Butlletí de la
Institució Catalana d’Història Natural
88
(1-4)
Barcelona 2024


88
Barcelona 2024
Barcelona 2024
Editor en cap
Juli Pujade-Villar, Universitat de Barcelona, Facultat de Biologia, Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals (Secció invertebrats), Barcelona.
Coeditors
Albert Masó, Universitat de Barcelona, Facultat de Biologia, Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals (Secció ecologia), Barcelona.
Joan Pino, Centre de Recerca Ecològica i Aplicacions Forestals (CREAF), Universitat Autònoma de Barcelona, Bellaterra.
Llorenç Sáez, Unitat de Botànica, Facultat de Ciències, Universitat Autònoma de Barcelona, Bellaterra.
Amador Viñolas, Consorci del Museu de Ciències Naturals de Barcelona, Laboratori de Natura, Coŀlecció d’artròpodes, Barcelona.
L’edició d’aquest Butlletí ha estat possible gràcies al suport de l’Institut d’Estudis Catalans
Agraïm la coŀlaboració de Florenci Vallès i Sala per la correcció del català.
Figura de la coberta: Flor de Potentilla supina (Rosaceae). Dibuix d’Amador Viñolas.
Aquesta publicació es diposita, per donar compliment a l’Esmena als articles 8, 9, 10, 21 i 78 del Codi Internacional de Nomenclatura Zoològica (ed. 1999), referents a l’ampliació i perfeccionament dels mètodes de publicació en els repositoris en línia Internet arxive (http://www.archive.org) i Biotaxa (http://www.biotaxa.org/index/index), amb enllaços a la pròpia pàgina de la publicació, en el lloc web: https://ichn2.iec.cat/Butlleti_85.htm i https://publicacions.iec.cat/PopulaFitxa.do?moduleName=revistes_cientifiques&subModu leName=&idColleccio=162
Data de publicació online volum 88 (1): 31 de març de 2024. Pàgines: 3-32.
Data de publicació online volum 88 (2): 30 de juny de 2024. Pàgines: 33-124.
Data de publicació online volum 88 (3): 30 de setembre de 2024. Pàgines: 125-148.
Data de publicació online volum 88 (4): 30 de desembre de 2024. Pàgines: 149-204.
Data de publicació volum 88 (1-4): 30 de desembre de 2024.
© Els autors dels articles
Aquesta edició és propietat de la Institució Catalana d’Història Natural (filial de l’Institut d’Estudis Catalans)
Carrer del Carme, 47. 08001 Barcelona
Compost per Amador Viñolas
Imprès a Ediciones Gráficas Rey, SL
ISSN (edició digital): 2013-3987
ISSN (edició impresa): 1133-6889
Dipòsit Legal: B 36100-1974

Els continguts del Butlletí de la Institució Catalana d’Història Natural estan subjectes —llevat que s’indiqui el contrari en el text o en el material gràfic— a una llicència Reconeixement - No comercial - Sense obres derivades 3.0 Espanya (by-nc-nd) de Creative Commons, el text complet de la qual es pot consultar a https://creativecommons.org/licenses/by-nc-nd/3.0/es/deed.ca. Així doncs, s’autoritza el públic en general a reproduir, distribuir i comunicar l’obra sempre que se’n reconegui l’autoria i l’entitat que la publica i no se’n faci un ús comercial ni cap obra derivada.
Factors ambientals clau per a la reproducció del gripau corredor als secans de Mas de Melons
Neus Oromí*, Sebastià Camarasa*, Delfí Sanuy*,**, Marc Fibla*, Eduard Reig*, Albert Montori***
* Escola Tècnica Superior d’Enginyeria Agrària, Departament de Ciència Animal (Fauna Silvestre), Universitat of Lleida, Av. Rovira Roure 177, 25188 Lleida, Catalonia, Spain
** Forest Science and Technology Centre of Catalonia (CTFC), 25280 Solsona, Catalonia, Spain
*** CREAC. Centre de Recerca i Educació ambiental de Calafell. GRENP/Aj. 43882 Calafell. Tarragona. Spain
Autor de correspondència: Neus Oromí Farrús. A/e:
noromi@xtec.cat
Rebut: 09.08.2023; Acceptat: 08.12.2023; Publicat: 31.03.2024
Resum
L’estudi que es presenta analitza el comportament reproductor del gripau corredor (Epidalea calamita) als secans de Mas de Melons. Es va fer el seguiment d’un total de sis punts de reproducció amb característiques diferents –per ex. en l’hidroperíode i en el tipus de substrat–des del 2012 fins al 2020. Els resultats indiquen que el comportament reproductiu del gripau corredor a Mas de Melons està condicionat per la temperatura i la precipitació. El principal factor desencadenant de l’inici de la reproducció sembla ser el règim de precipitacions, que proporciona la suficient humitat al sòl per a l’activitat dels gripaus i els assegura la disponibilitat de punts d’aigua on reproduir-se. Es constata, però, una variació notable en la duració de l’activitat reproductiva dels gripaus –tant en l’inici com en el final de la reproducció– entre basses dins un mateix any. Aquest fet podria ser degut a la variabilitat d’hàbitats i també a l’efecte crida inicial dels primers mascles que comencen a cantar quan troben un lloc on reproduir-se. L’efecte crida podria explicar també la variabilitat en el nombre d’individus totals capturats en els diferents anys en un mateix punt de reproducció. A més, el gripau corredor de la zona de Mas de Melons presenta un tipus de comportament en què la reproducció es produeix en diversos episodis dins un mateix període reproductiu.
Paraules clau: Epidalea calamita, reproducció, zones semiàrides, variables climàtiques.
Abstract
Environmental key factors for the reproduction of the natterjack toad in the drylands of Mas de Melons
The present study analyses the reproductive behaviour of the natterjack toad (Epidalea calamita) in the drylands of Mas de Melons. A total of six different breeding ponds e.g. characterized by hydroperiod or subtract type- were monitored from 2012 to 2020. The results indicate that the reproductive behaviour of the natterjack toad in Mas de Melons is influenced by temperature and precipitation. The main factor determining the start of reproduction season seems to be the rainfall pattern, which provides sufficient moisture in the soil for the activity of the toads and ensures the availability of water points for the reproduction. However, there is a notable variation in the duration of the reproductive activity - at the start and at the end of reproduction- between ponds within the same year. This fact could be due to the variability of habitats and also to the initial call effect of the first males which start singing when they find a place to reproduce. The call effect could also explain the variability in the number of total individuals caught in the different years at the same breeding site. In addition, the natterjack toad in the Mas de Melons area presents a type of behaviour in which reproduction occurs in several episodes within the same reproductive period.
Key words: Epidalea calamita, Reproduction, semiarid zones, climatic variables.
Introducció
Els amfibis, en ser animals ectoterms, depenen directament del medi extern per a la regulació de la seva temperatura corporal, ja que no tenen mecanismes fisiològics efectius destinats a tal fi (Brattstrom, 1963; Hutchinson & Dupré, 1992).
L’estudi de les poblacions d’amfibis en un territori és essencial per conèixer les seves estratègies, especialment sota condicions ambientals limitadores (Sinsch, 1992a). L’estudi dels amfibis en ambients semiàrids –caracteritzats per presentar
llargs períodes amb temperatures elevades, poca pluviometria i manca d’humitat– pot esdevenir clau per tal d’entendre la seva biologia i la seva capacitat d’adaptació a condicions extremes.
El concepte de població fa referència al grup d’individus de la mateixa espècie que es reprodueixen simultàniament en un mateix lloc. Ara bé, en la majoria de les espècies d’amfibis, és difícil definir el concepte de població donada l’estructura metapoblacional que presenten (Sinsch, 1992a). Aquest és el cas del gripau corredor (Epidalea calamita), que utilitza una
N. OROMÍ ET
xarxa de diferents punts d’aigua, sovint temporanis i reproductivament connectats, a causa de la seva capacitat de dispersió (Sinsch et al., 2012). Aquest potencial dispersiu està vinculat a la disponibilitat d’aigua i a altres característiques de l’hàbitat que ocupa (Sinsch et al., 2012). Es considera que el gripau corredor, a l’àrea mediterrània, té un comportament reproductor explosiu (Jakob et al., 2003; Richter-Boix et al., 2007; Tejedo, 1988; Wells, 1977) el qual està molt associat a episodis de pluja (Gómez-Mestre, 2014). Els períodes de sequera seguits per episodis de pluja inciten els mascles a acudir a les basses amb rapidesa, i en molt pocs dies es formen grans concentracions i cors de mascles (Beebee, 1983; Bank & Beebee, 1986; Tejedo, 1988). Cal tenir en compte que Epidalea calamita és un anur abundant en les zones semiàrides de la vall de l’Ebre i presenta algunes diferències biològiques en comparació amb altres poblacions de la mateixa espècie de la resta d’Europa on no ocupen zones semiàrides (Miaud et al., 2000; Leskovar et al., 2006; Sinsch et al., 2007).
Estudis previs realitzats en la mateixa zona, utilitzant radiotransmissors que a més a més mesuraven la temperatura corporal, han evidenciat que l’espècie no presenta limitacions importants relacionades amb aquesta variable i que el seu comportament termoregulador està condicionat principalment per les característiques tèrmiques dels refugis que permeten el manteniment del balanç hídric corporal (Oromi et al., 2010; Oromí, 2011). Aquestes dades indiquen que el factor limitador de l’activitat del gripau corredor en ambients semiàrids és la precipitació i la humitat, i no pas la temperatura. Ara bé, l’inici de la reproducció es produeix a finals d’hivern i a la primavera, moments en els quals les temperatures no són prou elevades per posar en risc l’equilibri hídric dels individus però sí que poden ser prou baixes per a fer que els animals no s’activin. Per això, cal veure quina de les dues variables o quina combinació d’elles és el factor que desencadena l’inici de la reproducció. El present estudi pretén analitzar el comportament reproductor del gripau corredor als secans de Mas de Melons. L’objectiu principal és determinar quin és o quins són els factors ambientals desencadenants de la reproducció i com les variables meteorològiques influeixen en la durada del període reproductor. També es pretén analitzar la variació de la població reproductiva al llarg dels anys i entre els diferents punts de reproducció.
La població de gripau corredor (Epidalea calamita) estudiada està situada a l’espai natural de Mas de Melons-Alfés, entre les comarques del Segrià i les Garrigues (província de Lleida). L’espai es troba inclòs dins la Directiva d’Hàbitats de la Xarxa Natura 2000 (ES0000021). L’àrea està caracteritzada per ser una zona semiàrida en la qual predomina el paisatge agrícola de secà amb guarets i cultius de cereals d’hivern (Conesa et al., 1994). Les dades meteorològiques s’han obtingut dels registres de 2007 a 2021 de les estacions meteorològiques de Castelldans (lat: 41,5291; long: 0,75303), desmantellada l’any 2018, i de Lleida-la Femosa (lat: 41,58462; long: 0,64172), posada en funcionament l’any 2018. Ambdues estacions són molt pròximes a la zona estudiada (Servei Meteorològic de Catalunya - XEMA). En aquest període, la mitjana anual de temperatura ha estat de 14,6 ºC, amb un registre màxim de 43,1 ºC al mes de juny i un registre mínim de -4,8 ºC al mes de gener. La precipitació acumulada mitjana anual ha estat de 357,6 mm, amb un màxim de 499,8 mm l’any 2013 i un mínim de 264,2 mm l’any 2015. El període d’aridesa s’estén de juny a setembre i les pluges es concentren principalment a la primavera i, secundàriament, a la tardor (Fig. 1).
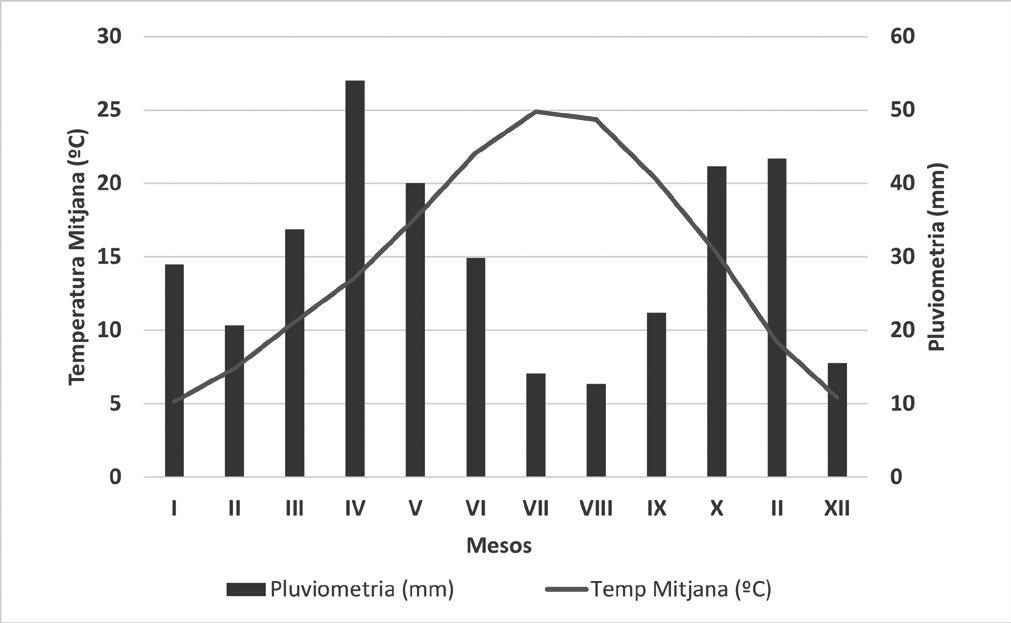
Figura 1. Climograma combinat dels anys 2007 al 2021 de les estacions de Castelldans i Lleida-la Femosa (Servei Meteorològic de Catalunya, 2019).
Taula 1. Coordenades UTM i principals característiques de les diferents basses estudiades. BA: bassa amb substrat principalment argilós formada per un petit canyissar amb marges amb matolls i brolles. CA: tolles d’aigua formades en les depressions que es produeixen en les roques. T: tolla de substrat bàsicament argilós formada per la pluja. L’hidroperíde s’ha determinat seguint el criteri de Richter (2006) i Richter et al. (2007). AO: tòtil (Alytes obstetricans). BS: gripau comú (Bufo spinosus). PC: gripau d’esperons (Pelobates cultripes). PP: granoteta de punts (Pelodytes punctatus). GV: granota verda (Pelophylax perezi).
Nom Latitud Longitud Hidroperíode Descripció Presència d’altres amfibis
Artesa 41°32’53”N 0°41’50”E Permanent
Astor 41°31’26”N 0°40’35”E Temporània/Permanent
BA. El 2015 va ser tapada i va desaparèixer. GV
BA. Serveix d’abeurador de bestiar. AO, BS, GV, PC, PP. Cadolles 41°31’09”N 0°41’12”E Efímera
CA, Aflorament rocallós amb bassots a la matriu rocosa.Circuit 41°32’05”N 0°41’30”E Temporània
BA. Serveix d’abeurador de bestiar. BS, PC, PP. Bassa Nova 41°32’46”N 0°40’19”E Temporània
BA. Serveix d’abeurador de bestiar. BS, PC, PP. Pedrera 41°31’16”N 0°40’09”E Temporània CA, T PC, PP.
Taula 2. Individus capturats en cada bassa al llarg dels anys. (-): sense seguiment. (*): bassa destruïda. (—): sense reproducció
Taula 3. Test post-Hoc de Bonferroni per a SVL dels mascles entre basses i valors estimats de SVL per bassa. *: valors estadísticament significatius. IC: interval de confiança.
Bassa Nova Artesa Circuit Pedrera Cadolles
Nova
Taula 4. Test post-Hoc de Bonferroni per a SVL dels masces entre anys. i valors estimats de SVL per any. *: valors estadísticament significatius. IC: interval de confiança.
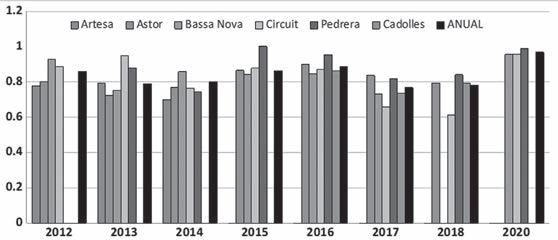
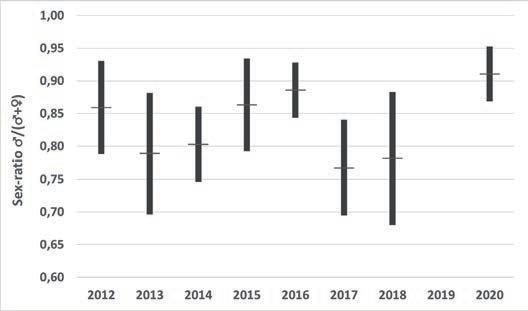
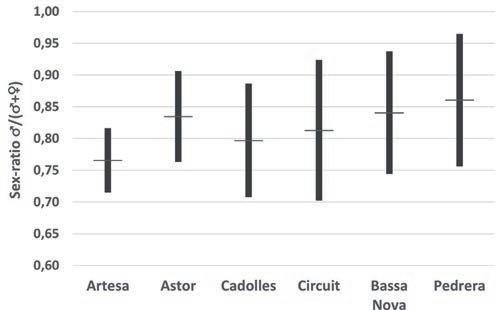
Figura 2. Sex-ratio (♂♂/(♂♂+♀♀)) estimada pels diferents anys de seguiment i les diferents basses. a) per anys i bassa, b) per anys considerant totes les basses com a una unitat i c) per bassa considerant tots els anys junts. En b) i c) línia horitzontal= mitjana, barres negres =mitjana ± desviació estàndard.
El seguiment de les poblacions es va realitzar en un total de sis punts de reproducció de l’espècie. En aquestes localitats, s’inclouen basses de característiques diferents tant des del punt de vista de l’hidroperíode com de l’ús i del substrat (Taula 1). El seguiment de la reproducció del gripau corredor es va realitzar des del 2012 i fins al 2020. Totes les sortides de camp van efectuar-se en dies de pluja o en dies immediatament posteriors (després de la posta de sol), des de les primeres pluges del mes de febrer fins a la finalització del període reproductor. A cada sortida s’anotaven el nombre de mascles i femelles de cada bassa, la longitud corporal (SVL) (amb un calibrador de 0,1 mm de precisió), el nombre d’am-
Taula 5. Anàlisi estadística dels efectes de les variables meteorològiques sobre l’inici de la reproducció. *: variables significatives. TM: temperatura mitjana diària. Tmax: temperatura màxima diària. Tmin: temperatura mínima diària. HRM: humitat relativa màxima diària. HRX: humitat relativa màxima diària. HRN: humitat relativa mínima diària.
Variable DF Chi-quadrat P>Chi-quadrat
TM 1 2,81
Tmax 1 7,54
0,0938
0,006*
Tmin 1 0,66 0,4168
HRM 1 7,87
0,005*
HRX 1 1,43 0,2326
HRN 1 14,64 0,0001*
PPT 1 7,46
0,0063*
plexus observats i la presència de postes, de larves i d’altres espècies d’amfibis. Es va considerar que l’inici de la reproducció tenia lloc quan es produïen els primers cors de mascles i, el final, quan ja no es trobaven femelles a les basses ni se sentien cants. Per a cada bassa i any es va considerar que hi va haver reproducció quan s’observaven postes i/o larves. De cada exemplar capturat se’n va mesurar la longitud corporal o SVL (longitud del musell a la cloaca) amb un peu de rei (precisió de 0,1 mm). La sex-ratio es va estimar calculant la proporció del nombre de mascles respecte el total d’exemplars [♂♂/(♂♂+♀♀)] seguint el criteri de Wilson & Hardy (2002).
Per tal d’analitzar les interaccions del sexe i la mida corporal (SVL) en funció de l’any i la bassa, es va realitzar una anàlisi de variància (ANOVA) amb un test a posteriori de comparació de mitjanes (test HSD de Bonferroni) per com-
parar diferències entre basses i anys en cada sexe i en funció de la SVL. Prèviament es va comprovar que la variable SVL seguia una distribució normal. Per determinar els efectes de les variables meteorològiques sobre la reproducció (dies en que s’inicia la reproducció), es va realitzar una regressió logística (General Lineal Model- Logistic Regression) en què la reproducció es defineix com a una variable depenent qualitativa dicotòmica amb dues categories: reproducció o no reproducció. Per la realització d’aquesta anàlisi es van considerar com a variables independents les variables meteorològiques següents: temperatura mitjana diària, temperatura màxima diària, temperatura mínima diària, humitat relativa mitjana diària, humitat relativa màxima diària i humitat relativa mínima diària). Per determinar la influència de les variables meteorològiques considerades en la durada del període reproductor, es van correlacionar (Rs) aquestes variables amb els dies transcorreguts des que es detecten els primers cors fins que ja no se’n detecten. Per testar la influència de les variables meteorològiques (temperatura màxima, mínima i mitjana; humitat relativa, pressió atmosfèrica i pluviometria) en l’inici del període de reproducció, es van considerar els 20 dies previs a l’inici de la reproducció dividits en quatre períodes de cinc dies, corresponents a 1-5, 5-10, 11-15 i 16-20 dies anteriors a l’inici de la reproducció. En cada un d’aquests períodes es va estimar el valor mitjà de cada variable i l’anàlisi es va realitzar mitjançant una ANOVA Kruskal-Wallis utilitzant el període com a variable d’agrupació. En l’anàlisi univariant es va utilitzar una prova no paramètrica perquè algunes variables no complien la condició de normalitat (test de KolmogorovSmirnov). Totes les anàlisis es van realitzar mitjançant els paquets estadístics Statistica 10.0 i JMP 9.0.1. (SAS Institute, 2011) amb un nivell de significació alfa=0,05.
Taula 6: ANOVA de Kruskal-Wallis entre períodes de 5 dies previs a l’inici de l’activitat reproductora. Només es mostren les variables significatives (en negreta). Es mostren les anàlisis creuades entre setmanes (estadístic z i valor p).
Comparacions múltiples z i p valors entre períodes
Kruskal-Wallis test: H (3, N= 32), p Període de 5 díes 2 3
Resultats
Des de 2012 fins a 2020 es van realitzar 57 sortides de camp. Es varen capturar un total de 6417 individus, 5340 mascles i 1077 femelles (Taula 2). El nombre d’individus capturats va variar considerablement entre anys i entre els diferents punts de reproducció.
En la figura 2 s’observa com la sex-ratio varia al llarg dels anys i entre basses, i que els mascles són molt més nombrosos que les femelles. La sex-ratio estimada per al conjunt de basses i anys és de 0,819, amb una desviació estàndard de 0,072. La màxima proporció de mascles es va donar a la bas-
sa de la Pedrera, amb un 0,953 l’any 2016, i la mínima a la bassa d’Artesa l’any 2014, amb una proporció de 0,700. En totes les basses i tots els anys sempre el nombre de mascles va ser molt superior al de femelles.
El nombre d’individus és molt variable entre anys i basses. A la figura 3 podem observar el nombre d’individus capturats de 2012 a 2020, relativitzats al nombre de campanyes realitzades en els diferents punts de reproducció estudiats. Si analitzem les dades sense tenir en compte el 2012 (any amb menys recursos i efectivitat en la captura d’individus) i els dos darrers anys (2019 i 2020), en els quals el nombre d’individus reproductors va ser molt baix (conseqüència de la forta
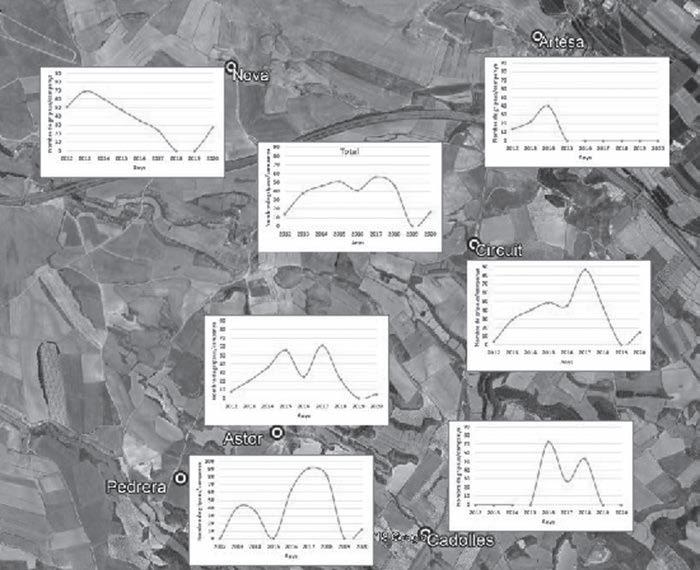
Figura 3. Mitjana d’individus capturats per campanya, any i bassa de 2012 a 2020. Cada gràfic correspon a la bassa que té el nom més proper a ell, excepte en el gràfic central superior (Total), on es representa el nombre de gripaus totals capturats per campanya al llarg dels anys i per a tots els punts d’aigua en conjunt.
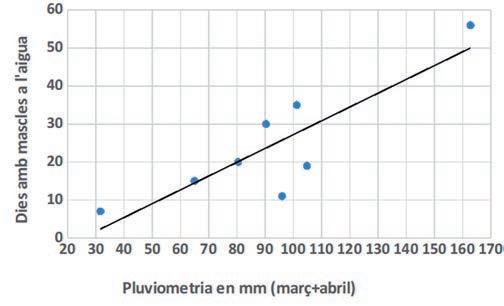
Figura 4. Relació entre la durada de la reproducció (dies amb mascles a l’aigua) i la pluviometria acumulada dels mesos de març i abril per a cada any.
sequera que va patir la zona a l’hivern i principis de primavera), el nombre d’individus/campanya a cada bassa mostra una dinàmica en dents de serra força clara al llarg dels anys a les diferents basses.
L’anàlisi de variància realitzat indica que la mida corporal (SVL) varia significativament entre sexes (F1, 1767=13,237, p=0,0003). Les femelles (77,5 +/- 7,37 mm) són en general més grosses que els mascles (75,41 +/- 6,35 mm). S’han trobat diferències significatives en la longitud del cos dels mascles, entre punts d’aigua (F(5, 1513)=27,971, p<0,001) i entre anys (F(7, 1511)=23,623, p<0,001), però això no ha succeït en el cas de les femelles (ni entre basses [F4, 245=2,0367, p=0,0899] ni entre anys [F7, 242=1,6334, P=0,1266]). Ara bé, el test post-
Hoc de Bonferroni mostra que aquestes diferències trobades en els mascles no es donen entre totes les basses (Taula 3) ni entre tots els anys (Taula 4).
Com es pot observar a la Taula 3, la Bassa Nova i la del Circuit són les basses més diferenciades de les altres. La Bassa Nova, que té hidroperíode llarg, mostra diferències significatives amb les tres basses més temporànies.
Pel que fa a la reproducció, la data més primerenca en la qual es va detectar reproducció va ser el 3 de març (2020) i la més tardana el 30 d’abril (2013). Els resultats de l’anàlisi de l’efecte de les variables meteorològiques sobre l’inici de la reproducció mostren que els factors que tenen un efecte significatiu són la temperatura màxima, la humitat relativa mitjana i la mínima, i la precipitació (Taula 5).
S’ha estimat que els mascles són presents a les basses una mitjana de 20,9 dies, a la qual correspon una desviació estàndard de 17,1 dies. En aquest sentit, s’ha obtingut una correlació positiva i significativa entre el nombre de dies amb mascles a l’aigua cantant i la pluviometria de març i abril (Fig.4, R=0,857; R2=0,748; P=0,0066). El nombre mínim de dies de presència de mascles a les basses va ser de 7 dies, l’any 2015, i el màxim va ser de 56, l’any 2013.
En les basses més permanents, sempre que hi ha hagut reproducció, s’ha completat el cicle de desenvolupament de les postes fins a la metamorfosi. Els punts amb major variabilitat reproductiva han estat les basses temporànies: Cadolles, Pedrera, Circuit i, alguns anys, Bassa Nova. En el cas de les Cadolles, que és un punt d’aigua molt temporani (efímer) que s’asseca cada any, en cap dels anys de seguiment s’ha observat que els capgrossos es desenvolupessin fins a completar la
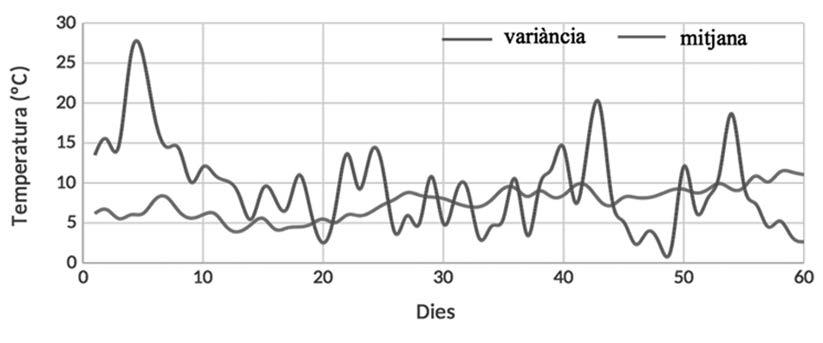
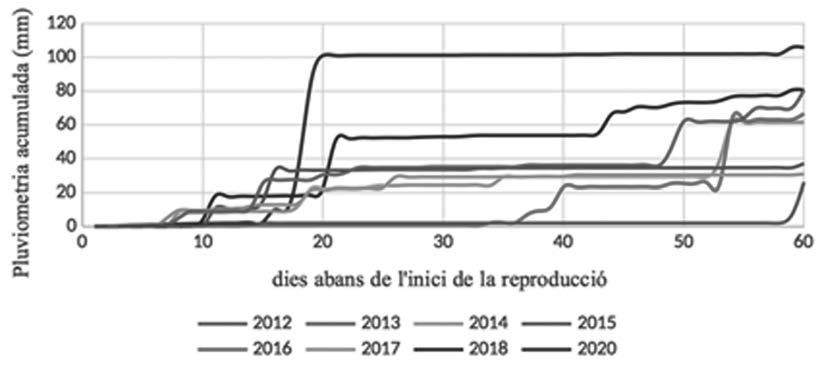
Figura 5. a) Evolució de les temperatures mínimes interanuals. En taronja: temperatura mínima diària. En blau:. variància diària interanual de la temperatura mitjana. b) Evolució de la pluviometria acumulada 60 dies abans de l’inici de la reproducció. En tots dos casos, el dia 60 correspon al primer dia amb cors de mascles a les basses.
metamorfosi. S’observa també que hi han hagut anys sense reproducció en aquest punt, així com en el de la Pedrera. Els resultats obtinguts en el test ANOVA Kruskal-Wallis aplicat a períodes de cinc dies previs a l’inici de l’activitat reproductora relacionats amb les variables meteorològiques considerades es mostren a la Taula 6. Només la temperatura mínima i el nombre de dies amb temperatures mínimes sota zero són significatius. Ara bé, el test de múltiples comparacions indica que només existeixen diferències significatives entre el primer i tercer període de cinc dies previs a l’inici de la reproducció. En la figura 5, s’observa l’evolució de les temperatures mitjanes i la variància interanual dels 60 dies previs a l’inici de la reproducció. Tots els anys excepte dos l’inici de la reproducció coincideix amb episodis de pluja el mateix dia. En els altres dos casos va ploure el dia anterior.
Discussió
La població reproductiva de Mas de Melons és nombrosa malgrat les limitacions propiciades per l’aridesa de la zona. La tipologia d’hàbitats amb sòls argilosos i pedregosos i la climatologia probablement limiten l’establiment de pobla-
(1), 2024
cions abundants d’altres espècies d’amfibis, més lligades a punts de reproducció permanents o a ambients més humits. Per aquest motiu, el gripau corredor és el més abundant a la zona i no sembla presentar, de moment, competidors importants. Els pocs punts d’aigua presents a la zona i l’elevada capacitat de dispersió de l’espècie (Sinsch et al., 2012) fan que es concentrin molts individus en un mateix punt de reproducció.
El nombre de mascles comptabilitzats a les basses és major que el de femelles, amb una sex-ratio significament elevada i desviada cap als mascles. Aquesta tendència és característica de l’espècie, en la qual els mascles són més abundants i romanen durant més temps a les basses que les femelles i, a diferència d’elles, es podent aparellar més d’un cop durant un mateix període reproductiu (Miaud et al., 2000; Sinsch, 1992a; Tejedo, 1992). Si ens fixem en els individus capturats, el nombre varia al llarg dels diferents anys de mostreig en cada bassa (Taula 2). S’observa també una estabilitat en la dinàmica del nombre d’individus totals capturats per campanya des de 2013 fins a 2018 (Fig. 3, Total), que indica que l’ocupació d’un determinat punt d’aigua per part dels mascles depèn d’altres factors que no són la pluviometria o la temperatura. Probablement, de la mateixa manera que succeeix en altres
espècies d’amfibis (Sinsch, 1992b; Diego-Rasilla & Luengo, 2004), l’efecte crida que té el cant dels primers mascles de gripau corredor que arriben a les basses serveix d’orientació per a altres mascles. No obstant, el comportament reproductiu sembla deure’s a un patró en què tant la precipitació com la temperatura juguen un paper important. Quan les temperatures son càlides, amb mínimes mitjanes superiors als 10 ºC, i hi ha precipitació, es desencadena la reproducció (Fig. 5), la qual té lloc bàsicament entre març i abril. En tots els anys en què s’ha constatat que hi ha hagut reproducció sempre s’han superat els 20 mm de pluviometria acumulada (Fig. 5).
S’han observat diferències significatives en la mida dels mascles entre basses i anys, i també existeix una variabilitat en relació amb altres poblacions de l’àrea de distribució de l’espècie (Oromí, 2011). La variabilitat en la mida dels individus podria respondre a la variació de l’estructura d’edats de l’espècie al llarg dels anys. Estudis previs han analitzat els diferents mecanismes que poden determinar la variació de la mida de l’espècie al llarg del seu gradient latitudinal (Sinsch et al., 2010). Sembla que la mida corporal depèn del tipus de sòl i de la durada del període d’activitat. Si analitzem els estudis previs realitzats en l’espècie en la zona de Mas de Melons, que s’han basat en la descripció del seu comportament termoregulador i en la seva capacitat de dispersió i de creixement ossi (Oromí, 2011), s’observa un efecte notable de la durada del període d’activitat anual sobre el creixement dels individus, especialment durant el primer any de vida (Sinsch et al., 2010).
Els estudis sobre el comportament termoregulador de l’espècie en la zona evidencien que el factor determinant d’aquest comportament és el manteniment de la humitat en els refugis utilitzats durant la hibernació i els períodes de sequera (Oromí et al., 2010, 2011). Probablement, l’inici de la reproducció en les basses temporànies, és a dir, que s’assequen durant l’any, ve desencadenat per la formació de tolles per pluja o per la saturació i entollament de les basses (Cano & Cano, 2015). En aquest sentit, cal tenir en compte que no tots els individus es reprodueixen al mateix temps, sinó que ho fan en forma de pics reproductius que coincideixen amb les pluges. Això explicaria perquè, els anys més plujosos, la durada del període reproductor és més gran a la zona estudiada. Des que es van començar a estudiar les poblacions de gripau corredor de la zona de Mas de Melons (2004), alguns anys no s’ha detectar reproducció de l’espècie (observació personal NO i DS). Concretament, al llarg dels anys de seguiment d’aquest estudi, els gripaus no van reproduir-se durant el 2019. Aquest any, la primavera va ser seca i va haver-hi poques precipitacions durant els primers mesos de l’any. La precipitació acumulada de gener a març va ser només de 14,3 mm. En el 2020, la precipitació acumulada durant els 7 o els 15 dies abans de la reproducció va ser molt baixa (5,1 mm). Encara que la precipitació acumulada dos mesos abans va ser superior als 100 mm (Fig. 5), la reproducció no es va iniciar fins que les temperatures no van augmentar i es van estabilitzar. Aquesta combinació dels dos factors, temperatura i pluviometria, ens indica que l’inici de la reproducció del gripau corredor no ve únicament determinat per la pluviometria i
que ambdues variables determinen l’inici de la reproducció. De fet, són les temperatures mínimes i la manca de dies amb temperatures mínimes sota zero, associades a pluges en les 24 hores prèvies, els factors que s’han observat com a més importants en el desencadenament de la reproducció. Per altra banda, com s’observa a la figura 5a els dies previs a l’inici de la reproducció les temperatures s’estabilitzen (menys variància) amb mínimes per sobre dels 5ºC. Quan les temperatures van començar a ser més altes, amb una petita pluja n’hi va haver prou perquè s’iniciés la reproducció, ja que la humitat en del sòl era elevada i hi havia presència de punts d’aigua per reproduir-se. Aquesta mateixa situació es va observar durant els anys 2022 i 2023, que van ser especialment secs. La duració de l’activitat reproductora i la permanència de mascles als punts de reproducció estan relacionades amb la pluviometria acumulada en els mesos de març i abril per a cada any, i presenten major variabilitat en les basses temporànies. Les característiques d’aquests punts de reproducció, bàsicament tolles formades a causa de les irregularitats de les roques, fan que tinguin poca capacitat i que l’aigua s’hi evapori en poc temps, la qual cosa dificulta l’èxit reproductiu. Tot i que les basses són pròximes entre elles, alguns anys presenten variació en l’inici i el final de l’activitat reproductiva dels gripaus. Probablement, la variabilitat en l’hàbitat i les condicions ambientals entre els punts de reproducció (Aubry et al., 2012) poden explicar aquesta asincronia, però per altra banda podria ser deguda a l’efecte crida inicial dels primers mascles que comencen a cantar en trobar un punt d’aigua idoni on reproduir-se (Sinsch, 1992b; Diego-Rasilla & Luengo, 2004). La reproducció de les poblacions de gripau corredor de la zona es produeix a la primavera, a diferència d’altres poblacions, com les del Garraf i les de Collserola, que poden reproduir-se a la tardor (Richter-Boix, 2006). A més, la reproducció, tot i que és explosiva, sembla que es produeix en diversos episodis dins un mateix període reproductiu. Aquest tipus de comportament s’ha descrit també en altres poblacions, com les de Doñana (Díaz-Paniagua et al., 2005).
Estem molt agraïts als molts companys i companyes que de forma més o menys puntual ens van ajudar en les sortides de camp. Els permisos anuals per a la captura d’exemplars de gripau corredor van ser donats pel Servei de Fauna de la Generalitat de Catalunya.
Aubry, A., Bécart, E., Davenport, J., Lynn, D., Marnell, F., & Emmerson, M. C. 2012. Patterns of synchrony in natterjack toad breeding activity and reproductive success at local and regional scales. Ecography, 35: 1-11.
Banks, B., & Beebee, T. J. C. 1986. Climatic effects on calling and spawning of the natterjack toad Bufo calamita: discriminant analyses and applications for conservation monitoring. Biological conservation, 36: 339-350.
Beebee, T. J. C. 1983. The natterjack toad. Oxford University Press Brattstrom, B. H. 1963. A preliminary review of the thermal requirements of amphibians. Ecology, 44: 238-255.
Cano Barbacil, C., & Cano Sánchez, J. 2015. La lluvia, la charca y el sapo corredor, una relación de supervivencia en ambientes semiáridos. Agencia Estatal de Meteorologia. p. 262-269
Conesa, J. A., Moyoral, A., J., P., Recasens, J., & Pedro, J. 1994. The vegetables landscape of the space of Lleida’s natural interest: Southern area. Ed. I.E.I Diputació de Lleida Díaz-Paniagua, C., Gómez-Rodríguez, C., Portheault, A., & De Vries, W. 2005. Los anfibios de Doñana. Ed. Naturaleza y Parques Naturales. Serie Técnica. 181 p. Diego-Rasilla, F. J., & Luengo, R. M. 2004. Heterospecific call recognition and phonotaxis in the orientation behavior of the marbled newt, Triturus marmoratus. Behavioral Ecology and Sociobiology, 55: 556-560
Gómez-Mestre, I. 2014. Sapo corredor. Enciclopedia Virtual de los Vertebrados Españoles, 29 p. Hutchinson, V. H. & Dupré, R. K. 1992. Thermoregulation. Environmental physiology of the amphibians. Ed by M.E. Feder and W.W. Burggren, University of Chicago Press, Chicago, pp. 206-249.
Jakob, C., Poizat, G., Veith, M., Seitz, A., & Crivelli, A. 2003. Breeding phenology and larval distribution natterjack toads (Bufo calamita) vary between northern and sourthen latitudes. Hydrobiologia, 499: 51-61.
Leskovar, C., Oromi, N., Sanuy, D., & Sinsch, U. 2006. Demographic traits of reproductive Natterjack toads (Bufo calamita) vary between northern and southern latitudes. Amphibia-Reptilia, 27: 365-375.
Miaud, C., Sanuy, D., & Avrillier, J. N. 2000. Terrestrial movements of the natterjack toad Bufo calamita (Amphibia, Anura) in a semiarid, agricultural landscape. Amphibia-Reptilia, 21: 357-369.
Oromí, 2011. Latitudinal and altitudinal variation of life history traits in natterjack toads (Bufo calamita): genetic adaptation vs phenotypic plasticity. Doctoral Thesis. http://hdl.handle. net/10803/51584
Oromí, N., Camarasa, S., González, E., Alàs, J., Pavillard, M., & Sanuy, D. (2011). Comportament termoregulador del gripau corredor (Bufo calamita) en una zona semiàrida. Butlletí de la Institució Catalana d’Història Natural, 76: 95-104.
Oromi, N., Sanuy, D., & Sinsch, U. 2010. Thermal ecology of natterjack toads (Bufo calamita) in a semiarid landscape. Journal of Thermal Biology, 35: 34-40.
Richter-Boix, A. (2006). Structure of an anuran community in a Mediterranean area. Tesi doctoral. Universitat de Barcelona. 232 pàg. (http://www.tdx.cesca.es/TDX-0420106- 083313/).
Richter-Boix, A., Llorente, G. A., & Montori, A. 2006. Breeding phenology of an amphibian community in a Mediterranean area. Amphibia-Reptilia, 27: 549-559.
Richter-Boix, A., Llorente, G. A., & Montori, A. 2007. Hierarchial competition in pond-breeding anuran larvae in a Mediterranean area. Amphibia-Reptilia, 28: 247-261.
SAS Institute, 2011. JMP 9.0.1. Statistical Discovery. From SAS. USA: Author. A Business Unit of SAS Campus Drive Cary, NC 27513.
Sinsch, U., Oromi, N., Miaud, C., Denton, J., & Sanuy, D. 2012. Connectivity of local Bufo calamita populations: modelling the migratory range of radio-tracked toads. Animal Conservation, 15: 388–396
Sinsch, U. 1992a. Structure and dynamic of a natterjack toad metapopulation (Bufo calamita). Oecologia, 90: 489-499.
Sinsch, U. 1992b. Sex-biassed site fidelity and orientation behaviour in reproductive natterjack toads (Bufo calamita). Ethology Ecology & Evolution, 4:15-32
Sinsch, U., Marangoni, F., Oromi, N., Leskovar, C., Sanuy, D., & Tejedo, M. 2010. Proximate mechanisms determining size variability in natterjack toads. Journal of Zoology, 281:272-281.
Sinsch, U., Oromi, N. & Sanuy, D. 2007. Growth marks in Natterjack Toad (Bufo calamita) bones: histological correlates of hibernation and aestivation periods. Herpetological Journal, 17: 129-137.
Tejedo, M. 1988. Fighting for females in the toad Bufo calamita is affected by the operational sex ratio. Animal Behaviour, 36: 1765-1769.
Tejedo, M. 1992. Large male mating advantage in the natterjack toads, Bufo calamita-sexual selection or energetic constraints. Animal Behaviour, 44: 557-569.
Wells, K. F. 1977. The social behaviour of anuran amphibians. Animal Behaviour, 25: 666-693.
Wilson, K., & Hardy, I.C.W. 2002. Statistical analysis of sex ratios: an introduction. In: Hardy, I.C.W. (ed.): Sex ratios – concepts and research methods. Cambridge University Press, Cambridge. 2002, 48-92.
Butlletí de la Institució Catalana d’Història Natural, 88 (1): 11-12. 2024 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
Potentilla supina (Rosaceae) a Catalunya (NE de la península Ibèrica)
Potentilla supina (Rosaceae) in Catalonia (NE Iberian Peninsula)
Jordi Bassols Garcia* & Josep Gesti Perich**
* C/ Gran de Sant Andreu, 310. 08030 Barcelona. A/e: jbassolsg@gmail.com
** C/ Sant Ignasi, 42. 17430 Santa Coloma de Farners.
Autor de correspondència: Jordi Bassols. A/e: jbassolsg@gmail.com
Rebut: 14.12.2023. Acceptat: 11.01.2024. Publicat: 31.03.2024
Potentilla supina L. subsp. supina
Osona: Vic, les Casasses, al marge de la bassa permanent que hi ha vora el riu Gurri, 31TDG4045, 450 m, herbassar humit, 11-XI-2023, J. Bassols (Fig. 1).


Potentilla supina és una rosàcia de distribució principalment holàrtica i naturalitzada en altres regions, com ara Austràlia (POWO, 2023), que es caracteritza pel seu port prostrat, per les fulles pinnatisectes (com a mínim les basals), per les flors axil·lars solitàries i per la mida dels pètals, que són


J. BASSOLS GARCIA & J. GESTI PERICH
petits (2-3 mm). Alguns autors hi diferencien diversos tàxons subespecífics, d’entre els quals només la subsp. supina s’ha trobat a l’Europa occidental (Soják, 1987, 1993).
A la península Ibèrica s’ha indicat sobretot a les àrees central i oriental (Sánchez-Gómez & Güemes, 1994; Guillén & Rico, 1998; Benito et al., 1999; Valdés et al., 2001; López et al., 2016), per bé que durant els darrers 30 anys també s’ha anat trobant esparsament a l’oest (Ladero et al., 1995; Bariego & Rico, 2003), al nord (Sáenz et al., 2012) i al sud peninsulars (Martínez-Sagarra et al., 2019; Fuentes et al., 2020). L’aparició d’aquestes noves localitats, sovint ubicades en herbassars higro-nitròfils de marges d’embassaments o d’altres espais alterats i temporalment inundats, sembla indicar que l’espècie es troba en expansió al territori peninsular. Pel que fa a Catalunya, no tenim constància de cap citació prèvia; les localitats més pròximes són les de la Plana Alta, a Castelló (BDBCV, 2023), i les del Llenguadoc (INPN, 2023).
A les Casasses hem observat un sol individu de P. supina subsp. supina (Fig. 1) creixent en un herbassar humit sobre terreny llimós-argilós, al marge d’una bassa de règim permanent (alimentada majoritàriament per les aigües del freàtic), de nova construcció. Al seu voltant les espècies més abundants eren Atriplex prostrata DC., Cyperus eragrostis Lam., C. fuscus L., Dysphania ambrosioides (L.) Mosyakin & Clemants, Juncus articulatus L. subsp. articulatus, Lycopus europaeus L., Persicaria maculosa Gray, Ranunculus sceleratus L. i Sonchus asper (L.) Hill subsp. asper. Es trobava en plena floració i presentava abundants llavors a les flors marcides.
Desconeixem el vector de dispersió que va causar l’arribada d’aquesta espècie a la localitat, tot i que creiem que probablement va ser algun dels ocells migradors que la freqüenten i poden transportar llavors.
Tot i així, cal destacar que la zona ha estat recentment sotmesa a moviments de terra lligats als treballs de restauració ambiental que s’hi van dur a terme entre els mesos de novembre de 2022 i febrer de 2023, els quals han comportat la creació de la bassa i la plantació d’espècies autòctones. Per tant, no es pot descartar la possibilitat que les llavors d’aquesta espècie puguin haver arribat accidentalment a causa de les plantacions realitzades, o bé transportades per la maquinària d’obra. A l’entorn més proper de l’individu localitzat es van plantar les següents espècies: Alisma plantago-aquatica L., Eleocharis palustris (L.) Roem. & Schult., Limniris pseudacorus (L.) Fuss, Scirpoides holoschoenus (L.) Soják i Sparganium erectum L. Totes elles procedien d’un viver situat a la comarca del Vallès Occidental, on no tenim constància que es cultivi P. supina
Bariego, R. & Rico, E. 2003. Novedades florísticas para la Cuenca del Duero (España). Lazaroa, 24: 129-130.
BDBCV. 2023. Banc de Dades de la Biodiversitat de la Comunitat Valenciana. Generalitat Valenciana. Disponible a: www.bdb.gva. es [Data de consulta: 7 desembre 2023].
Benito, J. L., Martínez, J. M. & Pedrocchi, C. 1999. Potentilla supina L. (Rosaceae) reencontrada en Aragón un siglo después. Flora Montiberica, 11: 6-8.
Fuentes, J., Sánchez, R., Segura, J. M., Cueto, M., Ramírez, J. & Gutiérrez, L. 2020. Novedades corológicas destacables para la flora vascular de Andalucía, (sur de España) IV. Anales de Biología, 42: 63-73.
Guillén, A. & Rico, E. 1998. Potentilla. L. P. 96-140. In: Muñoz, F.; Navarro, C. (eds.). Flora iberica VI - Rosaceae. Real Jardín Botánico. CSIC. Madrid. 592 p. INPN. 2023. Inventaire national du patrimoine naturel. Muséum national d’Histoire naturelle. Office français de la biodiversité. Disponible a: https://inpn.mnhn.fr [Data de consulta: 7 desembre 2023].
Ladero, M., Amor, A., Pérez Chiscano, J. L. & Santos, M. T. 1995. Algunas plantas interesantes de la flora extremeña. Studia Botanica,14: 203-206.
López, S., Fabregat, C., Fabado, J. & Picornell, E. 2016. Nuevos datos para la flora de Aragón, II. Flora Montiberica, 64: 20-25. Martínez-Sagarra, G., Reyes, E., López-Tirado, J. & Devesa, J. A. 2019. Novedades florísticas para la provincia de Córdoba (España). II. Acta Botanica Malacitana, 44: 113-115. POWO. 2023. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Disponible a: http://www. plantsoftheworldonline.org [Data de consulta: 7 desembre 2023].
Sáenz, J. A., Aran, V. J., Barbadillo, P., Bariego, P., Barredo, J. J., Benito, J., Escalante, M. J., García-López, J. M., Marín, L., Mateo, G., Molina, C., Montamarta, G., Pérez, J. M., Patino, S., Pinto, M. Á. & Valencia, J. 2012. Anuario Botánico de Burgos, I. Adiciones y revisiones al atlas de la flora vascular silvestre de Burgos 2007-2012. Aula de MedioAmbiente de la Caja de Burgos. Burgos. 195 p.
Sánchez-Gómez, P. & Güemes J. 1994. Acerca de Potentilla supina L. en la Península Ibérica. Acta Botanica Malacitana, 19: 209210.
Sójak, J. 1987. Notes on Potentilla paradoxa and P. supina. Preslia, 59: 271-272.
Sójak, J. 1993. Taxonomische Bemerkungen zu einigen mediterranen Potentilla-Sippen. Preslia, 65: 117-130.
Valdés, A., Alcaraz, F. & Rivera, D. 2001. Catálogo de las plantasvasculares de la provincia de Albacete (España). Instituto de Estudios Albacetenses “Don Juan Manuel”. Serie I-Estudios, 127. Albacete. 304 p.
* Carrer Villar, 34-36. 08041 Barcelona. A/e: margaridagenera9@gmail.com
Rebut: 06.12.2023; Acceptat: 12.03.2024; Publicat: 31.03.2024
Resum
A través d’aquest article ens proposem retre un petit homenatge a la figura de Jordi Sabater Pi, per totes les seves aportacions relacionades, sobretot, amb l’evolució humana i precedents, no tan sols en l’àmbit estrictament de la recerca, sinó també en el de la sensibilització pel patrimoni natural. També per llurs qualitats com a gran divulgador científic. Una de les seves aportacions més innovadores fou la constatació que la cultura no és exclusiva dels humans, ampliant així el concepte de «fet cultural». Les seves recerques han demostrat que els ximpanzés no solament són capaços d’usar «bastons» per a atrapar tèrmits del termiter, sinó que, a més, transmeten llurs coneixements de generació en generació. Per tant, l’hem de considerar com el principal introductor de l’etologia al nostre país. Un altre aspecte remarcable que el distingeix, entre molts altres, és la seva habilitat pel dibuix, emprat com el llenguatge universal que permet no tan sols enregistrar, documentar i descriure determinats elements amb tot rigor, sinó també transmetre continguts de gran complexitat, de forma inteŀligible a la societat en general i impregnant-los de sentiments profunds del propi autor. El nostre objectiu, és, doncs, –encara que sigui a grans pinzellades– recordar algunes de les accions més rellevants del Dr. Jordi Sabater Pi en el camp de la ciència i de l’art, que han contribuït a avançar en els coneixements sobre els nostres orígens i a fomentar el sentiment d’estima per la preservació i conservació de l’entorn natural, lligat inexorablement amb l’emergència i el desenvolupament de la nostra espècie. Incloem també alguns episodis vitals que ens varen vincular personalment i van enfortir la nostra amistat.
Paraules clau: dibuix, primatologia, etologia, prehistòria, evolució, «fet cultural», Ikunde, historiografia, Lluís de Lassaletta.
Abstract
Jordi Sabater Pi (1922-2009). Remembering the figure of an excellent naturalist of our country
Through this article we intend to pay a small tribute to the figure of Jordi Sabater Pi, for all his contributions related, above all, to human evolution and precedents, not only in the strictly field of research, but also in the awareness of natural heritage. Also, for his qualities as a great science communicator. One of his most innovative contributions was the confirmation that culture is not exclusive to humans, thus expanding the concept of «cultural fact», His research has shown that chimpanzees are not only capable of using «sticks» to trap termites, but also transmit their knowledge from generation to generation. Therefore, we must consider him as the main introducer of ethology in our country. Another remarkable aspect that distinguishes him, among many others, is his ability to draw, used as the universal language that allows not only recording, documenting and describing certain elements with great rigor, but also transmitting contents of great complexity, in an intelligible way to society in general, impregnating them with deep feelings of the author himself. Our goal, therefore, is –albeit in broad brushstrokes– to recall some of the most relevant actions of Dr. Jordi Sabater Pi in the field of science and art, which have contributed to advancing knowledge about our origins and promoting the feeling of appreciation for the preservation and conservation of the natural environment, inexorably linked to the emergence and development of our species. We also include some vital episodes that linked us personally, thus strengthening our friendship.
Key words: drawing, primatology, ethology, prehistory, evolution, «cultural fact», Ikunde, historiography, Lluís de Lassaletta.
Introducció
Amb aquest text breu, però, molt ric en sentiments, ens sumem als actes que s’han anat celebrant, al llarg de l’any 2022, en commemoració del centenari del naixement del Dr. Jordi Sabater Pi. L’hem escrit, sobretot, en reconeixement a les seves aportacions relacionades amb l’evolució humana, no tan sols en l’àmbit estrictament de la recerca, sinó també en el de la sensibilització vers el patrimoni natural. El nos-
tre propòsit és, encara que sigui a grans pinzellades, recordar algunes de les accions més rellevants d’aquest investigador en els camps de la ciència, de l’art i de la socialització patrimonial, ja que –sens dubte– han contribuït, a fomentar el sentiment d’estima envers la preservació i conservació de l’entorn natural, lligat inexorablement amb l’emergència i el desenvolupament de la nostra espècie.
La recollida d’informació ha consistit a cercar les dades biogràfiques més significatives i d’alguns documents que ens
M. GENERA I MONELLS
confirmen episodis coneguts a partir de les pròpies vivències i records. Doncs, d’una banda jo havia residit –encara que per poc temps– amb els meus pares i les dues germanes, a la ciutat de Bata, Guinea equatorial, entre els anys 1955 i 1956, aleshores colònia espanyola. Molt temps després, vaig tenir el privilegi de conèixer personalment a Jordi Sabater i la seva muller Núria Coca i compartir, durant alguns anys, inquietuds professionals, amics i veïns del domicili de Barcelona.
Pel que fa als dibuixos, ens basem sobretot en la bibliografia fins ara publicada a més d’accedir a les principals colleccions que contenen el seu llegat. La més important és la que es troba dipositada en el CRAI (Centre de Recursos per a l’Aprenentatge i la Investigació) de la Facultat de Belles Arts de la Universitat de Barcelona, i n’hi ha una altra, molt més reduïda numèricament però no menys interessant, que es conserva en l’Arxiu del Museu Etnològic de Barcelona, ara Museu de les Cultures del Món.
Aquest estudi comprèn cinc apartats que es desglossen tot seguit.
La figura de Jordi Sabater Pi: algunes dades biogràfiques
Des del seu naixement a Barcelona, el 2 d’agost de 1922, fins al seu decés a la mateixa ciutat, el 5 d’agost de l’any 2009, distingim quatre grans etapes de la seva vida, tot i que aquí només ens referirem a les dues centrals, perquè són les que comprenen la seva activitat professional i científica que vàrem conèixer directament (Fig. 1).
Jordi Sabater, l’any 1940, va viatjar a l’illa de Fernando Poo, avui Malabo, amb la finalitat de treballar com a capatàs en una plantació familiar. Aquesta circumstància li permeté entrar en contacte directe, fins i tot de forma immersiva, en els ambients selvàtics. Els arribats de fora del continent africà, és a dir, els forasters que hi vàrem residir, designàvem aquests paratges, com el «bosc», no solament per tractar-se de sectors del nostre planeta molt densos en vegetació i pràcticament ignots, sinó també per considerar-los fins i tot mítics, impregnats de misterioses llegendes. Als pocs infants forasters que vivíem allí, ens fascinava veure’ns enmig de tanta verdor. Després d’uns anys, Jordi Sabater va traslladar-se a Rio Muni, en terres continentals, a l’àrea de Nkumadjap, que limita amb el Camerun. Pel que vaig aprendre del meu pare, que –a començaments dels anys cinquanta– cercava indrets idonis per a implementar noves factories, aquesta zona era summament interessant per a conèixer el món dels primats. Ell mateix –en més d’una ocasió– havia vist algun primat de dimensions extraordinàries durant els trajectes que feia endinsant-se en el bosc.
Després d’haver contactat amb científics de gran renom, com el Dr. J.P. Capin del Departament d’Ornitologia de l’American Museum of Natural History de Nova York, Jordi Sabater va coŀlaborar en la recerca dels «ocells de la mel» i, poc més tard, també va investigar amb el Dr. H.L. Shapiro, director del Departament d’Antropologia d’aquella mateixa institució. Així mateix, arran de la coneixença dels Drs. D. Stark, Schultz, Biegert, Osman Hill, es centrà en l’estudi dels goriŀles i els ximpanzés. De fet, la seva dedicació apassionada a l’estudi d’alguns primats es complementava amb la d’altres éssers del territori (humans nadius, animals i plantes...) (Fig. 2), que


anava documentant sobretot a través d’anotacions i de dibuixos, representant els trets essencials de cadascun, minuciosament detallats, en llibretes i quaderns de treball.
El 1960 va estudiar la granota gegant (Conraua goliath Boulenger, 1906) a les cascades del riu Mbia, i en va aportar dades inèdites fins aleshores. El 1966, tal com expliquem més endavant, tingué l’oportunitat de trobar-se amb un petit gorilla albí. Aquest fet el projectà en la societat amb un caràcter molt més popular i proper a la ciutadania, ampliant, així, la seva trajectòria científica que s’havia mantingut gairebé en silenci, malgrat el significat i la rellevància que realment tenien les seves descobertes.
Al 1969 tornà a Barcelona i fou nomenat conservador de primats i del terrari del zoològic de la ciutat. No obstant, després de rebre una beca de la National Geographic Society, va retornar a l’Àfrica, en aquesta ocasió a Rwanda, amb la finalitat d’estudiar els goriŀles de muntanya.
El 1975, un cop ja restablert de nou a Catalunya, Sabater va emprendre els cursos reglamentaris a la Universitat de Barcelona i, un cop els va haver finalitzats, l’any 1977 s’incorporà com a professor a la Facultat de Psicologia de la mateixa universitat. Poc després d’haver defensat la seva tesi doctoral titulada Aportación a la eto-ecología comparativa de los gorilas («Gorilla gorilla gorilla») y chimpancés («Pan troglodytes troglodytes») de Río Muni (Barcelona, 1981), assolí el merescut reconeixement obtenint una càtedra d’etologia i primatologia. (Sabater Pi, 1984 & 1985). L’any 1989 dirigí una nova expedició per a investigar el comportament dels bonobos al Zaire (actual República Democràtica del Congo), comptant amb el patrocini de la Universitat de Barcelona.
L’any 2000 va cedir a aquesta mateixa Universitat, llur arxiu personal, el qual consta d’una coŀlecció de dibuixos, car-
tes, articles i llibres, a més d’alguns objectes que constituïen el seu patrimoni més preuat.
En traspassar, va deixar un gran nombre d’alumnes, amics i seguidors que l’admiràvem, a més del seu gran llegat científic, artístic i etnològic, com ja hem dit abans.
L’any 1958 Sabater Pi fou contractat per l’Ajuntament de Barcelona com a conservador del centre d’Ikunde, situat a les proximitats de la ciutat de Bata. El fundador d’aquest centre, Lluís de Lassaletta Delclòs (Barcelona 1922- Bata 1957) (Fig. 3a), havia mort poc abans, el dia 28 desembre de 1957, a causa de les mossegades d’una serp (un escurçó del Gabon) *. Ell mateix havia pronosticat abans de morir que aquelles mossegades no tenien salvació. Durant les visites a aquell indret, se solien fer demostracions amb alguns animals, particularment amb serps (Fig. 3b). El dia de l’accident, en Lluís havia estat molt reticent a fer-les, com si hagués tingut un mal presagi. Sembla que, després de la mossegada, va romandre encara, unes hores amb vida, tot i haver perdut el coneixement abans (Lorenzo Gacia, 2001: 149-150), contràriament al que consta en altres fonts, on es diu que va morir en qüestió d’uns «20 minuts». (Nerín Abad, 2017: 46).
En Lluís, juntament amb el seu germà Josep Maria, havien adequat aquest espai com a reserva i lloc d’estada transitòria dels animals que havien caçat amb vida, per adaptar-los a un entorn en captivitat, durant les fases prèvies a ser traslladats a alguns dels zoològics europeus, en particular, els de Barcelona i Madrid, un cop ja adaptats.
Per als petits que vivíem en aquelles terres, aquest paratge constituïa un dels majors atractius als quals teníem accés –si
M. GENERA I MONELLS


3. a) Molt probablement, Lluís de Lassaletta, a la costa propera a Bata, fotografiat a mitjan anys cinquanta; b) Visita al centre d’Ikunde per part de la nostra família l’any 1956. [Fotografies de J. Genera i Bartolí].
no era fins i tot el millor i més adequat– per passejar els caps de setmana, ja que era l’únic proper a la nostra residència de Bata i, a més, oferia la possibilitat de penetrar en el fascinant món de la natura immersos en el propi entorn. Nosaltres hi solíem anar sovint.
Casualment, en el vol de retorn a casa, l’abril de l’any 1957, vàrem coincidir amb Lluís Lassaletta durant els trajectes Bata-Lagos-Madrid, en el que fou el seu darrer viatgea la península , ja que la seva vida es va truncar uns mesos després.

Amb tot el risc de cometre errors, recordo que calia recórrer un parell de quilòmetres en vehicle (una pick-up) per arribar al centre d’Ikunde. Un cop allí, s’accedia a un clos que delimitava un conjunt d’instaŀlacions força precàries, construïdes amb tècniques senzilles, seguint la tradició de l’arquitectura indígena, emprant materials sostenibles, fusta, fang i brancalls. Hi havia un espai central ampli, on es trobaven les gàbies dels rèptils amb les serps més perilloses. A l’entrada –a mà esquerra– voletejaven les aus (recordo un espècimen d’àguila (?) que coronava un petit monticle, tot

Figura 4. a) Detall d’alguns episodis domèstics en convivència amb primats. b) Exemple de tràfic i compravenda de primats, molt probablement davant de la factoria de Bata. A l’esquerra de la imatge es pot veure una gàbia. [Fotografies realitzades per J. Genera i Bartolí]. Ambdues escenes apareixen definides en el pròleg d’un dels primer llibres de Sabater, on diu que «no s’haurien de tolerar les exhibicions grotesques d’aquests animals disfressats d’humans, ni la seva explotació comercial».
guaitant aquell escenari) i hi havia més d’un grup de gallines «de Guinea» que a nosaltres ens resultaven pintoresques pel seu plomatge esquitxat de blanc i negre. En front hi havia una sèrie d’estructures de maçoneria –a mode de laboratoris, més que de magatzems– on gaudírem, tot aprenent sobre la vida i la seva gestació, immersos en un pluralisme cultural, replet de creences i rituals, per a tots nosaltres exòtics. Aquell en-
torn, era capaç d’encendre fins a l’esclat, tot el potencial creatiu que duem –de vegades ocult– al nostre interior, recorrent al dibuix, com a expressió del fet vital i a la praxis d’un procés d’aprenentatge extraordinari. Inspiradora era l’observació atenta i minuciosa de tots els elements que configuraven aquella versió de «laboratori de campanya», amb diverses rengleres de flascons de vidre ben ordenats que conservaven
les mostres extretes de les diferents espècies, molt útils per a la recerca i l’experimentació, activitats que allí es practicaven paraŀlelament, en el camp de la zoologia en particular. Tots aquells objectes captivaven la mirada encuriosida d’un infant de curta edat, i els que més l’atreien eren, els que contenien els fetus de diverses espècies animals o algunes de les despulles dels que havien perdut la vida prematurament.
Després d’un rastreig de la informació que hi ha sobre la trajectòria professional de Sabater Pi, hem comprovat que aquest investigador va assumir la direcció del centre d’Ikunde l’any 1959 i el va continuar dirigint, durant un lapse aproximat d’una dècada. La seva missió havia de ser el subministrament de gran part dels fons zoològics, com ja s’havia fet abans. Ara bé, a partir d’aquell moment, també va tenir com a objectiu la recollida d’elements d’interès etnogràfic, arqueològic i botànic per a les institucions barcelonines (AAVV, 2017).
Uns nadius, l’any 1966, van vendre a Sabater Pi un petit goriŀla albí amb ulls blaus, que al cap d’un temps el van portar al zoo de Barcelona. Molt poc després –aquest petit ésser tan especial– per no dir singular, és convertí en una de les icones més emblemàtiques del mateix zoològic (Sabater Pi &Tobaruela, 2003)
Aleshores la compra-venda de simis era un fet absolutament normal. Jo mateixa tenia un «tití» (així, designàvem tots els micos de petites dimensions), que el meu pare havia comprat a un venedor ambulant d’ous. Algunes famílies espanyoles tenien un ximpanzé «ensinistrat» (Fig. 4a), altres els traginaven a munt i avall (Fig. 4b), suposo que per a treure’n algun profit pecuniari.
Conèixer en Jordi va permetre’ns –en certa manera– reviure amb intensitat, la nostra estada a Guinea, els petits i grans records de les nostres «memòries d’Àfrica» repletes d’experiències, evidentment, amb durades i circumstàncies molt diferents.
La concepció moderna de la prehistòria investigada per equips pluridisciplinaris
La promoció de «prehistoriadors» de 1970-1975 de la Universitat de Barcelona, aleshores llicenciats i doctors en filosofia i lletres d’aquesta prestigiosa institució, vàrem tenir l’oportunitat de seguir l’anomenat «Pla Maluquer». Aquest pla permetia accedir a determinades matèries pròpies de la Facultat de Ciències, com alguns estudis preliminars sobre el Quaternari que s’impartien a Geològiques i l’antropologia física de Biologia. Entretant, després d’haver coŀlaborat en diverses excavacions dirigides pel professor H. de Lumley –sobretot a la Caune de l’Aragó a Talteüll i a la Cova de Lazaret a Niça– vàrem comprovar que en la investigació de la prehistòria, principalment en les etapes més antigues, era imprescindible que amb els historiadors coŀlaboressin equips pluridisciplinaris formats per especialistes experts en geologia, biologia, paleontologia, antropologia, dibuix, etc. Partim, conceptualment, del principi que l’estudi del paleolític no es limita solament a la consideració dels primers útils fa-
bricats per l’ésser humà, que ens permeten establir les diferents fases culturals d’aquest període, sinó que també té com a objectiu reconstruir l’ambient en el qual es desenvolupà llur existència, tant el constituït pels factors climàtics, ecològics, ..., com els derivats de l’impacte de la seva pròpia presència (Genera i Monells, 1975). És, justament, en aquella etapa, que alguns de nosaltres ens vàrem inscriure com a socis de la Institució Catalana d’Història Natural (ICHN), perquè era l’entitat constituïda per investigadors de prestigi en tots aquests àmbits científics associats a la recerca prehistòrica. A més, aquesta entitat estava vinculada a l’Institut d’Estudis Catalans (IEC). Dins d’aquest context inteŀlectual, les aportacions de Jordi Sabater Pi foren crucials i molt enriquidores també per als prehistoriadors.
La primera conferència impartida per aquest investigador a la qual vaig assistir va tenir lloc al Museu Arqueològic el mes de març de l’any 1978, arran d’un curs sobre primatologia. Aquest museu, en aquells moments, pertanyia a l’Institut de Prehistòria i Arqueologia de la Diputació de Barcelona, que era dirigit pel professor Eduard Ripoll Perelló. Les classes sempre les acompanyava amb dibuixos que feia a la pissarra mentre parlava. Un mètode altament pedagògic.
Molt poc més tard, Sabater Pi es convertí també en un gran coŀlaborador de les activitats que es duien a terme promogudes per l’ADAHUB (Associació de Diplomats en Arqueologia Hispànica Universitat de Barcelona) a través d’una exposició itinerant sobre «l’evolució de l’home». En termes actuals, hauríem de dir «l’evolució de l’ésser humà» En l’exposició, es mostraven –entre altres objectes– la coŀlecció de rèpliques d’homínids que jo acabava d’adquirir a Kènia –sobretot els cranis més significatius, descoberts a les formacions de Koobi-Fora, al Llac Turkana– durant la meva estada a Àfrica oriental, arran del VIII Congrés Panafricà, que va tenir lloc el setembre de 1977 (P.A.C.P.Q.S. Panafrican Congress of Prehistory & Quaternary Studies) (Genera i Monells, 1978: 345-347)
Aquesta exposició es complementava amb conferències i altres activitats divulgatives de gran qualitat. Una d’elles va anar a càrrec de Sabater Pi. Gràcies a disposar dels butlletins publicats pel propi ADAHUB (en particular el corresponent als mesos de gener- març de 1979), hem pogut comprovar els temes i les dates de cada sessió. La del professor Sabater i Pi, que llavors ja era conservador del Departament de Primats del Parc Zoològic de Barcelona i professor de psicologia animal de la Universitat de Barcelona, va tenir lloc el dia 9 de novembre de 1978. Si no m’erro, la va impartir al palau de la Virreina de Barcelona. Va tenir per títol El ximpanzé i els orígens de la cultura, com una de les seves obres publicades, i tractà de les capacitats cognoscitives d’aquest grans simis, tenint en compte els estudis recents d’aleshores (Sabater Pi, 1984 & 1985). Al llarg d’aquella presentació feu referència als aspectes que s’expliquen a continuació:
La capacitat de reconèixer l’esquema corporal i de tenir noció de la mort; la capacitat comunicativa generalitzada, preposicional i abstracta; la capacitat d’usar i fabricar eines elementals, l’escassa incidència de l’incest primari, una certa capacitat estètica...
M. GENERA I MONELLS
– La capacitat cooperativa, relacional i funcional que es manifesta en les caceres cooperatives que comporten la distribució d’aliments entre adults, si és soŀlicitada.
- La formació d’unitats familiars mínimes (mare fills i nets) molt estables.
Les següents sessions varen tractar sobre Australopithecus Raymond Dart, 1925 i Homo habilis Leakey, Tobias & Napier, 1964, i es van centrar en les darreres troballes (Genera i Monells, el 24.11). La de la Dra. Rutta Rosenstingl i d’Emília Solà va estar centrada en «La comunicación entre homínidos sin habla» (1.12.78).
A mitjan decenni dels anys vuitanta, va protagonitzar una sessió científica dins el curs sobre evolució que havíem organitzat en el centre de la UNED de Tortosa. Era el primer cop que es plantejava en aquella ciutat el tema de l’evolució humana mostrant una petita selecció de rèpliques de cranis (pertanyent a la coŀlecció ja esmentada) i utillatges lítics, a més dels bastonets usats pels ximpanzés que ell sempre portava. Tingué un gran èxit de públic, àdhuc entre els eclesiàstics assistents. Aprofitant aquell viatge a la capital del Baix Ebre, l’endemà vàrem desplaçar-nos a les terres del Priorat i Ribera d’Ebre, on vam visitar, juntament amb la seva muller, Núria, els jaciments prehistòrics que excavem des del 1976. L’abril de 1991, vaig assistir a un nou curs a càrrec del Dr. Sabater, que tenia per títol «Etoprimatologia i prehistòria». Aquest va ser organitzat pel Museu Arqueològic de Barcelona i per la Universitat de Barcelona, juntament amb la Societat Catalana d’Arqueologia i el Parc Zoològic de Barcelona. Finalment, comptant amb la seva participació, es va dur a terme un nou curs –els dies 27 i 28 de novembre de 1999– a la UNED. Aquest es va fer entre les poblacions de Terrassa i Talteüll, i va ser organitzat pel consorci del Centre Associat de la UNED de Terrassa, amb la coordinació científica de la que subscriu aquest article. Entre els ponents, es trobaven els següents investigadors: David Serrat, Domènech Campillo, el mateix Jordi Sabater, que va fer una dissertació sobre les conductes culturals dels primats no humans, Daniel Turbón, Margarida Genera, Victòria Cabrera, ... La veritat és que fou un èxit de participació, no tan sols pel que fa a l’alumnat al qual anava especialment adreçat, sinó també quant al públic en general.
Així, doncs, per haver mantingut llaços d’amistat, enfortits per les idees, però també per la coincidència d’haver viscut tots dos un temps en el continent africà (1956-1957) –sense aleshores haver-nos conegut– i d’haver estat, a Barcelona, veïns molt propers, els vincles d’estima recíproca van transcendir –fins i tot– als nostres familiars més íntims, a amics i a alumnes.
La seva aportació científica i pedagògica en l’àmbit de la sensibilització envers l’entorn natural
Una de les més grans aportacions de Jordi Sabater Pi, ha estat la constatació que la cultura no és exclusiva dels humans, ampliant així el concepte de fet cultural. També ha estat molt notable la seva capacitat com a divulgador científic. Les seves recerques han permès constatar que els ximpanzés no solament
són capaços d’usar bastonets per a atrapar tèrmits del termiter, sinó que a més transmeten llurs coneixements de generació a generació. Així mateix, va dissenyar tècniques que eviten l’alteració de l’estat anímic dels simis en determinades circumstàncies inhabituals, durant les recerques. Per tant, l’hem de considerar com el principal introductor de l’etologia i la primatologia a casa nostra, però també en l’àmbit mundial. Pels avenços assolits, tinc la convicció que ha estat mereixedor de molts reconeixements i premis que s’han atorgat a altres investigadors per temes de comportament dels primats. Si més no, opinem que haurien d’haver estat compartits.
Un altre aspecte que és molt remarcable en la figura del Dr. Jordi Sabater Pi, i que el distingeix entre molts altres investigadors, és la seva faceta com a dibuixant científic, naturalista i fotògraf (Pou, 2023)
Ha resultat fonamental la seva importantíssima labor professional en el camp del dibuix –massa sovint anònima– emprat com a llenguatge universal que permet no tan sols enregistrar, documentar i descriure determinats elements amb tot rigor, sinó també transmetre continguts de gran complexitat de forma inteŀligible per a la societat en general, impregnantlos de sentiments profunds del propi autor, sense cap mena de filtre idiomàtic, ni ideològic. A més, el dibuix aplicat a la investigació científica, en èpoques anteriors a la fotografia, ha resultat un mitjà molt eficaç de registre i difusió de les imatges d’experiències viscudes en uns determinats llocs i moments molt precisos.
Evocant algunes de les reflexions del propi Sabater, la investigació naturalista combina l’observació i el dibuix. Doncs, el fet d’observar constitueix la base del coneixement científic perquè pressuposa una millor comprensió de l’entorn que ens envolta. La seva fascinació en copsar minuciosament en que observava en un dibuix, va anticipar-se en la necessitat imminent de la conservació del medi natural, matèria que ara tant ens preocupa, sense saber implementar les solucions adequades.
El dibuix –com a traç per a plasmar una imatge sobre una superfície– és una acció altament expressiva i de comunicació pròpia dels éssers humans que permet transmetre idees i conceptes. Així mateix, tenint en compte les diferents facetes d’un dibuix i que la seva comprensió pot dependre del prisma a través del qual es contempli i de la llum projectada a cada instant, ens adonarem de l’amplitud de la temàtica que ara plantegem i de la multitud d’interpretacions que es poden originar (Genera i Monells, 2022a, b & 2023) (Fig. 5).
En el cas de Sabater Pi, és molt senzill, copsava amb tota minuciositat i rigor el que veia a través de la mirada neta i transparent pròpia d’un científic extraordinàriament humà. Ho sabia plasmar amb tot el respecte a la natura.
Es tracta, generalment, de dibuixos sobre paper com ja hem dit –quaderns de viatge– realitzats a llapis, ploma o pinzell –de vegades aplicant tècniques mixtes– i iŀluminats amb color. S’expressava a través d’uns dibuixos extraordinàriament eloqüents i, fins i tot les dedicatòries de molts dels seus llibres, les solia acompanyar d’un petit esbós –sovint amb la imatge d’un primat– que personalitzava encara més les seves afectuoses paraules escrites.

5. Dibuixos realitzats per Jordi Sabater i Pi en els seus quaderns de treball. [Fotografies de la autora. Fons: CRAI, UB].

