
Cones Support Alignment to an Inconsistent World by Suppressing Mouse Circadian Responses to the Blue Colors Associated with Twilight Joshua W. Mouland & Franck Martial & Alex Watson & Robert J. Lucas & Timothy M. Brown https://ebookmass.com/product/cones-supportalignment-to-an-inconsistent-world-by-suppressingmouse-circadian-responses-to-the-blue-colorsassociated-with-twilight-joshua-w-mouland-franckmartial-alex-watson-robert/

Download more ebook from https://ebookmass.com
More products digital (pdf, epub, mobi) instant download maybe you interests ...

Elsevier Weekblad - Week 26 - 2022 Gebruiker
https://ebookmass.com/product/elsevier-weekbladweek-26-2022-gebruiker/

Destination C1 & C2 : grammar & vocabulary with answer key 18 [printing]. Edition Malcolm Mann
https://ebookmass.com/product/destination-c1-c2-grammarvocabulary-with-answer-key-18-printing-edition-malcolm-mann/

Jock Seeks Geek: The Holidates Series Book #26 Jill Brashear
https://ebookmass.com/product/jock-seeks-geek-the-holidatesseries-book-26-jill-brashear/

The New York Review of Books – N. 09, May 26 2022
Various Authors
https://ebookmass.com/product/the-new-york-review-ofbooks-n-09-may-26-2022-various-authors/

Calculate with Confidence, 8e (Oct 26, 2021)_(0323696953)_(Elsevier) 8th Edition Morris Rn Bsn Ma Lnc
https://ebookmass.com/product/calculate-withconfidence-8e-oct-26-2021_0323696953_elsevier-8th-edition-morrisrn-bsn-ma-lnc/

1 st International Congress and Exhibition on Sustainability in Music, Art, Textile and Fashion (ICESMATF 2023) January, 26-27 Madrid, Spain Exhibition Book 1st Edition Tatiana Lissa
https://ebookmass.com/product/1-st-international-congress-andexhibition-on-sustainability-in-music-art-textile-and-fashionicesmatf-2023-january-26-27-madrid-spain-exhibition-book-1stedition-tatiana-lissa/

An Introduction to Quantitative Ecology Timothy E. Essington
https://ebookmass.com/product/an-introduction-to-quantitativeecology-timothy-e-essington/

Entrepreneurial Responses to Chronic Adversity Dean A. Shepherd
https://ebookmass.com/product/entrepreneurial-responses-tochronic-adversity-dean-a-shepherd/

An Insider’s Guide to the Japanese Martial Arts : A New Look at Japan’s Fighting Traditions Alexander Bennett
https://ebookmass.com/product/an-insiders-guide-to-the-japanesemartial-arts-a-new-look-at-japans-fighting-traditions-alexanderbennett/
ConesSupportAlignmenttoanInconsistentWorld bySuppressingMouseCircadianResponsestothe
BlueColorsAssociatedwithTwilight
Highlights
d Cone-derivedblue:yellowcolorsignalsinfluencecircadian entrainmentinmice
d Thebluecolorsassociatedwithtwilightsuppresscircadian lightresponses
d Colorsignalssupportcircadianentrainmenttolow-amplitude light:darkcycles
d Colorsignalsbuffertheclockagainstcloud-relatedchanges inlightlevels
Authors
JoshuaW.Mouland,FranckMartial, AlexWatson,RobertJ.Lucas, TimothyM.Brown
Correspondence timothy.brown@manchester.ac.uk
InBrief
Changesinthespectralcontentof ambientlightaredetectabletomost mammalsasablueshiftinthecolorof twilight.Moulandetal.showthatthese ‘‘blue’’colorssuppresscircadian responsestolight,supportingrobust circadianentrainmentwhen environmentalconditionsrenderlight intensityaweakorunreliableindicatorof timeofday.

Moulandetal.,2019,CurrentBiology 29,4260–4267 December16,2019 ª 2019TheAuthor(s).PublishedbyElsevierLtd. https://doi.org/10.1016/j.cub.2019.10.028
ConesSupportAlignmenttoanInconsistentWorld bySuppressingMouseCircadianResponses totheBlueColorsAssociatedwithTwilight
JoshuaW.Mouland,1 FranckMartial,1 AlexWatson,1 RobertJ.Lucas,1 andTimothyM.Brown1,2,* 1CentreforBiologicalTiming,FacultyofBiology,Medicine&Health,UniversityofManchester,OxfordRoad,ManchesterM139PT,UK 2LeadContact
*Correspondence: timothy.brown@manchester.ac.uk https://doi.org/10.1016/j.cub.2019.10.028
SUMMARY
Inhumans,short-wavelengthlightevokeslarger circadianresponsesthanlongerwavelengths[1–3]. Thisreflectsthefactthatmelanopsin,akeycontributortocircadianassessmentsoflightintensity,most efficientlycapturesphotonsaround480nm[4–8] andgivesrisetothepopularviewthat‘‘blue’’lightexertsthestrongesteffectsontheclock.However,inthe naturalworld,thereisoftennodirectcorrelationbetweenperceivedcolor(asreportedbythecone-based visualsystem)andmelanopsinexcitation.Accordingly,althoughthemammalianclockdoesreceive cone-basedchromaticsignals[9],theinfluenceofcoloroncircadianresponsestolightremainsunclear. Here,wedefinethenatureandfunctionalsignificance ofchromaticinfluencesonthemousecircadiansystem.Usingpolychromaticlightingandmicewith alteredconespectralsensitivity(Opn1mwR),we generateconditionsthatdifferincolor(i.e.,ratioof L-toS-coneopsinactivation)whileprovidingidentical melanopsinandrodactivation.Whenbiasedtoward S-opsinactivation(appearing‘‘blue’’),thesestimuli reliablyproduceweakercircadianbehavioralresponsesthanthosefavoringL-opsin(‘‘yellow’’).This influenceofcolor(whichisabsentinanimalslacking conephototransduction; Cnga3 / )alignswithnaturalchangesinspectralcompositionovertwilight, wheredecreasingsolarangleisaccompaniedbya strongblueshift[9–11].Accordingly,wefindthat naturalisticcolorchangessupportcircadianalignmentwhenenvironmentalconditionsrenderdiurnal variationsinlightintensityweak/ambiguoussources oftiminginformation.Ourdatathusestablishhowcolorcontributestocircadianentrainmentinmammals andprovideimportantnewinsighttoinformthe designoflightingenvironmentsthatbenefithealth.
RESULTS
ColorModulatesCircadianAssessmentofLightLevels
Cone-derivedcolorsignalsreachthesuprachiasmaticnuclei (SCN)andcaninfluenceclockphase[9],butitremainsunclear
whichcolorsmosteffectivelyengagecircadianresponsesand howsuchamechanismcontributestoentrainmentunderrealworldconditions.Giventhepredictableshiftsinambientlight spectraatdawnanddusk[4, 12],wehypothesizedthatlight whosecolorresembledtwilight(i.e.,blue)wouldproduce weakercircadianresponsesthanlightofequivalentintensity butwhosecolorwasassociatedwithdaytime(yellowtowhite). Totestthis,weassessedcircadianbehaviorunderpolychromaticlightingwhosespectralcompositioncouldbevariedto adjustcolorindependentlyoflightintensity(Figure1A).
Themammaliancircadiansystemtrackslightintensityviaa combinationofmelanopsinandouter-retinalsignalsrelayed byintrinsicallyphotosensitiveretinalganglioncells(ipRGCs) [4, 13–15].Usingtheprinciplesofsilentsubstitution[16],we thereforeaimedtogeneratestimuliwithequivalentbrightness formelanopsin,rods,andcones(termedhere‘‘equi-luminant’’) butdistinctspectra(andconsequentlycolor)forthedichromatic mousevisualsystem.Toenablethegenerationofsubstantial differencesincolorwhilecontrollingmelanopsinandrodactivation,weemployedavalidated[9, 17–24]mouseline(Opn1mwR; hereaftertermedred-cone)[25],wherethenativeM-coneopsin (lmax=511nm)isreplacedwiththehumanL-coneopsin (lmax=556nm).
Westartedbyestablishingahousingenvironmentthat provideddiffuseoverheadilluminationfromindependently controllablelight-emittingdiode(LED)sources(Figure1A).We thencalibratedapolychromaticlightingcondition(using385-, 460-,and630-nmprimaries)thatrecreatedawild-typemouse’s experienceofnaturaldaylight(i.e.,‘‘white’’light; FigureS1).By adjustingtheintensitiesofeachprimaryrelativetothisreference whitepoint,wethenproducedapairofexperimentalstimuli. ThefirstmaximizedL-opsinandminimizedS-opsinexcitation (L+S ;thereforeappearing‘‘yellow’’byanalogywiththehuman longversusshort-wavelengthcolorchannel).Thesecond minimizedL-opsinandmaximizedS-opsinactivation(L S+; thereforeappearing‘‘blue’’)torecapitulateawild-typemouse’s experienceoftwilight.Importantly,therewerenegligibledifferences(<0.01logunits)betweenL S+andL+S stimuliinmelanopsinandrodexcitationaswellasintheaverageilluminance formouseconeopsins(Figure1A).
Wefirstusedthisapproachtoevaluatetheimpactofcolor onthecircadianperiodofvoluntarywheelrunningunderconstantillumination,aparadigmusedextensivelytoassessthe impactoflightonthemammalianclock(e.g.,[13]).Here,redconemice(n=8)wereexposedtoalternating2-weekblocks ofconstantL S+(blue)andthenequi-luminantL+S (yellow)


Figure1.ColorModulatesCircadianAssessmentofLightLevels
(A)Schematicofexperimentalparadigm(left),spectralcompositionofL S+(blue)andL+S (yellow)stimuli(mid),andopsinsensitivitycurves(right)forred-cone micewithcorrespondingquantificationforstimuliatmaximumintensity(ND0).Seealso FigureS1 foradditionaldetailsofstimulusdesign. (B)Representativeactogramforred-conemouseunderconstantL S+(blue)orL+S (yellow)illuminationat0.01,0.1,and13 intensitylevelshownin(A)(ND2–ND0,respectively).
(C)Circadianperiodforred-conemiceunderL S+(blue)versusL+S (yellow)illuminationatvaryingintensity(n=7–8/intensity).Dataanalyzedbytwo-way repeatedmeasures(RM)ANOVAwithSidak’spost-tests:intensity,F2,20 =39.4;p<0.001;color,F1,20 =11.7;p=0.003;intensity 3 color,F2,20 =3.8;p=0.04.
(D)Activityboutduration(a),expressedasafractionofcircadianperiodlength,forred-conemiceasabove.Two-wayRMANOVA:intensity,F2,20 =17.0; p<0.001;color,F1,20 =2.98;p=0.10;intensity 3 color,F2,20 =0.1;p=0.92.
(E)Sameas(B)butforconelessmouse.
(F)Sameas(C)butforconelessmice.Two-wayRMANOVA:intensity,F2,14 =31.3;p<0.001;color,F1,14 =0.1;p=0.82;intensity 3 color,F2,14 =2.6;p=0.11.
(G)Sameas(D)forconelessmice(n=5–6)mice.Two-wayRMANOVA:intensity,F2,14 =9.1;p=0.003;color,F1,14 =2.4;p=0.14;intensity 3 color,F2,14 =0.9; p=0.44.
(H)Spectralcompositionofstimulithatmodulatedconeilluminance(m(L,S))withoutchangingcolorormelanopsin/rodexcitation.
(I)Representativeactogramsfortwored-conemiceexposedtoconstantL+S (yellow),L S+(blue),L S (dim),andL+S+(bright)stimuliatND1.
