Aquaculture
journal homepage: www.elsevier.com/locate/aquaculture
Effect of protein on the post-ingestive response of tilapia fed encapsulated diets
Leandro S. Costa a , Helder O. Guilherme b, * , Raphael N. Bahiense b , F´ abio A.C. Santos b , Pedro G. Gamarano b , Jose F.L. Olmeda c , Paula A.P. Ribeiro b , Ronald K. Luz b
a Departamento de Zootecnia, Universidade Federal de Viçosa (UFV), 36570-900, Avenida PH Rolfs, s/n, Campus Universitarios, Viçosa, Minas Gerais, Brazil
b Laboratorio de Aquacultura, Universidade Federal de Minas Gerais (UFMG), 31270- 901 Belo Horizonte, Minas Gerais, Brazil
c Department of Physiology, Faculty of Biology, Regional Campus of International Excellence “Campus Mare Nostrum” , University of Murcia, 30100 Murcia, Spain
ARTICLE INFO
Keywords:
Protein intake
Feeding behavior
Physiology
Nutrition
Fish
ABSTRACT
Two experiments were carried out with Nile tilapia (Oreochromis niloticus) to assess whether protein concentration influences post-ingestive signaling and the ability of fish to modulate protein intake from a practical diet. The animals were fed three types of encapsulated diet: control, cellulose-based diet (CD) = 0% protein; diet with 25% crude protein (D25); and diet with 42% crude protein (D42). In the first experiment, 20 fish were divided into two groups, one being fed CD and D25 and the other fed CD and D42. In the second experiment, 15 tilapia were fed D25 and D42. In the first experiment, the two groups of tilapia consumed more capsules containing D25 (62.83%) and D42 (55.54%), and the animals that received D25 noticed the difference between the capsules more quickly (7 days of the experiment) than did those that received D42 (11 days). In the second experiment, there was a preference for capsules containing D25, compared to D42, which was demonstrated after 23 days of the experiment. The animals adjusted both the amount of crude protein ingested daily (31.86%) and the rate of daily feed consumption to biomass (1.36%). Tilapia can select the diet that best meets their nutritional needs, adjusting the protein ingested and feed consumed.
1. Introduction
The term “nutritional wisdom” is used to describe the ability of fish to regulate nutrient intake (S´ anchez-V´ azquez et al., 1994; SanchezV´ azquez et al., 1995; Simpson and Raubenheimer, 2001; Geurden et al., 2005; Fortes-Silva et al., 2011a; Fortes-Silva et al., 2016; Santos et al., 2019; Abd El-Wahab et al., 2020). The control of nutrient intake, including protein, by fish is similar to that described for mammals, being done via chemical receptors present in the gastrointestinal tract (Rubio et al., 2003; Almaida-Pag´ an et al., 2006; Fortes-Silva et al., 2011a, 2011b; Fortes-Silva et al., 2016), liver tissue (Otero-Rodino et al., 2016) and brain tissue (Otero-Rodino et al., 2015; Babaei et al., 2017; Comesana et al., 2018; Comesana et al., 2020), after evaluating the nutritional constituents of the food. These chemical receptors originate signals (neural and hormonal activity) that inform brain centers about the nutritional properties of foods and, thus, modify eating behavior (Forbes, 2001). With this information, animals learn to regulate the intake of a particular nutrient, with its metabolic consequences, called
“nutritional reward” , at post-ingestion and post-absorption levels (Forbes, 2001).
Animals have evolved several mechanisms for regulating ingestion that enable the perception of the energy content, as well as the deficiency of some nutrients, of a given food. This set of mechanisms was called “specific hunger”: that is, the animal is able to sense nutrients in the diet and choose a diet with the required elements. (Fortes-Silva et al., 2016). However, these mechanisms are not unique in the decision to ingest or not certain foods (Simpson and Raubenheimer, 2001; FortesSilva et al., 2012). The preference for a particular food, and even the amount consumed, can be influenced by its “palatability” , a fixed effect of its sensory properties, including taste and texture (Sanchez-Vazquez et al., 1994; Simpson and Raubenheimer, 2001). However, classical ingestion control theories suggest that this event entails changes in some aspect of the animal’s physiology, which eventually becomes strong enough to inhibit or stimulate feeding (Forbes, 2001).
Studies using self-demand feeders have shown that fish are able to select macronutrients, such as proteins, lipids and carbohydrates, and
* Corresponding author at: Universidade Federal de Minas Gerais, Escola de Veterin´ aria, Departamento de Zootecnia, Laborat ´ orio de Aquacultura – LAQUA, Av. Antonio Carlos, 6627, CEP 31270–901 Belo Horizonte, MG, Brasil.
E-mail address: helderog@gmail.com (H.O. Guilherme).
https://doi.org/10.1016/j.aquaculture.2022.738289
Received 22 August 2021; Received in revised form 24 January 2022; Accepted 22 April 2022
Availableonline27April2022
0044-8486/©2022ElsevierB.V.Allrightsreserved.
thus regulate the amount of each element to be ingested (Vivas et al., 2006; Geurden et al., 2007; de Mattos et al., 2016; Fortes-Silva et al., 2017; Luz et al., 2018; Santos et al., 2019). Morais (2017) and Bertucci et al. (2019) demonstrated the existence of taste receptors and signaling elements in the gastrointestinal tract of fish, suggesting that, in addition to oral taste sensations and palatability, the sensory properties of a diet can also have functional effects. Thus, the methodology of using encapsulated foods challenges animals to select or self-compose a postingestion diet, without evaluating organoleptic characteristics such as flavor and texture (Almaida-Pag´ an et al., 2006; Fortes-Silva et al., 2011a; Santos et al., 2019; Almeida et al., 2021).
Proteins are extremely important to fish organism (NRC, 2011). These molecules compose part of muscle and act as hormones, antibodies, enzymes and molecules that store and transport substances (Tu et al., 2015). In addition, protein and amino acids play a role in controlling hunger and satiety, thus acting on food ingestion (Forbes, 2001). In aquaculture, the amount of protein supplied to animals must be carefully evaluated, as the cost of its inclusion in the diet will impact final production costs (Kabir et al., 2019). The supply of this nutrient below the animal’s requirement can affect its growth. At the same time, its excess in the diet leads to the catabolism of amino acids for energy supply, increasing ammonia excretion and directly impacting water quality (NRC, 2011).
The objective of this work was to study the post-ingestive response related to the control of consumption in tilapia fed with encapsulated diets of low and high protein content.
2. Material and methods
The experiment was performed at the Laborat ´ orio de Aquacultura (Laqua) of the Departamento de Zootecnia of the Universidade Federal de Viçosa, following the ethical norms approved by the Ethics Committee on the Use of Animals of the institution (protocol n◦ 0134/2019).
2.1. Animal facilities and maintenance
Juvenile Oreochromis niloticus, phenotypically inverted to males, were acquired from the 3D Aqua Ltda. (Belo Horizonte, Minas Gerais, Brazil) and housed in 40-L aquaria, maintained in a recirculation system with controlled temperature, constant aeration and a 12Light:12Dark photoperiod (Key West group DNI). The physical-chemical parameters of the water remained within ideal levels for the species, with the following mean values: dissolved oxygen 4.75 ± 1.34 mg L 1 , temperature 27.1 ± 0.8 ◦ C, pH 6.6 ± 0.2 (Probe model HI9146, Hanna instruments) and total ammonia 0.03 ± 0.02 ppm L 1 (Labcon Test).
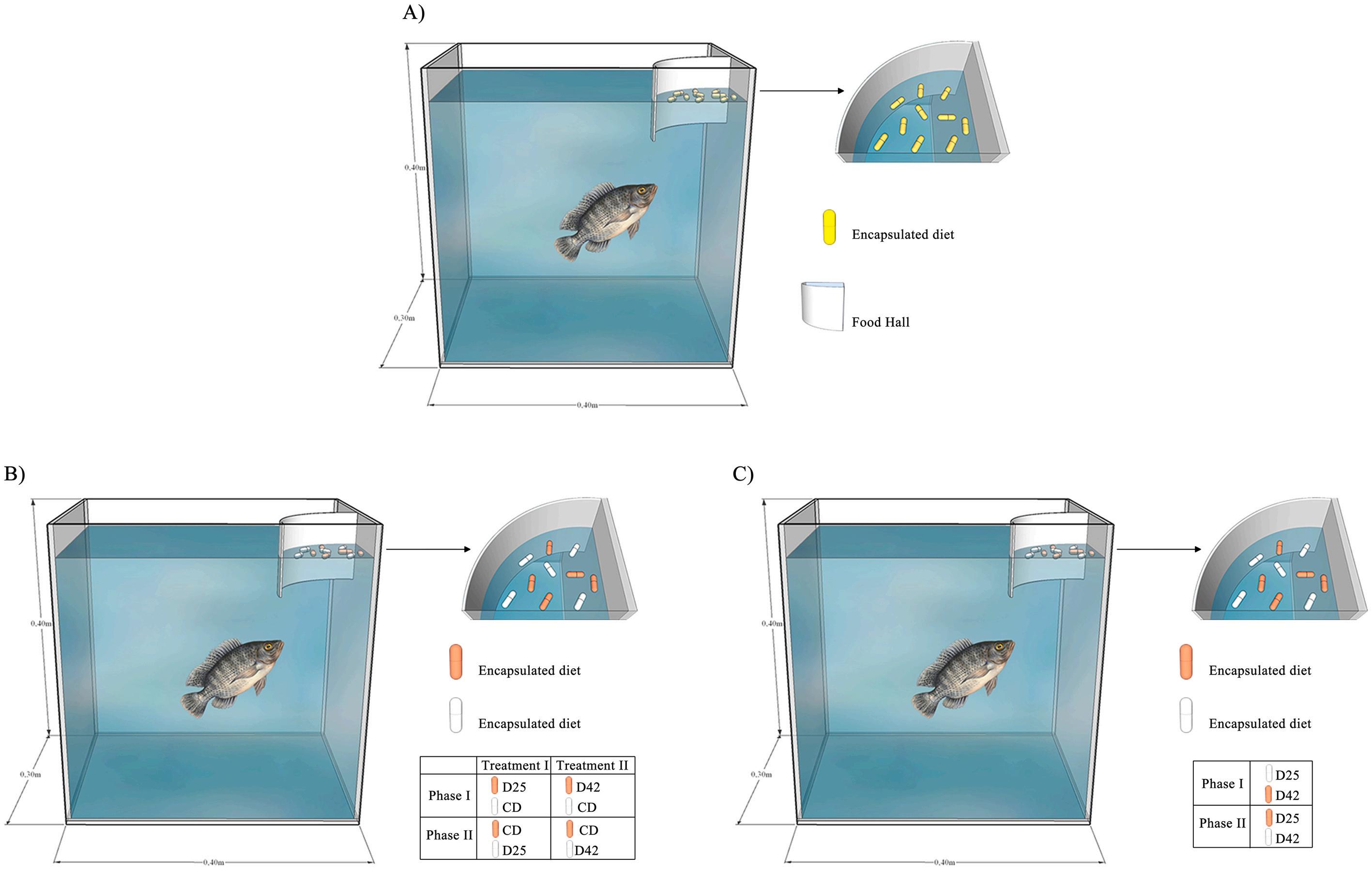
To avoid dominance and territoriality among animals, which could affect food ingestion, each aquarium contained a single animal (FortesSilva et al., 2011a; Fortes-Silva et al., 2012), adapted to the experimental facilities for 14 days. During this period, feeding training was carried out, with the supply of encapsulated diet (Fig. 1A) containing commercial feed with 32% crude protein and 3800 Kcal g 1 of crude energy, supplied twice a day (at 09:00 and 17:00 h). The capsules were offered to the animals and, after 5 min, the leftovers were collected. The time of 5 min was determined by testing the stability of the capsule in water to maintain its structural integrity, thus preventing its breakage and consequent release of contents into the aquarium.
