Historical review
An invertebrate (nonvertebrate) refers specifically to an animal without a backbone, in contrast to the vertebrates represented by fish, amphibians, reptiles, birds, and mammals. An invertebrate animal has no central skeleton and has a ventral nervous system, while the heart is dorsal in position, with an open circulatory system, and its integument is formed of secretory substances, generally tanned proteins and sometimes chitinized.
At present, about 1.5 million species of invertebrates have been named and about 8000–10,000 new species are added to the list each year. With the exception of the protozoans (about 30,000 species) and the vertebrates (about 43,000 species), all other animals are invertebrates, comprising altogether about 95% of the animals in the animal kingdom. Strictly speaking, the protozoans are also invertebrates, but a distinction is sometimes made. If we also consider the extinct species, the total number of species that have lived would be much greater than that of the existing forms alone. The numbers of species are just estimations and vary enormously amongst zoologists. Evolution of new species is a continuous process; in addition, names of new species added annually to the list might result from the discovery of forms that have long been in existence but have only just come to light.
The invertebrates are grouped into about 30 phyla, the number of which varies among different zoologists. A phylum is somewhat difficult to define, but it may be considered as a basic structural pattern of evolutionary descent from a common ancestry, with its members more or less homogeneous and bearing a unique combination of distinct characteristics. The members of a phylum are all constructed along the same general plan. Some of the phyla are major ones (according to the constituting number of species), while others are minor phyla: a minor phylum can be a group separated (e.g. by class and/or order) from a major phylum such as Nemertinia from Platyhelminthes.
Embryology and Reproduction. https://doi.org/10.1016/B978-0-12-814114-4.00001-1
Nemertinia was elevated to the rank of a phylum since its members have additional and/or different characters. A minor phylum can also be a newly discovered group of animals, which was established independently as a phylum, e.g. phylum Pogonophora.
According to embryology, the invertebrate phyla are grouped into two lines:
Annelid line Echinoderm line
Egg potency Mosaic Equipotent
Cleavage Spiral Radial
Blastomeres Determinate Indeterminate Gastrulation Delamination Invagination
Coelom Schizocoele Enterocoele
Site of mouth Protostome Deuterostome
Larva Trochophore (ciliated band around the equator) Pluteus (ciliated band around the mouth)
Embryology (from Greek έμβρύον, embryon, ‘unborn embryo’; and -λογίa, -logia). It may be defined as that branch of biological science that deals with the development of the individual organisms. It is concerned with successive series of changes, in form and function, through which the initial germ of the new individual is transformed into a sexually mature adult. Normally, the germ with which development commences is an egg that has been fertilized by a sperm. The sexually mature adult is an individual that has developed to a point where it can produce mature eggs if a female, or sperm if a male. Sometimes the word ontogeny is used as a synonym for embryology, but more often it is defined to include the entire life history of an individual from its origin to its death.
Many principles of embryology apply to both invertebrate animals and vertebrates. Therefore, the study
of invertebrate embryology has advanced that of vertebrate embryology, despite the presence of many differences. The most obvious is the release of a larva in numerous invertebrate species before the completion of development.
The history of embryology has passed through three phases: fact-finding or descriptive, comparative, and experimental embryology. More recently, the molecular studies have been established.
DESCRIPTIVE PHASE
The first famous name associated with this period is Aristotle.
Aristotle (384–322 BC)
With the possible exception of Plato, Aristotle is by far the most influential philosopher in the history of Western thinking. Aristotle studied natural sciences, and these studies dominated until early modern times. Aristotle’s influence was vast and extended far beyond philosophy. For example, he was the founder of biology, and Charles Darwin (1809–82) regarded him as the most important contributor to more than one subject.
The influence of Aristotle’s family (his father was a physician) initiated his strong interest in anatomy, and the structure of living organisms in general. He described the reproduction and development of many kinds of animals. He was the first to study and write the accounts of development of the hen’s egg. His Macedonian origin, being a student, and then a teacher in Plato’s academy, and his connections with the group of outstanding scientists surrounding Plato (who worked in a wide variety of subjects, ranging from medicine and biology to mathematics and astronomy), contributed to him developing remarkable observation skills. These media had greatly influenced and characterized the scientific thinking of Aristotle. He was able to reach and formulate some scientifically significant findings as follows:
1. He noted the different methods by which animals originate from eggs, and described them as oviparous, viviparous, and ovoviviparous animals, and divided them into the following categories:
● Mammals: because their young arose from a mixture of male and female fluids and are born alive. The eggs of mammals had not been discovered at that time.
● Sharks: arose from eggs that are retained within the body of the mother and are also born alive.
● Reptiles and birds: in which the eggs are complete ones.
● Amphibia and bony fishes: in which the eggs are incomplete (complete eggs are those with an albumen coat and a shell).
2. He identified the two major cell division patterns by which embryos are formed: complete division of the egg
(holoblastic), as in frogs and mammals; and incomplete division of the egg (meroblastic), as in chicks.
3. He was the first to figure out the functions of the placenta and the umbilical cord.
4. He also observed that the cephalic (anterior) part of the embryo grows faster than the posterior one (Fig. 1.1C and D). Therefore he discovered—as pointed out by Joseph Needham (one of the modern embryological pioneers) in 1934—the morphogenetic cephalocaudal gradient.
5. As a result of describing different embryos, he claimed the most valuable theory, which initiated massive arguments and discussions during later times, the epigenesis. This theory states that development proceeds from a simple formless beginning to the complex organization of the adult.
After Aristotle, there was hardly any progress in embryology for the next 2000 years. It was not until the 15th and 16th centuries that the science in general, and embryology in particular, was refreshed. The most famous man during that period was William Harvey.
