitamos adoptar varias estrategias, entre ellas el carbono azul como estrategia esencial. El carbono azul es el carbono orgánico que ha sido capturado y secuestrado por plantas marino-costeras. Los ecosistemas de pastos marinos, manglares y marismas son altamente productivos y actúan como los mayores depósitos de carbono en los ecosistemas costeros (Nellemann et al., 2009; Sappal et al., 2016). Los ecosistemas costeros con vegetación (bosques de mangles, pastos marinos, marismas o pantanos) son desproporcionadamente más importantes en el secuestro de dióxido de carbono que los ecosistemas terrestres.
Estos ecosistemas son excelentes sumideros de carbono y brindan varios servicios ecosistémicos, como ser refugio para una variedad de fauna como peces, cangrejos y aves migratorias. Según varios investigadores, los ecosistemas de carbono azul tienen un gran potencial para secuestrar carbono dentro de su biomasa viva y en el suelo subterráneo (sedimentos), donde se puede encontrar raíces, hojarasca y madera muerta. Se estima que el potencial para secuestrar carbono azul es decenal a corto plazo en la biomasa y milenario en seis escalas de tiempo más largos en sedimentos (Duarte et al. 2005a; Lo Iacono et al. 2008; Mcleod et al. 2011). El océano desempeña un papel vital en la captura y el reciclaje del CO2 atmosférico y absorbe globalmente más de un tercio de las emisiones antropogénicas de CO2 a través de procesos biológicos, físicos y químicos debido al intercambio gaseoso en la interfaz entre el océano y la atmósfera (Siegenthaler y Sarmiento, 1993; Tamis y Foekema, 2015). Los ecosistemas costeros representan menos del 5% de la superficie total de la Tierra, pero tienen un rol significativo en la regulación global del ciclo de carbono (Twilley et al., 1992). Estos ecosistemas almacenan alrededor de 3418,5 toneladas métricas de dióxido de carbono equivalente por hectárea (t CO2/ha) en su biomasa y carbono sedimentario.
Puntos Importantes
El cambio climático global incrementa las amenazas para las zonas costeras. El cambio
MANEJO AMBIENTAL
57
climático provoca el aumento del nivel del mar, tsunamis, etc dañando los ecosistemas costeros. Los ecosistemas de carbono azul como los manglares y marismas tienen un enorme potencial para secuestrar el carbono atmosférico y mitigar el cambio climático global. Lamentablemente, actividades naturales y antropogénicas (destrucción de manglares para el cultivo de camarón) provocan una gran destrucción del ecosistema de carbono azul. Como resultado, el contenido de carbono atmosférico aumenta día a día y la Tierra se calienta. La conservación y protección de los ecosistemas de carbono azul mitigará las emisiones de CO2. La conservación de los ecosistemas de carbono azul también ayudará en la conservación de la biodiversidad, la protección de comunidades a lo largo de las zonas costeras, la conservación de pesquerías de gran valor, la prevención de la erosión y también la degradación de las comunidades adyacentes.
Este carbono azul costero brinda beneficios para la mitigación del cambio climático debido a su capacidad de almacenar carbono en condiciones adversas. Debido a esto, ha llamado la atención de la comunidad científica en general. Una porción significativa del carbono biogénico llega al lecho marino, donde puede ser enterrado y encerrado de manera efectiva fuera de la atmósfera durante escalas de tiempo prolongadas, constituyendo un sumidero de CO2 y contribuyendo en la mitigación del cambio climático (Bowler et al., 2009). La reunión del Comité de Biodiversidad OSPAR (BDC) en marzo de 2015, consideró que los ecosistemas de carbono azul tienen un potencial colosal para mitigar el cambio climático global en el área marítima OSPAR (OSPAR, 2015). Los ecosistemas marinos y costeros juegan un papel crucial en el secuestro y almacenamiento de carbono y mitigan el cambio climático global (Duarte et al., 2005a; Nelleman et al., 2009; Murray et al. 2011). Los hábitats costeros con vegetación están ampliamente distribuidos y se estima que son globalmente responsables del entierro de 120-329 Tg C/año, lo que representa al menos la mitad de la estimación más baja del entierro global de carbono en los sedimentos marinos (Nellemann et al., 2009). En los últimos años, los sumideros de “carbono azul” se están perdiendo a un ritmo crítico, y se requieren medidas
urgentes para evitar una mayor degradación y pérdida de estos ecosistemas de carbono azul. Se debe dar una alta prioridad a la gestión de restauración y conservación para comprender los diferentes factores que influyen en la captura de carbono y mejorar la actividad científica para prevenir la degradación de estos ecosistemas.
Ecosistemas de carbono azul y biodiversidad: Los hábitats costeros de carbono azul son puntos calientes (hot spots en inglés) para la biodiversidad y brindan funciones ecosistémicas valiosas, lo que incluye tener una enorme capacidad de ser sumidero de carbono (Duarte et al., 2008, Duarte et al., 2009; Nellemann et al., 2009). Sin embargo, en los últimos años, la destrucción y degradación de este hábitat está aumentando debido a diferentes actividades antropogénicas. La restauración de estos hábitats da como resultado el desarrollo del carbono azul, así como la conservación de la biodiversidad de estos ecosistemas. Por lo tanto, la protección de los hábitats de carbono azul y la restauración a gran escala de sumideros de carbono azul perdidos es una estrategia beneficiosa o “win-win” para todos, ya que mitiga las emisiones de CO2, mejora los recursos costeros (Nellemann et al., 2009;) y la biodiversidad.
Ecosistemas de carbono azul y almacenamiento de carbono: Los ecosistemas de carbono azul eliminan el CO2 de la atmósfera a través de la fotosíntesis, devuelven algo a la atmósfera a través de la respiración y la oxidación, y almacenan el carbono restante en dos depósitos: biomasa viva (tanto sobre el suelo en la madera, las hojas y vegetación subterránea en raíces)
y carbono orgánico del suelo. La tasa de secuestro de carbono es la cantidad total de carbono que se agrega a la biomasa y al suelo anualmente. Estos ecosistemas suelen tener una vegetación madura que mantiene una biomasa constante, y todo lo captado termina enterrado en la reserva de carbono del suelo. Los ecosistemas de carbono azul son puntos calientes para el secuestro de carbono porque convierten el CO2 en biomasa vegetal que se encuentra en ambientes de depósito, y los suelos tienen altas tasas de acumulación que dan como resultado el rápido entierro de materia orgánica en condiciones anóxicas, acumulando partículas de carbono tanto autóctonas como alóctonas (Kennedy et al., 2010; Saintilan et al., 2013; Donato et al., 2011; Lo Iacono et al., 2008; Serrano et al., 2016a).Las tasas anuales de secuestro de carbono varían en los diferentes hábitats costeros, como marismas o pantanos y manglares. El secuestro de carbono promedio es de 6 a 8 toneladas/ CO2e/ha/año. Los pastos marinos tienden a secuestrar carbono en aproximadamente 4 t CO2e/ha/año (Lewis et al. 2009).
La tasa de carbono almacenado en la biomasa viva de las praderas marinas es de 0.4 a 18.3 t CO2/ha, y en pantanos de 12 a 60 t CO2/ha. Los manglares mantienen 237563 t CO2/ha en biomasa viva. Sin embargo, la preservación a largo plazo y la acumulación continua de carbono en los suelos de marismas, manglares y pastos marinos dan como resultado la formación de depósitos ricos en materia orgánica de varios metros de espesor (Mateo et al., 1997; Donato et al., 2011). Según muchos investigadores, la tasa de almacenamiento y secuestro de carbono en los diferentes ecosistemas de carbono
Tabla 1: Extensión y reservas (stocks) de carbono y tasas de depósito dentro del metro superior del suelo de ecosistemas de pantanos, manglares y pastos marinos, y macroalgas C enterradas en el océano (Fuente: Serrano et al., 2019).
- FEBRERO 2023 58 MANEJO AMBIENTAL
Pantanos Manglares Pastos marinos Macroalgas
Ecosistema Extensión Global (km2) Tasa global de entierro de C (Tg C/año) Stock global de C en el suelo (Pg C) 22,000 - 400,000 4.8 - 87.3 0.4 - 6.5 137,760 - 152,3615 22.5 - 24.9 5 -10.4 177,000 - 600,000 48 - 112 4.2 - 8.4 1,400,000 - 5,700,000 61 - 268 n/a
azul se muestra a continuación (Tabla 1). La tasa de almacenamiento de carbono en el suelo y la biomasa del ecosistema de carbono azul en pastos marinos es: 512 Mg CO2e/ha, marismas: 917 Mg CO2e/ha y manglares; 1028 Mg CO2e/ha. Los ecosistemas de carbono azul eliminan regularmente el CO2 de la atmósfera y lo secuestran en forma de carbono del suelo. La tasa promedio de secuestro de carbono de estos ecosistemas son pastos marinos: 138 gC/m2/año igual a 5.1 tCO /ha/año, marismas: 218 gC/m2/año igual a 8.0 tCO/ha/año, manglares: 226 gC/ m2/año igual a 8.3 t CO/ha/año.
Factores que influyen en el almacenamiento de carbono azul en ecosistemas: El carbono orgánico secuestrado en ecosistemas de carbono azul alcanza el suelo marino y se cubre con una capa de sedimentos. El secuestro de carbono ocurre cuando la tasa de captación de sedimentos es más significativa que la tasa a largo plazo de la erosión, bioturbación y descomposición. Muchos factores influyen el secuestro de carbono, como factores bióticos y abióticos que actúan en la columna de agua, el dosel, suelo y la historia del paisaje y registros de variación del nivel del mar. Los otros factores se detallan en la Tabla 2.
Carbono azul y mitigación del cambio climático:
El almacenamiento de carbono en el ecosistema de carbono azul es una de las soluciones más baratas, seguras y cómodas para reducir las emisiones de gases de efecto invernadero y promover la adaptación al cambio climático (Jones et al., 2012, Turner et al., 2009). Dos mecanismos primarios para reducir las emisiones de gases de efecto invernadero en esta pérdida continua de hábitats costeros y ecosistemas marinos son:
1) Conservar depósitos de carbono históricos.
2) Restaurar y reconstruir depósitos de carbono degradados.
Debido a la perturbación de los ecosistemas de carbono azul, la tasa a la que se pierde el carbono es mucho mayor que la tasa a la que se puede restaurar. Las otras ventajas de los ecosistemas de carbono azul son que los manglares actúan como barreras naturales, sirviendo como la primera defensa de las marejadas ciclónicas, estabilizando la costa, reduciendo riesgos para las comunidades
Tabla 2: Factores que influyen en el almacenamiento de carbono en los ecosistemas de carbono azul.
Factores:
1) Variabilidad espacial en la sedimentación
I. Mar abierto
II. Zonas costeras
III. Cañones submarinos
Influencia y discusión
La sedimentación es la velocidad a la que se hunden las partículas en suspensión y se acumulan en el suelo oceánico. La cantidad de sedimentos suspendidos y la tasa de deposición varían drásticamente en los diferentes sitios del océano como Tasa de sedimentación baja en mar abierto debido al viento
Zonas costeras - alta tasa de sedimentación debido al aporte de los ríos
Cañones submarinos: la tasa de almacenaje de carbono en los cañones submarinos es más significativa que la del talud continental adyacente.
2) Cambios humanos en los sistemas sedimentarios globales
I. Agricultura/preparación-limpieza de tierra
II. Presas
3) Densidad de la vegetación
Las diferentes actividades humanas han hecho que se modifique el ciclo de los sedimentos. El ciclo sedimentario global comenzó a cambiar cuando los humanos comenzaron a preparar y limpiar la tierra para la agricultura y construir represas. La limpieza de la tierra provoca un aumento en la tasa de erosión.
Las presas son trampas de sedimentos casi 100% eficientes.
La densidad de la vegetación en los manglares, praderas de pastos marinos y marismas es suficiente para cambiar los flujos de agua, lo cual es suficiente para reducir la erosión y aumentar la deposición de sedimentos porque ejercen un control primario sobre el almacenamiento de carbono a través de la producción de biomasa y el ciclo de nutrientes (Lavery et al., 2013; Serrano et al. 2014, 2016a; Kelleway et al., 2016a; Atwood et al., 2018; 2015)
4) Carga de nutrientes
Tanto los ecosistemas de manglares como los de pastos marinos, que han estado sujetos a altas cargas de nutrientes, aumentan la tasa de captura y secuestro de carbono.
- FEBRERO 2023 59 MANEJO AMBIENTAL
costeras (Barbier, 2007; Das y Vincent, 2009). Las praderas de pastos marinos también reducen la erosión de la costa al atrapar sedimentos en suspensión en sus sistemas de raíces (Barbier, 2007). Los ecosistemas costeros absorben contaminantes como metales pesados, nutrientes, materia en suspensión y patógenos, ayudan a mantener la calidad del agua, previenen la eutrofización y generan “zonas muertas”. Un ecosistema costero saludable también brinda diversas oportunidades recreativas, como el snorkel, la pesca recreativa, la navegación y el ecoturismo costero es uno de los sectores de más rápido crecimiento.
Amenazas al almacenamiento de carbono azul: las áreas de manglar, pastos marinos y pantanos ya se han perdido en las últimas décadas debido a la intervención de diferentes actividades humanas como la recuperación, deforestación, la ingeniería y la urbanización, la transformación en estanques de acuicultura (Green y Short 2003 Duarte et al. 2005b; Silliman et al. 2009. Durante las últimas décadas, el hábitat de pastos marinos disminuyó debido a la eutrofización costera, la sedimentación y el desarrollo (Green and Short 2003; Duarte et al. 2005b; Waycott et al. 2009), mientras que los hábitats de manglares y pantanos han sido dañados por dragados, rellenos, drenajes, cascadas tróficas y también por la introducción o invasión accidental de especies invasoras (Valiela et al. 2001; Alongi 2002; Silliman et al. 2005; 2009). El aumento del nivel y la temperatura de la superficie del mar debido al cambio climático es otra amenaza para el ecosistema costero que puede erosionar e inundar manglares y pantanos aumentando la profundidad del agua y reduciendo la disponibilidad de luz para apoyar la fotosíntesis (Björk et al. 2008; Woodroffe 1995; Silliman et al. 2009). La pérdida anual promedio mundial de pérdidas de ecosistemas de carbono azul reduce su capacidad de almacenamiento de carbono. Tiene graves implicaciones para las poblaciones humanas que dependen de estos ecosistemas para su alimentación, sustento y protección costera. Se deben abordar estrategias de conservación y gestión para proteger estos ecosistemas de carbono azul y controlar la dinámica del carbono en los sistemas costeros (Middleton y McKee, 2001; Kristensen et al., 2008).
Tabla 3: Políticas actuales necesarias para salvaguardar el carbono en los ecosistemas de carbono azul asociados (Fuente: Climate Focus, 2011; Conservación International, 2008; Pendleton et al., 2012).
Políticas y Oportunidades Descripción
Artículo 4 de la Convención Marco de las Naciones Unidas sobre el Cambio Climático (CMNUCC)
Promover la gestión sostenible, la conservación y la mejora de los sumideros y depósitos de todos los gases de efecto invernadero, que incluyen al océano y otros ecosistemas marino-costeros.
Reducción de emisiones por deforestación Un marco para fomentar y financiar actividades que reducen las emisiones o mejorar las absorciones de GEI de actividades relacionadas con los bosques
Las Acciones de Mitigación Nacionalmente Apropiadas (NAMAs por su sigla en inglés)
Parte de los mecanismos de la CMNUCC para que los países en desarrollo accedan al financiamiento del carbono y brinden oportunidades para incluir actividades de cambio de uso de la tierra, conservación y restauración en los ecosistemas costeros en los esfuerzos nacionales de mitigación.
Mecanismos de Desarrollo Limpio (MDL)
Los países en desarrollo pueden obtener financiamiento para proyectos elegibles con un beneficio neto de GEI, que incluye la captura de CO2 de los bosques. El MDL ha aprobado una metodología de restauración de manglares a gran escala.
Mercado Voluntario de Carbono
Brinda la posibilidad de generar apoyo financiero para actividades de conservación o restauración de ecosistemas de carbono azul y proporciona un marco para contabilizar las reducciones de emisiones de GEI en humedales costeros, manglares, marismas y pastos marinos, deltas, llanuras aluviales, entre otros.
Políticas actuales para salvaguardar ecosistemas de carbono azul asociados al carbono: diferentes políticas incentivan las actividades de mitigación basadas en la naturaleza para los ecosistemas de carbono azul mediante diferentes políticas de carbono y mecanismos financieros. Se destacan algunos ejemplos abajo en la Tabla 3.
Conclusión: En los últimos años los ecosistemas de carbono azul han recibido atención internacional por su papel potencial en mitigar las emisiones de CO2. La conservación y protección de los ecosistemas de carbono azul ayudarán a
mantener en el futuro el carbono global secuestrado y prevenir emisiones que relacionan a cambios de uso de la tierra. La conservación del carbono azul de los ecosistemas también ayudará en la conservación de la biodiversidad, protección de comunidades costeras, conservación de pesquerías, prevenir la erosión y también prevenir la degradación de comunidades adyacentes. Necesitamos crear una concienciación masiva sobre la importancia de los ecosistemas de carbono azul para que el ecosistema pueda sostenerse•
*Referencias pueden ser proporcionadas a petición
- FEBRERO 2023 60 MANEJO AMBIENTAL