2.2. Formulation and preparation of encapsulated diet
Two isoenergetic diets with ~4165 Kcal g 1 of gross energy were formulated based on the nutritional requirements of juvenile tilapia (NRC, 2011). Diets varied in protein content, the first with 25% crude protein and the second with 42% (Table 1). The diets were extruded, with pellets ranging from 6 to 8 mm in diameter (Imbramx40, Imbramaq Ltda., Ribeirao Preto, Sao Paulo, Brazil). Oil was sprayed after extrusion and the diets were dried and stored in a cold chamber ( 20 ◦ C) until use.
To eliminate the effect of palatability and organoleptic properties of the food, the diets were supplied inside gelatin capsules, (number 04, FAGRON – SM Empreendimentos Ltda, An´ apolis, Goi´ as, Brazil). The feed pellets were ground to reduce particle size and inserted into the gelatin capsules with the aid of a manual encapsulator (Ideal®, Aruj´ a, Sao Paulo, Brazil).
White- and orange-colored capsules were used in the experimental phase. This procedure allowed capsule recognition by the fish and facilitated identification when recording leftovers in the aquariums. After encapsulation of the experimental diets, a sample of 100 capsules was taken for individual weighing to obtain the average capsule weight for each diet. This value was used to calculate the consumption of each diet as well as the daily intake of protein and energy. Capsule mean weight and standard deviation for each diet were: D25 = 0.1769 ± 0.009 g; D42 = 0.1680 ± 0.007 g, and CD = 0.1245 ± 0.005 g.
2.3. Experiment 1
An experiment was carried out to evaluate the ability of juvenile O. niloticus to identify the internal content of capsules after ingestion and determine whether protein content affects the time required for this identification The experiment had a completely randomized design with two treatments and ten replications. Twenty fish (average weight 66.41 ± 2.85 g) were housed in 20 aquaria (40-L each). Four types of capsules with different contents were produced:
- Training capsule: yellow capsule, containing commercial feed and provided only during the training phase of the animals to consume the diet;
- Negative control capsule (CD): filled only with cellulose;
- Low protein capsule (D25): filled with experimental diet containing 25% crude protein;
- High protein capsule (D42): filled with experimental diet containing 42% crude protein.
For two weeks, the animals were trained to consume capsules filled with commercial feed (Fig. 1A), after which they were fasted for two days before the beginning of the first phase of Experiment 1. The capsules were initially offered to the animals at a total amount of 10% of live weight, the amount corresponding to 5% of each diet. This amount was periodically adjusted, according to consumption, so that there was always a surplus of both offered diets. The control diet (CD) was offered in white capsules to all animals and the experimental diets, D25 and D42, were supplied in orange capsules, each to 10 different animals (Fig. 1B). The first phase of the experiment was completed when the animals demonstrated a statistically significant and persistent consumption preference for one of the capsules (Fortes-Silva et al., 2011a), which occurred at day 16 of the experiment. At the end of the first phase, the animals were fasted for two days before the beginning of the second phase, during which the internal content of the capsules was inverted. Thus, the experimental diets, D25 or D42, were offered in white capsules and CD, with only cellulose, in orange capsules. This procedure was necessary to demonstrate that capsule color does not interfere with the food preference of the animals. The second phase was completed when the animals demonstrated a statistically different and constant percentage of consumption of one of the capsules (Fortes-Silva et al., 2011a), which occurred after another 15 days of the experiment. Experiment 1 had a total duration of 33 days and food preference was calculated in relation to the percentage of the offered diets consumed.
2.4. Experiment 2
A second experiment was carried out to evaluate the ability of tilapia to select and adjust dietary intake based on protein level. Fifteen fish (average weight 81.39 ± 4.49 g) were housed individually in 15 aquariums (40 L each). After capsule consumption training (same
Fig. 1. Experimental scheme. A) Training and maintenance of animals, B) Experiment 1, C) Experiment 2.
Table 1
Composition of experimental diets.
cComposition
dAmino acid profile (g/100 g)
∑ EAA/∑ NEAA 0.83 0.83
a Vitamin and mineral supplement composition: Vit. A. 1.200.000 UI; Vit. D3.200.000 UI; Vit. E. 12.000 mg; Vit. K. 2.400 mg; Vit. B1. 4.800 mg; Vit. B2.4.800 mg; Vit. B6. 4.000 mg; Vit. B12. 4.800 mg; Ac. folic. 1.200 mg; Ac. Pantothenic. 3.750 mg; Vit. C. 48.000 mg; Biotin. 48 mg; Hill. 65.000 mg;Niacin. 24.000 mg; Fe. 10.000 mg; Cu. 6.000 mg; Mn. 4.000 mg; Zn. 6.000 mg;I. 20 mg; Co. 2 mg; Se. 20 mg.
b Butil-Hidroxi-tolueno (antioxidant).
c Values analyzed in the laboratory. Expressed on the basis of dry matter.(Dry matter in greenhouse 105◦ KJeldahl method Energy through the calori-metric pump Ethereal extract through extraction with petroleum ether Mineralmatter in muffle at 600◦
d Values analyzed in laboratory; (HPLC and enzymatic).
procedure as in Experiment 1), the animals were fasted for two days prior to the beginning of the first phase of Experiment 2. In this phase, the fish were fed twice a day (at 09:00 and 17:00 h), with two types of capsules being made available (Fig. 1C):