(J)Circadianperioddeterminedforred-conemice(n=14)undertheconditionsillustratedin(I).Dataanalyzedbyone-wayRMANOVAwithDunnett’spost-tests: F3,39 =3.869;p=0.016.
*p<0.05,**p<0.01,and***p<0.001;ns=p>0.05.
illuminationacross3logarithmicallyspacedintensities(Figure1B).Asexpected,circadianperiodreliablylengthenedwith increasingintensity,butwealsoidentifiedasignificantimpact ofcolor,withlongercircadianperiodsunderL+S (yellow) versusL S+(blue)illumination(Figure1C),especiallyatintermediateintensities(Sidak’spost-test;p=0.006).Thesedata stronglysupportourhypothesisthatbluelightwillhaveaweaker effectontheclockthanequi-luminantyellowillumination.
Interestingly,anothercommonimpactofincreasinglightintensityonmousebehavior,compressionofactivityboutduration (a),wasnotsimilarlyimpacted.Hence,althoughtherewasa robustdecreasein a asafunctionofintensity,wedidnotdetect anysignificantinfluenceofcolor(Figure1D).Thismayreflect SCN-independentinfluencesonactivity[26]ortheinvolvement ofSCNneuronsthatprocessachromaticsignals[9].Ineither case,itseemsthatcolordoesnotgloballyimpactallbehavioral responsestolightbutinsteadmorespecificallyimpactsonclock speed.
BecauseourexperimentalstimuliselectivelymodulatetheratioofL-toS-coneopsinactivation,circadianbehaviorshouldbe indistinguishableunderequi-luminantL+S andL S+conditionsinanimalsthatlackedconephototransduction(Figure1E; Cnga3 / mice[27];hereaftertermedconeless).Accordingly, althoughconelessmice(n=7)retainedintensity-dependent increasesincircadianperiodandreductionin a duration,there werenodetectableeffectsofcolor(Figures1Fand1G).Indeed, atthetwohighestintensities,conelessmicewereatleastas likelytodisplaylongerfree-runningperiodsunderbluerather thanequi-luminantyellow(7outof11pairedmeasurements), whereasthisoccurredinonly1of15observationsfromredconemice(p=0.003;Fisher’sexacttest).Bycontrast,redconeandconelessdatawerequalitativelysimilaratthelowest intensity(whichfallsbelowtherangewherestrongcone-mediatedresponsesareobservable)[20].
Wenextsoughttoconfirmthatthereductioninthecircadian periodofred-conemiceunderblueilluminationathigherintensitieswasaspecificresultofcolorratherthanadifferencein effectiveconeilluminance.Tothisend,inaseparatebatchof red-conemice(n=14),wefirstpresented2-weekblocksof L+S (yellow)andthenL S+(blue)stimulifollowedbyblocks oftwoadditionalstimuliofintermediatecolor(equivalenttoa wild-typemouse’sexperienceofanovercastday)butvarying coneilluminance(Figure1H;L+S+(‘‘bright’’)andL S (‘‘dim’’)). Effectivephotonfluxformelanopsinandrodswas12.7logphotons/cm2/sforallstimuli.Ourexpectationwasthat,ifthe reducedcircadianperiodunderL S+(blue)illuminationsimply reflectedareductionineffectiveconeilluminance,circadian periodsshouldbeevenmorereducedundertheL S (dim) condition.Asabove,weonceagainfoundasignificant decreaseincircadianperiodunderL S+(blue)versus L+S (yellow)illumination(Figures1Iand1J;Dunnett’s post-test;p=0.04).Bycontrast,circadianperiodswerenot significantlydifferentfromL+S undereitherL S (dim)or L+S+(bright)conditions(p=0.44andp=0.91,respectively). Collectively,thesedataconfirmaspecificimpactofconederivedchromaticsignalsoncircadianperiod,withcolors resemblingthoseencounteredduringlatestagesoftwilight (blue)exertingaweakerimpactontheclockthancolorsassociatedwithdaytimeillumination.
ColorModulatesRe-entrainmentfollowing‘‘JetLag’’ Ourdataaboveindicatethatthetwilightblueshiftsubstantially attenuatescircadianresponsestolightandthusimplythat bluestimulishouldbelesseffectiveatresettingtheclockthan equi-luminantyellow.Totestthis,weinitiallyevaluatedchanges inthetimingofred-conemouse(n=16)behavioralrhythmsin responsetoacutepulsesofL+S (yellow)versusL S+(blue), presentedimmediatelyfollowingtransferfromalight:dark(LD) cycletoconstantdark.Wechosethisapproachtoavoidlongtermadaptationeffectsthatmightaccompanytestingunder constantdarkhousing(whereresettingresponsesaredominatedbyrodcontributions)[13].Withtheaimoffurther increasingconeinfluences,weemployedbrief(5-min)exposures[13, 28]atsub-saturatingintensitiesandpresentedthese eitherearlyorlateintheprojectednight(FiguresS2Aand S2B).Despiteatrendtowardsmallerphaseadvancesanddelaysfollowingbluestimuli,inneithercasewerethemeasured shiftssignificantlydifferentfromthoseevokedbyyellow(FigureS2C).Giventheinter-trialvariabilityassociatedwiththis kindofassay[29–31],itishardtodefinitivelyexcludeanyimpact ofcolor.Nonetheless,itseemsthat,underthespecificconditionsstudiedhere,colordoesnotexertamajorinfluenceon themagnitudeofacutelight-pulse-inducedresetting.
Becauseweidentifiedcleareffectsofcolorundermuch longerdurationsofilluminationthanthoseusedabove(Figure1), wenextaskedwhethercolorwouldmodulatetheabilityof micetore-entraintolargeshiftsinthetimingoftheLDcycle (jetlagparadigm).Here,red-conemice(n=8)experiencedat least7daysofaconventional12:12LDcycleandthentheonset ofthelightphasewasdelayedoradvancedby6handrendered aseitherL S+(blue)orL S+(yellow)(Figure2A).Wefoundthat changesinphase(activitymidpoint)producedbyL+S (yellow) stimuliweresignificantlymorerapidthanL S+(blue)forboth delayandadvanceshifts(Figure2B).Bycontrast,wecould notdetectasignificantinfluenceofcolorforeithershiftdirection inconelessmice(Figures2Cand2D).Collectively,thesedata supportourhypothesisthatcolorsignals,suppliedbycones, modulatecircadianresponsestolightsuchthatstimulithat appearbluearelesseffectiveatre-entrainingthecircadiansystemthanthosewithyellowcolor.
ColorSupportsCircadianEntrainmenttoUnreliable IntensityCues
Ourdataprovideastraightforwardmechanismbywhichcolor signalscouldaidcircadianentrainment—byreducingresponses tolightwhosecolorisindicativeoflatestagesoftwilight.To probetheecologicalsignificanceofthismechanism,wenext establishedanewhousingenvironmentthatallowedmore dynamiccontrolovertheintensityandcolorofillumination andfittedthiswithpassiveinfraredsensorsthatdetecteven smallwake-relatedbehaviorsratherthanjustdailyvariationsin locomotion[32].
Usingthissystem,wefirstaskedwhetheraprimaryfunctionof colorinputwastosupportcircadianentrainmentwhendiurnal changesinlightintensityaresmall.Inthenaturalworld,especiallyintheregionswheretheancestorsoflaboratorymice evolved,thisisanuncommoncircumstance.Nonetheless, suchapossibilityhasbeenproposedtoexplainhowsome animalsmaintainentrainmentduringthearcticsummer,where
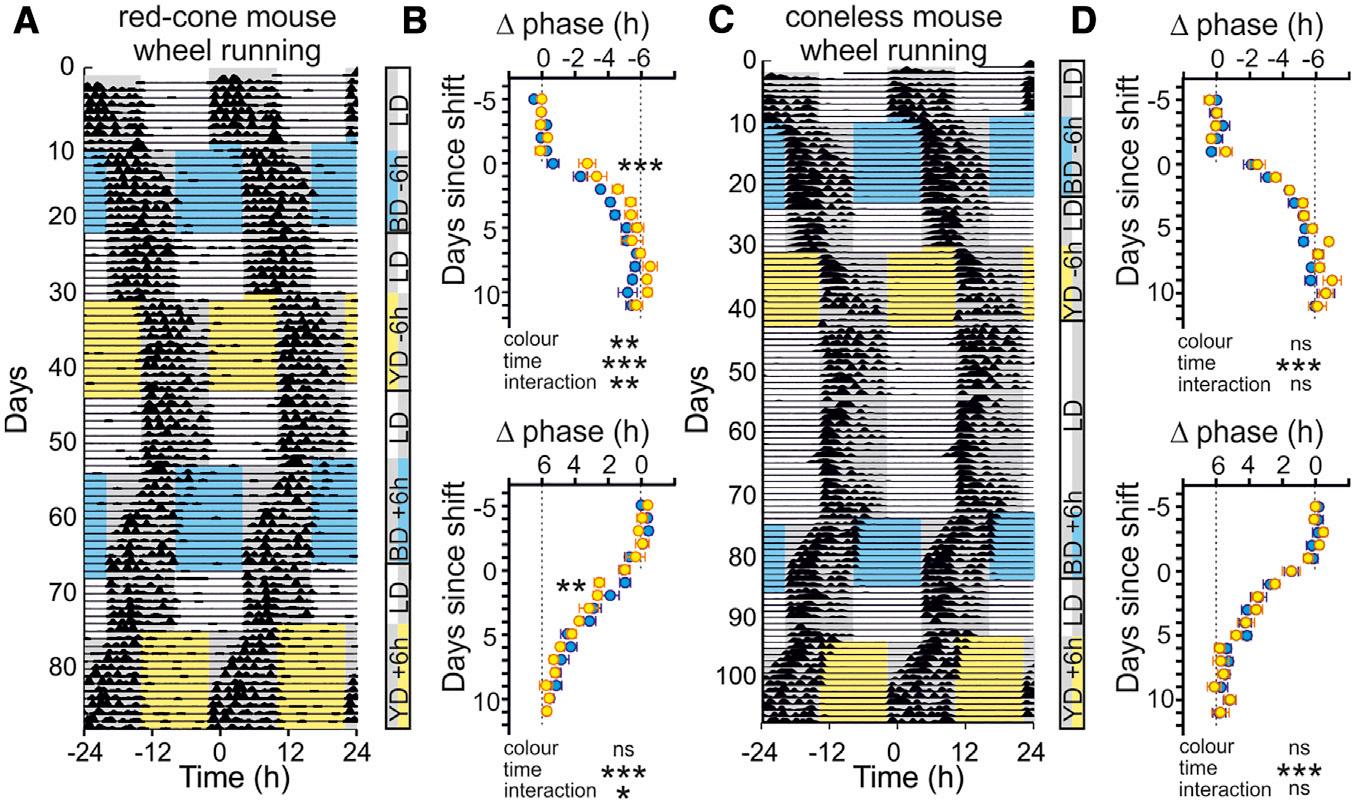
Figure2.ColorModulatesRe-entrainment followingJetLag
(A)Representativeactogramforred-cone mouseunder12:12LDcyclesandsubsequently exposedto6-hdelaysandadvanceswherethe lightphasewasrenderedinL S+(blue)or L+S (yellow)at0.1 3 intensitylevelsshownin Figure1A.
