All rights reserved. No part of this publication may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permissions may be sought directly from Elsevier’s Rights Department: phone: (+1) 215 239 3804 (US) or (+44) 1865 843830 (UK); fax: (+44) 1865 853333; e-mail: healthpermissions@elsevier.com. You may also complete your request on-line via the Elsevier website at http://www.elsevier.com/permissions.
Notice
Knowledge and best practice in this field are constantly changing. As new research and experience broaden our knowledge, changes in practice, treatment and drug therapy may become necessary or appropriate. Readers are advised to check the most current information provided (i) on procedures featured or (ii) by the manufacturer of each product to be administered, to verify the recommended dose or formula, the method and duration of administration, and contraindications. It is the responsibility of the practitioner, relying on their own experience and knowledge of the patient, to make diagnoses, to determine dosages and the best treatment for each individual patient, and to take all appropriate safety precautions. To the fullest extent of the law, neither the Publisher nor the Authors assume any liability for any injury and/or damage to persons or property arising out of or related to any use of the material contained in this book.
The Publisher
International Standard Book Number: 978-0-323-47978-3
Executive Content Strategist: James Merritt
Director, Content Development: Rebecca Gruliow
Publishing Services Manager: Patricia Tannian
Senior Project Manager: John Casey Design Direction: Ryan Cook
D EDICATION
To
Our Students, Our Colleagues, and Our Families
PREFACE
This ninth edition of Cellular and Molecular Immunology includes substantial revisions, which we made to keep the textbook current with scientific advances and, at the same time, maintain the clear and readable style that has been typical of previous editions. Whenever we have added new information, we have focused primarily on important concepts and have not increased the length of the book. We have also rewritten many sections for increased clarity, accuracy, and completeness.
A general theme in modern immunology is that the field is moving beyond establishing fundamental principles of the mechanisms of immune responses to applying these principles to understand human disease and develop new therapies. The revolution in immunological therapies over the last twenty years has been extraordinary. It is especially satisfying for immunologists that some of the most innovative and effective immunotherapies have been developed because the basic science has matured and the complex mechanisms of immune activation and regulation have been elucidated in increasing detail. In this edition of the book, we have paid special attention to the clinical relevance of immunology and have emphasized how newly developed therapies work and what their strengths and pitfalls are.
In addition to these translational aspects of immunology, we have also updated basic concepts wherever there has been significant new understanding. Some examples of these fundamental advances include current views on innate lymphoid cells, the biology of inflammasome activation, the role of follicular helper T cells in antibody responses in germinal centers, newly described memory lymphocyte subsets, and the protective and pathogenic roles of effector T cells.
As in previous editions, each chapter is written so that it can be read and understood on its own, without referring to other chapters. In order to do this, it is often necessary to repeat some basic concepts and general principles that are covered in other chapters. We feel such repetition is valuable because it enables the reader to consolidate learning and to understand the content of each chapter independently of the others. We also feel this is helpful for faculty teaching from the book, because they can consider each chapter the topic of one or two lectures.
We have also continued to improve our illustration program. All illustrations have been revised to provide more visual depth and clarity. New figures have been added, and previously used figures have been reviewed and often changed for accuracy. We have kept design features such as the use of bold italic text to highlight “take-home messages” to make the book easy to read. The lists of suggested readings continue to emphasize recent review articles that provide in-depth coverage of particular topics for the interested reader. We have divided the lists into sections based on themes to help readers find the most useful articles for their needs. This edition also includes a page listing the online resources available to instructors and students (page vii).
Individuals who have helped us with specific topics are (in alphabetical order) Drs. Mark Anderson, Jason Cyster, Andrew Gross, Richard Locksley, Miriam Merad, Michael Rosenblum, Wayne Shreffler, and Catherine Wu; all were generous with advice and comments. Our illustrators, David and Alexandra Baker of DNA Illustrations, remain full partners in the book and provide invaluable suggestions for clarity and accuracy. Several members of the
Elsevier staff played critical roles. Our editor, James Merritt, has been a source of support and encouragement. Our managing editor, Rebecca Gruliow, shepherded the book through its preparation and into production. Ryan Cook was responsible for managing the design, and John Casey was invaluable throughout the production stage. We also owe a debt of gratitude to our families for their unflagging support and their tolerance of our absences. Finally, our students were the original inspiration for the first edition of this book, and we remain continually grateful to them, because from them we learn how to think about the science of immunology and how to communicate knowledge in the clearest and most meaningful way.
ABUL K. ABBAS
ANDREW H. LICHTMAN
SHIV PILLAI
ONLINE RESOURCES FOR INSTRUCTORS AND STUDENTS
RESOURCES FOR INSTRUCTORS
The following resources for instructors are available for use when teaching via Evolve. Please contact your local sales representative for more information or go directly to the Evolve website to request access: https://evolve.elsevier.com. Note: It may take 1 to 3 days for account access setup and verification.
Image Collection
All figures from Cellular and Molecular Immunology, ed 9, are available as an image collection in three formats, with labels on/off: PowerPoint, JPEG, and PDF versions. Figures may be downloaded individually or by chapter.
Animations
The 11 animations that students can access via the online version of the book also are available to Instructors on Evolve. Topics for which animations are available are indicated by in the margin. The animations can be easily downloaded to your computer by following the links on the Evolve site.
Test Bank
Instructors can access and download 114 UMSLE-style multiple choice and matching questions from the test bank for use in classroom presentations and testing.
RESOURCES FOR STUDENTS
The following resources are available online to students with the purchase of Cellular and Molecular Immunology, ed 9, on StudentConsult.com.
Textbook online
The complete textbook is available online at StudentConsult.com. The online version is fully searchable and provides all figures from the print book, with enhanced functionality for many, including clickable enlargements and slideshow views of multiple-part images.
Glossary
The complete book glossary is available online at StudentConsult.com, with searchable terms linked to their discussion in the text. Readers may click on boldface highlighted key
terms in the text to view pop-up definitions from the Glossary as they read the chapters online.
Clinical Cases
Five clinical cases are available online and linked via icons from the corresponding textbook discussion, indicated by in the margin. These clinical cases cover various diseases involving the immune system and are meant to show how the basic science of immunology contributes to our understanding of human diseases. Each case illustrates typical ways in which a disease manifests, what tests are used in diagnosis, and common modes of treatment. Each case poses questions and provides answers with explanations to increase understanding.
