Lange Critical Care 1st Edition
John M. Oropello
https://ebookmass.com/product/lange-critical-care-1st-edition-john-moropello/
ebookmass.com
Laws of Depravity 1st Edition Eriq La Salle
https://ebookmass.com/product/laws-of-depravity-1st-edition-eriq-lasalle/
ebookmass.com
How to Calm Your Mind: Finding Peace and Productivity in Anxious Times Chris Bailey
https://ebookmass.com/product/how-to-calm-your-mind-finding-peace-andproductivity-in-anxious-times-chris-bailey/
ebookmass.com
Popular Culture, Voice and Linguistic Diversity: Young Adults On- and Offline 1st Edition Sender Dovchin
https://ebookmass.com/product/popular-culture-voice-and-linguisticdiversity-young-adults-on-and-offline-1st-edition-sender-dovchin/
ebookmass.com
Plumbing, Electricity, Acoustics: Sustainable Design Methods for Architecture, 1st Edition – Ebook PDF Version
https://ebookmass.com/product/plumbing-electricity-acousticssustainable-design-methods-for-architecture-1st-edition-ebook-pdfversion/
ebookmass.com
The Python Advantage: Python for excel in 2024 Hayden Van Der Post
https://ebookmass.com/product/the-python-advantage-python-for-excelin-2024-hayden-van-der-post/
ebookmass.com
Contents lists available at ScienceDirect
Aquaculture
journal homepage: www.elsevier.com/locate/aquaculture
Genetic parameters of fillet fatty acids and fat deposition in gilthead seabream (Sparus aurata) using the novel 30 k Medfish SNP array
S.S. Horn a, * , M.L. Aslam a , G.F. Difford a , K. Tsakoniti b , S. Karapanagiotis b , B. Gulzari c , J.W.M. Bastiaansen c , C. Penaloza d , R. Houston d , B. Ruyter a , A.K. Sonesson a
a Nofima, Norwegian Institute for Food, Fisheries and Aquaculture Research, NO-1433 Ås, Norway
b Galaxidi Marine Farm S.A., Galaxidi 330 52, Greece
c Wageningen University & Research, Animal Breeding and Genomics, PO Box 338, 6700 AH Wageningen, the Netherlands
d The Roslin Institute and Royal (Dick) School of Veterinary Studies, University of Edinburgh, Midlothian EH25 9RG, United Kingdom
ARTICLE INFO
Keywords:
Omega-3
Gilthead seabream
Lipid metabolism
Lipid deposition
Genetic parameters
ABSTRACT
Lipid-related traits are important candidates for a breeding goal for gilthead seabream, because they affect both fish and human health, as well as production efficiency. However, to date there have been very few estimates of genetic parameters for these traits, and the genetic relationship between fatty acids and other important traits have never been reported for gilthead seabream. Therefore, the aim of this study was to estimate genomic heritability and genetic relationships of fat deposition traits and individual muscle fatty acids in a commercial population of gilthead seabream using the novel ~30 k MedFish SNP array.
In total 967 gilthead seabream fed with a commercial feed were genotyped with the MedFish SNP chip which included ~30 K informative markers for this species. On average, the fish weighed 372 g. The mean content of eicosapentaenoic acid (EPA) + docosahexaenoic acid (DHA) was 822 mg per 100 g fillet. The heritability of muscle fat, viscera weight and percentage viscera were in the range of 0.34–0.46. The genetic correlation of body weight with muscle fat was 0.12, indicating that genetic variation in muscle fat is largely independent of the weight of the fish. The heritability of the product of endogenous fatty acid synthesis (n = 240), palmitoleic acid (16:1n-7), was high (0.43). The estimated heritability of EPA (%) and DHA (%) was 0.39 and 0.33, respectively. Both EPA and DHA had low, non-significant genetic correlations with body weight, and DHA had a negative genetic correlation with muscle fat ( 0.53).
It is possible to increase EPA and DHA content in gilthead seabream fillets by selective breeding. The high heritability of 16:1n-7, a marker of de novo lipogenesis, suggests that there is a strong genetic component to this metabolic pathway in gilthead seabream. Muscle fat deposition and body weight seem to be independent traits, and selective breeding for faster growth is not likely to influence the proportional content of EPA and DHA.
1. Introduction
Gilthead seabream is one of the major aquaculture species in Europe, ranked as number four, with a total production of almost 200,000 tons in 2019 (FEAP, 2020), but is at an early stage regarding selective breeding. The first commercial breeding programs of seabream were initiated in the early 2000's and have primarily focussed on improving growth rate (Brown, 2004; Janssen et al., 2017; Thorland et al., 2007).
* Corresponding author.
Lipids are important in production of all farmed animals because they are linked to production efficiency, health, and product quality. Excess dietary lipids that are not deposited in the edible muscle or used as energy source for growth are considered a loss and an indicator of low efficiency. Constant energy excess may lead to excessive fat deposition in and around internal organs and thereby increase the risk of metabolic disorders, oxidative stress and inflammation (Fontana et al., 2007; Todorcevi´ c et al., 2010).
E-mail addresses: siri.storteig.horn@nofima.no (S.S. Horn), Luqman.Aslam@Nofima.no (M.L. Aslam), gareth.difford@nofima.no (G.F. Difford), k.tsakoniti@ galaxidimarine.farm (K. Tsakoniti), s.karapanagiotis@galaxidimarine.farm (S. Karapanagiotis), benan.gulzari@wur.nl (B. Gulzari), john.bastiaansen@wur.nl (J.W.M. Bastiaansen), Carolina.Penaloza@roslin.ed.ac.uk (C. Penaloza), ross.houston@roslin.ed.ac.uk (R. Houston), Bente.Ruyter@Nofima.no (B. Ruyter), anna. sonesson@nofima.no (A.K. Sonesson).
https://doi.org/10.1016/j.aquaculture.2022.738292
Received 22 December 2021; Received in revised form 22 April 2022; Accepted 23 April 2022
Availableonline27April2022
0044-8486/©2022TheAuthors.PublishedbyElsevierB.V.ThisisanopenaccessarticleundertheCCBYlicense(http://creativecommons.org/licenses/by/4.0/).
The lipid content of muscle tissue affects organoleptic and processing qualities of fish fillets. In seabream, it has been shown that the taste, colour, and juiciness of fillets are affected by the fat content (Grigorakis et al., 2003; Grigorakis, 2007). Muscle fat also affects the nutritional quality of fillets. Fish is an important source of the essential omega-3 long-chain polyunsaturated fatty acids (LC-PUFAs), eicosapentaenoic acid (EPA, 20:5n-3) and docosahexaenoic acid (DHA, 22:6n-3) in human diets. EPA and DHA have several beneficial health effects, such as preventing and attenuating a range of inflammatory disorders, including cardiovascular disease, immune dysfunction, and obesity (Calder, 2015; Thota et al., 2018; Todorcevic and Hodson, 2015). Hence, improving the omega-3 content of fish fillet is a point of focus in aquaculture. As a significant part of the Mediterranean diet, seabream is an important source of these essential and health-promoting fatty acids.
In general, omega-3 fatty acid content of fish muscle is largely determined by the dietary fatty acid composition, which is well documented also for gilthead seabream (Ballester-Lozano et al., 2011; Grigorakis et al., 2002; Izquierdo et al., 2003; Mnari et al., 2007). In addition to feed, other factors including environmental factors (e.g. water temperature) have also been shown to influence the omega-3 fatty acid content of seabream muscle (Ibarz et al., 2005). Moreover, metabolic factors are likely to play a role, as higher DHA level in muscle compared to the feed have been observed in gilthead seabream (Izquierdo et al., 2003). Omega-3 content in fish fillets can also be regulated genetically: a recent study in gilthead seabream showed that there is a heritable component to variation in muscle content of EPA and DHA in gilthead seabream (Vallecillos et al., 2021). Dong et al. (2016) estimated a heritability of 0.44 for the omega-3 levels of fillets of the marine fish species large yellow croaker. Studies in Atlantic salmon have revealed medium to high heritability for the omega-3 levels of fillets and showed that genetic variation is possibly linked to omega-3 bioconversion capacity (Horn et al., 2018; Leaver et al., 2011). Although marine species have very limited capacities for omega-3 bioconversion (conversion of the shorter chain alpha-linolenic acid (ALA) to the longer chain EPA and DHA), some species like gilthead seabream and European seabass do express several of the required genes. Fatty acid elongase genes, as well as Delta-6 desaturase have been described, and are upregulated by dietary factors in the same manner as in salmonids (Seiliez et al., 2003; Tocher and Ghioni, 1999). Still, in gilthead seabream, the omega-3 fatty acid bioconversion capacity is shown to be limited due to a deficiency in Delta5 desaturase activity (Tocher and Ghioni, 1999). There are, however, other metabolic factors that can influence the omega-3 fatty acid composition in fish, such as fatty acid uptake, deposition, and beta-oxidation (Henderson and Sargent, 1985; Horn et al., 2019; Vegusdal et al., 2004). Although knowledge is limited in gilthead seabream, studies have shown a preferential retention of arachidonic acid (20:4 n-6) and DHA over EPA (Izquierdo, 1996; Kalogeropoulos et al., 1992; Koven et al., 2001).
