9A1. An Overview of the Donor Area: Basic Principles Jean Devroye
11B. Hair Survival of Partial Follicles: Implications for Pluripotent Stem Cells
and Melanocyte Reservoir Jung-Chul Kim and Yung-Chul Choi 11C. Methods of Studying Hair Survival and Growth
11C1. A Technique for Counting Hairs in a Research Study Box Michael L Beehner
11C2. The Cross-Section Trichometer Bernard H Cohen
11C3. Enhanced Patient Selection: A New Method for Measuring Hair Growth Parameters—
The TrichoScan® Jerzy R Kolasinski 11C4. Commentary Walter P Unger
12. The Recipient Site
12A1. The History of Follicular Unit Transplantation Bobby L Limmer
12A2. Commentary Walter P Unger
12B1. The Pitfalls of FUT Incisions and How to Avoid Them Jennifer H Martinick
12B2. Commentary Walter P Unger
12C1. High-Density Follicular Unit Transplant Thomas C Nakatsui
12C2. Commentary Walter P Unger
12D1. Mega Session Follicular Unit Transplantation Jerry Wong
12D2. Commentary Walter P Unger
12E. Recombinant Follicular Unit Transplantation James A Harris
12F1. The Hairline Zone and “Egg” Walter P Unger
12F2.
12G. The Anterior Temporal Area
12H. Transplanting the Vertex Jerry Wong
12I1. Combination Grafting Indications and Techniques Vance W
12I2.
12I3.
13A.
13B1. “Stick-And-Place”
DVD CONTENTS
The title of each video is listed below; each video can be accessed by clicking on the appropriate title in the list on the opening page of the accompanying DVD. For ease of cross-reference, the appropriate text chapter section number is provided.
6A.1. Making Recipient Site Incisions
Walter P Unger
6A.2. Outlining the Recipient Area
Walter P Unger
8B. Tumescent Anesthesia in Hair Transplantation
John DN Gillespie
8C. Supraorbital Nerve Blocks in Hair Restoration Surgery
Cam Simmons
9A3.1. Donor Strip Excision in Previously Harvested Area
Walter P Unger
9A3.2. The Collapse Closure
Thomas Kohn and Shadi Zari
9B. Evaluation of Scalp Elasticity
Melvin L Mayer
9D1.1. Managing Wide Donor Strips
Jerry Wong
9D1.2. Scalp Laxity Exercises
Jerry Wong
9E1. Skin Hook Technique
Damkerng Pathomvanich
9E2. The Haber Spreader
Robert S Haber
9F1. Trichophytic Closure Overview
Paul T Rose
9F2. Trichophytic Closure in Asians
Dae-Young Kim
9G1. Conventional FUE
James A Harris
9G2.1. Beard-to-Scalp FUE: Manual Needle
Jack M Yu and Amy Y Yu
9G2.2. Beard-to-Scalp FUE: Motorized Punch
James A Harris
10C. Preparing Grafts for High FU Density FUT
Thomas C Nakatsui
10D. Preparation of Multi-Unit Grafts
Vance W Elliott
12B. Pearls of FUT Incising
Jennifer H Martinick
12D. Mega Session FUT
Jerry Wong
12F1. The Hairline Zone
Walter P Unger
12I. Combination Grafting Indications and Techniques
Marc R Avram MD Department of Dermatology, Weill Cornell Medical College/New York Presbyterian Hospital, and Private practice, New York, New York, USA
Marco N Barusco MD Founder and Medical Director, Tempus Hair Restoration, PA, Port Orange, Florida, USA
Alan Bauman MD, ABHRS Founder and Medical Director, Bauman Medical Group, Boca Raton, Florida, USA
Michael L Beehner MD Saratoga Hair, Saratoga Springs, New York, USA
Guillermo Blugerman MD Private practice, Buenos Aires, Argentina
MJ Kristine S Bunagan MD Department of Dermatology, Davao Medical Center, Davao City, Philippines, and Hair Transplant Clinic Philippines, Davao City, Philippines
Glenn M Charles MD Charles Hair, Boca Raton, Florida, USA
Hangrae Cho MD PhD Ohkims Dermatologic Clinic, Seoul, Korea
Yung-Chul Choi MD Togo Clinic, Seoul, Korea
Bernard H Cohen MD Voluntary Professor, Department of Dermatology and Cutaneous Surgery, University of Miami, Miller School of Medicine, Miami, and The Hair Loss Center, Coral Gables, Florida, USA
John Cole MD For Hair, Alpharetta, Georgia, USA