Self-Assessment Questions
Students can test and score themselves with 135 interactive multiple choice questions available on StudentConsult.com
Animations
Animations are available online at StudentConsult.com to illustrate the following topics:
Clonal selection
Steps in maturation of lymphocytes
Capture and presentation of protein antigens by dendritic cells
Induction and effector phases of cell-mediated Immunity
T cell–mediated immune reactions
Sequence of events in helper T cell–dependent antibody responses
Antibody-mediated opsonization and phagocytosis of microbes
Pathways of complement activation
Induction of CD8 T cell responses against tumors
Immediate hypersensitivity
Topics for which animations are available are indicated by in the margin of the text.
CONTENTS
CHAPTER 1 Properties and Overview of Immune Responses 1
CHAPTER 2 Cells and Tissues of the Immune System 13
CHAPTER 3 Leukocyte Circulation and Migration into Tissues 39
CHAPTER 4 Innate Immunity 57
CHAPTER 5 Antibodies and Antigens 97
CHAPTER 6 Antigen Presentation to T Lymphocytes and the Functions of Major Histocompatibility Complex Molecules 117
CHAPTER 7 Immune Receptors and Signal Transduction 145
CHAPTER 8 Lymphocyte Development and Antigen Receptor Gene Rearrangement 179
CHAPTER 9 Activation of T Lymphocytes 209
CHAPTER 10 Differentiation and Functions of CD4+ Effector T Cells 225
CHAPTER 11 Differentiation and Functions of CD8+ Effector T Cells 243
CHAPTER 12 B Cell Activation and Antibody Production 251
CHAPTER 13 Effector Mechanisms of Humoral Immunity 275
CHAPTER 14 Specialized Immunity at Epithelial Barriers and in Immune Privileged Tissues 299
CHAPTER 15 Immunologic Tolerance and Autoimmunity 325
CHAPTER 16 Immunity to Microbes 351
CHAPTER 17 Transplantation Immunology 373
CHAPTER 18 Immunity to Tumors 397
CHAPTER 19 Hypersensitivity Disorders 417
CHAPTER 20 Allergy 437
CHAPTER 21 Congenital and Acquired Immunodeficiencies 459
Glossary 489
Appendices
I Cytokines 519
II Principal Features of Selected CD Molecules 523
III Laboratory Techniques Commonly Used in Immunology 531
Properties and Overview of Immune Responses
INNATE AND ADAPTIVE IMMUNITY, 2
INNATE IMMUNITY: THE EARLY DEFENSE, 3
ADAPTIVE IMMUNITY, 4
Cardinal Features of Adaptive Immune Responses, 4
Overview of Humoral and Cell-Mediated Immunity, 5
Initiation and Development of Adaptive Immune Responses, 8
Humoral Immunity, 10
Cell-Mediated Immunity, 10
SUMMARY, 11
The term immunity is derived from the Latin word immunitas, which referred to the protection from legal prosecution offered to Roman senators during their tenures in office. Historically, immunity meant protection from disease and, more specifically, infectious disease. The cells and molecules responsible for immunity constitute the immune system, and their collective and coordinated response to the introduction of foreign substances is called the immune response. The physiologic function of the immune system is defense against infectious microbes; however, even noninfectious foreign substances and products of damaged cells can elicit immune responses. Furthermore, mechanisms that normally protect individuals from infection and eliminate foreign substances also are capable of causing tissue injury and disease in some situations. Therefore, a more inclusive definition of the immune response is a reaction to microbes as well as to molecules that are recognized as foreign, regardless of the physiologic or pathologic consequence of such a reaction. Under some situations, even self molecules can elicit immune responses (so-called autoimmune responses). Immunology is the study of immune responses in this broader sense and of the cellular and molecular events that occur after an organism encounters microbes and other foreign macromolecules.
Historians often credit Thucydides, in the fifth century BC in Athens, as having first mentioned immunity to an infection that he called plague (but that was probably not the bubonic plague we recognize today). The concept of protective immunity may have existed long before, as suggested by the ancient Chinese custom of making children resistant to smallpox by having them inhale powders made from the skin lesions of patients recovering from the disease. Immunology, in its modern form, is an experimental science in which explanations of immunologic phenomena are based on experimental observations and the conclusions drawn from them. The evolution of immunology as an experimental discipline has depended on our ability to manipulate the function of the immune system under controlled conditions.
Historically, the first clear example of this manipulation, and one that remains among the most dramatic ever recorded, was Edward Jenner’s successful vaccination against smallpox. Jenner, an English physician, noticed that milkmaids who had recovered from cowpox never contracted the more serious smallpox. On the basis of this observation, he injected the material from a cowpox pustule into the arm of an 8-year-old boy. When this boy was later intentionally inoculated with smallpox, the disease did not develop. Jenner’s landmark treatise on vaccination (Latin vaccinus, of or from cows) was published in 1798. It led to the widespread acceptance of this method for inducing immunity to infectious diseases, and vaccination remains the most effective method for preventing infections (Table 1.1). An eloquent testament to the importance of immunology was the announcement by the World Health Organization in 1980 that smallpox was the first disease that had been eradicated worldwide by a program of vaccination.
Since the 1960s, there has been a remarkable transformation in our understanding of the immune system and its functions. Advances in cell culture techniques (including monoclonal antibody production), immunochemistry, recombinant DNA methodology, x-ray crystallography, and creation of genetically altered animals (especially transgenic and knockout mice) have changed immunology from a largely descriptive science into one in which diverse immune phenomena can be explained in structural and biochemical terms. Some of the most
TABLE 1.1 Effectiveness of Vaccines for Some
This table illustrates the striking decrease in the incidence of selected infectious diseases in the United States for which effective vaccines have been developed.
Data from Orenstein WA, Hinman
AR, Bart KJ, Hadler SC: Immunization. In Mandell GL, Bennett JE, Dolin R (eds): Principles and practices of infectious diseases, ed 4, New York, 1995, Churchill Livingstone; and Morbidity and Mortality Weekly Report 64, No. 20, 2015.
important advances in immunology have come since the 1990s, with the development of therapies targeting different components of the immune system that are based on fundamental science and are dramatically altering the progression of human inflammatory diseases and cancers.
In this chapter, we outline the general features of immune responses and introduce the concepts that form the cornerstones of modern immunology and that recur throughout this book.
INNATE AND ADAPTIVE IMMUNITY
Defense against microbes is mediated by sequential and coordinated responses that are called innate and adaptive immunity (Fig. 1.1 and Table 1.2). Innate immunity (also called natural immunity or native immunity) is essential for defending against microbes in the first few hours or days after infection, before adaptive immune responses have developed. Innate immunity is mediated by mechanisms that are in place even before an infection occurs (hence innate) and that facilitate rapid responses to invading microbes.