Figura 6. Imatge on es pot veure el fill de Jordi Sabater Pi, Oriol Sabater Coca, pronunciant unes paraules en record del seu pare, en l’acte d’atorgaments dels premis Nat, en la seu del Museu de Ciències de Barcelona, a finals del desembre de 2022. [Fotografia de la autora].
Després del seu decés, s’han presentat diverses exposicions que, tot i que han estat de petit format, han divulgat l’obra d’aquest investigador, en particular per al públic que no el va arribar a conèixer personalment. Entre les més recents, cal citar: «L’art d’un naturalista de camp», en el Museu d’Història de Catalunya, «El viatge iŀlustrat. Quaderns de camp: del llapis i el pinzell al món digital», en el Museu Etnològic
o de les Cultures del Món (18.06.22-06.11.22); “Dibuixant de la natura” (5 al 8 d’octubre de 2022) en el marc de la VIII Iberian Primatologica Conference i en commemoració del centenari del seu naixement. Més recentment encara, del 13 de desembre de 2022 al 2 d’abril de 2023, el Museu de Ciències Naturals de Granollers va realitzar la mostra «L’art de la ciència».
M. GENERA I MONELLS
A través dels seus treballs, podem percebre la seva magnífica formació tant artística com científica, una dualitat que va propiciar la qualitat del seu llegat, el qual ara recordem amb tanta admiració. Conscientment, deixem per a una altra publicació els resultats de la seva tasca com a documentalista de l’art corporal mitjançant el tatuatge, al qual també va dedicar molta atenció. (Sabater Pi, 1993 & 2001; Sabater i Pi & Sabater i Coca, 1992).
Entre els reconeixements i premis que va rebre, destaquen els nomenaments de doctor honoris causa per la Universitat Autònoma de Barcelona (1997) i per la Universitat Autònoma de Madrid (1993). També va rebre el premi de la Fundació Catalana per a la Recerca (1991), la Medalla d’Or al Mèrit Científic de l’Ajuntament de Barcelona (1996), la Creu de Sant Jordi (2000) i la Medalla Narcís Monturiol al mèrit científic i tecnològic de la Generalitat de Catalunya (2005). A les darreries del 2022, el Museu de Ciències Naturals li atorgava la menció d’honor dels Premis Nat, a títol pòstum. (Fig. 6)
Consideracions finals
Com a cloenda, remarquem que Jordi Sabater Pi fou un gran investigador –que, malgrat la seva condició d’autodidacte–, va desenvolupar una trajectòria brillant de forma esteŀlar, sobretot pel que fa a l’estudi dels goriŀles i l’ús d’eines entre els ximpanzés. De fet, l’hem de considerar un dels pioners de la primatologia en l’àmbit mundial.
També és el paradigma del dibuixant naturalista extraordinari i del fotògraf. Va emprar el dibuix com una eina molt eficaç de documentació i transmissió de coneixements científics. A través dels seus treballs, podem percebre la seva magnífica formació, que incrementa la qualitat del seu llegat.
El dibuix realitzat sobre una base científica rigorosa és una eina pedagògica i didàctica de valor incalculable. Ens permet visualitzar idees i conceptes sovint molt difícils de comprendre per tractar-se de continguts abstractes d’interpretació complexa. Per altra banda, és capaç d’explicar el procés a través del qual s’ha desenvolupat.
El nivell d’exceŀlència assolit pel professor Sabater Pi a través dels seus dibuixos de primats propicia que l’espectador gaudeixi de la seva gran bellesa plàstica i senti l’emoció d’aprofundir en el coneixement d’aquests éssers tan pròxims a nosaltres.
En definitiva, la sensibilitat i tendresa de les representacions desperten -encara més- el desig de protegir i conservar la natura, tal com prodigà el seu autor amb vehemència i convicció en el decurs de tota la seva vida.
Com a petit homenatge, cloem aquests paràgrafs, recordant les seves pròpies paraules:
«Si observes, coneixes si coneixes, estimes, i si estimes, protegeixes.»
Hem insinuat que dibuixar pot ser una manera d’estimar la natura, i opinem que el dibuix també pot esdevenir una forma de memoritzar la trajectòria terrenal d’una determinada persona fins a l’eternitat.
Al CRAI de Belles Arts i al Museu d’Etnologia o de les Cultures del Món, per facilitar-nos informació referent als dibuixos custodiats en els seus respectius arxius. A en Josep Maria Enric, per les seves aportacions referents a la vida a la Guinea Equatorial durant els anys cinquanta, així com a Margarita Monells Borrell, per autoritzar-nos a publicar les fotografies de l’arxiu familiar.
*En tot allò referent a la mort de Lluís de la Salletta, hauríem d’aprofundir més documentalment, ja que hi ha diverses versions.
AAVV. 2017. IKUNDE. Barcelona, metròpoli colonial, catàleg de l’exposició temporal. Museu de Cultures del Món/Museu Etnològic de Barcelona. Ajuntament de Barcelona, 1ª edició, Vanguard Gràfic, SA, 288 p.
Genera i Monells, M. 1975. Métodos de estudio del paleolítico inferior, aplicados a la problemàtica de las culturas con industrias líticas más arcaicas en Cataluña mediterrània, Tesi de llicenciatura, Universitat de Barcelona. 225 p.
Genera i Monells, M. 1978. VIII Congreso Panafricano de Prehistoria. Pyrenae, 13-14 (1977-1978): 345-347.
Genera i Monells, M. 2022a. “The figure of Jordi Sabater Pi. Some bio-historiographic data regarding his activity as a scientific divulgator of the origins of Humanity”, Llibre de resums de la VIII Conference Iberian Primatological, Barcelona. p. 38
Genera i Monells, M. 2022b. “Human evolution through the look some paradigmatic drawers” Llibre de resums de la VIII Conference Iberian Primatological, Barcelona. p. 39.
Genera i Monells, M. 2023. “Seguint el traç esteŀlar d’un gran dibuixant de la natura” dedicat a la figura de Jordi Sabater Pi. Darwin News, l’informatiu dels primats, 19: 5-11.
Lorenzo Gacia, A. 2001. Vivències de Guinea. Viena memòria, 1ª edició, Barcelona. 228 p.
Nerín Abad, G. 1998. Guinea equatorial. Història en blanc i negre. Dones negres i homes blancs a la Guinea Equatorial, 1843-1968, 46, Biblioteca Universal Empúries, Barcelona. 267 p.
Pou, T. 2023. Jordi Sabater Pi, l’últim naturalista, Ajuntament de Barcelona-Universitat de Barcelona edicions, Barcelona. 312 p.
Sabater Pi, J. 1984. Gorilas y chimpancés del África occidental. Estudio comparativo de su conducta. 1ª edició, F.c.e. México. 279 p.
Sabater Pi, J. 1985. Etología de la vivienda humana. De los nidos de chimpancés a la vivienda humana. Monografías Labor Universitaria, 1ª edició, Barcelona, 126 p.
Sabater Pi, J. 1993: L’art d’un naturalista, Universitat de Barcelona, Fundació la Caixa, Publicacions de la Universitat de Barcelona. 77 p.
Sabater Pi, J. 2001. El traç de la natura, 1ª edició, Edicions 62, Barcelona 156 p.
Sabater i Pi, J. & Sabater i Coca, O. 1992. Els tatuatges dels fang de l’Àfrica occidental. Art, simbolisme i biologia en una manifestació artística poc coneguda. Ajuntament de Barcelona, Treballs del Museu Etnològic/1, 250 p.
Sabater Pi, J. & Tobaruela, P. 2003. Floquet per sempre, el goriŀla. la blanc vist pel seu descobridor. Edicions 62, Barcelona. 94 p.
Cristian R. Altaba*
* Conselleria d’Agricultura, Pesca i Medi Natural, Govern de les Illes Balears, 07009 Palma; Grup de Recerca en Evolució i Cognició Humana, Universitat de les Illes Balears, 07071 Palma. A/e: cristianr.altaba@uib.cat
Rebut: 17.02.2024; Acceptat: 14.03.2024; Publicat: 31.03.2024
Resum
Es descriu un nou gènere de la família Strombidae Caenogastropoda: Stromboidea): Javastrombus n. gen., del Miocè superior de Java occidental (Indonèsia). Es considera proper morfològicament a Persististrombus, del qual probablement deriva; i se’n distingeix perquè és més petit, té una estructura més delicada, un desenvolupament notable dels sinus aperturals, una quilla supracolumeŀlar i una delicada escultura espiral. S’hi inclouen sis noves espècies: J. praegracilis n. sp. (espècie tipus), J. pinguis n. sp., J. cylindratus n. sp., J. kecil n. sp., J. subinermis n. sp i J. sondaicus n. sp. El nou gènere ocupa una posició geogràfica allunyada tant d’Afristrombus, actualment limitat a les costes africanes occidentals tropicals (però estès en el Neogen al Paratetis i la Mediterrània), com de Persististrombus, distribuït per l’àrea mediterrània durant l’Oligocè i el Miocè i per les costes tropicals atlàntiques i pacífiques d’Amèrica des del Miocè. El nou gènere representa una branca del grup format per Persististrombus, Afristrombus i ell mateix que s’hauria diversificat en aïllament, en un medi favorable, estable i complex, al llarg del Miocè mitjà (Languià i Serravaŀlià, entre 16,0 i 11,6 Ma).
Paraules clau: Gastropoda, Strombidae, Javastrombus, Afristrombus, Persististrombus, Java, Indonèsia, Miocè mitjà, taxonomia, biogeografia, diversificació.
Abstract
A new fossil genus of the family Strombidae (Gastropoda: Caenogastropoda: Stromboidea) from the Middle Miocene of Java
A new genus of the family Strombidae (Caenogastropoda: Stromboidea) is described: Javastrombus n. gen., from the Middle Miocene of Western Java (Indonesia). It is considered to be phylogenetically close to Persististrombus, from which it probably derived. It is distinguished by its small size and delicate structure, the notable development of the apertural sinuses, a supracolumellar keel, and the delicate spiral sculpture. Six new species are included therein: J. praegracilis n. sp. (type species), J. pinguis n. sp., J. cylindratus n. sp., J. kecil n. sp., J. subinermis n. sp, and J. sondaicus n. sp. The new genus occupies a geographical position distant from both Afristrombus, currently limited to the tropical West African shores (but extending during the Neogene throughout the Paratethys and the Mediterranean), and Persististrombus, distributed throughout the Mediterranean area during the Oligocene and Miocene, as well as the tropical Atlantic and Pacific coasts of America since the Miocene. The new genus represents a branch of the group it constitutes together with Persististrombus and Afristrombus diversified in isolation in a favorable, stable and complex environment during the Middle Miocene (Langhian and Serravalliian, between 16.0 i 11.6 Ma).
Key words: Gastropoda, Strombidae, Javastrombus, Afristrombus, Persististrombus, Java, Indonesia, Middle Miocene, taxonomy, biogeography, diversification.
urn:lsid:zoobank.org:pub: 1DFFFEBA-BE5C-4F21-8496-00BBD28C5937
Introducció
La família Strombidae presenta diferents exemples de radiacions evolutives que van tenir lloc al llarg del Cenozoic i mostra una excepcional diversitat en la morfologia de les conquilles (Roy et al., 2001; Savazzi, 2007). La filogènia de les nombroses espècies actuals s’ha començat a aclarir mitjançant anàlisis morfològiques i moleculars (Simone, 2005; Latiolais et al., 2006; Li et al., 2022). Un resultat prou clar d’aquestes anàlisis és que cal acceptar una notable diversitat de gèneres, tant per reconèixer els diversos llinatges evolu-
tius, com per permetre una comunicació sobre aquesta diversitat biològica (Simone, 2005; Maxwell et al., 2020; Dekkers, 2023; Wieneke et al., 2024).
Un dels escenaris més suggeridors, i alhora desconeguts, de la diversificació dels estròmbids es troba al Miocè mitjà de les illes de la Sonda, majoritàriament a l’actual Indonèsia, i a regions veïnes. Ja en els treballs clàssics de Martin (1879-1880, 1881-1906) aquest autor descrivia nombroses espècies desconegudes fins llavors de moŀluscs marins fòssils d’aquests jaciments. Entre aquestes, n’hi ha diverses d’estròmbids, les quals encara plantegen qüestions que cal-
C. R. ALTABA
dria investigar. Ara bé, als darrers anys hi ha hagut un interès renovat per aquestes faunes, que presenten estats de conservació francament excepcionals i una riquesa molt destacable (Hoek Ostende et al., 2002). Així, s’han publicat nombroses novetats del Miocè mitjà i superior de Java (Robba, 1996, 2013; Kase et al., 2015; Landau et al., 2020a, b; Dovesi & Parsons, 2021; Merle et al., 2021; Clezard & Dovesi, 2023) que inclouen alguns estromboïdeus (Dekkers et al., 2020; Liverani & Wieneke, 2022).
La història geològica de Java és complexa (Smyth et al., 2005; Clements & Hall, 2007), i presenta diverses fases de deposició de carbonats durant el Cenozoic (Wilson, 2002). Les formacions del Miocè mitjà destaquen per contenir una enorme abundància de conquilles molt ben conservades, que visqueren en ambients poc profunds. La més ben estudiada és la formació Nyalindung, la qual consisteix bàsicament en sediments siliciclàstics, pertany a l’estatge Seravaŀlià i té una edat aproximada de 12 milions d’anys. Condicions semblants a les del Seravaŀlià existiren des del Languià, un estatge al qual pertanyen altres jaciments de l’est de Java (Syarifin, 2011; Robba, 2013; Aswan et al., 2017; Kapid et al., 2019; Dekkers et al., 2020; Aswan, 2021; Dovesi & Parsons, 2021; Kesuma & Aswan, 2022).
Al present treball es donen a conèixer un total de sis espècies, agrupades en un nou gènere. Es tracta d’espècies força rares; de fet, fins ara no s’havia documentat cap espècimen atribuïble a aquest grup.
Material i mètodes
Els jaciments que han fornit els exemplars estudiats aquí es troben a la regió més occidental de l’illa de Java (Indonèsia), i són els que venen a continuació.
1) Golden Park, Suradita, Kecamatan (districte) Cisauk, Kapubaten (regència) Tangerang, Propinsi (província) Banten. 6º 20’ 58’’ S, 106º 39’ 08’’ E. Altitud 22 m.
2) Barangbang, Ciwarak, Kecamatan (districte) Jatiwaras, Kabupaten (regència) Tasikmalaya, Propinsi (província) Jawa Barat. 7º 31’ 10’’ S, 108º 12’ 36’’ E. Altitud 303 m.
3) Curug Orok, Ci Kandang, Kecamatan (districte) Cikajung, Kabupaten (regència) Garut, Propinsi (província) Jawa Barat. 7º 23’ 14’’ S, 107º 44’ 09’’ E. Altitud 1.132 m.
4) Curug Rahong, Mekarjaya, Kecamatan (districte) Sukaraja, Kabupaten (regència) Tasikmalaya, Propinsi (província) Jawa Barat. 7º 27’ 24’’ S, 108º 08’ 44’’ E. Altitud 291 m.
Tots aquests jaciments semblen atribuïbles a un interval del Miocè mitjà que abasta des de la zona NN5 fins a la NN7, pertany als estatges Languià i Serravaŀlià i té una antiguitat d’entre 16,0 i 11,6 Ma. Aquesta datació és orientativa, perquè falten dades adequades.
Els fòssils es recolliren amb eines manuals senzilles, amb les quals s’extreien de la matriu sorrenca més o menys consolidada. Després foren rentats i embolicats amb paper i plàstic, deixant que s’assequessin molt lentament. Un cop eixuts, es varen estabilitzar submergint-los en una solució al 3 % de resina sintètica Paraloid B72.
El material estudiat consta de 24 exemplars en total. Es troba a la coŀlecció malacològica de l’autor (CRA), reconeguda com a coŀlecció museològica pel Consell de Mallorca. Es conserven en calaixeres metàŀliques, dins de capsetes de poliestirè transparent, amb cotó fluix i etiquetes de paper sense àcid escrites amb tinta xinesa, en un medi amb temperatura i humitat controlades.
La terminologia de les sinuositats de l’extrem abapical del llavi segueix la proposta de Dekkers (2021). El sinus estromboide és un tret exclusiu dels estròmbids (i famílies molt properes), per on l’animal treu l’ull dret (els ulls són grossos i estan situats a l’extrem de peduncles prou llargs); el sinus anterior, que és homòleg del canal sifonal dels neogastròpodes, allotja l’ull esquerre. Ambdós sinus estan separats per un sortint del marge de l’obertura, anomenat lòbul estromboide
Les mesures de longitud s’han pres en paraŀlel a l’eix d’enrotllament, amb un peu de rei. Donat que l’extrem anterior està quasi sempre una mica trencat i que l’àpex està lleument erosionat en alguns exemplars, la longitud total original devia ser entre 1 i 3 mm superior a la mesurada.
Classe Gastropoda Cuvier, 1795
Subclasse Caenogastropoda L.R. Cox, 1960
Ordre Littorinimorpha Golikov & Starobogatov, 1975
Superfamília Stromboidea Rafinesque, 1815
Família Strombidae Rafinesque, 1815
Javastrombus n. gen. urn:lsid:zoobank.org:act:CB8E28FA-6E10-4049-A79FB50E7E6F109F
Diagnosi
Javastrombus n. gen. es caracteritza per tenir la mida petita, un gruix moderat de la conquilla, una corona de punxes agudes i fortes i (quasi sempre) unes punxes molt més febles a la part abapical del dors, l’espira cònica, l’escultura espiral fina i limitada (o més evident) a la rampa subsutural, l’obertura llisa amb columeŀla engruixida i delimitada adapicalment per una quilla que la separa de la rampa subsutural, i el sinus estromboide i el sinus anterior ben marcats. Entre aquests dos sinus, té un lòbul estromboide que és ben arrodonit i més o menys prominent i està inclinat cap endavant.
Descripció. Conquilla de mida moderada o bastant petita en el context de Strombidae, bastant gràcil i moderadament sòlida, amb una filera de 4 o 5 punxes primes, arrodonides, més o menys potents, ortogonals a l’eix d’enrotllament o lleugerament inclinades adapicalment o abapical, i regularment espaiades a l’espatlla de la darrera volta; la primera d’aquestes és menuda, mentre que les centrals són les més desenvolupades. Generalment hi ha també entre una i tres punxes baixes, a la part abapical del dors; que són clarament menys prominents, estan alineades en espiral amb el sinus estromboide i en alguns casos estan unides per una tènue carena espiral. L’espira és cònica, té unes 7 o 8 voltes, forma un angle d’entre 45º i 66º, i presenta un creixement regular fins
al darrer quart de volta, on s’expandeix. El cos de la darrera volta, abans d’aquesta expansió del llavi, varia des d’estret fins a inflat. El llavi és en forma d’ala, poc o moderadament expandit i més o menys engruixit en part, però té el marge arrodonit i molt prim als extrems; té una inclinació prosoclina més o menys marcada i un perfil que va de quasi rectilini a fortament corbat; té l’extrem posterior poc o gens prominent, formant un canal posterior bastant o força profund, després del qual s’insereix a la darrera volta; a l’extrem anterior hi té un sinus estromboide molt marcat i fortament sinuós, adjacent al lòbul estromboide, que és més o menys prominent i poc o molt retorçat cap endavant. El sinus anterior és obert, breu i lleugerament o moderadament reflectit. La columeŀla és ampla, engruixida i llisa; està limitada posteriorment per un caire angulós, resultat dels nòduls de l’espira que esdevenen baixos, estrets i amples fins a formar aquesta quilla a l’espatlla de l’inici de la darrera volta. Adapicalment, la columeŀla forma un caŀlus que s’uneix al llavi, tot recobrint la rampa subsutural de l’inici de la darrera volta en grau variable. La superfície externa és quasi llisa, sense cap àrea granulosa; està solcada per estries de creixement colabrals, que són irregulars i febles, i estan entrecreuades per cordes espirals fines i nombroses, força regulars. Aquestes cordes espirals són més marcades a la rampa subsutural, fins a la base de les punxes de la corona, com també a l’espira de la teleoconquilla; molt afeblides, poden aparèixer a zones del dors. La protoconquilla és pauciespiral, prima i cònica, i consta de poc més de dues voltes llises (malgrat el bon estat de conservació, no s’hi aprecia microescultura). L’inici de la teleoconquilla està marcat per l’aparició de costelletes axials, ortoclines, i molt regulars, entre les quals apareixen les cordetes espirals. Aquesta escultura axial arriba fins a un límit entre dues i quasi quatre voltes de l’obertura; a partir d’aquest límit, hi ha una transició ràpida cap a l’escultura de nòduls angulosos,
també regularment espaiats, situats més o menys per damunt la sutura, que esdevenen la quilla supracolumeŀlar i després punxes a la darrera mitja volta.
Etimologia
El nom del nou gènere deriva d’ajuntar el de l’illa de Java amb el del gènere típic de la família.
Espècie tipus
L’espècie tipus de Javastrombus n. gen. és Javastrombus praegracilis, n. sp.
Espècies incloses
Dins Javastrombus n. gen. s’hi inclouen sis espècies noves: J. praegracilis n. sp. (espècie tipus), J. pinguis n. sp., J. cylindratus n. sp., J. kecil n. sp., J. subinermis n. sp i J. sondaicus n. sp.
Distribució
Els escassos exemplars coneguts de Javastrombus spp. procedeixen tots de petits afloraments del Miocè mitjà a l’illa de Java (Indonèsia), en els quals s’hi ha trobat una abundant i diversa fauna malacològica fòssil, pròpia d’ambients tropicals poc profunds i de fons tous.
Comparació
Es distingeix de Persististrombus Kronenberg & Lee, 2007 i d’Afristrombus Bandel, 2007, els gèneres més propers morfològicament, per la seva mida reduïda i estructura delicada, el desenvolupament notable dels sinus aperturals, una quilla supracolumeŀlar i la delicada escultura espiral.
Javastrombus praegracilis n. sp.
urn:lsid:zoobank.org:act:BA5CC060-AC9A-4659-B8BA-9B51D1F18D84
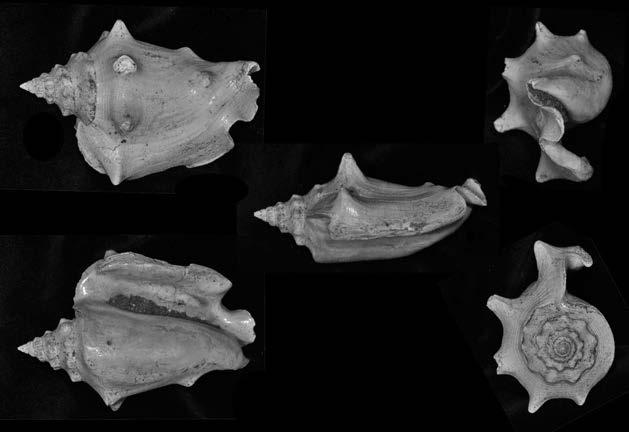
Material tipus
HOLOTIP: CRA-23102-1, de Serpong (Suradita) (Fig. 1).
PARATIPS 1 i 2: CRA-23102-2 i 3, de Serpong (Suradita).
PARATIP 3: CRA-23171, de Curug Orok (Ci Kandang).
Diagnosi
Espècie mitjana de Javastrombus, amb espira aguda i ornada d’una corona de punxes baixes i amples, aproximadament ortogonals a l’eix d’enrotllament, nòduls ben definits (però petits i baixos) per damunt de la sutura, llavi engruixit amb canal posterior poc profund i sinus estromboide profund, darrera volta gràcil, 5 o 6 punxes a l’espatlla, arrodonides i escassament inclinades adapicalment, 2 o 3 punxes dorsals baixes (o alguna només incipient) i ben separades al llarg d’una tènue carena espiral, caŀlus columeŀlar poc reflectit i recobrint fins a la meitat de la rampa subsutural de l’inici de la darrera volta, i sinus anterior fortament recorbat.
Descripció
Conquilla de mida moderada (holotip: 54,6 mm; paratips: entre 49,3 i 49,6 mm), força gràcil (com indica el seu nom específic) i moderadament sòlida, amb una corona de 5 o 6 punxes primes, arrodonides, breus i amples, només lleugerament inclinades adapicalment; hi ha 2 o 3 punxes molt baixes, molt menys prominents (alguna d’aquestes molt escassament desenvolupada) i alineades a la part abapical del dors, al llarg d’una incipient carena espiral, amb el sinus estromboide. L’espira és cònica i elevada, relativament llarga i estreta, amb 8,2-8,6 voltes de creixement regular i forma un angle d’uns 50º; el cos de la darrera volta, abans de l’expansió del llavi, és força estret. El llavi és en forma d’ala, poc expandit, i força engruixit al mig, prosoclí però quasi recte a tota la part central; el marge és arrodonit, però molt prim al sinus posterior, el qual és ample, poc profund i allargassat cap endavant. L’extrem posterior del llavi és poc prominent i
s’insereix a la darrera volta, on recobreix fins a la meitat de la rampa subsutural; a l’extrem anterior el sinus estromboide és molt marcat i profund, adjacent al lòbul estromboide, el qual és relativament gran i ben torçat prosoclinament; el sinus anterior és menys incís, un xic allargassat i ben recorbat. La columeŀla és engruixida i llisa, unida adapicalment al llavi per damunt de la quilla a l’espatlla. La superfície externa és quasi llisa, amb estries de creixement irregulars, cordes espirals molt fines, un xic més marcades al dors, i en canvi bastant fortes a la rampa subsutural, on n’hi ha 11 repartides de manera bastant regular que incrementen el seu gruix envers la sutura (i 5-7 de molt més febles, intercalades); aquestes es continuen per l’espira (a la part més adapical, visibles als espais entre les costelletes axials). La protoconquilla consta de 2,4-2,5 voltes aparentment llises. L’inici de la teleoconquilla està marcat per l’aparició de costelletes axials, ortoclines i molt regulars, que s’estenen al llarg de 3,2-4,2 voltes, i s’aturen a 2,5 voltes de l’obertura; els nòduls subtriangulars (10 o 11 a la penúltima volta) estan regularment espaiats i se situen clarament per damunt la sutura.
Etimologia
El nom específic praegracilis és un adjectiu singular, propi del llatí postclàssic, superlatiu de gracilis («prim» o «estret»).
Distribució
Miocè mitjà, als jaciments de Serpong (Suradita) i Curug Orok (Ci Kandang).
Javastrombus pinguis n. sp. urn:lsid:zoobank.org:act:28C4243B-AB23-4B50-89AD-9DD17F1BEE49
Material tipus
HOLOTIP: CRA-23130-1, de Serpong (Suradita) (Fig. 2). PARATIPS 1-3: CRA-23130-2 a 4, de Serpong (Suradi-
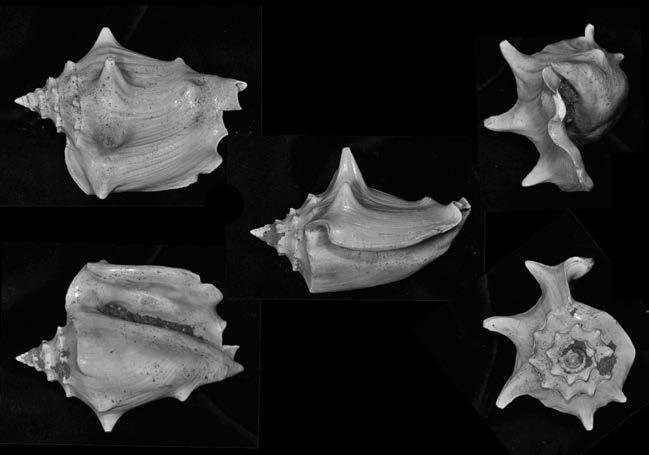
ta). PARATIPS 4 i 5: CRA-23169-1 i 2, de Curug Orok (Ci Kandang).
Diagnosi
Espècie mitjana de Javastrombus, quasi llisa, amb espira molt ampla i un xic còncava, amb una corona de 5 punxes ortogonals, llargues, esmolades i orientades una mica abapicalment, nòduls petits que sobresurten lleugerament per damunt de la sutura, llavi engruixit adapicalment, ample i baix, amb canal posterior poc profund i sinus estromboide poc marcat, darrera volta fortament inflada, espatlla amb quilla supracolumeŀlar molt marcada, 3 punxes dorsals baixes i ben separades, caŀlus columeŀlar poc reflectit i ben estès per la rampa subsutural de l’inici de la darrera volta, i sinus anterior sobtadament recorbat.
Descripció
Conquilla de mida moderada (holotip: 48,2 mm; paratips: entre 50,6 i 46,2 mm), inflada (d’aquí el seu nom específic) i moderadament sòlida, ornada amb una corona formada per 5 punxes robustes i agudes, arrodonides, bastant llargues i eixamplades a la base, lleugerament inclinades abapicalment; hi ha 2 punxes molt baixes, alineades en espiral amb el sinus estromboide, a la part abapical del dors. L’espira és cònica i elevada, eixamplada cap a la base, amb 8,8 voltes de creixement bastant regular i formant un angle general d’uns 66º; el cos de la darrera volta, abans de l’expansió del llavi, és fortament inflat, sobretot adapicalment. El llavi és en forma d’ala, bastant expandit, fortament corbat i prosoclí, i força engruixit just davant del sinus posterior; el marge és arrodonit, però molt prim al sinus posterior, el qual és ample, poc profund i senzill (o molt poc allargassat cap endavant). L’extrem posterior del llavi és molt poc prominent i s’insereix a la darrera volta, recobrint-hi fins a dos terços de la rampa subsutural; a l’extrem anterior el sinus estromboide és relativament poc marcat, a l’igual que el lòbul estromboide. El sinus anterior
és obert, curt i sobtadament recorbat a l’extrem. La columeŀla és engruixida i llisa, unida adapicalment al llavi per damunt de la quilla a l’espatlla; el caŀlus columeŀlar s’estén per més de la meitat de l’amplada de la rampa subsutural a l’inici de la darrera volta. La superfície externa és quasi llisa, lluent, amb estries de creixement irregulars i cordes espirals molt fines i esvaïdes, excepte a la rampa subsutural, on n’hi ha entre 17 i 22 de gruix irregular, més atapeïdes abapicalment. La protoconquilla consta de 2,3 voltes aparentment llises. L’inici de la teleoconquilla està marcat per l’aparició de costelletes axials, ortoclines i molt regulars, que s’estenen al llarg de 3,0 voltes, i s’aturen a 3,5-3,6 voltes de l’obertura; els nòduls subtriangulars (10 a la penúltima volta) són petits i regularment espaiats, i sobresurten lleugerament per damunt la sutura.
Etimologia
El nom específic fa referència a la forma bombada: pinguis és un adjectiu comú al llatí de totes les èpoques, que significa «gras» o «gruixut».
Distribució
Miocè mitjà dels jaciments a Serpong (Suradita) i Curug Orok (Ci Kandang).
Javastrombus cylindratus n. sp. urn:lsid:zoobank.org:act:C243E236-BD98-4685-BAB8-0B149FD94801
Material tipus
HOLOTIP: CRA-23153, de Barangbang (Ciwarak) (Fig. 3). PARATIP: CRA-23128, de Serpong (Suradita).
Diagnosi
Espècie mitjana de Javastrombus, amb espira moderadament ampla i corona de 5 punxes relativament baixes, bastant esmolades però de base ampla, i lleugerament inclinades
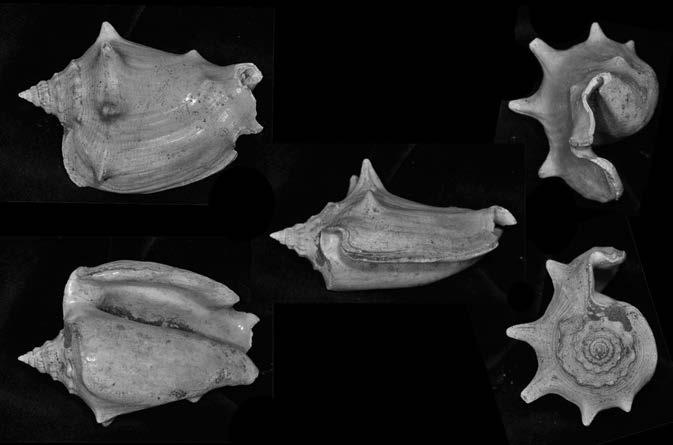
C. R. ALTABA
adapicalment, nòduls molt febles i quasi enfonsats a la sutura, llavi engruixit a la banda adapical, amb canal posterior ben marcat i sinus estromboide petit, cos de la darrera volta cilindroide, 2 punxes dorsals molt baixes i ben separades, caŀlus columeŀlar fortament reflectit i recobrint quasi tota la rampa subsutural de l’inici de la darrera volta, i sinus anterior a penes recorbat.
Descripció
Conquilla de mida moderada (holotip: 51,6 mm; paratip: 50,2 mm), d’aspecte vagament cilindroide (com indica el seu nom específic) i moderadament sòlida, amb una corona de 5 punxes primes, baixes, bastant esmolades però ben eixamplades a la base, i lleugerament inclinades adapicalment. Al dors hi ha 2 punxes baixes i poc marcades, alineades en espiral amb el sinus estromboide al llarg d’una carena molt tènue. L’espira és cònica i elevada, bastant ampla, amb 7,3 voltes de creixement molt regular, amb un angle d’uns 55º; el cos de la darrera volta és moderadament estret, quasi cilíndric i només estretit a l’extrem abapical. El llavi és en forma d’ala moderadament expandida, una mica prominent adapicalment, engruixit a la part central i amb el marge arrodonit; ortoclí a quasi tota la seva longitud, s’aprima al sinus posterior, que és ample i profund, amb l’extrem posterior prominent i recobrint la rampa subsutural fins a arran de la sutura mateixa. El sinus estromboide és petit, però el lòbul estromboide és ben desenvolupat. El sinus anterior és ben eixamplat, breu i molt poc recorbat. La columeŀla és fortament engruixida, llisa, unida adapicalment al llavi superant la forta quilla supracolumeŀlar, la qual forma quasi un angle recte i esbossa una carena. La superfície externa és quasi llisa, amb estries de creixement irregulars i ben marcades, intersectades per cordes espirals molt fines, quasi imperceptibles als extrems del dors, però apreciables a la rampa subsutural, on n’hi ha 14; d’aquestes, les més properes a la sutura són més fortes i
regulars. La protoconquilla consta de 2,3 voltes aparentment llises. L’inici de la teleoconquilla està marcat per l’aparició de costelletes axials, ortoclines i molt regulars, que s’estenen al llarg de només 2,0 voltes, i s’aturen a 3,0 voltes de l’obertura; els nòduls subtriangulars (11 o12 a la penúltima volta) no són gaire regulars i emergeixen lleugerament per damunt de la sutura.
Etimologia
L’epítet cylindratus és un adjectiu i indica una forma general similar a la d’un cilindre.
Distribució
Miocè mitjà dels jaciments a Barangbang (Ciwarak) i Serpong (Suradita).
Javastrombus kecil n. sp. urn:lsid:zoobank.org:act:991035D4-F5FD-4379-8A76-76E706D9AA91
Material tipus
HOLOTIP: CRA-23154-1, de Barangbang (Ciwarak) (Fig. 4). PARATIP 1: CRA-23154-2, de Barangbang (Ciwarak) (Fig. 5). PARATIPS 2 i 3: CRA-23125-1 i 2, de Serpong (Suradita). PARATIPS 4 i 5: CRA-23170, de Curug Orok (Ci Kandang).
Diagnosi
Espècie petita de Javastrombus, amb espira aguda i corona amb 5 punxes fortes, de base ampla però esmolades a l’extrem i inclinades abapicalment, nòduls prominents, aguts i separats de la sutura, llavi rectilini a la seva major part, una mica prosoclí i regularment engruixit, canal posterior i sinus estromboide molt profunds i evidents, darrera volta eixamplada adapicalment, una o dues punxes dorsals baixes i peti-
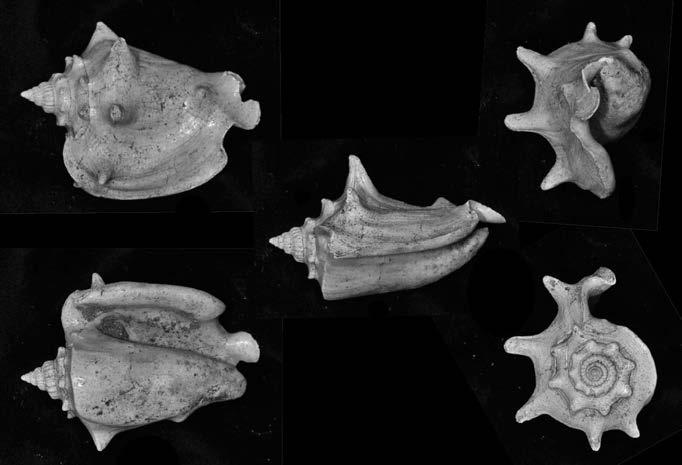
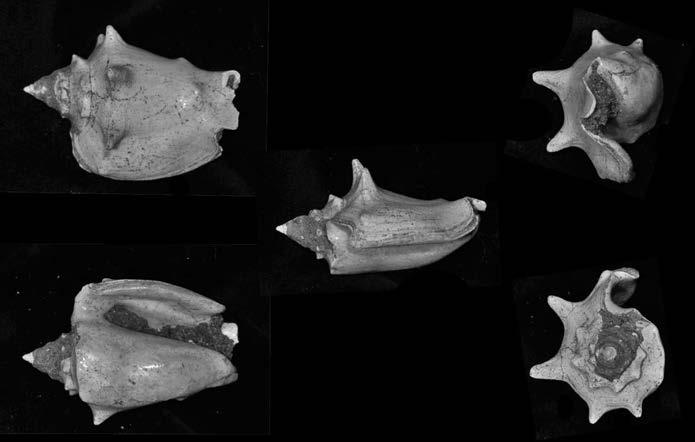
tes, caŀlus columeŀlar engruixit i ben reflectit però poc estès damunt la rampa subsutural de l’inici de la darrera volta, i sinus anterior breu, estret i sobtadament recorbat.
Descripció
Conquilla de mida petita (holotip: 43,2 mm; paratips: entre 35,0 i 42,0 mm), rabassuda i moderadament sòlida, amb una corona formada per 5 punxes amples a la base però esmolades a l’extrem, orientades abapicalment però en el cas de les més llargues (dorsals) lleugerament recargolades adapicalment a l’extrem; hi ha només una o dues punxes baixes al dors, petites però ben marcades. L’espira és cònica i ben elevada, molt estreta, amb només 6,7 voltes de creixement regular i forma un angle d’uns 50º; el cos de la darrera volta, abans de l’expansió del llavi, és cònic, fortament eixamplat
adapicalment. El llavi és en forma d’ala, moderadament expandit i regularment engruixit, amb orientació lleugerament prosoclina i quasi recte a la seva major part; el marge és arrodonit, però molt prim al sinus posterior, el qual és mitjanament ample però ben profund. L’extrem posterior del llavi és força prominent i recobreix fins a un terç o la meitat de la rampa subsutural; a l’extrem anterior el sinus estromboide és molt marcat i conspicu, com ho és també el lòbul estromboide. El sinus anterior és estret, curt i sobtadament recorbat. La columeŀla és molt eixamplada, ben engruixida i llisa, unida adapicalment al llavi per damunt de la quilla supracolumeŀlar, la qual és molt marcada i forma una carena una mica prominent. La superfície externa és quasi llisa, amb estries de creixement irregulars i molt fines, que s’entrecreuen amb cordes espirals també molt primes; aquestes són apreciables
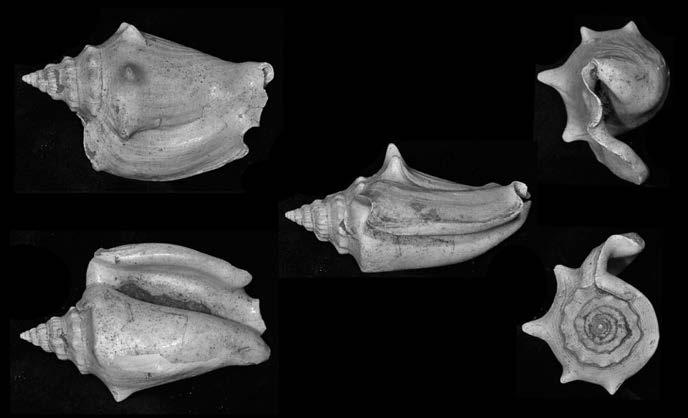
C. R. ALTABA
a la meitat adapical de la darrera volta, incloent la rampa subsutural, a la qual n’hi ha entre 14 i 16, irregulars. La protoconquilla consta de 2,1 voltes aparentment llises. L’inici de la teleoconquilla està marcat per l’aparició de costelletes axials, ortoclines i molt regulars, que s’estenen al llarg de 2,7 voltes, i s’aturen a només a 1,6-2,4 voltes de l’obertura; els nòduls són prominents, aguts i subcònics (8 a la penúltima volta), estan regularment espaiats i se situen per damunt la sutura, de la qual en sobresurten marcadament.
Etimologia
Kecil, l’epítet específic d’aquesta espècie, és un adjectiu pres directament de l’indonesi, que significa ‘petit’.
Distribució
Miocè mitjà dels jaciments a Barangbang (Ciwarak), Serpong (Suradita) i Curug Orok (Ci Kandang).
Javastrombus subinermis n. sp. urn:lsid:zoobank.org:act:DBCFB470-DA3F-4909-9AE3D1EFEDB100C0
Material tipus
HOLOTIP: CRA-23172 (Fig. 6).
Diagnosi
Espècie mitjana de Javastrombus, amb espira molt aguda i corona amb només 4 punxes baixes, arrodonides i ortogonals, nòduls clarament atenuats i molt propers a la sutura, llavi rectilini a la seva major part, ortoclí i regularment engruixit, canal posterior i sinus estromboide moderadament profunds, darrera volta poc eixamplada adapicalment, amb una sola punxa dorsal només incipient, caŀlus columeŀlar ben reflectit i adherent damunt tota la rampa subsutural de l’inici de la darrera volta, i sinus anterior breu, estret i recorbat només a l’extrem.
Descripció
Conquilla de mida mitjana (50,0 mm), esvelta i moderadament sòlida, amb només 4 punxes petites, baixes i de base estreta, còniques però ben arrodonides a l’extrem, ortogonals a l’eix d’enrotllament. El dors només presenta un esbós de punxa, molt baix, adjacent al sinus estromboide. L’espira és cònica i ben elevada, molt estreta, amb 7,7 voltes de creixement sumament regular, i forma un angle de només 45º; el cos de la darrera volta, abans de l’expansió del llavi, és subcilíndric, poc eixamplat adapicalment. El llavi és en forma d’ala, moderadament expandit i més engruixit al mig, amb orientació lleugerament ortoclina, quasi recte excepte als extrems; el marge és arrodonit, però molt prim al sinus posterior, el qual és molt profund i eixamplat anteriorment. L’extrem posterior del llavi no és pas prominent, però s’estén per tota la rampa subsutural i és adherent excepte a l’extrem adapical; a l’extrem anterior el sinus estromboide és moderadament ample i profund, però el lòbul estromboide és molt menys marcat. El sinus anterior és estret, molt curt però recorbat. La columeŀla és eixamplada, engruixida i llisa, unida adapicalment al llavi per damunt de la quilla supracolumellar; aquesta és ben marcada, però sense destacar en el conjunt de l’espira. La superfície externa és quasi llisa, amb estries de creixement irregulars, en general molt fines, però formant lleus solcs colabrals regularment espaiats; a més, té fines cordes espirals, perceptibles a tot el dors i una mica més fortes a la rampa subsutural, on n’hi ha 13 bastant iguals. La protoconquilla consta de 2,2 voltes aparentment llises. A l’inici de la teleoconquilla apareixen costelletes axials, ortoclines i molt regulars, que s’estenen al llarg de 2,8 voltes, i s’aturen a 2,7 voltes de l’obertura; els nòduls de l’espira són roms però elevats, i estan arrenglerats amb poc ordre just per damunt la sutura (13 a la penúltima volta).
Etimologia
L’epítet específic denota la reduïda armadura d’aquesta conquilla: subinermis és un adjectiu que significa desarmat, o sense blindatge.
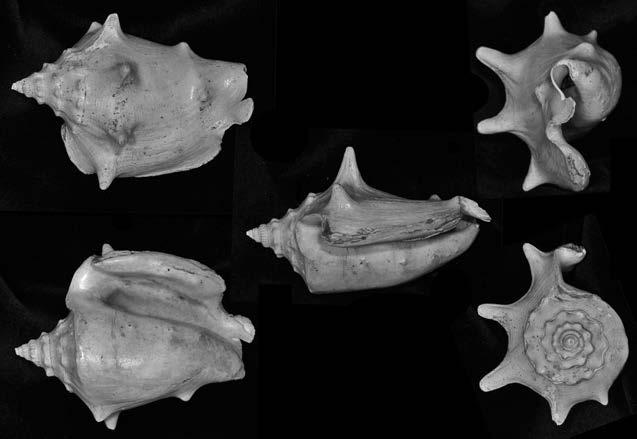
Distribució
Miocè mitjà del jaciment a Curug Orok (Ci Kandang).
Javastrombus sondaicus n. sp. urn:lsid:zoobank.org:act:B532BB94-3DD9-4C94-8E99DE3938B7DAE1
Material tipus
HOLOTIP: CRA-23090, de Curug Rahong (Mekarjaya) (Fig. 7). PARATIPS 1 i 2: CRA-23103-1 i 2, de Serpong (Suradita). PARATIP 3: CRA-23168, de Curug Orok (Ci Kandang).
Diagnosi
Espècie robusta i grossa de Javastrombus, amb espira molt ampla i ornada de nòduls suprasuturals sobresurten poc per damunt de la sutura, llavi engruixit amb canal posterior profund i sinus estromboide ample, darrera volta túmida, corona amb 5 punxes arrodonides i orientades abapicalment, 2 punxes dorsals molt baixes i ben separades, cal·lus columeŀlar reflectit que recobreix els nòduls de l’inici de la darrera volta, i sinus anterior lleugerament recorbat.
Descripció
Conquilla de mida grossa (holotip: 54,5 mm; paratips: entre 55,7 i 47,3 mm), robusta i moderadament sòlida, amb una filera de 5 punxes aprimades distalment, arrodonides a l’extrem, amb base ampla i només un xic inclinades abapicalment, tot i que ho semblen més; hi ha 2 punxes dorsals, molt baixes però conspícues, alineades en espiral amb el sinus estromboide, al llarg d’una carena espiral summament tènue a la part abapical del dors. L’espira és cònica i elevada, una mica estretida apicalment, amb 8,5 voltes de creixement ben regular, i forma un angle d’uns 62º; el cos de la darrera volta, abans de l’expansió del llavi, és voluminós, túmid. El llavi és en forma d’ala, moderadament expandit, i força engruixit al tram central, prosoclí i quasi recte; el marge és arrodonit, però molt prim al sinus posterior, el qual és força ample, molt profund i allargassat cap endavant. L’extrem posterior del llavi és notablement prominent, però només recobreix una petita fracció de la rampa subsutural; a l’extrem anterior, el sinus estromboide és ben marcat i ample, adjacent al lòbul estromboide, el qual és prominent i fortament torçat. El sinus anterior és ample, curt i a penes recorbat. La columeŀla és ben engruixida i llisa, unida adapicalment al llavi per damunt de la quilla supracolumeŀlar, la qual és fortament marcada i esbossa una carena. La superfície externa és quasi llisa, amb estries de creixement irregulars, intersectades a la part adapical per cordes espirals molt fines, les quals són perceptibles a la zona del vessant abapical de les punxes, però molt més conspícues a la rampa subsutural; a aquesta zona n’hi ha 13 de principals, i a partir de la quarta des de la sutura n’apareixen d’altres de secundàries, molt més primes, intercalades entre les principals. La protoconquilla consta de 2,4 voltes aparentment llises. L’inici de la teleoconquilla està marcat per l’aparició de costelletes axials, ortoclines i molt regulars, que s’estenen al llarg de 2,3 voltes, i s’aturen a 3,8 voltes
de l’obertura; els nòduls de l’espira (12 o 13 a la penúltima volta) són molt petits, roms i molt baixos, estan regularment espaiats i amb prou feines sobresurten de la sutura.
Etimologia
El nom específic d’aquesta espècie, sondaicus, és un adjectiu gentilici de la regió insular de la Sonda.
Distribució
Miocè mitjà dels jaciments a Curug Rahong (Mekarjaya), Serpong (Suradita) i Curug Orok (Ci Kandang).
Discussió
La diversitat descoberta dins el nou gènere Javastrombus és un indicador de la riquesa de les faunes del Miocè mitjà de Java, com també de l’esforç que encara hi manca per assolirne una comprensió adequada. L’ordre de les diferències entre les morfoespècies identificades és el mateix que el de les diferències que existeixen entre espècies vivents pertanyents a gèneres diversificats dins la família dels estròmbids. D’altra banda, hi ha diverses espècies que procedeixen del mateix jaciment i que vivien juntes, la qual cosa es pot interpretar com una prova de l’existència d’espècies en simpatria. Per tot això, considerar-les com a espècies distintes és raonable; en tot cas, ha de permetre’n un estudi més aprofundit. Per facilitar-ne la comparació i determinació, es pot seguir la següent clau dicotòmica.
1 Dors amb només un esbós de punxa, però amb línies espirals evidents .................................. J. subinermis
– Amb 2 o 3 punxes baixes al dors, on no hi ha línies espirals o hi són molt tènues ..................................2
2 Cos de la darrera volta ben inflada, punxes de la corona orientades abapicalment (almenys a la base).......3
– Cos de la darrera volta estreta, punxes de la corona orientades ortogonalment o lleugerament adapicalment, punxes dorsals al llarg d’una tènue corda espiral ........................................................................... 5
3 Mida relativament grossa, rampa subsutural amb poques cordes desiguals J. sondaicus – Mida mitjana o petita, rampa subsutural amb cordes nombroses i força iguals.........................................4
4 Espira molt ampla amb nòduls que sobresurten lleugerament per damunt de la sutura, quilla supracolumeŀlar ben marcada, superfície llisa, mida mitjana ......................... J. pinguis
– Espira molt estreta amb nòduls forts i ben separats de la sutura, quilla supracolumeŀlar poc marcada, teleoconquilla breu, mida petita ............................ J. kecil
5 Esvelt, espira molt estreta, rampa subsutural amb cordes més fortes cap a la sutura, sinus estromboide ben marcat i canal anterior fortament recorbat .......................................................... J. praegracilis
C. R. ALTABA
– Cos de la darrera volta cilindroide, espira moderadament estreta, rampa subsutural amb cordes un xic més fortes arran de la corona de punxes, sinus estromboide petit i canal anterior a penes recorbat......
J. cylindratus
L’àrea on es dipositaren els sediments que han fornit exemplars de Javastrombus devia ser, al llarg del Miocè, una plataforma marina extensa i d’escassa fondària, amb àrees carbonàtiques (Hall, 2013; Morley et al., 2016). La disparitat en l’escultura de les diferents espècies descrites aquí podria indicar una intensitat variable en la pressió dels depredadors; en el cas d’altres membres de la mateixa família, s’ha atribuït a aquesta causa el desenvolupament iteratiu d’espines (Landau et al., 2011). En aquell medi, doncs, hi havia interaccions de la fauna marina que eren tan intenses i variades com les que hi ha a les mars tropicals actuals, o potser més.
L’origen del gènere Javastrombus cal cercar-lo en el grup format pels gèneres Persististrombus i Afristrombus, tot i que Javastrombus ocupa una posició geogràfica allunyada dels altres dos gèneres citats. Afristrombus actualment es troba tan sols a les costes africanes occidentals tropicals, però durant el Neogen estava estès pel Paratetis d’Europa central i la regió mediterrània. En canvi, Persististrombus es trobava distribuït per l’àrea mediterrània durant l’Oligocè i el Miocè, però des del Miocè també es troba a les costes tropicals atlàntiques i pacífiques d’Amèrica. Persististrombus i Afristrombus estan relacionats filogenèticament, però la seva divergència és anterior a la diversificació de la majoria d’estròmbids americans (Latiolais et al, 2006). Cal destacar que Afristrombus fou descrit vàlidament per Bandel (2007), de manera que el nom de substitució Tethystrombus Dekkers, 2008 és un sinònim posterior. El nou gènere Javastrombus representa una branca d’aquest grup que s’hauria diversificat en aïllament, en un medi favorable, estable i complex, al llarg del Miocè mitjà, durant el Languià i el Serravaŀlià (entre 16,0 i 11,6 Ma).
La relació de Javastrombus amb els gèneres propers esmentats podria indicar que es tracta de tàxons parafilètics, però això no ha de representar un problema insalvable si considerem les classificacions en diacronia (Cela-Conde & Altaba, 2002; Carter et al., 2011, 2015). En qualsevol cas, identificar i anomenar la radiació evolutiva dins Javastrombus sembla útil i correcte.
La diversificació de Javastrombus i gèneres propers sembla atribuïble, almenys en bona part, a processos geològics que afectaren la continuïtat o l’aïllament dels seus hàbitats. Així, la complexa reorganització del Paratetis en el context europeu (Palcu & Krijgsman, 2021; Palcu et al., 2023) sembla el motor evolutiu de la família Strombidae en el Miocè de la mar Paratetis al sud i centre d’Europa (Harzhauser & Kronenberg, 2013). A l’altre extrem del Vell Món, l’aparició d’una geografia canviant i dinàmica deguda al xoc de la placa indoaustraliana amb la placa euroasiàtica (Aribowo et al., 2022) sembla la responsable de l’extrema diversificació de molts grups marins que hi va haver a la zona d’Insulíndia (Williams & Duda, 2008). Altres casos de diversificació
comparable dins de la família Strombidae s’han documentat a l’istme de Panamà (Landau et al., 2011) i a la regió dels Everglades, al sud de la península de Florida (Petuch, 2019; Petuch & Berschauer, 2021). De tota manera, cal tenir present que, a banda d’aquests processos de vicariança, hi ha hagut dispersió a llarga distància que ha jugat un paper important. Això últim ho posa de relleu l’existència d’espècies de moŀluscs amb distribució amfiatlàntica al llarg del Neogen (Manganelli & Spadini, 2010). Destriar els processos biogeogràfics requerirà una anàlisi filogenètica i un coneixement adient de la situació estratigràfica dels diferents Javastrombus
La complexitat filogenètica descoberta recentment és tan sols un primer pas. Cal més material i un estudi adequat dels jaciments per conèixer la distribució temporal de les diverses espècies i els patrons filogenètics i adaptatius dins aquesta radiació evolutiva.
Els exemplars fòssils foren recoŀlectats amb entusiasme per Rais M. Razes i Muhamad Yusuf, excepte l’holotip de J. sondaicus, que me’l va cedir gentilment Josep Juárez-Ruiz. Ed Petuch va atendre amablement diferents consultes sobre faunes actuals i fòssils.
Aribowo, S., Husson, L., Natawidjaja, D. H., Authemayou, C., Daryono, M. R., Puji, A. R., Valla, P. G., Pamumpuni, A., Wardhana, D. D., de Gelder, G., Djarwadi, D. & Lorcery, M. 2022. Active back-arc thrust in North West Java, Indonesia. Tectonics, 41: e2021TC007120. https://doi.org/10.1029/2021TC007120
Aswan, E. S. 2021. Depositional environmental evolution of Nyalindung Formation based on paleontology molluscan study, Ciodeng area, Sukabumi, West Java, Indonesia. Riset Geologi dan Pertambangan, Indonesian Journal of Geology and Mining, 31: 117-130.
Aswan, E. S., Kistiani, D., Abdurrahman, I. Y., Santoso, W. D., Rudyawan, A. & Oo, T. Z. 2017. Late Miocene molluscan stage of Jawa insight from new field studies. IOP Conference Series: Earth and Environmental Science, 71: 012031. doi:10.1088/1755-1315 /71/1/012031
Bandel, K. 2007. About the larval shell of some Stromboidea, connected to a review of the classification and phylogeny of the Strombimorpha (Caenogastropoda). Freiberger Forschungshefte, C, 524: 97-206.
Carter, J. G., Altaba, C. R., Anderson, L. C. et al. 2011. A synoptical classification of the Bivalvia (Mollusca). Paleontological Contributions, 4: 1-47.
Carter, J. G., Altaba, C. R., Anderson, L. C., Campbell, D. C., Fang, Z., Harries, P. J. & Skelton, P. W. 2015. The paracladistic approach to phylogenetic taxonomy. Paleontological Contributions, 12: 1-9.
Cela-Conde, C. & Altaba, C. R. 2002. Multiplying genera versus moving species: a new taxonomic proposal for the family Hominidae. South African Journal of Science, 98: 1-4.
Clements, B. & Hall, R. 2007. Cretaceous to Late Miocene stratigraphic and tectonic evolution of West Java. P. 617-634. In: 31st
Annual Convention of the Indonesian Petroleum Association. Indonesian Petroleum Association (IPA). Jakarta. Indonèsia. 1192 p.
Clezard, A. & Dovesi, M. 2023. A New species of fossil Pseudosimnia (Gastropoda: Ovulidae) from the Miocene of West Java, Indonesia. The Festivus, 55:154-164.
Dekkers, A. M. 2002. About abnormalities on the number of eyes and the evolution of the possible eye-sight related shell aspects in Strombidae; introducing new shell terms in Strombidae morphology (Gastropoda: Stromboidea, Strombidae). The Festivus, 53: 163-181.
Dekkers, A. M. 2023. A treatise on the Taxonomy of Strombidae. 2023. Acta Malacologica Inquisitionis, Journal of Malacological Research, 1: 1-44.
Dekkers, A. M., Liverani, V., Ćorić, S., Maxwell, S. J. & Landau, B. M. 2020. A new genus for Indo-Pacific fossil strombids, and two new species from the Miocene of Java and Borneo (Caenogastropoda, Strombidae). Basteria, 84: 1-9.
Dovesi, M. & Parsons, J. 2021. Two new species of Barycypraea (Gastropoda: Cypraeidae) from the Miocene of Java, Indonesia. The Festivus, 53: 116-126.
Hall, R. 2013. The palaeogeography of Sundaland and Wallacea since the Late Jurassic. Journal of Limnology, 72: 1-17.
Harzhauser, M. & Kronenberg, G. C. 2013. The Neogene strombid gastropod Persististrombus in the Paratethys Sea. Acta Palaeontologica Polonica, 58: 785-802.
Hoek Ostende, L. W. van den, Leloux, J., Wesselingh, F. P. & Winkler Prins, C. F. 2002. Cenozoic Molluscan types from Java (Indonesia) in the Martin Collection (Division of Cenozoic Mollusca), National Museum of Natural History, Leiden. NNM Technical Bulletin, 5: 1-130, 3 pls.
Kapid, R., Santoso, W. D., Ikhsan, B., Jambak, M. A. & Irawan, D. E. 2019. The Mid Miocene Climatic Optimum (MMCO) indication at low latitude sediment case study: the Miocene Cibulakan Formation, Bogor Basin, Indonesia. International Journal on Advanced Science Engineering Information Technology, 2: 594-600.
Kase, T., Kurihara, Y., Aguilar, Y. M., Pandita, H., S. Fernando, A, G & Hayashi, H. 2015. A new cerithioidean genus Megistocerithium (Gastropoda; Mollusca) from the Miocene of Southeast Asia: a possible relict of Mesozoic “Eustomatidae”. Paleontological Research, 19: 299-311.
Kesuma, N. B. & Aswan. 2022. Paleocurrent study of Nyalindung Formation through mollusks. Paleontological aspects in Ci Galasar River, Sukabumi, West Java. Bulletin of Geology, Institut Teknologi Bandung, 6: 919-933.
Landau, B. M., Beu, A. G., Breitenberger, A. & Dekkers, A. M. 2020a. Middle Miocene tonnoidean gastropods from near Wonosari, Yogyakarta, Java, Indonesia. Basteria, 84: 10-25.
Landau, B., Kronenberg, G. C., Herbert, G. & Silva, C. M. da 2011. The genus Strombus (Mollusca: Caenogastropoda: Strombidae) in the Neogene of the Bocas del Toro area, Panama, with the description of three new species. Journal of Palaeontology, 85: 337-352.
Landau, B. M., Raven, J. G. M., Breitenberger, A. E. & Dekkers, A. M., 2020b. Semiricinula preturbinoides spec. nov., a new species from the Miocene of Java (Gastropoda: Muricidae). Basteria, 84: 131-134.
Latiolais, J. M., Taylor, M. S., Roy, K. & Hellberg, M. E. 2006. A molecular phylogenetic analysis of strombid gastropod morphological diversity. Molecular Phylogenetics and Evolution, 41: 436-444.
Li, F., Zheng, J., Ma, Q., Gu, Z., Wang, A.,Yang, Y. & Liu, C. 2022. Phylogeny of Strombidae (Gastropoda) based on mitochondrial genomes. Frontiers in Marine Science, 9: 930910. doi: 10.3389/ fmars.2022.930910
Liverani, V. & Wieneke, U. 2022. Some more species in the genus Spinatus Dekkers et al., 2020 (Gastropoda: Strombidae). Conchylia, 52: 81-87.
Manganelli, G. & Spadini, V. 2010. Onustus plioextensus (Sacco 1896) (Gastropoda: Caenogastropoda): a Mediterranean Pliocene xenophorid with western Atlantic relationships? Journal of Conchology, 40: 333-338.
Martin, K. 1879-1880. Die Tertiärschichten auf Java, nach den Entdeckungen von Fr. Junghuhn. Brill, Leiden. 164 p., 28 pls., 1 mapa. Martin, K. 1891-1906. Die Fossilien von Java auf grund einer Sammlung von Dr. R. D. M. Verbeek und von anderen I. Band. Gasteropoda. Mit einem Anhange über: Die Foraminiferen führenden Gesteine. Sammlungen des geologischen Reichs-Museums in Leiden, Neue Folge, 1: 1-331.
Maxwell, S. J., Dekkers, A. M., Rymer, T. L. & Congdon, B. C. 2020. Towards resolving the American and West African Strombidae (Mollusca: Gastropoda: Neostromboidae) using integrated taxonomy. The Festivus, 52: 3-38.
Merle, D., Landau, B. M. & Breitenberger, A.E. 2021. New Muricidae (Mollusca, Gastropoda) from the Miocene of Java (Indonesia). Basteria 85: 21-33.
Morley, R. J., Morley, H. P. & Swiecicki, T. 2016. Mio-Pliocene palaeogeography, uplands and river systems of the Sunda region based on mapping within a framework of VIM depositional cycles. P. 1303-1328. In: 40th Annual Convention of the Indonesian Petroleum Association. Indonesian Petroleum Association (IPA). Jakarta. Indonèsia. 2005 p.
Palcu, D. V. & W. Krijgsman, W. 2021. The dire straits of Paratethys: Gateways to the anoxic giant of Eurasia. Geological Society, London, Special Publications, 523: 111-139.
Palcu, D. V., Mariș, I., de Leeuw, A., Melinte-Dobrinescu, M., Anton, E., Frunzescu, D., Popov, S., Stoica, M. & Jovane, L. 2023. The legacy of the Tethys Ocean: Anoxic seas, evaporitic basins, and megalakes in the Cenozoic of Central Europe Wout Krijgsman. Earth-Science Reviews, 246: 104594. https://doi. org/10.1016/j.earscirev.2023.104594
Petuch, E. J. 2019. Cenozoic seas. The view from Eastern North America. CRC Press, Boca Raton. 324 p.
Petuch, E. J. & Berschauer, D. P. 2021. Ancient seas of southern Florida: the geology and paleontology of the Everglades Region. 2nd ed. CRC Press, Boca Raton & Oxon. 284 p.
Robba, E. 1996. The Rembangian (Middle Miocene) mollusk-fauna of Java, Indonesia. I. Archaeogastropoda. Rivista Italiana di Paleontologia e Stratigrafia, 102: 267-292.
Robba, E. 2013. Tertiary and Quaternary fossil pyramidelloidean gastropods of Indonesia. Scripta Geologica, 144: 1-191.
Roy, K., Balch, D. P. & Hellberg, M. E. 2001. Spatial patterns of morphological diversity across the Indo-Pacific: analyses using stromboid gastropods. Proceedings of the Royal Society of London, B, 268: 2503-2508.
Savazzi, E. 2007. Constructional morphology of strombid gastropods. Lethaia, 24: 311-331.
Smyth, H., Hall, R., Hamilton, J. & Kinny, P. 2005. East Java: Cenozoic basins, volcanoes and ancient basement. P. 251266. In: 30th Annual Convention of the Indonesian Petroleum Association. Indonesian Petroleum Association (IPA). Jakarta. Indonèsia. 1192 p.
Simone, L. R. L. 2005. Comparative morphological study of representatives of the three families of Stromboidea and the Xenophoroidea (Mollusca, Caenogastropoda), with an assessment of their phylogeny. Arquivos de Zoologia. São Paulo, 37(2): 141-267.
Syarifin, 2011. Paleontologi formasi Nyalindung. Bulletin of Scientific Contribution, 9: 17-27.
C. R. ALTABA
Wieneke, U., Stoutjesdijk, H. & Simonet, P. (eds.) 2024. Gastropoda Stromboidea. Disponible en: http://www.stromboidea.de/ [Data de consulta: 12 febrer 2024]
Wilson, M. E. J. 2002. Cenozoic carbonates in Southeast Asia: implications for equatorial carbonate development. Sedimentary Geology, 147: 295-428.
Williams, S. T. & Duda, T. F. 2008. Did tectonic activity stimulate Oligo-Miocene speciation in the Indo-West Pacific? Evolution, 62: 1618-1634.
Juli Mauri1, Andreu Ubach2, Sergi Herrando3,4, Marc Anton4, Marc Fusellas4 i Constantí Stefanescu2
1 Doctorand en el programa de Gestió de la Biodiversitat de la Universitat Autònoma de Barcelona. Edifici C Facultat de Cièncias i Biocièncias. 08193 Bellaterra, Barcelona.
2 Grup de Recerca BiBio. Museu de Ciències Naturals de Granollers. C/ Francesc Macià, 51. 08402 Granollers.
3 CREAF, Centre de Recerca Ecològica i Aplicacions Forestals. Campus de Bellaterra (UAB) Edifici C. 08193 Cerdanyola del Vallès, Barcelona.
4 Institut Català d’Ornitologia. Nat- Museu de Ciències Naturals de Barcelona. Pl. Leonardo da Vinci, 4-5. 08019 Barcelona.
Autor de correspondència: Juli Mauri i de los Rios, jmaurir@gmail.com
Rebut: 01.04.2024; Acceptat: 06.05.2024; Publicat: 30.06.2024
Resum
Aquest article analitza l’estat i la tendència de la biodiversitat a la Regió Metropolitana de Barcelona (RMB) durant vint anys (20022022), utilitzant dades estandarditzades de dos grups faunístics, els ocells i les papallones , que són bioindicadors efectius dels canvis en hàbitats, ecosistemes i paisatges. Aquests grups són àmpliament utilitzats, tant a Catalunya com a Europa, en xarxes de seguiment de biodiversitat basats en ciència ciutadana, que permeten estudiar els canvis de les poblacions d’animals salvatges. L’article examina les tendències poblacionals d’aquests grups en relació amb el grau de protecció legal, i proporciona així informació rellevant sobre l’eficàcia de la protecció d’espais naturals per a la conservació de la biodiversitat. En els últims 20 anys, la RMB ha tingut una estabilitat en les poblacions d’ocells, llevat dels que habiten espais oberts naturals. En contrast, les poblacions de papallones han experimentat un declivi, particularment en zones protegides, amb una pèrdua notable d’espècies especialistes. Això reflecteix un possible deteriorament dels hàbitats. Els espais del Pla d’Espais d’Interès Natural (PEIN), destinats a la conservació de la biodiversitat, no han aconseguit millorar la tendència general de les poblacions d’ocells i papallones.
Paraules clau: Ocells, papallones, biodiversitat, PEIN (Pla d’Espais d’Interès Natural), boscos, espais oberts, conservació, tendències poblacionals, bioindicadors, Regió Metropolitana de Barcelona, NE península Ibèrica, Conca Mediterrània.
Abstract
20 years of changes in the biodiversity of birds and butterflies in the Metropolitan Region of Barcelona
This article analyses the state and trend of biodiversity in the Barcelona Metropolitan Region (RMB) for twenty years (2002-2022), using standardised data from two different taxa, birds and butterflies, which are effective bioindicators of habitats, ecosystems and landscapes changes. These groups are widely used, both in Catalonia and in Europe, in biodiversity monitoring networks based on citizen science. The article examines the population trends of these groups in relation to the legal protection, thus providing relevant information on the effectiveness of the protection of natural areas for the conservation of biodiversity. In the last 20 years, the RMB has seen a stability in bird populations, except for those that inhabit natural open spaces. In contrast, butterflies have experienced a decline, particularly in protected areas, with a noticeable loss of specialist species. This reflects possible habitat deterioration. Protected Areas, intended for the conservation of biodiversity, have not improved the situation of birds or butterflies.
Key words: Birds;, butterflies, biodiversity, PEIN (Plan of Areas of Natural Interest), forests, open areas, conservation, population trends, bioindicators, Metropolitan Region of Barcelona, NE Iberian Peninsula, Mediterranean Basin. Butlletí de la
Introducció
En les darreres dècades, més de la meitat de la població mundial s’ha concentrat en zones urbanes, una proporció que es preveu que sigui del 66 % el 2050 (Nacions Unides, 2014). Aquest creixement suposa que des d’inicis del segle XXI el 85% dels europeus viu en una ciutat (Boada et al., 2019). Hom pot veure com l’ocupació territorial i el desplaçament progressiu de la població cap a les àrees urbanes sotmeten cada dia més els espais oberts urbans i periurbans a fricci-
ons i pressions molt importants, amb una tendència a la banalització i l’empobriment dels hàbitats i ecosistemes. Així doncs, a les regions densament poblades com les àrees metropolitanes, la pèrdua de biodiversitat es veu molt acusada per fenòmens de pertorbació ambiental i la fragmentació dels hàbitats per efecte de barrera. Aquest és de fet un dels factors de canvis ambientals que, juntament amb d’altres com la intensificació de l’agricultura, la destrucció de les selves tropicals i el canvi climàtic, fan que molts experts afirmin que ens
J. MAURI ET AL.
trobem davant la sisena gran extinció (Barnosky et al., 2011). Segons dades de l’UICN, aquesta extinció ha provocat ja la disminució significativa de molts taxons com insectes, molluscs i amfibis (Almond et al., 2020. Veure també https:// www.iucnredlist.org/).
La conca mediterrània i les muntanyes alpines circumdants estan entre les regions biogeogràfiques biològicament més riques del món i la seva fauna d’insectes i d’ocells és especialment diversa (Kudrna et al., 2011, Keller et al., 2020), però sense estar exempta d’aquestes pressions. La Regió Metropolitana de Barcelona (RMB) està situada a la zona central de la Depressió Litoral i la Serralada Prelitoral de Catalunya amb una extensió de 323.499 ha i una població estimada en més de cinc milions d’habitants l’any 2019 (Llobet et al., 2020). A la RMB el canvi en els usos del sòl ha estat molt rellevant a nivell ecològic i de paisatge, i ha comportat un fenomen greu de pèrdua d’espais oberts així com l’aïllament progressiu dels espais naturals que resten sense urbanitzar. Tot i que es va elaborar el Plan de ordenación de Barcelona y su zona de influencia (1953), no és fins l’any 1956 que s’aprova la primera Llei Estatal del Sòl (a nivell de tota Espanya) i fins el 1975 no s’aprova la Llei del Sòl a Catalunya; finalment, el primer instrument regulador d’aquesta regió s’instaura el 1976 amb el Pla General Metropolità de Barcelona. Per tant, és normal que aquest enorme creixement que transforma els hàbitats de la RMB s’hagi fet durant molts anys sense una planificació conservacionista des del punt de vista de la biodiversitat. Un dels resultats d’aquest desenvolupament és la “creació” de múltiples bosses d’espais oberts, principalment aïllats, que resten sense urbanitzar i, sovint, sense una planificació que defineixi els seus usos, almenys fins fa ben poc. Per altra banda, el 1985 la Generalitat de Catalunya aprova el Pla d’Espais d’Interès Natural (PEIN) com una mesura territorial de conservació dels hàbitats naturals, del paisatge i de la biodiversitat (a nivell de tota Catalunya). La superfície PEIN dins la RMB és de 61.609 ha, el que suposa la protecció ambiental del 19,04 % del seu territori.
Les espècies comunes són molt importants en la conservació dels hàbitats i dels ecosistemes, ja que fins i tot declivis percentualment petits de la seva abundància poden provocar grans pèrdues absolutes d’individus i de biomassa, que esdevenen fenòmens que pertorben significativament l’estructura dels ecosistemes (Gaston et al., 2008). Aquestes espècies comunes són essencials per a la configuració dels ecosistemes, proporcionant estructura i serveis. Sovint, aquestes espècies són també les «dominants» o «estructurals» que defineixen els diferents tipus d’hàbitats (Gaston et al., 2008). Justament en aquesta regió, els ocells i les papallones diürnes que són comunes han estat utilitzats a les darreres dècades com a bioindicadors dels canvis en els hàbitats en programes de seguiment de llarga trajectòria i ben consolidats, com el Seguiment d’Ocells Comuns de Catalunya –amb sèries de dades des de 2002–, i el Catalan Butterfly Monitoring Scheme –que atresora dades de seguiment des de 1994–.
Els ocells són coneguts indicadors ambientals que han estat molt utilitzats en l’avaluació de la resposta dels organismes enfront les modificacions de l’entorn (Furness &
Greenwood, 1993). Aquesta capacitat per indicar els canvis que es produeixen en el medi és deguda, entre d’altres coses, a estar a la part alta de les cadenes tròfiques, essent doncs particularment sensibles a les alteracions de qualsevol esglaó inferior. Per altra banda, el fet que hi hagi espècies adaptades a diferents hàbitats, tant oberts com tancats, de climes secs o humits, d’ambients antròpics o allunyats de la presència humana, els dona en conjunt una gran capacitat indicadora dels canvis ambientals (Gregory & van Strien, 2010).
Els ropalòcers o papallones diürnes es troben entre els grups de fauna silvestre més sensibles als canvis ambientals, tant els paisatgístics com els climàtics (Thomas, 2005). Es caracteritzen per tenir un cicle de vida curt, una alta especialització alimentària (sobretot en la fase larvària) i una forta afinitat amb hàbitats específics durant diferents etapes del seu cicle vital. En síntesi, les papallones depenen en gran mesura de la qualitat de l’hàbitat i de la disponibilitat de plantes, ja sigui de plantes nutrícies de les fases larvàries, com de la disponibilitat de plantes nectaríferes, però també de la presència d’herbes que serveixen de refugi tant als adults com a les larves. Per altra banda s’ha demostrat que les papallones especialistes d’hàbitat es veuen molt més afectades que les generalistes pels canvis paisatgístics (Stefanescu et al., 2011). En aquest context de canvi amb declivis importants d’amplis grups d’espècies, l’estudi de les tendències de grups bioindicadors en una regió fortament humanitzada com la RMB pot aportar informació important per entendre la problemàtica que afecta la pèrdua de la biodiversitat en el marc de les conurbacions europees i especialment mediterrànies. Tenint en compte totes les consideracions prèvies, aquest treball pretén avaluar: 1) les tendències de la biodiversitat d’ocells i papallones en el conjunt de la RMB en els darrers 20 anys, 2) els patrons de riquesa mitjana d’ocells i papallones a la RMB dins i fora dels espais naturals protegits, 3) les possibles diferències en aquestes tendències entre hàbitats diferents i dins i fora dels espais naturals protegits de la RMB.
Materials i mètodes
Obtenció de les dades i situació geogràfica
Per al desenvolupament dels indicadors d’ocells i papallones de la RMB, s’han obtingut les dades del Seguiment d’Ocells Comuns de Catalunya (SOCC), i del Catalan Butterfly Monitoring Scheme (CBMS), ambdós basats en la recollida de dades de camp mitjançant ciència ciutadana.
El SOCC és un projecte científic iniciat l’any 2002 que recull dades d’ocells d’itineraris fixats (vegeu https://socc.cat/ socc/projecte/ ) i es basa en el transsecte com a mètode de camp. Durant el cens l’observador anota tots els ocells que va detectant, ja sigui visualment o auditiva, al llarg d’un itinerari de 3 km. El SOCC permet estudiar tant les poblacions nidificants com les que són presents a l’hivern. Per aquest motiu, cada any es fan 2 censos en cada estació. Els indicadors que es mostren en aquest estudi es desenvolupen exclusivament a partir de les dades del SOCC de primavera (ocells nidificants).
El 1994 s’inicia a Catalunya el CBMS (vegeu catalanbms. org/ca), un programa de seguiment de papallones a llarg termini basat tant en la metodologia com en l’aproximació del BMS britànic (1976), que consisteix en l’obtenció de dades detallades de la diversitat i abundància de papallones mitjançant censos visuals setmanals seguint transsectes fixes dividits en seccions que es corresponen amb els diferents hàbitats (Pollard & Yates, 1994). A Catalunya, el període de mostreig es prolonga des de març fins a setembre, ambdós mesos inclosos. Els projectes SOCC i CBMS són una font d’informació adequada per valorar si les tendències poden estar associades a la protecció dels espais naturals perquè hi ha un bon nombre d’itineraris ubicats tant dins com fora d’aquests espais. S’entén, en aquest estudi, que els espais protegits són tots aquells situats a la RMB que s’inclouen en el Pla d’Espais d’Interès Natural (PEIN). Això inclou pràcticament tota l’àrea natural protegida de la RMB, però queden fora d’aquesta catalogació espais naturals catalogats sota algun altre tipus de figura de protecció d’àmbit exclusivament local. D’altra banda, els espais situats fora de PEIN se situen normalment en ambients més o menys urbanitzats o antròpics.
Així doncs, més enllà de l’anàlisi realitzada per al conjunt de la RMB, s’han establert dos subconjunts d’itineraris per ambdós taxons; un subconjunt comprès per itineraris que estan fora dels espais PEIN i un altre amb itineraris inclosos en espais PEIN (Figura 1). En el cas dels ocells s’accepten com a PEIN els itineraris que tenen més del 60 % de la seva longitud en un PEIN o més d’un 60 % de l’àrea de l’itinerari amb un buffer de 500 metres dins de PEIN. A més, s’hi ha inclòs itine-
raris totalment fora de PEIN pels dos criteris anteriors però que se situen al costat d’espais PEIN i que segueixen bé els seus ocells; aquest seria el cas, per exemple, d’un itinerari localitzat fora de PEIN però que ressegueix un curs fluvial inclòs en un PEIN. Per altra banda, els itineraris del CBMS són, en termes generals, més curts en longitud que els del SOCC i es comptabilitzen només els exemplars vistos en un radi molt menor que el SOCC. Per tant, en el cas de les papallones no ha calgut establir criteris per destriar si els itineraris estan dins o fora de PEIN, ja que en tots els casos la inclusió o no es compleix de forma absoluta a partir de la pròpia localització de l’itinerari. Aplicant aquests criteris s’han obtingut en el cas dels ocells 122 itineraris, 61 dins de PEIN i 61 fora de PEIN, mentre que per a les papallones, s’ha tingut en compte un total de 69 itineraris, 39 en espais fora de PEIN i 30 considerats dins de PEIN (veure taules suplementàries 3 i 4).
Anàlisi de dades
Per a l’anàlisi de les tendències de sengles taxons (ocells i papallones) al llarg del temps a la RMB s’ha comparat la riquesa de les comunitats de papallones i d’ocells al principi i al final de la sèrie i s’ha elaborat un indicador global d’abundància d’aquesta regió utilitzant, les dades de seguiments estandarditzats des de 2002 fins el 2022, que és el període de mostreig comú per als dos projectes. A més s’han calculat una bateria d’indicadors que incorpora, per a cadascun dels dos taxons: 1) dos indicadors de tendència poblacional del conjunt de les espècies, un per dins i l’altre per fora dels
1: Mapa de l’àrea d’estudi. S’hi indiquen els límits de la Regió Metropolitana de Barcelona (RMB), les àrees incloses en els PEIN (terrestre) i la localització dins o fora d’aquestes àrees protegides dels itineraris SOCC i CBMS inclosos en l’estudi.
espais PEIN, i 2) quatre indicadors d’espècies forestals i de prats, també calculats de manera separada amb els itineraris de dins i fora del PEIN,que ha permès avaluar quin resultat ha donat la protecció del medi natural d’aquest territori per a ambdós taxons.
Per al càlcul dels indicadors específics d’hàbitats es té en compte les espècies veritablement especialistes forestals i de prats presents a la regió. Per a les papallones les preferències d’hàbitat es calculen anualment amb dades del CBMS, que emanen de les dades de caracterització de la vegetació del projecte. Pels ocells, seguint els estàndards recomanats per la European Bird Census Council i la selecció d’hàbitat elaborada en el segon atles d’ocells nidificants de Catalunya (Estrada et al., 2004).
Per a l’elaboració dels indicadors multi específics s’ha seguit la metodologia bàsica descrita per Soldaat et al (2017), que parteix del càlcul d’una mitjana geomètrica dels índex d’abundància regionals de les espècies, calculats per als itineraris disponibles dins d’aquesta regió. En el cas de les papallones els càlculs s’han fet utilitzant el paquet rbms (Schmucki et al., 2022), i en el cas dels ocells amb el paquet rTRIM (Bogaart et al.,2018).
Les espècies incloses tant d’ocells com de papallones en els indicadors globals i en els específics d’hàbitats, són les que apareixen en els itineraris i de les quals es tenen dades suficients com per a calcular-ne una tendència regional, d’acord amb els criteris de cadascun dels programes de seguiment (Taula 1). Tanmateix, cal tenir en compte que en els indicadors de prats no hi ha inclosos els ocells especialistes d’ambients agrícoles (degut a què a diferència de les papallones hi ha ocells «de bosc» que ocupen espais agrícoles arboris) i, per tant, s’utilitzen tan sols les espècies autènticament especialistes de prats, matollars baixos i brolles naturals. Com a conseqüència de la diferent disponibilitat d’aquest hàbitat dins i fora dels espais naturals protegits, el nombre d’espècies per a les que es disposa de dades és molt diferent en l’indicador dins de PEIN (9 spp) i fora de PEIN (només 3 spp). En papallones, per al càlcul d’aquest indicador es combinen les dades poblacionals d’espècies característiques tant de prats com d’altres hàbitats oberts, i el nombre d’espècies és el mateixa (4 ssp tant dins com fora de PEIN). Tant per als ocells com per a les papallones, les dades disponibles d’aquestes espècies especialistes són representatives de la xarxa estudiada ja que tenen un bon nombre de poblacions a la RMB.
Taula 1. Nombre d’itineraris totals emprats en l’estudi i nombre d’espècies incloses en els diferents indicadors emprats per a avaluar les tendències de les poblacions d’ocells i papallones en la Regió Metropolitana.
Origen de les sèries de dades utilitzades
Grup faunístic Indicador global Indicador segons Indicador ambients Indicador ambients protecció del territori de bosc de prat
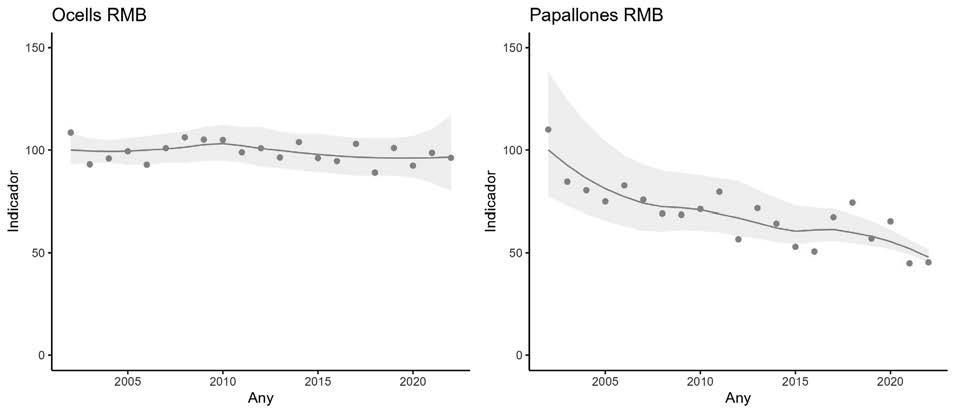
Figura 2. Indicador global d’abundància d’ocells i de papallones a la Regió Metropolitana de Barcelona. Els ocells mostren una tendència estable, mentre que les papallones mostren una regressió clara.
L’Indicador global d’abundància mostra que les poblacions d’ocells de la RMB s’han mantingut estables en els darrers 20 anys, mentre que les de papallones han patit una reducció estimada en prop d’un 50 %. No hi ha hagut cap variació significativa en aquesta tendència general en els darrers 10 anys (Figura 2).
La riquesa mitjana d’ocells (respecte totes les espècies detectades; tant les de la taula 1 de les quals es té prou dades per a calcular-ne les tendències i per tant, incloses en els indicadors, com les que no ho estan) dins les àrees incloses en el PEIN és més baixa que fora dels espais protegits (riquesa mitjana ± desviació estàndard: PEIN = 29,75 ± 8,55, no PEIN = 34,77 ± 7,98; riquesa total d’espècies presents: PEIN = 202 espècies, no PEIN = 211 espècies). Per contra, la riquesa mitjana d’espècies de papallones (respecte totes les espècies detectades; tant les de la taula 1 de les quals es té prou dades per a calcular-ne les tendències i per tant, incloses en els indicadors, com les que no ho estan) a les zones situades dins del PEIN és més alta que fora d’aquestes (PEIN = 64,75 ± 22,69, no PEIN = 47,82 ± 15,12) essent també superior el nombre total d’espècies observades als espais protegits (riquesa total d’espècies presents: PEIN = 133 espècies, no PEIN = 107 espècies). No obstant això, la diferència de riquesa mitjana entre dins i fora de PEIN en el cas dels ocells no és tan gran com en el cas de papallones.
Pel que fa a les tendències dins i fora dels espais naturals de la RMB, en general, dins els PEIN la situació és estable en el cas dels ocells i de regressió moderada en les papallones. A les àrees no incloses dins el PEIN, la situació és estable en tots dos grups biològics, per bé que les papallones han tingut oscil·lacions importants i en els darrers anys de la sèrie mostren valors molt baixos (Figura 3).
Respecte a les tendències a nivell d’hàbitats dins i fora de PEIN, primerament destaca una pitjor evolució de l’indicador de prats que del forestal per ambdós grups, tant a dins com fora PEIN (Taula 2). Els ocells i les papallones especialistes de prats estan en clara regressió a les àrees protegides PEIN, amb una pèrdua mitjana del nombre d’exemplars de prop del 60% en ocells i del 45 % en papallones, respectivament. Fora de les àrees protegides, la situació és estable pel que fa als ocells i incerta en papallones, per bé que aquestes últimes mostren valors molt baixos els darrers anys (Figura 4).
En el cas dels hàbitats forestals, els ocells especialistes de bosc mostren un increment significatiu de les seves poblacions, tant dins com fora de PEIN. En clar contrast amb aquests resultats, les papallones especialistes de bosc mostren una davallada dins de PEIN i una estabilitat fora, per bé que altra vegada, amb valors baixos els darrers anys (Figura 4).
Figura 3. Indicador Global d’Abundància d’ocells i de papallones dins i fora PEIN a la Regió Metropolitana de Barcelona. Els ocells mostren una tendència estable tant ens els espais PEIN com els de fora PEIN, mentre que les papallones mostren una regressió moderada en els espais PEIN i una tendència estable fora de PEIN.
Taula 2. Tendències observades en les poblacions d’ocells i papallones en la Regió Metropolitana de Barcelona. La taula agrupa les tendències que mostra cada indicador, fent dos enfocs concrets: un dels deu darrers anys i l’altre dels 20 darrers anys. Així doncs, hom pot observar els canvis de tendència ocorreguts al llarg d’aquest període i constatar que les tendències poblacionals tant en ocells com en papallones no ha millorat en cap cas en els darrers deu anys, ni tan sols en els espais PEIN. La columna «% de canvi 2002-2022» indica la diferència del valor de l’indicador entre el final i l’inici de la sèrie (és a dir, el percentatge en el canvi d’abundància), mostrant si han experimentat un increment (positiu) o bé una regressió (negatiu).
prats
bosc
Figura 4. En l’indicador específic d’espècies d’ambients de bosc els ocells mostren una increment moderat tant ens els espais PEIN com els de fora PEIN, mentre que les papallones mostren una regressió moderada dins de PEIN però estabilitat fora dels PEIN; en l’indicador específic d’espècies d’ambients de prats els ocells mostren una regressió moderada en els espais PEIN i una tendència estable en els de fora PEIN, mentre que les papallones mostren una regressió moderada en els espais PEIN però una tendència incerta en els de fora de PEIN.
Discussió
Patrons generals
Les poblacions de papallones i ocells mostren, a la RMB, una dinàmica poblacional similar a les seves respectives tendències en tot el conjunt català, essent estable en el cas dels ocells (Franch et al., 2021) i regressiva en les papallones (Melero et al., 2016; Colom et al., 2019).
L’elevat nombre d’espècies utilitzades en ocells per aquest indicador (72 spp) genera un estret interval de confiança i accentua la validesa dels resultats. D’altra banda, aquesta estima es pot considerar robusta i ben representativa del conjunt de les comunitats de papallones de la regió, ja que es basa en les dades d’un nombre elevat d’espècies (59), monitoritzades
Butlletí ICHN
(2), 2024
a partir d’un nombre important de poblacions i plenament representatiu de les comunitats de papallones de la RMB.
En el cas dels ocells cal contextualitzar els resultats amb escenaris similars però d’escales geogràfiques més àmplies. En concret a escala europea, les darreres actualitzacions disponibles mostren una davallada general de les poblacions, estimada i sostinguda en el temps, en un 19 % per al període 1980-2020 (168 espècies incloses; PECBMS, 2022). Per tant, es pot considerar que l’estabilitat observada al conjunt de la RMB indica una situació menys negativa que la del conjunt d’Europa. Aquest fet podria estar associat a un menor impacte local dels dos grans factors que estan fent disminuir les poblacions d’ocells a Europa, la intensificació agrícola i, en menor mesura, el canvi climàtic (Rigal et al., 2023). A dife-
rència del que s’observa en papallones, els efectes del canvi climàtic en molts ocells semblen, per ara, poc clars dins l’àrea d’estudi (Herrando et al., 2019).
Fins i tot, aquest resultat sembla una mica millor que el que ja s’ha publicat per al conjunt de Catalunya, que presenta una lleu disminució en les poblacions totals d’ocells (ICO, 2023). Malgrat tot, això pot emmascarar que en un entorn tan humanitzat com la RMB els descensos de les poblacions s’haguessin produït amb anterioritat a la implementació dels sistemes de seguiment d’ocells i que s’observi una imatge ja empobrida des de l’inici de la sèrie temporal. Tampoc es pot descartar que les tendències de les espècies d’ocells siguin en conjunt més negatives fora de la RMB perquè hi hagi factors més negatius afectant espècies més especialistes poc o gens representades dins la RMB per manca dels seus hàbitats preferents (espècies d’aiguamoll, alpines o estèpiques, entre d’altres).
Pel que fa a les papallones, es troben paraŀlelismes molt notables amb el que s’ha documentat en altres països occidentals. Per exemple, les poblacions de papallones també estan minvant fortament al centre i nord d’Europa (Warren et al., 2021) i en àmplies zones de Nord Amèrica (Forister et al., 2021). De fet, sembla un problema molt estès arreu del planeta que no només afecta les papallones sinó a molts altres grups d’insectes, inclosos els principals grups de pollinitzadors (Wagner, 2020; Goulson, 2021). La magnitud d’aquest declivi no difereix gaire del que s’ha calculat amb les dades del CBMS pel conjunt de Catalunya des de 1994, que situa la disminució de les poblacions de papallones en un 44%. Encara que les dades no són estrictament comparables perquè inclouen períodes i mostres diferents, en conjunt aquests càlculs evidencien una situació preocupant del grup de les papallones.
La protecció dels espais de la RMB: PEIN i fora PEIN
Respecte el rol de les àrees protegides en la conservació de la biodiversitat (en el cas d’aquest estudi, les zones del PEIN), els resultats d’aquest treball a la RMB són bastant concordants amb els desenvolupats a la Xarxa Natura 2000 al conjunt d’Europa (Pellissier et al., 2020), en el sentit que els ocells sembla que han estat més afavorits per la protecció dels espais naturals que les papallones. En conjunt, les poblacions d’ocells mostren una estabilitat numèrica, tant en zones protegides com no protegides, que es tradueix en un patró general estable dins espais naturals protegits. Malgrat tot, aquest fet amaga una dinàmica molt diferent per a espècies d’ocells pròpies d’ambients oberts, que estan en clara regressió, i espècies forestals, que estan en increment clar, tal com passa al conjunt de Catalunya (Herrando et al., 2016). Els resultats mostren que els espais naturals protegits (PEIN) allotgen en conjunt, un nombre similar d’espècies d’ocells (70) que els espais que no ho són (59) (Taula 1), dins el conjunt d’espècies incloses en els indicadors de les quals es té prou dades per a calcular-ne les tendències. Alguns ocells només presents als PEIN són, per exemple, espècies de bosc madur com Dryocopus martius o de prats com Anthus campestris
Els resultats per als ocells indiquen que fora dels espais naturals protegits hi ha una major diversitat mitjana a escala local (d’itinerari), cosa que contrasta amb els patrons de nombre total d’espècies dins i fora PEIN que s’ha comentat abans. Això segurament es relaciona amb uns patrons de diversitat ambiental que són dependents de l’escala. A escala local les zones situades fora de PEIN tenen un fort grau de fragmentació; es tracta de paisatges humanitzats, molt diversos paisatgísticament a aquesta escala. Aquesta diversitat ambiental queda reflectida en els transsectes de 3 km dels transsectes SOCC. En altres paraules, els espais naturals protegits tenen taques de majors dimensions de paisatges homogenis i allotgen un conjunt més gran d’espècies format en part per ocells especialistes d’hàbitats no presents fora dels espais naturals protegits, mentre que fora dels PEIN hi predominen espècies generalistes amb comunitats localment més riques a causa de la combinació (en àrees de mida reduïda) d’ambients urbans i periurbans, a més de retalls de bosc, camps de conreu i rius. Pel que fa als resultats en el grup de les papallones, la davallada de l’indicador d’abundància dins dels espais PEIN i l’estabilitat fora d’aquests espais coincideix amb un patró similar pel que fa a la riquesa mitjana per itinerari, que ha disminuït significativament en les estacions PEIN en els darrers 20 anys però s’ha mantingut sense canvis a fora (Figura suplementària 1). Tot i això, la riquesa d’espècies continua sent més elevada dins de les estacions PEIN que fora d’elles. Aquests fets combinats suggereixen que la pèrdua d’espècies ja s’havia produït amb anterioritat fora dels espais PEIN a causa de l’alteració i destrucció dels hàbitats relacionades amb la urbanització del paisatge. En tot cas, aquests resultats indiquen que la protecció del territori no ha estat suficient per revertir la pèrdua de biodiversitat en aquest grup, precisament a les zones que allotgen encara més diversitat d’espècies, malgrat que alguns d’aquests espais protegits de la regió metropolitana ja ho estaven a finals de segle XX (p. ex. Sant Llorenç del Munt i Serra de l’Obac).
Aquesta forta davallada que s’ha constatat dins de les àrees protegides té igualment un paral·lelisme notable amb el que Hallmann et al. (2017) van trobar a Alemanya per al conjunt d’insectes voladors. Aquests autors van documentar un descens superior al 75% de la biomassa en 27 anys (entre 1989 i 2016) en un conjunt de 63 espais naturals protegits. Els resultats són per tant, concloents i apunten a una pèrdua general de la biodiversitat de papallones a la RMB, més forta als espais protegits que als no protegits, posant clarament en dubte la idoneïtat de la gestió associada als espais presents a la xarxa PEIN en aquesta regió pel que fa als ropalòcers i possiblement, la majoria de la resta dels insectes pol·linitzadors (Bosch et al., 2022). Tot fa indicar que aquestes comunitats estan sotmeses a un procés de degradació en gran part originat per la pèrdua dels espais oberts (vegeu més avall) i cada cop més, per la incidència negativa de les sequeres. L’erosió de la diversitat de papallones és més avançada a les zones no protegides, que ja han perdut un cert nombre d’espècies especialistes que encara mantenen poblacions a les zones protegides. La major dominància de les espècies generalistes que en resulta, converteix aquestes comunitats en més resilients.
MAURI
Tendència en funció del tipus d’hàbitats
A trets generals, les poblacions d’ocells i papallones mostren una tendència negativa en els ambients de prats; els indicadors d’espècies especialistes d’aquests hàbitats, sobretot dins dels PEIN, indiquen una clara regressió. En bona mesura aquest fet es pot atribuir a l’abandonament de les activitats agro-ramaderes a mitja muntanya que ha produït un tancament constant de la vegetació (Herrando et al., 2016). En canvi, en els hàbitats forestals les tendències són divergents: els ocells experimenten clarament un increment i per contra les papallones pateixen una regressió en els espais forestals PEIN i una tendència estable fora dels PEIN.
En el cas dels indicadors d’ocells forestals i de prats (o d’ambients oberts) de la RMB, segueixen la mateixa evolució que la tendència regional catalana pels mateixos ambients (ICO, 2023). No obstant això, els ocells d’ambients oberts fora de PEIN es mantenen estables malgrat que ho fan amb un interval de confiança considerable, la qual cosa posa en dubte la robustesa d’aquesta estabilitat i això pot estar relacionat amb l’escàs nombre de les espècies representatives que hi queden, ja que la majoria haurien desaparegut prèviament. Malgrat no hi ha cap anàlisi exhaustiva, això és el que podria suggerir la regressió observada en la distribució d’algunes espècies d’ocells d’espais oberts a la RMB entre el primer i el segon atles d’ocells nidificants de Catalunya (Estrada et al., 2004), que cobreix les dues darreres dècades del segle XX, previ a l’inici del projecte SOCC. De fet, la regressió va començar abans dels anys 80 del segle passat, com suggereixen les regressions documentades al Penedès i àrees properes per a unes quantes espècies (Muntaner et al., 1983), actualment molt escasses o completament desaparegudes.
Els indicadors d’ocells que aquí es mostren per a la RMB semblen ser capaços de detectar variacions en part de l’àrea d’estudi, com posen de manifest un parell d’exemples. L’itinerari 165 del SOCC al Parc Natural del Garraf és el paradigma de la pèrdua de les poblacions d’ocells de prats dins dels espais naturals protegits de la RMB. Aquesta àrea va patir grans incendis els anys 1986 i 1994 i en poc temps es va convertir en una zona important per a espècies d’espais oberts (Herrando et al., 2002). Posteriorment, però, els intents d’establir ramaderia extensiva a la zona han estat poc reeixits i els hàbitats s’han tancat ràpidament. Actualment, la vegetació és massa desenvolupada per a la gran majoria de les espècies de prats, amb l’excepció de Lanius meridionalis, que encara manté una població a la zona. Un altre cas és la interrupció de la regressió de l’indicador d’ocells d’espais oberts dins de PEIN entre 2007 i 2015, que podem atribuir a l’efecte positiu dels incendis per a aquest grup d’espècies, principalment en el Parc Natural de Sant Llorenç del Munt i l’Obac, on es va recuperar hàbitat obert de forma temporal en aquest període. Però, com va passar al Garraf, quan les zones cremades tornen a recuperar el bosc, les espècies d’espais oberts tornen a disminuir.
Fora de les àrees protegides, les espècies d’ocells forestals també augmenten, per la qual cosa la protecció no sembla ser el factor que expliqui l’increment d’aquestes espècies, almenys en termes generals. Cal tenir present que l’augment
de la cobertura arbrada i la seva maduració és l’element clau que explica l’increment de les espècies d’ocells forestals, i que això s’ha produït també fora dels espais naturals protegits (Brotons et al., 2020).
Un apunt final és que, almenys en part, els indicadors globals estables d’ocells estan influenciats tant per la situació més aviat positiva dels ocells forestals, que presenten un increment moderat a curt termini, com per les espècies d’espais oberts, que tenen una situació clarament negativa. L’estabilitat del conjunt pot amagar processos ecològics importants i és per aquest motiu que és important disposar d’aquesta mena d’indicadors d’espècies especialistes d’hàbitats.
En el cas de les papallones, les davallades aparentment no estan tan lligades a l’hàbitat perquè afecten tant les zones de prats com les forestals. Tot i així, creiem que el tancament de la vegetació és en darrer terme el major responsable de la forta davallada de les comunitats de papallones, sobretot als espais PEIN. Es podria destacar el cas de l’itinerari CBMS de La Conreria, al Parc Natural de la Serra de Marina, com a paradigma del que està passant en alguns dels espais protegits en relació a les papallones. Aquest itinerari, amb una sèrie de 21 anys, és un dels que ha patit transformacions més profundes i negatives de la comunitat de papallones de tota la xarxa del CBMS. Això queda reflectit molt ràpidament per les disminucions molt significatives tant de la riquesa com de l’abundància. Els indicadors de comunitat, a més, informen dels motius d’aquest empobriment, ja que denoten un problema evident de tancament amb efectes molt negatius sobre un gran nombre de papallones lligades als ambients oberts. Aquestes davallades, a més, han afectat desproporcionadament les papallones de caràcter especialista respecte a les generalistes. El seguiment de la vegetació que es fa en aquest itinerari com a part del protocol del CBMS confirma aquest tancament, ja que en els darrers 20 anys els matollars alts i l’alzinar han passat d’una cobertura del 63% a una del 84%, mentre que els matollars baixos (que inclouen timonedes i altres comunitats vegetals molt interessants per a les papallones) han vist reduïda la cobertura d’un 30% a un 7%. La situació crítica d’aquest itinerari queda palesa pel fet que de les 29 espècies amb poblacions actuals ben establertes, 14 (48%) es troben en declivi i cap en augment. A més, fins a 7 espècies han patit extinció local: al 2023 s’han afegit 4 més (Anthocharis cardamines, Melitaea deione, Melanargia lachesis i Thymelicus acteon) a les que ja figuraven com extintes (Leptidea sinapis, Argynnis paphia, Polyommatus icarus). El cas de Melanargia lachesis és molt il·lustratiu atès que justament és una espècie que forma part de l’indicador de prats per a papallones. Aquesta situació tan preocupant segurament es veu agreujada per l’especial afectació de la zona del litoral barceloní pel que fa a la sequera extrema dels darrers 3 anys. Aquest exemple s’emmarca en el fenomen del tancament de la vegetació i de l’augment de la massa forestal, molt estès a Catalunya a partir de mitjan del segle passat i amb repercussions molt negatives sobre un grup molt associat als espais oberts com són les papallones (Ubach et al., 2020). Aquesta estreta associació afecta fins i tot les papallones que habiten els boscos i que han estat utilitzades en el càlcul de l’indiJ.
cador forestal. Malgrat que moltes d’elles utilitzen arbres i arbusts típics dels nostres boscos per a desenvolupar-se com a larves, els adults igualment requereixen d’espais mínimament oberts i d’ecotons. En efecte, aquests són necessaris ja sigui per trobar recursos alimentaris com ara fonts de nèctar, com per a l’establiment de territoris en el cas dels mascles, o fins i tot per a regular-se a nivell tèrmic. Per tant, paradoxalment, l’augment de la cobertura forestal als espais PEIN en les darreres dècades ha repercutit negativament sobre aquestes espècies, perquè ha suposat una homogeneïtzació del paisatge i la pèrdua d’una configuració en mosaic que també necessiten les papallones forestals. No sabem si l’estabilitat que mostren aquestes espècies en els darrers 10 anys (Taula 2) es podria relacionar amb un lleuger augment de la maduresa dels boscos als espais PEIN (dades dels inventaris forestals nacionals, D. Carrera com. pers.), que hauria millorat la seva estructura de forma beneficiosa per les papallones forestals en crear més clarianes, o bé al fet que les poblacions s’hagin estabilitzat al voltant d’uns valors crònicament baixos. Els indicadors d’hàbitat mostren una situació lleugerament millor fora que dins dels espais PEIN en el cas de les papallones, per bé que els intervals de confiança són molt amplis i posen un grau d’incertesa a les tendències. Aquest fet molt possiblement es relaciona amb una major variabilitat del paisatge en les zones no protegides, que són sotmeses a pertorbacions més grans que als espais PEIN. En paral·lel, això es tradueix en respostes poblacionals més fluctuants de les papallones. A més, la incertesa en les tendències dels indicadors s’ha vist multiplicada per uns valors inusualment baixos els dos darrers anys (Fig. 4), relacionats amb l’impacte addicional negatiu que ha suposat la situació d’extrema sequera.
Conclusions
En els darrers 20 anys, les poblacions d’ocells a la RMB ha mantingut una tendència estable en quant a l’abundància a excepció dels ocells d’espais oberts naturals, mentre que les papallones han sofert una regressió significativa, especialment en els espais protegits. Les espècies pròpies de prats són les que han sofert una davallada més acusada, i es constata la pèrdua d’espècies especialistes per contra de les més generalistes, fet que segurament evidencia un deteriorament ecològic dels hàbitats.
Hi ha evidències que apunten a què el desenvolupament dels espais PEIN com a figura de conservació de biodiversitat no ha servit per evitar la caiguda de les poblacions de papallones. En el cas dels ocells tampoc es veu una millora dins PEIN. Aquesta dinàmica poblacional és similar a la que es produeix globalment a Catalunya i coincideix amb la protecció i desenvolupament de masses forestals per contra de prats i espais oberts. Cal remarcar, però, el paper del PEIN en el manteniment de la biodiversitat ja que moltes espècies només es troben actualment dins la xarxa d’espais protegits.
Tant ocells com papallones són grups considerats com a excel·lents bioindicadors dels canvis en els hàbitats. L’evolució dels hàbitats de la RMB o la gestió dels mateixos, té una relació directa amb les tendències poblacionals descrites
i una gestió acurada dels espais oberts de la RMB podria contribuir a una recuperació de les espècies d’espais oberts, tant d’ocells com de papallones. Cal tenir en compte que bona part d’aquestes espècies tenen una distribució marcadament mediterrània (Keller et al., 2020; Kudrna et al., 2011) i, per tant, la nostra responsabilitat en la seva conservació es pot considerar major que sobre la que tenim en altres espècies que trobem a casa nostra en els límits de la seva distribució.
Agraïm sincerament a totes les persones que voluntàriament han aportat dades durant el període d’estudi (20222022) als programes de Seguiment d’Ocells Comuns de Catalunya i al Catalan Butterfly Monitoring Scheme. També volem expressar la nostra gratitud a l’Institut Català d’Ornitologia i al Museu de Granollers per la cessió de les dades i la seva inestimable col·laboració. Finalment, agraïm al Centre de Recerca Ecològica i Aplicacions Forestals per la seva col·laboració en aquest projecte. Tota aquesta contribució ha estat essencial per a la realització d’aquesta recerca.
Bibliografia
Almond, R.E.A., Grooten M. & Petersen, T. (Eds). 2020. Informe Planeta Vivo 2020: Revertir la curva de la pérdida de biodiversidad. Resumen. WWF, Gland, Suiza.
Barnosky AD, Matzke N, Tomiya S, WoganGOU, Swartz B, Quental TB, MarshallC, McGuire JL, Lindsey EL, Maguire KCet al. 2011. Has the Earth’s sixth mass extinction already arrived? Nature 471:51-57.
Boada, M., Maneja, R., Marlès, J. 2019. Renaturalització de la ciutat; Biodiversitat-sostenibiilitat urbana. Diputació de Barcelona.
Bogaart, P., van der Loo, M. & Pannekoek, J. 2018. rtrim: Trends and Indices for Monitoring Data. R package version 2.0.6. Disponible a: https://CRAN.R-project.org/package=rtrim [1 abril 2024].
Bosch J, Stefanescu C, Roquer-Beni L et al. 2022. Els pol·linitzadors silvestres a Catalunya; Informe sobre estatus, amenaces i àmbits prioritaris d’actuació per a la seva conservació. Barcelona. Generalitat de Catalunya. Departament d’Acció Climàtica, Alimentació i Agenda Rural.
Brotons, L., Pou, N., Herrando, S., Bota, G., Villero, D., Garrabou, J., Ordóñez, J. L., Anton, M., Gual, G., Recoder, L., Alcaraz, J., Pla, M., Sainz de la Maza, P., Pont, S. i Pino, J. 2020. Estat de la Natura a Catalunya 2020. Departament de Territori i Sostenibilitat. Generalitat de Catalunya. Barcelona.
Colom, P., Carreras, D., Stefanescu, C. 2019. Long-term monitoring of Menorcan butterfly populations reveals widespread insular biogeographical patterns and negative trends. Biodiversity and Conservation, 28: 1837-1851.
Departamento de Asuntos Económicos y Sociales, Nacions Unides, 2014. World Urbanization Prospects 2014. Disponible a: https:// www.un.org/es/desa/world-urbanization-prospects-2014 [01 abril 2024].
Estrada, J., Pedrocchi, V., Brotons, L. & Herrando, S. (eds.). 2004. Atles dels ocells nidificants de Catalunya 1999-2002. Institut Català d’Ornitologia (ICO)/Lynx Edicions. Barcelona. 638 p.
Forister M. L., Halsch C. A., Nice C. C., Fordyce J. A., Dilts T. E., Oliver J. C., Prudic, K. L., Shapiro, A. M., Wilson, J. K., Glassberg, J. 2021. Fewer butterflies seen by community scientists across the warming and drying landscapes of the American West Science, 371: 1042-1045.
Franch, M., Herrando, S., Anton, M., Villero, D. & Brotons, L. 2021. Atles dels ocells nidificants de Catalunya: Distribució i abundància 20152018 i canvi des de 1980. Institut Català d’Ornitologia / Cossetània Edicions. Barcelona. 640 p. Furness, R.W. and Greenwood, J.J.D. 1993. Birds as Monitors of Environmental Change. Chapman & Hall, London. 356 p. Gaston, K. J., & Fuller, R. A. 2008. Commonness, population depletion and conservation biology. Trends in Ecology and Evolution, 23(1), 14–19.
Goulson D. 2021. Silent Earth: Averting the Insect Apocalypse Random House. New York. 336 p. Gregory, R., & van Strien, A. 2020. Wild bird indicators: using composite population trends of birds as measures of environmental health. Ornithological Science, 9, 3–22.
Hallmann CA, Sorg M, Jongejans E, Siepel H, Hofland N, Schwan H, et al. 2017. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 12 (10): e0185809.
Herrando, S., Brotons, L., Anton, M., Páramo, F., Villero, D., Titeux, N., Quesada, J., & Stefanescu, C., 2016. Assessing impacts of land abandonment on Mediterranean biodiversity using indicators based on bird and butterfly monitoring data. Environmental Conservation, 43 (1): 69-78.
Herrando, S., & Brotons, L. 2002. Forest bird diversity in Mediterranean areas affected by wildfires: A multi-scale approach. Ecography, 25: 161-172.
Herrando, S., Titeux, N., Brotons, L. et al. 2019. Contrasting impacts of precipitation on Mediterranean birds and butterflies Scientific Reports 9, 5680.
Institut Català d’ornitologia. 2023. Disponible a: https://socc.cat/ socc/resultats/tendencies/ [1 abril 2024].
Juvillà, E. Et al. 2019. Renaturalització de la Ciutat. Diputació de Barcelona.
Keller, V., Herrando, S., Voříšek, P., Franch, M., Kipson, M., Milanesi, P., Martí, D., Anton, M., Klvaňová, A., Kalyakin, M.V., Bauer, H.-G. & Foppen, R.P.B. 2020. European Breeding Bird Atlas 2: Distribution, Abundance and Change. European Bird Census Council & Lynx Edicions, Barcelona. 967 p.
Kudrna O., Harpke A., Lux K., Pennerstorfer J., Schweiger O., Settele J. & Wiemers M. 2011. Distribution Atlas of Butterflies in Europe. Geselschaft fur Schmetterlingsschutz. Halle, 576 p. Llobet, J., coordinador. Equip d’investigadors de l’Institut d’Estudis Regionals i Metropolitans de Barcelona (IERMB). 2020. La metròpoli en 100 indicadors. L’AMB EN XIFRES 2019. Institut d’Estudis Regionals i Metropolitans de Barcelona.
Melero, Y., Stefanescu, C., Joan Pino, J. 2016. General declines in Mediterranean butterflies over the last two decades are modulated by species traits. Biological Conservation 201: 336-342. Muntaner, Jordi; Ferrer, Xavier; Martínez i Vilalta, Albert. 1983. Atlas dels ocells nidificants de Catalunya i Andorra. Ketres. Barcelona, p. 322.
Pan-European Common Bird Monitoring Scheme(PECBMS). Czech Society for Ornithology, 2022. Disponible a: https:// pecbms.info/ [1 abril 2024].
Pellissier, V., Schmucki, R., Pe’er, G., Aunins, A., Brereton, T.M., Brotons, L., Carnicer, J., Chodkiewicz, T., Chylarecki, P., del Moral, J.C., Escandell, V., Evans, D., Foppen, R., Harpke, A., Heliölä, J., Herrando, S., Kuussaari, M., Kühn, E., Lehikoinen, A., Lindström, Å., Moshøj, C.M., Musche, M., Noble, D., Oliver, T.H., Reif, J., Richard, D., Roy, D.B., Schweiger, O., Settele, J.,
Stefanescu, C., Teufelbauer, N., Touroult, J., Trautmann, S., van Strien, A.J., van Swaay, C.A.M., van Turnhout, C., Vermouzek, Z., Voříšek, P., Jiguet, F. and Julliard, R. 2020. Effects of Natura 2000 on nontarget bird and butterfly species based on citizen science data. Conservation Biology, 34: 666-676.
Pollard, T.J. Yates. 1994. Monitoring Butterflies for Ecology and Conservation. Springer Science & Business Media. London, 274 p. Prudic K. L., Shapiro A.M., Wilson J.K., Glassberg, J., 2021. Fewer butterflies seen by community scientists across the warming and drying landscapes of the American West. Science, 371 (6533): 1042-1045.
Rigal, S., Dakos, V., Alonso, H., Auniņš, A., Benkő, Z., Brotons, L., Chodkiewicz, T., Chylarecki, P., de Carli, E., del Moral, J. C., Domșa, C., Escandell, V., Fontaine, B., Foppen, R., Gregory, R., Harris, S., Herrando, S., Husby, M., Ieronymidou, C., Devictor, V., 2023. Farmland practices are driving bird population decline across Europe. Proceedings of the National Academy of Sciences of the United States of America, 120 (21): e2216573120.
Schmucki R., Harrower C.A., Dennis E.B. 2022. rbms: Computing generalised abundance indices for butterfly monitoring count data. R package version 1.1.3. Disponible a: https://github.com/ RetoSchmucki/rbms [1 abril 2024].
Soldaat L.L., Verweij R., Van Turnhout C.A.M., Visser H., Van Strien A.J… 2017. A Monte Carlo Method to Account for Sampling Error in Multispecies Indicators. Ecological Indicators, 81: 340-347.
Stefanescu C., Torre, I., Jubany, J., Páramo, F., 2011. Recent trends in butterfly populations from north-east Spain and Andorra in the light of habitat and climate change. Journal of Insect Conservation, 15: 83-93.
Thomas, J.A., 2005. Monitoring change in the abundance and distribution of insects using butterflies and other indicator groups. Phil. Trans. R. Soc. B, 360: 339–357.
Ubach, A., Páramo, F., Gutiérrez, C., Stefanescu, C, 2020. Vegetation encroachment drives changes in the composition of butterfly assemblages and species loss in Mediterranean ecosystems Insect Conservation and Diversity, 13 (2): 151-161.
Van Swaay, C. A M., Dennis, E.B., Schmucki, R., Sevilleja, C.G., Åström, S., Balalaikins, M., Barea-Azcón, J.M. , Bonelli, S., Botham, M., Cancela, J.P., Collins, S., De Flores, M., Dapporto, L., Dopagne, C., Dziekanska, I., Escobés, R., Faltynek Fric, Z., Fernández-García, J.M., Fontaine, B., Glogovčan, P., Gracianteparaluceta, A., Harpke, A., Harrower, C., Heliölä, J., Houard, X., Judge, M., Kolev, Z., Komac, B., Kühn, E., Kuussaari, M., Lang, A., Lysaght, L., Maes, D., McGowan,D., Mestdagh, X., Middlebrook, I., Monasterio, Y., Monteiro, E., Munguira, M.L., Musche, M., Olivares, F.J., Õunap, E., Ozden, O., Pavlíčko, A., Pendl, M., Pettersson, L.B., Rákosy, L., Roth, T., Rüdisser, J., Šašić, M., Scalercio, S., Settele, J., , Sielezniew, M., Sobczyk-Moran, G., Stefanescu, C., Švitra, G., Szabadfalvi, A., Tiitsaar, A., Titeux, N., Tzirkalli, E., Ubach, A., Verovnik, R., Vray, S., Warren, M.S., Wynhoff, I., & Roy, D.B. 2022. European Grassland Butterfly Indicator 1990-2020 Technical report. Butterfly Conservation Europe & ABLE/eBMS (www.butterflymonitoring.net) & Vlinderstichting report VS2022.039 Disponible a: https://butterflymonitoring.net/sites/default/files/Pdf/Indicators/ VS2022.039%20European_Grassland_Butterfly_Indicator_19902020_v3.pdf [1 abril 2024].
Wagner DL. 2020. Insect declines in the Anthropocene. Annu. Rev. Entomol. 65, 457–480.
Warren M., Maes D., Chris Van Swaayc, C., Goffartd P., Van Dycke H., Bournf N., Wynhoff I., Hoaref D., and Ellis S. 2021. The decline of butterflies in Europe: Problems,significance, and possible solutions. Proceedings of the National Academy of Sciences of the United States of America 118 (2): e2002551117.
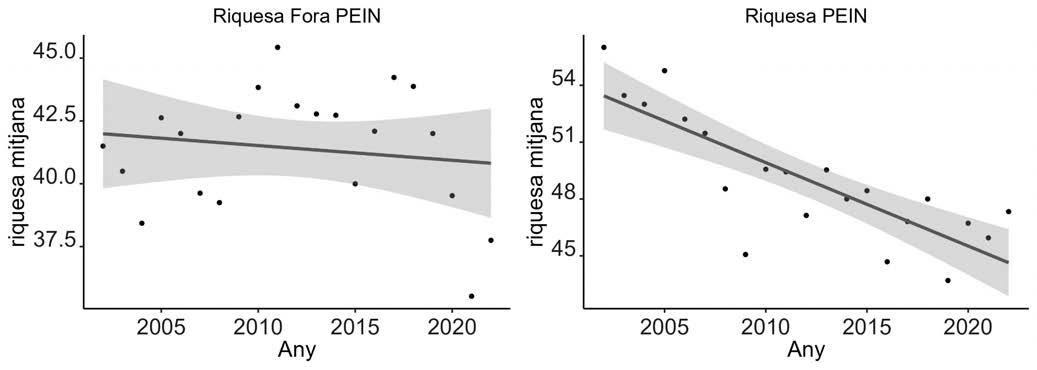
Figura complementària 1. Evolució de la riquesa mitjana de papallones: en els darrers 20 anys, les papallones mostren una tendència negativa significativa en les estacions PEIN, fet que no ha succeït fora de PEIN. Malgrat aquest descens, la riquesa d’espècies continua sent clarament més elevada dins de PEIN que fora PEIN.
Taula complementària 1. llista d’espècies d’ocells presents a la RMB tingudes en compte per a l’estudi. Taula 1.1 espècies presents dins de PEIN i Taula 1.2 espècies presents fora de PEIN.
Nom científic Nom comú català Present Present en PEIN fora PEIN
Accipiter gentilis Astor x x
Accipiter nisus Esparver comú x x
Acrocephalus arundinaceus Balquer x x
Acrocephalus melanopogon Boscarla mostatxuda x x
Acrocephalus schoenobaenus Boscarla dels joncs x x
Acrocephalus scirpaceus Boscarla de canyar x x
Actitis hypoleucos Xivitona x x
Aegithalos caudatus Mallerenga cuallarga x x
Agapornis roseicollis Agapornis de coll rosa x
Aix galericulata
Ànec manderí x
Alauda arvensis Alosa comuna x x
Alcedo atthis Blauet x x
Alectoris rufa Perdiu roja x x
Amandava amandava Múnia roig x
Amazona ochrocephala Amazona frontgroga x
Anas crecca Xarxet comú x x
Anas platyrhynchos Ànec collverd x x
Anser anser Oca comuna x x
Anthus campestris Trobat x x
Anthus cervinus Piula gola-roja x
Anthus pratensis Titella x x
Anthus spinoletta Grasset de muntanya x x
Anthus trivialis Piula dels arbres x x
Apus apus Falciot negre x x
Apus pallidus Falciot pàl·lid x x
Aquila chrysaetos Àguila daurada x x
Aquila fasciata Àguila cuabarrada x x
Aratinga nenday Aratinga nandai x
Ardea alba Agró blanc x x
Ardea cinerea Bernat pescaire x x
Ardea purpurea Agró roig x x
Ardeola ralloides Martinet ros x x
Athene noctua Mussol comú x x
Aythya ferina Morell cap-roig x x
Aythya nyroca Morell xocolater x
Bubo bubo Duc x
Bubulcus ibis Esplugabous x x
Buteo buteo Aligot comú x x
Cairina moschata f. domestica Ànec mut x x
Calandrella brachydactyla Terrerola comuna x x
Calidris alba Territ tresdits x
Calidris ferruginea Territ becllarg x Calidris minuta Territ menut x Calidris pugnax Batallaire x Calidris temminckii Territ de temminck x
Caprimulgus europaeus Enganyapastors x x Carduelis carduelis Cadernera x x
Cecropis daurica Oreneta cua-rogenca x x Certhia brachydactyla Raspinell comú x x Cettia cetti Rossinyol bord x x
Charadrius dubius Corriol petit x x
Charadrius hiaticula Corriol gros x x
Chlidonias hybrida Fumarell carablanc x x Chloris chloris Verdum x x Ciconia ciconia Cigonya blanca x x Circaetus gallicus Àguila marcenca x x
Circus aeruginosus Arpella comuna x x
Circus cyaneus Arpella pàl·lida x x
Circus pygargus Esparver cendrós x x
Cisticola juncidis Trist x x
Clamator glandarius Cucut reial x x
Coccothraustes coccothraustes Durbec x x Columba livia Colom roquer x x Columba oenas Xixella x x
Columba palumbus Tudó x x
Coracias garrulus Gaig blau x x
Corvus corax Corb x x
Corvus corone
Cornella negre x x
Corvus monedula Gralla x x
Coturnix coturnix
Guatlla x x
Cuculus canorus Cucut x x
Curruca communis Tallareta comuna x x
Curruca conspicillata
Tallarol trencamates x Curruca hortensis
Curruca iberiae
Curruca melanocephala
Curruca undata
Cyanistes caeruleus
Cyanoliseus patagonus
Tallarol emmascarat x x
Tallarol de garriga x x
Tallarol capnegre x x
Tallareta cuallarga x x
Mallerenga blava x x
Cotorra de Patagònia x Delichon urbicum
Oreneta cuablanca x x
Dendrocopos major Picot garser gros x x
Dryobates minor Picot garser petit x x
Dryocopus martius Picot negre x
Egretta garzetta Martinet blanc x x
Emberiza calandra Cruixidell x x
Emberiza cia Sit negre x x
Emberiza cirlus
Emberiza citrinella
Gratapalles x x
Gafarró comú x x
Emberiza hortulana Hortolà x x
GEA, FLORA ET FAUNA
Erithacus rubecula Pit-roig
Estrilda astrild Bec de corall senegalès
Estrilda troglodytes Bec de corall cuanegre
Euplectes afer Bisbe groc
Falco columbarius Esmerla
Falco eleonorae Falcó de la reina
Falco peregrinus Falcó pelegrí
Falco subbuteo Falcó mostatxut
Falco tinnunculus Xoriguer comú
Falco vespertinus Falcó cama-roig
Ficedula hypoleuca Mastegatatxes
Fringilla coelebs Pinsà comú
Fringilla montifringilla Pinsà mec
Fulica atra Fotja comuna
Fulica cristata Fotja banyuda
Galerida cristata Cogullada vulgar
Galerida theklae Cogullada fosca
Gallinago gallinago Becadell comú
Gallinula chloropus Polla d’aigua
Garrulus glandarius Gaig
Gelochelidon nilotica Curroc
Grus grus Grua
Gyps fulvus Voltor comú
Haematopus ostralegus Garsa de mar
Hieraaetus pennatus Àguila calçada
Himantopus himantopus Cames llargues
Hippolais polyglotta Bosqueta vulgar
Hirundo rustica Oreneta vulgar
Ixobrychus minutus Martinet menut
Jynx torquilla Colltort
Lanius collurio Escorxador
Lanius meridionalis Botxí meridional
Lanius senator Capsigrany
Larus argentatus Gavià argentat de potes
Larus audouinii Gavina corsa
Larus fuscus Gavià fosc
Larus melanocephalus Gavina capnegre
Larus michahellis Gavià argentat de potes grogues
Larus ridibundus Gavina vulgar
Leiothrix lutea Rossinyol del Japó
Linaria cannabina Passerell comú
Locustella luscinioides Boscaler comú
Locustella naevia Boscaler pintat gros x
Lophophanes cristatus Mallerenga emplomallada
Loxia curvirostra Trencapinyes x
Lullula arborea Cotoliu
Luscinia megarhynchos Rossinyol x
Mareca strepera Ànec griset x
Merops apiaster Abellerol
Milvus migrans Milà negre
Milvus milvus Milà reial
Monticola saxatilis Merla roquera x
Monticola solitarius Merla blava
Morus bassanus Mascarell x
Motacilla alba Cuereta blanca x x
Motacilla cinerea Cuereta torrentera x x
Motacilla flava Cuereta groga x x
Muscicapa striata Papamosques gris x x
Myiopsitta monachus Cotorreta pitgrisa
Neophron percnopterus Aufrany
Netta rufina Xibec
Numenius arquata Becut
Nycticorax nycticorax Martinet de nit
Nymphicus hollandicus Cacatua nimfa
Oenanthe hispanica Còlit ros
Oenanthe leucura Còlit negre
Oenanthe oenanthe Còlit gris
Oriolus oriolus Oriol
Otus scops Xot
Pandion haliaetus Àguila pescadora x x
Parus major Mallerenga carbonera
Passer domesticus Pardal comú x x
Passer montanus Pardal xarrec x x
Pavo cristatus Paó blau x
Periparus ater Mallerenga petita x x
Pernis apivorus Aligot vesper x x
Petronia petronia Pardal roquer x x
Phalacrocorax aristotelis Corb marí emplomallat x
Phalacrocorax carbo Corb marí gros x x
Phasianus colchicus Faisà x x
Phoenicurus ochruros Cotxa fumada x x
Phoenicurus phoenicurus Cotxa cua-roja x x
Phylloscopus bonelli Mosquiter pàl·lid x x
Phylloscopus collybita Mosquiter comú x x
Phylloscopus ibericus Mosquiter ibèric x x
Phylloscopus sibilatrix Mosquiter xiulaire x x
Phylloscopus trochilus Mosquiter de passa x x
Pica pica Garsa x x
Picus sharpei Picot verd x x
Picus viridis Picot verd x x
Platalea leucorodia Becplaner x x
Plegadis falcinellus Capó reial x x
Pluvialis squatarola Pigre gris x
Podiceps cristatus Cabussó emplomallat x Podiceps nigricollis Cabussó collnegre x Poecile palustris Mallerenga d’aigua x x
Poicephalus senegalus Lloro del Senegal x Porphyrio porphyrio Polla blava x x
Prunella collaris Cercavores x x
Prunella modularis Pardal de bardissa x x
Psittacara acuticaudatus Aratinga capblava x
Psittacara mitratus Aratinga mitrada x x
Psittacula krameri Cotorra de kramer x x
Ptyonoprogne rupestris Roquerol x x
Pyrrhocorax pyrrhocorax Gralla de bec vermell x Pyrrhula pyrrhula Pinsà borroner x x
Rallus aquaticus Rascló x x
Recurvirostra avosetta Bec d’alena x
Regulus ignicapilla Bruel x x
Regulus regulus Reietó x x
Remiz pendulinus Teixidor x x
Riparia riparia Oreneta de ribera x x
Saxicola rubetra Bitxac rogenc x x
Saxicola rubicola Bitxac comú x x
Serinus canaria Canari x
Serinus serinus Gafarró x x
Sitta europaea Pica-soques blau x x
Spatula clypeata Ànec cullerot x x
Spatula querquedula Xarrasclet x
Spinus spinus Lluer x x
Streptopelia decaocto Tòrtora turca x x
Streptopelia turtur Tòrtora comuna x x
Strix aluco Gamarús x x
Sturnus unicolor Estornell negre x x
Sturnus vulgaris Estornell vulgar x x
Sylvia atricapilla Tallarol de casquet x x
Sylvia borin Tallarol gros x x
Sylvia conspicillata Tallarol trencamates x Tachybaptus ruficollis Cabusset x x
Tachymarptis melba Ballester x x
Tadorna tadorna Ànec blanc x x
Thalasseus sandvicensis Xatrac becllarg x x
Tringa erythropus Gamba roja pintada x
Tringa glareola Valona x x
Tringa nebularia Gamba verda x x
Tringa ochropus Xivita x x
Tringa totanus Gamba roja x x
Troglodytes troglodytes Cargolet x x
Turdus merula Merla x x
Turdus philomelos Tord x x
Turdus pilaris Griva cerdana x x
Turdus viscivorus Griva x x
Tyto alba Òliba x
Upupa epops Puput x x
Vanellus vanellus Fredeluga x x
Taula complementària 2. llista d’espècies de papallones presents a la RMB tingudes en compte per a l’estudi. Taula 2.1 espècies presents dins de PEIN i Taula 2.2 espècies presents fora de PEIN.
Nom científic
Nom comú català Present Present en PEIN fora PEIN
Danaus chrysippus Papallona tigre x
Danaus plexippus Monarca x
Carcharodus alceae Capgròs comú x x
Carcharodus baeticus Capgròs del malrubí x x
Carcharodus floccifera Capgròs fosc x
Carcharodus lavatherae
Capgròs pàl·lid x x
Erynnis tages Fúria x x
Gegenes nostrodamus Sageta negra x x
Hesperia comma Dard de taques blanques x x
Muschampia proto Fals merlet x x
Ochlodes sylvanus Dard ros x x
Pyrgus alveus
Pyrgus armoricanus
Pyrgus cirsii
Pyrgus carthami
Pyrgus malvoides
Pyrgus serratulae
Merlet major x
Merlet ruderal x x
Merlet rogenc x x
Merlet reial x
Merlet comú x x
Merlet olivaci x
Spialia sertorius Murri de la pimpinella x
Thymelicus acteon Daurat dosc x x
Thymelicus lineola Daurat de punta negra x
Thymelicus sylvestris Daurat de punta taronja x x
Libythea celtis
Aricia agestis
Aricia cramera
Cupido alcetas
Celastrina argiolus
Callophrys avis
Cacyreus marshalli
Cupido minimus
Cupido osiris
Callophrys rubi
Papallona del lledoner x x
Moreneta septentrional x x
Moreneta meridional x x
Cuetes del melgó x x
Blaveta de l’heura x x
Verdeta d’ull ros x x
Barrinadora del gerani x x
Cupido menut x x
Cupido blau x x
Verdeta d’ull blanc x x
Cupido argiades Cuetes de taques taronges x
Glaucopsyche alexis Turquesa europea x x
Glaucopsyche melanops
Hamearis lucina
Turquesa mediterrània x x
Papallona de la prímula x Iolana debilitata Blaveta de l’espantallops x Lycaena alciphron Coure tornassol x Lampides boeticus Blaveta dels pèsols x x
Lycaena phlaeas Coure comú x x
Leptotes pirithous Blaveta estriada x x Laeosopis roboris Morada del freixe x x Phengaris arion Formiguera gran x Favonius quercus Morada x x
Polyommatus amandus Blaveta de la garlanda x Plebejus argus Blavet argiu x x
Lysandra bellargus Blaveta lluent x x
Lysandra coridon Griseta de muntanya x x
Polyommatus escheri Blaveta de l’astràgal x x
Polyommatus fulgens Griseta de vellut x x
Lysandra hispana Griseta mediterrània x x
Polyommatus icarus Blaveta comuna x x
Pseudophilotes panoptes Blaveta de la farigola x x
Polyommatus ripartii
Cyaniris semiargus
Polyommatus thersites
Satyrium acaciae
Satyrium esculi
Satyrium ilicis
Scolitantides orion
Satyrium spini
Satyrium w-album
Tomares ballus
Thecla betulae
Marroneta de vellut x x
Cobalt x
Blaveta de la trepadella x x
Marroneta de l’aranyoner x
Marroneta de l’alzina x x
Marroneta del roure x x
Blaveta del crespinell x x
Marroneta de taca blava x x
Marroneta de l’om x x
Coure verdet x x
Tecla x x
Fabriciana adippe
Speyeria aglaja
Apatura ilia
Araschnia levana
Argentada de punts vermells x x
Argentada de muntanya x x
Tornassolada petita x x
Teranyina x Argynnis paphia
Argentada comuna x x Aglais urticae
Papallona de les ortigues x x Brenthis daphne
Boloria dia
Vanessa cardui
Charaxes jasius
Euphydryas aurinia
Perlada de l’esbarzer x x
Donzella violeta x x
Migradora dels cards x x
Papallona de l’arboç x x
Brocat variable x x
Aglais io Paó de dia x x
Issoria lathonia Mirallets x x
Limenitis camilla Nimfa de bosc x x Limenitis reducta Nimfa mediterrània x x
Melitaea celadussa
Damer boscà x Melitaea cinxia
Damer puntejat x x
Melitaea diamina Damer de la valeriana x Melitaea deione Damer dels conillets x x
Melitaea didyma Damer roig x x Melitaea parthenoides Damer de prat x x
Melitaea phoebe Damer de la centàurea x x Melitaea trivia Damer de la blenera x x Nymphalis antiopa Vellutada del salze x x Nymphalis polychloros Nimfa dorment x x Polygonia c-album Papallona de la c blanca x x Argynnis pandora Pandora x x Vanessa atalanta Atalanta x x Iphiclides feisthamelii Reina zebrada x x Papilio machaon Papallona reina x x Zerynthia rumina Arlequí x x Anthocharis cardamines Aurora x x Aporia crataegi Blanca de l’arç x x Anthocharis euphenoides Aurora groga x x Colias alfacariensis Safranera pàl·lida x x Colias crocea Safranera de l’alfals x x Euchloe belemia Marbrada zebrada x Euchloe crameri Marbrada comuna x x Gonepteryx cleopatra Cleòpatra x x Gonepteryx rhamni Llimonera x x Leptidea reali Angelet de muntanya x Leptidea sinapis Angelet comú x x Pieris brassicae Blanca de la col x x Pontia daplidice Pòntia comuna x x Pieris mannii Blanqueta de mitja lluna x x Pieris napi Blanqueta perfumada x x Pieris rapae Blanqueta de la col x x Arethusana arethusa Fals faune x Aphantopus hyperantus Papallona dels ullets x Brintesia circe Bruixa x x Coenonympha arcania Lleonada de matollar x x Chazara briseis Bruixa petita x Coenonympha dorus Lleonada de garriga x x Coenonympha pamphilus Lleonada comuna x x Erebia meolans Muntanyesa comuna x Hipparchia hermione Faune petit x Hipparchia fagi Faune gran x x Hipparchia fidia Faune ziga-zaga x x Hipparchia semele Faune lleonat x x Hipparchia statilinus Faune bru x x Lasiommata maera Margenera gran x Lasiommata megera Margenera comuna x x Maniola jurtina Bruna de prat x x Melanargia lachesis Escac ibèric x x Melanargia occitanica Escac ferruginós x x Pararge aegeria Bruna de bosc x x
Pyronia bathseba Saltabardisses cintada x x Pyronia cecilia Saltabardisses de solell x x Pyronia tithonus Saltabardisses europea x x Satyrus actaea Sàtir petit x x
J. MAURI ET AL.
Taula complementària 3. llistat dels itineraris SOCC tinguts en compte en l’estudi, indicant si pertanyen o no a un PEIN així com les coordenades.
IITINERARI SOCC PEIN X
3 SÍ 407919,1269 4568093,491
5 NO 423049,1308 4577111,579
7 NO 403446,3922 4599200,267
11 SÍ 436403,5934 4592800,846
13 SÍ 453873,847 4608061,76
15 NO 413565,5996 4584672,313
16 SÍ 391748,1401 4566172,552
20 SÍ 409066,9816 4600803,466
22 NO 433660,2011 4613156,62
25 SÍ 462648,7673 4613778,527
26 SÍ 428432,9493 4592502,626
35 NO 448715,0038 4615034,083
39 NO 386474,7994 4577941,91
40 NO 418248,4115 4582871,871
53 NO 416184,7591 4568953,759
55 NO 432021,6193 4581971,155
58 NO 431047,5051 4605414,918
60 SÍ 445274,7926 4598086,355
63 NO 433789,0128 4592503,755
66 SÍ 464065,2385 4613247,572
77 SÍ 396455,2223 4564090,427
84 SÍ 419968,8783 4616902,313
85 SÍ 421808,5904 4617127,483
86 SÍ 422997,1215 4615827,822
87 NO 385288,2758 4587078,526
88 NO 425267,0449 4574484,152
91 NO 418660,316 4578966,862
93 SÍ 476676,7194 4614204,123
94 SÍ 403069,9637 4605134,922
95 SÍ 384724,3083 4568862,2
104 SÍ 433083,892 4616311,431
107 NO 411611,7937 4591673,291
110 NO 421311,9725 4600006,052
111 NO 416798,4121 4603923,13
112 NO 425405,053 4594076,111
113 SÍ 409810,4591 4603288,967
114 SÍ 433488,8285 4600422,11
119 SÍ 428742,0689 4587994,434
126 SÍ 402849,9186 4583045,85
128 NO 452964,1225 4616038,852
140 NO 423650,1608 4597721,876
143 NO 454500,2999 4599535,618
146 SÍ 455176,2623 4623298,632
147 SÍ 444663,8019 4623768,23
148 SÍ 440049,8955 4625517,48
149 SÍ 393176,5051 4574626,195
160 SÍ 402692,0991 4573751,554
165 SÍ 408590,2385 4570678,441
168 NO 457134,8627 4614391,245
184 SÍ 412068,9082 4607182,861
185 SÍ 415566,423 4608143,401
186 SÍ 418418,6086 4609718,882
190 SÍ 420684,5009 4613200,059
192 SÍ 463923,5017 4612220,166
205 NO 411175,1356 4594065,887
215 NO 418781,2562 4569880,481
216 NO 432826,2783 4582152,469
218 NO 434315,0252 4596039,498
226 NO 432360,5706 4589933,606
227 SÍ 437755,3224 4600366,872
228 NO 392585,8395 4586690,609
230 SÍ 445094,333 4600116,296
231 NO 429503,6541 4583057,748
232 NO 430692,9828 4580279,64
233 NO 431769,5623 4587860,115
234 SÍ 424769,7164 4582443,535
236 NO 382213,9416 4576253,202
244 SÍ 457196,1017 4624003,678
245 NO 401377,9813 4590122,306
246 NO 433547,2356 4585939,745
248 NO 383744,7833 4593006,62
256 NO 436838,1962 4603204,54
257 SÍ 420226,3759 4569990,872
258 SÍ 426946,2116 4584677,836
276 NO 430471,1699 4579912,3
277 SÍ 415190,3455 4579758,871
291 SÍ 439680,5481 4600611,293
312 SÍ 440869,7579 4600979,574
321 NO 462008,2337 4605580,178
322 NO 425220,1225 4604739,032
323 NO 454686,337 4600249,724
348 NO 471096,1983 4607259,638
382 SÍ 425987,7728 4571491,341
388 SÍ 455012,2155 4604193,822
389 SÍ 441011,2349 4594417,855
410 SÍ 458241,2699 4614743,51
414 NO 444578,6525 4592821,984
415 SÍ 447851,4909 4596549,61
418 NO 433267,915 4594702,828 425 NO 432580,5836 4588495,572
426 SÍ 407152,0687 4599185,533
488 SÍ 433871,4124 4591667,278
490 SÍ 448458,996 4597424,292
499 NO 466125,8783 4606058,075
501 NO 410825,3489 4575831,665
520 SÍ 451136,1247 4628064,575
531 NO 418159,8513 4574405,404
539 SÍ 454306,0517 4624123,372
540 NO 437107,7208 4589137,187
546 SÍ 449693,6763 4620596,105
551 SÍ 401240,2499 4571576,5
556 SÍ 385848,9466 4569118,291
557 NO 413325,1 4597922,45
561 NO 424822,6 4593128,5
563 SÍ 411833,1051 4573704,295
564 NO 432721,9363 4590241,093
567 SÍ 388651,15 4567448,35
577 SÍ 408610,2161 4568309,942
584 NO 405045,1 4596012,775
588 NO 410577,4508 4601324,71
590 NO 431282,7191 4594100,187
591 SÍ 424075,5986 4586594,996
592 NO 414682,3895 4569796,979
594 NO 461001,4344 4618235,85
595 NO 422384,4197 4574758,683
596 SÍ 416820,3978 4569630,649
597 NO 426339,8079 4600885,86
605 NO 399513,5841 4582025,389
608 NO 418231,019 4600927,944
610 NO 422903,9936 4599621,271
Taula complementària 4. llistat dels itineraris CBMS tinguts en compte en l’estudi, indicant si pertanyen o no a un PEIN així com les coordenades.