Jerry E Cooley MD Carolina Dermatology Hair Center, Charlotte, North Carolina, USA
Paul C Cotterill MD Private practice, Toronto, Ontario, Canada
Jean Devroye MD Hair Transplant Surgery, Brussels, Belgium
Eric L Eisenberg MD, FRCP(C) Division of Dermatology, Toronto Western Hospital, University Health Network, University of Toronto, Ontario, and Private practice, Mississauga, Ontario, Canada
Vance W Elliott MD CCFP ABHRS Private practice, Sherwood Park, Alberta, Canada
Jeffrey Epstein MD Director, Foundation for Hair Restoration, and Division of Facial Plastic Surgery, Department of Otolaryngology, University of Miami, Miami, Florida, USA
Bessam Farjo MD Farjo Medical Center, Manchester, UK
Nilofer Farjo MD Farjo Medical Center, Manchester, UK
Patrick Frechet MD Private practice, Geneva, Switzerland
John DN Gillespie MD Gillespie Clinic, Calgary, Alberta, Canada
Alex Ginzburg MD Hair Transplant, Raanana, Israel
Robert S Haber MD Clinical Associate Professor, Dermatology & Pediatrics, Case Western Reserve University School of Medicine, Cleveland, Ohio, USA, and Haber Dermatology and Cosmetic Surgery, South Euclid, Ohio, USA
James A Harris MD FACS Clinical Instructor, Department of Otolaryngology/Head and Neck Surgery, University of Colorado School of Medicine, Aurora, Colorado and the Hair Sciences Center of Colorado, Greenwood Village, Colorado, USA
Jen-Chih Hsieh PhD Aderans Research Institute, Philadelphia, Pennsylvania, USA
Sungjoo Tommy Hwang MD PhD Private practice, Director, Dr. Hwang’s Hair-Hair Clinic, Seoul, Korea
Francisco Jimenez MD Private practice, Las Palmas de Gran Canaria, Canary Islands, Spain
Sheldon S Kabaker MD FACS Clinical Professor, University of California at San Francisco, Oakland, California, USA
Emina Karamanovski MD Lam Institute for Hair Restoration, Plano, Texas, USA
Sharon A Keene MD Chief Medical Officer, HairDx and solo practitioner Physician's Hair Institute, Tucson, Arizona, USA
Dae-Young Kim MD Yonsei Hair Clinic, Seoul, Korea
Jung-Chul Kim MD PhD Professor, Hair Transplantation and Research Center, Kyungpook National University Hospital, Daegu, Korea
Moon-Kyu Kim MD PhD Hair Transplantation and Research Center, Kyungpook National University Hospital, Daegu, Korea
Russell G Knudsen MB BS Private practice, Sydney, New South Wales, Australia
Thomas Kohn MD Assistant Professor, Division of Dermatology, McGill University, Montreal, Canada
Yuha Y Kojima MD PhoenixBio Ltd, Hiroshima, Japan
Jerzy R Kolasinski MD Klinika Kolasinski–Hair Clinic Poznan, Swarzedz, Poland
Walter Krugluger MD Moser Medical Group, Vienna, Austria
Melike Kulahci MD Transmed Hair and Cosmetic Surgery Clinic, Istanbul, Turkey
Tina Lardner MD Hair Sciences Center of Colorado, Greenwood Village, Colorado, USA
Matt L Leavitt DO Medical Director, Medical Hair Restoration, Maitland, Florida, USA
Young-Ran Lee MD PhD ABHRS Anamo Hair Transplant Center, Seoul, Korea
Bobby L Limmer MD Private practice, San Antonio, Texas, USA
E Antonio Mangubat MD Southcenter Cosmetic Surgery, Seattle, Washington, USA
Jennifer H Martinick MBBS FACCS New Hair Clinic, Perth, Western Australia, Australia
Mario Marzola MB BS Norwood Day Surgery, Adelaide, Australia
Melvin L Mayer MD Bosley Medical, San Diego, California, USA
Paul J McAndrews MD Department of Dermatology, University of Southern California School of Medicine and Private practice, Beverly Hills and Pasadena, California, USA
Michael Morgan MD University of South Florida, Tampa, Florida, USA
Masahisa Nagai MD Nagai Clinic, Fukuoka-Shi, Japan
Thomas C Nakatsui MD Division of Dermatology, University of Alberta, Edmonton, Alberta, Canada
Mike Neff Private practice, New Port Richey, Florida, USA
