Instant digital products (PDF, ePub, MOBI) ready for you
Download now and discover formats that fit your needs...
The Diversity of Fishes: Biology, Evolution and Ecology, 3rd Edition Douglas E. Facey
https://ebookmass.com/product/the-diversity-of-fishes-biologyevolution-and-ecology-3rd-edition-douglas-e-facey/
ebookmass.com
Myxomycetes: biology, systematics, biogeography, and ecology Elsevier.
https://ebookmass.com/product/myxomycetes-biology-systematicsbiogeography-and-ecology-elsevier/
ebookmass.com
Neuropsychology: Science and Practice 3rd Edition Sandra Koffler (Editor)
https://ebookmass.com/product/neuropsychology-science-andpractice-3rd-edition-sandra-koffler-editor/
ebookmass.com
Binding 13 Chloe Walsh
https://ebookmass.com/product/binding-13-chloe-walsh/
ebookmass.com
Philemon Iko-Ojo Omede
https://ebookmass.com/product/nigerian-consumer-credit-law-regulationand-market-insights-philemon-iko-ojo-omede/
ebookmass.com
SCALLOPS
Biology,Ecology,Aquaculture, andFisheries
THIRDEDITION
Editedby SANDRA E.SHUMWAY
DepartmentofMarineSciences,UniversityofConnecticut, Groton,Connecticut,USA
G.JAY PARSONS
FisheriesandOceansCanada,Aquaculture,Biotechnology andAquaticAnimalHealthScienceBranch,Ottawa,Ontario,Canada
ForAndyBrandandNeilBourne,scallopafficionados,mentors,andfriends.
EffectsonMarineEcosystems589 EffectsonDifferentSeabedTypes594 Management599 Conclusion603 Acknowledgements604 References604
15.Dynamics,Assessment,andManagement ofExploitedNaturalScallopPopulations
J.M.(LOBO)ORENSANZ,ANAM.PARMA,ANDSTEPHENJ.SMITH
Introduction611 PopulationStructureandDynamics613 AssessmentofAbundanceandItsSpatialDistribution632 TheFishingandDepletionProcesses640 OverfishingandEcologicalEffectsofFishing654 Management661 Acknowledgements676 Acronymsusedinthetext676 References677
16.ScallopsoftheWestCoastofNorthAmerica
G.JAYPARSONS,RAYMONDB.LAUZIER,ANDNEILF.BOURNE Foreword697 Introduction697 Fisheries698 Aquaculture709 Future713 Acknowledgements714 References714
17.FisheriesSeaScallop, Placopectenmagellanicus
KEVIND.E.STOKESBURY,CATHERINEE.O’KEEFE,AND BRADLEYP.HARRIS
Foreword719 Introduction719 LifeHistory721 Ecosystem723
TheFishingFleetsofCanadaandtheUnitedStates724 StockAssessments726 FisheriesManagement729 Summary731 Acknowledgements732 References732
18.ScallopAquacultureandFisheriesinEastern NorthAmerica
SHAWNM.C.ROBINSON,G.JAYPARSONS,LESLIE-ANNEDAVIDSON, SANDRAE.SHUMWAY,ANDNORMANJ.BLAKE
Introduction737
SeaScallop(Placopectenmagellanicus)737
BayScallop(Argopectenirradians)758 CalicoScallop, Argopectengibbus 761 AquacultureandEnhancement765 Future767 Acknowledgements767 References767
19.TheEuropeanScallopFisheriesfor Pecten maximus, Aequipectenopercularis,Chlamysislandica, and Mimachlamysvaria
PETERF.DUNCAN,ANDREWR.BRAND,ØIVINDSTRAND,AND ERICFOUCHER
Introduction781 BiologyandEcology782 Fisheries801 TheFuture838 Acknowledgements841 References841
20.EuropeanAquaculture ØIVINDSTRAND,ANGELESLOURO,ANDPETERF.DUNCAN
Introduction859 HistoricalOverviewofEuropeanScallopAquaculture859 SpatProduction862 Grow-OutCulture867 SeabedRanchingandStockEnhancement875 BiologicalConstraintstoScallopCulture879 MarketandEconomics881 StockProtectionSecurityandLegalIssues882 FutureProspects883 Acknowledgements884 References884
21.ScallopFisheriesandAquaculture inJapan
YOSHINOBUKOSAKA
Introduction891 Patinopecten(Mizuhopecten)yessoensis 891 Future926 Pectenalbicans 931 Chlamys(Mimachlamys)nobilis 932 Acknowledgements932 References933
22.ScallopsandScallopAquacultureinChina XIMINGGUOANDYOUSHENGLUO
Introduction937 TheChineseScallop938 OtherScallopSpecies943
Fishery945 Aquaculture945
Harvest,Processing,andMarketing951
Acknowledgements951 References951
23.ScallopsofNorthwesternPacificRussian Federation
VICTORV.IVIN,OLGAG.SHEVCHENKO,AND TATIANAYU.ORLOVA
Introduction953 BiologyandEcology953 FishingandAquaculture975
Acknowledgements991 References991
24.AquacultureoftheScallop Nodipectennodosus inBrazil
GUILHERMES.RUPPANDG.JAYPARSONS