In contrast to innate immunity, there are other immune responses that are stimulated by exposure to infectious agents and increase in magnitude and defensive capabilities with each successive exposure to a particular microbe. Because this form of immunity develops as a response to infection and adapts to the infection, it is called adaptive immunity (also called specific immunity or acquired immunity). The adaptive immune system recognizes and reacts to a large number of microbial and nonmicrobial substances, called antigens. Although many
FIGURE 1.1 Innate and adaptive immunity. The mechanisms of innate immunity provide the initial defense against infections. Adaptive immune responses develop later and require the activation of lymphocytes. The kinetics of the innate and adaptive immune responses are approximations and may vary in different infections. Only selected cell types are shown. ILC, Innate lymphoid cell; NK, natural killer.
TABLE 1.2 Features of Innate and
Innate
Characteristics
Specificity For molecules shared by groups of related microbes and molecules produced by damaged host cells
Diversity Limited; recognition molecules encoded by inherited (germline) genes
Memory None or limited
Nonreactivity to self Yes
Components
Cellular and chemical barriers
Skin, mucosal epithelia; antimicrobial molecules
Blood proteins Complement, various lectins and agglutinins
pathogens have evolved to resist the innate immune response, adaptive immune responses, being stronger and more specialized, are capable of eradicating even these infections. There are also numerous connections between innate and adaptive immune responses. The innate immune response to microbes provides early danger signals that stimulate adaptive immune responses. Conversely, adaptive immune responses often work by enhancing the protective mechanisms of innate immunity, making them more capable of effectively combating microbes.
Every individual’s immune system is able to recognize, respond to, and eliminate many foreign (nonself) antigens but does not usually react against that individual’s own (self) antigens and tissues. Different mechanisms are used by the innate and adaptive immune systems to prevent reactions against healthy self cells.
Because of the ability of lymphocytes and other immune cells to circulate among tissues, immunity is systemic, meaning that even if an immune response is initiated in one site it can provide protection at distant sites. This feature is, of course, essential for the success of vaccination—a vaccine administered in the subcutaneous or muscle tissue of the arm can protect from infections in any tissue.
Immune responses are regulated by a system of positive feedback loops that amplify the reaction and by control mechanisms that prevent inappropriate or pathologic reactions. When lymphocytes are activated, they trigger mechanisms that further increase the magnitude of the response. This positive feedback is important to enable the small number of lymphocytes that are specific for any microbe to generate a large response needed to eradicate that infection. Many control mechanisms become active during immune responses, which prevent excessive activation of lymphocytes that could cause collateral
Adaptive
For microbial and nonmicrobial antigens
Very large; receptor genes are formed by somatic recombination of gene segments in lymphocytes
Yes
Yes
Lymphocytes in epithelia; antibodies secreted at epithelial surfaces
Antibodies
Lymphocytes
damage to normal tissues, and also prevent responses against self antigens.
Mechanisms for defending the host against microbes are present in all multicellular organisms. The phylogenetically oldest mechanisms of host defense are those of innate immunity, which are present even in plants and insects. Approximately 500 million years ago, jawless fish, such as lampreys and hagfish, developed an immune system containing lymphocyte-like cells that may function like lymphocytes in more advanced species and even respond to immunization. The antigen receptors on these cells are proteins with limited variablity that are capable of recognizing many antigens but are distinct from the highly variable antibodies and T cell receptors that appeared later in evolution. The more specialized defense mechanisms that constitute adaptive immunity are found in vertebrates only. Most of the components of the adaptive immune system, including lymphocytes with highly diverse antigen receptors, antibodies, and specialized lymphoid tissues, evolved coordinately within a short time in jawed vertebrates (e.g., sharks) approximately 360 million years ago.
INNATE IMMUNITY: THE EARLY DEFENSE
The innate immune system responds almost immediately to microbes and injured cells, and repeated exposures invoke virtually identical innate immune responses. The receptors of innate immunity are specific for structures that are common to groups of related microbes and do not distinguish fine differences between microbes. The principal components of innate immunity are (1) physical and chemical barriers, such as epithelia and antimicrobial chemicals produced at epithelial surfaces; (2) phagocytic cells (neutrophils, macrophages), dendritic cells (DCs),
Adaptive Immunity
mast cells, natural killer (NK cells) and other innate lymphoid cells, and mast cells; and (3) blood proteins, including components of the complement system and other mediators of inflammation. Many innate immune cells, such as macrophages, DCs, and mast cells, are always present in most tissues, where they function as sentinels to keep watch for invading microbes. The innate immune response combats microbes by two main reactions—by recruiting phagocytes and other leukocytes that destroy the microbes, in the process called inflammation, and by blocking viral replication or killing virus-infected cells without a need for an inflammatory reaction. We will discuss the features, mechanisms, and components of innate immunity in Chapter 4
ADAPTIVE IMMUNITY
The adaptive immune response is mediated by cells called lymphocytes and their products. Lymphocytes express highly diverse receptors that are capable of recognizing a vast number of antigens. There are two major populations of lymphocytes, called B lymphocytes and T lymphocytes, which mediate different types of adaptive immune responses. We will first summarize the important properties of the adaptive immune system and then return to the different types of adaptive immune responses.
Cardinal Features of Adaptive Immune Responses
The fundamental properties of the adaptive immune system reflect the properties of the lymphocytes that mediate these responses.
• Specificity and diversity. Immune responses are specific for distinct antigens and often for different portions of a single complex protein, polysaccharide, or other macromolecule (Fig. 1.2). The parts of complex antigens that are specifically recognized by lymphocytes are called determinants or epitopes. This fine specificity exists because individual lymphocytes express membrane receptors that can distinguish subtle differences in structure between distinct epitopes. Clones of lymphocytes with different specificities are present in unimmunized individuals and are able to recognize and respond to foreign antigens (Fig. 1.3). This fundamental concept is called clonal selection. It was clearly enunciated by Macfarlane Burnet in 1957, as a hypothesis to explain how the immune system could respond to a large number and variety of antigens. According to this hypothesis, which is now a proven feature of adaptive immunity, antigen-specific clones of lymphocytes develop before and independent of exposure to antigen. An introduced antigen binds to (selects) the cells of the pre-existing antigen-specific clone and activates them. As a result, the cells specific for the antigen proliferate to generate thousands of progeny with the same specificity, a process called clonal expansion. The total number of antigenic specificities of the lymphocytes in an individual, called the lymphocyte repertoire, is extremely large. It is estimated that the immune system of an individual can discriminate 107 to 109 distinct antigenic determinants. This ability of the lymphocyte repertoire to recognize a very large number of antigens, called diversity, is the result of variability in the structures of the antigenbinding sites of lymphocyte receptors for antigens. In other words, there are many different clones of
FIGURE 1.2 Specificity, memory, and contraction of adaptive immune responses. Antigens X and Y induce the production of different antibodies (specificity). The secondary response to antigen X is more rapid and larger than the primary response (memory). Antibody levels decline with time after each immunization (contraction, the process that maintains homeostasis). The same features are seen in T cell–mediated immune responses.