The English Physician William Harvey (1578–1657) (Before the invention of the microscope)
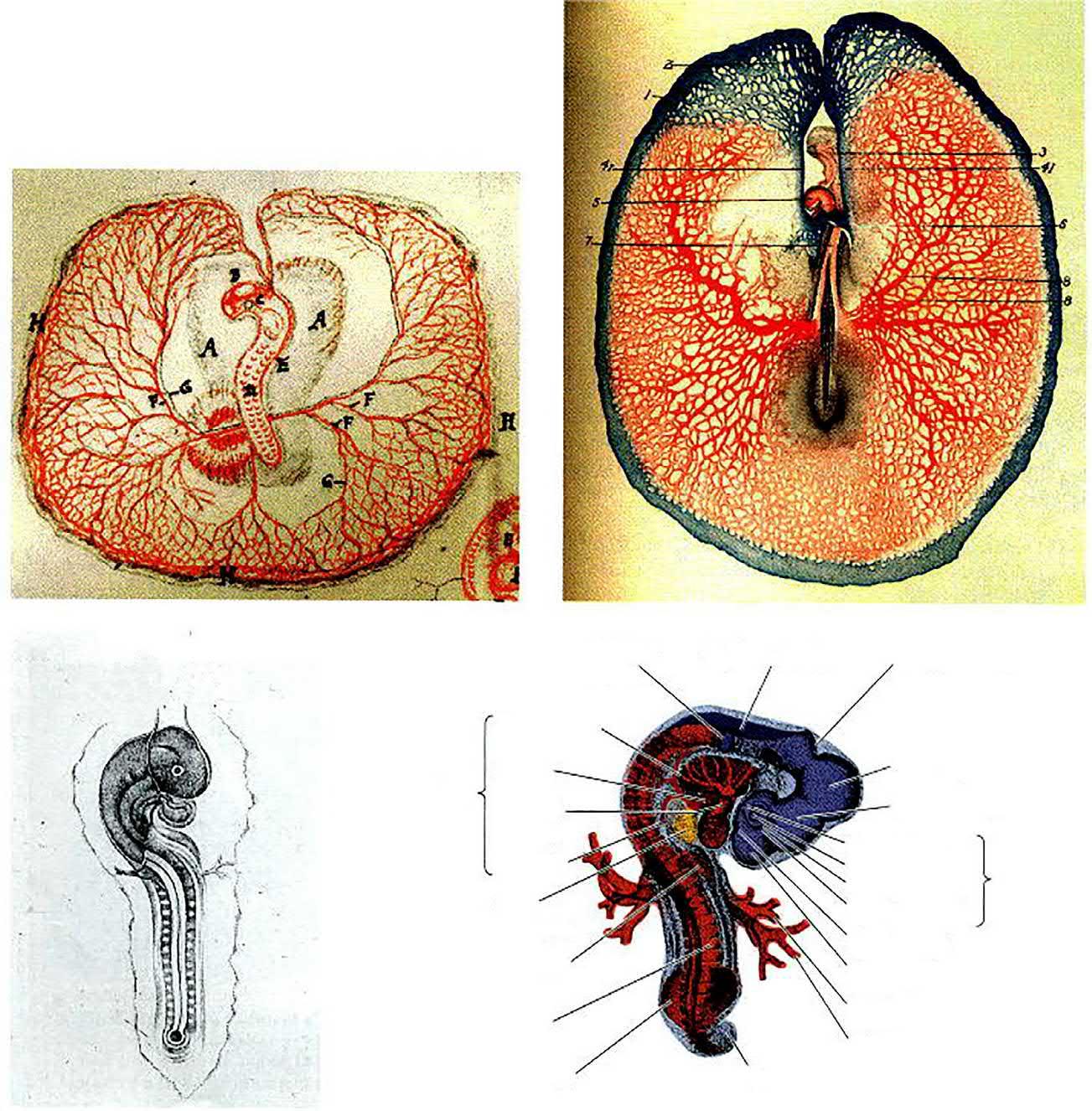
Harvey described the development of the chick in great detail. He also contributed a study of the development of the deer, and compared the two with each other. He was the first to observe the blastoderm of the chick embryo—that small region of the egg that contains the yolk-free cytoplasm which gives rise to the embryo. He was also the first to notice that islands of blood cells form before the heart does (Fig. 1.1A and B). Harvey suggested that the amniotic fluid might function as a shock absorber for the embryo. His most important finding was his conclusion that all animals arise from eggs, but this was based on theoretical considerations. He famously stated, ‘Omne vivum ex ovo’. He finally published a book, following his numerous studies and results, Exercitationes de generatione animalium (On Animal Generation), published in 1651. His doctrine of omne vivum ex ovo (all life comes from the egg) was the first definite statement against the idea of spontaneous generation.
In addition to all this work, Harvey’s main line of research was dealing with the blood circulation in the human body, and he became an embryologist, in addition to being the founder of modern physiology and medicine. He published his theories in a book titled Exercitatio Anatomica de Motu Cordis et Sanguinis in Animalibus (An Anatomical Study of the Motion of the Heart and of the Blood in Animals), where he explained how the heart propelled the blood in a circular course through the body. His discovery was received with great interest in England, although it was greeted with some scepticism on the Continent (www. en.wikipedia.org/wiki/William_Harvey and www.bbc.co.uk/history/historic_figures/harvey_william. shtml).
Auditory vesicle
Aor tic arches
Atrium Hear t
Tr uncus
ar terious
Ventricle
Liver
r udiment
Forelimb bud
Somite
Hindlimb bud
Myelencephalon
Metencephalon
Mesencephalon
Diencephalon
Choroid fissure
Lens Eye
Sensor y retina
Pigmented retina
Olfactory pit
Telencephalon
Vitelline vein
Vitelline ar tery
Tail
FIG. 1.1 Depictions of chick developmental anatomy. (A) Dorsal view (looking ‘down’ at what will become the back) of a 2-day chick embryo, as depicted by Marcello Malpighi in 1672. (B) Ventral view (looking ‘up’ at the prospective belly) of a chick embryo at a similar stage, seen through a dissecting microscope and rendered by F.R. Lillie in 1908. (C) Eduard D’Alton’s depiction of a later stage 2-day chick embryo in Pander (1817). (D) Modern rendering of a 3-day chick embryo after Carlson (1981). Details of the anatomy will be discussed in later chapters. (From Gilbert (2000).)
Discovery of the microscope
Leeuwenhoek, Anton van [lay'-vuhn-hook, ahn' -tohn vahn] (1632–1723)
Leeuwenhoek was a Dutch biologist and microscopist. He was a businessman, and became interested in science when he developed an interest in lenses and started building
simple microscopes as a hobby. Each microscope consisted of a flat brass or copper plate in which a small, single glass lens was mounted. The lens was held up to the eye, and the object to be studied was placed on the head of a movable pin, just on the other side of the lens. Leeuwenhoek built over 400 microscopes, whose focuses were amazingly accurate and clear. Many of these microscopes still exist. The most
powerful of these instruments can magnify objects up to 275 times. Although future microscopes were to contain more than one lens (compound microscopes), Leeuwenhoek’s single lens had such perfection that he was able to make great advances and thus to draw attention to his field.
Leeuwenhoek was the first person to observe single-celled animals (protozoans) with a microscope. He described them in a letter to the Royal Society, which published his detailed pictures in 1683. Leeuwenhoek was also the first person to observe clearly, using a microscope, and to describe the red blood cells (corpuscles) in humans and other animals, as well as sperm cells. In addition, he studied the structure of plants, the compound eyes of insects and the life cycles of fleas, aphids, and ants.