Introduction999
Aquacultureof Nodipectennodosus inBrazil1001
Acknowledgements1015 References1015
25.ScallopsBiology,Fisheries,andManagement inArgentina
GASPARSORIA,J.M.(LOBO)ORENSANZ,ENRIQUEM.MORSAN, ANAM.PARMA,ANDRICARDOO.AMOROSO
Introduction1019
TheTehuelcheScallop, Aequipectentehuelchus
1019
ThePatagonianScallop, Zygochlamyspatagonica 1032 Acknowledgements1041 References1041
26.ScallopFisheryandAquacultureinChile: AHistoryofDevelopmentsandDeclines
ELISABETHVONBRAND,ALEJANDROABARCA, GERMANE.MERINO,ANDWOLFGANGSTOTZ
Introduction1047
SpeciesDescription1047 Fisheries1050
HowScallopAquacultureStartedinChile1053
AquacultureProduction1055
Acknowledgements1068 References1068
27.ScallopAquacultureandFisheriesinVenezuela CE ´ SARJ.LODEIROS,LUISFREITES,JOSE ´ J.ALIO ´ ,MAXIMIANONU ´ NEZ, ANDJOHNH.HIMMELMAN
Introduction1073
Distribution,Habitat,andReproduction1073 Fisheries1077 Aquaculture1078 PerspectivesforCulture1084 Acknowledgements1085 References1085
28.ScallopFisheryandCultureinPeru
JAIMEMENDO,MATTHIASWOLFF,TANIAMENDO,ANDLUISYSLA
Introduction1089
SpeciesDescription1089 ProductionChainandMarketing1094 FisheriesandAquacultureProduction1095 ChallengesandProjectionsofPeruvianScallopProduction1104 Acknowledgements1105 References1105
29.ScallopFisheriesandAquacultureinMexico
CESARA.RUIZ-VERDUGO,VOLKERKOCH,ESTEBANFELIX-PICO, ANAISABELBELTRAN-LUGO,CARLOSCA ´ CERES-MARTI ´ NEZ, JOSE MANUELMAZON-SUASTEGUI,MIGUELROBLES-MUNGARAY, ANDJORGECACERES-MARTI ´ NEZ
Introduction1111 DescriptionofMexicanScallops1112 Fisheries1114 Aquaculture1119 References1123
30.ScallopFisheries,Mariculture,andEnhancement inAustralasia
MICHAELC.L.DREDGE,ISLAYD.MARSDEN,ANDJAMESR.WILLIAMS
Introduction1127 Biology1129 FisheriesandFisheryManagement1140 Culture1161
FutureDirectionsandConclusions1164 Acknowledgements1166 References1166
GeneralIndex1171 SpeciesIndex1189
ListofContributors
AlejandroAbarca DepartamentodeAcuicultura, UniversidadCato ´ licadelNorte,Coquimbo,Chile
Jose ´ J.Alio ´ InstitutoNacionaldeInvestigacionesAgrı´colas, Cumana ´ ,EstadoSucre,Venezuela
RicardoO.Amoroso SchoolofAquaticandFishery Sciences,UniversityofWashington,Seattle,WA,USA
BruceJ.Barber EckerdCollege,St.Petersburg,Florida, USA
AnaIsabelBeltran-Lugo DepartamentodeIngenierı ´ aen Pesquerı´as,UniversidadAuto ´ nomadeBajaCaliforniaSur Ap.,LaPaz,BajaCaliforniaSur,Me ´ xico
PeterG.Beninger LaboratoiredeBiologieMarine,Faculte ´ desSciences,Universite ´ deNantes,Nantes,France
NormanJ.Blake CollegeofMarineScience,Universityof SouthFlorida,St.Petersburg,Florida,USA
NeilF.Bourne FisheriesandOceansCanada,Pacific BiologicalStation,Nanaimo,BritishColumbia,Canada
SusanM.Bower FisheriesandOceansCanada,Pacific BiologicalStation,Nanaimo,BritishColumbia, Canada
AndrewR.Brand UniversityofLiverpool,PortErin,Isleof Man,UnitedKingdom
V.MonicaBricelj DepartmentofMarineandCoastal SciencesandHaskinShellfishResearchLaboratory,School ofEnvironmentalandBiologicalSciences,Rutgers University,PortNorris,NJ,USA
JorgeCaceres-Martı ´ nez CentrodeInvestigacio ´ nCientı´ficay Educacio ´ nSuperiordeEnsenada(CICESE),Ensenada, BajaCalifornia,Me ´ xico;InstitutodeSanidadAcuı´cola, Ensenada,BajaCalifornia,Me ´ xicoInstitutodeSanidad Acuı´cola,Ensenada,BajaCalifornia,Me ´ xico
CarlosCa ´ ceres-Martı ´ nez DepartamentodeIngenierı ´ aen Pesquerı´as,UniversidadAuto ´ nomadeBajaCaliforniaSur Ap.,LaPaz,BajaCaliforniaSur,Me ´ xico
PeterD.Chantler UnitofVeterinaryMolecularandCellular Biology,ComparativeBiomedicalSciences,RoyalVeterinary College,UniversityofLondon,London,UnitedKingdom
SimonM.Cragg InstituteofMarineSciences,Universityof Portsmouth,Portsmouth,UnitedKingdom
PeterJ.Cranford FisheriesandOceansCanada,Coastal EcosystemSciencesDivision,BedfordInstituteof Oceanography,Dartmouth,NovaScotia,Canada
Leslie-AnneDavidson FisheriesandOceansCanada, Moncton,NewBrunswick,Canada
MichaelC.L.Dredge Tasmania,Australia
PeterF.Duncan FacultyofScience,Health,Educationand Engineering,UniversityoftheSunshineCoast, Queensland,Australia
