ListofContributors
SelmaAybek LaboratoryforBehavioralNeurologyand ImagingofCognition,DepartmentofNeurosciencesand ClinicofNeurology,UniversityMedicalCenter,Geneva, Switzerland
ClaudioL.Bassetti DepartmentofNeurology,University Hospital,Bern,Switzerland
OlafBlanke LaboratoryofCognitiveNeuroscience,Brain MindInstitute,SchoolofLifeSciences,E ´ cole PolytechniqueFe ´ de ´ raledeLausanne,Lausanne, Switzerland;CenterforNeuroprosthetics,E ´ cole PolytechniqueFe ´ de ´ raledeLausanne,Lausanne, Switzerland;DepartmentofNeurology,University Hospital,Geneva,Switzerland
HalBlumenfeld DepartmentsofNeurology,Neurobiology, andNeurosurgery,YaleUniversitySchoolofMedicine, NewHaven,CT,USA
MelanieBoly DepartmentofPsychiatry,Universityof Wisconsin-Madison,Madison,WI,USA;Departmentof Neurology,UniversityofWisconsin-Madison,Madison, WI,USA
Marie-Aure ´ lieBruno ComaScienceGroup,Neurology DepartmentandGIGA,UniversityofLie ` ge,Lie ` ge, Belgium
ChrisButler NuffieldDepartmentofClinical Neurosciences,UniversityofOxford,JohnRadcliffe Hospital,Oxford,UK
CamilleChatelle DepartmentofNeurology,Massachusetts GeneralHospital,Boston,MA,USA;Departmentof PhysicalMedicineandRehabilitation,Spaulding RehabilitationHospitalandHarvardMedicalSchool, Boston,MA,USA
AthenaDemertzi ComaScienceGroup,Neurology DepartmentandGIGA,UniversityofLie ` ge,Lie ` ge,Belgium
SebastianDieguez LaboratoryforCognitiveand NeurologicalSciences,DepartmentofMedicine,Ho ˆ pital deFribourg,Universite ´ deFribourg,Fribourg, Switzerland
BrianL.Edlow DepartmentofNeurology,Massachusetts GeneralHospital,Boston,MA,USA;AthinoulaA. MartinosCenterforBiomedicalImaging,Massachusetts GeneralHospital,Charlestown,MA,USA
AndreasK.Engel DepartmentofNeurophysiologyand Pathophysiology,UniversityMedicalCenterHamburgEppendorf,Hamburg,Germany
NathanFaivre LaboratoryofCognitiveNeuroscience,Brain MindInstitute,SchoolofLifeSciences,E ´ colePolytechnique Fe ´ de ´ raledeLausanne,Lausanne,Switzerland;Center forNeuroprosthetics,E ´ colePolytechniqueFe ´ de ´ ralede Lausanne,Lausanne,Switzerland
JosephJ.Fins WeillCornellMedicalCollegeandthe RockefellerUniversity,NewYork,NY,andYaleLaw School,NewHaven,CT,USA
PascalFries ErnstStru ¨ ngmannInstituteforNeuroscience inCooperationwithMax-Planck-Society,Frankfurt, Germany;DondersInstituteforBrain,Cognitionand Behaviour,RadboudUniversityNijmegen,Nijmegen,The Netherlands
MichaelS.Gazzaniga DynamicalNeuroscience,University ofCalifornia,SantaBarbara,CA,USA;Psychologicaland BrainSciences,UniversityofCalifornia,SantaBarbara, CA,USA
JosephT.Giacino DepartmentofPhysicalMedicineand Rehabilitation,SpauldingRehabilitationHospitaland HarvardMedicalSchool,Boston,MA,USA
OliviaGosseries DepartmentofPsychiatry,Universityof Wisconsin-Madison,Madison,WI,USA;ComaScience Group,NeurologyDepartmentandGIGA,Universityof Lie ` ge,Lie ` ge,Belgium
ChristofKoch AllenInstituteforBrainScience,Seattle, WA,USA
AndreaKubler InstituteofPsychology,Universityof Wurzburg,Wurzburg,Germany
RonKupers DepartmentofNeuroscienceandPharmacology, UniversityofCopenhagen,Copenhagen,Denmark
StevenLaureys ComaScienceGroup,Neurology DepartmentandGIGA,UniversityofLie ` ge,Lie ` ge, Belgium
NicoleL.Marinsek DynamicalNeuroscience,Universityof California,SantaBarbara,CA,USA
GeorgeA.Mashour DepartmentofAnesthesiology, NeuroscienceGraduateProgram,Centerfor ConsciousnessScience,UniversityofMichiganMedical School,AnnArbor,MI,USA
MarcelloMassimini DepartmentofBiomedicaland ClinicalSciences“LuigiSacco,”UniversityofMilan, Milan,Italy
DonatellaMattia FondazioneSantaLucia,IRCCS,Rome, Italy
MichaelB.Miller DynamicalNeuroscience,Universityof California,SantaBarbara,CA,USA;Psychologicaland BrainSciences,UniversityofCalifornia,SantaBarbara, CA,USA
LionelNaccache INSERMU1127,InstitutduCerveauetde laMoelleEpinie ` re,PICNICLab,Paris,France,Faculte ´ de Me ´ decinePitie ´ -Salpe ˆ trie ` re,Universite ´ PierreetMarie Curie,Paris,France;DepartmentsofNeurologyandof ClinicalNeurophysiology,AssistancePublique Ho ˆ pitaux deParis,GroupehospitalierPitie ´ -Salpe ˆ trie ` reCharlesFoix, Paris,France
PaoloNichelli DepartmentofBiomedical,Metabolicaland NeuralSciences,UniversityofModenaandReggio Emilia,Emilia-Romagna,Italy
Marie-ChristineNizzi PsychologyDepartment,Harvard University,Cambridge,MA,USA
AdrianM.Owen TheBrainandMindInstitute,The UniversityofWesternOntario,London,ON,Canada
PietroPietrini ClinicalPsychologyBranch,Departmentof Neuroscience,UniversityofPisaMedicalSchool,Pisa, Italy
BradleyR.Postle DepartmentsofPsychologyand Psychiatry,UniversityofWisconsin-Madison,Madison, WI,USA
MauricePtito DepartmentofNeuroscienceand Pharmacology,UniversityofCopenhagen,Copenhagen, Denmark
GeraintRees FacultyofLifeSciences,SchoolofLifeand MedicalSciences,UniversityCollegeLondon,London,UK
MarioRosanova DepartmentofBiomedicalandClinical Sciences“LuigiSacco,”UniversityofMilan,Milan,Italy; FondazioneEuropeadiRicercaBiomedica,FERBOnlus, Milan,Italy
EricSalmon CyclotronResearchCentre,Universityof Liege,Belgium
NicholasD.Schiff DepartmentofNeurologyand Neuroscience,WeillMedicalCollegeofCornell University,NewYork,NY,USA
CarolineSchnakers DepartmentofPsychologyand Neurosurgery,UniversityofCalifornia,LosAngeles,CA, USA
FrancescaSiclari CenterforInvestigationandResearchon Sleep,UniversityHospitalandUniversityofLausanne, Switzerland
GiulioTononi DepartmentofPsychiatry,Universityof Wisconsin-Madison,Madison,WI,USA
NaotsuguTsuchiya SchoolofPsychologicalSciences, FacultyofBiomedicalandPsychologicalSciences,Monash University,VIC,Australia
PatrikVuilleumier LaboratoryforBehavioralNeurology andImagingofCognition,DepartmentofNeurosciences andClinicofNeurology,UniversityMedicalCenter, Geneva,Switzerland
SusanWhitfield-Gabrieli MartinosImagingCenterat McGovernInstituteforBrainResearch,Massachusetts InstituteforTechnology,Cambridge,MA,USA
AdamZeman DepartmentofNeurology,Universityof ExeterMedicalSchool,Exeter,UK
NeuroanatomicalBasisofConsciousness
HalBlumenfeld
DepartmentsofNeurology,Neurobiology,andNeurosurgery,YaleUniversitySchoolofMedicine, NewHaven,CT,USA
OUTLINE
Introduction3
TheConsciousnessSystem5
SubcorticalNetworksandConsciousness6
TheThalamusandConsciousness8
GlutamatergicandRelatedArousalSystems9
CholinergicArousalSystems9
GABAergicArousalSystems12
NoradrenergicArousalSystems12
SerotoninergicArousalSystems13
DopaminergicArousalSystems13
HistaminergicArousalSystems14
OrexinergicArousalSystems15
AdenosineandArousal15
AmygdalaandArousal15
AttentionandAwareness:RolesofSubcortical ArousalSystems,TectalRegion,BasalGanglia, Claustrum,andCerebellum15
INTRODUCTION
Consciousnessisofgreatimportancetonormal humanqualityoflife.Thenatureofconsciousnessand thebestwaytounderstandanddefineithavelong generatedlivelydebateamongscientists,philosophers, clinicians,andthegeneralpublic.Fromaneurological perspective,consciousnessisclassicallydescribedas emergingfrombrainsystemsthatmakeupthe content of consciousness,regulatedbydistinctsystemsthatcontrol the level ofconsciousness(PlumandPosner,1982).
Thecontentofconsciousnessisthesubstrateupon whichlevelsofconsciousnessact.Thiscontentincludes
CorticalNetworksandConsciousness16 TheCortexandArousal16 AttentionandConsciousness16 HemisphericDominanceofAttention17 Affect,Motivation,andAttention18 TheBindingProblem18 Top-DownandBottom-UpAttentionNetworks19 Task-PositiveandTask-NegativeNetworks19 MemorySystemsandConsciousness20 VolitionandConsciousFreeWill21 Self-AwarenessandEmbodiment22 Awareness:ConsciousReportand ContrastiveAnalysis22 Acknowledgments23 References23
allthevarioustypesofinformationprocessedby hierarchicallyorganizedsensory,motor,emotional,and memorysystemsinthebrain(Figure1.1).Muchof neuroscienceisdedicatedtounderstandingthenormal functioningofthesesystems.Selectivedeficitsincontentsofconsciousness,suchaslossofaportionofone’s visualfield,orsuddenimpairmentinspokenlanguage, arealsothemainsubjectmatterofclinicalneurology. However,levelofconsciousnesscanaffectallof thesespecificfunctions.Thelevelofconsciousnessis controlledbyspecializedcorticalandsubcorticalsystemsthatdeterminetheamountofalertness,attention, andawareness(mnemonic,AAA)(Blumenfeld,2002).
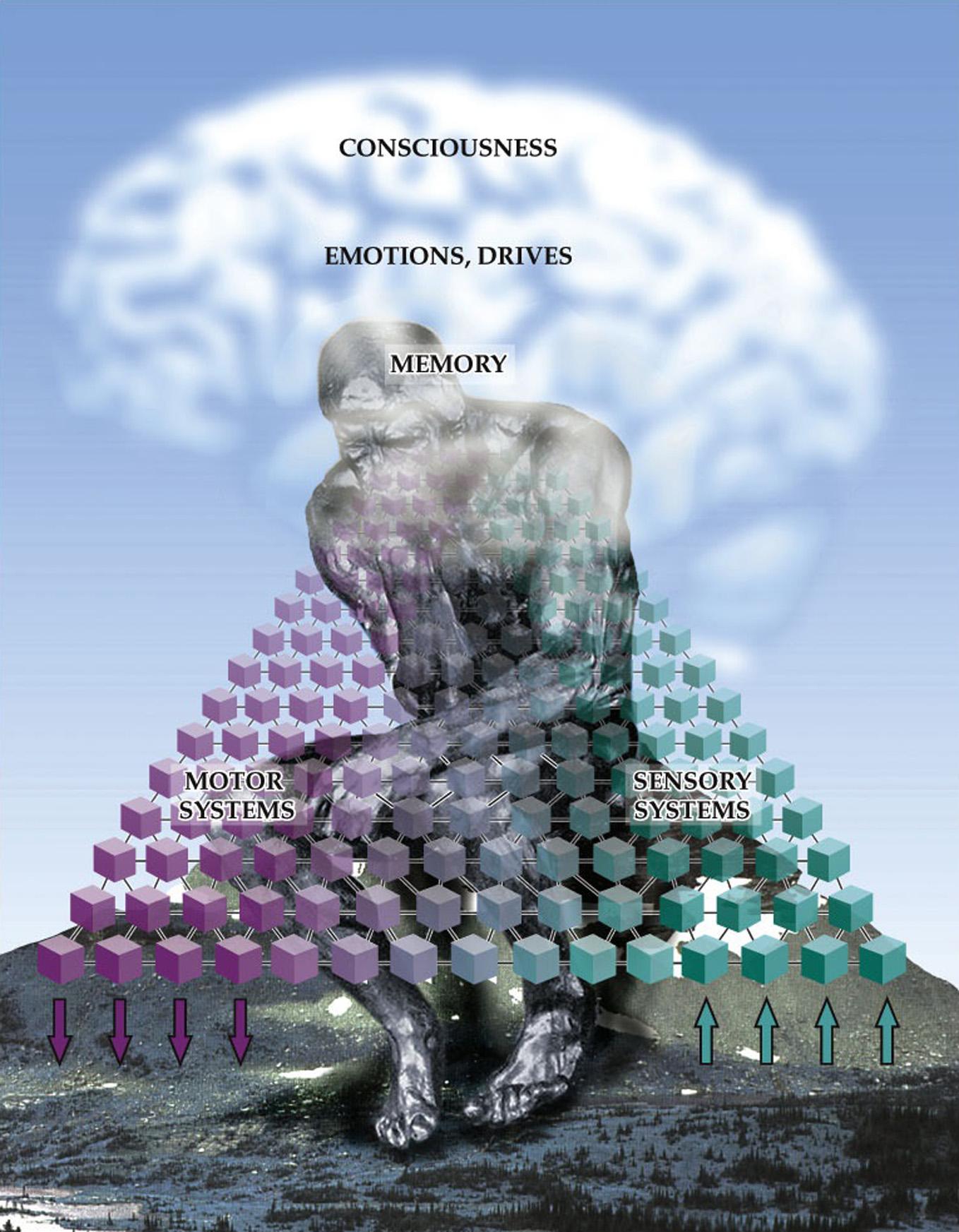
FIGURE1.1 Thecontentofconsciousness.Parallelinterconnectedandhierarchicallyorganizedsensoryandmotorsystemsreceiveinputs, generateoutputs,andperforminternalprocessingonmultiplelevels,fromrelativelysimpletohighlyabstract.Threeadditionalspecial systems—mediatingmemory,emotionsanddrives,andconsciousnessitself—actontheothersystemsinawidelydistributedmanner, especiallyatthehighestlevelsofprocessing. Source:Modifiedwithpermissionfrom Blumenfeld(2010)
Basicalertness(arousal,wakefulness)isnecessaryfor anymeaningfulresponsestooccur.Attentionenables selectiveorsustainedinformationtobeprocessed. Finally,awarenessistheabilitytoformexperiences thatcanpotentiallybereportedlater.Thischapterwill reviewtheneuroanatomicalbasisofbrainsystemsthat controlthelevelofconsciousness.Inanalogywith othercortical-subcorticalsystemssuchasthesensory, motororlimbicsystems,thebrainnetworksdedicated toregulatingthelevelofconsciousnesscanbereferred toasthe“consciousnesssystem”(Blumenfeld,2010, 2012).Thischapterbeginswithanoverviewofthe maincorticalandsubcorticalstructuresthatconstitute theconsciousnesssystem.Next,themajorsubcortical networksthatregulatelevelofconsciousnessareeach
discussedinturn,includingthethalamusandsubcorticalarousalnucleiactingthroughmultipleneurotransmitters(glutamate,acetylcholine,gammaamino butyricacid(GABA),norepinephrine,serotonin,dopamine,histamine,orexin)thatarisefromtheupper brainstem,basalforebrain,andhypothalamus.The secondhalfofthechapterreviewsimportantcortical networksforcontrollingthelevelofalertness,attention,andawareness,includingsystemsthatselectand encodeconsciousexperiencesintomemoriesforsubsequentreport.Thisneuroanatomicalreviewofthe corticalandsubcorticalsystemsthatcontrollevelof consciousnesswillserveasageneralintroductiontothe normalfunctionsaswellasdisordersofconsciousness discussedintheremainingchaptersinthisbook.
