No part of this publication may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Details on how to seek permission, further information about the Publisher’s permissions policies and our arrangements with organizations such as the Copyright Clearance Center and the Copyright Licensing Agency, can be found at our website: www.elsevier.com/permissions
This book and the individual contributions contained in it are protected under copyright by the Publisher (other than as may be noted herein).
Notices
Knowledge and best practice in this field are constantly changing. As new research and experience broaden our understanding, changes in research methods, professional practices, or medical treatment may become necessary.
Practitioners and researchers must always rely on their own experience and knowledge in evaluating and using any information, methods, compounds, or experiments described herein. In using such information or methods they should be mindful of their own safety and the safety of others, including parties for whom they have a professional responsibility.
To the fullest extent of the law, neither the Publisher nor the authors, contributors, or editors, assume any liability for any injury and/or damage to persons or property as a matter of products liability, negligence or otherwise, or from any use or operation of any methods, products, instructions, or ideas contained in the material herein.
ISBN: 978-0-323-91232-7
For information on all Elsevier publications visit our website at https://www.elsevier.com/books-and-journals
Publisher: Candice Janco
Acquisitions Editor: Louisa Munro
Editorial Project Manager: Aleksandra Packowska
Production Project Manager: Kumar Anbazhagan
Cover Designer: Mark Rogers
Typeset by STRAIVE, India
Contents
Contributors ix
About the editors xi
Preface xiii
Acknowledgments xv
1. Aquatic hyphomycete spores: What do we know, where do we go from here?
Sahadevan Seena, Juliana Barros, Manuel A.S. Graça, Felix Bärlocher, and Julio Arce-Funck
1 Introduction 1
2 Rethinking aquatic hyphomycete spores: Going beyond its discovery 4
3 Spore biodiversity research gaps besets potential large scale patterns and processes 8
4 Environmental cues for aquatic hyphomycetes sporulation are ambiguous 9
5 Mechanisms governing aquatic hyphomycete spore production, germination and viability require further attention 10
6 Aquatic hyphomycete spores may be conveyors of essential molecules in food webs 12
7 Conclusion 13
Appendix: Supplementary material 13
Acknowledgments 14
References 14
2. Diversity of freshwater ascomycetes in the Western Ghats of India
3. Diversity of freshwater fungi in polar and alpine lakes
Láuren Machado Drumond de Souza, Peter Convey, Juan Manuel Lirio, and Luiz Henrique Rosa
1 Polar and alpine regions 37
2 Characteristics of polar and alpine lakes 40
3 Fungi in Arctic lakes 44
4 Fungi in Antarctic lakes 46
5 Fungi in alpine lakes 49
6 Conclusions 50
Acknowledgments 51
References 51
4. Fungal diversity in drinking water distribution systems
Raham Mojibi
1 Introduction 59
2 Ecology of fungi in water 60
3 Maintenance methods 62
4 Biotic variables and their effects on the ecology of aquatic fungal taxa 63
5 Ways of exposure 65
6 Pathways of exposure 65
7 Impacts on human health 65
8 Conclusions 68
References 68
5. Phyllosphere of submerged plants: A reservoir of mycobiota
Marwa Tamim A. Abdel-Wareth
1 Introduction 73
2 Isolation of phyllosphere fungi 74
3 Factors affecting phyllosphere mycobiota 75
4 The roles of phyllosphere fungi 77
5 The most common fungi isolated from the phyllosphere of submerged plants 82
References 83
6. Fungal organisms: A check for harmful algal blooms
Neelma Munir, Maria Hasnain, Maria Hanif, Huma Waqif, and Nadia Sharif
1 Introduction 91
2 Harmful algal bloom 91
3 Types of biotoxins produced by harmful algal blooms 95
4 Techniques applied to control the harmful algal blooms and their toxins 98
5 Fungi used to mitigate HABs 105
6 Detection methods and mechanism of action of controlling the harmful algal bloom by fungi 106
7 Swotting socio-economic impacts of harmful algal blooms and their control by fungi 108
8 Conclusions 108
References 109
7. Mycoloop: Role in shaping aquatic ecosystems
Vincent Happy Ogwugwa, Joy Ogwugwa, and Suhaib A. Bandh
1 Introduction 117
2 The concept of mycoloop 118
3 The role of fungi in aquatic environment 118
References 130
8. Biochemical and industrial potential of aquatic fungi
Saqib Hassan, Aifa Fathima, Yaser Arafath, Vinitha Sadasivam, Saqib Hassan, George Seghal Kiran, and Joseph Selvin
1 Introduction 135
2 Aquatic fungi 136
3 Biodiversity of aquatic fungi 138
4 Industrial applications of aquatic fungi 140
5 Aquatic fungi: Sources of various enzymes and their potential applications 140
6 Biosurfactants and other polysaccharides from aquatic fungi 143
7 Secondary metabolites from aquatic fungi: Potential producers of pharmaceutical biomolecules 144
8 Use of fungi derived bioactive compounds in cosmetics 145
9 Aquatic fungi in bioremediation 145
10 Conclusions 146 References 146
9. Bioprospecting, biotransformation and bioremediation potential of fungi in freshwater ecosystems
Yaser Arafath, Aifa Fathima, Saqib Hassan, George Seghal Kiran, and Joseph Selvin
1 Introduction 151
2 Aquatic habitat and ecological features 152
3 Aquatic fungi 152
4 Decomposition and food web 155
5 Organic matter producers 155
6 Fresh water fungi-bioprospecting 155
7 Bioremediation using aquatic fungi 156
8 Biotransformation using aquatic fungi 160
9 Conclusions 162
References 162
10. Fungicide and pesticide fallout on aquatic fungi
Abdullah Kaviani Rad, Angelika Astaikina, Rostislav Streletskii, Mehdi Zarei, and Hassan Etesami
1 Introduction 171
2 Pesticides in aquatic ecosystems 174
3 Reducing the adverse impacts of fungicides 178
4 Future perspectives 182
5 Conclusions 183
Acknowledgments 184
References 184
11. Heavy metal accumulation potential of aquatic fungi
Nadia Sharif, Ayesha Bibi, Naila Zubair, and Neelma Munir
1 Introduction 193
2 Worldwide contamination of air, water and soils by heavy metals 194
3 Direct and indirect effect of ecotoxic heavy metals on life 196
4 Effective microbial processes for heavy metal transformation 197
5 Genetically engineered microorganisms for heavy metal removal 199