EstebanFe ´ lix-Pico CentroInterdisciplinariodeCiencias Marinas,LaPaz,BajaCaliforniaSur,Mexico
EricFoucher IFREMER LaboratoireHalieutiquedePort enBessin,PortenBessin,France
LuisFreites InstitutoOceanogra ´ ficodeVenezuela, UniversidaddeOriente,Cumana ´ ,EstadoSucre, Venezuela
RodmanG.Getchell DepartmentofMicrobiologyand Immunology,CollegeofVeterinaryMedicine,Cornell University,Ithaca,NY,USA
HelgaE.Guderley De ´ partementdeBiologie,Universite ´ Laval,Que ´ becCity,Que ´ bec,Canada
XimingGuo HaskinShellfishResearchLaboratory, DepartmentofMarineandCoastalSciences,Rutgers University,PortNorris,NJ,USA
BradleyP.Harris Fisheries,AquaticScienceand TechnologyLaboratory,AlaskaPacificUniversity, Anchorage,AK,USA
JohnH.Himmelman De ´ partementdeBiologie,Universite ´ Laval,Que ´ becCity,Que ´ bec,Canada
LeighM.Howarth SchoolofOceanSciences,Bangor University,MenaiBridge,Anglesey,UnitedKingdom
VictorV.Ivin InstituteofMarineBiology,FarEastern BranchoftheRussianAcademyofSciences,Vladivostok, Russia
VolkerKoch DeutscheGesellschaftfu ¨ rInternationale Zusammenarbeit(GIZ),LaPaz,BajaCaliforniaSur,Me ´ xico YoshinobuKosaka AomoriPrefecturalIndustrial TechnologyResearchCenter,FoodResearchInstitute, Aomori,Japan
MaureenK.Krause DepartmentofBiology,Hofstra University,Hempstead,NY,USA
FranciscoJ.Lagreze-Squella FederalUniversityofParana ´ , PontaldoParana ´ ,Parana ´ ,Brasil
RaymondB.Lauzier FisheriesandOceansCanada,Pacific BiologicalStation,Nanaimo,BritishColumbia,Canada
thescallopaquacultureindustryinBritishColumbia.Neil,thankyouforyourfriendship,insightsand contributions.
ThecurrenteditionhasbenefittedgreatlyfromtheeffortsofNoreenBlaschikwhohastirelesslyhelpedwith proofreading,referencecross-checkingandotherthanklesstasks.ThetalentsofEricHeupelwerekeyinredraftingmanyofthefiguresandaregratefullyacknowledged.Gettingauthorstoagreetoparticipateinventuressuch asthisismucheasierthanextractingthefinaltextsandweextendourdeepestgratitudetotheauthorsfortheir skill,patienceandperseverance.ThankstoSharon,MichaelandChristopherfortheirsupport,cajolingand patience.GusandZeusspentmanyeveningspawingovertexts,andtheirassistancemadethetasksathand moretolerable.
AspecialthankstoPatriciaOsborne,ElsevierSciencePublisherswhoinitiatedandencouragedtheeffortfora ThirdEditionandtoJaclynTruesdell,DebbieClarkandKarenMillerfortheirpatienceandguidanceduringthe production.
Forsuccessfulcultureandmanagement,agoodknowledgeofthebiology,ecology,physiologyandfisheriesof thespeciesisnecessaryandwehopethatthisThirdEditionofScallops:Biology,Ecology,Aquaculture,and Fisherieswillserveasasolidreferencebaseforyearstocome.
SandraE.Shumway1 andG.JayParsons2
1DepartmentofMarineSciences,UniversityofConnecticut,Groton,CT,USA 2FisheriesandOceansCanada,Aquaculture,BiotechnologyandAquaticAnimalHealthScienceBranch, Ottawa,ON,Canada
theimportanceofgeneticstofisheriesandaquacultureofscallopsinChapter12.InChapter13,Orensanz,Parma andIribanediscusspopulationdynamicsandthemanagementofnaturalstocks.ThecommercialspeciesofscallopsarecoveredinChapter14accordingtogeographiclocation.MuchofthedatapresentedinChapter14is beingreportedforthefirsttime,includingsuchelementarymeasuresasgrowth,meatweightandlandings. ThisworkwouldnothavebeenpossiblewithouttheeffortsofDr.ScottSiddall.Hisexpertiseinthefieldof desktoppublishingcoupledwithhisknowledgeofshellfishbiologymademytaskpossible.Hiscontinuedinterest,supportandinterjectionsofhumourwhenmostneededmadethetaskenjoyable.JanBarter,Martha Hernandez-Davis,JohnHedleyandJimRollinshelpedinvariouswaysandtheirwillingnesstoundertakesuch arduoustasksasproofreading,crosscheckingreferences,retypingofmanuscriptsandredrawingandrepairof submitteddiagramsaregratefullyacknowledged.AspecialthanksmustgotoBobGoodman,ElsevierScience PublishingCompany,forhisconsiderablepatiencein‘meeting’deadlinesandhisinsightandsupport.Finally, Iwouldliketothanktheauthorsfortheirskill,patience,andperseverance.