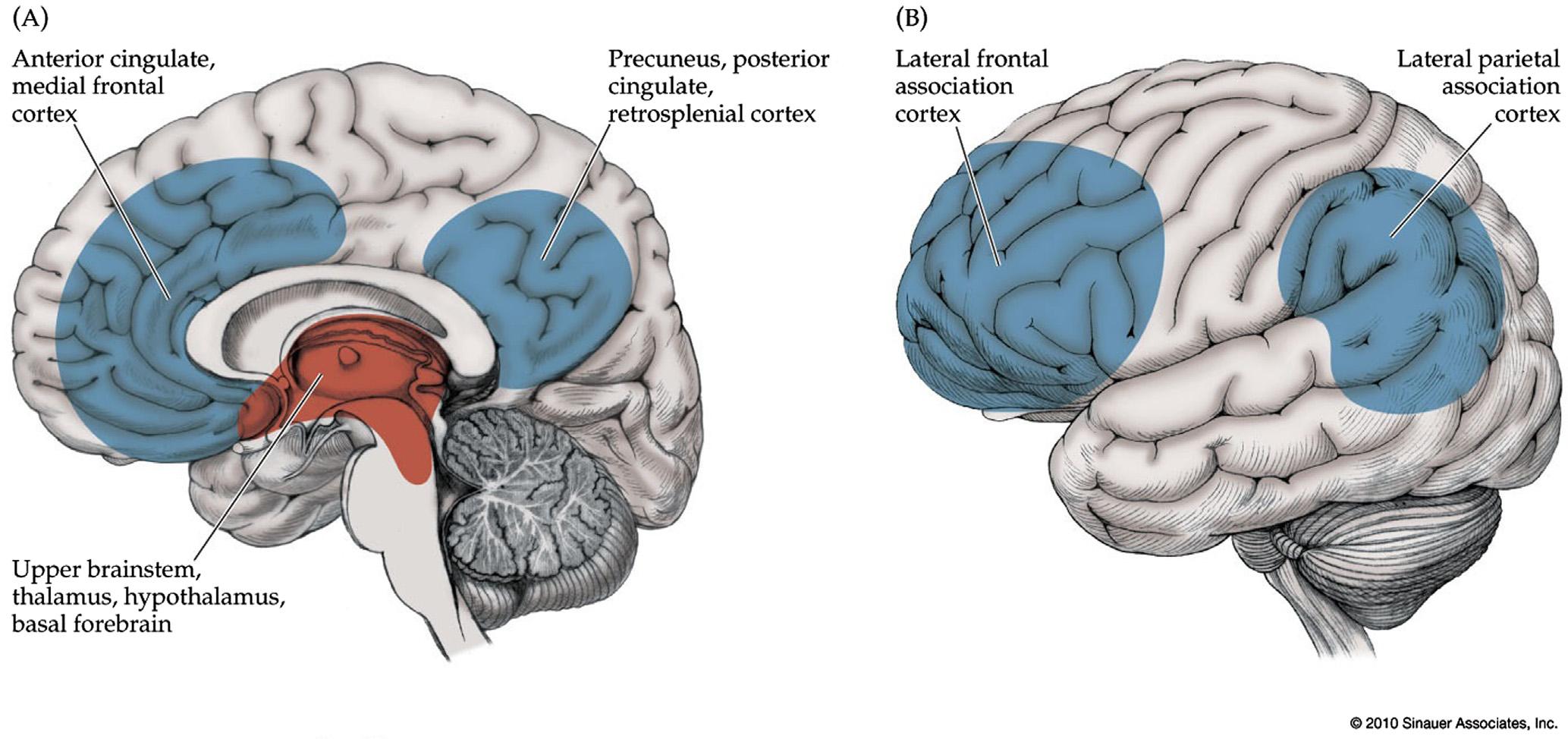
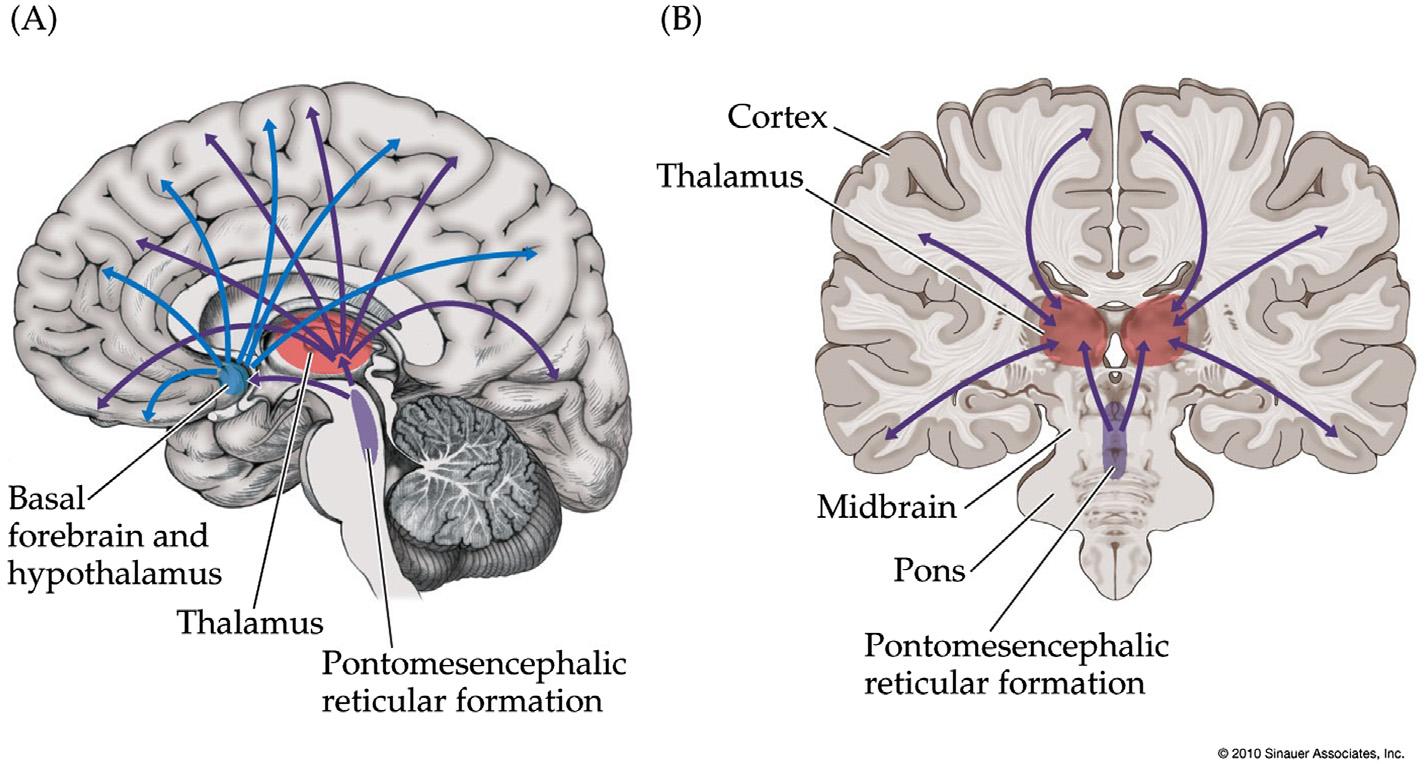
FIGURE1.2 Theconsciousnesssystem.Anatomicalstructuresinvolvedinregulatingthelevelofconsciousness,specificallycontrollingthe levelofalertness,attentionandawareness.(A)Medialviewshowingcortical(blue)andsubcortical(red)componentsoftheconsciousnesssystem.(B)Lateralcorticalcomponentsoftheconsciousnesssystem.Notethatothercircuitsnotpicturedhere,suchastheanteriorinsula,claustrum,basalganglia,amygdala,andcerebellum,mayalsoplayaroleinattentionandotheraspectsofconsciousness. Source:Reproducedwith permissionfrom Blumenfeld(2010)
THECONSCIOUSNESSSYSTEM
Thespecializedbrainnetworkscontrollingthelevel ofconsciousnesscanbereferredtoasthe“consciousnesssystem”(Blumenfeld,2009,2010)(Figure1.2). Ithaslongbeenrecognizedthroughstudiesbased onhumanbraindisorders(Penfield,1950;Plumand Posner,1972;VonEconomo,1930)aswellasexperimentalanimalmodels(Bremer,1955;MoruzziandMagoun, 1949;SteriadeandMcCarley,2010)thatthelevelof consciousnessdependscriticallyonbothcorticaland subcorticalstructures.Hereweprovideabriefoverview ofthecorticalandsubcorticalnetworkscomprising theconsciousnesssystem,whichwillbediscussedin greaterdetailintheremainingsectionsofthechapter.
Corticalcomponentsoftheconsciousnesssystem includethemajorregionsofthehigher-order“heteromodal”(Mesulam,2000)associationcortex(Figure1.2;see also Figure1.11).Onthemedialbrainsurface,important componentsarethemedialfrontal,anteriorcingulate, posteriorcingulate,andmedialparietal(precuneus, retrosplenial)cortex(Figure1.2A).Onthelateral surface,majorconsciousnesssystemnetworksinclude thelateralfrontal,anteriorinsula,orbitalfrontal,and lateraltemporal-parietalassociationcortex(Figure1.2B).
Itisimportanttorecognizethatindividual componentsofthehigher-orderassociationcortexplay importantandwell-studiedrolesinspecificcognitive functionsinthedominantandnon-dominanthemispheresasdescribedinthebehavioralneurology
literature(HeilmanandValenstein,2003;Mesulam, 2000).Recentlythesesameassociationcortexregions havealsobeendescribedasparticipatingineithersocalledtask-positivenetworksbasedontheiractivation duringexternallyorientedattention(Asplundetal., 2010;BuschmanandMiller,2007;Dosenbachetal., 2007;Vanhaudenhuyseetal.,2011)ortask-negative networks,alsoknownasthe“defaultmode”basedon activityatrest(Foxetal.,2005;Raichleetal.,2001). Regardlessoftheheterogeneousfunctionsofindividual regionsornetworks,itisthe collective activityofwidespreadareasofbilateralassociationcortexthatdeterminesthelevelofconsciousness.Takenasawhole,the higher-orderassociationcortexinteractswithsubcorticalarousalsystems(SteriadeandMcCarley,2010)to exertpowerfulcontrolovertheoveralllevelofarousal, attention,andawareness.
Subcorticalcomponentsoftheconsciousnesssystem includetheupperbrainstemactivatingsystems,thalamus,hypothalamus,andbasalforebrain(Figure1.2A). Itislikelythatothersubcorticalstructures(notshown) alsoparticipate,includingportionsofthebasalganglia,cerebellum,amygdala,andclaustrum.Multiple parallelneurotransmittersystemsparticipatein subcorticalarousalincludingacetylcholine,glutamate, gammaaminobutyricacid(GABA),norepinephrine, serotonin,dopamine,histamine,andorexin(Cooper etal.,2003;Saperetal.,2005;Steriadeetal.,1997; SteriadeandMcCarley,2010).Likethediversecortical regionsalreadydiscussed,thesesubcorticalpathways
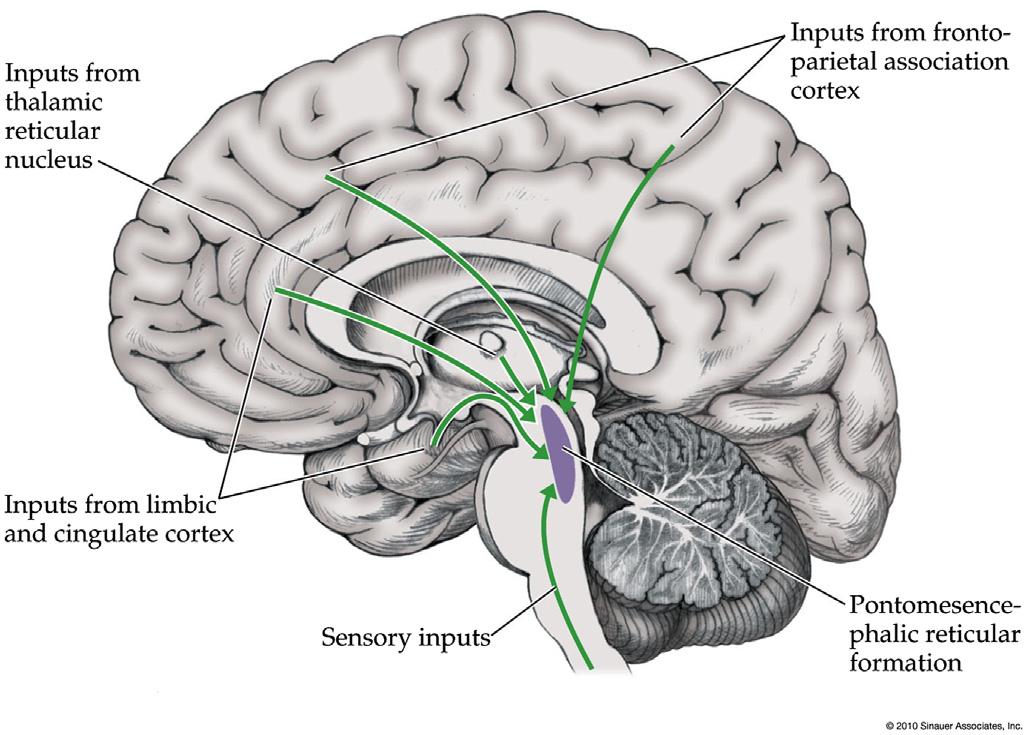
FIGURE1.3 Arousalcircuitsofthepontomesencephalicreticularformation,thalamus,hypothalamusandbasalforebrain.(A)Midsagittal view;(B)coronalview.Widespreadprojectionstothecortexarisefromoutputsofthepontomesencephalicreticularformationrelayedviathe thalamicintralaminarnuclei,basalforebrain,andhypothalamus. Source:Reproducedwithpermissionfrom Blumenfeld(2010)
eachcarryoutindividualroles,butitisthecollective andparallelactionsofallofthesesystemsthat togethercontrolthelevelofconsciousness.
Understandingconsciousnessdependsnotonlyon neuroanatomybutalsoonneurophysiology.Although thischapterwillfocusonthe“where”ofconsciousness, equallyimportantis“how”thesenetworksinteractto formconsciousness.Recentproposedphysiological mechanismsforconsciousnessincludesynchronized oscillations(BuzsakiandWang,2012;Llina ´ sandPare ´ , 1997;Singer,1998),slowcorticalpotentials(Lietal., 2014),connectivity(Bolyetal.,2011;Rosanovaetal., 2012;RubinovandSporns,2010),information integration(Tononi,2005;TononiandKoch,2008),populationneuroenergetics(Shulmanetal.,2003),and recurrentorglobalneuronalprocessing(Dehaeneetal., 1998;LammeandRoelfsema,2000;SergentandDehaene, 2004)amongothers.Muchadditionalworkisneeded beforethephysiologicalmechanismsofconsciousness aremoredefinitelyknown.Bycontrast,whenitcomesto neuroanatomy,thepastcenturyofresearchhasatleast ledtoabasicunderstandingofthemostimportantbrain structurescontributingtoconsciousness.Wenowturnin greaterdetailtothesemajorcorticalandsubcorticalnetworksthatconstitutetheconsciousnesssystem.
SUBCORTICALNETWORKS ANDCONSCIOUSNESS
Themainsubcorticalcomponentsoftheconsciousnesssystemincludethemidbrainandupperpons,thalamus,hypothalamus,andbasalforebrain(Figure1.2). Thesestructurescontributeimportantlytomaintaining
alertnessandarousal.Attentionandawarenessarealso facilitatedbythesamemidlinearousalsystems,aswellas byothersubcorticalnetworksincludingthesuperiorcolliculi,cerebellum,amygdala,basalganglia,claustrum,and thalamicreticularnucleus(CrickandKoch,2005;Dreher andGrafman,2002;Krauzlisetal.,2013;O’Halloranetal., 2012;ZikopoulosandBarbas,2012).