Lymphocyte clones mature in generative lymphoid organs, in the absence of antigens
Clones of mature lymphocytes specific for diverse antigens enter lymphoid tissues
Antigen-specific clones are activated ("selected") by antigens
Antigen-specific immune responses occur
Lymphocyte precursor Mature lymphocyte
Antigen X
Anti-X antibody
FIGURE 1.3 Clonal selection. Each antigen (X) selects a preexisting clone of specific lymphocytes and stimulates the proliferation and differentiation of that clone. The diagram shows only B lymphocytes giving rise to antibody-secreting effector cells, but the same principle applies to T lymphocytes.
lymphocytes and each clone has a unique antigen receptor and therefore a singular antigen specificity, contributing to a total repertoire that is extremely diverse. The expression of different antigen receptors in different clones of T and B cells is the reason why these receptors are said to be clonally distributed. The molecular mechanisms that generate such diverse antigen receptors are discussed in Chapter 8. Diversity is essential if the immune system is to defend individuals against the many potential pathogens in the environment.
• Memory. Exposure of the immune system to a foreign antigen enhances its ability to respond again to that antigen. Responses to second and subsequent exposures to the same antigen, called secondary immune responses, are usually more rapid, greater in magnitude, and often qualitatively different from the first, or primary, immune response to that antigen (see Fig. 1.2). Immunologic memory occurs because each exposure to an antigen generates long-lived memory cells specific for the antigen. There are two reasons why the secondary response is typically stronger than the primary immune response—memory cells accumulate and become more numerous than the naive lymphocytes specific for the antigen that exist at the time of initial antigen exposure, and memory cells react more rapidly and vigorously to antigen challenge than do naive lymphocytes. Memory enables the immune system to mount heightened responses to
persistent or recurring exposure to the same antigen and thus to combat infections by microbes that are prevalent in the environment and are encountered repeatedly.
• Nonreactivity to self (self tolerance). One of the most remarkable properties of every normal individual’s immune system is its ability to recognize, respond to, and eliminate many foreign (nonself) antigens while not reacting harmfully to that individual’s own (self) antigens. Immunologic unresponsiveness is also called tolerance. Tolerance to self antigens, or self-tolerance, is maintained by several mechanisms. These include eliminating lymphocytes that express receptors specific for some self antigens, inactivating self-reactive lymphocytes, or suppressing these cells by the actions of other (regulatory) cells. Abnormalities in the induction or maintenance of self-tolerance lead to immune responses against self (autologous) antigens, which may result in disorders called autoimmune diseases. The mechanisms of self-tolerance and its failure are discussed in Chapter 15
Overview of Humoral and Cell-Mediated Immunity
There are two types of adaptive immunity, called humoral immunity and cell-mediated immunity, which are induced by different types of lymphocytes and function to eliminate different types of microbes (Figs. 1.4 and 1.5). Humoral immunity is mediated by molecules in the
blood and mucosal secretions, called antibodies, which are produced by B lymphocytes. Antibodies recognize microbial antigens, neutralize the infectivity of the microbes, and target microbes for elimination by phagocytes and the complement system. Humoral immunity is the principal defense mechanism against microbes and their toxins located outside cells (e.g. in the lumens of the gastrointestinal and respiratory tracts and in the blood) because secreted antibodies can bind to these microbes and toxins, neutralize them, and assist in their elimination.
Cell-mediated immunity, also called cellular immunity, is mediated by T lymphocytes. Many microbes are ingested by but survive within phagocytes, and some microbes, notably viruses, infect and replicate in various host cells. In these locations the microbes are inaccessible to circulating antibodies. Defense against such infections is a function of cell-mediated immunity, which promotes the destruction of microbes inside phagocytes and the killing of infected cells to eliminate reservoirs of infection.
Protective immunity against a microbe may be provided either by the host’s response to the microbe or by the transfer of antibodies that defend against the microbe (Fig. 1.6). The form of immunity that is induced by exposure to a foreign antigen is called active immunity because the immunized individual plays an active role in responding to the antigen. Individuals and lymphocytes that have not encountered a particular antigen are said to be naive, implying that they are immunologically inexperienced. Individuals who have responded to a microbial antigen and are protected from subsequent exposures to that microbe are said to be immune Immunity can also be conferred on an individual by transferring antibodies from an immunized individual into an individual who has not encountered the antigen (see Fig. 1.6). The recipient of such a transfer becomes immune to the particular antigen without ever having been exposed to or having responded to that antigen. Therefore, this form of immunity is called passive immunity. A physiologically important example of passive immunity is the transfer of maternal antibodies
FIGURE 1.4 Types of adaptive immunity. In humoral immunity, B lymphocytes secrete
Antigen recognition
Effector functions
through the placenta to the fetus, which enables newborns to combat infections for several months before they develop the ability to produce antibodies themselves. Passive immunization is also a medically useful method for conferring resistance rapidly, without having to wait for an active immune response to develop. Passive immunization against potentially lethal toxins by the administration of antibodies from immunized animals or people is a lifesaving treatment for rabies infection and snake bites. Patients with some genetic immunodeficiency diseases are passively immunized by transfer of pooled antibodies from healthy donors. The first demonstration of humoral immunity was provided by Emil von Behring and Shibasaburo Kitasato + Microbe
Cytokines B lymphocyte
Antibody
Helper T lymphocyte
Microbial antigen presented by antigenpresenting cell
Neutralization of microbe, phagocytosis, complement activation
Activation of macrophages
Inflammation
Activation (proliferation and differentiation) of T and B lymphocytes
Cytotoxic T lymphocyte (CTL)
Regulatory T lymphocyte
Infected cell expressing microbial antigen Regulatory T lymphocyte
Responding T lymphocyte
Killing of infected cell
Suppression of lymphocytes
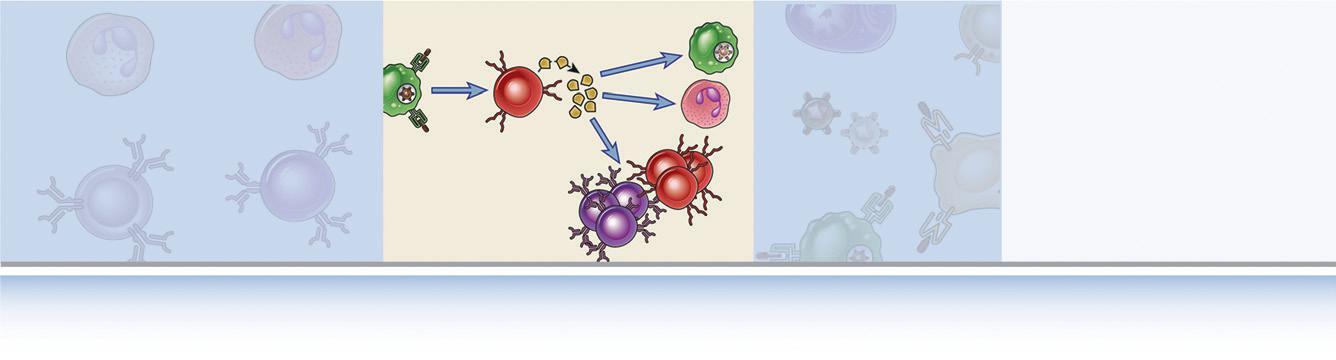
FIGURE 1.5 Classes of lymphocytes. B lymphocytes recognize many different types of antigens and develop into antibody-secreting cells. Helper T lymphocytes recognize antigens on the surfaces of antigenpresenting cells and secrete cytokines, which stimulate different mechanisms of immunity and inflammation.