1
ReconcilingMorphologicalandMolecular ApproachesinDevelopingaPhylogenyforthe Pectinidae(Mollusca:Bivalvia)
JeanneM.Serb
INTRODUCTION
Scallops(PectinidaeRafinesque,1815)areoneofthemostmorphologically,behaviourally,andbiologically diversefamilyofbivalves.AsamajormolluscancladeofMesozoicandCenozoiceras,thePectinidaehashada tumultuous245-million-yearhistory,survivingtwomassextinctioneventsattheTriassic-Jurassicand Cretaceous-Paleogeneboundaries(Hautmann,2010;NewellandBoyd,1995;Waller,2006a).Overtheintervening quarterofabillionyears,itislikelythatscallopsunderwentseveraladaptiveradiationsthatplayedamajorrole ingeneratingthebiologicaldiversityseentoday.Currently,about270extantspeciesofscallopsarerecognised (Dijkstra,2014)andaredistributedacrosspolar,temperate,andtropicalseas,withthegreatestbiologicaldiversityinthetropicalCaribbeanandIndo-Pacificoceans(RainesandPoppe,2006).Mostscallopspeciesarefound intheshallowsofsublittoralreefs,sandybays,andseagrassbedsofthecontinentalshelves,withasmallernumberofspeciesrestrictedtodeeperwaterincludingtheabyssaldepthsat7000m.Whilethetypicalscallopshell shapeisthefamiliarroundeddiskandstraighthingelineformedbytwoear-likeauricles,scallopsexhibitawide rangeofmodificationsonthisgeneralisedphenotype,withstrikingshape,texture,andcolourvariantsofthe shell.Thisvarietyhasmadescallopsafavouriteofamateurshellcollectorsandscientistsalikeand,asaresult, theirshellsarewellrepresentedinthecollectionsofnaturalhistorymuseums(e.g. Dijkstra,1999; Dijkstraand Koehler,2008).Theon-goingchallengeforscallopbiologistsistoorganisetheimpressivebiologicaldiversityof thePectinidaeinaphylogeneticcontext.
Underaphylogeneticclassificationsystem,taxaareorganisedhierarchicallyintogroupsthatreflectahypothesisofevolutionaryrelationshipderivedfromaphylogenetictree.Aphylogenetictree(alsocalleda‘phylogeny’ or‘topology’)isabifurcatingdiagramthatdepictstherelationshipsbetweentaxabasedonsharedhistory throughcommondescent(Gregory,2008).Notonlycanthephylogenyhelpidentifygroupsoftaxa(clades),but phylogenetictreesindicaterelationshipsbetweenthesegroups,forexample,howtribesarerelatedtooneanother inasubfamily.Phylogenetictreesalsoprovideahistoricalcontextandarelativeorderforwhenlineagesarose anddiversified.Moreover,thephylogenycanbeusedtostudytheevolutionoftraits,suchaswhetheracharacterstateisancestral(plesiomorphic)orderived(i.e.traitpolarity),ortotesthypothesesregardingtherateor modeoftraitevolution.
Basingaclassificationsystemonphylogenyincorporatestheevolutionaryhistoryofthosetaxaandallthe associatedinformationintotheclassification.Speciesorlineagesthataredescendantsofasingleancestorrepresent‘naturalgroups’,andtherecognitionofnaturalgroupsgivespredictivepowertoaclassificationsystem becausethesharedcharactersmayalsoindicatesimilaritiesinlifehistorytraits,habitatrequirements,orecologicalfunctionduetosharedhistoryoftheorganisms.Thus,modernclassificationsystemsareinformationretrieval resources,whereanindividualspeciescanbeusedtorepresentabroadersetofspeciesinsomecontexts,suchas
possessthealivincular-alateligament.Incontrast,thectenoliumisuniquetothePectinidaeandispresentatleastin theearlypost-larval(dissoconch)growthstages.Thectenoliumisarowofhook-shapeddenticlesalongtheventral edgeofbyssalnotch(Waller,1978,1984)(Figure1.1).Asaproteinbyssusissecretedbythebyssalglandwithinthe muscularfoot,itpassesoverthedenticlesofthectenoliumwhichkeepsthearrayofthreadsflatalongtheedgeof theshell(Waller,1984:Figure4b,c).Thecombinationofabyssalthreadsheetandthetwo-pointcontactofshelltoa surface(see Stanley,1972)stabilisestheanimal’sconnectiontothesubstrateandgreatlyreduceshowmuchtheshell canrotatearoundthebyssus.Asaresult,muchmoretorquemustbeappliedtobreakthebyssalattachment(Waller, 1984)andthismayprovideanadditionallevelofprotectionfrompredatorsthatmustdislodgeandmanipulateprey itemsbeforeconsumption(Caroetal.,2008).
TheRelationshipBetweenLifeHabitandShellShape
Scallopsmetamorphosefromaswimmingpediveligertoacrawling,benthicpost-larva.Atthistime,themantlebeginssecretionofthedissoconchshell(HodgsonandBurke,1988),whosemicrosculpturepatternshavebeen usedascharactersinsomeclassificationsystems(Waller,1991).Allnewlymetamorphosedscallopspossessactenoliumandcanproduceabyssustoattachtoasubstrate.Thisattachmentistemporaryastheanimalcanrelease thebyssusandthensecreteanewbyssustoattachtoanothersubstrate.Astheanimalgrowsandmatures,the ctenoliummaybeovergrownbytheadvancingfrontoftheanteriorauricle(Waller,1984:Figures2a c,3b).This resultsintheprogressivelossoftheabilitytoattachusingabyssusasbodysizeincreases(Caddy,1972).Thus, somescallopspeciesonlyemployabyssusasjuvenilesandlosetheabilitytoproduceabyssusasadults,becoming‘free-living’.