Intermsofalertnessandarousal,muchhasbeen learnedaboutthebasicanatomyofconsciousnessby understandingwhichbrainlesionscancausecoma. Comaisasastateofunarousableunresponsivenessin whichtheeyesareclosedandnopurposefulresponses canbeelicited(Fisher,1969;PlumandPosner,1972). Comaoccurseitherthroughbilateraldamagetowidespreadcorticalareas,orvialesionsinacoresetofstructureslyinginupperbrainstemandmedialdiencephalon. Thesecriticalsubcorticalarousalstructureswereinitially identifiedbasedonstrokesandotherlocalizeddisorders inhumanpatients(Penfield,1950;PlumandPosner, 1972;VonEconomo,1930)aswellaslesion,disconnection,andstimulationexperimentsperformedinanimal models(Bremer,1955;MoruzziandMagoun,1949; SteriadeandMcCarley,2010).Inthebrainstem,the subcorticalarousalsystemsbeginintheupperpons andextendtothemidbrain.Lesionsinthissmallbut criticalregionoftheupperponsandmidbrainproduceprofoundcoma,whereaslesionsinthelower ponsormedulladonottypicallydisruptconsciousness(Figure1.3).Thecorebrainstemarousalsystems lieinthetegmentumandincludeavarietyofnuclei embeddedwithinthebrainstemreticularformation. Thetegmentumissandwichedbetweenthemore ventralbrainstembasis—containingascendingand descendingwhitematterpathways;andthemore
TABLE1.1 WidespreadProjectionSystemsintheNervousSystem
Projection systemLocation(s)ofcellbodiesMaintarget(s)
Reticular formation
Intralaminar nuclei
Midline thalamicnuclei
MidbrainandrostralponsThalamicintralaminarnuclei, hypothalamus,basalforebrain
Neurotransmitter receptor(s)a,b Function(s)c
Unknown (glutamate?)d
Alertness
ThalamicintralaminarnucleiCortex,striatum(Glutamate?)Alertness
MidlinethalamicnucleiCortex(Glutamate?)Alertness
NorepinephrinePons:locusceruleusandlateral tegmentalarea
DopamineMidbrain:substantianigra parscompactaandventral tegmentalarea
SerotoninMidbrain,pons,andmedulla: raphenuclei
HistamineHypothalamus:tubero-mammillary nucleus;midbrain:reticular formation
Orexin (hypocretin)
Striatum,limbiccortex, amygdala,nucleusaccumbens, prefrontalcortex
D1 5
EntireCNS5-HT1A F,5-HT2A C, 5-HT3 7
EntirebrainH1 4
PosteriorlateralhypothalamusEntirebrainOX1,OX2
AcetylcholineBasalforebrain:nucleusbasalis, medialseptalnucleus,andnucleus ofdiagonalband
Pontomesencephalicregion: pedunculopontinenucleusand laterodorsaltegmentalnucleus
Cerebralcortexincluding hippocampus
Thalamus,cerebellum, pons,medulla
Muscarinic(M1 5), nicotinicsubtypes
Muscarinic(M1 5), nicotinicsubtypes
Alertness,attention,mood elevation
Movements,initiative, workingmemory
Alertness,moodelevation, breathingcontrol
Alertness
Alertness,foodintake
Alertness,memory
Alertness,memory
aManyoftheneuronsreleasingtheneuromodulatorytransmitterslistedherealsoreleaseavarietyofpeptides,whicharelikelytoplayaneuromodulatoryroleaswell. bAdditionalreceptorsubtypesareconstantlybeingadded. cFunctionslistedarehighlysimplifiedhere;seereferencesattheendofthischapterforadditionaldetails. dEntriesinparenthesiswithquestionmarkareuncertain.
Source:Modifiedwithpermissionfrom Blumenfeld(2010).
dorsaltectum—lyingdorsaltothecerebralaqueduct orfourthventricle.Lesionsoutsidethebrainstem tegmentuminthebasisortectumdonotproduce coma.Bilaterallesionsofthethalamus,particularly intheintralaminarandmidlinethalamicnuclei,can alsoproduceprofoundsuppressionofarousal.
Subsequentworkhasrevealedthatthesubcortical arousalsystemsconsistofmultipleparallelneurotransmittersystemsandpathways(Figure1.3; Table1.1). Unlikemostpathwaysinthenervoussystemwhich projecttorelativelynarrowtargetregions,thesubcorticalarousalsystemsbelongtoasetofwidespread projectingsystems(Table1.1)thatreachmanystructuresoreventheentirenervoussystem.Interestingly theprojectionsystemsarisingfromtheupperbrainstemincludingthemidbrainandupperpons(pontomesencephalicreticularformation; Figure1.3)tend toprojectupwardtothecortex,diencephalon,andbasal forebrainwhilethoseinthelowerponsandmedullaprojectdownwardtothebrainstem,cerebellum, andspinalcord.Theupwardprojectingsystemswere
originallycalledthe“ascendingreticularactivatingsystem”(ARAS)(MoruzziandMagoun,1949)recognizing theirimportantroleinarousal.Althoughtheterm “ARAS”isstilloccasionallyused,inrealitythese arousalsystemsarisefromavarietyofspecificnuclei (Table1.1)ratherthanfromwhatwasformerlyconsideredasinglediffuselyorganizedsystem.
Thesubcorticalarousalsystemsinthemidbrainand upperponshavethreemaintargets(Figure1.3): (i)Putativeglutamatergicneuronsfromthereticular formationandcholinergicneuronsfromthepedunculopontinetegmentalnucleusandlaterodorsal tegmentalnucleus(LDT)projectmainlytothethalamus,particularlytothethalamicintralaminarnuclei which,inturn,increasecorticalarousal.(ii)Other neuronsprojecttothenucleusbasalisandhypothalamus,whichagainrelayarousalinfluencestothecortex. (iii)Finally,themonaminergicneurotransmitter systems(norepinephrine,dopamine,serotonin;not shownin Figure1.3)projectdirectlytotheentireforebrainincludingthecortexandsubcorticalstructures.
Majorinputstothepontomesencephalicreticular formationandrelatedstructures. Source:Reproducedwithpermission from Blumenfeld(2010)
Inadditiontotheseascendingconnections,thesubcorticalarousalsystemsarealsohighlyinterconnected andstronglyinfluenceeachother’sfunctionthrough multipleconnectionswithinandbetweenthebrainstem,thalamus,hypothalamus,andbasalforebrain.
Theupperbrainstemarousalsystemsareinfluenced byavarietyofinputsincludingnumerousregionsofthe associationcortexandlimbiccortex,aswellassensory pathwayssuchastheanterolateralpaintransmission pathways(Figure1.4).Inhibitoryinfluencesarisefrom thethalamicreticularnucleus(nottobeconfusedwith thereticularformation)aswellasotherGABAergic inputs(ParentandSteriade,1981,1984;Ropertand Steriade,1981).Thearousalsystemsarealsostrongly regulatedbybrainstemandhypothalamiccircuitscontrollingcircadiansleeprhythms(Saperetal.,2005,2010).
Thesubcorticalarousalsystemswillnoweachbe discussedingreaterdetailtomorefullyappreciate thefunctionalanatomyofthesecomplexparallel arousalsystemsandtheircontributionstoconsciousness.Itshouldbenotedthat,unlikegrosslesionsof thebrainstem-diencephalicarousalsystems,lesions orpharmacologicalblockadeoftheindividualprojectingneurotransmitters ystemsdonotcausecoma. Blockadeofsomeneurotransmitters,especiallyacetylcholineorhistamine,producessevereconfusion anddrowsiness,butnotcoma.Thus,thenormal awake,consciousstatedoesnotdependonasingle projectionsystem,butratherontheparallelactionof multipleanatomicalandn eurotransmittersystems actingtogether(Table1.1).Afterdiscussingeachof themajorsubcorticalarousalsystems,emphasizing theirroleinmaintainingthealertstate,wewillthen brieflydiscusstheirroletogetherwithothersubcorticalstructures(tectalregion,basalganglia,claustrum, cerebellum)inattentionandawareness.
TheThalamusandConsciousness
Nearlyallinformationdestinedforthecortexfirst reachesthethalamus.Thethalamustransmitsthis informationandthenreceivesanevengreaternumber ofreciprocalconnectionsbackfromthecortex. Thereforethethalamusplaysakeyroleinallaspects offorebrainfunctionincludingconsciousness.The thalamusrelaysthecontentofconsciousness,andalso controlsthelevelofconsciousnessthroughspecialized circuitsthatregulatethelevelofarousalandare crucialforselectiveattention.
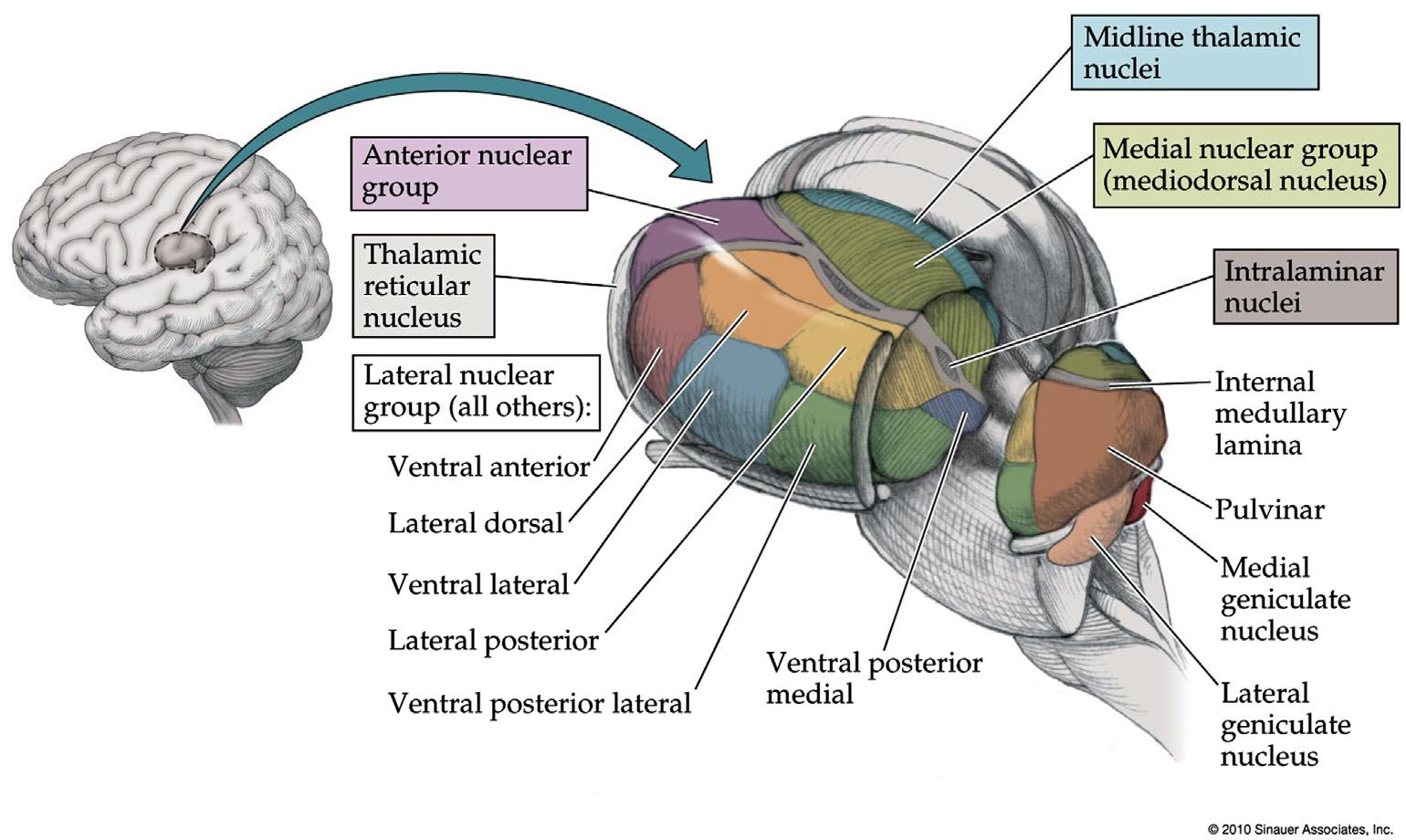
Organizationofthethalamuscanbedescribed basedonregionsorbasedonprojections.Theregional organizationofthethalamusdividesthethalamicsubnucleiproceedingfromlateraltomedial(Figure1.5) intothethalamicreticularnucleuslocatedmost laterally,followedbythelateralnucleargroupwhich containsthelargestnumberofthalamicrelaynuclei (Table1.2).Continuingmedially,nextcomesthe Y-shapedwhitematterinternalmedullarylamina whichseparatesthelateral,anterior,andmedial nucleargroupsfromeachother(Figure1.5).Embedded withintheinternalmedullarylaminalietheintralaminarnuclei.Finallyathinlayerofmidlinethalamic nucleiarelocatedmostmedially,adjacenttothethird ventricle(Figure1.5).
Projectionpatternscanalsobeusedtoclassifythe thalamicsubnuclei(Table1.2).Some,suchasthe ventralposteriorlateralnucleus,asomatosensory relayinthelateralthalamus,projecttoarelatively localizedregionofcortex,andarereferredtoas specificrelaynuclei.Others,suchasthethalamicintralaminarnuclei,havemorewidespreadprojectionsto manycorticalareas,andarecalleddiffuselyorwidely projecting(“nonspecific”)nuclei.
Thespecificthalamicrelaynucleicommunicatewith thecortexregardingeachsensoryandmotorfunction, andarethereforeresponsibleforalltheindividual contentsofconsciousness.Ontheotherhand,the widelyprojectingthalamicnucleiinfluencetheoverall levelofcorticalarousal,andthereforecontrolthe levelofconsciousness.Therostralintralaminarnuclei (centrallateral,paracentral,centralmedialnuclei; Table1.2)andmidlinethalamicnucleiarethoughtto beparticularlyimportantforactivatingthecortex (Figure1.3).Aswasalreadydiscussed,theintralaminar thalamusplaysakeyroleintransmittingarousalinfluencesfromthemidbrainandupperpontinecholinergic andglutamatergicsystemstothecortex.
Thethalamicreticularnucleusformsathinshellof predominantlyGABAergicinhibitoryneuronsonthe lateralthalamus(Figure1.5).Asaxonstraversethis nucleustravelingfromthalamustocortexorfrom cortexbacktothalamustheygiveoffcollateral
FIGURE1.4
FIGURE1.5 Thethalamus.Mainnucleardivisionsandnucleiareshown(seealso Table1.2).Theposteriorportionofthereticularnucleus hasbeenremoved. Source:Reproducedwithpermissionfrom Blumenfeld(2010)
branchestothethalamicreticularneurons.The thalamicreticularneurons,inturn,projecttothethalamusandinhibitthespecificthalamocorticalneurons correspondingtoindividualcorticothalamicloops. Thereciprocalconnectionsbetweenthalamicrelay nucleiandthethalamicreticularnucleusarethought toplayanimportantroleingeneratingcorticothalamicrhythmsduringnormalsleepandwaking activity,aswellasinpathologicalrhythmssuchas epilepsy(McCormick,2002;McCormickandBal,1997; McCormickandContreras,2001;Steriadeetal.,1993b). Thesephysiologicalrhythmsarecrucialforregulating thelevelofconsciousness.Inaddition,thethalamic reticularnucleusinfluencesarousalthroughlong-range inhibitoryprojectionstothepontomesencephalicreticularformation(ParentandSteriade,1984).Selective attentionmayalsobemediatedthroughtheparticular arrangementofreticularthalamicneuronsandtheir directedinhibitoryprojectionstothethalamus,which arecapableofgeneratinganinhibitorysurround arounda“searchlight”offocusedattentioninanarrow bandofthalamocorticalchannels(Crick,1984;Mayo, 2009;Pinault,2004).
GlutamatergicandRelatedArousalSystems
Glutamateisthemostprevalentexcitatoryneurotransmitterofthecentralnervoussystem.Formanyof thearousalsystemsthemostlikelyneurotransmitteris glutamate,althoughithasnotbeenidentifiedwith certainty(SteriadeandMcCarley,2010).Arousal systempathwaysprobablymediatedbyglutamate
includethosearisingfromthemidbrainandupper pontinereticularformationthatprojecttothethalamus andbasalforebrain(Steriade,2004;Steriadeetal., 1993a),aswellasthewidespreadprojectionsfromthe thalamicintralaminarnucleitothecortex(Figure1.3). Itisnotknownwhetherotherexcitatoryaminoacid neurotransmitterssuchasaspartatemightalsoplaya significantroleinarousal.