Joan Pedrol Solanes*& Antoni Mayoral Arqué*
* Institut d’Estudis Ilerdencs. Secció de Botànica. Pl. Catedral s/n. 25002 Lleida.
Autor de correspondència: Joan Pedrol. A/e: jpedrolsolanes@gmail.com
Rebut: 17.04.2024. Acceptat: 13.05.2024. Publicat: 30.06.2024
Genista teretifolia Willk.
Aquesta lleguminosa és un endemisme dels Pirineus centrals i occidentals. Descrita dels voltants de Pamplona a mitjan segle XIX, la seva àrea coneguda tradicionalment comprenia Àlaba, Navarra, nord de Saragossa i oest d’Osca. No va ser fins a la darrera dècada del segle XX quan es van donar a conèixer les poblacions de la Ribagorça aragonesa (Montserrat, 1988; Villar et al., 1997), de les quals la del Montsec d’Estall és la més propera a Catalunya. L’única referència catalana d’aquesta planta era el plec MA 433233: «[Montsec d’Ares] Encima de la ermita de la virgen de Fabregada, 1200 m, calizas, 25-VI-1986, P. Catalán, I. Aizpuru, F. Muñoz Garmendia & J. Pedrol». La indicació provincial de Lleida en Flora iberica (Talavera, 1999) es basa en aquest material.
S’ha realitzat una exploració més detallada del territori i n’hem localitzat dues noves poblacions, que estan situades a l’obaga del Montsec d’Ares. A continuació, en donem les referències.
Pallars Jussà: obaga del Montsec d’Ares, per sobre de la Mare de Déu de la Fabregada, carena entre el barranc de Coma-roia i el de la Pua, CG1360, 1095 m, clariana de carrascar sobre calcarenites, 18-V-2023, J. Pedrol 11859JP (HBIL19553); pista de la Fabregada al coll d’Ares, collada del Comú, CG1259, 1200 m, marge de pista sobre calcarenites, 16-VI-2023, J. Pedrol 11902JP (HBIL19554).
La primera d’aquestes localitats correspon, malgrat les diferències d’altitud, amb quasi total seguretat a la de la recollecció de 1986. Si és així, cal ressaltar que la indicació de substrat que llavors es va donar és errònia, perquè en realitat la planta creix sobre un sòl format a partir d’un substrat de calcarenites. En aquestes poblacions del Montsec d’Ares (Fig. 1) la planta forma pulvínuls en què les tiges prostrades sovint queden enterrades i les ascendents no superen els 15 cm d’alçada. Apareix en llocs oberts, com clarianes del carrascar i zones desforestades, però prospera molt bé en les cunetes de les pistes, on sembla que actua com a colonitzadora, sempre sobre calcarenites del Maastrichtià que originen sòls de reacció àcida. A la Baixa Ribagorça, Gil & Montserrat (1992) esmenten que la planta també viu a les crestes for-
DOI: 10.2436/20.1502.01.177

mades per conglomerats, un hàbitat on de moment no l’hem detectada a Catalunya.
En la primera població s’han comptat uns 10 individus, mentre que en la segona s’hi distingeixen tres subpoblacions, aparentment amb un total d’uns 15 individus. Cal tenir present però, que la planta presenta tiges prostrades que queden enterrades. Això en dificulta el recompte, i per tant el nombre total d’individus total pot ser menor.
Puccinellia rupestris (With.) Fernald. & Weath.
Aquesta gramínia anual no ha estat recollida com a planta catalana a Flora iberica (Ruiz de Clavijo & Devesa, 2020) ni a la Checklist de Sáez & Aymerich (2021), i tampoc apareix al BDBC (Font, 2023), malgrat que Mayoral (1998) ja en va publicar una localitat de la Plana d’Urgell fa força anys. Ruiz de Clavijo & Devesa (2020) la consideren una espècie difosa sobretot pel N de la península Ibèrica. A la vall de l’Ebre presenta unes poques localitats (Julià, 1991), de les quals la més propera a Catalunya és de la Serreta Negra de Fraga (Montserrat & Montserrat, 1986).

A Catalunya, a la primera referència de Mayoral (1998) –entre Linyola i Vallfogona de Balaguer (CG2021, 231 m)–, n’hi hem pogut afegir unes poques més, repartides per les comarques de la Noguera i el Segrià. Alguns dels plecs dipositats a l’herbari HBIL són els que figuren a la relació següent.
Noguera: entre Linyola i Vallfogona de Balaguer, CG22, Coronopodo-Sclerochloetum durae, 14-VIII-1992, A. Mayoral (HBIL 7447); id., depressió salina nitrogenada, Coronopodo-Sclerochloetum durae, 23-V-1994, A. Mayoral (HBIL 12101); id., depressió salina, 1-V-2003, A. Mayoral (HBIL 15707); Penelles, Castell del Remei, CG3220, 250 m, llocs trepitjats i més o menys salins, 30-VI-1998, J. Pino & J. Pedrol 6382JP (HBIL 12737); entre Térmens i Bellvís, CG1619, 197 m, terreny salí que es fa anar com a femer, 30-IV-2023, A. Mayoral (HBIL19571). Segrià: Alcoletge, cap a Bell-lloc, CG1012, 170 m, fondos salins, 2-V-2022, J. Pedrol & A. Tarragó 11732JP (HBIL 16388).
Puccinellia rupestris es diferencia fàcilment dels seus congèneres pel fet de presentar una panícula (Fig. 2) molt més compacta. Com aquests, es troba en depressions salines, però la seva ubicació en indrets més trepitjats, vores de camins, camps fortament adobats i, fins i tot, en terrenys que es fan servir com a femers, fa que l’haguem de considerar més nitròfila, més ruderal. D’ençà l’any 1992, ha estat observada amb regularitat a les terres de Ponent i les seves poblacions han experimentat osciŀlacions que van de pocs a més d’un centenar d’individus. El seu òptim de floració esdevé entre mitjan abril i primers de juny, i en alguna ocasió s’han vist exemplars florits a l’agost.
Thlaspi alliaceum L. (Mummenhoffia alliacea (L.) Esmailbegi & Al-Shehbaz)
Segrià: Lleida, la Mitjana, 130 m, 5-V-1999, F. Royo (MTE 13351); Lleida, la Mitjana, 31TCG0411, 150 m, herbassar, 1-IV-2023, J. Pedrol 11821 JP (HBIL19281).
Thlaspi alliaceum és una crucífera anual de fins a 60(80) cm, en general unicaule. Segons Pujadas (1993) es distribueix pel centre i sud d’Europa i oest d’Àsia, mentre que a la

península Ibèrica apareix dispersa. En aquest territori, té unes poques localitats comprovades al N (Palència, Cantàbria i Àlaba) i al centre (Madrid) però amb referències bibliogràfiques més nombroses no comprovades.
Malgrat que no està recollida com a planta catalana (Sáez & Aymerich, 2021), ja havia estat coŀlectada per en Ferran Royo l’any 1999 a la Mitjana de Lleida, on nosaltres la vam veure també entre els anys 2011-2013 i on durant l’any 2023 l’hem tornat a localitzar i n’hem fet el seguiment.
El Parc de la Mitjana de Lleida és un espai d’unes 90 ha integrat en l’àrea periurbana de la ciutat que correspon a un espai fluvial recorregut pel riu Segre. Està destinat a usos d’esbarjo i educatius, i té una gran quantitat d’ambients naturals, molts d’ells relacionats amb la presència d’aigua (Conesa et al., 1994). Dins d’aquest espai, durant la primavera de 2023, en vam trobar dos rodals separats per uns 300 m. Cadascun tenia prop de mig centenar d’individus, que es van desenvolupar en els mesos de març i abril. L’hàbitat correspon a herbassars de teròfits de primavera, sovint molt atapeïts, amb, entre d’altres, Avena barbata Link, Bromus diandrus Roth, Cardaria draba (L.) Desv., Erodium cicutarium (L.) L’Hér., Euphorbia helioscopia L. o Melilotus spicatus (Sm.) Breistr. Pensem que a la Mitjana hi està ben establerta almenys des de l’any 1999. Donat que el parc té extensions importants d’herbassars de primavera, a més d’àrees de reserva d’accés restringit que no hem explorat aquest any, cal suposar que la planta és més abundant encara. La seva fenologia, amb un cicle molt curt, i la semblança amb Th. perfoliatum L. (Microthlaspi perfoliatum (L.) F.K. Mey., Noccaea perfoliata (L.) Al-Shehbaz) (Fig. 3) –de
la qual es diferencia bé pel seu port més estilitzat, la presència de pèls patents a la base de la tija i l’ornamentació de les sements–, pot ser la causa del fet que hagi passat força desapercebuda. Per determinar la seva àrea real caldrà explorar també hàbitats similars al llarg del curs del Baix Segre.
Pel que fa a l’estatus d’aquesta espècie, a la península Ibèrica Pujadas (1993) la considera autòctona, malgrat la dispersió de les seves localitats. A França Tisson & Foucault (2014) també la consideren autòctona al sud-oest, Provença i vall de la Loira, mentre que l’assenyalen com a naturalitzada i en expansió a l’Alvèrnia i al Lionès. Nosaltres, amb les dades de què disposem fins ara, no ens atrevim a pronunciar-nos al respecte, pensem que cal observar més detingudament el seu comportament al nostre territori.
Aprofitem per indicar que la referència que apareix a GBIF (2024) de Th. alliaceum d’Andorra [pr. Canillo, 1690 m, 42º 34’ 16” N, 01º 35’ 54” E, C. Aedo, I. Aizpuru, P. Bariego, J. Pedrol & C. Molina, 29-V-2004. ARAN 66209] correspon a Th. perfoliatum, a l’igual que els duplicats revisats de la mateixa recoŀlecció (MA 714996, HBIL 16372). Cal excloure doncs Th. alliaceum de la flora andorrana.
Agraïments
A Alfredo Pérez Meler, per la seva ajuda i entusiasme en la recerca al camp de Genista teretifolia. Albert Tarragó és l’autor de les fotografies de Thlaspi alliaceum que ens ha cedit amablement. Iñaki Aizpuru i Leopoldo Medina ens han ajudat amb diverses recerques als herbaris ARAN i MA respectivament. Amb Carlos Aedo hem discutit diversos aspectes de la morfologia i corologia de Thlaspi alliaceum
Bibliografia
Conesa, J. A., Mayoral, A., Pedrol, J. & Recasens, J. 1994. El paisatge vegetal dels espais d’interès natural de Lleida: àrea meridional. Institut d’Estudis Ilerdencs. Lleida. 194 p.
Font, X. 2023. Mòdul Flora i Vegetació. Banc de Dades de Biodiversitat de Catalunya. Generalitat de Catalunya i Universitat de Barcelona. Disponible online: http://biodiver.bio.ub.es/biocat/ homepage.html [Data de consulta: 8 març 2024]
GBIF. 2024. Global Biodiversity Information Facility. Disponible online: https://www.gbif.org. Consultat mitjançant https://www. gbif.org/es/species/3052738 [Data de consulta: 10 d’abril 2024]
Gil, E. & Montserrat, G. 1992. Notas sobre distribución y ecología de Genista teretifolia Willk. P. 353-358. Actes del Simposi Internacional de Botànica ‘Pius Font i Quer’, Vol. 2 (Fanerogàmia). Lleida. 496 p.
Julià, M. A. 1991. El gènere Puccinellia Parl. (Poaceae) a la península Ibèrica. Barcelona. 288 p. Mayoral, A. 1998. Notes florístiques de la Plana d’Urgell, V. Ilerda «Ciències», 52: 25-28.
Montserrat, G. 1988. Genista teretifolia y Medicago coronata, novedades para el Prepirineo central. Anales del Jardín Botánico de Madrid, 45: 342-343.
Montserrat, G. & Montserrat, J. M. 1986. Notas citotaxonómicas sobre el género Puccinellia (Poaceae) en la Península Ibérica. Collectanea Botanica (Barcelona), 16: 341-349.
Pujadas, A. 1993. Thlaspi L. P. 255-264. In: Castroviejo, S., Aedo, C., Gómez Campo, C., Laínz, M., Montserrat, P., Morales, R., Muñoz Garmendia, F., Nieto Feliner, G., Rico, E., Talavera, S. & Villar, L. (eds.). Flora iberica IV. Real Jardín Botánico-CSIC. Madrid. 730 p.
Ruiz de Clavijo, E. & Devesa, J. A. 2020. Puccinellia Parl. P. 598612. In: Devesa, J. A., Buira, A., Romero Zarco, C., Quintanar, A. & Aedo, C. (eds.). Flora iberica XIX(I). Real Jardín BotánicoCSIC, Madrid. 738 p. Sáez, L. & Aymerich, P. 2021. An annotated Checklist of the Vascular Plants of Catalonia (northeastern Iberian Peninsula). Kit-book Serveis Editorials. Barcelona. 717 p. Talavera, S. 1999. Genista L. P. 45-119. In: Talavera, S., Aedo, C., Castroviejo, S., Romero Zarco, C., Sáez, L., Salgueiro, F.J. & Velayos, M. –eds.–. Flora iberica VII(I). Real Jardín BotánicoCSIC, Madrid. 578 p. Tison, J. M. & de Foucault, B. (coords.) 2014. Flora Gallica. Flore de France. Biotope. Mèze. xx +1.196 p. Villar, L., Sesé, J. A. & Ferrández, J. V. 1997. Flora del Pirineo Aragonés, vol. I. Consejo de Protección de la Naturaleza de Aragón – Instituto de Estudios Altoaragoneses. Huesca. 648 p.
* Museu de Ciències Naturals de Barcelona. Laboratori de Natura. Coŀlecció d’artròpodes. Passeig Picasso, s/n. 08003 Barcelona. A/e: lluisfilba@gmail.com
Rebut: 28.10.2023; Acceptat: 20.05.2024; Publicat: 30.06.2024
Resum
Els tricòpters hipogeus constitueixen una part important del poblament dels artròpodes de l’anomenada associació parietal de les cavitats subterrànies. En l’àrea iberobalear es coneixen 11 espècies de tricòpters de caràcter subtroglòfil. Aquest article parla de la seva posició sistemàtica i la seva taxonomia, i dona dades de la seva distribució. S’acompanya el treball amb imatges que faciliten la identificació dels diferents taxons. En diferents taules es dona el nombre de localitats per unitat geogràfica, el nombre de cavitats d’on procedeixen les mostres de cada espècie, el període d’estada subterrània i la concurrència d’espècies en una mateixa cavitat.
Paraules clau: Insecta, Trichoptera, subtroglòfils, grup Stenophylax, iberobalears.
Abstract
Catalog of the Ibero-Balearic hypogean caddisflies (Insecta: Trichoptera) from the collection of the Natural Sciences Museum of Barcelona
Hypogean caddisflies constitute an important part of the arthropod population of the so-called parietal association of subterranean cavities. In the Iberian-Balearic area, 11 species of subtroglophile trichoptera are known, of their systematic position taxonomy and distribution data are provided. The work is accompanied by images that facilitate the identification and recognition of different taxa. The provided tables indicate the number of localities per geographical unit, the number of cavities from which the samples come for each species, and the period of underground dwelling ,and the ocurrence of species in the same cavity.
Key words: Insecta, Trichoptera, subtroglophiles, Stenophylax group, Ibero-Balearic area.
Introducció
La coŀlecció de tricòpters hipogeus del Museu de Ciències Naturals de Barcelona té els seu origen en les recerques bioespeleològiques de la dècada de 1950, que es van fer sota la direcció del Dr. Francesc Español quan exercia el càrrec de conservador del Museu de Zoologia de Barcelona. Les primeres recoŀleccions les va fer el mateix Español i persones del seu cercle de coŀlaboradors, com Lluís Auroux, Joan Senent i, principalment, Isidre Gonzàlez-Urgellés, que inicià l’estudi d’aquest grup amb la determinació d’alguns taxons.
L’any 1956, F. Español envià a l’entomòleg de la ciutat suïssa de Lausanne Fernand Schmid uns exemplars de tricòpters recoŀlectats per ell mateix. Entre aquests espècimens hi havia uns exemplars d’una nova espècie que, en honor a l’entomòleg català, el professor Schmid va descriure com a Stenophylax espanioli Schmid, 1957.
Fruit d’aquesta relació i de l’intercanvi de mostres entre els dos entomòlegs, el Dr. F. Espanyol s’avingué a publicar un treball que F. Schmid tenia tot just acabat sobre els tricòpters cavernícoles (Font: Arxiu MCNB). Efectivament, el
1957, aparegué un opuscle que és de referència per a la taxonomia d’aquest grup (Schmid, 1957).
Entrats els anys de 1970, una època àlgida de la bioespeleologia a Catalunya, es multipliquen les recerques de fauna subterrània, juntament amb la recoŀlecció d’organismes troglobis, i són també objecte d’interès els artròpodes que colonitzen les entrades de les cavitats. Des d’aquesta època fins a finals del s. XX, bioespeleòlegs com M. Nebot, X. Bellés, J. Comas, C. Ribera i O. Escolà, juntament amb les agrupacions espeleològiques Grup d’Investigacions Espeleològiques de Montblanc i Secció d’Activitats Subterrànies, són els autors de més del 70 % de les recoŀleccions. És de destacar l’aportació d’Oleguer Escolà (conservador del Museu de Zoologia de Barcelona a l’època), recoŀlector de prop del 40 % del total de mostres de la coŀlecció. Als precedents, cal afegir els noms de J. Mederos i F. Fadrique, que han fet recoŀleccions en el segle actual.
Material i mètodes
Les mostres de tricòpters troglòfils procedeixen dels re-
L. FILBÀ ESQUERRA
gistres establerts al Museu de Ciències Naturals de Barcelona de recoŀleccions fetes entre els anys 1951 i 2019, i han estat revisades o determinades entre el 2018 i el 2021 (veure apèndix amb les dades especificades). Anteriorment al procés d’actualització, l’autor d’aquest catàleg havia publicat uns resultats en notes sobre la presència de les espècies de tricòpters cavernícoles iberobalears (Filbà Esquerra, 1974a, 1974b, 1977).
Ara, en ocasió de la redacció del present catàleg, s’han revisat i s’han inclòs els registres citats en aquests treballs inicials, amb la intenció de donar una unitat informativa al catàleg i també per entrar-hi algunes esmenes. També s’han completat algunes dades de localització de les cavitats. En la majoria de casos, s’ha afegit l’altitud a la ubicació.
Les dades primàries de camp dels registres procedeixen de srotcel ŀ ocer slep setef de les mostres, les quals es limiten al nom de la cavitat, la data de recoŀlecció i el nom del recoŀlector, el qual és seguit per l’abreviació leg., que és la del mot llatí legitur. En alguns casos hi falta el nom del recoŀlector, i a voltes també la data.
En alguns casos, el registre de les mostres ha servit per revisar el nom de la cavitat, per corregir la seva ubicació en la unitat geogràfica o municipal o per anotar-ne l’altitud. En el cas de Catalunya, ha estat d’inestimable servei el catàleg d’Espeleoíndex i, en el de Navarra, el Catàleg Espeleològic de Navarra.
El caràcter subtroglòfil dels tricòpters hipogeus
La zona vestibular de les cavitats subterrànies constitueix l’habitat de determinats artròpodes.
Algunes espècies d’insectes tricòpters (Insecta: Trichoptera), són habituals en les parets i sostres a l’entrada de coves, avencs, o mines artificials, en la zona de penombra, amb unes condicions climàtiques marcades per un alt grau de saturació d’humitat ambiental i a recer de corrents d’aire permanents. Els tricòpters, juntament amb lepidòpters, araneids, opilions i himenòpters, constitueixen l’anomenada “associació parietal” de les coves (Jeannel, 1926).
L’estada sota terra en la fase adulta del grup de tricòpters de la família Limnephilidae, anomenat el «grup Stenophylax» (Schmid, 1957), és admès que va lligada a la diapausa que experimenten, i és considerada com un fenomen indispensable per al desenvolupament normal del seu cicle biològic (Bouvet &Ginet, 1969), de manera que les coves actuen de refugi a l’espera de la revinguda de les aigües als rierols on fan la posta (Bouvet, 1971). Els adults no s’alimenten, i únicament succionen aigua de les parets (Salavert, 2012).
A inicis de primavera fan el vol de migració cap als refugis subterranis i, cap a final de tardor (amb un pic al mes de novembre), retornen a les aigües on es desenvolupen les fases immadures del cicle.
Respecte a la migració dels adults cap a l’exterior, el desencadenant per iniciar la sortida és la inversió tèrmica cavitat-exterior (Salavert, 2012).
Pel comportament explicat, els tricòpters del grup Stenophylax se situen dins la categoria ecològica dels subtroglòfils,
(2), 2024
que colonitzen temporalment el medi cavernícola perquè això afavoreix el desenvolupament del seu cicle biològic (Bellés, 1987).
Posició sistemàtica
Les onze espècies de tricòpters hipogeus de caràcter subtroglòfil de l’àrea iberobalear s’inclouen dins el grup Stenophylax (Schmid, 1955), de la familia Limnephilidae. Són insectes de mida grossa dins de l’ordre (longitud de l’ala anterior entre 18 i 22 mm.).


1. a) Stenophylax sp. (Cueva de los Arnales, Tella-Sin (Sobrarbe), [fotografia d’A. Meseguer]. b) Mesophylax aspersus (la Mina dels Ports, Tortosa), [fotografia d’A. Meseguer].
Ordre Trichoptera Kirby, 1813
Família Limnephilidae Kolenati, 1848
Gènere Stenophylax Kolenati, 1848 (Fig. 1a)
S. espanioli Schmid, 1957
S. fissus (McLachlan, 1875)
S. malatestus (Schmid, 1957)
S. mitis McLachlan, 1875
S. mucronatus McLachlan, 1880
S. nycterobius (McLachlan, 1875)
S. permistus McLachlan, 1875
S. sequax (McLachlan, 1875)
S. testaceus (Gmelin, 1788)
S. vibex (Curtis, 1834)
Gènere Mesophylax McLachlan, 1882
M. aspersus (Rambur, 1842) (Fig. 1b)
Nomenclatura taxonòmica
En la revisió dels gèneres Stenophylax (Kolenati 1848) i Micropterna (Stein, 1874), Schmid (1957) assenyala que «Micropterna només es diferencia de Stenophylax per un sol caràcter: a les potes anteriors del mascle el protars és molt breu i més curt que el segon artell».
El mateix autor observa, no obstant, que aquest caràcter és inconstant, per la seva diferent longitud segons l’espècie i pel fet de no presentar-se en tres de les espècies del gènere. Reconeix també que «aquest sol caràcter sexual secundari no és suficient per requerir una separació genèrica de Stenophylax».
L’autor reconeix la validesa de Micropterna ja que malgrat aquest feble caràcter que el separa de Stenophylax, ambdós gèneres constitueixen dues unitats filètiques diferents.
Taula 1. Nombre de localitats de cada espècie. S’indica la regió i la unitat geogràfica on estan situades.
Nombre de localitats de cada especieper unitat geográfica
Tricòpters subtroglòfils del grup Stenophylax
Unes dècades després, Botosaneanu (1992), en un treball sobre els tricòpters de la Mediterrània oriental, proposà alguns canvis de nomenclatura, entre els quals hi ha la inclusió del gènere Micropterna dins el gènere Stenophylax.
Malicky (1993) es mostra en desacord amb aquests canvis i opina que, malgrat el caràcter del protars a Micropterna, «durant més de trenta anys la majoria d’autors han mantingut els dos gèneres per raó d’una continuïtat en la nomenclatura, considerant que un canvi de nom en uns insectes tan ben coneguts no aporta cap avenç a la ciència, sinó confusió».
A causa dels dos criteris exposats, actualment es mantenen les dues nomenclatures. El catàleg de Fauna Ibèrica (González et al., 1992) suprimeix el gènere Micropterna en favor de Stenophylax. El mateix criteri segueixen els autors González & Martínez (2011), mentre que el projecte Fauna Europaea manté el gènere Micropterna. En el present catàleg s’ha adoptat la nomenclatura seguida a Fauna Ibèrica.
Elements de la coŀlecció
El catàleg comprèn 671 mostres corresponents a sengles localitats (tàxon, lloc, data i leg.) de les 11 espècies (Taula 1), procedents de 374 cavitats subterrànies iberobalears (Taula 2, Annex). En la Taula 1 s’exposa el nombre de localitats de cada espècie per unitat geogràfica, i s’observa un clar predomini de les localitats de l’Aragó i de Catalunya, a causa de la gran activitat de recerca radiada des de la capital catalana.
Les dades de recoŀlecció i de determinació dels espècimens estudiats en la realització d’aquest catàleg, es poden consultar a GBIF.org (10 January 2024) GBIF Occurrence download https://doi.org/10.15468/dl.nz2r29
Període d’estada subterrània
La durada i l’època de la fase hipogea és variable segons l’espècie, com queda exposat en la Taula 3. S’observa que, en aquelles espècies que compten amb més registres, el període de vol abasta quasi tot l’any. És el cas de S. mucronatus, M. aspersus, S. fissus i S. malatestus. En les espècies restants es pot notar que en S.espanioli, S. sequax i S. testaceus l’es-
tada s’inicia tard a la primavera i s’allarga a la tardor. S. mitis, S. permistus i S. vibex tenen un període més curt que es concentra a l’estiu. S. nycterobius és l’espècie que té el pic d’afluència més tard.
Concurrència d’espècies en una mateixa cavitat
En un terç aproximat de les cavitats estudiades se citen dues o més espècies de tricòpters subtroglòfils. La presència de les espècies en una mateixa cavitat s’ha determinat a partir de totes les captures fetes en el període temporal que abasta les mostres. Aquesta concurrència no suposa forçosament la seva presència simultània.
Considerem els casos de concurrència de quatre o més espècies (15 cavitats) com un fet singular, que es podria atribuir tant al nombre d’espècies de tricòpters subtroglòfils que poblen un determinat massís muntanyós com a la situació d’aïllament geogràfic de la cavitat que, juntament amb les dimensions del seu pòrtic i les condicions climàtiques requerides per a la diapausa dels adults, serien els factors que podrien atraure el vol de migració cap a un refugi comú.
A la Taula 4 es dona la relació d’aquestes cavitats i el nom de les espècies que hi concorren.
És de ressaltar el cas únic de la presència de 9 espècies, que no hi concorren simultàniament sinó en una estada diferenciada entre els mesos de juny (3 sp.), juliol (8 sp.) i agost (5 sp.). Es dona a la cova d’Allará (Val d’Echo, Pirineu axial).
Els caràcters distintius de cada espècie
Un dels propòsits a l’hora de redactar el present catàleg és facilitar les orientacions per a una correcta determinació de les espècies. El tricòpters del grup Stenophylax són insectes de mida grossa dins l’ordre. Aquest fet en facilita la manipulació i fa innecessària la preparació de genitàlies. L’observació de l’aparell copulador en visions lateral i ventral són suficients per distingir les espècies. Les fotografies s’acompanyen en algun cas d’un dibuix per destacar caràcters poc visibles i que són determinants. Aquests caràcters són ressaltats sovint mitjançant unes indicacions gràfiques.
de cavitats: 374).
Nombre de cavitats amb registres de tricòpters subtroglòfils
Nord peninsular Costa Est peninsular
Massís Galaic 1
Serralada Cantàbrica 17
Sistema Central ibèric 1
Muntanyes Basques 10
Sistema Ibèric 7
Serralada Transversal Catalana 9
Serralada Prelitoral Catalana 113
Serralada Litoral Catalana 29
Depresió Central Catalana 5
Llevant peninsular 26
Pirineu axial 67 182
Prepirineu 83 186
Sud peninsular Penibètica 3 Illes
Subbètica 1
Illes Balears 2
Taula 3. Període d’estada subterrània. S’indica el nombre d’exemplars mascle (total 1.138) i femella (total 711) de cada espècie per cada mes de l’any. S’indica també el nombre de localitats de les quals provenen les dades.
Taula 4. Llista de les cavitats on concorren d’entre 4 a 9 espècies
Espècies que concorrent a cada cavitat
Nom de la cavitat Nombre (Municipi) d’espècies
Avenc de la Suor 4
(Alfare de Carles)
Avenc de les Clivelleres (num. 1 (Mediona) 4 X
Cueva de la Galiana (Ucero) 4
Cueva del Beral (Ansó) 4
Graller del Boixeguer 4
(St. Esteve de la Sarga) (Casbas de Huesca)
Solencio de Bastaras 4
Tub de Captació (Canfranc) 4
Cova de les Rondes (La Llacuna) 4
Bora Fosca (Tavertet) 5
Cova de la Font Mentidora 5
(Conca de Dalt)
Forat de l’Aire (Baells) 5
Mina de Can Montsant (Tordera) 5
Avenc del GIEM (La Febró) 5 X
Surgència de Poblet (Mont-ral() 6
Cova d’Allará (Val d’Echo) 9
Els trets distintius de cada espècie venen explicats en l’apartat següent.
Característiques i distribució de les espècies
Stenophylax espanioli Schmid, 1957
Espècie descrita amb un holotipus mascle de l’avenc dels Pinyerets (Montmell, comarca del Baix Penedès) i un alotipus femella de la cova de les Tremolises (actualment anomenada espluga de les Tremolises) la qual pertany a la loca-
De cada una de les espècies del catàleg, es dona una descripció dels caràcters més distintius de les genitàlies masculina i femenina, acompanyada d’imatges amb indicacions gràfiques. Per a la distribució general de cada espècie, fora de l’àrea estudiada, s’han consultat les dades de PESI (2024) i de GBIF.org
litat de l’Espluga de Serra, que pertany al terme municipal de Tremp. Aquesta espècie és coneguda també de la cova de Mendikute (Tolosa). En tots els casos, els exemplars van ser recoŀlectats pel Dr. F. Español.
Explica l’autor que va descriure l’espècie que totes les femelles provenien de localitats diferents de les dels mascles. Malgrat això, comenta, «les fàcies dels dos sexes són tant semblants que no crec haver-me equivocat assignant-los el mateix nom».
Efectivament, l’assignació s’ha comprovat que és correcta amb la captura la mateixa data de 15 ♂ i 10 ♀ en una mateixa cavitat (Forat d l’Os, Os de Balaguer). Les femelles són distintes de les de l’altra espècie recollida en la mateixa localitat i la mateixa data, que és Stenophylax malatestus
Stenophylax espanioli es distribueix per la península Ibèrica i el marge mediterrani africà (Salavert et al. 2011). Les localitats del catàleg abasten l’Aragó (límit nord en una cavitat de Fanlo) i Catalunya (límit més al sud a la Vall d’Albaida, Cova de la Sarsa). És citada de Córdoba, Granada i Jaén. (Salavert et al. 2011); també de Terol (Martinez et al. 2018) i de Guadalajara (González et al. 1992). A la coŀlecció del Museu n’hi ha 31 localitats (Taula 1).
Mascle
Es distingeix pels apèndixs superiors marcadament còncaus (Fig, 2: 1) i per tenir apèndixs intermedis divergents i disposats horitzontalment a l’àpex (Fig. 2: 2).
Femella
A la figura 3 s’anomenen les peces que conformen l’estructura externa de l’aparell genital femení del grup Stenophylax.
A S. espanioli l’escata dorsal en visió caudal és estreta i prominent (Fig. 3: 2), i lateralment s’uneix per sota amb l’escata ventral (Fig. 3: 4). Els lòbuls laterals (Fig. 3: 3) són poc prominents i sovint presenten una petita protuberància superior (no és apreciable en l’exemplar representat aquí).
Stenophylax fissus McL
És de distribució principalment mediterrània, però arriba fins a Suïssa i Alemanya. Schmid (1957) la cita també d’Algèria. Citada en cavitats de Granada i de Jaén (Salavert et al 2011). Les localitats del catàleg procedeixen del nord i l’est peninsular. A la coŀlecció del Museu hi ha 42 localitats (Taula 1) d’aquesta espècie.
Mascle
Els apèndixs intermedis presenten l’extrem apical agut, són amples i girats cap a l’exterior (Fig. 4: 1). La part lliure dels apèndixs inferiors té una forma de bec subagut (Fig. 4: 2)
Femella
En visió ventral, l’escata dorsal té un perfil còncau (Fig. 5: 1) i els lòbuls laterals són molt poc prominents (Fig. 5: 2).
Stenophylax malatestus Schmid, 1957
Espècie descrita a Algèria central, amb paratipus femella procedents d’Anglesola (comarca de l’Urgell). Citada a Itàlia i a
l’illa de Sardenya. Les localitats ibèriques procedeixen de Cantàbria, Euskadi, Pirineus i serralades mediterrànies. A la coŀlecció del Museu hi ha 30 localitats d’aquesta espècie (Taula 1)
Mascle
Apèndixs intermedis amb la carena lateral superior dentada (Fig. 6: 1), i l’àpex còncau (Fig. 6: 2)
Femella
Els lòbuls laterals (Fig. 7: 1) són llargs (visió lateral), i amples a la base (Fig. 7: 1a) (visió ventral). La part ventral del IX segment és prominent, l’espai entre els segments és esclerotitzada (Fig. 7: 2a) i els dos lòbuls són contigus (Fig. 7: 2).
Stenophylax mitis McL., 1875
Espècie de distribució centreeuropea i mediterrània. La majoria de localitats del catàleg procedeixen del Prepirineu i del Pirineu axial. La resta són de les serralades catalanes, i tres són d’Euskadi. A la coŀlecció del Museu hi ha 29 localitats d’aquesta espècie (Taula 1) És l’espècie més aïllada del gènere, per la forma de les peces de l’armadura genital en ambdós sexes.
Mascle
Apèndixs superiors estrets, més del doble de llargs que d’amples (Fig. 8: 1); els inferiors més amples al centre que a l’àpex (Fig. 8: 2)
Femella
Els lòbuls laterals són molt llargs, i en visió lateral són recorbats cap a avall i aguts a l’àpex (Fig. 9: 1).
Stenophylax mucronatus McL., 1880
Espècie distribuïda per la Mediterrània occidental i Centreeuropa. És la segona en nombre de localitats del catàleg, la majoria de les quals són del Prepirineu i de la serralada Prelitoral Catalana. També és present a Navarra i al País Valencià. També és la segona espècie en nombre de localitats a Andalusia (Salavert et al. 2011). A la coŀlecció del Museu hi ha 87 localitats d’aquesta espècie (Taula 1).
Morfologia genital
L’atribució dels exemplars d’aquesta espècie a S mucronatus (McLachlan 1880) o bé a S crossotus (McLachlan 1884) va portar dubtes a l’autor en els anteriors treballs (Filbà Esquerra 1974a, 1974b, 1977 i 1986), de manera que, si bé en el primer estudi els va assignar tots a S mucronatus, en els següents els va atribuir tots a S. crossotus. Aquesta rectificació va ser motivada pel fet que S mucronatus es considerava una espècie no coneguda al sud dels Pirineus
Respecte a les dues espècies, el mateix Schmid apunta que «S crossotus es distingeix de S mucronatus sobretot per la seva coloració rogenca, per les llargues sedes del nervi costal de les ales anteriors i per una lleugera diferència en la forma dels apèndixs genitals».
També Frochot i Fotius-Jaboulet (1962) esmenta que «poden acostar encara més les dues espècies posant en evidència els nombrosos punts en comú tant de les ales com de les peces genitals de mascles i de femelles».
Més endavant, Malicky (2014) reconeix la dificultat per atribuir uns caràcters diferenciadors entre les dues espècies donada la variabilitat de l’extrem dels apèndixs inferiors i l’aparentment manca de divisió geogràfica marcada entre les formes. En conclusió, deixa les dues espècies com a pràcticament indistingibles i n’estableix la sinonímia.
D’altra banda, l’examen d’uns exemplars de la Grotte de Lameza (Seix, Arieja, França) – coŀlecció MCNB- no ha aportat criteris que permetin distingir les dues espècies citades. Per tot el que s’ha exposat, es considera com a espècie vàlida Stenophylax mucronatus McL. 1880.
Mascle. Stenophylax mucronatus es distingeix de les seves espècies pròximes S. espanioli, S. vibex i S. permistus per la major amplada de la part apical dels apèndixs inferiors, que tenen l’extrem engruixit i més corbat (Fig. 10: 1). Els apèndixs intermedis són prims, estan dirigits horitzontalment cap enfora i tenen l’àpex agut i en forma de ganxo (Fig. 10: 2). Són més o menys divergents segons la retracció de les peces genitals.
Femella
Els lòbuls laterals són estrets i molt llargs (Fig. 11: 1), i l’escata ventral és molt ampla (Fig. 11: 2).
Stenphylax nycterobius McL., 1875
D’àmplia distribució europea, és l’espècie del gènere Stenophylax que concentra més localitats al Pirineu axial i Prepirineu (un total de 32). Citada a l’avenc GESM de Tolox, Málaga (Salavert et al. 2011), una cavitat situada a una altitud de 1.710 m. Hi ha també un bon nombre de citacions de Sierra Nevada (Andalusia), que corresponen a exemplars epigeus en estat adult els quals es troben a altituds que van dels 1.400 als 2.420 m. El total de localitats de la coŀlecció del Museu és de 44 (Taula 1).
Mascle
En visió ventral, els apèndixs intermedis (Fig. 12: 1) són paraŀlels i molt pròxims a la seva part terminal. Els apèndixs inferiors (Fig. 12: 2) són acabats en forma de bec rom.
Femella
La part dorsal del IX segment abdominal forma uns angles laterals molt prominents (Fig. 13: 1). Els lòbuls laterals (Fig. 13: 2) són molt grossos i de forma triangular.
Stenophylax permistus McL., 1875
És l’espècie amb una més ample distribució europea. Les localitats ibèriques del catàleg se situen al nord (Aragó, Navarra, Euskadi, Castella-Lleó i Astúries, i la més meridional correspon a una cavitat de la serra de l’Obac [Catalunya]). La citació de l’espècie a Sierra Nevada (M. Sáinz Bariáin, 2014) correspon a exemplars en estat larvari. Caldria la captura d’exemplars adults per confirmar que hi és present. A la coŀlecció del Museu hi ha 13 localitats d’aquesta espècie (Taula 1).
Mascle
La forma dels apèndixs superiors (Fig. 14: 1) és pròxima a la de S mucronatus, S. espanioli i S. vibex, però es diferencia de la de S. permistus per la seva disposició en forma de tenalles.
Femella
Es pot distingir pels lòbuls laterals del X segment abdominal, que són triangulars i acabats en pic (Fig. 15: 1). L’escata dorsal és sinuosa (Fig. 15: 2).
Stenophylax sequax McL., 1875
Espècie d’àmplia distribució europea. Les localitats del catàleg procedeixen d’Aragó, Catalunya, Euskadi, Navarra i Burgos. N’hi ha 30, 18 de les quals són dels Pirineus (Taula 1).
Mascle
Es caracteritza per la vora apical del VIII segment abdominal densament recoberta d’espínules (Fig. 16: 1). Un altre caràcter típic és la terminació dels apèndixs inferiors en un llarg esperó recorbat amunt (Fig. 16: 2).
Femella
Pròxima a S. nycterobius. La part dorsal del IX segment abdominal forma uns angles laterals prominents (Fig. 17: 1). Els lòbuls laterals (Fig. 17: 2) són triangulars i lleugerament arrodonits a l’àpex.
Stenophylax testaceus Gmelin, 1788
Distribuïda pel centre i sud d’Europa. Hi ha un total de 33 localitats a la coŀlecció del Museu, la majoria procedents dels Pirineus (Taula 1). Citada a Granada i a Lleó (González et al. 1992).
Mascle
Es distingeix per la presència d’una dent a la part basal dels apèndixs superiors (Fig. 128: 1).
Femella
Lòbuls laterals prominents i aguts (Fig. 19: 1). L’escata dorsal és molt reduïda (no està representada al dibuix), i l’escata ventral és llarga (Fig. 19: 2).
Stenophylax vibex Curt., 1834
Distribuïda pel centre i l’est d’Europa i també per la Gran Bretanya. Set localitats són dels Pirineus, una del Sistema Ibèric (Sierra de Albarracín) i dues del massís català del Cabrerès. Schmid (1957) la indica a la Sierra de Guadarrama. També és citada en diverses localitats del centre i nord de la península Ibèrica (González et al. 1992).
D’altra banda, és citada en una cavitat de Jaén (Salavert et al., 2011). És l’espècie amb menys registres del catàleg. Només hi té 12 localitats (Taula 1).
Mascle
Pròxim a S.mucronatus, S. espanioli i S permistus. Els apèndixs superiors en la visió lateral són molt prominents (Fig. 20: 1). En la visió ventral es pot distingir en la seva vora inferior interna una petita ala (Fig. 20: 1a).
En els apèndixs inferiors, el seu extrem apical intern no és recte com en les altres tres espècies pròximes, sinó que és lleugerament corbat endins i té l’extrem arrodonit (Fig. 20: 2).
Femella
Els lòbuls laterals són robustos (Fig. 21: 2) i, en vista ven-
tral, són molt amples a la base i presenten al costat intern una petita dent (Fig. 21: 1).
Mesophylax aspersus Ramb., 1842
Espècie coneguda de la Bètica, zona mediterrània, Pirineus, serralada Cantàbrica, Eivissa i Mallorca. El focus de distribució de M. aspersus és la regió mediterrània i és l’espècie més freqüent en l’àrea estudiada. A la coŀlecció del Museu compta amb 320 localitats (Taula 1).
Es pot reconèixer per la coloració bruna de les ales (Figura 1b), que a la resta d’espècies del grup són de coloració ocrosa. S’iŀlustra en les figures 22a i 22b la genitàlia del ♂ en visions lateral i ventral
S’iŀlustra en les figures 23a i 23b la genitàlia de la ♀ en visions lateral i ventral
Conclusions
La coŀlecció de tricòpters subtroglòfils del MCNB s’ha format per l’activitat de la pròpia institució així com pel treball de recoŀlecció dels bioespeleòlegs. Les localitats procedeixen en un 85 % de Catalunya i Aragó, i en el conjunt d’aquestes dues regions hi són presents les 11 espècies de l’àrea
La serralada Cantàbrica i els sistemes muntanyosos mediterranis alberguen entre 4 i 6 espècies del grup. En la resta de regions, on les captures són més escasses, el nombre d’espècies localitzades és entre 1 i 3.
Mesophylax aspersus és l’espècie més freqüent en l’àrea iberobalear. Es troba en el 47% de les localitats. El predomini d’aquesta espècie a la zona mediterrània peninsular podria indicar que aquesta regió és el seu nucli de radiació.
Stenophylax mucronatus és la segona espècie en nombre de localitats, mentre que Stenophylax vibex i Stenophylax permistus són les espècies amb menys registres.
La resta de tàxons del grup tenen una presència més o menys uniforme, donat que tenen un nombre de localitats similar.
Agraïments
Estic molt agraït al Dr. Hans Malicky (Lunz am See, Àustria) per les seves orientacions en la qüestió de la nomenclatura del grup Stenophylax. Als autors de les imatges, Agustí Meseguer i Jorge Mederos. A Neus Brañas, pel seu treball en la depuració de la base de dades, i a Glòria Masó i Berta Caballero, pel seu inestimable ajut.
Bibliografia
Bellés, X. 1987. Fauna cavernícola i intersticial de la Península Ibèrica i les Illes Balears CSIC-Editorial Moll. Palma de Mallorca. 208 p.
Botosaneanu, L. 1992. Trichoptera of the Levant (Fauna Palestina, Insecta VI). Journal of the North American Benthological Society, 13 (4): 1-293.
Bouvet, I. 1971. La diapause des Trichoptères cavernícoles. Bulletin de la Societé. Zoologique de France, 96 (4): 375-384.
Bouvet, I. & Ginet, R., 1969. Données biològiquesxt et biogeographiques sur le “groupe de Stenophylax” cavernicoles de France. Bulletin de la Societé Linnéenne de Lyon, 10: 334-349.
Filbà Esquerra L. 1974a. Contribución al conocimiento de los Tricópteros cavernícolas de Catalunya. IV Simposium de Bio Espeleologia. Barcelona. 1: 20-35. Filbà Esquerra L. 1974b. Nuevas estaciones ibéricas de tricópteros cavernícoles. Actas del III Congreso Nacional de Espeleologia. Madrid. (1): 1-5.
Filbà Esquerra L. 1977. Noves localitzacions ibèriques de Tricòpters cavernícoles. Comunicacions del 6è. Simposium d’Espeleologia. Terrassa. (1): 131-134
Filbà Esquerra L. 1986. Tricòpters recoŀlectats al Montseny. Jornades de recerca naturalista al Montseny: 61-63. Diputació de Barcelona.
Frochot, B. & Fotius-Jaboulet, M. C., 1964. Revision de Stenophylax mucronatus McL. avec description de la femella adulte et de la larve. Travaux du Laboratoire de zoologie et de la Station aquicole Grimaldi de la Faculté des sciences de Dijon, 57: 1-28
GBIF.org. 2024. https://www.gbif.org/occurrence/download/0050224231120084113126
González, M. A., Terra, L. S. W, Garcia de Jalon, D., Cobo, F. 1992 Lista faunistica y geografica de los Tricópteros (Trichoptera) de la Peninsula Ibérica e Islas Baleares. Asociación Española de Limnologia 11.
González González, M. A., Martínez Menéndez, J. 2011. Checklist of the caddisflies of the Iberian Peninsula and Balearic Islands. Zoosymposia 5: 115-135.
Jeannel, R. 1926. Faune cavernicole de la France, avec une étude des conditions d’existence dans le domaine souterrain. Encyclopédie entomològiques, 7. Lechevalier, París. 334 p.
Malicky, H. 1993. Comments on the book “Trichoptera of Levant” by L. Botosaneanu. Braueria 20: 24-25
Malicky, H. 2014. Neue Beiträge zur Kenntnis asiatischer und mediterraner Köcherfliegen (Trichoptera). Braueria, 41: 43-50. Martinez, J., Martin, L., González, M. A. 2018. Tricópteros (Insecta:Trichoptera) de la provincia de Teruel (Aragon, España). Revista Teruel. 95-96: 373-401.
PESI. 2024. Pan-European Species directories Infrastructure Sáinz-Bariáin, M. 2014. Diversidad, estrategias vitales y filogeografía de especies sensibles al cambio climático: Tricópteros en el Parque Nacional de Sierra Nevada. Tesis doctoral. Universidad de Granada. 401 p. [http://hdl.handle. net/10481/34652]
Salavert, V., Zamora-Muñoz, C. & Tinaut, A. 2011. Distribución de los tricópteros troglófilos (Trichoptera, Limnephilidae) en cuevas andaluzas (Andalucia, España). Boletin de la Asociación Española de Entomologia, 35 (3-4): 325-344.
Salavert, V. 2012. Estrategias vitales e implicaciones evolutivas de Tricópteros de cursos de agua temporales. Tesi doctoral. Universitat de Granada. 1-265.
Schmid, F. 1955. Contributions à l’étude des Limnophilidae (Trichoptera). Mitteilungen der Schweizerischen Entomologischen, 28 Beiheft. 245 p.
Schmid, F. 1957. Les genres Stenophylax Kol., Micropterna St. et Mesophylax McL. (Tricopt. Limnoph.). Trabajos Museo de Zoologia, Nueva Serie. Zoologica., 2 (2): 1-52

Figura 2 Part distal de l’abdomen de Stenophylax espanioli Sch. ♂: a) visió lateral, b) visió caudal en què s’assenyalen els apèndix superiors (1) i els apèndix intermedis (2). [Fig. 1b: fotografia d’A. Meseguer].
3 Part distal de l’abdomen de Stenophylax espanioli Sch. ♀: a) visió lateral, b) visió caudal. [Fotografies de Jorge Mederos].
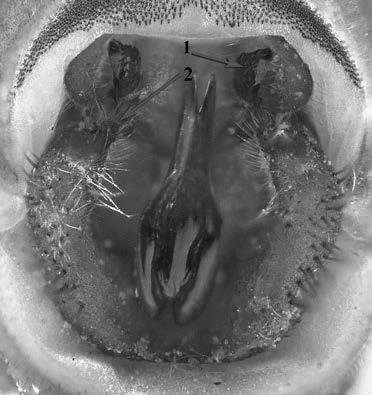
Figura 4. Part distal de l’abdomen de Stenophylax fissus McL. ♂ en visió caudal. S’hi assenyalen els apèndixs intermedis (1) i els apèndix inferiors (2). [Fotografia d’A. Meseguer].
Figura 5. Part distal de l’abdomen de Stenophylax fissus McL. ♀: a) visió lateral i b) vista ventro-caudal. S’hi assenyalen l’escata dorsal (1) i els lòbuls laterals (2). [Fotografia de Jorge Mederos]
Figura 6. Part distal de l’abdomen de Stenophylax malatestus Sch. ♂ en visió caudal. S’hi assenyalen la carena lateral (1) i l’àpex dels apèndix intermedis (2). [Fotografia d’A. Meseguer]
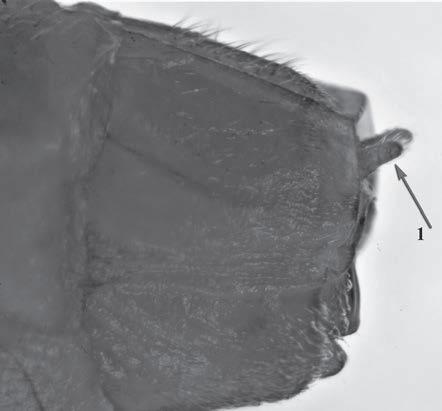
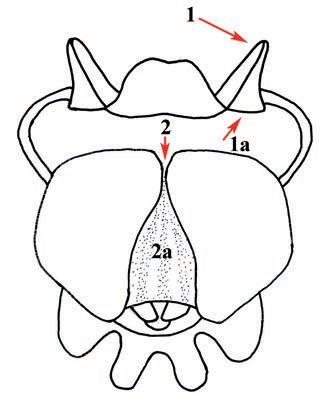
Figura 7. Part distal de l’abdomen de Stenophylax malatestus Sch. ♀: a) visió lateral i b) visió caudal. S’hi assenyalen els lòbuls laterals prominents (1) i amples a la base (1a), els lòbuls ventrals contigus a la part superior (2) i l’espai esclerotitzat que hi ha entre els segments (2a)
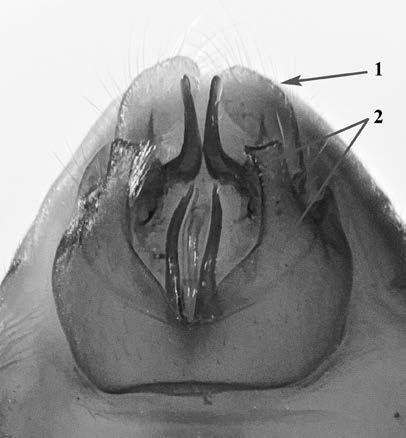
8. Part distal de l’abdomen de Stenophylax mitis McL. en visió ventro-caudal. S’hi assenyalen els apèndixs superiors (1) i els apèndixs inferiors (2). [Fotografia de Jorge Mederos].

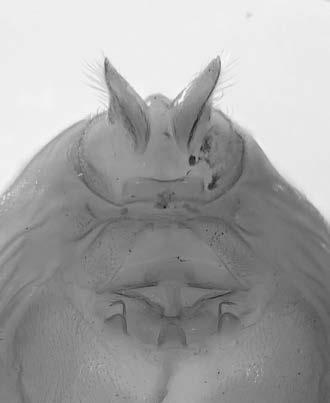
9. Part distal de l’abdomen de Stenophylax mitis McL. ♀: (a) visió lateral i (b) visió ventral. S’hi assenyalen els lòbuls laterals (1). [Fig. 9b: fotografia de Jorge Mederos].
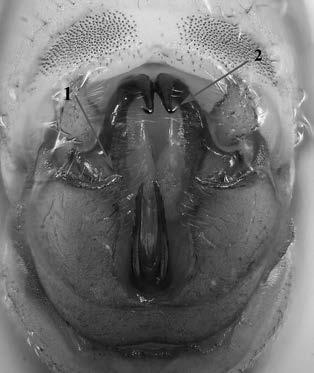
Figura 10. Part distal de l’abdomen de Stenophylax mucronatus McL. ♂ en visió caudal. S’hi assenyalen la part apical dels apèndixs inferiors (1) i la part apical dels apèndixs intermedis (2). [Fotografia d’A. Meseguer].
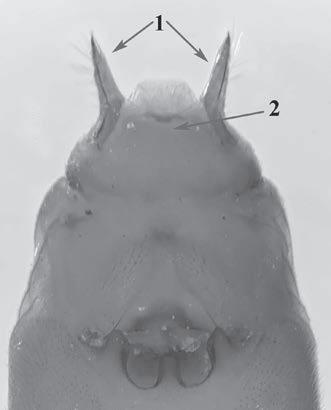
11. Part distal de l’abdomen de Stenophylax mucronatus McL. ♀ en visió ventral. S’hi assenyalen els lòbuls laterals (1) i l’escata ventral (2). [Fotografia de Jorge Mederos].
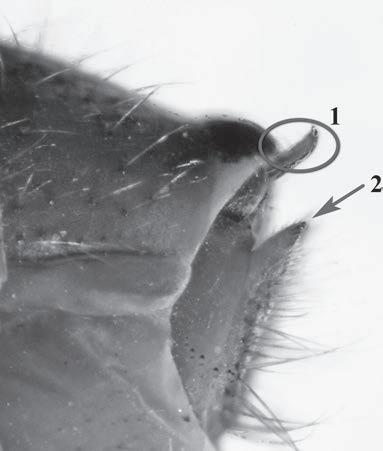

12. Part distal de l’abdomen de Stenophylax nycterobius McL. ♂: a) visió lateral i b) visió ventro-caudal. S’hi assenyalen els apèndixs intermedis (1) i els apèndixs inferiors (2). [Fig. 12b: fotografia d’A. Meseguer].

13. Part distal de l’abdomen de Stenophylax nycterobius McL.
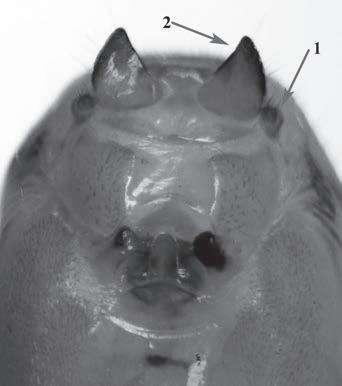
lateral i b) visió ventral. S’hi assenyalen els angles laterals del IX segment (1) i els lòbuls laterals (2)


Figura 14. Part distal de l’abdomen de Stenophylax permistus McL.: a) visió lateral, b) visió caudal. S’hi assenyalen els apèndixs superiors (1). [Fig. 14b: fotografia d’A. Meseguer].


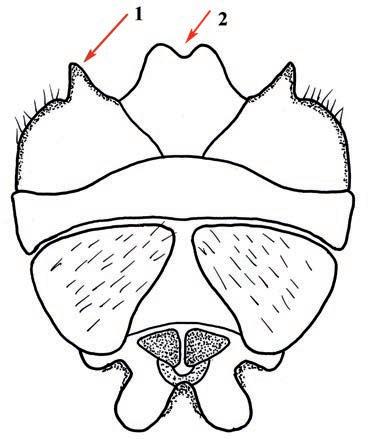
15. Stenophylax permistus McL. ♀: a) visió lateral, b-c) visió ventral. S’hi assenyalen els lòbuls laterals del X segment (1) i l’escata dorsal (2). [Fig. 15b: fotografia de Jorge Mederos]
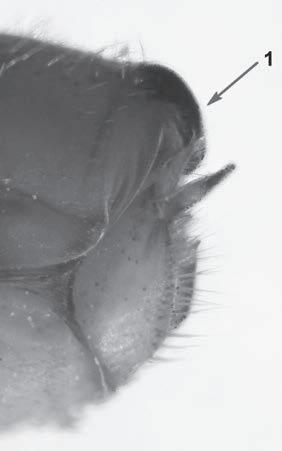

Figura 16. Part distal de l’abdomen de Stenophylax sequax McL. ♂: a) visió lateral i b) visió caudal. S’hi assenyalen la vora apical del VIII segment (1) i l’esperó dels apèndixs inferiors (2). [Fig. 16b: fotografia d’A. Meseguer].

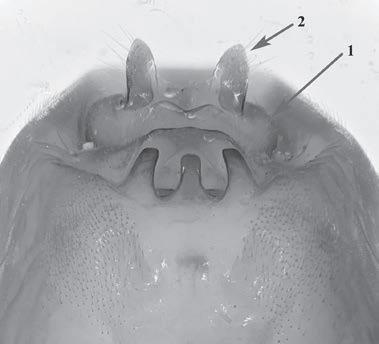
Figura 17. Part distal de l’abdomen de Stenophylax sequax McL. ♀: a) visió lateral i b) visió caudal. S’hi assenyalen la part dorsal del IX segment (1) i els lòbuls laterals (2). [Fig. 17b: fotografia de Jorge Mederos].
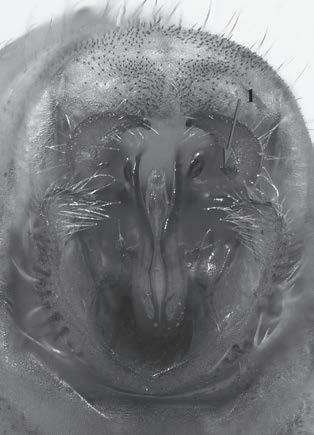
Figura 18. Part distal de l’abdomen de Stenophylax testaceus Gmelin. ♂ en visió caudal. S’hi assenyala la dent dels apèndixs superiors (1). [Fotografia d’A. Meseguer].
(2), 2024
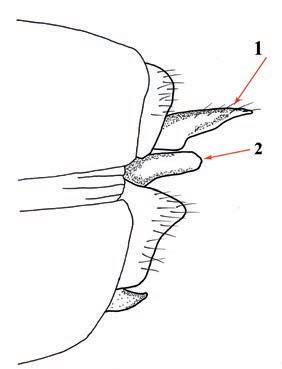

Figura 19. Stenophylax testaceus Gmelin. ♀: a) visió lateral i b) visió ventral. S’hi assenyalen els lòbuls laterals (1) i l’escata ventral (2). [Fig. 18b: fotografia de Jorge Mederos].

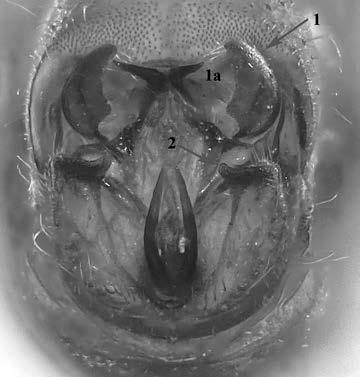
Figura 20. Part distal de l’abdomen de Stenophylax vibex Curt. ♂: a) visió lateral, i b) visió caudal. S’hi assenyalen els apèndixs superiors (1), la petita ala a la seva vora interna (1a) i l’extrem apical dels apèndixs inferiors (2). [Fig. 20b: fotografia d’A. Meseguer].
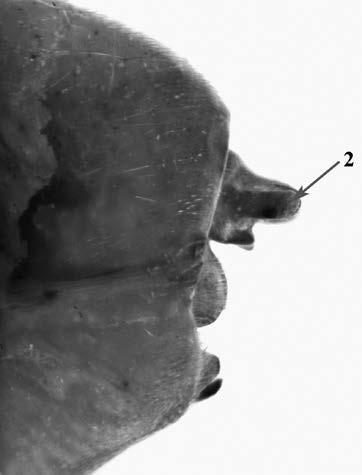
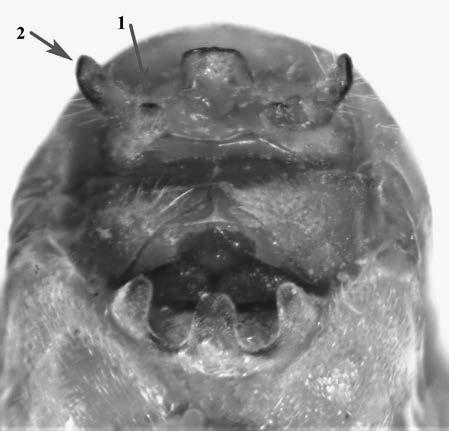
Figura 21. Part distal de l’abdomen de Stenophylax vibex Curt. ♀: a) visió lateral en la qual s’assenyalen els lòbuls laterals (2) i b) visió caudal en la qual s’assenyala una dent de la part inferior dels lòbuls laterals (1).