ESTADÍSTICAS ÍNDICE Edición 150 - Diciembre 2022 63 68 72 Exportaciones de camarón y tilapia Reporte de mercado de EE. UU. Reporte de mercado de China
COMERCIO EXTERIOR
CAMARÓN: EVOLUCIÓN DE EXPORTACIONES 2010 -2022
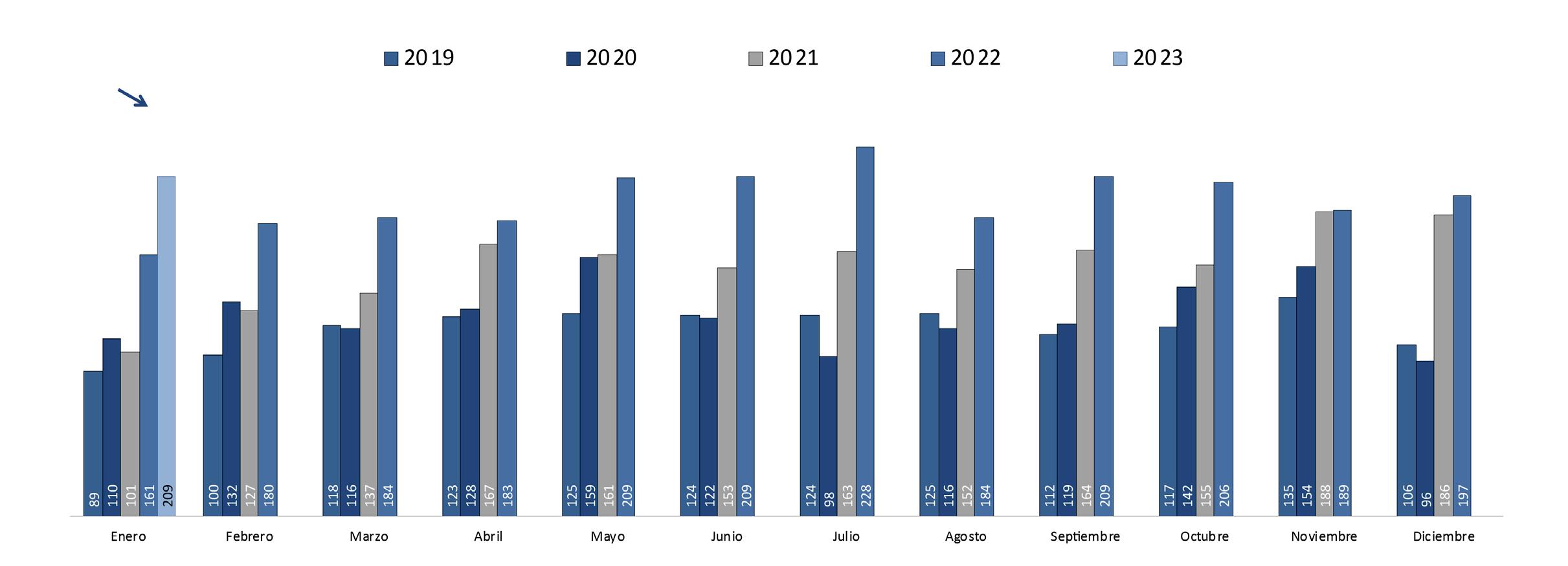
Fuente: Banco Central del Ecuador
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: COMPARATIVO MENSUAL (Millones de Libras) 2019 - 2023
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
- FEBRERO 2023 63 ESTADÍSTICAS
COMERCIO EXTERIOR
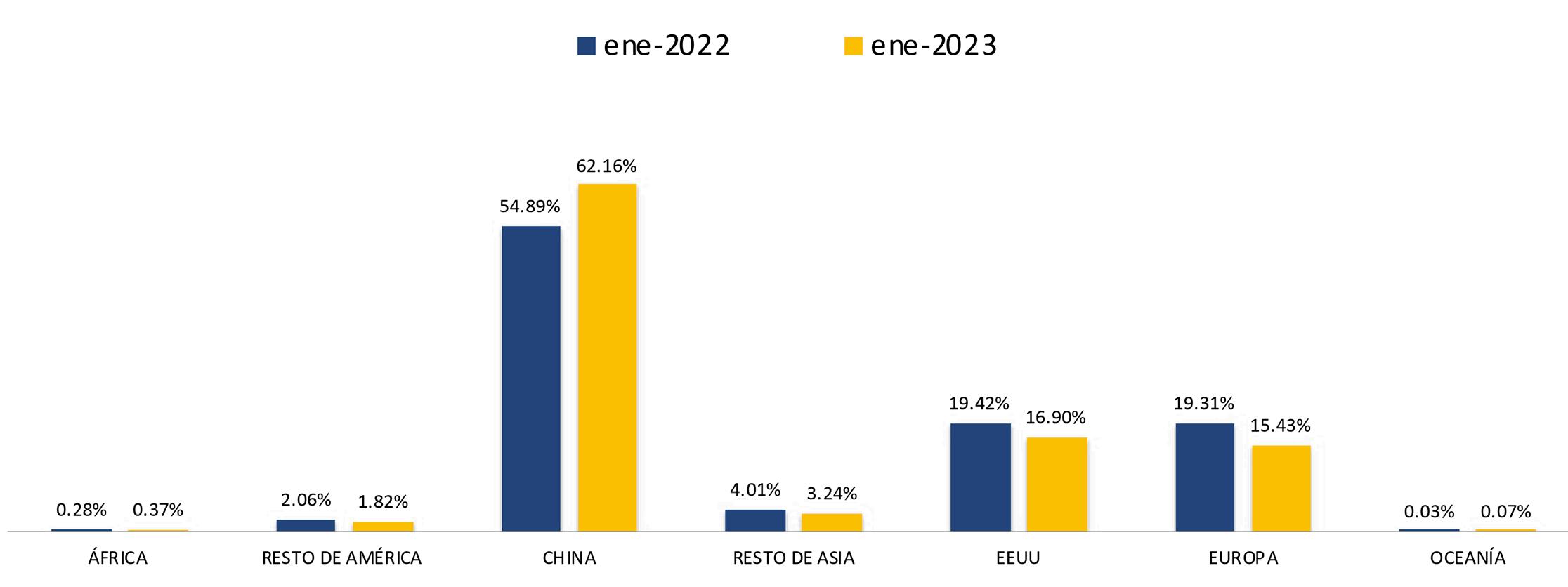
CAMARÓN: PARTICIPACIÓN POR DESTINO (Libras)
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: PRINCIPALES PAÍSES DESTINOS DE EXPORTACIÓN (Millones de Libras)
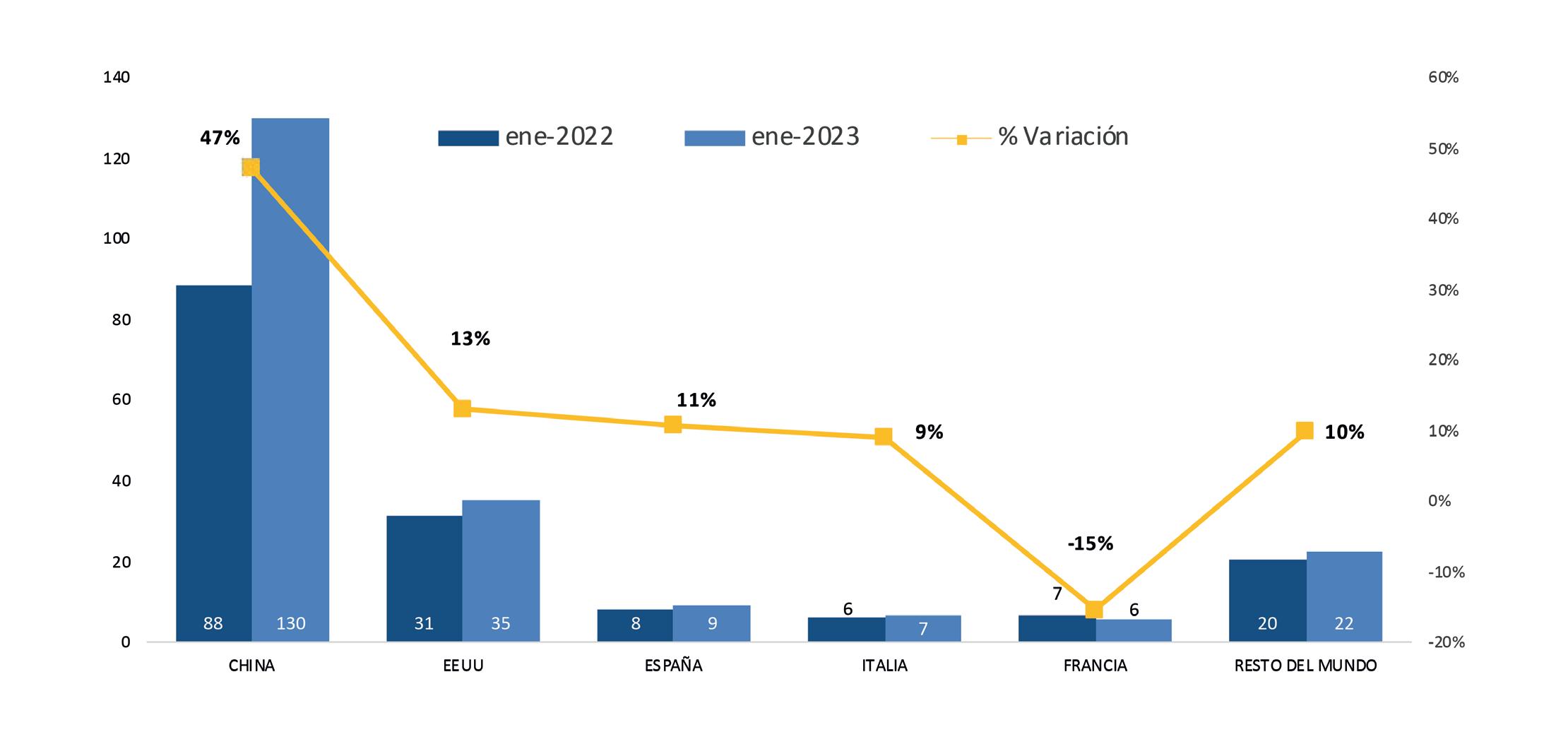
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
- FEBRERO 2023 64 ESTADÍSTICAS
COMERCIO EXTERIOR
CAMARÓN: EXPORTACIONES POR PARTIDA ARANCELARIA
Enero - Diciembre 2022
Fuente: Banco Central del Ecuador
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: EVOLUCIÓN DEL PRECIO PROMEDIO ANUAL DE EXPORTACIÓN (Libras)