CholinergicArousalSystems
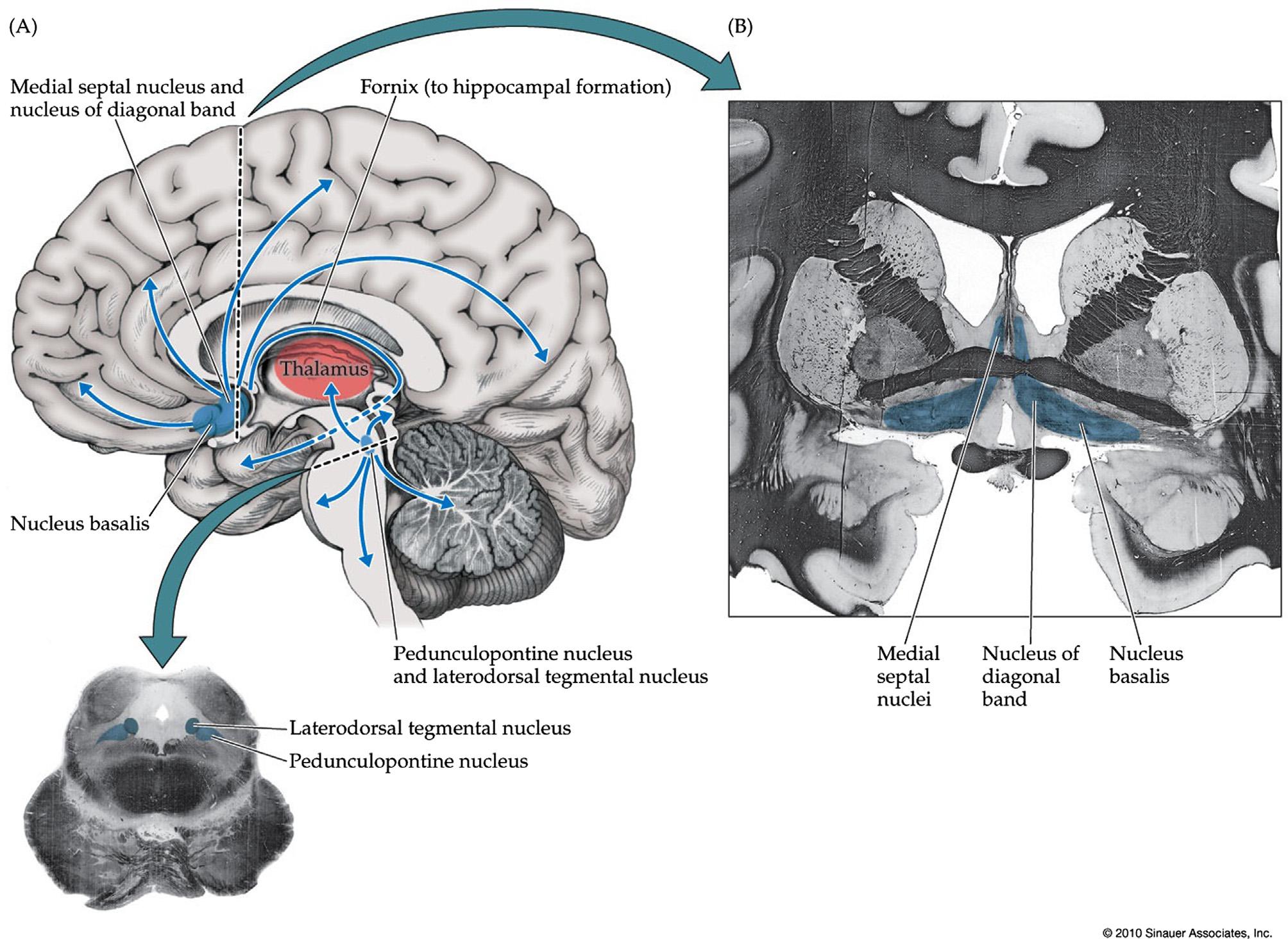
Acetylcholineisthemajorneurotransmitterofthe peripheralnervoussystem,butinthecentralnervoussystemithasamoreneuromodulatoryfunction,whereits roleinarousalhasbeenstudiedextensively.Thetwo mainsourcesofcholinergicprojectionsneuronsinthe centralnervoussystemlieinthebrainstempontomesencephalicreticularformationandinthebasalforebrain (Figure1.6; Table1.1).Atthejunctionofthemidbrainand pons,thepedunculopontinenucleusislocatedinthelateralreticularformation,whilethelaterodorsaltegmental nucleusliesintheperiaqueductalgray(Mesulametal., 1983).Thepedunculopontinenucleusstretchesfromthe caudalmidbrainsubstantianigra pars reticulataintothe rostralponstowardsthesuperiorcerebellarpeduncle (Mena-Segoviaetal.,2009;Ryeetal.,1987).Thenucleus hasagradientofincreasingcholinergicanddecreasing GABAergicneuronsasitextendscaudally,andalsocontainsglutamatergicneurons(WangandMorales,2009). Cholinergicneuronsfromthesebrainstemnucleiproject tothethalamus,includingtheintralaminarnuclei,playinganimportantroleinarousal.Brainstemcholinergic
Nucleia Maininputsb
RELAYNUCLEI
Lateralnucleargroup
Ventralposteriorlateral nucleus Mediallemniscus, spinothalamictract
Ventralposteromedial nucleus Trigeminallemniscus, trigeminothalamictract, tasteinputs
Mainoutputs
Diffusenessof projectionsto cortexc Proposedfunctions
Somatosensorycortex 1 Relayssomatosensoryspinal inputstocortex
Somatosensoryandtaste cortex 1
Relayssomatosensorycranial nerveinputsandtasteto cortex
LateralgeniculatenucleusRetinaPrimaryvisualcortex 1 Relaysvisualinputstocortex
MedialgeniculatenucleusInferiorcolliculusPrimaryauditorycortex 1 Relaysauditoryinputsto cortex
VentrallateralnucleusInternalglobuspallidus,deep cerebellarnuclei,substantia nigraparsreticulata
VentralanteriornucleusSubstantianigraparsreticulata, internalglobuspallidus,deep cerebellarnuclei
PulvinarTectum(extrageniculatevisual pathway),othersensoryinputs
Motor,premotor,and supplementarymotor cortex 1
Relaysbasalgangliaand cerebellarinputstocortex
Widespreadtofrontal lobe,including prefrontal,premotor, andsupplementary motorcortex 111 Relaysbasalgangliaand cerebellarinputstocortex
Parietotemporo-occipital associationcortex 11
Behavioralorientationtoward relevantvisualandother stimuli
LateraldorsalnucleusSeeanteriornucleus 11 Functionswithanteriornuclei
LateralposteriornucleusSeepulvinar 11 Functionswithpulvinar
VentralmedialnucleusMidbrainreticularformationWidespreadtocortex 111 Mayhelpmaintainalert, consciousstate
Medialnucleargroup
Mediodorsalnucleus(MD)Amygdala,olfactorycortex, limbicbasalganglia
Anteriornucleargroup
Frontalcortex 11 Limbicpathways,majorrelay tofrontalcortex
AnteriornucleusMammillarybody,hippocampal formation Cingulategyrus 1 Limbicpathways
Midlinethalamicnuclei
Paraventricular,parataenial, interanteromedial,intermediodorsal,rhomboid, reuniens(medialventral)
INTRALAMINARNUCLEI
Rostralintralaminarnuclei
Paracentralnucleus
Hypothalamus,basalforebrain, amygdala,hippocampus
Amygdala,hippocampus, limbiccortex 11 Limbicpathways
CentralmedialnucleusDeepcerebellarnuclei,globus pallidus,brainstemascending reticularactivatingsystems (ARAS),sensorypathways
Cerebralcortex,striatum 111 Maintainalertconsciousness; motorrelayforbasalganglia andcerebellum
Centrallateralnucleus
Caudalintralaminarnuclei
CentromediannucleusGlobuspallidus,ARAS,sensory pathways
Parafascicularnucleus
RETICULARNUCLEUS
ReticularnucleusCerebralcortex,thalamicrelay andintralaminarnuclei,ARAS
Striatum,cerebralcortex 111 Motorrelayforbasalganglia
Thalamicrelayand intralaminarnuclei,ARAS
NoneRegulatesstateofother thalamicnuclei
aSomeadditional,smallernucleihavenotbeenincludedhere. bInadditiontotheinputslisted,allthalamicnucleireceivereciprocalinputsfromthecortexandfromthethalamicreticularnucleus.Modulatorycholinergic,noradrenergic,serotonergic, andhistaminergicinputsalsoreachmostthalamicnuclei.
c 1 representsleastdiffuse(specificthalamicrelaynuclei); 11 representsmoderatelydiffuse; 111 representsmostdiffuse. Source:Modifiedwithpermissionfrom Blumenfeld(2010).
FIGURE1.6 Cholinergicprojectionsystems.(A)Overviewandinsetshowingaxialsectionthroughcaudalmidbrain.(B)Coronalsection throughbasalforebrain.Seealso Table1.1 Source:Reproducedwithpermissionfrom Blumenfeld(2010)
arousalisthoughttoactsynergisticallywithnoncholinergicputativeglutamatergicpontomesencephalic neuronsthatprojecttointralaminarthalamusandbasal forebrain(Figure1.3)(Rasmussonetal.,1994,1996; Steriade,2004;Steriadeetal.,1993a).Insleep,pontogeniculatewavesarisefromcholinergicbrainstemneurons projectingtothalamocorticalneuronsinthelateralgeniculatenucleus(Steriadeetal.,1989,1990).Thepedunculopontinenucleusalsohasnumerousascendingand descendingmotorprojectionsandisinvolvedin controllinglocomotion(InglisandWinn,1995).
Interestingly,thebrainstemhasveryfewdirect cholinergicprojectionstothecortexandnearlyall facilitatoryeffectsofthebrainstemcholinergicsystems oncortexaremediatedviathethalamus(Beninatoand Spencer,1987;Cornwalletal.,1990;Hallangeretal., 1987;HallangerandWainer,1988;Heckersetal.,1992; JonesandWebster,1988;SatohandFibiger,1986).The majorsourceofcholinergicinputtothecortexisthe basalforebrain(Figure1.6; Table1.1).Cholinergic neuronsinthenucleusbasalisofMeynertand
surroundingregions(substantiainnominata,globus pallidus,andpreopticmagnocellularnucleus)notonly projecttoalmosttheentireneocortex(Mesulametal., 1983;Ryeetal.,1984)butalsoinnervatesomenucleiin thethalamus(reticularthalamic,mediodorsal,anteroventral/anteromedial,andventromedialnuclei) (Heckersetal.,1992;Parentetal.,1988;Steriadeetal., 1987).Thehippocampalarchicortex,however, receivescholinergicinputsmainlyfromthemedial septalnucleiandnucleusofthediagonalbandof Broca(Ryeetal.,1984).Additionalcholinergicneuronslieinthemedialhabenula,andshort-rangecholinergicneuronsarepresentinthestriatumandtoa morelimitedextentwithinthecortexitself.Likethe brainstemcholinergicnuclei,thebasalforebraincontainsbothcholinergicandnon-cholinergicneurons, includingGABAandglutamateastransmitters amongothers(Brashearetal.,1986;AlvaroDuque etal.,2007).
Thebrainstemandbasalforebraincholinergic systemsworktogethertoabolishcorticalslowwave
activityandpromoteanalertstate(Dringenbergand Olmstead,2003;Steriade,2004).Cholinergicarousalin thecentralnervoussystemismediatedpredominantly bymuscarinicacetylcholinereceptors,althoughnicotinicreceptorsmayalsoplayanimportantrolein arousalandattention(Bloemetal.,2014).Aswouldbe expectedfromtheconnectionsofcholinergicneurons describedhere,pharmacologicalblockadeofcentral cholinergicneurotransmissionproducesanacutestate ofdeliriumandmemoryloss.However,despitethe importanceofacetylcholineinconsciousness,selective damagetocholinergicneurotransmissiondoesnotproducecoma(Blanco-Centurionetal.,2006,2007;Buzsaki etal.,1988;Fulleretal.,2011),presumablybecause ofthemultipleparallelneurotransmittersystems participatinginconsciousnessasalreadydiscussed.
GABAergicArousalSystems
Foundinlocalinhibitoryinterneuronsthroughout thecortexandsubcorticalstructures,GABAisthe mostprevalentinhibitoryneurotransmitterinthecentralnervoussystemandplaysamajorroleinregulatingarousal.Severallong-rangeGABAergicprojection systemsalsocontributetocontrollingarousal.Some GABAergicneuronsinthebasalforebrainarethought topromotearousalbecausetheseinhibitoryneuronsin turnprojecttocorticalinhibitoryinterneurons(Freund andMeskenaite,1992).However,theoveralleffectsof basalforebrainGABAergicneuronsonarousalmay beheterogeneousbecauseofvariablefiringpatterns withrespecttocorticalactivationandsleep-wake cycles(Hassanietal.,2009;Jones,2004;Mannsetal., 2000);andbecauseparvalbumin-containingGABAergic neuronsarerelatedtocorticaldesynchronywhereas neuropeptideY-containingneuronsmayhavethe oppositeeffect(AlvaroDuqueetal.,2000,2007).
Otherimportantlong-rangeGABAergicprojections mainlyinhibitarousal.Theseincludeneuronsinthe
ventrallateralpreopticnucleus,whichhavewidespreadinhibitoryprojectionstovirtuallyallsubcortical arousalsystems(Saperetal.,2010);lateralseptal GABAergicneuronsthoughttoinhibitthebasalforebrainandhypothalamus(MesulamandMufson,1984; VaroqueauxandPoulain,1999);andthethalamic reticularnucleuswhichcontainsGABAergicneurons projectingbothtotheremainderofthethalamusand tothebrainstemreticularformation(Parentand Steriade,1984;Steriadeetal.,1984).Inaddition, GABAergicneuronsintheglobuspallidusinternal segmentinhibitregionsofthethalamusincludingthe intralaminarnuclei.IthasbeenproposedthatparadoxicalarousaleffectsofGABAagonistssuchaszolpidem inminimallyconsciousstate,orbenzodiazepinesin catatoniamayoccurwhentheseagentsinhibittheglobuspallidus,therebyremovingtonicinhibitionofthe intralaminarthalamus(Brownetal.,2010;Giacino etal.,2014).ActivationofthesemultipleGABAergic inhibitoryprojectionsconvergingonthesubcortical arousalsystemshasalsobeenproposedasthemechanismforlossofconsciousnessinpartialseizures (Blumenfeld,2012;EnglotandBlumenfeld,2009).
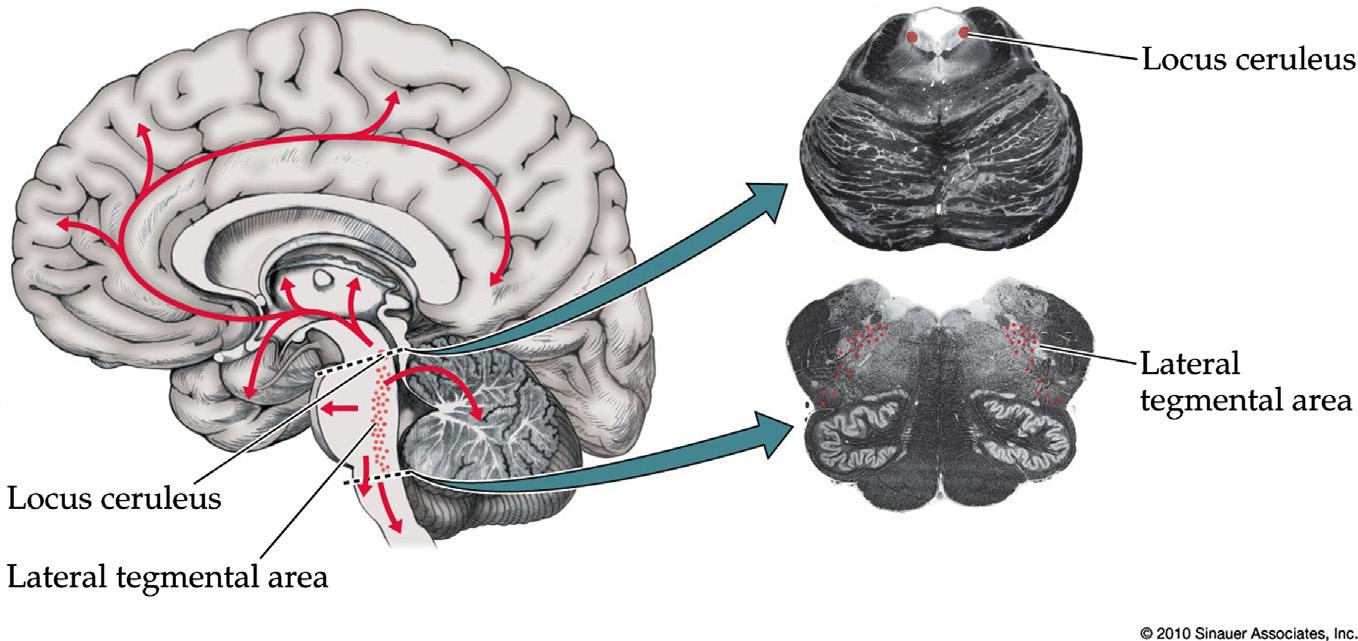
NoradrenergicArousalSystems
Norepinephrine(noradrenaline)-containingneurons arelocatedinthelocusceruleusintherostralpons adjacenttothefourthventricle,aswellasinthenearby lateraltegmentalareaextendingintothemorecaudal ponsandmedulla(Figure1.7; Table1.1).Ascending noradrenergicprojectionsreachthecortex,thalamus andhypothalamus(Footeetal.,1983;Morrisonetal., 1981;Pickeletal.,1974)toregulatesleep-wakecycles, attention,andmood,whiledescendingprojectionsto thebrainstem,cerebellum,andspinalcordmodulate autonomicfunctionandgatingofpain.Manyattentionenhancingdrugssuchasamphetaminesaugment noradrenergicfunction.Norepinephrineisthoughtto
FIGURE1.7 Noradrenergicprojectionsystems.Seealso Table1.1 Source:Reproducedwithpermissionfrom Blumenfeld(2010)
Table1.1 Source:Reproducedwithpermissionfrom Blumenfeld(2010)
playanimportantroleinpromotingarousal(Berridge, 2008;Berridgeetal.,2012;ConstantinopleandBruno, 2011).Forexample,selective α-2agonistssuchas clonidineortheanestheticagentdexmedetomidine markedlydepressarousalpossiblybyinhibitinglocus ceruleusneurons(Correa-Salesetal.,1992;DeSarro etal.,1987;ScheininandSchwinn,1992).However, selectiveremovalorblockadeofnoradrenergicneurons affectsarousalbutdoesnotproducecoma(Berridge etal.,1993;Blanco-Centurionetal.,2004,2007;Cirelli andTononi,2004;HunsleyandPalmiter,2003)again reinforcingthenotionofmultipleparallelsystems promotingconsciousness.