The first publicly available combined-species SNP array for European seabass and gilthead seabream, the new Axiom® MedFish SNP array, was recently developed (Penaloza et al., 2021). This array enables the accurate high-throughput genotyping of ~ 30 K SNPs distributed throughout the seabream genome. The availability of genotype data on individuals allows the computation of genomic relationship matrix (GRM) where the coefficient of relationships between relatives are described more precisely with possibility of covering the deviations in relatedness caused by Mendelian sampling (Houston et al., 2020; Nielsen et al., 2009). Although heritability can be underestimated or overestimated in influential regions with high or low linkage disequilibrium (LD), GRM reflects more precise relatedness among individuals than the pedigree-based relationships (Da et al., 2014). The genetic variance components estimated using GRM are therefore possibly more precise than pedigree-based estimates, especially when the available pedigree is not very deep.
A key factor in the rate of genetic improvement possible for any given trait in a breeding programme is its heritability, and its genetic
correlations with other key traits in the breeding goal. Only a few studies have reported genetic parameters of fat deposition traits in gilthead seabream (Elalfy et al., 2021; García-Celdr´ an et al., 2015b; Navarro et al., 2009b), but never using high-throughput genotyping. One study has reported heritability of muscle fatty acids (Vallecillos et al., 2021), but the genetic correlations between fatty acids and other important traits have never been reported for gilthead seabream.
The aim of the current study was to estimate genomic heritability and genetic relationships of fat deposition traits and individual muscle fatty acids in gilthead seabream using the novel ~30 k MedFish SNP array.
2. Material and methods
Gilthead seabream originating from the selective breeding program of Galaxidi Marine Farm SA (Galaxidi, Greece) were used in this experiment. All fish used in this study were the offspring of 33 males and 20 females mass spawned in a single broodstock tank over one day. The fish were stocked in a sea cage at an average weight of 2.6 g. During the sea phase, the fish were fed commercial feed and kept in commercial sea cages in Galaxidi, Greece. They were slaughtered in September 2019, after 15 months at sea and fasted prior to slaughter. From the average body weight of ≈165 g to 270 g, the fish were fed a diet with a protein/ lipid content of 45/20, that only contained fish oil. From six to eight months, the fish were fed a second experimental diet of a 45/16 protein/ lipid content, that included some sunflower oil. During the last twoweek phase of the trial before slaughter, the fish returned to the first diet. Due to the logistics of sampling, slaughtering took place over several days, therefore the number of days fasted prior to slaughter varied from 4 to 12, which is considered in the statistical analysis of the data.
At the time of slaughter, body weight, viscera weight, and muscle fat measured by using Distell meter were recorded on 959 fish. Total viscera weight was measured directly after slaughter by removing and weighing the entire viscera, including liver and heart. The liver and heart were also weighed separately, so for the analyses in the current study, the viscera weight was calculated by subtracting heart weight and liver weight from total viscera weight. Visceral fat was not measured directly, but viscera percentage of body weight (viscera weight/body weight*100) was used as an indicator of amount of visceral fat in the current study (Viscera %).
2.1. Lipid measurements
Two methods of measuring muscle fat percentage were applied on the fish studied, the non-invasive Distell fat meter (Distell (Model-FM 692, www.distell.com), and the gold standard laboratory Folch method (MFATF). Directly after slaughter, a non-invasive fat measurement was made on each of the 959 fish using the Distell fat meter, a handheld microwave dielectric spectrometer. Two measurements were recorded above the lateral line and two below, repeated on both sides of the fish. The average of these eight measurements was used as a measure of the total muscle fat percentage (MFATD). A calibration equation for muscle fat in sea bream using the internal “custom calibration” setting was generated as per the manufacturer's specifications (Distell, 2011). The resulting calibration equation on 20 fish had a coefficient of determination of R2 = 0.79. The average of the eight measurements per fish after use of the custom calibration was used as measure of the total muscle fat percentage (MFATD).
The whole skinless fillet of 240 of the 959 fish were individually homogenized, and total lipids were extracted using the Folch extraction method (Folch et al., 1957) to record total muscle fat percentage (MFATF). The fatty acid composition of the total lipids was determined using the Mason & Waller method by means of methyl ester gas chromatography separation and flame ionization detection (Mason and Waller, 1964). This gave fatty acid composition estimates for both
S.S. Horn et al.
proportional (% of total fatty acids) and quantitative (mg per g fillet, based on internal standard) content of each fatty acid. For the data analysis, outliers and error datapoints were removed through the Interquartile Range (IQR) method. The results of the current study will focus on the five most prevalent fatty acids of the muscle (18:1n-9, 16:0, 18:2n-6, 22:6n-3 (DHA) and 16:1n-7), as well as the omega-3 fatty acids 20:5n-3 (EPA) and 18:3n-3 (ALA).
2.2. DNA extraction and genotyping
DNA was extracted from fin clips by IdentiGEN (Dublin, Ireland), using a crude DNA isolation method consisting of PK/Chelex extraction followed by a dilution step. The fish were genotyped using the MedFish array, which contains approximately 29,800 validated SNPs for gilthead seabream (Penaloza et al., 2021).
Genotype data were filtered using Plink software (Purcell et al., 2007), excluding SNPs with minor allele frequency (MAF) lower than 2%, missing call rates exceeding 10%, and variants which had HardyWeinberg equilibrium exact test p-value below 1e-15. In total 25,919 SNPs passed filters and quality control.
2.3. Statistical analysis
The genomic relationship matrix was generated with the GCTA software, following the method by Yang et al. (2011), using the following equation to estimate the genetic relationship between individuals j and k:
Gjk = 1 N
N i=1 ( xij 2pi )(xik 2pi ) 2pi (1 pi ) ,
where xij is the number of copies of the reference allele for the ith SNP of the jth individual, xik is the number of copies of the reference allele for the ith SNP of the kth individual, and pi is the frequency of the reference allele, estimated from the observed allele frequencies.
Variance and covariance components were estimated by residual maximum likelihood procedures using the CGTA program. Bivariate analyses were performed to estimate genetic correlations between traits, using the following bivariate animal model (Henderson, 1984):
Y = XB + U + E
where Y is a matrix of phenotypic records for individuals i = 1, 2, , n and traits j = 1, 2, X is a matrix of the fixed effects of animal i on trait j, B is a matrix of fxed efect solutions. U is a matrix containing the random effects of animal i on trait j, with variance with variance G0⊗ G, where G is the genomic relationship matrix between individuals and G0 is a genetic variance–covariance matrix among traits. E is a matrix of residual effects, that is assumed to have a variance of
R = [
2 e1 σ e1,e2 σ e1,e2 σ 2 e2 ], where 1 and 2 indicate traits.
Univariate analyses were performed to estimate heritability for all traits. For the univariate analyses, matrices Y, B, U and E were reduced to vectors and matrices G and R were reduced to scalars. Heritability (narrow sense) was estimated as the ratio of additive genetic variance to total phenotypic variance. Slaughter day (representing number of days fasted) was included as a fixed effect for all traits.
Genetic coefficient of variation was determined based on the method suggested by Burton and Devane (1953) as follows:
GCV = σ2 √ g μ *100,
Where μ is the mean of the trait, and σ 2g is the genetic variance.
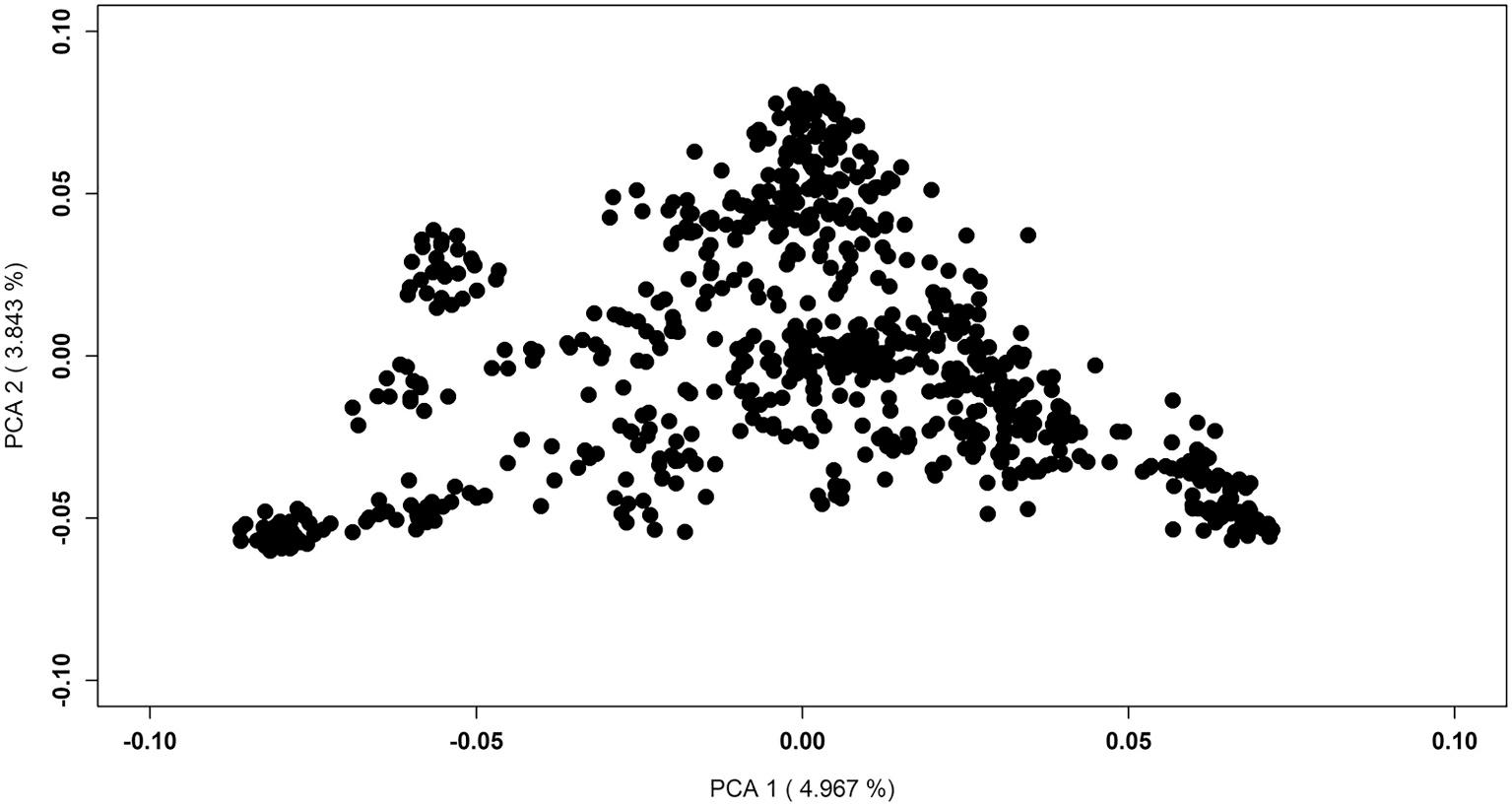
As parent fish were not genotyped, it was not possible to perform parentage assignment and develop pedigree, therefore, principal component analysis (PCA) was performed to evaluate population
structure and get an idea on number of clusters possibly representing full and/or half-sib families. The PCA based cluster analysis showed that the population is quite homogenous (little to no cluster differentiation), with relatively low variation explained by first two PCAs, 4.9% and 3.8% variation explained by PCA1 and PCA2, respectively (Fig. 1). The cluster analysis showed ~42 clusters with number of individuals ranging from 2 to 53 individuals per cluster with a mean cluster size of 23 individuals.