- White capsule, filled with an experimental diet containing 25% crude protein;
- Orange capsule, filled with an experimental diet containing 42% crude protein.
The first phase lasted 30 days, the time necessary for the animals to demonstrate a constant consumption of the diets. The fish were then fasted for two days before the beginning of the second phase, when the internal content of the capsules was changed, which lasted for 15 days. At the end of the experimental period, the daily and final consumption percentage of each diet, the average daily percentage of protein ingested by the animal, and the daily food consumption rate to biomass, were calculated.
2.5. Data analysis
The results for daily and final consumption percentage of capsules in the two phases of the two experiments, as well as the average daily percentage of protein ingested by the animal and the daily food consumption rate to biomass, are presented as mean and standard error (SEM). All data were submitted to Levene’s homoscedasticity test and the Shapiro-Wilk normality test. Data on percentage of daily and final consumption of capsules were submitted to Student’s t-test (p < 0.05). The data referring to the average daily percentage of protein ingested by the animal and the daily food consumption rate to biomass were submitted to ANOVA and, when significant, Scott-Knott test (p < 0.05). Statistical analyses were performed with InfoStat® (version 2016, University of Cordoba, Cordoba, Argentina).
3. Results
3.1. Experiment 1
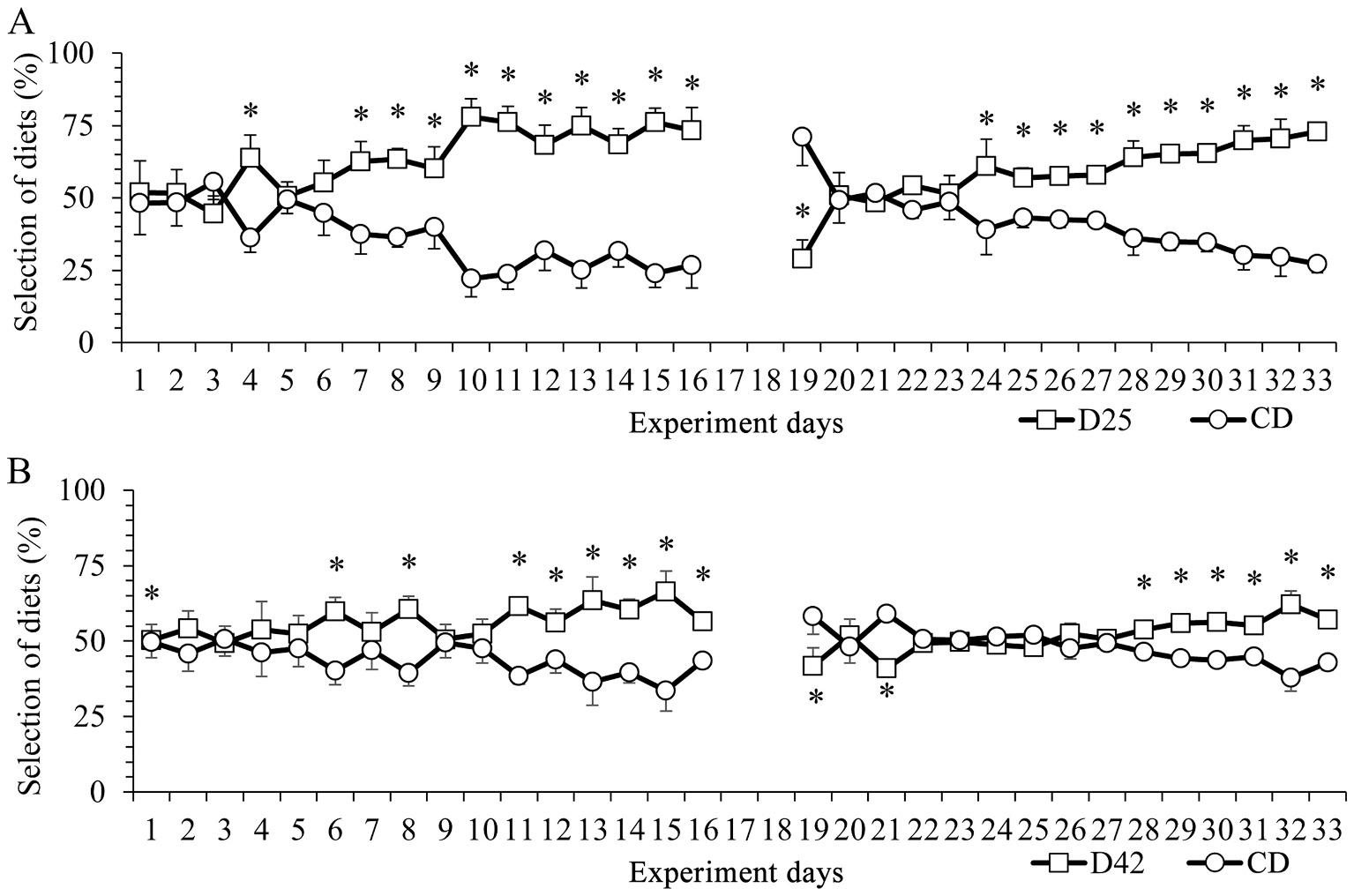
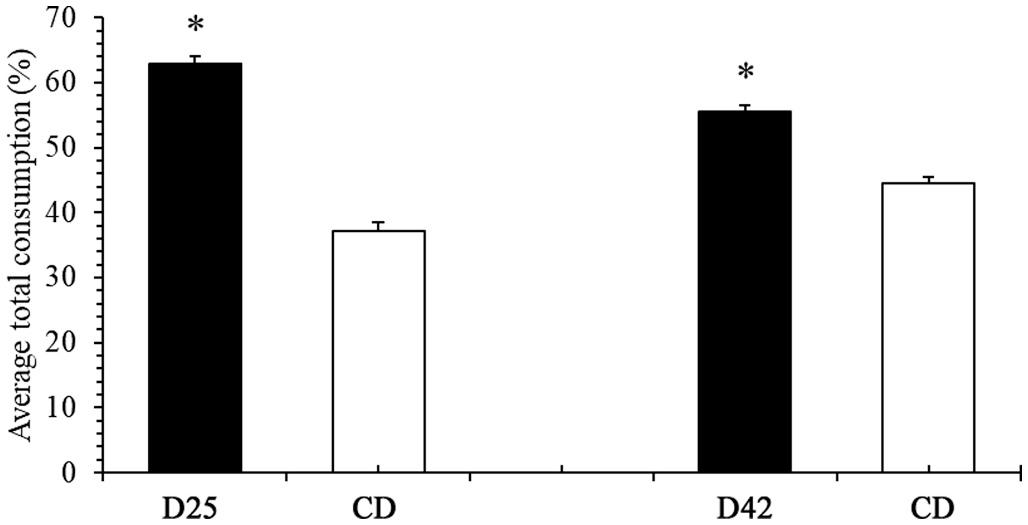
The tilapia did not show a preference for any of the diets provided at the beginning of Experiment 1. However, as the experiment progressed, the animals increased their consumption of capsules containing the experimental diets (Fig. 2A and B). The animals in the group that received D25 and CD diets showed a consistent and higher intake of D25 on the 7th day of the experiment, which continued until the end of the first phase (P < 0.05) (Fig. 2A). Fish in the group that received D42 and CD diets showed higher consumption of D42 capsules only on the 11th day, which continued until the end of the first phase (p < 0.05) (Fig. 2B). In the second phase of Experiment 1, when there was an inversion of capsule contents, the animals of both groups (D25 vs CD and D42 vs CD) had greater consumption of capsules containing the control diet on the 1st day (p < 0.05). This preference, however, did not continue throughout the evaluation period. The group of animals that received D25 and CD diets (Fig. 2A) did not show a diet preference between the 2nd and 5th day of the second phase, that is, from the 20th to the 23rd day of the experiment. However, on the 6th day of the second phase, the animals showed a statistically higher consumption of the D25 diet (p < 0.05). This preference remained until the end of Experiment 1. A similar result was observed for the group of animals that received D42 and CD diets (Fig. 2B). Consistent consumption preference for one of the diets was only demonstrated ten days after the beginning of the second phase, that is, on the 28th day of the experiment, and remained this way until the end of the experiment (p < 0.05). Fig. 3 shows the total consumption of capsules at the end of the experiment. In both treatments, the animals consumed significantly more capsules containing the formulated diet when compared to the control diet resulting in D24 (62.83%) vs. CD (37.17%) and D42 (55.54%) vs. CD (44.46%).
3.2. Experiment 2
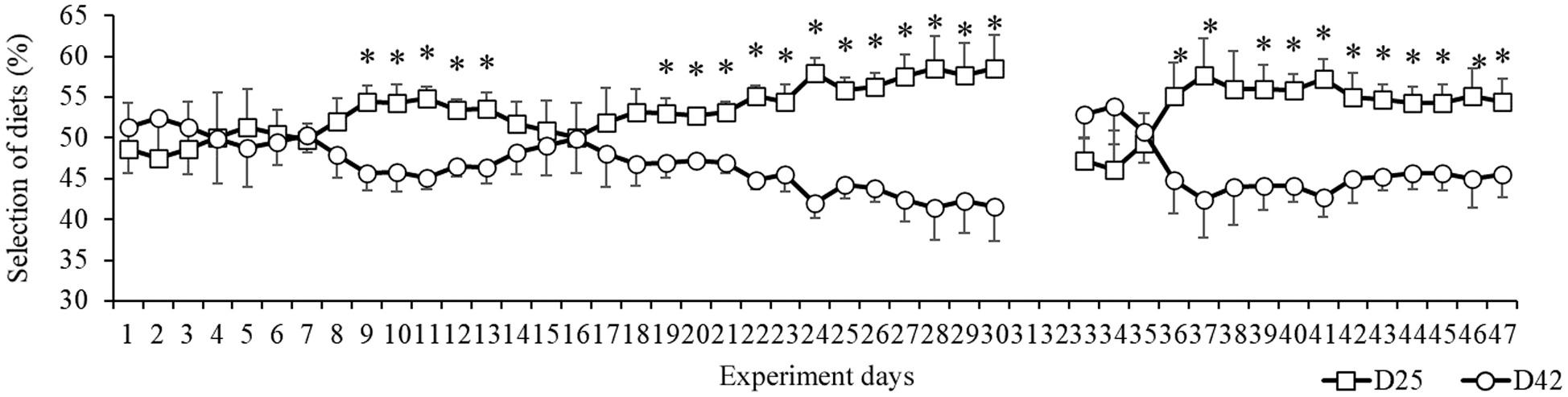
Initially, the animals did not show a consumption preference between the offered capsules, a behavior that remained until the 8th day of the experiment (Fig. 4). From the 9th to the 13th day of the experiment, there was greater consumption of D25 than of D42 (p < 0.05). This same preference was not observed from the 14th to the 18th day, a period in which the animals consumed both capsules (p > 0.05). From the 19th day of the experiment until the end of the first experimental phase, the animals returned to show preference for D25 (p < 0.05). In the second phase, after inversion of the color of the capsules, the animals resumed the selection of the D25 diet on the 4th day and continued to do so until the end.
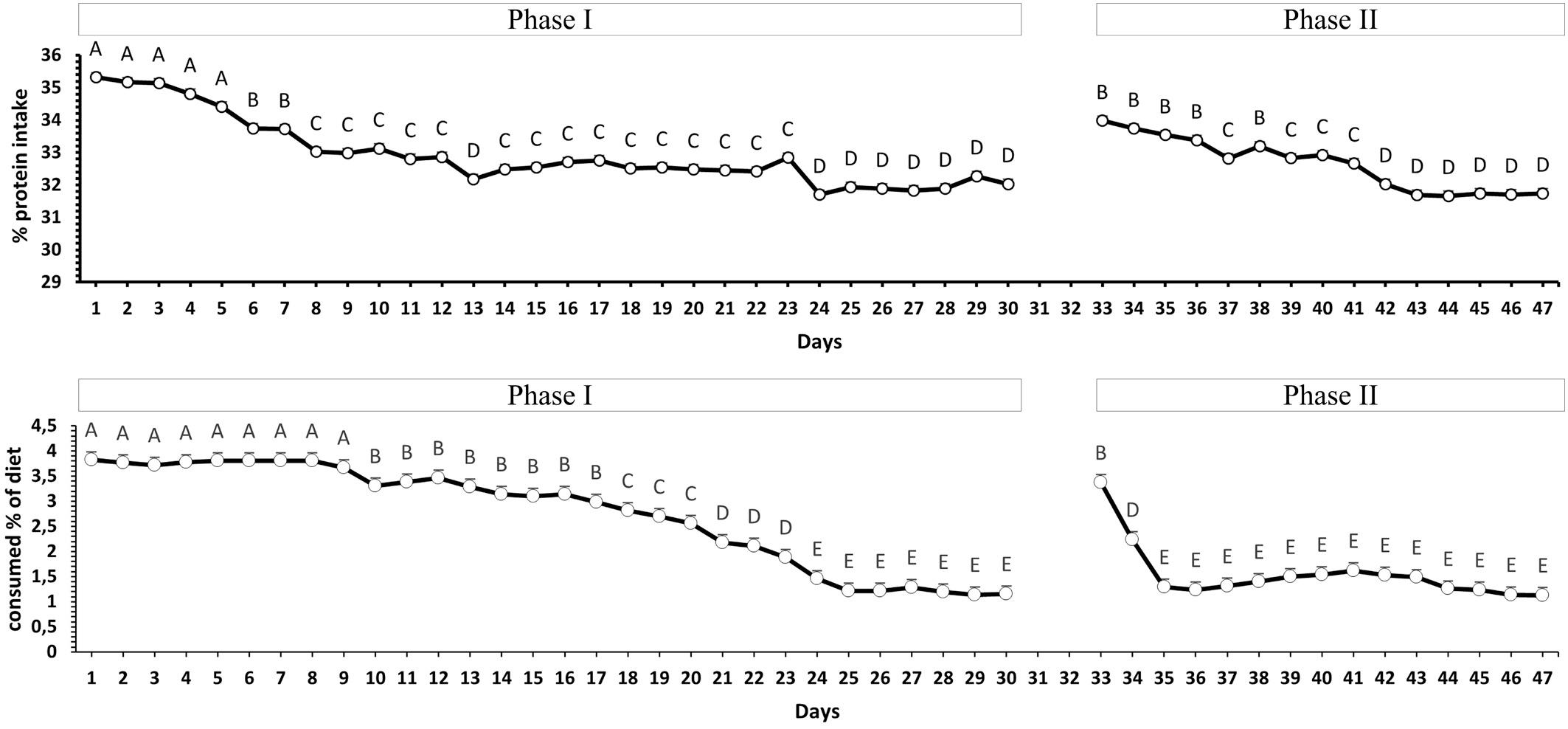
The average daily percentage of protein ingested by the animal was calculated and presented in Fig. 5A. Tilapia could adjust protein consumption during the experimental period. From the beginning of the experiment until the fifth day, the animals consumed the highest protein, with a daily average of 34.97%. This value decreased, with
-CD: Negative control capsule, filled only with cellulose;
-D25: Low protein capsule, filled with experimental diet containing 25% crude protein;
-D42: High protein capsule, filled with experimental diet containing 42% crude protein.
average daily consumption of 32.79% between the 6th and 23rd day, remaining stable between the 24th and 30th day of the experiment with an average consumption of 31.97% of crude protein in this period. In the second phase of the experiment, when the color of the diet’s capsules was alternated, the animals consumed the highest protein content between the 33rd and 38th days, with an average of 33.66% of crude protein. However, this value decreased until consumption stabilized from the 42nd day onwards, remaining at that level until the end of the experiment with an average consumption of 31.76% of crude protein, a value like that of the final of the first phase.