Basedontheabilitytoattachandmodeoflocomotion,adultscallopscanbeplacedintosixfunctionalgroups orlifehabits(sensu Stanley,1970).Theselifehabitsareassociatedwithspecificshellformsofscallops(Stanley, 1970).Themajorityofscallopspecieshavethebyssalattachinglifehabit,whereabilitytoattachtemporarilytoa substrateviabyssusismaintainedaftermaturity.Byssalattachershaveachlamydoidform,wheretheshellhasa strongasymmetryalongtheanterior posterioraxissuchthattheauriclesareofdifferentsizesandtheshell outlineisoblique(Figure1.2A).Additionally,therightvalvehasadeepbyssalnotchwithawell-developed ctenolium(Figure1.1).Ithasbeenhypothesisedthatthechlamydoidformistheplesiomorphicconditionforthe Pectinidae(Waller,1993).Afewbyssalattachingspecies,like Pedumspondyloidium (Gmelin,1791),exhibitthe nestlinglifehabit.Thesespeciesareobligateassociateswithlivingscleractiniancorals,wherethepost-larvapreferentiallysettleson,andbyssallyattachesto,coralpolypswhichgrowaroundthescallopencasingit(Yonge, 1967;Waller,1972a).
Otherspecies,suchas Crassadomagigantea (Gray,1825),permanentlyattachtohard,oftenabiotic,substrates bysecretingnewshellmaterial.Thiscementinglifehabitisassociatedwithashellthattransitionsfromthe
FIGURE1.1 Somemorphologicalfeaturesofthepectinidshell.Rightvalveof Volachlamystranquebaria orientedwithanterior(ant.)towards rightandposterior(post.)towardsleft.
Ctenolium
Ant. auricle Post. auricle Byssal notch Umbo
Right valve
4 1.RECONCILINGMORPHOLOGICALANDMOLECULARAPPROACHESINDEVELOPINGAPHYLOGENYFORTHEPECTINIDAE(MOLLUSCA:BIVALVIA)
FIGURE1.2 Fourcommonformsofscallopshells.Three-dimensionalsurfacescansoftheleftvalveofrepresentativescallopspeciesfor thechlamydoid(A);aequipectinoid(B);pectinoid(C),andamusioid(D)forms.Thecementingformisnotshown.Anterior(ant.)towards rightandposterior(post)towardsleft.
chlamydoidformtohighlyvariableshellshapeasgrowthoftherightvalveconformstothesubstrate. Morphologicalchangesincludeventralmigrationoftheligamentsystemandanincreaseddistancebetweenthe palliallineandshellmargin(Yonge,1951;Waller,1972a;Harper,1991),creatingathickenedanddistortedshell thatsuperficiallyresemblestheshellofoyster.Whilenotaspecificshellshape,thisshellformisoftenassociated withthename Hinnites (asin Hinnitespusio (Linnaeus,1758)).
Somepectinidspeciesdonotattachtoasubstrateofanykindasadults.These‘free-living’specieseitherpassivelyoccupyapositionon,orarepartiallycoveredin,asoftsubstrate(‘recliners’ sensu Stanley,1970).Shellsthat reflectthereclininglifehabitarelike Aequipectenopercularis (Linnaeus,1758)(aequipectinoidform),wherethe valvesaremoreequilateralandflaringthanthechlamydoidformandtherightvalvehasashallowbyssalnotch thatpersistslateinontogeny(Figure1.2B).Likeaequipectinoidform,eachvalveinthepectinoidform(asin Pecten Mu ¨ ller,1776, sensustricto)isstronglysymmetricalongtheanterior posterioraxisandishighlyflared alongthedorsal ventralaxis(Figure1.2C).Incontrast,thereisadramaticshapeasymmetrybetweentheleft andrightvalvesnotpresentintheaequipectinoidform,wheretheleftvalveisflatorslightlyconcaveandthe rightvalveisstronglyconvex.Thepectinoidformisassociatedwiththerecessinglifehabit,wherescallopsconstructasaucer-shapeddepressioninsoftsubstratesbydirectingwaterjetsfromitsmantlecavity(Baird,1958; SakuraiandSeto,2000).Speciesofthisformoftenlackabyssalnotchasadults.