3. Results and discussion
The body size and muscle fat level were within the normal range for commercially produced harvest sized gilthead seabream, with a mean body weight of 372 g, ranging from 124 to 546 g (Table 1). The coefficient of variation for all production traits surpassed 0.15, showing that there was substantial individual variation in these traits.
3.1. Genetic parameters of lipid related production traits
The heritability of muscle fat, viscera weight and viscera % were all above 0.3, showing that there is a substantial genetic component to these traits (see Table 2). Viscera % had an especially high heritability estimate of 0.46, which was almost equal to the estimated heritability of viscera weight with body weight included as covariate in the statistical model (0.45 ± 0.06, result not shown). This result is in line with the previously reported heritability of 0.5 for visceral fat in gilthead seabream by Navarro et al. (2009b), although García-Celdr´ an et al. (2015a) reported a heritability of 0.2 for visceral fat with body weight as a covariate in the model. In the current study, the observed variation in viscera weight was assumed to mainly be due to the size of the fish and the amount of lipids stored here; hence the viscera weight expressed as % of body weight was used as an indicator of amount of visceral fat. The genetic correlation between body weight and viscera weight was 0.72, supporting that there is some genetic variation in viscera weight that is independent of the weight of the fish, which could be variation in distribution of lipid deposition.
Previously reported heritability of muscle fat ranges from 0.05 to 0.31 in gilthead seabream (Elalfy et al., 2021; García-Celdr´ an et al., 2015a; Navarro et al., 2009b; Vallecillos et al., 2021). In the current study, both methods of recording muscle fat showed heritability slightly higher than previous estimates: MFATD had a heritability of 0.46, and MFATF of 0.34. The considerably smaller dataset is reflected in the larger standard error for MFATF (Table 2). The differences in heritability estimates between studies could be partly related to the accuracy of the recording technique. Moreover, there are differences in the statistical models used. García-Celdran et al. (2015a) and Vallecillos et al. (2021) included body weight as a covariate. In the current study, body weight was not included as a covariate in the statistical model, but when tested, body weight as covariate did not have a significant effect on heritability estimates of the muscle fat traits (results not shown). This is due to that the genetic correlation between body weight and muscle fat was very low (rg = 0.12 for MFATD and rg = 0.13 for MFATF), which was in accordance with the low phenotypic correlations observed (Fig. 2 & Table 3). A low genetic correlation between body weight and muscle fat is in agreement with the estimate reported by Navarro et al. (2009a) (rg = 0.12). However, estimates reported by García-Celdr´ an et al. (2015b) and Vallecillos et al. (2021) are higher (0.29 and 0.59, respectively). This could explain the differences in statistical models and heritability estimates. Furthermore, these differences in estimates of heritability and genetic correlations may be due to genetic differences among the studied populations, as well as genotype-by-environment interactions (Gulzari et al., 2022; Kause et al., 2002).
The low genetic correlation between body weight and muscle fat observed in the current study of gilthead seabream is contrary to findings of salmonid species such as Atlantic salmon where muscle fat percentage is highly genetically correlated with body weight, i.e. rg =
Table 1
Descriptive statistics for production traits.
SD = standard deviation. CV = coefficient of variation. Viscera % = (viscera weight/body weight), MFATD = muscle fat % by Distell meter, MFATF = muscle fat % by Folch chemical analysis.
Table 2
Heritability (h2) of production traits.
Trait N Va Ve h2 GCV %
Body weight 967 1435 (271) 2842 (187) 0.34 (0.05) 10
Viscera weight 911 14.87 (2.73) 23.31 (1.7) 0.39 (0.06) 17
Viscera % 911 0.58 (0.09) 0.67 (0.05) 0.46 (0.05) 12 MFATD 960 1.03 (0.16) 1.21 (0.09) 0.46 (0.05) 13 MFATF 238 1.13 (0.46) 2.2 (0.38) 0.34 (0.12) 11
Standard errors in brackets. Va: Additive genetic variance. Ve: Residual variance. GCV = Genetic coefficient of variation. Viscera % = (viscera weight/body weight), MFATD = muscle fat % by Distell meter, MFATF = muscle fat % by Folch chemical analysis.
0.45–0.84 (Powell et al., 2008; Tsai et al., 2015; Vieira et al., 2007). There are similar findings in the salmonids Rainbow trout (rg = 0.30–0.52) (Blay et al., 2021) and European white fish (rg = 0.62–0.64) (Janhunen et al., 2017). This difference is likely explained by the inherent differences in the mechanisms of lipid deposition between the fish species. For example, in Atlantic salmon, the muscle is a main lipid storage site, while in gilthead seabream very little fat is stored in muscle, and the viscera is the main lipid storage site (McClelland et al., 1995). These differences in lipid storage are likely due to differences in behavioural adaptation to their natural habitats; seabream is a bottomdwelling sedentary fish species, while Atlantic salmon is an active surface feeder species, and needs energy available in their locomotory muscles (McClelland et al., 1995).

Fig. 2. Scatter plot of body weight against muscle fat, including the regression line and correlation coefficient, n = 238.
Table 3
Genetic (upper triangle) and phenotypic (bottom triangle) correlations.
There was a low genetic correlation between the two fat deposition traits MFATD and Viscera % (rg = 0.23). This agrees with Navarro et al. (2009a), who found an even lower correlation of 0.05, and used a direct measure of visceral fat - manual removing and weighing of
Standard errors in brackets.
* Only converged with no covariates in the statistical model.
** p > 0.01.
fat
Fig. 1. PCA plot of the population structure.
deposits. This shows that deposition of fat in these two lipid deposits (muscle and viscera) are genetically different traits, similar to what has been described in Rainbow trout (Kause et al., 2007).
The genetic correlation between the two methods of measuring muscle fat was high; 0.82 (Table 3), but lower than reported by a recent study on European seabass, where the genetic correlation between the same two methods on the same trait was close to unity at 0.96 ± 0.03 (Difford et al., 2021). Pearson's correlation coefficient for the two methods was 0.6, showing that the Distell meter was not highly accurate for phenotypic recording of muscle fat percentage in this dataset.
3.2. Muscle fatty acid composition
The fatty acid composition of muscle was recorded as both proportional content (% of total fatty acids) and quantitative content (mg/g tissue) on 240 fish (Table 4). The phenotypic variation in proportional content was low, while the variation in quantitative content was higher, reflecting the variation in total muscle fat content. The major fatty acids in the muscle, constituting more than 60% of muscle fatty acids, were oleic acid (18:1n-9), palmitic acid (16:0), and linoleic acid (18:2n-6). Oleic and linoleic acid are the major fatty acids from plant oils in fish feed, while palmitic acid is the most common saturated fatty acid found in all animals. The mean proportional content of EPA and DHA in the gilthead seabream muscle was 2.43% and 6.95% of total lipids, respectively. The mean quantitative content of EPA and DHA was 2.14 and 6.08 mg/g, respectively, i.e. 822 mg per 100 g fillet in total. Considering the recommendations on daily intake of omega-3 FAs for the general human public of 250–500 mg EPA + DHA (Cunnane, 2004; GOED, 2014), farmed seabream is a good source of the essential nutrients EPA and DHA.
3.3. Heritability and genetic correlations of fatty acids in gilthead seabream fillets
The estimated heritability of the proportional content of both EPA and DHA was relatively high, at 0.39 and 0.33, respectively (Table 5). These estimates are substantially higher than those recently reported by Vallecillos et al. (2021), at 0.05 for EPA and 0.11 for DHA. This difference is likely due to the different statistical models used, as Vallecillos et al. (2021) used a Bayesian model, which is quite conservative, and included body weight as a covariate in the model, which could remove some of the genetic variation in the fatty acid traits. In the current study, we chose not to include body weight as a covariate for fatty acids, as both % EPA and % DHA had low and non-significant genetic correlations
Table 4
Descriptive statistics of proportional (% of total fatty acids) and quantitative (mg/g tissue) content of the top five most prevalent fatty acids in muscle, and the omega-3 fatty acids EPA (20:5n-3) and ALA (18:3n-3).