(After the invention and general use of the microscope)
As might be expected, the study of embryology was based on speculation, until the invention of the microscope, which allowed detailed observations. One of the founders of the study of microscopic anatomy was Marcello Malpighi.
The Italian Physiologist Marcello Malpighi (1628–94)
Malpighi was among the earliest scientists to study the minute structures of living organisms with the new tool then, the microscope. He studied what he thought to be the unincubated egg. He described the various parts in the blastoderm and/or organs of the embryo. The primary contribution of Malpighi was his successful presentation of a visible evidence for the first time, of the detailed constitution of the young embryo. Evidence made by one of the new tools was as certain in the 17th as in the 20th century, which drew popular enthusiasm. However, Malpighi believed that the organs of the embryo preexisted in the egg and these structures became visible as they increased in size. He formulated his theory of development, although he did not use the word preformation. A question was raised as to what he meant by the preexistence of the animal in the egg. He expressed his notions only tentatively, and he was, in fact, uncertain whether new structures existed before he observed them (according to Adelmann’s interpretation).
According to preformation theory, all the organs of the adult were prefigured in miniature form within the sperm, or, more usually the egg.
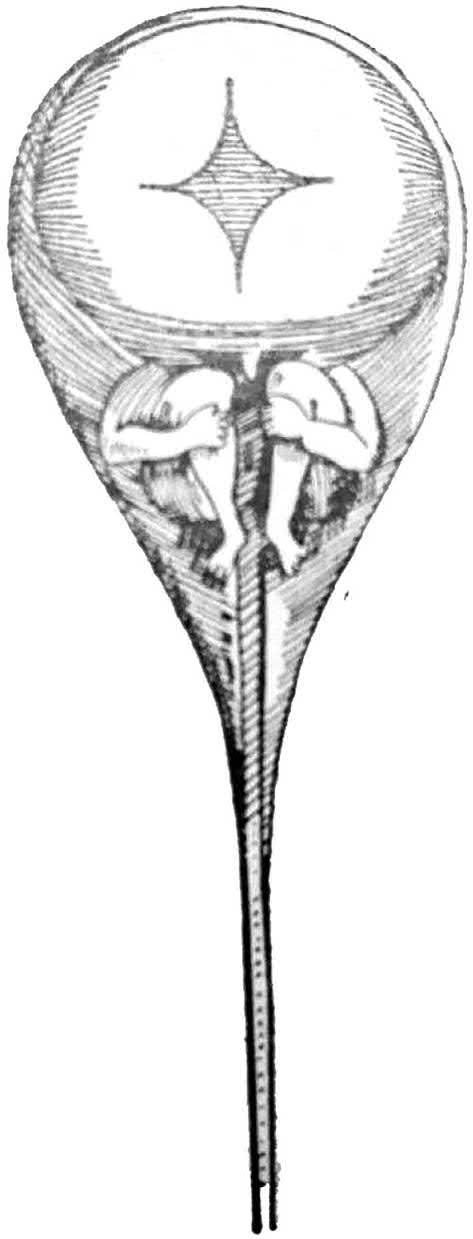
Consequently, many discussions and arguments took place during that period, and the scientists were split into ovists and animalculists. Leeuwenhoek, who co-discovered the sperm in 1678, and the other co-discoverer of sperm, Nicolas Hartsoeker, illustrated a drawing of what they hoped to find as a preformed human (‘homunculus’) within the human sperm (Fig. 1.2).
However, this theory of preformation opposed that of epigenesis by Aristotle, which resulted in a massive debate among the philosophically minded scientists of the 18th century. The most famous scientist who strongly disputed the preformation and revived Aristotle’s epigenesis was Caspar Friedrich Wolff.
FIG. 1.2 A hypothetical drawing of the human sperm by Hartsoeker (1694), showing a miniature of the embryo preformed in the sperm. (From the Arabic version of Comparative Embryology of the Vertebrates, the original by Huettner (1949).)
The German Caspar Friedrich Wolff (1733–94) Wolff was an anatomist and embryologist working in St Petersburg. He is often considered a founder of modern embryology at that time. He revived the view of one of the predecessors, Maître-Jan, who pointed out and explained, in 1722, the egg examined by Malpighi. His opinion was that Malpighi had studied an unincubated egg, after exposure to the heat of Bologna, not a fresh egg. After careful observation of the development of chick embryos, Wolff reached the first proof of epigenesis. He observed and identified that the blood vessels of the chick blastoderm are not present from
the beginning. He also studied plant development; he was able to compare the development of some internal animal organs; and he interpreted the development of the intestine, and probably the central nervous system, which are formed by the folding of homogeneous tissue layers into tubes. This meant that the embryonic organs develop from tissues that have no counterpart in the adult organism, and he foreshadowed the germ layers. At the same time, Wolff supported epigenesis, and he is considered to be the father of epigenetic descriptive embryology. He marked the point when embryonic development began to be described in terms of the proliferation of cells, rather than preformed miniature. He postulated an unknown force, essential force, which acts like gravity or magnetism that organizes embryonic development. In his Theoria Generationis (1759), he tried to explain the emergence of the organism by the actions of a ‘vis essentialis’, an organizing, formative force, and declared that ‘all believers in epigenesis are vitalists’.
Several scientists and/or philosophers believed in this view, and added contributions to support it. Then, a mechanical, goal-directed force called developmental force (Bildungstrieb) was postulated—not theoretically, but it could be shown to exist by experimentation, e.g. regeneration of Hydra when cut. Thus, development could proceed through a predetermined force inherent in the matter of the embryo. This hypothesis, in which epigenetic development is directed by preformed instructions, is not far from the view held by modern biologists that most of the instructions for forming the organism are already present in the egg.
COMPARATIVE PHASE
The end of preformationism did not come until the 1820s, when a revolution and evolution in descriptive embryology was created by the new microscopic investigators. Among the most talented of these was a group of three students (born in three consecutive years), of the philosophy of nature (Naturphilosophie) at Würzburg. They came from the Baltic region, but studied in northern Germany. They began to build constructively on Wolff’s concepts, including the epigenesis and the results of his microscopic examinations. With the combination of new staining techniques, improved microscopes, and institutional reforms in the German universities, these students were able to document the epigenesis of anatomical structures and to carry on the work of their teachers. These students were the German Heinz Christian Pander (born 1794), German-Russian Karl Ernst von Baer (born 1792) and German-Prussian Martin Heinrich Rathke (born 1793), and they became known as the three Founders of Modern Embryology.