(B)Mean±SEMphasechange(mid-pointbetweenactivityonsetsandoffsets,normalizedto pre-shiftaverageforeachmouse)forred-cone mice(n=8)duringL S+(blue)andL+S (yellow) shifts.Dataanalyzedbytwo-wayRMANOVA withSidak’spost-testsareshown.Delays(top panel):time,F16,112 =103.5;p<0.0001;color, F1,7 =24.2;p=0.002;color 3 time,F 16,112 = 2.3,p=0.007.Advances(bottompanel):time, F16,112 =99.3;p<0.0001;color,F1,7 =2.45;p= 0.16;color 3 time,F16,112 =1.7;p=0.049.
(C)Sameas(A)butforconelessmouse. (D)Sameas(B)butforconelessmice.Two-way RMANOVAisshown.Delays(toppanel;n=7): time,F 16,96 =143.8;p<0.0001;color,F 1,6 =5.17;p=0.06;color 3 time,F 16,96 =1.60;p=0.08.Advances(bottompanel;n=8):time,F 16,112 =133.2;p< 0.0001;color,F1,7 =0.05;p=0.84;color 3 time,F16,112 =0.56;p=0.91. *p<0.05,**p<0.01,and***p<0.001,respectively;ns=p>0.05.Seealso FigureS2 fordetailsofresponsestoacutepulsesofL S+(blue)andL+S (yellow) stimuli.
dailyvariationsinlightintensityareverymarkedlyreduced[12]. Moreover,giventhereducedexposuretonaturallightassociatedwithmodernlife,suchaneffectofcolor(ifpresent)could havesubstantialpracticalsignificance.
Ininitialexperiments,weevaluatedwhethermicecouldmaintainentrainmentinthepresenceoflargediurnalvariationsin colorwithoutanyassociatedchangeinlightintensity.Accordingly,wefirstentrainedmicetoaconventional12h:12hLDcycle andthenreplacedthelightphasewitheitherL+S (yellow)or equi-luminantL S+(blue)andthedarkphasewiththeopposite color(Figure3A;n=6/condition;spectrain FigureS3A).In bothcases,miceimmediatelylostentrainmentandfreeran withanelongatedperiod(Figure3B).Thus,evenfairlylargevariationsincolordonotactasanindependentzeitgeberforthe circadiansystem,implyingthatcolorinsteadexertsitseffects bymodulatingresponsestovariationsinlightintensity.
Wenexttheninvestigatedwhetherdailychangesincolor wouldfacilitateentrainmenttoverylowamplitudediurnal variationsinlightintensitybygeneratingtwonewsetsoflightingconditions.Thefirstprovidedamodest(0.75logunit)daily variationinintensityformelanopsinandrods(‘‘mel/rod’’; FigureS3 B),withnochangeincolororconeilluminance.The secondprovidedanequivalentdailyvariationinmelanopsin/ rodactivationwhilepresentingasimultaneouschangein color(‘‘col+mel’’;bluecoloralignedwiththedimphase; FigureS2 C).Asexpected,ontransitiontothemel/rodcondition, red-conemice(n=6)immediatelylostentrainment(Figures 3Cand S2D)andbegantofreerunwithalongperiod(Figure3D).Bycontrast,thisdisruptiveeffectofareduced diurnalvariationinlightintensitywasamelioratedbyinclusion ofcolorchanges.Specifically,althoughnoanimalsshowed un-interruptedentrainmentfollowingtheswitchformLDto col+mel,twoanimalsretained 24-hrhythmsindicativeof partialentrainment(Figures3Cand S2 D;remaininganimals freeran; Figure3C).Asaresult,acrossthegroup,wedid
notdetectasignificantperiodlengthening(Figure3D),asin theotherconditionstestedhere.
Insummary,thesedataprovidesomesupportfortheidea thatcolormayaidentrainmenttolow-amplitudelightdarkcycles butsuggestthat,forrobustentrainment,changesinlightintensitygreaterthanthoseachievableherearerequired.Infact, evenduringthearcticsummer,thediurnalchangeinlight intensitywillbeatleastdoublethatwhichweemployed(FigureS3E),althoughmuchofthediurnalcolorchangewould alsobelost(atleastformice).Accordingly,althoughwedonot discounttheideathatsomeanimalsusecolortohelpentrainmentundersuchconditions,itseemsunlikelythatthisistheprimaryroleofcolorinputtotheclockformostmammals.Instead, amoregloballyrelevantpotentialbenefitofusingcoloristo compensateforstochasticfluctuationsinthediurnalrhythmof lightintensity,e.g.,duetovariationsincloudcover[11, 12].
Cloudscanreduceambientlightlevelsby>10-fold,rendering thetimingofsunrise/sunsetambiguousforasystemthatrelies simplyonlightintensity.Thetwilightblueshift,however,isretainedirrespectiveofclouds[9–11].Totestwhetherthiscolorinformationbuffersclockentrainmentagainstweather-related changesinillumination,wedesignedanexperimentalparadigm toprovidenaturalisticcyclesofcolorand/orlightintensitythat incorporatedstochasticvariationstosimulatetheimpactof clouds(Figure4A).Ourlightingsystemallowedustorecreate (forred-conemice)muchofthenaturalvariationincolorand lightintensitythatawild-typemousewouldexperiencearound dawnandduskonclearandcloudydays(FiguresS4A–S4D). Wethenpresentedsuchstimuliascyclesof3days,modeled onanorthernlatitudesummer,withcontinuouslyvarying changesincloudcover(FiguresS4DandS4F;‘‘natural’’).For comparison,wefollowedthesewithmatchedcyclesproviding identicaldailychangesinlightintensitybutwherethecolor wasfixedthroughouttoresembleday(FiguresS4EandS4F;‘‘intensityonly’’).

Red-conemice(n=12)weretheninitiallyentrainedtoa16 h:8hLDcycle(providingstabledailychangesincolorand lightintensity)andthereafterexperiencednaturalisticdailyvariationsinlightintensitywithsimulatedcloudsthatincludedor lackedtheassociatedvariationsincolor(Figures4Aand4B). Analysisofthemeandailyactivitypatternsrevealedconsistent changesundernaturalversusintensity-onlycycles.Specifically,althoughtheoveralltimingwassimilarunderbothconditions(Figures4Cand S4G),themagnitudeofthediurnalvariationinactivitywascompressedintheabsenceofcolorsignals (Figure4C).Toquantifythiseffect,weusedanestablished metricofcircadianrhythmrobustness(‘‘interdailystability’’) [33],confirmingasignificantimpairmentunderintensity-only versusnaturaldays(Figure4E).
Thisobservationcouldreflecteitherasimplereductionin themagnitudeofdailyrhythmsintheabsenceofcolororan increaseintheday-dayvariabilityoftheseactivitypatterns. Subsequentanalysesimplicatedbothfactors.Hence,thepercentageofdailyactivitythatoccurredoutsidethenightwas increasedunderintensity-only(Figure4F),whilebin-binvariationinactivity(‘‘intradailyvariability’’)[33]wasequivalent underbothconditions(Figure4G).Thus,theintensity-only conditionwasassociatedwithareductionintheamplitude ofday-nightvariationsinactivitywithoutanysubstantive increaseinthefragmentationofactivitypatternsacross thedailycycle.Importantly,however,whenweanalyzedthe day-daysimilarityofactivitypatterns(bycalculatingthe
Figure3.ColorIsNotanIndependent TimingCuefortheCircadianClock
(A)Representativepassiveinfrared(PIR)-derived actogramsfortwored-conemicetransferredfrom 12:12LDtoalignedL+S :L S+(yellow:blue)or L S+:L+S (blue:yellow)cycles(spectraprovidedin FigureS3A).
(B)PeriodofactivityrhythmsunderLDand L+S :L S+(yellow:blue;top)orL S+:L+S (blue:yellow;bottom).Data(n=6inbothcases) arecomparedagainstanexpectedperiodof24h (one-samplettests)andbetweenconditions (pairedttests),showinganincreaseinperiod, above24h,inbothcases.
(C)PIR-derivedactogramsfortwored-conemice transferredfrom12:12LDtoalignedcycles providingmodestdailychangesinilluminationjust formelanopsinandrods(mel/rod; FigureS3B)or withsuperimposedchangesincolor(col+mel; FigureS3C).Note,twomiceretainedpartial entrainmentundercol+mel(showninleftpanel and FigureS3D)althoughotheranimalsfreeran withalongcircadianperiod(representative exampleinrightpanel).
(D)PeriodofactivityrhythmsunderLDandsubsequentmel/rod(top)orcol+melcycles(bottom). Data(n=6inbothcases)areanalyzedwithonesamplettestsandpairedttestsasabove.
***p<0.001.Spectralpowerdistributionsforall stimuliareprovidedin FigureS3
meanpairwisecorrelations),wefoundasignificantreduction underintensity-onlydays(Figure4H),indicatingthatdaily activitytimingwasmorevariableintheabsenceofcolor signals.
Insummary,naturalisticdiurnalvariationsincolorconfer enhancedrobustnessandstabilitytodailyactivitypatterns inthefaceofweather-relatedfluctuationsinlightintensity. Todeterminewhetherthisresultsfromaspecificimpacton thecircadiancontrolofactivity,interspersedbetweenblocks ofnaturalandintensity-onlydays,wealsoincluded24-h epochsofconstantdimilluminationofintermediatecolor(Figures4Aand4B).Importantly,intheabsenceofenvironmental cues,meandailyactivitypatternsexhibitedafurthercompressioninamplitudewhenanimalshadpreviouslyexperienced intensity-onlyversusnaturaldays(Figure4D).Accordingly, subsequentquantificationrevealedasimilarsetofchanges tothosedescribedabovebutofgreatermagnitude.Specifically,interdailystabilitywassignificantlyreducedfollowing intensity-onlyversusnaturaldays(Figure4E),andthiseffect wasassociatedwithsignificantincreasesinthepercentage ofdailyactivityoccurringoutsidetheprojectednight(Figure4F),increasedintradailyvariability(Figure4G),anda reductioninmeanday-daycorrelation(Figure4H).These datatherebyconfirmthatnaturalisticvariationsincolorsubstantiallyenhancetheamplitudeandstabilityofclock-driven behavioralrhythmswhenthediurnalvariationinlightintensity providesunreliabletiminginformation.

Figure4.DailyChangesinColorSupportStableEntrainmentintheFaceofWeather-RelatedVariationinLightIntensity
(A)Schematicofthelightexposureparadigmthatincludednaturalisticchangesincolorandintensitywithsuperimposedstochasticvariationstosimulateclouds. Leftandrightpanels,respectively,providequantificationofapparentcolorandconcurrentchangesinlightintensity;see FigureS4 foradditionaldetailsofstimuli.
(B)RepresentativePIR-derivedactogramsforared-conemouseunderthelightingscheduleshownin(A).Symbolsadjacenttothetracesindicate24-h epochs thatwereusedforsubsequentanalysis.
(C)Mean±SEMnormalizedactivitywaveformsforred-conemice(n=12)underdaysprovidingnaturalchangesincolorandintensityormatchedintensity-only days.
(D)Sameas(C)butfor24-hepochsofconstantdimilluminationfollowingnaturalorintensity-onlydays.
(E–H)Quantificationofrhythmrobustnessandstabilityforred-conemice(n=12)undernaturalorintensity-onlydays(diurnal)andsubsequentconstantroutine (circadian),analyzedthroughoutbypairedttest;(E)interdailystability(diurnal:p=0.02;circadianp=0.001);(F)percentactivityoccurring duringthe‘‘day’’/ projectedday(diurnal:p=0.004;circadianp=0.0002);(G)intradailyvariability(diurnal:p=0.26;circadian=0.02);(H)meanday-daycorrelationinactivity patterns(diurnal:0.025;circadian:0.005).See STARMethods forfurtherdetailsofanalysisprocedures.*p<0.05,**p<0.01,and***p<0.001.