Fatty acid
N = 240. CV = coefficient of variation. SD = standard deviation.
with body weight (Table 6). However, DHA (but not EPA) was significantly correlated with muscle fat, which is in agreement with Vallecillos et al. (2021). Thus, including muscle fat as a covariate in the model would likely reduce the heritability estimate for DHA in this data material.
One limitation of this study is the lack of pedigree, which may have biased the estimation of genetic parameters, as population stratification can inflate estimates of genetic relatedness (Dandine-Roulland et al., 2016). However, the evaluation of population structure by principal component analysis showed that the population is quite homogenous with little to no cluster differentiation (Fig. 1). Also, previous studies have shown that genomic information can more accurately reflect the relationships between individuals than pedigree information (Da et al., 2014; Wang and Da, 2014). The increased accuracy of genomic prediction compared with pedigree prediction is evident in a range of aquaculture species (reviewed in Houston et al. (2020)).
The high heritability for both EPA and DHA estimated in the current study demonstrate that selective breeding is a promising tool for increasing muscle content of these essential nutrients in the fillet of this species. This agrees with studies in Atlantic salmon that have shown significant heritability of EPA and DHA content of muscle (Leaver et al., 2011; Horn et al., 2018). Wang et al. (2019) also showed that there is a genetic component to omega-3 content of Asian seabass fillets. Heritability estimates of DHA in tilapia and common carp have been close to zero, which could be due to the very low quantities of DHA detected (Nguyen et al., 2010; Prchal et al., 2018). The estimates for both fish species indicated that EPA had a considerably higher heritability than DHA, while in Atlantic salmon, the heritability of EPA was lower than DHA (Horn et al., 2018).
Alpha-linolenic acid (18:3n-3; ALA) had the lowest heritability of all fatty acids (0.03). The ratio of DHA/ALA was included in the analysis as an indicator-trait for the conversion of ALA to DHA (omega-3 bioconversion), but is in this case likely reflecting variation in DHA due to the very low variation in ALA. This low variation, together with the low general content of ALA in the muscle of gilthead seabream, indicates that omega-3 bioconversion of ALA to DHA is not substantial, and not very important for the overall DHA content in the muscle of this species. This implies that the phenotypic and genetic variation observed in EPA and DHA levels are linked to other metabolic factors, such as deposition or beta-oxidation, rather than omega-3 bioconversion. This is in accordance with a study reporting that the omega-3 bioconversion capacity is limited in gilthead seabream due to a deficiency in Delta5 desaturase activity, catalyzing conversion of 20:4n-3 to EPA (Tocher and Ghioni, 1999).
The estimates of genetic correlations showed that the proportional content of each fatty acid was differently correlated with muscle fat (Table 6). The standard errors on the estimates were large due to the small dataset on fatty acids, but the genetic correlations agreed with the phenotypic correlations. The fatty acids with the strongest positive correlations with muscle fat were 18:1n-9 and 16:1n-7 (Table 6), both of which are products of de novo lipogenesis, synthesis of fatty acids, although 18:1n-9 is also found in the feed. This indicates that a genetic predisposition for higher muscle fat is due to a higher inherent de novo lipogenesis activity. The high heritability of % 16:1n-7, which is almost exclusively a product of de novo lipogenesis, also indicates that the rate of de novo lipogenesis in gilthead seabream muscle is a heritable trait (Table 5).
The genetic correlations indicated that the observed genetic variation in % EPA and % DHA is independent of body weight, as both % EPA and % DHA had low and non-significant genetic correlations with body weight (Table 6). The proportional content of EPA had a weak positive (non-significant) correlation with muscle fat, while DHA had a negative genetic correlation. This difference between EPA and DHA likely reflects the metabolic differences between the two fatty acids, and their roles in muscle tissue (Calder, 2006; Stillwell and Wassall, 2003). The observed relationship between DHA and muscle fat is seen in several species and
Table 5
Heritability (h2) of fatty acid traits.
% = proportional content of fatty acids, mg/g = quantitative content of fatty acids, Va: Additive genetic variance. Ve: Residual variance. Standard errors in brackets. GCV = genetic coefficient of variation. N = 240.
Table 6
Phenotypic (rP) and genetic (rG) correlations between production traits and fatty acids.
Trait
MFATF * % 18:1n-9
MFATF * % 16:1n-7
(0.20)
(0.17)
(0.20)
MFATF * 18:2n-6 0.07** 0.20 (0.29)
MFATF * 16:0 0** 0.15 (0.30)
DHA * EPA 0.42 0.55 (0.19)
DHA * 18:2n-6 0.22 0.37 (0.29)
DHA * 16:0 0.23 0.48 (0.25)
DHA * 16:1n-7 0.04** 0.05 (0.26)
DHA * 18:3n-3 0.22 nc
DHA * 18:1n-9 0.63 nc
MFATF = Muscle fat % by Folch chemical analysis, BW = body weight. SE = standard error. EPA = 20:5n-3. DHA = 22:6n-3. Nc = no convergence. ** = p > 0.01.
tissues and can be explained by DHA being a PUFA with important roles in cell membranes that is therefore found in phospholipids (PL) to a larger degree than in triglycerides (TG). When muscle fat increases, the proportion of PL relative to TG decreases, and therefore the % DHA decreases (Tocher, 2003).
3.4. Selection strategies for increased omega-3 fatty acid content of fillets
The heritability of EPA and DHA indicated that the content of healthy omega-3 fatty acids in seabream fillets can be increased through selective breeding, and there are two possible strategies to achieve this: by increasing the quantitative content (mg per g muscle), or by increasing the proportional content (% of total muscle fatty acids).
As muscle fat percentage increases, so does the quantitative content of all fatty acids. Therefore, the heritability of the quantitative content of all fatty acids mirrored the heritability of muscle fat and were all close to 0.34 (Table 5). This was confirmed in the estimates of genetic correlations, where the quantitative content of all fatty acids had a genetic correlation to muscle fat close to unity (results not shown). This implies that an increase in the quantitative content of EPA and DHA can be achieved by selective breeding for increased muscle fat content, which could be achieved by e.g. the Distell fat meter for non-invasive phenotypic recording. It should then be considered that increased muscle fat would influence the organoleptic qualities of the fish (Grigorakis, 2007). It should also be considered that selective breeding for increased muscle fat content would result in an increase in all fatty acids, including the less desirable omega-6 fatty acids. This is critical from a nutritional health perspective, as it is not only the amount of EPA and DHA that is important, but also the ratio of omega-3 to omega-6 fatty acids (Saini and Keum, 2018; Simopoulos, 2002). In addition, the genetic
correlations indicated that increasing muscle fat would actually reduce the proportional content of DHA in the muscle, leading to a less desirable fatty acid profile of the seabream fillet (Table 6). On the other hand, the genetic correlations between proportional contents of muscle fatty acids showed a potential to select for an overall healthier fatty acid profile in muscle (Table 6). DHA was positively correlated with EPA (rg = 0.55), and negatively correlated with the main saturated fatty acid (16:0) and the main omega-6 fatty acid (18:2n-6) in muscle (rg = 0.48 and 0.37, respectively). Thus, according to these results, selection for increased proportional content of DHA will result in a fish fillet with a higher proportion of marine omega-3 fatty acids, and lower proportion of saturated fatty acids and pro-inflammatory omega-6 fatty acids. However, a lower selection response may be expected using this strategy as the genetic coefficients of variation was lower for proportional compared to quantitative content of EPA and DHA (Table 5).
Even though both quantitative and proportional content of EPA and DHA are highly heritable, getting genetic response in fatty acid traits is not straight forward. With low phenotypic and genetic coefficients of variation it is difficult to get a reasonable selection response, even when heritability is high. We recommend future research into economic weights and selection indices which are outside the scope of this paper. Another challenge with implementing selection for fatty acid traits is that phenotyping requires costly and time-consuming chemical analyses, which make large-scale data recording challenging. New rapid methods for predicting fatty acid composition in fish fillet are being developed, such as Raman and near infrared spectroscopy (e.g. EWOS SalmoNIR technology from Cargill) (Afseth et al., 2006; Bekhit et al., 2014; Blay et al., 2021), which can make practical implementation of selection for these traits more feasible.
4. Conclusions
In the gilthead seabream, muscle fat deposition and body weight seem to be independent traits, and selective breeding for faster growth is not likely to influence the proportional content of EPA and DHA. The fatty acid 16:1n-7, a marker of de novo lipogenesis, had a high heritability (0.43), indicating that there is a strong genetic component to this metabolic pathway in seabream.
It is possible to increase EPA and DHA content in gilthead seabream fillets by selective breeding, as the estimated heritability of EPA (%) and DHA (%) was 0.39 and 0.33, respectively, and there was a positive genetic correlation between the two fatty acids (0.55). There is a potential to select for a healthier overall fatty acid profile in muscle of seabream by selection for increased proportional content of DHA, as DHA was negatively correlated with the major omega-6 fatty acid (18:2n-6) and the major saturated fatty acid (16:0).