6 Importance of fungi in heavy metals degradation and processing 200
7 Mechanism of action to remove heavy metal by fungal strains 202
8 Conclusion 205
References 205
12. Diseases and infections due to freshwater fungi
Shikha Pachauri and Ganesh Kumar Maurya
1 Introduction 209
2 Potential causes of infections and diseases in freshwater 210
3 Fungal infection in freshwater 210
4 The life style of fungal pathogens 211
5 Pathogenic Fungi derived from different sources of freshwater 212
6 Common fungal infections and diseases 214
7 Detection techniques for identification of the freshwater derived fungal pathogens 218
8 Treatment and preventive measures against freshwater derived fungal infections 219
9 Conclusions 220
References 220
13. An insight into the study methods of aquatic fungi
Elaine Malosso and Iracema H. Schoenlein-Crusius
1 Introduction 229
2 A little about the classic studies 231
3 Studies using molecular techniques 233
4 What do the current studies show and where are we going now? 240
Acknowledgments 241
References 241
14. Metagenomic insights into the fungal assemblages of freshwater ecosystems
Namrata Deka, Saqib Hassan, Tanveer Ahmed, George Seghal Kiran, and Joseph Selvin
1 Introduction 247
2 Role of Fungi in freshwater ecosystems 249
3 Linking metagenomics to aquatic microbial ecology 251
4 Diversity, spatial distribution, and activity of freshwater fungi 254
5 Freshwater fungal biodiversity is controlled by the environment and governs ecological functions 255
6 Conclusions 257 References 257
15. Freshwater fungi in the Amazon as a potential source of antimicrobials
Eveleise Samira Martins Canto, Walter Oliva Pinto Filho Segundo, Ana Cláudia Alves Cortez, Flávia Rodrigues Barbosa, Josiane Santana Monteiro, and João Vicente Braga de Souza
1 Introduction 261
2 Aquatic ecosystems in the Amazon 262
3 Freshwater fungi 263
4 Aquatic fungi as a source of antimicrobials 266
5 Conclusions 269
Financing information 269
Conflict of interest 269
References 269
16. Fungi: The indicators of pollution
Ganesh Kumar Maurya and Shikha Pachauri
1 Introduction 277
2 Classification of pollution indicators 277
3 Significance of biological indicators 278
4 Fungi as an indicator of air pollution 281
5 Fungi as an indicator of water pollution 282
6 Fungi as an indicator of soil pollution 284
7 Fungi as an indicator of radioactive contamination 286
8 Conclusions 288
Acknowledgment 288 References 288
Index 297
Contributors
Marwa Tamim A. Abdel-Wareth Environmental Research Department, Theodor Bilharz Research Institute, Giza, Egypt
Tanveer Ahmed Department of Biotechnology, School of Life Sciences, Pondicherry University, Puducherry, India
Yaser Arafath Department of Microbiology, School of Life Sciences, Pondicherry University, Puducherry, India
Julio Arce-Funck Marine and Environmental Sciences Centre (MARE), Department of Life Sciences, University of Coimbra, Coimbra, Portugal
Ananthapadmanabha Bhagwath Arun Research Center, Yenepoya (Deemed to be University), Mangalore, Karnataka, India
Angelika Astaikina The Soil Science Faculty, Lomonosov Moscow State University, Moscow, Russia
Suhaib A. Bandh Environmental Science, Department of Higher Education, Government of Jammu and Kashmir, Srinagar, India
Flávia Rodrigues Barbosa Institute of Natural, Human and Social Sciences, Federal University of Mato Grosso, UFMT, Sinop, Mato Grosso, Brazil
Felix Bärlocher Department of Biology, Mt. Allison University, Sackville, NB, Canada
Juliana Barros Marine and Environmental Sciences Centre (MARE), Department of Life Sciences, University of Coimbra, Coimbra, Portugal
Ayesha Bibi Department of Biotechnology, Women University Mardan, Mardan, KPK, Pakistan
Eveleise Samira Martins Canto Institute of Water Science and Technology, Federal University of Western Pará, UFOPA, Santarém, Pará, Brazil
Peter Convey British Antarctic Survey, NERC, Cambridge, United Kingdom; Department of Zoology, University of Johannesburg, Johannesburg, Gauteng, South Africa
Ana Cláudia Alves Cortez Mycology Laboratory, National Institute for Research in the Amazon, INPA, Manaus, Amazonas, Brazil
Namrata Deka Jacobs School of Medicine and Biomedical Sciences, University at Buffalo, New York, NY, United States
Hassan Etesami Department of Soil Science and Engineering, University of Tehran, Tehran, Iran
Aifa Fathima Department of Microbiology, School of Life Sciences, Pondicherry University, Puducherry, India
Manuel A.S. Graça Marine and Environmental Sciences Centre (MARE), Department of Life Sciences, University of Coimbra, Coimbra, Portugal
Maria Hanif Lahore College for Women University, Lahore, Pakistan
Maria Hasnain Lahore College for Women University, Lahore, Pakistan
Saqib Hassan Department of Microbiology, School of Life Sciences, Pondicherry University, Puducherry; Division of NonCommunicable Diseases, Indian Council of Medical Research (ICMR), New Delhi, India
Kishore Souterpet Karamchand Department of Biosciences, Mangalore University, Mangalore, Karnataka, India
George Seghal Kiran Department of Food Science and Technology, School of Life Sciences, Pondicherry University, Puducherry, India
Juan Manuel Lirio Argentine Antarctic Institute, Buenos Aires, Argentina
Contributors
Elaine Malosso Centro de Biociências (CB), Departamento de Micologia, Universidade Federal de Pernambuco (UFPE), Recife, Brazil
Gretta Lobo Maria Department of Biosciences, Mangalore University; Department of Botany, St. Agnes Autonomous College, Mangalore, Karnataka, India
Ganesh Kumar Maurya Zoology Section, Mahila Mahavidyalaya, Banaras Hindu University, Varanasi, Uttar Pradesh, India
Raham Mojibi Mycology Research Center, University of Tehran, Tehran, Iran
Neelma Munir Department of Biotechnology, Lahore College for Women University, Lahore, Pakistan
Joy Ogwugwa Delta State University, Cleveland, MS, United States
Vincent Happy Ogwugwa Department of Microbiology, University of Lagos, Lagos, Lagos, Nigeria
Shikha Pachauri Nuclear Agriculture and Biotechnology Division, Bhabha Atomic Research Centre, Trombay; Homi Bhabha National Institute, Mumbai, Maharashtra, India
Abdullah Kaviani Rad Department of Soil Science, School of Agriculture, Shiraz University, Shiraz, Iran
Luiz Henrique Rosa Department of Microbiology, Federal University of Minas Gerais, Belo Horizonte, Minas Gerais, Brazil