Anexa.
Regió Gran unitat Localitat
Andalusia
Penibètica
Subbètica
Aragó
Pirineu axial
Gènere Espècie ♂ ♀ Recoŀlector
Cova Benaocaz Benaocaz 26/03/1986 1000 Stenophylax mucronatus 0 1 O. Escolà
Sima de los Derrubios Carratraca 08/02/1970 900 Mesophylax aspersus 1 0 Gil
Sima del Republicano Villaluenga del Rosario 31/07/1970 800 Stenophylax mucronatus 1 0 C. Ribera
Cueva Nacimiento del Jaen 15/09/1981 1300 Stenophylax nycterobius 0 2 A. Lista Guadalquivir
Av. Del Dinar Val d’Echo 12/08/1980 2000 Stenophylax nycterobius 1 4 O. Escolà
Av. Del Dinar Val d’Echo 12/08/1980 2000 Stenophylax testaceus 2 2 O. Escolà
Avenc BA-2 (Baladrás) Biescas 01/07/1985 1800 Stenophylax nycterobius 2 2 Inglés, Picanyol
Avenc BA-2 (Baladrás) Biescas 07/08/1985 1800 Stenophylax testaceus 0 1 Inglés, Picanyol
Avenc BA-2 (Baladrus) Biescas 01/07/1985 1800 Stenophylax testaceus 0 1 Inglés, Picanyol
Avenc C-61 Puértolas 06/08/1991 1200 Stenophylax mitis 1 0 O. Escolà
Avenc Camí Ara (A-31) Torla 04/10/1986 1100 Stenophylax testaceus 1 0 O. Escolà
Avenc de Sevil Aínsa-Sobrarbe 18/05/1970 900 Mesophylax aspersus 0 1 Ll. Auroux
Avenc del Torrent. Cema Blanca Val d’Echo 08/10/1988 2000 Stenophylax nycterobius 1 0 O. Escolà
Avenc E.R.E. Ansó 01/09/1985 2300 Stenophylax testaceus 1 0 O. Escolà
Avenc E.R.E. Ansó 07/07/1990 2300 Stenophylax testaceus 0 1 O. Escolà
Avenc E.R.E. Ansó 09/07/1990 2200 Stenophylax testaceus 0 1 O. Escolà
Avenc E-9 (Peña Telera) Biescas 14/07/1985 Stenophylax nycterobius 1 0 O. Escolà
Avenc F0-4 Ansó 04/08/1993 2350 Stenophylax testaceus 1 0 Bosch, Escolà
Avenc F-143 Ansó 01/08/1995 Stenophylax testaceus 3 4 O. Escolà
Avenc G-3 (Gamueta) Ansó 22/08/1982 2000 Stenophylax mitis 2 0 O. Escolà
Avenc num. 5 Escuain 1550 Stenophylax mitis 1 0 R. Canela
Avenc R-3. La Reclusa Ansó 11/08/1980 2300 Stenophylax nycterobius 1 1 O. Escolà
Avenc R-3. La Reclusa Ansó 08/06/1981 2300 Stenophylax nycterobius 1 0 O. Escolà
C-74 Tella 06/08/2007 940 Stenophylax mitis 0 1 R. Canela
Cova AS-1 (Boca del Infierno) Val d’Echo 03/05/1987 1000 Mesophylax aspersus 2 0 O. Escolà
Cova AS-1 (Boca del Infierno) Val d’Echo 01/05/1988 1000 Mesophylax aspersus 1 1 O. Escolà
Cova Barber Tella 15/08/1971 1550 Stenophylax mitis 0 1 R. Canela
Cova C-46 Escuain 15/08/1995 1200 Stenophylax nycterobius 1 0 Alfambre
Cova F-168 Val d’Echo 05/08/1998 900 Stenophylax nycterobius 0 1 O. Escolà
Cova Fuendeselva Val d’Echo 23/07/1991 1000 Mesophylax aspersus 4 3 O. Escolà
Cova Mora Jaca 27/06/1982 900 Mesophylax aspersus 1 0 O. Escolà
Cova Mora Jaca 11/07/1983 900 Mesophylax aspersus 3 1 O. Escolà
Cova Mora Jaca 11/07/1983 900 Stenophylax mucronatus 1 0 O. Escolà
Cueva Allará Val d’Echo 03/07/1969 1600 Mesophylax aspersus 2 0 O. Escolà
Cueva Allará
Cueva Allará
Cueva Allará
Cueva Allará
Cueva Allará
Cueva Allará
Cueva Allará
Cueva Allará
Cueva Allará
Cueva Allará
Cueva Allará
Cueva Allará
Cueva Allará
Val d’Echo 12/08/1980 2000 Mesophylax aspersus 0 2 O. Escolà
Val d’Echo 22/07/1991 1600 Mesophylax aspersus 1 1 O. Escolà
Val d’Echo 03/07/1969 1600 Stenophylax fissus 4 2 O. Escolà
Val d’Echo 03/07/1969 1600 Stenophylax mitis 2 0 O. Escolà
Val d’Echo 12/08/1980 1600 Stenophylax mitis 1 1 O. Escolà
Val d’Echo 22/07/1991 1600 Stenophylax mitis 0 1 O. Escolà
Val d’Echo 12/08/1980 1600 Stenophylax mucronatus 0 3 O. Escolà
Val d’Echo 03/07/1969 1600 Stenophylax nycterobius 0 3 O. Escolà
Val d’Echo 12/08/1980 1600 Stenophylax nycterobius 21 23 O. Escolà
Val d’Echo 08/06/1981 1600 Stenophylax nycterobius 0 2 O. Escolà
Val d’Echo 22/07/1991 1600 Stenophylax nycterobius 7 9 O. Escolà
Val d’Echo 03/07/1969 1600 Stenophylax permistus 1 0 O. Escolà
Val d’Echo 22/07/1991 1600 Stenophylax sequax 0 3 O. Escolà
Cueva Allará Val d’Echo 03/07/1969 1600 Stenophylax testaceus 1 0 O. Escolà
Cueva Allará Val d’Echo 22/07/1991 1600 Stenophylax testaceus 1 0 O. Escolà
Cueva Allará Val d’Echo 03/07/1969 1600 Stenophylax vibex 2 1 O. Escolà
Cueva Allará Val d’Echo 08/06/1981 1600 Stenophylax vibex 1 3 O. Escolà
Cueva Allará Val d’Echo 22/07/1991 1600 Stenophylax vibex 0 1 O. Escolà
Cueva Bujaruelo Bujaruelo 18/09/1970 2000 Mesophylax aspersus 1 0 GIEM
Cueva de los Arnales Tella-Sin 24/08/2021 1155 Stenophylax sequax 2 0 A. Messeguer
Cueva de los Gloces Fanlo 06/06/1981 1500 Stenophylax mitis 1 0 J. Comas
Cueva del Beral Ansó 15/08/1971 2000 Stenophylax mitis 1 3 O. Escolà
Cueva del Beral Ansó 15/08/1971 2000 Stenophylax nycterobius 1 0 O. Escolà
Cueva del Beral Ansó 15/08/1971 2000 Stenophylax vibex 4 3 O. Escolà
Cueva del Ibon de Izagra Aísa 13/07/1980 1800 Stenophylax nycterobius 3 4 O. Escolà
Cueva del Ibon de Izagra Aísa 03/08/1980 1800 Stenophylax nycterobius 9 10 O. Escolà
Cueva del Ibon de Izagra Aísa 03/08/1980 1800 Stenophylax permistus 1 1 O. Escolà
Cueva del Ibon de Izagra Aísa 03/08/1980 1800 Stenophylax testaceus 3 1 O. Escolà
Cueva del Molino de Aso Fanlo 16/09/1981 940 Mesophylax aspersus 0 2 O. Escolà
Cueva del Molino de Aso Fanlo 01/09/1973 940 Stenophylax nycterobius 1 0 T. Cuñé
Cueva Grande de Pardina Fanlo 06/07/1985 900 Mesophylax aspersus 1 0 O. Escolà
Cueva Grande de Pardina Fanlo
Cueva Inferior de las Gloces Fanlo
06/07/1985 900 Stenophylax mitis 0 1 O. Escolà
13/07/1983 1240 Stenophylax espanioli 0 1 O. Escolà
Cueva Inferior de las Gloces Fanlo 13/07/1983 1240 Stenophylax sequax 0 1 O. Escolà
Cueva Lomenera Fanlo
11/07/1985 1600 Stenophylax mitis 2 7 O. Escolà
Cueva Lomenera Fanlo 11/07/1985 1600 Stenophylax nycterobius 2 2 O. Escolà
Cueva Otal Torla 20/09/1986 2200 Stenophylax testaceus 1 0 O. Escolà
Cueva Superior de Castillo Viejo Val d’Echo 03/06/1989 1200 Mesophylax aspersus 1 0 O. Escolà
Asturies
Prepirineu
Cueva Vieja de Villanúa Villanua 12/09/1965 1000 Mesophylax aspersus 1 0 Ll. Auroux
Cueva Vieja de Villanúa Villanúa 12/09/1966 1000 Stenophylax fissus 1 0 Ll. Auroux
Cueva Villacampa (Secorun) Sabiñánigo 06/10/1986 1050 Mesophylax aspersus 1 0 O. Escolà
Cuevas de Tendeñera 15/10/2007 2300 Stenophylax nycterobius 1 1 J. Ferreres
Cv. Santa Elena Biescas 14/08/1980 1198 Stenophylax nycterobius 1 4 O. Escolà
Cv. Santa Elena Biescas 14/08/1980 1198 Stenophylax sequax 2 0 O. Escolà
F 0-21 Ansó 05/08/1993 Stenophylax testaceus 0 1 O. Escolà
Sima Alano Espelunga Ansó 04/08/1992 1800 Stenophylax sequax 1 0 O. Escolà
Sima Cema Blanca Val d’Echo 17/09/1988 2000 Stenophylax nycterobius 0 1 O. Escolà
Sima Cema Blanca Val d’Echo 08/10/1988 2000 Stenophylax testaceus 1 0 O. Escolà
Sima del Ibon de la Reclusa Val d’Echo 08/08/1964 1700 Stenophylax mitis 1 0 O. Escolà
Sima del Ibon de la Reclusa Val d’Echo 10/08/1980 1700 Stenophylax mitis 9 20 O. Escolà
Sistema Sabadell Hoz de Jaca 10/07/1982 2175 Stenophylax testaceus 1 0 O. Escolà
Tub de captació Canfranc 15/09/1981 1000 Mesophylax aspersus 3 5 O. Escolà
Tub de captació Canfranc 15/09/1981 1000 Stenophylax mitis 2 4 O. Escolà
Tub de captació Canfranc 15/09/1981 1000 Stenophylax mucronatus 0 1 O. Escolà
Avenc de Can Pallàs (Betesa) Areny de Noguera 25/09/1982 1200 Stenophylax nycterobius 1
Avenc de la Sia Sta. Marguerida Purroi de la Solana 13/12/1980 875 Mesophylax aspersus
Avenc de la Sia Sta. Marguerida Purroi de la Solana 26/04/1981 875 Mesophylax aspersus
Avenc de la Sia Sta. Marguerida Purroi de la Solana 26/04/1981 875 Stenophylax fissus 4
Avenc de Sant Salvador de Pinyana Castillonroi 20/01/1971 700 Mesophylax aspersus 3
Avenc de Sant Salvador de Pinyana Castillonroi 25/08/1979 700 Mesophylax aspersus
Avenc de Sant Salvador de Pinyana Castillonroi 20/01/1979
Avenc de Sant Salvador de Pinyana Castillonroi
Avenc del Toscar Foradad del Toscar 30/10/1976 1000 Mesophylax aspersus
Avenc Tossal de Felis Purroi de la Solana 26/04/1981 875 Stenophylax malatestus
Caricau del Puço Bonansa 10/06/1973 1300 Stenophylax mucronatus 1
Cova Celleret del Ramon Beranui 03/03/1997 1200 Mesophylax aspersus 0 2 O. Escolà
Cova del Salaber Peralta de Calasanz 25/03/1967 700 Mesophylax aspersus 0 1 sense dades
Cova Ortiguers El Turbon 14/08/1979 Stenophylax nycterobius 0 1 J. Santamaria
Cova Ovis Areny de Noguera (Betesa) 11/07/1976 1150 Stenophylax sequax 5 2 O. Escolà
Cueva de la Higuera Peralta de Calasanz 25/03/1967 600 Mesophylax aspersus 1 0 O. Escolà
Cueva Hoyo de la Madre (Belsué). H-5 Nueno 1100 Stenophylax fissus 0 1 Ph. Chapman
Cueva Hueso Santo Pueyo de Araguás (Oncins) 11/08/1991 1200 Stenophylax espanioli 0 1 O. Escolà
Forat de l’Aire (o de les Guitarres) Baells (Sant Quilis) 30/11/1978 600 Mesophylax aspersus 8 1 J. Comas
Forat de l’Aire (o de les Guitarres) Baells (Sant Quilis) 21/01/1979 600 Mesophylax aspersus 9 0 O. Escolà
Forat de l’Aire (o de les Guitarres) Baells (Sant Quilis) 30/11/1980 700 Mesophylax aspersus 12 0 O. Escolà
Forat de l’Aire (o de les Guitarres) Baells (Sant Quilis) 26/04/1981 700 Mesophylax aspersus 0 1 O. Escolà
Forat de l’Aire (o de les Guitarres) Baells (Sant Quilis) 20/09/1981 700 Mesophylax aspersus 1 0 O. Escolà
Forat de l’Aire (o de les Guitarres) Baells (Sant Quilis) 25/09/1968 700 Stenophylax espanioli 1 1 I. Gonzàlez
Forat de l’Aire (o de les Guitarres) Baells (Sant Quilis) 25/09/1966 700 Stenophylax fissus 1 1 Ll. Auroux
Forat de l’Aire (o de les Guitarres) Baells (Sant Quilis) 25/09/1968 700 Stenophylax malatestus 6 4 I. Gonzàlez
Forat de l’Aire (o de les Guitarres) Baells (Sant Quilis) 26/04/1981 700 Stenophylax malatestus 10 2 O. Escolà
Forat de l’Aire (o de les Guitarres) Baells (Sant Quilis) 25/09/1968 700 Stenophylax mucronatus 1 0 I. Gonzàlez
Forat del Minguilli Camporrells 08/11/1970 500 Mesophylax aspersus 9 2 O. Escolà
Sia de Santa Ana Purroi de la Solana 12/10/1981 675 Stenophylax fissus 3 4 O. Escolà
Sima del Cubilillo Las Peñas de Riglos 20/03/1982 950 Stenophylax malatestus 0 1 O. Escolà
Sima Esteban Felipe Nueno (Belsué) 27/11/1988 1149 Mesophylax aspersus 2 0 O. Escolà
Sima Esteban Felipe Nueno (Belsué) 24/03/1978 1149 Stenophylax malatestus 1 0 S. Gimenez
Solencio de Bastaras Casbas de Huesca 13/09/1973 665 Mesophylax aspersus 3 5 Ll. Filbà
Solencio de Bastaras Casbas de Huesca 26/06/1977 665 Mesophylax aspersus 1 0 O. Escolà
Solencio de Bastaras Casbas de Huesca 665 Stenophylax mitis 0 1 sense dades
Solencio de Bastaras Casbas de Huesca 13/09/1973 665 Stenophylax mucronatus 1 1 Ll. Filbà
Sistema Ibèric
Solencio de Bastaras Casbas de Huesca 26/06/1977 665 Stenophylax mucronatus 1 0 O. Escolà
Cueva Ajo y Agua Calcena 10/09/1972 825 Stenophylax mucronatus 4 9 Ll. Filbà
Cueva de la Ubriga El Vallecillo 02/08/1971 1396 Mesophylax aspersus 1 0 Poderós, Ubach
Cueva de la Ubriga El Vallecillo 02/08/1971 1396 Stenophylax vibex 0 1 Poderós, Ubach
Cueva de las Baticambras Molinos 25/04/1964 600 Mesophylax aspersus 0 1 O. Escolà Cueva Juaco Aguaviva de Bergantes 22/08/1971 600 Stenophylax malatestus 0 1 X. Bellés
Serralada Cantàbrica
Cueva del Infierno San Esteban de Cuñaba 09/08/1974 1300 Stenophylax mitis 2 0 J. Comas
Cueva del Infierno San Esteban de Cuñaba 17/08/1978 1300 Stenophylax mitis 2 1 O. Escolà
Cueva del Infierno San Esteban de Cuñaba 09/08/1974 1300 Stenophylax nycterobius 1 0 J. Comas
Cueva del Rinoceronte Benia de Onís 30/07/1978 1100 Stenophylax fissus 1 0 O. Escolà
Cueva del Triumbio Cangas de Onís 04/08/1976 1000 Stenophylax permistus 1 0 J. Comas
Sima Budrio la Peña Cangas de Onís 11/08/1978 1150 Stenophylax fissus 1 2 O. Escolà
Sima Budrio la Peña Cangas de Onís 11/08/1978 1150 Stenophylax permistus 0 2 O. Escolà
Cantabria
Serralada Cantàbrica
Cueva Coquisera Matienzo 24/03/1974 500 Stenophylax fissus 1 0 O. Escolà
Cueva Cullalvera Ramales 25/03/1991 90 Mesophylax aspersus 1 0 O. Escolà
Cueva Villegas Alfoz de Lloredo (Cobreces) 31/03/1972 Stenophylax fissus 2 2 Ll. Filbà
Cv. Num. 4 Monte Castillo Puente Viesgo 15/03/1983 150 Stenophylax fissus 2 2 Ll. Auroux
Cv. Num. 4 Monte Castillo Puente Viesgo 15/03/1983 150 Stenophylax permistus 0 1 Ll. Auroux
Sima Cervajera num. 9 Guriezo 22/02/1976 552 Stenophylax fissus 1 0 Cantero
Torca Hoyon 2 Rasines 31/03/1991 150 Stenophylax fissus 0 1 O. Escolà
Castella-La Manxa
Sistema Central
Cueva del Tornero Checa 29/09/1971 1500 Stenophylax nycterobius 1 1 J. Pallisé
Castella-Lleó
Serralada Cantàbrica
Cueva Calderón Velilla 15/08/1971 1230 Stenophylax nycterobius 2 0 S.A.S. (Cueva del Triangulo)
Cueva Canales (Arreba) Valle de Manzanedo 24/07/1973
Cueva Canales (Arreba) Valle de Manzanedo 24/07/1973
650 Stenophylax permistus 1 0 O. Escolà
650 Stenophylax sequax 11 6 O. Escolà
Cueva de Agudin 2 Cardaño 15/08/1971 1250 Stenophylax nycterobius 1 0 S.A.S.
Cueva del Embudo Valle de Valdebezana 25/07/1973 850 Stenophylax fissus 0 2 T. Serra
Cueva del Embudo Valle de Valdebezana 25/07/1973 850 Stenophylax nycterobius 0 1 T. Serra
Cueva del Embudo Valle de Valdebezana 25/07/1973 900 Stenophylax permistus 0 1 T. Serra
Cueva Fuentemolinos Belorado (Puras de Villafranca) 02/11/2019 Mesophylax aspersus 1 0 sense dades
Cueva Honseca Velilla del rio Carrion 15/08/1971 1200 Stenophylax nycterobius 1 1 S.A.S.
Sima Alberich Cardaño 15/08/1971 1250 Mesophylax aspersus 3 4 S.A.S.
Sistema Ibèric
Cova del bosc Ucero 14/04/1979 1000 Mesophylax aspersus 1 1 Ll. Filbà
Cueva de la Culebra Tajueco 02/03/1974 1000 Mesophylax aspersus 1 0 sense dades
Cueva de la Galiana (Cueva Ucero 28/07/1973 900 Mesophylax aspersus
grande de Ucero)
Cueva de la Galiana (Cueva Ucero
grande de Ucero)
Cueva de la Galiana (Cueva Ucero 19/07/1979
Mesophylax aspersus 4 4
Filbà grande de Ucero)
Cueva de la Galiana (Cueva Ucero 19/07/1979
Stenophylax fissus 0 2 Ll. Filbà grande de Ucero)
Cueva de la Galiana (Cueva Ucero 28/07/1973
Stenophylax nycterobius 7 5 Ll. Filbà grande de Ucero)
Cueva de la Galiana (Cueva Ucero 19/07/1979
Stenophylax nycterobius 13 12 Ll. Filbà grande de Ucero)
Cueva de la Galiana (Cueva Ucero 28/07/1973
Stenophylax permistus 1 0 Ll. Filbà grande de Ucero)
Catalunya
Depressió Central Catalana
Avenc de Castellcir Castellcir 20/10/1979 790 Mesophylax aspersus 6 0 O. Escolà
Cova de la Pollosa Collsuspina 09/05/1965 833 Mesophylax aspersus 2 3 I. Gonzàlez
(Forat negre de Puigantic)
Cova de la Pollosa Moià 20/10/1979 833 Mesophylax aspersus 1 0 O. Escolà
(Forat negre de Puigantic)
Cova de la Pollosa Collsuspina 28/06/1981 833 Mesophylax aspersus 1 0 O. Escolà
(Forat negre de Puigantic)
Cova de la Pollosa Collsuspina 05/09/1982 833 Mesophylax aspersus 2 0 O. Escolà
(Forat negre de Puigantic)
Cova de la Pollosa Collsuspina 13/03/1983 833 Mesophylax aspersus 0 1 O. Escolà
(Forat negre de Puigantic)
Cova de la Pollosa Collsuspina 13/03/1983 833 Stenophylax malatestus 1 0 O. Escolà
(Forat negre de Puigantic)
Cova de la Pollosa Collsuspina 09/05/1965 833 Stenophylax mucronatus 5 0 I. Gonzàlez
(Forat negre de Puigantic)
Cova de la Pollosa Collsuspina 01/05/1983 833 Stenophylax mucronatus 7 0 O. Escolà (Forat negre de Puigantic)
Cova de l’Espluga L’Espluga de Francolí 02/10/1966 410 Mesophylax aspersus 1 0 M. Nebot
Mina num. 2 Santa Coloma Castellcir 31/07/1982 880 Stenophylax sequax 5 4 O. Escolà
Mina num. 2 Santa Coloma Castellcir 31/07/1982 550 Stenophylax testaceus 0 1 O. Escolà
Mina Santa Coloma Castellcir 13/11/1983 880 Mesophylax aspersus 1 0 O. Escolà Pirineu axial
Avenc del Paradell Alt Urgell 21/07/1970 2000 Stenophylax nycterobius 2 0 O. Escolà
Avenc num. 9 de la Boixeda Camprodon 26/11/1972 1215 Mesophylax aspersus 1 0 O. Escolà
Baborell de Montsec Fòrnols i la Vansa 12/10/1969 1606 Stenophylax fissus 1 0 O. Escolà
Baborell de Montsec Fòrnols i la Vansa 12/10/1969 1606 Stenophylax malatestus 1 0 O. Escolà
Cauva deth Marimanha Naut Aran 20/07/1982 2175 Stenophylax vibex 3 2 sense dades
Cigalera de l’obaga de Baleran Alt Àneu 27/07/1982 2160 Stenophylax mitis 1 1 O. Escolà
Cigalera de l’obaga de Baleran Alt Àneu 27/07/1982 2160 Stenophylax vibex 0 1 O. Escolà
Cova A d’Olopte Isòvol 14/08/1973 1169 Stenophylax sequax 1 0 I. Gonzàlez
Cova de les Pubilles Gombrèn 15/06/1966 1650 Mesophylax aspersus 0 1 M. Nebot
Cova de les Pubilles Gombrèn 27/07/1971 1650 Mesophylax aspersus 1 0 Ll. Filbà
Cova de les Pubilles Gombrèn 15/06/1966 1650 Stenophylax mucronatus 1 0 M. Nebot
Cova Tuta Castellar de n’Hug 22/10/1967 1587 Mesophylax aspersus 2 0 Escolà, Ubach
Embut del Puigmal Queralbs 03/10/1971 2280 Stenophylax nycterobius 7 10 Ll. Filbà
Embut del Puigmal Queralbs 14/09/1975 2280 Stenophylax testaceus 0 4 Ll. Filbà
Forat de la neu Josa i Tuixén 18/10/1970 2430 Mesophylax aspersus 1 1 O. Escolà
Forat de la neu Josa i Tuixén 18/10/1970 2430 Stenophylax malatestus 2 1 O. Escolà
Forat de l’Estaca (Grallera gran La Vansa i Fòrnols 21/07/1970 2443 Stenophylax nycterobius 1 1 O. Escolà de Cornellana?)
Forat dels Eugassers Guardiola de Berguedà 01/11/1978 1993 Mesophylax aspersus 1 0 O. Escolà
Forat dels Eugassers Guardiola de Berguedà 16/08/1969 1993 Stenophylax nycterobius 1 1 sense dades
Grallera petita del Boixeder Josa i Tuixén 06/09/1968 2374 Mesophylax aspersus 1 0 O. Escolà (Grallera petita de Josa)
Mina Saragossa Queralbs 20/03/1977 1700 Mesophylax aspersus 1 0 O. Escolà
Mina Saragossa Queralbs 24/10/1999 1700 Mesophylax aspersus 1 0 O. Escolà
Tuta de la Fou de Bor Bellver de Cerdanya (Bor) 09/08/1982 1150 Stenophylax sequax 1 0 J.. Nebot
Tuta de la Fou de Bor Bellver de Cerdanya (Bor) 05/06/1965 1150 Stenophylax vibex 1 0 C. Ribera
Prepirineu Av. del Jabalí Baix Pallars 08/04/1966 0 Stenophylax malatestus 1 0 Ll. Auroux
Av. Pla del Vi Beuda
18/12/1983 1028 Mesophylax aspersus 0 1 O. Escolà
Avenc de la Mina Alòs de Balaguer 16/11/1969 1000 Mesophylax aspersus 4 0 Ll. Auroux
Avenc de la Roca del Corb Peramola 29/11/1970 866 Mesophylax aspersus 1 0 J. Comas
Avenc de la Roca del Corb Peramola 26/09/1993 866 Stenophylax mucronatus 2 1 O. Escolà
Avenc de la Serra de la Garriga Odèn
Avenc de la Serra de la Garriga Odèn
24/07/1968 1025 Stenophylax mucronatus 1 0 O. Escolà
24/07/1968 1025 Stenophylax sequax 1 0 O. Escolà
Avenc del Bages Guixers 01/10/1972 1405 Mesophylax aspersus 4 1 O. Escolà, M. Ubach
Avenc del Bages Guixers 01/10/1972 1405 Stenophylax fissus 1 1 O. Escolà, M. Ubach
Avenc del Bages Guixers 01/10/1972 1405 Stenophylax malatestus 1 0 O. Escolà, M. Ubach
Avenc del Bages Guixers 01/10/1972 1405 Stenophylax mucronatus 0 1 O. Escolà, M. Ubach
Avenc del barranc de la Conqueta Les Avellanes 02/11/1968 570 Mesophylax aspersus 1 0 Escolà, Romero, i Santa Linya Ubach
Avenc del Capolatell Navès 26/07/1967 1307 Mesophylax aspersus 0 1 O. Escolà
Avenc del Castell de Tarabau Baronia de Rialb 14/05/1972 940 Mesophylax aspersus 1 1 C. Ribera (Pou fons del Castell)
Avenc del Castell de Tarabau Baronia de Rialb 14/05/1972 940 Stenophylax mucronatus 0 1 C. Ribera (Pou fons del Castell)
Avenc del cingle de la Corba, Castellar del Riu 27/10/1968 1591 Mesophylax aspersus 3 0 O. Escolà Avenc del Sixto
Avenc del Mort Peramola 950 Mesophylax aspersus 2 0 C. Ribera
Avenc del Mort Peramola 01/11/1969 950 Mesophylax aspersus 3 0 Ribera, Roca, D.Romero
Avenc del Mort Peramola 25/10/1970 950 Mesophylax aspersus 5 0 C. Ribera
Avenc del Serrat de les Bruixes Vidrà (Collfred) 23/05/1971 1230 Stenophylax mucronatus 0 1 O. Escolà
Avenc d’en Jarillo 2 Os de Balaguer 07/11/1970 700 Mesophylax aspersus 6 0 O. Escolà
Avenc d’en Jarrillo 2 Os de Balaguer 07/11/1970 700 Stenophylax mucronatus 1 0 O. Escolà
Avenc Fred de Vallcebre Vallcebre 18/07/1970 1440 Stenophylax fissus 1 0 Roca
Avenc Fred de Vallcebre Vallcebre 18/07/1970 1440 Stenophylax nycterobius 2 0 Roca
Avenc num. 3 del Capolatell Navès 12/10/1980 1307 Mesophylax aspersus 2 0 O. Escolà
Baborell de Montsec Fòrnols i la Vansa 12/10/1969 1606 Mesophylax aspersus 2 0 O. Escolà
Bauma de Bruguer Terrades 01/11/1970 259 Mesophylax aspersus 1 1 O. Escolà
Bauma de Bruguer Terrades 20/04/1975 259 Mesophylax aspersus 2 1 J. Comas
Bauma de Talaixà Terrades 01/11/1970 234 Mesophylax aspersus 1 0 GIEM (o Avenc de Talaixà)
Bòfia de la Corba Castellar del Riu 27/10/1968 1602 Mesophylax aspersus 18 2 O. Escolà (Avenc de l’Heura)
Bòfia de la Corba Castellar del Riu 23/04/1978 1602 Mesophylax aspersus 0 1 O. Escolà (Avenc de l’Heura)
Bòfia de la Corba Castellar del Riu 23/04/1978 1602 Stenophylax fissus 1 0 O. Escolà (Avenc de l’Heura)
Bòfia de la Corba Castellar del Riu 23/04/1978 1602 Stenophylax malatestus 5 0 O. Escolà (Avenc de l’Heura)
Bofia de les Esglevades Navès 08/10/1978 1160 Mesophylax aspersus 1 0 O. Escolà
Bòfia de l’Obaga d’Espades o La Nou del Berguedà 02/09/2010 1388 Stenophylax espanioli 1 0 sense dades Cova d’Espades
Bòfia de l’Obaga d’Espades o La Nou del Berguedà 02/09/2010 1388 Stenophylax mucronatus 1 sense dades
Cova d’Espades
Bòfia de Torremàs (Avenc Odèn 28/09/1963 1175 Stenophylax mucronatus 0 1 J. Senent Montserrat Ubach)
Bòfia de Torremàs (Avenc Odèn 16/06/1974 1175 Stenophylax mucronatus 1 0 J. Comas Montserrat Ubach)
Bòfia dels Prats Coll de Nargó 07/09/1969 1655 Mesophylax aspersus 1 1 O. Escolà
Bòfia Port del Comte La Coma i la Pedra 21/09/1975 2050 Stenophylax nycterobius 1 0 J. Riart, C. Molinero
Bora Major Terrades 13/04/1974 347 Mesophylax aspersus 2 6 Bellés, Comas
Bora Major Terrades 04/11/1979 347 Mesophylax aspersus 1 0 O. Escolà
Bora Major Terrades 23/07/2004 347 Mesophylax aspersus 1 1 E.G.A
Bora Major Terrades 13/04/1974 347 Stenophylax mucronatus 0 1 Bellés, Comas
Botet de Casa Rei Tremp (Esplugafreda) 26/08/1986 940 Mesophylax aspersus 1 1 Bellés, Comas
Cova 3 de Rotgers Borredà 19/07/1969 912 Stenophylax sequax 2 4 O. Escolà
Cova Cosp (Palmerola) Les Lloses 07/10/1984 900 Mesophylax aspersus 2 0 O. Escolà
Cova Cuberes Serradell 19/07/1975 1135 Stenophylax sequax 1 0 X. Bellés
Cova de la Font Mentidora Conca de Dalt 13/06/1976 1400 Mesophylax aspersus 0 2 O. Escolà
Cova de la Font Mentidora Conca de Dalt 03/08/2015 1400 Mesophylax aspersus 0 3 Ll. Filbà
Cova de la Font Mentidora Conca de Dalt 13/06/1976 1400 Stenophylax mucronatus 2 0 O. Escolà
Cova de la Font Mentidora Conca de Dalt 03/08/2015 1400 Stenophylax mucronatus 4 4 Ll. Filbà
Cova de la Font Mentidora Conca de Dalt 13/06/1976 1400 Stenophylax nycterobius 0 1 O. Escolà
Cova de la Font Mentidora Conca de Dalt 03/08/2015 1400 Stenophylax nycterobius 0 2
Cova de la Font Mentidora Conca de Dalt 03/08/2015 1400 Stenophylax sequax 1 0 Ll. Filbà
Cova de la Font Mentidora Conca de Dalt 03/08/2015 1400 Stenophylax sequax 1 1 Ll. Filbà
Cova de la Font Mentidora Conca de Dalt 13/06/1976 1400 Stenophylax testaceus 0 2 O. Escolà
Cova de la Mosquera (Mina Beuda 05/07/1981 315 Stenophylax espanioli 3 0 Ll. Filbà de can Trompa)
Cova de la Mosquera (Mina Beuda 05/07/1981 315 Stenophylax mucronatus 0 1 Ll. Filbà de can Trompa)
Cova de les Lloberes Les Lloses 25/07/1970 1337 Stenophylax mucronatus 1 1 O. Escolà
Cova de les Lloberes Les Lloses 25/07/1970 1337 Stenophylax sequax 1 2 O. Escolà
Cova de l’Orri Sales de Llierca 15/02/1971 578 Stenophylax mucronatus 0 1 R. Canela
Cova del Calvari Vilada 05/06/1988 1186 Stenophylax espanioli 1 0 O. Escolà
Cova del Pla de Principi Albanyà 28/07/1972 980 Stenophylax mitis 2 6 Ll. Filbà
Cova del Pla de Principi Albanyà 06/05/1973 1066 Stenophylax mucronatus 2 0 Ll. Filbà
Cova del Pla de Principi Albanyà 28/07/1972 1066 Stenophylax sequax 3 0 Ll. Filbà
Cova del Rabeig del Cavall Navés 14/05/1967
Cova del Sanat
Cova del Sanat
Cova Fonda de Tragó de Noguera
950 Mesophylax aspersus 0 1 O. Escolà
Tremp (LaTorre de Tamúrcia) 25/09/1977 1670 Stenophylax mucronatus 1 0 O. Escolà
Tremp (LaTorre de Tamúrcia) 25/09/1977 1670 Stenophylax nycterobius 1 0 O. Escolà
26/05/1978 855 Mesophylax aspersus 1 0 O. Escolà Os de Balaguer
Cova Fonda de Tragó de Noguera
26/05/1978
855 Stenophylax espanioli 1 0 O. Escolà Os de Balaguer
Cova Fonda de Tragó de Noguera
26/05/1978
855 Stenophylax mucronatus 0 1 O. Escolà Os de Balaguer
Cova Fosca de Vilanova Vilanova de Meià
31/12/1963
836 Mesophylax aspersus 1 0 J. Senent
Cova Joan d’Os Les Avellanes i Santa 31/12/1968 830 Mesophylax aspersus 2 0 F. Español Linya (Tartareu)
Cova la Molsa Guardiola de Berguedà Stenophylax sequax 2 1 sense dades
Cova num 2 (Cova de la Paret num. 2) Àger
Cova num. 4 de Blancafort Ós de Balaguer 25/10/1970
815 Stenophylax testaceus 0 1 Ll. Auroux
400 Mesophylax aspersus 6 1 O. Escolà (Pantà de Canelles) (Tragó de Noguera)
Cova num. 4 de Blancafort Ós de Balaguer 25/10/1970
400 Stenophylax fissus 0 1 O. Escolà (Pantà de Canelles) (Tragó de Noguera)
Espluga Llorna Conca de Dalt (Serradell) 24/09/1977 1350 Mesophylax aspersus 0 O Escolà
Forat de la Grallera Llimiana 28/12/1968 1069 Stenophylax mucronatus O. Escolà
Forat de Safor Tremp (Llastarri) 29/12/1984 1340 Mesophylax aspersus 1 0 O. Escolà
Forat del Mollons Senterada (Lluçà) 23/09/1984 980 Mesophylax aspersus 1 1 O. Escolà
Forat del Toscà Tremp 26/12/1975 935 Mesophylax aspersus 1 0 O. Escolà
Forat d’Os (Forat Bufador) Os de Balaguer 27/05/1978 595 Mesophylax aspersus 9 8 O. Escolà
Forat d’Os (Forat Bufador) Os de Balaguer 27/05/1978 595 Stenophylax espanioli 15 10 O. Escolà
Forat d’Os (Forat Bufador) Os de Balaguer 27/05/1978 595 Stenophylax malatestus 4 3 O. Escolà
Forat La Bou Conca de Dalt 11/06/1978 1059 Mesophylax aspersus 0 1 O. Escolà
Forat La Bou Conca de Dalt 31/05/1992 1059 Mesophylax aspersus 1 0 O. Escolà
Graller de Badià Àger 16/03/1967 1185 Mesophylax aspersus 3 0 Ll. Auroux
Graller de Badià Àger 19/07/1974 1185 Mesophylax aspersus 0 1 O. Escolà
Graller de Castellar Tremp (Sapeira) 31/05/1971 1280 Stenophylax mucronatus 1 0 O. Escolà
Graller de Castellet Tremp 18/05/1980 1280 Stenophylax mucronatus 3 0 O. Escolà
Graller de Gurb Tremp 18/07/1971 1382 Stenophylax mucronatus 0 2 O. Escolà
Graller de la Pleta de l’Ós Saldes 30/06/1968 2075 Stenophylax testaceus 1 1 O. Escolà
Graller de les Planelles Pont de Suert (Corroncui) 27/07/1986 1365 Stenophylax sequax 1 1 O. Escolà
Graller del Boixeguer Sant Esteve de la Sarga 09/12/1967 1370 Mesophylax aspersus 1 3 O. Escolà
Graller del Boixeguer Sant Esteve de la Sarga 09/12/1967 1370 Stenophylax fissus 1 1 O. Escolà
Graller del Boixeguer Sant Esteve de la Sarga 09/12/1967 1370 Stenophylax malatestus 2 1 O. Escolà
Graller del Boixeguer Sant Esteve de la Sarga 09/12/1967 1370 Stenophylax nycterobius 3 0 O. Escolà
Grallera de Montsec Llimiana 15/08/1966 1069 Stenophylax espanioli 1 0 Ll. Auroux
Grallera de Prat Rodó Fígols i Alinyà 19/07/1970 2245 Mesophylax aspersus 1 0 O. Escolà (Forat de la Grallera)
Mina inferior de la Canal (Llastarri) Tremp 06/07/1980 1245 Mesophylax aspersus 0 1 O. Escolà
Mina inferior de la Canal (Llastarri) Tremp 29/12/1984 1245 Mesophylax aspersus 1 0 O. Escolà
Mina inferior de la Canal (Llastarri) Tremp 06/07/1980 1245 Stenophylax mucronatus 1 0 Ll. Filbà
Mina inferior del Canal (Llastarri) Tremp 06/07/1980 1200 Stenophylax malatestus 0 1 O. Escolà
Mina inferior del Canal (Llastarri) Tremp 06/07/1980 1200 Stenophylax mitis 0 1 O. Escolà
Mina La Pilar Alós de Balaguer 02/11/2019 800 Mesophylax aspersus 1 0 sense dades
Mina superior de la Canal (Llastarri) Tremp 06/07/1980 1249 Mesophylax aspersus 4 0 Ll. Filbà
Pou del Coll d’Alzina Odèn 23/09/1963 1065 Mesophylax aspersus 2 1 O. Escolà
Pou del Coll d’Alzina Odèn 22/11/1963 1065 Mesophylax aspersus 1 0 J. Senent
Pou del Coll d’Alzina Odèn 23/09/1963 1065 Stenophylax fissus 0 1 O. Escolà
Pou del Coll d’Alzina Odèn 23/09/1963 1065 Stenophylax malatestus 1 1 O. Escolà
Querant de Picalts Vilanova de Meià 03/01/1964 750 Mesophylax aspersus 2 0 J. Senent
Surgència del Cingle Vilanova de Meià 14/04/1995 800 Mesophylax aspersus 0 1 O. Escolà
Surgència del Cingle Vilanova de Meià 14/04/1995 800 Stenophylax sequax 0 1 O. Escolà
Serralada Litoral Catalana
Avenc de la Fragata Sitges 16/05/1964
Avenc de la Roser Vallirana 26/05/1974
Avenc de les Aranyes Tordera 28/10/2020
Avenc de l’Esquerda La Palma de Cervelló 16/02/1975
Avenc de l’Esquerda num. 1 La Palma de Cervelló 06/04/1975
Avenc de Sant Cristofol Begues 16/10/1966
Avenc de Sant Marçal Olesa de Bonesvalls 01/11/1973
499 Mesophylax aspersus 1 0 M. Nebot
460 Stenophylax fissus 1 0 Ll. Filbà
400 Mesophylax aspersus 0 1 EGA
230 Mesophylax aspersus 9 0 Bellés, Comas
230 Mesophylax aspersus 1 0 O. Escolà
516 Mesophylax aspersus 0 1 O. Escolà
362 Mesophylax aspersus 0 1 O. Escolà
Avenc de Sant Marçal Olesa de Bonesvalls 21/06/1990 362 Mesophylax aspersus 0 1 Sibila
Avenc del Curset Vallirana 27/11/1966 465 Mesophylax aspersus 1 0 O. Escolà
Avenc del Karst Muntanyes de l’ordal 21/04/1963 Mesophylax aspersus 1 1 O. Escolà (cavitat desapareguda)
Avenc del Karst Muntanyes de l’ordal 20/06/1982 Mesophylax aspersus 0 1 O. Escolà (cavitat desapareguda)
Avenc del Mall Begues 30/11/1986 545 Mesophylax aspersus 1 0 O. Escolà
Avenc del Puig de Marc La Riba 12/10/1971 685 Stenophylax mucronatus 1 0 GIEM
Avenc del Vermell Begues 12/03/1986 422 Mesophylax aspersus 1 0 O. Escolà
Avenc del Vermell Begues 29/03/1987 422 Mesophylax aspersus 0 1 O. Escolà
Avenc dels Guerrillers Gavà (cavitat desapareguda) 05/09/1971 345 Mesophylax aspersus 1 2 Ll. Filbà
Avenc dels Sellons Olesa de Bonesvalls Mesophylax aspersus 1 1 Casado
Avenc d’en Roca Cervelló 12/09/1971 503 Stenophylax mucronatus 0 1 X. Bellés
Avenc d’Olèrdola Olèrdola 05/05/1969 323 Mesophylax aspersus 0 1 O. Escolà
Avenc Emili Sabaté Sitges 19/05/1963 390 Mesophylax aspersus 0 1 I. Gonzàlez
Avenc Font i Sagué (Arcada Gran) Olesa de Bonesvalls 31/05/1970 381 Mesophylax aspersus 1 0 Ll. Auroux
Avenc GIESC Begues 26/12/1978 563 Mesophylax aspersus 1 0 O. Escolà
Avenc Lluís Solà Begues 08/09/1974 512 Mesophylax aspersus 0 1 J. Comas
Avenc Nostre (Avenc de Can Rafel) Cervelló 31/05/1973 458 Stenophylax mucronatus 1
Avenc Nostre (Avenc de Can Rafel) Cervelló 01/05/1974 458 Stenophylax mucronatus 1
Avenc Pomar Vallirana 26/05/1974 431 Mesophylax aspersus 1
Avenc Pomar Vallirana 26/05/1974 431 Stenophylax mucronatus 0
Avenc Pompeu Fabra Vallirana 08/09/1974 453 Stenophylax espanioli 1 0 J. Comas
Cova de la Moneda Mont-ral 29/06/1967 1000 Mesophylax aspersus 0
Cova de la Moneda Mont-ral 29/06/1967
Cova de Mont-ral Mont-ral 15/06/1965 1000 Stenophylax espanioli 2
Cova del Bosc cremat Pallejà 13/04/1974
Cova del Bosc cremat Pallejà
Cova del Pla de les Comes Cervelló
(Cova del Lledoner)
Cova del Pla de les Comes Cervelló
(Cova del Lledoner)
Mina de can Montsant Tordera 14/09/1985
Mina de can Montsant Tordera 14/04/1985
Mina de can Montsant Tordera 16/06/1985
Mina de can Montsant Tordera 28/09/1986
Mina de can Montsant Tordera 14/04/1985
Mina de can Montsant Tordera 16/06/1985
Mina de can Montsant Tordera 28/09/1986
Mina de can Montsant Tordera 28/09/1986
Stenophylax fissus 1 1 O. Escolà
Comas, Escolà
Escolà
Surgència Poblet Mont-ral 15/06/1965 957 Mesophylax aspersus 9 10 I. Gonzàlez
Surgència Poblet Mont-ral 15/06/1965 957 Stenophylax espanioli 0 1 I. Gonzàlez
Surgència Poblet Mont-ral 15/06/1965 957 Stenophylax fissus 0 2 I. Gonzàlez
Surgència Poblet Mont-ral 15/06/1965 957 Stenophylax malatestus 3 2 I. Gonzàlez
Surgència Poblet Mont-ral 15/06/1965 957 Stenophylax mitis 0 2 I. Gonzàlez
Surgència Poblet Mont-ral 15/06/1965 957 Stenophylax sequax 1 0 I. Gonzàlez
Serralada Prelitoral Catalana Av. de la Lludriga Capafonts 24/06/1971 900 Stenophylax sequax 0 1 GIEM Av. de la Lludriga Capafonts 24/06/1971 900 Stenophylax testaceus 2 0 GIEM Av. D’en Viumala La Llacuna 11/11/1979 716 Mesophylax aspersus 3 2 O. Escolà
Avenc B Montsià 15/09/1972 300 Mesophylax aspersus 1 0 D. Schibi
Avenc circa Mina del Port Mas de Barberans 14/04/1984 400 Mesophylax aspersus 2
Avenc Costa Dreta El Figueró i Montmany 26/04/1967 500 Mesophylax aspersus 1 0 I. Gonzàlez
Avenc de Can Castellet de dalt Sentmenat 16/06/1968 627 Mesophylax aspersus 5 11 O. Escolà
Avenc de Can Castellet de dalt Sentmenat 16/06/1963 627 Stenophylax mucronatus 1 0 O. Escolà
Avenc de Can Pobla Matadepera 03/10/1976 1008 Stenophylax nycterobius 0 1 O. Escolà
Avenc de Castellsapera Terrassa 29/10/1989 875 Mesophylax aspersus 0 1 sense dades
Avenc de la Brega Mura 08/09/1968 725 Mesophylax aspersus 1 1 sense dades
Avenc de la Canal de la Revella Sant Llorenç Savall 27/08/1977 832 Mesophylax aspersus 0 1 sense dades
Avenc de la Canal de l’Esquirol Matadepera 27/06/1976 816 Mesophylax aspersus 1 1 O. Escolà
Avenc de la canal de Mura Vacarisses 31/07/1977 808 Mesophylax aspersus 1 0 sense dades
Avenc de la canal de Mura Vacarisses 10/02/1980 808 Mesophylax aspersus 1 0 O. Escolà
Avenc de la canal de Mura Vacarisses 06/06/2019 808 Mesophylax aspersus 0 1 F. Fadrique, S. Gago
Avenc de la Carbonera Margalef 08/04/1966 845 Mesophylax aspersus 0 1 Ll. Auroux
Avenc de la Carbonera Margalef 08/04/1966 845 Stenophylax malatestus 1 1 Ll. Auroux
Avenc de la Codoleda Matadepera 15/10/1967 855 Mesophylax aspersus 1 1 A. Messeguer
Avenc de la Codoleda Matadepera 24/10/1971 855 Mesophylax aspersus 6 1 X. Bellés
Avenc de la Codoleda Matadepera 28/09/1975 855 Mesophylax aspersus 2 5 O. Escolà
Avenc de la Codoleda Matadepera 23/10/1977 855 Mesophylax aspersus 3 2 O. Escolà
Avenc de la Falconera Mura 19/10/1968 805 Mesophylax aspersus 1 0 Quintana
Avenc de la Falconera Mura 24/02/1977 805 Mesophylax aspersus 1 0 O. Escolà
Avenc de la Falconera Mura 28/06/2012 805 Stenophylax mucronatus 0 1 M. Nebot, J. Pastor
Avenc de la Febró La Febró 07/08/1966 976 Mesophylax aspersus 1 0 O. Escolà
Avenc de la Febró La Febró 07/08/1966 976 Stenophylax testaceus 9 2 O. Escolà
Avenc de la Solana Pontons 28/11/1971 650 Mesophylax aspersus 2 5 D. Schibi
Avenc de la Solana Pontons 12/10/1971 650 Mesophylax aspersus 3 0 Español, Schibi, –Cantons
Avenc de la Solana La Llacuna 28/11/1971 650 Mesophylax aspersus 1 0 X. Bellés
Avenc de la Solana Pontons 12/10/1971 650 Stenophylax espanioli 1 0 Español, Schibi
Avenc de la Suor Alfara de Carles 14/08/1972 1218 Mesophylax aspersus 1 0 O. Escolà
Avenc de la Suor Alfara de Carles 13/04/1974 1218 Mesophylax aspersus 0 1 O. Escolà
Avenc de la Suor Alfara de Carles 14/08/1972 1218 Stenophylax mucronatus 9 0 O. Escolà
Avenc de la Suor Alfara de Carles 14/08/1972 1218 Stenophylax testaceus 0 1 O. Escolà
Avenc de l’Arboçar Margalef 06/04/1966 908 Stenophylax fissus 3 0 Ll. Auroux
Avenc de l’Arboçar Margalef 08/04/1966 908 Stenophylax malatestus 2 0 Ll. Auroux
Avenc de l’Auferí Margalef 08/04/1966 897 Stenophylax fissus 2 0 Ll. Auroux
Avenc de l’Auferí Margalef 08/04/1966 897 Stenophylax malatestus 1 1 Ll. Auroux
Avenc de l’Engany Tortosa 20/04/1973 1315 Stenophylax fissus 1 1 D. Schibi
Avenc de l’Engany Tortosa 02/04/1970 1315 Stenophylax mucronatus 1 0 Montsonís, Schibi
Avenc de les Clivelleres
Mediona 12/03/1974 606 Mesophylax aspersus 0 1 O. Escolà
Avenc de les Clivelleres Mediona 09/06/1974 606 Mesophylax aspersus 0 1 O. Escolà
Avenc de les Clivelleres num. 1 Mediona 01/05/1973 592 Mesophylax aspersus 1 0 O. Escolà
Avenc de les Clivelleres num. 1 Mediona 24/02/1974 592 Mesophylax aspersus 1 0 O. Escolà
Avenc de les Clivelleres num. 1 Mediona 12/03/1974 592 Mesophylax aspersus 0 1 O. Escolà
Avenc de les Clivelleres num. 1 Mediona 01/05/1975 592 Mesophylax aspersus 2 2 O. Escolà
Avenc de les Clivelleres num. 1 Mediona 07/11/1976 606 Mesophylax aspersus 1 1 O. Escolà
Avenc de les Clivelleres num. 1 Mediona 01/05/1977 592 Mesophylax aspersus 3 7 O. Escolà
Avenc de les Clivelleres num. 1 Mediona 06/11/1977 592 Mesophylax aspersus 9 0 O. Escolà
Avenc de les Clivelleres num. 1 Mediona 03/12/1978 592 Mesophylax aspersus 1 1 O. Escolà
Avenc de les Clivelleres num. 1 Mediona 29/04/1980 606 Mesophylax aspersus 1 0 O. Escolà
Avenc de les Clivelleres num. 1 Mediona 23/10/1988
Avenc de les Clivelleres num. 1 Mediona 09/06/1974
Avenc de les Clivelleres num. 1 Mediona 01/05/1977
592 Mesophylax aspersus 6 0 O. Escolà
592 Stenophylax espanioli 2 0 O. Escolà
592 Stenophylax espanioli 1 0 O. Escolà
Avenc de les Clivelleres num. 1 Mediona 09/06/1974 592 Stenophylax mucronatus 1 0 O. Escolà
Avenc de les Clivelleres num. 1 Mediona 01/05/1977
Avenc de les Clivelleres num. 2 Mediona 09/03/1975
Avenc de les Clivelleres num. 2 Mediona 06/11/1977
592 Stenophylax mucronatus 6 8 O. Escolà
610 Mesophylax aspersus 2 0 J. Comas
610 Mesophylax aspersus 29 3 O. Escolà
Avenc de les Clivelleres num. 2 Mediona 03/12/1978 610 Mesophylax aspersus 12 2 O. Escolà
Avenc de les Clivelleres num. 3 Mediona 09/06/1974
Avenc de les Clivelleres num. 3 Mediona 23/10/1988
Avenc de les Clivelleres num. 3 Mediona 23/04/1989
Avenc de les Clivelleres num. 3 Mediona 01/05/1977
Avenc de les Clivelleres num.3 Mediona 03/03/1973
606 Mesophylax aspersus 1 0 O. Escolà
606 Mesophylax aspersus 1 0 O. Escolà
606 Mesophylax aspersus 1 0 O. Escolà
606 Stenophylax mucronatus 1 0 O. Escolà
606 Mesophylax aspersus 4 0 J. Comas
Avenc de les Pedreres Gualba 12/05/1971 481 Mesophylax aspersus 0 1 O. Escolà
Avenc de les Pedreres Gualba 12/05/1974 481 Stenophylax mucronatus 1 0 O. Escolà
Avenc de l’Espadella Tivissa 10/09/1993 517 Mesophylax aspersus 0 1 O. Escolà
Avenc de l’obaga de l’Illa Castellar del Vallés 01/11/1964 518 Mesophylax aspersus 6
Avenc de l’Orellut Montblanc 21/10/1990
aspersus
Avenc de Sant Jaume de la Mata Mura 20/10/1968 873 Mesophylax aspersus 2 0 sense dades
Avenc del Bac Sant Martí de Centelles 27/10/1984
Avenc del Bac Sant Martí de Centelles 28/10/1984
Mesophylax aspersus 5 0 J. Comas
Mesophylax aspersus 4 0 O. Escolà
Avenc del Batlle vell Pontons 25/07/1971 705 Mesophylax aspersus 1 1 O. Escolà (Etiquetat Vallebell)
Avenc del Batlle vell Pontons 25/07/1971
Stenophylax espanioli 1 1 O. Escolà (Etiquetat Vallebell)
Avenc del Club Matadepera 13/06/1971
Avenc del Club Matadepera 26/10/1975
Avenc del Club Matadepera 27/11/1977
Mesophylax aspersus 0 1 T. Cuñé
Avenc del Club Matadepera 23/11/1980 795 Mesophylax aspersus 5 0 J. Comas
Avenc del Club Matadepera 13/06/1968 795 Stenophylax espanioli 1 0 O. Escolà
Avenc del Figuerot Matadepera 18/06/1978
Mesophylax aspersus 5 3 O. Escolà
Avenc del Figuerot Matadepera 07/10/1979 742 Mesophylax aspersus 3 3 O. Escolà
Avenc del Figuerot Matadepera 18/06/1978 742 Stenophylax mucronatus 2 1 O. Escolà
Avenc del GIEM La Febró 27/03/1970 975 Mesophylax aspersus 2 1 GIEM
Avenc del GIEM La Febró 08/04/1971 975 Mesophylax aspersus 0 1 GIEM
Avenc del GIEM La Febró 29/05/1977 975 Mesophylax aspersus 5 2 O. Escolà
Avenc del GIEM La Febró 12/02/1978 975 Mesophylax aspersus 0 1 GIEM
Avenc del GIEM La Febró 18/10/1979 975 Mesophylax aspersus 0 1 O. Escolà
Avenc del GIEM La Febró 02/09/1979 975 Stenophylax espanioli 1 0 O. Escolà
Avenc del GIEM La Febró 02/09/1979 975 Stenophylax mitis 1 0 O. Escolà
Avenc del GIEM La Febró 29/05/1977 975 Stenophylax mucronatus 2 1 O. Escolà
Avenc del GIEM La Febró 29/05/1977 975 Stenophylax testaceus 9 2 O. Escolà
Avenc del GIEM La Febró 02/09/1979 975 Stenophylax testaceus 13 10 O. Escolà
Avenc del Julivert Vimbodí i Poblet 06/07/1969 652 Mesophylax aspersus 3 2 I. Gonzàlez
Avenc del Julivert Vombodí i Poblet 09/11/1980 652 Mesophylax aspersus 3 0 sense dades
Avenc del Llest Mura 01/04/1973 852 Mesophylax aspersus 1 1 Ll. Auroux
Avenc del Mascar Alfara de Carles 07/06/1976 1058 Mesophylax aspersus 0 1 O. Escolà
Avenc del Muronell Castellar del Vallès 01/12/1963 506 Mesophylax aspersus 2 0 O. Escolà
Avenc del Pèndol Tortosa 14/04/1974 1252 Mesophylax aspersus 2 0 O. Escolà
Avenc del Pèndol Tortosa 12/04/1974 1252 Mesophylax aspersus 2 0 O. Escolà
Avenc del Púdol Margalef 15/03/1966 826 Stenophylax malatestus 9 0 Ll. Auroux
Avenc del Puig de Marc La Riba 29/12/1968 685 Mesophylax aspersus 1 0 Masdeu
Avenc del Roc de les Abelles Mont-Ral (Farena) 12/04/1952 800 Stenophylax mucronatus 0 1 F. Español
Avenc del Salany Tortosa 30/04/1972 1243 Mesophylax aspersus 0 1 O. Escolà
Avenc del Tortosí Mura 29/05/1975 750 Mesophylax aspersus 1 0 O. Escolà
Avenc del Tortosí Mura 03/08/1968 750 Stenophylax mucronatus 1 0 O. Escolà
Avenc del Tortosí Mura 26/07/1980 750 Stenophylax mucronatus 2 1 O. Escolà
Avenc del Tortosí Mura 29/05/1975 750 Stenophylax permistus 0 1 O. Escolà
Avenc dels Mamelons Tortosa 21/04/1973 1222 Stenophylax mucronatus 1 0 D. Schibi
Avenc dels Pinyerets El Montmell 25/07/1978 755 Mesophylax aspersus 3 3 O. Escolà
Avenc d’en Daniel Capafonts 01/11/1969 800 Mesophylax aspersus 1 0 GIEM
Avenc d’En Toni El Figueró i Montmany 09/05/1976 523 Mesophylax aspersus 0 1 J. Comas
Avenc d’Francesc Margalef 08/04/1966 914 Stenophylax fissus 1 0 Ll. Auroux
Avenc d’Francesc Margalef 08/04/1966 914 Stenophylax malatestus 4 0 Ll. Auroux
Avenc Enfonsat Seva 24/09/1985 550 Stenophylax sequax 1 0 O. Escolà
Avenc Gran Margalef 08/04/1966 826 Stenophylax fissus 1 0 Ll. Auroux
Avenc Gran Margalef 08/04/1966 826 Stenophylax malatestus 2 1 Ll. Auroux
Avenc Gran Margalef 08/04/1966 826 Stenophylax mucronatus 0 1 Ll. Auroux
Avenc Gran de Rojalons Montblanc 30/03/1969 840 Mesophylax aspersus 1 0 I. Gonzàlez
Avenc Nyaca-Nyaca Muntanyes de Prades 04/03/1973 Stenophylax malatestus 1 0 D. Schibi
Avenc Subils-Godoy Sant Llorenç del Munt 12/09/1966 670 Mesophylax aspersus 0 1 O. Escolà
Avenc Tossals (Avenc de la Tossa) Prat de Comte 20/03/1981 580 Mesophylax aspersus 0 4 O. Escolà
Cau dels Llimbs Sant Feliu de Codines 16/10/1988 550 Mesophylax aspersus 1 0 O. Escolà
Cova Balaguer Alfara de Carles 05/03/1978 1148 Mesophylax aspersus 1 0 O. Escolà
Cova Balaguer Alfara de Carles 05/03/1978 1148 Stenophylax mucronatus 0 1 O. Escolà
Cova Balaguer Alfara de Carles 02/07/1978 1148 Stenophylax mucronatus 1 1 O. Escolà
Cova Cambra Tortosa 25/04/1970 1300 Mesophylax aspersus 1 0 O. Escolà
Cova Dakotes Cabrera d’Anoia 08/04/1996 308 Mesophylax aspersus 0 1 O. Escolà
Cova de la Mola Colldejou 28/05/1971
Cova de la Mola Colldejou 28/05/1971
Cova de la Moneda Mont-ral 29/09/1951
Cova de la Moneda Mont-ral
880 Stenophylax espanioli 1 0 GIEM
880 Stenophylax mucronatus 1 0 GIEM
1000 Mesophylax aspersus 3 0 F. Español
1000 Stenophylax malatestus 1 0 Ll. Auroux
Cova de la Rabosa El Perelló 20/08/1972 400 Mesophylax aspersus 0 1 Bley
Cova de la Sensada Nova Orpí 15/11/1981 568 Mesophylax aspersus 1 0 O. Escolà
Cova de la Solana Roquetes 16/06/1984 1198 Mesophylax aspersus 1 0 sense dades
Cova de les Animetes Matadepera 11/03/1984 915 Mesophylax aspersus 5 3 O. Escolà
Cova de les Rondes La Llacuna 16/03/1980 750 Mesophylax aspersus 1 0 J. Comas
Cova de les Rondes La Llacuna 29/11/1981 750 Mesophylax aspersus 0 1 O. Escolà
Cova de les Rondes La Llacuna 16/05/1982 750 Mesophylax aspersus 1 1 O. Escolà
Cova de les Rondes La Llacuna 24/09/1970 750 Stenophylax espanioli 0 1 O. Escolà
Cova de les Rondes La Llacuna 23/03/1980 750 Stenophylax malatestus 1 0 J. Espinosa; X.Bellés
Cova de les Rondes La Llacuna 24/09/1970 750 Stenophylax mucronatus 2 2 O. Escolà
Cova de Mont-ral Mont-ral 30/08/1970 1000 Stenophylax espanioli 2 2 sense dades
Cova de Sant Jordi Montblanc (Prenafeta) 08/01/1960 500 Stenophylax mucronatus 1 0 J. Senent
Cova de Vallmajor Albinyana 21/03/1966 334 Mesophylax aspersus 2 6 Ll. Auroux
Cova de Vallmajor Albinyana 10/07/1966 334 Mesophylax aspersus 2 0 Casado
Cova de Vallmajor Albinyana 23/11/1969 334 Mesophylax aspersus 2 0 E. Roca
Cova de Vallmajor Albinyana 07/04/1984 334 Mesophylax aspersus 3 0 J. Comas
Cova de Vallmajor Albinyana 29/06/1975 334 Stenophylax espanioli 1 0 J.M. Parera
Cova de Vallmajor Albinyana 07/04/1984 334 Stenophylax mucronatus 0 1 J. Comas
Cova del Bolet Mediona 05/03/1995 630 Stenophylax malatestus 0 1 O. Escolà
Cova del Codó Mont-ral 18/06/1967 980 Stenophylax espanioli 1 0 O. Escolà
Cova del Codó Mont-ral 07/09/1971 980 Stenophylax espanioli 0 1 GIEM
Cova del Codó Mont-ral 28/03/1965 980 Stenophylax malatestus 1 0 I. Gonzàlez
Cova del Conill Horta de Sant Joan 23/11/1969 425 Mesophylax aspersus 3 0 O. Escolà
Cova del Drac Vilaverd 02/11/1952 460 Mesophylax aspersus 2 1 F. Español
Cova del Fondal Matadepera 03/07/1977 930 Mesophylax aspersus 1 0 O. Escolà
Cova del Garrofet Querol 12/09/1970 776 Stenophylax espanioli 0 1 GIEM
Cova del Gat Figuerola del Camp 08/01/1960 445 Mesophylax aspersus 1 0 J. Senent
Cova del Gat Figuerola del Camp 08/01/1960 445 Stenophylax mucronatus 2 0 J. Senent
Cova del Gurugú Montblanc 05/06/1969 500 Stenophylax mucronatus 0 1 GIEM
Cova del Llorito (etiquetat com Avenc) Tarragona 20/02/1966 90 Mesophylax aspersus 3 0 I. Gonzàlez
Cova del Mandil (Cova del Diable) Querol 19/06/1977 510 Stenophylax espanioli 1 0 O. Escolà
Cova del Pasteral La Cellera de Ter 03/01/1981 213 Mesophylax aspersus 3 1 F. Vallhonrat, De Gregorio
Cova del Racó de l’Hospital Montblanc 26/05/1969 593 Mesophylax aspersus 0 1 GIEM
Cova del Racó de l’Hospital Montblanc 14/05/1969 593 Mesophylax aspersus 0 1 GIEM
Cova del Racó de l’Hospital Montblanc 04/10/1969 593 Mesophylax aspersus 2 0 GIEM
Cova del Racó de l’Hospital Montblanc 25/06/1971 593 Mesophylax aspersus 1 0 GIEM
Cova del Racó de l’Hospital Montblanc 25/04/1971 593 Mesophylax aspersus 1 0 Topoy, Espelt
Cova del Racó de l’Hospital Montblanc 31/10/1971 593 Mesophylax aspersus 3 0 GIEM
Cova del Racó de l’Hospital Montblanc 26/05/1969 593 Stenophylax mucronatus 1 0 GIEM
Cova del Racó de l’Hospital Montblanc 25/06/1971 593 Stenophylax testaceus 0 1 GIEM
Cova del Traça Cabra del Camp 09/11/1980 564 Mesophylax aspersus 6 3 O. Escolà
Cova del Traça Cabra del Camp 21/12/1980 564 Mesophylax aspersus 1 0 O. Escolà
Cova del Traça Cabra del Camp 09/07/1978 564 Stenophylax espanioli 1 0 O. Escolà
Cova del Triangle Sant Llorenç Savall 04/05/1969 887 Mesophylax aspersus 0 1 O. Escolà
Cova dels Calaixos de l’Ereldo El Vilosell 996 Stenophylax espanioli 0 1 I. Gonzàlez
Cova dels Calaixos de l’Ereldo El Vilosell 996 Stenophylax espanioli 1 0 I. Gonzàlez
Cova dels Ermitans Caldes de Montbui 17/11/1974 520 Mesophylax aspersus 2 1 X. Bellés
Cova dels Ermitans Caldes de Montbui 20/12/1974 520 Mesophylax aspersus 2 0 Bellés, Comas
Cova Freda Collbató 23/11/1969 502 Mesophylax aspersus 3 0 Ll. Auroux
Cova Freda (Avenc de les Torres) Torrelles de Foix 02/11/1980 685 Mesophylax aspersus 4 1 O. Escolà
Cova Gran del Bonretorn L’Albiol 29/01/1989 535 Mesophylax aspersus 1 0 O. Escolà
Cova Llúdriga Tivissa 14/08/1993 495 Mesophylax aspersus 1 0 O. Escolà
Cova Mal Pas Alfara de Carles 02/07/1978 1142 Stenophylax mucronatus 3 1 Ll. Filbà
Cova Nerola Vombodí i Poblet 02/10/1970 597 Mesophylax aspersus 0 1 sense dades
Cova Nerola Vimbodí i Poblet 30/12/1971 597 Mesophylax aspersus 1 0 GIEM
Cova petita de la Moneda Mont-ral 1000 Mesophylax aspersus 1 0 Ll. Auroux
Cova Rojalons num. 1 Montblanc 30/03/1969 852 Stenophylax fissus 1 0 I. Gonzàlez (Avenc gran de Rojalons)
Cova Simanya petita Sant Llorenç Savall 17/10/1971 895 Mesophylax aspersus 1 0 X. Bellés
Cova Simanya petita Sant Llorenç Savall 26/03/1972 895 Mesophylax aspersus 1 0 Bley
Coves de Mura Mura 28/07/1968 689 Mesophylax aspersus 3 1 O. Escolà
Coves de Mura Mura 20/06/1971 689 Mesophylax aspersus 0 2 R. Lloses
Coves de Mura Mura 05/10/1975 689 Mesophylax aspersus 4 0 Comas, Enriquez
Coves de Mura Mura 02/11/2002 689 Mesophylax aspersus 1 0 G. Masó
Coves de Mura Mura 20/06/1971 689 Stenophylax mucronatus 1 0 R. Lloses
Coves del Mamut Cabrera d’Anoia 08/10/1967 295 Mesophylax aspersus 2 3 Español, Auroux
Coves del Mamut Cabrera d’Anoia 05/04/1970 295 Mesophylax aspersus 2 0 Escolà, Auroux
Coves del Rufino Arbolí 14/01/2001 732 Mesophylax aspersus 1 0 Fadrique, Escolà
Coves del Salnitre Collbató 15/11/1973 535 Mesophylax aspersus 10 2 Bley, Auroux
Forat (Cova) del Rastre Roquetes 22/05/1972 1237 Stenophylax mucronatus 1 0 O. Escolà
Forat Calent Caldes de Montbui 02/02/1992 200 Mesophylax aspersus 2 0 O. Escolà
Forat del riu Algars Alfara de Carles 07/05/2012 1035 Mesophylax aspersus 1 2 J. Mederos, J. Pastor
Forat del riu Algars Alfara de Carles 07/05/2012 1035 Mesophylax aspersus 0 1 F. Fadrique
Forat del riu Algars Alfara de Carles 07/05/2012 1035 Stenophylax mucronatus 2 0 J. Mederos, J. Pastor
Forat del Vent-Cova d’Artús Albinyana 19/01/1992 360 Mesophylax aspersus 3 0 O. Escolà
Mina de Pontons (marge dret del Foix)Pontons 01/08/1971 600 Mesophylax aspersus 0 1 D. Schibi
Mina del Port Mas de Barberans 08/06/1980 400 Mesophylax aspersus 3 3 O. Escolà
Mina del Port Mas de Barberans 14/04/1984 400 Mesophylax aspersus 2 1 O. Escolà
Mina del Port Mas de Barberans 24/04/1988 400 Mesophylax aspersus 1 0 O. Escolà
Mina del Port Mas de Barberans 15/09/1993 400 Mesophylax aspersus 2 3 O. Escolà
Galicia
Mina del Port Mas de Barberans 21/10/1996
Mina del Port Mas de Barberans 30/10/1997
Mina del Port Mas de Barberans 19/04/1998
Mina Font de les Canals La Llacuna (Pla d’Ancosa) 25/07/1971
Mina Horts num. 2 Torre de l’Espanyol 24/06/1998
Mina num. 1 de Pontons Pontons 28/08/1996
Pou de Costa Dreta Collbató 12/01/1986
Pouetons de les Agulles El Bruc 24/04/1971
Pouetons de les Agulles El Bruc 07/10/1973
Pouetons de les Agulles El Bruc 31/05/1987
Pouetons de les Agulles El Bruc 30/05/1965
Serralada Transversal Catalana
Avenc de Can Plana Sant Martí de Llémana 21/06/1992
Avenc de Can Plana Sant Martí de Llémana 21/06/1992
aspersus
Mesophylax aspersus 3
Fadrique
Escolà
Stenophylax mucronatus 1 1 O. Escolà
Avenc de Can Roca Roja num.3 Sant Feliu de Pallarols 08/11/1981 970 Mesophylax aspersus 1 0 O. Escolà
Avenc del Bloc (Cova Sant Aniol) Sant Aniol de Finestres 15/07/1967 456 Mesophylax aspersus 1 0 R. Viñas
Avenc num. 1 de can Roca Roja Sant Feliu de Pallerols 23/05/1976
Bora Fosca Tavertet 01/05/1971
Bora Fosca
Bora Fosca
Bora Fosca
Tavertet 20/06/1971
Tavertet 27/05/1973
Tavertet 24/10/1976
Bora Fosca Tavertet 18/06/1972
Mesophylax aspersus 1
Mesophylax aspersus 1 0 J.M. Parera
Mesophylax aspersus 6 15 Ll. Filbà
Mesophylax aspersus 1
Parera
Stenophylax fissus 2 1 Ll. Filbà
Bora Fosca Tavertet 18/06/1972 650 Stenophylax mucronatus 1 5 Ll. Filbà
Bora Fosca Tavertet 03/09/1972 650 Stenophylax mucronatus 6 1 Ll. Filbà
Bora Fosca Tavertet 07/07/1974 650 Stenophylax mucronatus 0 1 J.M. Ventura
Bora Fosca Tavertet 27/05/1975 650 Stenophylax mucronatus 1 0 Ll. Filbà
Bora Fosca Tavertet 03/09/1972 650 Stenophylax vibex 0 1 Ll. Filbà
Cova de les Guilloteres Amer 19/05/1974
Cova del Serrat del Vent Tavertet 12/07/1981
Cova del Serrat del Vent Tavertet 12/07/1981
Mesophylax aspersus 0 1 J. Comas
Stenophylax mitis 5 5 O. Escolà
1000 Stenophylax sequax 8 5 O. Escolà
Cova del Serrat del Vent Tavertet 12/07/1981 1000 Stenophylax vibex 5 5 O. Escolà
Cova Stop (Cova del Far) Susqueda 27/10/1984
Mesophylax aspersus 1 0 O. Escolà
Forat del Far Susqueda 21/10/1979 1071 Mesophylax aspersus 1 0 O. Escolà
Forat del Far Susqueda 19/11/1983 1071 Mesophylax aspersus 2 0 Bellés, Comas
Forat del Far Susqueda 30/04/2014 1071 Mesophylax aspersus 1 1 Biosp.
Forat del Far Susqueda 21/05/1983 1071 Stenophylax mucronatus 2 1 X. Bellés Euskadi
Muntanyes Basques
Cova Azkoiti Ceanuri 29/07/1962 300 Stenophylax fissus 1 1 E. Nolte
Cueva del Viento Mendaro 15/07/1998 247 Stenophylax mitis 4 2 Salg., Fresn., Escolls, Escolà
Cueva la Laina Karrantza 26/05/1974 200 Stenophylax permistus 2 0 GEV
Cueva Laguanaz 3 num. 6 Turtzioz (Trucios) 16/08/1976
200 Stenophylax sequax 2 0 Cano
Cueva Mendikute Albiztur 15/07/1998 710 Stenophylax testaceus 0 1 Salg., Fresn., Escolls, Escolà
Cueva Peña Rota Zigoiotia 16/04/1965 1000 Stenophylax fissus 0 1 J.M. Vitoria
Cueva Peña Rota Zigoiotia 16/04/1965 1000 Stenophylax permistus 1 0 J.M. Vitoria
Cueva Saldias Gaztelu 17/07/1998 300 Stenophylax mitis 1 1 O. Escolà
Cueva Saldias Gaztelu 13/07/1998 300 Stenophylax mitis 4 0 O. Escolà
Cueva Saldias Gaztelu 13/07/1998 300 Stenophylax permistus 1 0 O. Escolà
Sima Agarre-Erdikoa Ibarranguelu 03/05/1969 350 Stenophylax fissus 1 0 sense dades
Sima Lezandi Lemoa 20/06/1962 1091 Stenophylax fissus 2 2 I. Gonzàlez
Massís Galaic
Cueva del rey Cintolo Mondoñedo 28/06/1974 280 Stenophylax fissus 0 1 Rosaura Illes Balears
Illes Balears
Avenc des Gel Escorca 08/12/1976 1070 Mesophylax aspersus 1 0 O. Escolà
Cova gran des Robiols Puigpunyent 07/12/1976 309 Mesophylax aspersus 1 1 O. Escolà Murcia
Llevant peninsular
Cueva del Puerto Calasparra 13/04/1979 503 Mesophylax aspersus 0 1 P. Cantons
Cueva del Puerto Calasparra 13/04/1974 503 Stenophylax malatestus 1 0 J. Comas Navarra
Pirineu axial
Cova de Benasa Roncal-Salazar 13/08/2019 693 Stenophylax sequax 2 0 Sendra, Pallisé
Cova Lezegalde Iribas 14/10/1984 650 Stenophylax sequax 0 1 O. Escolà
Cova Orobe num. 2 Olazagutia 16/07/1998 830 Stenophylax mucronatus 0 1 Salgado, Fresneda, Escolà
Cova Orobe num. 2 Olazagutia 16/07/1998 830 Stenophylax sequax 4 4 Salgado, Fresneda, Escolà
Cueva del Ibon de Isaba Otxindundua 26/06/1982 833 Stenophylax vibex 1 0 O. Escolà
Sima Ormazarreta I Aralar 30/08/1984 1200 Stenophylax mucronatus 0 2 Auroux, Macià
Sima Ormazarreta I Aralar 30/08/1984
Sima Urbasa III Comunal de Urbasa 27/07/1984
Sima Urbasa V Comunal de Urbasa 27/07/1984
Sima Urbasa V Comunal de Urbasa 25/07/1984
Sima Urbasa VII Urdiain 27/07/1984
1200 Stenophylax testaceus 1 2 Ll. Auroux
500 Stenophylax nycterobius 1 0 F. Pauné
500 Stenophylax mucronatus 1 0 M. Nebot
500 Stenophylax permistus 0 1 M. Nebot
500 Stenophylax vibex 0 1 C. Marin
País Valencià
Llevant peninsular
Avenc de les Casetes Vallibona 10/10/1971 1100 Mesophylax aspersus 5 6 Ll. Filbà
Avenc de les Casetes Vallibona 10/10/1971
1100 Stenophylax fissus 1 0 Ll. Filbà
Avenc de Santa Bárbara Tírig
22/07/1974 590 Mesophylax aspersus 1 0 C. Ribera
Avenc de Santa Bárbara Tirig 14/09/1993 590 Mesophylax aspersus 3 2 O. Escolà
Avenc de Santa Bárbara Tirig 14/09/1993 590 Stenophylax testaceus 0 1 O. Escolà
Avenc del Cap de la Solana Vallibona 18/07/1969 1140 Mesophylax aspersus 1 0 Ll. Filbà
Avenc del Flare Tarbena 21/04/1978 600 Stenophylax fissus 0 1 Crespo, Zaragoza
Avenc del Gos Barx 24/05/1980 400 Stenophylax mucronatus 1 0 Comas, Escolà
Avenc del Pla de l’Om Tirig 14/09/1993 450 Mesophylax aspersus 3 2 O. Escolà
Cova Carrasquilla Enguera 25/05/1980 535 Mesophylax aspersus 1 0 Comas, Escolà
Cova Castellets País Valencià 28/03/1970 Mesophylax aspersus 4 5 I. Gonzàlez
Cova de la Sarsa Bocairent 01/09/1978 700 Stenophylax espanioli 1 0 J. Comas
Cova de l’Alt del Pi Serra 16/04/1965 685 Stenophylax fissus 1 0 I. Gonzàlez
Cova de l’Alt del Pi Serra 08/08/1976 685 Stenophylax mucronatus 4 0 I. Gonzàlez
Cova de Sant Pere Traiguera 08/04/1993 300 Stenophylax mucronatus 0 1 O. Escolà
Cova del Toro Alcudia de Veo 24/04/1977 471 Mesophylax aspersus 1 0 J.V. Gonzalez
Cova dels Encenalls Sant Mateu 07/04/1993 400 Stenophylax malatestus 1 1 O. Escolà
Cova dels Encenalls Sant Mateu 07/04/1993 400 Stenophylax testaceus 0 1 O. Escolà
Cova Font de la Llobera Xixona 09/11/1977 890 Mesophylax aspersus 2 1 O. Escolà
Cova Fosca El Verger 29/04/1978 70 Mesophylax aspersus 0 1 C. Ribera
Cova Joliana Alcoi 15/11/1966 675 Mesophylax aspersus 0 1 Barrachina
Cova Masies d’Abad Les Coves de Vinromà 01/07/1971 200 Mesophylax aspersus 1 1 Viñas, Español
Cova Negra Montanejos 29/06/1973 637 Stenophylax espanioli 2 0 J. Comas
Cova Puntassa Tinença de Benifassà 23/12/1993 1300 Mesophylax aspersus O. Escolà (Coratxà)
Cova Santa La Serra d’En Galceran 26/05/1980 550 Mesophylax aspersus 0 1 Comas, Escolà
Cova Tornera Vallibona 11/10/1971 650 Mesophylax aspersus 3 2 Ll. Filbà
Cueva de Confrides Confrides 31/10/1977 875 Mesophylax aspersus 4 0 O. Escolà
Cueva del Torcacho Iglesuela del Cid 20/03/1970 1292 Mesophylax aspersus 0 1 F. Fadrique
Cueva Oscura Atzeneta del Maestrat 03/05/1998 170 Mesophylax aspersus 1 0 F. Fadrique
Cueva Somo Tárbena 30/04/1978 853 Mesophylax aspersus 0 1 Blas, Ribera i Zaragoza
Cueva Somo Tárbena 30/04/1978 853 Stenophylax fissus 1 0 Blas, Ribera, Zaragoza .
Clàudia Pla-Narbona*, Andreu Ubach*, Constantí Stefanescu*
* Museu de Ciències Naturals de Granollers. Grup de Recerca en Lepidòpters - Biodiversitat i Bioindicadors (BiBio). C/ Francesc Macià, 51. 08402 Granollers. A/e: cplanarbona@mcng.cat
Rebut: 21.02.2024; Acceptat: 23.05.2024; Publicat: 30.06.2024
Resum
El mud-puddling, o beure del fang, és un comportament que tenen algunes papallones i altres insectes d’adquirir nutrients que es troben a tolls enfangats, marges de rieres, fruita podrida, excrements, carronya i, fins i tot, suor. Aquest comportament, que produeix un efecte crida que genera «abeuradors de papallones», ha estat molt estudiat en zones tropicals, mentre que en latituds temperades és poc conegut. El treball que es presenta és un estudi sobre aquest comportament en dues localitats de Catalunya, situades a la regió alpina i la regió mediterrània humida. Aquest treball combina censos de papallones diürnes en els abeuradors escollits amb dades del Programa de seguiment de papallones de Catalunya (CBMS) i l’ús de càmeres de fototrampeig durant l’estiu del 2022. En els censos s’han observat 1462 individus de 70 espècies, pertanyents sobretot als licènids i hespèrids. Hem detectat també que els licènids, hespèrids i pièrids prefereixen beure del fang i d’excrements, mentre que els nimfalins i satirins combinen beure del fang i d’exsudats d’arbres o flors. Els resultats també mostren que els mascles joves són els que més duen a terme aquest comportament, que hi ha un pic d’activitat a mig matí a totes dues localitats i que l’activitat augmenta amb la temperatura exceptuant temperatures extremes (38-40 ºC). L’estudi destaca mancances de coneixement entorn a aquest comportament en latituds temperades que caldria tenir en compte en futurs treballs, com la funció dels abeuradors en períodes de sequera i la taxa de depredació que hi pateixen les papallones.
Paraules clau: Abeurador, mud-puddling, beure del fang, papallones diürnes, comportament, càmeres de fototrampeig, regió mediterrània, regió alpina, temperatura, precipitació, patró d’activitat.
Abstract
Mud-puddling: A study of butterfly puddling sites in Catalonia
Some butterflies and other insects acquire nutrients from puddles, stream banks, rotting fruit, carrion, excrement and even sweat. This behaviour, which is called mud-puddling, produces a calling effect generating butterfly clusters. It has been widely studied in tropical areas, however, there are still many gaps in knowledge in temperate latitudes. The work presented is a study of this behaviour in two locations in Catalonia, one in the Alpine region and another in the humid Mediterranean region. This work combines censuses of diurnal butterflies in the chosen puddles with data from the Catalan Butterfly Monitoring Scheme (CBMS) and the use of photo-trapping cameras, during the summer 2022. In the censuses, 1462 individuals of 70 species, mainly belonging to Lycaenidae and Hesperiidae, have been observed. Also, Lycaenidae, Hesperiidae and Pieridae prefer puddles and excrement, while Nymphalinae and Satyrinae combine puddles and flora exudates. Our results also show that the participants are mainly young males, that there is an activity peak between 12 a.m. and 2 p.m. in both localities and that butterflies’ activity leaking from puddles increases with the temperature except for extreme temperatures (38-40 ºC). The study highlights many gaps in knowledge related to this behaviour in temperate latitudes that should be taken into account in future works: such as puddles function in drought periods and predation rate suffered by butterflies.
Key words: Puddle, mud-puddling, butterflies, behaviour, photo-trapping cameras, Mediterranean region, Alpine region, temperature, precipitation, activity pattern.
Introducció
En el context de canvi global actual i per a la conservació de les espècies, és necessari conèixer a fons aspectes de la seva ecologia que els són crucials per a la supervivència. En aquest sentit, l’estudi de la disponibilitat de recursos i com són usats en diferents ambients i regions climàtiques del territori és molt rellevant. Les papallones diürnes, concretament, poden veure limitada l’obtenció de recursos de nèctar i
d’altres tipus en condicions de sequera, una situació cada cop més freqüent a causa del canvi climàtic (Phillips et al. 2018). El mud-puddling, o beure del fang, és el comportament que tenen algunes papallones, i altres insectes, d’adquirir nutrients que es troben a tolls enfangats, marges de rieres, fruita podrida, excrements, carronya i, fins i tot, suor (Molleman 2010). Aquest comportament produeix un efecte crida que provoca que en algunes zones es formin agrupacions d’individus, de vegades força nombroses, anomenades

«abeuradors de papallones» (Fig. 1). Aquestes agrupacions, quan estan formades per dos o més individus de la mateixa espècie, s’anomenen gremis (Prokopy & Roitberg 2001). Les primeres observacions d’abeuradors de papallones van ser publicades el 1863 per Henry W. Batesa a l’Amazònia, però no va ser fins més d’un segle més tard que es va dur a terme el primer experiment per estudiar-los (Arms et al. 1974).
Des d’aleshores, diferents estudis coincideixen en què són els mascles joves els que mostren més aquest comportament, tot i que també s’ha observat en individus vells i, més ocasionalment, en femelles (Boggs & Jackson 1991). També coincideixen en què el principal objectiu del mud-puddling que duen a terme els mascles és obtenir sals minerals i aminoàcids, escassos en les plantes i el nèctar de què s’alimenten les erugues i les papallones adultes, respectivament. Aquests seran després transmesos a les femelles en el moment de la còpula en forma de regal nupcial, i així augmentaran la seva fertilitat i longevitat (Pivnick & McNeil 1987). Aquest comportament és molt típic a les zones tropicals, on és comú trobar grans concentracions d’algunes espècies de papallones a les ribes dels rius, també en fonts geotermals (Phon et al. 2017) o inclús en platges (Tea et al. 2020). A regions temperades i àrides és un fenomen habitual però menys estudiat. A Catalunya, en particular, és molt notori als ambients pirinencs. A banda dels abeuradors de fang, a casa nostra també s’han observat individus bevent d’excrements, fruita madura i exsudats d’arbres o flors. La preferència per cada substrat varia segons la família i l’espècie (Vila, Stefanescu & Sesma 2018). Resten, però, molts buits de coneixement respecte l’ecologia d’aquest comportament, com per exemple quins costos ecològics té (p. ex. risc de predació), per quin motiu hi ha espècies que ho fan amb més freqüència que d’altres, perquè algunes espècies no ho fan mai o perquè les preferències de substrat i la tendència a formar gremis varien entre famílies i entre espècies. A més, tot i que s’ha suggerit que el mud-puddling en les regions àrides podria haver sorgit de la necessitat d’obtenir aigua (Adler & Pearson 1982), es desconeix si la tendència a exercir aquest comportament incrementa en episodis severs de sequera.
Per aprofundir en aspectes poc coneguts d’aquest comportament, hem dut a terme un estudi de les comunitats de papallones que trobem en abeuradors a dos punts del territori català. Els objectius del treball són, per una banda, estudiar la composició i demografia de les comunitats de papallones que beuen del fang en latituds temperades i, per una altra, observar com les condicions climàtiques afecten l’activitat en els abeuradors de papallones. Les nostres hipòtesis en relació amb aquests objectius són: (1) tal com passa a les regions tropicals, la majoria d’individus que beuen del fang a la regió temperada són mascles joves (Boggs & Jackson 1991); i (2) la temperatura i la precipitació tenen un paper rellevant a l’hora de regular aquesta activitat: esperem una major activitat a majors temperatures i, pel que fa a la precipitació, entenent les diferències en la disponibilitat d’aigua entre regions, esperem més activitat just després de la precipitació a la regió més seca degut a una major disponibilitat d’aigua als abeuradors, mentre que a les regions alpines, just després de la precipitació, esperem menys activitat per la creació espontània de més abeuradors que provocarà un efecte de dilució al nostre abeurador d’estudi.
Material i mètodes
Localitats d’estudi
Per a poder obtenir dades d’activitat bevent del fang i relacionar-les amb diferents condicions ambientals hem seleccionat dues localitats on hi ha estacions del programa Catalan Butterfly Monitoring Scheme (CBMS; www.catalanbms. org), per a les quals disposem d’informació molt completa sobre les comunitats de papallones, i que estan situades en dues regions climàtiques diferents. Les localitats són: (1) Gerri de la Sal (CBMS-85, Pallars Sobirà), situada al Prepirineu a 593 m d’altitud en un ambient que correspon a la regió climàtica mediterrània humida, on es realitza un seguiment de papallones des de l’any 2006 en un transsecte de 1468 m; i (2) Capçaleres del Catllar (CBMS-181, Ripollès), al Pirineu, a 1502 m d’altitud, en un ambient que correspon a la regió climàtica alpina, on s’hi fan censos des del 2020 en un transsecte de 1492 m (Fig 2). La delimitació de les regions

2. Mapa de Catalunya amb les regions climàtiques utilitzades per a l’estudi i les localitats d’estudi. A la dreta, el recorregut de cadascun dels itineraris CBMS dins de les localitats d’estudi i la ubicació exacta de l’abeurador on s’ha fet l’estudi.
climàtiques s’ha fet a partir del nombre d’hores per any en què se supera la temperatura de 21 ºC (21DDG), seguint un criteri (regió alpina i subalpina: ≤ 0-150 DDG; regió mediterrània humida: 150-400 DDG; regió mediterrània àrida: ≥ 400 DDG) i usant les dades proporcionades pel Servei Meteorològic de Catalunya (www.meteocat.cat). Aquestes regions es corresponen amb força exactitud amb les regions bioclimàtiques definides per Metzguer et al. (2013).
En aquestes estacions s’hi duen a terme comptatges de papallones seguint la metodologia BMS (Pollard & Yates 1993), la qual parteix de recomptes visuals d’exemplars adults de papallones diürnes al llarg d’un transsecte en què es compten les papallones que estan a una distància de 5 m per davant i als costats de l’observador. El transsecte consta de diverses seccions corresponents a hàbitats diferents i es recorre un cop per setmana, de l’1 de març al 30 de setembre, a una velocitat constant.
En cadascuna de les estacions s’han detectat zones propícies per al comportament objecte d’estudi a partir de l’observació de concentracions regulars de papallones bevent del fang en diferents anys. En tots dos casos són marges d’un riu que transcorre a través de l’itinerari. La zona d’estudi en ambdós casos ha estat una parceŀla (o abeurador) d’entre 3 i 5 m2 de sòl nu, amb predomini de pedres i fang.
Treball de camp: censos de papallones bevent del fang
Per tal d’estudiar la comunitat de papallones de cada localitat, s’han realitzat censos en els abeuradors de cada localitat entre el 9 de juny i el 24 d’agost de 2022 al Catllar, i entre el 10 de juny i el 25 d’agost de 2022 a Gerri de la Sal. Els censos es van fer entre les 10 i 16 h, coincidint amb el període de màxima activitat de papallones, cada 15 dies aproximadament, amb un total de sis censos per localitat. Cada cens es va dividir en quatre mostrejos de 15 minuts, cada dues hores, per tal de recollir els canvis d’abundància i riquesa al llarg del dia. A cada mostreig es van anotar les condicions climàtiques, el nombre d’individus que visitaven l’abeurador, la seva espècie, el seu sexe (femella, mascle o indeterminat), el desgast alar com a aproximació de l’edat (adults nous o E1, adults intermedis o E2, i adults gastats o E3) i quins formaven gremis, entenent per gremi les agrupacions de dos o més individus dins de l’abeurador, ja siguin de la mateixa o de diferents espècies.
Treball de camp: registres d’abundància a partir del fototrampeig
A cada localitat es va instaŀlar una càmera de fototrampeig model scout guard sentinel II 20 MP, marca ARCEA, enfocant l’abeurador i disparant fotografies amb una freqüència de 30 minuts, durant el període de temps que van durar els censos (12 setmanes). Posteriorment, les imatges obtingudes s’han analitzat manualment anotant l’abundància de papallones de cada imatge juntament amb la localitat, data, hora, nuvolositat i presència d’altres animals o persones. La identificació de les espècies no ha estat possible en la majoria de casos però sí l’abundància d’individus com a aproximació a l’activitat de l’abeurador.
Les dades d’activitat obtingudes ens han servit per interpretar els patrons d’activitat estacional i diària. Hem definit 19 franges horàries, de 30 minuts cadascuna, de 9:00 a 18:00. Per definir aquests patrons hem eliminat els comptatges de les imatges en què no hi havia sol, ja que l’activitat de les papallones disminueix ostensiblement en aquest cas. Pel mateix motiu, hem eliminat aquelles dades en què hi havia altres animals o en què es van fer mostrejos, ja que poden haver fet variar l’activitat a l’abeurador.
Inicialment vam decidir enregistrar la temperatura amb sensors iButton instaŀlats en llocs ombrívols a prop de la càmera, però el de Gerri de la Sal el vam perdre i el del Catllar va quedar sense memòria a la meitat del mostreig (setmana 7). Si bé és cert que les càmeres de fototrampeig registren la temperatura juntament amb la imatge, les mateixes càmeres marquen una temperatura molt més alta que la real a degut al sobreescalfament causat pel sol i per la pròpia càmera. Així doncs, davant la manca de dades completes dels sensors de temperatura, hem hagut d’utilitzar altres fonts d’informació meteorològica.
D’entre les estacions meteorològiques més properes, s’han establert correlacions de Pearson per determinar la més fiable d’acord amb els registres obtinguts per les càmeres de fototrampeig, i també l’iButton en el cas del Catllar, i s’han escollit les que tenien un valor més alt de correlació. Per al cas de Gerri de la Sal, hem seleccionat l’estació meteorològica automàtica (www.meteo.cat) de la Pobla de Segur (r= 0.644, n = 1413, p- valor <2e-16), que és a 10 km de l’abeurador i, per al cas del Catllar, l’estació local de la vall del Catllar (r= 0.872, n = 641, p-valor < 2e-16 per la sèrie incompleta de dades de l’iButton i r=0.902, n = 1413, p-valor < 2e-16 per les dades de la càmera), que es troba a menys d’1 km de l’abeurador. A banda de la temperatura, aquestes estacions meteorològiques també ens aporten dades de precipitació per a les dues localitats (PPT, en mm).
Anàlisi de dades
Per tipificar i comparar les comunitats de papallones associades als dos abeuradors estudiats s’han utilitzat les dades dels censos directes. Hem calculat l’abundància i la riquesa de cada localitat i hem testat si hi ha diferències a partir d’una anàlisi ANOVA. També, per a cada localitat, hem elaborat testos U-Mann Whitney per comprovar si les espècies més comunes a la zona (segons dades del CBMS a cada localitat) són també les més abundants als abeuradors. Això ens ha permès avaluar quines espècies són més propenses a beure del fang i quines, contràriament, no mostren aquest tipus de comportament. Aquesta comparativa s’ha fet restringint les dades dels censos del CBMS al període juny-agost del 2022 dels itineraris CBMS-85 i CBMS-181, i eliminant les dades de la secció on hi ha l’abeurador per evitar pseudoreplicació. Per tal d’analitzar les diferències entre sexes i edat hem fet un model lineal generalitzat mixt (GLMM) en què la variable resposta era el nombre d’individus, els factors fixos el sexe,
Taula 1. Llista de papallones que beuen del fang en les dues localitats mostrejades. Els percentatges de sexes (mascle (M), femella (F) o indeterminat (indet.)), i els estats alars (joves (E1), entremig (E2) i vells (E3)), estan calculats sense distingir entre localitats. Les famílies o subfamílies diferenciades són els hespèrids (HES), libiteíns, (LIB), licènids (LYC), nimfalins (NYM), pièrids (PIE), papiliònids (PAP) i satirins (SAT).
l’estat alar i la seva interacció, i el factor aleatori l’espècie, per tal de controlar el biaix que es produeix entre els individus de cada espècie. S’ha realitzat un model per a cada localitat per tal de reduir la complexitat del model.
La influència de les variables ambientals (meteorològiques i horàries) s’ha estudiat amb les dades d’abundància proporcionades per les càmeres de fototrampeig, a partir de models GLMM. En aquests models, el període temporal que explica la variació en l’abundància de papallones al llarg de la temporada, és a dir la fenologia, s’ha considerat com un factor aleatori. En cada localitat, s’ha ajustat un model GAM a la corba d’abundància del total de les espècies detectades i s’han identificat tres períodes que reflecteixen moments fenològics amb màximes abundàncies de papallones.
Hem dut a terme dos models GLMM: (1) En un primer model hem analitzat els patrons d’activitat en funció de la franja horària i de la temperatura, utilitzant com a variable resposta l’abundància d’individus observada a les fotos de fototrampeig, com a variables predictives la franja horària (1-19), el valor quadràtic de la franja horària, la temperatura, la localitat i la interacció d’aquestes dues últimes variables, i com a factor aleatori el període fenològic. (2) En un segon model hem analitzat com es veu afectada l’activitat diària als abeuradors en funció de la temperatura i la precipitació recent. En aquest model la variable resposta ha estat la màxima abundància enregistrada en un cens diari, les variables predictives la temperatura màxima del dia, la precipitació acumulada (PPT) del mateix dia, la PPT del dia anterior i la localitat, i com a factor aleatori el període fenològic. Per a tots dos models s’han fet servir les dades d’abundància del 10 de juny al 24 d’agost en el cas del Catllar, i del 28 de juny al 25 d’agost en el cas de Gerri de la Sal, i s’han redimensionat els valors de les variables.
L’anàlisi de les dades s’ha realitzat amb R versió 4.3.1. (R Core Team, 2023). S’han utilitzat la funció aov del paquet stats per a l’anàlisi ANOVA, el paquet mgcv (Wood, 2021) per a l’anàlisi GAM, el paquet lme4 (Bates, 2023) per a l’anàlisi GLMM, la funció cor del paquet stats per calcular les correlacions entre les diferents variables i el paquet ggplot2 (Wickham, 2023) per a l’elaboració de gràfics.
Resultats i discussió
Les comunitats de papallones que beuen del fang
Visió general
En els sis censos fets durant els mesos de juny, juliol i agost a les localitats del Catllar (regió alpina) i Gerri de la Sal (regió mediterrània humida) hem enregistrat 1462 individus de 70 espècies diferents bevent del fang. Al Catllar hem observat 819 individus i a Gerri de la Sal 643 individus, que corresponen a un 61 % (39 espècies) i un 78 % (53 espècies) de les espècies presents a la zona segons dades del CBMS en el mateix període de temps, respectivament (Taula 1). No hi ha diferències significatives pel que fa a l’abundància (n = 509, p-valor = 0.231) ni la riquesa (n = 12, p-valor = 1) entre les dues regions climàtiques segons les dades dels censos, però sí que s’ha observat una major activitat a la regió alpina
Taula 2. Llista d’espècies de la comunitat de papallones de l’estació del CBMS amb les seves abundàncies calculades fent la mitjana de juny a agost del 2022, i llista d’espècies censades a l’abeurador de la localitat amb la seva abundància en el mateix període a l’Estació CBMS-181, Catllar. Les famílies o subfamílies diferenciades són els LYC: licènids; NYM: nimfàlids; PIE: pièrids; SAT: satírids; HES: hespèrids; PAP: papiliònids.
Espècie
Família Abu. cens Abu. abeuradors CBMS-181
Taula 3. Llista d’espècies de la comunitat de papallones de l’estació del CBMS amb les seves abundàncies calculades fent la mitjana de juny a agost del 2022, i llista d’espècies censades a l’abeurador de la localitat amb la seva abundància en el mateix període a l’Estació CBMS-85, Gerri de la Sal
Les famílies o subfamílies diferenciades són els LYC: licènids; NYM: nimfàlids; PIE: pièrids; SAT: satírids; HES: hespèrids; PAP: papiliònids
Espècie Família Abu. cens Abu. abeurador CBMS-85
Carcharodus alceae HES 2 0
Erynnis tages HES 0 1
Hesperia comma HES 13 2
Pyrgus sp. (P. carthami i P. malvoides) HES 197 99.5
Thymelicus sylvestris HES 11 0
Libythea celtis LIB 8 5.5
Aricia agestis LYC 1 0
Aricia cramera LYC 0 2
Celastrina argiolus LYC 0 6
Cupido alcetas LYC 1 0
Cupido argiades LYC 2 0
Cupido minimus LYC 0 0.5
Cupido osiris LYC 5 2.5
Cyaniris semiargus LYC 1 0
Favonius quercus LYC 1 2
Glaucopsyche alexis LYC 0 0.5
Laeosopis roboris LYC 0 2
Lampides boeticus LYC 1 5.5
Leptidea sinapis LYC 6 0
Leptotes pirithous LYC 2 0
Lysandra bellargus LYC 17 19
Lysandra coridon LYC 155 274
Plebejus argus LYC 28 28
Polyommatus amandus LYC 3 2
Polyommatus damon LYC 2 0
Polyommatus escheri LYC 44 20
Polyommatus icarus LYC 15 14.5
Polyommatus thersites LYC 1 3
Satyrium esculi LYC 5 18
Thecla betulae LYC 0 2
Aglais io NYM 3 0
Apatura ilia NYM 5 15.5
Argynnis adippe NYM 2 2
Argynnis aglaja NYM 3 2
Argynnis niobe NYM 0 2
Argynnis paphia NYM 1 4
Argynnis sp. NYM 1 8
Boloria dia NYM 7 21
Brenthis daphne NYM 1 0
Limenitis camilla NYM 0 4
Limenitis reducta NYM 14 13.5
Melitaea cinxia NYM 0 0.5
Melitaea deione NYM 0 0.5
Melitaea diamina NYM 1 0
Melitaea parthenoides NYM 1 0.5
Melitaea phoebe NYM 2 2
Melitaea sp. NYM 3 4
Melitaea trivia NYM 0 8
Nymphalis polychloros NYM 4 2.5
Polygonia c-album NYM 4 7
Vanessa atalanta NYM 3 0
Vanessa cardui NYM 0 1
Iphiclides feisthamelii PAP 6 49.5
Papilio machaon PAP 2 2
Anthocharis cardamines PIE 0 0.5
Anthocharis euphenoides PIE 0 3.5
Aporia crataegi PIE 0 0.5
Colias alfacariensis PIE 0 2
Colias crocea PIE 0 12
Gonepteryx cleopatra PIE 1 6.5
Gonepteryx sp. PIE 2 6
Leptidea sinapis PIE
Pieris brassicae PIE
Pieris ergane PIE
Pieris napi PIE 2 1.5
Pieris rapae PIE 1 26
Arethusana arethusa SAT 1 0
Brintesia circe SAT 0 93
Coenonympha arcania SAT 11 20
Coenonympha dorus SAT 8 0
Coenonympha pamphilus SAT 0 6.5
Hipparchia fagi SAT 0 4
Hipparchia fidia SAT 3 11
Hipparchia hermione SAT 9 21.5
Espècie
Família Abu. cens Abu. abeurador CBMS-85
Hipparchia statilinus SAT 0 7.5
Lasiommata megera SAT 0 5.5
Maniola jurtina SAT 0 7
Melanargia lachesis SAT 3 44
Pararge aegeria SAT 0 8
Pyronia bathseba SAT 2 18
Pyronia cecilia SAT 0 10
Pyronia tithonus SAT 6 12
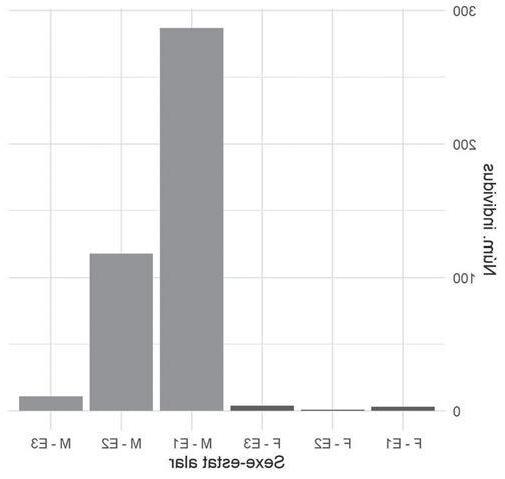
Figura 3. Observacions de papallones bevent del fang en funció del sexe (F-femelles, M-mascles) i l’estat alar (E1, E2, E3).
(Catllar) respecte a la regió mediterrània humida (Gerri de la Sal) pel que fa a les dades de les càmeres (vegeu secció «Patrons d’activitat i efectes climàtics»).
L’abundància de les espècies presents a les dues localitats (dades del CBMS) està fortament correlacionada amb la de les espècies censades als abeuradors (W=1300, pvalor=6.93e-8 al Catllar; W= 2527.5, p-valor=0.0013 a Gerri de la Sal), de manera que aquelles espècies que apareixen en major nombre als abeuradors són també les més abundants a la zona (Taula 2-3, Fig. 3). Les famílies que més beuen del fang són els licènids, hespèrids i nimfàlids, tant en nombre d’espècies com en nombre d’individus. Aquestes representen un 95,4 % de l’abundància (782 individus) i un 79 % de les espècies (30 spp.) a l’abeurador del Catllar, i un 88 % de l’abundància (568 individus) i 70 % de les espècies (37 spp.) al de Gerri de la Sal (Taula 2-3).
Al Catllar, les espècies amb una major presència als abeuradors són els licènids Cupido minimus (Füssly 1775) (n = 166), Lysandra coridon (Poda 1761) (n = 125), Lysandra bellargus (von Rottemburg 1775) (n = 97) i Aricia agestis (Denis & Schiffermüller 1775) (n = 56), l’hespèrid Thymelicus sylvestris (Poda 1761) (n = 33) i diverses espècies del gènere Pyrgus (Pyrgus serratulae (Rambur 1839), P. alveus (Hübner 1802) i P. malvoides (Elwes & Edwards 1897) (n = 69)).
També hi ha una abundància destacable del pièrid Leptidea
reali (Reissinger 1990) (n = 21). A Gerri de la Sal destaquen les espècies del gènere Pyrgus (n = 197), L. coridon (n = 155), Polyommatus escheri (Hübner 1823) (n = 44), Plebejus argus (Linnaeus 1758) (n = 25) i L. bellargus (n = 17). En totes dues localitats s’han observat força espècies de nimfàlids (13 i 14, al Catllar i a Gerri de la Sal, respectivament). A més, a Gerri de la Sal hi ha una proporció més destacable de satírids (8 espècies) i papiliònids (2 espècies) que al Catllar, on s’han observat 5 espècies de satírids i cap papiliònid (Taula 1).
També, cal destacar que hi ha espècies que han aparegut bevent del fang però que no han aparegut en els censos CBMS, com per exemple Plebejus argus (Linnaeus 1758) o Carcharodus alceae (Esper 1780) al Catllar (Taula 2) o Thymelicus sylvestris (Poda 1761), Leptidea sinapis (Linnaeus 1758) o Coenonympha dorus (Esper 1782) a Gerri de la Sal (Taula 3). Això reforça el fet que els abeuradors són punts d’atracció importants per a la fauna de la zona, fins al punt que hi apareixen espècies que no queden registrades als censos BMS.
Beure d’altres substrats
Hi ha espècies que, tot i ser presents i abundants a la zona (segons censos CBMS), no han acudit als abeuradors durant el nostre estudi, per exemple Pararge aegeria (Linnaeus 1758) i Argynnis paphia (Linnaeus 1758) al Catllar (Taula 2a) i Brintesia circe (Fabricius 1775) a Gerri de la Sal (Taula 2b). Tot i això, aquestes tres espècies s’han observat en repetides ocasions en abeuradors d’altres localitats. Sense anar més lluny, A. paphia s’ha observat a l’abeurador de Gerri de la Sal. El comportament de mud-puddling també es duu a terme en altres substrats. Durant els censos hem observat diverses espècies de nimfàlids bevent dels exsudats de l’escorça de salzes, entre les quals hi ha Nymphalis polychloros (Linnaeus 1758), Apatura ilia (Denis & Schiffermüller 1775), Polygo-
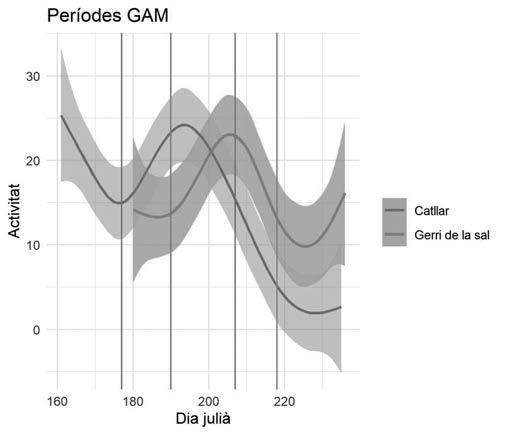
Figura 4. Gràfic iŀlustrant la corba d’abundància a partir del model GAM a les dues localitats. En vertical, les marques que separen cadascun dels 3 períodes a cada localitat, amb el color corresponent.
nia c-album (Linnaeus 1758), Limenitis reducta (Staudinger 1901) i Vanessa atalanta (Linnaeus 1758), i també diverses espècies de satírids fent el mateix, com B. circe, P. aegeria, Hipparchia hermione (Linnaeus 1764) i Maniola jurtina (Linnaeus 1758). També s’ha observat que P. c-album bevia dels exsudats de les flors dels cards (Oro et al. 2023) i s’han vist licènids com L. bellargus i L. coridon bevent d’objectes personals dels observadors, com la carpeta, un llibre o la cantimplora. Per últim, els excrements també són un substrat utilitzat per alguns pièrids (L. reali), alguns nimfalins (Melitaea sp.), alguns satirins (Erebia sp.) i alguns hespèrids (Pyrgus sp., Thymelicus acteon (von Rottemburg 1775), Carcharodus lavaterae (Esper 1783), Erynnis tages (Linnaeus 1758)), i per gairebé totes les espècies de licènids registrades en les nostres observacions.
Taula 4. Resultats del model GLMM: núm. individus ~ sexe*estat alar + (1|Espècie); en negreta s’indiquen els p-valors significatius.
Taula 5. Resultats dels 2 models GLMM elaborats indicant la fórmula i el p-valor de les variables testades en cada cas. En negreta s’indiquen els p-valors significatius.
Model 1 Model 2
Abundància ~ Temp. Mitj * Abundància màx ~ Temp. Localitat + Franja + Màx + PPT mateix dia + PPT Franja2 + (1|Període) dia anterior + Localitat + (1|Període)
Variables Coeficient p-valor Coeficient p-valor
Temperatura 0.652 < 2e-16
6.18e-11 PPT mateix dia – – -0.148 0.017 PPT dia anterior – -0.167 0.008 Franja horària -0.034 0.058 – –Franja2 -0.293 < 2e-16 – –Localitat -0.821 0.031 -1.322 0.011 Localitat*Temp. -0.284 6.32e-8
Segons les nostres observacions, sembla que en latituds temperades hi ha famílies que prefereixen beure del fang i d’excrements, com els licènids, hespèrids i pièrids, mentre que en els nimfalins i satirins les preferències per beure del fang o d’exsudats d’arbres i flors estan més equilibrades.
Els gremis
Burger & Gotchfield (2001) van suggerir que les papallones poden seguir dues estratègies a l’hora de beure del fang: (1) agrupar-se en grups grans o gremis per tal de reduir el risc de depredació individual, o (2) distanciar-se per aprofitar la
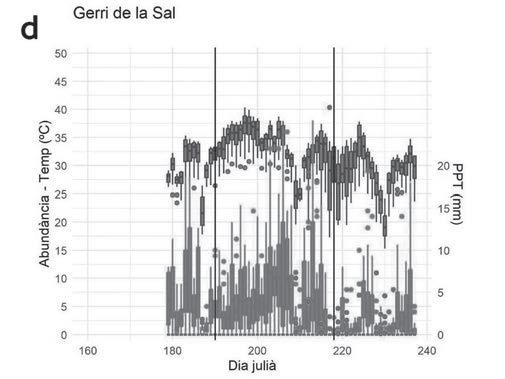
Figura 5. Diagrames de caixes que mostren els patrons diari (per franja horària, a-b) i estacional (per dia, c-d) d’activitat als abeuradors segons les imatges de fototrampeig: en blau, les dades del Catllar i, en verd, les de Gerri de la Sal. En els gràfics també s’hi representa la temperatura (ºC) i la precipitació diària acumulada (PPT, mm). Les franges horàries a-b) van de la 1 (9:00) a la 19 (18:00), i els períodes fenològics c-d) apareixen indicats amb marques negres verticals.
seva coloració críptica. En el nostre estudi, les famílies que creen gremis, o agrupacions de més de dos individu, són majoritàriament els licènids (sobretot L. coridon, C. minimus, L. bellargus) i els hespèrids (sobretot el gènere Pyrgus), així com gèneres concrets de nimfalins (Melitaea sp.) (>20 individus) i alguns pièrids (Pieris ergane (Geyer 1828) i L. reali), que formen gremis molt menors en nombre d’individus (4-12 individus). S’han observat puntualment alguns satirins fent gremis de pocs individus (2 o 3), però en poques ocasions. L’efecte del sexe i l’estat alar
Les nostres observacions confirmen un cop més que el comportament de beure del fang el duen a terme preferentment els mascles (p = 0.018), majoritàriament quan no són vells (p = 0.007) (Fig. 3, Taula 4). No obstant això, ocasionalment també hem observat algunes femelles (Erebia meolans (von Prunner 1798), n=2 E2 i E3; o Gonepteryx sp. (Leach 1815), n=2 E1 i E2) i mascles vells (Cupido osiris (Meigen 1829) o Polyommatus icarus (von Rottemburg 1775); Taula
Butlletí ICHN 88 (2), 2024
1) als abeuradors, tal com afirmen altres estudis (Sculley & Boggs 1996, Boggs & Jackson 1991).
Patrons d’activitat i efectes climàtics
Patrons d’activitat
Les dades recollides amb les càmeres de fototrampeig ens han permès observar el patró d’activitat diari (Fig. 5a-b) i estacional (Fig. 5c-d) de les papallones que visiten els abeuradors en les dues localitats estudiades. L’activitat en els abeuradors al llarg del dia segueix una distribució unimodal, de manera que el pic d’activitat es dona al migdia, entre les 11:30 i 15:00 (franges 6-12, Fig. 5a) al Catllar, i entre les 12:00 i 14:00 (franges 7-10, Fig. 5b) a Gerri de la Sal, i, tot i que després disminueix, no ho fa dràsticament en cap de les dues localitats. Això queda reflectit al model 1 (Taula 5), en què és el valor quadràtic de la franja horària el que té un efec-
te significatiu sobre l’activitat però no la franja horària. També hem observat més activitat a la regió alpina (Catllar) que a la regió mediterrània (Gerri de la Sal) en tots dos models (p = 0.031 model 1 i p = 0.011 model 2, Taula 5).
Pel que fa a l’activitat al llarg de la temporada d’estudi hem distingit tres períodes fenològics a cadascuna de les localitats. Els períodes resultants del model GAM per a cada localitat han estat per al Catllar: el període 1 els dies julians 161-177, el període 2 els dies 178-207 i el període 3 els dies 208-237; i per a Gerri de la Sal: període 1 els dies 180-190, el període 2 els dies 191-217 i el període 3 els dies 218-237 (Fig. 4, Fig. 5c-d). Aquests períodes reflecteixen moments fenològics amb màximes abundàncies de papallones al llarg de la temporada. Són més abundants els comptatges en el segon període, que comprèn del 27 de juny (dia 178) al 26 de juliol (dia 207, Fig. 5c) per al Catllar i del 10 de juliol (dia 171) al 5 d’agost (dia 217, Fig. 5d) per a Gerri de la Sal. En aquest darrer cas, les dades són més escasses degut a la pèrdua de la informació de les primeres dues setmanes de mostreig.
L’efecte de la temperatura
Les anàlisis ens indiquen un efecte significatiu de la temperatura, de manera que a major temperatura l’activitat als abeuradors també augmenta, però no ho fa de la mateixa manera en una regió i a l’altra (Taula 5). A la regió alpina (Catllar) l’activitat augmenta amb la temperatura, mentre que a la regió mediterrània (Gerri de la Sal) aquest efecte positiu de la temperatura es veu atenuat quan aquesta marca valors de 38-40 ºC. En aquests casos l’activitat disminueix o es manté però deixa d’augmentar. Aquestes diferències entre localitats es poden explicar per diversos motius. En primer lloc, les espècies poden tenir adaptacions locals i comportar-se d’una manera diferent en climes més càlids o més freds, tot i ser presents en les dues localitats. En segon lloc, la temperatura màxima assolida és gairebé 10 ºC més alta a Gerri de la Sal que al Catllar (Fig. 5a-b), i aquesta diferència podria provocar una reducció de l’activitat general en aquesta localitat o inclús entrades en diapausa els dies més càlids. Aquests resultats, per tant, confirmen parcialment la nostra hipòtesi inicial segons la qual l’activitat als abeuradors augmenta amb la temperatura (excepte quan les temperatures són extremament altes), un fet que es podria relacionar amb la necessitat de les papallones d’hidratar-se en episodis de molta calor a latituds temperades.
L’efecte de la precipitació
Per altra banda, els dies que plou l’activitat de les papallones als abeuradors disminueix considerablement (p = 0.017) i el mateix passa quan la pluja ha caigut el dia anterior (p = 0.008, Taula 5). Pel que fa a la disminució de l’abundància el mateix dia de la precipitació, aquesta l’associem a la disminució de l’activitat general de les papallones quan plou. En canvi, l’efecte de la pluja del dia anterior el podem associar a diverses casuístiques, com el fet que es creïn altres abeuradors temporals potencialment visitats per papallones de manera que l’activitat es redueix a l’abeurador mostrejat, no
perquè l’activitat general de la zona baixi, sinó per un efecte de dilució en què l’esforç es reparteix entre d’altres abeuradors temporals o en altres substrats. Altrament, la baixada podria ser deguda al fet que no necessiten obtenir l’aigua dels abeuradors perquè que ja n’hi ha de disponible al nèctar de les flors. Una última explicació podria ser que les sals del sòl es dissolen amb la pluja i, en conseqüència, les papallones visiten altres abeuradors més rics en sals, però això no ho hem pogut testar amb el nostre mètode perquè no hem analitzat el substrat de cada abeurador. De cara a futurs estudis, seria interessant comparar i analitzar diferents tipus de substrat. D’altra banda, la baixa quantitat de dades de la precipitació caiguda durant el període mostrejat, sobretot a la regió mediterrània, no ens permet extreure conclusions sòlides respecte a les diferències entre regions. En futurs estudis sobre aquest comportament recomanem dur a terme els mostrejos amb les càmeres de fototrampeig durant tota la temporada de vol de les papallones, de març a setembre, per aprofundir en l’efecte de la precipitació del dia anterior en cadascuna de les regions climàtiques, així com en els patrons d’activitat estacional.
Valoració del mètode d’estudi: l’ús de càmeres de fototrampeig
Podem afirmar que l’ús de càmeres de fototrampeig per estudiar l’activitat de les papallones als abeuradors ens aporta informació que complementa la dels censos i, tot i que és poc precisa, ens aporta una gran quantitat de dades de manera contínua difícil d’obtenir d’altres maneres. Tant és així que hem pogut observar diferències en l’activitat als abeuradors gràcies a les dades de les càmeres, que amb els censos no hem pogut detectar. En aquesta línia, les càmeres de fototrampeig poden servir per documentar i estudiar la depredació en abeuradors de papallones, un aspecte en el qual no hem aprofundit en aquest treball. Les nostres càmeres han captat la presència de possibles depredadors de manera aleatòria i, tot i que no han fotografiat cap depredació evident, han retratat animals (Fig. 6) que poden incloure les papallones a la seva dieta, com ara ocells (Burger & Gotchfield 2001, Stefanescu 2023) i rèptils (observació d’un atac d’una sargantana a una Celastrina argioulus en un abeurador a la Vall de Cabó, 2023; P. Roy, com. pers.). En un futur, seria interessant indagar aquest aspecte fent servir les càmeres de fototrampeig per detectar i gravar depredacions en els abeuradors.
Per tot això, valorem positivament l’ús d’aquesta metodologia sempre que es complementi amb treball de camp destinat a verificar les dades de les càmeres i obtenir més informació sobre les espècies mostrejades. D’altra banda, es podria fer un estudi de mercat comptant amb l’assessorament d’experts per escollir un model de càmera amb alta qualitat d’imatge que s’ajusti a aquest tipus d’estudi, ja que hi ha marge de millora pel que fa a la qualitat d’imatge de les càmeres utilitzades en aquest treball.
Pel que fa als censos, assumim un marge de millora pel fet que la informació obtinguda no és del tot completa, donat que espècies que s’han observat bevent del fang de manera regular en anys anteriors no han estat detectades durant els


nostres censos (p. ex. Pieris napi al Catllar, o papiliònids formant gremis a Gerri de la Sal).
Conclusions
Aquest treball ha servit per ampliar el coneixement del comportament de mud-puddling (que en català hem traduït com a «beure del fang») a latituds temperades. A Catalunya, les famílies de papallones diürnes que més duen a terme aquest comportament són els licènids i els hespèrids, que també són les que tenen més tendència a formar gremis. Els nimfalins i els satirins mostren una major tendència a combinar el beure del fang amb l’aprofitament d’exsudats de l’escorça dels arbres i de les flors de card, i pràcticament totes les famílies poden beure d’excrements. Ara bé, no totes les espècies de les famílies esmentades duen a terme aquest comportament, i se’n desconeixen els motius. També, als abeuradors s’han observat espècies que no han estat detectades pels censos CBMS, la qual cosa reforça la idea que els abeuradors són punts d’atracció importants per a la fauna de la zona. L’activitat observada a l’abeurador ha estat lleugerament major a la regió alpina respecte a la regió mediterrània, mentre que la proporció d’espècies que duu a terme aquest comportament a cadascuna de les regions ha estat força similar. Hem comprovat que són sobretot els mascles joves els que acudeixen als abeuradors, tal com passa en regions tropicals, i que l’activitat segueix una distribució unimodal amb un pic d’activitat a migdia que baixa gradualment a mesura que avança la tarda. La temperatura juga un paper molt important afavorint l’activitat de les papallones als abeuradors, excepte a temperatures extremament altes (38-40 ºC), les quals fan que es redueixi la seva activitat en general. Pel que fa a la precipitació recent, no hem pogut extreure conclusions sòlides sobre el seu efecte en l’activitat dels abeuradors a la regió mediterrània humida per la manca de dades degut a les poques precipitacions ocorregudes durant el període de mostreig.
Malgrat que aquest estudi ha contribuït a conèixer millor el fenomen del mud-puddling a la nostra regió, també posa en evidència buits de coneixement importants com, per exemple, quin efecte té la precipitació del dia anterior en l’activitat dels abeuradors, si aquest comportament pot tenir com a objectiu l’obtenció d’aigua per minimitzar el risc de deshidratació en latituds temperades, quina funció tenen els abeuradors en períodes de sequera a les regions àrides, què porta a les espècies a dur o no a terme aquest comportament, i quina és la taxa de depredació en abeuradors. Tots aquests aspectes constitueixen vies d’investigació per explorar en el futur en relació amb aquest fenomen.
Agraeixo a la Institució Catalana d’Història Natural (ICHN) l’atorgament del premi a la Dona Naturalista 2022 que m’ha permès liderar aquest treball i dur-lo a terme amb el suport de l’equip del Catalan Butterfly Monitoring Scheme (CBMS). També agraeixo el suport en el disseny experimental i el treball de camp a en Mike Lockwood, la Blanca Font i en Josep Piqué. Així mateix, estic agraïda a tota la xarxa de voluntaris del CBMS per nodrir la base de dades que ens permet dur a terme treballs científics d’alta qualitat.
Adler, P. H. & Pearson, D. L. 1982. Why do male butterflies visit mud puddles? Canadian Journal of Zoology, 60: 322-325. Arms, K., Feeny, P. & Lederhouse, R. C. 1974. Sodium: stimulus for puddling behavior by tiger swallowtail. Science, 185: 372-374. Bates, D., Mächler, M., Bolker, B. & Walker, S. 2023. lme4: Linear mixed-effects models using ‘Eigen’ and S4. Versió 1.1-27. Disponible a: https://github.com/lme4/lme4 [Data de consulta: 24 abril 2024].
Bates, H. W. 1863. The Naturalist on the river Amazons, a record of adventures, habits of animals, sketches of Brazilian and Indian life and aspects of nature under the Equator during eleven years of travel. London, J. Murray. Vol 1. 351 p. Boggs, C. L. & Jackson, L. A. 1991. Mud puddling by butterflies is not a simple matter. Ecological Entomology, 16: 123-127.
Burger, J. & Gotchfeld, M. 2001. Smooth-billed ani (Crotophaga ani) predation on butterflies in Mato Grosso, Brazil: risk decreases with increased group size. Behavioral Ecology and Sociobiology, 49: 482-492.
Metzguer, M. J., Bunce, R. G. H., Jongman, R. H. G., Sayre, R., Trabucco, A. & Zomer, R. 2013. A high-resolution bioclimate map of the world: a unifying framework for global biodiversity research and monitoring. Global Ecology and Biogeography, 22: 630-638.
Molleman, F. 2010. Puddling: From natural history to understanding how it affects fitness. Entomologia Experimentalis et Applicata, 134 (2): 107–113.
Oro, D., Stefanescu, C., Alba, M., Capitán, J., Ubach, A. & Genovart, M. 2023. Factors affecting survival and dispersal of the comma butterfly in a high mountain deciduous forest habitat. Animal Biodiversity and Conservation, 46 (1): 1-11.
Phillips, B. B., Shaw, R. F., Holland, M. J., Fry, E. L., Bardgett, R. D., Bullock, J. M. $ Osborne, J. L. 2018. Drought reduces floral resources for pollinators. Global Change Biology, 24: 3226-3235.
Phon C., Kirton L. G. & Yusoff, N., 2017. Monitoring butterflies using counts of puddling males: A case study of the Rajah Brooke’s Birdwing (Trogonoptera brookiana albescens). PLoS One, 13(4): e0196304.
Pollard, E., & Yates, T.J. 1993. Monitoring Butterflies for Ecology and Conservation. Chapman & Hall, London, UK. 274 p. Prokopy, R.J. & Roitberg, B.D. 2001. Joining and avoidance behaviour in non-social insects. Annual Review of Entomology, 46: 631-665.
R Core Team. 2023. stats: The R Stats Package. Versió 4.3.0. Disponible a: https://stat.ethz.ch/R-manual/R-devel/library/stats/ html/stats-package.html [Data de consulta: 24 abril 2024].
Sculley, C.E. & Boggs, C.L. 1996. Mating systems and sexual division of foraging effort affect puddling behaviour by butterflies. Ecological Entomology, 21:193–197.
Stefanescu, C. 2023. Feeding on migratory butterflies: opportunistic observations of birds preying on the Painted Lady, Vanessa cardui Catalan Journal of Ornithology, 39: 21-28.
Tea, Y.K., Soong, J.W., Gan, C.W. & Lohman, D.J. 2020. Beachgoing butterflies: marine puddling on black sand beaches of Tangkoko Batuangus Nature Reserve, North Sulawesi. Journal of the Lepidopterists Society, 74(2): 127–131.
Vila, R., Stefanescu, C. & Sesma, J.M. 2018. Guia de les papallones diürnes de Catalunya. Lynx. Barcelona. 509 p. Wickham, H. 2023. ggplot2: A system for declarative graphics in R. Versió 3.3.5. Disponible a: https://github.com/tidyverse/ggplot2 [Data de consulta: 24 abril 2024].
Wood, S.N. 2021. mgcv: Mixed GAM Computation Vehicle with GCV/AIC/REML smoothness estimation. Versió 1.8-36. Disponible a: https://CRAN.R-project.org/package=mgcv [Data de consulta: 24 abril 2024].
Pivnick, K.A. & McNeil, J.N. 1987. Puddling in butterflies: sodium affects reproductive success in Thymelicus lineola Physiological Entomology, 12: 461-472.
de dues espècies de lepidòpters
Leucoptera malifoliella
Juli Pujade-Villar*, Csaba Thuróczy** & Elisenda Olivella***
* Universitat de Barcelona. Facultat de Biologia. Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals. Avda. Diagonal, 645. 08028 Barcelona, Catalunya. A/e:: jpujade@ub.edu ** Malomárok 27. 9730 Kőszeg, Hungary. A/e: thuroczy.cs@freemail.hu *** C/ Tarragona, 106, 1r 1a. E- 08015 Barcelona, Catalunya. A/e:eolivell@xtec.cat
Autor de correspondència: Juli Pujade-Villar, A/e: jpujade@ub.edu
Rebut: 08.05.2024; Acceptat: 03.06.2024; Publicat: 30.06.2024
Resum
Han estat determinades, atacant larves de Leucoptera malifoliella i Phyllonorycter corylifoliella, 9 espècies i 9 morfotipus pertanyents a dues famílies de Chalcidoidea (Eulophidae i Pteromalidae), emergits de les mines de fulles de diferents conreus de pomeres de Lleida. Els Eulophidae pertanyen als gèneres Baryscapus (1 espècie), Chysocharis (1 espècie), Pediobius (1 espècie), Pnigalio (1 espècie i 4 morfotipus) i Sympiesis (4 espècies i 2 morfotipus). Els Pteromalidae són molt escassos. D’aquesta família, s’han obtingut Dibrachys sp, Pteromalus sp i un exemplar el gènere del qual no ha pogut ser determinat. Pediobius pyrgo es cita per primer cop a Catalunya. Finalment, Pediobius pyrgo i els gèneres Dybrachis i Pteromalus es citen per primer cop a Phyllonorycter corylifoliella
Paraules clau: parasitoides, minadors de fulles, Malus domestica, Lleida, Catalunya.
Abstract
Chalcidoides obtained from two species of miner lepidopteran: Leucoptera malifoliella (Lyonetiidae) and Phyllonorycter corylifoliella (Gracillariidae) in apple crops in the province of Lleida
We have determined 9 species and 9 morphotypes included in two different families of Chalcidoidea (Eulophidae and Pteromalidae) attacking larvae of Leucoptera malifoliella and Phyllonorycter corylifoliella emerging from leaf mines on apple tree fields in Lleida. The Eulophidae are identified as Baryscapus (1 species), Chysocharis (1 species), Pediobius (1 species), Pnigalio (1 species and 4 morphotypes), and Sympiesis (4 species and 2 morphotypes). The Pteromalidae are less abundant and were identified as Dibrachys sp, Pteromalus sp, and a specimen whose generic identity is unknown. Pediobius pyrgo is reported for Catalonia for the first time. Finally, Pediobius pyrgo and the genera Dybrachis and Pteromalus are reported for the first time associated with Phyllonorycter corylifoliella
Key words: Parasitoids, leaf miners, Malus domestica, Lleida, Catalonia. Butlletí de la Institució Catalana d’Història
Introducció
Les plantacions de fruiters a Lleida, i les de pomeres en particular, constitueixen un motor econòmic molt important de la plana lleidatana. Aquets tipus de conreu, com qualsevol altre, pot presentar problemes amb organismes que poden constituir plagues que repercuteix en l’economia de la plantació. Els arbres fruiters poden ser atacats per diverses espècies d’insectes i d’aràcnids (subclasse àcars) i, amb una importància menor, mamífers, aus i moŀluscs (Sarasúa et al. 2000).
Tot i que cada un dels fitòfags té enemics naturals (ja siguin depredadors, parasitoides o patògens), els sistemes de producció actuals constitueixen un potent fre al desenvolu-
pament d’aquests animals i conformen un fort obstacle per a la seva activitat reguladora de les poblacions de les plagues (Sarasúa et al., 2000). Així, en plantacions no tractades o sense insecticides, la fauna útil és més rica i variada que en les plantacions comercials on usen insecticides. Aquest fet fa que conèixer les espècies que ataquen aquestes espècies perjudicials sigui de gran interès, ja que qualsevol d’elles pot ser beneficiosa per a aconseguir el control biològic d’una plaga (Hawkins, 1993). De fet, en moltes finques comercials lleidatanes de pomera (i també de perera) s’apliquen tècniques de Control Integrat de Plagues i de Producció Integrada, la qual cosa sovint comporta un control natural de les plagues (Sarasúa et al., 2000).
J. PUJADE-VILLAR ET AL.
Entre les espècies perjudicials del cultiu de pomeres hi tenim els minadors de fulles, que s’alimenten de les cèŀlules del parènquima de les fulles o d’altres òrgans vegetals i que, amb major o menor intensitat, disminueixen la capacitat fotosintetitzadora de l’arbre. Moltes de les espècies minadores són oligòfagues i, per aquesta raó, la identificació de la planta hoste pot servir per a la determinació de l’espècie minadora. A Europa, es coneixen com a mínim 10 espècies que poden causar danys als conreus de fruiters. No es van detectar danys a les plantacions de pomeres produïts per microlepidòpters minadors de fulles a Europa fins a finals dels anys quaranta. Així, l’any 1946 va ser detectat un primer focus de minadors a Llombardia, Emília-Romanya i el Vèneto, i des d’allí es van estendre per tota la península italiana (Ciampolini, et al., 1988). Els microlepidòpters minadors de fulles són una plaga de certa importància als conreus de pomera de la plana de Lleida des de 1964. (Sarto i Monteys & Vives de Quadres 1991). Les principals espècies minadores de fulla de pomeres a Lleida són els lepidòpters Leucoptera malifoliella (Costa, 1836) (Lyonetiidae), que fa una mina circular, Phyllonorycter corylifoliella (Hübner, 1796) (Gracillariidae), que provoca la mina translúcida del feix també anomenada mina en placa de la fulla de pomera i, en menor mesura, Phyllonorycter mespilella (Hübner, 1905) (Gracillariidae), que forma la mina puntejada (Olivella, 1997a; Bellostas et al., 1998). Phyllonorycter corylifoliella és una de les espècies més abundants als conreus de pomera de la plana de Lleida. (Olivella, 1997b).
El parasitoidisme és molt freqüent entre els lepidòpters minadors, ja que encara que pot semblar que la mina protegeix de la depredació, en realitat existeixen famílies d’himenòpters (com els Eulophidae) especialitzats a parasitar minadors de fulles (Gauld & Bolton, 1988). La fauna parasitoide de P. corylifoliella i de P. mespilella ha estat parcialment estudiada per
Bellostas et al. (1998) i Sarasúa et al. (2000). L’única referència respecte al parasitoidisme de L. malifoliella a Lleida va ser feta en forma de llista d’espècies per Sarasúa et al. (2000).
En aquest estudi exposem la fauna parasitoide de les dues espècies de minadors de les fulles més importants a les pomeres lleidatanes: L. malifoliella i P. corylifoliella
Material i mètodes
Zona de mostreig
Les experiències de camp han estat realitzades en sis finques situades dins de la comarca del Segrià, a la plana de Lleida. Les finques són les següents: dues al terme municipal d’Alcarràs (Alcarràs-1 i Alcarràs-3), dues al terme municipal de Lleida (Lleida-1 i Lleida-2), una al de Benavent de Segrià (Benavent) i una al de Torrefarrera (Malpartit). A la figura 1 es dona l’emplaçament exacte de les finques dins la comarca del Segrià, i la d’aquesta dins de Catalunya. Ocasionalment s’han usat dades obtingudes a d’altres finques (Torregrossa, Barbens, La Bordera, Mollerussa, Puigvert, Torrefarrera i Torregrossa). La localització dels diferents indrets és la següent: Alcarràs (31TBG90), Alpicat (31TBG21), Benavent de Segrià (31TCG21), Barbens, (31TCG31), Lleida (31TCG01), Mollerussa (31TCG21), Menàrguens (31TCG12), Palau d’Anglesola (31TCG21), Torregrossa (31TCG10), Torrefarrera (31TCG01).
Les característiques de les principals finques estudiades són les següents (els tractaments fets estan resumits a l’apèndix): ALCARRÀS-1. Es tracta d’una finca de dues ha situada a 3,5 km d’Alcarràs, en la carretera que va cap a Vallmanya. Està formada per 12 fileres de 100 pomeres «Golden» cadascuna. Els arbres van ser plantats l’any 1987 i la seva alçada mitjana
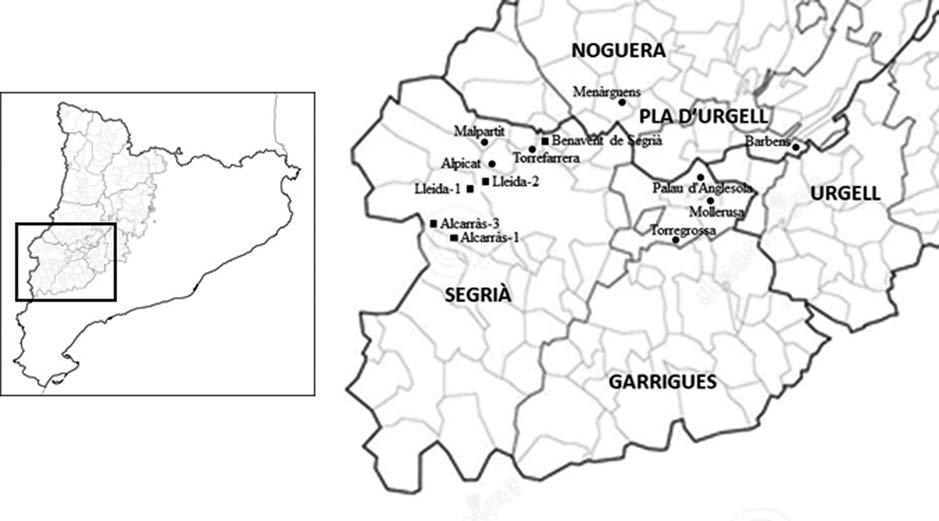
supera els quatre m. Les coŀlectes van ser realitzades en sis fileres de 22 arbres que no van rebre cap tractament insecticida durant el temps d’estudi.
ALCARRÀS-3. Es tracta d’una finca de 0,8 ha situada al km set de la carretera d’Alcarràs a Vallmanya, en una zona on el conreu majoritari són els presseguers i les pereres. Està formada per vuit fileres de 73 arbres, i hi alternen les de poma «Golden» amb les de poma «Starking». Les pomeres van ser plantades l’any 1984 i la seva alçada mitjana ronda els 3,5 m. En ella es realitza un control de plagues sota l’estratègia de lluita integrada.
BENAVENT DE SEGRIÀ. Aquesta finca està situada en una zona on el conreu majoritari són les pomeres. Està relativament aïllada de la resta de conreus (les pomeres més properes estan a 500 m i la resta a més d’un km). Té 0,5 ha i estava formada per set fileres de 15 pomeres «Golden» cadascuna, plantades a finals dels anys seixanta. Fou explotada comercialment fins a l’any 1992, quan es va signar un acord amb l’IRTA UDL per realitzar-hi un estudi sobre la biologia i la dinàmica de població del lepidòpter Cydia pomonella Linnaeus, 1758 (Tortricidae). Per aquest motiu, la finca deixà d’explotar se durant els anys 1993 i 1994, tot i que fou mínimament regada a l’estiu. Finalment, els arbres van ser arrencats a mitjan 1995 i es va començar una nova explotació amb pereres.
LLEIDA-1. És una finca d’una ha situada a la carretera que va de Lleida a Torres de Sanui, on es realitza lluita biològica contra plagues. És aïllada, i està formada per nou fileres de pomera «Golden» (895 arbres) i un bancal de 300 pereres
de la varietat ‘blanquilla’. Els arbres van ser plantats a finals dels anys setanta i tenen uns 3 m d’alçada. L. malifoliella va afectar notablement aquesta finca sobretot en els anys 1990 i 1991, un bienni en què els danys foren molt elevats.
LLEIDA-2. Es tracta d’una finca, aïllada i de 0,5 ha, abandonada des de l’any 1987. Està situada en la carretera que va de Lleida a Torres de Sanui. Conté 19 presseguers (ja quasi morts), 34 pereres i a l’entorn de 95 pomeres de la varietat «Golden». La finca es va utilitzar per a fer estudis del desenvolupament larvari de les tres espècies de minadores sense interferència humana (no va rebre insecticides ni adobs i no va ser regada durant el temps d’estudi). A la primavera del 1993 tots els arbres d’aquesta finca van ser talats i fins ara no s’hi ha plantat res més.
MALPARTIT. Es tracta d’una finca d’explotació comercial de sis ha. És aïllada i conté 1.200 pereres majoritàriament de la varietat «Conference», 2.892 pomeres de la varietat «Golden» i a l’entorn d’un miler de presseguers. L’any 1991 els experiments van ser realitzats en arbres (pomeres i pereres) plantats l’any 1977 (de 3,5 a 4,5 m d’alçada). Els anys 199293 els experiments es realitzaren en un bancal de 0,8 ha format per 12 fileres de 40 pomeres cadascuna, molt més joves (plantades l’any 1989) i amb altures inferiors als 3 m.
Leucoptera malifoliella (Fig. 2b) és una espècie minadora de les fulles de pomera, coneguda com la minadora circular