Fig. 5B shows the average daily feed consumption rate to the animal’s live weight. At the beginning of the experiment, there was a higher diet consumption, and this consumption decreased until it stabilized. The highest feed consumption occurred from the beginning of the experiment until the 9th day, with an average of 3.77% of the biomass. Between the 10th and 23rd days, consumption was 2.86% of the biomass, reaching stability between the 24th and 30th days of the
Fig. 2. Daily frequency of feeding, expressed as percentage of total diet consumption (Mean ± SEM), during 33 days of the experiment: G1 = D25 vs CD; G2 = D42 vs CD. The contents of the capsules were inverted after 16 days to verify that the color of the capsule did not influence consumption. Asterisk represents significant differences by Student’s t-test (p < 0.05, n = 10).
-CD: Negative control capsule, filled only with cellulose;
-D25: Low protein capsule, filled with experimental diet containing 25% crude protein;
-D42: High protein capsule, filled with experimental diet containing 42% crude protein.
experiment, staying at that level until the first phase’s end. During this period, the average feed consumption was 1.24% of the animal’s biomass. In the first two days of the second phase of the experiment, the animals increased consumption (2.80% of live weight). However, from the 3rd day of the second phase, which corresponds to the 35th day of the experiment, the consumption was adjusted and remained with an average of 1.35% of the biomass until the end of the experiment.
4. Discussion
Few works in the literature report the feeding behavior of fish using encapsulated diets, and in most of them, the study was done with purified diets. Supplying food inside gelatin capsules allows to isolate the effect of palatability and to evaluate if the animals can choose the food through pre- and post-ingestion signals. In our study, tilapia assimilated the color of the capsules as a pre-ingestive signal and the quality of their internal content as a post-ingestive signal, for, during experiment 1, significantly higher consumption of D25 and D42 diets was observed to the detriment of DC, even after changing the color of the capsule in the second phase of the experiment (Fig. 2). This identification ability was evidenced in studies with European sea bass (Dicentrarchus labrax) (Rubio et al., 2003), sharpsnout seabream (Diplodus puntazzo) (AlmaidaPag´ an et al., 2006), Nilotic tilapia (Oreochromis niloticus) (Fortes-Silva et al., 2011a; Fortes-Silva et al., 2012; Almeida et al., 2021), tambaqui (Colossoma macropomum) (Filho et al., 2018; Pereira et al., 2018); and pacama (Lophiosilurus alexandri) (Santos et al., 2019).
Despite the significant influence of palatability, fish also use indirect factors such as the physiological state to stimulate the consumption of some foods (Higuera, 2001), which may have driven the consumption of D25 and D42 diets to the detriment of DC, as the control diet consisted of cellulose, which does not provide nutrients to the animals, resulting in a decrease in the supply of nutrients to the organism. In studies where the effect of palatability was isolated, the post-ingestive signs indicate to the animal the quality of the ingested diet. Gastrointestinal receptors initially make identification of protein intake during protein digestion (Rubio et al., 2003; Almaida-Pag´ an et al., 2006; Fortes-Silva et al., 2011a), and the amino acids absorbed after this process are recognized by receptors present in liver tissue (Bernardis and Bellinger, 1996). Hepatic receptors communicate with brain centers via neurotransmitters or hormones, informing the nutritional properties of food and
Fig. 3. Final consumption of capsules (Mean ± SEM) of groups D25 vs DC and D42 vs DC, expressed as percentage of total intake, during 33 days of the experiment. Asterisk represents a significant difference in consumption within the same group (p < 0.05, n = 10).
Fig. 4. Daily frequency of feeding, expressed as percentage of total diet consumption (Mean ± SEM), during 47 days of the experiment. Provision of D25 vs D42. The contents of the capsules were inverted after 30 days to verify that the color of the capsule did not influence consumption. Asterisk represents significant differences by Student’s t-test (p < 0.05, n = 15).
-CD: Negative control capsule, filled only with cellulose;
-D25: Low protein capsule, filled with experimental diet containing 25% crude protein;
-D42: High protein capsule, filled with experimental diet containing 42% crude protein.
Fig. 5. (A) Average daily percentage of protein ingested by the animal during the 47 days of the experiment. The average daily percentage of protein was calculated by adding the amounts of protein consumed from each diet. Letters represent the significant difference between points by the Scott-Knott test (p < 0.05, n = 15). (B) Daily feed consumption rate to biomass during the 47 days of the experiment. The feed consumption rate was calculated as a percentage of feed consumed to live weight.
modifying the animal’s eating behavior (Forbes, 2001).
Protein and amino acids are vital for the functioning of the animal organism (Tu et al., 2015). The level of restriction of these nutrients, in this study, may have influenced the time taken by the animal to identify the internal content of the capsules since fish that were challenged with D25 vs. CD were able to distinguish the D25 capsules faster (Fig. 2A) significantly increasing its consumption compared to treatment D42 vs. DC (Fig. 2B), which showed a later significant difference in consumption. The D25 diet was formulated with protein deficiency and some essential amino acids such as histidine, methionine, threonine, and valine (Fig. 1), as indicated by the NRC (2011) To date, no studies have been found on the influence of protein concentration on the time of identification of a diet by animals.
Terrestrial animals show an appetite for specific nutrients (Forbes, 1998). A deficiency of these essential nutrients is known to reduce feed intake in fish (Kim et al., 1992a; Walton et al., 1986; Choo et al., 1991; Kim et al., 1992b). When animals are fed deficient diets, two opposite
responses can be induced: when the deficiency is mild, an increase in feed intake to reach the absolute level of requirement – as observed in poultry and swine regarding essential amino acids (Boorman, 1979; Henry, 1985), or an inhibitory response when the deficiency is more significant (Higuera, 2001). There was no significant difference in total consumption between the D25 and D42 capsules in our study. However, when we analyzed the total consumption of capsules within the treatments D24 vs. DC and D42 vs. DC (Fig. 3), we noticed that the animals consumed significantly more D25 and D42 capsules, thus increasing nutrient intake to meet their body’s needs.
In the second experiment, after adjusting for consumption, the animals ingested an average of 55.6% of D25 and 44.4% of D42 (Fig. 4). The adjustment also occurred to the crude protein intake and the feed consumption rate, resulting in 31.86% and 1.36%, respectively (Fig. 5A and B) after stabilization. Other studies have also reported this eating behavior of stabilizing the consumption of diets or nutrients (Boujard and M´ edale, 1994; Fortes-Silva et al., 2012; Santos et al., 2019). The way
fish regulate the intake and use of macronutrients is essential for aquaculture, as the animal can adjust the nutrient concentration in the body through variation in food intake (Atienza et al., 2004).
Feed selection by animals aims to satisfy nutritional requirements, although palatability also plays an important role (Higuera, 2001). A balanced nutrient intake is the main objective of feed selection. Animals can select, from a range of available rations, different proportions of each to ensure adequate intake of nutrients and energy, essential for normal bodily functions and growth (Simpson and Raubenheimer, 1999). In higher vertebrates, feed selection does not appear to be random or purposeless; it seems to aim to achieve a certain nutrient intake level (Kyriazakis, 1997). A similar fact was observed in our study. The animals were offered two rations, with different crude protein and amino acids levels, one deficient and the other with excess. The animal showed a significant preference for the consumption of D25. However, part of the diet consumed was composed of D42, which possibly supplied nutrients in low levels on D25.
Diets with a high protein concentration can decrease consumption and affect growth in some fish species (Gurure et al., 1995; Santinha et al., 1996). Such adverse effects have also been observed in higher vertebrates fed diets containing high protein and amino acids levels (Li and Anderson, 1983). The reduction in feed consumption by animals, in this situation, may work as a strategy to reduce the possible harmful effects of excessive nutrient intake (Higuera, 2001). In this study, the animals adjusted their daily protein intake by consuming different proportions of each diet and reducing, at the end of the experiment, the total food consumption. The average percentage of crude protein in the food ingested by the animals after the selection stabilization period was approximately 32% (Fig. 5A). This value is close to that recommended by the NRC (2011), which indicates a level of roughly 34% of crude protein, and by Santos et al. (2020), who report better performance of tilapia fed diets containing 31.12% crude protein. It should be emphasized that preferences do not need to coincide with a sensible and balanced diet formulated to provide maximum growth and efficient feed conversion (Higuera, 2001).
Feed consumption by fish is affected by climate, water temperature, dissolved oxygen, nutrient levels, and dietary energy (Higuera, 2001; Zhou et al., 2018; Mallekh et al., 2003). Our work observed that the feed consumption rate decreased throughout the experiment and stabilized at 1.36% of the biomass. Values similar to those found in our study were reported by Omasaki et al. (2017) (1.54%) and Tran-Duy et al. (2008), who reported consumption of 1.54 and 1.24% of the biomass, respectively. We emphasize that our research was carried out in a water recirculation system, with temperature, water quality, and photoperiod control, therefore, without these variables interfering in the food consumption rate.
5. Conclusion
Given the above, we conclude that Nilotic tilapia (O. niloticus) can select a diet that best meets its nutritional needs, and low-protein diets provide faster response in the selection process. The animals were able to adjust the consumption of crude protein by 31.86% through the selection of diets, with a consumption rate of 1.36% of the biomass, these values being recommended in the production of tilapia in a water recirculation system.
CRediT authorship contribution statement
Leandro S. Costa: Conceptualization, Methodology, Validation, Formal analysis, Investigation, Writing – original draft, Writing – review & editing, Visualization. Helder O. Guilherme: Conceptualization, Methodology, Validation, Formal analysis, Investigation. Raphael N. Bahiense: Conceptualization, Methodology, Validation, Formal analysis, Investigation. F ´ abio A.C. Santos: Conceptualization, Methodology, Validation, Formal analysis, Investigation. Pedro G. Gamarano:
Conceptualization, Methodology, Validation, Formal analysis, Investigation. Jose F.L. Olmeda: Conceptualization, Methodology, Validation, Formal analysis, Investigation. Paula A.P. Ribeiro: Conceptualization, Methodology, Validation, Formal analysis, Investigation, Resources, Data curation, Investigation, Writing – original draft, Writing – review & editing, Visualization, Supervision, Project administration, Funding acquisition. Ronald K. Luz: Conceptualization, Methodology, Validation, Formal analysis, Investigation, Funding acquisition.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We are grateful for the financial support granted to this research by CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnol ´ ogico - CNPq 401117 / 2014-6) and by CAPES (Coordenaçao de Aperfeiçoamento de Pessoal de Nível Superior), as well as the research grants granted to professors Ronald Kennedy Luz (CNPq n.308547/2018-7) and Paula Adriane Perez Ribeiro (CNPq n.308684/2017-6). We would also like to thank Departamento de Zootecnia da Universidade Federal de Lavras (UFLA), especiallyh Priscila Vieira Rosa, for facilitating access to the Enzimologia and Biologia Molecular laboratories.