Jonathan L Nelson MD Family Dermatology Center, Rye, New York, USA
Rolf EA Nordström MD PhD Professor, Plastic and Reconstructive Surgery, and Chief of Nordström Hospital, Helsinki, Finland
Aron G Nusbaum MD University of Miami Miller School of Medicine, Miami, Florida, USA
Bernard P Nusbaum MD Voluntary Faculty, Department of Dermatology and Cutoneous Surgery, University of Miami Miller School of Medicine, Miami, Florida, and Miami Hair, Coral Gables, Florida, USA
Nina Otberg MD Department of Dermatology and Skin Science, University of British Columbia, Vancouver, British Columbia, Canada, and Hair Clinic, Skin and Laser Center, Potsdam, Germany
William M Parsley MD Private practice, Louisville, Kentucky, USA
Nishit S Patel MS University of South Florida, Tampa, Florida, USA
Akio Sato MD Tokyo Memorial Clinic Hirayama, Tokyo, Japan
Diego Schavelzon MD Buenos Aires, Argentina
†Gerard E Seery MD FRCS
Jerry Shapiro MD FRCPC Department of Dermatology and Skin Science, University of British Columbia, Vancouver, British Columbia, Canada
Ronald Shapiro MD Medical Director, Shapiro Medical Group, Bloomington, Minnesota, USA
Richard C Shiell MB BS Retired Hair Transplant Surgeon, and Private practice, Sandringham, Victoria, Australia
Cam Simmons MD ABHRS Canadian Hair Transplant Centre, Toronto, Ontario, Canada
†Deceased.
Dow Stough MD The Stough Hair Clinic, Hot Springs, Arizona, USA
J Kevin Thompson PhD Department of Psychology, University of South Florida, Tampa, Florida, USA
Koh-ei Toyoshima MD PhoenixBio Ltd, Hiroshima, Japan
Arthur Tykocinski MD Private practice, São Paulo, Brazil
Mark Unger MD Private practice, Toronto, Ontario, Canada
Robin Unger MD Associate Professor (Dermatology), Mount Sinai Medical School, New York, New York, USA
Walter P Unger MD FRCP(C) FACP Clinical Professor of Dermatology, Co-Director of the Division of Cosmetic Dermatologic Surgery, Mount Sinai Medical School, New York, New York, USA, and Private practices in New York, New York, and Toronto, Canada
James E Vogel MD FACS Associate Professor of Surgery, Department of Surgery, Division of Plastic Surgery, Johns Hopkins Hospital and School of Medicine, Baltimore, Maryland, USA
Jeffrey E Vogel MD Candidate, Emory University School of Medicine, Atlanta, Georgia, USA
Mark A Waldman MD Private practice, Louisville, Kentucky, USA
Ken Washenik MD PhD Bosley, Aderans Research Institute, Beverly Hills, California, USA
Carlos K Wesley MD Private practice, New York, New York, USA
Bradley R Wolf MD Private practice, Cincinnati, Ohio, USA
Jerry Wong MD Hasson & Wong Aesthetic Surgery, Vancouver, British Columbia, Canada
Kuniyoshi Yagyu MD Kioicho Clinic, Tokyo, Japan
Amy Y Yu MD Huai’An, Jiang Su, China
Jack M Yu MD Huai’An, Jiang Su, China
Kenneth CY Yu MD Facial Plastic Surgery Fellow, Aesthetic Facial Plastic Surgery Medical Clinic, Oakland, California, USA
Ying Zheng PhD Aderans Research Institute, Philadelphia, Pennsylvania, USA
1
BASIC SCIENCE
1A. HAIR ANATOMY AND HISTOLOGY
Paul T Rose, Ronald Shapiro, Michael Morgan, and Nishit S Patel
EMBRYOLOGY
Hair follicles are derived from an interaction between the embryological ectoderm and the mesoderm beginning around the second month of pregnancy (1,2). The earliest event is the development of regularly spaced epidermal thickenings, called placodes, initiated by molecular signaling from the underlying mesenchyme (3). In response, the overlying epidermis sends molecular signals back inducing thickening of the mesenchyme. This complex epithelial–mesenchymal metabolic signaling and crosstalk occurs throughout follicular morphogenesis. The major embryological stages are described below.