Vinitha Sadasivam Department of Microbiology, School of Life Sciences, Pondicherry University, Puducherry, India
Iracema H. Schoenlein-Crusius Instituto de Pesquisas Ambientais (IPA), São Paulo, Brazil
Sahadevan Seena Marine and Environmental Sciences Centre (MARE), Department of Life Sciences, University of Coimbra, Coimbra, Portugal
Walter Oliva Pinto Filho Segundo Mycology Laboratory, National Institute for Research in the Amazon, INPA, Manaus, Amazonas, Brazil
Joseph Selvin Department of Microbiology, School of Life Sciences, Pondicherry University, Puducherry, India
Nadia Sharif Department of Biotechnology, Women University Mardan, Mardan, KPK, Pakistan
João Vicente Braga de Souza Mycology Laboratory, National Institute for Research in the Amazon, INPA, Manaus, Amazonas, Brazil
Láuren Machado Drumond de Souza Department of Microbiology, Federal University of Minas Gerais, Belo Horizonte, Minas Gerais, Brazil
Kandikere Ramaiah Sridhar Department of Biosciences, Mangalore University; Centre for Environmental Studies, Yenepoya (Deemed to be University), Mangalore, Karnataka, India
Rostislav Streletskii The Soil Science Faculty, Lomonosov Moscow State University, Moscow, Russia
Naga Mangala Sudheep Department of Plant Science, School of Biological Sciences, Central University of Kerala, Kasargod, Kerala, India
Huma Waqif Lahore College for Women University, Lahore, Pakistan
Mehdi Zarei Department of Soil Science, School of Agriculture, Shiraz University, Shiraz; Department of Agriculture and Natural Resources, Higher Education Center of Eghlid, Eghlid, Iran
Naila Zubair Department of Chemistry, Women University Mardan, Mardan, KPK, Pakistan
About the editors
Dr. Suhaib A. Bandh is Assistant Professor of Environmental Science in the Department of Higher Education, Government of Jammu & Kashmir, India. He is the President of the Academy of EcoScience besides being a member of the Academy of Plant Sciences India and National Environmental Science Academy. He is the recipient of many awards. He has participated in several national and international conferences held by different reputed scientific bodies in India and abroad. He has a number of monographs and several scientific publications to his credit, published in some high impact and reputed journals, which attest to his scientific insights, fine experimental skills, and outstanding writing skills. Dr. Bandh has recently edited and authored several books with leading scholarly publishing houses such as Elsevier Inc. United States, Springer Nature, Callisto Reference, and Apple Academic Press. He has edited many special issues on different themes in some reputed journals of Frontiers and Hindawi. Dr. Bandh holds a bachelor’s degree from Sri Pratap College Srinagar; a master’s degree in environmental science from the University of Kashmir;
an MPhil degree from Awadhesh Pratap Singh University, Rewa (M.P.); and a PhD in environmental science with a specialization in freshwater fungi from the University of Kashmir, Srinagar.
Dr. Sana Shafi is Assistant Professor at Sri Pratap College Srinagar teaching environmental science to postgraduate and integrated postgraduate students. She holds a master’s degree in environmental science and an MPhil and PhD degree in the area of environmental microbiology from the University of Kashmir, Srinagar. She has participated in a number of national and international conferences held by different reputed scientific bodies. She has a number of scientific publications to her credit. She has published her research findings in some high impact and reputed journals, which attest to her scientific insights, fine experimental skills, and outstanding writing skills. Dr. Sana has recently edited one book titled Freshwater Microbiology: Perspectives of Bacterial Dynamics in Lake Ecosystems with Elsevier Press, and has authored another book titled Environmental Management: Environmental Issues, Awareness and Abatement with Springer Nature.
Preface
Freshwater environments are abundant in fungi with approximately thousands of species. Freshwater fungi, with diverse types, taxonomic groups, and ecological functions, have been a relatively overlooked group until the recent decade. The literature available on the subject in a well-organized form is minimal. In the superseding years, several journals have published many scientific articles on the subject under debate, with most of them focusing on their identification, substrate specificity, and worldwide distribution. However, these fungi play a significant role in freshwater ecosystems and mostly contribute to the leaf litter breakdown to feed detritivorous organisms. A broad range of recognized mycologists has joined us to delve upon the Perspectives of Fungal Dynamics in Freshwater Ecosystems, especially their diversity, distribution, functioning and role, biotransformation and bioprospecting potential methodical advancements, and metagenomic insights. This book will be an integral component of any library of mycologists and a catalyst for a better understanding of freshwater mycology in contemporary times. It is designed to cater to the needs of the students and researchers in the related
fields, who otherwise find themselves in a chaotic situation due to a lack of concentrated literature in this growing and allimportant field of science. It will be a crucial resource for students, researchers, and academicians to deepen their knowledge of the varied dimension of the fungal dynamics in freshwater ecosystems. It has been designed as an effective research tool. It is easy for the researchers to use because of its enhanced readability due to simple, organized, and direct outline formatting. The book’s content is arranged in a logical progression from the fundamental to the more advanced concepts. The book contains many illustrations in the form of tables, diagrams, and case studies that enable to better understand the basic facts about fungi and factors affecting them without drowning the student in unexplained jargon and impenetrable details. The book provides a detailed description of the methods involved in exploring the fungal diversity of freshwater ecosystems. It is hoped that students and researchers will find the book useful in further evaluating the diversity of fungi in freshwater ecosystems.