de les fulles (Fig. 2a) i es una plaga important dels conreus de pomera del Segrià des de 1964 (SPV, 1964-1981 & 1989-1991). Està distribuïda per tot Europa central i meridional. A Catalunya l’espècie està amplament difosa pels conreus de pomera, tant de la plana de Lleida com de l’Empordà, i també a d’altres comarques on la pomera és un conreu minoritari (Vallès Oriental i Cerdanya). A la plana de Lleida, l’hem capturada a (Olivella, 1995): Alcarràs, Alpicat, Benavent, Barbens, Lleida, Mollerussa, Menàrguens, Palau d’Anglesola, Torregrossa i Torrefarrera. La larva nounata ha de travessar perpendicularment la fulla des del revers i a través del parènquima llacunós i del parènquima en palissada abans de començar la galeria sota l’epidermis de l’anvers, on continuarà el seu desenvolupament. La larva madura necessita les potes per poder abandonar la mina i desplaçar se fins a arribar a un lloc adient per pupar. La pupació es produeix sempre fora de la mina, en un capoll lligat per fils de seda al seu suport. L’oviposició es produeix al revers de les fulles. L’atac de la larva afecta totes les cèŀlules de la zona ja que estan interconnectades. Aquest procés provoca l’assecament de les epidermis superior i inferior, i va seguit d’un assecament de la zona minada, la qual cosa, si el nombre de mines és suficientment elevat, pot arribar a assecar tota la fulla. La mina té un contorn circular típic que fa que l’espècie rebi el nom vulgar de minadora circular de les fulles.
Phyllonorycter corylifoliella (Fig. 2d) és una espècie oligòfaga, coneguda com a minadora en placa de la fulla de pomera (Fig. 2c). Malgrat ser comuna en els conreus de pomera de la plana de Lleida, no provoca de moment danys d’importància (Olivella, 1997b) i és poc freqüent a les fulles de perera (Olivella 1997b). Està distribuïda per Europa des del nord de Finlàndia fins a Portugal (GBIF, 2024) A Catalunya, està amplament distribuïda per la plana de Lleida, i l’hem capturada en les mateixes localitats que l’espècie anterior. Les larves poden alimentar-se de diferents rosàcies (Kuznetsov 1981; Emmet et al. 1985): pomera (Malus domestica Borkh. i altres espècies del gènere Malus), perera (Pyrus communis L.), arç blanc (Crataegus spp., Fig. 3a), codonyer (Cydonia oblonga Mill.), nesprer (Mespilus germanica L.), moixera de guilla (Sorbus aucuparia L.), moixera (Aria edulis (Willd.) M. Roem. = Sorbus aria Crantz, Cotoneaster spp., Amelanchier spp. i Betula
spp. No es coneixen totes les plantes nutrícies d’aquesta espècie a la península Ibèrica, però a la plana de Lleida l’hem observat sobre les següents rosàcies: pomera, perera, codonyer i ametller (Prunus dulcis). Bona part del desenvolupament larvari d’aquesta espècie té lloc en un mina plana que no presenta plecs. Aquest tret característic li ha donat el nom comú. La larva (Fig. 3b) només s’alimenta de les cèŀlules del parènquima en palissada, respectant les cèŀlules del parènquima llacunòs i les dues capes epidèrmiques; per tant, no provoca l’assecament de la zona on és la mina i les fulles minades no presenten cap àrea més fosca a la banda contrària on aquesta es troba (Olivella 1997b). L’oviposició té lloc sobre l’anvers de la fulla. De l’ou emergeixen larves que perforen l’epidermis de l’anvers fins a situar-se sota l’epidermis de la fulla i començar la mina. Les restes de l’ou romanen adherides a l’epidermis de la fulla a fi de protegir l’entrada de la mina.
Recoŀlecció de material
La recoŀlecció de fulles es va fer trencant la fulla pel pecíol, o tallant llucs sencers (= brots anuals) amb tisores de podar convencionals o amb tisores de podar amb braç telescòpic de 3 m destinades a poder accedir als llucs de la part superior dels arbres.
Les fulles recoŀlectades van ser coŀlocades dintre d’una bossa de cartró, i aquesta va ser ficada en una altra de plàstic transparent. D’aquesta manera es reduïa la pèrdua d’humitat i al mateix temps es retardava el podriment de les fulles. Vam realitzar diverses recollides de fulles tant de l’arbre com del terra durant tot el període de mostreig, i les vam dipositar en insectaris per esperar l’aparició dels adults. Al final de cada generació, també es recolliren un mínim de 100 mines (amb larva viva o no) per obtenir els parasitoides.
Els adults obtinguts van ser conservats en alcohol de 70º fins al moment del seu estudi.
Determinació de parasitoides
El segon autor ha estat el responsable de determinar els 561 exemplars de Chalcidoidea (329 mascles i 233 femelles)
obtinguts de les mines de L. malifoliella i P. corylifoliella entre el 1991 i el 1995. El material està dipositat a la coŀlecció de Csaba Thuróczy i al Savaria Múzeum (Hongria).
Fotografies
Si no es diu el contrari, han estat fetes per la darrera autora. La imatge de les larves P. corylifoliella va ser obtinguda als Serveis Cinentífico-Tècnics de la UB (CCiTUB) amb el microscopi electrònic de rastreig Leica Cambridge S360 SEM (Oxford Instruments) a alt voltatge. Prèviament, les dues erugues van ser cobertes d’or.
Atacant larves de L. malifoliella i P. corylifoliella, han estat determinades 9 espècies i 9 morfotipus (que no han pogut ser identificats a nivell específic) pertanyents a dues famílies de Chalcidoidea: Eulophidae i Pteromalidae. Els parasitoides obtinguts són els següents:
Família Eulophidae
Baryscapus nigroviolaceus (Nees, 1834)
Material estudiat
Emergits de L. malifoliella: Alcarràs -1 2.x.1993: 1 ♂ & 1 ♀; Lleida 6.vi.1992: 15 ♀, 11.vii.1992: 3 ♀, 20.vi.1992: 1 ♂; Malpartit, 3.v.1992: 13 ♂ & 6 ♀, 14.xi.1992: 1 ♀; Torregrossa 5.x.1991: 47 ♂. Emergits de P. corylifoliella: Malpartit 17.iv.1992: 1 ♂.
Espècie distribuïda pel Paleàrtic occidental (Noyes, 2019).
Majoritàriament parasitoide primari de larves de sis famílies de lepidòpters (Heliozelidae, Lasiocampidae, Lyonetiidae, Tineidae, Tortricidae i Yponomeutidae), tot i que també ha estat citada en algunes espècies de Curculionidae (Coleoptera) i de Cecidomyiidae (Diptera), i fins i tot com a parasitoide secundari d’Elasmidae i Eulophidae (Noyes, 2019). Ha estat citada tant a L. malifoliella com a P. corylifoliella. A Catalunya només es coneix de la província de Lleida (Bellostas et al., 1998; Sarasúa et al., 2000).
Chrysocharis pentheus (Walker, 1839)
Material estudiat
Emergits de L. malifoliella: Alcarràs 2.ix.1991: 1 ♂, 30.vii1991: 1 ♀, 4.x.1991: 1 ♀, Alcarràs -1 1.ix.1992: 2 ♂; 9.vii.1993: 1 ♂, 5.viii.1993: 1 ♂ & 1 ♀, 9.viii.1993: 2 ♀, 20.viii.1993: 1 ♂ & 1 ♀, 4.ix.1993: 2 ♂ & 1 ♀, 16.ix.1992: 3 ♀, 17.ix.1993: 1 ♀, 1.x.1993 1 ♀, 4.x.1993: 1 ♂ & 3 ♀, 2.iv.1994: 1 ♂; La Bordeta 25.ix1992: 2 ♂; Lleida 6.vi.1992: 1 ♂, 24.vii.1992: 1 ♂ & 1♀, 1.viii.1992: 5 ♂ & 1 ♀, 9.viii.1992: 1 ♂, 22.viii.1992: 2 ♂, 8.ix.1992: 1 ♂ & 1 ♀, 16.ix.1992: 1 ♀, 25.vi.1993: 2 ♂, 23.vii.1993: 1 ♀, 5.viii.1993: 1 ♀, 4.ix.1993: 1 ♂ & 1 ♀, Benavent de Segrià 9.viii.1992: 2 ♀, 9.vii.1993: 1 ♀, 23.vii.1993: 1 ♂ & 1 ♀, 20.viii.1993: 1 ♀, 4.ix.1993: 7 ♂ & 2 ♀, 22.ix.1994: 1 ♀, 1.x.1993: 1 ♀, 10.vi.1994: 1 ♂ & 3 ♀, 26.vi.1994: 1 ♀, 13.vii.1994:
1 ♂ & 2 ♀, 27.x.1994: 1 ♂; Malpartit 22.viii.1991: 1 ♀, 16.ix.1992: 1 ♀, 1.x.1992: 1 ♀, 9.x.1992: 1 ♂, 28.xi.1992: 1 ♂, 19.ii.1993: 1 ♂, 4.ix.1993: 3 ♀, 15.x.1993: 1 ♂; Malpartit, Alpicat 4.ix.1993: 5 ♂ & 2 ♀, 16.ix.1992: 1 ♂. Emergits de P. corylifoliella: Alcarràs -1 6.vi.1992: 1 ♀, 1.viii.1992: 1 ♂, 1.x.1992: 1 ♂ & 1♀, 16.x.1992: 1 ♂, 25.x.1992: 2 ♂, 28.xi.1992: 1 ♂, 12.xii.1992: 1 ♂ & 1 ♀, 16.xii.1992: 1 ♂, 5.ii.1993: 1 ♂, 29.x.1993: 1 ♂, 11.xi.1993: 1 ♂, 28.xi.1993: 3 ♂, 1.v.1995: 1 ♂; Alcarràs -1 12.ix.1991: 2 ♀, 8.xi.1991: 1 ♂, 24.ii.1994: 1 ♂; Alcarràs -3 28.viii.1991: 1 ♀, 12.ix.1991: 1 ♂, 15.ix.1991: 1 ♂, 5.iii.1993: 1 ♀, 15.x.1993: 1 ♂ & 1 ♀; Lleida 18.vii.1992: 1 ♂, 24.vii.1992: 1 ♂, 1.viii.1992: 1 ♂, 24.ix.1992: 1 ♂, 19.ii.1993: 1 ♂; Benavent de Segrià 15.x.1993: 1 ♂, 24.x.1993: 1♀, 10.vi.1994: 1 ♀; Malpartit 28.viii.1991: 1 ♂, 8.ix.1991: 1 ♂, 19.x.1991: 1 ♂, 25.x.1991: 1♂,8.xi.1991: 1 ♂, 12.xii.1992: 1 ♂, 22.i.1993: 1 ♂ & 1♀, 28.xi.1993: 3 ♂, 24.ii.1994: 1 ♂; Mollerussa 4.x.1991: 1 ♂.
Espècie distribuïda per tot l’hemisferi nord (Noyes, 2019). Malgrat que ha estat citada com a hoste primari d’algunes espècies de coleòpters i lepidòpters, ataca fonamentalment larves de Diptera (Agromyzidae principalment, però també dues espècies de Cecidomyiidae) i espècies de 12 famílies de lepidòpters (Noyes, 2019), entre les quals hi ha L. malifoliella i P. corylifoliella. Citada a Catalunya només a la província de Lleida (Bellostas et al., 1998; Sarasúa et al., 2000).
Pediobius pyrgo (Walker, 1839)
Material estudiat
Emergits de L. malifoliella: Lleida 11.vii.1992: 1 ♂.
Espècie distribuïda per tot l’hemisferi nord (Noyes, 2019). Ataca principalment espècies de 14 famílies de lepidòpters, tot i que també ha estat citada com a hoste primari d’algunes espècies de coleòpters, dermàpters, dípters i himenòpters (Noyes, 2019). Ha estat obtinguda atacant L. malifoliella. Es cita per primer cop a P. corylifoliella, però ha estat referenciada com a parasitoide de P. blancardella. A Catalunya només es coneix de la província de Lleida (primera citació per a Catalunya).
Pnigalio pectinicornis (Linnaeus, 1758)
Material estudiat
Emergits de P. corylifoliella: Alcarràs -1 26.i.1992: 1 ♀, 20.vi.1992: 1 ♂, 28.xi.1992: 1 ♂ & 1 ♀, 20.viii.1993, 1 ♀; Alcarràs -2 29.v.1992: 1 ♂; Alcarràs -3 15.xi.1991: 1 ♀, 12.xii.1991: 1 ♀; Malpartit 12.ii.1994: 2 ♀.
Espècie de distribució paleàrtica també coŀlectada a Nova Zelanda (Noyes, 2019). Ataca principalment espècies d’11 famílies de lepidòpters, tot i que també ha estat citada com a hoste primari d’algunes espècies de coleòpters, dípters i himenòpters, i fins i tot com a parasitoide secundari del bracònid Apanteles circumscriptus (Nees, 1834) i de dues espècies d’eulòfids (Hymenoptera) (Noyes, 2013). Ha estat obtinguda atacant L. malifoliella i P. corylifoliella. A Catalunya només es coneix de la província de Lleida (Bellostas et al., 1998; Sarasúa et al., 2000).
Pnigalio sp. A
Material estudiat
Emergits de P. corylifoliella: Alcarràs -1 25.xii.1991 1 ♂, 1.viii.1992: 1 ♂, 9.x.1992: 1 ♂, 1.xi.1992: 3 ♂, 14.xi.1992: 2 ♂, 28.xi.1992: 1 ♂, 12.xii.1992: 1 ♂, 5.ii.1993: 1 ♀, 14.x.1993: 1 ♀, 24.x.1993: 1 ♀, 29.x.1993: 2 ♂; 11.xi.1993: 2 ♂; Alcarràs -3 3.xi.1991: 1 ♂, 9.x.1992: 1 ♂, 15.x.1993: 1 ♂; Barbens 25.x.1992: 1 ♂, 1.xi.1992: 3 ♂, 14.xi.1992: 1 ♂, 8.i.1993: 1 ♂, Malpartit 28.xi.1992: 1 ♂, 24.ii.1994: 2 ♀, 20.viii.1995: 1 ♀.
Pnigalio sp. B
Material estudiat
Emergits de P. corylifoliella: Alcarràs -1 8.xi.1991: 1♀, 12.vi.1992: 1♀; Malpartit 28.viii.1991: 3♀.
Pnigalio sp. C
Material estudiat
Emergits de P. corylifoliella: Alcarràs -1 20.vi.1992: 1 ♀, 18.vii.1992: 1 ♂, 1.viii.1992: 1 ♀, 15.viii.1992: 1 ♀, 1.xi.1992: 2 ♀, 20.viii.1993: 3 ♂ & 1 ♀; 18.vii.1992: 1 ♂; Benavent de Segrià 24.x.1993: 1 ♂; Malpartit 28.vii.1991: 1 ♂, 28.viii.1991: 1 ♂, 25.viii.1992: 1 ♂ & 1 ♀, 20.viii.1995: 1 ♀; Malpartit-Alpicat 4.ix.1993: 1 ♀.
Pnigalio sp.
Material estudiat
Emergits de L. malifoliella: Alcarràs 15.xi.1991: 1 ♂ & 1 ♀. Emergits de P. corylifoliella: Alcarràs -1 25.x.1992: 1 ♂, 22.i.1993: 1 ♂, 20.viii.1993: 1 ♂; Alcarràs -3 15.xi.1991: 1 ♂; 20.vi.1992: 1 ♂; Lleida 18.vii.1992: 1 ♂; Benavent de Segrià 22.ix.1994: 1 ♂; Malpartit 12.ii.1994: 1 ♂.
Sympiesis acalle (Walker, 1848)
Material estudiat
Emergits de P. corylifoliella: Alcarràs -1 12.vi.1992: 1 ♀, 4.vii.1992: 1 ♂, 18.vii.1992.18: 2 ♀,1.viii.1992: 1 ♀, 9.viii.1992: 2♀, 15.viii.1992: 2 ♀, 22.viii.1992: 1 ♀, 1.x.1992: 1 ♂, 9.x.1992: 1 ♂, 16.x.1992: 1 ♂ & 1 ♀, 14.xi.1992: 1 ♂, 28.xi.1992: 1 ♂, 8.i.1993: 1 ♀, 9.viii.1993: 1 ♂, 20.viii.1993: 2 ♂ & 1 ♀, 4.ix.1993: 1 ♂, 11.xi.1993: 1 ♂, 24.ii.1994: 1 ♀; Alcarràs -3 26.x.1991: 1 ♂ & 1 ♀, 15.xi.1991: 1 ♂, 25.xii.1991: 1 ♂; Barbens 22.i.1993: 1 ♂; Benavent de Segrià 25.vi.1992: 1 ♂, 5.viii.1993: 1 ♂, 20.viii.1993: 1 ♀; Lleida 12.vi.1992: 1 ♂, 20.vi.1992: 1 ♂, 26.vi.1992: 1 ♂, 11-vii.1992: 1 ♂, 1.viii.1992: 2 ♂ & 1 ♀, 15.viii.1992: 2 ♀, 9.viii.1992: 2 ♂, 14.xi.1992: 1 ♀, 19.ii.1993: 1 ♀, 9.viii.1993: 2 ♂; Benavent de Segrià 25.vi.1992: 1 ♀, 24.x.1992: 1 ♂, 25.vi.1993: 2 ♂ & 1 ♀, 23.vii.1993: 2 ♂, 5.viii.1993: 2 ♂ & 2 ♀, 15.x.1993: 1 ♂, 24.x.1993: 1 ♂, 29.x.1993: 1 ♂; Malpartit 24.vi.1991: 2 ♂ & 2 ♀, 4.vii.1991: 3 ♂ & 3 ♀, 22.viii.1991: 1 ♀, 28.viii.1991: 7 ♂ & 7 ♀, 4.iii.1992: 2 ♀, 17.iv.1992: 1 ♀, 12.vi.1992: 1 ♀, 26.vi.1992: 1 ♂ & 2 ♀, 11.vii.1992: 1 ♂ & 1 ♀, 4.vii.1992: 2 ♀, 15.viii.1992: 3 ♀, 14.xi.1992: 1 ♂, 28.xi.1992: 1 ♀, 12.xii.1992: 1 ♂, 19.iii.1993: 1 ♂, 4.ix.1993: 1 ♂ & 2 ♀, 20.viii.1993: 1 ♂ & 4♀, 24.x.1993: 1
Butlletí ICHN 88 (2), 2024
♂, 11.xi.1993: 1 ♂, 12.ii.1994: 1 ♂, 20.viii.1995: 2 ♂ & 1♀; Torreforta 9.viii.1992: 2 ♂ & 1 ♀.
Espècie distribuïda per la regió Neàrtica (Noyes, 2019). Ataca principalment espècies d’11 famílies de lepidòpters, tot i que també ha estat citada com a hoste primari d’algunes espècies de dípters i d’himenòpters, i fins i tot com a parasitoide secundari de bracònids (incloent el gènere Apanteles Förster, 1862) i Icneumònids (Noyes, 2013). Ha estat obtinguda atacant L. malifoliella i P. corylifoliella. A Catalunya només es coneix de la província de Lleida (Bellostas et al., 1998; Sarasúa et al., 2000).
Sympiesis gordius (Walker, 1839)
Material estudiat
Emergits de P. corylifoliella: Alcarràs -1 26.i.1992: 1 ♀, 11.vii.1992 ♂, 4.vii.1992: 1 ♀, 9.viii.1992: 1 ♀, 15.viii.1992: 1 ♂, 20.viii.1992: 2 ♀, 8.i.1993: 1 ♀, 20.viii.1993: 1 ♂, 4.ix.1993: 1 ♂ & 1 ♂, 24.ii.1994: 1 ♀, 9.vii.1995: 2 ♂; Alcarràs -3 26.ix.1991: 1 ♂; Lleida 26.vi.1992: 1 ♀, 24.vii.1992: 3 ♂, 1.viii.1992: 2 ♀, 15.viii.1992: 3 ♂, 28.viii.1992: 1 ♀; Benavent de Segrià 29.v.1993: 1 ♂, 25.vi.1993: 1 ♀, 20.viii.1993: 1 ♂, 10.vi.1994: 1 ♀; Malpartit 20.viii.1991: 1 ♀, 28.viii.1991: 8 ♂ & 10♀, 15.viii.1992: 2 ♂ & 2 ♀, 29.viii.1992: 1 ♀, 1.x.1992: 1 ♂, 4.ix.1993: 2 ♀, 1.xi.1993: 2 ♂, 20.viii.1995: 3 ♂; Mollerussa 14.viii.1991: 1 ♂, 29.xi.1991: 1 ♂.
Espècie de distribució paleàrtica també citada al Canadà (Noyes, 2019). Ataca principalment espècies de nou famílies de lepidòpters (sobretot de la família Gracillariidae), també algunes espècies de coleòpters i d’himenòpters, i és parasitoide secundari de bracònids (incloent el gènere Apanteles) i d’eulòfids (Noyes, 2013). Ha estat citada a L. malifoliella i a P. corylifoliella. A Catalunya només es coneix de la província de Lleida (Bellostas et al., 1998; Sarasúa et al., 2000).
Sympiesis sp. nr. gordius (Walker)
Material estudiat
Emergits de P. corylifoliella: Lleida 1.viii.1992: 1 ♀; Malpartit 4.ix.1993: 2♀.
Sympiesis gregori Boucek, 1959
Material estudiat
Emergits de P. corylifoliella: Alcarràs -1 20.vi.1992: 1 ♀; Lleida 26.vi.1992: 1 ♀; Benavent de Segrià 26.v.1994: 1 ♀, 10.vi.1994: 2 ♂ & 1 ♀.
Espècie de distribució paleàrtica occidental (Noyes, 2019).
Parasitoide primari de les famílies Gracillariidae i Nepticulidae (Lepidoptera). Ha estat citada a P. corylifoliella. A Catalunya es coneix de les províncies de Tarragona (Fibla & Martínez, 2000) i Lleida (Bellostas et al., 1998; Sarasúa et al. (2000).
Sympiesis sericeicornis (Nees, 1834)
Material estudiat
Emergits de P. corylifoliella: Alcarràs -1 26.i.1992: 1 ♂, 24.vii.1992: 1 ♀, 9.viii.1992: 1 ♂, 15.viii.1992: 1 ♂, 16.x.1992: 1 ♂, 28.xi.1992: 1 ♀, 22.i.1993: 1 ♂, 8.i.1993: 2
♂, 5.ii.1993: 1 ♂, 22.viii.1993: 1 ♀, 11.iii.1994: 1 ♂; Alcarràs -2 11.x.1991: 1 ♂; Alcarràs -3 20.ix.1991: 1 ♂, 26.x.1991: 1 ♀, 15.xi.1991: 2 ♀, 9.x.1992: 1 ♀; Lleida 17.iv.1992: 1 ♂, 12.vi.1992: 1 ♀, 20.vi.1992: 1 ♂, 11.vii.1992: 1 ♂, 18.vii.1992: 1 ♂, 1.viii.1992: 3 ♂, 22.viii.1992: 1 ♂, 28.xi.1992: 2 ♂, 12.xii.1992: 1 ♂, 16.xii.1992: 1 ♂, 5.iii.1993: 1 ♀, 20.viii.1993: 2 ♀; Lleida,-Benavent de Segrià 29.v.1993: 1 ♂, 5.viii.1993: 1 ♂, 15.x.1993: 1 ♂, 26.v.1994: 2 ♂, 27.x.1994: 2 ♂; Malpartit 8.viii.1991: 1 ♀, 28.viii.1991: 1 ♀, 19.x.1991: 1 ♂, 15.viii.1992: 1 ♂ & 1 ♀, 28.xi.1993: 1 ♀, 26.xii.1993: 1 ♀, 12.ii.1994: 1 ♂, 1994.II.24: 1 ♀; Puigvert 24.vi.1994: 1 ♀; Torrefarrera 1.viii.1992: 1 ♂, 9.viii.1992: 1 ♀.
Espècie distribuïda per la re gió Neàrtica (Noyes, 2019). Ataca principalment espècies d’11 famílies de lepidòpters (sobretot de la família Gracillariidae), i també algunes espècies de coleòpters i d’himenòpters. És parasitoide secundari de bracònids (incloent el gènere Apanteles) i d’eulòfids (Noyes, 2013). Ha estat citada a L. malifoliella i a P. corylifoliella. A Catalunya només es coneix de la província de Lleida (Bellostas et al., 1998; Sarasúa et al., 2000).
Sympiesis sp.
Material estudiat
Emergits de Phyllonorycter corylifoliella: Alcarràs -1 18.ii.1993: 1 ♂; Lleida-Benavent: 2 ♂, 10.vi.1994: 1 ♂.
Família Pteromalidae
Dibrachys sp.
Material estudiat
Emergits de L. malifoliella: Torregrossa 17.iv.1992: 1 ♂.
Pteromalus sp.
Material estudiat
Emergits de P. corylifoliella: Alcarràs -1 Lleida 28.xi.1992: 1 ♀; 12.xii.1992: ♀.
Pteromalidae sp.
Material estudiat
Emergits de P. corylifoliella: Malpartit 17.iv.1992: 1♀
Discussió
Els Eulophidae són amb diferència els calcidoideus més habituals parasitoides d’aquestes dues espècies minadores de fulles de pomeres. No sembla que hi hagi un parasitoide comú que ataqui les dues larves de lepidòpters que provoquen mines a les pomeres en les àrees d’estudi. Les úniques espècies que comparteixen són Baryscapus nigroviolaceus, Chysocharis pentheus i una espècie no identificada de Pnigalio. Segons Noyes (2019), aquests dos lepidòpters comparteixen Pnigalio pectinicornis i P. soemius. Per altra banda, Pediobius pyrgo només ha emergit de L. malifoliella i de Pnigalio pectinicornis, i totes les espècies del gènere Sympiesis estan associades només a P. corylifoliella
Pel que fa a L. malifoliella, malgrat que la major part de calcidoideus parasitoides corresponen a Eulophidae (amb 24 espècies), també han estat citades dues espècies de Pteromalidae i Trichomalus sp. (Noyes, 2019). De les mines d’aquest lepidòpter han estat identificades l’única espècie citada del gènere Baryscapus, una espècie de les 4 citades del gènere Chrysocharis i P. Pyrgo, que es cita per primer cop en aquest hoste. A més, una espècie no identificada del gènere Dibrachys (Pteromalidae, només coneguda D. microgastri en aquest hoste) i una espècie no identificada del gènere Pnigalio (Eulophidae, dues espècies del gènere esmentat es coneixen en aquest hoste: P. pectinicornis i P. soemius (Walker, 1839).
D’acord amb els nostres resultats, el percentatge de parasitisme més elevat es produeix a les generacions hivernants, però no és suficient per controlar la població de L. malifoliella. L’espècie més abundant és Chrysocharis pentheus,com també succeeix a Hongria a les zones de Szabolcs Szatmar, Bács Kiskun i Zala (Balázs, 1989), però no a Itàlia (a la zona del Piemont), on Chrysocharis nitetis (Walker, 1839) va ser l’espècie més àmpliament distribuïda i l’última en desaparèixer dels conreus on es fan tractaments insecticides (Santoro & Arzone, 1983).
Segons Noyes (2019), les larves de P. corylifoliella només són atacades pel que fa als calcidoideus per espècies de la família Eulophidae, tot i que en el nostre estudi també han emergit Pteromalidae. En aquestes larves, han estat citades 46 espècies d’Eulophidae i Euderus sp. D’aquestes espècies, a Lleida n’han estat identificades set: les 4 especies del gènere Sympiesis, l’única espècie del gènere Baryscapus, una espècie de les 5 citades de Pnigalio i una de les 4 espècies de Chrysocharis que parasiten P. corylifoliella. A més, hi ha diversos morfotips del gènere Pnigalio que no corresponen a cap de les espècies de Pnigalio citades en aquest hoste: P. agraules (Walker, 1839), P. incompletus (Bouček, 1971), P. pectinicornis, P. rotundiventris (Erdős, 1954) i P. soemius (Walker, 1839). També han emergit tres exemplars de Pteromalidae, dos dels quals són del gènere Pteromalus i no han pogut ser identificats. La família Pteromalidae es cita per primer cop com a parasitoide de P. corylifoliella.
Els estudis sobre els parasitoides que ataquen P. corylifoliella són més escassos, perquè aquesta espècie és menys freqüent en els camps de conreu europeus. Només s’han descrit els resultats obtinguts a Hongria, on aquesta espècie és una plaga secundària dels camps de conreu. A Hongria, els parasitoides dominants de P. corylifoliella són: S. gordius, S. sericeicornis, P. pectinicornis i Tetrastichus ecus (Nees, 1834). Els nostres estudis indiquen que, durant la generació d’estiu, els eulòfids són els parasitoides més abundants (S. acalle és un dels més freqüents) i que la major part del parasitoidisme d’eulòfids d’aquest minador és deu al gènere Sympiesis. Aquest fet no és favorable per al control biològic de les poblacions de P. corylifoliella perquè, durant l’estiu, augmenta la hiperparasitació d’Apanteles circumscriptus (Nees, 1834) (Braconidae) deguda bàsicament a diferents espècies de Sympiesis i, en conseqüència, disminueix la població d’Apanteles, la qual cosa pot provocar una explosió de la població del minador. També s’ha observat hiperparasitisme
entre diverses espècies d’eulòfids. No s’ha pogut identificar la larva del parasitoide hiperparasitada, perquè aquest hiperparasitisme només és observable quan s’està produint i no quan ja ha emergit l’eulòfid adult.
El control biològic d’aquestes espècies no es deu exclusivament a la presència d’eulòfids (Bellostas et al., 1998; Sarasúa et al., 2000), ja que també poden ser parasitades per icneumònids de les famílies Braconidae (com els dels gèneres Apanteles i Pholetesor Mason, 1981) i Ichneumonidae (com és el cas de Gelis), i per Pteromalidae (com s’ha mencionat anteriorment), però els Eulophidae són els seus principals paràsits. Cal tenir en compte també que algunes espècies de Sympiesis (gènere d’eulòfid més abundant en aquest estudi) poden actuar com parasitoides primaris o com a parasitoides secundaris atacant larves de bracònids (que sempre són parasitoides primaris), la qual cosa pot fer variar el control biològic de la plaga.
Bibliografia
Bellostas, J.J., Olivella, E., Verdú, M.J., Sarasua, M.J. & Avilla, J. 1998. Fauna parasitoidee de Phyllonorycter spp. en plantaciones de manzano de Lleida. Boletín de Sanidad Vegetal Plagas, 24 (2): 313-320.
Ciampolini, M.; Süss, L. & Trematerra, P. 1988. I Phyllonorycter Hb. (Lepidoptera Gracillariidae) most frequent and injurious to Pomoideae and Prunoideae in Italy. Bollelino di Zoologia agraria Bachicoltura, Series II, 20: 61 88.
Emmet, A. M., Watkinson, I. A. & Wilson, M. R. 1985. Gracillariidae. In: Heath, J. & Emmet, A. M. [ed.]. p. 244 362. The moths and butterflies of Great Britain and Ireland. Vol. 2. Colchester: Harley Books.
GBIF, 2024. GBIF—the Global Biodiversity Information Facility. https:// www.gbif.org/what-is-gbif [Data de consulta: 5 maig de 2024]
(2), 2024 GEA, FLORA ET FAUNA
Hawkins, B. A., 1993. Parasitoides species richness, host mortality and biological control. The American Naturalist, 141 (4): 634 641. https://doi.org/10.1086/285495
Kuznetsov, V. I., 1981. Gracillariidae (Lithocolletidae). In: MEDVEDEV, G. S. [ed.]. p. 199 411. Keys to the insects of the European part of the USSR. Reimpr. de 1990. Vol. IV, part II. Leiden: E. J. Brill.
Noyes, J.S. 2019. Universal Chalcidoidea Database. World Wide Web electronic publication. Accesible a: <http://www.nhm.ac.uk/ chalcidoids> [Data de consulta: 1 de maig de 2024].
Olivella, E. 1995. La minadora circular de les fulles Leucoptera malifoliella (Costa, 1836), als conreus de pomera de la plana de Lleida (Lepidoptera, Lyonetiidae) Treballs de la Societat Catalana de Lepidopterologia. 13-30
Olivella, E., 1997a. Phyllonorycter mespilella (Hübner, 1805) new species for the Iberian fauna. Shilap, Revista de Lepidopterología, 25: 37-42.
Olivella. E. 1997b. Estudi comparatiu de les espècies de microlepidòpters minadores de fulles presents als conreus de pomera de la Plana de Lleida (Lepidoptera, Gracillariidae, Lyonetiidae). Treballs de la Societat Catalana de Lepidopterologia, XIV (1995-1996): 7-25.
Sarasúa, M. J., Avilla, J., Torà, R. & Vilajeliu, M. 2000. Enemics naturals de plagues als conreus de fruita de llavor a Catalunya. Dossiers agraris ICEA, 6: 7-19.
Sarto i Monteys, V., Vives de Quadras, J. M. 1991. Introducció a l’estudi de les plagues de lepidòpters a Catalunya. Treballs de la Societat Catalana de Lepidopterologia, 11: 105-120.
SPV. 1964-1981. Servei de Protecció dels Vegetals (Generalitat de Catalunya). Memoria de la estación de avisos. 5. p. Lleida. Publicació inedita.
SPV. 1989-1991. Servei de Protecció dels Vegetals (Generalitat de Catalunya). Memoria de la estación de avisos. 5. p. Lleida. Publicació inedita.
SPV, 1992. Guia de productes fitosanitaris. Servei de Protecció dels vegetals. Departament d’Agricultura Ramaderia i Pesca de la Generalitat de Catalunya. Barcelona. 216pp.
L’any 1991 es van fer pocs tractaments amb insecticides perquè, com ja s’ha explicat abans, les temperatures sota zero del mes d’abril van gelar tota la poma als arbres. A cap finca, a excepció d’Alcarràs 3, es va fer tractament amb ovicides o inhibidors de creixement (com el Diflubenzuró) que són els productes indicats contra els minadores, tan sols es van aplicar insecticides convencionals per a controlar les plagues de les pomeres: organofosforats i carbamats. Aquests compostos presenten entre mitjana i alta toxicitat contra els himenòpters parasitoides i altres depredadors de les plagues: bàsicament neuròpters, dípters, hemípters, coleòpters i àcars carnívors (SPV, 1992). A la taula 1 s’indiquen els tractaments amb insecticides per a cada finca, durant l’any 1991. Les dades de toxicitat dels insecticides sobre la fauna útil (artròpodes parasitoides i depredadors de les plagues) han estat subministrades pel SPV de la Generalitat de Catalunya (SPV, 1992).
L’any 1992 als tractaments amb insecticides van augmentar respecte els de l’any anterior. A Alcarràs 1 no es va fer cap tractament amb insecticides a la zona experimental. A la zona experimental de Malpartit tampoc es va fer, aquest any, cap tractament amb ovicides. En canvi, es van fer diversos tractaments amb insecticides organofosforats i carbamats. A la taula 2 s’indiquen els tractaments amb insecticides realitzats l’any 1992, a cadascuna de les finques.
A Alcarràs 3 els tractaments realitzats van ser més específics perquè és una finca que està dins d’un projecte de Control Integrat de Plagues (CIP) en pomeres, de l’IRTA-UDL. En
aquest sistema de lluita es dona prioritat als elements naturals de control (lluita biològica), però no es descarta la utilització de plaguicides, si bé únicament quan sigui estrictament necessari i, utilitzant els productes que menys pertorbació causin als ecosistemes. En aquesta finca es fan seguiments setmanals de les plagues i no es tracta amb insecticides, sense haver valorat abans l’efecte que això pot provocar sobre la fauna útil, tractant en tot cas amb el producte més adient per a cada ocasió. En les dates de 6 de maig i 7 de juliol es va tractar amb Teflubenzuró i Fenoxicarb respectivament, per controlar les poblacions de Cydia pomonella, plaga clau de la pomera a la zona. Els dos insecticides emprats són ovicides i inhibidors de creixement molt efectius, molt utilitzats per combatre plagues de lepidòpters. Per tant, també van afectar directament a la població de L. malifoliella, que en aquelles dates estava també en estadi d’ou o de larva nounata.
A la taula 3 es presenten les dates i els productes insecticides emprats durant 1993, a cadascuna de les finques. A Malpartit, el tractament amb Imidacloprid del 6 de juny va afectar greument a la població de minadors (sobretot a L. malifoliella). Aquest insecticida va ser emprat per controlar les poblacions d’àfids de la finca, però va resultar de gran efectivitat per les minadors. A Alcarràs 1 no es va fer cap tractament insecticida a la zona experimental.
A la taula 4, es presenten els tractaments insecticides realitzats fins el 13 de juliol de 1994. Aquest any, tant a Malpartit com a Alcarràs 1, les poblacions de minadors van quedar molt afectades pels tractaments amb Imidacloprid realitzats a la primavera.
Finca Data tractament Producte usat Toxicitat sobre fauna útil
Alcarràs 1 23 de febrer Polisulfur de calci Baixa
Alcarràs 1 30 d’abril Dimetoat Elevada
Alcarràs 1 15 de maig Dimetoat + Format Elevada
Alcarràs 1 28 de maig Dimetoat Elevada
Alcarràs 3 25 de febrer Polisulfur de calci Baixa
Alcarràs 3 29 d’abril Fenoxicarb Variable
Alcarràs 3 24 de maig Etiofencarb Mitjana-alta
Alcarràs 3 26 de maig Mercarbam Elevada
Alcarràs 3 13 de novembre Polisulfur de calci Baixa
Benavent 14 de març Polisulfur de calci Baixa
Benavent 2 d’abril Clorfenvinfòs Elevada
Benavent 7 de juny Clorfenvinfòs Elevada
Benavent 25 de juliol Ometoat + Carbaril Elevada
Lleida 1 20 de maig Pirimicarb Baixa
Malpartit 30-31 de juliol Clorfenvinfòs Elevada
Malpartit 14 d’agost Metil-Oxidemeton Elevada
Taula 2. Calendari de tractaments per l’any 1992. (–) Toxicitat desconeguda.
Finca Data tractament Producte usat Toxicitat sobre fauna útil
Alcarràs 3 6 de maig Teflubenzuró –
Alcarràs 3 7 de juliol Fenoxicarb Variable
Benavent 21 de març Metil-Paration Elevada
Benavent 30 d’abril Dimetoat Elevada
Benavent 6 de maig Teflubenzuró –
Benavent 27 de maig Metomil Elevada
Benavent 2 de juliol Fenoxicarb Variable
Benavent 1 d’agost Fenoxicarb Variable
Benavent 1 d’agost Etiofencarb Mitjana-alta
Lleida 1 10 de maig Diflubenzuró Mitjana
Lleida 1 10 de juliol Diflubenzuró Mitjana
Malpartit 13 de maig Diazinon Mitjana
Malpartit 28 de juny Metidation Elevada
Malpartit 19 de juny Metil-Azinfós Elevada
Malpartit 15 de juliol Metil-Oxidemeton Elevada
Malpartit 28 d’agost Metomil Elevada
Taula 3. Calendari de tractaments amb insecticides per l’any 1993. (–) Toxicitat desconeguda.
Finca Data tractament Producte usat Toxicitat sobre fauna útil
Lleida 1 3 de juliol Bacillus thuringiensis Baixa
Lleida 1 10 de juliol Bacillus thuringiensis Baixa
Lleida 1 18 de juliol Bacillus thuringiensis Baixa
Malpartit 5 de maig Metil-Oxidemeton Elevada
Malpartit 31 de maig Diazinon Mitjana
Malpartit 6 de juny Imidacloprid –
Malpartit 4 de juliol Metil-Oxidemeton Elevada
Malpartit 3 d’agost Clorfenvinfòs Mitjana
Malpartit 23 d’agost Lambda-Cihalotrín
Taula 4. Calendari de tractaments amb insecticides per l’any 1994. (-) Toxicitat desconeguda.
Finca Data tractament Producte usat Toxicitat sobre fauna útil
Malpartit 11 de març Oli + 5 % DNOC –
Malpartit 21 de març Imidacloprid –
Alcarràs 1 20 de febrer Oli + 5 % DNOC –
Alcarràs 1 8 d’abril Dimetoat Elevada
Alcarràs 1 22 d’abril Dimetoat Elevada
Alcarràs 1 6 de maig Dimetoat Elevada
Alcarràs 1 6 de maig Piretrines –
Alcarràs 1 21 de maig Imidacloprid –
Bolton,
Kiko Gómez*, Xavier Espadaler**, Maria Teresa Martínez-Ferrer*** & Josep Miquel Campos-Rivela***
* Independent researcher. Castelldefels, Barcelona, Spain.
** CREAF, Universitat Autònoma de Barcelona, 08193 Cerdanyola del Valles, Spain
*** IRTA-Amposta. Carretera de Balada, Km 1, 43870 Amposta, Tarragona, Spain
Autor de correspondència: Xavier Espadaler. A/e: xavierespadaler@gmail.com
Rebut: 25.05.2024; Acceptat: 05.06.2024; Publicat: 30.06.2024
Abstract
The presence of a population of Monomorium monomorium Bolton in a citrus orchard at the fluvial island of Gràcia (Tarragona, Spain) is reported, this being the second Iberian record for that very small shining black species. Soil nesting, polygyny and tending of honeydew producing insects was ascertained. We briefly discuss if this species should be considered as native or exotic in the Iberian Peninsula.
Key words: Monomorium monomorium, Ebro Delta, Iberian Peninsula.
Resum
Monomorium monomorium Bolton, 1987 a Espanya (Hymenoptera: Formicidae)
S’informa de la presència d’una població de Monomorium monomorium Bolton en un hort de cítrics de l’illa fluvial de Gràcia (Tarragona, Espanya), sent el segon registre ibèric d’aquesta espècie negre brillant i molt petita. Es va comprovar la nidificació al sòl, la poligínia i la cura d’insectes productors de melassa. Comentem breument si aquesta espècie s’ha de considerar autòctona o exòtica a la Península Ibèrica.
Paraules clau: Monomorium monomorium, Delta de l’Ebre, Península Ibèrica. Butlletí de la Institució Catalana d’Història Natural, 88 (2): 101-103. 2024
Introduction
During a study on the effect of ants on biological control on citrus orchards, the abundance and diversity of ant species occurring in a commercial mandarin orchard was studied from spring until autumn along with citrus key pests’ populations, natural enemies, and crop phenology.
Ants were collected using pitfall traps and among the 7 species, one of them is of biogeographic and faunistic interest, Monomorium monomorium Bolton, 1987. An old Iberian citation by Saunders (1888; as M. minutum Mayr, 1855), from Gibraltar has never been confirmed: in effect, M. monomorium workers, under the current species concept, are not present in the classic Saunders’ collection at the British Museum of Natural History and has not been detected in Gibraltar since that old mention in 1888 (R. Guillem, pers. com.). It is highly likely that Saunders’ material belonged in the newly described M. andrei two years later by himself (Saunders, 1890). The mention by Collingwood (1978) of M. monomorium (as M. minutum) in the south part of Spain is likely based on Saunders first paper, as no specimens of that species were present in the extense collections done by Prof.
H. Franz (Vienna) (Collingwood & Yarrow 1969) in his long trip in Spain and Portugal.
In Spain the species M. monomorium had already been detected at a public garden in Córdoba city, therefore being that one the single locality known for this species from the Iberian Peninsula (Reyes-López et al., 2008). The population in Córdoba seems well established since workers were regularly collected between 2003 and 2007. Despite the eight literature records in Iberia noted for M. monomorium in Antmaps (2024), a critical appraisal of them clearly boils down to a single record at a single locality (Córdoba city). Here we add a second, verified, Spanish population, also flourishing and well established.
Island of Gràcia, Deltebre, Tarragona province. The fluvial island (surface 125 ha approx.) lies within the Ebro delta region and is surrounded by the river fresh water. The habitat studied is a citrus grove (40.7221 N 0.6847 E; elevation 2 m) with peripheral small remnants of a mosaic of poplar woods

and cane community (Camarasa et al., 1977). Soil is alluvial and humid because of the close level of the water table, with very fine particles and easy to excavate.
The study was conducted in a 0.58 ha plot selected within a commercial mandarin Citrus clementina Hort. ex Tanaka. cv. ‘Nules’ orchard. Trees were grafted on citrange Carrizo rootstock with a tree spacing of 5.8 × 3 and a drip irrigation system. Twelve pitfall traps (Ø=9.5 cm; L=11 cm) containing 300 ml of water and liquid soap (0.2%) were installed in the soil of the orchard. Traps remained in the orchard for 72h. After this period, ants caught inside the traps were identified in the laboratory. This activity was conducted three times: late May, late July, and late September. A general field survey (18.vi.2018) was conducted by direct visual search on tree trunks. Ant identification was done using Bolton (1987). Vouchers have been deposited with the Museu de Ciències Naturals de Barcelona.
Running the workers through the keys in Bolton (1987), the name M. monomorium was obtained. Their morphology fits exactly with workers from Córdoba city and other samples from the Mediterranean coast of France and Italy in our dry collection; also, with images available at http://Antweb. org (accessed 11.ii.2020) (Fig. 1). This is the verified second record for the Iberian Peninsula. Other ant species found in pitfall traps apart from M. monomorium were: Lasius grandis
Forel, Pheidole pallidula (Nylander), Plagiolepis pygmaea (Latreille), Tapinoma erraticum (Latreille), Hypoponera eduardi (Forel) and Solenopsis monticola Bernard. The most abundant species were M. monomorium and P. pallidula representing 60 % and 36 % respectively of the overall individuals captured. Depending on the date, M. monomorium was the most abundant in late May and P. pallidula was the most abundant in late July and late September.
During the general field survey workers were seen foraging up the trunk on citrus trees, tending honeydew-producing insects. Nests were located by following workers returning to the nest. The entrance was a small hole, scarcely wider than worker body width size, and without any trace of soil or litter surrounding it. This fits exactly with Emery’s (1916) description for M. monomorium: «Formicai piccolo nel suolo, aventi una sola apertura non circondata da un cercine di tera». Three nests were located. Polygyny (two queens, three queens) was ascertained in two partially excavated nests. The soil where this small ant may be collected close to Muggia (Italy) was already described by Muller (1923) as: «… terreno alluvionale leggermente salmastro».
Other species detected in the same habitat during the general field survey were Cardiocondyla mauritanica Forel, Formica sp. and Plagiolepis pygmaea (Latreille). This last species was seen also foraging on citrus trees and could easily be confused with Monomorium. The dense and clear-cut trails produced by Monomorium and a check with a magnifier allowed distinction of both species at the field (Formicine with a single petiole vs. Myrmicine with petiole and postpetiole).
It is debatable if this species should be considered as native or exotic in the Iberian Peninsula. Its origin is unknown. The species seems indeed to be infrequent in its geographical extension in Southern Europe (Antmaps, 2024), and shows an Eastern tendency. In Spain, it was tentatively considered either as exotic (Reyes-López et al., 2008) or as native (Reyes-López & Carpintero, 2014). McGlynn (1999) treated M. monomorium as exotic in places as Hawaiian Islands, China, SE Asia, or Caribbean Islands. Subsequently, SchlickSteiner et al. (2008), based on McGlynn (1999) and likely because the ant is registered only indoors, treated it as exotic in Austria. Blatrix et al. (2018) consider it as native in France. Similar conclusion for Italy (Schifani, 2022), Croatia (Bračko, 2006), Albania (Wagner et al., 2018) and, seemingly, Crete (Salata et al., 2020). Surprisingly, Borowiec & Salata (2021) qualify it as a tramp species commonly found in tourist resorts or suburban and urban areas from Greece and many Greek islands. Karaman & Kiran (2018) considered it as native in Turkey although as introduced two years later (Kiran & Karaman 2020). No clear pattern. In short, the species seems to be constantly sparse and geographically restricted in several habitats. The extreme rarity in space and time of this species in Iberia and the fact that the two available records (city garden Córdoba; citrus grove Deltebre) are certainly anthropized habitats, further suggests accepting the exotic label for M. monomorium in Iberia.
It remains to be clarified an intriguing taxonomic question, the assertion from Bolton (1987: 288) stating «… Finally I suspect that southern European populations currently referred to as minutum Mayr may in truth consist of two separate species.». As far as we know, a characteristic and single phenotype is collected, albeit rarely, in Southern European countries. The population now discovered in Spain seems well posited for a focused sociometric study (Tschinkel, 1989; 2011).
We would like to express our sincere gratitude to Benito Medina, technical manager of Illa de Gràcia S.A. for generously allowing us access to their farm facilities and for facilitating our study. To Rhian Guillem, from the Gibraltar Botanic Gardens, for her comments on Saunders’ collection. A reviewer did a careful, and much needed, check of the whole text.
References
Antmaps. 2024. http://antmaps.org. (Accessed 5 May 2024). AntWeb. 2024. Version 8.106.1. California Academy of Science, online at https://www.antweb.org. (Accessed 5 May 2024).
Blatrix, R., Colin, T., Wegnez, P., Galkowski, C. & Geniez, P. 2018. Introduced ants (Hymenoptera: Formicidae) of mainland France and Belgium, with a focus on greenhouses. Annales de la Société Entomologique de France (N.S.), 54(4): 293-308. Bolton, B. 1987. A review of the Solenopsis genus-group and revision of Afrotropical Monomorium Mayr (Hymenoptera:
Formicidae). Bulletin of the British Museum of natural History (Entomology), 54: 263-452.
Borowiec, L. & Salata, S. 2021. Notes on ants (Hymenoptera: Formicidae) from Western Greece. Annals of the Upper Silesian Museum in Bytom Entomology, 30: 1-23.
Bračko, G. 2006. Review of the ant fauna (Hymenoptera: Formicidae) of Croatia. Acta Entomologica Slovenica, 14: 131156.
Camarasa, J.M., Folch, R., Masalles, R.M. & Velasco, E. 1977. El paisatge vegetal del delta de l’Ebre. Treballs de la Institució Catalana d’Història Natural, 8: 47-67.
Collingwood, C.A. 1978. A provisional list of Iberian Formicidae with a key to the worker caste (Hym. Aculeata) EOS, 52 (1976): 65-95.
Collingwood, C.A. & Yarrow, I.H.H. 1969. A survey of Iberian Formicidae (Hymenoptera). EOS, 44: 53-101.
Emery, C. 1916. Fauna entomológica italiana. I. Hymenoptera.Formicidae. Bullettino della Societa Entomologica Italiana, 47: 79-275.
Karaman, C. & Kiran, K. 2018. New tramp ant species for Turkey: Tetramorium lanuginosum Mayr (Hymenoptera: Formicidae). Trakya University Journal of Natural Sciences, 19(1), e1-e4.
Kiran, K. & Karaman, C. 2020. Additions to the ant fauna of Turkey (Hymenoptera, Formicidae). Zoosystema, 42(18), 285-329.
McGlynn, T. P. 1999. The worldwide transfer of ants: geographical distribution and ecological invasions. Journal of Biogeography, 26: 535-548.
Muller, G. 1923. Le formiche della Venezia Giulia e della Dalmazia. Bollettino della Società Adriatica di Scienze Naturali in Trieste, 28: 11-180.
Reyes-López J. & Carpintero, S. 2014. Comparison of the exotic and native ant communities (Hymenoptera: Formicidae) in urban green areas at inland, coastal and insular sites in Spain. European Journal of Entomology, 111(3): 421–428.
Reyes-López, J., Ordoñez-Urbano, C. & Carpintero-Ortega, S. 2008. Relación actualizada de las hormigas alóctonas de Andalucía (Sur de España). Boletín de la Asociación española de Entomología, 32 (1-2): 81-94.
Salata, S., Borowiec, L. & Trichas, A. 2020. Review of ants (Hymenoptera: Formicidae) of Crete, with keys to species determination and zoogeographical remarks. Monographs of the Upper Silesian Museum, 12, 5-96.
Saunders, E. 1888. On a collection of ants from Gibraltar and Tangier. Entomologist’s Monthly Magazine, 25: 17.
Saunders, E. 1890. Aculeate Hymenoptera collected by J. J. Walker, Esq., R.N., F.L.S., at Gibraltar and in North Africa. (Part IHeterogyna). Entomologist's Monthly Magazine, 26: 201-205, 289-291.
Schlick-Steiner, B.C., Steiner, F.M. & Pautasso, M. 2008. Ants and people: a test of two mechanisms potentially responsible for the large-scale human population-biodiversity correlation for Formicidae in Europe. Journal of Biogeography, 35: 2195-2206. Schifani, E. 2022. The new Checklist of the Italian Fauna: Formicidae. Biogeographia, 37, ucl006.
Tschinkel, W.R. 1989. Insect sociometry, a field in search of data. Notes from Underground, 2: 3-7.
Tschinkel, W.R. 2011. Back to basics: sociometry and sociogenesis of ant societies (Hymenoptera: Formicidae). Myrmecological News, 14: 49-54.
Wagner, H.C., Seifert, B., Borovsky, R. & Paill, W. 2018. First insight into the ant diversity of the Vjosa valley, Albania (Hymenoptera: Formicidae). Acta ZooBot Austria, 155: 315–321.
Amador Viñolas* & Josep Muñoz-Batet*
* Museu de Ciències Naturals de Barcelona. Laboratori de Natura. Coŀlecció d’artròpodes. Passeig Picasso, s/n. 08003 Barcelona. A/e: av.rodama@gmail.com
Rebut: 07.06.2024; Acceptat: 16.06.2024; Publicat: 30.06.2024
Resum
Con el presente trabajo se continua la revisión de las especies presentes en la Península Ibérica e Islas Baleares de la subfamilia Ernobiinae Pic, 1912, iniciado con el género Episernus C. G. Thomson,1863. Se realiza el estudio del género Ernobius C. G. Thomson, 1859 con catorce especies presentes en el área ibérica, de las que se relaciona el material estudiado y consultado, se da una redescripción de las mismas, se comenta su distribución y biología, y se complementa con el habitus de todas las especies, dibujos de las antenas de los machos, de los palpos maxilares y labiales, y de los edeagos con detalle de sus piezas.
Palabras clave: Coleoptera, Ptinidae, Ernobiinae, Ernobiini, Ernobius, revisión, Península Ibérica e Islas Baleares.
Abstract
The Ernobiinae of the Iberian Peninsula and Balearic Islands. 2nd note. The genus Ernobius C. G. Thomson, 1859 (Coleoptera: Ptinidae)
With the present study, the revision of the species present in the Iberian Peninsula and Balearic Islands of the subfamily Ernobiinae Pic, 1912, started with the genus Episernus C. G. Thomson, 1863, continues. The study of the genus Ernobius C. G. Thomson, 1859 is carried out with fourteen species present in the Iberian area, of which the studied and consulted material is related, a redescription of them is given, their distribution and biology are commented, and complemented with the habitus of all the species, drawings of the antennae of the males, of the maxillary and labial palps, and of the aedeagus with detail of their parts.
Key word: Coleoptera, Ptinidae, Ernobiinae, Ernobiini, Ernobius, review, Iberian Peninsula and Balearic Islands.
Resum
Els Ernobiinae de la Península Ibèrica i les Illes Balears. 2a nota. El gènere Ernobius C. G. Thomson, 1859 (Coleoptera: Ptinidae)
Amb aquest treball es continua la revisió de les espècies presents a la Península Ibèrica i Illes Balears de la subfamília Ernobiinae Pic, 1912, iniciat amb el gènere Episernus C. G. Thomson,1863. Es realitza l’estudi del gènere Ernobius C. G. Thomson, 1859 amb catorze espècies presents a l’àrea ibèrica, de les quals es relaciona el material estudiat i consultat, se’n dóna una redescripció, se’n comenta la distribució i la biologia, i es complementa amb l’habitus de totes les espècies, dibuixos de les antenes dels mascles, dels palps maxiŀlars i labials, i dels edeagus amb detall de les peces.
Paraules clau: Coleoptera, Ptinidae, Ernobiinae, Ernobiini, Ernobius, revisió, península Ibèrica i Illes Balears.
Introducción
En el año 2013 se inició una puesta al día del contenido genérico y específico de los Ptinidae presentes en la Península Ibérica e Islas Baleares con la revisión de las subfamilias Dorcatominae C. G. Thomson, 1859 y Mesocoelopinae Mulsant & Rey, 1864 (Viñolas, 2013a, b, c, d); en el año 2018 se inicia la revisión de la subfamilia Ernobiinae Pic, 1912 con el estudio del género Episernus C. G. Thomson, 1863 (Viñolas & Recalde-Iruzun, 1918) y en el 2019 se realiza la revisión de la familia Ptilininae (Viñolas & Muñoz-Batet, 2019). El contenido específico de la familia Ptinidae presente en Espa-
ñol (1992) había quedado desfasado por la descripción de nuevos taxones y a la localización por primera vez, en el área de estudio, de especies africanas y europeas, así como de otras introducidas, Aunque Zahradník (2014) en el Catálogo Paleártico contempla como válidas las tribus creadas por White (1982) (Calymmaderini, Gastrallini, Hadrobregnini, Nicobiini, Ozognathini, Prothecini, Stegobiini) y las continua incluyendo en el Catálogo Superior de las superfamilias Derodontoidea y Bostrichoidea (Zahradník & Háva, 2014), no se contemplan en el presente catálogo al estar invalidadas por el artículo 13.1 del Código de Nomenclatura Zoológica
A. VIÑOLAS & J. MUÑOZ-BATET
(Bouchard et al., 2011), así como la tribu Ochinini Zahradník, 2014 al ser un sin sentido al quedar anuladas las anteriores y su validez ser más que dudosa por los débiles caracteres empleados (grosor de los artejos de las antenas), variabilidad que presentan numerosos taxones incluidos en un mismo género (Viñolas, 2020). Así que las tribus indicadas en Viñolas & Recalde-Iruzun, 2019 no se deben de tener en cuenta, ya que en la actualidad no hay ninguna tribu indicada en la subfamilia (Bouchard et al., 2011).
En el presente trabajo se lleva acabo la revisión del género Ernobius complementándola con la clave específica. El género es el de mayor contenido específico, 14 especies, y requería una profunda revisión, sobre todo en lo concerniente a la representación gráfica de los edeagos, ya que los dibujos hasta ahora existentes daban pie a numerosas determinaciones erróneas al no representar fidedignamente las piezas internas y al no indicar, en numerosas ocasiones, si la visión era dorsal o ventral (Español, 1992). Para la correcta determinación del contenido específico del género es esencial el estudio del edeago, ya que el único carácter externo utilizable es la conformación de las antenas del macho, puesto que otros caracteres externos indicados, como la puntuación pronotal con puntos en forma de «C» (Español, 1992), no son correctos. La determinación de las hembras, con grandes diferencias en el tamaño de las antenas (Fig. 1, 2) sólo es posible en el grupo E. cupresi Choubaut, 1899 (Fig. 6) y prácticamente indiferenciables en el grupo E. parens (Mulsant & Rey, 1863).
Se relaciona el material estudiado o revisado de cada especie, se da su biología y distribución conocida, se representa el habi-
tus del macho de todas las especies, las antenas de los machos y los edeagos con separación de la pieza media de los mismos
En los ejemplares colectados del género Ernobius se procedió a la extracción del edeago, el cual tras tratamiento de limpieza, aclarado y extracción del aire se montó en preparación microscópica sobre una lámina de estireno transparente, de la marca Evergreen®, con líquido DMHF. En los ejemplares del género procedentes de las diferentes colecciones estudiadas, tras su reblandecimiento, se procedió de la misma forma que en los colectados. Todos los ejemplares se montaron en seco sobre cartulinas entomológicas.
Las fotografías se realizaron con una cámara Canon® modelo EOS 760D, con objetivo de microscopía y por el método de capas, con tratamiento de las imágenes mediante el programa Zerene Stacker®. Los dibujos se realizaron con el programa de Adobe® Ilustrator CS5, con la obtención de archivos PostScript® 3™
Acrónimos de las colecciones
MNCN Museo Nacional de Ciencias Naturales de Madrid MZB Museu de Ciències Naturals de Barcelona
IR José Ignacio Recalde Irurzun, Villaba- Atarrabia, Navarra
JM Josep Muñoz Batet, Girona
1-5. Antenas de Ernobius nigrinus: 1) macho; 2) hembra. Último artejo de los palpos: 3) maxilares; 4) labiales. 5) Abdomen de Ernobius Escalas: a, c = 0,5 mm; b = 0,01 mm.
JB Jamie Bookwalter, CREAF, Universitat Autònoma de Barcelona
AV Amador Viñolas, Barcelona
Subfamilia Ernobiinae Pic, 1912
Género Ernobius C. G. Thomson, 1859
Ernobius C. G. Thomson, 1859. Skandinaviens Coleoptera, Synoptiskt Bearbetade, Tome 1: 88
Conophoribium Chevrolat, 1862. Annales de la Société Entomologique de France, (4) 1 (1861): 391
Philoxylon LeConte, 1861. Smithsonian Miscellaneous Collection: 205
Liozoum Mulsant & Rey, 1863. Opuscules Entomologiques, 13: 92
Especie tipo: Dermestes mollis Linnaeus, 1758, designación original, por monotipia.
Cuerpo de contorno subparalelo, poco convexo y con la superficie cubierta de corta y densa pubescencia. Antenas de once artejos con maza terminal de tres artejos, muy desarrollada o poco indicada, las de las hembras más cortas y con la maza menos desarrollada (Figs. 1-2, 6-19). Ojos salientes. Último artejo de los palpos maxilares y labiales rectangulares (Figs. 3-4); Protórax transverso, con la máxima anchura en la base, ángulos posteriores redondeados y con los márgenes laterales explanados o no; con la superficie cubierta de punteado bien indicado. Élitros con un punteado fino, denso y desordenado. Abdomen (Fig. 5) con seis segmentos de la misma longitud, excepto el sexto algo más largo. Meso- y metasternón planos, coxas anteriores muy próximas, las posteriores más separadas. Tibias y tarsos gráciles. Edeago asimétrico, con el lóbulo medio muy bien caracterizado, así como los parámeros (Figs. 20-33).
Clave de las especies presentes en la Península Ibérica, válidas para los machos
1 La suma de los artejos del funículo más corta o igual que el primer artejo de la maza antenal (Figs. 6, 10, 13-14) .... ........................................................................................2 –
La suma de los artejos del funículo algo o mucho más larga que el primer artejo de la maza antenal (Figs. 7-9, 11-12, 15-19) ..................................................................6
2 La suma de los artejos de funículo más corta que el primer artejo de la maza antenal (Fig. 6); edeago según figura 20 E. cupresi Chobaut, 1899 – La suma de los artejos del funículo igual o algo más corta que el primer artejo de la maza antenal (Figs. 10, 13-14); edeago según figuras 24, 27-28 ......................................4
4 Talla grande entre 4,5 y 7,0 mm; antenas según figura 10; edeago según figura 24 E. laticollis Pic, 1927 –Talla menor entre 3,0 y 4 mm; antenas según figura 1314; edeagos según figuras 27-28 .....................................5
5 Antena según figura 13; edeago según figura 27 .............. E. nigrinus (Sturm, 1837) – Antena según figura 14; edeago según figura 28 ................................................E. pallidipennis Pic, 1902
6 Superficie dorsal del cuarto artejo de los tarsos con la excavación apical no sobrepasando, o apenas, la mitad del artejo ...............................................................................7
– Superficie dorsal del cuarto artejo de los tarsos con la excavación apical sobrepasando la mitad del artejo y alcanzando los tres cuartos de la longitud del mismo ...........10
7 Protibias curvadas hacia dentro; antenas según figura 8; edeago según figura 23 E. gigas (Mulsant & Rey, 1863)
Protibias rectas, con el ápice vuelto hacia fuera .............8
8 Puntuación protonal poco densa, pero muy marcada; antenas según figura 11; edeago según figura 25
.................................... E. lucidus (Mulsant & Rey, 1863)
– Puntuación protonal más densa y fina; antenas figuras 12 y 18; edeago según figuras 26, 32 9
9 Talla comprendida entre 5 y 6 mm; protórax anchamente explanado en los márgenes laterales; antenas según figura 18; edeago según figura 32
E, reflexus (Mulsant & Rey, 1863)
– Talla comprendida entre 3 y 5 mm; protórax poco o nada explanado en los márgenes laterales; antenas según figura 12; edeago según figura 26
..................................... E. mollis mollis (Linnaeus, 1758)
10 Antenas con el séptimo y octavo artejos del funículo juntos más cortos que el primer articulo de la maza antenal (Fig. 17); edeago según figura 31 ..................................... E. pruinosus (Mulsant & Rey, 1863) – Antenas con el séptimo y octavo artejos del funículo juntos igual o más largos que el primer artejo de la maza antenal (Figs. 7, 9, 15-16,19); edeago según figuras 21-22, 29-30, 33 11
11 Puntos del protórax muy marcados; antenas según figuras 16, 19; edeago según figuras 30, 33 ..............................12 – Puntos del protórax menos marcados; antenas según figuras 7, 9, 15; edeago según figuras 21, 29 13
12 Cuerpo de color moreno amarillento; antenas según figura 19; edeago según figura 33 E. rufus (Illiger, 1807) – Cuerpo de color moreno rojizo; antenas según figura 16: edeago según figura 30 ................... E. pini (Sturm, 1837)
13 Cuerpo de color rojizo; antenas según figuras 7, 15; edeago según figuras 21, 29 .................................................14
– Cabeza y protórax negruzcos, élitros rojizos; antena según figura 9; edeago según figura 22
E. gallicus Johnson, 1975
14 Antenas según figura 7; edeago según figura 21
E. fulvus C. Johnson, 1975 – Antenas según figura 15; edeago según figura 29 .....................................
E. parens (Mulsant & Rey, 1863)

16) E. pini; 17) E. pruinosus; 18) E. reflexus; 19) E, rufus. Escala = 0,5 mm,
Ernobius cupressi Chobaut, 1899 (Fig. 34)
Ernobius cupressi Chobaut, 1899. Bulletin de la Société
Entomologique de France: 104
Longitud de 1,8 a 2,8 mm. Cuerpo rojizo más o menos negruzco, superficie con la pubescencia muy corta y dispersa. Antenas con la suma de los artejos del funículo más corta que el primer artejo de la maza antenal, que está muy desarrollada (Fig. 6). Protórax transverso, 1,4 veces más ancho que largo, con la mayor anchura en la base, márgenes poco explanados y de contorno rectilíneo, superficie fuertemente punteada. Élitros 1,81 veces más largos que anchos tomados conjuntamente. Edeago (Fig. 20a) con los parámeros estrechos y aguzados y el lóbulo medio muy ancho (Fig. 20b)
Material estudiado
Butlletí ICHN
(8), 2024
1 ♂, etiquetado: «6-V-1989, Coll de Jou, Tarragona» (AV).
Material revisado
1 ♂, etiquetado: «19-VII-1974, Vall d’Aneu, Lleida, J. R. Duplà leg.» (MZB 83-7444).
Distribución
Especie conocida sólo de Chipre, España, Francia y Grecia. En el área peninsular sólo se ha localizado en Cataluña (Español, 1990; Johnson, 1975; Viñolas, 2020).
España: Girona (Cap de Creus); Lleida (Vall d’Aneu); Tarragona (Coll de Jou).
Biología
La serie típica fue recolectada en Cupressus sempervirens var. stricta Aiton (= fastigata De Cand.). En Francia se ha recolectado batiendo Cupressus sp.
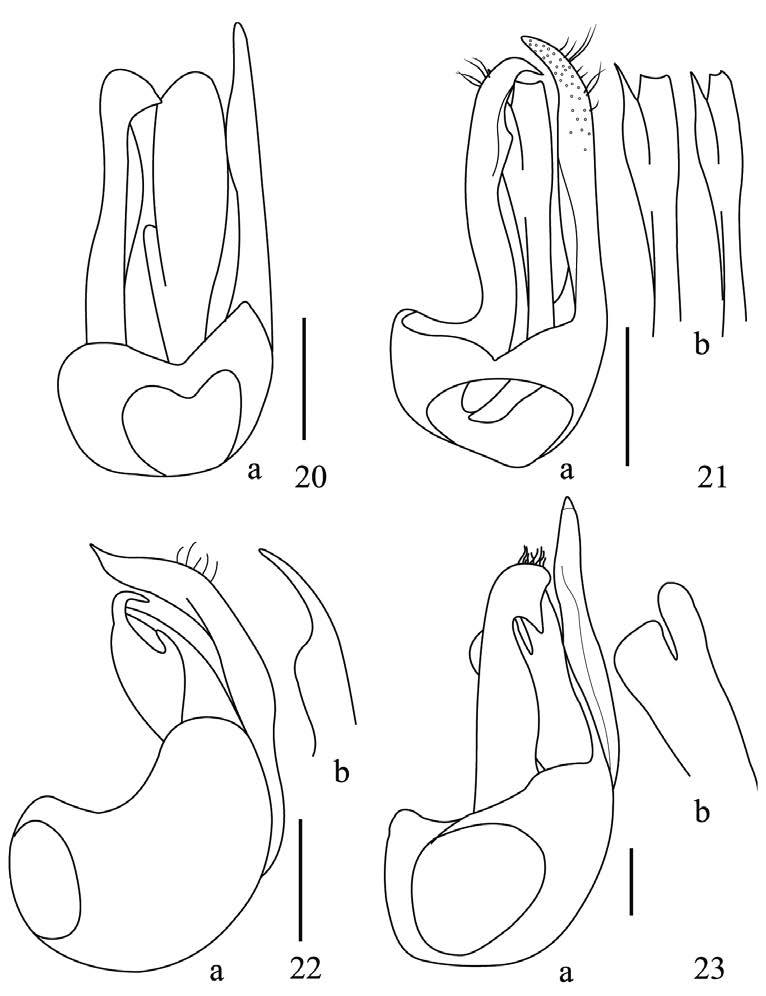
Figuras 20-23. a) Edeago en visión ventral y b) pieza media de: 20) Ernobius cupressi Chobaut, 1899; 21) E. fulvus Johnson, 1975; 22) E. gallicus Johnson, 1975; 23) E. gigas (Mulsant & Rey, 1863). Escala = 0,2 mm.
Ernobius fulvus Johnson, 1975 (Fig. 35)
Ernobius fulvus Johsnson, 1975. Entomologische Blätter, 71 (2): 89
Ernobius vinolasi Novoa & Baselga, 2000. The Coleopterits Bulletin, 54 (3): 403
Longitud de 2,5 a 4,0 mm. Cuerpo rojizo obscuro, con la pubescencia de la superficie larga. Antenas (Fig. 7) con la suma de los artejos del funículo más larga que la de la maza antenal. Protórax poco transverso, 1,33 veces más ancho que largo, con la mayor anchura cerca de la base, los márgenes poco explanados y los ángulos posteriores muy redondeados, superficie densamente punteada. Élitros 1,77 veces más largos que anchos, tomados conjuntamente. Edeago (Fig, 21a)
con los parámeros con distinta configuración y el lóbulo medio ancho y escotado en el ápice (Fig. 21b).
Material revisado
3 ♂, etiquetados: 1 ♂ «Corse» «Holotype» «Ernobius fulvus n. sp. C. Jonshon det.» (MZB 78-1297); 1 ♂ «4-VII-1963, Col de Bavella, Corse, G. Tempère leg.» «Ernobius fulvus C. Jonshon 1975, F. Español det.» (MZB 83-7455); 1 ♂ «Isla de San Martín, Vigo (Pontevedra), 13-VII-1997, 10 m, 29TNG0872, F. Novoa leg.» «Holotype, Ernobius vinolasi n. sp., F. Novoa & A. Balsega det.» (MNCN 8634).
Distribución
Conocida de las islas de Cerdeña, Córcega y Sicilia (Bar-
A. VIÑOLAS & J. MUÑOZ.BATET
nouin, 2014; Johnson, 1975). Su presencia en las Islas Cies (Pontevedra) se debió, seguramente, a una introducción en la repoblación forestal realizada con resinosas en la década de los 60 en el noroeste peninsular (Novoa & Baselga, 2000; Viñolas, 2002, 2018, 2020; Viñolas & Masó, 2013). España: Pontevedra (Isla de San Martiño, Islas Cíes).
Biología
Los ejemplares de la serie típica se obtuvieron por emergencia de Pinus nigra var. calabrica (Loudon) C.K. Schneid (Johsnson, 1975). En Córcega se ha recolectado sobre P. nigra subsp. salzmani var. laricio (Loudon) Hylander. En Pontevedra se capturo en madera de P. pinaster Aiton (Novoa & Baselga, 2000).
Ernobius gallicus Johnson, 1975 (Fig. 36) Ernobius gallicus Johnson, 1975. Entomologische Blätter, 71 (2): 89
Longitud de 2,0 a 3,0 mm. Cuerpo negruzco, con los élitros más o menos rojizos, superficies con la pubescencia larga y densa. Antenas (Fig.9) con la suma de los artejos del funículo ligeramente más larga que la suma de los artejos de la maza. Protórax ligeramente transverso, 1.33 veces más ancho que largo, con la mayor anchura cerca de la base, con los ángulos posteriores muy redondeados y la superficie fuertemente punteada. Élitros 1,73 veces más largos que anchos tomados conjuntamente. Edeago (Fig. 22a) con el parámero izquierdo ancho, corto y escotado en forma de «c» y el derecho largo y fino, lóbulo medio (Fig.22b) estrechado y aguzado en el ápice.
Material estudiado
2 ♀, etiquetadas: 1 ♀ «IV-2011, Turo de Balasc, Collserola, Barcelona, J. Mederos leg.» (AV); 1 ♀ «1/31-V-2011, Turo de Balasc, Collserola, Barcelona, J. Mederos leg.» (AV).
Material revisado
2 ♂ y 3 ♀, etiquetados: 1 ♂ y 3 ♀ «28-VII-1959, Montserrat, Barcelona, H. Freude leg.» (MZB, 83-4524, 83-7452, 837453, 83-7465); 1 ♂ «5-VII-1964, Montserrat, Barcelona, H. Freude leg.» (MZB, 83-4523).
Distribución
Especie descrita de Arcachon (Francia) (Viñolas & Masó, 2013), conocida sólo de España y Francia. En el área peninsular, de momento, sólo se ha localizado en Cataluña (Español, 1977, 1990, 1992; Viñolas, 2020) y con una excéntrica cita, a comprobar, de Portugal (Bercedo et al., 2008).
España: Barcelona (Matadepera, Montserrat, Parque Natural de Collserola, Parque Natural de Sant Llorenç de Munt i l’Obac); Lleida (Solsona). Portugal: Setúbal (Almada).
Biología
La mayoría de ejemplares conocidos de la especie se han capturado batiendo ramas de Pinus sp. Los del Parque Natural de Collserola con trampa Malaise situada en bosque de Pinus halepensis Miller. Los de Portugal se recolectaron con trampas de luz UV situadas en un bosque de P. pinaster Aiton.
Butlletí ICHN 88 (8), 2024 GEA, FLORA ET FAUNA
Ernobius gigas (Mulsant & Rey, 1863) (Fig. 37)
Liozoum gigas Mulsant & Rey, 1863. Opuscules Entomologiques, 13: 113
Ernobius gigas (Mulsant & Rey): Heyden, Reitter & Weise, 1891. Catalogus Coleopterorum Europae, Caucasi et Armeniae Rossicae: 462
Ernobius mollis var. subelongatus Pic, 1914. L’Échange, Revue Linnéenne, 30: 34
Ernobius mulsantianus Sharp, 1916. The Entomologist’s Monthly Magazine, 52: 221
Longitud de 4,0 a 8,0 mm. Cuerpo de color rojizo más o menos obscuro y con la pubescencia larga. Antenas largas (Fig. 8) con la suma de los artejos del funículo más larga que la de la maza antenal. Protórax transverso 1,36 veces más ancho que largo, con su máxima anchura cerca de la base, ángulos posteriores y márgenes muy redondeados, superficie con punteado disperso y poco marcado. Élitros largos, 2,05 veces más largos que anchos tomados conjuntamente. Edeago (Fig. 23a) con el parámero izquierdo ancho y el derecho alargado, lóbulo medio (Fig. 23b) ancho y escotado en el ápice.
Material estudiado
1 ♂, etiquetado: «16-III-1988, Sant Sadurni d’Osormort, Bellavista & Dantart leg.» (AV).
Material revisado
5 ♂♀, etiquetados: 1 ej. «Seva, Barcelona» (MZB 76-5241); 1 ej. «Menorca, Islas Baleares, Cuello leg.» (MZB 83-7488); 1 ej. «VIII-1937, Tortosa, Tarragona» (MZB 83-7482); 1 ej. «VIII-1965, Jaca, Huesca, L. Auroux leg. (MZB 83-7481); 1 ej. «25-VIII-1988, Sant Martí Sesgueioles, Anoia, Barcelona, J.J. Pérez de Gregorio leg.» (MZB 83-7466).
Distribución
La especie ha sido citada de los siguientes países: Alemania, Argelia, España, Francia, Gran Bretaña, Italia, Letonia, Lituania, Rusia, Suiza, Ucrania y Turquía. En la Península Ibérica ha sido recolectada en el noreste español, sur de Portugal e Islas Baleares (Español, 1977, 1995; Johnson, 1975; López-Colón & Agoiz-Bustamante, 2003; Viñolas, 2020; Viñolas & Muñoz-Batet, 2016).
España: Barcelona (Sant Martí Sesgueioles, Sant Sadurni d’Osormort, Seva); Girona (la Jonquera); Huesca (Jaca); Navarra (Valtierra); Tarragona (Ports de Tortosa).
Portugal: Faro (Lagos, Meia Praia).
Islas Baleares: Menorca (sin indicación).
Biología
Poco se sabe sobre su biología, aunque es una especie asociada a Pinus quemados o muertos.
Ernobius laticollis Pic, 1927 (Fig. 38)
Ernobius laticollis Pic, 1927. L’Échange, Revue Linnéenne, 43: 9
Ernobius mollis var. granulosus Pic, 1939. L’Échange, Revue Linnéenne, 55 (475): 17
Longitud de 4,5 a 7,0 mm. Cuerpo rojizo más o menos obscuro, superficie con la pubescencia larga y poco densa.
Antenas (Fig. 10) con la suma de los artejos del funículo igual de larga que la suma de los dos primeros artejos de la maza antenal. Protórax muy transverso, 1,58 veces más ancho que largo, con la máxima anchura cerca de la base, los ángulos posteriores muy redondeados, los márgenes ligeramente explanados y redondeados. Élitros 1,9 veces más largos que anchos, tomados conjuntamente. Edeago (Fig. 24a) con una conformación muy característica del parámero izquierdo, así como la del lóbulo medio (Fig, 24b)
Material estudiado
2 ♂ y 1 ♂, etiquetados: 1 ♂ «17-XII-2018, Villanueva de Huerva, Zaragoza, E. Piera leg.» (JM); 1 ♀ «28-XII-2018, Vi-
llanueva de Huerva, Zaragoza, E. Piera leg.» (JM); 1 ♂ «7-II2019, Villanueva de Huerva, Zaragoza, E. Piera leg.» (AV).
Material revisado
5 ♂, etiquetados: 3 ♂ «X-1940, Vallvidrera, Barcelona, F. Español leg.» (MZB 83-7501, 83-7502, 83-7503); 1 ♂ «10XI-1940, Vallvidrera, Barcelona, J. Mateu leg.» (MZB 837505); 1 ♂ «VII-1973, Valldoreix, Barcelona, F. Español leg.» (MZB 83-7504).
Distribución
Especie con una distribución conocida un poco irregular, citada de unos pocos países europeos y africanos: Austria,
GEA, FLORA ET FAUNA
A. VIÑOLAS & J. MUÑOZ.BATET
España, Francia, Grecia, Italia, Suiza y Túnez. En el área peninsular sólo se ha localizado en la zona norte, sin citas de Portugal (Español, 1977, 1992; Johnson, 1975; LópezColón, 2001; Recalde Irurzun & San Martín Moreno, 2017; Viñolas, 2020).
España: Barcelona (Valldoreix, Vallvidrera, Sant Cugat, Tavertet); Navarra (Garde); Salamanca (El Payo, Peñaparda, Robleda); Zaragoza (Villanueva de Huerva).
Biología
Se desarrolla en la gruesa corteza de Pinus halepensis Miller, P. nigra J.F. Arnold y P. sylvestris L Los ejemplares de Tavertet fueron recolectados sobre P. sylvestnis; los de Vallvidrea y Sant Cugat en una área boscosa de P. halepensis con algún pie de P. pinea L.; los de Zaragoza con trampa de interceptación de vuelo situadas en un pinar de P. halepensis
Ernobius lucidus (Mulsant & Rey, 1863) (Fig. 39)
Liozoum lucidum Mulsant & Rey, 1863. Opuscules Entomologiques, 13: 109 Ernobius lucidus (Mulsant & Rey): Kiesenwetter, 1877. Naturgeschichte der Insecten Deutschlands.: 123 Ernobius lucidipennis Pic, 1914. L’Échange, Revue Linnéenne, 30: 34
Longitud de 4,0 a 6,0 mm. Cuerpo más o menos rojizo obscuro, pubescencia de la superficie corta y densa, Antenas (Fig. 11) con la suma de los artejos del funículo más corta que la suma de los artejos de la maza antenal. Protórax muy transverso, 2 veces más ancho que largo, con la máxima anchura en la base, ángulos posteriores redondeados, superficie con la puntuación poco impresa. Élitros 2,0 veces más largos que anchos, tomados conjuntamente. Edeago (Fig. 25a) muy bien caracterizado por la conformación del parámero izquierdo y el lóbulo medio (Fig. 25b).
Material estudiado
13 ♂ y 12 ♀, etiquetados: 2 ♂ y 3 ♀ «26-VIII-1989, Siles, Jaén, J. Lencina leg.» (AV); 7 ♂ y 9 ♀ «10-VIII-2013, El Tillar, Serra de Prades, Tarragona, A. Viñolas leg.» (AV); 1 ♂ «12-IX-2017, Es Caló de Sant Agusti, Formentera, Baleares, M. Rondós & M. Macià leg.» (AV); 1 ♂ «6-IX-2018, Es Caló de Sant Agusti, Formentera, Baleares, J. Muñoz leg.» (AV); 1 ♂ «15-30-IX-2022, Bardenas Reales, Navarra, J.I. Recalde et al leg.» (IR); 1 ♂ «1-15-X-2022, Bardenas Reales, Navarra, J.I. Recalde et al leg.» (IR).
Material revisado
2 ♂ y 2 ♀, etiquetados: 1 ♂ «Mallorca, G. Jordá leg.» (MZB 83-7506); 1 ♀ «Santa Eulàlia de Ronsana, Barcelona, J. Ribes leg.» (MZB 83-7508); 1 ej. «VIII-1974, Eivissa, N. Ferrer leg.» (MZB 83-7509); 1 ♂ y 1 ♀ «21-IX-1987, La Joya, Almería, J. Mateu leg.» (MZB 83-7511, 83-7512) .
Distribución
Conocida de España, Francia meridional, Portugal e Islas Baleares, citada con dudas del norte de Italia. El catálogo paleártico la cita de Alemania (Zahradník, 2014), localización poco creíble al estar el área de distribución de esta especie
Butlletí ICHN 88 (8), 2024
circunscrita al extremo sur occidental de Europa. Con una amplia distribución peninsular y balear (Audisio et al., 1995; Johnson, 1975; Español, 1977, 1992; Gottwald, 1977; Laclos et al., 2007; López-Colón, 2001; Viñolas, 2020; Viñolas & Muñoz, 2017; Viñolas et al., 2015).
España: Almeria (La Joya); Barcelona (Granollers, Santa Eulàlia de Ronçana); Jaén (Siles); Madrid (Sierra de Guadarrama); Málaga (Torremolinos); Navarra (Bardenas Reales, Cascante); Segovia (Navafría); Tarragona (Vimbodí i Poblet); Zaragoza (Valmadrid).
Portugal: Leiria (São Pedro de Moel).
Islas Baleares: Eivissa (Sant Joan de Labritja, sin indicación); Formentera (Es Caló de Sant Agustí); Mallorca (Pollença, sin indicación); Menorca (sin indicación).
Biología
Como todas las especies del género está asociada a coníferas, con preferencia a Pinus halepensis Miller y P. pinaster Aiton.
Ernobius mollis mollis (Linnaeus, 1758) (Fig. 40)
Dermestes mollis Linnaeus, 1758. Systema Naturae per regna tria nature: 355
Ptinus mollis (Linnaeus): Linnaeus, 1767. Systema Naturae per regna tria nature: 565
Anobium molle (Linnaeus): Fabricius, 1775. Systema entomologiae: 62
Anobium testaceus Kugelann, 1792. Neuestes Magazin für die Liebhaber der Entomologie, 1 (4): 489
Anobium sybaris Kugelann, 1792. Neuestes Magazin für die Liebhaber der Entomologie, 1 (4): 490
Anobium convexifrons Melsheimer, 1846. Proceedings of the Academy of Natural Sciences of Philadelphia, 2 (18441845): 309
Ernobius mollis (Linnaeus): C.G. Thomson, 1859. Skandinaviens Coleoptera, Synoptiskt Bearbetade, Tome 1: 88
Liozoum molle (Linnaeus): Mulsant & Rey, 1863. Opuscules Entomologiques, 13: 117
Liozoum sulcatulum Mulsant & Rey, 1863. Opuscules Entomologiques, 13: 111
Liozoum consimile Mulsant & Rey, 1863. Opuscules Entomologiques, 13: 117
Liozoum consimile var. laetum Mulsant & Rey, 1864. Histoire naturelle des Coléoptères de France. Térédiles: 171
Ernobius consimilis (Mulsant & Rey): Kiesenwetter, 1877. Naturgeschichte der Insecten Deutschlands.: 124
Ernobius tarsatus Kraatz, 1881. Deutsche Entomologische Zeitschrift, 25: 302
Ernobius reversus Sharp, 1916. The Entomologist’s Monthly Magazine, 52: 222
Longitud de 3,0 a 6,5 mm. Cuerpo de color rojizo amarillento, superficie con la pubescencia no muy larga y densa. Antenas (Fig.12) con la suma de los artejos del funículo ligeramente más corta que la suma de los artejos de la maza. Protórax convexo, 1,36 veces más ancho que largo, con la máxima anchura cerca de la base, los márgenes ligeramente explanados y de contorno curvado, ángulos posteriores re-
28-31. a) Edeago en visión ventral y b) pieza media de: 28) Ernobius pallidipennis Pic, 1902; 29) E. parens (Mulsant & Rey, 1863); 30) E. pini pini (Ratzeburg, 1837); 31) E. pruinosus (Mulsant & Rey, 1863). Escala = 0,3 mm.
dondeados, superficie fuerte y densamente punteada. Élitros 1,75 veces más largos que anchos tomados conjuntamente. Edeago (Fig.26a) con el parámero izquierdo muy bien caracterizado en el ápice, lóbulo medio ancho (Fig. 26b) y escotado en el ápice.
Material estudiado 17 ♂♀, etiquetados: 2 ej. «16/25-VI-2012, El Tillar, Vimbodí i Poblet, serra de Prades, Tarragona, F. Benvenuti leg.» (AV); 1 ej. «4/13-VII-2012, El Tillar, Vimbodí i Poblet, serra de Prades, Tarragona, A. Viñolas leg.» (AV); 3 ♂ y 2 ♀ «10-VII2013, El Tillar, Vimbodí i Poblet, serra de Prades, Tarragona, A. Viñolas leg.» (AV); 1 ♂ «30-VII-2017, Canya de la Rabassa, el Serrat, Ordino, Andorra, S07ILFa2, CREAF leg.»
(AV); 1 ej.. «25-VI-2021, Baga de Ventallola, Setcases, 1950 m, Ripollès, Girona, A. Planella & E. Piera leg.» (JM); 2 ej. «25-VII-2021, Baga de Ventallola, Setcases, 1950 m, Ripollès, Girona, A. Planella & E. Piera leg.» (JM); 4 ej. «28VIII-2021, Baga de Ventallola, Setcases, 1950 m, Ripollès, Girona, A. Planella & E. Piera leg.» (JM); 1 ej. «13-IX-2021, Baga de Ventallola, Setcases, 1950 m, Ripollès, Girona, A. Planella & E. Piera leg.» (JM).
Material revisado
10 ♂♀, etiquetados: 1 ej. «Capsanes, Tarragona, J. Vives leg.» (MZB 83-7692); 1 ej. «23-IV-1911, Masnou, Barcelona» (MZB 83-7567); 1 ej. «7/21-V-1956, Pego, Alicante, H. Coiffait leg.» (MZB 83-7645); 1 ej. «2-III-1962, Barcelona
A. VIÑOLAS & J. MUÑOZ-BATET
caja de cría, F. Español leg.» (MZB 83-7569); 1 ej. «14-V1974, Barcelona ciudad» (MZB 83-7695); 1 ♂ «12-IV-1979, Mediona, Barcelona, F. Español leg.» (MZB 76-6198); 1 ♂ y 1 ♀ «VIII-1981, Seròs, Lleida, T. Yélamos leg.» (MZB 83-4526, 83-7676); 1 ♂ «13-V-1989, Revolcadores, Cañada de la Cruz, Murcia, X. Vázquez leg.» (MZB 83-7660); 1 ♀ «VIII-2010, Margalef, Tarragona, J. J. de Gregorio & F. Vallhonrat leg.» (MZB 2014-1833).
Distribución
Con una amplia distribución paleártica (Europa, África y Asia) ha sido introducida en las regiones Afrotropical, Australiana, Neártica, Neotropical y Oriental, actualmente se la puede considerar una especie cosmopolita. Presente en toda el área peninsular y balear (Corrêa de Barros, 1907; Español, 1964, 1977; Fuente, 1932; Johnson, 1975; Pérez Moreno & Moreno Grijalba, 2009; Sakai, 2002; Viñolas & Ghahari, 2017; Viñolas, 2020, 2022; Viñolas et al., 2013, 2015, 2020, 2022; Viñolas & Bookwalter, 2018; Viñolas & Muñoz-Batet, 2022).
Andorra: Ordino (Besalí, el Serrat).
España: Alicante (Pego); Barcelona (Barcelona, el Bruc, Masnou, Mediona); Bizkaia (Sarria), Girona (Setcases), La Rioja (Lumbreras, Villoslada de Cameros); Lleida (Seròs); Murcia (Revolcadores); Tarragona (Capsanes, Cornudella de Montsant, Margalef, Vimbodí i Poblet), Valencia (Cortes de Pallás).
Portugal: Vila Real (Castedo).
Islas Baleares: Mallorca (Palma); Menorca (sin indicación).
Biología
Se desarrolla en la corteza de resinosas, en las ramas muertas y en sus piñas. Se ha citado de las siguientes especies Larix decidua Mill., 1768, Picea abies (L.) H.Karst. 1881, Pinus heldreichii Christ, P. sylvestris L., P. nigra J.F.Arnold, P. pinaster
Aiton, P. strobus L., P. taeda L. P. uncinata Ramond ex A.DC y Pseudotsuga menziesii (Mirb.) Franco. Algunas especies de Ernobius pueden atacar la madera muerta siempre que conserve la corteza. En el área peninsular se ha recolectado en P. sylvestris, P. uncinata y en piñas mediante caja de cría
Ernobius nigrinus (Sturm, 1837) (Fig. 41)
Byrrhus mollis Geoffroy, 1785. Entomologia parisiensis: 26
Anobium nigrinum Sturm, 1837. Die Insekten. Deutschland Insekten, Käfer. Vol. 11: 126
Anobium politum Redtenbacher, 1847, Fauna Austriaca: 346 (non Duftschmid, 1825)
Liozoum fuscum Mulsant & Rey, 1863. Opuscules Entomologiques, 13: 131
Liozoum nigrinum (Sturm): Mulsant & Rey, 1863. Opuscules Entomologiques, 13: 133
Ernobius canaliculatus C. G. Thomson, 1871. Opuscula Entomologica, 4: 380
Ernobius nigrinus (Sturm): Kiesenwetter, 1877. Naturgeschichte der Insecten Deutschlands.: 129
Ernobius fuscus (Mulsant & Rey): Kiesenwetter, 1877. Naturgeschichte der Insecten Deutschlands : 129
Butlletí ICHN 88 (8), 2024 GEA, FLORA ET FAUNA
Ernobius pueli Lavagne, 1914. Bulletin de la Société Entomologique de France: 137
Ernibius pueli var. berardi Lavagne, 1914. Bulletin de la Société Entomologique de France: 137
Ernobius pallidipennis var. mayeti Pic, 1914. L’Échange , Revue Linnéenne, 30: 27
Ernobius nigrinus var. rufescens Pic, 1916. L’Échange, Revue Linnéenne, 32: 3 (nota)
Ernobius nigriclava Roubal, 1917. Archiv für Naturgeschichte, A 82 (3) (1916): 51
Longitud de 3,1 a 4,0 mm. Antenas, cabeza y protórax de color negruzco, élitros rojizos más o menos negruzcos, superficie con la pubescencia corta y densa. Antenas (Fig. 13) con la suma de los artejos del funículo inferior a la suma de los dos primeros artejos de la maza, muy desarrollados. Protórax convexo, 1,5 veces más ancho que largo, con la máxima anchura en la base, márgenes explanados junto a los ángulos posteriores que están redondeados. Élitros 1,9 veces más largos que anchos tomados conjuntamente. Edeago (Fig. 27a) con el parámero izquierdo ancho con el ápice en forma de pico, el derecho ensanchado en el ápice, lóbulo medio (Fig. 27b) ancho y fuertemente escotado lateralmente.
Material estudiado
7 ♂ y 98 ♀, etiquetados: 1 ♀ «15/29-VI-2013, Garde, Navarra, I. Recalde leg.» (IR); 2 ♂ «2014, Iriso, Navarra, Robledal, Recalde & San Martín leg.» (AV, IR); 1 ♂ y 2 ♀ «30V/13-VI-2015, Mace, Isaba, Navarra, I. Recalde leg.» (AV, IR); 1 ♀ «20-V-2017, Isaba, Navarra, I. Recalde leg» (IR); 2 ♀ «8/17-VI-2017, Isaba, Navarra, I. Recalde leg.» (AV); 1 ♀ «12-VII-2017, Bordes de la Mollera, Llorts, Ordino, Andorra, S10ILMa1, CREAF leg.» (AV); 1 ♀ «13-VII2017, Pleta de Llom, El Serrat, Ordino, Andorra, S12IHFb1, CREAF leg.» (AV); 1 ♂ «13-VII-2017, Canya de la Rabassa, El Serrat, Ordino, Andorra, S07ILMa1, CREAF leg.» (AV); 1 ♂ «13-VII-2017, Besalí, El Serrat, Ordino, Andorra, S07IHFa1, CREAF leg.» (AV); 1 ♂ «12-VII-2017, Comis Vell, Arcalis, Ordino, Andorra, CREAF leg.» (AV); 1 ♂ y 1 ♀ «1-X-2017, Pleta de Llors, El Serrat, Ordino, Andorra, CREAF leg.» (JB); 31 ♀ «25-VI-2021, Baga de Ventallola, Setcases, 1950 m, Ripollès, Girona, A. Planella & E. Piera leg.» (AV, JM.); 57 ♀ «8-VII-2021, Baga de Ventallola, Setcases, 1950 m, Ripollès, Girona, A. Planella & E. Piera leg.» (AV, JM); 27 ♀ «25-VII-2021, Baga de Ventallola, Setcases, 1950 m, Ripollès, Girona, A. Planella & E. Piera leg.» (JM); 1 ♀ «13-IX-2021, Baga de Ventallola, Setcases, 1950 m, Ripollès, Girona, A. Planella & E. Piera leg.» (JM)
Distribución
Especie conocida de numerosos países europeos, citada de: Alemania, Armenia, Austria, Azerbaiján, Bélgica, Bosnia, Bulgaria, Bielorrusia, Croacia, Dinamarca, Eslovaquia, Eslovenia, España, Estonia, Finlandia, Francia, Gran Bretaña, Georgia, Grecia, Holanda, Hungría, Italia, Letonia, Lituania, Noruega, Polonia, República Checa, Rumanía, Rusia, Suecia, Suiza, Turquía y Ucrania. En el área de estudio sólo se conoce del norte peninsular, con pocas y recientes citas (Español, 1977, 1992; Johnson, 1975; Pérez Moreno & Moreno Grijal-
13-14. a) Edeago en visión ventral y b) pieza media de: 13) Ernobius reflexus (Mulsant & Rey, 1863); 14) E. rufus (Illiger, 1807). Escala = 0,3 mm.
ba, 2009; Viñolas, 2017, 2020, 2022; Viñolas & Bookwalter, 2018, Viñolas & Muñoz-Batet, 2022). El catálogo paleártico cita la especie de las Islas Canarias, localización poco creíble y que debería de obviarse (Viñolas, 2017).
Andorra: OrdinO (Arcalis, El Serrat, Llorts).
España: Girona (Setcases), Huesca (Selva de Oza, Valle de Hecho), La riOja (Lumbreras), LLeida (Àger), Navarra (Iriso, Isaba), Segovia (San Rafael).
Biología
Se desarrolla en las ramas delgadas de Pinus cembra L., P. sylvestris L., P. uncinata Raymond ex A.DC. y Picea abies (L.) H. Karst., después del ataque primario de las mismas por especies de Curculionidae de los géneros Magdalis Germar, 1817 y Hylastes Erichson 1836.
Ernobius pallidipennis Pic, 1902 (Fig. 42)
Ernobius pallidipennis Pic, 1902. L’Échange, Revue Linnéenne, 18: 79
Ernobius diversepunctatus Pic, 1914. L’Échange, Revue Linnéenne, 30: 33
Longitud de 3,1 a4,2 mm. Cuerpo rojizo, con la cabeza y protórax negruzcos, superficie con la pubescencia muy corta y densa. Antenas (Fig. 14) con la suma de los artejos del funículo ligeramente más larga que la longitud del primer artejo de la maza. Protórax transverso, 1,35 veces más ancho que largo, con la máxima anchura en la base, márgenes ligeramente explanados en la mitad basal, superficie con el punteado bien marcado y nada contiguo. Élitros 2,82 veces más largos que anchos tomados conjuntamente. Edeago (Fig. 28a) con el parámero izquierdo corto y con el borde interno
sinuado, el derecho fino y ligeramente curvado, lóbulo medio (28b) no muy ancho y con el ápice curvado.
Material estudiado
6 ♂ y 8 ♀, etiquetados: 1 ♂ y 1 ♀ «1-VI-1983, Robledo, Salamanca, O. Calvo & J.L. López Colón leg.» (AV); 1 ♂ «24V-1984, Carche, Jumilla, Murcia, J.L. Lencina leg.» (AV); 1 ♂ «25-VII-1988, Jumilla, Murcia, J. L. Lencina leg.» (AV); 1 ♂ y 6 ♀ «26-VII-1989, Siles, Jaén, J. I. López Colón leg.» (AV); 1 ♂ y 1 ♀ «16-IV-1998, Puerto de Tordigas, Cuenca, J.L. Lencina leg.» (AV); 1 ♂ «11-VI-2013, Val de Boquero el Ciego, La Retuerta, Zaragoza, R. Macià leg.» (AV). Material revisado
23 ♂♀, etiquetados: 12 ♂♀ «3-VII-1973, Sierra de Gata, Cáceres, J. López Colón leg.» (MZB 83-7726, 83-7727, 83-7728, 83-7729, 83-7730, 83-7731); 11 ♂♀ «13-V-1989, Revolcadores, Cañada de la Cruz, Murcia, X. Vázquez leg.» (MZB 83-7734, 83-7735, 83-7736, 83-7737, 83-7738).
Distribución
Especie con una distribución mediterránea discontinua, citada de: Argelia, España, Grecia y Marruecos. Con pocas citaciones ibéricas y al parecer presente en toda el área peninsular española (Español, 1995; Johnson, 1975; Viñolas, 2020).
España: Alicante (Callosa de Ensarriá); Cáceres (Sierra de Gata); Cuenca (Puerto de Tordigas); Granada (La Sagra, sin indicación); Málaga (Ronda); Múrcia (Carche, Jumilla, Revolcadores); Pontevedra (Los Masos); Salamanca (Robledo); Tarragona (Cornudella de Montsant, Vimbodí i Poblet); Zaragoza (La Retuerta).
A. VIÑOLAS & J. MUÑOZ-BATET
Biología
Se desconoce, aunque es de suponer que, como todas las especies del género, dependerá del arbolado de resinosas.
Ernobius parens (Mulsant & Rey, 1863) (Fig. 43)
Liozoum parens Mulsant & Rey, 1863. Opuscules Entomologiques, 13: 118
Liozoum crassicorne Mulsant & Rey, 1864. Histoire naturelle des Coléoptères de France. Térédiles: 175
Ernobius parens (Mulsant & Rey): Kiesenwetter, 1877. Naturgeschichte der Insecten Deutschlands.: 125
Ernobius obscuriventris Pic, 1902. L’Échange, Revue Linnéenne, 18: 79
Ernobius bipartitus Pic, 1935. L’Échange, Revue Linnéenne, 51 (460): 5
Longitud de 1,6 a 3,1 mm. Cuerpo de color rojizo claro, ligeramente amarillento, superficie con la pubescencia larga y densa. Antenas (Fig. 15) con la suma de los artejos del funículo más larga que la suma de los artejos de la maza, poco desarrollados. Protórax convexo, 1,71 veces más ancho que largo, con la mayor anchura en la base, mitad basal de los márgenes y ángulos posteriores explanados, superficie con el punteado denso y poco marcado. Élitros 1,64 veces más largos que anchos tomados conjuntamente, Edeago (Fig.29a) con el parámero izquierdo fuertemente explanado lateralmente, lóbulo medio (Fig, 29b) estrecho y con un pequeño saliente lateral en la zona apical.
Material estudiado
20 ♂ y 14 ♀, etiquetados: 1 ♀ «4-IV-2015, Bosc Duna Continental, Torruella de Montgri, Girona, A. Viñolas leg.» (AV); 18 ♂ y 7 ♀ «16-VI-2018, Turó del Vilar, BP 1.1-BP 2.1-BP 2.2-BP 3.1-BP 3.2-BP 4.1-CV 2.1-CV 2.2-CV 2.3-CV 3.1CV 3.2-NI 3.3, Blanes, Girona, P. Pons & E. Lucha leg.» (AV, IR, JM); 2 ♂ y 5 ♀ «6-IX-2018, Es Caló de Sant Agusti, Formentera, Baleares, J. Muñoz leg.» (JM, AV); 1 ♀ «8-VIII2019, Puigmoltó, Sant Pere de Ribes, Barcelona, M. RocaCusachs leg.» (AV).
Material revisado
22 ♂♀, etiquetados: 2 ♂ y 1 ♀ «00-00-0000, Pollença, Mallorca, Illes Balears, G. Jordà leg.» (MZB 83-7757, 83-7759, 83-7762); 1 ej. «00-00-0000, Felanitx, Mallorca, Illes Balears, Palace leg.» (MZB 83-7760); 1 ej. «20-VIII-1940, Sidamunt, Mallorca, Illes Balears, A. M. leg.» (MZB 83-7758); 2 ej. «X-1950, Sant Joan de Labritja, Evissa, Illes Balears, F. Español leg.» (MZB 83-7756, 83-7776); 1 ej. «1-V-1956, Son Espanyolet, Mallorca, Illes Balears, R. López leg.» (MZB 83-7777); 1 ej. «V-1956, Sierra Bermeja, Málaga, M. González leg.» (MZB 83-7764); 9 ej. «VIII-1982, Menorca, Illes Balears, Cuello leg.» (MZB 83-7768, 83-7769, 83-7770, 83-7766); 1 ♂ «23-VII-1990, Alfara de Carles, Tarragona, X. Vázquez leg.» (MZB 83-7767); 2 ♂ «26-VII-1997, Fraga, Huesca, T. Yélamos leg.»; 1 ej. «25-VIII-1998, Sant Martí Sesgueioles, Barcelona, J.J. Pérez de Gregorio leg.» (MZB 83-7775).
Butlletí ICHN 88 (8), 2024 GEA, FLORA ET FAUNA
Especie con una distribución europea discontinua, conocida también del norte de África, citada de: Alemania, Argelia, España, Francia, Hungría, Marruecos, Túnez y Ucrania. Distribución que deberá ser revisada. En la Península Ibérica se conoce de la mitad oriental y también de las Islas Baleares (Español, 1977; GONHS, 2017; Johnson, 1975; Viñolas, 2002, 2020; Viñolas & Muñoz-Batet, 2016). Aunque el Catálogo Paleártico la cita de Portugal (Zahradník, 2014) no se han localizado datos sobre su presencia en el país.
España: Barcelona (Sant Marti Sesgucioles, Sant Pere de Ribes); Girona (Blanes, Torrroella de Montgrí); Huesca (Fraga); Madrid (Sierra de Guadarrama); Málaga (Sierra Bermeja); Segovia (El Espinar); Tarragona (Alfara de Carles, Cornudella de Montsant): Teruel (Moscardón).
Gibraltar: (sin indicación).
Islas Baleares: Mallorca (Felanitx, Pollença, Portals Nous, Son Espanyolet); Menorca (sin indicación); Eivissa (Sant Antoni de Portmany, Sant Joan de Labritja, Sant Josep de sa Talaia); Formentera (Es Caló de Sant Agustí).
Biología
En Francia citan la especie en Pinus halepensis Mill. y P. pinaster Aiton. En Eivissa y Formentera se capturaron ejemplares con trampa de luz UV situada en bosque de P. halepensis y en la primera también se recolecto sobre troncos recién apeados de P. halepensis. En Blanes con trampas de interceptación de vuelo situadas en un bosque quemado de P. pinea L., Quercus ilex L. y Q. suber L. En Torroella de Montgrí con trampa de interceptación de vuelo situada en un bosque de P. pinaster Aiton con presencia de ejemplares aislados de P. halepensis y P. pinea.
Ernobius pini pini (Ratzeburg, 1837) (Fig.44)
Anobium pini Ratzeburg, 1837. Erster Theil Die Käfer: 43
Anobium pini Sturm, 1837. Die Insekten. Deutschland Insekten, Käfer. Vol. 11: 121
Liozoum pini (Sturm): Mulsant & Rey, 1863. Opuscules Entomologiques, 13: 124
Liozoum crassiusculum Mulsant & Rey, 1864. Histoire naturelle des Coléoptères de France. Térédiles: 175
Ernobius crassiusculus (Mulsant & Rey): Kiesenwetter, 1877. Naturgeschichte der Insecten Deutschlands.: 125
Ernobius mollis (Linnaeus): Kiesenwetter, 1877. Naturgeschichte der Insecten Deutschlands.: 128 (non Linnaeus, 1758)
Ernobius angusticollis tauri Pic, 1907. L’Échange, Revue Linnéenne, 23 (276): 185
Ernobius oblitus Sharp, 1916. The Entomologist’s Monthly Magazine, 52: 179
Ernobius pini galiberti Pic, 1918. L’Échange, Revue Linnéenne, 34: 9
Ernobius pini (Sturm): Fauconnet, 1892. Fauna analytique des Coléoptères de France: 303
Longitud de 1,7 a 3,1 mm. Cuerpo de color rojizo, más o menos claro, superficie con la pubescencia corta y densa. Antenas (Fig.16) con la suma de los artejos del funículo aproxi-

A. VIÑOLAS & J. MUÑOZ-BATET
madamente igual a la suma de los artejos de la maza antenal, no muy desarrollados. Protórax muy convexo, transverso y 1,5 veces más ancho que largo, superficie con el punteado fuerte y muy denso. Élitros 1,86 veces más largos que anchos tomados conjuntamente, Edeago (Fig. 30a) con el parámero izquierdo con un desarrollo interno lateral muy característico, lóbulo medio (Fig. 30b) con el ápice escotado.
Material estudiado
23 ♂♀, etiquetados: 1 ♂ y 1 ♀ «5-V-2011, Avinyonet del Penedès, Barcelona, M. Goula leg.» (AV); 1 ♂ y 1 ♀ «V-2011, Rada, Navarra, I. Recalde leg.» (AV); 1 ♂ «7-VI-2012, coll d’Heures, Moianès, A. Viñolas leg.» (AV); 1 ej. «25-VI/4VII-2012, El Tillar, Vimbodí i Poblet, serra de Prades, Tarragona, A. Viñolas leg» (AV); 1 ej. «4/13-VII-2012, El Tillar, Vimbodí i Poblet, serra de Prades, Tarragona, A. Viñolas leg.» (AV); 1 ♂ «1/30-XI-2012, turó de can Balasc, Barcelona, J. Mederos leg.» (AV); 1 ♀ «5-V-2013, Cortes de Pallás, Valencia» (AV); 1 ♂ «V-2013, Traibuenas, Rada, Navarra, I. Recalde leg.» (IR); 3 ♂ y 1 ♀ «6-VI-2014, Sant Bartomeu del Grau, Barcelona, R. Macià leg.» (AV); 2 ♂ y 5 ♀ «26-V-2017, Tivissa, Tarragona, A. Cervelló leg.» (AV); 1 ♀ «24-VI-2017, Mura, Barcelona, S. Trócolí leg.» (AV); 1 ♀ «5-VIII-2017, Isaba, Navarra, I. Recalde leg.» (AV); 1 ♂ «16-VI-2019, Turó del Vilar, NI 3.3, Blanes, Girona, P. Pons leg.» (AV).
Material revisado
6 ♂♀, etiquetados: 1 ♂ «3-VI-1908, Montgat, Barcelona, Codina leg.» (MZB 83-7802); 2 ej. «15-III-1909, Montsant, Tarragona» (MZB 83-7796); 1 ej. «3-V-1915, Castelldefels, Barcelona» (MZB 83-7798); 1 ej. «21-IV-1941, Dehesa, Valencia, Torres & Perris leg.» (MZB 83-7800); 1 ♂ «16-VI1991, Sierra de Baza, Granada, X. Vázquez leg.» (MZB 766252).
Distribución
Subespecie con una amplia distribución, citada de: Alemania, Argelia, Austria, Bélgica, Chipre, Croacia, Dinamarca, Eslovaquia, Estonia, España, Finlandia, Francia, Georgia, Gran Bretaña, Grecia, Holanda, Hungría, Italia, Letonia, Polonia, República Checa, Rumanía, Rusia, Suecia, Suiza, Túnez y Turquía. El ejemplar de Lugo (Diéguez Fernández, 2013) se deberá de revisar, ya que dicha localización está totalmente fuera del área de distribución conocida de la especie, que está localizada en la región ibérica oriental (Echave & Trócoli, 2018; Español, 1977; Español & Viñolas, 1992; Johnson, 1975; Pérez Moreno & Moreno Grijalba, 2009; Viñolas, 2020, 2022; Viñolas et al., 2013, 2015).
España: Barcelona (Avinyonet del Penedès, Castelldefels, Moià, Montgat, Mura, Parque Natural de Collserola, Sant Llorenç del Munt, Tiana); Granada (Sierra de Baza); La Rioja (Villoslada de Cameros); Lleida (Aigüestortes); Lugo (Monforte de Lemos); Navarra (Isaba, Rada); Tarragona (Tivissa, Vimbodí i Poblet); Valencia (Dehesa de la Albufera).
Biología
Las larvas se desarrollan en los brotes de diferentes especies del género Pinus, después de que han sido atacados por
Butlletí ICHN
(8), 2024 GEA, FLORA ET FAUNA
especies de Scolytinae. Otros autores indican su presencia en tocones y árboles debilitados por el ataque de Scolytinae. En la Península se ha citado de Pinus nigra J. F. Arnold y P. sylvestris L. Los ejemplares de Mura se han recolectado mediante trampas de luz en una área de Pinus halepensis Miller.
Ernobius pruinosus (Mulsant & Rey, 1863) (Fig. 45)
Liozoum pruinosus Mulsant & Rey, 1863. Opuscules Entomologiques, 13: 101
Ernobius pruinosus (Mulsant & Rey): Kiesenwetter, 1877. Naturgeschichte der Insecten Deutschlands.: 122
Ernobius normandi Pic, 1899. Bulletin de la Société Zoologique de France, 24: 24
Ernobius cedri Reitter, 1901. Verhandlungen des naturforschenden Vereines in Brünn, 40: 18
Ernobius thery Pic, 1902. L’Échange, Revue Linnéenne, 18: 79
Ernobius incisus Pic, 1904. L’Échange, Revue Linnéenne, 20: 2
Ernobius beauprei Pic, 1908. L’Échange, Revue Linnéenne, 24: 57
Ernobius minutus Pic, 1921. L’Échange, Revue Linnéenne, 37: 2
Ernobius tuniseus Pic, 1921. L’Échange, Revue Linnéenne, 37: 2
Longitud 2,3 a 3,5 mm. Cuerpo de color rojizo obscuro, antenas y cabeza negruzcos, con la pubescencia de la superficie muy densa. Antenas (Fig.17) con la suma de los artejos del funículo más corta que la de los artejo de la maza antenal, que son proporcionalmente anchos. Protórax convexo y transverso, 1,52 veces más ancho que largo, con la máxima anchura en la base, márgenes explanadas, superficie con el punteado bien indicado, nada contiguo. Élitros 1,74 veces más largos que anchos tomados conjuntamente. Edeago (Fig. 31a) parámero izquierdo con el ápice dilatado lateralmente, lóbulo medio (Fig. 31b) ancho y con el ápice particularmente conformado.
Material revisado
3 ♂♀, etiquetados: 1 ej. «Denia, Alicante, Torres-Sala leg.» (MZB 83-7784); 1 ej. «Mallorca, G. Jordá leg.» (MZB 837792); 1 ♂ «13-XI-1982, Serra Finestres, Llorà, Girona, J.J. Pérez de Gregorio leg.» (MZB 83-7792).
Distribución
Con una distribución europea discontinua y a revisar, citada de: Alemania, Argelia, Eslovaquia, España, Francia, Grecia, Italia, Líbano, Marruecos y Túnez. En la Península Ibérica sólo hay unas pocas citas del área mediterránea, conocida también de las Islas Baleares (Español, 1977a; Fuente, 1932; Johnson, 1975; Viñolas, 2020).
España: Alicante (Denia); Almería (Tíjola); Girona (Llorà).
Islas Baleares: Eivissa (Sant Antoni de Portmany); Mallorca (Palma, sin indicación).
Biología
Asociada, como todas las especies del género a resinosas. Sólo se ha citado de Pinus pinaster Ait.