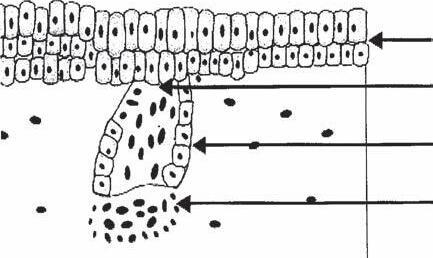
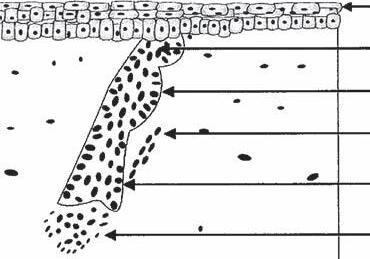
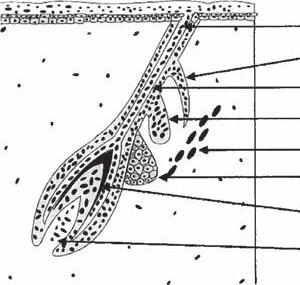
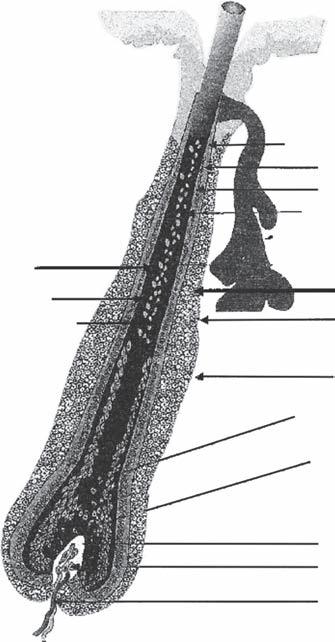
Pregerm Stage (Fig. 1A-1a)
The first signs of a hair follicle are an aggregation of mesenchymal cells in the superficial level of the dermis and a simultaneous thickening of basal epidermal cells immediately above it (4,5).
Germ Stage (Fig. 1A-1b)
At this point, the basal epidermal cells become elongated and start to bulge downward as the “hair germ,” while at the same time the underlying mesenchymal cells begin to replicate to form the rudiment of what will become the dermal papilla.
Hair-Peg Stage (Fig. 1A-1c)
During this stage, the epithelial cells of the hair germ grow downward and form a column or “hair peg” that seems to propel the mesenchymal aggregate downward.
Bulbous-Peg Stage (Fig. 1A-1d)
Subsequently, three areas of swelling appear along the hair peg as the column extends downward. The most superior swelling will become the apocrine gland. The middle swelling develops into the sebaceous gland, while the lowest becomes the bulge area to which the arrector pili (AP) muscle will attach. The AP muscle forms near the developing sebaceous gland, but its development is independent of the hair follicle. During this stage, the epidermal cells at the advancing base of this column surround a portion of the underlying mesodermal cells, forming the dermal papilla (4,5).
The First Primordial Hair (Fig. 1A-1e)
At this point the cells at the base of the column that surround the dermal papilla begin to actively proliferate, forming the early matrix, and this initial hair shaft moves upward. Above
this, the central cells of the follicular peg appear to degenerate, and the emerging hair seems to push out the plug so a hair canal can be formed.