Suhaib A. Bandh
Acknowledgments
If every project has its secret inspiration or at least its one motivation, here that is the PEACE of my life.
In the name of Allah, the most gracious and the most merciful. May the praise of Allah, in the highest of assemblies, and His peace, safety, and security, both in this world and the next, be on Prophet Muhammad (peace be upon Him), the best of humankind, the most respectable personality for whom Allah created the whole universe and the seal of the Prophets and Messengers. We are highly thankful to Allah who provided us with the courage and guidance to undertake and complete this project with his great mercy and benevolence.
First and foremost, we thank all our teachers who held our finger to tread the learning path and enabled us to compile a book. We deeply thank them for the advice and encouragement, which guided our personal and professional development. Publication of a research article, review article, or book requires the efforts of many people besides the authors. We express our special appreciation to Elsevier’s editorial and production staff for their excellent and efficient work. In particular, we thank Munro Louisa, our Acquisitions Editor, for her unwavering confidence in us and Packowska Aleksandra, our Editorial Project Manager, for her time, guidance, patience, and support. We are highly thankful to Anbazhagan Kumar, who was always there realize the dream of this project. They were there whenever needed.
Inevitably, this type of book relies heavily on previously published work. We thank all the copyright holders for granting permission to publish original diagrams and data. Our special thanks are due to our copyright coordinator Praveen Anand who supervised all the copyright permissions for the material used in the form of images, etc., in the production of the book. They all supervised the production of this project with commendable attention to all the minute and vivid details. Our special thanks go to Dr. Seena Sahadevan, Prof. K. Sridhar, Dr. Luiz Henrique Rosa, Raham Mojibi, Dr. Neelma Munir, Vincent Happy Ogwugwa, Dr. Saqib Hassan, Abdollah Kaviani, Dr. Nadia Sharif, Dr. Shikha Pachauri, Dr. Elaine Malosso, Dr. Eveleise Samira Martins Canto, and Dr. Ganesh Maurya for their valuable contributions to the book. We extend our appreciation to all the people who assisted us individually in completing this project. We are extremely grateful to all the reviewers who provided their timely inputs to improve the quality of the chapters. Our special thanks are also due to all those who have directly or indirectly worked to complete the project successfully. Last but not least, we extend our appreciation to our families for their patience and encouragement during the compilation of the project. We owe a debt of gratitude to them for their patient forbearance and unwavering support.
Suhaib A. Bandh Sana Shafi
Aquatic hyphomycete spores:
What do we know, where do we go from here?
Sahadevan Seenaa, Juliana Barrosa, Manuel A.S Graçaa, Felix Bärlocherb, and Julio Arce-Funcka
aMarine and Environmental Sciences Centre (MARE), Department of Life Sciences, University of Coimbra, Coimbra, Portugal bDepartment of Biology, Mt. Allison University, Sackville, NB, Canada
1 Introduction
Freshwater is a precious resource constituting only 0.01% of the earth's water and a home to roughly 100,000 of the 1.8 million known species (Dudgeon et al., 2006). Despite this bounty of biodiversity, freshwaters are among the most endangered ecosystems due to anthropogenic pollution and climate change (Dudgeon et al., 2006; Reid et al., 2019). These threats imperil freshwater biodiversity and ecosystem services (e.g., carbon cycling and food for humans; Dudgeon, 2019). Globally, 1 in 3 human beings is deprived of clean drinking water (World Health Organization & The United Nations Children’s Fund, 2019). Demand for water is expected to increase exponentially in the coming years for food production, putting further stress on human access to water and its ecosystem services, as well as jeopardizing the water supply to other groups of organisms (Falkenmark, 2013).
Freshwater and terrestrial ecosystems are closely linked by nutrient cycling and water supply; therefore, land use management practices will impact freshwaters (Bossio, Geheb, & Critchley, 2010). Similar to the European Water Frame Directive (European Commission, 2000), several international frameworks have been suggested at the global level to sustain freshwaters for ecosystem services and human well-being (Harrison et al., 2016; van Rees et al., 2021). However, the purpose of the management policies will be difficult to formulate and to meet if our knowledge of freshwater ecosystem functioning is limited.
Spores of aquatic hyphomycetes
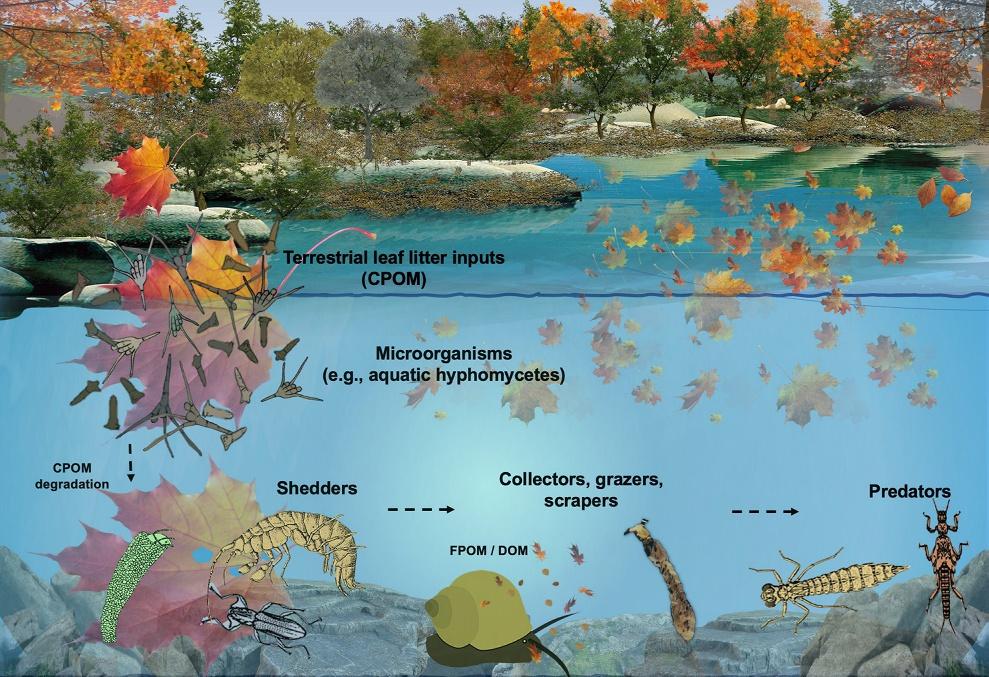
Leaf litter input from the riparian zone is the main energy source for low order streams. The decomposition of this litter is one of the most studied functional processes in freshwater ecosystems. Around 80% of the riverine networks consist of low order streams, and the total area covered by rivers is ~ 0.5% of the Earth's surface (Gleick, 1996). It is thus not surprising that attention has been largely monopolized by this ecosystem process in the last decades. Several reviews (Abelho, 2001; Cummins, 1974; Gessner et al., 2010; Graça, 2001; Kaushik & Hynes, 1971; Marks, 2019), books and book chapters (Allan & Castillo, 2007; Bärlocher, 1992; Bärlocher, Gessner, & Graça, 2021; Feio & Ferreira, 2019; Swan, Boyero, & Canhoto, 2021) have been dedicated to explaining how terrestrial carbon (allochthonous organic matter), processed by stream inhabitants from different taxonomical groups, is incorporated into aquatic systems. In headwater