The German Heinrich Christian von Pander (1794–1865)
Von Pander was an anatomist. He was the first to discover and demonstrate the three germ layers, in the embryo
of the chick. He had studied its development for 2 years (before becoming a palaeontologist). He thus realized that preformation could not be true. At the same time, his observations strengthened the belief in epigenesis since he discovered that organs are formed through interactions between simple structures. That is the view that we now call induction. Pander published a book, with glorious engravings (Fig. 1.1C), done by the artist Eduard D’Alton (a friend of Goethe who was one of the scientists of natural philosophy (Naturphilosophen), and whose influence on embryology was quite strong). However, Pander did not carry on, and von Baer took it upon himself to continue, and eventually broaden the base by additional examinations.
The German-Russian Karl Ernst von Baer (1792–1876)
Von Baer was a German-Russian biologist and the second of the three Founders of Modern Embryology of that time, but he was so famous that he was known as the ‘father of the modern embryology’. In 1828, he reported: “I have two small embryos preserved in alcohol that I forgot to label. I am unable to determine the genus to which they belong. They may be lizards, small birds or even mammals.” This quandary pushed von Baer to extend his study and examinations to many different vertebrates. Through his detailed study of chick development, and then comparing the results to the embryos of these different vertebrates, von Baer:
1. generalized Pander’s germ layers to other animals, and thus generalized the science of embryology itself; 2. was the first famous embryologist to establish a new branch known as comparative embryology;
3. reached many valuable results through his detailed studies. He discovered the notochord, then the neural tube. The most important discovery was that of the mammalian egg in 1827. He observed and identified the egg cell, differentiating it from the fluid-filled sac or Graffian follicle that surrounds it. He therefore, fulfilled and revived the theoretical consideration of Harvey (1578–1657) that ‘Life begins with an egg’; and
4. derived four generalizations (now often known as ‘von Baer’s laws’):
(a) The more general characteristics of any large group of animals appear in the embryo earlier than the more special characteristics, e.g. in all vertebrate embryos, the notochord appear first, shortly after gastrulation.
(b) After the more general characteristics, those that are less general arise, and so on until the most specific characters appear, e.g. all vertebrates initially have the same type of skin, and only later, the skin would develop: fish scales, reptilian scales, bird feathers, or the hair, claws, etc. of mammals.
(c) The embryo of any particular kind of animal grows unlike the forms of other species rather than passing
through them, e.g. human embryos never pass through a stage equivalent to an adult fish, frog, or bird.
(d) The embryo of a higher species may resemble the embryo of a lower species, but never resembles the adult of that species, e.g. human embryos initially share common characteristics with those of lower species, but none of them passes through the stages of the others.
Von Baer formulated these generalizations prior to Darwin’s theory of evolution in 1856. ‘Lower animals’ would be those appearing earlier in life’s history. His results initiated many discussions and arguments with others, especially philosophers. Afterwards, he understood the biogenetic law, which was clarified by Meyer (1935) as cited in Holmes (1944), and which was based on the recapitulation theory of Haeckel (1834–1919).
The German-Prussian Martin Heinrich Rathke (1793–1860)
Rathke was the last of the three Founders of Modern Embryology. His quality of mind, in particular, was in many ways similar to that of von Baer. He made distinguishable advances in demonstrating the existence of the germ layers in the invertebrates. He studied carefully the development of many different species especially the crayfish. On the other hand, Rathke looked at the development of some vertebrates (frogs, salamanders, fish, birds, and mammals). He emphasized the similarities in the development of all these vertebrate groups. These studies contributed significantly to a variety of topics. He was the first to discover the embryonic precursors of gills in embryos of higher animals that lack gills as adults, as well as, in mammals. In 1838, he published an important study of the pituitary gland. This embryologist is now known by his associated eponyms: Rathke’s pouch—a depression in the roof of the embryonic mouth in front of the buccopharyngeal membrane; and Rathke’s pouch cysts—groups of epithelial cells forming small colloid-filled cysts in the pars intermedia of the pituitary gland.
These were thus the three Founders of Modern Embryology: von Baer had been a student at Göttingen with von Pander. Rathke studied natural history and medicine at the University of Göttingen in 1814. Later, in 1818, he moved to Berlin to receive his MD degree. He became a physician, and was appointed in 1829 as a professor of physiology and pathology at the University of Dorpat. While in Dorpat, Rathke established contact with Karl Ernst von Baer, who was then a professor at Königsberg. When Baer left Königsberg for St Petersburg in 1834, Rathke succeeded him as a professor of zoology and anatomy. From there, these three scientists met, and started sharing their interest in embryology. They complemented each other’s researches and added to their achievements.
Embryology and other subjects
Since the time of von Baer up to the present time, biological sciences, in general, have greatly improved. The comparative embryology has been influenced by different special studies and theories that are associated with it, such as:
● cell theory;
● chromosome theory;
● genetics and gene theory;
● phylogeny and recapitulation theory; and
● retention theory.
Cell theory
This theory was developed during the time of von Baer, by the German physiologist Theodor Schwann (1810–82) and the German botanist Matthias Jacob Schleiden (1804–81). They are both credited for formulating the cell theory, which believed that all living organisms are composed of cells. Schwann studied medicine at the University of Berlin. His most important work was the microscopic study of animal tissue, which led to the idea that all living organisms are made up of units-cells, and that each contains essential components such as a nucleus. He published this idea in Microscopic Researches into accordance in the Structure and Growth of Animals and Plants (1839).
Schleiden, after studying law, concentrated on botanical studies. In 1838, 1 year before Schwann, he developed the idea that the cell is the basic unit of plants and that growth consisted of production and development of new cells. Schleiden’s two-volume textbook Principles of Scientific Botany (1842–43) was for a long time a model for modern botanical works. The cell theory was extended by other scientists, and it drew the attention of biologists to the study of the cell contents. Embryologists, on the other hand, started to base their field on the concept of the cell.
Chromosome theory
This theory, which deals with inheritance, was announced at the beginning of the 20th century by Walter Stanborough Sutton (1877–1916). He was an American surgeon and geneticist, and his work formed the basis of the chromosome theory of heredity. The latter greatly influenced the course of the field of genetics. In 1902, Sutton proposed that chromosomes during division make the basis of Mendelian inheritance.
Genetics and gene theory
The American zoologist Thomas Hunt Morgan (1866–1945) was a professor of experimental zoology at Columbia University (1904–28), where he conducted extensive breeding experiments on the fruit fly, Drosophila melanogaster. In 1927, he published a book entitled Experimental Embryology. From his research, he concluded that the individual units of heredity, the genes, are arranged in a line
on the chromosomes. This theory, known as the chromosome theory of heredity, was presented in 1915, in The Mechanism of Mendelian Heredity, which he wrote with his co-workers, A.H. Sturtevant and C.B. Bridges. He claimed that embryology is fundamentally the study of heredity in action and he gave a support to Mendel’s laws of inheritance; however, these were rejected for some time.