Hair development begins at about 9 weeks and by 22 weeks the entire initial population of follicles is completed, including those on the scalp. About five million hair follicles cover the human body at birth with approximately 100,000 being scalp hairs. It is generally believed that no “new” hair follicles develop after birth.
On the scalp, fine “lanugo” hairs present at birth are shed in a more or less synchronized wave pattern by the third or fourth month after birth (6). This synchronization is lost with the successive population of hairs and, by the end of the first year of life, an asynchronous population of follicles in all stages of the growth cycle exists (7,8).
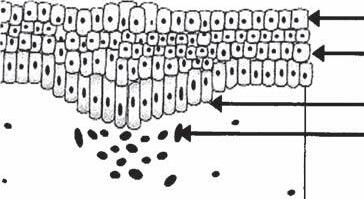
GROSS ANATOMY OF THE TERMINAL HAIR
The hair follicle is anatomically divided into an upper, middle, and lower region. These three regions are respectively termed the infundibulum, the isthmus, and the inferior segment (Fig. 1A-2). The infundibulum and isthmus comprise the permanent portions of the hair follicle while the inferior segment is transient and undergoes cyclical regeneration (Box 1A-1).
Infundibulum
The upper region, or infundibulum, of the follicle consists of the area from the opening of the sebaceous duct to the follicular opening on the surface of the skin. It contains the fully formed hair shaft as it exits the epidermis. The epithelium of the epidermis is contiguous with the wall of the infundibulum.
Isthmus
The middle region, or isthmus, begins at the opening of the sebaceous duct and continues down until it reaches the insertion of the AP muscle. At the level of the insertion of the AP muscle is the bulge area which is very obvious in the embryonic follicle, but only intermittently obvious in the adult follicle. The bulge, considered a part of the outer root sheath (ORS), contains follicular stem cells that are critical for the regeneration of the follicular unit (FU) (Box 1A-2) (9). Recent immunohistochemical studies using stem cell markers such as CD34 and keratin15 have suggested that follicular stem cells may be more widely distributed along the ORS (9,10). The stem cells of the bulge area may, in addition, be recruited to generate epidermal and sebaceous gland cells (11). Interestingly, the bulge area appears to be inundated with nerve endings, as well as Merkel cells. Such a network may be involved in providing signals for hair growth and cycling.
Periderm
Intermediate layer
Basal layer
Dermal layer
Dermal papilla Hair peg
Periderm
Intermediate layer
Hair germ
Dermal papilla
Periderm
Hair canal
Sebaceous gland
Arrector muscle
Bulge
Dermal papilla
e)
1A-1
Hair canal
Apocrine rudiment
Inner root sheath
Sebaceous gland
Arrector muscle
Bulge
Hair
Dermal papilla
of embryology: (a) pregerm stage, (b) germ stage, (c) hair-peg stage, (d) bulbous-peg stage, and (e) first primordial hair.
Inferior Segment
This inferior segment of the hair follicle is the area from the base of the follicle to the insertion of the AP muscle. This area is transient and cyclically regenerates itself in a controlled fashion.
The bottom-most portion of the inferior segment is referred to as the hair bulb and contains the rapidly dividing undifferentiated matrix cells and melanocytes which are of epithelial origin. The hair bulb surrounds the dermal papilla. The combination of the hair bulb and the dermal papilla, with its associated nerves and blood vessels, constitutes the hair root.