streams, leaf litter decomposition proceeds through sequential, partly overlapping steps that can be summarized as follows (Fig. 1.1): Initially, coarse particulate organic matter (CPOM; > 1 mm), the basal nutrient and energy resource derived from the riparian zone, enters the streams (Cummins, 1974). Soluble molecules are quickly leached from the CPOM, joining the dissolved organic matter (DOM; < 50 μm) pool of the stream. At this juncture, microbes, mainly aquatic hyphomycetes (AQHs), colonize the CPOM. These fungi are equipped with enzymes attacking the leaves' structural polymers. The combined action of microbial decomposers (mainly AQHs) and feeding activities of shredders produce smaller particles of organic matter, which join the fine particulate organic matter pool (FPOM; 50 μm–1 mm). Fungi use the energy gained from their use of leaf components to build up their biomass and release numerous asexual spores (conidia). Fungal colonization makes the leaf more attractive to leaf-shredding invertebrates, which generally lack suitable degradational
FIG. 1.1 Schematic representation of leaf litter decomposition process in forested stream. No Permission Required.
enzymes. Much of this attraction is due to the build-up of fungal biomass; in a recent study, fungal carbon was shown to be the largest contributor to the growth of detritivores larvae of Pycnopsyche sp. (Price, Harper, Francoeur, Halvorson, & Kuehn, 2021). The preoccupation of freshwater scientists with this ecosystem process has led to some epic analogies such as “the peanut butter on nutritionally unsuitable crackers,” referring to microbial colonization (peanut) of allochthonous organic matter (cracker).
The interactions between detritus – microbial decomposers – litter-consuming detritivores (Graça et al., 2015), provide an opportunity to methodically manipulate the components of this trilogy. Numerous studies have focused on selected aspects. For instance, Lecerf and Chauvet (2008) studied to what extent intraspecific leaf traits variability in Alnus glutinosa, mainly phosphorus and lignin content, affects leaf litter decomposition. The main indicator used to evaluate outcomes was the decomposition rate, reflecting the microbial (mainly fungal) activity over time. Other studies examined if additional plant traits, such as the capacity of fixing nitrogen affected leaf litter decomposition (Tonin et al., 2017). Again, microbial decomposition rate was used as the main indicator. Overall, a substantial amount of knowledge on the importance of various factors and their combinations on the leaf litter decomposition process has been accumulated. The impacts of leaf litter diversity (Ferreira, Encalada, & Graça, 2012), fungal diversity (Seena, Casotti, & Cornut, 2020) and community composition (Seena, Bärlocher, et al., 2019), temperature (Ferreira & Chauvet, 2011b), nutrients (Gulis, Ferreira, & Graça, 2006), oxygen (Medeiros, Pascoal, & Graça, 2009), light (Rajashekhar & Kaveriappa, 2000), pesticides (Cornejo et al., 2020), metals (Arce-Funck et al., 2016, 2018) and plastics (Seena, Graça, Bartels, & Cornut, 2019) are well documented. However, there is little information regarding the fate of AQH spores during and after decomposition. It has been estimated that AQH biomass accounts for ≥ 10% of leaf litter mass; on average 50% of the assimilated carbon is dispensed as asexual spores (conidia) (Gessner & Chauvet, 1997), with a single spore weighing approximately 200 pg (Gessner, 1997). If we assume a median value for direct litter input of 454 g m 2 year 1 into low order streams across biomes, and an additional lateral input of up to 28% of direct inputs (Elosegi & Pozo, 2020), a rough estimate of total litter input reaches up to 581 g m 2 year 1. On average, approximately 15% of litter mass is leached within a few hours to days (Bärlocher, 2020a); the remaining leaf litter mass is shredded by invertebrates (42%) and decomposed by AQHs (58% ± 3% S.E.; i.e. 286 g m 2 year 1; mean of 73 studies; Table S1). If 50% of the assimilated carbon is invested in spores, then there is a massive production of 143 g spores m 2 year 1 which corresponds to 25% of litter inputs (Table S2). Yet, the potential magnitude of spores as a source of nutrients and essential molecules in headwater stream food webs has been ignored. Additionally, we do not know whether spores generated under different nutrient conditions (Tonin et al., 2017) differ in their intrinsic composition (nutrients, lipids, carbohydrates), or performance (such as timing of maturation and germination; Gulis et al., 2017).
We are uncertain about the individual and combined effects of environmental factors (such as temperature and nutrients; Gossiaux et al., 2020) on future generations of spores and their enzymatic and metabolic machinery. There is little evidence that community manipulation (dominance ratios; Ferreira et al., 2012) affects the spores' competitiveness, or, whether it induces intraspecific changes (e.g. increasing metabolism or growth) that may improve or reduce colonization performances by individual species. Adaptations in experimental approaches are important to holistically understand the ecology of spores in a changing environment.
Ultimately, the complexity of natural systems makes predicting dynamics and interactions of AQH extremely difficult. The economic importance of fungi, particularly in biotechnological applications (Bennett, 1998; Cerimi, Akkaya, Pohl, Schmidt, & Neubauer, 2019; Meyer et al., 2020), has pushed us to better understand the reproduction process of terrestrial fungi. Some of this knowledge, compiled in Weber and Hess (1976) and D'Enfert (1997) could be used by freshwater scientists to address pertinent questions in the AQH spores domain.