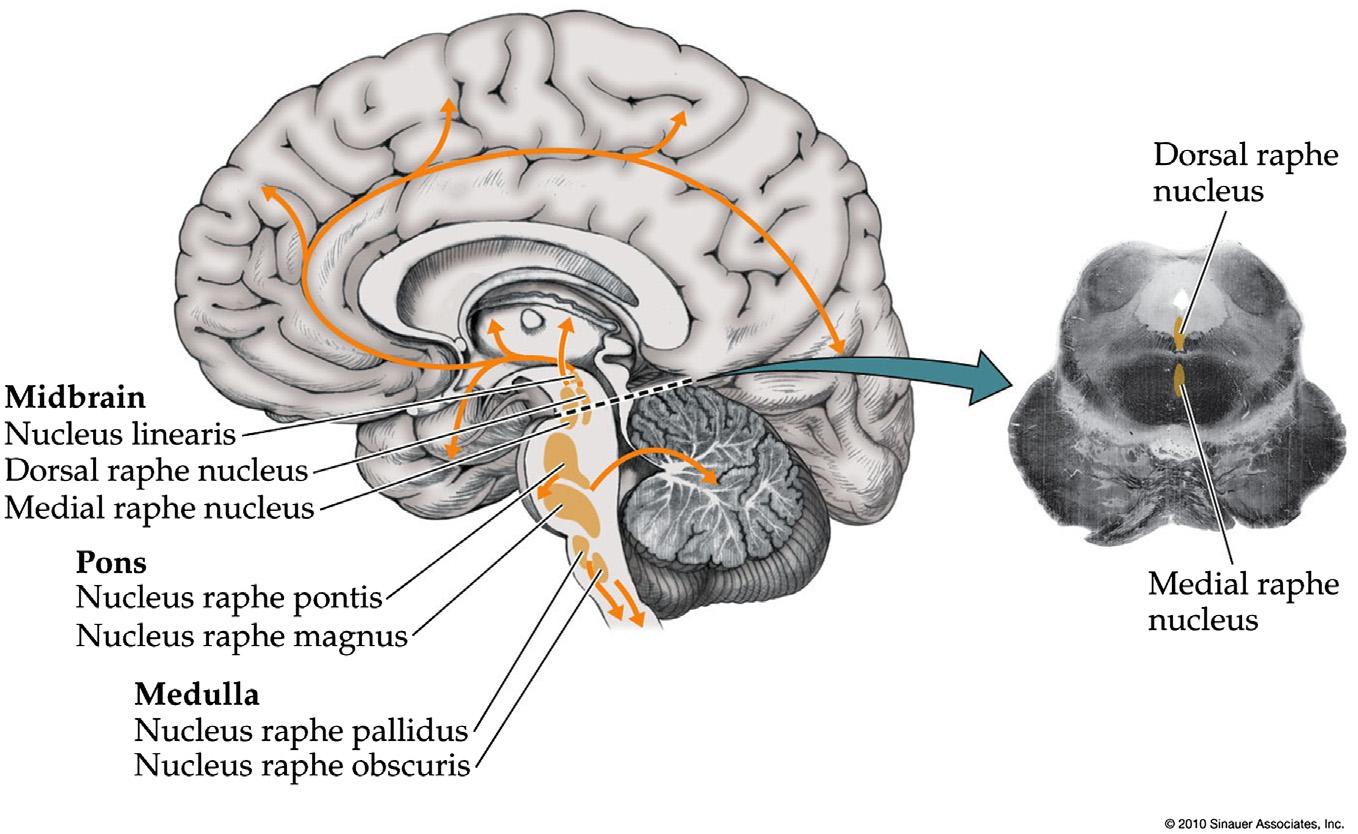
SerotoninergicArousalSystems
Serotonergicneuronsarefoundpredominantlyinthe midlineraphenucleiofthemidbrain,pons,andmedulla (Figure1.8; Table1.1).Themorerostralserotonergic neuronsinthemidbrainandupperpontineraphe nucleiprojecttotheentireforebrain,participatingin sleep-wakeregulation;dysfunctionofserotonergic systemsisthoughttoplayaroleinanumberof psychiatricdisordersincludingdepression,anxiety, obsessive-compulsivedisorder,aggressivebehavior,and eatingdisorders.Morecaudalserotonergicneuronsinthe ponsandmedullaareimportantformodulationof breathing,pain,temperature control,cardiovascular,and motorfunction.
Themostimportantrostralraphenucleiparticipating inarousalarethedorsalrapheandmedianraphe (JacobsandAzmitia,1992;Wiklundetal.,1981).Therole ofserotonergicneuronsinarousaliscomplex,possibly becausethelargediversityofserotoninreceptors (HannonandHoyer,2008)leadstoeffectsthateither
promoteorinhibitarousalindifferentbrainregions (Dugovicetal.,1989;Dzoljicetal.,1992;Kumaretal., 2007;Lemoineetal.,2007;LeonardandLlina ´ s,1994; Luebkeetal.,1992;MoncktonandMcCormick,2002; Murakietal.,2004;RogawskiandAghajanian,1980). Rostralbrainstemserotonergicneuronshavebeenproposedtopromotearousalinresponsetohypoventilation andincreasedcarbondioxidelevels,perhapsplayingan importantroleinpreventingsuddeninfantdeathsyndromeandsuddenunexplaineddeathinepilepsy (BuchananandRicherson,2010;Kinneyetal.,2009; RichersonandBuchanan,2011;Sowersetal.,2013).
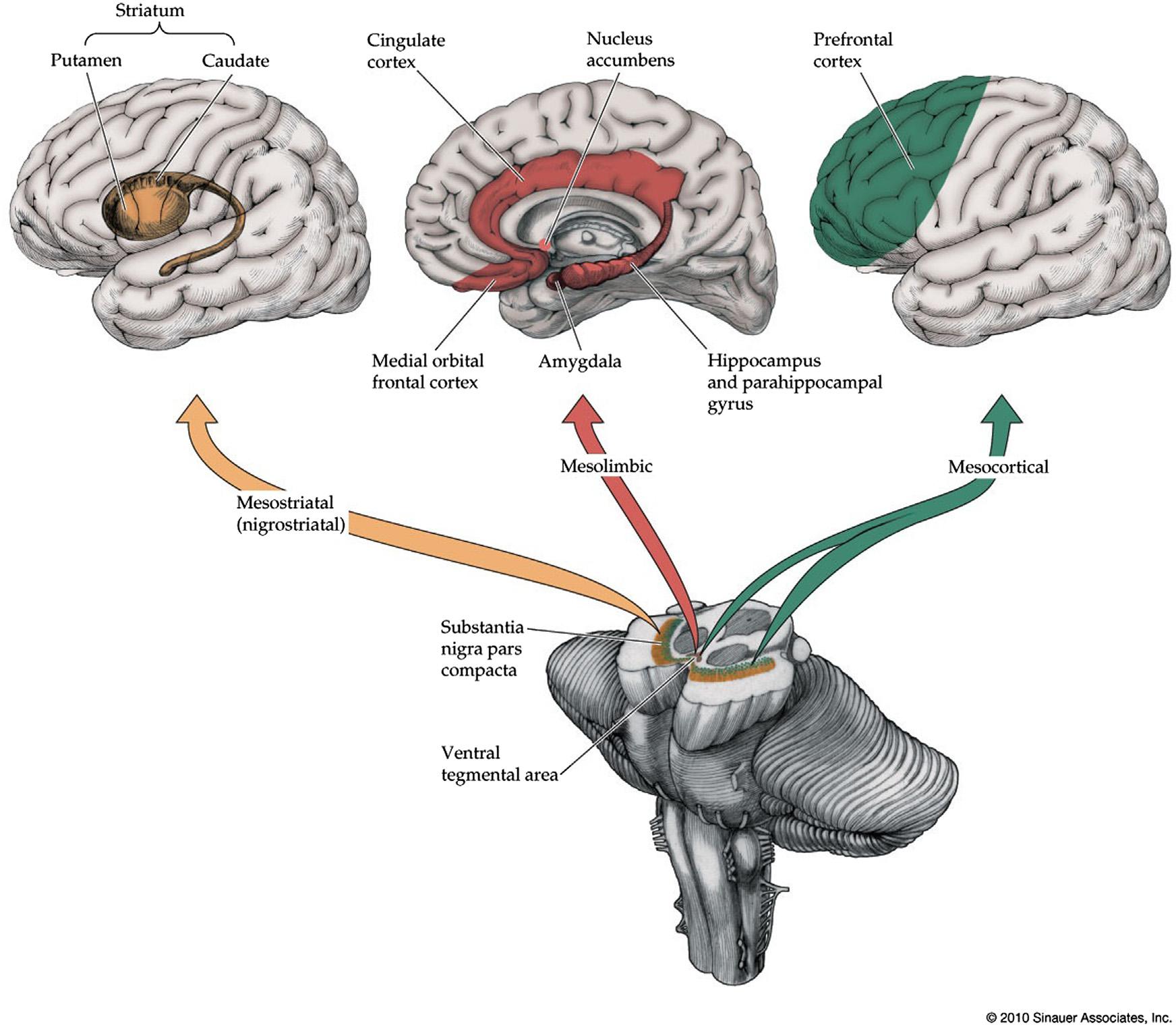
DopaminergicArousalSystems
Mostdopaminergicneuronsarelocatedinthe ventralmidbrain,eitherinthesubstantianigrapars compactaorintheadjacentventraltegmentalarea (Figure1.9; Table1.1).Thesemesencephalicnucleigive risetothefollowingthreeascendingdopaminergic projectionsystems:(i)themesostriatal(nigrostriatal) pathwayprojectsfromthesubstantianigratothe caudateandputamen(Matsudaetal.,2009);(ii)the mesolimbicpathwayarisesmainlyfromtheventral tegmentalareaandprojectstolimbicstructures includingthemedialtemporallobe,amygdala,cingulategyrus,septalnuclei,andnucleusaccumbens (Fallon,1981;OadesandHalliday,1987);(iii)the mesocorticalpathwayarisesmainlyfromtheventral tegmentalareaaswellasscatteredneuronsinthe vicinityofthesubstantianigraandventralperiaqueductalgray,projectingtotheprefrontalcortex (Figure1.9)aswellastothethalamus(Garcia-Cabezas etal.,2009;Groenewegen,1988;Luetal.,2006; Sanchez-Gonzalezetal.,2005).Dopaminergicneurons
FIGURE1.8 Serotonergicprojectionssystems.Seealso
intheventraltegmentalareaalsoprojectcaudallyto variousbrainstemnucleiandtothespinalcord(Oades andHalliday,1987).
Dopaminemaycontributetomaintainingthe wakingstateatleastinpartthrougheffectsonother subcorticalarousalcircuits(Deutchetal.,1986; Luetal.,2006;Neylanetal.,1992;Quetal.,2008; Volkowetal.,2009).Effectsofdopamineonthe thalamusandcortexcanbeeitherexcitatoryorinhibitory(BandyopadhyayandHablitz,2007;Govindaiah etal.,2010;LavinandGrace,1998;Penit-Soriaetal., 1987).Impaireddopaminergictransmissiontotheprefrontalcortexhasbeenproposedtobeimportantfor theapatheticnegativesymptomsofschizophrenia,and mayalsocontributetostatesofmarkedlyreduced motivation,initiativeandaction/intentionseenin frontallobedisorders,abulia,andakineticmutism (Combarrosetal.,2000;Kimetal.,2007;Yangetal., 2007).Amantadineimprovesarousalinchronicdisordersofconsciousness,althoughitisunclearwhether themechanismisthroughenhanceddopaminergic
neurotransmissionoreffectsofthismedicationon otherneurotransmittersystems(Giacinoetal.,2012).
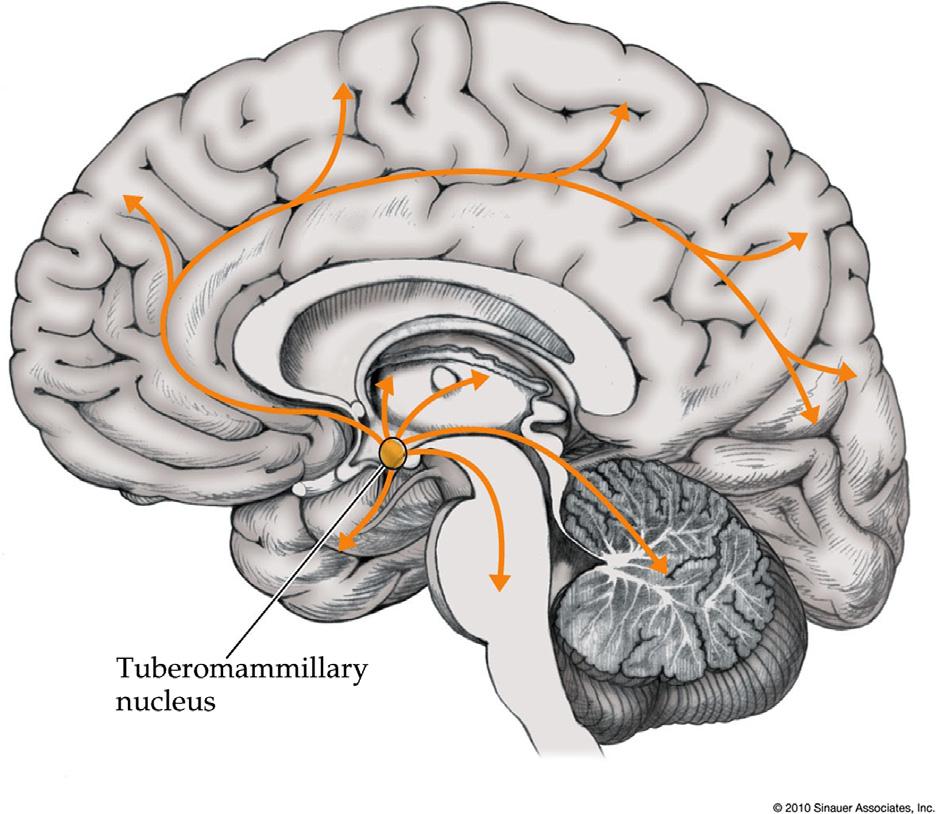
HistaminergicArousalSystems
Histamine-containingneuronsarefoundmainlyin thetuberomamillarynucleus(Panulaetal.,1984)of theposteriorhypothalamus(Figure1.10; Table1.1), althoughafewscatteredhistaminergicneuronsare alsofoundinthemidbrainreticularformation. Widespreadascendingprojectionsofhistaminergic neuronsfromthetuberomamillarynucleusreach nearlytheentireforebrainincludingcortexandthalamus,whiledescendingprojectionstargetthebrainstem andspinalcord(Brownetal.,2001;HongandLee, 2011;Linetal.,1996;Panulaetal.,1989).