In 1926, Morgan wrote The Theory of the Gene, in which he stated that genes are held together in linkage groups, and that there is an interchange of alleles, genes, affecting the same trait. This interchange, known as crossing-over, allowed Morgan to construct chromosome maps showing the relative location of many genes.
For his work in the field of genetics, Morgan was awarded the Nobel Prize for physiology or medicine in 1933.
Phylogeny and recapitulation theory
This theory was published by Ernst Heinrich Haeckel (1834–1919), a German naturalist who made important discoveries in zoology. He is best known for his enthusiastic early support to Darwin’s theory of evolution, and being its main expounder on the European continent. He presented strong evidence of evolution in 1866, in his book General Morphology, and is famous for popularizing the phrase ‘ontogeny repeats phylogeny’. This phrase is a brief form of his recapitulation theory, later known as the biogenetic law. This theory states that ontogeny is a brief and incomplete recapitulation of phylogeny, or that an animal passes through stages in its development that are comparable to those which its ancestors passed through during their evolution. Haeckel also made significant contributions to knowledge of invertebrates and influenced the course of embryological research.
Retention theory
This theory has been proposed by the British biologist and Darwinian scholar, De Beer.
Sir Gavin De Beer (1899–1972)
De Beer is one of the modern embryology pioneers. In addition to being a zoologist and morphologist, he also contributed to experimental embryology, anatomy and evolution. He refuted the germ-layer theory, and developed the concept of paedomorphism: the retention of juvenile characteristics of ancestors in mature adults. This retention theory explains that such structures found in the bodies of some animals correspond to similar parts used for the same or different purposes in the body of some distant ancestor. Thus the vestigial tail of the human embryo, for example, arises in the same place and manner as the tails of other vertebrates, and therefore there is no doubt that some remote prehuman ancestor made use of a tail.
De Beer also developed the theory of piecemeal or mosaic evolution, on the basis of his work on the fossil
Archaeopteryx, which had reptilian and avian traits, explaining the combination of both bird and reptile features. He elucidated the embryological origins of many tissues, wrote the authoritative Charles Darwin: Scientific Biography (1963) and served as a director of the British Natural History Museum (1950–60).
EXPERIMENTAL PHASE
A new generation of embryologists then felt that embryology should not merely be a guide to study anatomy and evolution, and that it should answer the question: ‘How does an egg become an adult?’ Embryologists were to study the mechanisms of organ formation (morphogenesis) and differentiation. This new branch of study was given a German name that means ‘causal embryology’, ‘physiological embryology’, or ‘developmental mechanics’. These scientists began working with this new concept in the 1870s, and the first who led the group in this was Wilhelm His.
The Swiss-German Wilhelm His (1831–1904)
His was a researcher and teacher, who contributed to the fields of anatomy, histology and embryology. In addition to his investigations on some organs, he was the first to attempt a causal explanation of embryonic development. His was the most vocal against Haeckel theory and the most explicit. He claimed, with confidence, that during development of an embryo, any organ develops according to its future physiological function; and each stage—e.g. cleavage, blastulation, gastrulation, etc.—triggers the development of the following stage. According to this thinking, development is a mechanical and/or physiological process and never a recapitulation of the ancestral history.
One of His’ remarkable achievements was his invention of the microtome (1870), a machine that can make very thin sections for the material that complements the micro-technique. It makes very accurate and better serial sections, even for a whole embryo that facilitates the detailed observation.
Around the same era, there was another scientist whose studies were related to and/or flourished the experimental embryology.
The German August Weismann (1834–1914)
Weismann was a German biologist who made fundamental contributions to the understanding of the processes of heredity and evolution. He proposed a hypothesis of development based on the fate map of the frog embryo. Weismann claimed that when the first cleavage division separated the future right half of the embryo from the future left half, there would be a separation of ‘right’ determinants from ‘left’ determinants in the resulting blastomeres. This led him to distinguish ‘germplasm’ (hereditary elements carried by sex cells) from ‘somatoplasm’ (the rest of the body). He thus proposed a model of inheritance that included a
strict separation, in the early embryo, of a germ-line from the soma. He noted that the germplasm, on passing from generation to generation, remains largely unaffected by the environment (this work was based on the scant knowledge of fertilization available at that time).
Among Weismann’s important works were Studies in the Theory of Descent (2 vols. 1882), The Germ-plasm: A Theory of Heredity (1893), and The Evolution Theory (2 vols. 1904).
One of the most eminent pioneers in the analysis of development at the same time as Weismann was Boveri.
The German Theodor Heinrich Boveri (1862–1915)
Boveri mostly preferred the sea urchin embryos, on which he experimented during the summer months at the Marine Biological Station in Naples-Italy. His work on sea urchins showed that it was necessary to have all chromosomes present, for proper embryonic development to take place. This discovery was an important part of the BoveriSutton chromosome theory (see above). His other significant discovery was the ‘centrosome’ (1888), which he described as the especial organ of cell division
As some of Boveri’s countrymen appreciated the importance of worms, he started in 1880, to study the embryogenesis of the nematode Ascaris megalocepha (=Parascaris equorum), a parasite found in the intestine of horses. The females produce millions of eggs, releasing more than 100,000 per day. In fixed and stained early cleavage stages, Boveri found a peculiar but reproducible division pattern accompanied by loss of chromatin in most cells. The first division generates two cells, one of which, at the following division, passes through a process called ‘chromatin diminution’ (see Chapter 9). Since Boveri was the first to discover this phenomenon, it should be his associated eponym.
Cell theory was formulated in 1838 and 1839, and was well accepted by scientists working on different lines of science. Wilhelm His disputed Haeckel’s theory violently. Weismann proposed his germ-plasm theory with a new line of research. In the late 1800s, these three significant events explored different approaches of study in the field of embryology leading to the appearance of different schools of embryologists. Each school specialized in a definite major technique such as:
● defect, destroying a portion of the embryo;
● isolation, removing a portion of the embryo;
● recombination, replacing an original part with a part from a different region of the embryo;
● transplantation, whereby a portion of the embryo is replaced by a portion from a different embryo; and
● fate mapping experiments, tracing the fate of a portion and/or a cell of the embryo.
The work of these schools was parallel and/or succeeded each other, and their researches complemented each other,
which greatly extended and flourished this new branch of experimental embryology. Each school had a leader or one famous scientist who planned and created a firm basis for the school’s line of work. One of the most famous of these leaders was Wilhelm Roux.