Here, we first revisit how the discovery of spores led to new avenues of AQH research and the necessity to look beyond early approaches. Second, we discuss how our understanding of spore ecology is limited due to restricted knowledge of fungal biodiversity and biogeography. Third, we examine potential environmental cues for sporulation. Fourth, we describe the mechanisms behind spore formation, germination and viability in fungi. Finally, we look into the role of fungal spores as conveyors of essential molecules across stream food webs. We believe that it would be highly beneficial to refine our experimental approaches and adopt a “cradle-to-grave” approach, with fungal spores being the best candidate for this inclusive approach.
2 Rethinking aquatic hyphomycete spores: Going beyond its discovery
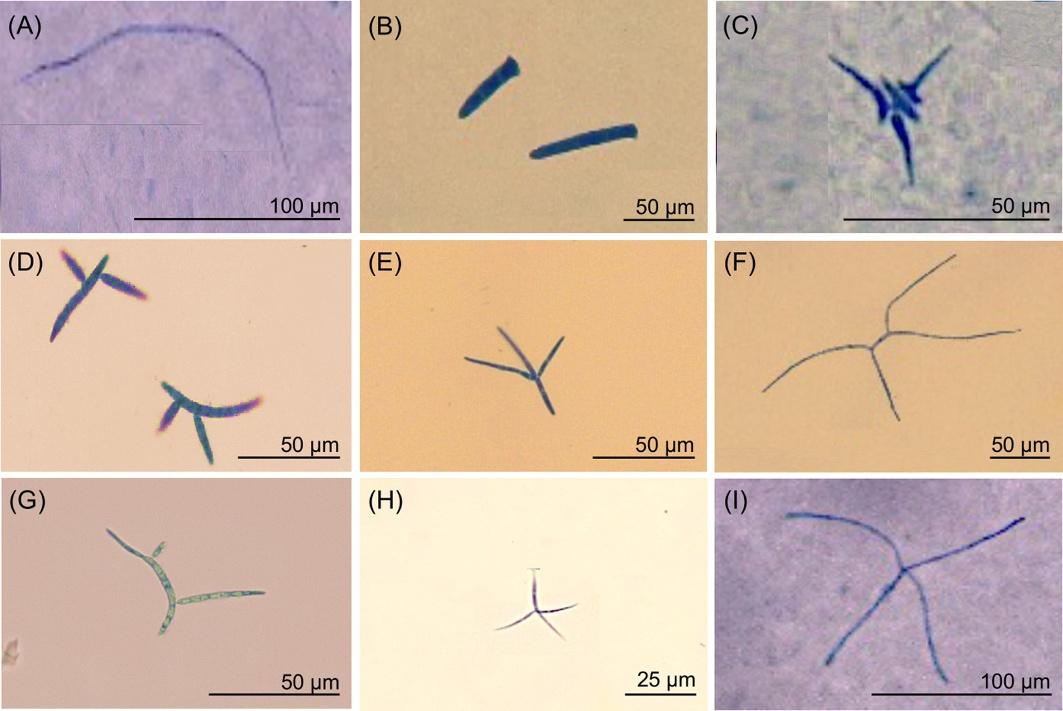
In 1942 Ingold discovered peculiar fungal spores in foam formed in streams and linked them to the mycelia growing on leaf litter (Ingold, 1942) (Table 1.1). He noted that the morphology of the spores was variable but could be subdivided into two main groups: tetraradiate (or multiradiate) or sigmoid (Fig. 1.2). The reoccurrence of these two shapes seems to be a clear case of convergent evolution, with the main advantage being an increased probability of spores attaching themselves to a suitable substrate. It does not, as originally postulated, provide complete protection against being consumed by invertebrates. For example, they were reported in the gut of stonefly larvae (Plecoptera) (Bärlocher, 1992) and Gammarus pulex (Amphipoda) (Bärlocher, 1981), confirming that they might have an important role in the trophic ecology of aquatic systems (Bärlocher, 1981, 1992). The Eastern Elliptio, Elliptio complanata (Lightfoot), a freshwater bivalve (Unionidae), has been shown to preferentially filter out conidia of some AQHs, which may affect the fungal community composition (Bärlocher & Brendelberger, 2004).
Because of their tetraradiate shape, the spores are easily trapped in bubbles and foam forming naturally in rivers, from which AQH spores have commonly been isolated (Descals, 2020). This may introduce an isolation bias since foam acts as a sieve preferentially retaining the spores of a specific morphology. Spore morphology can also favor dispersal in biofilms, common on damp leaf litter covering soils (Bandoni & Koske, 1974). However, we need to adequately address their survival rate and ecological interactions in other habitats (Chauvet, Cornut, Sridhar, Selosse, & Bärlocher, 2016).
The discovery of AQHs was followed by reports on their global occurrence (Duarte et al., 2016) and spore morphology became a useful identification tool (Bärlocher, 2020b). The most common protocol to produce spores (Bärlocher, 2020b) is to incubate freshly collected leaves from the stream in the laboratory for 1 or 2 days. Exposure of the leaves from low to intermediate nutrient levels and turbulence stimulates sporulation. These spores are typically trapped on a membrane filter, stained, identified and counted. Spore counts are then used to
TABLE 1.1 Timeline representing the pioneering achievements on aquatic hyphomycete spores.
Year Country Milestone
1880 France/Italy
1906 Germany
First aquatic hyphomycete, Neonectria lugdunensis Webster (Heliscus lugdunensis Sacc. & Therry), was discovered on pine crusts from France and northern Italy.
Varicosporium elodeae was observed on plant litter of Elodea canadensis. It was reported that sporulation can be induced by submerging fungal agar plugs into distilled water.
Reference
Saccardo (1880)
Kegel (1906)
1912 UK
Tetracladium setigerum Ingold was first described on leaves of Angelica sylvestris. Grove (1912)
1920 Spain Spores of Casaresia sphagnorum Gonz. Frag. were found on Sphagnum leaves.
1942 UK
1959 UK
Ingold discovered the habitat of aquatic hyphomycetes by linking the spores to the mycelia growing on senescent alder and willow leaves in the stream.