References
Abd El-Wahab, A., Visscher, C., Teitge, F., Steinhagen, D., 2020. Choice preference of diets with different protein levels depending on water temperature in Nile tilapia. J. World Aquacult. Soc. 51, 512–526. https://doi.org/10.1111/jwas.12651
Almaida-Pagan, P.F., Rubio, V.C., Mendiola, P., De Costa, J., Madrid, J.A., 2006. Macronutrient selection through post-ingestive signals in sharpsnout seabream fed gelatine capsules and challenged with protein dilution. Physiol. Behav. 88, 550–558. https://doi.org/10.1016/j.physbeh.2006.05.027
Almeida, C.A.L., de Almeida, C.K.L., de Fatima Ferreira Martins, E., Bessonart, M., Pereira, R.T., Paulino, R.R., Fortes-Silva, R., 2021. Coping with suboptimal water temperature: modifications in blood parameters, body composition, and postingestive-driven diet selection in Nile tilapia fed two vegetable oil blends. Animal 15. https://doi.org/10.1016/j.animal.2020.100092
Atienza, M.T., Chatzifotis, S., Divanach, P., 2004. Macronutrient selection by sharp snout seabream (Diplodus puntazzo). Aquaculture 232, 481–491. https://doi.org/10.1016/ S0044-8486(03)00468-X
Babaei, S., S´ aez, A., Caballero-Solares, A., Fern´ andez, F., Baanante, I.V., Met ´ on, I., 2017. Effect of dietary macronutrients on the expression of cholecystokinin, leptin, ghrelin and neuropeptide Y in gilthead sea bream (Sparus aurata). Gen. Comp. Endocrinol. 240, 121–128. https://doi.org/10.1016/j.ygcen.2016.10.003
Bernardis, L.L., Bellinger, L.L., 1996. The lateral hypothalamic area revisited: ingestive behavior. Neurosci. Biobehav. Rev. 20 (2), 189–287
Bertucci, J.I., Blanco, A.M., Sundarrajan, L., Rajeswari, J.J., Velasco, C., Unniappan, S., 2019. Nutrient regulation of endocrine factors influencing feeding and growth in fish. Front. Endocrinol. (Lausanne). 10, 1–17. https://doi.org/10.3389/ fendo.2019.00083
Boorman, K.N., 1979. Regulation of protein and amino acid intake. In: Boorman, K.N., Freeman, B.M. (Eds.), Food Intake Regulation in Poultry. Longman, Edinburgh, pp. 87–126
Boujard, T., Medale, F., 1994. Regulation of voluntary feed intake in juvenile rainbow trout fed by hand or by self-feeders with diets containing two different protein/ energy ratios. Aquat. Living Resour. 7, 211–215. https://doi.org/10.1051/alr: 1994023
Choo, P.S., Smith, T.K., Cho, C.Y., Ferguson, H.W., 1991. Dietary excesses of leucine influence growth and body composition of rainbow trout. J. Nutr. 121, 1932–1939. https://doi.org/10.1093/jn/121.12.1932
Comesana, S., Velasco, C., Ceinos, R.M., L ´ opez-Patino, M.A., Míguez, J.M., Morais, S., Soengas, J.L., 2018. Evidence for the presence in rainbow trout brain of amino acidsensing systems involved in the control of food intake. Am. J. Phys. Regul. Integr. Comp. Phys. 314, R201–R215. https://doi.org/10.1152/ajpregu.00283.2017
Comesana, S., Conde-Sieira, M., Velasco, C., Soengas, J.L., Morais, S., 2020. Oral and preabsorptive sensing of amino acids relates to hypothalamic control of food intake in rainbow trout. J. Exp. Biol. 223 https://doi.org/10.1242/jeb.221721.
de Mattos, B.O., Nascimento-Filho, E.C.T., dos Anjos-Santos, A., S´ anchez-V´ azquez, F.J., Fortes-Silva, R., 2016. Daily self-feeding activity rhythms and dietary self-selection of pirarucu (Arapaima gigas). Aquaculture 465, 152–157. https://doi.org/10.1016/j. aquaculture.2016.09.005
Filho, E.C.T.N., Mattos, B.O., Santos, A.A., Barreto, K.A., Albinati, R.C.B., Oliveira Vidal, L.V., Fortes-Silva, R., 2018. Geometric approach to evaluate the energy/
L.S. Costa
protein balance in tambaqui (Colossoma macropomum): can fish ensure nutritional intake targets by postingestion signals? Aquac. Nutr. 24, 741–747. https://doi.org/ 10.1111/anu.12602
Forbes, J.M., 1998. Voluntary Food Intake and Diet Selection in Farm Animals. International Oxford, Cabi Forbes, J.M., 2001. Consequences of feeding for future feeding. Comp. Biochem. Physiol. - A Mol. Integr. Physiol. 128, 461–468. https://doi.org/10.1016/s1095-6433(00) 00328-7
Fortes-Silva, R., Martínez, F.J., S´ anchez-V´ azquez, F.J., 2011a. Macronutrient selection in Nile tilapia fed gelatin capsules and challenged with protein dilution/restriction. Physiol. Behav. 102, 356–360. https://doi.org/10.1016/j.physbeh.2010.11.036
Fortes-Silva, R., S´ anchez-V´ azquez, F.J., Martínez, F.J., 2011b. Effects of pretreating a plant-based diet with phytase on diet selection and nutrient utilization in European sea bass. Aquaculture 319, 417–422. https://doi.org/10.1016/j. aquaculture.2011.07.023.
Fortes-Silva, R., Rosa, P.V., Zamora, S., S´ anchez-V´ azquez, F.J., 2012. Dietary selfselection of protein-unbalanced diets supplemented with three essential amino acids in Nile tilapia. Physiol. Behav. 105, 639–644. https://doi.org/10.1016/j. physbeh.2011.09.023
Fortes-Silva, R., Oliveira, I.E., Vieira, V.P., Winkaler, E.U., Guerra-Santos, B., Cerqueira, R.B., 2016. Daily rhythms of locomotor activity and the influence of a light and dark cycle on gut microbiota species in tambaqui (Colossoma macropomum). Biol. Rhythm. Res. 47, 183–190. https://doi.org/10.1080/09291016.2015.1094972
Fortes-Silva, R., Costa, L.S., Paulino, R.R., Rodrigues, E.J.D., Sanchez-Vazquez, F.J., Rosa, P.V., 2017. Hybrid fish model (Pseudoplatystoma reticulatum♀ × Leiarius marmoratus♂) to study feeding behaviour: protein source self-selection and demandfeeding rhythms. Aquac. Res. 48, 3705–3718. https://doi.org/10.1111/are.13196
Geurden, I., Cuvier, A., Gondouin, E., Olsen, R.E., Ruohonen, K., Kaushik, S., Boujard, T., 2005. Rainbow trout can discriminate between feeds with different oil sources. Physiol. Behav. 85, 107–114. https://doi.org/10.1016/j.physbeh.2005.03.010
Geurden, I., Corraze, G., Boujard, T., 2007. Self-feeding behaviour of rainbow trout, Oncorhynchus mykiss, offered diets with distinct feed oils. Appl. Anim. Behav. Sci. 108, 313–326. https://doi.org/10.1016/j.applanim.2006.12.006
Gurure, R.M., Moccia, R.D., Atkinson, J.L., 1995. Optimal protein requirements of young Arctic charr (Salvelinus alpinus) fed practical diets. Aquac. Nutr. 1, 227–234 Henry, Y., 1985. Dietary factors involved in feed intake regulation in growing pigs: a review. Livest. Prod. Sci. 12, 339–354. https://doi.org/10.1016/0301-6226(85) 90133-2.
Higuera, M., 2001. Effects of nutritional factors and feed characteristics on feed intake. In: Food Intake in Fish, 827, pp. 250–268
Kabir, K.A., Verdegem, M.C.J., Verreth, J.A.J., Phillips, M.J., Schrama, J.W., 2019. Effect of dietary protein to energy ratio, stocking density and feeding level on performance of Nile tilapia in pond aquaculture. Aquaculture 511, 634200. https://doi.org/ 10.1016/j.aquaculture.2019.06.014
Kim, K. Il, Kayes, T.B., Amundson, C.H., 1992a. Requirements for sulfur amino acids and utilization of D-methionine by rainbow trout (Oncorhynchus mykiss). Aquaculture 101, 95–103. https://doi.org/10.1016/0044-8486(92)90235-D
Kim, K. Il, Kayes, T.B., Amundson, C.H., 1992b. Requirements for lysine and arginine by rainbow trout (Oncorhynchus mykiss). Aquaculture 106, 333–344. https://doi.org/ 10.1016/0044-8486(92)90265-M
Kyriazakis, I., 1997. The nutritional choices of farm animals: To eat or what to eat? In: Forbes, J.M., Lawrence, T.L.J., Rodway, R.G., Varley, M.A. (Eds.), Animal Choices. British Society of Animal Science, Edinburgh, pp. 55–65 Li, E.T.S., Anderson, G.H., 1983. Amino acids in the regulation of food intake. In: Nutrition Abstract and Reviews in Clinical Nutrition, 53, pp. 169–178 Luz, R.K., Boluda-Navarro, D., S´ anchez-V´ azquez, F.J., Fortes-Silva, R., Portella, M.C., Zamora Navarro, S., Madrid, J.A., 2018. European seabass (Dicentrarchus labrax) ability to discriminate between diets made with different dietary fat sources. Aquac. Nutr. 24, 65–73. https://doi.org/10.1111/anu.12534
Mallekh, R., Lagard` ere, J.P., Eneau, J.P., Cloutour, C., 2003. An acoustic detector of turbot feeding activity. Aquaculture 221, 481–489. https://doi.org/10.1016/S00448486(03)00074-7.