Perhapsthemostintriguingscalloplifehabitisgliding(‘swimming’ sensu Stanley,1970).Whileallnon-permanentlyattachedscallopspecieshavetheabilitytoswimtoescapepredators(Himmelmanetal.,2009)orseek favourablehabitat(BuddenbrockandMoller-Racke,1953;HamiltonandKoch,1996),theseactivitiesarenot stronglydependentonshellshape.Glidingisatypeofswimmingbehaviourthatincludes:(i)agreatdistance travelledperswimmingeffort(5 30mpereffort; Brand,2006);(ii)themaintenanceofanear-horizontaltrajectoryabovethesubstrate(Morton,1980;Joll,1989;Anselletal.,1998);and(iii)aglidingcomponent,wherethe animalcontinuestomoveforwardwhilethevalvesareheldclosed(ManuelandDadswell,1993;Chengetal., 1996;Anselletal.,1998).Neitherhorizontaltrajectorynoraglidingcomponentisobservedinscallopsthatswim forshortdistances(Marshetal.,1992;Anselletal.,1998;Donovanetal.,2002).Theglidinglifehabitisassociated withtheamusioidform(asin Amusium Roding,1798, sensustricto).Animalswiththisformhavethin,lightweightshellswithasmoothexteriorandcircularoutlineconducivetoreducingdragandincreasinglift (Figure1.2D).Fromtheside,theshellsareslightlyconvexandcreateagapalongthemarginsventraltotheauriclessothatwatercanbeexpelledduringtheglidingphase(Hayami,1991:Figure1).Bothauriclesaresymmetricalandaregreatlyreducedinsizecomparedtoothershellforms.Specieswithanamusioidformrarelyattach, andassuchtherightvalvehasasmallbyssalnotchorlacksonealltogether.
SinceallfiveshellshapeshaveevolvedinarepetitivefashioninthehistoryofthePectinidae,theydonot denotehigher-leveltaxa.Rather,ithasbeendemonstratedthatshellformsreflectlifehabitratherthanhomologoustraits(Stanley,1970;Alejandrinoetal.,2011;Serbetal.,2011).Thus,theutilityofshellformasthesole basisofanyscallopclassificationsystemisquestionable(Waller,1991,2006a).Indeed,80%oftherecentandfossilspeciesandsubspecieswereoriginallyplacedintoonlyfivegenera, Chlamys, Aequipecten, PectenAmusium,and Hinnites,basedsolelyonshellform.Asaresult,thesefivegenerawererecognisedaspolyphyletic,and
group
FIGURE1.3 Waller’s1991 phylogenetichypothesisforscallopgroupscontainingcommercialspecies.Phylogenyisplottedagainsttimein millionsofyears(basedon Harlandetal.,1982).Numbersonbranchesrefertomorphologicalcharacterswhichdefineclades:(1)resilium withnon-calicifedcore;(2)ctenolium;(3)earlypre-radialstageofleftdissoconchwithantimarginalgroovesandridgeletsinmicrosculpture, resilial,anddorsalhingeteetharenearlyequallydeveloped;(4)earlypre-radialstageofmicrosculpturemaybesimilarto(1)inbeingpredominatelyantimarginalorpatternmaybeobscuredbyclosed-spacedcommarginallirae;tendencyforreductionofresilialhingeteethwhile dentalpatternbecomesdominatedbydorsal,infradorsal,orsupradorsalteeth;(5)earlypre-radialstageofmicrosculptureofleftvalveis eitherveryfinelypittedorcompletelysmooth,hingedominatedbydorsaland/orintermediateteeth,internalribcarinaebecomesnearlyuniversalinthe Decatopecten and Pecten groups;(6)prismaticstageofrightvalveissecondarilyextendedtoaheightofseveralmillimetres(in contrasttomoreancestralpectinids,whereitislimitedto ,2mm),leftpre-radialstageisgenerallysmooth,rarelyfinelypitted;(7)earlypreradialstageofmicrosculptureofleftvalveisverycoarselypittedandthesepitsarenearlyequidimensionalremnantsofoncecontinuousantimarginalgrooves,hingeteethsimilarto Chlamys group;and(8)earlypre-radialstageofmicrosculptureofleftvalveisverycoarselypitted, hingedentitionbecomesdominatedbyresilialteeth,atleastinearlygrowthstages.