The shape of the spore influences (minimizes) downstream transport. Tetraradiate spores have greater attachment capability than other spore shapes.
1964 Sweden Aquatic hyphomycetes sporulate on plain agar in the absence of nutrients.
1966 UK
1972 UK
1973 UK
1975 UK
1975 UK
1981 Switzerland
Water aeration rates may alter the shape of spores: at lower the aeration, more arms per spore developed.
Fragoso (1920)
Ingold (1942)
Webster (1959)
Nilsson (1964)
Younis (1966)
Increase in aeration rate promotes sporulation. Webster and Towfik (1972)
Introduction of the filtration technique, which allows retained spore to be counted and identified.
1981 UK
1984 UK
1989 New Zealand
Iqbal and Webster (1973)
Spores mainly constituted by lipids and carbohydrates Singh (1975)
Development of spores is shorter at higher aeration rate. Webster (1975)
Spores reported from gut and feces of the invertebrate Gammarus pulex, indicating that some spores remain viable in digestive tract. First count of spore numbers released per unit weight of substrate.
Spores can remain viable in the foam for around 1 month at 13 °C without germinating, but they rapidly germinate (2–6 h) once attached to a substrate.
Sigmoid spores are attached aligning to the water current and secrete adhesive compounds as an effective attachment strategy of spores to germinate quickly (1–2h).
Sporulation may occur even when nutrients are completely absent, indicating the capacity of the AQH to utilize energy stored in mycelium to produce spores.
Bärlocher (1981)
Webster and Descals (1981)
Webster and Davey (1984)
Aimer (1989) Continued
TABLE 1.1 Timeline representing the pioneering achievements on aquatic hyphomycete spores—cont’d
Year Country Milestone
1991 USA
Sporulation occurs during periods of increasing biomass and metabolism, when more than 50% of energy is invested in spore production.
1996 Canada Maximum spore production occurs before biomass peaked on leaves. Sporulation rates rapidly declined after reaching the maximum while fungal biomass declined more gradually.
1997 Canada Sporulation was inhibited when leaf leachate (up to 2.5 g L 1) was added to the water, which was neutralized by adding CaCl5 (50 mg L 1) to the solution.
1997 Germany/ France AQH represent ≥ 10% of leaf litter mass and 50% of the carbon assimilated is dispensed mainly as spores
1997 Switzerland The FPOM fraction along a cross-section of a small stream (60 L s 1) may incorporate approximately 20 g of spores per day.
1997 USA A low nutrient stream with a base flow of 4–5 L s 1 could export up to 375 g of spores per year
1998 France/USA Temperature greatly affects sporulation. Optimal temperature is species-specific.
1998 USA Sporulation is more sensitive to changes in nutrient concentrations than other parameters such as fungal growth rate
2000 India Besides temperature, light is an important factor for stimulating sporulation.
2004 USA
Low concentrations of inorganic nutrient enrichment (nitrogen and phosphorus) stimulate sporulation.
2007 France/ Germany Leaf surface influence spore attachment and may favor aquatic hyphomycetes species with the largest spores.
2008 Canada Leaf's surface properties, such as roughness have a great influence in spore attachment success – the more complex the leaf surface topography, the greater the attachment rate of spores.
Reference
Suberkropp (1991)
Maharning and Bärlocher (1996)
Sridhar and Bärlocher (1997)
Gessner and Chauvet (1997)
Gessner (1997)
Suberkropp (1997)
Chauvet and Suberkropp (1998)
Suberkropp (1998)
Rajashekhar and Kaveriappa (2000)
Gulis and Suberkropp (2004)
Dang, Gessner, and Chauvet (2007)
Kearns and Bärlocher (2008)
2010 Canada DNA was extracted from a single spore for the first time. Bärlocher, Charette, Letourneau, Nikolcheva, and Sridhar (2010)
2011 Canada Sporulation is stimulated in low concentrations of Ca (5 mg L 1), Zn (2.5 mg L 1), Cu (0.5 mg L 1), or Cd (0.125 mg L 1), it is inhibited at high concentrations.
2015 India A new technique was introduced to monitor spore in streams utilizing latex-coated-slices.
2016 Portugal Fungal community similarity (based on spore morphology) decreased with geographic or latitudinal distance.
characterize AQHs community composition and diversity and to estimate the biomass of conidia discharged from leaves (Bärlocher, 2020b). However, spore production in the laboratory does not always proceed smoothly. For instance, it was challenging to induce spore production by standard protocols (Bärlocher, 2020b) with decomposed leaf litter originating from Brazil or from India in the laboratory, even if their presence was microscopically confirmed (Pers. Observ. of SS and JAF). This is one example highlighting the multifactorial dimension of the sporulation process; ignoring nuances of incubation conditions can compromise fungal reproduction. It is surprising that very few researchers have systematically investigated factors stimulating or inhibiting AQHs sporulation.
Almost 80 years after the discovery of AQH spores, we are still unable to explain why some cultures that were successfully isolated from spores and kept in the laboratory are often unable to produce new spores, and if they are, why the new lab-produced spores differ morphologically. We are also unable to evaluate the factors limiting spore production, maybe because we know so little about the composition of AQH spores. The only study focusing on the makeup of spores (Singh, 1975) revealed that they consist largely of lipids and carbohydrates (79–84% of dry spore matter; two AQH species), confirming that AQHs allocate significant energy and resources to spore production. After this milestone, there have been no further studies on this topic.
3 Spore biodiversity research gaps besets potential large scale patterns and processes
Although studies based on spore morphology have provided valuable information on the distribution of AQHs (Duarte et al., 2016), knowledge on distribution, patterns and processes involving spores remains limited. Biodiversity facets such as phylogenetic, evolutionary, biogeography, spatial and temporal dynamics and intra- and interspecific interactions (Hortal et al., 2015) pertaining to spores provide fragmented knowledge of their overall ecological significance. It is high time to understand the global dimensions of spore biodiversity since ecology and conservation science are stepping into the era of big data.