CRediT authorship contribution statement
S.S. Horn: Methodology, Writing – original draft, Visualization, Writing – review & editing, Formal analysis. M.L. Aslam: Methodology,
Writing – review & editing, Formal analysis. G.F. Difford: Methodology, Writing – review & editing, Formal analysis. K. Tsakoniti: Resources, Conceptualization, Data curation. S. Karapanagiotis: Resources, Data curation, Writing – review & editing. B. Gulzari: Writing – review & editing. J.W.M. Bastiaansen: Writing – review & editing. C. Penaloza: Resources, Writing – review & editing. R. Houston: Resources, Writing – review & editing. B. Ruyter: Data curation, Conceptualization, Writing – review & editing. A.K. Sonesson: Funding acquisition, Project administration, Supervision, Conceptualization, Methodology, Writing –review & editing.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
This study was made possible by the EU project MedAID (H2020 grant agreement No 727315). Authors would like to thank Galaxidi Marine Farm S.A. for their participation in the study and its staff for providing necessary help and facilities for raising the population and the experimental data collection. Authors are grateful to Målfrid Tofteberg Bjerke and Anne Marie Langseter for skilful technical assistance.
References
Afseth, N.K., Wold, J.P., Segtnan, V.H., 2006. The potential of Raman spectroscopy for characterisation of the fatty acid unsaturation of salmon. Anal. Chim. Acta 572 (1), 85–92
Ballester-Lozano, G.F., Benedito-Palos, L., Navarro, J.C., Kaushik, S., P´ erez-S´ anchez, J., 2011. Prediction of fillet fatty acid composition of market-size Gilthead Sea bream (Sparus aurata) using a regression modelling approach. Aquaculture 319, 81–88
Bekhit, M.Y., Grung, B., Mjos, S.A., 2014. Determination of omega-3 fatty acids in fish oil supplements using vibrational spectroscopy and chemometric methods. Appl. Spectrosc. 68 (10), 1190–1200
Blay, C., Haffray, P., Bugeon, J., D’Ambrosio, J., Dechamp, N., Collewet, G., Enez, F., Petit, V., Cousin, X., Corraze, G., et al., 2021. Genetic parameters and genome-wide association studies of quality traits characterised using imaging technologies in rainbow trout, Oncorhynchus mykiss Front. Genet. 12 (219)
Brown, R.C., 2004. Genetic Management and Selective Breeding in Farmed Populations of Gilthead Seabream (Sparus aurata)
Burton, G.W., Devane, D.E., 1953. Estimating heritability in tall fescue (Festuca arundinacea) from replicated clonal material 1. Agron. J. 45 (10), 478–481 Calder, P.C., 2006. n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 83 (6), 1505S–1519S Calder, P.C., 2015. Marine omega-3 fatty acids and inflammatory processes: effects, mechanisms and clinical relevance. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 1851 (4), 469–484
Cunnane, S., 2004. Recommedations for intakes of polyunsaturated fatty acids in healthy adults. ISSFAL Newslett. 11, 12–25
Da, Y., Wang, C., Wang, S., Hu, G., 2014. Mixed model methods for genomic prediction and variance component estimation of additive and dominance effects using SNP markers. PLoS One 9 (1), e87666
Dandine-Roulland, C., Bellenguez, C., Debette, S., Amouyel, P., G´ enin, E., Perdry, H., 2016. Accuracy of heritability estimations in presence of hidden population stratification. Sci. Rep. 6 (1), 26471
Difford, G.F., Díaz-Gil, C., S´ anchez-Moya, A., Aslam, M.L., Horn, S.S., Ruyter, B., Sonesson, A.K., 2021. Genomic and phenotypic agreement defines the use of microwave dielectric spectroscopy for recording muscle lipid content in European seabass (Dicentrarchus labrax). Front. Genet. 12 (1594). Distell, 2011. Technical Manual Distell Fish FatMeter. Available online at: http://www. distell.com (accessed Febuary 21, 2022).
Dong, L., Xiao, S., Wang, Q., Wang, Z., 2016. Comparative analysis of the GBLUP, emBayesB, and GWAS algorithms to predict genetic values in large yellow croaker (Larimichthys crocea). BMC Genomics 17 (1), 460 Elalfy, I., Shin, H.S., Negrín-Baez, D., Navarro, A., Zamorano, M.J., Manchado, M., Afonso, J.M., 2021. Genetic parameters for quality traits by non-invasive methods and their G x E interactions in ocean cages and estuaries on gilthead seabream (Sparus aurata). Aquaculture 537, 736462
FEAP, 2020. European Aquaculture Production Report 2014–2019: Federation of European Aqauculture Producers Folch, J., Lees, M., Stanley, G.H.S., 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 226 (1), 497–509
Fontana, L., Eagon, J.C., Trujillo, M.E., Scherer, P.E., Klein, S., 2007. Visceral fat adipokine secretion is associated with systemic inflammation in obese humans. Diabetes 56 (4), 1010
García-Celdran, M., Ramis, G., Manchado, M., Estevez, A., Afonso, J.M., Armero, E., 2015a. Estimates of heritabilities and genetic correlations of carcass quality traits in a reared gilthead sea bream (Sparus aurata L.) population sourced from three broodstocks along the Spanish coasts. Aquaculture 446, 175–180
García-Celdr´ an, M., Ramis, G., Manchado, M., Est´ evez, A., Navarro, A., Armero, E., 2015b. Estimates of heritabilities and genetic correlations of raw flesh quality traits in a reared gilthead sea bream (Sparus aurata L.) population sourced from broodstocks along the Spanish coasts. Aquaculture 446, 181–186
GOED, 2014. Global Recommendations for EPA and DHA Intake (Rev. 19 November 2014)
Grigorakis, K., 2007. Compositional and organoleptic quality of farmed and wild gilthead sea bream (Sparus aurata) and sea bass (Dicentrarchus labrax) and factors affecting it: a review. Aquaculture 272 (1), 55–75
Grigorakis, K., Alexis, M.N., Taylor, K.D.A., Hole, M., 2002. Comparison of wild and cultured gilthead sea bream (Sparus aurata); composition, appearance and seasonal variations. Int. J. Food Sci. Technol. 37 (5), 477–484
Grigorakis, K., Taylor, K.D.A., Alexis, M.N., 2003. Organoleptic and volatile aroma compounds comparison of wild and cultured gilthead sea bream (Sparus aurata): sensory differences and possible chemical basis. Aquaculture 225 (1), 109–119
Gulzari, B., Komen, H., Nammula, V.R., Bastiaansen, J.W.M., 2022. Genetic parameters and genotype by environment interaction for production traits and organ weights of gilthead seabream (Sparus aurata) reared in sea cages. Aquaculture 548, 737555
Henderson, C.R., 1984. Applications of Linear Models in Animal Breeding. Univerity of Guelph, Guelph
Henderson, R.J., Sargent, J.R., 1985. Chain-length specificities of mitochondrial and peroxisomal beta-oxidation of fatty-acids in livers of rainbow-trout (SalmoGairdneri). Comp. Biochem. Physiol. B-Biochem. Mol. Biol. 82 (1), 79–85
Horn, S.S., Ruyter, B., Meuwissen, T.H.E., Hillestad, B., Sonesson, A.K., 2018. Genetic effects of fatty acid composition in muscle of Atlantic salmon. Genet. Sel. Evol. 50
Horn, S.S., Sonesson, A.K., Krasnov, A., Moghadam, H., Hillestad, B., Meuwissen, T.H.E., Ruyter, B., 2019. Individual differences in EPA and DHA content of Atlantic salmon are associated with gene expression of key metabolic processes. Sci. Rep. 9 (1) (3889–3889)
Houston, R.D., Bean, T.P., Macqueen, D.J., Gundappa, M.K., Jin, Y.H., Jenkins, T.L., Selly, S.L.C., Martin, S.A.M., Stevens, J.R., Santos, E.M., et al., 2020. Harnessing genomics to fast-track genetic improvement in aquaculture. Nat. Rev. Genet. 21 (7), 389–409
Ibarz, A., Blasco, J., Beltr´ an, M., Gallardo, M.A., S´ anchez, J., Sala, R., Fern´ andezBorras, J., 2005. Cold-induced alterations on proximate composition and fatty acid profiles of several tissues in gilthead sea bream (Sparus aurata). Aquaculture 249 (1), 477–486
Izquierdo, M., 1996. Essential fatty acid requirements of cultured marine fish larvae. Aquac. Nutr. 2 (4), 183–191
Izquierdo, M.S., Obach, A., Arantzamendi, L., Montero, D., Robaina, L., Rosenlund, G., 2003. Dietary lipid sources for seabream and seabass: growth performance, tissue composition and flesh quality. Aquac. Nutr. 9 (6), 397–407
Janhunen, M., Nousiainen, A., Koskinen, H., Vehvilainen, H., Kause, A., 2017. Selection strategies for controlling muscle lipid content recorded with a non-destructive method in European whitefish, Coregonus lavaretus. Aquaculture 481, 229–238
Janssen, K., Chavanne, H., Berentsen, P., Komen, H., 2017. Impact of selective breeding on European aquaculture. Aquaculture 472, 8–16
Kalogeropoulos, N., Alexis, M.N., Henderson, R.J., 1992. Effects of dietary soybean and cod-liver oil levels on growth and body composition of gilthead bream (Sparus aurata). Aquaculture 104 (3–4), 293–308
Kause, A., Ritola, O., Paananen, T., Mantysaari, E., Eskelinen, U., 2002. Coupling body weight and its composition: a quantitative genetic analysis in rainbow trout. Aquaculture 211 (1–4), 65–79
Kause, A., Tobin, D., Mantysaari, E.A., Martin, S.A.M., Houlihan, D.F., Kiessling, A., Ruohonen, K., 2007. Genetic potential for simultaneous selection of growth and body composition in rainbow trout (Oncorhynchus mykiss) depends on the dietary protein and lipid content: phenotypic and genetic correlations on two diets. Aquaculture 271 (1), 162–172