Anti-histaminemedicationsareintendedtoacton peripheralhistaminereleasefrommastcells,butare well-knowntoinducedrowsinesspresumablythrough centralactions(WhiteandRumbold,1988).Histamine canproducearousaleffectsoncortex(Dringenberg
FIGURE1.9 Dopaminergicprojectionssystems.Seealso Table1.1 Source:Reproducedwithpermissionfrom Blumenfeld(2010)
FIGURE1.10 Histaminergicprojectionssystems.Seealso Table1.1. Source:Reproducedwithpermissionfrom Blumenfeld(2010).
andKuo,2003;McCormickandWilliamson,1989)and thalamus(McCormickandWilliamson,1991).Inadditiontothecortexandthalamus,arousalactionsof histaminemaybemediatedbyactivationofothersubcorticalarousalsystemsincludingotherhypothalamic nuclei(Brownetal.,2001;Linetal.,1994),thebasal forebrain(DringenbergandKuo,2003;Khatebetal., 1995;Zantetal.,2012),brainstemcholinergic(Khateb etal.,1990;Linetal.,1996),andnoradrenergicnuclei (Korotkovaetal.,2005).Effectsofhistamineare receptor-dependentasactivationofH1 receptorspromoteswakefulness,whereasH3-receptorsappearto havetheoppositerole(Huangetal.,2006;Khateb etal.,1990;Linetal.,1990,1996;Valjakkaetal.,1996).
OrexinergicArousalSystems
Orexin(hypocretin)isapeptideproducedin neuronsoftheperifornical,lateral,andposterior hypothalamus(Peyronetal.,1998;Sakuraietal.,1998), whichprojecttobothcortexandvirtuallyallsubcorticalarousalsystems(Chemellietal.,1999;Peyron etal.,1998)topromotetheawakestate.Thearousal effectsoforexinlikelyarisefrombothcorticaland subcorticalmechanisms(Bourginetal.,2000;Eriksson etal.,2001;Espan ˜ aetal.,2001;Haganetal.,1999; Horvathetal.,1999;Kiyashchenkoetal.,2002; Tsunematsuetal.,2011;vandenPoletal.,2002).
Abnormalitiesoftheorexinsystemarethought toplayaroleinnarcolepsy,adisordercharacterized byexcessivedaytimesleepinessandpathological transitionsintorapideyemovementsleep(Anacletetal., 2009;Chemellietal.,1999;Gerashchenkoetal., 2003;Haraetal.,2001;Linetal.,1999;Nishinoetal.,2000;
Peyronetal.,2000;Thannickaletal.,2000).Thebeneficial effectsofmodafenilinpreventingthesymptomsof narcolepsymayinpartbethroughactivationoforexin neurons(Chemellietal.,1999).
AdenosineandArousal
Althoughtheneuroanatomicalsourcesofadenosine arenotwellcharacterized,thisneuromodulatormay beimportantinmechanismsofconsciousarousal (Huangetal.,2014;LiuandGao,2007).Theeffectsof adenosineonarousalaregenerallyinhibitory,andcircadianfluctuationsinadenosinelevelspeakjustprior totheinitiationofsleep.Adenosinereceptorsare foundinbothcortexandthalamus,wheretheyhave anoverallinhibitoryfunctiononarousal.Caffeine blocksadenosinereceptorsandthismaybeoneimportantmechanismwherebycoffeepromotesalertness (Huangetal.,2011;Lazarusetal.,2011).
AmygdalaandArousal
Becausetheamygdalahaswidespreadandreciprocal cortical-subcorticalconnectionsthatcontributetoarousalparticularlyinresponsetoemotions,itisappropriatetoincludethiscomplexofnucleilocatedinthe anteromedialtemporallobeasanimportantsubcortical componentoftheconsciousnesssystem(Steriadeand McCarley,2010).Themaincomponentsoftheamygdaloidnuclearcomplexarethecorticomedial,basolateral, andcentralnuclei,aswellasthebednucleusofthe striaterminalis.Thebasolateralnucleusislargestin humansandhaswidespreaddirectandindirectconnectionstothecortex,basalforebrain,andmedialthalamus (Aggleton,2000;LeDoux,2007).Thesmallercorticomedialnucleusparticipatesinappetitivestatesviathe hypothalamus,aswellasinolfaction.Thecentral nucleus,althoughsmallest,hasimportantconnections withthehypothalamusandbrainstemparticipatingin arousalandautonomiccontrol(LeDoux,2007).
AttentionandAwareness:RolesofSubcortical ArousalSystems,TectalRegion,BasalGanglia, Claustrum,andCerebellum
Tocompleteourdiscussionofsubcorticalnetworks regulatingthelevelofconsciousness,itisimportantto againemphasizethefunctionsoftheconsciousness systemincontrollingalertness,attention,andawareness,andtobrieflymentionseveraladditionalsubcorticalstructuresparticipatinginthesefunctions.As wehavealreadydiscussed,thethalamusandother multipleparallelsubcorticalarousalsystemsinthe upperbrainstem,hypothalamus,andbasalforebrain (Table1.1)areessentialformaintainingthealertstate.
Thesesamesystemsalsoplayakeyroleincontrolling attentionandawarenessnotonlyinapermissivesense (e.g.,beinginacomaisnotcompatiblewithattention andawareness),butalsobyfacilitatingtheadditionalprocessingincorticalandsubcorticalnetworks necessaryforattentionandforawareness.
Severaladditionalsubcorticalstructuresalsoplay arole.Componentsofthetectalregion,specifically thesuperiorcolliculiandpretectalareaformanimportantcircuitalongwiththepulvinarofthethalamus todirectsaccadiceyemovementstowardssalient stimuli,andthesamecircuitsalsoparticipatein directedattention(Krauzlisetal.,2013).Thebasal gangliahavemajorreciprocalconnectionswiththe thalamicintralaminarnucleiandthiscircuitaswellas otherbasalgangliaconnectionsmaycontributeto arousalandattentionfunctions(DreherandGrafman, 2002;Hageretal.,1998;RingandSerra-Mestres,2002). Theclaustrumisathinlayerofneuronslocatedinthe whitematterbetweentheputamenandinsula,with widespreadcorticalconnectionsthathavebeenproposedtoplayanimportantroleintheattentionand awarenessaspectsofconsciousness(CrickandKoch, 2005).Finally,thecerebellumhasmajorreciprocal connectionswiththeprefrontalcortexandhasalso beenproposedtoparticipateinattention,although thisremainssomewhatcontroversial(Baumannand Mattingley,2014;Bischoff-Gretheetal.,2002;Dreher andGrafman,2002;O’Halloranetal.,2012).
CORTICALNETWORKS ANDCONSCIOUSNESS
Thecorticalcomponentsoftheconsciousnesssystem includewidespreadregionsofassociationcortexin thebilateralcerebralhemispheres,particularlyinthe lateralfrontal,anteriorinsula,lateralparietal(andadjacenttemporal-occipitalcortex),medialfrontal,medial parietal(precuneus)andcingulatecortex(Figure1.2). Ashasalreadybeendiscussed,individualcortical componentsoftheconsciousnesssystemhavespecific well-studiedfunctionsinbehavioralneurologywhich contributetothevariouscontentsofconsciousnessalong withspecificprimaryandsecondarysensorimotorand limbiccorticalareas(Figure1.11).However,itisthe collectiveactionofwidespreadbilateralassociation cortexregionsthatgivesrisetoregulationofthelevelof alertness,attention,andconsciousawareness.
Inthissectionwewillreviewthecontributionsof thecerebralcortextoarousalandthegenerationofan alert,awakestate.Wewillnextdiscussattentionin somewhatgreaterdetail,describingseveralformulationsofcorticalsystemsthatcontroldifferentaspects ofattention.Therelationshipbetweenmemory
systemsandconsciousnesswillthenbeconsidered,as wellastheroleofcorticalnetworksinself-awareness, planningvoluntaryactionandfreewill.Finally,we willreviewcorticalnetworksrevealedbycontrastive analysis(perceivedvsnon-perceivedstimuli)toplay animportantroleinconsciousawareness.
TheCortexandArousal
Themostimportantinputtosubcorticalarousal systems,includingthethalamus,hypothalamus, basalforebrain,andthemultiplebrainstemarousal systemsisthecerebralcortexitself.Ithaslongbeen knownthatstimulationofthehigher-orderheteromodalfrontoparietalassociationcortexincreasesarousal (Figure1.11)(Segundoetal.,1955).Conversely,ablationofthesesameregionsofthehigher-orderassociationcortexmarkedlydecreasesarousal(Ropertand Steriade,1981;SteriadeandMcCarley,2010;Watson, etal.,1977),althoughthesubcorticalarousalsystems alsoreceiveinputsfromprimarysensorimotorcortices (Catsman-BerrevoetsandKuypers,1981;Ropertand Steriade,1981;RossiandBrodal,1956).Infurthersupportoftheimportanceofthecerebralcortexinmaintainingconsciousnessitwasrecognizedbyclinicians thatunilateralcorticallesionsusuallydonotmarkedly depresslevelofconsciousness,butbilaterallesionsof theassociationcortexcanproducecoma(Plumand Posner,1972;Posneretal.,2007).Theparietalcortexof thenon-dominant(usuallyright)hemisphereappears toplayaparticularlyimportantroleinarousalwhere largelesions—althoughnotproducingcoma—dooften produceamarkedlydrowsyclinicalstatewithforced eyeclosure.Thus,inadditiontoitsimportantrolein producingthespecificindividualcontentsofconsciousness,thecerebralcortexisalsoamajordriverin regulatingtheoveralllevelofconsciousarousal.
AttentionandConsciousness
Therehasbeenrecentdebateontherelationship betweenattentionandconsciousness.Someviewattentionandconsciousnessasorthogonalfunctionsthat canbefullydissociatedandoperateindependently (KochandTsuchiya,2007).Othersconsiderattention andconsciousnesstobeessentiallyidentical,constitutingdifferentnamesforthesamesetoffunctions (Prinz,2000,2012).Stillotherspositthatattentionis necessaryforbutnotidenticaltoconsciousness becauseadditionalfunctionsareneededforconscious awareness(Dehaeneetal.,2006;KouiderandDehaene, 2007).Theseverydifferentunderstandingsoftherelationshipbetweenattentionandconsciousnessmay ariseatleastinpartfromdifferentmodelsfordefining andunderstandingattention.Therearealargenumber
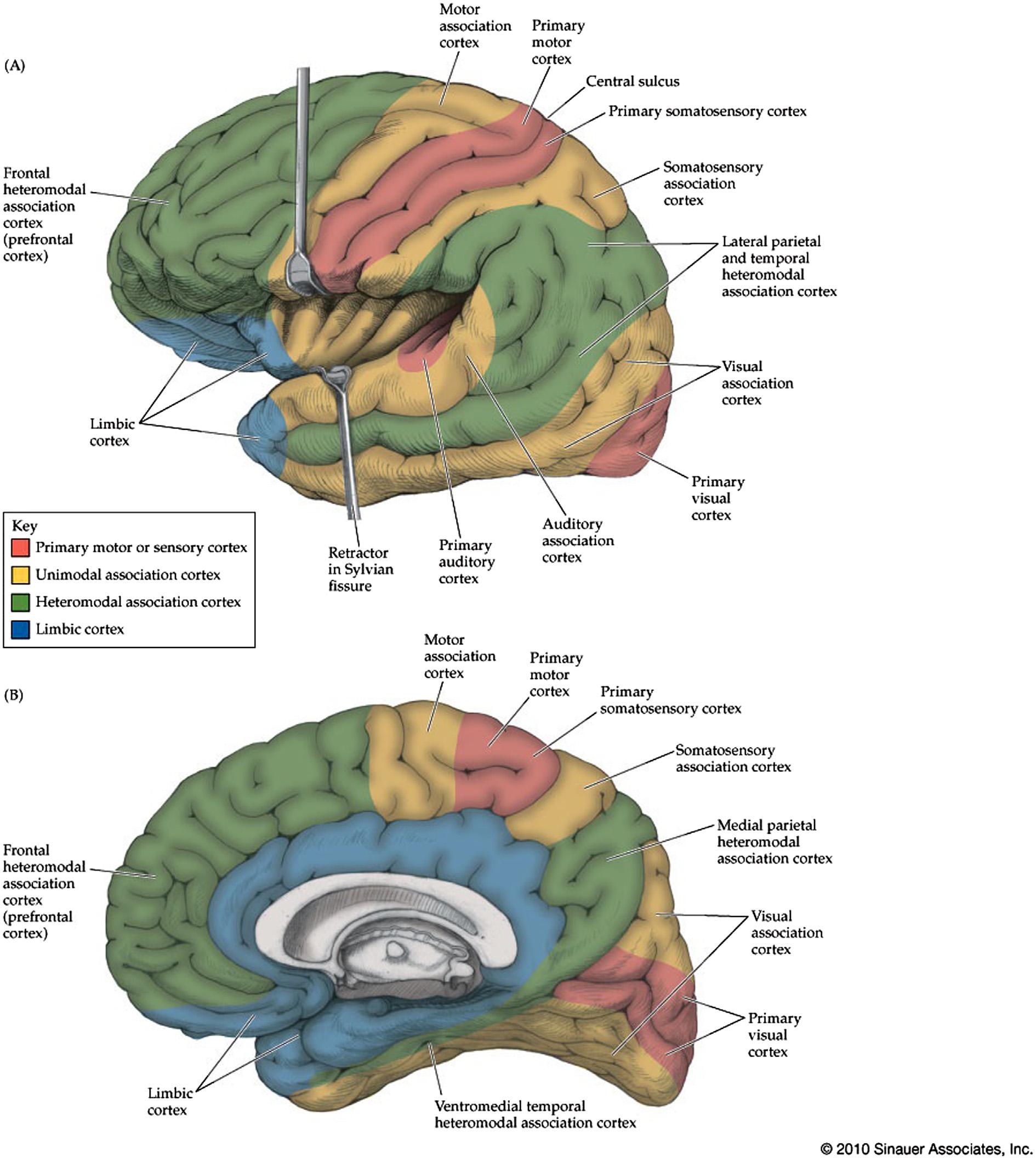
FIGURE1.11 Cerebralcortex.(A)Lateralview.(B)Medialview.Mainprimarycorticalandassociationcorticalareasareindicated. Unimodalassociationcortexismodality-specific,whereashigher-orderheteromodalassociationcortexcombinesinformationacrossmodalities.Regionslabeledhereaslimbiccortexincludeallocorticalregionssuchasthehippocampus(archicortex)andpyriformcortex(paleocortex) aswellastransitionalparalimbicandneocorticalareassuchastheparahippocampalgyrus,cingulategyrus,anteriorinsula,orbitofrontalcortex,andtemporalpole. Source:Reproducedwithpermissionfrom Blumenfeld(2010)
ofmodelsofattentionandacompletediscussionof theanatomyofattentionisfarbeyondthescopeofthis chapter.However,herewewillatleastdiscussafew ofthemajorcurrentformulationsofattentionespeciallyinhowtheircorticalanatomicalunderpinnings relatetounderstandingconsciousness.