Cytotoxic T lymphocytes recognize antigens on infected cells and kill these cells. Regulatory T cells suppress immune responses (e.g., to self antigens).
in 1890, using a passive immunization strategy. They showed that if serum from animals that had been immunized with an attenuated form of diphtheria toxin was transferred to naive animals, the recipients became specifically resistant to diphtheria infection. The active components of the serum were called antitoxins because they neutralized the pathologic effects of the diphtheria toxin. This result led to the treatment of otherwise lethal diphtheria infection by the administration of antitoxin, an achievement that was recognized by the award of the first Nobel Prize in Physiology or Medicine to von Behring. In the 1890s Paul Ehrlich postulated that immune cells use receptors, which he called side chains, to recognize microbial toxins and, subsequently, secrete
Active immunity
Microbial antigen (vaccine or infection)
Serum (antibodies) from immune individual Days or weeks
Passive immunity
Administration of serum to uninfected individual
FIGURE 1.6
Recovery (immunity)
Recovery (immunity)
Active and passive immunity. Active immunity is conferred by a host response to a microbe or microbial antigen, whereas passive immunity is conferred by adoptive transfer of antibodies or T lymphocytes specific for the microbe. Both forms of immunity provide resistance to infection and are specific for microbial antigens, but only active immune responses generate immunologic memory. Therapeutic passive transfer of antibodies, but not lymphocytes, is done routinely and also occurs during pregnancy (from mother to fetus).
these receptors to combat microbes. He also coined the term antibodies (antikörper in German) for the serum proteins that bound foreign substances, such as toxins, and the substances that generated antibodies were called antigens. The modern definition of antigens includes molecules that bind to specific lymphocyte receptors, whether or not they stimulate immune responses. According to strict definitions, substances that stimulate immune responses are called immunogens, but the term antigen is often used interchangeably with immunogen. The properties of antibodies and antigens are described in Chapter 5. Ehrlich’s concepts are a remarkably prescient model for the specificity of adaptive immunity. These early studies of antibodies led to the general acceptance of the humoral theory of immunity, according to which host defense against infections is mediated by substances present in body fluids (once called humors).
Élie Metchnikoff initially championed the cellular theory of immunity, which stated that host cells are the principal mediators of immunity. His demonstration of phagocytes surrounding a thorn stuck into a translucent starfish larva, published in 1883, was perhaps the first experimental evidence that cells respond to foreign invaders. Ehrlich and Metchnikoff shared the Nobel Prize in 1908, in recognition of their contributions to establishing these fundamental principles of immunity. Sir Almroth Wright’s observation in the early 1900s that factors in immune serum enhanced the phagocytosis of bacteria by coating the bacteria, a process known as opsonization, lent support to the belief that antibodies prepare microbes for ingestion by phagocytes. These early cellularists were unable to prove that specific immunity to microbes could be mediated by cells. The importance
of cellular immunity in host defense became firmly established in the 1950s, when it was shown that resistance to an intracellular bacterium, Listeria monocytogenes, could be transferred to animals with cells but not with serum. We now know that the specificity of cell-mediated immunity is due to T lymphocytes, which often function in concert with other cells, such as phagocytes, to eliminate microbes.
In the clinical setting, immunity to a previously encountered microbe is measured indirectly, either by assaying for the presence of products of immune responses (such as serum antibodies specific for microbial antigens) or by administering substances purified from the microbe and measuring reactions to these substances. A reaction to an antigen is detectable only in individuals who have previously encountered the antigen (the reaction at the time of the first encounter is usually too small to detect). These individuals are said to be sensitized to the antigen, and the reaction is an indication of sensitivity. Such a reaction to a microbial antigen implies that the sensitized individual is capable of mounting a protective immune response to the microbe.
Initiation and Development of Adaptive Immune Responses
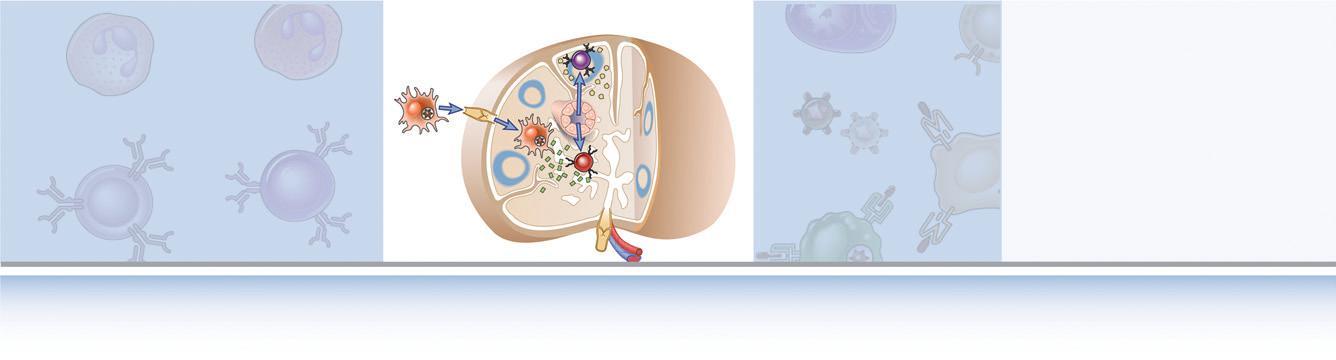
Adaptive immune responses develop in several steps, starting with the capture of antigen, followed by the activation of specific lymphocytes (Fig. 1.7).