DISCUSSION
Contrarytocommonbeliefs,itisyellowratherthanbluecolors thathavethestrongesteffectonthemammaliancircadiansystem.Thisrelationshipalignswithnaturalshiftsinthecolorof ambientillumination,detectableduringtwilightbymammals withdi-andtri-chromaticvisualsystems[12].Accordingly,we showthatthiscolorsignalsupportsrobustandstablecircadian-drivenbehaviorinthenaturalworld,wherestochasticvariationsinlightlevelsintroduceambiguitytointensityasasignalof timeofday.
Theoretically,reducedcircadianresponsestobluecolors couldariseindirectlyasaresultofchromaticchangesinpupil diameter;however,ourpreviousworkindicatesthatchromatic blue:yellowmodulationsdonotproduceobservablepupillary responsesinmice[17].Bycontrast,ouridentificationofasignificantproportionofcellswithintheSCNthatprocessconederivedchromaticsignals[9]providesasimpleanddirectneurobiologicaloriginfortheeffectsofcolorreportedhere.
Althoughtheobservedreductionsincircadianresponsesto stimulithatresembletwilightarelogicalfromanecological perspective,thiseffectissurprisinggiventheoverallpositive
relationshipbetweenSCNfiringandcircadianresetting[20, 34] andthefactthatmostchromaticSCNcellsareexcitedby bluecolors[9].Theimplicationthenisthatthese‘‘blue-ON’’ SCNcells,whoseresponsesalignwiththoseofarecentlyidentifiedsubtypeofchromaticipRGC[35],mayactivelyoppose circadianphaseresetting.Becausevasopressin-expressing SCNneuronsarebelievedtoopposelight-drivencircadian resetting[36],anintriguingpossibilityisthatthispopulationcorrespondstothosethatprocessblue-ONsignals[4].
Anespeciallypertinentquestion,however,iswhethertheeffectsofcolordescribedhereextendtoothermammals,such ashumans.Thequalitativerelationshipbetweensunposition andblue-yellowcolorshouldberetainedforanymammal capableofcolorvision[12],andtheoreticalstudiessuggest thatcolorcouldaidcircadianentrainmentinhumans[11]. ExistingevidenceforcoloropponencyinprimateipRGCsand melanopsin-dependentresponsesinman[37–40]givefurther reasonstobelievethattheeffectsofcolorreportedherecould extendalsotohumans.Todate,however,muchofourcurrent understandingofthespectralsensitivityofthehumancircadian systemhasbeeninferredbasedonacute‘‘non-visual’’responses,suchasmelatoninsuppression.Consistentwitha
veryrecentobservationthatS-coneselectivemodulationsdo notnoticeablyinfluencesuchresponses[41],acutesuppression ofmelatoninbylightappearstobeprimarilydrivenbymelanopsin[42].Suchresponsesdonotalwaysprovideareliable proxyforcircadianphotosensitivity[43],however.Indeed,direct investigationsofhumancircadianresettingrevealthatlow-intensity,short-wavelengthlight(460nm)producessmallerresponsesthanlongerwavelengthlight(555nm)ofequivalentmelanopicilluminance[3].Thesedataarethereforeconsistentwith thecircadianeffectsofcolorweidentifyinmice.
Suchanarrangementispotentiallyimportantforpracticalapproachesintendedtoadjustthecircadianimpactofartificial light.Currentapproachestypicallyrelyonmanipulatingtheratio ofshort-andlong-wavelengthlight,achievingmodestdifferencesinmelanopicilluminanceattheexpenseofperceptible changesincolor[44].Asaresult,stimuliwithhighmelanopsin excitationappear‘‘bluer’’(andviceversa).Astrongprediction ofourresearchisthatthesechangesincolormayopposeany benefitsobtainedfrommodulatingmelanopsinphotoncapture. Recentworkindicatesthatmelanopsin-directedmodulations thatlackperceptibledifferenceincolorexertbeneficialeffects [45, 46].Ourdatanowsuggestthatsupplementingsuchapproacheswithcolorchangesoftheappropriatedirectioncould beespeciallyeffectiveatmodulatingcircadianresponses.
STAR+METHODS
Detailedmethodsareprovidedintheonlineversionofthispaper andincludethefollowing:
d KEYRESOURCESTABLE
d LEADCONTACTANDMATERIALSAVAILABILITY
d EXPERIMENTALMODELANDSUBJECTDETAILS
B Animals
d METHODDETAILS
B LightSources
B LightStimuli
B BehavioralParadigms
d QUANTIFICATIONANDSTATISTICALANALYSIS
B 1.Effectofcoloronclockspeed
B 2.Jet-lagparadigm
B 3.Acutephasere-setting
B 4/5.Color-onlyandColorwithlowamplitudediurnal lightingchanges
B 6.NaturalEntrainmentparadigm
d DATAANDCODEAVAILABILITY
SUPPLEMENTALINFORMATION
SupplementalInformationcanbefoundonlineat https://doi.org/10.1016/j. cub.2019.10.028
ACKNOWLEDGMENTS
ThisworkwassupportedbyagrantfromtheBiotechnologicalandBiological SciencesResearchCouncilUK(B/N014901/1).
AUTHORCONTRIBUTIONS
T.M.B.,R.J.L.,andJ.W.M.designedtheexperiments.F.M.,J.W.M.,and T.M.B.constructedandcalibratedtheexperimentalapparatus.J.W.M.and
A.W.performedtheexperiments.J.W.M.,A.W.,andT.M.B.performedthe analysis.T.M.B.,R.J.L.,andJ.W.M.wrotethemanuscript.
DECLARATIONOFINTERESTS
Theauthorsdeclarenocompetinginterests.
Received:August1,2019
Revised:September19,2019
Accepted:October16,2019
Published:December16,2019
REFERENCES
1. Wright,H.R.,andLack,L.C.(2001).Effectoflightwavelengthonsuppressionandphasedelayofthemelatoninrhythm.Chronobiol.Int. 18, 801–808
2. Wright,H.R.,Lack,L.C.,andKennaway,D.J.(2004).Differentialeffectsof lightwavelengthinphaseadvancingthemelatoninrhythm.J.PinealRes. 36,140–144
3. Gooley,J.J.,Rajaratnam,S.M.,Brainard,G.C.,Kronauer,R.E.,Czeisler, C.A.,andLockley,S.W.(2010).Spectralresponsesofthehumancircadian systemdependontheirradianceanddurationofexposuretolight.Sci. Transl.Med. 2,31ra33
4. Brown,T.M.(2016).Usinglighttotellthetimeofday:sensorycodinginthe mammaliancircadianvisualnetwork.J.Exp.Biol. 219,1779–1792
5. Provencio,I.,andFoster,R.G.(1995).Circadianrhythmsinmicecanbe regulatedbyphotoreceptorswithcone-likecharacteristics.BrainRes. 694,183–190
6. Yoshimura,T.,andEbihara,S.(1996).Spectralsensitivityofphotoreceptorsmediatingphase-shiftsofcircadianrhythmsinretinallydegenerate CBA/J(rd/rd)andnormalCBA/N(+/+)mice.J.Comp.Physiol.A Neuroethol.Sens.NeuralBehav.Physiol. 178,797–802
7. Hattar,S.,Lucas,R.J.,Mrosovsky,N.,Thompson,S.,Douglas,R.H., Hankins,M.W.,Lem,J.,Biel,M.,Hofmann,F.,Foster,R.G.,andYau, K.W.(2003).Melanopsinandrod-conephotoreceptivesystemsaccount forallmajoraccessoryvisualfunctionsinmice.Nature 424,76–81
8. Bailes,H.J.,andLucas,R.J.(2013).Humanmelanopsinformsapigment maximallysensitivetobluelight(lmax z 479nm)supportingactivationof G(q/11)andG(i/o)signallingcascades.Proc.Biol.Sci. 280,20122987
9. Walmsley,L.,Hanna,L.,Mouland,J.,Martial,F.,West,A.,Smedley,A.R., Bechtold,D.A.,Webb,A.R.,Lucas,R.J.,andBrown,T.M.(2015).Colour asasignalforentrainingthemammaliancircadianclock.PLoSBiol. 13, e1002127
10. Spitschan,M.,Aguirre,G.K.,Brainard,D.H.,andSweeney,A.M.(2016). Variationofoutdoorilluminationasafunctionofsolarelevationandlight pollution.Sci.Rep. 6,26756
11. Woelders,T.,Wams,E.J.,Gordijn,M.C.M.,Beersma,D.G.M.,andHut, R.A.(2018).Integrationofcolorandintensityincreasestimesignalstability forthehumancircadiansystemwhensunlightisobscuredbyclouds.Sci. Rep. 8,15214
12. Spitschan,M.,Lucas,R.J.,andBrown,T.M.(2017).Chromaticclocks:coloropponencyinnon-image-formingvisualfunction.Neurosci.Biobehav. Rev. 78,24–33
13. Lall,G.S.,Revell,V.L.,Momiji,H.,AlEnezi,J.,Altimus,C.M.,Guler,A.D., Aguilar,C.,Cameron,M.A.,Allender,S.,Hankins,M.W.,andLucas,R.J. (2010).Distinctcontributionsofrod,cone,andmelanopsinphotoreceptorstoencodingirradiance.Neuron 66,417–428
14. Altimus,C.M.,Guler,A.D.,Alam,N.M.,Arman,A.C.,Prusky,G.T., Sampath,A.P.,andHattar,S.(2010).Rodphotoreceptorsdrivecircadian photoentrainmentacrossawiderangeoflightintensities.Nat.Neurosci. 13,1107–1112
15. Guler,A.D.,Ecker,J.L.,Lall,G.S.,Haq,S.,Altimus,C.M.,Liao,H.W., Barnard,A.R.,Cahill,H.,Badea,T.C.,Zhao,H.,etal.(2008).Melanopsin
cellsaretheprincipalconduitsforrod-coneinputtonon-image-forming vision.Nature 453,102–105
16. Spitschan,M.,andWoelders,T.(2018).Themethodofsilentsubstitution forexaminingmelanopsincontributionstopupilcontrol.Front.Neurol. 9, 941
17. Hayter,E.A.,andBrown,T.M.(2018).Additivecontributionsofmelanopsin andbothconetypesprovidebroadbandsensitivitytomousepupilcontrol. BMCBiol. 16,83
18. Brown,T.M.,Gias,C.,Hatori,M.,Keding,S.R.,Semo,M.,Coffey,P.J., Gigg,J.,Piggins,H.D.,Panda,S.,andLucas,R.J.(2010).Melanopsincontributionstoirradiancecodinginthethalamo-corticalvisualsystem.PLoS Biol. 8,e1000558
19. Brown,T.M.,Tsujimura,S.,Allen,A.E.,Wynne,J.,Bedford,R.,Vickery,G., Vugler,A.,andLucas,R.J.(2012).Melanopsin-basedbrightnessdiscriminationinmiceandhumans.Curr.Biol. 22,1134–1141