HISTOLOGY OF THE TERMINAL HAIR FOLLICLE
The dermal papilla and the surrounding hair matrix are located at the bottom of the hair follicle. Above this, the
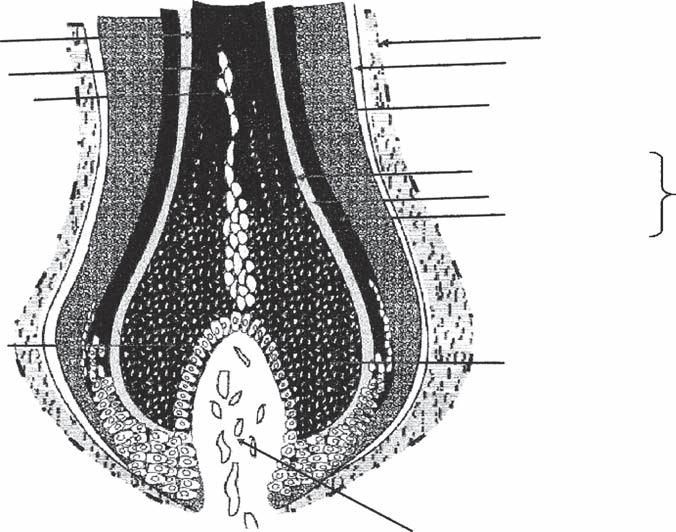
hair follicle consists (from inside out) of the following concentric layers: the hair shaft, the inner root sheath (IRS), the ORS, and the perifollicular dermal sheath. The histology of these components will be discussed in more detail below (Figs. 1A-3–1A-6).
Dermal Papilla
As the name suggests, the dermal papilla is of mesodermal origin. The papilla is surrounded by the matrix cells of the hair bulb. At the distal aspect of the hair bulb, the papilla connects with a fibrous dermal sheath that surrounds the hair follicle.
The dermal papilla contains specialized fibroblasts with powerful hair follicle inductive properties. The dermal papilla is needed for initiation of a new hair cycle and for normal hair growth (12). It is believed to regulate matrix cell division and the caliber of the hair produced (13). Androgen receptors
Figure
Stages
Inferior region
Box 1A-1 Anatomical Division of Hair Follicle
Upper region (infundibulum)—from the opening of the sebaceous duct to the follicular opening on the surface of the skin (permanent)
Middle region (isthmus)—from the opening of the sebaceous duct to the insertion of the arrector pili muscle (permanent)
Lower region (inferior segment)—from the base of the follicle to the insertion of the arrector pili muscle (cyclical)
Box 1A-2 Proposed Location of Stem Cells
Outer root sheath
have been clearly demonstrated in the dermal papilla (14). The papilla has been associated with the production of a large number of growth factors and growth factor receptors, which may play a role in hair growth (15).
Matrix Cells
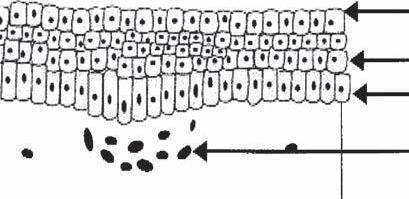
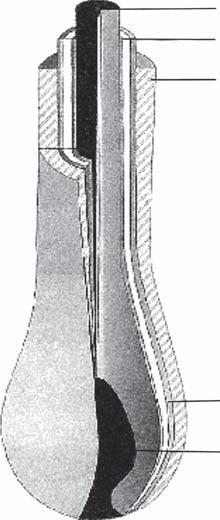
The matrix cells are located directly above the papilla. These cells are undifferentiated, actively dividing, and immunologically “privileged” (15). Melanocytes are present between the basal cells of the matrix. Melanin produced by the melanocytes is incorporated into new hairs by phagocytosis. The matrix cells differentiate into the multiple components of the hair follicle, including the hair shaft, the IRS, and the ORS. These three structures can be visualized as three concentric cylinders embedded within each other (Fig. 1A-6).
Hair Shaft
The hair shaft is the only part of the hair follicle to exit the epidermis. It in itself is also made up of three layers: (i)the medulla, (ii)the cortex, and (iii)the hair shaft cuticle.
1. Hair shaft medulla: The medulla is the most central layer of the hair shaft and probably arises from matrix cells directly above the papilla. In the suprabulbar region, the cells of the medulla begin to show vesicles in their cytoplasm. At the level of the epidermis, the cells of the medulla appear to dehydrate, and the vacuoles become air filled. Terminal hairs contain a medulla but vellus hairs and finer hairs do not.