Most microbes and other antigens enter through epithelial barriers and adaptive immune responses to these antigens develop in peripheral (secondary) lymphoid organs. The initiation of adaptive immune responses
FIGURE 1.7 Development of adaptive immune responses. Adaptive immune responses consist of distinct steps, the first three being the recognition of antigen, the activation of lymphocytes, and the elimination of antigen (the effector phase). The response contracts (declines) as antigen-stimulated lymphocytes die by apoptosis, restoring homeostasis, and the antigen-specific cells that survive are responsible for memory. The duration of each phase may vary in different immune responses. The y-axis represents an arbitrary measure of the magnitude of the response. These principles apply to humoral immunity (mediated by B lymphocytes) and cell-mediated immunity (mediated by T lymphocytes).
requires that antigens be captured and displayed to specific lymphocytes. The cells that serve this role are called antigen-presenting cells (APCs). The most specialized APCs are dendritic cells, which capture microbial antigens that enter from the external environment, transport these antigens to lymphoid organs, and present the antigens to naive T lymphocytes to initiate immune responses. Other cell types function as APCs at different stages of cell-mediated and humoral immune responses. We will describe the functions of APCs in Chapter 6 Lymphocytes that have not responded to antigen are said to be naive. The activation of these lymphocytes by antigen leads to the proliferation of these cells, resulting in an increase in the size of the antigen-specific clones, called clonal expansion. This is followed by differentiation of the activated lymphocytes into cells capable of eliminating the antigen, called effector cells because they mediate the ultimate effect of the immune response, and memory cells that survive for long periods and mount strong responses to repeat antigen encounter. Antigen elimination often requires the participation of other, nonlymphoid cells, such as macrophages and neutrophils, which are also sometimes called effector cells. These steps
in lymphocyte activation typically take a few days, which explains why the adaptive response is slow to develop and innate immunity has to provide protection initially.
After the adaptive immune response has eradicated the infection, the stimuli for lymphocyte activation dissipate and most of the effector cells die, resulting in the decline of the response. Memory cells remain, ready to respond vigorously if the same infection recurs.
The cells of the immune system interact with one another and with other host cells during the initiation and effector stages of innate and adaptive immune responses. Many of these interactions are mediated by cytokines. Cytokines are a large group of secreted proteins with diverse structures and functions, which regulate and coordinate many activities of the cells of innate and adaptive immunity. All cells of the immune system secrete at least some cytokines and express specific signaling receptors for several cytokines. Among the many functions of cytokines we will discuss throughout this book are promoting the growth and differentiation of immune cells, activating the effector functions of lymphocytes and phagocytes, and stimulating directed movement of immune cells from blood into tissues and within
tissues. A large subset of structurally related cytokines that regulate cell migration and movement are called chemokines. Some of the most effective drugs developed to treat immunologic diseases target cytokines, which reflects the importance of these proteins in immune responses. We will describe the functions of individual cytokines when we discuss immune responses in which these proteins play important roles.
Humoral Immunity
B lymphocytes that recognize antigens proliferate and differentiate into plasma cells that secrete different classes of antibodies with distinct functions. Each clone of B cells expresses a cell surface antigen receptor, which is a membrane-bound form of antibody, with a unique antigen specificity. Many different types of antigens, including proteins, polysaccharides, lipids, and small molecules, are capable of eliciting antibody responses. The response of B cells to protein antigens requires activating signals (help) from CD4+ T cells (which is the historical reason for calling these T cells helper cells). B cells can respond to many nonprotein antigens without the participation of helper T cells. Each plasma cell secretes antibodies that have the same antigen-binding site as the cell surface antigen receptor that first recognized the antigen. Polysaccharides and lipids stimulate secretion mainly of the antibody class called immunoglobulin M (IgM). Protein antigens induce the production of antibodies of different classes (IgG, IgA, IgE) from a single clone of B cells. These different antibody classes serve distinct functions, mentioned later. Helper T cells also stimulate the production of antibodies with increased affinity for the antigen. This process, called affinity maturation, improves the quality of the humoral immune response.
The humoral immune response combats microbes in many ways. Antibodies bind to microbes and prevent them from infecting cells, thus neutralizing the microbes. In fact, antibody-mediated neutralization is the only mechanism of adaptive immunity that stops an infection before it is established; this is why eliciting the production of potent antibodies is a key goal of vaccination. IgG antibodies coat microbes and target them for phagocytosis because phagocytes (neutrophils and macrophages) express receptors for parts of IgG molecules. IgG and IgM activate the complement system, and complement products promote phagocytosis and destruction of microbes. IgA is secreted from mucosal epithelia and neutralizes microbes in the lumens of mucosal tissues, such as the respiratory and gastrointestinal tracts, thus preventing inhaled and ingested microbes from infecting the host. Maternal IgG is actively transported across the placenta and protects the newborn until the baby’s immune system becomes mature. Most IgG antibodies have halflives in the circulation of approximately 3 weeks, whereas other classes of antibodies have half-lives of just a few days. Some antibody-secreting plasma cells migrate to the bone marrow or mucosal tissues and live for years, continuing to produce low levels of antibodies. The antibodies that are secreted by these long-lived plasma cells provide immediate protection if the microbe returns
to infect the individual. More effective protection is provided by memory cells that are activated by the microbe and rapidly differentiate to generate large numbers of plasma cells.
Cell-Mediated Immunity
T lymphocytes, the cells of cell-mediated immunity, recognize the antigens of cell-associated microbes, and different types of T cells help phagocytes to destroy these microbes or kill the infected cells. T cells do not produce antibody molecules. Their antigen receptors are membrane molecules distinct from but structurally related to antibodies (see Chapter 7). T lymphocytes have a restricted specificity for antigens; they recognize peptides derived from foreign proteins that are bound to host proteins called major histocompatibility complex (MHC) molecules, which are expressed on the surfaces of other cells. As a result, these T cells recognize and respond to cell surfaceassociated but not soluble antigens (see Chapter 6).