20. Brown,T.M.,Wynne,J.,Piggins,H.D.,andLucas,R.J.(2011).Multiplehypothalamiccellpopulationsencodingdistinctvisualinformation. J.Physiol. 589,1173–1194
21. Allen,A.E.,Brown,T.M.,andLucas,R.J.(2011).Adistinctcontributionof short-wavelength-sensitiveconestolight-evokedactivityinthemouse pretectalolivarynucleus.J.Neurosci. 31,16833–16843
22. Allen,A.E.,Procyk,C.A.,Howarth,M.,Walmsley,L.,andBrown,T.M. (2016).Visualinputtothemouselateralposteriorandposteriorthalamic nuclei:photoreceptiveoriginsandretinotopicorder.J.Physiol. 594, 1911–1929
23. Allen,A.E.,Storchi,R.,Martial,F.P.,Bedford,R.A.,andLucas,R.J.(2017). Melanopsincontributionstotherepresentationofimagesintheearlyvisualsystem.Curr.Biol 27,1623–1632.e4
24. Allen,A.E.,Storchi,R.,Martial,F.P.,Petersen,R.S.,Montemurro,M.A., Brown,T.M.,andLucas,R.J.(2014).Melanopsin-drivenlightadaptation inmousevision.Curr.Biol. 24,2481–2490
25. Smallwood,P.M.,Olveczky,B.P.,Williams,G.L.,Jacobs,G.H.,Reese, B.E.,Meister,M.,andNathans,J.(2003).Geneticallyengineeredmice withanadditionalclassofconephotoreceptors:implicationsfortheevolutionofcolorvision.Proc.Natl.Acad.Sci.USA 100,11706–11711
26. Shuboni,D.D.,Cramm,S.L.,Yan,L.,Ramanathan,C.,Cavanaugh,B.L., Nunez,A.A.,andSmale,L.(2015).Acuteeffectsoflightonthebrainand behaviorofdiurnalArvicanthisniloticusandnocturnalMusmusculus. Physiol.Behav. 138,75–86
27. Biel,M.,Seeliger,M.,Pfeifer,A.,Kohler,K.,Gerstner,A.,Ludwig,A., Jaissle,G.,Fauser,S.,Zrenner,E.,andHofmann,F.(1999).Selective lossofconefunctioninmicelackingthecyclicnucleotide-gatedchannel CNG3.Proc.Natl.Acad.Sci.USA 96,7553–7557
28. Dkhissi-Benyahya,O.,Gronfier,C.,DeVanssay,W.,Flamant,F.,and Cooper,H.M.(2007).Modelingtheroleofmid-wavelengthconesincircadianresponsestolight.Neuron 53,677–687
29. Altimus,C.M.,Guler,A.D.,Villa,K.L.,McNeill,D.S.,Legates,T.A.,and Hattar,S.(2008).Rods-conesandmelanopsindetectlightanddarkto modulatesleepindependentofimageformation.Proc.Natl.Acad.Sci. USA 105,19998–20003
30. Dobb,R.,Martial,F.,Elijah,D.,Storchi,R.,Brown,T.M.,andLucas,R.J. (2017).Theimpactoftemporalmodulationsinirradianceunderlight adaptedconditionsonthemousesuprachiasmaticnuclei(SCN).Sci. Rep. 7,10582
31. Daan,S.,andPittendrigh,C.S.(1976).Afunctionalanalysisofcircadian pacemakersinnocturnalrodents:II.Thevariabilityofphaseresponse curves.J.Compar.Physiol 106,253–266
32. Brown,L.A.,Hasan,S.,Foster,R.G.,andPeirson,S.N.(2016).COMPASS: continuousopenmousephenotypingofactivityandsleepstatus. WellcomeOpenRes. 1,2
33. vanSomeren,E.J.,Hagebeuk,E.E.,Lijzenga,C.,Scheltens,P.,deRooij, S.E.,Jonker,C.,Pot,A.M.,Mirmiran,M.,andSwaab,D.F.(1996). Circadianrest-activityrhythmdisturbancesinAlzheimer’sdisease.Biol. Psychiatry 40,259–270
34. Meijer,J.H.,Rusak,B.,andGanshirt,G.(1992).Therelationbetweenlightinduceddischargeinthesuprachiasmaticnucleusandphaseshiftsof hamstercircadianrhythms.BrainRes. 598,257–263
35. Stabio,M.E.,Sabbah,S.,Quattrochi,L.E.,Ilardi,M.C.,Fogerson,P.M., Leyrer,M.L.,Kim,M.T.,Kim,I.,Schiel,M.,Renna,J.M.,etal.(2018). TheM5cell:acolor-opponentintrinsicallyphotosensitiveretinalganglion cell.Neuron 97,150–163.e4
36. Yamaguchi,Y.,Suzuki,T.,Mizoro,Y.,Kori,H.,Okada,K.,Chen,Y.,Fustin, J.M.,Yamazaki,F.,Mizuguchi,N.,Zhang,J.,etal.(2013).Micegenetically deficientinvasopressinV1aandV1breceptorsareresistanttojetlag. Science 342,85–90
37. Dacey,D.M.,Liao,H.W.,Peterson,B.B.,Robinson,F.R.,Smith,V.C., Pokorny,J.,Yau,K.W.,andGamlin,P.D.(2005).Melanopsin-expressing ganglioncellsinprimateretinasignalcolourandirradianceandproject totheLGN.Nature 433,749–754
38. Spitschan,M.,Jain,S.,Brainard,D.H.,andAguirre,G.K.(2014).Opponent melanopsinandS-conesignalsinthehumanpupillarylightresponse. Proc.Natl.Acad.Sci.USA 111,15568–15572
39. Woelders,T.,Leenheers,T.,Gordijn,M.C.M.,Hut,R.A.,Beersma,D.G.M., andWams,E.J.(2018).Melanopsin-andL-cone-inducedpupilconstrictionisinhibitedbyS-andM-conesinhumans.Proc.Natl.Acad.Sci. USA 115,792–797
40. Cao,D.,Nicandro,N.,andBarrionuevo,P.A.(2015).Afive-primaryphotostimulatorsuitableforstudyingintrinsicallyphotosensitiveretinalganglion cellfunctionsinhumans.J.Vis 15,15.1.27
41. Spitschan,M.,Lazar,R.,Yetik,E.,andCajochen,C.(2019).Noevidence foranSconecontributiontothehumancircadianresponsetolight.Curr. Biol. 29,R1297–R1298
42. Prayag,A.S.,Najjar,R.P.,andGronfier,C.(2019).Melatoninsuppression isexquisitelysensitivetolightandprimarilydrivenbymelanopsininhumans.J.PinealRes. 66,e12562
43. Rahman,S.A.,StHilaire,M.A.,Gronfier,C.,Chang,A.M.,Santhi,N., Czeisler,C.A.,Klerman,E.B.,andLockley,S.W.(2018).Functionaldecouplingofmelatoninsuppressionandcircadianphaseresettinginhumans. J.Physiol. 596,2147–2157
44. Tahkamo,L.,Partonen,T.,andPesonen,A.K.(2019).Systematicreviewof lightexposureimpactonhumancircadianrhythm.Chronobiol.Int. 36, 151–170
45. Allen,A.E.,Hazelhoff,E.M.,Martial,F.P.,Cajochen,C.,andLucas,R.J. (2018).Exploitingmetamerismtoregulatetheimpactofavisualdisplay onalertnessandmelatoninsuppressionindependentofvisualappearance.Sleep(Basel) 41,zsy100
46. Souman,J.L.,Borra,T.,deGoijer,I.,Schlangen,L.J.M.,Vlaskamp,B.N.S., andLucassen,M.P.(2018).Spectraltuningofwhitelightallowsforstrong reductioninmelatoninsuppressionwithoutchangingilluminationlevelor colortemperature.J.Biol.Rhythms 33,420–431
47. Govardovskii,V.I.,Fyhrquist,N.,Reuter,T.,Kuzmin,D.G.,andDonner,K. (2000).Insearchofthevisualpigmenttemplate.Vis.Neurosci. 17, 509–528
48. Jacobs,G.H.,andWilliams,G.A.(2007).ContributionsofthemouseUV photopigmenttotheERGandtovision.Doc.Ophthalmol. 115,137–144
49. Sokolove,P.G.,andBushell,W.N.(1978).Thechisquareperiodogram:its utilityforanalysisofcircadianrhythms.J.Theor.Biol. 72,131–160
STAR+METHODS
KEYRESOURCESTABLE
REAGENTorRESOURCESOURCEIDENTIFIER
ExperimentalModels:Organisms/Strains
Mouse: Opn1mwR
Mouse: Cnga3 /
DrJeremyNathans,JohnsHopkins University
MGICat#2678771,RRID:MGI:2678771
MGICat#3723602,RRID:MGI:3723602 SoftwareandAlgorithms
TheChronobiologyKit:KitCollect, KitMonitor,KitAnalyzer
StanfordSoftwareSystems
https://query.com/chronokit/ MATLABR2017aMathWorks
https://uk.mathworks.com/products/ matlab.html
GraphPadPrism7.04GraphPad
http://www.graphpad.com/ Pythonv2.7.10Python https://www.python.org/downloads/ Other
RGBLEDstripsExpertElectricalSuppliesLEDST60RGB UVLEDStripsExpertElectricalSuppliesLEDST60UV/385NM RGBWbulbsLIFXLIFXColor1000 UVbulbsLedEnginLZ1-00UA00-00U7 405nmLEDThorlabsM405L4 460nmLEDThorlabsM455L4 630nmLEDThorlabsM625L4 PTFEdiffusingsheetDirectPlasticsPTS01004 Neutraldensitygel2110.9NDLeeFilters
AvianDCoating,WhitepaintAvianTechnologies
http://www.leefilters.com/lighting/ colour-details.html#211
https://aviantechnologies.com/product/ avian-d-white-reflectance-coating/ COMPASSPassiveinfraredsystem(PIR)LawrenceBrown[32]NA
LEADCONTACTANDMATERIALSAVAILABILITY
Thisstudydidnotgeneratenewuniquereagents.Furtherinformationandrequestsforresourcesshouldbedirectedtoandwillbe fulfilledbytheLeadContact,TimBrown(Timothy.Brown@manchester.ac.uk ).
EXPERIMENTALMODELANDSUBJECTDETAILS
Animals
AllexperimentsreceivedinstitutionalethicscommitteeapprovalandinaccordancewithUKAnimals(ScientificProcedures)Act 1986,andEuropeanDirective2010/63/EU.Adult(>8weeks)malemicefromaC57BL/6backgroundstrainwereusedthroughout. ForthemajorityoftheexperimentsweusedmiceexpressingthehumanL-coneopsininplaceoftheirnativeM-coneopsin (Opn1mwR [25];).Additionalexperimentsusedmicelackingthecone-specificcyclicnucleotidegatedchannelalphasubunit (Cgna3 / )[27]).
Micewereindividuallyhousedwith adlibitum foodandwater.Cageswerelocatedinlighttightcabinetswheretheambientlight couldbecarefullycontrolled.Underseveralparadigms(Constantlight,andbothphaseshiftsexperiments)micewerealsohoused withaccesstoarunningwheeltoassesslocomotoractivity.
METHODDETAILS
LightSources
Housingenvironment1
Onelighttightcabinet(usedtogeneratedatain Figures1 and 2)wasfittedwithfourparallelrowsofRGB(SMD5050)andultraviolet (385nm)LEDstripsLEDintensitieswithdriverstoprovidepulse-widthmodulationbasedregulationofLEDintensity(ExpertElectrical
,4260–4267.e1–e4,December16,2019
SuppliesLtd.;Rochdale,UK).A1.5mmthickPTFEdiffusingsheet(Directplastics;Sheffield,UK)wasinstalled 5cmbelowtheLEDs toprovideuniformillumination.Neutraldensity(ND)gelssheets(LeeFilters;Andover,UK)werefittedtothediffusertoreducelight intensityby10or100foldasrequired(ND1andND2respectively).