Source:Imagereprintedfrom Waller(1991) withpermission.
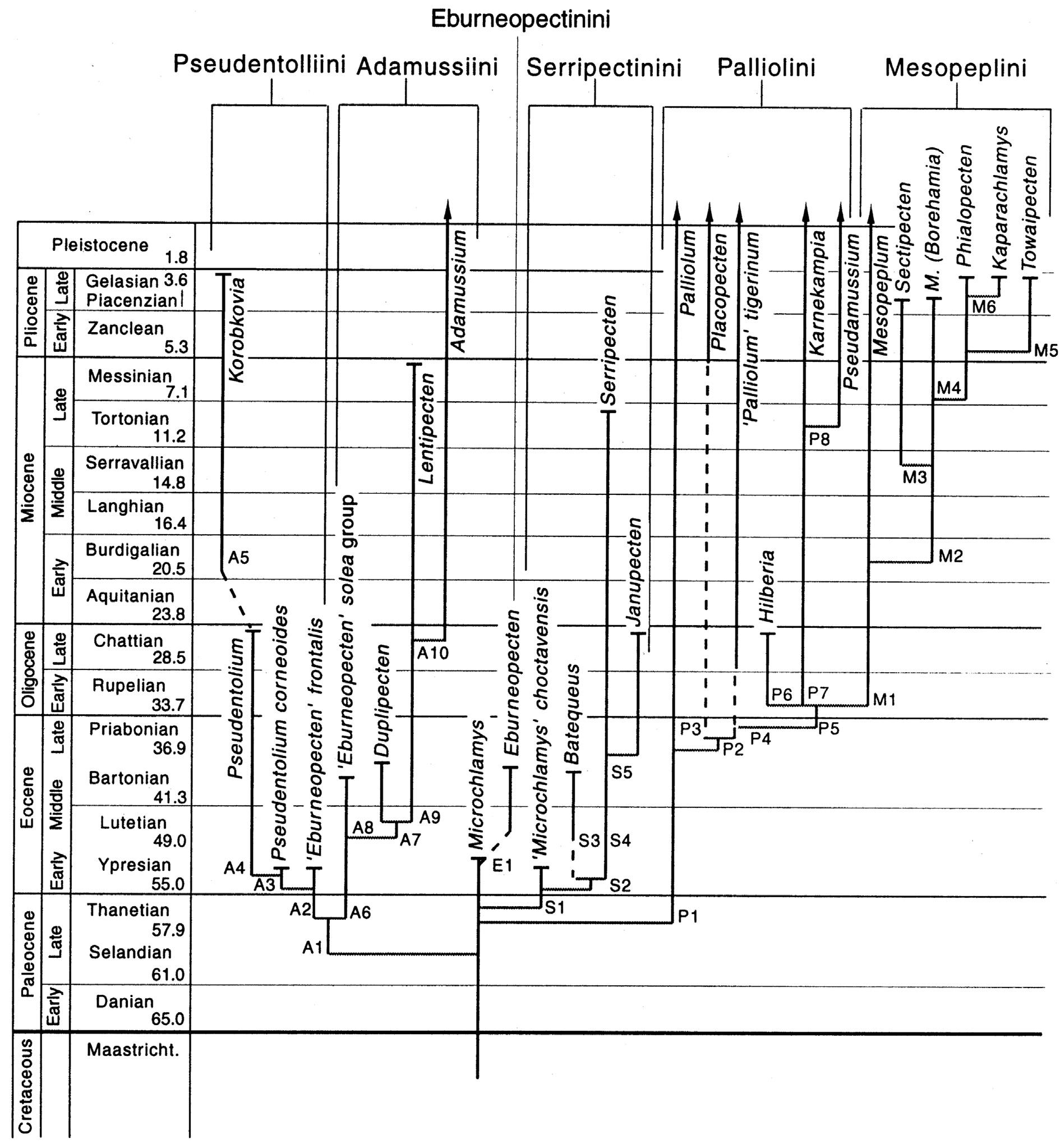
(e.g. RainesandPoppe,2006; Dijkstra,2013a).Inhis1993work, Waller(1993) focusedonrelationshipswithinthe Chlamydinae,whichhefurthersubdividedintofourtribes(Chlamydini,Crassadomini,Mimachlamydini,and Aequipectinini).Thispaperalsoincludedtheintroductionoftwonewextanttribes(Crassadomini, Mimachlamydini),threenewextantgenera(Caribachlamys Waller,1993, Laevichlamys Waller,1993 and Spathochlamys Waller,1993),andclearhypothesesfortaxonomicrelationshipsamonglineages,whichhereferred toas‘paraclades’,that‘cannotyetbecharacterizedonthebasisofuniqueapomorphies(=synapomorphies)’ (Waller,1993:p.199).Onemajorcontributionofhis1993workwastodividespeciespossessingthecommon chlamydoidformintodifferenttribes(Chlamydini,Crassadomini,Mimachlamydini),asWallerrecognisedthis shellshapeasplesiomorphic.Later, Waller(2006a) revisedhistaxonomicclassificationmovingthePalliolini fromthePectininae(Waller,1993:Figure1)andelevatedthetribetothesubfamily,Palliolinae(Figure1.4).The Palliolinaewasdividedintofivetribes,threeofwhich(Adamussiini,Mesopeplini,Palliolini)includeextanttaxa
8 1.RECONCILINGMORPHOLOGICALANDMOLECULARAPPROACHESINDEVELOPINGAPHYLOGENYFORTHEPECTINIDAE(MOLLUSCA:BIVALVIA)
FIGURE1.4 Waller’s2006phylogenetichypothesisforthesubfamilyPalliolinae.Dashedlinesindicatemissingfossilrecords.Branches endinginarrowsrepresentextantlineages,thoseendingincross-barsareextincttaxa.Letter-numberlabelsonnodesrefertocharacters describedin(Waller,2006a).Timeinmillionsofyearsfrom Berggrenetal.(1995). Source:Imagereprintedfrom Waller(2006a) withpermission.
fromthegenera Adamussium Thiele,1934, Palliolum Monterosato,1884, Placopecten Verrill,1897,and Pseudamussium Morch,1853.Basedonevidencefrommolecularphylogeneticstudies, Waller(2006a) createdthe tribeAmusiiniandmoved Annachlamys Iredale,1939and Flexopecten Sacco,1897fromtheDecatopectininitothe PectininiandAequipectinini,respectively(Table1.1)(Figure1.5).
Waller’sworkpresentsmanyfirsts.Hewasthefirstworkerinscallopstoroutinelyincorporatefossilandlivingtaxaintophylogenetichypotheses(Waller,1969,1991,2006a).Hewasthefirsttodevelopafamilialclassificationsystembasedonaphylogeny,notsimplybinningmorphologicallysimilarspeciesintogroups(Waller,1991, 2006a).Hewasthefirstresearchertoovercomeadependenceofusinghomoplasiouscharactersinscallopmorphologyandsetouttodevelopasetofcharacterstoavoidissueswithconvergenceintheadultshellshape
Mostrecently, RainesandPoppe(2006) compiledabeautifullyphotographediconographyof250speciesin thePectinidae.Theclassificationsystemtheyemployedprimarilyfollowstheoneby Hertlein(1969),butmodifiedbytheworksofDijkstra,Habe,Kafanov,Wagner,andWaller.TheclassificationofRainesandPoppeisthe onlysystemtorecognisethesubfamilyHemipectinae.Inaddition,theirsubfamilyChlamydinaeincludesmore tribesthanothermoderntaxonomictreatments(Table1.1).