Butlletí ICHN 88 (8), 2024

Ernobius reflexus (Mulsant & Rey, 1863) (Fig. 46)
Liozoum reflexus Mulsant & Rey, 1863. Opuscules Entomologiques, 13: 96
Ernobius reflexus (Mulsant & Rey): Kiesenwetter, 1877. Naturgeschichte der Insecten Deutschlands.: 121
Ernobius algiricus Pic, 1915. L’Échange, Revue Linnéenne, 3: 17
Longitud de 5,0 a 8,1 mm. Cuerpo rojizo amarillento, con la pubescencia de la superficie muy densa. Antenas (Fig, 18) con la suma de los artejos del funículo un poco más larga que la suma de los artejos de la maza antenal, que son muy alargados. Protórax fuertemente transverso, 1,91 veces más ancho que largo, con los márgenes redondeados y explanados, superficie con el punteado denso y poco indicado. Élitros 2,5 veces más largos que anchos tomados conjuntamente. Edeago (Fig. 32a) con el parámero izquierdo apicalmente dilatado y el lóbulo medio (Fig. 32b) con la parte apical ancha y escotada en el medio.
Material revisado
.5 ♂♀, etiquetados: 1 ej. «Mallorca, G. Jordá leg.» (MZB 837807); 1 ej. «12-X-1932, Balenyà, Barcelona, A. Vilarrubia leg.» (MZB 83-7805); 1 ej. «19-VII-1954, Prat de Llobregat, Barcelona, Monés leg.» (MZB 83-7808); 1 ej. «3-I-1965,
Sant Pol de Mar, Barcelona, M. Gras leg.» (MZB 83-7813); 1 ej. «X-1966, San Roque, Cádiz, J. Ramirez leg.» (MZB 837812).
Distribución
Especie propia del Mediterráneo occidental conocida de: Argelia, España, Francia, Marruecos y Túnez (Español, 1977, 1995; Johnson, 1975; Viñolas, 2020).
España: Barcelona (Balenyà, Barcelona, el Prat de Llobregat, Sant Cugat, Sant Pol de Mar); Cádiz (San Roque); Gipuzkoa (Sierra Aralar); Huesca (Valle de Añisclo); Madrid (El Pardo).
Islas Baleares: Mallorca (Esporles, Pollença, sin indicación).
Biología
Especie, al parecer, asociada con Pinus halepensis Miller, P. pinea L. y P. pinaster Aiton. Los ejemplares de Sant Cugat se recolectaron en pies de P. halepensis.
Ernobius rufus (Illiger, 1807) (Fig. 47)
Anobium rufum Illiger, 1807. Magazin für Insectenkunde, 6: 20
Anobium nitidulum Wollaston, 1871. Transactions of the Entomological Society of London, (2): 262
GEA, FLORA ET FAUNA
Ernobius rufus (Illiger): Pic, 1912. Anobiidae. Coleopterorum catalogus, pars 48: 23
Longitud de 2,5 a 3,2 mm. Cuerpo rojizo negruzco, con la parte apical de los élitros más clara, superficie con la pubescencia no muy densa. Antenas (Fig.19) con la suma de los artejos del funículo aproximadamente igual a la suma de los artejos de la maza antenal. Protórax transverso, 1,53 veces más ancho que largo, con la máxima anchura en la base y los márgenes ligeramente explanados, superficie con el punteado poco indicado. Élitros 1,62 veces más largos que anchos tomados conjuntamente. Edeago (Fig. 33a) con el parámero izquierdo fuertemente dilatado lateralmente, lóbulo medio (Fig, 33b) aguzado.
Material estudiado
12 ♂ y 4 ♀, etiquetados: 1 ♂ «19-V-2012, Rambla de la Sierra, 30SWF5799, Tabernas, Almería, R. Macià leg.» (AV); 1 ♂ «8-VI-2013, Cortes de Pallás, Valencia» (AV); 1 ♂ y 1 ♀ «12-IX-2017, Es Caló de Sant Agustí, Formentera, illes Balears, M. Rondós & R. Macià leg.» (AV); 1 ♂ «16-V-2018, La Bernardilla, Los Guájares, Granada, J. Muñoz leg.» (AV); 7 ♂ y 2 ♀ «6-IX-2018, Es Caló de Sant Agusti, Formentera, Baleares, J. Muñoz leg.» (AV, JM); 2 ♀ «14-VI-2022, El Lloar, Taragona, A. Viñolas leg.» (AV).
Material revisado
1 ♂, etiquetado: «10-III-1979, Estepona, Málaga, J. de Ferrer leg.» (MZB 83-7782).
Distribución
Especie descrita de Portugal y conocida de: España, Italia, Marruecos, Portugal e Islas Baleares y Madeira (Español, 1977, 1992; Johnson, 1975; Viñolas, 2020, 2022; Viñolas & Muñoz Batet, 2017). Con introducciones en la región Afrotropical.
España: Almería (Tabernas); Badajoz (Olivenza); Granada (Los Guájares); Madrid (Sierra de Guadarrama); Málaga (Estepona); Segovia (El Espinar); Tarragona (El Lloar); Valencia (Cortes de Pallás).
Portugal: Leiria (São Pedro de Moel).
Islas Baleares: Formentera (Es Caló de Sant Agustí).
Biología
Poco se conoce de su biología, como todas las especies del género asociada a Pinus sp.
Agradecimientos
A Mercedes París, del Museo Nacional de Ciencias Naturales de Madrid, y Glòria Masó y Berta Caballero del Museu Ciències Naturals de Barcelona el habernos permitido consultar las colecciones de las entidades. Y a todos los entomólogos la cesión del material colectado en sus campañas entomológicas.
Bibliografía
Butlletí ICHN 88 (8), 2024
Audisio, P., Gobbi, G., Liberti, G. & Nardi, G. 1995. Coleoptera Polyphaga IX (Bostrichoidea, Cleroidea, Lymexyloidea). In: Minelli, A., Ruffo, S. & La Posta, S. (eds.), Cheklist delle specie della fauna italiana, 54. Calderini, Bologna. 27 p. Barnouin, T. 2014. Subfamilias Dryophylinae, Ernobiinae. P. 458-461. In: Catalogue des Coléoptères de France. Tronquet, M. (coord.). Supplément au tome 23 - R.A.R.E. Association Roussillonnaise d’Entomologie. Perpignan. 1052 p. Bercedo, P., Arnáiz, L. & De Sousa Zuzarte, A. J. 2008. Coleópteros de Portugal: Ernobius gallicus Johnson, 1975 especie nueva para la fauna portuguesa; y presencia de Nicobium zuzartei Bercedo & Arnaiz, 2007 en la costa atlántica portuguesa (Coleoptera: Ptinidae: Ernobiinae, Anobiinae). Boletín Sociedad Entomológica Aragonesa, 43: 377-378.
Bouchard, P., Bousquet, Y., Davies, A. E., Alonso-Zarazaga, M. A, Lawrence, J. F., Lyal, C. H. C., Newton, A. F., Reid, C. A. M., Schmitt, M., Ślipiński, S. A. & Smith, A. B. T. 2011. Familygroup names in Coleoptera (Insecta). ZooKeys, 88: 1-972.
Corrêa de Barros, J. M. 1907. Quelques Coléoptères nouveaux pour la faune du Portugal. Bulletin de la Société Portugaise de Sciences Naturelles, 1 (3): 130-143.
Diéguez Fernández, J, M. 2013. Registros interesantes de coleópteros para España (Insecta: Coleoptera). 2ª nota. Arquivos Entomolóxicos, 8: 277-286
Echave, P. & Trócoli, S. 2018. Nuevas aportaciones al catálogo de los coleópteros del Parc Natural de Sant Llorenç del Munt i l‘Obac (Barcelona, España). Heteropterus, Revista de Entomología, 18 (1): 141-153.
Español, F. 1964. Notas sobre Anóbidos. 11: Los anóbidos de las Islas Canarias. 12: Afropetalium nuevo género de Dorcatominae del África tropical. 13: Xestobium austriacum Reitt. en los Cárpatos rusos. 14: Adiciones al catálogo de anóbidos de Marruecos. Publicaciones del Instituto de Biología Aplicada, Barcelona, 37: 95-121.
Español, F. 1977. Los Ernobius Thoms. de la fauna española (Col. Anobiidae, nota 77). Publicaciones del Departamento de Zoología de la Universidad de Barcelona, 2: 19-28.
Español, F. 1990. Nuevos datos sobre Anóbidos Ibéricos (Col. Anobiidae). Misceŀlània Zoològica, 12 (1988): 171-175.
Español, F. 1992. Coleoptera, Anobiidae. En: Fauna Ibérica, vol. 2, Ramos, M. A. et al. (Eds.). Museo Nacional de Ciencias Naturales. CSIC. Madrid. 195 p.
Español, F, 1995. Insecta: Coleoptera, 10. Familia: Anobiidae. Catalogus de la entomofauna aragonesa, 7: 3-5.
Español, F. & Viñolas, A. 1992. Coleòpters del Parc Nacional d’Aigüestortes i Estany de Sant Maurici. Generalitat de Catalunya. Departament d’Agricultura Ramaderia i Pesca. Lleida. 48 p. Fuente, J. M. de la 1932. Catálogo sistemático - geográfico de los Coleópteros observados en la península ibérica, Pirineos propiamente dichos y Baleares. Boletín de la Sociedad Entomológica de España, 15 (2-4): 38-53, 75-76.
GONHS (Gibraltar Ornithological of Natural History Society), 2017 Provisional List of the Coleoptera of Gibraltar. http://www.gonhs. org/ColeopteraofGibraltar.htm. (Consultado: 15-XII-2022) Gottwald, J. 1977. Die paläarktischen Xyletinus-Arten (Col. Anobiidae). Acta Entomologica Bohemoslovaca, 74: 158-177. Johnson, C. 1975. A review of the Palaearctic species of the genus Ernobius Thomson. Entomologische Blätter, 71 (2): 65-93.
Laclos, E. de, Brustel, H., Büche, B., Coache, A. Jiroux, E. & Ponel, P. 2007. Anobiidae nouveaux ou méconnus de la faune de France (Coleoptera). L’Entomologiste, 63 (3): 137-140.
López-Colón, J. I. 2001. Algunos datos corológicos sobre anóbidos ibéricos (Coleoptera, Anobiidae). Boletín Sociedad Entomológica Aragonesa, 28: 115-116.
López-Colón, J. I. & Agoiz-Bustamante, J. L. 2003. Más datos sobre anóbidos ibéricos (Coleoptera, Anobiidae). Boletín Sociedad Entomológica Aragonesa, 32: 246-247.
Novoa, F. & Baselga, A. 2000. A new species of Ernobius Thomson (Coleoptera: Anobiidae: Ernobiinae) from the Cíes Islands, Spain. The Coleopterits Bulletin, 54 (3): 403-407.
Pérez Moreno, I. & Moreno Grijalba, F. 2009. Los coleópteros saproxílicos del Parque Natural Sierra de Cebollera (La Rioja). 28 Ciencias de la Tierra. Instituto de Estudios Riojanos. Logroño. 180 p. Pic. M. 1912. Anobiidae. Coleopterorum catalogus, pars 48. Junk. Berlin. 92 p.
Recalde Irurzun, J. I. & San Martín Moreno, A. F. 2017. Presencia de Hallomenus (Hallomenus) axillaris (Illiger, 1807) en la Península Ibérica, confirmación de Ochina (Dulgieris) latreillii (Bonelli, 1812) y Platysoma (Cylister) lineare Erichson, 1834, y otros coleópteros destacables de un bosque sur-pirenaico de Pinus sylvestris (Insecta: Coleoptera). Revista gaditana de Entomología, 8 (1): 53-66.
Sakai, M. 2002. The Genus Ernobius Thomson of Japan (Coleoptera, Anobiidae, Ernobiinae). Japanese Journal of Systematic Entomology, 8 (1): 119-130.
Viñolas, A. 2013a. Els Dorcatominae de la península Ibèrica i illes Balears. 1a nota. Els gèneres Caenocara C.G. Thomson, 1859, Calymmaderus Solier, 1849 i Mizodorcatoma Hayashi, 1955 (Coleoptera: Ptinidae). Butlletí de la Institució Catalana d’Història Natural, 76 (2012): 117-132.
Viñolas, A. 2013b. Els Dorcatominae de la península Ibèrica i illes Balears. 2a nota. El gènere Dorcatoma Herbst, 1792. (Coleoptera: Ptinidae). Orsis, 27: 7-28.
Viñolas, A. 2013c. Els Dorcatominae de la península Ibèrica i illes Balears. 3a nota. El gènere Stagetus Wollaston, 1861, amb la descripció de S. confusus n. sp. (Coleoptera: Ptinidae). Orsis, 27: 95-121.
Viñolas, A. 2013d. Els Mesocoelopodinae de la península Ibèrica i illes Balears. Els gèneres Mesocoelopus Jacquelin du Val, 1860, Mesothes Mulsant & Rey, 1864 i Rhamna Peyerimhoff, 1913 (Coleoptera: Ptinidae). Butlletí de la Institució Catalana d’Història Natural, 76 (2012): 105-116.
Viñolas, A. 2002. Nova aportació al coneixement dels anòbids de la península Ibèrica (Coleoptera: Anobiidae). Butlletí de la Institució Catalana d‘Història Natural, 70: 73-77.
Viñolas, A. 2017. Nueva aportación al conocimiento de los Ptinidae (Coleoptera) de la Península Ibérica e Islas Canarias, con la descripción de un nuevo Stagetus Wollaston, 1861 de Navarra. Arquivos Entomoloxicos, 18: 137-148.
Viñolas, A. 2018. Nuevos datos sobre la validez específica de Ernobius vinolasi Novoa & Baselga, 2000 (Coleoptera: Ptinidae: Ernobiinae). Arquivos Entomolóxicos, 19: 75-80.
Viñolas, A. 2020. Catálogo comentado de los Ptinidae (Coleoptera) de la Península Ibérica, Islas Baleares e Islas Canarias. Monografies de la Institució Catalana d’Història Natural, 1. 179 p.
Viñolas, A. 2022. Addenda al catálogo comentado de los Ptinidae (Coleoptera) de la Península Ibérica, Islas Baleares e Islas Canarias. Butlletí de la Institució Catalana d’Història Natural, 86 (3): 99-103.
Viñolas, A. & Bookwalter, J. 2018. Six interesting species of Ptinidae (Coleoptera) from Andorra and Tarragona, Catalonia (Iberian Peninsula). Heteropterus, 18 (1): 97-106.
Viñolas, A. & Ghahari, H. 2017. A checklist of Ptinidae (Coleoptera: Bostrichoidea) from Iran. Redia, 100: 139-147.
Viñolas, A. & Masó, G. 2013. The collection of type specimens of the family Ptinidae (Coleoptera) deposited in the Natural History Museum of Barcelona, Spain. Arxius de Misceŀlània Zoològica, 11: 1-79.
Viñolas, A. & Muñoz-Batet, J. 2016. Noves dades sobre els Ptinidae Latreille, 1802 (Coleoptera) del Parc Natural dels Ports, Tarragona, Catalunya. Butlletí de la Societat Catalana d‘Història Natural, 80: 113-115.
Viñolas, A. & Muñoz Batet, J. 2017. Noves aportacions al conèixement de la fauna coleopterològica de la península Ibèrica. Nota 2a (Coleoptera). Butlletí de la Institució Catalana d’Història Natural, 81: 177-184.
Viñolas, A. & Muñoz-Batet, J. 2019. Els Ptilininae de la península Ibèrica i illes Balears (Coleoptera: Ptinidae). Butlletí de la Institució Catalana d’Història Natural, 83: 95-102.
Viñolas, A. & Muñoz-Batet, J. 2022. Nous o interessants Ptinidae (Coleoptera) per a Catalunya. Butlletí de la Institució Catalana d’Història Natural, 86 (1): 27-30.
Viñolas, A., Muñoz-Batet, J. & Soler, J. 2015. Els coleòpters saproxílics de la casa forestal del Tillar, serra de Prades, Tarragona. V Jornades sobre el bosc de Poblet i les muntanyes de Prades: 361-370.
Viñolas, A., Muñoz, J., Mencuccini, M. & Benvenuti, F. 2013. Nuevos datos sobre Rushia parreyssi (Mulsant, 1856), Melandrydae Leach, 1815 y otros coleópteros interesantes de la sierra de Prades, Tarragona (Coleoptera). Orsis, 27: 29-51.
Viñolas, A. & Recalde.Iruzun, J. I. 2018. Los Ernobiinae de la Península Ibérica e Islas Baleares.1a nota. El género Episernus C. G. Thomson, 1863 (Coleoptera: Ptinidae). Butlletí de la Institució Catalana d’Història Natural, 82: 97-107.
White, R. E. 1982. A catalog of the Coleoptera of America north of Mexico: Family Anobiidae. United States Department of Agriculture. Agriculture Handbook Number 529–70. i–xi + 58 p.
Zahradník, P. 2014. Family Ptinidae. In: Zahradník, P. & Háva, J. Catalogue of the world genera and subgenera of the superfamilies Derodontoidea and Bostrichoidea (Coleoptera: Derodontiformia, Bostrichiformia). Zootaxa, 3754 (4): 301-352.
Zahradník, P. & Háva, J. 2014. Catalogue of the world genera and subgenera of the superfamilies Derodontoidea and Bostrichoidea (Coleoptera: Derodontiformia, Bostrichiformia). Zootaxa, 3754 (4): 301-352.
Pere Aymerich*
* C. Onze de Setembre, 31. E08600 Berga. Catalonia..A/e: pere_aymerich@yahoo.es
Rebut: 06.05.2024; Acceptat: 30.07.2024; Publicat: 30.09.2024
Abstract
Noccaea cadinensis sp. nov. is described here. This species is known from a single locality in Serra del Cadí (Eastern Pyrenees, SW Europe), where it grows in limestone scree and stony meadows of the alpine vegetation belt. It is morphologically close to N. nevadensis from Sierra Nevada (S Iberian Peninsula), from which it can be separated by several characters (fruit length, style length, corymb elongation, petiole length, leaves margin, seed size). The new species is assessed as EN (Endangered) according to IUCN Red List Categories and Criteria.
Key words: taxonomy, endemism, mountain, Iberian Peninsula.
Resum
Una nova espècie de Noccaea (Brassicaceae) endèmica d’hàbitats alpins dels Pirineus
Es descriu Noccaea cadinensis sp. nov. Aquesta espècie es coneix actualment només d’una localitat de la serra del Cadí (Pirineus orientals), on viu en tarteres i prats pedregosos sobre calcària de l’estatge alpí. Morfològicament és similar a N. nevadensis de Sierra Nevada (sud de la península Ibèrica), de la qual es diferencia per diversos caràcters (longitude del fruit, longitud de l’estil, elongació del corimb, longitud del pecíol, marge de les fulles, mida de les llavors,). Seguint els criteris i categories de la IUCN, aquesta nova espècie és catalogada com a EN (En perill).
Paraules clau: taxonomia, endemisme, muntanya, península Ibèrica.
Introduction
Noccaea Moench (Brassicaceae) is a genus formerly nested in Thlaspi L. until Meyer (1979) split it into 6 genera. The splitting of Thaspi is now widely accepted, but there is still controversy over the number of genera into which it should be divided, ranging between 3 (Al-Shehbaz, 2014) and 13 (https://brassibase.cos.uni-heidelberg.de/). More recent and comprehensive work supports the acceptance of a low number of genera, but it is recognized that new phylogenetic approaches must be made with new information (Özüdoğru et al., 2019). In its broadest sense, Noccaea has about 136 species (AlShehbaz, 2022), some of very recent description (Güzel et al., 2018; Özgişi et al., 2018; Özgişi, 2020; Sefali et al., 2023) while in a more restricted delimitation 92 are accepted (https://brassibase.cos.uni-heidelberg.de/). The vast majority are of Eurasian distribution and the greatest diversity is found in the mountains and steppes of C-W Asia (Al-Shebahz, 2014). Narrow range endemic species are common, especially in the mountains, and the morphological differences between taxa are often subtle. Examples of species restricted to high mountain or alpine habitats in Europe are Noccaea rotundifolia (L.) Moench (Alps), N. corymbosa (J. Gay). F.K. Mey. (Alps W-C), N. cepaeifolia (Wulfen) Rchb. (Alps E),
N. minima (Ard.) F.K. Mey. (Alps E), N. nevadensis (Boiss. & Reut.) F.K. Mey. (Sierra Nevada), N. stylosa (Ten.) Rchb. (Apennines) and N. bellidifolia (Griseb.) F.K. Mey. (Balkans).
To date any Noccaea species restricted to alpine habitats was known in the Pyrenees. In this range the known species are N. brachypetala (Jord.) F.K. Mey, N. caerulescens J. Presl & C. Presl and N. occitanica (Jord.) F.K. Mey., which are mostly found in montane and subalpine vegetation belts, while N. perfoliata (L.) Al-Shebaz [Microthlaspi perfoliatum (L.) F.K. Mey.] has a wide distribution from the basal belt to the subalpine. The nearest known true high mountain (mainly alpine) Noccaea species were 500-600 km away, in the SW Alps and Sierra Nevada.
In 2022, a Noccaea population was discovered in the alpine belt at the western end of the Serra del Cadí, a peripheral (Pre-Pyrenean) range of the SE Pyrenees. These plants showed features that clearly distinguished them from the Noccaea already known in the area, especially its unwinged fruits and longer petals. This Noccaea population could not be identified with the available literature of the flora of Pyrenees or nearby regions, although these plants were fairly similar to N. nevadensis. A subsequent analysis revealed a set of characters that justify its description as a new species.
P. AYMERICH
The new species is identified on the basis of macromorphological characters. In a first approach, samples collected in the only known population the years 2022 and 2023, in various phenological stages, were compared with the information provided by the regional floras (Bolòs & Vigo, 1990; Pujadas, 1993; Tison & Foucault, 2014; Tison et al., 2014; Pignatti, 2017) for the Noccaea taxa known in SW Europe. In this bibliography the descriptions of these taxa are often partial and in some cases not coincident between different works. This bibliographic search pointed towards a morphological affinity between the plants of the Pyrenees and Noccaea nevadensis of Sierra Nevada (SE Iberian Peninsula). Also the use of the dichotomous keys provided by the synopsis of the genus Noccaea by Al-Shehbaz (2014) led to N. nevadensis For this reason, later the effort was focused on analyzing the differences between N. nevadensis and the Pyrenean plants. For the subsequent comparison, we used bibliographic information on N. nevadensis -based on Pujadas (1993), Blanca et al. (2009) and Al-Shehbaz (2014)- and N. nevadensis vouchers from the herbaria MA, GRA and BC. A list of revised vouchers is shown in Appendix 1.
Considering that the main characters that separate N. cadinensis and N. nevadensis are quantitative and that there is some overlap between these characters, a statistical analysis of the fruit characters has been made. Student’s T-test was used to test whether the measurements of two characters (fruit length, style length) are statistically significant or not. The sample analyzed was 36 ripe fruits of N. cadinensis and 32 of N. nevadensis (vouchers MA304335, MA306154, MA45048, MA304551, MA306154).
Noccaea cadinensis Aymerich sp. nov.
Holotype
Spain, Catalonia, Serra del Cadí, between Torreta de Cadí and Cap de la Costa Verda peaks, 42o16’60’’ N, 1o34’22’’ E, 2335 m, limestone scree, 18 July 2022, P. Aymerich (BCN170324).
Diagnosis
Noccaea cadinensis is morphologically close to N. nevadensis, but differs from it by the fruit usually 4-8 mm long (versus 7-12 mm in N. nevadensis), style usually 1-1.5 mm long (versus usually 1.5-3 mm), seeds 1.8-2.5 mm long (versus 1.4-1.8 mm), corymb not or weakly elongate in fructification to 25 mm (versus usually elongated, to 55 mm) and basal leaves with petiole usually shorter or equal to leaf blade (versus petiole usually longer than blade in N. nevadensis). Also the rosette leaves are usually marginally toothed in Nocaea cadinensis but usually non-dentate in N. nevadensis and ratio fruit body length / style length is usually higher in N. cadinensis.
A comparative table of the characters of Noccaea cadinensis and N. nevadensis is shown (Table 1). The characters of N. nevadensis are a compilation of bibliographical data and own observations.
Etimology
The specific name cadinensis is derived from an ancient name of Serra del Cadí range, Cadino.
Description
Perennial herb, 5-10 cm tall, glabrous, stoloniferous and caespitose (Fig. 1a,b). Basal leaves (Fig. 2) arranged in rosettes (10)15-22(31) mm diameter; deep green, glossy; thick and somewhat fleshy; petiole 2-7(12) mm long, usually shorter than leaf blade (sometimes equal, exceptionally long-




er); blade 3-11 x 2-5 mm, obovate, elliptic or rhombic, with weakly marked veins, apex obtuse, base tapering, margins usually with some coarse and obtuse teeth or rarely entire (0-5 teeth per leaf side). Stem ascending, simple, usually one per rosette, c. 1 mm diameter, with 4-8 leaves. Cauline leaves sessile, oblong, auriculate at base; 5-10 mm long; margins entire or with inconspicous teeth (0-4 per leaf side); apex obtuse to subacute; auricles with obtuse to subacute apexs. Inflorescence, a corymb with 11-20 flowers (Fig 1b, 3); 10-17 (25) mm diameter, 12-17 (20) mm height, diameterheight ratio 1.3-2.4; only weakly elongated in fruit, to 25 mm height (Fig. 4a). Flowers pedicels ascending, 2-5 mm long in anthesis (3-6 mm in fructification); sepals 2.5-4 × 1.5-2 mm, oblong-obovate, deep green and often partially purple tinged, but paler green and translucent in a wide marginal strip; petals oblong-obovate, 5-8 × c. 2 mm, colour initially
white, turning mauve with age; stamens 3-4 mm long, anthers yellow, 0.8-1.2 mm; Fruits an obovate silicula (Fig. 4a), 4-9 × 2-3.5 mm, glabrous, margin not winged, base cuneate, emarginate at apex, fruiting style 0.5-2.1 mm, exserted from the apex sinus. Seeds 1-3 per loculus, 1.8-2.5 × 0.7-1.5 mm, oblong, elliptic or obovate, smooth, light brown.
Relationships
The new species is morphologically close to Noccaea nevadensis. Although the morphological differences between the two are quantitative and overlap, they are considered sufficient to separate both taxa at species level. Differences in inflorescence elongation and fruits measurements are notorious at fruiting stage (Fig. 4a, b). The fruit and the style in Sierra Nevada plants are significantly longer (P<<0,01; t-test) than those of the Serra del Cadí plants. Table 2 summarizes the differences in fruit and style length of the populations from

Table 1 Comparative table of Noccaea cadinensis and N. nevadensis characters. In bold type, most useful characters to discriminate these species.
Character
Noccaea cadinensis
Height 5-10 cm
Basal leaves shape
Obovate, rhomboid or elliptic
Basal leaves dimensions 3-11 × 2-5 mm
Basal leaves margin
Basal leaves petiole
Cauline leaves
Inflorescence
Pedicels
Usually with coarse teeth, rarely entire
Noccaea nevadensis
4-15 cm
Obovate to elliptic
5-32 × 2-12 mm
Usually entire, sometimes with coarse teeth
Usually shorter (or equal) than leaf blade, Usually longer than leaf blade, exceptionally exceptionally longer shorter
Oblong, 4-10 mm long, base auriculate
Oblong, 5-15 mm long, base auriculate
Corymb not or weakly elongate in fructification, Corymb usually elongate in fructification, to to 2.5 cm 5.5 cm
2-5 mm long in flowers, 3-6 mm in fruits 1-6 mm in flowers, 3-6 mm in fruits
Sepals 2.5-4 mm long, often purple tinged 2-3.7 mm long, often purple tinged
Petals
Stamens
Style
Fruit
5-8 mm long, white to mauve 5-7 mm long, white or pinkish
3-4 mm long; anthers yellow 0.8-1.2 mm
3-4 mm; anthers yellow 0.7-1 mm
0.5-2.1 mm long, in fruit exserted from apical notch 1.2-3.2 mm long, in fruit exserted from apical notch
Unwinged; obovate, 4-9 × 2-3.5 mm; tappering at Unwinged; obovate-oblong, 6-12 × 2.3-3,5 base; apex emarginate mm; tappering at base; apex emarginate
Ratio L fruit body / L style 3.0-8.0 1.9-6.0 (in ripe fruit)
Seeds number 1-3 (1)2-3(4)
Seeds shape and colour
Seeds size
Oblong-obovate, light brown
Oblong-obovate, brown
1.8-2.5 × 0.7-1.4 mm 1.4-1.8 × 1.0-1.2 mm
Table 2. Comparison of fruits and style length in N. cadinensis and N. nevadensis. Mean ± standard deviation, and minimum and maximum values (in brackets) are shown, all of them in mm.
Character
Noccaea cadinensis
Noccaea nevadensis
Fruit length 6.2 ± 1.6 (4.0-9.0) 8.8 ± 1.7 (6.0-12.0)
Style lenght 1.3 ± 0.4 (0.5-1.6) 2.3 ± 0.5 (1.3-3.2)


4. Fruting corymbs of N. cadinensis (a) and N. nevadensis (b), MA45047. The differences in the elongation and density of the corymbs and in the lengths of the fruits and styles are visibles.


Sierra Nevada and the Pyrenees. The two taxa are geographically separated and the floristic affinities between the Pyrenees and Sierra Nevada are limited, the former more closely related to the Alps and the latter to the mountains of North Africa. In addition N. nevadensis is reported as attached to siliceous rocks, while N. cadinensis thrives in limestone.
In fact, to elucidate the affinities between the taxa of Noccaea is often difficult, because between groups of evolutionarily related taxa the morphological differences may be weak or subtle (Al-Shehbaz, 2014). The molecular techniques has improved the knowledge of the relationships between Noccaea taxa, but there are also groups of taxa that appear genetically close, with a weak divergence (Özüdoğru et al., 2109).
For example, most of the European mountain taxa related to N. rotundifolia (N. corymbosa, N. cepaeifolia, N. stilosa), which are now recognized as species, had previously been treated with varietal or subspecific ranks under N. rotundifolia. Morphologically they are very similar, with a few dis-
criminating characters and these often overlap. The current criterion is to treat them as species, on the basis of this weak differentiation and their allopatric distribution. Noccaea cadinensis also shows similarities with plants in this group, as a short style usually up to 1.5 mm (with the exception of N. stilosa), but these species have distinctly pink flowers and tipically more seeds per locule.
Without genetic information it is uncertain to assess the true relationships of N. cadinensis with N. nevadensis or other Noccaea. But the morphological similarity suggests that it may be evolutionarily more related to N. nevadensis than to the alpine taxa. Even a more widely distributed Iberian ancestor is possible, since the general shape of N. cadinensis is also reminiscent of Noccaea stenoptera (Boiss. & Reut.) F.K. Mey., an endemic species of the NW quadrant of the Iberian Peninsula with a wider ecology, not clearly linked to the high mountains. N. stenoptera was first considered silicicolous (Pujadas, 1993) but later is proved that it is indif-
P. AYMERICH

ferent to the rock substrate, since in the Cantabrian and Burgos mountains it is found on calcareous ridges reminiscent of N. cadinensis habitat, but at a lower altitude (Alejandre et al., 2006; Carlón et al., 2014). These three species have in common the pinkish-white (or withe turning pink) colour. Noccaea nevadensis and N. stenoptera have a lax and elongate fruiting corymb, unlike N. cadinensis Noccaea stenoptera clearly differs from N. cadinensis and N. nevadensis in having winged fruit valves, a character that is usually given taxonomic importance, but interestingly N. stenoptera and N. nevadensis are very close genetically despite this difference (Özüdoğru et al., 2109).
Phenology
Flowering time second half May to first half June. Fruits ripening and seeds dispersal July.
Habitat
Shallow soils on limestone rocks, in stabilized screes and stony meadows of the alpine zone, at an altitude of 2330-2350 m (Fig. 5a). Most Noccaea cadinensis individuals grow within or on the margins of cushions of Festuca gautieri (Hack.) K. Richt., which is the most common species (Fig. 5b). Other taxa observed in these places are Arenaria grandiflora L. subsp. grandiflora, Anthyllis vulneraria subsp. vulnerarioides (All.) Arcang., Botrychium lunaria (L.) Swartz, Carduus carlinoides Gouan subsp. carlinoides, Crepis pygmaea L., Helictotrihon sedenense (DC.) Holub subsp. sedenense, Iberis saxatilis L., Iberis spathulata DC. subsp. spathulata, Jasione crispa (Pourr.) Samp., Petrocallis pyrenaica (L.) R. Br. and Thymus nervosus Willk Taxa nomenclature is based in the Catalonia flora Checklist (Sáez & Aymerich, 2021).
Butlletí ICHN 88 (3), 2024
Distribution
Currently, this species is known only from the type locality. It is located at the foot of a west-facing slope between the peaks of Torreta de Cadí and Cap de la Costa Verda, in the western Cadí mountain range, SE Pyrenees (Figure 6). This mountain runs from west to east along about 30 km and has maximum altitudes around 2600 m a.sl.
Conservation status assessment
To date, N. cadinensis is known only from an area of less than 0.5 ha. A detailed census of the population is not available, but it has been estimated at tens or a few hundred individuals. A survey of nearby areas with suitable habitats has not allowed to locate other populations. Although the Serra del Cadí flora is fairly well-known (Vigo et al., 2003) it is possible that this species exists in other places and has been unnoticed, due to its early flowering and the weaker botanical knowledge of the western part of the mountain.
No current risk factors are known, because this area is rarely visited by people, is not used as pasture for domestic livestock and wild ungulates have low densities. Only climate warming is a potential threat, although it appears to be a relatively xeromorphic plant, which would probably not be among the most affected in a scenario of reduced rainfall and snow cover.
With the available data and the application of the IUCN (2012) methodology, the risk category of N. cadinensis is assessed as Endangered EN D. Only criterion D (Population size estimated to number fewer than 250 mature individuals) is applicable, as there are no signs of population or area decline, and extreme fluctuations seem unlikely.
We thank the curators of BC, BCN, MA and GRA herbaria for allowing the review of Noccaea nevadensis specimens. Also to Glòria Aymerich, Jana Colell, Anna Seuba, Llorenç Sáez, Ignasi Soriano and David Vilasís for his help in some aspects of this work.
Al-Shebahz, I. A. 2014. A synopsis of the genus Noccaea (Coluteocarpeae, Brassicaceae). Harvard Papers in Botany, 19: 25-51
Al-Shebahz, I. A. 2022. Noccaea klimesii (Coluteocarpeae; Brassicaceae), a new species from Ladakh, India. Phytotaxa, 555: 209-212.
Alejandre, J. A., Bariego, P., Benito, J., Escalante, M. A., García, J. M., Martín, L., Montamarta, G., Patino, S., Prieto, M. A. & Valencia, J. 2006. Atlas de la flora vascular silvestre de Burgos. Monografías de Botánica Ibérica 2. Jolube Consultor y Editor Ambiental. 925 p. Blanca, G., Cabezudo, B., Cueto, M., Fernández, C. & Morales, C. 2009. Flora vascular de Andalucía oriental. Vol. 3. Junta de Andalucía. Sevilla. 460 p.
Bolòs, O. & Vigo, J. 1990. Flora dels Països Catalans. Vol. 2. Ed. Barcino. Barcelona. 736 p.
Carlón, L., Laínz, M., Moreno, G., Rodríguez, J. M. & Sánchez, O. 2013. Contribuciones al conocimiento de la flora cantàbrica, IX. Documentos del Jardín Botánico de Gijón, 10: 1-153. Güzel, Y., Özüdoğru, B., Kayikçi, S. & Özgişi, K. 2018. Noccaea aliathanii (Brassicaceae): a new species from southern Anatolia. Turkish Journal of Botany, 42: 780-789.
Meyer, F. K. 1979. Kritisch Revision der “Thlaspi” – Arten Europas, Afrikas und Vorderasiens, I. Geschichte, Morphologie und Chorologie. Feddes Repertorium, 90: 129-154.
Özgişi, K. 2020. A new species of Noccaea (Brassicaceae) from central Anatolia. Phytotaxa, 432: 95-103.
Özgişi, K., Ocak, A. & Özüdoğru, B. 2018. Noccaea birolmutlui, a new crucifer species from South West Anatolia, Turkey. Phytotaxa, 345: 59-67.
Özüdoğru, B., Özgişi, K.,Tarıkahya-Hacıoğlu, B., Ocak, A., Mummenhoff, K. & Al-Shebahz, I. A. 2019. Phylogeny of the Genus Noccaea (Brassicaceae) and a Critical Review of its Generic Circumscription. Annals of the Missouri Botanical Garden, 104: 339–354.
Pignatti, S. 2017. Flora d’Italia. Vol. 2. Edagricole. Milà. 1196 p. Pujadas, A. 1993. Thlaspi L. P. 255-264. In: Castroviejo, S.; Aedo, C.; Gómez C.; Laínz, M.; Montserrat, P.; Morales, R.; MuñozGarmendia, F.; Nieto, G.; Rico, E.; Talavera, S.; Villar, L.(eds.). Flora iberica, Vol. 4. Real Jardín Botánico-CSIC. Madrid. 730 p.
Sáez, L. & Aymerich, P. 2021. An annotated checklist of the vascular plants of Catalonia (north-eastern Iberian Peninsula). Kitbook. Barcelona. 717 p.
Sefali, A., Yapar, Y. & Demir, I. 2023. Noccaea anatolica Sp. Nov. (Brassicaceae): A new species from Eastern Anatolia, Türkiye. Bitlis Eren University Journal of Science and Technology, 13: 187-197.
Tison, J.M. & Foucault, B. 2014. Flora Gallica. Flore de France Biotope. Mèze. 1195 p.
Tison, J.M., Jauzein, P. & Michaud, H. 2014. Flore de la France méditerranéenne continentale. Naturalia Publications. Tulliers. 2078 p.
Vigo, J., Soriano, I., Carreras, J., Aymerich, P., Carrillo, E., Masalles, R.M., Font, X. & Ninot, J.M. 2003. Flora del Parc Natural del CadíMoixeró i de les serres veïnes. Museu de Ciències NaturalsInstitut Botànic de Barcelona. Barcelona. 406 p.
Appendix 1. Noccaea nevadensis vouchers used for the comparative study
Spain: Sierra Nevada, Peñón de Jerez, ad 3000 m, supra Jerez del Marquesado, in lapidosis, 19-VII-1923, leg. Font Quer (BC109110). Sierra Nevada, puerto de Trevélez, in schistosis, ad 3000 m, 23-VII-1923, leg. P. Font Quer (BC109111). Granada, Sierra Nevada, carretera que sube al Mulhacén, ladera pr. a la Laguna del Peñón Negro, undated, leg. A.B. Robles & P. Sánchez, (GDA19851). Granada, Sierra Nevada, Peñones de San Francisco, 14-V-1944, leg. Muñoz Medina (GDA3389). Sierra Nevada, Hoya de la Mora, pedregales esquistosos, leg. A. Segura, 30-5-1967 (MA301963). Granada, subida a Sierra Nevada, cerca del refugio de la Mora, alt. 2550 m, 18-VII-1976, leg. A. Barra & al. (MA304535). Granada, subida a Sierra Nevada, alt. 2200-2550 m, sobre esquisto, 17-VII-1976, leg. A. Barra & al. (MA304551). Granada, Sierra Nevada, Pico Veleta, leg. P. Cubas, 1-VII-1978, G. López & M. Moreno (MA306154). Granada, Sierra Nevada, Hoya de San Juan, 2600 m, 5-VII1971, leg. Fernández Casas (MA409192). Sierra Nevada, región nival, 1-VII-1866, leg. P. del Campo (MA45047). Granada, Sierra Nevada, Cueva de los Panderones, 23-VI1858, leg. V. López Seoane, (MA45048). Granada, Sierra Nevada, undated, leg. P. Sáinz Gutérrez (MA45049). Granada, Sierra Nevada, entre enebros rastreros, 5-VIII-1976, leg. M. Pascual (MA620006). Granada, Cauchiles del Monachil, 2800 m, 25-VI-1970, leg.?, (MA753103). Granada, Sierra Nevada, faldas del pico Veleta cerca del viejo observatorio astronómico, matorrales y pastos psicroxerófilos, en sustrato ácido, esquistos, 4-VII-2010, leg. E. Glazkova & A. Quintanar (MA836384). Almería, puerto de la Ragua, 2100 m, pastizal, 18-IV-2014, leg. C. Aedo (MA89290).
Josep Gesti Perich*
* C/ Sant Ignasi, 42. 17430 Santa Coloma de Farners. A/e: josepgesti@tutamail.com
Rebut: 14.07.2024; Acceptat: 30.08.2024; Publicat: 30.09.2024
Resum
S’aporten dades sobre la flora vascular del massís de les Guilleries, i es constata la presència de diversos tàxons autòctons que no s’hi coneixien (Cuscuta approximata subsp. approximata, Diplotaxis virgata subsp. virgata, Genista linifolia subsp. linifolia, Ranunculus trichophyllus i Veronica cymbalaria) o que no s’hi havien retrobat des d’antic (Descurainia sophia, Persicaria amphibia) i de diverses espècies aŀlòctones noves o poc reportades fins ara en aquest massís (Austrocylindropuntia subulata, Berberis julianae, Clerodendrum bungei, Commelina communis, Cucurbita ficifolia, Eschscholzia californica, Fagopyrum esculentum, Kalanchoe ×houghtonii, Nassella tenuissima, Opuntia leoglossa, O. phaeacantha, Oxalis pes-caprae i Spiraea cantoniensis).
Paraules clau: flora vascular, massís de les Guilleries, Catalunya, península Ibèrica.
Abstract
New data on the vascular flora of the Guilleries massif (northeastern Catalonia)
We provide data on the vascular flora of the Guilleries massif, confirming the presence of several autochthonous taxa that were not known in this territory (Cuscuta approximata subsp. approximata, Diplotaxis virgata subsp. virgata, Genista linifolia subsp. linifolia, Ranunculus trichophyllus and Veronica cymbalaria) or not found since ancient times (Descurainia sophia, Persicaria amphibia) and several new or rarely reported non-native species (Austrocylindropuntia subulata, Berberis julianae, Clerodendrum bungei, Commelina communis, Cucurbita ficifolia, Eschscholzia californica, Fagopyrum esculentum, Kalanchoe ×houghtonii, Nassella tenuissima, Opuntia leoglossa, O. phaeacantha, Oxalis pes-caprae and Spiraea cantoniensis).
Key words: vascular plants, Guilleries massif, Catalonia, Iberian Peninsula. Butlletí de la Institució
Introducció
En el marc dels treballs de catalogació de la flora vascular de les Guilleries (https://floraguilleries.cat), en aquest article donem informació sobre la presència de diverses plantes que no havien estat indicades anteriorment al massís i també sobre algunes espècies de les quals es tenien molt poques dades o que no s’hi s’havien retrobat des d’antic. Hi apleguem tant tàxons autòctons com, sobretot, plantes aŀlòctones que s’afegeixen a la llarga llista d’espècies introduïdes que s’han reportat en els darrers anys al massís (Gesti, 2023).
Materials i mètodes
Totes les observacions que aportem provenen del massís de les Guilleries i han estat obtingudes entre els anys 2022 i 2024.
La llista dels tàxons es presenta segons l’ordre alfabètic dels noms científics (seguint Sáez & Aymerich, 2021), els quals van precedits per un asterisc (*) quan es tracta d’espècies aŀlòctones. Per a cadascun, donem localitats concretes
amb indicació de la comarca, el municipi i el lloc, el quadrat UTM d’1 km de costat (sempre a la zona 31T, sistema de referència ETRS89), l’altitud, l’hàbitat i la data d’observació. Així mateix, per a cada tàxon indiquem la seva distribució general a Catalunya (basada en Font, 2024), l’àrea de procedència nadiua si són aŀlòctons (seguint Gómez-Bellver, 2023 i POWO, 2024), les localitats conegudes properes a les Guilleries (quan n’hi ha) i una breu descripció de l’estat de les poblacions sobre les quals aportem informació.
Resultats i discussió
*Austrocylindropuntia subulata (Muehlenpf.) Backeb. Selva: Riudarenes, sota la Mare de Déu d’Argimon, DG6931, 460 m, rocams, 24-XII-2023 (Fig. 1a).
Cactàcia nativa de l’Amèrica del Sud que ha estat àmpliament indicada a l’àrea meridional de Catalunya i de manera més esparsa al litoral i prelitoral central i septentrional. Les citacions més properes a les Guilleries es situen a la Garriga (Sáez et al., 2015) i a Canet de Mar (C. Gómez-Bellver & N.
J. GESTI PERICH
Marquès in Font, 2024). N’hem vist una població formada per aproximadament una desena de peus, en un vessant rocós inclinat situat sota l’esplanada de l’ermita d’Argimon. En tractar-se d’un turó elevat i allunyat dels nuclis de població, creiem probable que l’espècie hi hagués estat plantada i que després s’hagi estès vegetativament pendent avall per arrelament de fragments.
*Berberis julianae C.K. Schneid.
Selva: Amer, sobre la presa del Pasteral, DG6648, 190 m, talús humit, 27-XI-2022 (Fig. 1b).
Berberidàcia nativa de l’Àsia de l’Est que a Catalunya només ha estat indicada a Ripoll i a Sant Joan de les Abadesses (Aymerich, 2020) i a Sant Quirze Safaja (Aymerich & Sáez, 2021). N’hem vist un petit rodal (que podria correspondre a rebrots d’un únic individu) on s’ha observat floració (març) i fructificació (novembre) els dos anys en què s’ha visitat (2022 i 2023).
*Clerodendrum bungei Steud.
Selva: Santa Coloma de Farners, riba de la riera de Santa Coloma, entre la Font Picant i la resclosa de Sant Salvador, DG7134, 130 m, plantació de plàtans (Platanus orientalis L. var. acerifolia Aiton), 25-III-2024 (Fig. 1c).
Lamiàcia originària del sud-est asiàtic que s’utilitza com a ornamental i de la qual només ha estat publicada una citació a Catalunya, a Arbúcies, en una localitat ubicada dins dels límits de les Guilleries (Aymerich, 2016). Afegim, doncs, una nova localitat d’aquesta espècie al massís, en aquest cas al sector oriental. A la riera de Santa Coloma n’hi hem observat una densa colònia, florint el mes de juliol, formada per més d’un centenar de tiges que ocupen una superfície d’uns 50 m2 d’una plantació de plàtans no gestionada. La densitat i la mida regular de les tiges fa pensar quepodrien ser rebrots fruit d’una extensió vegetativa.
*Commelina communis L.
Osona: Tavèrnoles, Fucimanya, DG4647, 510 m, talús ombrívol, 22-IX-2023 (Fig. 1d).
Commelinàcia originària de l’Àsia oriental i meridional coneguda d’unes poques localitats a Catalunya, a Cantallops (Font, 2000; Font & Vilar, 2000), la Pobla de Segur (Aymerich, 2019) i al riu Tordera a Sant Feliu de Buixalleu i Fogars de la Selva (Barnola in Font, 2024; Verloove & Aymerich, 2020). N’hem vist un petit rodal, en flor el mes de setembre, que recobria densament una superfície d’uns 8 m2, en un talús proper a un dels carrers de la urbanització.
*Cucurbita ficifolia C.D. Bouché
Selva: Anglès, carretera de Sant Amanç, prop de can Soldat, DG7042, 280 m, talús, 31-III-2024; Arbúcies, prop del Molí Sec, DG5930, 310 m, herbassar ruderal, 8-XII-2023; Hostalric, marge esquerra de la Tordera, prop del pont de la carretera BV5122, DG7021, 55 m, sorrals fluvials, 22-X-2023 (Fig. 1e).
Cucurbitàcia originària de Bolívia i del Perú de la qual no coneixem citacions anteriors a Catalunya com a espècie al-
Butlletí ICHN 88 (3), 2024
lòctona, per bé que es cultiva esporàdicament per a l’aprofitament de la polpa del fruit en confiteria (Fernandes, 2005).
A les tres localitats observades, totes elles situades a la zona més perifèrica del massís, hi hem vist únicament exemplars aïllats creixent en indrets molt ruderalitzats. A Hostalric es trobava florida el mes d’octubre.
Cuscuta approximata Bab. subsp. approximata
Osona: Seva, sota la collada de Sobrevia, DG4333, 700 m, matollar, 29-VI-2024 (Fig. 1f).
Convolvulàcia àmpliament distribuïda per les àrees temperades d’Euràsia, d’Àfrica i de l’Amèrica del Nord que a Catalunya ha estat citada sobretot a les terres interiors i pirinenques, i de manera molt més escassa a la resta del territori. Les localitats que s’han reportat més a prop de les Guilleries són les de Castellterçol i Sant Quirze de Safaja (Mercadé, 2016). A Seva l’hem vist, en flor al mes de juny, parasitant Astragalus monspessulanus L. subsp. gypsophilus Rouy i Hippocrepis scorpioides Benth.
Descurainia sophia (L.) Prantl
Osona: Sant Julià de Vilatorta, sota el Pla de les Cabres, DG4342, 570 m, herbassar ruderal al marge d’un camí, 28V-2023 (Fig. 1g).
Brassicàcia originària d’Euràsia i del Nord d’Àfrica que s’ha reportat profusament a les àrees continentals i pirinenques de Catalunya però de manera molt més esparsa a la resta del país. Fou citada a les Guilleries per Vayreda (1902) que la va observar a “Monsolí” (Mansolí, Sant Hilari Sacalm) i no s’hi havia retrobat des d’aleshores. A Sant Julià de Vilatorta n’hem vist només dos peus, en flor el mes de maig, en un espai freqüentat i ruderal, la qual cosa fa pensar que hi podria haver arribat accidentalment.
Diplotaxis virgata (Cav.) DC. subsp. virgata
Selva: Massanes, sota de can Tos de la via, DG7122, 55 m, herbassar al marge d’un camp, 25-II-2023.
Brassicàcia de distribució ibèrica que a Catalunya es coneix principalment de la meitat occidental i que, en canvi, ha estat escassament reportada del quadrant nord-oriental, a Barcelona i al Vallès Oriental (Martínez Laborde, 1992; Panareda et al., 2023). No tenim constància que hagi estat citada abans a les Guilleries. La indicació més propera és a Sant Celoni (Panareda et al., 2023). N’hem vist una població formada per algunes desenes d’individus, en flor al febrer, resseguint el marge d’un sembrat.
*Eschscholzia californica Cham.
Osona: Vilanova de Sau, prop de mas Arbell, DG4944, 520 m, herbassar ruderal, 10-V-2024 (Fig. 1h).
Papaveràcia nadiua de la part occidental de l’Amèrica del Nord que s’ha estès com a aŀlòctona pel seu us ornamental. A Catalunya s’ha reportat principalment a l’àrea litoral i prelitoral, però no tenim constància de citacions anteriors al massís de les Guilleries. Les localitats reportades més a prop són

1. a) Austrocylindropuntia subulata (Riudarenes); b) Berberis julianae (Amer); c) Clerodendrum bungei (Santa Coloma de Farners); d) Commelina communis (Tavèrnoles); e) Cucurbita ficifolia (Hostalric); f) Cuscuta approximata subsp. approximata (Seva); g) Descurainia sophia (Sant Julià de Vilatorta); h) Eschscholzia californica (Vilanova de Sau); i) Fagopyrum esculentum (Massanes).
a Riells i Viabrea (Barnola et al., 2021) i a Maçanet de la Selva (HGI 13355, leg. T. Hernández, 6-VIII-1997). A Vilanova de Sau n’hem vist quatre peus, en flor el mes de maig, en un marge de camí entre masos (des dels quals es podria haver estès).
*Fagopyrum esculentum Moench
Selva: Massanes, entre ca l’Aulet i can Pererol, DG6924, 175 m, camp sense cultivar, 19-V-2024 (Fig. 1i).
Poligonàcia originària de l’Himàlaia i la Xina que havia
estat cultivada en el passat a Catalunya però que actualment es troba en desús, raó per la qual la major part de les citacions catalanes son antigues. No n’hi ha referències del massís de les Guilleries per bé que se sap que es cultivava ben a prop, entre l’Empalme (estació de ferrocarril de Maçanet-Massanes) i Sils (Llensa (1945). L’hem vist abundant i en flor el mes de maig, al marge d’un camp que havia estat llaurat però no sembrat. Desconeixem si l’espècie hi ha arribat (cultivada o accidentalment) els anys anteriors o si es tracta de poblacions que s’hi han mantingut des de molt més antic.
J. GESTI PERICH
Genista linifolia L. subsp. linifolia
Selva: Riudarenes, turó de les Marfulledes, a tocar del camí d’Argimon, DG7031, 275 m, brolla esclarissada, 16-VI-2024 (Fig. 2a).
Fabàcia de distribució mediterrània occidental que a Catalunya es fa al llarg del litoral i prelitoral del quadrant nordoriental. No tenim coneixement que hagués estat reportada a les Guilleries, per bé que havia estat observada per Zeller (1958) no massa lluny del límit oriental del massís, entre Sils i Tordera. Al paratge d’Argimon n’hi hem vist cinc exemplars en una àrea d’uns 50 m2; dos d’ells de port gran i fructificats i els altres tres més petits i no reproductius.
*Kalanchoe ×houghtonii D.B. Ward
Selva: Santa Coloma de Farners, esplanada sobre el parc de Sant Salvador, DG7134, 130 m, mur, 8-III-2024.
Crassulàcia producte de la hibridació artificial entre K. daigremontiana i K. tubiflora que es troba en clara expansió a Catalunya (Gómez-Bellver, 2023), molt especialment al llarg de tota la franja litoral i als ambients periurbans. A les Guilleries se’n conexia una localitat de Sant Sadurní d’Osormort (Aymerich, 2023), a la qual n’afegim una segona, que es troba al sector oriental del massís. N’hem vist una petita població, amb dos individus grossos florits i amb abundants propàguls foliars, envoltats de petits exemplars originats a partir d’aquests propàguls.
*Nassella tenuissima (Trin.) Barkworth
Osona: Vilanova de Sau, riba esquerra del pantà de Sau, sobre l’església de Sant Romà, DG4947, 420 m, erm, 7-VII2023 (Fig. 2b).
Gramínia originària del continent americà emprada en xerojardineria que a Catalunya ha estat reportada com a espècie aŀlòctona a Viladecans (Álvarez et al., 2016), a la Zona Franca (BC 932580, leg. S. Pyke, 23-IX-2013), a Reus (Verloove et al., 2019) i a Salt (Verloove & Aymerich, 2020). Al pantà de Sau n’hi hem vist tres mates, en flor el mes de juliol, en un erm amb escassa vegetació, sobre els sòls argilosos dessecats després de la baixada del nivell de l’aigua.
*Opuntia leoglossa Font & M. Köhler
Selva: Riudarenes, pista forestal a l’oest de can Murgadella, DG6930, 225 m, talús sorrenc, 3-XII-2023 (Fig. 2c); Sant Hilari Sacalm, antigues feixes al Serrat, DG5935, 850 m, herbassar ruderal, 7-XII-2022.
Cactàcia d’origen desconegut (possible híbrid artificial) descrita recentment com a espècie (Köhler & Font, 2021) que només es coneix naturalitzada a Austràlia i a la península Ibèrica. A Catalunya s’ha reportat principalment al litoral, i són molt escasses les citacions a l’interior (Gómez-Bellver, 2023). A Sant Hilari Sacalm n’hem vist una petita colònia en una feixa situada sota d’un carreró on també hi havia alguns exemplars, presumiblement plantats, que suposem que deuen ser el focus de dispersió. A Riudarenes hi hem observat un sol rodal a tocar d’una pista forestal, on suposem que hi de-
Butlletí ICHN 88 (3), 2024 GEA, FLORA ET FAUNA
via arribar algun fragment accidentalment o amb algun abocament.
*Opuntia phaeacantha Engelm.
Selva: Hostalric, vessant oriental del turó del Castell d’Hostalric, DG6921, 130 m, prats secs, 10-III-2024 (Fig. 2d).
Cactàcia nadiua del sud-oest dels Estats Units i nord de Mèxic que ha estat reportada esparsament a la meitat meridional de Catalunya, al Bages, Baix Camp, Baix Llobregat i Montsià. Si bé de vegades s’ha confós amb O. engelmannii Salm-Dyck (Gómez-Bellver, 2023; Verloove & Guiggi, 2019), a les plantes que hem observat a Hostalric tant els caràcters morfològics (mida dels cladodis, nombre i densitat d’areoles, espines i gloquidis) com el port cespitós, de menys de 80 cm d’alçada, s’adiuen amb O. phaeacantha. Forma una densa colònia quasi contínua que cobreix una superfície d’uns 250 m2, la qual es trobava profusament fructificada el mes de març. De l’anàlisi d’ortoimatges aèries històriques (VISSIR, 2024) concloem que aquesta planta és present al turó com a mínim des de l’any 2009 i que s’hi ha anat estenent lentament des d’aleshores.
*Oxalis pes-caprae L.
Selva: Santa Coloma de Farners, esplanada sobre el parc de Sant Salvador, DG7134, 130 m, herbassar ruderal, 8-III-2024 (Fig. 2e).
Oxalidàcia originària de l’Àfrica del Sud que es troba en expansió a Catalunya, on ha estat localitzada al llarg de tota la franja litoral i també en alguns sectors prelitorals, sobretot al sud del país. A Santa Coloma de Farners n’hem vist alguns exemplars, en flor el mes de març, creixent en un espai fortament ruderalitzat on s’observaven abocaments de restes vegetals, que podrien ser el vector d’arribada de l’espècie.
Persicaria amphibia (L.) Delarbre
Osona: Viladrau, presa del Molí Espatllat, a la riera de Font Savellà, DG4433, 690 m, aigües permanents de curs lent, 8-VI-2024 (Fig. 2f).
Poligonàcia de distribució subcosmopolita indicada en molts punts de Catalunya, sobretot a la meitat septentrional. Havia estat documentada per Llensa (1945) al sector meridional de les Guilleries (Massanes), i també per Bolòs (1983) al límit entre el Montseny i les Guilleries (vora el riu Gurri, sota la carretera de Seva a Viladrau). La localitat que aportem es troba força a prop d’aquesta darrera, també a cavall entre el Montseny i les Guilleries, tot i que ja no a la conca del Gurri sinó en la d’un tributari de la riera Major. Cobria una superfície d’uns 250 m2 de la làmina d’aigua i es trobava en plena floració al mes de juny.
Ranunculus trichophyllus Chaix
Osona: Viladrau, presa del Molí Espatllat, a la riera de Font Savellà, DG4433, 690 m, aigües permanents de curs lent, 8-VI-2024 (Fig. 2g).
Ranunculàcia aquàtica subcosmopolita àmpliament reportada a Catalunya per bé que a les Guilleries només ens consta

2. a) Genista linifolia subsp. linifolia (Riudarenes); b) Nassella tenuissima (Vilanova de Sau); c) Opuntia leoglossa (Riudarenes); d) Opuntia phaeacantha (Hostalric); e) Oxalis pes-caprae (Santa Coloma de Farners); f) Persicaria amphibia (Viladrau); g) Ranunculus trichophyllus (Viladrau); h) Spiraea cantoniensis (Vilanova de Sau); i) Veronica cymbalaria (Massanes).
una citació de Folgueroles que correspon al grup (Pérez-Haase et al., 2013, sub. R. gr. trichophyllus Chaix). A la localitat que indiquem, que es troba en el límit entre Montseny i les Guilleries, s’hi feia abundantment tot convivint amb l’espècie anterior; en flor i fruit el mes de juny.
*Spiraea cantoniensis Lour.
Osona: Vilanova de Sau, sota la presa de Sau, DG5146, 400 m, bardissa, 17-V-2023 (Fig. 2h). Selva: Massanes, prop de can Remilans, DG7124, 80 m, vorada de camí, 2-VI-2024.
Rosàcia nadiua del sud-est de la Xina que ha estat àmpliament utilitzada com a ornamental i que a Catalunya s’ha citat ocasionalment com a aŀlòctona al Baix Ebre (Royo, 2006), al Berguedà (Aymerich, 2013, 2017), al Ripollès (Vigo, 1983) i a la Selva (Aymerich, 2017). A Sau hi hem observat diversos individus que semblen subespontanis i que s’hi deuen haver establert a partir de fruits procedents de plantes cultivades prop de la presa. A Massanes no podem assegurar si els exemplars observats són subespontanis o si es tracta de rebrots de plantes cultivades en el passat.
GEA, FLORA ET FAUNA
Veronica cymbalaria Bodard
Selva: Massanes, camí del Cambrerol, prop de la zona esportiva, DG7123, 125 m, herbei al voral del camí, 10-III-2024 (Fig. 2i).
Plantaginàcia de distribució circummediterrània que a Catalunya ha estat molt poc reportada i de manera molt esparsa. No tenim coneixement que hagués estat citada a les Guilleries. Les indicacions més properes són l’estació del ferrocarril de Caldes de Malavella (HGI 5882, leg. L. Vilar, 15-V-1991) i la de Fontmartina (Fogars de Montclús; BC 925429, leg. N. Escué, 18-III-1990). A Massanes n’hem vist una nodrida població, en flor al mes de març, repartida al llarg d’uns 10 metres d’un voral viari.
Bibliografia
Álvarez, H., Ibáñez, N. & Gómez-Bellver, C. 2016. Noves aportacions al coneixement de la flora aŀlòctona de la comarca del Baix Llobregat (Catalunya, Espanya). Collectanea Botanica (Barcelona), 35: e007.
Aymerich, P. 2013. Plantas alóctonas de origen ornamental en la cuenca alta del río Llobregat (Cataluña, noreste de la Península Ibérica). Bouteloua, 16: 52-79.
Aymerich, P. 2016. Contribució al coneixement de la flora aŀlòctona del nord i el centre de Catalunya. Orsis, 30: 11-40.
Aymerich, P. 2017. Notes sobre flora aŀlòctona a Catalunya. Butlletí de la Institució Catalana d’Història Natural, 81: 97-116.
Aymerich, P. 2019. Notes sobre flora aŀlòctona a Catalunya. II. Butlletí de la Institució Catalana d’Història Natural, 83: 3-21.
Aymerich, P. 2020. Notes sobre flora aŀlòctona a Catalunya. III. Butlletí de la Institució Catalana d’Història Natural, 84: 101124.
Aymerich, P. & Sáez, L. 2021. Aportacions a la flora al·lòctona catalana. Butlletí de la Institució Catalana d’Història Natural, 85: 151-162.
Aymerich, P. 2023. Notes sobre flora aŀlòctona a Catalunya. IV. Butlletí de la Institució Catalana d’Història Natural, 87: 225-238. Barnola, P., Panareda, J. M. & Gesti, J. 2021. Aportacions al coneixement de la flora vascular del sector oriental del massís del Montseny (Catalunya, NE de la península Ibèrica). Butlletí de la Institució Catalana d’Història Natural, 85: 199-207. Bolòs, O. 1983. La vegetació del Montseny. Diputació de Barcelona, Servei de Parcs Naturals. Barcelona. 170 p. Fernandes, R. B. 2005. Cucurbita L. P. 466-470. In: Castroviejo, S.; Aedo, C.; Cirujano, S.; Laínz, M.; Montserrat, P.; Morales, R.; Muñoz Garmendia, F.; Navarro, C.; Paiva, J.; Soriano, C. (eds.). Flora iberica III - Plumbaginaceae (partim)-Capparaceae. Real Jardín Botánico. Consejo Superior de Investigaciones Científicas. Madrid. 657 p.
Font, J. 2000. Estudis botànics de la serra de l’Albera. Catàleg florístic general i poblament vegetal de les basses de l’Albera Tesi doctoral. Universitat de Girona. Font, J. & Vilar, L. 2000. Plantes vasculars del Quadrat UTM 31T DG99, Sant Climent Sescebes. ORCA: Catàlegs Florístics Locals, 10. Institut d’Estudis Catalans (Secció de Ciències Biològiques). Barcelona. 76 p.
Font, X. 2024. Mòdul de Flora i Vegetació. Banc de Dades de Biodiversitat de Catalunya. Generalitat de Catalunya i Universitat
de Barcelona. Disponible a: http://biodiver.bio.ub.es/biocat/ homepage.html [Data de consulta: 20 juny 2024].
Gesti, J. 2023. Flora de les Guilleries orientals. Catàleg, caracterització i espècies singulars. Càtedra de l’Aigua, Natura i Benestar - Eumo. Sant Hilari Sacalm - Vic. 218 p. Gómez-Bellver, C. 2023. Flora aŀlòctona vascular ocasional, naturalitzada i invasora al territori comprès per Catalunya, el País Valencià i les Illes Balears. El cas d’estudi del neòfit recent invasor Kalanchoe ×houghtonii. Tesi doctoral. Universitat de Barcelona. Barcelona. 1.233 p. Köhler, M. & Font, F. 2021. Opuntia leoglossa sp. nov. (Cactaceae): a new identity for the aloctone “Lion’s Tongue” cactus. Phytotaxa, 510: 281-287.
Llensa, S. 1945. Inventario razonado de la Flora de Hostalrich y su comarca. Anales de la Escuela de Peritos Agrícolas y Superior de Agricultura y de los Servicios Técnicos de Agricultura, 5: 121-290. Martínez Laborde, J. B. 1992. Sobre la corología de Diplotaxis DC. (Cruciferae, Brassiceae). Anales del Jardín Botánico de Madrid, 50: 276-278.
Mercadé, A. 2016. Estudis de flora i vegetació del Moianès i àrees properes. Tesi doctoral. Universitat de Barcelona. 619 p. Panareda, J. M., Barnola, P. & Gesti, J. 2023. Noves aportacions al coneixement de la flora vascular del massís del Montseny (Catalunya, NE de la península Ibèrica). Butlletí de la Institució Catalana d’Història Natural, 87 (1): 29-34.
Pérez-Haase, A., Mercadé, A., Batriu, E. & Blanco, J. M. 2013. Aportació al coneixement florístic de l’Espai Natural de les Guilleries-Savassona. Grup de Recerca de Geobotànica i Cartografia de la Vegetació. Universitat de Barcelona. Barcelona. 238 p.
POWO. 2024. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Disponible a: http://www. plantsoftheworldonline.org [Data de consulta: 20 juny 2024].
Royo, F. 2006. Flora i vegetació de les planes i serres litorals compreses entre el riu Ebro i la serra d’Irta. Tesi doctoral. Universitat de Barcelona. Barcelona. 1.134 p.
Sáez, L. & Aymerich, P. 2021. An annotated Checklist of the Vascular Plants of Catalonia (northeastern Iberian Peninsula) Kit-book Serveis Editorials, S.C.P. Barcelona. 717 p.
Sáez, L., Galán de Mera, A., Pyke, S., Pié, G. & Carnicero, P. 2015. New data on vascular plants from Montseny massif (northeastern Iberian Peninsula). Orsis, 29: 205-230.
Vayreda, E. 1902. Plantas de Cataluña. Anales de la Sociedad Española de Historia Natural, 10 (30): 491-582.
Verloove, F., Aymerich, P., Gómez-Bellver, C. & López-Pujol, J. 2019. Chorological notes on the non-native flora of the province of Tarragona (Catalonia, Spain). Butlletí de la Institució Catalana d’Història Natural, 83: 133-146.
Verloove, F. & Guiggi, A. 2019. Further records of cacti (Cactaceae) from Tarragona province (Catalonia), Spain. Haseltonia, 26: 2335.
Verloove, F. & Aymerich, P. 2020. Chorological novelties for the alien flora of northeastern Catalonia (Iberian Peninsula). Butlletí de la Institució Catalana d’Història Natural, 84: 137-153.
Vigo, J. 1983. Flora de la vall de Ribes. I. Generalitats. Catàleg florístic. Acta Botanica Barcinonensia, 35: 1-793.
VISSIR. 2024. Visualitzador avançat de geoinformació. Institut Cartogràfic i Geològic de Catalunya. Disponible a: http://srv.icgc. cat/vissir3 [Data de consulta: 3 juliol 2024].
Zeller, W. 1958. Ètude phytosociologique du chêne-liège en Catalogne. Pirineos, 47-50: 1-194.
Butlletí ICHN
(3), 2024
Andreu Cera1, Maria José Chesa2, Àlex Cortada3, Joana Mendes4, Antonio Gómez-Bolea5,6.
1 UMR 950 EVA, INRAE, Université de Caen-Normandie. Caen, França.
2 Alichenology, Travessera de les Corts, 365. 08029 Barcelona, Espanya.
3 Carrer Picasso, 37. Maó, Espanya.
4 Institut Menorquí d’Estudis (IME). Maó, Espanya.
5 Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals, Universitat de Barcelona. Barcelona. Espanya.
6 Institut de Recerca de la Biodiversitat (IRBio). Universitat de Barcelona. Barcelona, Espanya.
Autor de correspondència: Andreu Cera. A/e: andreucera@outlook.com
Rebut: 01.02.2024; Acceptat: 10.09.2024; Publicat: 30.09.2024
Resum
Ramalina lusitanica i R. pusilla són dos líquens epífits de distribució litoral a la regió mediterrània, al litoral sud-oest de la península Ibèrica i a la regió macaronèsica. A Catalunya hi ha poques citacions, i la majoria són antigues. En aquest treball hem verificat la presència d’aquests líquens en les quadrícules UTM 10 × 10 km on van ser citats anteriorment, i hem prospectat noves quadrícules de la costa de Catalunya on es podrien trobar les condicions idònies per a la seva presència. Hem seleccionat i visitat 15 quadrícules. Com a resultat d’aquest treball, el nombre de quadrícules amb presència de R. pusilla passen de tres a sis i el de R. lusitanica d’una a tres. Hem caracteritzat biològicament i microclimàticament una població amb presència de les dues espècies de Ramalina. Arran d’aquest treball, hem proposat la inclusió de R. lusitanica com a espècie en perill d’extinció al Catàleg de Flora amenaçada de Catalunya, considerant que d’aquesta forma també queda protegida R. pusilla, perquè totes dues espècies formen part de la mateixa comunitat.
Paraules clau: autoecologia, litoral, mediterrani, temperatura, humitat relativa, espècies amenaçades.
Abstract
Data on chorology and autoecology of Ramalina lusitanica and Ramalina pusilla in Catalonia
Ramalina lusitanica and R. pusilla are two epiphytic lichens with a coastal distribution in the Mediterranean region, southwest coast of the Iberian Peninsula and the Macaronesian region. On the coast of Catalunya, there are very few known populations, and most of them are old references. In this study, we have verified the presence of these lichens in the UTM 10 × 10 squares where they were previously cited and have prospectively identified new squares on the Catalan coast where the optimal conditions for their presence could be found. We have selected and visited 15 squares. As a result of this work, the number of squares with the presence of R. pusilla increased from three to six, while that of R. lusitanica increased from one to three. We have biologically and microclimatologically characterised a population with the presence of both Ramalina species. After this work, we have therefore proposed the protection of R. lusitanica as endangered species to the Catalogue of Threatened Flora of Catalunya, on the grounds that this will also benefit R. pusilla, which is part of the same community.
Key words: autoecology, litoral, mediterranean, temperature, relative humidity, threatened species.
Introducció
Ramalina lusitanica H. Magn i R. pusilla Duby són dos líquens de distribució litoral a la regió mediterrània, al sud-oest de la península Ibèrica i a la regió macaronèsica. Viuen com a epífits a les branques i branquillons d’arbres i arbustos de la zona litoral. És en aquest ambient on troben suficient humitat atmosfèrica, llum i temperatura, donat que són considerades espècies aerohigròfiles, heliòfiles i termòfiles (Roux, 2020). Les dues espècies han estat catalogades en diferents llistes d’espècies amenaçades de regions properes a Catalunya. A Itàlia, les dues espècies estan catalogades com a vulnerables
a la llista vermella dels líquens epífits (Nascimbene et al., 2013). A França, R. lusitanica està considerada en perill crític d’extinció en el catàleg de líquens (Roux, 2020), en canvi R. pusilla no està amenaçada. Al País Valencià, R.lusitanica no hi és present i R. pusilla està considerada en perill d’extinció en el catàleg de flora amenaçada (Atienza & SegarraMoragues, 2006), perquè només hi ha dues poblacions de R. pusilla amb molts pocs taŀlus. Tanmateix, fins a l’any 2022, a Catalunya, R. lusitanica només havia estat citada una vegada a l’Escala l’any 2017 (segons el portal gbif.org), i R. pusilla havia estat citada a l’Escala també l’any 2017, i a Begur i a Tossa de Mar durant la dècada dels anys 1980 (segons el
A. CERA ET AL.
portal gbif.org). A altres regions de la costa mediterrània peninsular el nombre de poblacions també és escàs (segons el portal gbif.org).
Per tot el que s’ha exposat anteriorment, es fa necessari un estudi de la presència d’aquestes dues espècies a Catalunya. L’objectiu d’aquest treball ha sigut la localització de les poblacions anteriorment citades, la cerca de noves poblacions de R. lusitanica i R. pusilla a Catalunya, i la caracterització microclimàtica d’una localitat amb presència de les dues espècies.
Metodologia
En el transcurs dels anys 2022, 2023 i 2024 hem seleccionat i visitat 15 quadrícules UTM 10 × 10 km, de les 48 que hi ha a la costa de Catalunya (Taula 1). Per un cantó, hem seleccionat les quadrícules que tenien citacions antigues de més d’una dècada segons el portal gbif.org (Taula 1), i per un altre totes les quadrícules potencials, considerant com a tals aquelles amb vegetació arbòria o arbustiva que estan més o menys arrecerades del vent i que, per tant, presumiblement tenen una alta humitat relativa. Hem descartat les zones exposades a forts vents, com ara el vessant nord del Cap de Creus o les serres costaneres de les muntanyes de Tivissa-Vandellòs. També s’ha descartat la costa del Maresme i la del Barcelonès per estar molt urbanitzades i exposades (Visió amb el Google Earth Pro). La prospecció s’ha fet fins a l’alçada que hem pogut arribar estant a peu pla.
Taula 1. Resultats de les visites a les quadrícules UTM de 10 × 10 km on potencialment hi hauria R. lusitanica i R. pusilla. * Aquesta citació va ser introduïda al GBIF posteriorment a l’any 2022, quan ja s’havia començat el treball de camp realitzat per nosaltres.
UTM 31T Citacions anteriors Visitades 2022-2024
EG17 R. pusilla
EG16 R. pusilla, R. lusitanica 2017* No visitada*
EG14 R. pusilla: 1985, 1986 R. pusilla
EG13 R. lusitanica, R. pusilla
DG91 R. pusilla: 1985, 1986 R. pusilla
DF06 No trobades
CF65 No trobades
CF55 No trobades
CF01 No trobades
CF11 R. lusitanica R. pusilla
CF21 No trobades
CF10 No trobades
CF00 No trobades
CE09 No trobades
BE89 No trobades
BE88 No trobades
A les localitats on hi havia Ramalina lusitanica i/o R. pusilla hem anotat el foròfit i hem quantificat, aproximadament, el nombre de taŀlus de les dues espècies. Per a la caracterització microclimàtica, hem escollit la localitat del Garxal (Deltebre, Baix Ebre, 1-2 m s.n.m., 31TCF11), ja que presenta les dues espècies de Ramalina estudiades amb els taŀlus més ben desenvolupats. Aquesta loca-
Butlletí
2024
litat és un sistema dunar proper al mar i amb inundacions estacionals. En aquesta localitat hem ubicat dos sensors HOBO Temperature/RH Data Logger (MX2301A, ONSET, USA). El primer (Garxal 1) prop del taŀlus de Ramalina lusitanica a 15 cm del terra i el segon (Garxal 2) a prop del taŀlus de Ramalina pusilla a 15 cm del terra. Aquests sensors han recollit dades durant els 365 dies que van des de l’1 de maig de 2022 al 30 d’abril de 2023. Les nostres dades les hem comparat amb les recollides, en el mateix període, per l’estació meteorològica oficial més propera, que és a l’illa de Buda (https:// www.meteo.cat/observacions/xema/dades?codi=DL).
Resultats i discussió
Hem detectat la presència de R. lusitanica a dues quadrícules en les que també hi havia R. pusilla (EG13 i CF11), i de R. pusilla a cinc quadrícules, incloses les dues anteriors (EG17, EG14, EG13, DG91 i CF11).
Localitats amb presència de R. lusitanica i R. pusilla són:
Cala Pedrosa, Palafrugell, Baix Empordà, 1-80 m s.n.m. 22/02/2022. 31TEG13.
Una vintena de taŀlus de R. lusitanica en branquetes de tres peus d’Erica arborea L., al vessant nord de la cala. Centenars de taŀlus petits de R. pusilla tant sobre Erica arborea, com sobre branques i branquillons de Pinus halepensis Mill
Cala Castell, Palamós, Baix Empordà, 2-50 m s.n.m. 24/03/2024. 31TEG13.
Dos taŀlus de R. lusitanica sobre Quercus ilex L. i dos més sobre Pistacia lentiscus L. Centenars de petits taŀlus de R. pusilla sobre Quercus ilex, Pistacia lentiscus i branquillons de Pinus halepensis.
La Vinya d’en Gervasi, Palafrugell, Baix Empordà, 100 m s.n.m. 22/02/2022. 31TEG13.
Deu taŀlus de R. lusitanica sobre dos Quercus suber. Petits taŀlus de R. pusilla sobre Quercus suber i altres arbustos.
El Garxal, Deltebre, Baix Ebre, 1-2 m s.n.m. 08/02/2022. 31TCF11.
Dos taŀlus de R. lusitanica sobre un arbust de Thymelaea hirsuta Endl. Més d’una cinquantena de taŀlus de R. pusilla sobre Thymelaea hirsuta, dels quals 10 són de mida grossa i la resta són de mida petita (Figura 1).
Localitats amb només presència de R. pusilla són:
Cala Montjoi, Roses, Alt Empordà ,13-35 m s.n.m. 16/05/2022. 31TEG17
Un únic taŀlus gros de R. pusilla sobre Phillyrea angustifolia L., i també tres taŀlus a la vall de Montjoi sobre Olea europea
Sa Riera, Begur, Baix Empordà, 15-20 m s.n.m. 24/02/2024. 31TEG14.
En 12 arbres de les espècies Fraxinus sp. i Acer sp hi havia 10 taŀlus visibles i identificables per arbre. L’observació confirma la presència de R. pusilla en aquesta quadrícula de
les muntanyes de Begur, després de les citacions de GómezBolea (1985) i Boqueras & Gómez-Bolea (1986).
El Far de Sant Sebastià, Calella de Palafrugell, Baix Empordà, 150 m s.n.m. 05/01/2022. 31TEG13.
Desenes de taŀlus de R. pusilla sobre Olea europaea L. Cala Bona, Tossa de Mar, la Selva, 1-40 m s.n.m. 22/02/2022. 31TDG91.
Quasi una vintena de taŀlus de R. pusilla sobre Rhamnus alaternus L., Phillyrea angustifolia, Quercus suber, Erica arborea i Pinus halepensis. Observació que confirma la seva presència després de les citacions de Gómez-Bolea (1985) i Boqueras & Gómez-Bolea (1986).
El caràcter heliòfil d’aquestes espècies fa que es trobin com a epífites sobre arbres i arbusts molt defoliats, on arriba prou llum, però també a les branques i branquillons de les capçades, on nosaltres no hem pogut accedir. Alternativament, és freqüent trobar branques i branquillons caiguts a terra on podem observar aquesta comunitat. Una prospecció més complerta comportaria poder accedir a aquestes alçades mitjançant cordes o bé fent caure les branques més altes, dues tècniques que sempre seran destructives. L’ús de binocles l’hem provat, però no ens dona una resolució prou bona per poder identificar les espècies. La situació és ben diferent al Baix Ebre, on els foròfits tenen poca alçada i, consegüentment, les nostres observacions s’apropen més a la presència real d’aquestes espècies.
Caracterització biològica i microclimàtica de la comunitat de R. lusitanica i R. pusilla a la localitat del Garxal
Es tracta d’un sistema dunar proper al mar i amb inundacions estacionals. El principal foròfit de les dues Ramalina és Thymelaea hirsuta. Els líquens epífits acompanyants són Buellia mediterranea Giralt, Diploicia canescens (Dicks.) A. Massal., Flavoparmelia soredians (Nyl.) Hale, Hyperphyscia adglutinata (Flörke) H. Mayrhofer & Poelt., Lecanora conizella Nyl., Lecanora lividocinerea Bagl., Lecanora strobilinoides Giralt & Gómez-Bolea, Physcia adscendens H. Olivier, Polyblastia rouxiana Vězda & Vivant, Pyrrhospora quernea (Dicks.) Körb., Ramalina canariensis J. Steiner i Xanthoria parietina (L.) Th. Fr.
Els sensors (Garxal 1 i Garxal 2) han registrat dades des de l’1 de maig de 2022 fins al 30 d’abril de 2023. En aquests 365 dies, la T mitjana va ser de 19,4 °C (Garxal 1) i 19,3 °C (Garxal 2), quasi un grau més que a l’estació meteorològica de l’illa de Buda on, en el mateix període, la T mitjana va ser de 18,5 °C. L’amplitud tèrmica anual, calculada a partir de la mitjana mensual, va ser de 20,7 °C (Garxal 1) i 20,4 °C (Garxal 2), i només de 16,6 °C a l’illa de Buda (Figura 2). Si considerem, com diuen Lange et al. (1977) per a Ramalina maciformis (en una localitat costanera del desert de Negev, Israel), que una humitat relativa (Hr) de l’aire superior al 80% seria el valor límit inferior per a una activitat fotosintètica neta respecte a la respiració, en el Garxal 1 hem observat un total de 2.998 hores anuals en què se supera el 80% de Hr, i 2.910 en el Garxal 2 (Figura 3). Aquests valors són molt més alts a l’estació meteorològica de l’illa de Buda, on es registren un total de 4.321 hores en què la Hr és superior al 80%. Aquestes diferències entre l’estació meteorològica i els sensors posen en relleu la importància de tenir dades de microhàbitat per descriure l’autoecologia d’aquestes espècies.