T lymphocytes consist of functionally distinct populations, the best defined of which are helper T cells and cytotoxic (or cytolytic) T lymphocytes (CTLs). The functions of helper T cells are mediated mainly by secreted cytokines, whereas CTLs produce molecules that kill other cells. Some T lymphocytes, which are called regulatory T cells, function mainly to inhibit immune responses. We will return to a more detailed discussion of the properties of lymphocytes in Chapter 2 and in later chapters. Different classes of lymphocytes can be distinguished by the expression of cell surface proteins, many of which are designated by a unique “CD” number (see Chapter 2), such as CD4 or CD8.
Upon activation in secondary lymphoid organs, naive T lymphocytes differentiate into effector cells, and many of them leave and migrate to sites of infection. When these effector T cells again encounter cell-associated microbes, they are activated to perform the functions that are responsible for elimination of the microbes. Some CD4+ helper T cells secrete cytokines that recruit leukocytes and stimulate production of microbicidal substances in phagocytes. Thus, these T cells help phagocytes to kill the infectious pathogens. Other CD4+ helper T cells secrete cytokines that help B cells to produce a type of antibody called IgE and activate leukocytes called eosinophils, which are able to kill parasites that may be too large to be phagocytosed. Some CD4+ helper T cells stay in the lymphoid organs and stimulate B cell responses.
CD8+ CTLs kill cells harboring microbes in the cytoplasm. These microbes may be viruses that infect many cell types or bacteria that are ingested by macrophages but escape from phagocytic vesicles into the cytoplasm (where they are inaccessible to the killing machinery of phagocytes, which is largely confined to vesicles). By destroying the infected cells, CTLs eliminate the reservoirs of infection. CTLs also kill tumor cells that express antigens that are recognized as foreign.
In the remainder of the book, we describe in detail the recognition, activation, regulation, and effector phases of innate and adaptive immune responses. The principles introduced in this chapter recur throughout this book.
SUMMARY
Y Protective immunity against microbes is mediated by the early reactions of innate immunity and the later responses of adaptive immunity. Innate immune responses are stimulated by molecular structures shared by groups of microbes and by molecules expressed by damaged host cells. Adaptive immunity is specific for different microbial and nonmicrobial antigens and is increased by repeated exposures to antigen (immunologic memory).
Y Many features of adaptive immunity are of fundamental importance for its normal functions. These include specificity for different antigens, a diverse repertoire capable of recognizing a wide variety of antigens, memory of antigen exposure, and the ability to discriminate between foreign antigens and self antigens.
Y Immunity may be acquired by a response to antigen (active immunity) or conferred by transfer of antibodies or effector cells (passive immunity).
Y Lymphocytes are the only cells capable of specifically recognizing antigens and are thus the principal cells of adaptive immunity. The total population of lymphocytes consists of many clones, each with a unique antigen receptor and specificity. The two major subsets of lymphocytes are B cells and T cells, and they differ in their antigen receptors and functions.
Y The adaptive immune response is initiated by the recognition of foreign antigens by specific lymphocytes. Specialized APCs capture microbial antigens and display these antigens for recognition by lymphocytes. Lymphocytes respond by proliferating and by differentiating into effector cells, whose function is to eliminate the antigen, and into memory cells, which show enhanced responses on subsequent encounters with the antigen. The elimination of
antigens often requires the participation of various effector cells.
Y Humoral immunity is mediated by antibodies secreted by B lymphocytes and is the mechanism of defense against extracellular microbes. Antibodies neutralize the infectivity of microbes and promote the elimination of microbes by phagocytes and by activation of the complement system.
Y Cell-mediated immunity is mediated by T lymphocytes and their products, such as cytokines, and is important for defense against intracellular microbes. CD4+ helper T lymphocytes help macrophages to eliminate ingested microbes and help B cells to produce antibodies. CD8+ CTLs kill cells harboring intracellular pathogens, thus eliminating reservoirs of infection.
SELECTED READINGS
Historical Ideas
Burnet FM. A modification of Jerne’s theory of antibody production using the concept of clonal selection. Australien J Sci 1957;20:67-69.
Cohn M, Mitchison NA, Paul WE, et al. Reflections on the clonal selection theory. Nat Rev Immunol. 2007;7:823-830.
Jerne NK. The natural-selection theory of antibody formation. Proc Natl Acad Sci USA. 1955;41:849-857.
Silverstein AM. Cellular versus humoral immunology: a centurylong dispute. Nat Immunol. 2003;4:425-428.
Evolution of the Immune System
Boehm T, Swann JB. Origin and evolution of adaptive immunity. Annu Rev Anim Biosci. 2014;2:259-283.
Flajnik MF, Du Pasquier L. Evolution of innate and adaptive immunity: can we draw a line? Trends Immunol. 2004;25: 640-644.
Litman GW, Rast JP, Fugmann SD. The origins of vertebrate adaptive immunity. Nat Rev Immunol. 2010;10:543-553.
Cells and Tissues of the Immune System
CELLS OF THE IMMUNE SYSTEM, 13
Phagocytes, 14
Mast Cells, Basophils, and Eosinophils, 17
Lymphocytes, 21
Natural Killer Cells and Cytokine-Secreting Innate Lymphoid Cells, 27
ANATOMY AND FUNCTIONS OF LYMPHOID TISSUES, 27
Bone Marrow, 28
Thymus, 28
The Lymphatic System, 30
Lymph Nodes, 32
Spleen, 35
Cutaneous and Mucosal Immune Systems, 36
SUMMARY, 36
The cells of the innate and adaptive immune system are normally present as circulating cells in the blood and lymph, in lymphoid organs, and as scattered cells in virtually all tissues. The anatomic arrangement of these cells in lymphoid tissues and their ability to circulate and exchange among blood, lymph, and tissues are of critical importance for the generation of immune responses. The immune system faces numerous challenges to generate effective protective responses against infectious pathogens. First, the system must be able to respond rapidly to small numbers of many different microbes that may be introduced at any site in the body. Second, in the adaptive immune response, very few naive lymphocytes specifically recognize and respond to any one antigen. Third, the effector mechanisms of the adaptive immune system (antibodies and effector T cells) may have to locate and destroy microbes at sites that are distant from the site where the immune response was induced. The capacity of the immune system to meet these challenges and to optimally perform its protective functions is dependent on the remarkably rapid and varied responses of immune cells, the way these cells are organized in lymphoid tissues, and their ability to migrate from one tissue to another.