The German Wilhelm Roux (1850–1924)
Roux was a young German embryologist and was considered the founder of the branch of experimental embryology. He strongly believed in the ideas and thoughts of His, in spite of the fact that Roux was a student of Haeckel, and considered himself his disciple. He unarguably acted as an intermediator between Darwinism and the new causal analytical embryology. The attempts at mechanical explanations of development were started by His and remained lacking in experimentation until Wilhelm Roux, who was able to succeed where His had apparently failed. This recalled a statement in the introduction of T.H. Morgan’s book Experimental Embryology: ‘The recognition of the advantage of applying the method of experiments to problems of development may be said to have begun with Wilhelm Roux in 1883’.
Roux was one of the first scientists to test Weismann’s hypothesis. He followed the defect experiments. In 1888, Roux published the results of a series of defect experiments on frog embryos. He freed one blastomere in the two-celled amphibian egg, from the influence of the other, by killing the latter. He observed that the surviving cell developed into a half-larva. We now know that Roux’s experiment was technically flawed. He thought that he had demonstrated the independent differentiation of the surviving cell. He interpreted the egg, for the first time, as a mechanism mechanically analyzable by outside interference. He concluded that the frog embryo was a mosaic of self-differentiating parts and that it was likely that each cell received a specific set of determinants and differentiated accordingly.
Roux not only performed the experiment, but was also able to generalize its significance. His ability to mobilize thoughts around it was due to his own qualities of mind and personality.
Some experimental embryologists, however, especially Driesch (see below), opposed Roux in his explanation that development simply proceeds mechanically. Eventually, the branch of experimental embryology was well established and differentiated into more specifications. The results and arguments tallied with the view of Roux. Many scientists, including cytologists, such as Boveri, Driesch, Delage, Brachet, and Flemming from Europe and Conklin, Harrison, Wilson, Loeb, Morgan, and Whiteman from the United States, agreed with the opinion and explanation of Roux.
The German Hans Adolf Eduard Driesch (1867–1941)
Driesch was a German experimental embryologist and philosopher. It is likely that nobody appreciated Roux’s work and the experimental approach to embryology more than
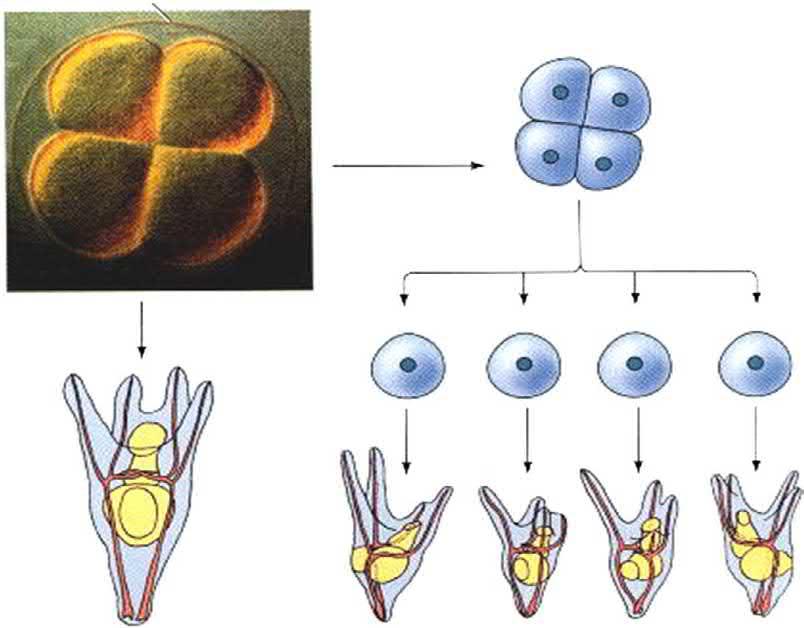
Hans Driesch. However, contrary to Roux, Driesch’s goal and attempts were to explain development, in terms of the laws of physics and mathematics. While Roux’s studies were ‘defect’ experiments, those of Driesch were ‘isolation’ ones. In 1891, Driesch and Fiedler carried out, independently, the separation of the two-cell sea urchin blastomeres from each other by vigorous shaking (or later, by placing them in calcium-free seawater). To Driesch’s surprise, each of the blastomeres of the embryo developed into a miniature, nearly complete larva. Similarly, when Driesch separated the blastomeres from fourand eight-cell embryos, some of the isolated cells produced entire pluteus larvae (Fig. 1.3). Driesch, then, reached a result that completely opposed the predictions of Weismann and Roux. He identified that each isolated blastomere regulated its development so as to produce a complete organism, rather than self-differentiating into its future embryonic part.
On the other hand, Driesch was a philosopher and one of the leading vitalists. He attempted to explain life processes through the theory of vitalism. At the same time, he was a teleologist. He believed that many of the basic problems of biology could not be solved simply by philosophy. In 1894, after publishing his papers on experiments on sea urchin eggs, Driesch wrote a theoretical essay, titled Analytische Theorie der organischen Entwicklung, in which he declared that his studies in developmental biology pointed to a ‘blueprint’ or teleology, an Aristotlean entelechy, a scientific demonstration of Immanuel Kant’s notion that the
organism develops as if it has a purposeful intelligence. The History and Theory of Vitalism (1905) was his main work.
In the same year of Driesch’s experiments on sea urchin eggs, C. Chun reported the results of experiments he had started in 1877, in which he traced the development of separated blastomeres of two-cell ctenophore embryos. He chose the eggs of the ctenophore, e.g. Beroë ovata, because it is excellent for experimental work, as it is large, clear, and does not disintegrate when cut. He observed that each cell of the two developed, giving rise to a half-embryo that possessed exactly half of the normal set of adult structures. These results provided a view of embryos as being composed of a ‘mosaic’ of individual parts, which was completely different from Driesch’s view. However, Driesch, being a powerful and arrogant man, refused and ridiculed Chun’s results, and made personal attacks on Chun’s integrity. T.H. Morgan collaborated with Driesch, repeated Chun’s experiments and observed the same results as Chun in 1895.
There is an explanation that allows for each of the two experiments of Driesch and Chun and their results being accurate. The embryos of many animals (including those of amphibians) possess remarkable regulative abilities, while the embryos of many other animals do not. This is what is now known as indeterminate and determinate blastomeres.
The discovery of regulation led to the discovery of the ‘organizers’ by Spemann.