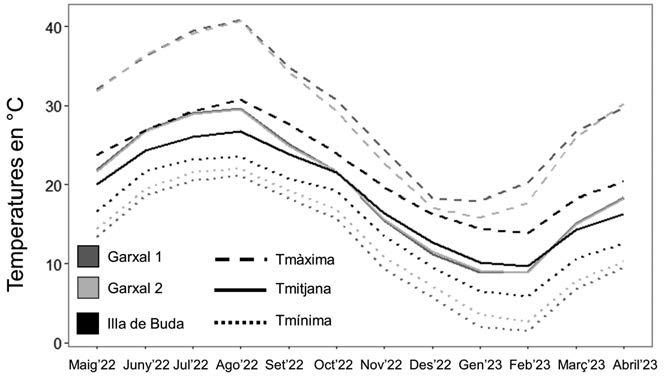
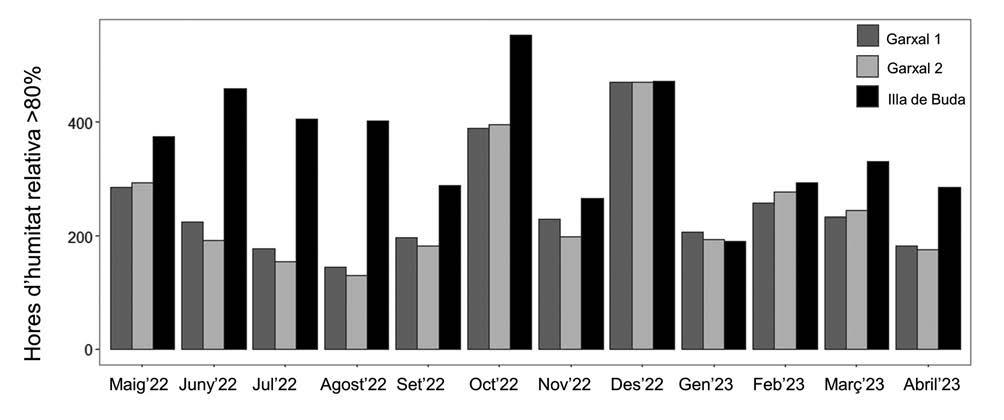
El context biogeogràfic i català de R. lusitanica i R. pusilla
R. lusitanica i R. pusilla formen part d’una comunitat litoral de líquens fruticulosos epífits en què hi ha altres espècies de Ramalina, amb una distribució més amplia, com és el cas de R. lacera, que al País Valencià està catalogada en la llista vermella (Atienza & Segarra-Moragues, 2006), i R. canariensis que l’hem trobat a quasi totes les localitats visitades i és la de més amplia distribució de totes elles. Aquestes espècies de Ramalina tenen el seu nínxol ecològic dins la zona litoral, i a la vegada són líquens aerohigròfils (Roux, 2020). La seva distribució a la península Ibèrica ocupa la costa alentejana (Carvalho et al., 2002) i la desembocadura del Tajo (Sérgio et al., 2016), on es troben la majoria de poblacions d’aquestes espècies, però també les tenim a la costa mediterrània peninsular on hi ha molt poques citacions (segons el portal gbif. org). En canvi, a les illes mediterrànies, com per exemple a l’illa de Menorca, hi ha una gran abundància d’aquestes espècies de Ramalina, fins i tot a les àrees rurals i aforestades de l’illa (Cera et al., 2022).
Butlletí ICHN
(3), 2024
A Catalunya, R. lusitanica sembla més estenoica, o bé es troba al límit de la seva àrea de distribució. El cas és que té un àrea de distribució molt més reduïda que R. pusilla. Com a hipòtesi, afirmem que és possible que el seu sistema de reproducció expliqui aquesta diferència. R. lusitanica es reprodueix exclusivament de forma sexual, mitjançant la formació d’ascòspores, en canvi R. pusilla, a més de la reproducció sexual, es podria reproduir també asexualment per grups d’algues liquenificades que s’alliberarien a través de les fenestracions que presenta el taŀlus.
A partir dels resultats d’aquesta investigació hem proposat la inclusió de Ramalina lusitanica com a espècie en perill d’extinció al Catàleg de flora amenaçada de Catalunya (DOGC Número 9047-23.11.2023) per la seva distribució puntual i amb poblacions de molt pocs individus, considerant que d’aquesta forma també queden protegides R. pusilla, R.
lacera i R. canariensis, espècies que formen part de la mateixa comunitat.
Agraïments
Volem expressar el nostre agraïment a la Institució Catalana d’Història Natural per la concessió del premi TorrasFoulon de l’any 2022, fet que ha permès el finançament del present treball. Agraïm als gestors dels espais naturals protegits que ens hagin autoritzat fer les nostres prospeccions: Parc Natural del Cap de Creus, espais naturals protegits de la demarcació de Girona (Tossa de Mar i Palafrugell), Espai d’Interès Natural protegit Tamarit-Punta de la Móra-Costes del Tarragonès, Parc Natural del Delta de l’Ebre. També estem agraïts a la Generalitat de Catalunya per la cessió de dades meteorològiques de l’illa de Buda.
Bibliografia
Atienza, V., & Segarra-Moragues, J. G. 2006. Evaluation of the Conservation Status of Threatened Lichens List from Coastal areas of the Valencian Community (Eastern Spain). Cryptogamie Mycologie, 27, 167-183. Boqueras, M., & Gómez-Bolea, A. 1986. Líquens epífits, i els seus fongs paràsits, observats sobre Quercus suber, a Catalunya. Folia Botanica Miscellanea, 5: 49-69.
Carvalho, P., Figueira, R., Jones, M., Sérgio, S., & Sim-Sim, M. 2002. Biodiversidade da Vegetaão liquénica no litoral alentejano, área de Sines. Portugaliae Acta Biológica, 20: 225-248.
Cera, A., Mendes, J., Cortada, À., & Gómez-Bolea, A. 2022. Cover of Ramalina species as an indicator of habitat quality in threatened coastal woodlands. The Bryologist, 125: Article 3. https://doi. org/10.1639/0007-2745-125.3.424
GBIF Secretariat: GBIF Backbone Taxonomy. Checklist dataset https://doi.org/10.15468/39omei accessed via GBIF.org on 202409-09
Gómez-Bolea, A. 1985. Líquenes epífitos en Catalunya [PhD thesis]. Universitat de Barcelona. 259 p. Lange, O. L., Geiger, I. L., & Schulze, E.-D. 1977. Ecophysiological investigations on lichens of the Negev desert. Oecologia, 28: Article 3. https://doi.org/10.1007/BF00751603
Nascimbene, J., Nimis, P. L., & Ravera, S. 2013. Evaluating the conservation status of epiphytic lichens of Italy : A red list. Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology, 147: Article 4. https://doi.org/10.1080/112635 04.2012.748101
Roux, C. 2020. Catalogue des lichens et champignons lichénicoles de France métropolitaine (3nd Edition). Association française de lichénologie. 1769 p.
Sérgio, C., Carvalho, P., Garcia, C. A., Almeida, E., Novais, V., Sim-Sim, M., Jordão, H., & Sousa, A.J. 2016. Floristic changes of epiphytic flora in the Metropolitan Lisbon area between 1980–1981 and 2010–2011 related to urban air quality. Ecological Indicators, 67: 839-852. https://doi.org/10.1016/j. ecolind.2016.03.022
Butlletí de la Institució Catalana d’Història Natural, 88 (3): 145-147. 2024
Primeres gales de cinípids mexicans parasitades per Eupelmus pulchriceps (Cameron, 1904) (Hym., Eupelmidae)
First Mexican cynipid oak galls parasitized by Eupelmus pulchriceps (Cameron, 1904) (Hym., Eupelmidae)
Júlia
Cazorla-Vila* & Juli Pujade-Villar*
* Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals. Facultat de Biologia. Universitat de Barcelona. Avda. Diagonal, 643, 08028 Barcelona, Catalunya. A/e: juliacavi1503@gmail.com i jpujade@ub.edu
Rebut: 25.08.2024. Acceptat: 15.09.2024. Publicat: 30.09.2024
Els eupèlmids (Hymenoptera: Chalcidoidea: Eupelmidae) són una família de calcidoideus que comprèn ectoparasitoides de larves o prepupes d’insectes, generalment amagats en els teixits vegetals o en capolls, i també endoparasitoides o depredadors d’insectes o ous d’aranyes (Gibson 1993, 1997). Encara que es menciona que els hostes originals dels eupèlmids són coleòpters, la majoria de les espècies d’aquesta família ataquen dípters, himenòpters o lepidòpters (Gibson 1995).
A Amèrica es coneixen 4 espècies d’Eupelmus que emergeixen de gales de Cynipini en roures (Hymenoptera: Cynipidae: Cynipini), dues de les quals s’han trobat a Mèxic: Eupelmus cynipidis Ashmead, 1882 (citada recentment per Cazorla-Vila, 2023) y Eupelmus pulchriceps (Cameron, 1904).
Eupelmus pulchriceps és una espècie polífaga àmpliament distribuïda pel continent americà des del Canadà fins l’Argentina (Catalog of American Eupelmidae obtained from oak cynipid galls, in prep.). Es tracta d’un parasitoide primari o secundari que ataca 34 espècies diferents de 14 famílies pertanyents a 5 ordres d’insectes (Gibson, 2011).
Segons Forbes et al. (2016) i Gibson (2011), el parasitoidisme d’aquesta espècie en gales de cinípids només ha estat mencionat als Estats Units en gales de Belonocnema treatae Mayr, 1881 (forma asexuada) i de Dryocosmus kuriphilus Yasumatsu, 1951 (forma asexiada). A Mèxic, aquesta espècie ha estat citada per Pérez-Benavides et al. (2020) amb la denominació d’Eupelmus aff. pulchriceps com a parasitoide de Bruchinae (Coleoptera) i per Ruíz-Cancino et al. (2004) i González-Hernández (2004) sense especificar ni l’hoste ni la localitat.
En aquest estudi citem amb seguretat per primer cop E. pulchriceps en gales de cinípids de Mèxic i es donen els primers hostes en els quals l’hem trobat en aquest país (Fig. 1): Amphibolips hidalgoensis ǒ, Andricus sp. (tumoral) ǒ i una gala indeterminada coŀlectada a Quercus liebmanii citant-la per primer cop als estats de México, Morelos i Puebla.
Material estudiat: 4 ♀
MEX-278, Felipe Neri, Municipio de Felipe Neri (Morelos), Ex. Amphibolips hidalgoensis Pujade-Villar & Melika, 2011


2. Eupelmus pulchriceps (♀): a) habitus lateral; b) ala anterior; c) ovopositor; d) potes en visió ventral.
asex. sobre Q. rugosa, (29.v.2012) viii.2012: 1♀ (R.D. García-Martinón leg.). P-390, Las Cruces, San Luis Huexotla, Municipio de Texcoco (México), Ex. Andricus sp. (tumoral) asex. sobre Q. microphylla, (05.vii.2018) vi.2019: 2♀ (V. D. Cibrián-Tovar leg.). P-749, Jardín Botánico de Puebla (Puebla), Ex. gala asexuada indeterminada sobre Q. liebmanii, (03.vii.2023) 01-15.vii.2023: 1♀ (JP-V leg.).
Eupelmus pulchriceps es caracteritza per presentar una coloració fosca (Fig. 2a) amb reflexes verdosos, blavosos i de color de coure; antenes completament fosques, tot i que en alguns casos l’escap és taronja; ales (Fig. 2b) hialines presentant línia calva; ovopositor (Fig. 2c) llarg i presentant una
Butlletí ICHN 88 (3), 2024
coloració bandejada, amb la base de color marró fosc, el centre groguenc-blanquinós i l’àpex marró clar; segon parell de potes (Fig. 2d) amb dues fileres de dents de color negre que estan distribuïdes de manera simètrica; dents apicals de la tíbia (Fig. 2d) del segon parell de potes també negres.
Aquesta investigació va ser finançada per «PID2021128146NB-I00/MCIN/AEI/10.13039/501100011033/» i «FEDER una manera de hacer Europa».
Bibliografia
Cazorla-Vila, J., Martínez-Romero, A., Equihua-Martínez, A., Estrada-Venegas, E. G. & Pujade-Villar. 2023. Primer registro de Eupelmus cynipidis Ashmead, 1882 (Hymenoptera: Chalcidoidea: Eupelmidae) en México. Dugesiana, 30(2): 111-114. https://doi. org/10.32870/dugesiana.v30i2.7311.
Forbes, A. A., Hall, C. M., Lund, J. A., Hood, G. R., Izen, R., Egan, S. P. & Ott, J. R. 2016. Parasitoids, hyperparasitoids and inquilines associated with the sexual and asexual generations of the gall former, Belonocnema treatae (Hymenoptera: Cynipidae). Annals of the Entomological Society of America, 109(1): 49-63. https:// doi.org/10.1093/aesa/sav112.
Gibson, G. A. P. 1993. Chapter 16 Superfamilies Mymarommatoidea and Chalcidoidea. Pp. 570-634. In: Goulet H, Huber JT (Eds.) Hymenoptera of the world: an identification guide to families. Agriculture Canada, Publication 1894/E, Ottawa, Ontario, 668 p. Gibson, G. A. P. 1995. Parasitic wasps of the subfamily Eupelminae: classification and revision of world genera (Hymenoptera: Chalcidoidea: Eupelmidae). Memoirs on Entomology, Interna
tional 5: v+421 p.
Gibson, G. A. P., 1997. Chapter 11. Eupelmidae. In: Annotated keys to the genera of Nearctic Chalcidoidea (Hymenoptera). Pp: 430476. Gibson, G.A.P., Huber, J.T. & Woolley, J.B. (Edit.). National Research Council Research Press, Ottawa, Canada. 794 p. Gibson, G. A P. 2011. The species of Eupelmus (Eupelmus) Dalman and Eupelmus (Episolindelia) Girault (Hymenoptera: Eupelmidae) in North America north of Mexico. Zootaxa, 2951: 1-97. https://doi.org/10.11646/zootaxa.2951.1.1. González-Hernández, A. 2004. Actualización del catálogo de autoridades taxonómicas: Hymenoptera-Parasítica. Depto. de Zoología de Invertebrados. Facultad de Ciencias Biológicas, UANL. Base de datos SNIB-Conabio proyecto BE008, México. D.F. Ruíz Cancino, E., Coronado Blanco, J. M., Myartseva, S. N. & Luna Salas, J. F. 2004. Adenda a Chalcidoidea (Hymenoptera). In: Bousquets, J.L., J.J. Morrone, O.Y. Ordóñez & I.V. Fernández (Ed.). Biodiversidad, Taxonomía y Biogeografía de artrópodos de México: Hacia una síntesis de su conocimiento. Volumen IV. 725-734.
Butlletí de la Institució Catalana d’Història Natural, 88 (4): 149-151. 2024 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
Primer registro de Aedes vittatus (Bigot, 1861) y Aedes vexans (Meigen, 1830) (Diptera: Culicidae) en la provincia de Tarragona
First record of Aedes vittatus Bigot, 1861 and Aedes vexans (Meigen, 1830) (Diptera: Culicidae) in the province of Tarragona
Carlos Pradera* & Alberto Bernués-Bañeres*
* Anticimex 3D Sanidad Ambiental, SA. 08174 Sant Cugat del Vallès, Barcelona. A/e: carlos.pradera@anticimex.com.es, alberto.bernues@anticimex.com.es Autor de correspondencia: Carlos Pradera. A/e: carlos.pradera@anticimex.com.es
Rebut: 18.09.2024. Acceptat: 02.10.2024. Publicat: 30.12.2024
A pesar de que en Cataluña existe una tradición en el estudio y control de culícidos, se observa una falta de estudios de campo que den cuenta de la riqueza y distribución de las distintas especies presentes (Bueno-Marí et al., 2012). Un ejemplo es Aedes mariae (Sergent & Sergent, 1903) que se desarrolla en colecciones de agua salada en rocas costeras cuya ecología fue estudiada por Margalef (1949), pero de la que no había un conocimiento detallado de su distribución hasta recientemente (Pradera et al., 2023). Por tanto, se valoran los datos recabados de especies poco estudiadas y, especialmente, de aquellas que puedan tener un impacto sobre la salud de las personas como los que se reportan en esta nota.
Se reportan los primeros datos registrados sobre la presencia de Aedes vittatus (Bigot, 1861) y Aedes vexans (Meigen, 1830) para la provincia de Tarragona, especies que fueron detectadas de manera fortuita (Fig. 1). Fueron capturados adultos de ambas especies con la ayuda de un aspirador entomológico al posarse sobre personas para picar en la mañana del 15 de septiembre de 2024 (Fig. 2). Ocurrió durante el curso de una excursión en dos ubicaciones bien separadas en el término municipal de Mas de Barberans (Tarragona), al sur del parque natural dels Ports:
1. Barranco de Lledó (40.742200, 0.369800), 350 m.s.n.m. Ae. vexans: 9 hembras. Ae. vittatus: 2 hembras, 1 macho. Carlos Pradera leg.
2. Barranco de Lloret (40.7737, 0.3571), 290 m.s.n.m. Ae. vittatus: 5 hembras, 1 macho. Carlos Pradera leg. En Cataluña, ambas especies han sido citadas sólo en las provincias de Barcelona y Gerona (Bueno-Marí et al., 2012). De ambas, la que cuenta con menos registros en España es A. vittatus (Díez-Fernández et al., 2018); aunque se encuentra distribuida por el sur de Europa, Asia y, especialmente, en África donde juega un importante rol vectorial en la transmisión de arbovirus que causan brotes epidémicos de fiebre amarilla (Díez-Fernández et al., 2018). Es una especie mamofílica, estenógama y exofílica, que pica a personas de manera agresiva (Becker et al., 2020) cerca de sus biotopos larva-
rios, de los que no se aleja para alimentarse (Schaffner et al., 2001). En ambos puntos de detección, salieron las hembras de la vegetación para picar, aunque parece tener una actividad principalmente crepuscular (Becker et al., 2020). Parece que el foco de cría preferente son oquedades rocosas junto a ríos y sus márgenes (Bueno Marí & Jiménez Peydró, 2010), aunque también puede hacerlo en agujeros de árboles o recipientes artificiales (Becker et al., 2020) como contenedores y bidones. El 5 de junio de 2016, junto al río Ter en el término municipal de Cellera de Ter (Gerona), se encontraron adultos y larvas en charcos de rocas (41.9847, 2.5810).
Aedes vexans es una especie con más registros en España (Bueno-Marí et al., 2012) y una amplia distribución mundial: América del Norte, África del Norte, Europa, Asia y Australia (Wilkerson et al., 2021). Es una especie multivoltina ligada a cuerpos de agua temporales por la crecida de ríos y lagos o cultivos de inundación (Becker et al., 2020). Pica a mamíferos y a personas siendo una importante molestia. En el Barranco de Lledó salieron de la vegetación bastantes hembras de las que se capturó un pequeño número. Su presencia se ve favorecida por la existencia de granjas de animales en las cercanías, aunque la hembra puede desplazarse hasta 15 km desde su lugar de cría (Becker et al., 2020). Existen zonas donde está muy presente siendo una molestia importante para las personas, especialmente cuando se produce una eclosión sincrónica de huevos, situación vivida con un gran número de hembras la tarde del 8 de octubre de 2016 en Cruïlles (Gerona) junto al río Daró donde resulta común (41.9428, 3.0093). Estas características hacen que Ae. vexans sea considerado como uno de los vectores tipo de distintas enfermedades; detectándose ejemplares infectados de forma natural por arbovirus (Reinert, 1973) como la encefalomielitis equina, encefalitis de California (Sudia et al. 1971), virus Tahyna (Lundström, 1994), virus West Nile o incluso la mixomatosis, además de potencial transmisor de tularemia (Encinas Grandes, 1982; Schaffner et al. 2001). Entre los parásitos que es capaz de transmitir destaca Dirofilaria immitis (Lewandowski et al., 1980).
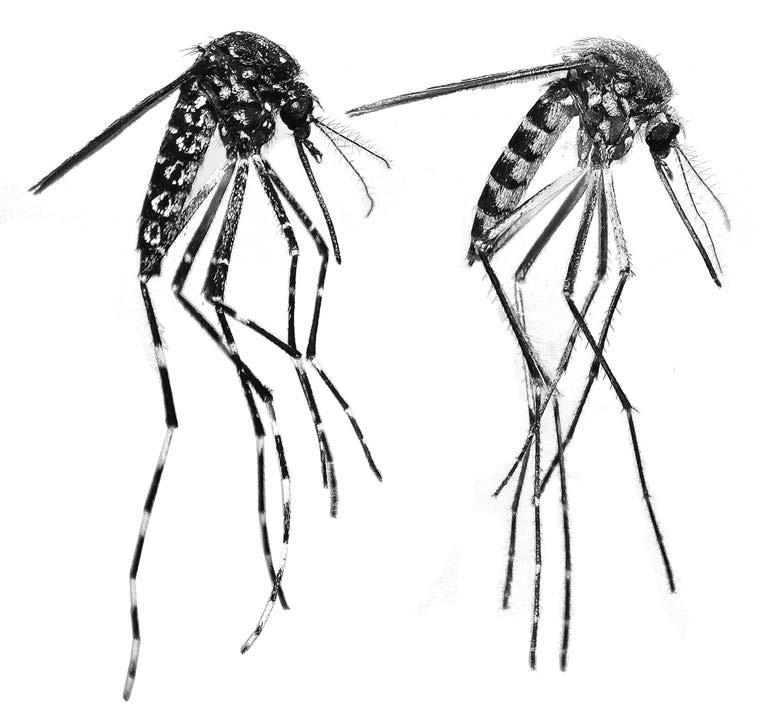

Butlletí ICHN 88 (4), 2024
La identificación de los ejemplares recolectados se realizó siguiendo los criterios taxonómicos de Becker et al. (2020). Los ejemplares se conservan en la colección particular de uno de los autores (C.P.).
Bibliografía
Becker, N., Petric, D., Zgomba, M., Boase, C., Madon, M. B., Dahl, C. & Kaiser, A. 2020. Mosquitoes and their control. Third Edition. Springer Heidelberg Dordrecht, London-New York. 570 pp. Bueno-Marí, R., Bernués-Bañeres, A. & Jiménez-Peydró, R. 2012. Updated checklist and distribution maps of mosquitoes (Diptera: Culicidae) of Spain. European Mosquito Bulletin, 30: 91-126. Bueno Marí, R., & Jiménez Peydró, R. 2010. Revisión y datos nuevos de Aedes vittatus (Bigot, 1861) para España (Diptera: Culicidae). Dugesiana, 17 (2): 143-144.
Díez-Fernández, A., Martínez-de la Puente, J., Ruiz, S., Gutiérrez-López, R., Soriguer, R. & Figuerola, J. 2018. Aedes vittatus in Spain: current distribution, barcoding characterization and potential role as a vector of human diseases. Parasites Vectors, 11: 297. Encinas Grandes, A., 1982. Taxonomía y biología de los mosquitos del área salmantina (Diptera, Culicidae). CSIC. Centro de edafología y Biología aplicada. Ed. Universidad de Salamanca, 437 p.
Lewandowski, Jr. H. B., Hooper, G. R. & Newson, H. D. 1980. Determination of some important natural potential vectors of dog heartworm in central Michigan. Mosquito News, 40: 73-79. Lundström, J. O. 1994. Vector competence of western European mosquitoes for arboviruses: A review of field and experimental studies. Bulletin of the Society for Vector Ecology, 19(1): 43-48. Margalef, R. 1949. Sobre la ecología de las larvas del mosquito Aëdes mariae. Publicaciones del Instituto de Biología Aplicada, 6: 83-101.
Pradera, C., Bengoa Paulis, M. & Ollé, A. 2023. Distribución de focos larvarios de Aedes mariae (Diptera: Culicidae) en Cataluña y su gestión. Butlletí de la Institució Catalana d’Història Natural, 87 (4): 239-244.
Reinert, J. F. 1973. Contributions to the mosquito fauna of Southeast Asia-XVI. Genus Aedes Meigen, subgenus Aedimorphus Theobald in Southeast Asia. Contributions of the American Entomological Institute, 9 (5): 1-218.
Schaffner, F., Angel, G., Geoffroy, B., Hervy, J. O. & Rhaeim, A. 2001 The mosquitoes of Europe / Les moustiques d’Europe. IRD Éditions and EID Méditerranée
Sudia, W. D., Newhouse, W. F., Calisher, C. H. & Chamberlain, R. W. 1971. California group arboviruses: isolations from mosquitoes in North America. Mosquito News, 31 (4): 576-600.
Wilkerson, R. C., Linton Y.-M. & Strickman, D. 2021. Mosquitoes of the World. Volume 1. Johns Hopkins University Press. 600 p.
Butlletí de la Institució Catalana d’Història Natural, 88 (4): 153-155. 2024 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
Stricticimex Ferris & Usinger, 1957 (Heteroptera: Cimicidae): primer registro de este género en Europa
Stricticimex Ferris & Usinger, 1957 (Heteroptera: Cimicidae): first record of this genus in Europe
Carlos Pradera*, Antonio Jesús Reina-Muñoz**, Juan Quetglas*** & Amador Barambio-Zarco****
* Av. Ramon Berenguer IV, 52. 08924, Santa Coloma de Gramenet, Barcelona. A/e: desinsectador@yahoo.es ** Av. de la Estación, 97. 18230, Atarfe, Granada. A/e: zujairas@hotmail.com *** Urb. El Señorío, 25. 41807, Espartinas, Sevilla. A/e: jqmurcielagos@gmail.com **** Av. General Avilés, 50. 46015, Valencia. A/e: amadorbz@gmail.com
Autor de correspondencia: Carlos Pradera. A/e: desinsectador@yahoo.es
Rebut: 03.10.2024. Acceptat: 14.10.2024. Publicat: 30.12.2024
El 7 de agosto de 2024 fue realizado un tratamiento rutinario contra chinches de cama, Cimex lectularius Linnaeus, 1758, en un apartamento turístico de la ciudad de Granada. La propiedad recibió la queja de un huésped que vio una chinche de cama sobre la almohada antes de acostarse de madrugada Esto motivó la solicitud de tratamiento que se llevó a cabo dos días después. Antes del tratamiento, se observaron manchas de sangre en las sábanas y varios ejemplares sobre paredes cerca de la ventana No se detectó ningún foco cerca de la cama. El aspecto de los ejemplares, diferente al de C. lectularius y apreciable en el tamaño de la cabeza y del pronoto (Fig. 1), motivó a recoger una muestra (Antonio Jesús Reina-Muñoz leg ). Fue realizado un tratamiento químico mediante la pulverización de
un insecticida emulsionable (DIPTRON con etofenprox 10 % y butóxido de piperonilo 20 %). La eliminación quedó resuelta con una única aplicación y no hubo reclamación. En la muestra había dos hembras, una incompleta a la que faltan cabeza y cuatro patas y la otra entera (Fig. 2). Siguiendo las claves de Usinger (1966), se han identificado las dos hembras como pertenecientes al género Stricticimex Ferris & Usinger, 1957 de la subfamilia Cacodminae Kirkaldy, 1899. Los cimícidos de esta subfamilia se caracterizan porque el seno paragenital, solo presente en las hembras, se encuentra en el dorso que, en el caso concreto de Stricticimex, está en el margen posterior del terguito III cerca del margen derecho del abdomen (Usinger, 1966).

1. Hembra del género Stricticimex de Granada (izquierda) y hembra de Cimex lectularius capturada en Ourense (derecha) en marzo de 2024, sobre papel milimetrado. Stricticimex se diferencia a simple vista por tener la cabeza más grande en relación al cuerpo y tener el pronoto y los rudimentos alares más cortos.

Se conocen nueve especies del género Stricticimex, descritas en África y Asia como parásitas de murciélagos. En la monografía de Usinger (1966) se recogen seis especies: S. pattoni (Horváth, 1925) en India, S. transversus Ferris & Usinger, 1957, S. antennatus Ferris & Usinger, 1957 en Sudáfrica, S. namru Usinger, 1960 en Egipto, S. intermedius Ferris & Usinger, 1959 en Kenia y S. brevispinosus Usinger, 1959 en Burundi. Ueshima (1968) describió S. parvus de Tailandia, taxón con el cual Klein (1970) sinonimizó S. khmerensis Klein, 1969, descrito de Camboya. Fain & Elsen (1972) describieron S. puylaerti de Camerún y Fain (1972) describió S. anciauxi de Zaire.
Como especies de importancia médica destacan S. antennatus y S. parvus, ambas reportadas picando activa y agresivamente personas en cuevas en Sudáfrica y Tailandia, respectivamente (Overal & Wingate, 1976; Hamlili et al., 2023). Stricticimex parvus es posible vector en la transmisión del virus Kaeng Khoi a humanos (Williams et al., 1976). A nivel geográfico, la especie más cercana a la península ibérica es S. namru, descrita de Egipto, pero también presente en Irán e India (Ghahari et al., 2016).
Con los datos recabados e identificado el género, se informa a la propiedad del apartamento, la cual nos comenta que hace tres años fueron detectados murciélagos en un cajón de persiana. No se ha podido realizar ninguna acción posterior ni comprobación. La detección de este cimícido genera preguntas para las que no tenemos respuesta. Creemos que podría estar asociado a la presencia de murciélagos. En las ciudades de Andalucía las especies comunes urbanas son del género Pipistrellus Kaup, 1829, principalmente Pipistrellus pygmaeus (Leach, 1825), y Eptesicus isabellinus (Temminck, 1839), pero en Granada ciudad se conocen colonias de Tadarida teniotis (Rafinesque, 1814) e Hypsugo savii (Bonaparte, 1837) (Grupo de Murciélagos de la Estación Bioló-
gica de Doñana, J. Quetglas, datos inéditos). Los cimícidos detectados en el sur peninsular son Cacodmus vicinus Horváth, 1934 en refugios de Pipistrellus (Quetglas et al., 2012; J. Quetglas, datos inéditos) y Cimex pipistrelli Jenyns, 1839 asociado a Nyctalus lasiopterus (Schreber, 1780) (Balvín et al., 2012; J. Quetglas, datos inéditos). En nuestro continente, los cimícidos asociados a murciélagos son cuatro: C. lectularius, C. pipistrelli, Cimex emarginatus Simov, Ivanova & Schunger, 2006 y Cacodmus vicinus (Balvín et al., 2014). Este sería el primer caso de Stricticimex fuera de cuevas en una vivienda y en Europa, donde no consta su presencia.
A Marta Goula, Jan Tomàs y Jesús Tanco por la revisión y mejoras aportadas al texto.
Balvín, O., Bartonička, T., Simov, N., Paunović, M. & Vilímová, J. 2014. Distribution and host relations of species of the genus Cimex on bats in Europe. Folia Zoologica, 63 (4): 281-289.
Balvín, O., Ševčík, M., Jahelková, H., Bartonička, T., Orlova, M. & Vilímová, J. 2012. Transport of bugs of the genus Cimex (Heteroptera: Cimicidae) by bats in western Palaearctic. Vespertilio, 16: 43-54.
Fain, A. 1972. Notes sur les Punaises parasites de Chiroptères de la République du Zaïre avec description de deux espèces et d’une sous-espèce nouvelle. Revue de Zoologie et de Botanique Africaines, 85 (3-4): 187-202.
Fain, A. & Elsen, P. 1972. Un nouveau Cimicide du Cameroun: Stricticimex puylaerti sp. n. Revue de Zoologie et de Botanique Africaines, 85 (1-2): 142-146.
Ghahari, H., Moulet, P. & Ostovan, H. 2016. An annotated catalog of the Iranian Cimicidae and Largidae (Hemiptera: Heteroptera) and in memoriam Carl Walter Schaefer (1934–2015). Zootaxa, 4111 (2): 194-200.
Hamlili, F.Z., Bérenger, J.M. & Parola, P. 2023. Cimicids of Medical and Veterinary Importance. Insects, 14(4): 392.
Klein, J. M. 1969. Nouvelles Punaises du Cambodge: Crassicimex apsarae n. sp. et Stricticimex khmerensis n. sp. [HEM. CIMICIDAE]. Bulletin de la Société entomologique de France, 74 (3-4): 87-96.
Klein, J. M. 1970. Cimicides du Cambodge. (III). Description des mâles de Crassicimex apsarae, Stricticimex parvus et Aphrania thnotae n. sp. [Hemiptera, Cimicidae]. Annales de la Société entomologique de France (N.S.), 6 (3): 713-719.
Overal, W. L. & Wingate, L. R. 1976. The biology of the Batbug Stricticimex antennatus (Hemiptera: Cimicidae) in South Africa. Annals of the Natal Museum, 22 (3): 821-828.
Quetglas, J., Balvín, O., Lucan, R. K. & Benda, P. 2012. First records of the bat bug Cacodmus vicinus (Heteroptera: Cimicidae) from Europe and further data on its distribution. Vespertilio, 16: 243-248.
Ueshima, N. 1968. New Species and Records of Cimicidae with Keys (Hemiptera). The Pan-Pacific Entomologist, 44(4): 264-279.
Usinger, R. L. 1966. Monograph of Cimicidae (HemipteraHeteroptera). Entomological Society of America. Maryland. 582 p. Williams, J. E., Imlarp, S., Top, F. H., Cavanaugh, D. C. & Russell, P. K. 1976. Kaeng Khoi virus from naturally infected bedbugs (Cimicidae) and immature free-tailed bats. Bulletin of the World Health Organization, 53 (4): 365-369.
Butlletí de la Institució
* Anticimex 3D Sanidad Ambiental SA.08174 Sant Cugat del Vallès, Barcelona. A/e: carlos.pradera@anticimex.com.es
** Consultor Externo. Ministerio de Sanidad. Madrid. A/e: antricola@me.com
Autor de correspondencia: Carlos Pradera: A/e: carlos.pradera@anticimex.com.es
Rebut: 08.10.2024; Acceptat: 17.10.2024; Publicat: 30.12.2024
Resumen
Se aportan los resultados de diversos muestreos realizados entre 2023 y 2024 para la detección de la garrapata Hyalomma lusitanicum en Cataluña. Se ha detectado en 70 nuevos municipios de las provincias de Barcelona y Tarragona que, sumados a los 31 en los que se había detectado en 2022, ascienden a 101. También se aportan dos registros de Hyalomma marginatum en la provincia de Gerona.
Palabras clave: garrapata, conejo, jabalí, corzo, vector, salud pública.
Abstract
New data on the distribution of the tick Hyalomma lusitanicum (Acari, Ixodidae) in Catalonia
The results of various samplings carried out between 2023 and 2024 for the detection of the Hyalomma lusitanicum tick in Catalonia are provided. It has been detected in 70 new municipalities in the provinces of Barcelona and Tarragona, which, added to the 31 in which it was detected in 2022, amount to 101. Two records of Hyalomma marginatum in the province of Gerona are also provided.
Key words: tick, rabbit, wild boar, roe deer, vector, public health.
Abstract
Noves dades sobre la distribució de la paparra Hyalomma lusitanicum (Acari, Ixodidae) a Catalunya
S’aporten els resultats de diversos mostrejos realitzats entre el 2023 i el 2024 per a la detecció de la paparra Hyalomma lusitanicum a Catalunya. S’ha detectat a 70 nous municipis de les províncies de Barcelona i Tarragona que, sumats als 31 en què s’havia detectat el 2022, ascendeixen a 101. També s’aporten dos registres de Hyalomma marginatum a la província de Girona.
Paraules clau: paparra, conill, senglar, cabirol, vector, salut pública.
Introducción
Un estudio del año 2022 daba cuenta de la presencia de la garrapata Hyalomma lusitanicum Koch, 1844, en 31 municipios de Cataluña, a la par que se mencionaban sus implicaciones potenciales para la salud pública (Pradera & Estrada-Peña, 2022) (Fig. 1). El muestreo se llevó a cabo en la zona periurbana de municipios del área de Barcelona, la cual está muy urbanizada y cuyo medio natural aparece fragmentado, permitiendo que ungulados como el jabalí, Sus scrofa Linnaeus, 1758, proliferen (Castillo-Contreras et al., 2018). El muestreo también se concentró en zonas alrededor de núcleos urbanos, los cuales se presentan alterados y degradados por la actividad humana, y se convierten en espacios en los que prolifera el conejo común, Oryctolagus cuniculus (Linnaeus, 1758). Ambas especies forman un buen binomio de hospedadores para
10.2436/20.1502.01.189
la proliferación de H. lusitanicum (González et al., 2016). Las larvas y ninfas se alimentan sobre conejo y los adultos sobre ungulados domésticos y silvestres (Valcárcel et al., 2016).
En la actualidad, la zona de estudio tiene un alto riesgo para la salud pública, debido a la gran proliferación de H. lusitanicum alrededor de núcleos urbanos (Fig. 2). Hasta donde conocemos, esta problemática no se había detectado en el área de Barcelona hasta fechas recientes (Pradera & Estrada-Peña, 2022), siendo solamente objeto de los planes de control de plagas urbanas la garrapata Rhipicephalus sanguineus (Latreille, 1806). Dada la implicación de H. lusitanicum como vector de microorganismos patógenos que pueden afectar a la salud pública (Valcárcel et al., 2020), y con la voluntad de ahondar en el conocimiento de su distribución en Cataluña, se han continuado los muestreos durante los años 2023 y 2024.


Material y métodos
Se ha llevado a cabo una prospección de H. lusitanicum de carácter documental (sin pretender estimar la densidad) en 2023 (de abril a julio) y 2024 (de abril a septiembre) en municipios de las provincias Barcelona y Tarragona, y puntualmente en Gerona, guiados por los resultados de los muestreos llevados a cabo en 2022 (Pradera & Estrada-Peña, 2022). La prospección se planificó seleccionando espacios de entornos urbanos degradados y con poca vegetación en los que había presencia de conejo común. Se ha buscado en caminos por los que había indicios de jabalí o eran idóneos para su paso. Este método está enfocado a la detección de H. lusitanicum, de la que es conocida la preferencia por zonas con las caracte-
rísticas mencionadas. La captura de ejemplares ha sido principalmente directa por avistamiento de garrapatas subidas a las plantas, moviéndose sobre el suelo o atraídas por la exhalación de la respiración del colector en márgenes de caminos, método sencillo y práctico para la captura de garrapatas «cazadoras» como Hyalomma Koch, 1844 (Cuadrado-Matías et al., 2024). En menor medida, también se ha utilizado el método por arrastre (bandera). Los ejemplares recolectados se identificaron con las claves aportadas por Estrada-Peña et al. (2004) y se conservaron en etanol de 70º.
Se han capturado 1.274 individuos adultos de H. lusitanicum en 81 municipios, 55 de la provincia de Barcelona y 26 municipios de Tarragona (Tabla 1). En 11 de estos municipios de la provincia de Barcelona ya se habían recolectado adultos de H. lusitanicum en los muestreos de 2022, en los que se encontró en 31 municipios, 30 de la provincia de Barcelona y 1 de Tarragona (Pradera & Estrada-Peña, 2022). Por tanto, tenemos registros de H. lusitanicum en 101 municipios de Cataluña (Fig. 3). En los muestreos también han sido capturados 118 adultos de otras especies como Dermacentor marginatus (Sulzer, 1776), Hyalomma marginatum Koch, 1844, Ixodes ventalloi Gil Collado, 1936, Rhipicephalus pusillus Gil Collado, 1936 y R. sanguineus (Tabla 2). Destacamos dos capturas de H. marginatum en Gerona por sus implicaciones para la salud pública (Fig. 1) (Valcárcel et al., 2020).
La continuación de los muestreos iniciados en 2022 en las provincias de Barcelona y Tarragona nos ha permitido per-
Provincia Municipio Año Mes Día Coordenadas Ind.
Barcelona Abrera 2024 7 26 41.498, 1.926 9
Barcelona Argentona 2024 5 11 41.556, 2.407 3
Barcelona Avinyonet del Penedès 2024 8 6 41.361, 1.786 1
Barcelona Badalona 2023 6 29 41.463, 2.233 100
Barcelona Badalona 2024 6 7 41.555, 2.282 33
Barcelona Badia del Vallès 2023 7 18 41.515, 2.115 4
Barcelona Barberà del Vallès 2024 6 7 41.512, 2.118 3
Barcelona Begues 2023 4 2 41.289, 1.912 12
Barcelona Caldes de Montbui 2023 7 13 41.623, 2.165 3
Barcelona Caldes de Montbui 2024 6 10 41.613, 2.174 2
Barcelona Canovelles 2023 7 7 41.622, 2.286 3
Barcelona Castellar del Vallès 2024 8 3 41.584, 2.084 2
Barcelona Castellbell i el Vilar 2024 6 24 41.631, 1.856 5
Barcelona Castellet i la Gornal 2024 7 28 41.253, 1.589 19
Barcelona Castellví de la Marca 2024 7 28 41.328, 1.615 29
Barcelona Cubelles 2024 7 14 41.220, 1.680 5
Barcelona Esparreguera 2024 7 26 41.536, 1.878 3
Barcelona Franqueses del Vallès (Les) 2023 7 7 41.643, 2.293 40
Barcelona Gavà 2024 8 9 41.298, 1.998 11
Barcelona Gavà 2024 8 9 41.290, 1.988 4
Barcelona Lliçà d'amunt 2023 7 11 41.602, 2.245 2
Barcelona Martorell 2024 8 24 41.499, 1.915 6
Barcelona Matadepera 2024 8 10 41.594, 2.029 1
Barcelona Mataró 2024 5 4 41.560, 2.426 7
Barcelona Montmeló 2023 7 10 41.546, 2.251 1
Barcelona Montornès del Vallès 2024 6 6 41.554, 2.278 6
Barcelona Olèrdola 2024 8 6 41.320, 1.709 15
Barcelona Pacs del Penedès 2024 8 4 41.346, 1.662 1
Barcelona Palau-Solità i Plegamans 2023 6 27 41.562, 2.173 4
Barcelona Papiol (El) 2023 7 7 41.439, 2.006 50
Barcelona Polinyà del Vallès 2023 6 27 41.558, 2.165 60
Barcelona Polinyà del Vallès 2023 6 27 41.571, 2.149 25
Barcelona Polinyà del Vallès 2023 10 19 41.559, 2.151 10
Barcelona Premià de Dalt 2024 5 10 41.494, 2.338 4
Barcelona Ripollet 2024 6 14 41.509, 2.150 34
Barcelona Roca del Vallès (La) 2024 7 30 41.577, 2.314 21
Barcelona Rubí 2023 4 6 41.490, 2.014 7
Barcelona Rubí 2023 7 7 41.489, 2.013 50
Barcelona Sabadell 2023 7 7 41.515, 2.094 50
Barcelona Sant Adrià del Besòs 2024 6 23 41.432, 2.212 3
Barcelona Sant Boi de Llobregat 2023 7 7 41.350, 2.044 1
Barcelona Sant Boi de Llobregat 2023 7 20 41.343, 2.034 1
Barcelona Sant Feliu de Llobregat 2023 7 7 41.371, 2.036 1
Barcelona Sant Martí Sarroca 2024 7 28 41.337, 1.659 32
Barcelona Sant Pere de Ribes 2024 7 14 41.254, 1.768 10
Barcelona Sant Quirze del Vallès 2023 7 7 41.521, 2.033 50
Barcelona Sant Sadurní d'Anoia 2024 8 4 41.421, 1.788 1
filar la distribución de H. lusitanicum. Pero también nos ha permitido de alguna manera observar su expansión. En el estudio exponemos los datos de su presencia, pero no los de su ausencia por la dificultad de afirmar con seguridad que no se encuentra en un lugar. Sin embargo, en varias ocasiones hemos encontrado H. lusitanicum en lugares donde previamente no se había hallado, quizás por su carácter expansivo, o quizás por defectos en la planificación del muestreo (bien fuera de la época de actividad o sin una comprensión completa de sus necesidades abióticas). Un ejemplo es el municipio de Mataró, en el que en julio de 2022 se muestreó en varios puntos con presencia de conejo junto al Cementerio Municipal Torrent de les Valls sin encontrarse la garrapata. En mayo de 2024, se regresó a la zona y se encontraron 5
Provincia Municipio Año Mes Día Coordenadas Ind.
Barcelona Sant Vicenç de Castellet 2024 6 24 41.675, 1.863 8
Barcelona Santa Coloma de Cervelló 2024 8 9 41.366, 2.027 3
Barcelona Santa Coloma de Gramenet 2024 5 1 41.465, 2.211 46
Barcelona Santa Margarida i els Monjos 2024 7 28 41.329, 1.660 1
Barcelona Santa Maria de Martorelles 2024 6 9 41.522, 2.254 3
Barcelona Santa Perpètua de la Mogoda 2023 7 14 41.528, 2.200 50
Barcelona Sentmenat 2024 6 10 41.613, 2.148 8
Barcelona
Ulldecona 2024 5 14 40.598, 0.431 7
Tarragona Vendrell (El) 2023 6 28 41.188, 1.549 100
Tarragona Vendrell (El) 2024 7 2 41.182, 1.505 46
adultos (Tabla 1). Otro ejemplo es el margen derecho del río Besós a la altura de Sant Adrià del Besós, en el que existe una población elevada de conejos que los hace visibles durante el día. En 2022 no se encontró H. lusitanicum, pero en junio de 2024 se capturaron 6 adultos (Tabla 1). Podríamos poner otros ejemplos que dan cuenta, a nuestro parecer, de la expansión de esta garrapata.
Creemos que en lugares del área de Barcelona con abundancia de conejo en los que la garrapata no se ha encontrado, se detectará en los próximos años. Hyalomma lusitanicum está bien adaptada a la vegetación mediterránea (EstradaPeña et al., 2004) y las hembras son capaces de realizar una puesta de varios miles de huevos, asegurando un alto número de ejemplares en la generación siguiente (Valcárcel et al.,
D.
R. pusillus
R. pusillus
R.
Figura 3. Municipios donde se ha encontrado Hyalomma lusitanicum en Cataluña: en rojo los 31 municipios del estudio 2022 y en naranja los 70 del actual estudio de 2023-2024.
2020). La zona muestreada cuenta con abundantes ungulados silvestres que recorren ampliamente el territorio y pueden dispersar la garrapata mientras se alimenta sobre ellos (CastilloContreras et al., 2018). Si la superpoblación de jabalíes es un problema Cataluña, es preciso añadir la expansión del corzo, Capreolus capreolus (Linnaeus, 1758) (Griffiths et al., 2016). Es una especie históricamente extinguida en Cataluña, pero reintroducida a partir de la década de 1970 y en expansión (Montané et al., 2001). Junto al jabalí, el corzo está cada vez más presente y cercano a poblaciones, lo cual se observa en el censo de capturas de 2022: 70.736 registros de jabalíes y 11.804 de corzos (IDESCAT, 2024). Dadas las necesidades de H. lusitanicum como parásito estricto y la ingesta de sangre de sus hospedadores, su abundancia parece venir condicionada por la pobre o nula gestión de conejos y ungulados silvestres en Cataluña. Si la tendencia del clima favorece el asentamiento de algunas especies exóticas, la ausencia de una estrategia de gestión de sus hospedadores eleva considerablemente su abundancia y los riesgos para la salud pública. Para dotarlas de un adecuado marco de interpretación, debemos dejar anotadas algunas de las situaciones de las que hemos sido testigos o se nos han informado acerca de la presencia/abundancia de H. lusitanicum. Tenemos constancia de la retirada de conejos durante el año 2023 en dos municipios cercanos a Barcelona como medida para reducir la población de garrapatas. También en otros dos ayuntamientos se han llevado a cabo tratamientos mediante aplicación de acaricidas en zonas con alta densidad. Se ha visto eficaz la pulverización de piretroides (ECOREX CIPER EW, cipermetrina 40/60 al 10.80%) contra H. lusitanicum realizada de manera dirigida en franjas cerca de caminos o bien alrededor de instalaciones municipales (para tratamientos en superficies mayores y en madrigueras remitimos a González et al., 2018). En este problema de sobrepoblación de garrapatas que pueden afectar seriamente la salud pública, hemos notado que los centros de educación primaria y secundaria son especialmente sensibles cuando han sido situados en zonas periurbanas rodeados de campos, en las que proliferan los hospedadores de la garrapata, y en las que sus poblaciones pueden ser altas. No es necesario indicar que la población humana en dichos centros es extraordinariamente frágil frente a los vectores transmitidos por garrapatas. Otra casuística son las instalaciones deportivas, viviendas e incluso cementerios en zona periurbana donde la presencia cercana de la garrapata durante la primavera ha representado quejas por parte de la ciudadanía. También destacamos el caso de parques urbanos donde prolifera el conejo y la aparición de garrapatas ha alterado su normal funcionamiento.
Dos años después de haber mostrado el problema creciente de H. lusitanicum en Cataluña, no hemos visto avance en las autoridades de la salud pública. Se trata de una garrapata que puede ser responsable de la circulación, amplificación y transmisión de determinados patógenos (para los que aún no existen datos representativos) con el consiguiente problema, actualmente ignorado, para la salud pública. En su momento apuntábamos que se debería iniciar un programa de monitorización para conocer la distribución de esta y otras garrapatas, al menos a nivel regional; aunque a nivel estatal sí se ha ini-
ciado su estudio mediante el Proyecto GARES del Ministerio de Sanidad, la escala de este proyecto impide que se muestreen en profundidad las diferentes zonas de interés (o riesgo para los humanos) en una región.
A Eduard Marqués, Carme Freire, Eduard Durany, Marta Reig, Julen Zuberogoitia, Dídac González y, especialmente, a Vanessa Ramírez por la ayuda en la recolección. Y también a Félix Valcárcel por la revisión y mejora del texto.
Castillo-Contreras, R., Carvalho, J., Serrano, E., Mentaberre, G., Fernández-Aguilar, X., Colom, A. & López-Olvera, J. R. 2018. Urban wild boars prefer fragmented areas with food resources near natural corridors. Science of the Total Environment, 615: 282-288.
Cuadrado-Matías, R., Casades-Martí, L., Peralbo-Moreno, A., BazFlores, S., García-Manzanilla, E. & Ruiz-Fons, F. 2024. Testing the efficiency of capture methods for questing Hyalomma lusitanicum (Acari: Ixodidae), a vector of Crimean-Congo hemorrhagic fever virus. Journal of Medical Entomology, 61 (1): 152-165. Estrada-Peña, A., Bouattour, A., Camicas, J. L. & Walker, A. R. 2004. Ticks of Domestic Animals in the Mediterranean Region A guide to identification of species. Universidad de Zaragoza. 131 p. Montané, J., Marco Sánchez, I., López, L., Manteca Vilanova, X., & Lavín González, S. 2001. Captura y manejo postcaptura del corzo (Capreolus capreolus). Medicina Veterinaria, 18 (2): 341-351. Pradera, C. & Estrada-Peña, A. 2022. Hyalomma lusitanicum (Acari: Ixodidae) como potencial problema de salud pública en el área de Barcelona. Butlletí de la Institució Catalana d’Història Natural, 86 (3): 111-116.
González, J., Valcárcel, F., Pérez-Sánchez, J. L., Tercero-Jaime, J. M. & Olmeda, A. S. 2016. Seasonal dynamics of ixodid ticks on wild rabbits Oryctolagus cuniculus (Leporidae) from Central Spain. Experimental and Applied Acarology, 70 (3): 369-380. González, J., Valcárcel, F., Tercero, J. M., Cutuli, M. T., Sánchez, M., González, M.G. & Olmeda, A.S. 2018. Ejemplo práctico del control alternativo de garrapatas en una explotación extensiva. Albéitar, 217: 6-10.
Griffiths Romano, C., Laura Mingorance Perez, L., Rubio Asensi, C. & Valls Maurel, O. 2016. Estudi de la població de cabirol (Capreolus capreolus) a la vall d’Alinyà i anàlisi dels conflictes socioeconòmics originats per la reintroducció de l’espècie a Catalunya. TFG. Universitat Autònoma de Barcelona. 169 p. Institut d’Estadística de Catalunya (IDESCAT). 2024. Caça. Captures. Per espècies. Províncies. En línea: <https://www.idescat.cat/i ndicadors/?id=aec&n=15201&lang=CAT> [Consulta: octubre de 2024].
Valcárcel, F., González, J., Pérez Sánchez, J. L., Tercero, J. M. & Olmeda, A.S. 2016. Long-Term Ecological Study of Host-Seeking Adults of Hyalomma lusitanicum (Acari: Ixodidae) in a Meso-Mediterranean Climate. Journal of Medical Entomology, 53 (1): 221-4.
Valcárcel, F., González, J., González, M. G., Sánchez, M., Tercero, J. M, Elhachimi, L., Carbonell, J. D. & Olmeda, A. S. 2020. Comparative Ecology of Hyalomma lusitanicum and Hyalomma marginatum Koch, 1844 (Acarina: Ixodidae). Insects, 11 (5): 303.
Moitessieria canyellesensis sp. nov. (Gastropoda: Moitessieriidae), una nova espècie estigòbia de Castellar del Vallès (Barcelona, Catalunya)
Vicenç Bros*, **, Jordi Cadevall*, **, Glòria Guillén*, **, Albert Orozco*, **, Miquel Capdevila*, ** & Jordi Corbella*, **
* Associació Catalana de Malacologia (ACM). Museu Blau., Plaça Leonardo da Vinci 4-5. 08019 Barcelona, Spain. ** Museu de Ciències Naturals de Barcelona, Laboratori de Natura. Passeig Picasso s/n. 08003 Barcelona, Spain.
Autor de correspondència. Vicenç Bros Caton. A/e: vbrosc@gmail.com
Rebut: 06.08.2024; Acceptat: 18.10.2024; Publicat: 30.12.2024
Resum
Es descriu un nou moitessièrid (Gastropoda: Moitessieriidae) present a tres fonts situades al municipi de Castellar del Vallès, a partir de material conquiliològic obtingut mitjançant el triatge de sediments: Moitessieria canyellesensis sp. nov. Es detallen les característiques conquiliològiques que el diferencien de les restants espècies del gènere Moitessieria Bourguignat, 1864, i en particular de les més properes geogràficament i conquiliològica: Moitessieria olleri Altimira, 1960 i Moitessieria punctata Alba et al., 2010.
Paraules clau: Mollusca, Truncatelloidea, diversitat subterrània, fauna estigòbia, península Ibèrica.
Abstract
Moitessieria canyellesensis sp. nov. (Gastropoda: Moitessieriidae), a new stygobie species from Castellar del Vallès (Barcelona, Catalonia)
A new moitessièrid (Mollusca: Gastropoda: Truncatelloidea) present in three springs located in the municipality of Castellar del Vallès is described, based on conchological material obtained through sediment screening: Moitessieria canyellesensis sp. nov. The conchiliological characteristics that differentiate it from the remaining species of the genus Moitessieria Bourguignat, 1864 are detailed, and in particular the closest geographically and conchologically: Moitessieria olleri Altimira, 1960 and Moitessieria punctata Alba et al., 2010.
Key words: Mollusca, Truncatelloidea, subterranean diversity, stygobiont fauna, Iberian Peninsula.
Introducció
La família Moitessieriidae Bourguignat, 1864, està composta per microgastròpodes estigobis i s’inclou en la superfamília Truncatelloidea Gris, 1840 (MolluscaBase, 2021). Els estudis filogenètics de Falniowski et al. (2019) van mostrar Moitessieriidae com un clade diferent i ben recolzat molecularment, amb la qual cosa van confirmar la seva monofília. Aquests estudis, però, no incloïen els gèneres Palaospeum Boeters, 1999, Baldufa Alba et al., 2010 i Tarracospeum Quiñonero-Salgado et al., 2021, presents a la península Ibèrica i que, a manca de més estudis filogenètics moleculars que els incloguin, en aquest treball es consideren Moitessieriidae seguint els criteris morfològics dels seus autors respectius. Pel que fa al gènere Spiralix Boeters, 1972, també present a la península Ibèrica, Ritchling et al. (2016) ja havien considerat que la Moitessieria cf. puteana Coutagne, 1883 de diversos treballs anteriors (e.g. Wilke et al., 2001, Wilke et al.,
2013) s’havia d’anomenar Spiralix puteana (Coutagne, 1883), seguint Boeters & Falkner, 2009. Per tant, el gènere Spiralix Boeters, 1972 ja estava implícitament inclòs en la monofília molecular proposada per Falkniowski et al. (2019).
De fet, tal i com exposa Wilke (2019), la fixació genètica de les diferències conquiliològiques dins Moitessieriidae sovint no està prou clarificada. La família està mancada encara de força informació molecular.
A la península Ibèrica s’hi han descrit i/o citat els següents gèneres inclosos en la família Moitessieriidae: Moitessieria Bourguignat, 1864; Baldufa Alba et al., 2010; Palaospeum Boeters, 1999; Sardopaladilhia Manganelli et al., 1998; Spiralix Boeters, 1972 i Tarracospeum Quiñonero-Salgado et al., 2021. En el seu conjunt representen una fracció molt important de la biodiversitat dels medis subterranis i un patrimoni biològic d’alt valor, que cal conservar (Glöer, 2022).
El gènere Moitessieria Bourguignat, 1864 inclou fins ara 43 espècies acceptades. Es distribueixen pel nord-est de
V. BROS ET AL.
la península Ibèrica, el sud de França, el nord-oest d’Itàlia i l’illa de Sardenya (Bodon & Giusti, 1991; Corbella et al., 2020; Glöer, 2022 i Prié et al., 2024). D’aquestes, 24 són presents a Catalunya i 17 d’elles en són endèmiques (Cadevall et al., 2020).
En el present treball s’analitzen els resultats de les prospeccions dutes a terme, durant el juny del 2023, en dues fonts de la vall del torrent de Canyelles, a Castellar del Vallès (Barcelona) (Fig. 1), que van permetre obtenir nombroses conquilles en bon estat (però no animals vius) de microgastròpodes estigobis del gènere Moitessieria. L’estudi d’aquests materials ha possibilitat la descripció en base conquiliològica d’una nova espècie, que es presenta aquí. Aquest estudi també ha permès revisar i esmenar l’atribució específica del moitessièrid de la font de les Bassetes de Castellar del Vallès, que a Bros et al. (2022) va ésser identificat com a M. punctata Alba et al., 2010 i que aquí atribuïm a la nova espècie que descrivim.
Material i mètodes
Context geogràfic
El torrent de Canyelles està localitzat a la Serralada Prelitoral Catalana, en el municipi de Castellar del Vallès. És afluent del riu Ripoll, el qual, al seu torn, és tributari del riu Besòs. El sistema hidràulic del torrent de Canyelles ha tingut un paper històricament molt important per al desenvolupament de Castellar del Vallès. A la seva llera, als rodals del poble, hi ha terrenys d’ús hortícola regats amb les aigües d’aquest sistema, que hi arriben conduïdes mitjançant una canonada, fins a la caseta de repartiment del xaragall del Fuió (o d’en Foió). La canonada segueix després fins al poble, on havia alimentat les fonts públiques (Vergés i Vernís, 1987; Prat Paz, 2005; Font, 2016).
La geologia de la conca del torrent de Canyelles inclou, a la capçalera, dipòsits paleògens de l’Eocè (conglomerats del Cuisià, amb bretxes i intercalacions de lutites i gresos vermells). Més avall, a la zona on hi ha les fonts del Bassal i de la Noguera, hi predominen materials triàsics fortament enca-
valcats de fàcies Muschelkalk (calcàries, dolomies, gresos i lutites), de natura carbonàtica. La zona més baixa, propera a la desembocadura al riu Ripoll i on se situa la font de les Bassetes, està ocupada per dipòsits aŀluvials quaternaris amb graves amb matriu llimosa o sorrenca vermellosa (Casanovas-Vilar et al., 2011; Martínez i Rius et al., 2011).
El cobriment vegetal de la zona és un bosc mixt constituït bàsicament per pins blancs (Pinus halepensis) i alzines (Quercus ilex), amb una regeneració ja molt desenvolupada de l’alzinar litoral (Quercetum ilicis galloprovinciale), especialment en el fons de la vall del torrent de Canyelles. A les vores del torrent hi trobem de manera puntual algunes espècies vegetals característicament de ribera. Els conreus i les hortes dominen el paisatge a les zones més properes a l’aiguabarreig amb el riu Ripoll, sota el poble de Castellar del Vallès.
Material estudiat i obtenció
El material estudiat consta de 81 conquilles procedents de la font del Bassal (Fig. 2, a-b i d) (38 conquilles) i de la font de la Noguera (Fig. 2, c i e) (43 conquilles), totes dues fonts situades a la vall del torrent de Canyelles, dins el terme municipal de Castellar del Vallès, a la comarca del Vallès Occidental (Barcelona, Catalunya) [31TDG2408] (Fig. 1) (VB, GG i JC leg., 20/06/2023). A més, es van estudiar novament 11 conquilles del moitessièrid present a la font de les Bassetes (Fig. 2, f i g), que amb anterioritat (Bros et al. 2022) havien estat atribuïdes a M. punctata
L’obtenció dels espècimens es va fer a partir dels sediments obtinguts a l’interior dels conductes soterrats de captació i conducció de l’aigua de les dues fonts cap la zona d’horta. La metodologia de triatge mitjançant tamisos per extreure les conquilles de les espècies de moŀluscs estigobis dels sediments és la mateixa que està descrita en diferents articles anteriors dels autors (e.g. Tarruella et al., 2008; Corbella et al., 2019 i 2020; Bros et al., 2022).
Mostra de comparació
Per tal d’obtenir dades complementàries d’algunes espè-

2. Entorn i detalls dels punts de mostreig al torrent de Canyelles, a Castellar del Vallès (31/12/2023). a) tubs de conducció de la font del Bassal; b) el Bassal, en el propi curs del torrent de Canyelles; c) font de la Noguera; d) pericó sota la font del Bassal; e) pericó de la font de la Noguera; f) font de les Bassetes; g) brollador i piló de la font de les Bassetes
cies ja conegudes del gènere Moitessieria (dades necessàries i no sempre disponibles en les fonts bibliogràfiques), s’ha examinat un seguit de conquilles de cinc espècies (de les 43 incloses actualment al gènere (vegeu, més endavant, l’apartat «Comparativa amb les altres espècies del gènere Moitessieria») que, en conjunt, formen la mostra de comparació d’aquest estudi. La mostra inclou el material següent: Moitessieria garrotxaensis Quiñonero-Salgado & Rolán,
2017: 7 conquilles procedents de la font de la Teula, Sant Feliu de Pallerols (Girona, Espanya) (localitat tipus), 05/01/2020 (GG i JC leg. i det.).
Moitessieria pesanta Quiñonero-Salgado & Rolán, 2019: 10 conquilles procedents de la font de les Molleres, Sant Joan les Fonts (Girona, Espanya) (localitat tipus), 08/12/2014 (GG i JC leg. i det.).
Moitessieria racamondi Callot-Girardi, 2013: 3 paratips
Taula 1 Estadística descriptiva de les variables mètriques de Moitessieria canyellesensis sp. nov.
Abreviatures: n = mida mostral; SD = desviació estàndard; L = longitud (mm); A = amplada (mm); L/A = índex longitud / amplada; LDV = longitud de la darrera volta (mm); LDV/L = índex longitud de la darrera volta / longitud de la conquilla (%); NV = nombre de voltes; IS = inclinació de la sutura (graus sexagesimals); LE = nombre de línies espirals a la darrera volta; DE = densitat d’estriació (estries/mm).
Població de la font de la Noguera
Població de la font del Bassal Les
(Callot-Girardi leg., 19/10/2012) i 10 topotips, tots ells procedents de la font Dame (Fontdama), Salses (Pirineus Orientals, França) (localitat tipus), 04/11/2013 (GG i JC leg. i det.).
Moitessieria servaini (Bourguignat, 1880): nombroses conquilles procedents d’una font sense nom a Puendeluna (Osca, Espanya), 05/12/2011 (GG i JC leg. i det.).
Moitessieria simoniana (Saint-Simon, 1848): diverses conquilles procedents d’una font sense nom de la carretera D-117 prop de Baliar, Saint-Girons (Sent Gironç) (Arieja, França), 09/07/2007 (GG i JC leg. i det.). També s’han uti-
litzat fotografies dels possibles sintips d’aquesta espècie que conté la Coŀlecció Coutagne (núm. 943), conservada al Museu de les Confluències (ref. 46030809) de Lió (França).
Microscòpia
Les variables mètriques emprades en l’estadística descriptiva (Taula 1) es van mesurar mitjançant un estereomicroscopi Leica Wild M3Z, proveït d’un micròmetre ocular amb una precisió de 0,1 mm.
Les conquilles foren fotografiades mitjançant una càmera iPhone 14 ProMax, muntada sobre un microscopi òptic Kyowa Unilux-12 83-483D, amb apilament d’imatges i processament amb el programari Helicon Focus 7 (JCD 11/3).
Les micrografies es van realitzar mitjançant un microscopi electrònic de rastreig Zeiss ambiental EVO/MA10, al Servei de Microscòpia dels Serveis Cientificotècnics de la Universitat Autònoma de Barcelona (UAB).
Variables mètriques
Les variables mètriques emprades en l’estadística descriptiva (Taula 1) i en la comparativa amb les altres espècies del gènere són les següents (Fig. 3):
– Longitud (L): distància en miŀlímetres entre l’àpex i l’extrem inferior del peristoma.
– Amplada (A): distància en miŀlímetres entre el costat esquerre de la darrera volta i l’extrem dret del llavi extern.
– Índex L/A: proporció entre la longitud i l’amplada, que valora la major o menor esveltesa de la conquilla.
– Longitud de la darrera volta (LDV): distància en miŀlímetres entre l’inici de la darrera volta i l’extrem inferior del peristoma.
– Índex LDV/L: proporció (%) entre la longitud de la darrera volta (LDV) i la longitud total de la conquilla (L).
– Voltes (NV): nombre total de voltes de la conquilla. Als textos s’indica amb un arrodoniment a quarts de volta.
– Inclinació de la sutura (IS): inclinació, en graus sexagesimals, dels segments rectilinis definits pels punts d’inici i final del tram de sutura superior de cada volta respecte de la línia vertical de l’eix columeŀlar. El valor indicat és la mitjana dels valors obtinguts a les sutures superiors de la V.
BROS
penúltima i l’antepenúltima voltes.
– Línies espirals (LE): nombre d’estries espirals presents a la darrera volta entre l’umbilic i la sutura.
– Densitat d’estriació (DE): nombre de línies espirals presents a la darrera volta, per miŀlímetre lineal de longitud
(indicat en estries/mm).
Tal i com s’ha fet en diversos altres treballs (e.g. Coutagne, 1883; Boeters & Gittenberger, 1980; Boeters & Falkner, 2001; Bertrand, 2001; Girardi, 2003 i Corbella et al., 2020), en aquest estudi no han estat utilitzades ni la longitud de

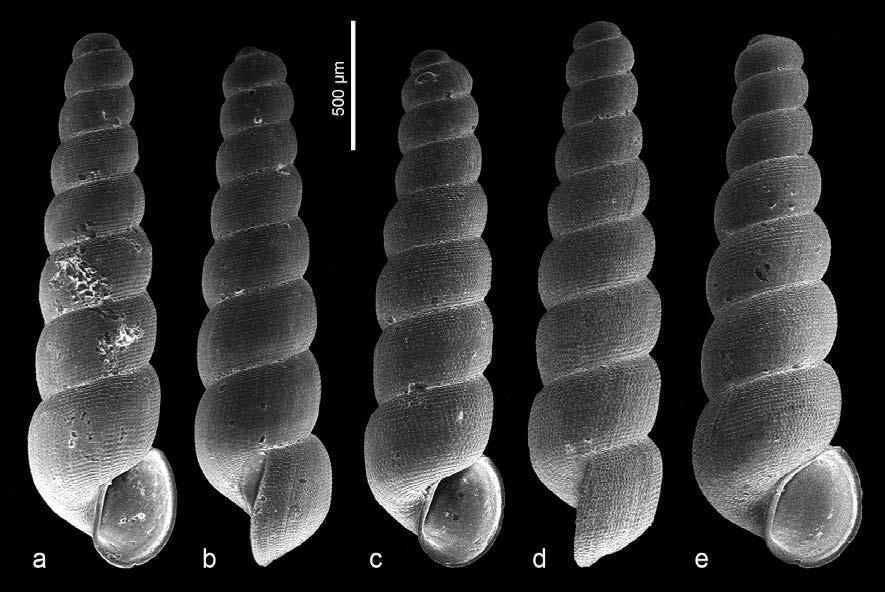
nov. a-b) paratips de la font del Bassal (a:
Abreviacions: BA: font del Bassal, NO: font de la Noguera).
b:
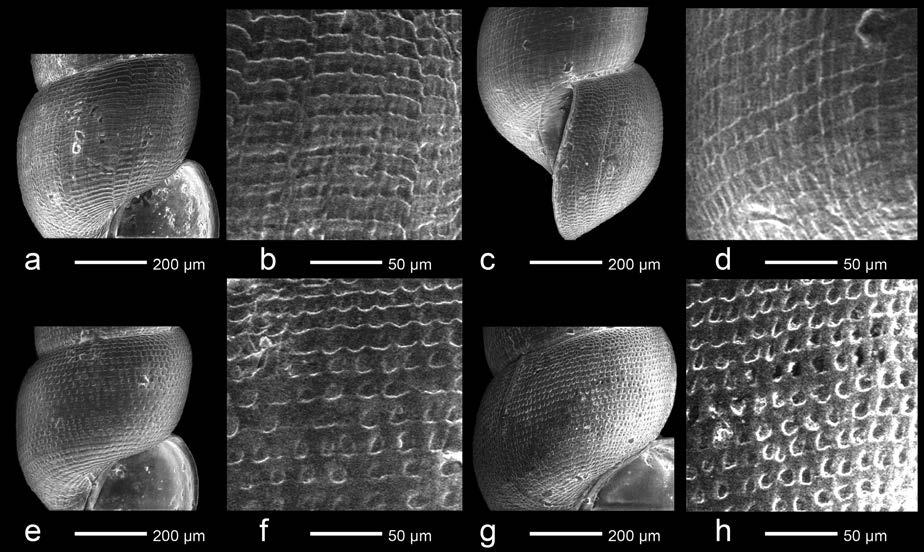
Figura 7. Micrografies de l’ornamentació de la teleoconquilla a la darrera volta de quatre paratips de Moitessieria canyellesensis sp. nov. a-d) paratips de la font del Bassal (a-b: BA-28, c-d: BA-29); e-h) paratips de la font de la Noguera (e-f: NO-41, g-h: NO-43). Abreviacions: BA: font del Bassal, NO: font de la Noguera).

Figura 8. Micrografies de detall de la protoconquilla de tres paratips de Moitessieria canyellesensis sp. nov. a-b) paratips de la font del Bassal (a: BA-27, b: BA-29); c) paratip de la font de la Noguera (NO-42). Abreviacions: BA: font del Bassal, NO: font de la Noguera)
l’obertura (distància en miŀlímetres entre el punt superior d’inici del llavi extern i l’extrem inferior del peristoma), ni l’amplada de l’obertura (prenent com a referència la perpendicularitat a l’eix columeŀlar, distància en miŀlímetres entre els dos punts laterals del peristoma més allunyats entre si) a causa de l’excessiva irregularitat intraespecífica d’aquestes dues variables dins el gènere Moitessieria.
Sistemàtica
La sistemàtica emprada segueix MolluscaBase (2021) i GBIF (2023).
Fílum Mollusca Cuvier, 1795
Classe Gastropoda Cuvier, 1795
Subclasse Caenogastropoda Cox, 1960
Ordre Littorinimorpha Golikov i Starobogatov, 1975
Superfamília Truncatelloidea Gris, 1840
Família Moitessieroiidae Bourguignat, 1864
Gènere Moitessieria Bourguignat, 1864
Moitessieria canyellesensis sp. nov.
(Figures 4, – 9b, 9e i 9h)
Holotip
Dipositat al Museu de Ciències de Barcelona amb el nú-
V. BROS ET AL.

Figura 9. Comparativa entre Moitessieria olleri (topotips de la Cova del Toll, Moià), Moitessieria canyellesensis sp. nov. (paratips de la font del Bassal, Castellar del Vallès) i Moitessieria punctata (paratips de les Fonts, Rellinars). a-c) exemplars complets a escala (a: M. olleri, b: M. canyellesensis sp. nov., c: M. punctata); d-f) darreres voltes, a igual proporció (escales diverses) (d: M. olleri, e: M. canyellesensis sp. nov., f: M. punctata); g–I: textures a escala de les darreres voltes (g: M. olleri, h: M. canyellesensis sp. nov., i: M. punctata).
Butlletí ICHN 88 (4), 2024
mero de catàleg MZB 2024-3746 (Figs. 4 i 5), Font del Bassal; longitud 2,35 mm; amplada 0,70 mm; 7 ¾ voltes.
Paratips
Un paratip dipositat al MCNB amb el número de catàleg MZB 2024-3747, Font de la Noguera. Resta de paratips dipositats a les coŀleccions dels autors (fonts del Bassal i de la Noguera) (Figs. 4 i 6–9).
Localitat típica
Font del Bassal (375 m), Castellar del Vallès (el Vallès Occidental, Barcelona). 31TDG2408.
Distribució
L’espècie ha estat localitzada, fins ara, a més de a la localitat típica, a d’altres fonts del torrent de Canyelles, a Castellar del Vallès, concretament a la font de la Noguera (365 m, 31TDG2408), i la font de les Bassetes (310 m, 31TDG2307).
Etimologia
L’epítet específic fa referència al torrent de Canyelles, dins la conca del qual s’ubiquen les fonts on s’ha localitzat la nova espècie.
Descripció
Conquilla despigmentada, turriculada i allargada, de forma subcilíndrica a cilindrocònica i creixement regular, tot i que amb certa freqüència presenta la darrera volta reinflada, sobresortint de la resta de l’espira (Figs. 4–6 i 9). La seva mida és gran, amb una longitud (L) mitjana de 2,14 mm (rang 1,65−2,68 mm) i una amplada (A) mitjana de 0,63 mm (rang 0,53−0,75 mm). Té 7 ¾ voltes (NV) de mitjana (rang 6½–10) i la conquilla és força esvelta, amb un índex longitud/amplada (L/A) mitjà de 3,41 (rang 2,71–4,46).
Les voltes de l’espira presenten un perfil entre convex i planoconvex. La sutura és profunda i molt inclinada respecte de l’eix columeŀlar, amb una inclinació (IS) mitjana de 19,9o (rang 15,0o–24,5°) (Figs. 4–6 i 9). La darrera volta pot ser no reinflada o bé reinflada en grau variable, és de perfil convex a planoconvex (Figs. 4–6 i 9), té una longitud (LDV) mitjana de 0,79 mm (rang 0,68−0,95 mm), ocupa de mitjana el 37 % de la longitud total de la conquilla (LDV/L) (rang 31 %–45 %), presenta el seu tram final no ascendent ni descendent i està completament enganxada a la resta de l’espira (Figs. 4−6). El llavi extern de l’obertura no és gens o és una mica prominent, amb la zona inferior moderadament convexa en vista lateral, i forma un sínul poc acusat a la zona d’inserció parietal (Figs. 5, 6 i 9). El peristoma és continu i poc o gens expandit. Sovint té el vorell superior molt adherit a la volta anterior, tot i que algunes vegades en queda quasi separat (Figs. 5−7 i 9), i el vorell columeŀlar és moderadament reflectit (Figs. 5 i 6).
L’umbilic pot ser des de quasi obliterat fins a obert en forma de fenedura estreta (Figs. 4–6 i 9).
L’ornamentació de la teleoconquilla està constituïda, en part, per solcs i crestes espirals d’amplades variables que, en creuar-se amb les línies de creixement, formen una malla de depressions quadrangulars. Aquesta microescultura és dèbil
(fins i tot desdibuixada) en la quasi totalitat de la teleoconquilla, excepte al tram final de la darrera volta, on és present i pot arribar a ésser ben marcada. Pel que fa a la resta de la teleoconquilla, presenta una microescultura a base de puntejadures arrenglerades espiralment de manera regular. Les puntejadures són de mida mitjana, de forma arrodonida i/o eŀlipsoidal, poden quedar obertes apicalment en major o menor grau (tot i que no sempre), i es disposen amb una densitat variable (Figs. 5−7). A la darrera volta, entre l’umbilic i la sutura, hi ha de mitjana 28 línies espirals (LE) (rang 20–39). Això fa que la densitat d’estriació (DE) mitjana sigui de 63 estries/mm (rang 42–80 estries/mm). L’ornamentació de la protoconquilla és aparent i variable, a base de papiŀles, rugositats i cordons disposats espiralment o irregular. Predomina un tipus o altre de microescultura segons els espècimens (Fig. 8).
L’opercle, l’anatomia reproductora, la ràdula i les dades moleculars no s’han pogut obtenir per manca de material viu.
Mesures
Vegeu la taula 1.
Diagnosi diferencial
La nova espècie es diferencia d’una bona part de les espècies del mateix gènere descrites fins ara per tenir una conquilla esvelta (índex L/A mitjà > 2,7), un nombre mitjà de voltes entre moderat i elevat (≥ 6 ½ ) i la darrera volta curta (relació mitjana LDV/L = 37 %), i per presentar una ornamentació sense puntejadures a la protoconquilla.
De les espècies en què les quatre característiques esmentades no són suficients per fer-ne la diferenciació [= M. garrotxaensis Quiñonero Salgado & Rolán, 2017, M. notenboomi Boeters, 2003, M. olleri Altimira, 1960, M. ouvezensis Boeters & Falkner, 2009, M. prioratensis Corbella et al., 2009, M. punctata Tarruella et al., 2010, M. robresia Boeters, 2003, M. sanctichristophori Corbella et al., 2011, M. servaini (Bourguignat, 1880) i M. tatirocae Tarruella et al., 2015], la nova espècie se’n diferencia, segons el cas, per les seves dimensions (L i/o A), la disposició de la darrera volta, la convexitat de les voltes, l’adherència i el grau d’expansió del peristoma, la prominència del llavi extern, el grau de pronunciament del sínul, la inclinació de la sutura, el tipus de microescultura de la teleoconquilla, el nombre mitjà de línies espirals a la darrera volta damunt de l’obertura i la densitat d’estriació mitjana a la darrera volta (vegeu l’apartat «Discussió» per a més detalls).
Distribució
Fins ara, la nova espècie es coneix de tres fonts (font del Bassal, font de la Noguera i font de les Bassetes) situades a la vall del torrent de Canyelles, al terme de Castellar del Vallès (el Vallès Occidental, Barcelona).
Hàbitat
L’espècie ha estat trobada, tot i que només conquilles, als conductes soterrats que, des dels brolladors respectius, també soterrats, capten i condueixen cap a una zona d’horta l’aigua
Taula 2 Comparativa del nou tàxon amb les dues espècies que li són més properes geogràficament i morfològica, M. olleri i M. punctata
Moitessieria canyellesensis sp. nov. Moitessieria olleri Moitessieria punctata
Distribució coneguda torrent Canyelles, Castellar del Vallès Moianès, Lluçanès i Vallès Occidental Bages i Vallès Occidental (Vallès Occidental)
Expansió del peristoma poc expandit poc expandit expandit
Sínul poc acusat poc acusat acusat
Disposició tram final de la darrera volta no ascendent no ascendent ascendent
Presència d’ornamentació reticular a la present només en la darrera volta present en tota la superfície No present teleoconquilla
Presència de puntejadures sempre no sempre presents sempre presents
Mida de les puntejadures mitjana petita mitjana
Disposició de les puntejadures poc densa poc densa molt densa
Densitat mitjana d’estriació 63 estries/mm 71 estries/mm 79 estries/mm
de les fonts del Bassal i de la Noguera, i als sediments del piló de la font de les Bassetes. L’espècie, doncs, té hàbitat estigobi.
Discussió
Atribució a la família Moitessieriidae
La família Moitessieriidae està constituïda per espècies de micromoŀluscs dulciaqüícoles estigobis. Tant Boeters & Gittenberger (1990) com Boeters et al. (2013) consideraren que l’única característica conquiliològica diferenciadora de la família (enfront de la família Hydrobiidae W. Stimpson, 1865) és la presència a la superfície de la teleoconquilla d’una ornamentació a base de línies espirals, paraŀleles a la sutura. L’hàbitat, la mida i l’ornamentació de la teleoconquilla permeten, doncs, atribuir la nova espècie a la família Moitessieriidae.
Atribució al gènere Moitessieria
D’entre els gèneres inclosos fins ara a la família Moitessieriidae (vegeu Corbella et al., 2020), a Catalunya només ha estat publicada la presència de Moitessieria, Baldufa Alba et al., 2010 i Tarracospeum Quiñonero-Salgado et al., 2021. Aquest darrer gènere va ésser atribuït a dita família tot i que a la microescultura de la teleoconquilla només presenta línies de creixement irregulars (Quiñonero-Salgado et al., 2021) i no té estriació espiral.
Baldufa té, entre d’altres, una conquilla cònicotrocoïdal, amb poques voltes, molt robusta i amb una ornamentació molt peculiar (recorda les circumvolucions cerebrals) a la protoconquilla (Alba et al., 2010). Aquestes característiques exclouen el gènere Baldufa per a la nova espècie.
Per la seva banda, Tarracospeum presenta el peristoma molt reflectit i desenganxat de la resta de la darrera volta, el llavi extern molt prominent lateralment i la superfície de la
Butlletí ICHN
(4), 2024
conquilla sense estriació espiral (Quiñonero-Salgado et al., 2021). Aquests trets exclouen l’atribució de la nova espècie a Tarracospeum. Tant l’hàbitat estigobi com les seves característiques conquiliològiques permeten, en canvi, l’atribució de la nova espècie al gènere Moitessieria. Així, M. canyellesensis sp. nov. té una conquilla despigmentada, molt petita, esvelta, turriculada subcilíndrica, amb una sutura profunda, un sínul a la zona d’inserció parietal del llavi extern, un umbilic molt estret o obliterat i una ornamentació espiral marcada a la teleoconquilla. La presència conjunta de totes aquestes característiques només es dona, entre els moitessièrids ibèrics, en espècies del gènere Moitessieria (vegeu, per exemple, Boeters, 2003).
Comparativa amb les altres espècies del gènere Moitessieria
Tot seguit discutim la distinció de la nova espècie respecte de les restants espècies vàlides que s’inclouen actualment en el gènere Moitessieria, mitjançant agrupacions morfològiques (conquiliològiques) d’espècies. Aquells caràcters que no s’han pogut obtenir de les referències bibliogràfiques que se citen, s’han observat o mesurat directament a la mostra de comparació esmentada a l’apartat «Material i mètodes». Moitessieria canyellesensis sp. nov. es diferencia d’un nombrós grup d’espècies de Moitessieria pel fet de tenir una mitjana en la relació entre la longitud i l’amplada (L/A) superior a 2,7. Aquest fet li dona una forma més esvelta que la que presenten les espècies de l’esmentat grup, format per M. atlantica Prié, 2024, M. aurea Tarruella et al , 2012, M. barrinae Alba et al., 2007, M. canfalonensis Corbella et al., 2020, M. collellensis Corbella et al., 2006, M. dexteri Corbella et al., 2012, M. fontsaintei Bertrand, 2001, M. foui Boeters, 2003, M. guilhemensis Callot-Girardi & Boeters, 2017, M. hedraensis Quiñonero-Salgado & Rolán, 2017, M. heideae Boeters & Falkner, 2003, M. juvenisanguis Boeters & Gittenberger, 1980, M. lludrigaensis Boeters, 2003, M. locardi
(Coutagne, 1883), M. magnanae Girardi, 2009, M. meijersae Boeters 2003, M. pesanta Quiñonero-Salgado & Rolán, 2019, M. racamondi Callot-Girardi, 2013, M. rhodani (Bourguignat in Coutagne, 1883), M. ripacurtiae Tarruella et al., 2013 i M. vidourlensis Prié, 2024. Totes elles presenten valors mitjans de l’índex L/A ≤ 2,7 (Alba et al., 2013; Bernasconi, 1994; Bertrand, 2001; Boeters, 2003; Boeters & Falkner, 2008; Boeters & Gittenberger, 1980; Callot-Girardi, 2013; Callot-Girardi & Boeters, 2017; Corbella et al., 2012 i 2020; Corbella et al., 2006; Girardi, 2003 i 2009; Girardi & Wienin, 2000; Prié et al., 2024; Quiñonero-Salgado & Rolán, 2017 i 2019; Tarruella et al., 2012 i 2013 i mostra de comparació).
El nombre mitjà de voltes de la conquilla (NV), igual o superior a 6 ½ en la nova espècie, també permet diferenciar-la de M. audiberti Callot-Girardi, 2015, M. bourguignati Coutagne, 1883, M. calloti Girardi, 2003, M. larzacensis Prié, 2024, M. massoti Bourguignat, 1864, M. pasterae Corbella et al., 2009 i M. seminiana Boeters, 2003, que presenten totes elles un nombre mitjà de voltes inferior a 6 ½ (Boeters, 2003; Callot-Girardi, 2015a; Corbella et al., 2006, Corbella et al., 2009; Coutagne, 1883; Girardi, 2003 i Prié, 2024).
Moitessieria guadelopensis Boeters, 2003, M. mugae Corbella et al., 2006, M. nezi Boeters & Bertrand, 2001 i M. simoniana (Saint-Simon, 1848) presenten un valor mitjà de la proporció entre la longitud de la darrera volta i la longitud total de la conquilla (LDV/L) ≥ 44 % (Boeters, 2003; Boeters & Bertrand, 2001 i mostra de comparació). La nova espècie, en canvi, presenta per dita relació un valor mitjà del 37%, que permet distingir-la de les quatre espècies suara esmentades.
El tipus de microescultura que presenta la protoconquilla és útil per diferenciar la nova espècie de M. boetersi CallotGirardi, 2015 i M. rolandiana Bourguignat, 1864. En aquestes dues espècies, la protoconquilla té micropuntejadures dispostes espiralment (Callot-Girardi, 2015b i Girardi, 2003), mentre que a Moitessieria canyellesensis sp. nov. l’ornamentació de la protoconquilla és una combinació de papiŀles, crestes i cordons, variable d’uns espècimens a uns altres, però mai amb presència de puntejadures.
Pel que fa a la distinció amb la resta de les espècies conegudes del gènere, convé fer-la de manera individualitzada. Tot seguit s’indica entre parèntesis la característica present a la nova espècie o bé la comparativa entre les dues espècies: Moitessieria garrotxaensis Quiñonero Salgado & Rolán, 2017 presenta una inclinació mitjana de la sutura (IS) menor (16o vs. 20o), el sínul acusat (vs. poc acusat) i no té puntejadures a la teleoconquilla (vs. puntejadures presents) (Quiñonero-Salgado & Rolán, 2017 i mostra de comparació).
Moitessieria notenboomi Boeters, 2003 és de longitud mitjana (L) i amplada mitjana (A) superiors (2,5 mm i 0,8 mm versus 2,2 mm i 0,6 mm), té un nombre mitjà de línies espirals (LE) inferior (18 vs. 28), una densitat d’estriació (DE) mitjana també inferior (40 estries/mm vs. 63 estries/mm), la darrera volta ascendent al seu tram final (vs. no ascendent) i el sínul acusat (vs. poc acusat) (Boeters, 2003).
Moitessieria olleri Altimira, 1960 (Fig. 9a, Taula 2) presenta una microescultura reticular en tota la superfície de la teleoconquilla (vs. només ben definida a la darrera volta), una
densitat d’estriació (DE) major (71 estries/mm vs. 63 estries/ mm) i, quan hi són presents, les puntejadures de mida petita (vs. mida mitjana) (Altimira, 1969, Corbella et al., 2006 i Tarruella et al., 2008).
Moitessieria ouvezensis Boeters & Falkner, 2009 té el peristoma poc adherent (vs. ben adherent), el llavi extern prominent (vs. poc o gens prominent) i una inclinació de la sutura (IS) menor (17o vs. 20o) (Boeters & Falkner 2009).
Moitessieria prioratensis Corbella et al., 2009 té les puntejadures grans (vs. mitjanes), no presenta microescultura reticulada (vs. present a la darrera volta) i la convexitat de les seves voltes és acusada i, sovint, desplaçada cap a la part basal de cada volta (vs. poc acusada i centrada en cada volta) (Corbella et al., 2009).
Moitessieria punctata Alba et al., 2010 (Fig. 9c, Taula 2) presenta el tram final de la darrera volta ascendent (vs. no ascendent), un sínul acusat (vs. poc acusat), el peristoma expandit (vs. poc expandit), les puntejadures disposades molt densament (vs. poc densament), la densitat d’estriació (DE) mitjana més elevada (78-80 estries/mm vs. 63 estries/mm) i no presenta microescultura reticulada (vs. present a la darrera volta) (Alba et al., 2010, Tarruella et al., 2015).
Moitessieria robresia Boeters, 2003 presenta un sínul acusat (vs. poc acusat), el peristoma poc adherent (vs. ben adherent) i la darrera volta ascendent al seu tram final (vs. no ascendent) (Boeters, 2003).
Moitessieria sanctichristophori Corbella et al , 2011 és de dimensions mitjanes més petites, tant en longitud (L) (1,7 mm vs. 2,1 mm) com en amplada (A) (0,5 mm vs. 0,6 mm) i no presenta microescultura reticulada (vs. present a la darrera volta) (Corbella et al., 2011).
Moitessieria servaini (Bourguignat, 1880) té el sínul acusat (vs. poc acusat), un nombre mitjà de línies espirals (LE) inferior (17 vs. 28) i presenta les puntejadures poc atapeïdes (vs. generalment atapeïdes) (Boeters, 2003; Bourguignat, 1880; Corbella et al., 2012 i mostra de comparació).
Finalment, M. tatirocae Tarruella et al., 2015 també té el sínul acusat (vs. poc acusat), no presenta microescultura reticulada (vs. present a la darrera volta) i les seves puntejadures són de mida gran (vs. mitjanes) (Tarruella et al., 2015).
En la Taula 2 se sintetitzen les característiques que distingeixen el nou tàxon de M. olleri i M. punctata, les dues espècies que li són més properes geogràficament i més similars morfològicament.
Conclusions
Es descriu una nova espècie de moitessièrid estigobi, M. canyellesensis sp. nov., a partir de material conquiliològic de dues fonts ubicades a la vall del torrent de Canyelles, a Castellar del Vallès: la font del Bassal i la font de la Noguera. Amb la descripció d’aquesta, s’eleva a 44 el nombre d’espècies atribuïdes al gènere Moitessieria, de les quals 25 són presents a Catalunya, i almenys 18 d’elles han de considerarse’n, per ara, endèmiques.
El present estudi també esmena l’atribució específica que Bros et al. (2022) van assignar a la Moitessieria present a la font de les Bassetes de Castellar del Vallès, que en dit treball es va atribuir a M. punctata. En realitat es tracta de la M. canyellesensis sp. nov. que ací es descriu.
Com succeeix amb la resta d’organismes estigobis, la distribució geogràfica restringida i l’elevat grau d’endemisme de la majoria de les espècies les fan vulnerables a les alteracions dels seus hàbitats. Tal i com ja es va indicar a Corbella et al. (2020), la conservació de les espècies de Moitessieria requereix de més investigacions, per tal d’augmentar els coneixements respecte les característiques dels hàbitats que condicionen la supervivència de les poblacions i, a més, aplicar mesures que evitin de manera eficient la contaminació de les aigües subterrànies i la sobreexplotació dels aqüífers.
Volem palesar el nostre agraïment a les següents persones i entitats, que han contribuït en la materialització d’aquest treball. A Henri Callot-Girardi (Montfavet, Valclusa, França), que ens ha proporcionat paratips de M. racamondi. A Cédric Audibert (Museu de les Confluències, Lió), per permetre’ns consultar els possibles sintips de M. simoniana de la Coŀlecció Coutagne (20/07/2011) i proporcionar-nos-en fotografies per poder-los estudiar. A Carles Ros i Miquel Àngel López per llur coŀlaboració en la localització sobre el terreny de les fonts, actualment un mica amagades, del torrent de Canyelles. A Jordi Martínez-Vilalta (Universitat Autònoma de Barcelona i C.R.E.A.F.), que ens possibilità l’accés al Servei de Microscòpia i Difracció de Raigs X (SMiDRX) dels Serveis Cientificotècnics de la UAB, i a en Martí de Cabo, tècnic superior de dits Serveis, que va realitzar les micrografies d’ESEM ambiental. També a Jorge Mederos, investigador associat del Museu de Ciències Naturals de Barcelona, pel seu ajut en les fotografies amb microscopi òptic. Finalment, agraïm a Josep Quintana Cardona i David M. Alba la seva valuosa aportació en la revisió del manuscrit.
Bibliografia
Alba, D. M., Corbella, J., Prats, L., Guillén, G. & Tarruella, A. 2013. Conchological characterization of Moitessieria foui Boeters, 2003 (Gastropoda: Moitessieriidae), Spira, 5 (1-2): 91-98. Alba, D. M., Tarruella, A., Prats, L., Guillén, G. & Corbella, J. 2010. Els moitessièrids (Gastropoda: Moitesieriidae) de Rellinars (el Vallès Occidental, Catalunya, Espanya). Spira, 3 (3): 159-186. Altimira, C. 1960. Notas Malacológicas. Contribución al conocimiento de los moluscos terrestres y de agua dulce de Cataluña. Misceŀlània Zoològica, 1 (3): 9-15. Bernasconi, R. 1994. Le genre Moitessieria Bgt., 1863 en France : Révisions, inventaire et description de M. Corsica n. sp. (Mollusca Gastropoda Prosobranchia Hydrobiidae). Mémoires de Biospéologie, 11: 7-20.
Bertrand, A. 2001. Moitessieria fontsaintei sp. nov. (Gastropoda : Prosobranchia : Moitessieriidae) des Pyrénées ariégeoises. Documents Malacologiques, 2: 39-41.
(4),
Bodon, M. & Giusti, F. 1991. The genus Moitessieria in the island of Sardinia and Italy. New data on the systematics of Moitessieria and Paladilhia (Prosobranchia: Hydrobiidae) (Studies on the Sardinian and Corsican malacofauna, IX). Malacologia, 33: 1–30.
Boeters, H. D. 1988. Moitessieriidae und Hydrobiidae in Spanien und Portugal (Gastropoda: Prosobranchia). Westeuropäische Moitessieriidae, 2 und Westeuropäische Hydrobiidae, 7. Archiv für Molluskenkunde, 118: 181-261.
Boeters, H. D. 2003. Supplementary notes on Moitesieriidae and Hydrobiidae from the Iberian Peninsula (Gastropoda, Caenogastropoda). Basteria, 67: 1-41.
Boeters, H. D. & Bertrand, A. 2001. A remarkably rich prosobranch fauna endemic to the French Pyrenées. Basteria, 65: 1-15.
Boeters, H. D. & Falkner, G. 2001. Beiträge zur Nomenklatur der europäischen Binnenmollusken, XIII. The identity of Paludina simoniana Saint Simon 1848 (Gastropoda: Moitessieriidae). Heldia, 3 (2/3): 81-84, pl. 11.
Boeters, H. D. & Falkner, G. 2003. Unbekannte westereuropäische Prosobranchia, 14. Neue und alte Grundwasserschnecken aus Frankreich (Gastropoda: Moitessieriidae et Hydrobiidae). Heldia, 5 (1/2): 7-18.
Boeters, H. D. & Falkner, G. 2009. Unbekannte westeuropäische Prosobranchia, 15.) Neue und alte Quell-und Grundwasserschnecken aus Frankreich (Gastropoda: Moitessieriidae et Hydrobiidae). Heldia, 5 (6): 149-162.
Boeters, H. D. & Gittenberger, E. 1980. Unbekannte westeuropäische Prosobranchia, 4. Basteria, 44: 65-68.
Boeters, H. D. & Gittenberger, E. 1990. Once more on the Moitessieriidae (Gastropoda Prosobranchia), with the description of Clameia brooki gen. et spec. nov. Basteria, 54: 123-129.
Boeters, H. D., Glöer, P. & Pesic, V. 2013. Some new freshwater gastropods from southern Europe (Mollusca: Gastropoda: Truncatelloidea). Folia Malacologica, 21: 225-235.
Bourguignat, J.R. 1880. Description de diverses espèces de Coelestele et de Paladilhia découvertes en Espagne par le Dr. G. Servain. Imprimerie P. Lachèse et Dolbeau, Angers.
Bros, V., Cadevall, J., Guillén, G., Prats, L., & Corbella, J. 2022. Diversitat i distribució dels moŀluscs estigobis i estigòfils (Moitessieriidae i Hydrobiidae) en el massís de Sant Llorenç del Munt i la serra de l'Obac (Barcelona, NE península Ibèrica). Spira, 8: 1-19.
Cadevall, J., Corbella, J., Bros, V., Orozco, A., Guillén, G., Prats, L., Capdevila, M. 2020. Els moŀluscs continentals de Catalunya i Andorra (península ibèrica). Llista comentada. Spira, 7 (3-4): 117-159.
Callot-Girardi, H. 2013. Moitessieria racamondi sp. nov., nouvelle espèce de la résurgence de Font Dame, et Moitessieria juvenisanguis Boeters et Gittenberger, 1980 (Gastropoda : Moitessieriidae), nouvelle présence à la source du lavoir (Salses-le-Château, Pyrénées-Orientales, France). Spira, 5 (1-2): 37-52.
Callot-Girardi, H. 2015a. Moitessieria audiberti, nouvelle espèce stygobie du ruisseau de Lachein-Bajet à Alas dans le département de l’Ariège, France. Bulletin de Malacologie Avenionia, 1: 1320.
Callot-Girardi, H. 2015b. Moitessieria boetersi, nouvelle espèce cavernicole pour la malacofaune française, dans les ruisseaux souterrains. D’Amiel et de Cabéou à Penne, Tarn, France (1ère partie). Bulletin de Malacologie Avenionia, 1: 30-43.
Callot-Girardi, H. & Boeters, H. D. 2017. Moitessieria guilhemensis, nouvelle espèce de la résurgence du Cabrier à Saint-Guilhemle-Désert, Hérault, France. (Mollusca : Caenogastropoda : Moitessieriidae). Bulletin de Malacologie Avenionia, 2: 42-64.
Casanovas-Vilar, I., Alba, D. M., Robles, J. M. & Moyà-Solà, S. 2011. Registro paleontológico continental del Mioceno de la V. BROS
cuenca del Vallès-Penedès. Paleontologia i Evolució, memòria especial, 6: 55-80.
Corbella, J., Alba, D. M., Tarruella, A., Prats, L. & Guillén, G. 2006. Dues noves espècies del gènere Moitessieria Bourguignat, 1863 (Neotaenioglossa : Rissoidea : Moitessieriidae) de la Font d’en Collell (Albanyà, l’Alt Empordà, Catalunya, Espanya). Spira, 2 (2): 71-111.
Corbella, J., Alba, D. M., Tarruella, A., Guillén, G. & Prats, L. 2009. Noves espècies de Moitessieria Bourguignat, 1863 (Neotaenioglossa: Rissooidea: Moitessieriidae) de la Font del Racó de la Pastera (Ulldemolins, el Priorat, Catalunya, Espanya). Spira, 3 (1-2): 59-86.
Corbella, J., Bros, V., Guillén, G., Prats, L. & Cadevall, J. 2020. Moitessieria canfalonensis sp. nov. (Gastropoda: Moitessieriidae), una nova espècie estigòbia de Monistrol de Calders (Barcelona, NE península Ibèrica). Spira, 7: 101-116.
Corbella, J. Guillén, G., Prats, L. & Bros, V. 2019. Estudi de la fauna malacològica estigòbia del Parc Natural de Sant Llorenç del Munt i l'Obac. Informe tècnic. Oficina Tècnica de Parcs Naturals, Diputació de Barcelona. 43 p.
Corbella, J., Guillén, G., Prats, L., Tarruella, A. & Alba, D. M. 2012. Els Moitessieriidae (Gastropoda: Rissooidea) de Les Deus de Sant Quintí de Mediona (l’Alt Penedès, Catalunya, Espanya); descripción de Moitessieria dexteri sp. nov. Spira, 4 (3-4): 121-138.
Coutagne, G. 1883. Révision sommaire du genre Moitessieria (suite). Feuille de Jeunes Naturalistes, 156: 143-147.
Falniowski, A., Prevorčnik, S., Delić, T., Alther, R., Altermatt, F., & Hofman, S. 2019. Monophyly of the Moitessieriidae Bourguignat, 1863 (Caenogastropoda : Truncatelloidea). Folia Malacologica, 27 (1): 61-70.
Font, J. O. 2016. Dels orígens de l’aprofitament de l'aigua a Castellar del Vallès fins als nostres dies: un viatge a través de la història de la Comunitat de Regants de les aigües del torrent de Canyelles. Plaça Vella, 57: 5-18.
GBIF Secretariat 2023. GBIF Backbone Taxonomy. Checklist dataset https://doi.org/10.15468/39omei accessed via GBIF.org on 2024-06-23.
Girardi, H. 2003. Moitessieria calloti espèce nouvelle : redescriptions de Moitessieria rhodani (Bourguignat, 1893) et autres Moitessieries du Gard et de l’Ardèche, France (Gastropoda : Moitessieriidae). Documents Malacologiques, 4: 59-65.
Girardi, H. 2009. Contributions à la connaissance des mollusques d’eau douce de France. Paladilhia subconica et Moitessieria magnanae, nouvelles espèces de la grotte des Châtaigniers à SaintMartin-de-Londres, Hérault, France et autres observations sur les Moitessieria (Mollusca : Caenogastropoda : Moitessieriidae). Documents Malacologiques, Hors série, 3: 109-118.
Girardi, H. et Wienin, M. 2000. Les Hydrobiidae (Mollusca : Gastropoda : Prosobranchia) du massif karstique de Saint-Julien-desRosiers (Gard). Documents Malacologiques, 1: 3-10.
Glöer, P. 2022. The freshwater gastropods of the West-Palaearctis. Identification key, Anatomy, Ecology, Distribution. Vol. II, Moitessieriidae, Bythinellidae, Stenothyridae. P. Glöer Ed., Hetlingen. 386 p.
Martínez i Rius, A., López Blanco, M., Florensa, R. M., Puig i Civera, C., Casanova i Petanas, J. & Picart i Boira, J. 2011. Mapa geològic de Catalunya 1:25.000, Castellar del Vallès 392-2-1 (7229). Institut Geològic de Catalunya (IGC) i Institut Cartogràfic de
Catalunya (ICC), Barcelona. MolluscaBase eds. 2021. MolluscaBase. Moitessieria Bourguignat, 1864. Accessed at : https://www.molluscabase.org/aphia. php?p=taxdetails¬id=715909 on 2024-06-24.
Prat Paz, E. 2005. Sistema hidràulic del torrent de Canyelles. L’aigua a Castellar del Vallès. Una història a través de la hidràulica. Plaça Vella, 50: 13-119.
Prié, V., Alonso, C., Bou, C., Galassi, D. M. P., Marmonier, P. & Dole-Olivier, M. J. 2024. The Cent Fonts Aquifer: An Overlooked Subterranean Biodiversity Hotspot in a Stygobiont-Rich Region. Diversity: 16, 50.
Quiñonero-Salgado, S. & Rolán, E. 2017. Two new species of the genus Moitessieria (Gastropoda, Moitessieriidae) from Spain. Iberus, 35 (2): 115-121.
Quiñonero-Salgado, S. & Rolán, E. 2019. A new species of the genus Moitessieria Bourguignat, 1863 (Gastropoda: Moitessieriidae) from Catalonia, Spain. Nemus, 9: 105-111.
Quiñonero-Salgado, S., Ruiz-Jarillo, R., Alonso, A. & Rolán, E. 2021. Tarracospeum raveni, a new genus and a new species of stygobiotic mollusc (Gastropoda: Moitessieriidae) for Spain. Nemus, 11: 120-126.
Richling, I., Malkowsky, Y., Kuhn, J., Niederhöfer, H. J., & Boeters, H. D. 2017. A vanishing hotspot—the impact of molecular insights on the diversity of Central European Bythiospeum Bourguignat, 1882 (Mollusca: Gastropoda: Truncatelloidea). Organisms Diversity & Evolution, 17 (1): 67-85.
Tarruella, A., Alba, D. M., Prats, L., Guillén, G. & Corbella, J. 2008. Caracterització conquiliològica i noves dades de distribució de Moitessieria olleri Altimira, 1960 (Neotaenioglossa: Rissooidea: Moitessieriidae). Spira, 2: 223-262.
Tarruella, A., Corbella, J., Guillén, G., Prats, L. & Alba, D. M. 2013. Moitessieria ripacurtiae sp. nov. (Gastropoda: Moitessieriidae), una nova espècie de gastròpode estigobi del Pont de Suert (l'Alta Ribagorça, Catalunya, Espanya). Spira 5 (1-2): 15-26.
Tarruella, A., Corbella, J., Prats, l., Guillén, G. & Alba, D.M. 2012. Una nova espècie de Moitessieria (Gastropoda: Moitessieriidae) del Forat de l’Or (Camarasa, La Noguera, Catalunya, Espanya). Spira, 4 (3-4): 109-119.
Tarruella, A., Corbella, J., Prats, L l., Guillén, G. & Alba, D. M. 2015. Els moitessièrids (Gastropoda : Moitessieriidae) de Monistrol de Montserrat (el Bages, Catalunya, Espanya). Spira, 5 (4): 163-180.
Vergés i Vernis, P. 1987. Topografía médica de Castellar (San Esteban) o del Vallés. Edició en facsímil de l'obra de 1895. Arxiu d’Història de Castellar. 214 p. Wilke, T. 2019. Moitessieriidae Bourguignat, 1863. P. 122-125. In: Lydeard, C. & Cummings, K. S. (Eds.). Freshwater Mollusks of the World: A Distribution Atlas. Johns Hopkins University Press. Baltimore. 242 p.
Wilke, T., Davis, G. M., Falniowski, A., Giusti, F., Bodon, M. & Szarowska, M. 2001. Molecular systematics of Hydrobiidae (Mollusca: Gastropoda: Rissooidea): testing monophyly and phylogenetic relationships. Proceedings of the Academy of natural Sciences of Philadelphia, 151(1): 1-21.
Wilke, T., Haase, M., Hershler, R., Liu, H-P., Misof, B. & Ponder, W. 2013. Pushing short DNA fragments to the limit: Phylogenetic relationships of «hydrobioid» gastropods (Caenogastropoda: Rissooidea). Molecular Phylogenetics and Evolution, 66 (3): 715-736.
Euophryum rufum (Broun, 1880) nueva cita para Cataluña (Curculionidae: Cossoninae; Pentarthrini)
Euophryum rufum (Broun, 1880) new record for Catalonia (Curculionidae: Cossoninae; Pentarthrini)
Amador Viñolas* & Josep Muñoz-Batet*
* Museu de Zoologia. Laboratori de Natura. Coŀlecció d’artròpodes. Passeig Picasso s/n. 08003 Barcelona.
Autor de correspondencia: Amador Viñolas. A/e: av.rodama@gmail.com
Rebut: 04.11.2024. Acceptat: 09.11.2024. Publicat: 30.12.2024 Butlletí de
El género Euophryum Broun, 1909 esta representado por tres especies, dos de Nueva Zelanda y una de Chile. De las dos especies de Nueva Zelanda se han reportado numerosas introducciones en diferentes países europeos, Euophryum confine (Broun, 1881) es la más extendida y E. rufum (Broun, 1880) se ha citado de Alemania, España, Francia, Gran Bretaña, Irlanda y Suiza (Alonzo-Zarazaga et al., 2017).
Material estudiado
8 ejemplares etiquetados: «barri de Ciutat Vella, Barcelona, Catalunya, 24-VIII-2024, V. Rubio leg.». Depositados en las colecciones de los autores.
Para su determinación se ha seguido la revisión del género realizada por Thompson (1989), pudiendo constatar que dichos ejemplares pertenecen a E. rufum (Fig. 1).
La primera y única cita conocida de E. rufum en la Península Ibérica es la de Bilbao (Alonso-Zarazaga, 1988), con la presente tenemos la segunda del área peninsular y la primera para Cataluña.
Bibliografía
Alonso-Zarazaga, M. A. 1988. Presencia en España de Euophryum rufum (Broun, 1880) (Col., Curc., Cossoninae), una plaga de la madera puesta en obra. Boletín de la Asociación española de Entomología, 12: 361.
Alonso-Zarazaga, M. A., Barrios, H., Borovec, R., Bouchard, P., Caldara, R., Colonnelli, E., Gültekin, L., Hlaváč, P., Korotyaev, B., Lyal, C. H. C., Machado, A., Meregalli, M., Pierotti, H., Ren, L. Sánchez-Ruiz, A., Sforzi, Silfverberg, H., Skuhrovec, J., Trýzna, M., Velázquez de Castro, A. J. & Yunakov, N. N. 2017. Cooperative Catalogue of Palaearctic Coleoptera Curculionoidea. 2nd. Edition. Monografías electrónicas S.E.A., vol. 14, 780 p. Thompson, R. T. 1989. A preliminary study of the weevil genus Euophryum Broun (Coleoptera: Curculionidae: Cossoninae). New Zealand Journal of Zoology,16: 65-79.
DOI: 10.2436/20.1502.01.191

Butlletí de la Institució Catalana d’Història Natural, 88 (4): 179-184. 2024
* Museu de Ciències Naturals de Barcelona. Laboratori de Natura. Coŀlecció d’artròpodes. Passeig Picasso, s/n. 08003 Barcelona. Catalunya. A/e: av.rodama@gmail.com; mederos@gmail.com
** Universidad Autónoma del Estado de Morelos. Av. Universidad 1001, Col. Chamilpa, C.P. 62209. Cuernavaca, Morelos, México
Autor de correspondència: Amador Viñolas, av.rodama@gmail.com
Rebut:14.11.2024; Acceptat: 22.11.2024; Publicat: 30.12.2024
Resum
Se estudian los coleópteros recolectados en dos campañas realizadas en Cuba, la primera en el Parque Nacional Alejandro de Humboldt, provincia de Guantánamo y la segunda en el Parque Nacional Marino de Punta Francés, situado en la Isla de la Juventud. De cada Parque se comenta la geografía, vegetación y climatología. De cada una de las especies recolectadas se aportan los datos conocidos sobre su presencia en la isla y distribución, así como su biología conocida. Se describe una nueva especie de la familia Monotomidae perteneciente al género Europs Wollaston, 1854, acompañada de la representación gráfica del habitus, detalles morfológicos, edeago y mapa de los Parques Nacionales con ubicación de los lugares de recolección.
Palabras clave: Coleoptera, Monotomidae, Europs baracoaensis Viñolas & Mederos n. sp., P. N. Alejandro de Humboldt, Guantánamo, Cuba.
Abstract
A new species of the genus Europs Wollaston, 1854 from Cuba (Coleoptera: Monotomidae: Monotominae: Europini)
Coleoptera collected in two campaigns in Cuba are studied, the first in the Alejandro de Humboldt National Park, Guantanamo province and the second in the Punta Frances National Marine Park, located on Juventud Island. Geography, vegetation and climatology are discussed in each Park. Of each species collected data on the known presence on the island and distribution as well as its known biology are given. A new species of the family Monotomidae belonging to the genus Europs Wollaston, 1854, is described, accompanied by the graphic representation of the habitus, morphological details, aedeagus and map of the National Parks with location of the collection sites.
Key word: Coleoptera, Monotomidae, Europs baracoaensis Viñolas & Mederos n. sp., Alejandro de Humboldt N. P., Guantánamo, Cuba.
Resum
Una nova espècie del gènere Europs Wollaston, 1854 de Cuba (Coleoptera: Monotomidae: Monotominae: Europini)
S’estudien els coleòpters recoŀlectats en dues campanyes realitzades a Cuba, la primera al Parc Nacional Alejandro de Humboldt, província de Guantánamo i la segona al Parc Nacional Marí de Punta Francès, situat a l’illa de la Joventut. De cada Parc es comenta la geografia, vegetació i climatologia. De cadascuna de les espècies recoŀlectades s’aporten les dades conegudes sobre la seva presència a l’illa i distribució, així com la seva biologia coneguda. Es descriu una nova espècie de la família Monotomidae pertanyent al gènere Europs Wollaston, 1854, acompanyada de la representació gràfica de l’habitus, detalls morfològics, edeagus i mapa dels Parcs Nacionals amb ubicació dels llocs de recoŀlecció.
Paraules clau: Coleoptera, Monotomidae, Europs baracoaensis Viñolas & Mederos n. sp., P. N. Alejandro de Humboldt, Guantánamo, Cuba.
urn:lsid:zoobank.org:pub: C420724A-1708-4D22-9F37-6A9C453DD602
Introducción
Se han efectuado una serie de recolecciones en la región oriental de Cuba dentro del proyecto «Colecciones Zoológicas, su conservación y manejo II» del Instituto de Ecología y Sistemática, con el objetivo de contribuir al enriquecimiento de la biodiversidad taxonómica y geográfica de las colecciones zoológicas, así como a la adecuada conservación y manejo de los ejemplares y su información asociada para el desarrollo de estudios taxonómicos sobre la diversidad faunística
DOI: 10.2436/20.1502.01.192
A. VIÑOLAS ET AL.
cubana. El proyecto centró sus esfuerzos de recolección en áreas de reconocida biodiversidad, poco estudiadas o amenazadas por el desarrollo económico social.
El estudio se realizó en el Parque Nacional Alejandro de Humboldt (provincias de Holguin y Guantánamo), en la Meseta de Iberia a orillas del arroyo del Pez Pega, Guantánamo (Fig. 1). El parque es el núcleo de la Reserva de la Biósfera Cuchillas del Toa y cubre un área de 684 km2. Junto con el resto de las Cuchillas de Toa fue declarado Patrimonio Mundial por la UNESCO en el año 2001 y es una de las más importantes reservas de la biósfera en la cuenca del Caribe. Debido a la toxicidad natural de la roca serpentinita que predomina en el área, al gradiente altitudinal, a las abundantes precipitaciones y al largo tiempo de emersión de esta parte de la isla, se alcanza un 70 % de endemismos en su flora. El parque atesora la mayor extensión de bosque húmedo bien conservado del Caribe insular.
La vegetación del parque es una pluvisilva submontana sobre suelo de mal drenaje. El estrato arbóreo es muy denso, con una cobertura entre el 90 y 100 %, su altura es irregular fluctuando entre 8 y 25 m, alcanzando algunos ejemplares los 30 m (Reyes, 2012). Las especies más abundantes que definen la fisonomía arbórea son: Bonnetia cubensis (Britton) Howard, Byrsonima biflora Griseb., Clusia tetrastigma Vesque, Cyrilla nipensis Urb., Podocarpus ekmanii Urb. y Sideroxylon jubilla (Ekman ex Urb.) T.D.Penn. La cobertura del estrato arbustivo fluctúa entre el 10-60 %, siendo más frecuentes los valores del 40 % y 60 %. Este estrato es relativamente pobre en especies constantes, pues sólo Cyathea parvula (Jenman) Domin,, Hieronyma nipensis Urb. y Podocarpus ekmanii Urb. se comportan como tales, sin embargo las más frecuentes son: Bonnetia cubensis (Britton) Howard, Clusia tetrastigma Vesque, Cyrilla nipensis Urb., Ilex hypaneura Loes., Sideroxylon jubilla (Ekman ex Urb.) T.D.Penn. y una especie del género Callicarpa L.
Cuba.
También están presentes, abundantemente, las Euphorbiaceae Pera ekmanii Urb. y Tabebuia dubia (C. Wright ex Sauvalle) Britton ex Seibert, la Melastomataceae Calycogonium grisebachii Triana y la Myrsinaceae Myrsine coriacea (Sw.) R. Br. ex Roem. & Schult., y en menor frecuencia la Arecaceae Bactris cubensis Burret, la Lauraceae Ocotea spathulata Mez, la Melastomataceae Miconia dodecandra Cogn., la Phyllanthaceae Hieronyma nipensis Urb. y la Sapindaceae Matayba domingensis (DC.) Radlk.
En el Parque Nacional Marino de Punta Francés, Isla de la Juventud (Fig. 2), se efectuaron unas recolecciones para el conocimiento de la biodiversidad de la zona financiadas por la Universidad de Oeste de Inglaterra (UWE), de Brístol.
Situado en la parte sur occidental de la Isla de la Juventud y con una extensión de 7,04 km2, de ellos 5,15 en el mar, cuenta con uno de los ecosistemas costeros mejor conservados de Cuba.
La zona terrestre está formada por una llanura de rocas calcáreas con una alta carstificación, donde se localizan numerosas cuevas con agua en su interior y en la línea de la costa farallones costeros con cuevas por donde penetra el mar formando playas subterráneas y lagunas costeras, con una gran variedad de flora y fauna, y playas con dunas de arena fina y blanca cubriendo una extensión de tres kilómetros de longitud. Cuenta con una frondosa vegetación, con más de 500 especies de plantas y 13 de helechos, aproximadamente
105 especies son endémicas de la zona. Su arrecife coralino, de corales duros, y los manglares sirven de refugio, alimentación y anidación a aun gran número de especies. El arbolado dominante está formado por palmerales y manglares, mezclados con ellos se pueden localizar, entre otros, Annona glabra L., Conocarpus erecta L. y Coccoloba uvifera L. en cuyas raíces, cuando llueve, crece el hongo Cantharellaceae Cantharellus cinnabarinu (Schwein.).
La recolección de los ejemplares estudiados se realizó en Punta Francés, en suelo arenoso, la vegetación se puede dividir en dos zonas, por un lado la vegetación propia de la costa arenosa compuesta por la C. uvifera y el Thrinax radiata Lodd. ex Schult. & Schult. junto con herbáceas típicas del hábitat y al otro lado el manglar formado por Avicennia germinans L., C. erecta, Languncularia racemosa (L.) C.F. Gaertn. y Rhizophora mangle L., con presencia de Bursera simaruba (L.) Sarg. y Metopium brownei (Jacq.) Urb.
Material y métodos
En el Parque Nacional Alejandro de Humboldt se han realizado diversas colectas con metodologías diferentes, empleando también trampas Malaise. Al no existir especialistas foráneos en muchos grupos los resultados de las recolecciones han permanecido ignorados. Los trabajos más completos en el conocimiento de varios grupos zoológicos y botánicos forman parte de los llamados «Inventarios Biológicos Rápidos» realizados con la participación del Field Museum de Chicago y del Centro Oriental de Ecosistemas y Biodiversidad (BIOECO) de Cuba.
Las trampas se colocaron el 28 de octubre de 2015 y se mantuvieron en el lugar 72 horas. Durante las primeras 42
horas el tiempo alternó entre soleado y ligeramente nublado, en las últimas 24 horas estuvo casi siempre nublado y llovió frecuentemente, con especial intensidad durante la noche.
En la Isla de la Juventud en Punta Francés, a nivel del mar, se colocaron diversas trampas Malaise durante el mes de marzo de 2012 y por periodos que oscilaron entre las 24 y 48 horas. El material estudiado corresponde a una trampa Malaise colocada en la zona el 30 de marzo y mantenida en el lugar por un periodo de 40 horas.
En los ejemplares estudiados se ha procedido a la extracción del edeago o de la espermateca, según el sexo, los cuales después de un tratamiento de limpieza y extracción del aire se montaron en preparación microscópica sobre una lámina de polistireno transparente, de la marca Evergreen®, con líquido DMFH. Las fotografías se realizaron con una cámara Canon® modelo EOS 760D, con objetivo de microscopía y por el método de capas, con tratamiento de las imágenes mediante el programa Zerene Stacker®. Las coordenadas de recolección y los mapas de los parques naturales se han obtenido mediante el programa Google Earth© de Google Inc.
Del muestreo realizado en el Parque Nacional Marino de Punta Francés, Isla de la Juventud, hemos podido estudiar las siguientes especies:
Familia Bostrichidae Latreille, 1802
Subfamilia Bostrichinae Latreille, 1802
Tribu Apatini Billberg, 1820
Apate monachus Fabricius, 1775
2 ♀. Especie africana, introducida por la actividad humana en numerosos países y citada de Cuba (Peck, 2005).
Tribu Xyloperthini Lesne, 1921
Xylomeira tridens (Fabricius, 1792)
1 ♂. Conocido de las Antillas occidentales, Méjico y Estados Unidos, citada de Cuba de Los Indios, Isla de la Juventud.
Familia Nitidulidae Latreille, 1802
Subfamilia Nitidulinae Latreille, 1802
Tribu Nitidulini Latreille, 1802
Stelidota ruderata Erichson, 1843
1 ♂ y 2 ♀. Especie conocida de Cuba, Hispaniola, Puerto Rico y Guadalupe. Peck (2005) de Cuba no da localidades precisas, aunque indica una amplia distribución
Familia Coccinellidae Latreille, 1807
Subfamilia Coccinellinae Latreille, 1807
Tribu Diomini Gordon, 1999
Diomus roseicollis (Mulsant, 1853)
1 ♂ y 1 ♀. Especie citada de Cuba, Puerto Rico, Antillas Menores y Florida (Estados Unidos). Peck (2005) de Cuba la
cita de Nueva Gerona, Isla de la Juventud.
Familia Curculionidae Latreille, 1802
Subfamilia Scolytinae Latreille, 1804
Tribu Cryphalini Lindemann, 1877
Hypothenemus gossypii (Hopkins, 1915)
1 ♂. Conocido de Cuba, Méjico y Florida (Estados Unidos). De Cuba se ha citado de Cayamas, localidad típica de la especie (Peck, 2005).
Tribu Xyleborini LeConte, 1876
Xylosandrus compactus (Eichhoff, 1875)
1 ♀. Especie nativa de África, introducida en el continente Americano y citada del Brasil, Islas Vírgenes, Estados Unidos (Florida y Texas) y Cuba. Ampliamente extendida en Cuba en donde es plaga de la variedad robusta de cafetos (Peck, 2005).
Del Parque Nacional Alejandro de Humboldt se ha podido estudiar la siguiente especie que es nueva para la ciencia.