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
CAMARÓN: EVOLUCIÓN DEL PRECIO PROMEDIO MENSUAL DE EXPORTACIÓN (Libras)
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
- FEBRERO 2023 65 ESTADÍSTICAS
COMERCIO EXTERIOR
TILAPIA: EVOLUCIÓN DE EXPORTACIONES MENSUALES A EE. UU.
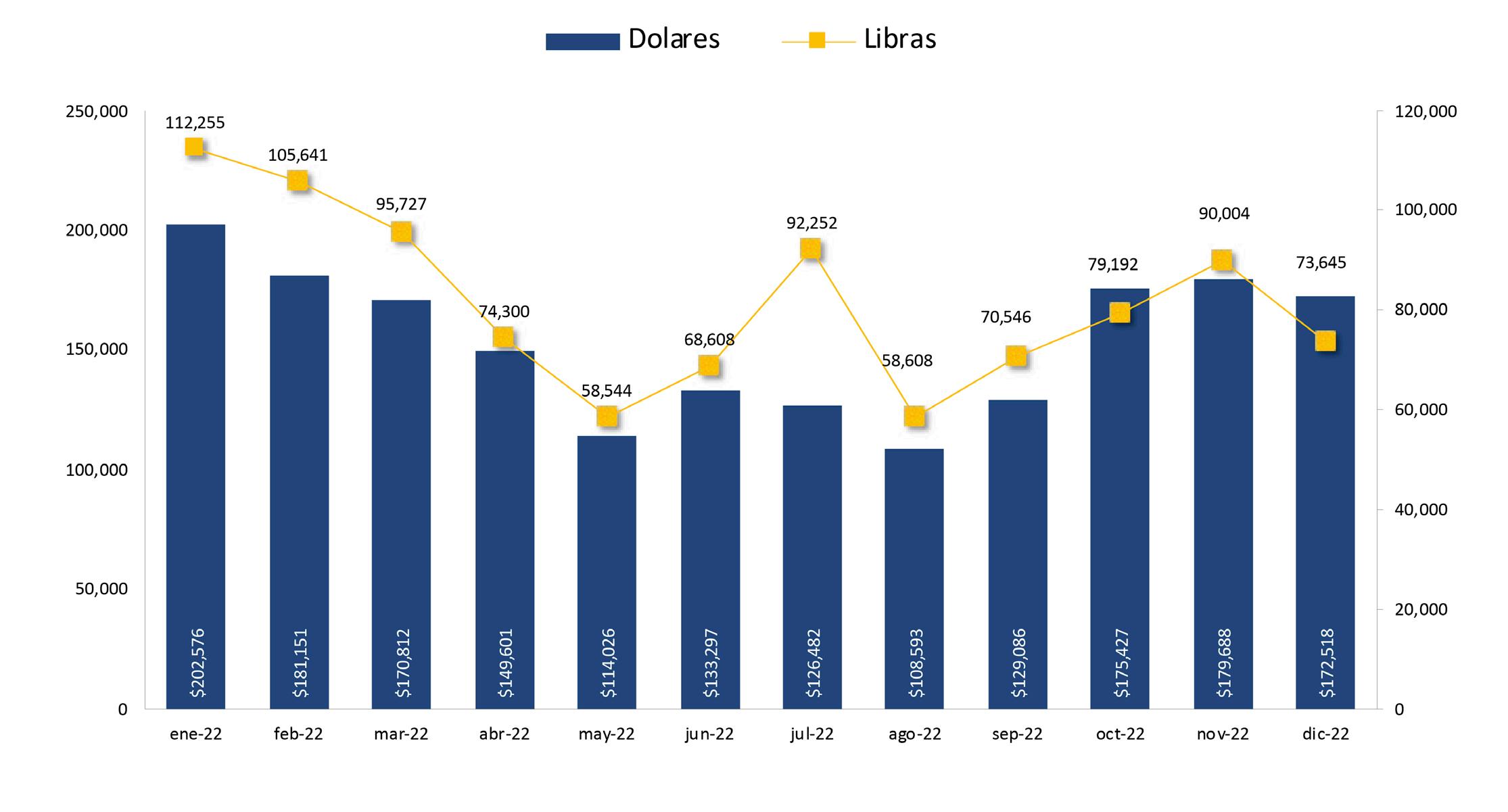
Fuente: National Oceanic and Atmospheric Administration - NOAA
Elaborado por: Cámara Nacional de Acuacultura
TILAPIA: EVOLUCIÓN DEL PRECIO PROMEDIO MENSUAL DE EXPORTACIÓN A EE. UU. (Libras)
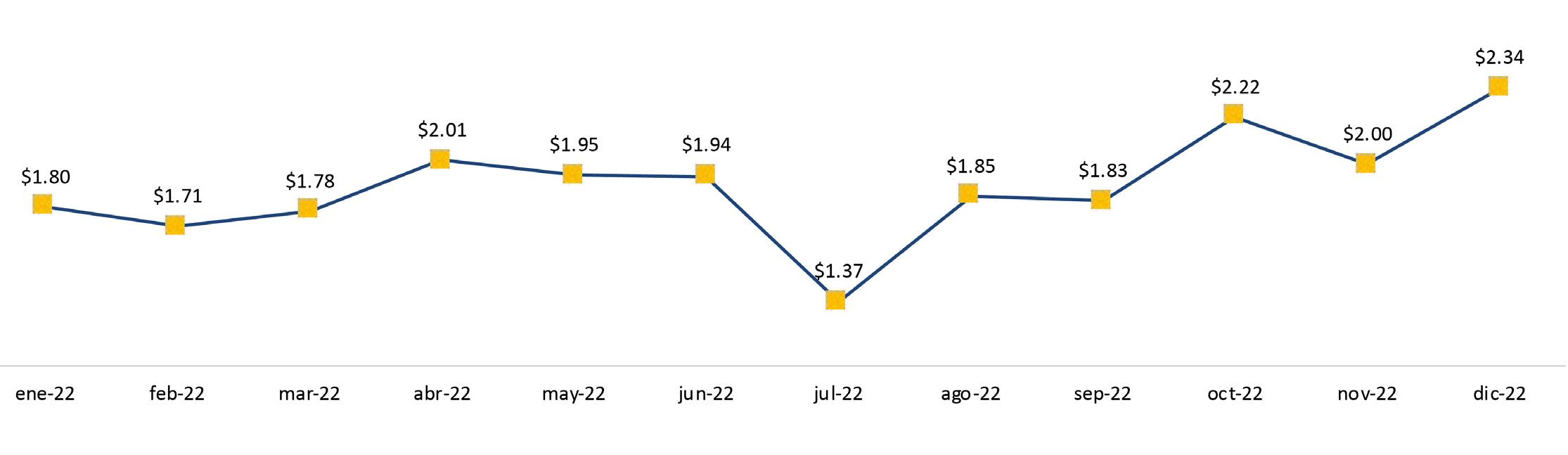
Fuente: National Oceanic and Atmospheric Administration - NOAA
Elaborado por: Cámara Nacional de Acuacultura
- FEBRERO 2023 66 ESTADÍSTICAS
Importación de camarón de China
Autor: Sander Visch Kontali Shrimp sander@kontali.no www.shrimp.kontali.no
Precios
Los precios de importación han estado a la baja desde septiembre y continuaron cayendo en diciembre. Los precios disminuyeron $0.43 desde noviembre y alcanzaron los $5.98/kg, el punto de precio más bajo observado durante 2022. En comparación al año anterior, los precios fueron $0.70 más bajos.