Driesch’s demonstration of regulative development. (A) An intact four-cell sea urchin embryo generates a normal pluteus larva. (B) When one removes the four-cell embryo from its fertilization envelope and isolates each of the four cells, each cell can form a smaller, but normal, pluteus larva. (All larvae are drawn to the same scale.) Note that the four larvae derived in this way are not identical, despite their ability to generate all the necessary cell types. Such variations are also seen in adult sea urchins formed in this way. (From Gilbert (2000).)
tilization envelope
Plutei developed from single cells of 4-cell embr yo
Nor mal plutcus lar va
FIG. 1.3
The German Hans Spemann (1869–1941)
Spemann was a German zoologist, the first graduate student of Boveri (see above) who completed his PhD thesis on nematodes. He won the 1935 Nobel Prize for physiology or medicine, for his work on the ‘organizers’ in the amphibian embryo. Using frog and newt embryos, Spemann found that development is determined by interactions between various parts of the embryo, with special areas serving as organizers for the development of nearby related tissues.
Another line of his experimental research was ‘fate mapping the embryo’, which was based on the cell theory. Following individual cells during development to see what they would become was one of the most important programs in embryological studies. Tracing the fate of the blastomeres of an embryo leads one to define the cell lineages of that embryo, and to construct a fate map to the gastrula or blastula of such embryo. What is most remarkable in this field is the zeal and commitment of those embryologists (Conklin, Wilson, Boveri, etc.) who patiently studied the ‘cell lineage’ of the invertebrate eggs without any other help than the microscope.
The American Edwin Grant Conklin (1863–1952)
Conklin spent most of his career as a distinguished academician and scientist in the Department of Biology at Princeton University. When he was a student, he was greatly interested in biology. Conklin decided to enter the graduate school at Johns Hopkins University, an extraordinary place to obtain a graduate education in biology. At that time, most American biologists were naturalists or morphologists, while those in Europe (e.g. Roux, Boveri, Spemann, Driesch) had applied experimental methods to embryological research. Conklin’s graduate advisor, William K. Brooks, was not satisfied with that new experimental approach, while his fellow graduate students and contemporaries, including Morgan, Harrison and Wilson, were enthusiastic about this line, and actively tried to incorporate these methods into their dissertation research. The efforts of these graduate students were to be the roots of developmental biology in America. However, afterwards these individuals drifted in other directions as their careers matured.
Conklin, during the summer of 1890, was at Woods Hole in search of a suitable material for his doctoral dissertation. He chose the marine snail Crepidula, and discovered that its eggs divide according to a fixed pattern of spiral cleavage, which enabled him to follow the individual cells and their descendants to their final positions in the tissues and organs of the larva. In this he performed one of the first cell lineage analyses, which occurred at Woods Hole for the next decade. With this event began a lifelong love of the Marine Biological Laboratory at Woods Hole, where Conklin subsequently spent almost every summer to extend his cell lineage studies. In 1905, he studied the development of the
egg of the tunicate sea squirt Styela partita. He patiently followed the fates of each cell in the embryo (the descendants of each particular cell) until they differentiated into particular structures. He was helped only by the peculiarity of the Styela egg, wherein the different regions contain different pigments. The most peculiar of these regions was the yellow crescent, or myoplasm, which was deposited in the tail muscle cells of the tadpole larva. Conklin had discovered a natural cell lineage marker.
He then constructed a linear version of the fate map of such species (Fig. 1.4). This was to be his most important work and his associated eponym. Conklin’s fate map was confirmed by cell removal experiments, which is another line of studies. The removal of a certain cell in early cleavage produces an embryo without its descendants, i.e. the organ of that cell. For example, the removal of the B41 cell (which should produce all the tail musculature) resulted in the absence of tail muscles (Reverberi and Minganti, 1946).
Most embryos are not so accommodating as to have cells of different colours. In addition, not all embryos have as few cells as tunicates. The local staining method was introduced later by Vogt. In 1929, Vogt traced the fate of each blastomere in the developing amphibian embryo. He was thus able to establish a correct and precise ‘map’ of the cleaving embryo. A variety of methods have been used to study the fates of early blastomeres. One of the earliest studies was carried out by Hörstadius in 1939. He used the vital dye Nile Blue to mark the surface of individual cells in sea urchin embryos (Fig. 1.5). He found that all the mesomeres and some macromeres give rise to the ectoderm of the larva, and that micromeres become skeletogenic cells. Another fate mapping process was the radioactive labeling method. One of the problems with the vital dyes and radioactive labels is that they become diluted at each cell division. One way around this problem was the creation of fluorescent dyes, which could be injected into an individual cell of the early embryo. The descendants of that cell could then be seen under ultraviolet light. ‘Dil’, as an example, is a powerfully fluorescent molecule that becomes incorporated into lipid membranes, and has been used to follow the fates of cells and their progeny.
More recent studies have confirmed and considerably extended these observations, using intracellular injections of fluorescent tracer dyes in the sea urchin Strongylocentrotus purpuratus (Cameron and Davidson, 1991). Sea urchin blastomeres through at least the 32-cell stage have specific, predictable fates (Fig. 1.6), a condition common to many phyla called ‘invariant cell lineage’.
Many other experimental embryologists extended this branch by their studies on different subjects, such as F.R. Lillie (1870–1947) on fertilization, and J. Loeb (1859–1924) on artificial parthenogenesis. Experimental embryology, which began in the late 19th century and reached its golden period during the early 20th century, changed the meaning
Notochord Endoder m Neural ectoder m
Anterior
Anterior 2-Cell stage 4-Cell stage 8-Cell stage 16-Cell stage 32-Cell stage 64-Cell stage Mesenchyme (A)
Endoder m Mesenchyme Vegetal
A7.1
A7.2
A7.3
b6.5
b6.6
a7.10
a7.11
a7.12
a7.13
a7.14
a7.15
a7.16
B7.1
B7.2
B7.3
B7.4
B7.5
B7.6
B7.7
B7.8
b7.9
b7.10
b7.11
b7.12
b7.13
b7.14
b7.15
b7.16
Derivative
Endoder m
Endoder m
Notochord
Brain stem
Endoder m
Notochord
Notochord
Spinal cord.