Morais, S., 2017. The physiology of taste in fish: potential implications for feeding stimulation and gut chemical sensing. Rev. Fish. Sci. Aquac. 25, 133–149. https:// doi.org/10.1080/23308249.2016.1249279
National Research Council (NRC), 2011. Nutrient Requirement of Fish. Committee on Animal Nutrition, Board on Agriculture, National Academic Press, Washington, DC, pp. 102–134
Omasaki, S.K., Janssen, K., Besson, M., Komen, H., 2017. Economic values of growth rate, feed intake, feed conversion ratio, mortality and uniformity for Nile tilapia. Aquaculture 481, 124–132. https://doi.org/10.1016/j.aquaculture.2017.04.013
Otero-Rodino, C., Libran-Perez, M., Velasco, C., Lopez-Patino, M.A., Míguez, J.M., Soengas, J.L., 2015. Evidence for the presence of glucosensor mechanisms not dependent on glucokinase in hypothalamus and hindbrain of rainbow trout (Oncorhynchus mykiss). PLoS One 10. https://doi.org/10.1371/journal. pone.0128603
Otero-Rodino, C., Libr´ an-P´ erez, M., Velasco, C., Alvarez-Otero, R., L ´ opez-Patino, M.A., Míguez, J.M., Soengas, J.L., 2016. Glucosensing in liver and Brockmann bodies of rainbow trout through glucokinase-independent mechanisms. Comp. Biochem. Physiol. Part - B Biochem. Mol. Biol. 199, 29–42. https://doi.org/10.1016/j. cbpb.2015.09.008
Pereira, R.T., Paulino, R.R., de Almeida, C.A.L., Rosa, P.V., Orlando, T.M., FortesSilva, R., 2018. Oil sources administered to tambaqui (Colossoma macropomum): growth, body composition and effect of masking organoleptic properties and fasting on diet preference. Appl. Anim. Behav. Sci. 199, 103–110. https://doi.org/10.1016/ j.applanim.2017.10.004
Rubio, V.C., Sanchez-Vazquez, F.J., Madrid, J.A., 2003. Macronutrient selection through postingestive signals in sea bass fed on gelatine capsules. Physiol. Behav. 78, 795–803. https://doi.org/10.1016/S0031-9384(03)00082-9
Sanchez-Vazquez, F.J., Martínez, M., Zamora, S., Madrid, J.A., 1994. Design and performance of an accurate demand feeder for the study of feeding behaviour in sea bass, Dicentrarchus labrax L. Physiol. Behav. 56, 789–794. https://doi.org/10.1016/ 0031-9384(94)90244-5
Sanchez-Vazquez, F.J., Madrid, J.A., Zamora, S., 1995. Circadian rhythms of feeding activity in sea bass, Dicentrarchus labrax L.: dual phasing capacity of diel demandfeeding pattern. J. Biol. Rhythm. 10, 256–266. https://doi.org/10.1177/ 074873049501000308
Santinha, P.J.M., Gomes, E.F.S., Coimbra, J.O., 1996. Effects of protein level of the diet on digestibility and growth of gilthead sea bream, Sparus aurata L. Aquac. Nutr. 2, 81–87
Santos, F.A.C., Fortes-Silva, R., Costa, L.S., Luz, R.K., Guilherme, H.O., Gamarano, P.G., Ribeiro, P.A.P., 2019. Regulation of voluntary protein/energy intake based practical diet composition for the carnivorous neotropical catfish Lophiosilurus alexandri Aquaculture 510, 198–205. https://doi.org/10.1016/j.aquaculture.2019.05.038
Santos, W.M., Costa, L.S., L ´ opez-Olmeda, J.F., Costa, N.C.S., Santos, F.A.C., Gamarano, P. G., Ribeiro, P.A.P., 2020. Effects of dietary protein levels on activities of protease and expression of ingestion and protein digestion-related genes in Nile tilapia juveniles. Aquac. Res. 51, 2973–2984. https://doi.org/10.1111/are.14635
Simpson, S.J., Raubenheimer, D., 1999. Assuaging nutritional complexity: a geometrical approach. Proc. Nutr. Soc. 58, 779–789. https://doi.org/10.1017/ S0029665199001068
Simpson, S.J., Raubenheimer, D., 2001. A framework for the study of macronutrient intake in fish. Aquac. Res. 32, 421–432. https://doi.org/10.1046/j.13652109.2001.00593.x
Tran-Duy, A., Schrama, J.W., van Dam, A.A., Verreth, J.A.J., 2008. Effects of oxygen concentration and body weight on maximum feed intake, growth and hematological parameters of Nile tilapia, Oreochromis niloticus Aquaculture 275, 152–162. https:// doi.org/10.1016/j.aquaculture.2007.12.024
Tu, Y., Xie, S., Han, D., Yang, Y., Jin, J., Liu, H., Zhu, X., 2015. Growth performance, digestive enzyme, transaminase and GH-IGF-I axis gene responsiveness to different dietary protein levels in broodstock allogenogynetic gibel carp (Carassius auratus gibelio) CAS III. Aquaculture 446, 290–297. https://doi.org/10.1016/j. aquaculture.2015.05.003
Vivas, M., Rubio, V.C., S´ anchez-V´ azquez, F.J., Mena, C., García García, B., Madrid, J.A., 2006. Dietary self-selection in sharpsnout seabream (Diplodus puntazzo) fed paired macronutrient feeds and challenged with protein dilution. Aquaculture 251, 430–437. https://doi.org/10.1016/j.aquaculture.2005.06.013
Walton, M.J., Cowey, C.B., Coloso, R.M., Adron, J.W., 1986. Dietary requirements of rainbow trout for tryptophan, lysine and arginine determined by growth and biochemical measurements. Fish Physiol. Biochem. 2, 161–169. https://doi.org/ 10.1007/BF02264084.
Zhou, C., Xu, D., Lin, K., Sun, C., Yang, X., 2018. Intelligent feeding control methods in aquaculture with an emphasis on fish: a review. Rev. Aquac. 10, 975–993. https:// doi.org/10.1111/raq.12218
Other documents randomly have different content
one of my kinsfolk.” He failed to obtain the Priory at once, though he made repeated efforts. On June 27th of this year he wrote[137]: “Though your suit for the Priory of St. Thomas in my behalf cannot stand, yet as you mind my preferment to the farm of the demesnes, I thank you. I desire them only for quietness, not for advantage”; and he wrote again on April 3rd, 1537.
The Priory of Stone contained many tombs of the Staffords, and Lord Stafford evidently hoped the house would escape. But the glory of his family had departed and he had no real influence. The Prior was William Smith, and he does not appear to have had any suspicion that his house was soon to come to an end. Even while the Visitors were making their investigations, if, indeed, any investigation at all was made in the great majority of cases, he was engaged in the business of his house. In his financial transactions with his Bishop he found the latter more worldly-wise than he was himself. Bishop Roland Lee sold him timber out of Blore Park and received the payment. But, being better informed of the trend of events, he prevented many of the trees from being felled and delivered to the dying Priory. On February 19th William Smith wrote urgently to Lee, [138] “Touching the timber in Blore Park which I bought and paid for to my lord, 40 trees are still standing, as the bearer can show. If I have not the said timber I know not where to be provided for my great work now in hand. I shall intreat you for your pains.” Several months later, Henry Lord Stafford wrote to Cromwell telling him “that the Prior of Stone hathe good hope that his howse schall stand, whereof all the contree is right glad, and praye fulle hertily for your lordeship therfore.” The Earl of Shrewsbury, however, had designs on it, and sought the assistance of Scudamore in obtaining it, bringing himself to address his letter “To my hertly biloved fellow John Skydmore, oon of the gentylmen vsshers of the Kynge’s most honourable Chamber.”
In these circumstances there was much uncertainty as to the extent to which the Act would literally and fully be carried out, and how far influence might succeed in nullifying it.
In due course another band of royal agents was let loose upon the land to carry out the work of dissolution. The “Instructions for the King’s Commissioners” are exceedingly minute. For each county an Auditor and Receiver was to be appointed, with one of the clerks of the late visitation, and to these were to be joined “three other discreet persons to be named by the King.” These were to visit each condemned house and exhibit the Statute of Dissolution to the head and his brethren. The inmates were then to be required to make on oath a full disclosure of the state of their affairs, to surrender their charters and seal, plate, and other effects. Such of the monks as were willing to take “capacities” were to be referred to the Archbishop of Canterbury and the Lord Chancellor, and were to be rewarded for their complacency; the rest were to be transferred to other houses. Orders were to be given that the monks were to cease receiving any income except such as was absolutely necessary, but they were to continue “to sow and till their grounds as they have done before,” and the Superior was referred to “the Chancellor of the Augmentation for his yearly stipend and pension.” But evidently the agents were prepared to accept favourable offers. Henry Lord Stafford, writing to Cromwell on March 12th, 1537, says: “The Commissioners will be in Staffordshire on Sunday next. The Prior of Stone thinks his house shall stand, whereof the country is glad; so my suit is in vain unless your Lordship help me to the Priory of Rontone, for which I was first suitor: howbeit Sir Simon Harcourt makes great labour for it”; and he proceeds again to plead his poverty and his large family, mentioning that he had twelve children. [139]
The Commissioners in Staffordshire dissolved three out of the nine houses which came within the scope of the Act, namely Ronton, Stone, and Trentham. The majority were spared, some, as we have seen, through the intervention of powerful or interested friends, but all on payment of large sums.[140] For instance, Hulton, having paid £66 13s. 4d., obtained a grant of exemption on October 1st, 1536, the Abbot being Edward Wilkyns; Rocester’s grant was made on March 11th, 1537, and its payment £100; Tutbury, per Arthur
Meverell, received its license on May 3rd; Croxden, per Thomas Chawner, on the 2nd of July, by payment of £100; St. Thomas’, Stafford, per Richard Whyttewall, on July 4th, by payment of £133 6s. 8d. On April 3rd, Robert Burgoyn had written to the Bishop, who so earnestly had desired the Priory: “According to your desire ... I have forwarded letters from the Chancellor of the Augmentations to Mr. Scudamore to survey the lead of the late house of Canons beside Stafford,”[141] which shows how narrowly the house escaped, even for a time, the clutches of Roland Lee and Cromwell, for though he spoke of it as “the late house,” it did not surrender till October, 1538. Lee kept up his persistent begging for it to the end.
The Grant of Exemption obtained by Croxden is given in the Appendix. It gives the King’s new title, “SupremumCaputAnglicanae Ecclesiae,” in its most offensive form, without the qualifying words, but it will be noticed that it is couched throughout in general terms. There is nothing in it which is peculiar to the particular house concerned. Everything would apply equally well to any other house. It looks as though the Commissioners went on their circuit provided with a supply of such general forms, having blanks for the names of houses and of abbots, which they were prepared to issue whenever they thought fit, that is, whenever a sufficiently large pecuniary inducement could be offered. That there was no genuine intention to allow any of the religious houses to continue permanently was speedily shown.
But for the present the elaborate grants for continuance served their turn, and allayed public dismay. No echo of the Pilgrimage of Grace was heard in Staffordshire, although that dangerous rising began near the north-eastern border of the county. It rolled northwards, and Lord Stafford was so entirely satisfied that he carried out the difficult and costly process of transferring his family monuments from the dissolved Priory at Stone to the Austin Friary at Stafford. He evidently had no idea that Commissioners would destroy that house, too, in a few months.
It was, of course, entirely to the interest of the agents of the Dissolution to conceal the real scope of their intentions, for the threatened monks naturally tried sometimes to keep back part of their cherished possessions and to save the sacred vessels and vestments from the profane uses to which they were likely to be put. The Act of Dissolution had foreseen the danger and had made all sales and leases of lands effected “within one year next before the making of this Act,” “utterly void and of none effect”; while all ornaments, jewels, goods and chattels which the houses possessed “at the first day of March in the year of our Lord God 1535 or any time sithen whensoever” were declared the property of the King. The Prior of Stone and some of his neighbours, who, as we have seen, resented the suppression of the house, attempted to save something.
“Articles and instructions” for special commissioners were issued “concerning the embezzling and taking [away] of certain plate, jewels, ornaments, goods, and chattels of the late monastery of Stone ... whereupon as well William Smyth, late Prior of the said house, James Colyer, James Atkyn, Sampson Greswike, Geoffrey Walkeden, and Hugh Rathebone, and all such other persons as Thomas Woodall, bringer hereof, shall name, are to be examined.”[142] The specific charges are as follows, so far as they can be traced, for the document unfortunately exists in a fragmentary form only: “Whether Colyer received a shrine, four standing cups, and two silver salts; whether Atkyn received certain sheep and cattle since the said fourth day of February; whether Greswike since the fourth day of February hath of the said house ..., ” and there the paper ends. February 4th was the day on which the Session of Parliament which had passed the Act of Dissolution had opened.