HemisphericDominanceofAttention
Evenmorethanthelateralityoflanguage,spatial attentionisstronglylateralizedandthelargemajority ofindividualshavedominantspatialattentioninthe righthemisphere(HeilmanandValenstein,2003; Heilmanetal.,2000;Mesulam,2000).Contralateral
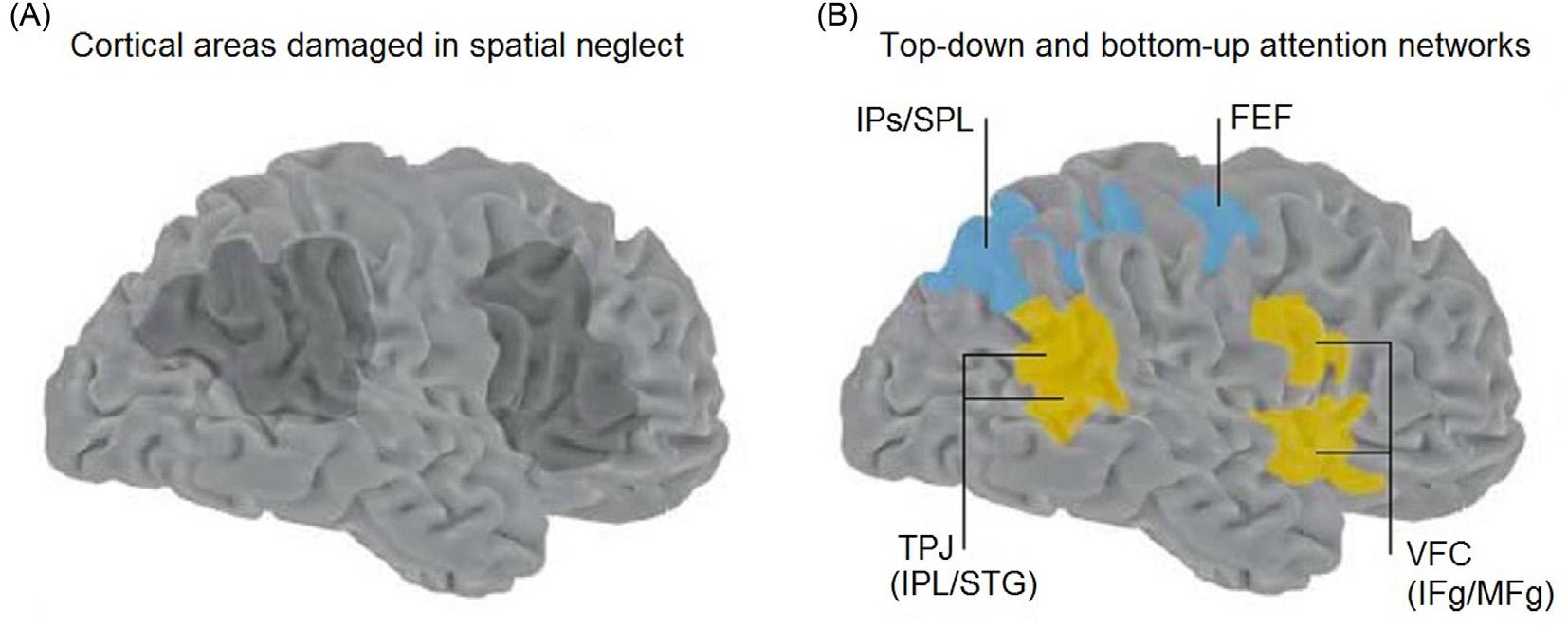
FIGURE1.12 Top-down,bottom-upattentionnetworksandhemineglect.(A)Areascommonlydamagedintherighthemisphere thatcausethecontralateralhemineglectsyndrome.(B)Dorsalandventralfrontoparietalattention.Areasinblueindicatethedorsalfrontoparietal“top-down”attentionnetwork.FEF,frontaleyefield;IPs/SPL,intraparietalsulcus/superiorparietallobule.Areasinorangeindicate thestimulus-drivenventralfrontoparietal“bottom-up”attentionnetwork.TPJ,temporoparietaljunction(IPL/STG,inferiorparietallobule/ superiortemporalgyrus);VFC,ventralfrontalcortex(IFg/MFg,inferiorfrontalgyrus/middlefrontalgyrus).Theareasdamagedinneglect (A)bettermatchtheventralnetwork. Source:Modifiedwithpermissionfrom CorbettaandShulman(2002)
attentionalneglectismorecommonandusuallyfar moreseverewithlesionsoftherighthemisphere, especiallywithdamagetotherightlateralparietal cortex,althoughhemineglectcanalsobeseenwith rightfrontallesions(Figure1.12A).Unilateralneglect profoundlyimpairstheaffectedindividual’sabilityto attendtovisual,auditory,andtactilestimuli,tomotor intentionandindeedeventotheveryexistenceofthe entirecontralateralhalfoftheirbodyandtheexternal worldoppositetothesideoftheirlesion.Althoughthe impairedattentionisusuallymostprofoundcontralateraltorighthemispherelesions,thereisalsosome attentionalimpairmentwhichextendstotheipsilateralsidesothatoverallattentionisglobally decreased(Heilmanetal.,2000).Milderdegreesof unilateralcontralateralneglectareusuallyseenwith leftlateralparietalorleftfrontallesions.Overall,these featuresoftheneglectsyndromesupportamodelin whichattentionfunctionsaredistributedbetweenthe associationcorticesinbothhemispheres,butwitha dominantroleplayedbytherightlateralparietaland frontalassociationcortex.
Some,evenasearlyasDescartes,havearguedthat languageisnecessaryforconsciousthought(Descartes, 1637/1988,1649/1970)andbecausethelefthemisphere isusuallydominantforlanguageitcouldbeviewed asmoreimportantforconsciousnessthantheright. However,clearlynon-verbalthoughtscanstillgenerateconsciousexperiences(Devinsky,2000).Therefore, languagemightbestbeviewedasanimportantcomponentofthecontentofconsciousness(alongwith otherspecificfunctionssuchasvisualperception, mathematicalability,judgmentofdistance,andsoon), ratherthanasanecessaryregulatoroftheoverall
levelofconsciousness.Insomestudiesimpaired consciousnessismorecommonwhenthelefthemisphereisimpairedforexample,duetostroke,orduringselectiveadministrationofbarbiturateanesthetics toonehemisphereatatimeintheangiogramWada test(Albertetal.,1976;Franczeketal.,1997;Schwartz, 1967),orwithfocalseizuresoriginatingintheleft hemisphere(Englotetal.,2010;Gabretal.,1989;Inoue andMihara,1998;Luxetal.,2002).However,itisdifficulttodeterminewhetherabiasoftestingmethods whichareheavilyweightedtowardsverbalratherthan non-verbalquestionsandresponsesmightstrongly influencethesefinding.
Affect,Motivation,andAttention
Oneimportantaspectofcorticalattentionnetworks thatshouldnotbeoverlookedisthemajorroleof affectandmotivationaldrives(DamasioandCarvalho, 2013;HeilmanandValenstein,2003;Heilmanetal., 2000;SatzandHeilman,1983).Subjectswhoare emotionallymotivated,forexample,byseekinga reward,areclearlymoresuccessfulinattentiontasks (HeilmanandValenstein,2003).Theorbitalfrontalcortexandotherlimbiccircuitshavebeenimplicatedand arelikelytobecrucialforthemotivationalaspectsof attention(Faw,2003).
TheBindingProblem
Aclassicalquandaryinunderstandingattention andconsciousnessisthequestionofwhereandhow thediverseaspectsofanyparticularperceptcome togethertoformaunifiedconsciousexperience.How dothelook,smell,andsoundofyourfriendcome togetherinyourbrainas“Adam.”Evenwithina
singlemodality,itischallengingtounderstandhow forexample,thedifferentpartsofavisualsceneare unifiedaswholeimage.Thisquestionhasbeen referredtoasthebindingproblem(Singer,2001). Manypotentialsolutionshavebeenproposedforhow differentcomponentsofsensoryinputarebound togetherintoasinglepercept,includingcoherenthighfrequencyoscillations,lateralconnectionsbetween neuronsinparticularcorticallayers,amongothers (Singer,2001;ZmigrodandHommel,2013).Intermsof corticalanatomysomeinsightsintohowvisualbindingoccursmaybegleanedfromaparticulardisorder referredtoasBalintsyndrome(Barton,2011;Heilman andValenstein,2003;MichelandHenaff,2004).
PatientswithBalintsyndromeexperiencesimultagnosia,meaningthattheyperceiveindividualfragments orcomponentsofavisualsceneoneatatimewithout assemblingthemintoacoherentwhole.Balintsyndromeiscausedbybilaterallesionsatthejuncture betweenthedorsolateralparietalandoccipitalcortices (oftenduetostrokesinthewatershedterritory betweentheposteriorandmiddlecerebralarteries).It hasbeenproposedthatsimultagnosiaoccursbecause theselesionsdisconnectvisualinputfromparietaland otherassociationcorticesneededtostitchtogetherthe individualcomponentsintoaunifiedimage.This suggeststhatonemechanismforattentionalbinding maybetheinteractionofprimarycorticesorunimodal associationcortexwithhigher-order(heteromodal) parietalorotherassociationcortices(Figure1.11).In thiscase,onewouldpredictthatsimultagnosiashould occurnotjustinvisualbutinother(e.g.,auditoryor tactile)modalitieswhenunimodalcorticesaredisconnectedfromthehigher-orderassociationcortex bilaterally,atopicforpotentialadditionalinvestigation infuturework.
Top-DownandBottom-UpAttentionNetworks
Emergingfromotherrecentformulationsof attention,includingthoseproposedbyMesulam, Heilman,andPosner(HeilmanandValenstein,2003; Mesulam,2000;PosnerandDehaene,1994;Posnerand Petersen,1990),themodelof CorbettaandShulman (2002) emphasizestwoseparatebutinteractingsystemsmediatingthetop-downandbottom-upaspects ofattention(Figure1.12B).Inthisscheme,goalorientedselectionofstimuliandresponsesiscontrolledbydorsalregionsofthefrontalandparietal associationcortex,includingthefrontaleyefieldsand intraparietalsulcus.Thistop-downattentionsystemis bilaterallydistributedinbothhemispheres.Incontrast, asecondattentionsystemservesastimulus-driven “circuit-breaking”roleforgrabbingandreorienting attentioninresponsetosalientorchangingsensorystimuli.Thisbottom-upattentionsystemislocalized
moreventrallyinthetemporal-parietaljunctionandthe ventralfrontalcortex(includingthefrontaloperculum), andisstronglylateralizedtotherighthemisphere.One importantareaofongoingresearchishowthetwo systemsinteract,forexample,duringstimulus-driven reorientinginvisualsearchwheresensoryinputsaffect goal-orientedattentionandvice-versa.
Task-PositiveandTask-NegativeNetworks
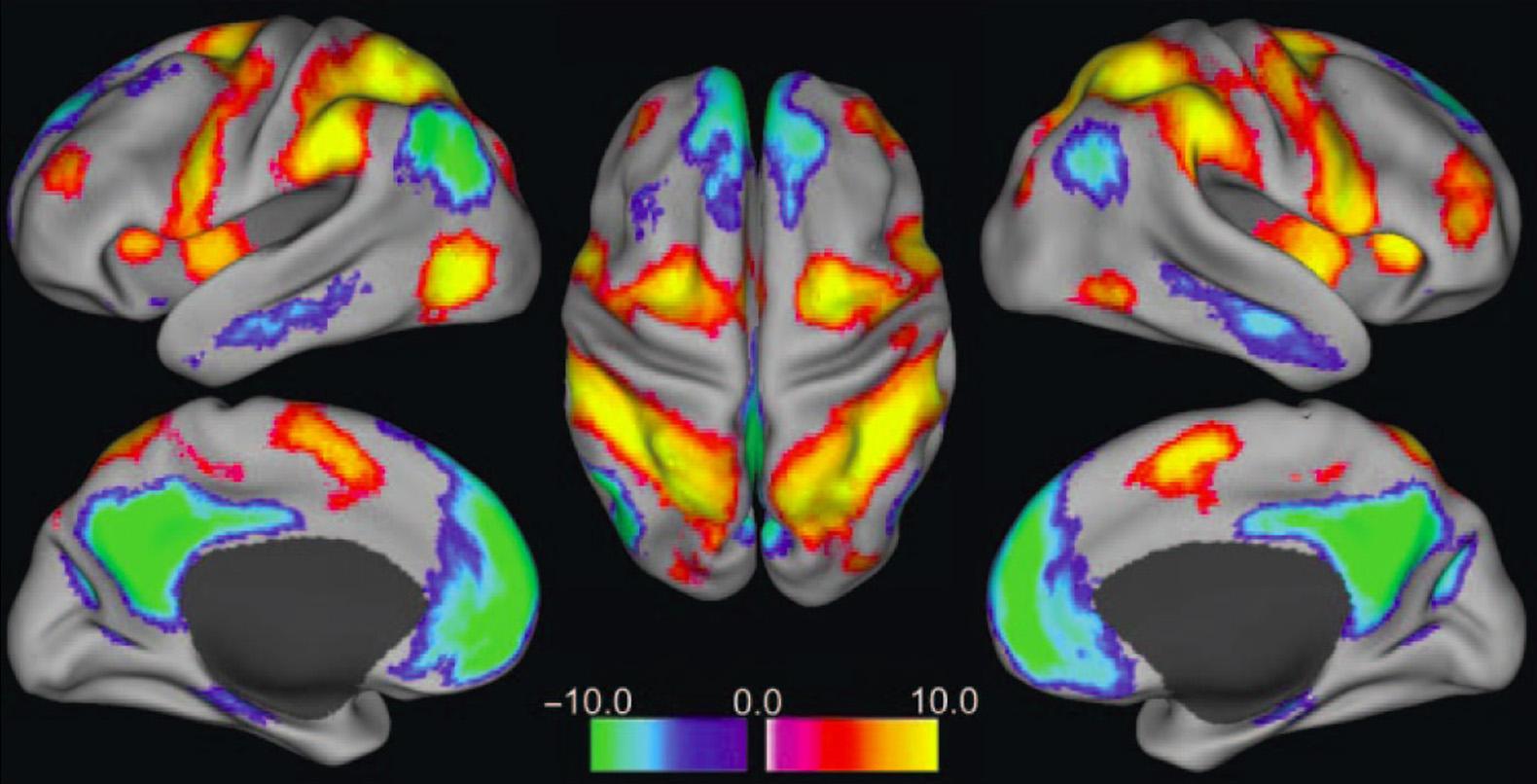
Anotherimportantwayofdescribingcortical attentionnetworkshasarisenfromfunctionalneuroimagingstudiesduringtaskscomparedtorest,aswell astheanalysisofrestingfunctionalconnectivity.Ina seriesofobservationsby Raichleetal.(2001), Shulman etal.(1997) andsubsequentlyconfirmedbyother groupsitwasnotedthat,regardlessofthespecific task,aparticularsetofregionstendstoshowreduced activityduringtaskblockswhenfunctionalneuroimagingdataareanalyzedbyconventionalblockdesignanalysescontrastingtaskversusrest.These brainregionsweredubbedthe“default-modenetwork”(hypothesizedtoberelativelyactiveatrestby default)or“task-negativenetworks.”Ontheother hand,regionsshowingrelativelyincreasedactivity duringtaskblocksshowgreatervariabilitydepending onthespecifictask,butdoshowsomegeneralsimilaritiesbetweenstudies—particularlythoseinvolving attention—andhavebeenreferredtoas“task-positive networks.”Ofinterest,likemanyotherimportant brainnetworks(Biswaletal.,1995,1997),thedefaultmodeandtask-positivenetworkscanbedemonstrated infunctionalneuroimagingstudiesbasedontheirhigh within-networkconnectivityusingrestingdatainthe absenceoftask(Foxetal.,2005;Fransson,2005; Greiciusetal.,2003).Atrest,thedefault-modeand task-positivenetworksdemonstratehighwithinnetworkfunctionalconnectivity(correlationsovertime) andthesetwonetworksareoverallnegativelycorrelatedwitheachother(Figure1.13).Basedonbothtaskrelatedanalyses,andrestingconnectivityanalyses,the default-modenetworkgenerallyincludesthefollowing corticalregionsbilaterally:precuneus/posteriorcingulate,posterior-inferiorparietallobule(angulargyrus), ventral-anteriormedialfrontal,middletemporalgyrus, andmedialtemporalcortex(Figure1.13)(Foxetal., 2005).Thetask-positivenetworksincludethefollowing corticalregionsbilaterally:anteriorinsula/frontaloperculum,supplementarymotor/dorsalmedialfrontal lobe,lateralpremotorcortex(includesfrontaleye fields),anteriormiddlefrontalgyrus,superiorparietal lobule/anteriorinferiorparietallobule,lateralinferior posteriortemporalgyrus(lateralarea37)(Figure1.13).