Housingenvironment2
Fourcustombuiltlightboxeswereaffixedabovetheroofofthesecondcabinet.EachlightboxconsistedoftwosmartRGBWbulbs (LIFXA60;LIFX,Cremorne,Australia)and6violetbulbs(405nm,LedEnginLZ1-00UA00-00U7;RSComponents,ManchesterUK),to allowfor5-primaryillumination.APTFEdiffuserwasmountedtoflooroflightboxes/roofofthecabinetandtheinteriorofthecabinet waspaintedwhitetoprovideuniformillumination.TheLIFXbulbswereconnectedwirelesslyoveralocalnetwork,UVbulbswere connectedtoLEDdrivers(T-Cube;Thorlabs,Ely,UK)viaamultichannelanalogoutputmodule(NI9264;NationalInstruments, TX,USA).LEDintensitieswerethencontrolledonasecondbysecondbasisusingaPCrunningPython(2.7.10).Neutraldensity gelswereaddedtoeachboxtoadjusttheoverallbrightnessasrequired.
Phasere-settingchamber
Stimuliwerepresentedviaacustomlightsource(componentsfromThorlabs)consistingofthreeindependentlycontrollableLEDS (405nm,460nmand630nm)combinedbydichroicmirrors.Lightstimuliwerethenandprojected(viaaconcavelens)ontoa1.5mm thickPTFEdiffusingsheet(Directplastics)thatsatontopofacylindricalchambercoatedwithspectrallyneutralreflectivepaint(Avian Dcoating,AvianTechnologiesLLC,NH,USA).
LightStimuli
Stimuliweredesignedandcalibratedasdescribedpreviously[9, 17]usingcalibratedspectroradiometers(DMc150;Bentham InstrumentsLtd,UK,andSpectroCal,CambridgeResearchSystems,UK).Inbrief,photonabsorptionforeachphotopigmentwas calculatedusingGovardovskiinomograms([47]withpeakabsorbanceat:S-opsin,365nm;L-opsin:556nm;melanopsin,480nm; rhodopsin,498nm)adjustedforlenstransmission[48].Inmostcases,stimuliweredesignedtoprovideidenticalexcitationof melanopsinandrodsandtoprovidethesameaverageilluminanceforL-andS-coneopsinbuttodifferintherationofL-versus S-opsinexcitation(i.e.,color).Inothercases,stimuliweredesignedtomodulateilluminanceforcones(orrod/melanopsinexcitation) withoutchangingcolor,ortosimultaneouslymodulatecolorandintensity.Spectraandrelevantquantificationforallstimuliare provideintherelevantfiguresorassociatedsupplementalfigures.
BehavioralParadigms
Forexperimentalparadigms1-3outlinedbelow(datashownin Figures1 and 2 andassociatedsupplementalfigure),micewere housedwitharunningwheeltoassesslocomotoractivity.Wheelrevolutionswereacquiredin60sbinsusingTheChronobiology Kit(StanfordSoftwareSystems,SantaCruz,CA).Micewerehousedwiththerunningwheelforatleastoneweekbeforestarting theexperimentalprotocols.Inmostcases,lightexposureanddataacquisitionwasperformedinhousingenvironment1withthe exceptionofacutephaseresettingassays(seebelow).Forexperimentalparadigms4-6,weusedhousingenvironment2and measuredcageactivityviaapassiveinfrared(PIR)system,asdescribedpreviously[32].
1.Effectofcoloronclockspeed
Eight Opn1mwR miceaged23weeksand7 Cgna3 / miceaged14-19weekswereputintoconstantlightconditionsatND2. EverytwoweeksthelightsalternatedbetweenL-S+(‘blue’)andL+S-(‘yellow’)conditions(seespectraandquantificationin Figure1A), andevery4weeksanNDgelwasremovedfromthelightsincreasingtheirradianceby10-fold.Asecondbatchofmice(14 Opn1mwR miceaged10-15weeks)werehousedunderconstantlightconditions(atND1)andreceived2successive2weekblocksofL+S-(‘yellow’),L-S+(‘blue’),L-S-(‘dim’)andL+S+(‘bright’)coneselectivestimuli(see Figure1H).
2.Jet-lagparadigm
Eight Opn1mwR miceaged14-22weeksand8 Cgna3 / miceaged9-17weekswerehousedunderaconventional12h:12hLDcycle (fluorescentlighting;effectivephotonflux=14.5,14.7,15and13logphotons/cm2/sformelanopsin,rhodopsin,L-coneandS-cone opsinrespectively).Afteroneweekthemicewerethensubjectedtoa6hourphasedelayorphaseadvancewiththelightphase renderedineitherL-S+(‘blue’)orL+S-(‘yellow’)asin Figure1A.Followingre-entrainment(>12days)tothenewLDcycle,mice werereturnedtoconventionalLDforatleast1weekpriortothenextstimulus.
3.Acutephasere-setting
Sixteen Opn1mwR miceaged6-16weekswerehousedunderstandard12h:12hLDfor2weeks.Subsequently,duringearly(ZT1415)orlatenight(ZT21-22)miceweretransferredtoatestchamber(describedabove)wheretheywereilluminatedfor5minuteswith L-S+(‘blue’)orL+S-(‘yellow’)illumination(spectraandquantificationin FigureS2A).TransferwasperformedindarknessusingIR goggles(ATNNVG-7,ArmasightInc.,NH,USA)andsubsequentlymicewerereturnedtotheirhomecageunderconstantdarkness forafurther10days.Micewerethenreturnedto12h:12hLDfortwoweekspriortoreceivinganotherteststimulus.
4.Color-onlyentrainmentparadigm
Opn1mwR mice(n=6/condition)aged16-19weekswerehousedforaweekundera12h:12hLDcycle(lightphaseeffectivephoton flux=13.9,13.8,14,12.9logphotons/cm2/sformelanopsin,rhodopsin,L-coneandS-coneopsinrespectively).Afterentrainingto theseconditions,theLDcyclewaschangedtoa12h:12h‘yellow’:’blue’or‘blue’:’yellow’cycle(spectraandquantificationin FigureS3A).
5.Colorwithlowamplitudediurnallightingchanges
Six Opn1mwR miceaged10-11weekswerehousedforaweekundera12h:12hLDcycle(daycomponentwasthe‘mel/rod+’stimuli from FigureS3B).SubsequentlythedarkphaseoftheLDcyclewasreplacedbya‘mel/rod-‘stimulus(FigureS3B)suchthatthe diurnallightcycleprovidedamodestdifferenceinmelanopsinandrodilluminationbutnochangeincolororconeilluminance.After 2weeks,micewerereturnedtoLDtore-entrainfor10days.Finally,miceweretransferredtoanewdiurnalcyclethatprovidedan identicalchangeinmelanopsinandrodexcitationtothatusedabovebutwhichalsoincorporatedlargechangesincolor(spectraand quantificationin FigureS3C).
6.NaturalEntrainmentparadigm
Twobatchesof6 Opn1mwR mice(11-13&15-18weeks)wereusedforthisexperiment.Micewerehousedundera16h:8h lightingcyclethatprovidedadailyvariationincolorandintensityresemblingchangesoccurringbetweensolarelevationsof+6 and 4degreesrelativetothehorizon(FiguresS4AandS4B).Micesubsequentlyexperienced5repeatingepochsconsistingofi) a3dayblockofnaturalisticcolorandintensitychangeswithsmoothtwilighttransitionsandcontinuouslyvaryingfluctuationssimulatingcloudsorii)amatched3-dayblockprovidingidenticalchangesinlightintensitybutwherecolorwasfixedtoresembleday(See Figures4Aand S4D–S4F).Eachblockwasterminatedbya24hepochofconstantdimilluminationofintermediatecolor.Forthenaturalandintensityonlycyclesdescribedabovefluctuationssimulatingcloudswererandomlygenerated apriori suchthatindividual daysweredistinctfromanyotherdayunderthesamecondition.
QUANTIFICATIONANDSTATISTICALANALYSIS
Allstatisticalanalyseswereperformedusing(GraphPadPrism7.04;GraphPadSoftwareInc.,CA,USA),withcriteriaforsignificance setatp<0.05.Samplesizeandtheotherrelevantstatisticaldetailsareprovidedinthemaintext.
1.Effectofcoloronclockspeed Circadianperiodwasdeterminedby c2-periodogram[49],performedon2weekblocksofdataundereachexperimentalcondition. Activityboutduration(a)wasdefinedasthefactionofeachcircadiandaythatactivitywasabovethemean(byreferencetoperiod asdeterminedabove).ForsubsequentstatisticalcomparisonofeffectsofL-S+(‘blue’)versusL+S-(‘yellow’)stimuliatvaryingirradiance(Figures1B–1G),datawerethenanalyzedby2-wayRMANOVAwithcolorasarepeatedfactorandSidak’spost-testswhere ANOVArevealedsignificantmaineffectsofcolororinteractionwithintensity.Dataanalyzedforeachintensityexcludedthosecases whereanindividualexhibitednegligibleorinterruptedwheelrunningunderoneorbothofthetwotestedcolors(duetoblocked wheels).Thiswasthecasefor1red-conemouse(ND1dataexcluded)andtwoconelessmice(foroneindividualND2andND1 dataexcluded,forthesecondND0andND2dataexcluded).Oftheremainingconelessdata,oneanimalexhibitedanunexpectedly shortfree-runningperiodunderhighintensity‘yellow’illumination.Intheabsenceofanyoverttechnicalreasontorejectthisdata,it isincludedintheanalysis.Ofnote,however,re-analysis(2-wayRMANOVA)excludingND0datafromthatindividualproduced anoutcomeequivalenttothatreportedinthetext:asignificanteffectofirradiance(F2,13 =36.2;p<0.0001)butnotcolor(F1, 13 =0.84;p=0.38)orinteractionwithirradiance(F2,13 =1.94;p=0.18).
Forcomparisonofcircadianperiodeffectsofcolorand-coneilluminancesignals(performedatND1only, Figures1Iand1J)data wasanalyzedbyone-wayRMANOVAwithDunnett’spost-tests(nodataexcluded).
2.Jet-lagparadigm
Circadianphasewasassessedasthemidpointbetweenactivityonsetandoffsetoneachday(respectivelydefinedasthestartof a30minepochwhereactivityexceedthedailymeanandthestartofa90minepochwhereactivityfellbelowthedailymean).For analysis,circadianphasemarkersforeachmousewerenormalizedbysubtractingthemeanforthe5-dayepochprecedingashift intheLDcycle.ThedataforL-S+(‘blue’)andL+S-(‘yellow’)stimuliundereachconditionwerethenanalyzedbytwo-wayRMANOVA, withSidak’spost-testswherewedetectedmaineffectsofcolororcolorXtimeinteractions.Foranalysesofred-conemicethere werenoexclusions.Foranalysesofconelessmice,oneanimalwasexcludedfromanalysisofdelayshiftsduetopoorrunningduring thefirstLDepochthatpreventedreliabledeterminationofstartingphasepriorto‘blue’delay.Analysisperformedasdescribedabove butusingjustonsetsoroffsetsrevealedqualitativelyequivalentimpactofcolortothatreportedinthemanuscript:RedconeDelay (Onsets)-Time(F16,112 =83.8;p<0.0001),color(F1,7 =6.5;p=0.038),interaction(F16,112 =2.5;p=0.0032);RedconeDelay(Offsets)-Time(F16,112 =43.8;p<0.0001),color(F1,7 =30.8;p=0.0009),interaction(F16,112 =2.0;p=0.022);RedconeAdvance(Onsets)-Time(F16,112 =186.8;p<0.0001),color(F1,7 =7.4;p=0.03),interaction(F16,112 =1.3;p=0.19);RedconeAdvance(Offsets)Time(F16,112 =26.2;p<0.0001),color(F1,7 =0.04;p=0.85),interaction(F16,112 =2.2;p=0.0085).