Koven, W., Barr, Y., Lutzky, S., Ben-Atia, I., Weiss, R., Harel, M., Tandler, A., 2001. The effect of dietary arachidonic acid (20: 4n 6) on growth, survival and resistance to handling stress in gilthead seabream (Sparus aurata) larvae. Aquaculture 193 (1–2), 107–122
Leaver, M.J., Taggart, J.B., Villeneuve, L., Bron, J.E., Guy, D.R., Bishop, S.C., Tocher, D. R., 2011. Heritability and mechanisms of n-3 long chain polyunsaturated fatty acid deposition in the flesh of Atlantic salmon. Comp. Biochem. Physiol. D-Genomics Proteomics 6 (1), 62–69
Mason, M.E., Waller, G.R., 1964. Dimethoxypropane induced transesterification of fats + oils in preparation of methyl esters for gas chromatographic analysis. Anal. Chem. 36 (3), 583
McClelland, G., Weber, J.-M., Zwingelstein, G., Brichon, G., 1995. Lipid composition off tissue and plasma in two mediterranean fishes, the gilt-head sea bream (Chrysophyrys auratus) and the European seabass (Dicentratchus labrx). Can. J. Fish. Aquat. Sci. 52 (1), 161–170
Mnari, A., Bouhlel, I., Chraief, I., Hammami, M., Romdhane, M.S., El Cafsi, M., Chaouch, A., 2007. Fatty acids in muscles and liver of Tunisian wild and farmed gilthead sea bream, Sparus aurata Food Chem. 100 (4), 1393–1397
Navarro, A., Zamorano, M.J., Hildebrandt, S., Gin´ es, R., Aguilera, C., Afonso, J.M., 2009a. Estimates of heritabilities and genetic correlations for body composition
S.S. Horn
traits and G× E interactions, in gilthead seabream (Sparus auratus L.). Aquaculture 295 (3–4), 183–187
Navarro, A., Zamorano, M.J., Hildebrandt, S., Gines, R., Aguilera, C., Afonso, J.M., 2009b. Estimates of heritabilities and genetic correlations for growth and carcass traits in gilthead seabream (Sparus auratus L.), under industrial conditions. Aquaculture 289 (3), 225–230
Nguyen, N.H., Ponzoni, R.W., Yee, H.Y., Abu-Bakar, K.R., Hamzah, A., Khaw, H.L., 2010. Quantitative genetic basis of fatty acid composition in the GIFT strain of Nile tilapia (Oreochromis niloticus) selected for high growth. Aquaculture 309 (1–4), 66–74
Nielsen, H.M., Sonesson, A.K., Yazdi, H., Meuwissen, T.H.E., 2009. Comparison of accuracy of genome-wide and BLUP breeding value estimates in sib based aquaculture breeding schemes. Aquaculture 289 (3), 259–264
Penaloza, C., Manousaki, T., Franch, R., Tsakogiannis, A., Sonesson, A.K., Aslam, M.L., Tsigenopoulos, C.S., 2021. Development and testing of a combined species SNP array for the European seabass (Dicentrarchus labrax) and gilthead seabream (Sparus aurata). Genomics 113 (4), 2096–2107
Powell, J., White, I., Guy, D., Brotherstone, S., 2008. Genetic parameters of production traits in Atlantic salmon (Salmo salar). Aquaculture 274 (2), 225–231
Prchal, M., Vandeputte, M., Gela, D., Dolezal, M., Blichtova, H., Rodina, M., Kocour, M., 2018. Estimation of genetic parameters of fatty acids composition in flesh of market size common carp (Cyprinus carpio L.) and their relation to performance traits revealed that selective breeding can indirectly affect flesh quality. Czech J. Anim. Sci. 63 (7), 280–291
Purcell, S., Neale, B., Todd-Brown, K., Thomas, L., Ferreira, M.A., Bender, D., Daly, M.J., 2007. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 81 (3), 559–575
Saini, R.K., Keum, Y.-S., 2018. Omega-3 and omega-6 polyunsaturated fatty acids: dietary sources, metabolism, and significance — a review. Life Sci. 203, 255–267
Seiliez, I., Panserat, S., Corraze, G., Kaushik, S., Bergot, P., 2003. Cloning and nutritional regulation of a Δ6-desaturase-like enzyme in the marine teleost gilthead seabream (Sparus aurata). Comp. Biochem. Physiol. B: Biochem. Mol. Biol. 135 (3), 449–460
Simopoulos, A.P., 2002. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 56 (8), 365–379
Stillwell, W., Wassall, S.R., 2003. Docosahexaenoic acid: membrane properties of a unique fatty acid. Chem. Phys. Lipids 126 (1), 1–27
Thorland, I., Papaioannou, N., Kottaras, L., Refstie, T., Papasolomontos, S., Rye, M., 2007. Family based selection for production traits in gilthead seabream (Sparus aurata) and European sea bass (Dicentrarchus labrax) in Greece. Aquaculture 272, S314
Thota, R.N., Ferguson, J.J.A., Abbott, K.A., Dias, C.B., Garg, M.L., 2018. Science behind the cardio-metabolic benefits of omega-3 polyunsaturated fatty acids: biochemical effects vs. clinical outcomes. Food Funct. 9 (7), 3576–3596
Tocher, D.R., 2003. Metabolism and functions of lipids and fatty acids in teleost fish. Rev. Fish. Sci. 11 (2), 107–184
Tocher, D.R., Ghioni, C., 1999. Fatty acid metabolism in marine fish: low activity of fatty acyl Δ5 desaturation in gilthead sea bream (Sparus aurata) cells. Lipids 34 (5), 433–440
Todorcevic, M., Hodson, L., 2015. The effect of marine derived n-3 fatty acids on adipose tissue metabolism and function. J. Clin. Med. 5 (1).
Todorcevi´ c, M., Skugor, S., Krasnov, A., Ruyter, B., 2010. Gene expression profiles in Atlantic salmon adipose-derived Stromo-vascular fraction during differentiation into adipocytes. BMC Genomics 11 (1), 39
Tsai, H.Y., Hamilton, A., Guy, D.R., Tinch, A.E., Bishop, S.C., Houston, R.D., 2015. The genetic architecture of growth and fillet traits in farmed Atlantic salmon (Salmo salar). BMC Genet. 16 (1), 1–11
Vallecillos, A., Marín, M., Bortoletti, M., Lopez, J., Afonso, J.M., Ramis, G., Armero, E., 2021. Genetic analysis of the fatty acid profile in gilthead seabream (Sparus aurata L.). Animals 11 (10)
Vegusdal, A., Ostbye, T.K., Tran, T.N., Gjoen, T., Ruyter, B., 2004. beta-oxidation, esterification, and secretion of radiolabeled fatty acids in cultivated Atlantic salmon skeletal muscle cells. Lipids 39 (7), 649–658
Vieira, V.L., Norris, A., Johnston, I.A., 2007. Heritability of fibre number and size parameters and their genetic relationship to flesh quality traits in Atlantic salmon (Salmo salar L.). Aquaculture 272, S100–S109
Wang, C., Da, Y., 2014. Quantitative genetics model as the unifying model for defining genomic relationship and inbreeding coefficient. PLoS One 9 (12), e114484
Wang, L., Chua, E., Sun, F., Wan, Z.Y., Ye, B., Pang, H., Yue, G.H., 2019. Mapping and validating QTL for fatty acid compositions and growth traits in Asian seabass. Mar. Biotechnol. 21 (5), 643–654
Yang, J., Lee, S.H., Goddard, M.E., Visscher, P.M., 2011. GCTA: a tool for genome-wide complex trait analysis. Am. J. Hum. Genet. 88 (1), 76–82
S.S. Horn et al.
Other documents randomly have different content
this change, it may be asked, less hazardous than one which purposes only to guard against the corruption, and to correct those abuses which have almost imperceptibly crept into the representation?—As the existence of corruption in the representative is too probable, so is its prevalence in the constituent body too notorious:—election by ballot, it has been supposed, will provide a remedy against this evil;—it will remain unknown to the candidates for whom the voters may poll, the application of bribery therefore will be ineffectual, as, notwithstanding his acceptance of a bribe, the voter may still poll, according to his inclination or his conscience, without the fear of a discovery.
In regard to the universal suffrage, it must be acknowledged that every individual, in the state, has an interest in the proper administration of its affairs, and that Government will, sometimes, need the support, even of its meanest subjects; if, therefore, they are interested in, and contribute to the support of Government, they are entitled to a participation in its privileges;—the privilege they claim is the Elective Franchise, and as the lower house is called the Commons House of Parliament, why should not the members, instead of representing property, be in fact the representatives of the People?—At the same time such material changes adopted on a sudden, might produce tumult and disorder, they might occasion convulsions, attended with far more dreadful effects, than the evils they purpose to remedy;—but when a minister acknowledges corruption in his own conduct;—when the sale of Boroughs is notoriously practised;—when, in some instances, the representatives of a County are returned by the influence alone of a few powerful individuals;—when the late convictions, in the House of Commons, for bribery, prove that it is frequently, if not generally practised by the candidates for seats in that house, and that it has a pernicious influence upon the constituent body;—it cannot be disputed, but that some remedy ought instantly to be applied, in order to eradicate, or at least to check the wide spreading infection.
Lord Grey, Lord Grenville, the Whig party, even the members of the opposition are collectively abused by this Author; Lord Grey and Lord Grenville, however, together with many other individuals whom
he severely censures, are men of acknowledged talents and information, as is evident from their general conduct, and the speeches they have delivered in various Parliamentary debates; but they differ in opinion from his Reverence, if however, every man, who thinks differently from the learned Author, be a profligate or a blockhead, we suspect that, besides himself, he will scarcely find a wise or a virtuous individual throughout the great Empire.