In the Augmentation Accounts for 1538 we find “A parcel of £20 due from James Colyer for the surplus value of a shrine of silver-gilt mortgaged to him by the Prior of Stone, part of which was paid 5 of June, 30 Henry VIII, £13 6s. 8d.”[143] The Abbot of Dieulacres
secured blank forms with the Convent seal before it was taken from him, and on these he subsequently made out ante-dated leases. Bishop Lee reported to Cromwell that the Prior of St. Thomas at Stafford was making “unreasonable waste,” which probably means that he had realized the uselessness of economy in the face of imminent dissolution.
The fines for continuance were exceedingly heavy and must have pressed very hardly on the houses which were called upon to raise such large sums. They appear to have been roughly calculated at a year’s income,[144] and no doubt they account in no small measure for the indebtedness which subsequently was charged against some of the houses.
Moreover, the officials looked for bribes and presents, and we may be sure they required to be well entertained when they visited the monasteries, to judge from their own large expenditure on “cates.”[145] The houses were impoverished by direct methods as well. Prior Richard, of St. Thomas’s, Stafford, was ordered to give the lease of a church at Audlem in Cheshire, belonging to the Priory, to a nominee of Cromwell’s. He protested against the unfairness, though he was unable to avoid compliance. “It is,” he says,[146] “in the occupation of five poor farmers there by lease,” but he had to give Cromwell’s nominee a fifty years’ lease in reversion, in consideration of Cromwell’s “goodness,” and the lessee was to pay six shillings and eightpence, whereas they could have had 40 marks from another. He adds that last Midsummer he paid Cromwell £60 and now sends £20 more.
The same policy of crippling the Abbeys was pursued even towards Burton, which did not come under the Act. On August 15th, 1538, the Abbot of Burton-on-Trent wrote to Cromwell[147]: “On the 12th of August I received the King’s letters and yours in favour of Mr. Robert Everest, one of the servers of the Chambers, for the tithe of the parsonage of Allstrye, Warwickshire. That tithe is so necessary for our house that we cannot do without it, and was appropriated
under the broad seal of England because we had not corn sufficient for hospitality. You write that Sir Thomas Gresley, lately deceased, had it. But that is 34 years ago, and he only had it then because the Abbot was indebted to him.”
The following letter from the Abbot of Burton is addressed “to the Ryght Worshipful Maister Holcroft the Kynge comycyoner at Lenton delyver this:”[148]
“Mayster Holcroft I enterlye recomend me vnto you beseching God that I may once be able to surrendre vnto you condygne thanks for thys youre goodness wyche have dymynysshed parte of the charges wyche by yor (scored through) comyssyon you myght have put me to, And as touchyng youre request of this brother and the lame chylde, god wyllyng I shall so accomplysshe hyt as shall both please yor mastership & content the partyes beyng not only in this thyng but also in all other redy at my prynces comandement and to my small power shew yor mastershippe pleasure pryng you accordyng to yor w’tyng of good word and lawfull favor Thus oure lorde have yor mastershippe in his kepyng to his pleasure and youre comfort from Burton the xviiith day of Maye “Yors assuryd
“Willm Abbot there.”
The friaries for some time were left to themselves. They were poor and had few inmates, and their houses were not settled in pleasant situations, with broad estates reaching “to my lord’s park pale.” They were, indeed, within or near the walls of the towns, and, consequently, were of little interest to the aristocracy.
But their poverty was no permanent security. On February 6th, 1538, Dr. Ingworth, the renegade Prior of the richest house of the Black Friars in England, and lately made Suffragan Bishop of Dover, was commissioned to visit all the friaries, and he rapidly carried out his work. We have very full particulars of his campaign in Staffordshire, and some remarkable details. On August 7th he was at the house of the Grey Friars at Lichfield, on August 9th he decided the fate of both the friaries at Stafford, and next day he was equally effective at Newcastle-under-Lyme;[149] and this in spite of the fact that he was obliged to confess that “the Friars in these parts have
many favourers, and great labour is made for their continuance. Divers trust to see them set up again, and some have gone up to sue for them.” But he tells Latimer (on August 23rd) that such strong expressions of popular opinion had had no terrors for him; he had visited 18 places, including Lichfield, Stafford, and Newcastle, and had only left one house standing.[150]
The Staffordshire friaries were without exception poor places in every respect, and the Bishop cleverly made that a strong argument against them. In spite of poverty, the friars clung to their old homes and work, although he invariably offered them money payments to depart. Writing from Lichfield, he told Cromwell that “divers of the Friars are very loath to forsake their houses, and yet they are not able to live.” The house at Lichfield, for instance, he says, “is in that taking, and yet loath to give up.”[151] The day after he had been there he wrote Cromwell a full account of his visit.[152] He announced that in spite of their wishes he had induced them all to surrender. The warden was in a pitiable state of ill-health, with a loathsome disease on his face. He had been little at home for the past six months, “yet now he came home and was loath to give up his house, though it is more in debt than all the stuff that belongs to it will pay, chalice, bells, and all, by 20 nobles.” The certificate of surrender is dated August 7th.[153] It states that the house was surrendered voluntarily, without any counsel or constraining, for very poverty a manifest falsehood, as the Bishop’s own letters testify. The witnesses of this surrender were Richard Wetwode, “Master of the Guild there,” and the two constables, Alexander Grene and Thomas Lont. The Visitor delivered the house and goods to these three, gave every friar a letter, and departed. An inventory of the goods so delivered follows, and comprises articles in kitchen, brewhouse, choir, and sextry. There is also a statement of the debts owing by the house, which were partly for malt and rye, with 30 shillings which had been borrowed “for byldyng of the quere” and 20 shillings due to the Bishop for five years’ rent. Four days later Dr. Legh wrote to Cromwell, on his own account and also at the instance
of the Bishop, to both of whom Wetwode had shown “great pleasure,” asking that Wetwode should have the preferment.[154] It is evident that just as great lords and enterprising country squires were interested parties in the suppression of monasteries, so the rising tradesmen in the towns cast longing eyes on the houses of the friars. This accounts for the co-operation of the municipal authorities in the work of dissolution.
A very full and detailed account of the Bishop’s procedure in regard to the friars is given in a memorandum referring to the visitation of the two houses at Stafford. This most interesting and valuable document is as follows:[155]
“Mem. This 9 day of August in the 30 year of our most dred Sovereign lord King Henry VIII., Richard Bishop of Dover, visitor under the Lord Privy Seal for the King’s Grace, was in Stafford in the Grey Friars and also in the Austen Friars, where that the said Visitor said to the heads and brethren of both places these words: Brethren, where that I understand ye have had information that I should come, by the King’s Commission, to suppress your house and put you out, fear not, for I have no such commission, nor I use no such fashion in any place. I am sent to reform every man to a good order and to give injunctions for preservation of the same. If ye can be content and think yourself able here to live and to be reformed and to observe such reasonable injunctions as I shall leave with you, the which or that I require your answer, ye shall here and see in writing, then I am and shall be content that ye shall with the King’s favour continue as before ye have do. If that ye be not able to live and observe the same then if ye of your own minds and wills give your houses into the King’s hands I must receive them. The said injunctions were read to them which were reasonable. The said heads with all the brethren with one assent, without any counsel or co-action, gave their houses into the Visitor’s hands to the King’s use. The Visitor received the same, and of the houses and implements made inventories and delivered them to such as should keep them to the King’s use, and so delivered to each friar a letter to visit his friends and so departed. This witnesseth John Savage and Thos. Russell, Bailiffs of the borough of Stafford; Wm. Stamforde and Ric. Warde, gentlemen, with divers others.”
The mean trickery as well as absolute perversion of truth in the Bishop’s conduct and statement could not be better shown than in this interesting record. The wretched friars were already trembling for their own safety, as they saw the monks on every side dispossessed and impoverished, and impoverished themselves at any rate by the check which the events of recent years must inevitably have given to bequests and alms to all religious institutions. Disheartened by long uncertainty, they fell easy victims to the bullying and falsehoods of the plausible Visitor and his coadjutors, the vulgar and rich shopkeepers who accompanied him, the latter eager for the site and buildings, adjoining perhaps their own places of business, and certainly convenient for warehouses and storerooms. It was not till the spring of 1539 that Parliament passed an Act recognising the faitaccompliand giving the King all the religious houses.
The Inventory of the possessions of the Austin Friars at Stafford, which were placed in charge of William Stamforde, of Rowley, and Master Richard Warde, of Tylynton, is full of interesting details throwing much light on the ecclesiastical and domestic arrangements of the time.[156] In the vestry there was a cross of copper gilt “with an image silver of parcel gilt,” a copper censer, four “suits” (i.e., sets) of vestments, one black set for requiems, and one with “images” of the Blessed Virgin, two green copes, one black “chamlet,” etc. In the choir were two old altar-cloths, two small candlesticks, a sacring-bell, and a “pair of organs.” There were two bells in the steeple. In the church were two stained cloths, an alabaster table, two ladders, and two benches. The contents of hall, kitchen, recreation-house, etc., are also given. There was little or no lead, and the yearly rents amounted only to 51s. 8d. The Visitor took into his own keeping the chalice, which weighed 13 oz., and he ordered that the servants should be paid ten shillings of their wages at the next Michaelmas.
The Inventory of the house of the Grey Friars is similarly detailed.[157] In the sextry there were five “suits” without albs; a
suit for requiems, one each of dun silk, yellow sey, and branched green silk. There were six copes, two being of linen cloth “stained with image work.” There were six altar-cloths, a pyx of latten, etc. In the church were four alabaster tables, a pair of large candlesticks, a cross, and a censer of latten, two missals, one printed and one written, “a pair of small organs,” etc. There was much lead, for half the choir was leaded and one of the chapels. The rents only came to 26s. 8d. Again the chalice was taken by the Visitor, with six spoons: 16 oz. in all.
Next day he was at Newcastle-under-Lyme.[158] He found it owed £14, for which all its substance was in pledge, yet was insufficient to meet the debt. The Inventory was again signed by the town officials, John Lymforde, Mayor, and Thomas Brodsha and Richard Smyth. The Inventory of the goods showed that in the vestry there were “suits” of blue silk, of silk with roses, and of green silk. There were eleven chasubles, five copes, and two old tunicles. In the choir there were two pairs of candlesticks of copper and latten, one cross of copper and gilt with a “Mary and John.” “A pair of organs” is mentioned and an alabaster table on the High Altar. In the steeple were two bells. In the house were two old feather beds, one old bolster, and five old coverlets, an old chest, and a green covering of say. The usual articles are mentioned in kitchen, brewhouse, hall, and buttery. The choir and cloisters were roofed with lead, and the rents came to 40s. a year. The Bishop of Dover took possession of the chalice, which was a small one, five spoons, and “two narrow bands of masers” (14 oz.), but he sent up to Cromwell three boxes “of evidence,” one of the King’s, one of other gentlemen’s, and one of the Convent’s.