A. VIÑOLAS
Familia Monotomidae Laporte, 1840
Subfamilia Monotominae Laporte, 1840
Tribu Europini Sen Gupta, 1988
Europs baracoaensis Viñolas & Mederos n. sp.
urn:lsid:zoobank.org:pub:6F9FCC59-A37E-4D14-B7CDCD7EE8473525
Diagnosis
El primer esternito abdominal con el surco de las coxas triangular lo aleja de E, striatulus Fall & Fall, 1907 y de su grupo (Fig. 4a), a parte de otros caracteres externos; separado también de E., sulcicollis Bousquet, 2003 porque la gruesa puntuación de la superficie del protórax no esta alineada. La puntuación pronotal y el color de cuerpo lo separan de las otras especies conocidas de Florida y México (Bousquet, 2003).
Descripción
Longitud de 2,17 a 2,31 mm. Cuerpo de contorno paralelo y poco convexo de color castaño rojizo, con los élitros amarillento con dos manchas suturales negruzcas (Fig. 3a, b). Cabeza grande tan ancha como el protórax; antenas de 10 artejos, con maza terminal de dos, el noveno muy transverso y el décimo redondeado, con las sienes anchas y salientes (Figs. 3a y 4b); los ojos redondeados, no muy grandes y salientes; surco gular muy marcado y profundo (Fig. 4c); superficie con el punteado disperso y tan grande como el del protórax (Fig. 4b). Protórax ligeramente transverso, 1,06 veces más ancho que largo, con su mayor anchura en el ápice, tan ancho como los élitros; con los márgenes ligeramente curvados y entrantes hacia la base (Fig. 4b) y provistos en la base de dos pequeños dientes; ángulos anteriores subrectos, muy marcado, los posteriores redondeados; superficie con fuerte punteado, igual al de la cabeza, dejando una zona media libre. Élitros cortos (Fig. 3a), 1,67 más largos que anchos tomados conjuntamente, de contorno paralelo y con su mayor anchura en la parte media, de color amarillento con una mancha negruzca triangular en la base y una mayor redondeada en el ápice, dejando libre el pigidio; húmeros angulosos y salientes hacia delante; Estrías de puntos gruesos muy marcadas y completas. Prosternón con una serie de pliegues horizontales en ápice (Fig. 4c); mesosternón muy estrecho; metasternón con fuerte punteado marginal. Abdomen con el primer esternito (Fig. 3b) ancho tan largo como los tres siguientes, que tienen la misma anchura, el último más desarrollado; superficie con punteado menos marcado y disperso. Coxas separadas. Patas gráciles. Edeago tipo cápsula (Fig. d), con la parte apical provista de pequeños dientes irregularmente dispersos; no se ha podido comparar con el edeago de las especies americanas y de México al no encontrar representaciones de los mismos.
Hembra
Desconocida, ya que los dos ejemplares estudiados son machos.
.Material estudiado
Holotipo
1 ♂, etiquetado: «P. N. Alejandro de Humboldt | Arroyo Pez Pega | Meseta de Iberia, Baracoa | Guantánamo. Cuba | O. C. Bello González leg.» «Malaise trap, 580 m | 28/30X-2015 | 72 horas | 20º28’31”N 74º43’46”O» «Holotipo | Europs | baracoaensis n. sp. | A. Viñolas & J. Mederos det. 2016».
Paratipo
1 ♂, etiquetado: «P. N. Alejandro de Humboldt | Arroyo Pez Pega | Meseta de Iberia, Baracoa | Guantánamo. Cuba | O. C. Bello Gonzalez leg.» «Malaise trap, 580 m | 28/30-X2015 | 72 horas | 20º28’31”N 74º43’46”O» «Paratipo | Europs | baracoaensis n. sp. | A. Viñolas & J. Mederos det. 2016». Holotipo y paratipo depositados en la colección de A. Viñolas de Barcelona.
Etimología
El nombre hace referencia a la localidad en la que se han colectado los ejemplares, Baracoa, provincia de Guantánamo, Cuba.
Biología
No se tienen datos sobre su biología, aunque se supone sigue el régimen alimentario del resto de especies del género asociadas a la vegetación del entorno.
Distribución
Sólo se conoce de la localidad típica de Arroyo Pez Pega, situada en la Meseta de Iberia término municipal de Baracoa, provincia de Guantánamo, Parque Nacional Alejandro de Humboldt, Cuba (Fig. 1).
Agradecimientos
A Norvis Hernández y al resto del personal vinculado a la conservación en el Parque Nacional Alejandro de Humboldt, especialmente en el sector Baracoa. A Alejandro Barro Cañamero del Departamento de Biología Animal y Humana de la Facultad de Biología de la Universidad de La Habana, la cesión para estudio de los coleópteros recolectados, en una de sus trampas, en los estudios realizados sobre la biodiversidad en la Isla Juventud de Cuba, durante el año 2012.
Bibliografía
Bousquet, Y. 2003. Review of the genus Europs Wollaston (Coleoptera: Monotomidae) of America, north of Mexico. Pan-Pacific Entomologist, 79: 11-22.
Peck, S. B. 2005. A Checklist of the Beetles of Cuba with Data on Distributions and Bionomics (Insecta: Coleoptera). Arthropods of Florida and Neighboring Land Areas. Volumen 18. Florida Department of Agriculture and Consumer Services. Gainesville. 241 p.
Reyes, O. J. 2012. Clasificación de la vegetación de la Región Oriental de Cuba. Revista del Jardín Botánico Nacional, 32-33 (2011-2012): 59-71.
Butlletí de la Institució Catalana d’Història Natural, 88 (4): 185-192. 2024
Samay Bravo-Cuautle1, Rosa D. García-Martiñón2, Uriel M. Barrera-Ruiz3, Betzabeth C. Pérez-Torres1, Agustín Aragón-García1, Armando Equihua-Martínez2, Edith G. Estrada-Venegas2, Mónica Rangel-Villafranco4, Silvia Romero-Rangel5, Víctor Cuesta-Porta6 & Juli Pujade-Villar6
1 Benemérita Universidad Autónoma de Puebla. Instituto de Ciencias. Centro de Agroecología, Maestría en Manejo Sostenible de Agroecosistemas. Edificio VAL 1. Km 1.7 Carr. a San Baltazar Tetela, San Pedro Zacachimalpa. 72960 Puebla, México. A/e: samay.bravocuatle.icuap@viep.com.mx, betzabeth. perez@correo.buap.mx, agustin.aragon@correo.buap.mx
2 Colegio de Postgraduados de Montecillo. Posgrado en Fitosanidad, Entomología y Acarología. Carretera México-Texcoco, Km 36.5. 56230 Montecillo, Estado de México (México). A/e: rosa.garcia@colpos.mx, equihuaa@colpos.mx, estradae@colpos.mx
3 Universidad Autónoma Chapingo. División de Ciencias Forestales. Carretera Federal México-Texcoco Km 38.5. 56230 Montecillo. Estado de México (México). A/e: barrera.uriel@colpos.mx
4 Universidad Intercultural del Estado de México. Laboratorio de socioecosistemas para la sustentabilidad Desarrollo sustentable Lib. Francisco Villa S/N, Col. Centro. 50640 San Felipe del Progreso, México. A/e: monica.rangel@uiem.edu.mx
5 Universidad Nacional Autónoma de México. Facultad de Estudios Superiores Iztacala. Laboratorio de Ecología y Taxonomía de Árboles y Arbustos. Avda. de los Barrios, 1. Los Reyes Iztacala. Tlalnepantla de Baz, Estado de México. A/e: sromero@unam.mx
6 Universitat de Barcelona. Facultat de Biologia. Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals. Avda. Diagonal, 645. 08028 Barcelona. Cataluña. A/e: victorcp93@gmail.com, jpujade@ub.edu
Autor de correspondencia: Juli Pujade-Villar. A/e: jpujade@ub.edu
Rebut: 17.11.2024; Acceptat: 25.11.2024; Publicat: 30.12.2024
Resumen
Se comparan las agallas de Plagiotrochus quercusilicis del paleártico occidental y de Neuroterus fusifex de México, como un ejemplo de convergencia evolutiva. Se menciona por primera vez que las agallas de N. fusifex pueden encontrarse en las hojas. Se citan nuevos huéspedes para N. fusifex y se amplía su área de distribución. Se exponen nuevos datos de la biología de N. fusifex
Palabras clave: Hymenoptera, Cynipini, Neuroterus, Plagiotrochus, agallas, convergencia evolutiva.
Abstract
A case of evolutionary convergence in galls of Old and New World cynipids (Hymenoptera Cynipidae)
The galls of Plagiotrochus quercusilicis from the Western Palearctic and Neuroterus fusifex from Mexico are compared, as an example of evolutionary convergence. It is mentioned for the first time that the galls of N. fusifex can be found in the leaves. New hosts are found for N. fusifex and its distribution area is mentioned. New data on the biology of N. fusifex are presented.
Key words: Hymenoptera, Cynipini, Neuroterus, Plagiotrochus, galls, evolutionary convergence.
Resum
Un cas de convergència evolutiva de gales de cinípids del vell i nou món (Hymenoptera Cynipidae)
Es comparen les gales de Plagiotrochus quercusilicis del Paleàrtic occidental i de Neuroterus fusifex de Mèxic, com un exemple de convergència evolutiva. Es menciona per primera vegada que les gales de N. fusifex poden trobar-se a les fulles. Se’n citen nous hostes per a N. fusifex i s’amplia la seva àrea de distribució. S’hi exposen noves dades de la biologia de N. fusifex.
Paraules clau: Hymenoptera, Cynipini, Neuroterus, Plagiotrochus, gales, convergència evolutiva.
Introducción
Los Cynipidae incluyen una amplia gama de himenópteros inductores de agallas y de inquilinos de algunas de éstas, aunque también se han citado ocasionalmente como inquilinos en agallas de Cecidomyiidae (Lobato-Vila & PujadeVillar, 2019; Lobato-Vila et al., 2022; Nastasi et al., 2024). La tribu Cynipini induce agallas en diversos géneros de Fagaceae, principalmente en Quercus L. (Pujade-Villar et al., 2001; Stone et al., 2002, 2009; Ronquist et al., 2015). Como
S. BRAVO-CUAUTLE
fitófagos siguen la distribución de su planta hospedadora por lo que los Cynipini se encuentran principalmente en la zona templada del hemisferio norte (región holártica), extendiéndose a áreas tropicales de América (neotrópico) y el sudeste asiático (región oriental). Agrupa más de 1.000 especies en todo el mundo y son, con diferencia, la tribu más prolífica de la familia (Melika & Abrahamson, 2002; Csóka et al., 2005; Ronquist et al., 2015; Pénzes et al., 2018; Buffington et al., 2020; Fang et al., 2020).
Las agallas se pueden encontrar en cualquier órgano de la planta (hojas, yemas, flores, bellotas, ramas, ramitas o raíces). Algunas agallas son específicas de una determinada región del órgano (haz o envés de las hojas, flores masculinas o femeninas, cúpula o nuez de bellota) y raramente pueden presentarse en dos órganos distintos.
En lo que se refiere a la biología, la mayoría los Cynipini experimentan ciclos de vida complejos con la alternancia de generaciones, donde existe una sucesión obligada entre una generación sexual y otra asexual (Pujade-Villar et al., 2001; Stone et al., 2002; Csóka et al., 2005), ya sea en la misma sección de Quercus (heterogonia), o que cada generación produzca agallas en una sección distinta de Quercus (heteroecia). La generación sexual suele emerger durante el verano y principios del otoño, y la generación asexual pasa el invierno dentro de la agalla y emerge en primavera. Para cada generación, la mayoría de las especies exhiben diferencias notables en la morfología, tanto de los adultos como de las agallas (Pujade-Villar et al., 2001; Stone et al., 2002; Csóka et al., 2005), y cada generación induce agallas habitualmente en diferentes órganos de la planta hospedadora.
Algunas agallas son características de una especie o género, pero muchas de sus morfologías han aparecido repetidamente de forma independiente varias veces a lo largo de la filogenia de estas avispas (Stone et al., 2002, Csóka et al., 2005). El Paleártico occidental es el área con las relaciones más inequívocas entre agallas y adultos, ya que observando la agalla sabemos de qué especie/generación se trata, lo cual probablemente se debe a una menor diversidad de los robles. En el Paleártico oriental y sobre todo en el Neártico existe una mayor diversidad de robles, y algunas morfologías de agallas son similares entre géneros, lo que dificulta la identificación de especies de avispas gallícolas (Stone et al., 2002, Csóka et al., 2005). Así, por ejemplo, en América, casi todas las especies de Atrusca Kinsey, 1930, producen agallas similares o idénticas (agallas esféricas frágiles con la cámara larval sujeta con fibras irradiantes), las agallas pubescentes de Striatoandricus Pujade-Villar, 2020, pueden confundirse con algunas agallas de especies del género Druon Kinsey, 1937, y en la mayoría de especies de los Andricus Hartig, 1840, tumorales sucede lo mismo que en Atrusca, por lo que sin la obtención del adulto es imposible saber de qué especie gallícola se trata.
El mecanismo de formación de las agallas aún es desconocido, aunque recientemente se ha descartado que sea debida a una relación simbiótica entre un virus y el himenóptero (Hearn et al., 2019). Además, Hearn et al. (2019) afirman que en Biorhiza pallida (Olivier, 1791) muchos de los genes ex-
Butlletí ICHN 88 (4), 2024
presados en las larvas jóvenes tendrían señales peptídicas de secreción, que son exportadas a los tejidos vegetales como parte del sistema de inducción, pudiendo actuar sobre algunas proteínas del hospedador para inducir un estado similar a la embriogénesis somática. Markel et al. (2024) mencionan que los cinípidos pueden cambiar radicalmente el perfil de metabolitos y reestructurar la pared celular para permitir la formación de agallas, proporcionando información sobre el mecanismo de inducción de éstas; sin embargo, no queda claro hasta qué punto las plantas pueden reprogramarse por completo para formar las agallas. Asimismo, Chen (2024) compara el perfil metabólico de dos especies de agallas de hojas de roble morfológicamente distintas inducidas por cinípidos y concluye que este perfil es diferente, por lo que cada forma de agalla tendría que tener su propio perfil metabólico que defina cada morfología de la agalla.
La convergencia evolutiva se da cuando dos estructuras similares han evolucionado independientemente a partir de estructuras ancestrales distintas por procesos de desarrollo muy diferentes. El desarrollo de estructuras similares en organismos no emparentados puede ser el resultado de la adaptación a ambientes parecidos y/o formas de vida semejantes (Fontdevila & Moya, 2003). Las estructuras similares que evolucionaron por convergencia se denominan análogas mientras que las estructuras homólogas son semejantes debido a una ascendencia evolutiva común.
La evolución convergente puede ocurrir por dos causas principales (Gómez Pompa et al., 1980): condiciones similares en el entorno o por presentar un nicho compartido. En el primer caso, los organismos que tienen estructuras análogas y que no están filogenéticamente emparentados, las desarrollan para poder adaptarse en un hábitat determinado; en el segundo, dos especies se adaptan a un nicho particular, aunque tengan orígenes diferentes.
El caso que presentamos es Plagiotrochus quercusilicis (Fabricius, 1798), especie que forma agallas en el paleártico occidental en la sección Cerris versus Neuroterus fusifex Pujade-Villar & Ferrer-Suay, 2016, del Neártico con agallas en la sección Quercus. Ambas especies producen agallas pluriloculares similares en hojas y amentos, y adultos cromáticamente parecidos, a pesar de pertenecer a géneros distintos y producir agallas en secciones que Quercus también distintas.
El material fue colectado por los autores en distintas anualidades. Las agallas fueron preservadas en el laboratorio hasta la obtención de los adultos, los cuales fueron depositados en alcohol absoluto. Se conservaron tanto las agallas como los adultos para su estudio posterior. Los adultos fueron examinados bajo la lupa binocular y parte de ellos se usó para un análisis molecular.
El análisis molecular se realizó con ocho individuos obtenidos de diferentes muestras de agallas de hojas y amentos recolectados entre 2014 y 2023, incluyendo un espécimen del material tipo de N. fusifex. El ADN fue extraído usando el DNeasy Blood & Tissue. Cada mezcla para la reacción en

Figura 1. a) Agallas jóvenes de Plagiotrochus quercusilicis en hojas de Q. ilex (imagen cedida de Biodiversidad virtual, autor: Francisco Rodríguez). b-c) Agallas maduras de P. quercusilicis sobre Q. coccifera (imágenes cedidas por Sisco Guasch depositadas en Biodoversidad virtual). d) Agallas de Neuroterus fusifex en hojas de Q. liebmanii (imagen: S. Bravo-Cuautle). e-f) Agallas de N. fusifex en hojas y amentos de Q. laeta (imagen: U.M. Barrera-Ruiz). g) Agallas de N. fusifex en hojas de Q. laeta (Foto R. D. García-Martiñón). h) Agallas de N. fusifex en amentos de Q. liebmanii (imagen: S. Bravo-Cuautle). i) Agallas de N. chinanteco sobre Q. macdougallii (imagen: R. Clark-Tapia).
cadena de la polimerasa (PCR) consistió en Taq polimerasa (MyTaq™ DNA Polymerase) 0.2 μL, MyTaq™ Red Reaction Buffer 0.4 μL, primers 0.4 μL a 10 μM y 2 μL de ADN, y se completó a 20 μL con agua. Las condiciones de los ciclos fueron 94 °C durante 5 minutos, seguidas de 35 ciclos de 94 °C durante 30 segundos, 45 °C durante 35 segundos y 72 °C durante 45 segundos, con un paso final de 72 °C durante 5 minutos. Los primeros utilizados para el gen mitocondrial del citocromo oxidasa I (COI) fueron: L1490 y H2198 (Fol-
mer et al. 1994). La secuenciación se llevó a cabo en Macrogen utilizando un secuenciador ABI3730XL DNA y el método de Sanger. Las secuencias fueron alineadas manualmente con el software MEGA 11 (Tamura, et al. 2021), y con el mismo programa se obtuvieron las distancias por pares de las secuencias.
Para explorar y visualizar las relaciones entre muestras en función de sus similitudes genéticas, se realizó un análisis de Coordenadas Principales (PCoA) usando el software
S. BRAVO-CUAUTLE
R (R Core Team, 2019) y la librería «vegan» (Oksanen et al., 2020). Se utilizó el método cmdscale con el argumento eig = TRUE para extraer los valores propios, que representan la varianza explicada en cada dimensión.
Las fotografías que ilustran este estudio fueron realizadas por los autores o bien, fueron cedidas por Francisco Rodríguez (Fig. 1a) y Sisco Guasch (Figs. 1b-c), las cuales están depositadas en Biodiversidad Virtual (https://www.biodiversidadvirtual.org/).
Plagiotrochus quercusilicis fue descrita formando agallas en hojas de un Quercus arbustivo y perennifolio, Q. coccifera L. (subgénero Cerris, sección Ilex), probablemente atacando el nervio principal de la hoja, pero al ser la hoja de pequeño tamaño y la agalla plurilocular, esta agalla ocupa toda la superficie foliar (Figs. 1a-b). Más tarde se describió Plagiotrochus fusifex Mayr, 1882, induciendo agallas en los amentos también en Q. coccifera (Fig. 1c). Ambas especies también pueden inducir agallas en Q. ilex L. (subgénero Cerris, sección Ilex), pero en este caso las agallas de las hojas presentan una superficie pubescente (Fig. 1a), al ser la hoja de Q. ilex pubescente. De estas dos especies se describen hasta ocho variedades o subespecies atendiendo a diferencias cromáticas de los adultos (Dalla Torre & Kieffer, 1910; Tavares, 1926). Plagiotrochus quercusilicis, P. fusifex y todas sus subespecies fueron sinonimizadas por Pujade-Villar & Ros-Farré (1998) bajo el nombre P. quercusilicis
Neuroterus fusifex fue descrita de México formando agallas en los amentos (Fig. 1f) del encino caducifolio Q. laeta Liebm. (subgénero Quercus, Sección Quercus, subsección Leucomexicanae, Grupo Glaucoideae). El epíteto ‘fusifex’ hacía referencia a que las agallas eran indistinguibles de la especie del paleártico P. quercusilicis que atacaba amentos (Pujade-Villar et al., 2016). La única diferencia era la coloración, las agallas mexicanas no siempre viran a rojizas (Figs. 1f, h), mientras que las agallas del paleártico eran verdes cuando eran inmaduras virando a rosa o rojizo en la madurez (Figs. 1b-c). Aunque aún no se ha demostrado su eficacia, muchas agallas cambian de color durante el desarrollo, normalmente de verde a rojo (Stone et al., 2002). Se desconoce hasta qué punto la larva del cinípido controla la síntesis de
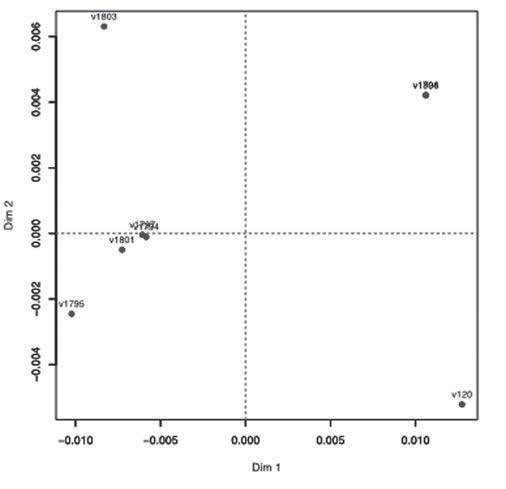
2. PCoA de las similitudes genéticas de las muestras de agalla (ver fig. 1 para la determinación numérica de las muestras).
pigmentos, pero hay evidencias de que las hembras de los parasitoides reconocen el color y lo utilizan para evaluar la calidad del hospedador (Askew, 1961; Czeczuga, 1977). De esta forma, cuando las larvas son muy jóvenes las agallas son verdes, por lo que se ha sugerido que la coloración podría disuadir el parasitoidismo en algunas etapas del desarrollo larvario (Pujade-Villar et al., 2024). Por otro lado, el color rojizo indica que existe una gran cantidad de taninos concentrados en la agalla. Los taninos son generalmente toxinas que reducen significativamente el crecimiento y la supervivencia de muchos herbívoros cuando se añaden a su dieta, ya que pueden inactivar algunas enzimas digestivas, por lo que las plantas con agallas ricas en taninos (rojas) son menos consumidas por los herbívoros (Margalef, 1977).
Recientemente hemos detectado en México agallas en la nerviación principal de las hojas de Q. laeta y Q. liebmanii Oerat. ex Trel., de las que se obtenían adultos aparentemente idénticos a los de las agallas en los amentos (N. fusifex, Figs. 1d-e, g). Las distancias genéticas entre las muestras analizadas se encuentran entre 0–2,5 % con una distancia de
1. Distancias por pares del gen de la citocromo oxidasa I (COI) de las muestras de Neuroterus. La primera columna indica el órgano vegetal donde se encontró la agalla de la que emergieron las muestras, la segunda (ID) corresponde al código de muestra (UB) y al código del woucher.
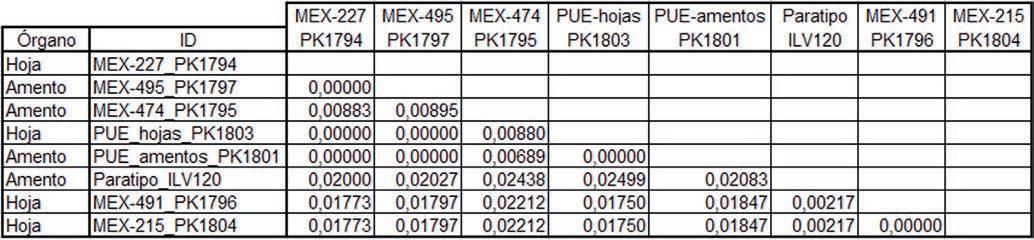
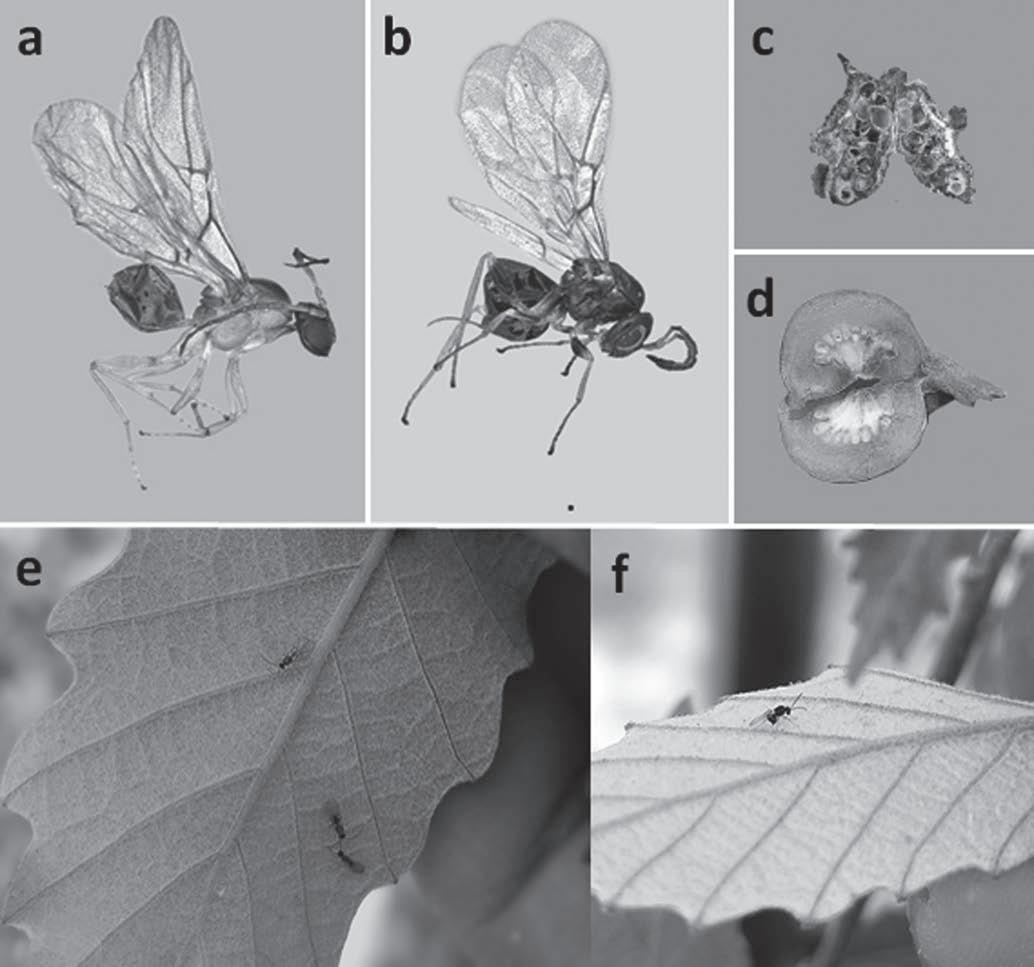
por S. Bravo-Cuautle].
1,2 % de media (Tabla 1). Las muestras se organizan en dos subgrupos distanciados por más de 1,75 % de distancia entre subgrupos y con baja distancia interna (siempre inferior al 1 %; Tabla 1). Cada uno de los subgrupos contiene muestras de agallas en hojas y en amentos, con lo que el órgano vegetal atacado no parece seguir ninguna estructura genética. Los resultados apuntan a que ambas morfologías de agallas pluriloculares (Figs. 3c-d) corresponden a la misma especie. Además, el análisis de PCoA muestra una varianza total explicada baja en las dos primeras dimensiones (Dim 1: 0.0006, Dim 2: 0.0001), lo que sugiere que no hay diferencias genéticas entre las muestras. A pesar de que se observan agrupamientos en los cuadrantes (Fig. 2), estos son mínimos
y limitados en comparación con la varianza total. En resumen, estos datos también apoyan que las muestras estudiadas constituyen una única especie desde un punto de vista de la variación de la varianza.
Como las hojas de los robles (Q. laeta y Q. liebmanii) son de mayor tamaño en comparación con las de Q. coccifera, no desaparece el limbo foliar por la presencia de la agalla (Figs. 1d-e, g), a no ser que sea atacada en sus fases muy iniciales de desarrollo (Fig. 1d), como sí sucede con P. quercusilicis, sino que se produce una doblez cóncava (Fig. 1d, g). Por otro lado, en la especie mexicana (N. fusifex) el color rojizo en la agalla no se produce siempre a diferencia de la especie paleártica occidental (P. quercusilicis).
En definitiva, nos encontramos con dos especies de Cynipini que ocasionan agallas idénticas en los amentos y en las hojas. Estas especies pertenecen a dos géneros distintos (Plagiotrochus Mayr, 1881 y Neuroterus Hartig, 1840), los cuales filogenéticamente están situados en clados distintos (Nicholls, per. com.). Además, atacan Quercus de secciones distintas. Esto tiene implicaciones muy interesantes si se considera el estudio de Chen (2024) mencionado anteriormente, ya que el perfil metabólico de dos especies, a pesar de ser filogenéticamente distantes, ha de ser muy parecido para inducir agallas tan similares en los mismos dos tipos de órganos, en encinos de secciones diferentes y en áreas geográficas distintas.
Por todo lo mencionado, nos encontramos con un caso extraordinario de convergencia evolutiva, donde dos especies de distintos géneros producen dos modelos de agallas idénticas en las hojas y en los amentos. Por otro lado, ambas especies pertenecen a las formas sexuadas de las que se desconoce la forma asexuada. Será interesante saber algún día cual es la forma asexuada de cada una de ellas por si también las agallas asexuales fuesen coincidentes en forma y órgano atacado.
Además, los adultos de N. fusifex se caracterizan por presentar machos ambarinos y hembras oscuras (Fig. 3a-b, e-f). Este tipo de cromatismo es coincidente con alguna de las subespecies invalidadas de P. quercusilicis (= fusifex) colectadas sobre Q. ilex, en las que las hembras son más oscuras que los machos. Por lo tanto, el cromatismo de los adultos también podríamos considerarlo como una convergencia evolutiva más entre P. quercusilicis y N. fusifex
Finalmente mencionar que existe otra especie en México recientemente descrita, Neuroterus chinanteco Pujade-Villar & Clark, 2023 (Pujade-Villar et al., 2023), correspondiente a una generación sexual que también produce agallas pluriloculares en las hojas, pero en un encino endémico de Oaxaca y en peligro de extinción, Q. macdougallii Martínez (sección Quercus, Serie Leucomexicanae) cuyas agallas (Fig. 1i) recuerdan las de las especies mencionadas; no provocan la torsión de la hoja como sucede con N. fusifex, ni presentan una coloración roja como ocurre en P. quercusilicis y a veces en N. fusifex, por lo que podríamos considerarla como una fase intermedia entre los dos modelos de agallas mencionados, en el que los machos también son amarillentos y las hembras oscuras. En este caso, no hemos encontrado agallas en los amentos.
Material estudiado de N. fusifex
Jardín Botánico, Ciudad Universitaria de la Benemérita (Universidad Autónoma de Puebla, México), Q. liebmannii en amentos, (20.iii.2024) 22.iii.2º24: 17 ♂ & 13 ♀ (3 ♂ & 3 ♀* JP-V col.) (S. Bravo-Cuautle leg.); mismos datos agallas en hojas: 18 ♂ & 12 ♀ (2 ♂ & 2 ♀* JP-V col.) (S. BravoCuautle leg); mismos datos (29.vi.24) 30.vi.24: 2 ♂ & 1♀ (1 ♂ & 1 ♀ JP-V col.) (JP-V & S. Bravo-Cuautle leg); Q. glaucoides en hojas (29.vi.2024) 30.vi.2014: 1 ♂ & 1 ♀ (JP-V col.) (JP-V & S. Bravo-Cuautle leg). Plateros-arenales (Es-
(4),
tado de México), Q. laeta en amentos, (19.iv.2014) 25-30. iv.2014: 18 ♂ & 55 ♀ (18 ♂ & 30 ♀* JP-V col., MEX-227) (García-Martiñón leg., 61N y 64N); mismos datos, Q. laeta en hojas, (19.x.2013) 23.i.2014: 3 ♀ (2 ♀ JP-V col., MEX226) (García-Martiñón leg., 33BN y 60N); Q. glabrescens en hojas, (19.iv.2014) 14-21.iv.14: 8 ♀ (5 ♀* JP-V col., MEX215) (García-Martiñón leg., 76N); mismos datos, Q. glabrescens en hojas, (21.vi.2014) 24.vi.2014: 3 ♀ (3 ♀ JP-V col., MEX-224) (García-Martiñón leg., 69N); mismos datos, (12.V.2014) 20-31.VII.2014 1 ♀ (JP-V col., MEX-226) (leg. García-Martiñón, 77N). Sta. Fe (Ciudad de México), Q. laeta en amentos, (25-iii.2019) iii 19: 103 ♂ & 272 ♀ (CibriánTobar y Barrera-Ruiz leg.); mismos datos, 8.iv.19: 3m & 3f (2 ♂ & 2 ♀ JP-V col, MEX-474a); mismos datos (27.iii.19) iv.19: 16 ♂ & 17 ♀* (JP-V col., MEX-495) (Barrera-Ruiz leg.); mismos datos, 27.iv.19: 5 ♂ 7 ♀ (3 ♂ & 4 ♀* JP-V col., MEX-474) (Barrera-Ruiz leg.). Rancho Concepción Chico (Comunidad Mazahua), San Felipe del Progreso (Estado de México), Q. obtusata amentos, (22.vi.2023) 22-30.vi.2023: 1 ♀ (JP-V col., MEX-685) (M. Rangel, J. Domingo & JP-V leg). Ctra. Federal Morelia-Naravatio desviación Yerbabuena km. 12 (Michoacán), Q. laeta en hoja, (14.vi.19) 14-30.vi.19: 3 ♂* (JP-V col., MEX-491) (A. Equihua, E. Estrada-Venegas & JP-V leg.). Miahuatlán de Porfirio Díaz (Oaxaca),Q. obtusata hoja, (05.iii.2018) 20.v.2018: 3 ♂ & 5 ♀ (JP-V col., MEX-541) (R. Clark leg.); Q. obtusata amento, (05.iii.2018) 20.v.2018: 2 ♂ & 3 ♀ (JP-V col., MEX-532) (R. Clark leg.).
UNSIJ (Universidad de la Sierra Juárez), Ixtlán de Juárez (Oaxaca), Q. glaucoides amento, (10.v.2023) 15.v.2023: 7 ♂ (JP-V col., MEX-761) (R. Clark leg.).
NOTA: el «*» de algunas de las muestras estudiadas indica que se han utilizado ejempares para el estudio molecular
Atendiendo a los datos del material estudiado, y según lo mencionado en Martínez-Romero et al. (2022), se amplía la distribución de N. fusifex en México, mencionando esta especie por primera vez en los Estados de Michoacán y Puebla. Además, se cita esta especie por primera vez en Q. liebmannii, Q. glabrescens y Q. glaucoides (pertenecientes a la sección Quercus). También informamos que hemos colectado agallas parecidas en los amentos de dos especies de encinos de la sección Lobatae, pero hasta obtener y examinar los adultos que emergan preferimos no adjudicar estas colectas a N. fusifex
Las formas asexuales de estas especies son desconocidas, pero se ha observado que las hembras sexuadas de N. fusifex ovipositan en el limbo foliar de Q. liebmannii (Fig. 3f).
Agradecimientos
Este estudio ha sido financiado por «PID2021-128146NB-I00/MCIN/AEI/10.13039/501100011033/» y «FEDER una manera de hacer Europa». Agradecemos muy sinceramente a nuestro amigo Sisco Guasch, recientemente fallecido, el habernos cedido las imágenes de las agallas en las hojas y amentos de P. quercusilicis sobre Q. coccifera (Figs. 1b-c) depositadas en Biodiversidad Virtual (https://www.bio-
diversidadvirtual.org/). También a Francisco Rodríguez por cedernos la imagen de las agallas jóvenes de P. quercusilicis sobre Q. ilex (Fig. 1a) depositada en Biodiversidad Virtual. Así mismo, a Jordi Clavell por los trámites para la cesión de Imágenes de Biodoversidad Virtual. Finalmente, los autores agradecen a CONAHCyT, por el apoyo económico brindado al primer autor para realizar la Maestría en Manejo Sostenible de Agroecosistemas, en el Centro de Agroecología, del Instituto de Ciencias, de la Benemérita Universidad Autónoma de Puebla.
Askew, R. R. 1961. A study of the biology of the species of the genus Mesopolobus Westwood (Hymenoptera: Pteromalidae) associated with cynipid galls on oak. Transactions of the Royal Entomological Society of London, 113: 155-168.
Buffington, M. L., Forshage, M., Liljeblad, J., Tang, C.-T. & van Noort, S. 2020. World Cynipoidea (Hymenoptera): A key to higher-level groups. Insect Systematics and Diversity, 4 (4): 1. https://doi.org/10.1093/isd/ixaa003
Chen, J. 2024. It’s gall relative: Metabolic profiling of two morphologically distinct oak leaf galls induced by cynipid wasps. Plant Physiology, 195 (1): 248-250. https://doi.org/10.1093/ plphys/kiae032
Csóka, G., Stone, G. N. & Melika, G. 2005 The biology, ecology and evolution of Gall-inducing Cynipidae. P. 573-642. In: Raman C, Schaefer W, Withers TM, editors. Biology, ecology and evolution (1) xxi + 429 p; ill.; no index. (2) xxi + pp 431-817.
Czeczuga, B. 1977. Carotenoids in leaves and their galls. Marcellia, 40: 177–80.
Dalla Torre, K.W.von & Kieffer, J. J. 1910. Cynipidae. Das Tierreich Friedlander & Sohn, Berlin, 891 p. https://doi.org/10.5962/bhl. title.1077
Fang, Z., Tang, C.-T., Nicholls, J. A., Zhu, Y., Xiong, T., Hearn, J., Sinclair, F., Melika, G., Nieves-Aldrey, J. L., Csóka, G., Mikolajczak, K. M., Stone, G. N. & Fang, S. 2020. A New Genus of Oak Gallwasp, Heocynips Fang, Nieves-Aldrey, and Melika (Hymenoptera: Cynipidae: Cynipini), from China. Proceedings of the Entomological Society of Washington, 122 (4): 787-804. https://doi.org/10.4289/0013-8797.122.4.787
Folmer, O., Black, M., Hoeh, W., Lutz, R. & Vrijenhoek, R. 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology, 3 (5): 294-299. Fontdevila, A. & Moya, A. 2003. Evolución: Origen, adaptación y divergencia de las especies. Editorial Síntesis. España. 591 p. Gómez-Pompa, A., Barrera, A., Gutiérrez-Vázquez, J. & Halffter, G. 1980. Biología: Unidad, Diversidad y Continuidad de los Seres Vivos. Editorial Consejo Nacional para la Enseñanza de la Biología. Ciudad de México. 942 p. Hearn, J., Blaxter, M., Schönrogge, K., Nieves-Aldrey, J.-L., PujadeVillar, J., Huguet, E., Drezen, J.-M., Shorthouse, J. D. & Stone, G. N. 2019. Genomic dissection of an extended phenotype: Oak galling by a cynipid gall wasp. PLoS Genet, 15 (11): e1008398. https://doi.org/10.1371/journal.pgen.1008398
Lobato-Vila, I. & Pujade-Villar, J. 2019. Revision of world Ceroptresini (Hymenoptera: Cynipidae) with the description of a new genus and five new species. Zootaxa, 4685: 1-67. https://doi. org/10.11646/zootaxa.4906.1.1
Lobato-Vila, I., Bae, J., Roca-Cusachs, M., Kang, M., Jung, S., Melika, G., Pénzes, Z. & Pujade-Villar, J. 2022. Global phylogeny of the inquilinous gall wasp tribe Synergini (Hymenoptera: Cynipoidea: Cynipidae): first insights and establishment of a new cynipid tribe. Zoological Journal of the Linnean Society, 195: 1338-1354. https://doi.org/10.1093/zoolinnean/zlab085. Margalef, R. 1977. Ecología. Omega. (2ª edición). Barcelona. 951p.
Markel, K., Novak, V., Bowen, B. P., Tian, Y. & Shih, P. M. 2024. Cynipid wasps systematically reprogram host metabolism and restructure cell walls in developing galls. Plant Physiology, (in press). https://doi.org/10.1093/plphys/kiae001
Martínez-Romero, A., Cuesta-Porta, V., Equihua-Martínez, A., Estrada-Venegas, E. D., Barrera-Ruiz, U. M., Cibrián-Tovar, D. & Pujade-Villar, J. 2022. Aportación al conocimiento de las especies de Cynipini (Hymenoptera: Cynipidae) en los estados mexicanos. Revista Mexicana de Biodiversidad, 93: e933998. https://doi.org/10.22201/ib.20078706e.2022.93.3998
Melika, G. & Abrahamson, W. 2002. Review of the world genera of oak cynipid wasps (Hymenoptera: Cynipidae, Cynipini). In: G. Melika and C. Thuróczy (Eds), Parasitic Wasps: Evolution, Systematics, Biodiversity and Biological Control. Agroinform, Budapest, P. 150-190.
Nastasi, L. F., Smith, C. N., Davis, C. K., Ward, A. K. G., Brown, G., Zhang, Y. M., Rollins, S., Friesen, C., Tribull, C. M., Forbes, A. A. & Deans, A. R. 2024. One must imagine Sisyphus happy: Integrative taxonomic characterization of 22 new Ceroptres species (Hymenoptera: Cynipidae: Ceroptresini). Zootaxa, 5508 (1): 1-63. https://doi.org/10.11646/zootaxa.5508.1.1
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O’Hara, R. B., Simpson, G. L., Solymos, P., Stevens, M. H. H. & Wagner, H. 2015. The vegan package. Community ecology package. R package version 2.3-2. http://CRAN.R-project.org/ package=vegan
Pénzes, Z., Tang, C.-T., Stone, G. N., Nicholls, J .A., Schwéger, S., Bozsó, M. & Melika, G. 2018. Current status of the oak gallwasp (Hymenoptera: Cynipidae: Cynipini) fauna of the Eastern Palaearctic and Oriental Regions. Zootaxa, 4433 (2): 245-289. https://doi.org/10.11646/zootaxa.4433.2.2
Pujade-Villar, J., García-Martiñón, R. D., Equihua-Martínez, A., Estrada-Venegas, E. D. & Ferrer-Suay, M. 2016. Neuroterus fusifex Pujade-Villar and Ferrer-Suay n. sp. (Hymenoptera: Cynipidae): first record of galls on catkins in Mexico. Folia Entomológica Mexicana (nueva serie), 2 (3): 75-83.
Pujade-Villar, J., Bellido, D., López, G. S. & Melika, G. 2001. Current state of knowledge of heterogony in Cynipidae (Hymenoptera, Cynipoidea). Sessió Conjunta d’Entomologia, 87-107.
Pujade-Villar, J., Melika, G. & Cuesta-Porta, V. 2024. Structure and function of oak gall wasps. In: Plant galls: structure and funcions. Oliveira, D.C. & Isaias, R.M.S. (Edts). Springer (in press).
Pujade-Villar, J. & Ros-Farré, P. 1998. Inquilinos y parasitoides de las agallas del género Plagiotrochus Mayr colectadas en el Nordeste de la Península Ibérica. Boletín de la Asociación Española de Entomología, 22 (1-2): 115-143.
Pujade-Villar, J. Clark, R. & Melika, G., 2023. Description of the first species of gall wasp (Hym., Cynipidae: Cynipini) and other unknown galls on Quercus macdougallii (Fagaceae). Butlletí de la Institució Catalana d’Història Natural, 87 (3): 153-160.
R Core Team, 2019. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Vienna, Austria. https://www.R-project.org/ [Fecha de consulta 10 July 2020].
Ronquist, F., Nieves-Aldrey, J.-L., Buffington, M. L., Liu, Z., Liljeblad, J. & Nylander, J. A. A. 2015. Phylogeny, Evolution and Classification of Gall Wasps: The Plot Thickens. PLOS ONE, 10 (5): e0123301. https://doi.org/10.1371/journal.pone.0123301
Stone, G. N., Hernandez-Lopez, A., Nicholls, J. A., Di Pierro, E., Pujade-Villar, J., Melika, G. & Cook, J. M. 2009. Extreme host plant conservatism during at least 20 million years of host plant pursuit by oak gallwasps. Evolution: International Journal of Organic Evolution, 63 (4); 854-869. https://doi.org/10.1111/ j.1558-5646.2008.00604.x
Stone, G. N., Schönrogge, K., Atkinson, R. J., Bellido, D. & Pujade-Villar, J. 2002. The population biology of oak gall wasps (Hymenoptera: Cynipidae). Annual review of entomology, 47 (1): 633-668. https://doi.org/10.1146/annurev. ento.47.091201.145247
Tamura, K., Stecher, G. & Kumar, S. (2021) MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Molecular Biology and Evolution, 38: 3022-3027. https://doi.org/10.1093/molbev/msab120
Tavares, J. da S. 1926. Os Cynipides da Península Ibérica. Broteria, ser. zool., 23: 16-78.
Butlletí de la Institució Catalana d’Història Natural, 88 (4): 193-194. 2024 ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
Primeras citas del género Colon Herbst, 1797 (Coleoptera: Colonidae) para Navarra, Península Ibérica
First records of the genus Colon Herbst, 1797 (Coleoptera: Colonidae) for Navarra, Iberian Peninsula
Amador Viñolas* & José Ignacio Recalde-Ururzum**
* Museu de Ciències Naturals. Laboratori de Natura, Coŀlecció d’artròpodes. Passeig Picasso, s/n. 08003 Barcelona. A/e: av.rodama@gmail.com ** C/ Andreszar, 21. 31610 Villava-Atarrabia, Navarra. A/e: jirecalde93@gmail.com
Autor de correspondencia A. Viñolas. A/e: av.rodama@gmail.com
Rebut: 06.12.2024. Acceptat: 15.12.2024. Publicat: 30.12.2024
Viñolas & Muñoz-Batet (2015) realizaron la revisión de las especies del género Colon Herbst, 1797 presentes en la Península Ibérica e islas Baleares, incluyendo un total de ocho taxones. El material estudiado por dichos autores o citado del ámbito ibérico se limita a las siguientes provincias españolas: Almería, Asturias, Barcelona, Baleares, Cádiz,

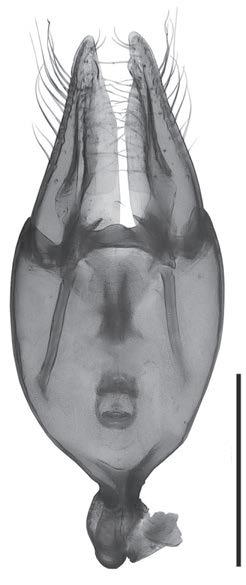
Ciudad Real, Córdoba, Girona, Guadalajara, Huesca, Málaga, Murcia y Teruel, si bien algunas de las antiguas citas son merecedoras de ser comprobadas (Viñolas & Muñoz-Batet, 2015, 2018; Muñoz-Batet, Piera, Calaf. & Viñolas, 2023), pues sólo se basaron en caracteres externos.
Durante recientes campañas de muestreo de coleópteros realizadas en Navarra se han obtenido una serie de ejemplares del género que, tras su estudio, se ha podido comprobar que pertenecen todas ellas a la especie Colon (Myloechus) pubescens Lucas, 1846.
Material estudiado
1 ♂ y 1 ♀, etiquetados: «V.2024, Lantz, Navarra, Beunza & Recalde leg.». Depositados en la colección de A. Viñolas.
4 ♂ y 3 ♀, etiquetados: «23.VI.2023, Caídas de la Negra, Balsa del Viso, P. N. de las Bardenas Reales, Navarra, PérezMoreno, Agoiz, San Martin & Recalde leg.» Depositados en las colecciones de los autores.
Colon (M.) pubescens es una buena especie bien caracterizada por la conformación de los metafémures y metatibias (Fig. 1), por la morfología del edeago (Fig. 2), así como por otros caracteres externos (Viñolas & Muñoz-Batet, 2015). Esta especie está extendida por el área iberobalear habiendo podido estudiar ejemplares de las siguientes provincias: Almería, Barcelona, Baleares (Mallorca), Girona, Guadalajara, Huesca, Navarra y Teruel.
Agradecimientos
A Ignacio Pérez-Moreno (Logroño), Jorge Agoiz (Tudela),
Antonio San Martín (Pamplona) y Rubén Beunza (Lantz), co-recolectores del material estudiado. Al ayuntamiento de Lantz, a Alejandro Urmeneta y la Comunidad de Bardenas Reales por las facilidades ofrecidas para la realización de los trabajos de campo.
Viñolas, A. & Muñoz-Batet, J. 2015 El género Colon Herbst, 1797, en la Península Ibérica (Coleoptera: Leiodidae: Coloninae). Butlletí de la Institució Catalana d’Història Natural, 79: 113-134
Viñolas, A. & Muñoz-Batet, J. 2018. Nuevas aportaciones al conocimiento de la fauna coleopterológica de la Península Ibérica. Nota 4a. (Coleoptera). Revista Gaditana de Entomología, 9 (1): 163-175.
Viñolas, A., Soler, J, & Muñoz-Batet, J. 2024. Los coleópteros del Parque Natural del Montseny. Monografies de la Institució Catalana d’Història Natural, 6. 201 p.
Muñoz-Batet, J., Piera, E.. Calaf, J. & Viñolas, A. 2023. Los coleópteros del Parque Natural de la Muntanya de Montserrat. Monografías de la Institució Catalana d’Història Natural. vol. 5. 108 p.
(4), 2024
Butlletí de la Institució Catalana d’Història Natural, 88 (4): 195-204. 2024
ISSN 2013-3987 (online edition): ISSN: 1133-6889 (print edition)
(Barcelona 6.09.1935- Sant Cugat del Vallès 1.10.2023)
Xavier Ferrer Parareda*, Adolf de Sostoa* & Jordi Serra-Cobo*, **
* Institut de Recerca de la Biodiversitat (IRBIo). Facultat de Biologia, Avda. Diagonal, 643. 08028 Barcelona. ferrer.xavier@gmail.com, asostoa@gmail com ** Universitat de Barcelona. Facultat de Biologia. Departament de Biologia Evolutiva, Ecologia i Ciències Ambientals. Avda. Diagonal, 643. 08028 Barcelona, jordi.serra.cobo@ub.edu
Autor de correspondència: Xavier Ferrer Parareda. A/e: ferrer.xavier@gmail.com
Rebut: 04.10.2024. Acceptat: 04.11.2024. Publicat: 30.12.2024

Introducció
«L’honestedat científica i personal, la fidelitat, la discreció, i la lleialtat formaren part essencial de la seva escala de valors». «Persona sàvia i bona, amable mirada i paraules acollidores». «Entranyable i disposat sempre a ajudar; mai em va fer sentir inferior». «Ha marxat un dels grans». «Ha tingut sempre com a primordials, en totes les activitats de la seva vida, els valors ètics i humans». «Tenia una exquisida educació i una notable capacitat de lideratge, uns trets que li permeteren canviar i potenciar la zoologia acadèmica catalana». Aquests són alguns dels missatges i pensaments dels
DOI: 10.2436/20.1502.01.195
seus familiars, professors, alumnes, amics o coneguts (Anònim, 2023).
Jacint Nadal ens explicava que de molt petit ja criava ocells i altres mascotes a casa seva, una torre amb força jardí del barri de Sant Gervasi de Barcelona. Aquesta infantesa, en contacte amb els animals, l’educació, la seva família (petita i gran), i una joventut dura (orfe de pare als 15 anys i de mare als 21), el forjaren com a home de principis auster, estable i emprenedor. En efecte, el seu pare era doctor en farmàcia, i metge; un patriarca amb esperit de servei que ajudava tothom i que va fer que els avis quan eren grans s’instaŀlessin amb ells a la torre. Posteriorment recollí també la Conxita, una
X. FERRER PARAREDA ET AL.
tieta de Jacint Nadal que havia quedat vídua amb la seva filla. Voltes de la vida, va ser la seva tieta Conxita qui va prendre el rol de fer de mare (i pare) dels sis germans Nadal Puigdefàbregas, amb Jacint, el baró més gran de tots, el tercer a la jerarquia. Va estudiar amb una beca del Coŀlegi de Metges (per ser orfe de metge) als Jesuïtes de Sarrià (Barcelona) on connectà amb bona part de la burgesia catalana. Posteriorment, els contactes personals li facilitaren molt l’establiment de projectes i coŀlaboracions amb empreses de línies de recerca en zoologia aplicada que sempre li van interessar. Els biòlegs, en funció de la seva dedicació professional, podríem dir coŀloquialment que es classifiquen en tres grups; de bata, de bota i gestors. Jacint Nadal va practicar amb èxit les tres disciplines al llarg dels seus 88 anys de vida. La major part de les seves notes expliquen les dificultats de la gestió acadèmica i molt poc sobre les grans realitzacions i del rol fonamental que jugà en el coneixement i la conservació de la biodiversitat animal a Iberoamèrica i a la Mediterrània. Format en un context clàssic i jeràrquic, va saber evolucionar adaptant-se als nous temps democràtics (Fig. 1).

dem que, sense un bagatge humà sòlid, les projeccions fins i tot professionals són curtes o no deixen escola. Aquest no ha estat el cas de J. Nadal, que va tenir com a primordial en la seva vida els valors ètics i humans, i que deixà una petjada de profunda humanitat.
Jacint Nadal ha estat prolífic en publicacions de recerca i divulgació alhora que entrevistat, per exemple (Anònim, 2023), (Vendrell Simón, 2003). La major part d’aquesta informació personal està recollida al CRAI_UB (Centre de Recursos per l’Aprenentatge i la Investigació); (Fons bibliogràfic Jacint Nadal Puigdefàbregas, CRAI, 2008).
Soci numerari de la ICHN des de 1976, en J. Nadal tingué poca activitat institucional. Als inicis dels anys 90 del S. XX la vida de la Institució era magra i ell escriu «Era una època de vaques flaques perquè tots els diners s’abocaven a fer el diccionari de l’IEC». El 4/3/1992 fou elegit president de la ICHN i va succeir Xavier Llimona Pagès en aquest càrrec, el qual el va exercir fins al 16/3/1994 (Camarasa, 2000). Jacint Nadal deixà la seva empremta a la ICHN en dos elements que el caracteritzaren: aconseguir finançament i millorar aspectes editorials. En efecte, les relacions que J. Nadal mantenia amb ICONA (Instituto para la Conservación de la Naturaleza, del Ministerio de Agricultura) eren exceŀlents. Així, ell va aconseguir l’ajut d’un milió de pessetes concedit pel sotsdirector de l’ICONA Anton Novás. Amb aquests diners va impulsar un canvi important al Butlletí de la ICHN en la vessant formal d’estructura i en la maquetació (que va encarregar a professionals). Aquest canvi fou generat i proposat per J.Ma. Camarasa Castillo, el qual, en el número 61 editat el 1993, va escriure un article anònim on s’explica que el “nou Butlletí” té sis seccions i contempla la publicació de, no solament descripcions de fauna, flora i gea, sinó també de materials de gestió naturalista, obituaris, etc. Jacint Nadal feu possible la modernització de les publicacions de la ICHN en uns anys difícils.
En una existència llarga, una vocació naturalista primerenca i una curiositat innata per molts aspectes de la vida el dugueren a conrear diverses àrees de coneixement i restar obert a les sobtades oportunitats que du la vida. Sovint obli-
Butlletí ICHN 88 (4), 2024
En la justificació del Doctorat Honoris Causa que l’any 2009 li va atorgar la Universidad Autónoma Metropolitana de México, es reconeixia que el Dr. Nadal havia primat la trajectòria acadèmica sobre la científica. Un exemple del seu interès acadèmic va ser el seu nomenament l’any 2005 com a membre de la Reial Acadèmia de Ciències i Arts de Barcelona (Anònim, 2024). En l’acte d’ingrés presentà la conferència titulada «Origen i evolució dels cordats». El Dr. Nadal assistia assíduament a les reunions de la Reial Acadèmia. Quan l’any 1975 va esdevenir catedràtic de la UB, era conscient que a Catalunya tradicionalment hi ha hagut una mancança de mentors en el coneixement de la fauna vertebrada, i el seu gran i genèric objectiu fou crear i potenciar l’especialitat dels animals vertebrats, poc desenvolupada a la Universitat de Barcelona (UB). De fet, abans que J. Nadal ocupés la càtedra de zoologia (vertebrats) de la UB, els en-
senyaments i la recerca de la zoologia a la Facultat de Biologia versaven tradicionalment sobre invertebrats (artròpodes i no artròpodes). Aquest objectiu romangué actiu durant tota la vida de J. Nadal, i a més a més, el seu tarannà, l’atzar i el contacte amb altres professionals li aportaren dos objectius més: la projecció a la societat i la gestió acadèmica. Aquests tres objectius sovint s’encavalcaren en el temps i li suposaren un esforç humà considerable, en especial durant el període en què fou degà a la Facultat de Biologia de la UB. El Dr. Nadal tenia un concepte obert de la zoologia, tant en la manera d’abordar la disciplina com en la d’entendre-la. El fet de venir del món de la fisiologia animal (per la seva tesi doctoral), li permeté copsar que estudiar els animals no consisteix solament a capturar-los, mesurar-los i determinar-los. A mitjan anys 1970, al Departament de Zoologia de la UB (dirigit llavors pel catedràtic Enrique Gadea Buisan), els animals vius estaven oficiosament prohibits i els animals es conservaven en alcohol o formol, o dissecats; aquestes eren les úniques vies d’entrada d’animals en l’esmentat departament, on quasi exclusivament es feien tesis doctorals d’anatomia i filogènia. Jacint Nadal, per contra, creia també en la validesa de les tècniques no invasives com les observacions (àmpliament utilitzades en l’ornitologia), i considerava també com a continguts zoològics el comportament, l’autoecologia, les adaptacions dels animals a l’acció de l’home i del canvi climàtic, etc. A (Jordà Sanuy & Ferrer Parareda 2020), s’explica el paper de J. Nadal com a transformador de les disciplines zoològiques a la UB (Fig.2).

i per a formar equips potents i pluridisciplinaris. Va aconseguir una ampliació important en el nombre de places i contractes per a professors de vertebrats a partir del 1977. Ell també va potenciar i facilitar en gran manera la direcció de tesis doctorals. Ell mateix en dirigí 35, alguna de les quals va comportar una ampliació a noves temàtiques, com per exemple la de les ascídies (cordats). El seu gran treball per impulsar la zoologia acadèmica es veu amb les tesis doctorals d’ornitologia defensades a la UB. Des de 1952, data de creació de la carrera de Biologia a la UB, van transcórrer 25 anys fins que fou defensada a Catalunya la primera tesi d’ornitologia, el 1977, dirigida pel Dr. Enrique Balcells Rocamora. Amb l’empenta de J. Nadal s’incrementà el nombre i la freqüència d’aquest tipus de tesis. En alguns anys, com el 1991 i el 1994, a la UB es defensaren fins a quatre tesis anuals d’ornitologia dirigides per ell mateix i/o pels diferents professors o doctors que ell potencià. Entre el 1977 –l’any en què J. Nadal començà a dedicar-se de ple a desenvolupar la unitat de vertebrats– i el 2006 es van llegir a la UB un total de 267 tesis sobre vertebrats. S’entén perfectament que la seva feina acadèmica arrossegués un formidable desenvolupament del coneixement de grups bandera, com els ocells o els cetacis, i que la seva tasca fos l’origen del gran coneixement que es té en la actualitat d’aquests animals (Fig.3).
El Dr. Nadal posà un interès especial a crear un equip de professors i investigadors especialistes en els diferents grups dels vertebrats, com ara els peixos, els ocells, els amfibis, els rèptils, els cetacis, els micromamífers, els ratpenats o quiròpters, etc. Aquesta iniciativa va potenciar la posada en marxa de projectes de recerca específics i pluridisciplinaris en el camp dels vertebrats en els àmbits local, nacional i internacional. Jacint Nadal en l’exercici de la docència, importantíssima per a ell, impulsà les estades de doctorands fora de la UB mitjançant la facilitació econòmica i/o potenciant contactes a l’estranger per a l’adquisició de coneixements als laboratoris

Les disciplines que el Dr. Nadal treballà foren biologia i ecologia de vertebrats, faunística, fisiologia animal, anatomia animal, zoologia sanitària, filogeografia, recursos baleners, etc. Globalment, el tàxon que més li va interessar fou probablement el dels mamífers. Jacint Nadal restà obert a la transversalitat amb d’altres disciplines, com la química, la genètica, etc., creant coneixement i equip de recerca sobre zoologia aplicada. Alhora, ell trobà finançament en moltes empreses (agroalimentàries, editorials, farmacèutiques, energètiques), institucions públiques i privades, nacionals i internacionals. En aquesta faceta de la seva feina, fou un acadèmic atípic durant les dècades dels anys 70, 80 i 90 del segle XX.
Projecció
Polifacètic, Jacint Nadal publicà al voltant de 200 treballs científics entre articles i llibres, entre els quals ell destacava sempre la monografia Vertebrados (Nadal, 2001) que utilitzà en les seves classes de vertebrats i que publicà set anys després que la concebés. Vist en perspectiva històrica, pensem que va tenir una major projecció el llibre Guión para trabajos prácticos. Zoología-Cordados (Nadal et al., 1968), un manual d’identificació de vertebrats ibèrics publicat en una època òrfena de guies d’identificació de vertebrats. El Dr. Nadal publicà treballs sobre molts tàxons, en especial sobre mamífers com els cetacis, els rosegadors, els quiròpters, etc. També va potenciar altres grups d’estudi com els ocells, els peixos continentals, els invertebrats dels arrossars, les sargantanes, les serps, els paràsits, etc. Un exemple destacat d’article científic en el qual va participar fou el relatiu a les zoonosis emergents en quiròpters (Serra-Cobo et al., 2013) (Fig.4).

4.
de la medalla i placa Narcís Monturiol al mèrit científic i tecnològic del 1992, la qual va ser lliurada pel president de la Generalitat Jordi Pujol Soley.
De mentalitat oberta i practicant del «viure i deixar viure», a J. Nadal li agradava la ciència ciutadana i, per primer cop a les universitats catalanes, un acadèmic acceptava i potenciava aquest tipus de recerca, en concret amb ocells vius i silvestres (Ferrer, 2023). L’any 1981 fou el principal signant
Butlletí ICHN 88 (4), 2024
acadèmic del contracte amb la Caixa de Barcelona destinat a elaborar el primer Atlas dels ocells nidificants de Catalunya i Andorra. El Dr. Nadal permeté també que l’equip s’ubiqués a la seva unitat i que les justificacions i factures les gestionés la seva administració, sense acceptar cap recompensa, ni econòmica ni de crèdits d’autoria. En la mateixa línia d’iniciar un contacte estret entre el món acadèmic i els zoòlegs aficionats, J. Nadal va establir al Departament de Zoologia, a principis de la dècada de 1990, la base de dades d’amfibis i rèptils de l’Asociación Herpetológica Española
Una activitat en la qual Jacint Nadal va esmerçar molt de temps va ser l’ajut gratia et amore a moltíssims biòlegs joves que li demanaven consell per crear una empresa, cercar feina, buscar contactes, etc Ell sempre tenia la porta oberta i estava disposat a donar un cop de mà a tothom. La realitat és que molts dels deixebles i altres persones a qui va ajudar actualment gestionen empreses privades (de consultoria ambiental, energètiques) o públiques (museus, zoos, aeroports, ports. ), organismes públics (ajuntaments, diputacions, ens comarcals, autonomies, parcs naturals, parcs nacionals), etc (Fig.5)

o el 1983. (Fot. Àlex Aguilar).
Jacint Nadal sempre va tenir clar que calia fer arribar a la societat els coneixements i les opinions dels naturalistes (professors, aficionats, professionals, etc.) i, ja durant el 1970, va publicar el seu primer article divulgatiu a la revista Muntanya. Per a ell va ser molt important la divulgació de la ciència (principalment dels animals i dels seus hàbitats) i de la conservació,i, a partir dels anys 1960, va coŀlaborar com a publicista amb moltes editorials (principalment Planeta, Reverté, RBA, Océano i Salvat.), així com amb revistes i diaris (Fig.6).
Una de les activitats divulgatives més importants en què va participar es va fer del 1989 al 1993 al Pabellón de la Naturaleza de Expo-92, de Sevilla, que estava situat al Sector del Jardí Americà de l’illa de la Cartuja. Amb aquesta participació va fer realitat un dels seus anhels: l’ensenyament i el bescanvi de coneixements sobre biodiversitat amb Iberoamèrica. Jacint Nadal va promoure el projecte aconseguint diners i patrocini per als continguts, i va engrescar el rector de la UB, que el 30/06/1989 signà un conveni de coŀlaboX. FERRER

ració segons el qual la UB aportava el projecte, la direcció científica i part dels continguts; l’ICONA posava els diners, la realització i part dels continguts; i la Sociedad Estatal para la Exposición Universal de Sevilla aportava els permisos i els 2.053 m2 on es construí el pavelló.
El Pavelló de la Natura estava constituït per quatre sales; la més gran de les quals era la “bombolla de selva amazònica” que representava els vuit primers metres d’altura de la selva amb organismes vius com bromeliàcies, grimpadores, iguanes, etc. La sala de la Naturaleza Latinoamericana fou també espectacular pels materials gràfics i escènics; per exemple, en el terra, hi havia una maqueta de la geografia física d’Amèrica Llatina a escala 1:1.000.000 coberta per una tarima de vidre sobre la qual el visitant podia passejar. Naturaleza por descubrir fou una sala de cinema de gran format Showscan (70 mm a 60 imatges per segon), on es projectaven filmacions originals de parcs nacionals espanyols i americans. La gran sala Conservemos la Naturaleza presentà continguts originals gràfics i de guió, creats expressament per l’equip de J. Nadal amb exemples iberoamericans, principalment. Per últim, a l’entrada del Pavelló de la Natura s’instaŀlà un petit àmbit expositiu intitulat Los Naturalistas Españoles en América. El Pavelló de la Natura fou un dels quatre pavellons temàtics de l’EXPO 92. Va requerir més de quatre anys de
feina original i va rebre prop d’un milió de visitants en total (unes 6000 persones/dia, de mitjana) (Fig.7).

Sobre la temàtica conservacionista, J. Nadal publicà el 1969 el seu primer article, i fou un dels fundadors de pes de DEPANA l’any 1976 (Lliga per a la Defensa del Patrimoni Natural). Sobretot va participar en l’entitat en els primers anys de la seva existència, quan no dubtà a signar alguns manifestos compromesos.
Gestió acadèmica
El Dr. Nadal va treballar principalment com a gestor acadèmic des del 1970 (quan fou nomenat catedràtic de Zoología Vertebrados de la Facultad de Ciencias de León) fins a principis del 1983 (final del seu Deganat a la Facultat de Biologia de la UB). Jacint Nadal, l’any1975, oficialment per trasllat, esdevé el nou catedràtic de Zoologia (Vertebrats) de la Facultat de Biologia de la UB, amb dedicació parcial fins a l’octubre de 1977 per formar part d’una comissió de servei a Madrid com a cap de perfeccionament del professorat al Instituto Nacional de Ciencias de la Educación (INCIE). A Madrid, J. Nadal farà amistat amb dos alts funcionaris del Ministeri d’Educació i Ciència (MEC), José Fernández Vega i Paco Arances, que seran clau en el procés de la seva gestió acadèmica a la Facultat de Biologia de la UB. També jugà un paper important en aquesta gestió el catedràtic Ramon Parés Farràs, que per a J. Nadal fou mentor i conseller important en tot moment. Ell qualificava aquella etapa de la seva vida com a «activa, moguda i fecunda», alhora que agitada políticament i socialment, ja que la major part va coincidir amb la transició social i política. En aquella etapa, la persona estable i serena del Dr. Nadal fou fonamental i molt apreciada; com un far en aigües tempestuoses. Per exemple, en el cop d’estat del 23/02/1981, escriu J. Nadal circa 2013: ”En sortir de la
Universitat veig la Gran Via deserta, el mateix que passava a la pròpia Universitat; vaig anar al Deganat on estava l’Enrique, el bidell, i aquest em pregunta: què faig?, tot informantme que havia desaparegut tothom, que estava tot desert. Donada la situació li dic que tanqui. «Em va donar la impressió que érem els dos últims en deixar la Universitat». També pel caràcter i els principis ètics del Dr. Nadal, en José Fernández Vega li demanà d’entrar en política com a sotsdirector de «Relaciones con las Cortes» en el partit Unión del Centro Democrático del llavors president Adolfo Suárez González. Coneguda l’estimació de J. Nadal per la docència, inclús crearen una càtedra de zoologia a la Universitat d’Alcalà de Henares per tal que pogués compatibilitzar la seva feina a l’INCIE amb la docència i restés al MEC. Jacint Nadal, als voltants del 2013, sintetitzava aquest període com: «[Vaig tenir una] activitat important durant l’època de la transició com a gestor acadèmic; una etapa que per a mi va ésser decisiva pel reconeixement científic i social de la Facultat, i en la que vaig esmerçar absoluta dedicació i les bones relacions que tenia amb personalitats del ministeri [MEC]» (Fig.8).

Figura 8. Jacint Nadal a la darrera classe de cordats de la facultat de biologia de la UB del curs 1977-78; Juny 1978 (Fot. Lluis Soria, Arxiu G. A. Llorente).
Apleguem aquí algunes informacions referent al paper fonamental que va jugar J. Nadal en la formació i configuració de la Facultat de Biologia de la UB. Quan l’any 1978 el Dr. Nadal va assumir el càrrec de degà, la Facultat de Biologia s’ubicava encara a l’edifici històric de plaça Universitat. Aquest càrrec finalitzà el 1983. Ell va ser elegit degà de la Facultat sense haver-se presentat per a ocupar el càrrec, tot i que ja era vicedegà. Les obres del nou edifici, l’actual Ramon Margalef, estaven aturades des de feia set o vuit anys a causa de l’elevat pressupost que comportava la construcció d’un edifici tan singular i de qualitat, i per no haver tingut en compte que Barcelona és zona semisísmica, per la qual cosa, en ser un edifici oficial i públic, havia de tenir una fonamentació especialment adequada. Jacint Nadal va desencallar la situació ajudat per Paco Arances, llavors director general de Programació i Inversions del MEC, de qui depenien els fons econòmics per acabar l’edifici. Jacint Nadal va aconseguir que el MEC en pagués una part important, fent així possible
Butlletí ICHN 88 (4), 2024
la seva finalització. També ell va aconseguir que el mateix MEC pagués la construcció d’un nou i modèlic animalari (l’anomenat Rata Hilton pel professorat), inicialment destinat a animals de laboratori i salvatges, encara que després es destinà quasi exclusivament a animals de laboratori. També el Dr. Nadal va iniciar la construcció de l’aulari (actual edifici Durfort), ja que l’edifici de la Facultat programat inicialment s’havia fet petit a causa de l’increment del nombre d’alumnes (els baby boomers). La inauguració de l’aulari es va fer quan ja no era degà. També en aquella època, ell va obtenir finançament i la cessió a la UB, per part de l’ICONA, del CRAM (Centre de Recerca d’Alta Muntanya), actual ECAM (Estació Científica d’Alta Muntanya), que està situat en un edifici del municipi de Vielha-Mig Aran, a uns 100 m de l’Hospital de Vielha, que es troba a l’entrada de la boca sud del túnel de Vielha. Aquesta estació de recerca de camp, que llavors es va estrenar, fou la primera de la UB, una idea inicial del catedràtic Ramon Margalef López que el Dr. Nadal va fer realitat (Fig.9).

Figura 9. Nomenament l’abril de 2005, de l’edifici principal de la Facultat de Biologia amb el nom Ramon Margalef. D’esquerra a dreta, Ramon Parés, Antoni Prevosti, Maria Monclús, Josep Pons i Jacint Nadal
El 31 d’octubre de 2023, l’actual Departament de Biologia Evolutiva, Ecologia i Ciències ambientals (BEECA) va proposar que la Facultat reconegués l’important paper de J. Nadal donant el seu nom a algun espai de la Facultat. En la Junta de Facultat del 19/12/2023 es va aprovar formalment batejar l’aula de graus amb el seu nom i el 02/05/2024 la proposta es va fer realitat amb la designació Aula de Graus Jacint Nadal.
El 1977, arrelat definitivament a Barcelona, Jacint Nadal abandonà la gestió acadèmica de la Facultat per dedicar-se plenament a la càtedra, estudiant, investigant i potenciantne el professorat. La gran energia, contactes i educació de J. Nadal van tenir un efecte activador i revulsiu en l’estructura de la llavors Càtedra de Zoologia. Conscient dels migrats recursos per a la docència, ell va implantar a la Unitat de Vertebrats una norma no escrita: la no utilització per a la recer-
ca dels diners del capítol II (el pressupost que la Universitat destinava a la Unitat de Vertebrats). Així, materials de docència com l’alcohol o el formol mai es desviaren a la recerca, perquè J. Nadal sempre va considerar essencials la docència i la millora de les pràctiques docents. La seva primera publicació en matèria docent és del 1967, i la primera que va fer de temàtica científica és del 1964. També, com a norma no escrita, Jacint Nadal impulsà i empenyé els seus professors a espavilar-se per trobar finançament per a les seves recerques fora de la Universitat, i ell fou el primer a facilitar-los els contactes necessaris (Fig.10).

siques sobre grups llavors poc coneguts en el món científic, com les espècies d’ocells marines i les cinegètiques (perdius, guatlles, ànecs i tinamús), els quiròpters i els cetacis. També impulsà la recerca de la biologia i ecologia dels peixos continentals en conques fluvials, com per exemple en el projecte de les comunitats de peixos del riu Matarranya, que es va fer en coŀlaboració amb la Universitat de Georgia (EE.UU) i va ser subvencionat pel Comité Conjunto Hispano-Norteamericano. Va participar en diversos projectes d’estudis dels virus ràbics en poblacions de quiròpters espanyols, els quals van ser finançats pel Ministeri de Sanitat i Consum o per la Conselleria de Salut i Consum del Govern Balear (Fig.11).
El Dr. Nadal fou un formidable generador de projectes per als diferents equips de zoologia de vertebrats i sobretot un generador de finançament per a la recerca, la gestió i la conservació de la biodiversitat tant a la Mediterrània com a Iberoamèrica. Ell tenia importants contactes en el context ibèric i els va utilitzar hàbilment per treure’n diners públics i privats. Jacint Nadal actuà com a investigador principal o com a coordinador en més de 40 projectes de gran abast. També, com a profund convençut de la importància dels equips, J. Nadal va delegar i potenciar en gran mesura les tasques i objectius dels seus professors, per exemple en recerques bà-
Dins del món de la recerca, l’objectiu més important de J. Nadal va ser la zoologia ambiental, i la va iniciar per influència del Dr. Enrique Balcells Rocamora, un dels seus mentors, entre el 1965 i el 1970, de manera que l’any 1972 va fer la seva primera publicació en el tema ambiental. El seu objectiu fou la creació i l’impuls de la seva principal línia de recerca: l’estudi dels animals vertebrats com a prospectors i integradors d’informació de la qualitat ambiental. El Dr. Nadal, que va fer la tesi doctoral a Barcelona sobre fisiologia animal, va construir un potent equip de bata i bota que va treballar sobre els efectes d’importants temes ambientals d’interès tan científic com social i econòmic referents als animals vertebrats. Ell sempre va estar convençut que la manera com determinats compostos químics (com els contaminants) afectaven els animals havia de ser analitzada per part dels biòlegs o per equips mixtos de químics i biòlegs, a fi de poder interpretar els efectes biològics que se’n derivaven. Aquesta visió pluridisciplinària era molt interessant, ja que generava resultats de ciència bàsica i aplicada. La recerca pluridisciplinària obrí la porta a nous àmbits de coneixement: l’ecotoxicologia, l’autoecologia, la química ambiental, el comportament animal, la biologia alimentària, els efectes dels impactes ambientals, l’avaluació del recursos pesquers, la biologia de poblacions i la genotoxicitat, entre d’altres. Algunes de les temàtiques treballades per J. Nadal foren el botulisme, els plaguicides organoclorats (per exemple, sobre els fumarells [Chlidonias sp] durant el 1972, al delta de l’Ebre), els metalls pesants (a les sargantanes de les Balears l’any 1979, i en musaranyes en la dècada de 1990, a Cercs), i els efectes de la contaminació atmosfèrica (per exemple, els de la pluja àcida de la central de Cercs sobre ratolins i ocells passeriformes). També han estat importants l’estudi de biomarcadors intrínsecs com els isòtops estables que serveixen per conèixer la dieta i les migracions de cetacis i tortugues marines, per esbrinar si els ocells marins s’alimenten de rebuigs pesquers, i per saber si mengen en superfície o en fondària. També es va endinsar en la problemàtica dels impactes ambientals generats per les explotacions hidroelèctriques en cursos fluvials d’alta muntanya, amb subvencions de FECSAOCIDE (Oficina de Control i Direcció d’Empresa), projecte d’investigació electrotècnic PIE 121.43. Des del 1991 fins als darrers anys de la seva activitat professional, s’interessà per l’ecoepidemiologia, especialment per les zoonosis emergents en quiròpters, participant en diferents projectes amb el Ministeri de Sanitat i Consum i amb la Conselleria de Salut i Consum del Govern Balear. Les converses sobre el tema sempre
X.
eren interessants, especialment per la contribució que feia el Dr. Nadal en els estudis ecoepidemiològics aportant els seus coneixements fisiològics.

Escriu Jacint Nadal circa 2013: «En l’etapa inicial d’estar a Barcelona com a catedràtic tinguérem dos projectes extraordinaris, un de cetacis i altres com ara els projectes Maire i Pelagus, sobre l’impacte de les arts de pesca d’arrossegament i semi-pelàgiques en les poblacions de peixos a la costa mediterrània, subvencionats pel Ministerio de Agricultura, Ganadería y Pesca» Allí constava la Càtedra de Zoologia (Vertebrats) de la UB com a primer coŀlaborador de la Subsecretaría de Pesca (...) Desprès varen venir un munt d’ajuts i de projectes: subvencions de Granges Castelló, [FECSA, Grups de Recerca Consolidats, de la GENCAT] ICONA, Ministeri de Educació i Ciència, tractat de bases amb EUA, projectes de la U.E., etc,. En conjunt, més de 4 milions d’euros, dels quals la UB rebia els corresponents overheads [despeses administratives generals de l’organització, llavors el 10%]”. La Unitat de Zoologia Vertebrats era una de les més productives de la UB com a generadora d’overheads, dels quals solament una petita part es retornava a la unitat.
Jacint Nadal escriu al 2013 resumint la seva història: «[Els projectes] no és el més important, per a mi [ell sempre fou
Butlletí ICHN 88 (4), 2024
discret i humil amb els seus èxits], són les 35 tesis doctorals dirigides, les publicacions fetes, entre elles el llibre de Vertebrados, l’haver organitzat per part de la UB el «Pabellón de la Naturaleza» d’Expo 92 (...). També la projecció americana del Departament de Biologia Animal (Vertebrats) i l’equip de vertebratòlegs format (4 catedràtics, 9 professors titulars), i d’altres nivells, a més dels que estan en altres universitats, CSIC, diputacions, centres de recerca, etc, i la quantitat d’amics assolits.
El Dr. Nadal sempre va destacar per la seva inteŀligència emocional, un atribut bastant desconegut, força escàs i poc valorat entre el professorat, així com per la curiositat per quasi tot. Jacint Nadal tenia altres qualitats escasses avui dia (la de saber escoltar i ser sensible a les problemàtiques dels interlocutors, per exemple), que amb la seva exquisida educació li permeteren ser un formidable diplomàtic, requerit en moltes meses, comissions, comitès avaluadors i tribunals.
La vinculació, amor i dedicació a la família van ser primordials per a ell. En la seva vida va tenir un paper clau la Isabel Lorenzo Busquets, quasi dos anys menor que Jacint, companya de promoció primer i esposa després. Ell diu “m’ha donat pau, tranquiŀlitat i set fills, l’obra de la que n’estic més joiós. I onze nets, i tant fills com nets inteŀligents i ben plantats. Tenir la vida familiar sense trasbalsos m’ha permès dedicar-me amb intensitat a la vida universitària”. Aquesta dona forta, inteŀligent, maca, d’ulls vius, amb molt de seny i molta personalitat, l’ajudà sempre en les petites i grans decisions de la vida (Fig.12)

El Dr. Nadal tenia una formació global, sobretot fisiològica, mèdica, veterinària i zoològica. Sempre estava llegint, formant-se, i posant-se al dia de l’activitat científica. L’impuls al jovent era una altra de les seves característiques. Cap al 2013 escrigué, en relació al jovent durant la seva vida a la Facultat: «(...) no em va mancar el recolzament dels estudiants, la joventut és noble, congruent i valenta. Gràcies».
IN MEMORIAM, JACINT NADAL PUIGDEFÀBREGAS
Indica aquesta actitud la darrera entrevista que li van fer, l’abril del 2023, uns cinc mesos abans del seu traspàs, quan el nomenaren Coŀlegiat d’Honor 2023 (CBC, 2023):
Quin creu és el paper del biòleg a la societat? Com es pot incidir?
«Posant de manifest les incògnites que ens queden per aclarir, que són moltes i enormes, i que al mateix temps són un estímul per aventurar-nos en un millor coneixement del món, la nostra casa. I incidirem estimulant la gent a fer-se constantment preguntes, que no ens quedem amb unes simples respostes, ja que aquestes mateixes engendren altres preguntes. Sempre hi ha un més enllà. I estimulant-nos a respectar l’entorn en què vivim».
Quina part de la seva activitat actual està lligada a la biologia?
«Vora els 88 anys, m’agradaria que fos estimular la societat a que se sentís responsable de la bona formació dels seus integrants. Amb una bona formació totes les altres coses sortirien rodades».
Què li aconsellaria a un futur biòleg?
«Que busqui assolir una bona consteŀlació de coneixements que li permetran la interpretació correcta dels fenòmens naturals».
Jacint Nadal sempre fou un convençut que un ambient humà de qualitat és un substrat base per propiciar nivells professionals d’exceŀlència. Per tant, per a ell, crear una atmosfera de treball còmoda i humana era clau alhora que consubstancial amb el seu caràcter. L’honestedat científica i personal, l’austeritat en tot, la fidelitat, la discreció i la lleialtat formaren part essencial de la seva escala de valors. Molt esportista, en tennis i futbol (aferrissat culer), i caminador. J. Nadal sempre va ser respectuós amb les idees i la feina d’altri (professionals, coŀlegues, etc.), amb independència del seus pensaments, creences i afinitats polítiques. Parafrasejant al periodista Jacinto Antón, podríem dir que Jacint Nadal fou un home d’ordre que disposava del sentit natural de l’autoritat dels nascuts en la púrpura social i econòmica. Alguns dels pòsits que va deixar en la gent que el van conèixer retraten aspectes de la seva personalitat: «El més palès era el seu exquisit tracte personal, sempre atent amb tothom, sabent ser i sabent estar a cada lloc i en cada moment, amb veneració i respecte envers els seus mestres i consideració i protecció amb els seus companys i deixebles. I tanmateix, també pragmàtic, tenint un instintiu olfacte per a les ocasions i situacions aprofitables». «Recordo el seu gran respecte escoltant l’altre abans d’exposar la seva opinió». «Sempre tenia les portes obertes del seu despatx a tothom fent seus els nostres problemes». «Mestre de la vida ens va orientar en el camí a seguir». «Dotat d’una bonhomia, generositat, honradesa, esperit de servei, serenor, sense crits ni murmuracions ni crítiques personals, i discreció» (Fig.13).
El setembre de 2003 un catedràtic de zoologia de la Universitat de Lleó en la seva dedicatòria d’un llibre sintetitzava el personatge:“Profesor Nadal, aprecio siempre tu sabiduría zoológica y un saber estar social magnífico. Ojalá, en la universidad y en la calle, cunda el ejemplo, ese seny de catalán honesto”

13. Málaga, setembre 1990, tesi doctoral d’Ana Carmen Duran, la primera per l’esquerra; desprès Mercè Durfort, Valentí Sans, Jacint Nadal, vicerectora de recerca, Joaquim Gosàlbez i Ramon Muñoz-Chapuli. (Foto M. Durfort).
Descansi en pau Jacint Nadal Puigdefàbregas que prengué la drecera cap el més enllà el matí de l’1 d’octubre de 2023, un dia noble i assenyalat. Ha estat un privilegi gaudir del seu mestratge.
El present article intenta recollir les opinions i sentiments orals o escrits de moltes persones que han posat el seu gra de sorra anònim per bastir-lo. A totes elles, el nostre més profund agraïment. Per la informació donada que ha completat algunes dades del manuscrit, per l’aportació de materials, i per correccions, esmenes i suggeriments sobre l’article, donem el nostre agraïment a: Àlex Aguilar Vila, Josep Ma Camarasa Castillo, família Nadal Puigdefàbregas, Gustavo A. Llorente Cabrera, Joan Jofre Torroella, Joandomènec Ros Aragonès i Dolors Vinyoles Cartanyà. També estem agraïts a altres persones que no citem. Finalment, a l'Albert Masó Planes per haver-nos cedit, entre d'altres, la imatge que encapçala aquest In Memoriam
Anònim, 2024. Jacint Nadal i Puigdefàbregas. Viquipèdia, l’Enciclopèdia Lliure. Accessible a: https://ca.wikipedia.org/ wiki/Jacint_Nadal_i_Puigdef%C3%A0bregas. [Data de consulta: 16 d’agost 2024].
Serra-Cobo, J., López-Roig, M., Seguí, M., Sánchez, L.P., Nadal, J., Borrás, M., Lavenir, R. & Bourhy, H. 2013. Ecological Factors Associated with European Bat Lyssavirus Seroprevalence in Spanish Bats. PLOS ONE , 8(5): e64467.
Camarasa, J. Mª. 2000. Cent anys de passió per la natura : una història de la Institució Catalana d’Història Natural, 1899-1999. Institució Catalana d’Història Natural. Col·lecció Memòria, 14: 183 pp. Barcelona
CBC, Coŀlegi de Biòlegs de Catalunya. 2023. Entrevista al Dr Jacint Nadal i Puigdefàbregas. Accessible a: https://cbiolegs. cat/2023/10/03/dr-jacint-nadal-bioentrevista [Data de consulta: 24 d’abril 2023].
CRAI, 2008. Fons bibliogràfic Jacint Nadal Puigdefàbregas. Accessible a: https://crai.ub.edu/recursos-d-informacio/patrimoni- bibliografic/ colleccions-especials/j-nadal [data de consulta: 24 d’abril 2023].
Ferrer, X. 2023. Obituaris: Jacint Nadal Puigdefàbregas Impulsor de l’ornitologia acadèmica catalana. L’abellerol (Butlletí de Contacte d l’Institut Català d’Ornitologia), 60: 16.
Jordà Sanuy, C. & Ferrer Parareda, X. 2020. Roser Nos. La zoologia al servei de la ciutadania. Butlletí de la Institució Catalana d’Història Natural, 84: 235-242.
Nadal, J., Vericat, J. R., Vidal, A. & Martínez-Rica, J. P. & Balcells, E. 1968. Guión para trabajos prácticos. ZoologíaCordados. Publicación Especial del Centro Pirenaico de Biología Experimental, 350 p. Jaca.
Nadal, J. 2001. Vertebrados, origen, organización, diversidad y biologia. Edicions de la Universitat de Barcelona: Omega cop. 858 pp. Barcelona.
Vendrell Simón, B. 2003. Entrevista a Jacint Nadal Puigdefàbregas. Omnis Cellula, 2003 (oct.-nov.): 34- 41.
Butlletí ICHN
(4), 2024
GEA, FLORA ET FAUNA
NEUS OROMÍ, SEBASTIÀ CAMARASA, DELFÍ SANUY, MARC FIBLA, EDUARD REIG, ALBERT MONTORI
Factors ambientals clau per a la reproducció del gripau corredor als secans de Mas de Melons
Environmental key factors for the reproduction of the natterjack toad in the drylands of Mas de Melons 3
MARGARIDA GENERA I MONELLS
Jordi Sabater Pi (1922-2009). Recordant la figura d’un exceŀlent naturalista del nostre país
Jordi Sabater Pi (1922-2009). Remembering the figure of an excellent naturalist of our country 13
CRISTIAN R. ALTABA
Un nou gènere fòssil de la família Strombidae (Gastropoda: Caenogastropoda: Stromboidea) del Miocè mitjà de Java
A new fossil genus of the family Strombidae (Gastropoda: Caenogastropoda: Stromboidea) from the Middle Miocene of Java 21
JULI MAURI, ANDREU UBACH, SERGI HERRANDO, MARC ANTON, MARC FUSELLAS I CONSTANTÍ STEFANESCU
20 anys de canvis en la biodiversitat d’ocells i papallones a la Regió Metropolitana de Barcelona 20 years of changes in the biodiversity of birds and butterflies in the Metropolitan Region of Barcelona 33
LLUÍS FILBÀ ESQUERRA
Catàleg dels tricòpters (Insecta: Trichoptera) hipogeus iberobalears de la coŀlecció del Museu de Ciències Naturals de Barcelona
Catalog of the Ibero-Balearic hypogean caddisflies (Insecta: Trichoptera) from the collection of the Natural Sciences Museum of Barcelona 53
CLÀUDIA PLA-NARBONA, ANDREU UBACH, CONSTANTÍ STEFANESCU
Beure del fang: un estudi dels abeuradors de papallones a Catalunya
Mud-puddling: A study of butterfly puddling sites in Catalonia 79
JULI PUJADE-VILLAR, CSABA THURÓCZY & ELISENDA OLIVELLA
Calcidoideus obtinguts de dues espècies de lepidòpters minadors: Leucoptera malifoliella (Lyonetiidae) i Phyllonorycter corylifoliella (Gracillariidae) en cultius de pomeres de la província de Lleida
Chalcidoides obtained from two species of miner lepidopteran: Leucoptera malifoliella (Lyonetiidae) and Phyllonorycter corylifoliella (Gracillariidae) in apple crops in the province of Lleida 91
KIKO GÓMEZ, XAVIER ESPADALER, MARIA TERESA MARTÍNEZ-FERRER & JOSEP MIQUEL CAMPOS-RIVELA
Monomorium monomorium Bolton, 1987 in Spain (Hymenoptera: Formicidae)
Monomorium monomorium Bolton, 1987 a Espanya (Hymenoptera: Formicidae)
AMADOR VIÑOLAS & JOSEP MUÑOZ-BATET
Los Ernobiinae de la Península Ibérica e Islas Baleares. 2a nota. El género Ernobius C. G. Thomson, 1859 (Coleoptera: Ptinidae)
The Ernobiinae of the Iberian Peninsula and Balearic Islands. 2nd note. The genus Ernobius C. G. Thomson, 1859 (Coleoptera: Ptinidae)
PERE AYMERICH
A new Noccaea (Brassicaceae) species endemic from alpine habitats of the Pyrenees
Una nova espècie de Noccaea (Brassicaceae) endèmica d’hàbitats alpins dels Pirineus
JOSEP GESTI PERICH
Noves dades sobre la flora vascular del massís de les Guilleries (nord-est de Catalunya)
New data on the vascular flora of the Guilleries massif (northeastern Catalonia)
ANDREU CERA, MARIA JOSÉ CHESA, ÀLEX CORTADA, JOANA MENDES, ANTONIO GÓMEZ-BOLEA
Dades per a la corologia i l’autoecologia de Ramalina lusitanica i R. pusilla a Catalunya
Data on chorology and autoecology of Ramalina lusitanica and Ramalina pusilla in Catalonia
CARLOS PRADERA & AGUSTÍN ESTRADA-PEÑA
Nuevos datos sobre la distribución de la garrapata Hyalomma lusitanicum (Acari, Ixodidae) en Cataluña
101
105
125
133
139
New data on the distribution of the tick Hyalomma lusitanicum (Acari, Ixodidae) in Catalonia 157
VICENÇ BROS, JORDI CADEVALL, GLÒRIA GUILLÉN, ALBERT OROZCO, MIQUEL CAPDEVILA & JORDI CORBELLA
Moitessieria canyellesensis sp. nov. (Gastropoda: Moitessieriidae), una nova espècie estigòbia de Castellar del Vallès (Barcelona, Catalunya)
Moitessieria canyellesensis sp. nov. (Gastropoda: Moitessieriidae), a new stygobie species from Castellar del Vallès (Barcelona, Catalonia) 163
AMADOR VIÑOLAS, JORGE MEDEROS & ORESTES CARLOS BELLO GONZÁLEZ
Una nueva especie del género Europs Wollaston, 1854 de Cuba (Coleoptera: Monotomidae: Monotominae: Europini)
A new species of the genus Europs Wollaston, 1854 from Cuba (Coleoptera: Monotomidae: Monotominae: Europini) 179
SAMAY BRAVO-CUAUTLE, ROSA D. GARCÍA-MARTIÑÓN, URIEL M. BARRERA-RUIZ, BETZABETH C. PÉREZ-TORRES, AGUSTÍN ARAGÓN-GARCÍA, ARMANDO EQUIHUA-MARTÍNEZ, EDITH G. ESTRADA-VENEGAS, MÓNICA RANGEL-VILLAFRANCO, SILVIA ROMERO-RANGEL, VÍCTOR CUESTA-PORTA & JULI PUJADE-VILLAR
Un caso de convergencia evolutiva en agallas de cinípidos del viejo y nuevo mundo (Hymenoptera Cynipidae)
A case of evolutionary convergence in galls of Old and New World cynipids (Hymenoptera Cynipidae) 185
NOTES BREUS
JORDI BASSOLS GARCIA & JOSEP GESTI PERICH
Potentilla supina (Rosaceae) a Catalunya (NE de la península Ibèrica)
Potentilla supina (Rosaceae) in Catalonia (NE Iberian Peninsula)
JOAN PEDROL SOLANES& ANTONI MAYORAL ARQUÉ
Tres plantes rares de Ponent, Catalunya (nord-est ibèric)
Three rare plants from W Catalonia (northeastern Iberian Peninsula)
JÚLIA CAZORLA-VILA & JULI PUJADE-VILLAR
Primeres gales de cinípids mexicans parasitades per Eupelmus pulchriceps (Cameron, 1904) (Hym., Eupelmidae)
First Mexican cynipid oak galls parasitized by Eupelmus pulchriceps (Cameron, 1904) (Hym., Eupelmidae)
CARLOS PRADERA & ALBERTO BERNUÉS-BAÑERES
Primer registro de Aedes vittatus (Bigot, 1861) y Aedes vexans (Meigen, 1830) (Diptera: Culicidae) en la provincia de Tarragona
First record of Aedes vittatus Bigot, 1861 and Aedes vexans (Meigen, 1830) (Diptera: Culicidae) in the province of Tarragona
CARLOS PRADERA, ANTONIO JESÚS REINA-MUÑOZ, JUAN QUETGLAS & AMADOR BARAMBIO-ZARCO
Stricticimex Ferris & Usinger, 1957 (Heteroptera: Cimicidae): primer registro de este género en Europa
Stricticimex Ferris & Usinger, 1957 (Heteroptera: Cimicidae): first record of this genus in Europe
AMADOR VIÑOLAS & JOSEP MUÑOZ-BATET
Euophryum rufum (Broun, 1880) nueva cita para Cataluña (Curculionidae: Cossoninae; Pentarthrini)
Euophryum rufum (Broun, 1880) new record for Catalonia (Curculionidae: Cossoninae; Pentarthrini)
AMADOR VIÑOLAS & JOSÉ IGNACIO RECALDE-URURZUM
Primeras citas del género Colon Herbst, 1797 (Coleoptera: Colonidae) para Navarra, Península Ibérica
11
49
145
149
153
177
First records of the genus Colon Herbst, 1797 (Coleoptera: Colonidae) for Navarra, Iberian Peninsula 193
IN MEMORIAM
XAVIER FERRER PARAREDA, ADOLF DE SOSTOA & JORDI SERRA-COBO
In Memoriam, Jacint Nadal Puigdefàbregas (Barcelona 6.09.1935- Sant Cugat del Vallès 1.10.2023)
195