A pesar de una caída en los precios, el fortalecimiento del volumen de importación en diciembre resultó en un aumento en el valor de las importaciones, finalizando en $582 millones. El valor total para 2022 alcanzó un poco menos de $5.7 billones, un 53% más que el año anterior.
Proveedores
Durante diciembre, las importaciones desde Ecuador aumentaron un 5% respecto a noviembre y alcanzaron las 62,224 toneladas. Esto corresponde a un aumento interanual del 44%. China se abrió a inicios de diciembre y eliminó una gran parte de sus estrictas regulaciones por la COVID-19. El camarón ecuatoriano tiene una gran demanda en China durante todo el año, particularmente durante las celebraciones del Año Nuevo chino, donde el camarón HOSO es muy popular. Los precios cayeron $0.35 con respecto al mes anterior, llegando a $5.86/kg. Ecuador se convirtió en el mayor proveedor de China en 2022, y el volumen total importado en 2022 alcanzó las 564,747 toneladas, un 49% por encima del volumen importado de Ecuador en 2021. El 64% del volumen importado de China en 2022 fue suministrado por Ecuador, un 2% más que en 2021.
Las importaciones de India se fortalecieron considerablemente durante diciembre después de haber estado en una trayectoria descendente desde agosto. Las importaciones aumentaron un 81% con respecto a noviembre, alcanzando las 13,549 toneladas. Esto fue un 8% más alto que el año anterior.
La mayor parte del camarón procedente de la India se destina a la industria de reprocesamiento y, dado que China abrió en diciembre, se esperaba que se abrieran más fábricas y aumentaran sus actividades. Los precios de la India también cayeron en diciembre en $0.68 a $5.66/kg. El volumen total de importación de India alcanzó las 137,275 toneladas, un aumento del 18% con respecto a 2021. Sin embargo, a pesar de este aumento, la participación de mercado de India en el mercado chino disminuyó del 19% en 2021 al 16% en 2022.