Caudal muscle
Brain
Brain
Palps
Epider mis
Sense organ
Epider mis
Epider mis
Epider mis
Endoder m
Endoder m
Mesenchyme
Muscle
Muscle
Muscle
Mesenchyme
Muscle
Epider mis
caudal muscle
Epider mis
Epider mis
Epider mis
Epider mis
Epider mis
Epider mis
Epider mis
FIG. 1.4 Fate map of the tunicate embryo. (A) The one-cell embryo (left), shown shortly before the first cell division, with the fate of the cytoplasmic regions indicated. The eight-cell embryo on the right shows these regions after three cell divisions. (B) A linear version of the fate map, showing the fates of each cell of the embryo. ((A) After Nishida (1987) and Reverberi and Minganti (1946); (B) After Conklin (1905) and Nishida (1987). From Gilbert (2000).)
of embryology. Not only did its aim become descriptive or comparative, but also it deals with much-detailed phenomena and processes. It deals with the protoplasmic organization of the egg and the sperm, their genetic constitution and the nature of the process by which the sperm initiates development
in the egg. This modern embryology also traces the history of the different cleaved cells and their fates. It tries to explain the way in which the differentiation of each cell occurs. It is interested mainly in the mechanics of each process of each stage and how the different organs arise in the adult.
Ectoder m
m
Animal
Ectoderm
Endoderm
Vegetal
Ciliary tuft
Oral ectoder m
Vegetal plate
Stomodeum
Skeletal rods
Secondar y mesenchyme cells
Anus Anus
Primar y mesenchyme cells
Skeletal rods
Primary mesenchyme cells
Invagina ting endoder m (gut)
Mouth
Skeletal rods
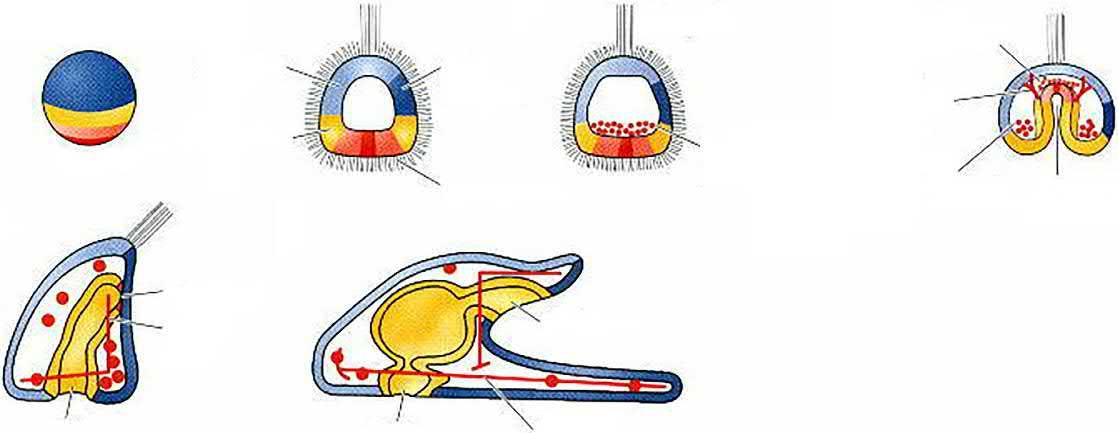
FIG. 1.5 Normal sea urchin development, following the fate of the cellular layers of the blastula. (A) Surface view of the blastula. (B) Late blastula with ciliary tuft and flattened vegetal plate. (C) Blastula with primary mesenchyme. (D) Gastrula with secondary mesenchyme. (E) Prism-stage larva. (F) Pluteus larva. Fates of the zygote cytoplasm can be followed through the colour pattern (Courtesy of D. McClay. From Gilbert (2000).)
Large micromere
Small micromere
Mesomere
Mesomere
Macromere
Aboral ectoder m
Oral ectoder m
Endoder m
Secondar y mesenchyme
Skeletogenic cells
Coelom
Micromere
Large micromere
Small micromere
FIG. 1.6 Fate map and cell lineage of the sea urchin Strongylocentrotus purpuratus. (A) The 60-cell embryo is shown, with the left side facing the viewer. Blastomere fates are segregated along the animal-vegetal axis of the egg. (B) Cell lineage map of the embryo. For simplicity, only one-quarter of the embryo is shown beyond second cleavage. The veg1 tier gives rise to both ectodermal and endodermal lineages, and the coelom comes from two sources: the second tier of micromeres, and some veg2 cells. (After Wray (1999). From Gilbert (2000).)
(F)
Vegetal pole
Animal pole
Gradually, it became largely replaced by molecular embryology. Molecular biology, in general, began in 1967, when Francis Crick co-discovered the DNA and unravelled its helical structure. Since that time, most embryology researches have dealt with the ontogeny of molecular diversity and with how different biochemical substances direct and control development. The origin of cellular diversity (molecular differentiation) in an individual, which is composed of genetically identical cells, is one of the central themes of embryological research. Developmental biology has thus emerged as a field of study that attempts to correlate the genes with morphological change, and to determine which genes are responsible for each morphological change that takes place in an embryo, and how these genes are regulated.
REFERENCES
Cameron, R.A., Davidson, E.H., 1991. Cell type specification during sea urchin development. Trends Genet. 7 (7), 212–218.
Carlson, B.M., 1981. Patten's Foundations of Embryology. McGraw-Hill, New York. Cited in Gilbert (2000).
Conklin, E.G., 1905. The organisation and cell lineage of the ascidian egg. J. Acad. Nat. Sci. Phila. 13, 1–119.
Gilbert, S.F., 2000. Developmental Biology, sixth ed. Sinauer Association, Inc. Publishers, Sunderland, MA.
Hartsoeker, N., 1694. Essai de dioptrique, Paris.
Holmes, S.J., 1944. Recapitulation and its supposed causes. Quart. Rev. Biol. 19 (4), 319–331.
Huettner, A.F., 1949. Fundamentals of Comparative Embryology of the Vertebrates. Macmillan Company, New York. (Translated by Sidra, S. and Abdel Wahab, A. Dar Nahdat Misr for Print and Publishing.)
Needham, J., 1934. A History of Embryology. Cambridge University Press, Cambridge, England. Cited in Willier et al., 1956.
Nishida, H., 1987. Cell lineage analysis in ascidian embryos by intracellular injection of a tracer enzyme. III. Up to the tissue-restricted stage. Dev. Biol. 121, 526–541.
Pander, C., 1817. Beitrage zur Entwicklung-eschichte des Hungers im Eye. Bonner, Wurzburg. Cited in Gilbert (2000).
Reverberi, G., Minganti, A., 1946. Fenomeni di evocazione nello sviluppo dell'uovo di Ascidie. Risultati dell'indagine sperimentale sull'ouvo di Asidiella aspersa e di Ascidia malaca allo stadio di 8 blastomeri. Pubbl. Sta. Zoo. Napoli 20, 199–252.
Wray, G.A., 1999. Introduction to sea urchins. In: Moody, S.A. (Ed.), Cell Lineage and Fate Determination. Academic Press, New York, pp. 3–9.