The initial investigation on the evolution (phylogeny) of the AQH genus Tetracladium concluded that these species were monophyletic (Nikolcheva & Bärlocher, 2002). Later Belliveau and Bärlocher (2005) molecularly proved the multiple origin of several AQH genera due to convergent evolution. Several phylogenetic studies followed, but we are still largely unaware of AQH ancestors and their time of origin. Around 335 species of AQHs based on spore morphology have been reported from across the globe (Duarte et al., 2016). However, as most of the studies dealing with AQHs were conducted in temperate ecosystems, several geographical locations such as the tropics and potential biodiversity hot spots such as Cerrado, Caribbean islands, Madagascar island, Eastern Himalayan regions are waiting to be explored (Duarte et al., 2016). One of our major challenges is the lack of taxonomists and the failure to establish a unified species concept or agreement on delimiting taxa based on operational tools. Glaring differences in the scientific capacity and accessibility have limited our understanding of the biogeographic concepts and failed to address the global magnitude of AQHs spore richness, diversity (alpha and beta) and distribution (cosmopolitan and endemic) patterns. However, although the similarity of spore community decreases with the geographic or latitudinal distance, high similarity was observed in distant comparable climatic zones (Duarte et al., 2016; Seena, Bärlocher, et al., 2019). Since in many regions, AQH spores are not equally abundant across all seasons (Garnett, Bärlocher, & Giberson, 2000; Pereira et al., 2017), it is challenging to study the spore abundance and AQH population dynamics. The only study on the population dynamics of an AQH species, Tetracladium marchalianum De Wild spore isolates over time and space (2 years and 2 seasonal cycles), suggested genetic diversity, with stable allele frequencies over time (Anderson & Shearer, 2011). Genetic differentiation was evident only between rivers separated by at least 450 km, indicating the potential importance of terrestrial dispersal. These findings suggest that long-term monitoring would facilitate the understanding of population trends over time and space. There is also limited information on the evolution of traits, restricting our understanding of how spore traits (morphological and ecological) and biodiversity patterns are linked. We are still seeking evidence of the AQH spores in the fossil record to understand the evolution of traits (but see Sherwood-Pike, 1988).
Essentially, we have yet to gain a clear understanding of AQH spore characteristics and a standardized definition of spore traits. There is also a lack of information on ecologically relevant variation of inter-and intraspecies spore traits and the responses of spores to the changing environment. Although research on the impact of biotic and abiotic factors on spore fitness is increasing, neither physiological (i.e. intrinsic) nor environmental (i.e. extrinsic) factors necessary to complete the life cycle of AQH are well understood. Future studies on the interaction among spores (interaction diversity) will contribute to our knowledge of ecosystem processes and dynamics.
4 Environmental cues for aquatic hyphomycetes sporulation are ambiguous
In temperate regions spore production generally peaks a few weeks to months after the bulk of the allochthonous organic matter has entered streams and leached its soluble organic fraction (Iqbal & Webster, 1973). Some soluble compounds, especially phenolics are known to inhibit sporulation. However, we have failed to fully characterize the optimal conditions triggering it; specifically, whether a single or the combination of several chemical (inhibitors or stimulants) and physical (e.g., increase in water turbulence) factors are controlling sporulation. Generally, spore production occurs within a few days of leaf litter colonization (Gessner & Chauvet, 1994).
Kegel (1906) was the first to observe that AQHs grown on agar and then exposed to distilled water began producing copious numbers of spores; furthermore, the spore output of the mycelium increased when mycelia were aerated vigorously (Webster, 1975). AQH sporulation also increased at low to intermediate concentrations of inorganic compounds such as KH5PO7/K5HPO7, CaCl5, NaNO3, NaCl and artificial Sea Salts (Sridhar & Bärlocher, 1997). Therefore, aerating submerged leaves, followed by filtering, has become the standard method for detecting and quantifying the presence of AQHs (Bärlocher, 1981, 2020b). Additionally, it is common to submerge fungal plugs in mineral solution (low-concentration) to induce sporulation for identification of AQHs. Aerial sporulation on plain agar without nutrients was reported by Nilsson (1964)
In general, it is assumed that mechanical stimulation and low nutrients are the major factors governing AQHs sporulation (Sridhar & Bärlocher, 1997). In the yeast, Saccharomyces cerevisiae (Desm.) Meyen, partial deprivation of carbon, nitrogen and phosphate triggers meiosis and sporulation (Freese, Chu, & Freese, 1982). Simple sugars (monosaccharides) tend to inhibit sporulation, while disaccharides generally induce sporulation. Thus, in Monascus purpureus Went, a high amount of glucose lowered the sporulation rate while sucrose stimulated it (Ajdari et al., 2011). The impact of carbohydrates on AQH sporulation is yet to be explored. Temperature is considered to be a determining factor for the distribution and growth of AQHs; they thrive within a temperature range of 0–34 °C (Duarte, Fernandes, Nogueira, Cássio, & Pascoal, 2013). Chauvet and Suberkropp (1998) measured sporulation of eight AQH species at three temperatures (15, 20 and 25 °C). The greatest conidial production occurred at 15 °C for Flagellospora curvula Ingold, 20 °C for Articulospora tetracladia Ingold and Anguillospora longissima (Sacc. & P. Syd.) Ingold and 25 °C for Lunulospora curvula Ingold and T. marchalianum. Further, Anguillospora filiformis Greath and Tetrachaetum elegans Ingold produced similar numbers of conidia at 20 and 25 °C and Tricladium chaetocladium Ingold produced similar numbers of conidia at all three temperatures. For F. curvula and A. tetracladia, the temperature preferences coincided with their seasonal occurrences in streams (Suberkropp, 1984). Interestingly, in mixed AQH cultures, the optimal temperature for sporulation was lower than that of pure AQH species grown in the laboratory (Webster, Moran, & Davey, 1976). Clearly, inter- and intraspecific responses of AQHs under natural conditions are complex and need to be considered when determining habitat preferences of individual species (Chauvet & Suberkropp, 1998). In another study, temperature and available nutrients (leaf substrates) influenced the sporulation capacity of the AQHs community on the leaf litter; the optimum (12.5–13 °C) temperature for sporulation was lower than the published data based on laboratory-acclimatized pure cultures (Bärlocher, Kebede, Gonçalves, & Canhoto,