Becausetheconceptoftwolargeanti-correlated networksmediatingattentionswitchinginthebrain maybeover-simplified,anumberofgroupshave
FIGURE1.13 Task-positiveanddefault-mode(task-negative)networksbyrestingfunctionalconnectivityanalysis.Aconjunctionanalysis wasperformedbyincludingvoxelssignificantlypositivelyornegativelycorrelatedwiththreeseedregionsinthetask-positivenetwork(intraparietalsulcus;frontaleyefields;lateralposterior-inferiortemporalregion)andthreeinthedefault-modenetwork(ventralmedialprefrontal cortex;posteriorcingulate/precuneus;lateralparietalcortex).Warmcolorsshowregionscorrelatedwiththetask-positiveseedsornegatively correlatedwithdefault-modeseeds.Coolcolorsshowregionscorrelatedwiththedefault-modeseedsornegativelycorrelatedwithtaskpositiveseeds. Source:Reproducedwithpermissionfrom Foxetal.(2005)
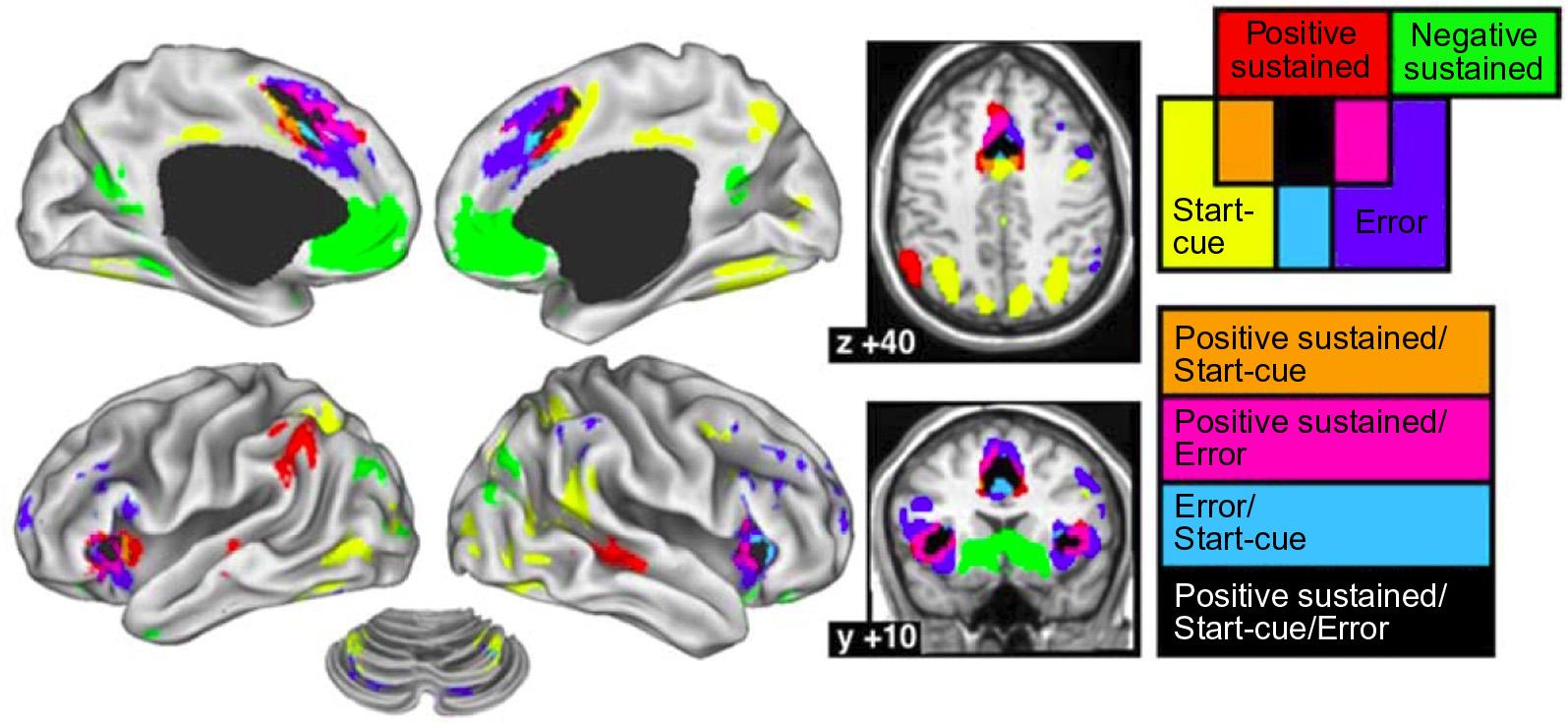
attemptedtofurtherrefinetheseideasandthefield isstillverymuchinevolution.Ithasbeenproposed thatcertaincomponentsofthetask-positivenetwork,referredtoasthefrontalparietalcontrol network,mightplayaroleinswitchingbetween externallydirectedattentionmediatedbythedorsal attentionnetwork(similartotop-downattentionareas ofCorbettaandShulmandiscussedabove)and internallydirectedattentionmediatedbythedefaultmodenetwork(GaoandLin,2012;Vincentetal., 2008).Thereisevidencethatthemorelateral componentsofthetask-positivenetwork,particularly thedorsolateralfrontalandparietalcorticesaremore importantforexternallydirectedattention,whilethe medialcomponentsofthedefault-modenetwork participateininternallydirectedattention(Demertzi etal.,2013;Vanhaudenhuyseetal.,2011).Inanother seriesofstudies, Dosenbachetal.(2006,2007,2008) identifiedcertainbilateralcorecomponentsofthetaskpositivenetwork,namelythedorsalanteriorcingulate/medialsuperiorfrontalcortex(andadjacent supplementarymotorarea),andanteriorinsula/frontal operculumwhichconstituteacommonnetwork involvedin:(i)taskinitiation,(ii)sustainedactivity, and(iii)errordetection(Figure1.14).Incontrast,they foundthatotherareasinthetask-positiveanddefaultmodenetworksareactivatedorinactivatedlessconsistentlyinthesethreedifferentconditions.Againit isunclearifthefunctionalimagingsignalsinanterior insula,whichplayaprominentroleinthisnetwork, couldalsohaveacontributionfromthenearby
claustrum,asubcorticalstructurediscussedabovethat mightparticipateinattentionandconsciousness.In additionalanalysesbasedonrestingfunctionalconnectivityandgraphtheory,thesamegroupagain emphasizedtheimportanceofwhattheyrefertoas thecingulo-opercularnetwork,asdistinctfromthe othertask-positiveanddefault-modenetworknodes (Dosenbachetal.,2007).Anotherveryinteresting approachhasbeenmodel-freeanalysisoffunctional neuroimagingdatabysimplyusingmassiveaveraging oflargedatasetsduringasimpleattentiontask (Gonzalez-Castilloetal.,2012).Thistypeofanalysis revealedthatvirtuallytheentirebrainshowssignals thatvaryovertimeinrelationtothetask,withacombinationoftransientorsustainedincreasesand/or decreasesdependingonthespecificbrainregion.
Insummary,alargenumberofcorticalnetworks havebeenshowntoparticipateindifferentaspectsof attentionandtomodulatetheiractivityinrelationto onsetandendofattentiontasks.Thedetailedroles thatdifferentcomponentsplayinattentionand consciousness,includinganteriorinsula/frontaloperculum,dorsalattention,default-mode,task-positive, bottom-up,top-downandothernetworks,willbe importantsubjectsforfurtherinvestigation.
MemorySystemsandConsciousness
Aswehavediscussed,thelevelofconsciousness dependsonregulationofalertness,attention,and awareness.Alertness(arousal,wakefulness)canbe
FIGURE1.14 Corecomponentsofhumantask-relatednetworks.Conjunctionanalysisofthefixed-effectsmapsforpositivesustained, start-cueanderror-relatedactivity.Thedorsalanteriorcingulate/medialsuperiorfrontalcortex(andadjacentsupplementarymotorarea),and anteriorinsula/frontaloperculumcarriedpositivesustained,start-cueanderror-relatedactivityacrossmostofthetasksincludedintheanalysis(conjunctionregionsshowninblack). Source:Reproducedwithpermissionfrom Dosenbachetal.(2006)
testedbasedonresponsestosimplequestionsor commands,andattentionbasedontasksrequiring selectiveresponses.Awareness,ontheotherhand,is demonstratedwhenasubjectreportsbeingawareofa particularstimulusorevent.Becausethisreport inevitablyoccursaftertheevent,testingofconscious awarenessrequiresmemory.Memorythereforeplaysa crucialroleintheabilitytoreportconsciousexperiences.Infactitmaybeusefultodefineawarenessas theattentiveandotherprocessesnecessaryforevents tobeselected,handedoffandencodedintomemory forsubsequentreport.
Dependingonwhetherreportofconsciousawarenessoccursfollowingashortorlongdelay,different memorysystemsmayparticipate.Immediaterecallor iconicmemorymostlikelyinvolvesactivityinprimaryandunimodalcorticesandwhereeventsare initiallyprocessed(Coltheart,1980;Loftusetal.,1992). Slightlymoreenduringmemorieslastingseveral secondscanenterworkingmemorysystemsinvolving thedorsolateralfrontalandparietalassociationcortices (Funahashietal.,1993;IkkaiandCurtis,2011).Some havedefinedconsciousnessbasedonthetransferof attendedeventsintoworkingmemory(Prinz,2012). Long-termmemorystorageoffactsandautobiographicaleventsrequirestheactionofmedialtemporal andmedialdiencephalicmemorycircuits(Squireand Wixted,2011;Tulving,2002).Episodicordeclarative memory,whichdependsontheseanatomicalsystems, canbeconsideredconsciousmemoryincontrastto unconsciousproceduralmemorythatinvolveslearning skillsorunconsciouspriming.Becausemedialtemporal andmedialdiencephalicmemorysystemsaresospecificallylinkedtotheencodingofconsciousexperiencesfor
subsequentreport,investigatingthe“gateway”leading intothesesystemsmaybeaproductiveapproachfor understandingthemechanismsofconsciousawareness.
VolitionandConsciousFreeWill
Althoughmuchresearchhasbeendoneinthefield ofconsciousperceptionparticularlyemphasizingthe visualsystem,anequallyimportantareaofinvestigationisthemechanismsofconsciousactionincludingthosegoverningplanningandinitiationof voluntarymovement(Roskies,2010).Understanding consciousactionhasimportantimplicationsfordeterminingmoralandlegalresponsibility,aswellasphilosophicalrelevanceincludingthedefinitionoffree will.Consciousactioncanbedividedintoplanning, “premeditation”orinitiationoffutureactivities,and awarenessofongoingorcompleteddeeds.Thelatteris moresimilartoconscioussensoryperception,inthe sensethatitinvolvessubsequentreportofeventsthat havealreadytakenplace.Consciousplanningand initiationofvoluntarymovementaswellasdecisionmakingontheotherhand,likelyinvolveother neuroanatomicalcircuits,aswewillbrieflydiscuss furtherbelow.
Motorplanningdependsonadistributednetwork includingthepremotor,supplementarymotorand otherfrontalcorticalcircuits,interactingwithparietal associationcortexandsubcorticalnetworksespecially inthebasalganglia(Roskies,2010).Decisionmakingis likewiseagrowingfieldinneuroscience,andrecent workhasagainimplicateddistributedfrontoparietal andsubcorticalcircuits(Glimcher,2003;Lauand Glimcher,2008;Levyetal.,2010;PlattandGlimcher,
1999;Roskies,2010).Basedonstimulationmapping, someinterestingdissociationshavebeenfound. Stimulationoftheparietalcortexisaccompaniedbyan awarenessofvoluntarymovementinitiationorurgeto moveevenifnoactualmotiontakesplace(Desmurget etal.,1999,2009;DesmurgetandSirigu,2009).Onthe otherhand,stimulationofthepremotorcortexofthe frontallobecanproduceactualmovementevenwhen thesubjectisunawareofthemovement(Desmurget etal.,2009;Roskies,2010).Finally,stimulationofthesupplementarymotorareaproducesanurgetomovethat mayfeelcompulsoryorinvoluntary(Friedetal.,1991).
Spontaneousvoluntarymovementsarepreceded 1 2searlierbya“readinesspotential”or“Bereitschafts potential”thatcanberecordedfromthescalpnearthe midlineandhassubsequentlybeenlocalizedtothe supplementarymotorareabasedonintracranialmeasurements(Friedetal.,2011).Inathought-provoking study,BenjaminLibetfamouslyexaminedthetimingof thereadinesspotentialandprovidedevidencethatit mayprecedethemomentwhenindividualsbecome consciouslyawareoftheirdecisionorwilltomoveby severalhundredmilliseconds(Libetetal.,1982,1983a,b). Theinterpretationofthisstudyanditsrelationtofree willhasbeenhotlydebatedinlaterwork(Herrmann etal.,2008;Joetal.,2013;Schurgeretal.,2012).
Self-AwarenessandEmbodiment
Althoughsomeconsiderself-awarenesstobethe defining sinequanon ofconsciousness,othersview awarenessofselftobejustoneexampleofthemany thingsthatanindividualcanbeawareof,andthereforenotnecessaryorsufficientforconsciousness (Zeman,2005).Acloselyrelatedtopicistheabilityto haveafirst-personperspectiveorsenseofembodiment arisingfromone’sownindividualpointofview, whichclearlymakesanimportantcontributiontothe subjectivefeelingofawareness.
Awarenessofselfcanbedrasticallyandselectively impairedincertainneurologicaldisorders,whichmay providesomeinsightintotheneuroanatomicalbasisof self-awareness.Forexample,patientswithlesionsinthe non-dominant(usuallyright)hemisphereparticularlyin thelateralparietalcortexareoftenagnostictothevery existenceoftheentireleftsideoftheirownbodies,and whenspecificallyaskedsometimesconsidertheirleft limbstobelongtosomeoneelse(Heilmanetal.,2000). Frontallobedysfunctioncommonlyleadstoimpaired self-monitoringorself-awareness,andisthoughtto underliepatient’sinabilitytorecognizetheirowndeficits,forexamplewhensomepatientsconfabulateinstead ofadmittingtoamnesia(Kopelman,2014).Onespecializedformofself-awarenessisawarenessofone’sown consciousawareness.Thisformofmeta-cognitionisalso
animportanttopicofinvestigationthatmayshed insightsintowhatmanyconsiderthehighestformsof consciousness(ZemanandCoebergh,2013).
Importantadvanceshaverecentlybeenmadein understandingthemechanismsthatprovideafirstpersonperspectivebytheinnovativestudiesofOlaf Blankeandcolleagues(Blanke,2004;Blankeetal.,2002; Eastonetal.,2009;Heydrichetal.,2010;Lenggenhager etal.,2009;Lopezetal.,2010).Aseriesofinvestigations usingpatientswithout-of-bodyexperience,orfunctional neuroimagingandbehavioralinterventionstocreatean out-of-bodyexperienceinnormalsubjects,haverevealed thattherighttemporal-parietaljunctionplaysacrucial roleinthisunusualcondition.Additionalinsightsmay comefromongoinginvestigationofothersituations wherethefirst-personperspectiveisaltered,forexample,duringearlydevelopmentorinpsychiatricdisorderswheredepersonalizationcanoccur(Bunningand Blanke,2005;Lewis,2011;SierraandDavid,2011).
Awareness:ConsciousReportand ContrastiveAnalysis
Philosophersandscientistshavelongenjoyeda debateaboutwhetherornotconsciousnesscanbe understoodthroughscientificinvestigation.Thecruxof theargumentrestsonthedefinitionsof“consciousness” and“understanding.”Somedefinethequalitativephenomenalaspectsofconsciousnessorqualia,asinternal subjectivefeelingsofawarenessaccessibleonlythrough first-personexperience,anddefineunderstanding consciousnessashavingthatfirst-personexperience (Chalmers,1996).Thesedefinitionsautomatically excludethepossibilityofunderstandingconsciousness throughscientificmeansbecausesciencerequiresexternalsecond-personobservations.Ontheotherhand,if onedefinesconsciousawarenessasanexperiencethat canbedescribedorreportedtoothers,andifone definesunderstandingconsciousnessasidentifyingthe necessaryandsufficientphysiologicalmechanismsfor suchanexperience,thenconsciousnessfallsclearlyin therealmofscientificinvestigation.Thekeytothis secondapproachisthatexternalreportofconsciousnessisallowedasamethodforinvestigatingconsciousness.Reportofconsciousnesshasitslimitationsof course,butsodoallscientificmethodsandaslongas thelimitationsarerecognizedandresultsinterpreted cautiously,muchprogresscanbemade.
Takingthisapproach,alargenumberofstudieshave investigatedthecontrastbetweenbrainactivitywhena consciouseventisreportedorisnotreportedunder verysimilarcircumstances.Thiscontrastiveanalysis hasbeenperformedinavarietyofsituationswhere eventsthatareorarenotreportedcanbeobtained (DehaeneandChangeux,2011;Lamme,2006;Tononi