3.Acutephasere-setting
Phaseshiftsweremeasuredmanuallyfromhighmagnificationactogramsbythreeexperiencedinvestigators(blindedtostimulus), byextrapolationofalineofbestfitthroughactivityonsetstothedayofthepulse.Valuesreportedinthemanuscriptaretheaverage ofthoseobtainedbythethreeinvestigators(whichwerereliablyinclosecorrespondence;mean±SDinter-ratervariability=12.7± 9.4min).Subsequentanalysisperformedbyunpairedttest.Datawereexcludedfromthisanalysisinafewcaseswhenvariabilityin
activityonsetsmadephase-shiftshardtoreliablymeasure(asassessedbyatleasttwooftheinvestigators);thiswasthecasefor1 individualfollowing‘blue’delay,1individualfollowing‘blue’advanceand3individualsfollowing‘yellow’delays(noindividuals excludedfollowing‘yellow’advances).
4/5.Color-onlyandColorwithlowamplitudediurnallightingchanges
Inallcases,circadianperiodwasdeterminedby c2-periodogram(asabove).Forstatisticalanalysesperiodestimatesobtainedunder LDconditionsandfollowingthesubsequentexperimentalmanipulationswerecomparedagainsteachotherbypairedttestsand againstanexpectedperiodof24hbyone-samplettests.Nodatawasexcluded.
6.NaturalEntrainmentparadigm
Analysesofdiurnalactivitypatternswerebasedonthelast48hofeach3dayblock(toreduceeffectsassociatedwiththetransition fromprecedingstimuli),foratotalof10daysofmatched‘natural’and‘intensity-only’cyclesforeachindividual.Analysesofcircadian activitypatternswerebasedonthe24hepochsofconstantroutineimmediatelyfollowing‘natural’and‘intensity-only’stimulus blocks,providingatotalof5daysforeachconditionperindividual.Todeterminemeanactivitywaveforms,PIRreportedactivity profilesforeachindividual(10minbins)werefirstsmoothedbya2hrunningmean.Wethenaveragedacrosstherelevantdays (asindicatedabove)andnormalizedbysubtractingthedailymeananddividingbythemaximaoftheresultingtimeseries.Onsets andoffsetsweredeterminedfromdailymeancrossingsofthe2hsmoothedtime-series,allotheranalyseswereperformedonunsmootheddataandanalyzedbypaired-testbetween‘natural’and‘intensity-only’conditions(orsubsequentconstantroutineas appropriate).InterdailyStability(ameasureofrhythmsrobustness)wasassessedbyquantifyingthefractionofvariancethatwas accountedforbyastable24hrhythm[33]usingtheformula(with n representingthetotalnumberofdatapoints, P thenumberof timebins):
Rhythmamplitudewasdeterminedbycalculatingthepercentageofdailyactivitythatoccurredoutsideofthe8hnight(orprojected night)epochacrosstherelevantselectionofdays.IntradailyVariability(ameasureofthefragmentationofactivitypatterns[33],)was assessedusingtheformula:
Theday-daysimilaritybetweenactivitypatternswasassessedbycalculatingthemeancorrelationcoefficient(Pearson’sr)between everypossiblepairofdaysundertherelevantexperimentalconditions(fordiurnalcycles45pairs,forconstantconditions10pairs).
DATAANDCODEAVAILABILITY
RawdataandanalysiscodewillbeprovideduponrequestbytheLeadContact,TimBrown(timothy.brown@manchester.ac.uk).
Another random document with no related content on Scribd:
"No sattuupa nyt kummia", sanoi hän. "Nyt juuri herratar – ei maaherra – on tarjonnut minulle laulutunteja. Kyllä he sanovat oikein kotona, kun väittävät, että Tunturihiiri on kuin kissa, hän putoaa aina jaloilleen."
"Te tulette luonnollisesti meille", sanoi Per Borting varmasti. "Ellette mieluummin valitse – Veyeriä", lisäsi hän pisteliäästi.
"Niin, teille minä mieluummin tahdon", sanoi Petra iloisesti, mutta sitten hän vaikeni ja seisoi hetken ääneti.
"Ei", sanoi hän päättäväisesti. "Minulla on paikka täällä. Ja kun hän tahtoo pitää minut luonaan tänä talvena, niin minun täytyy jäädä hänen luokseen. Etenkin nyt, kun hän on suruissaan ja yksin. Niin, sillä Veyer, hän matkustaa pois. Ulkomaille. Ja ajatelkaa, hän on muuttanut mieltään niin, että hän nyt pitää minusta, luullakseni, –vaikka minä nostin hänet ilmaan ihastuksissani. Eikä hän varmaankaan ole tottunut siihen. Niin että ei kiitos. Minun täytyy kyllä jäädä tänne."
Per Borting ei sanonut mitään. Mutta hänen avoimet pojankasvonsa ilmaisivat niin selvästi, mitä hän ajatteli, että sanat olivatkin aivan tarpeettomia.
"Mutta keväällä. Silloin kai te asutte kotona. Silloin minä tulen teille, ja silloin te ja minä istumme lammen luona, kun toiset nukkuvat päivällisuntaan – niin, ja sitten teidän äitinne ja minä laulamme yhdessä", lisäsi hän.
"Tuhat kiitosta", sanoi Per Borting onnellisena. Sillä Petrahan oli ensin ajatellut lampea ja häntä ja sitten vasta laulua ja äitiä. Ja
olihan hän, Borting, kaupungissa talvella – ja Veyer aikoi matkustaa pois.
Mutta Petra meni keittiöön Jennyn luo.
"On kai hirveän syntistä olla iloinen, kun täällä on suru", sanoi hän, "mutta minä en voi sille mitään. Minä olen sellainen onnen porsas, Jenny."
"Vai niin, se kesakkoinenko se sitten lopulta olikin", vastasi Jenny ja alkoi hihittää.
* * * * *
Lumi pyrysi ristiin rastiin raskaalta, valkean harmaalta taivaalta ja laskeutui valkeaksi, utuiseksi kanneksi sen harmaan peitteen ylitse, joka vielä oli maassa viime lumisateen jälkeen. Oli niin pimeää, että lyhdyt ulkona vielä paloivat, vaikka kello oli melkein kymmenen aamulla.
Oli kaksi päivää ennen jouluaattoa.
Petra seisoi nenä matkakirstussa ja järjesteli tavaroitaan. Maaherratar ilmestyi ovelle.
"Tässä minulla on joululahja – jos ehkä tahdotte käyttää sitä matkalla", sanoi hän ja ojensi Petralle pienen käsilaukun.
"Voi tuhannet kiitokset", sanoi Petra ja aukaisi sen. Sen sisässä oli neliskulmainen punainen seteli.
"Te olette unohtanut tämän", sanoi hän ja ojensi sen Letta rouvaa kohti.
"Se on teidän. Voitte kai käyttää sen ostoksiinne", sanoi maaherratar.
"Tehän olette itse voittanut sen, muistattehan."
Petra seisoi silmät selällään.
"Sata kruunua. Minä luulen, että olette hullu. Sillä te olisitte voinut ostaa joululahjoja puolelle kaupunkia. Ja te, joka sitäpaitsi olette antanut minulle koko laulun."
Ja ennenkuin maaherratar ehti äännähtääkään, lensi laukku matkakirstuun ja hän itse oli Petran sylissä kuin pihtien puristamana.
"Te olette kovin väkevä", oli ainoa, minkä hän sanoi, kun taas pääsi irti. Ja sitten hän hymyili.
Maaherratar oli oppinut hymyilemään aivan kuin muutkin ihmiset. Aluksi se ei ikäänkuin sopinut hänen kasvoihinsa, mutta nyt se jo oli siinä aivan kuin paikoillaan.
Niin sanoi myöskin vanha tohtori Sonderberg, kun hän tuli tavalliselle viikkokäynnilleen enemmän ystävänä kuin lääkärinä.
"Te olette opettanut herrattaren hymyilemään, te pieni", – sanoi tohtori Sonderberg.
Mutta Petra nauroi ja sanoi, että asianlaita varmaankin vain oli niin, että herratar oli unohtanut sen joksikin aikaa, kun hänellä oli ollut niin paljon ikävyyksiä ja suruja. Sillä kaikki ihmiset ovat toki syntyneet sellaisiksi, että osaavat hymyillä.
Maaherratar hymyili. Ja hän hymyili vielä enemmän, kun Petra veti esiin koruompelun ja sanoi:
"Ja tässä on minun joululahjani teille. Se on salvetti. Mutta tuo yksi kulma minun pitää ommella, kun tulen takaisin. Minulta koruompelu sujuu niin hitaasti, sillä se on minusta niin tympäisevää työtä."
"Kiitos. Minä luulen melkein, että pidän salvetin tällaisenaan, Petra, Näin se on aivan te itse", hymyili herratar. "Te olette hyvin kiltti. Se on varmaankin tuottanut teille monta ikävää hetkeä."
"Niin on", hymyili Petra avomielisesti. "Minun lahjojeni omituisuutena on muuten, etteivät ne koskaan ole valmiita, sillä kotona minä aina keksin niin paljon tekemistä ulkosalla. Tässä on se lahja, jonka olen aikonut Veyerille. Minun olisi pitänyt koristaa se koruompelulla – se on tarkoitettu liiveiksi. Mutta eikö se teistäkin ole yhtä sievä ilman koruompelua? Tai melkein ainakin?"
"Aivan yhtä sievä", vastasi Letta rouva vallan vakavana. "Vaikka luulenkin, että Vilhelm olisi pannut erikoista arvoa pariin teidän kätenne tekemään neulanpistokseen, niin hän kyllä tulee iloiseksi nähdessään, että te yleensä olette ajatellut häntä. Hänelle varmaankin on oleva suuri pettymys, ettette ole kotona hänen palatessaan."
"Kyllä varmaankin", sanoi Petra hyvillään.
"Ettekö voisi odottaa huomiseen, niin voisimme joka tapauksessa syödä aikaisen päivällisen yhdessä hänen kanssaan."
"Minä olen luvannut ylioppilas Bortingille matkustaa yhdessä hänen kanssaan. Hänen vanhempansahan asuvat nyt meidän pappilassamme. Ja minun veljeni matkustaa samalla kertaa."
Maaherratar kävi vakavaksi. Hän istuutui.
"Kuulkaa nyt, pikku Petra", sanoi hän. "Te tiedätte, että minä sanon suoraan, jos jokin ei minua miellytä. Enkä minä pidä siitä tavasta, millä te kohtelette molempia näitä nuoria ihmisiä. Tehän olette vain lapsi ettekä ajattele kihlausta ettekä muuta sellaista, –mutta on sääli näitä kahta poikaa, paitsi sitä ettei se myöskään ole aivan sopivaa. Toisen kanssa olette niin innokkaassa kirjevaihdossa, että – että – en tiedä mitä – ja –"
"Minusta on niin hienoa saada kirjeitä ulkomailta", puolustautui Petra.
"Sitäpaitsi se on hauskaa."
"Ja toisen kanssa te käytte konserteissa ja kävelette, ja nyt te matkustatte kotiinkin hänen kanssaan", jatkoi maaherratar tyytymättömänä. "Minun mielestäni se ei ole oikein. Minun mielestäni teidän pitäisi antaa toisen heistä tietää, että te pidätte enemmän toisesta."
"Niin, mutta se olisi toisesta ikävää. Enkä minä tiedä, kuka on toinen", sanoi Petra. "Minä pidän hirveän paljon kumpaisestakin –aina sen mukaan, kenen kanssa olen yhdessä."