Después de una caída de las importaciones en noviembre de Vietnam, el volumen de importación se fortaleció durante diciembre y alcanzó las 2,606 toneladas.
Este fue un aumento del 24% con respecto al mes pasado y un 61% superior al año anterior. A lo largo de 2022, el volumen total importado de Vietnam se duplicó desde 2021 alcanzando las 39,107 toneladas.
El bajo suministro de Vietnam a China durante el 2021 se debió al impacto general de los brotes de la COVID-19 en Vietnam, lo que provocó que los procesadores vietnamitas no pudieran entregar
- FEBRERO 2023
MERCADO
68
camarón debido a dificultades logísticas y a la baja disponibilidad de materia prima. Los precios de Vietnam también cayeron en diciembre en $0.57 para llegar a $5.71/kg.
Después de alcanzar su volumen más alto en un mes, durante los últimos tres años en noviembre, las importaciones de Tailandia cayeron nuevamente en diciembre a 2,613 toneladas, una caída del 23% respecto al mes anterior. En comparación con el año anterior, esto todavía fue un 19% superior a las importaciones de diciembre de 2021. Si observamos más de cerca los patrones históricos, podemos ver que las importaciones de Tailandia a China tienden a fortalecerse en el cuarto trimestre. Esto está relacionado con la mayor demanda de camarón de valor agregado de Tailandia antes de las celebraciones en China durante el primer trimestre.
Las importaciones totales de Tailandia en 2022 alcanzaron las 24,485 toneladas, un aumento del 10% con respecto a 2021. Después de que los precios alcanzaran su punto máximo durante septiembre, los precios han bajado y continuaron cayendo en