Thefourmostcomprehensiveclassificationsystemsof Hertlein(1969), Waller(1986,1991,1993,2006a), Raines andPoppe(2006),and Dijkstra(2014) arecomparedin Table1.1.Differencesamongtheclassificationsystems includewhetherextincttaxaareintegrated(Hertlein,Waller),ifallcurrentlyrecognisedspeciesareincluded (RainesandPoppe,Dijkstra),andiftheauthorgivesahypothesisofhowhighertaxaarerelatedtooneanother (Waller).Thesemodernclassificationsystemshaveinfluencedandsubsequentlyhavebeeninfluencedby(e.g. Waller,2006a)themolecular-basedphylogeneticstudiesdescribedbelow.
THEINFLUENCEOFMOLECULARPHYLOGENETICSONSYSTEMATICS
Inmolecularphylogenetics,speciesarerepresentedbynucleotidesequencesthatencodegenes,oraminoacid sequencesthatencodeproteins.Thesedataarealignedascharacterstatesinamatrixandanalysedunderamathematicalmodeldescribingevolutionarychange.Themethodsoftreeestimationincludeavarietyofapproaches andthereisalargebodyofliteratureonthesubject(Hillisetal.,1996;Swoffordetal.,1996;Felsenstein,2004). Currentlyacceptedphylogeneticmethodsutilisesometypeofoptimalitycriterion,whichisameasureoffitof thedata(charactermatrix)toagivenhypothesis(phylogenetictree).Thesemethodsinclude:Maximum Parsimony(MP: Fitch,1971);MaximumLikelihood(ML: Felsenstein,1981);andBayesianInference(BI: Yangand Rannala,1997).MP,ML,andBIarecharacter-basedmethods,wheretheoptimalitycriterionisappliedtoeach position(nucleotideoraminoacid)alongthealignment.Amongthese,MLandBIuseexplicitmodelsof sequenceevolutiontosearchforthebesttree(ML)ortrees(BI)thatareconsistentwithboththemodelanddata. Incontrast,distance-basedmethods,suchasneighbour-joining(SaitouandNei,1987),summarisethemolecular charactermatrixintoadistancematrix,wheresequencesinthealignmentarecomparedinapairwisefashionto calculatethefractionofsimilarity,thenaseriesofalgorithmsareappliedtoconstructasingletree.Theusefulnessofanyofthesemethodsdependsonitsphylogeneticaccuracy,theabilityofamethodtorecoveratopology thatbestfitsthedataathand.Computersimulation(Hall,2005;OgdenandRosenberg,2006),observationalbased(Yang,1996),andexperimentalevolution(Hillisetal.,1994)studieshaveagreedthattheapproachesthat areexplicitlymodel-based(BIandML)aremorephylogeneticallyaccuratethanMPestimates.Distance-based approaches,suchasneighbour-joiningorUPGMA,aretheleastaccurate.Adescriptionofthesimilaritiesand differencesamongtheoptimalitycriteriaandotherapproachescanbefoundin Hall(2008).Itisworthnoting thatphylogeneticinferencemethodsareundercontinualevaluationandre-developmenttoimprovetheaccuracy andspeedofcomputation.
Molecularphylogeneticsisapowerfultoolindevelopingaclassificationsystembecause:(i)molecularmarkers (e.g.genes)canbeselectedthatareindependentofshellmorphologyand,presumably,thesemarkerswillbeless influencedbyconvergentevolutionsothattheresultingtreeisamoreaccuraterepresentationofthedata; (ii)molecularmatriceshavealargenumberofcharactersformorerobustandaccuratephylogenetichypotheses; (iii)molecularcharactershavediscretestates(4nucleotidesor23aminoacids)whicharelesssubjectiveand reduceerrorinstatecalling;and(iv)selectionofappropriateoutgrouptaxacanbeusedtoidentifytheoldest lineageofthestudygroup(i.e.‘ingroup’),totestthemonophylyoftheingroupandtopolarisethehistorical sequenceofallsubsequentevolutionaryevents.Thus,themajorlimitationsinamolecularphylogeneticstudy aretheavailabilityofDNAorRNAsamplesforthetaxaofinterestandthenumberofmolecularmarkersdevelopedforthatparticulargroupoforganisms.
MolecularPhylogeneticStudiesofScallops
Earlymolecularphylogeneticstudiesthatincludedpectinidtaxawereinvestigatingrelationshipswithin Bivalvia.ManyofthesestudiesusedDNAsequencefromasinglenuclearlocus,18SrRNA(Riceetal.,1993; KenchingtonandRoddick,1994;SteinerandMuller,1996;Frischeretal.,1998;GiribetandCarranza,1999; SteinerandHammer,2000).Areviewofthesepapersisgivenin Waller(2006a).Primarily,thesestudiesexaminedthephylogeneticplacementofthePectinidaewithintheBivalvia.Becauseofthisbroadtaxonomicfocus,