This chapter describes the cells and tissues that compose the immune system. In Chapter 3, we describe the traffic patterns of lymphocytes throughout the body and the mechanisms of migration of lymphocytes and other leukocytes.
CELLS OF THE IMMUNE SYSTEM
The cells that serve specialized roles in innate and adaptive immune responses are phagocytes, dendritic cells (DCs), antigen-specific lymphocytes, and various other leukocytes that function to eliminate antigens. These cells were introduced briefly in Chapter 1. These cells are almost all derived from hematopoietic stem cells (HSCs) in the bone marrow, which differentiate along branching lineages. Based on their common precursors, immune cells are broadly classified as either myeloid cells, which include phagocytes and most DCs, or lymphoid cells, which include all lymphocytes. The numbers of some of these cell types in the blood are listed in Table 2.1 Although most of these cells are found in the blood, the responses of lymphocytes to antigens usually occur in lymphoid and other tissues and therefore may not be reflected by changes in the numbers of blood lymphocytes.
The expression of various membrane proteins is used to distinguish distinct populations of cells in the immune system. For instance, most helper T cells express a surface protein called CD4, and most cytotoxic T lymphocytes (CTLs) express a different surface protein called CD8. These and many other surface proteins are often called markers because they identify and discriminate between (mark) different cell populations. These markers not only delineate the different classes of cells in the innate and adaptive immune systems, but the proteins also have many functions in the cell types in which they are expressed. The most common way to determine if a particular marker is expressed on a cell is to test if antibodies specific for the marker bind to the cell. In this context, the antibodies are used by investigators or clinicians as analytical tools. There are available hundreds of different pure antibody preparations, called monoclonal antibodies, each specific for a different molecule and labeled with probes that can be readily detected on cell surfaces by use of appropriate instruments. (Monoclonal
antibodies are described in Chapter 5, and methods to detect labeled antibodies bound to cells are discussed in Appendix III.) The cluster of differentiation (CD) nomenclature is a widely adopted uniform method for naming cell surface molecules that are characteristic of a particular cell lineage or differentiation stage, have a defined structure, and are recognized by a group (cluster) of monoclonal antibodies. Thus, all structurally defined cell surface molecules are given a CD number designation (e.g., CD1, CD2). Although originally devised to define circulating immune cell (leukocyte) subtypes, CD markers are found on all cell types in the body. CD molecules have important functions in immune responses and are the targets of many therapeutic antibodies used in the treatment of inflammatory diseases and cancer. Appendix II provides a current list of leukocyte CD markers that are mentioned in this book.
Phagocytes
Phagocytes, including neutrophils and macrophages, are cells whose primary function is to ingest and destroy microbes and remove damaged tissues. The functional responses of phagocytes in host defense consist of sequential steps: recruitment of the cells to the sites of infection, recognition of and activation by microbes, ingestion of the microbes by the process of phagocytosis, and destruction of ingested microbes. In addition, through direct contact and by secreting cytokines, phagocytes communicate with other cells in ways that promote or regulate immune responses.
Blood neutrophils and monocytes are both produced in the bone marrow, circulate in the blood, and are recruited to sites of inflammation. Although both are actively phagocytic, they differ in significant ways (Table 2.2). The neutrophil response is more rapid and the lifespan of these cells is short, whereas monocytes become macrophages in the tissues, can live for long periods, and so the macrophage response may last for a prolonged time. Neutrophils mainly use cytoskeletal rearrangements and enzyme assembly to mount rapid, transient responses, whereas macrophages rely mostly on new gene transcription. These functions of phagocytes are important in innate immunity, as we will discuss in
Chapter 4, and also in the effector phase of some adaptive immune responses, as we will discuss in Chapter 10. As a prelude to more detailed discussions of the role of phagocytes in immune responses in later chapters, here we will describe the morphologic features of neutrophils and macrophages and briefly introduce their functional responses.
Neutrophils
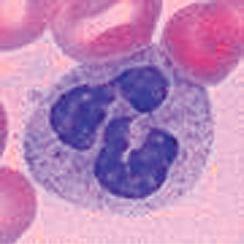
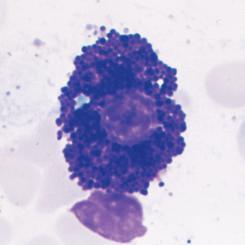
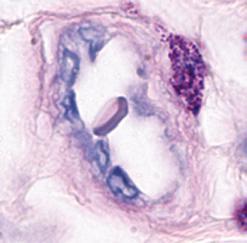
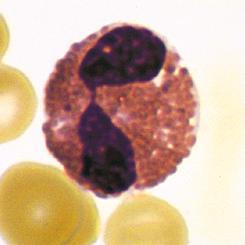
Neutrophils are the most abundant population of circulating white blood cells and the principal cell type in acute inflammatory reactions. Neutrophils circulate as spherical cells approximately 12 to 15 µm in diameter with numerous membranous projections. The nucleus is segmented into three to five connected lobules (Fig. 2.1A). Because of their nuclear morphology, neutrophils are also called polymorphonuclear leukocytes (PMNs). The cytoplasm contains two types of membrane-bound granules. The majority of these granules, called specific granules, are filled with enzymes, such as lysozyme, collagenase, and elastase. These granules do not stain strongly with either basic or acidic dyes (hematoxylin and eosin, respectively), which distinguishes neutrophils from two other
FIGURE 2.1 Morphology of neutrophils, mast cells, basophils, and eosinophils. A, The light micrograph of a WrightGiemsa–stained blood neutrophil shows the multilobed nucleus, because of which these cells are also called polymorphonuclear leukocytes, and the faint cytoplasmic granules. B, The light micrograph of a WrightGiemsa–stained section of skin shows a mast cell (arrow) adjacent to a small blood vessel, identifiable by the red blood cell in the lumen. The cytoplasmic granules in the mast cell, which are stained purple, are filled with histamine and other mediators that act on adjacent blood vessels to promote increased blood flow and delivery of plasma proteins and leukocytes into the tissue. (Courtesy of Dr. George Murphy, Department of Pathology, Brigham and Women’s Hospital, Boston, Massachusetts.) C, The light micrograph of a Wright-Giemsa–stained blood basophil shows the characteristic blue-staining cytoplasmic granules. (Courtesy of Dr. Jonathan Hecht, Department of Pathology, Brigham and Women’s Hospital, Boston, Massachusetts.) D, The light micrograph of a WrightGiemsa–stained blood eosinophil shows the characteristic segmented nucleus and red staining of the cytoplasmic granules.