He wrote from Lichfield an account of his journey to that point, and supplemented it on August 13th by another letter written at Shrewsbury.[159] He apologises for not being able to send at once all the Inventories, but he had no leisure for such work, and, moreover, his servants were ill. Perhaps they could not stand the rapidity with which he travelled, “but I trust to se yower lordschype within a veke,
and be that tyme I trust to make an ende in all Walys.” He continues: “Sumwhat to certyfye yower lordeschype of the state off suche as I have receyveyd sythe that I wrote to yow towcheynge Stafforde, the Austen Fryeres ther ys a pore howse, with small implementes, no jwelles but on lytyll chales, no led in the howse, in rentes by yere lis. xiiid. The Graye Fryeres ther, halfe the quere ledeyd and a chapell, small implementes, no plate but a chales and vi. small sponys, in renttes xxvis. iiiid. The Blacke Fryeres in Newecastell Underlyne, all in ruyne, and a pore howse, the quere ledeyd and the cloeyster led redy to fall downe, the reste slate and schyngyll; in fermys by yere xls. On master Broke hathe of late fownde the menys with the prior to gett of hym the more parte of they howseys and grownde ther by iii. leseys, and that for lytyll money; he wolde a gyve me golde to a grantteyd to hys leseys, but I toke no peny of hym nor of non other, nor non woll. Iff he have thoys leseys there ys lytyll besyde, for he hathe lyberte allmost in all. Ther ys a proper wode, but he hathe all in lese. No sylver above xiii. ounce.” The property had been fully mortgaged to Mr. Broke, possibly, but not probably—for the general poverty of the house amply accounts for it—with a view to the threatened visit of Dr. Ingworth. But the Bishop’s elaborate assertion of his superiority to bribery must have been amusing reading to Cromwell: he “doth protest too much, methinks.” He was not chary of sycophancy, however. He proceeds, after describing the ruined state of the Austin Friars at Shrewsbury: “My synguler good lorde, I beseche youe pardon me of my rude wrytynge, and yf that I do not my dewte as I owte to do I beseche youe pardon me, for my hart and intente ys to do that thynge that shulde specyally plese God, the kynges grace, and yower lordschype, accordeynge to my dewte.” He then goes on to “beseche” his Lordship, the son of the drunken brewer, “that yf before my cumynge there be any order taken for Newecastell Underlyne, that ye wolde be good lorde to on Master Johan Bothe, a servant of the kynges graces, the whyche is a grett bylder in theys partes, that he myght for money have the slate and schyngyll ther; for ther ys no other to be don with the more parte of that howse, but save the lede and slate, and take the profete of the grownde.
That master Bothe for yower sake sheuyd me many plesures and gave me venyson; wherefor I may no lesse do but wryght to yower lordeschype besecheynge yow to be good lorde to hym, an I ever yower orator to Jhesu, who preserve yower lordschype.”
In this year Bishop Lee issued a series of injunctions to the clergy of the diocese.[160] In these he ordered sermons to be preached at least quarterly in “all monasteries.” There were few remaining by the end of the year, and only one—Burton-on-Trent, in Staffordshire. No monks or friars were to have any “cure or servyce,” “except they be lawfully dispensed withall or licensed by the ordinary.” Confessions to monks and friars were forbidden, though ordered to be made to parish priests, before Communion, and the wearing of secular dress was sternly condemned.
Cromwell also issued injunctions to the clergy, on September 5th, 1538,[161] ordering, among other things, “that such Images as ye know in any of your cures to be so abused with pilgrimages or offerings of anything made thereunto ye shall for avoiding of that most detestable offence of Idolatry furthwith take down and deley ... admonyishng your parishioners that Images serve for no other purpose but as to be bookes of unlearned men that can no letters.... And therefor the kinges highnes graciously tenderyng the weale of his subjectes sowles hath in parte alredy and more will hereafter travail for the abolishing of suche Images as might be occasion of so greate an offence to god and so gret daunger to the sowles of his loving subjectes.” No charge of superstition had been brought against the image of St. Modwen at Burton by Layton and Legh at their visitation, but none the less it was pulled down. Sir William Bassett, of Meynell Langley, a few miles from Burton, wrote as follows to Cromwell:[162] “Ryght honorabull my inesspeyciall gud lord, accordyng to my bownden dewte and the teynor of youre lordschypys lettres lately to me dyrectyd, I have sende unto yowre gud lordschyp by thys beyrer, my brother, Francis Bassett, the ymages off sentt Anne off Buxtone and sentt Mudwen of Burtun apon Trentt, the wych ymages I dyd take frome the place where
they dyd stande, and browght them to my owne howss within xlviiie howres after the contemplation of yowre seyd lordschypis lettres, in as soober maner as my lyttull and rude wytt wollde serve me. And ffor that there schullde no more idollatre and supersticion be there usyd, I dyd nott only deface the tabernaculles and placis where they dyd stande, butt allso dyd take away cruchys, schertes, and schetes, with wax offeryd, being thynges thatt dyd alure and intyse the yngnorantt pepull to the seyd offeryng; allso gyffyng the kepers of bothe placis admonicion and charge thatt no more offeryng schulld be made in those placis tyll the kynges plesure and yowre lordschypis be ffurther knowen in that behallf.... And, my lord, as concerning the opynion off the pepull and the ffonde trust that they dyd putt in those ymages and the vanyte of the thynges, thys beyrer my brother can telle yowre lordschyp much better att large then I can wryte, for he was with me att the doing of all.” The said Francis Bassett was in the service of Cranmer, and we shall meet with him again; “There cam nothyng with theym but the bare imagis.” Bishop Lee saved from the spoilers the jewels of St. Chad’s Shrine at the Cathedral for “necessary uses.” Prebendary Arthur Dudley was one of the authorized commissioners for holding such Church goods as were seized by the Crown, but he apparently reverenced holy things, and gave the bones of St. Chad to some female relatives of his. The latter handed them to two brothers named Hodgetts, and eventually some of them have been deposited in the Roman Catholic Cathedral at Birmingham. The shrine disappeared, and as the relics had gone the Cathedral was spared such sacrilege as was witnessed elsewhere.
CHAPTER VIII
THE GENERAL SUPPRESSION: SECOND STAGE
Meanwhile the harrying of the houses continued, and the feeling of uncertainty deepened. It became more and more evident that the whole monastic edifice was falling. The dissolution of the lesser monasteries and the sharing of their spoils had served the useful purpose of creating an appetite for more. On February 20th, 1538, Francis Lord Hastings wrote to Cromwell begging the Abbey of Burton, apologizing for not having written sooner, and explaining that he should have done so but that he had been suffering from measles.[163] On August 23rd, Cranmer wrote urging on Cromwell the suppression of Tutbury, and desiring that Commissions should be sent to Rocester and Croxden.[164] The three had paid large sums for their continuance only a year before, but Cranmer was interested in obtaining them, or one of them, for his servant, Francis Bassett. Again, on December 14th, he wrote begging for Croxden to be given to Bassett.[165]
Bishop Roland Lee had continued his pleading for the Priory at Stafford, and when time went by without seeming to bring him nearer obtaining it he began to suspect Legh of playing him false. The latter, however, assured him he was as interested as the Bishop himself in the matter, though he advised an application direct to Cromwell. “I have spoken,” he wrote,[166] “to Mr. Strete for the suppression of St. Thomas’s, but I would your lordship should write to my Lord Privy Seal (Cromwell) for your own matter, and to thank him, for he told me he would move the King for you and your heirs to have St. Thomas’s, and no doubt the King will be content, and, indeed, it is all one. Remember to write to my lord to put away sinister suspicion, and be not light of credit against me: mistrust without cause is very unpleasant.” Thus encouraged, Lee returned to the charge, adding fresh inducements. He suggests that the longer
the matter is delayed the less there will be to confiscate, “as the Prior makes unreasonable waste.” He asks that the Priory may be let to him “at an easy rent, that the poor boys, my nephews, may have some relief thereby”; and he begs that Cromwell will write to the surveyors that he may buy what things belonging to the house he desires.[167] The latter request he obtained. Again, on December 13th, he wrote to Cromwell on the same subject. He even ventured to apply direct to the King, both in person and by letter. There is a letter of his written to the King on December 26th, which says: “Where at my being with your Majesty, I moved the same for the late Priory of St. Thomas, I was minded to pay a certain sum as your Grace should determine. I am so much bound to your Majesty that I can crave no more; but, being charged with eight poor children of my sister’s, now fatherless and motherless, I am forced to show the truth.”[168]
Other petitions had flowed in, and gradually matters were arranged. Some houses were granted as they stood, sites, buildings, furniture and other contents, stores, animals, farm implements, etc., to a single recipient in return for a single money payment. No doubt influential petitioners like Bishop Roland Lee and Lord Derby (who obtained Dieulacres) met with less rivalry than more obscure suitors who made efforts for the smaller houses. There appears to have been keen competition for the little nunnery at Brewood.
When the houses and belongings were to be sold en bloc, Dr. Legh, with whom went William Cavendish as auditor, appears to have had the management, while Scudamore conducted the business where other arrangements had been made, and the contents of the houses were sold by public auction.
The religious houses found that their attempts to secure a further lease of life for themselves by authorized payments to the Royal Treasury, or by irregular bribes to Cromwell and his friends, had all failed. The large fines recently paid served to prolong the houses for a twelvemonth only, and as the autumn of 1538 drew on the news probably reached all the houses that they were doomed.
The harvest having been safely gathered in by the monks, in accordance with the royal injunctions, the royal agents began to close round them once more, and the last agony began.
John Scudamore was appointed “Receiver-General unto the King’s Majesty of the dissolved possessions” in Staffordshire and elsewhere. He received his authorization on August 23rd, 1538, from Sir Richard Riche, the Solicitor-General and Chancellor of Augmentations. His instructions were to survey all the lands of surrendered houses and to make a return of their yearly value, with such pensions and corrodies, etc., as they might be burdened with. The bells and superfluous houses were to be sold, and the lead melted into “plokes” and sows and marked with the Royal mark, and delivered under indenture to the constables of neighbouring castles. [169] On September 27th, the goods of both houses at Stafford were sold, on October 4th the friary at Lichfield, on the 15th Scudamore was at Croxden, and next day at Rocester; on the 21st he was at Hulton. The details of all these sales are extant and are full of interest.
The sales were evidently conducted in a most wasteful way, as Robert Burgoyne, who acted as auditor at the sales at Stafford, testified. He told Scudamore, “I have sold in some ffrire houses all the buyldynges, the cause was for that they were so spoyled and torne by suche as sold the goodes, that in manner they were downe, and yff they shuld nott have ben sold, the kyng shuld have hadd nothyng theroff.”[170]
Although there had been loud discontent at the threatened dissolutions of the friaries at Stafford, the townspeople did not hesitate to profit by the sales any more than the country people did in the case of the monasteries. They knew that the end of the alms and easy rents had come, and it was only human nature to make the best of the sorry business. The town bought the stone wall of the Grey Friars and a pair of candlesticks at Stafford; and the churchwardens a Corporas.[171] The “warden of the sayd [Grey] Fryers” bought two brass pots for eight shillings and six plates for