diciembre a $10.06/kg, una caída de $0.14 respecto al mes anterior y $0.57 menos que el año anterior.
Perspectivas
De acuerdo con nuestras fuentes, se espera que la demanda de productos del mar caiga un poco después de las festividades en China, el año nuevo y el festival de los faroles. Sin embargo, a partir del 8 de enero, la Administración Estatal de Regulación del Mercado de China ha “suavizado” las pruebas de la COVID-19 en todos los alimentos refrigerados y congelados importados, de esta manera tanto importadores como los exportadores enfrentan menos riesgos.
Además, dado que se espera que la demanda de EE.UU. y la UE sea relativamente lenta en los próximos meses, los exportadores podrían establecer precios más bajos para enviar productos a China, brindando una oportunidad para que los importadores se abastezcan de camarón de menor precio. Por lo tanto, esperamos que el volumen de camarón importado se mantenga saludable, pero que los precios pagados por él serán más bajos.
En cuanto a la producción local, los productores chinos de camarón se apresuran a vender su oferta restante a medida que la demanda y los precios están cayendo. Además de eso, también se prevé que la futura oferta de camarón sea un poco menor, ya que la cantidad de PL sembradas en las últimas semanas ha sido considerablemente menor que el año anterior. Adicionalmente, los bajos precios del mercado hacen que los productores duden en sembrar y tomar riesgos; este es especialmente el caso de los productores que alquilan áreas para cultivo.
En los principales países proveedores, los precios en puerta de finca en Ecuador son bajos y los márgenes están presionados por el recorte del subsidio al diésel, especialmente para operaciones medianas. Con una demanda de mercado baja y - precios más bajos para productos procesados -, como las solicitudes del mercado estadounidense, el suministro de producto HOSO a China vuelve a ser más interesante. Sin embargo, dado que la demanda podría ser ligeramente menor en los próximos meses para el camarón HOSO ecuatoriano, los precios hacia el mercado chino también disminuirán aún más.
- FEBRERO 2023 69 MERCADO
Importaciones chinas en 2020, 2021 y 2022 (volúmenes y precio promedio/kg)
Volumen
Ene
Volumen Volumen Volumen (Toneladas)
Precio Precio Precio Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Precio de Importación (USD/kg)
Comparación de volúmenes de importación de los principales países proveedores (2021 vs 2022)
Dado que India está comenzando la nueva temporada de siembra, la primera cosecha llegará al mercado a principios del segundo trimestre. Por lo tanto, se espera que la oferta de India, al igual que en años anteriores, sea relativamente baja en los próximos meses.