"Niin niin. Olkaa vain varovainen, lapsi kulta", sanoi maaherratar leppyneenä ja nousi paikaltaan. "Joka tapauksessa sekä Vilhelm että minä kaipaamme teitä kovasti jouluna."
"Sepä hauskaa", vastasi Petra ilosta säteillen.
* * * * *
Lunta tuprutteli yhä tiheämmin ja tiheämmin. Oli mahdotonta eroittaa muuta vaununikkunasta kuin yksityisiä pieniä kaukaisia valoja, jotka välähtivät esiin pimeästä ja katosivat yhtä nopeasti. Kattolamppu loisti himmeästi ja leimuellen, juna jyskytteli eteenpäin. Mutta kolme matkustajaamme ei huomannut mitään siitä, mikä heidän ympärillään tapahtui. Pappila ulkopuoleltaan ja pappila sisäpuoleltaan oli kaiken aikaa puheenaiheena, niin että Per Borting vihdoin sanoi, että siellä hän nyt osaisi liikkua vaikka sokeana.
Hämäristä saakka oli ilma selkeää, ja kun juna iltamyöhällä pysähtyi vanhalle tutulle asemalle, hohtivat tähdet niin kirkkaina, että köyhät vilkkuvat rautatielyhdyt näyttivät vieläkin kurjemmilta ja nolommilta kuin muuten.
"Tunturihiiri", kaikui Finnin ääni, "onko sinulla uudet suksirakset minulle, niinkuin lupasit?" Ja rakennuksen takana ravistelivat hevoset kulkusiaan, joiden ääni kaikui kumahdellen ja sitten äkkiä lakkasi kuulumasta.
"Se on Musta. Se nykii aina noin, kun se on sidottuna", sanoi Petra ja kiiti läpi lumen kahden ison reen luo.
Toisessa istui turkkeihin kääritty herra.
"Isä."
Petra lensi hänen kaulaansa, puristeli häntä ja suuteli häntä ainoaan paljaaseen paikkaan – nenälle.
"No. Ei ole vaikeaa arvata, kuka te olette, sydämellinen nuori neiti", sanoi naurunsekainen ääni turkin sisästä. "On luonnollista,
että erehdyitte, sillä ovathan sekä hevonen että reki vanhoja tuttujanne."
"Ai! Anteeksi", sanoi Petra ällistyneenä. Mutta sitten hän nauroi ja käänsi taas päänsä turkkia kohti.
"Sitäpaitsi minä olen tottunut suutelemaan tämän pitäjän pappia."
"Toivon, ettette luovu siitä tavasta, pikku ystävä. Per'in isänä toki minäkin voinen vaatia vähän ystävyyttä. Toivomme, että saamme ilon nähdä teidät kaikki meillä ensimmäisenä joulupäivänä. – No, hyvää päivää ja tervetuloa, poikani."
"Tunturihiiri! Täällä on Musta. Jouduhan vähän", vinkui Finn. "Herman jo istuu reessä."
Tunturihiiri tuli hyppien. Per Borting seurasi perässä ja kääri hänet vällyihin.
Pari minuuttia myöhemmin reet lähtivät nytkähtäen liikkeeseen. Kulkuset soivat tahdikkaasti ja kaviot kapsahtelivat. Ensi tienhaarassa erottiin.
"Hyvästi ja tervetuloa."
"Huomenna", huusi Per.
Petra istui kasvot ylöspäin käännettyinä, onnellisena ja turvallisena äitinsä vanhassa tutussa turkissa.
"Tähdet loistavat aivan toisella tavalla täällä kuin kaupungissa", sanoi hän.
Joen rannalla olevan pienen valkean rakennuksen kaikki ikkunat olivat valaistut. Portailla seisoi Maren, kädet kudotun kaulahuivin alla. Hän oli vielä harmaampi ja ryppyisempi ja vinompi kuin miksi Petra hänet muisti. Hän käänsi Petran kasvot käytävän lamppua kohti.
"Enkö minä jo arvannut. Paljon laihempi eikä yhtä ruskea. Niin, se kaupunki!" sanoi hän harmissaan.
Pappi tunnusteli hapuillen Petraa toisella kädellä ja piti toisella Hermanista kiinni.
"Lapsi kultani. Rakas pikku Tunturihiiri", sanoi hän. Hän vapisi, eivätkä silmät enää nähneet mitään. Hän oli paljon vanhentunut.
"Voi isä, miten sinä olet käynyt – kauniiksi", sanoi Petra, mutta hänen äänensä ei ollut niin varma, että hän olisi uskaltanut sanoa sen enempää hetken aikaan.
Hän juoksi läpi talon – kaikki oli tarkastettava.
"Huonekalut ovat muuttuneet niin isoiksi minun mielestäni", sanoi hän Marenille, joka uskollisesti seurasi häntä eikä voinut katsella häntä kyllikseen.
"Niin, näetkös, pappilan suojathan olivat avarammat. Mutta näitä voikin paljon paremmin pitää puhtaina", sanoi Maren.
Petra seisoi pienen ullakkohuoneensa ikkunan luona ja riisuutui. Puiden latvojen takaa hän näki pappilan valot. Hän seisoi kauan katsellen niitä.
"Paikkoja ikävöi melkein enemmän kuin ihmisiä", huokasi hän ja veti uutimen alas.
Hän heräsi siihen, että Maren laski halkosylyksen jymisten lattialle ja alkoi panna puita uuniin.
"Niin hyvin kai sinä et ole nukkunut, sitten kun menit kaupunkiin?" kysyi hän.
"Minä nukun kaikkialla, Maren", sanoi Petra.
"Hoh, niin sinä varmaankin vain luulet", sanoi Maren, jonka epäluulo kaikkea vastaan, mikä kantoi kaupungin nimeä, oli syvälle juurtunut.
Petra makasi katsellen tulen leimuelevaa heijastusta seinällä. Uusi öljytty seinä. Ikävä seinä. Hän muisti vanhoja harmaita seinäpapereita, joille keltaiset sadetahrat muodostivat kuvioita, kukon ja miehen, joka ratsasti katon harjalla. Ja reikää, jonka läpi raontäyte näkyi. Petra muisti varsin hyvin, että hän itse oli tehnyt sen nähdäkseen, mitä paperin alla oli – kauan, kauan sitten.
Ja äkkiä pieni ruskea pää pujahti peitteen alle. Peite hytkähteli ja sen alta kuului tukahdutettua nyyhkytystä.
* * * * *
"Ei, minä en tahdo sinua mukaan, Finn."
Petra seisoi sovitellen suksia jalkaansa heti päivällisen jälkeen.
"Se on hävytöntä. Rakset ovat jo suksissa. Katso vain", sanoi Finn innokkaasti.
"Sinä voit mennä Hermanin kanssa. Minä tahdon mennä yksin", sanoi Petra ja hiihti tiehensä.
"Petra on muuttunut niin tyttömäiseksi", sanoi Finn suutuksissaan Hermanille. "Nyt hän tahtoo mennä yksin kuin täysikasvuinen ainakin."
Kuivaa lunta putoili hiljalleen. Se rapisi puissa ja pisteli Petraa kasvoihin, kun hän hiihti jokea pitkin. Hän meni maantien poikki, niityn yli ja pappilaan kuuluvan kivikon läpi.
Ison koivun luona hän pysähtyi, taputti runkoa ja nyökäytti päätään katsellen ylös kuiviin riippuviin oksiin. Sitten hän hiihti eteenpäin lampea kohti. Sukset liukuivat kahisten runkojen lomitse. Oli niin hiljaista, että hän kuuli, miten lumi putoili.
Lampi oli lumen peitossa, ainoastaan hiukan maata alempana. Kuivia kaisloja ja korsia pisti esille sen ympärillä.
Petra meni vähän syrjään ja jäi katselemaan pappilaa, joka kohosi siinä valkeana ja rauhallisena molempine ikkunariveineen ja näytti ikäänkuin kuuluvan yhteen puhtaan uuden lumen kanssa. Hän seisoi hetkisen ja katseli.
Sitten hän palasi takaisin, kiersi lammen ympäri ja pysähtyi taas tuijottamaan kauas entisiin aikoihin. Ja lumi putoili hiljalleen ja tiheään ja kasaantui valkeaksi huipuksi hänen lakkiinsa ja pieniksi valkeiksi tyynyiksi hänen olkapäilleen.
Suhahti läpi lehdon.
Notkea olento ilman lakkia, yllään ainoastaan tavalliset sisävaatteet, hiihti alaspäin liian pienillä suksilla. Hän kiepahti Petran
rinnalle.
"Minä ymmärsin, ette te aioitte mennä lammelle. Minä näin teidän hiihtävän koivikkoon ja silloin sieppasin renkipojan sukset ja livahdin perästä. No, mitä te pidätte uudesta kodistanne? Minä olen mielestäni melkein kuin rikoksentekijä, kun asun pappilassa."
"Se ei ole mikään koti. Nyt vielä", sanoi Petra. "Sellaiset tornihuvilat leikeltyine kuistikkoineen ovat ruminta, mitä tiedän. Ne näyttävät yhtä uusilta aina siihen hetkeen saakka, jolloin ne hajoavat. Niillä ei ikäänkuin ole mitään kasvoja. Vain siksi, että minun ihmiseni asuvat siellä, on tämä talo toisenlainen kuin muut rumat talot. Ovat sellaiset asiat kuitenkin omituisia. Siksi vain, että te nyt asutte pappilassa, olette te jotain muuta kuin tavallinen ystävä."
Per Borting kävi hehkuvan punaiseksi.
"Jotain muuta kuin tavallinen ystävä? Mitäsitten, Petra?"
"Mitä? Niin, sitä minä en tiedä", vastasi Petra ikäänkuin hyvin kaukaa. "Tiedättekö", lisäsi hän äkkiä vilkkaasti. "Herratar sanoi eilen, että on väärin, että minä pidän sekä teistä että Vilhelm Veyeristä. Mutta nyt minä hyvin tiedän, että te olette ainoa, jonka soisin asuvan täällä – jonka kanssa tahtoisin pitää kaikkea tätä yhteisenä."
"Petra", sanoi hän vain.
Hänen silmänsä syttyivät. Hän olisi niin kernaasti sulkenut Petran syliinsä, mutta hän ei uskaltanut säikäyttää häntä.
Petra sai tulla itsestään, kun hänen hetkensä oli tullut. Per Borting tiesi nyt voittaneensa niin paljon kuin oli voitettavissa. Vastaiseksi.
*** END OF THE PROJECT GUTENBERG EBOOK PIKKU PETRA ***
Updated editions will replace the previous one—the old editions will be renamed.
Creating the works from print editions not protected by U.S. copyright law means that no one owns a United States copyright in these works, so the Foundation (and you!) can copy and distribute it in the United States without permission and without paying copyright royalties. Special rules, set forth in the General Terms of Use part of this license, apply to copying and distributing Project Gutenberg™ electronic works to protect the PROJECT GUTENBERG™ concept and trademark. Project Gutenberg is a registered trademark, and may not be used if you charge for an eBook, except by following the terms of the trademark license, including paying royalties for use of the Project Gutenberg trademark. If you do not charge anything for copies of this eBook, complying with the trademark license is very easy. You may use this eBook for nearly any purpose such as creation of derivative works, reports, performances and research. Project Gutenberg eBooks may be modified and printed and given away—you may do practically ANYTHING in the United States with eBooks not protected by U.S. copyright law. Redistribution is subject to the trademark license, especially commercial redistribution.