He again resumes his attacks upon the Catholics, we shall, however, in the first instance, notice three questions, one particle of which he so confidently defies them to refute:
1st. “Does not a Papist kneel down before, pray and bow to images, pictures, and pieces of old wood representing our Blessed Saviour, the Virgin Mary, and many Saints, and does he not do this for the purpose either of paying adoration to these identical pieces of wood, old sheets of oil cloth, with faces smeared on them, almost as hideous as most of those you will see at the Catholic Board, on reading one of Robert Peele’s Anti-catholic speeches, or to their likenesses?”—In answer to this question we beg leave to refer to the catechisms of the catholics, in which occur the following questions and answers:—Is it lawful to adore the blessed Virgin or the saints? —No: for by adoration is here meant the honor due to God alone.— Do the commandments forbid us to honor the Saints?—No: for the honor we give them is different from that which we pay to God, we honor the Saints as friends and faithful servants to God.—If it is forbid in Catholics to worship the Saints, can it be supposed that they will pay adoration to their image, pictures or representations?— Or, is it consistent with probability that the same Church will direct them, in their riper years, to practise, what it has taught them when pupils, to reject and avoid?—The council of Trent has, without doubt, declared, that due honor and respect be given to images of Christ, the Virgin Mary, and the other Saints;—and who can deny that that degree of honor and respect, to which they are entitled, should be paid these, or any other representations?—It is natural to be affected at the Representation of a dear departed friend and benefactor; the Catholics look upon them as their best of benefactors, they therefore pay them a due degree of honor and respect, should they, however,
exceed this degree, they are not only culpable, but obnoxious likewise, to the censures of their own Church:—the Protestants themselves regard King William III. as their benefactor, of course they honor and drink to his memory, some even have been so far inflamed with enthusiasm, as to drink to his glorious and immortal memory on their bare knees, are not these Protestants equally idolatrous with the Catholics?
Secondly, “He wishes to know whether a Papist does not pray to Saints and Angels, and invoke their intercession, thereby making Gods, not only of Angels, but even of dead men, although expressly informed, by God himself, that there is but one mediator with the Father, not only of redemption but of intercession also, which is our blessed Saviour, and in doing so is he not guilty of idolatry?” That the Roman Catholics intreat of Saints and Angels to forward petitions in their behalf, for obtaining the divine mercy, is acknowledged, but that, in doing so, they mean to worship them as Gods, or to incur the guilt of idolatry, is as confidently denied:—they dread through an humble confidence of their own demerits, to offer from themselves an immediate address to the Deity:—and as Christ himself has given an example of praying for others, even his persecutors, in those memorable words, “Pardon them O Lord, for they know not what they do;” so the Catholics may probably imagine that the Saints and Angels whom they address, will petition in their behalf, and as they are pure in comparison with themselves, these petitions will have greater effect, than the immediate addresses of a polluted sinner— we think also, that the answer to the first, is a sufficient answer to this question.
Thirdly, “He is induced to enquire whether a Papist does not pay divine adoration to a mixture of flour and water, made up by the hands of an illiterate and possibly profligate priest, contrary to the figurative sense and meaning of the Holy Scripture, and in direct opposition to reason and common understanding? does he not believe that a Popish Priest has the power of making a God? and does he not bow down and worship this worse than golden image?”—It may be asked the Revd. Baronet if he ever enquired before administering the holy Communion to his own flock, whether
the bread provided for the purpose, were made by pure and undefiled hands, whether the wine were genuine, or brewed by some rascally vintner;—and is not the term profligate, equally applicable to a minister of the establishment, as to a Popish Priest, particularly, if the minister devotes much of his time to the joys of the chase, and (we may naturally suppose) its consequent festivity; whilst the Priest is laboriously and almost constantly employed in the conscientious discharge of his sacred functions? as to the adoration paid to the Host, the Catholic believes that the Body and Blood of Christ (acknowledged to partake of the Godhead) are actually present, and can he justly be called idolatrous for paying adoration to the presence of Divinity?—the learned gentleman may also be requested to explain what he means to impress upon the mind of a Catechumen, by the following words: “the Body and Blood of Christ are verily and indeed taken and received by the faithful in the Lord’s Supper.”—Let it be understood, that we by no means, recommend these doctrines, they may possibly give occasion of offence to such tender consciences as the Baronet possesses, they are matters of belief, and therefore left to every man’s own determination;—since, however, some particles of his questions have been absolutely and with truth denied, whilst others have been so far answered, we hope, as to exculpate the Catholics from all criminality either in act or intention, we may conclude, that should a Cardinal’s hat be now engaged to the Revd. Baronet, he will perhaps, not only be almost, but altogether inclined to renounce his own faith, and turn Papist.
In treating of the monstrous doctrines ascribed by Sir Harcourt Lees to the Catholics, we may premise that their own general conduct sufficiently refutes his accusation:—his strong assertions however, may seem to demand some farther enquiry.
The 4th Lateran council was held in the year 1215, at which were present, most of the christian sovereigns;—this council therefore may be properly termed a general congress of the temporal, as well as spiritual Powers of Christendom; they assembled for the purpose of suppressing the heresy of the Manchæans, or Albigenses, whose doctrines were (according to Mosheim) not only subversive of morality, decency, and good order, but even destructive to the
human species,—it was supported by the Counts of Thoulouse, Cominges, Foix, and aided by numerous bodies of banditti, hired for this purpose.—The heresy was condemned by the spiritual authority of the church; and the Fiefs of the princes encouraging it, were declared forfeitures to their liege lords, by the authority of the sovereigns, there assembled:—the censures of this council or rather congress were never promulgated, and scarcely known in this island, but were directed (we believe, solely) against the Manchæans or Albigenses, and the princes above mentioned, who encouraged and protected the votaries of this pernicious heresy
The council of Constance, held in the year 1414, expressly declares that it is heretical to affirm it lawful for a subject to kill his prince, on any pretence whatsoever, session 15. One solitary instance of the contrary doctrine being maintained, occurs in Mariana, whose book was condemned, and publicly burnt by a provincial council of her own order; this Mariana was a Spaniard, born at Talavera, in the year 1537, who became a Religious in the year 1554;—he was also condemned by the parliament of Paris, and by the doctors of Sorbonne, and his book burnt by the hands of the common hangman.
The council of Trent assembled in the year 1545, and continued to the year 1563, declares, that to violate the least point of public faith given to heretics, is a crime punishable by the laws of God and Man; session, 15 and 18. The doctrine then of the Catholics teaches them that no power on earth can absolve them from allegiance to their sovereigns and civil magistrates, and obliges them to keep faith with all men:—when therefore, the Revd. Author asserts, that according to the 4th Lateran council, oaths taken contrary to the interest of the Popish church are not to be called oaths, but perjuries, and that it is still in force, we must question the accuracy of his information, in regard to its being a received doctrine among the catholics;—the council of Constance, and subsequently the council of Trent decreed the very reverse, and the doctrine of the last general council must now be like the prevailing one:—the oaths taken by the priests and Bishops regard only spirituals, that part which refers to the Rights, Honors, State and Power of the Pope is confined (as the Catholics
assert) to such as live under the Pope’s temporal jurisdiction, or the jurisdiction of a Catholic Sovereign;—Our own sovereigns are bound (the author proceeds) by the coronation oath “to the utmost of their power to maintain the true profession of the gospel,” but by gospel, we are enjoined to do unto all men as we would they should do unto us; persecution therefore, in the slightest degree, is a departure from this precept,—“and the Protestant reformed religion as established by the laws”—whether will the Protestant Religion be better maintained by an observance of, or a deviation from, the doctrine contained in the Gospel? “to preserve the Bishops and Clergy of this Realm, and to the churches committed to their charge, all such rights and privileges as by law, do or shall appertain to them, or any of them,” Catholic Emancipation will not encroach upon these rights and privileges, it will only procure an equal participation of civil rights and privileges for the Catholics, which in compliance with the Gospel, the Protestants are bound to grant, as they in a like situation, would wish to obtain the same privileges for themselves.
As Sir Harcourt Lees is so very liberal in his application of censure upon individuals, upon Parties, upon Sects, who may differ from himself in opinion, with respect to the three natural points which concern human Life, Religion, Morals and Politics, we presume that he bears in mind and accords with the assertion of Demosthenes,[1] “that all men are, by nature, prone to delight in detraction and invective,” but that, through the multiplicity of his studious pursuits, the remainder of the sentence, in which this assertion is contained, has entirely escaped his recollection, “but to be disgusted with those who praise themselves,” as he is no less liberal in self Commendation;—The Athenian Orator indeed recounts to his Audience the services he had rendered the State, during his Administration, but handsomely, apologizes for this conduct, by premissing that he was obliged to pursue this method, in order to refute the Calumnies of his Adversary, and that therefore the odium, naturally attendant upon self Praise, ought to rest upon the Individual, who had compelled him to bring forward such a Relation: Sir Harcourt Lees, however had no adversary to contend with; no one disputed his merits; of course he had no Calumnies to refute; we
must conclude therefore, that he expects some material Reward for his present and former Exertions, and therefore points out his peculiar claims lest they should remain unknown and extinguished; his success we wish not to prevent; our aim is to guard the Public (since men are naturally inclined to delight in censure) from imbibing prejudice through the agreeable medium of invective.