Ene Volumen (Toneladas) Precio (USD/Kg) Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic

Vietnam ha comenzado a enfocarse más en los mercados de alta gama en China (y Japón) ya que requiere menos procesamiento y costos de transporte más bajos que abastecer a la UE o los EE.UU. Por lo tanto, podríamos ver un repunte en los productos provenientes de Vietnam. Sin embargo, China aún enfrenta niveles elevados de casos de COVID, y muchas personas no están regresando por completo a la misma vida que tenían antes de la pandemia. Se mantienen voluntariamente alejados de lugares públicos con una gran cantidad de personas, como los restaurantes de lujo y, por lo tanto, es posible que el servicio de alimentos regrese parcialmente a la normalidad en los próximos meses•
Este informe fue escrito originalmente en inglés por Seafood TIP. El informe fue traducido por la Cámara Nacional de Acuacultura.
Para más información sobre este artículo escriba a: sander@kontali.no
- FEBRERO 2023 70 MERCADO
Precios promedio de importación de los principales países proveedores (2021 vs 2022) Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic

Importación de camarón de Estados Unidos
Autores: Jim Kenny jkenny@urnerbarry.com
Gary Morrison gmorrison@urnerbarry.com

 Urner Barry
Urner Barry
Importaciones de todos los tipos, por tipo
La data actualizada de importación de camarón terminó con fuerza como el resto del año. Las importaciones de diciembre fueron un récord: 194.1 millones de libras, o un 32.7% por encima del mismo mes del año pasado. Como resultado, las importaciones anuales totales fueron un 19.8% superiores a las 1.970 millones de libras; otro máximo histórico y apenas por debajo de los dos billones de libras.
Los cinco principales socios comerciales de EE.UU. se mantuvieron igual. Los cuatro primeros enviaron significativamente más que el año pasado en esta época. India (+23.0%) se mantuvo en primera posición. Las grandes ganancias en Ecuador (+87.9%) lo llevaron al número dos, en un vaivén continuo con Indonesia (+29.3%). Vietnam (+52.8%) también envió más, mientras que Tailandia (-3.5%) envió menos después de un mes de ganancias.
Hay mucho que tener en cuenta sobre el establecimiento del registro de data en este informe y, francamente, de todo 2021. En términos de máximos de diciembre, nuestros cuatro principales socios comerciales establecieron una nueva marca. Indonesia envió un récord mensual de 37.361 millones de libras a EE.UU. Anualmente, estos mismos cuatro superaron las cifras del año pasado y lograron su mejor año hasta la fecha. India envió más de 750 millones de libras, un 25.2% más que el año pasado, que casi alcanzó los 600 millones de libras. También destaca Ecuador, que consolidó el segundo lugar con 405.293 millones de libras en el año.
Mirando por el tipo, todas las categorías principales fueron nuevamente más altas que el año pasado. El camarón con cáscara (+38.6%) fue nuevamente impulsado por Ecuador y casi todos los tamaños. También aumentaron las importaciones de camarón pelado (+41.6%) y empanizado (+37.1%). El cocido (+4.6%) también se envió más.
Ciclos de importación mensual por país (todos los tipos)
India: Hubo otro aumento y récord mensual de India (+23.0%), que envió más de 750 millones de libras en importaciones de 2021. Nueve meses consecutivos de máximos para esos períodos ayudaron a establecer el máximo histórico en libras anuales enviadas a los EE.UU. En diciembre se enviaron casi 69 millones de libras desde la India. Se envió más camarón pelado (+37.7%) y cocido (+11.2%), superando al camarón con cáscara (-14.4%).
Indonesia: Si bien las importaciones de camarón de Indonesia (+29.3%) aumentaron significativamente, estableciendo un récord histórico en diciembre de 37.361 millones de libras, no fue suficiente para contener el tonelaje enviado desde Ecuador. Estos dos países importadores han ido de un lado a otro en la clasificación durante gran parte del año. Sin embargo, cuando todo estuvo dicho y hecho, el esfuerzo de este último superó a Indonesia tanto en diciembre como en el calendario. Hubo más envíos de camarón con cáscara (+8.6%), pelado (+69.8%) y empanizado (+168.2%); pero menos del cocido (3.0%).

- FEBRERO 2023
72
Ecuador: El esfuerzo concentrado de Ecuador para hacer de los EE.UU. uno de sus principales socios comerciales brilló a fines de 2021. El total de diciembre ganó un 87.9% con respecto al año pasado a un récord de diciembre de 41.793 millones de libras. Pero la estadística más drástica fue la cifra anual de 405.293 millones de libras para cimentar a este país como el claro número dos. Al principio parecía completamente por necesidad, pero incluso cuando China reanudó las compras, todavía hubo un esfuerzo real para satisfacer a ambos clientes. A medida que aumenta la inversión en infraestructura, aumentan las expectativas para la región.
Vietnam y Tailandia: Vietnam (+52.8%) envió más, superando a los últimos meses mientras que Tailandia (-3.5%) retrocedió a la baja. El primero vio una gran ganancia anual, mientras que el segundo estuvo marginalmente por debajo del año pasado.
Importaciones de camarón con cáscara, cíclicos y por tamaño
Las importaciones de camarón con cáscara, que incluye el tipo pelado fácil, fueron nuevamente más altas (+38.6%) en diciembre. La afluencia de los últimos dos meses ayudó a llevar las importaciones anuales ligeramente por encima (+2.3%) de 2020.
Ecuador (+137.7%) y con menor volumen
Vietnam (+253.9%) lideraron el camino. Pero hubo ganancias de un amplio grupo de países, incluyendo entre otros, Indonesia (+8.6 %), México (+11.9 %), Tailandia (+67.7
%) y Perú (+32.3%). India (-14.4%) fue el país que envió menos. Al igual que el mes pasado, todos los tamaños, excepto el 51-60, que eran marginalmente más bajos y el 61-70, fueron todos más altos que el año pasado.
Los valores de reemplazo (importación $/ lb.) se movieron a la inversa de la ganancia positiva en noviembre. Los precios se vieron presionados por la menor demanda de los consumidores. El precio promedio en diciembre fue de $4.34 por libra, por debajo de los $4.42 por libra.
Valor agregado, importación de camarón pelado
La data de camarón de valor agregado se mantuvo en tendencia. Las importaciones de camarón pelado (+41.6%) fueron nuevamente muy superiores. Los top cuatro fueron nuevamente los que impulsaron esto. India (+37.7%), Ecuador (+47.0%), Indonesia (+69.8%) y Vietnam (+94.6%) superaron al
resto de países en territorio negativo. Los valores de reemplazo (importación $/lb.) para el camarón pelado bajaron de $4.36 por libra a $4.32 por libra.
La importación de camarón cocido (en agua tibia) finalmente se encontró entre los ganadores. Las importaciones de diciembre aumentaron un 4.6%. Las mayores importaciones de India y Vietnam ayudaron a impulsar las ganancias.
Las importaciones de camarón empanizado aumentaron un 37.1% en el mes de diciembre, con Indonesia (+168.2 %) a la cabeza.
Importaciones de camarón cocido, empanizado y otros
A pesar de que las importaciones aumentaron en todos los ámbitos y la demanda general de camarón fue ligeramente menor en diciembre, la idea de facilidad asociativa con
Detalle de importaciones de camarón YTD por tipo


- FEBRERO 2023 73 REPORTE DE MERCADO
Importaciones YTD de todos los tipos de camarón por año de EE.UU. y promedio Importación $/lb.
Fuente: USDOC. Urner Barry
Fuente: USDOC. Urner Barry
REPORTE DE MERCADO
el camarón cocido ayudó a que los precios subieran ligeramente a $5.03 por libra.
Línea de tiempo del precio del camarón; anuncios minoristas

Venta minorista: A medida que las importaciones continuaron siendo récord y los minoristas intentaron capturar iniciativas saludables para comenzar el año, los productos de los minoristas alcanzaron un máximo desde marzo de 2021. Fueron incluso un 17% más altos que el promedio de tres años para enero. Los precios promedio de los productos se desplomaron para impulsar el negocio. Enero promedió $7.51 por libra, por debajo de $8.24 por libra.

Suministro de camarón a EE.UU. y situación del Golfo
Casi todas las áreas, excepto Florida (costa oeste), experimentaron una disminución significativa en los desembarques. El Servicio Nacional de Pesca Marina reportó los desembarques de diciembre de 2021 (todas las especies, sin cabeza) de 4.358 millones de libras, frente a una ganancia positiva de 8.955 millones de libras el año pasado. El total anual de 71.158 millones de libras está significativamente por debajo
de los 102.348 millones de libras del año anterior.
Exportación de camarón ecuatoriano
Un tono firme afloró en el mercado de camarón blanco de origen latino al que se le dio fuerza en origen. Mientras tanto, el sentimiento del mercado para el camarón blanco y el camarón tigre negro importado de origen asiático se mantiene apenas estable/débil ahora que existe un inventario sustancial en el país.
Si bien persisten las preocupaciones logísticas, y ahora que los precios de reemplazo en India se han desconectado del mercado, ambos suelen ser factores de apoyo y los vendedores aún tienen en cuenta los inventarios en el país.
La demanda de camarón se ha desacelerado estacionalmente, pero una ráfaga de demanda minorista ha brindado cierto grado de apoyo•
- FEBRERO 2023

REUNIONES DIPLOMÁTICAS
Visita del Embajador de Cánada
Reunión con jefes de seguridad empresarial

Fortalecer las relaciones comerciales con Canadá y apoyar el proceso de negociación del acuerdo comercial con Ecuador fueron los objetivos a cumplir en la reunión entre el Embajador de Canadá en Ecuador, Stephen Potter, y directivos de la CNA. El diplomático visitó la entidad y fue recibido por la Directora Ejecutiva de la CNA, Yahira Piedrahita, y el Gerente de Comercio Exterior, Daniel Pesantes, quienes le dieron detalles sobre la cadena de valor y analizaron las oportunidades comerciales del camarón ecuatoriano en ese importante destino.

Visita parlamentarios UE
Ecuador recibió la visita de 7 diputados del Parlamento Europeo que conforman la Comisión de Pesca, con el objetivo de conocer los avances del sector pesquero en combatir la pesca ilegal, no declarada y no reglamentada.
En el marco de esta visita, los eurodiputados se reunieron con José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura (CNA); Daniel Pesantes, Gerente de Comercio Exterior de la CNA, y Pamela Nath, Directora de Sustainable Shrimp Partnership, quienes donde pudieron conocer de primera mano las cifras de la industria, su evolución y enfoque en materia de sostenibilidad y trazabilidad.

Adicionalmente se planteó a los europarlamentarios la necesidad de revisar el mecanismo mediante el cual, la UE otorga contingentes sin arancel a la importación de camarón por ser considerada una práctica que genere competencial desleal al camarón ecuatoriano.
Identificar las nuevas zonas donde se han desplazado los delitos, es una prioridad para la Cámara Nacional de Acuacultura, que a través de su director de Seguridad, Luis Herrera, lideró una reunión en la que participaron los directores de seguridad de empresas relacionadas a la cadena de valor del camarón.
La CNA solicitó a las Fuerzas Armadas fortalecer los controles CAMEX (control de armas, municiones y explosivos) en zonas identificadas como de mayor ocurrencia del delito, y que las autoridades de control replanteen sus estrategias para desacelerar los índices de violencia que se registran en corredores viales
Mesa de seguridad productiva
Con el objetivo de ser parte y gestionar acciones en las mesas de seguridad del sector productivo, la Cámara Nacional de Acuacultura, a través de su director de Seguridad, Luis Herrera, participó en el encuentro junto con la Viceministra del Interior, el Ministro de la Producción, Comercio Exterior, Inversiones y Pesca, representantes de la Policía Nacional, Secretaría Nacional de Aduanas del Ecuador (SENAE), la Corporación de Promoción e Inversiones (CORPEI) y demás miembros del sector productivo-exportador.
Durante la reunión se dio a conocer el plan piloto que se desarrolla en la provincia de Los Ríos, en el eje vial que conecta Babahoyo-Quevedo, con el fin de establecer parámetros de atención en seguridad a transportes para replicar en las vías geo-referenciadas.
Se conformaron 3 mesas de atención prioritarias: Catastro de logística, Seguridad de transportes en ejes viales y Extorsión (vacunas).
La CNA solicitó ver la factibilidad de que existan consolas que tengan conectividad directa con la plataforma del Ecu 911 para generar alertas en tiempo real y tener apoyo oportuno de Policía y Marina.
Habilitación sanitaria
La Subsecretaría de Calidad e Inocuidad (SCI) convocó a los representantes de los establecimientos procesadores y exportadores de camarón para darles a conocer el proceso de renovación del registro de habilitación en el sistema CIFER de la Administración General de Aduanas de China.
Por su parte, la Cámara Nacional de Acuacultura, a través de su gerente de Comercio Exterior, Daniel Pesantes, participó en el encuentro, con el objetivo estar bien informado y colaborar como ente coordinador-gestor a nivel público y privado.

- FEBRERO 2023 76 NOTICIAS
Reunión interinstitucional para garantizar el servicio eléctrico en el cantón Durán
Nuevas autoridades del Directorio de la Cámara Nacional de Acuacultura
Período 2023 -2025
José Antonio Camposano, Presidente Ejecutivo de la CNA, participó de una mesa de trabajo para revisar el plan de obras de electrificación 2023 – 2024, con el objetivo de gestionar el abastecimiento de energía eléctrica en el cantón Durán, donde se asientan empresas que pertenecen a la cadena de valor del camarón.

La reunión fue liderada por Enith Carrión, Viceministra de Electricidad y Energía Renovable, y participaron funcionarios de la Corporación Nacional de Electricidad (CNEL EP) y la Corporación Eléctrica del Ecuador (CELEC EP), con el propósito de dar seguimiento a la problemática en Durán, anteriormente denunciada por la CNA.


El 2 de febrero pasado, la Cámara Nacional de Acuacultura (CNA) eligió a sus nuevas autoridades. El ingeniero Marcelo Vélez fue electo Presidente del Directorio de la CNA, el ingeniero Luis Francisco Burgos, primer Vicepresidente, y el ingeniero Fabricio Vargas, segundo Vicepresidente.
El nuevo directorio de la institución se mantendrá en funciones hasta febrero de 2025.
- FEBRERO 2023 NOTICIAS
Alimento balanceado enfocado en la biorremediación
La multinacional BioMar amplió su oferta de alimentos con una nueva categoría de productos. Se trata de una solución de biorremediación para garantizar condiciones ambientales óptimas para los camarones, desde la precría hasta el engorde.
"La salud y el bienestar de los camarones comienzan con el ambiente ideal para que prosperen. Estas condiciones ayudarán a mejorar las tasas de supervivencia y un mayor rendimiento", aseguró Henrik Aarestrup, vicepresidente de LATAM Shrimp & Hatchery, BioMar Group.

“Esta es una extensión natural del programa de salud preventiva SmartCare de BioMar, donde se adopta un enfoque holístico de la salud y el bienestar”, comentó Danny Vélez, gerente general de BioMar Ecuador.
BioMar suministra dietas con probióticos que han sido aprobadas por la Comisión Europea para su inclusión en los alimentos acuícolas desde el 2010.
Nueva tecnología en detección múltiple de patógenos
En el marco de Aqua Expo Santa Elena 2023, Cargill presentó una nueva tecnología para la detección molecular multiplex de patógenos a través de PCR. El sistema puede detectar simultáneamente tres patógenos en un tiempo de respuesta de seis horas, lo que significa un ahorro de hasta el 60% versus la inversión que se realiza normalmente con PCRs tradicionales.

“Con este avance se vería beneficiado el proceso para tomar decisiones a tiempo en cuanto se obtiene un resultado. Esta nueva tecnología apoya las diferentes etapas del cultivo larvario, ya que se puede diagnosticar desde las post larvas, los nauplius, y se pueden determinar inclusive los reproductores que vamos a utilizar para producir esas post larvas, y que al final se obtenga una post larva de calidad, validada y libre de patógenos”, indicó Violeta Medina, líder de sanidad acuícola de Cargill.
Entre los patógenos que pueden ser diagnosticados en el kit que se ha desarrollado están: Mancha Blanca, AHPND (necrosis hepatopancreática aguda) y NHP (hepatopancreatitis necrotizante), pero también se han venido adicionando otros que son de importancia ya que son patógenos emergentes como EHP, y que para el caso del sector larvario, es relevante su diagnóstico.
- FEBRERO 2023 78 NOTICIAS EMPRESARIALES