To give advice, without giving offence, is a serious difficulty;—it has been pertinently observed, that to give advice is to seize an occasion of displaying our own wisdom, at the expence of others, and however readily a superiority in all other respects, may be admitted, the assumption of superior sense and understanding is universally offensive; the learned Baronet has made a pompous display of his diligence, his research, his consequent information, and urges his claim to attention, from the mortifying circumstance of our inferiority in understanding, when compared with his own:— making no pretensions to superiority of intellect, claiming only the free exercise of reason and common sense, and suspecting that some ingredients in the Rev. Baronet’s prescription (should it be adopted) might prove injurious to the general system; We are anxious to preserve the constitution sound and vigorous, and being members of the public, we look upon ourselves as included in the number of patients, we shall therefore take the liberty, without further ceremony, of examining (as far as we can understand it) into the composition of his Antidote.
The Revd. Author thus expresses himself: “to the superintending care of an energetic, firm, and most popular administration, I with confident security resign the civil and constitutional difference of my country; the established government in state, may in such hands be saved from destruction;” we must beg leave in this particular, to dissent from his prescription (of which we deem this an ingredient, since he recommends it by the authority of his own example) as, however energetic, firm, popular, and even strenuously devoted to public liberty the present ministers may be, they are but men, of
course liable to error and assailable by the temptations of power and interest we would, on the contrary, recommend a jealous vigilance with respect to every measure of every administration;—the immediate interests of a minister may be distinct from those of the people, he may therefore, in some instances be induced to sacrifice all considerations for the public good, to the prevailing motives of avarice and ambition:—and here we will repeat our dissent from a Right Honorable Gentleman, should he have asserted (which however, we gather only from a News-paper report) that the people have no right to enter into discussions concerning civil Polity in general, or the immediate measures of government; his legal experience we presume, will inform him that a Trustee is accountable for the proper management of his trust;—both the executive and legislative bodies are trustees of the people, to whom they are responsible, for the due discharge of the sacred trust reposed in them; the more they enquire into the effects resulting from the measures of every administration, the more competent will they become in appreciating the merits of their respective trustees, and the more clearly will they discern the propriety of conferring upon, or withholding from them their farther confidence.
He considers “unrestricted Emancipation to be impossible, so long as the Constitution in Church and State shall be Protestant” and asserts “that the necessity of excluding Papists from Parliament, and from the great offices of State, is imperious, in order to preserve both from destruction.” Neither can we subscribe to this opinion of the Revd. Author;—We have endeavored to prove (and hope with success) that the Authorities he quotes are unfounded, or obsolete and at present of no force:—an equal Participation in the same rights and privilege produces union and harmony, even among those who, from the circumstance of receiving different educations, may have imbibed different religious tenets; whilst a partial preference, and distinct privileges annexed to the profession of a particular faith, encourage the growth of jealousy and discussion:—Divide and impera is the boasted maxim of a crooked policy;—a well constituted Government consults for, and endeavours to promote the common welfare of all its subjects:—what have been the measures of policy pursued with regard to Ireland? The bulk of its population is Catholic,
and has during a century groaned under the arbitrary sway of a Protestant Government, partial to professors of its own faith, and arming them with distinct privileges;—the Catholics have been subject to an Oligarchy, composed of their declared opponents;— they have been debarred from acquiring real property;—their Estates have been liable to confiscation, for refusing to swear contrary to their consciences;—they have been restricted in the free exercise of their religion, and from educating their children in that faith which they esteemed the true one;—can it be expected, that these circumstances should attach them to the constitution? they acknowledge its excellence, and that its subjects enjoy a greater portion of liberty, than the subjects of any other state in Europe;— they look upon themselves, however, not as its subjects, but its slaves:—instead of interminable hostility against the Catholics, we would earnestly recommend conciliation;—let them be once admitted to all the privileges of subjects, they will (we dare affirm) become equally zealous in their attachment to the Constitution, as the Protestant himself.—In maintaining the contrary opinion, we regret that the Revd. Sir Harcourt Lees, should forget his character as a gentleman and a scholar, for, however he may vent his spleen in gross and vulgar abuse, a sinner against good-breeding, and the laws of decency will no more be esteemed a good author, than will a sinner against grammar, good argument, or good sense.
He is scarcely less abusive in his animadversions upon the Fanatics or Evangelical, and if they, in reality, maintain such tenets as he attributes to them, we must acknowledge that they are objectionable; they tend to diminish the general benevolence of mankind, and to render them regardless of moral rectitude; but persecution is still more dangerous, and in proposing a strong test, in addition to the Thirty-nine Articles he seems actuated, in some measure, by a spirit of persecution; even Bishop Burnet (whose orthodox we presume will not be disputed) confesses that the requiring subscription to the Thirty nine Articles, is a great imposition: —and, would you, Revs. Sir, impose a more grievous test of orthodoxy?—be assured, that the strength of your test, and its embracing a multitude of objects will not tend to confirm your establishment:—it may and probably will flourish, in power and
wealth, with the government of which it forms a part, but the superior excellence of the doctrines you profess, your own exemplary deportment in private life, together with a zealous exertion in the discharge of your sacred functions, will also render its influence extensive, or its duration permanent:—here we would recommend (but with the utmost deference and humility) the co-operation of Government,—were the gradations in preferment allotted to gradations in merit,—were the dignities of the Church, exclusively, appropriated to eminence in virtue, piety, learning, the clergy in early life would be stimulated to exertion for the attainment of these good qualities, their exertions would, in general, be crowned with success, and thus, at a more advanced period, they would become illustrious ornaments to their profession; whilst the people instructed by the impressive exhortations, and influenced by the upright conduct of their clergy, would be far more inclined to the practise of morality and good-order.
In regard to the radical reformers, their views may possibly be directed to tumult and anarchy, but the ostensible object of their contemplation do not appear to us visionary and impracticable, such material changes, however, admitted at once into the system, might be attended with danger and convulsion, we wilt by no means venture to deny that it might be so, yet Burnet (zealously devoted to Government in his time) recommends the annual Election of Representatives as an effectual expedient to stem the progress of corruption, which had, even then, made great inroads among all ranks of people: Election by ballot would defeat all the purposes which, at present, induce many of the candidates to resort to bribery; whilst universal suffrage would disable them from tampering with all the voters:—corruption has moreover in some instances, and at some periods, insinuated itself into the representative body;—the expediency of reform, therefore, can be no longer doubtful;—and that reform is necessary, the most eminent characters in our Empire have, long since, acknowledged, they have likewise on several occasions attempted to introduce it;—whilst Foreigners contemplating the excellence of the British Constitution, and enumerating the beneficial consequences, resulting to the several Nations of Europe, from the example of a free and independent
State, flourishing among themselves, have deeply lamented the existence of a vice in the system, which must gradually undermine it; —O Honte (says Raynal) l’Homme riche achete les suffrages de ses Commettans, pour obtenir l’Honneur de les representer; la Cour achete les suffrages des Representans gouverner plus despotiquement; une Nation sage ne travailleroit elle pas a prevenir l’une & l’autre corruption? N’est il pas etonnant que cela ne se soit pas fait, le jour qu’un Representant eut l’impudence de faire attendre ses Commettans dans son Antichambre, & de leur dire ensuite, je ne sais ce que vous voulez, mais je n’en ferai qu’a ma tete; je vous ai achetés fort cher, & j’ai bien resolu de vous vendre le plus cher que je pourrai:—we here confidently recommend a radical reform in ourselves, for the purpose of insuring to the body politic a sound and perfect recovery—Let every member of the state correct his own vices—and the voters should not only correct themselves; but it is their duty to examine into the qualifications which can entitle a candidate to their approbation;—is he frugal in his expences without meanness?—is he liberal, without profusion or ostentation?—is his private life marked by no destructive vice?—they may safely conclude that such a representative, unassailable by corruption, will discharge his trust with fidelity.
In consequence of the demise of our late and regretted Monarch, the period is at hand when the people will have a legal opportunity of freely and openly expressing their sentiments, in regard to the measures lately adopted by Administration;—if, in their opinion, the good and loyal subject is farther protected, by these means, in the unmolested enjoyment of his rights and privileges;—if the cottage of the meanest peasant still remains his castle, from which he may legally repel all violent intrusion;—if the Press restrained only in its licentiousness, is confirmed in the full exercise of its liberty (the great Bulwark of the British Constitution)—then will their former members, who supported such enactments, be returned to Parliament, as deserving objects of their preference:—but should they think that encroachments on civil liberty are substituted as safeguards to the Constitution;—that the habitation of every British subject is exposed, even, to nightly visitations, at the capricious will of a Magistrate;— that the liberty of the Press is essentially violated, under a specious
pretext of correcting its abuses, and preventing the dissemination of blasphemous and seditious libes;—the people will be aroused to a consideration of the danger impending over themselves, and therefore be cautious in their choice of Representatives;—in either case, however, we finally recommend inviolable abstinence from corruption:—Let no views of present emolument, no promises of further patronage,—let neither threats, nor intreaties prevail on you to depart from the conscientious discharge of your duty as voters:— the corruption of the constituent, will serve as an apology for the corruption of the Representative body:—exhibit in yourselves a generous example of reform:—your Representatives, chosen for the good qualities you have discerned in them, will copy the example and thus will the pernicious disorder, preying upon your very vitals, be in some measure checked, if not entirely eradicated from your constitution.
We have thus far obtruded upon the Public, in consequence of our objections to the Antidote of Sir Harcourt Lees, and conclude with an anxious wish that, whatever application may be adopted by them, it may most conduce to the advancement of civil and religious liberty, and may best contribute to the present and future welfare of the people.
PHILODEMUS.