2. Hair shaft cortex: Lateral to the medulla are the cortical cells of the hair shaft, which are fusiform in shape and arranged longitudinally parallel to the shaft. They become tightly packed and keratinized as they move upward. It is the cortex cells that impart the mechanical properties of hair. Pigment is present in some cortical cells. This pigment was initially acquired from melanosomes in the matrix.
3. Hair shaft cuticle: The cuticle is the outermost layer of the hair shaft and is composed of 6 to 10 layers of overlapping cuticle cells. They are arranged in a shingled type of array and later become keratinized and point upward, so they can interlock with the downward projecting cuticle cells of the surrounding IRS.
Inner Root Sheath
The IRS surrounds the hair shaft. It exists only in the inferior segment of the hair follicle traveling from the bulb up to the beginning of the isthmus. Beyond the erector pili muscle, the keratinized IRS deteriorates and is replaced by keratin from ORS. The IRS is also made up of three layers: (i) the IRS cuticle, which is one cell thick; (ii) Huxley’s layer, which is three or four cells thick; and (iii) Henle’s layer, which is also one cell thick. As the cells in the IRS keratinize, they give rigidity to this sheath, supporting the IRS function as a mold for the enclosed and developing hair shaft. Racial differences in shaft cross-section shape and size are determined by the IRS. The IRS is tightly moored to the hair shaft by the overlapping of cells from their adjacent cuticle layers. The hair shaft and IRS move upward together, slipping along the surrounding innermost layer of the ORS.
Outer Root Sheath
The ORS surrounds the IRS and consists of multiple layers of epithelial cuboidal cells containing large quantities of glycogen. It travels from the bottom of the hair bulb to the opening of the sebaceous duct. It is thinnest at the level of the hair bulb and thickest at the isthmus. The ORS contains melanocytes, neurosecretory cells (Merkel cells), and Langerhans cells and is also a proposed site of follicular stem cells (16,17).
Dermal Sheath
Outside the ORS, a vitreous, periodic acid Schiff stain-positive, hyaline membrane layer is evident. Collagen bundles from the dermis surround this layer to form the perifollicular dermal sheath.
KERATINIZATION
Keratinization of the hair follicle occurs in an organized fashion that helps mold and shape the hair shaft. First the IRS
Bulge area
Matrix cells
Isthmus
Infundibulum
Figure 1A-2 Major regions of the hair follicle.
Cuticle of hair shaft
Cortex of hair shaft
Medulla of hair shaft
Hair bulb
Huxley’s layer of IRS
Henle’s layer of IRS
Cuticle layer of IRS
Inner root sheath
Outer root sheath (ORS)
Hyaline basement membrane
Bulge area
Keratogenous zone
Germinative cells of hair matrix
Dermal papilla
Papilla cells
Papilla core
Figure 1A-3 Diagram of hair follicle histology (longitudinal view).
Cuticle of hair
Cortex of hair
Medulla of hair
Melanocytes
Fibrous sheath
1A-4
Hyaline basement membrane
Outer root sheath (ORS)
Cuticle of IRS
Huxley’s layer of IRS
Henle’s layer of IRS
Inner root sheath
Undifferentiated matrix cells
Papilla
of bulb area histology (longitudinal view).
Figure
Diagram
Hair shaft
Inner root sheath
Outer root sheath
Connective tissue
keratinizes with tychohylaline granules starting with Henle layer and followed by the cuticle and Huxley layer. This creates a firm mold surrounding the cortex and medulla of the hair shaft. Next, the cells of the cortex cells degenerate and fill with keratin fibrils. The medulla is last to cornify.
INNERVATION AND BLOOD SUPPLY
Hair follicles are innervated by nerves that course from the bulb to the epidermis. Some hair follicles possess a “hair-end
organ.” This anatomical structure is made up of a “collar” of nerves that surrounds the hair follicle at the level of the sebaceous gland. Fibers may innervate local structures to form a plexus of nerves at a level between the papillary and reticular dermis (18). Hordinsky and Ericson have demonstrated that cells in and around the dermal papilla stain, with a panneuronal antibody, and that substance P can be detected at the bulge area, and in perifollicular nerves and vessels (19). Such observations provide more evidence that suggests there may be neural signals that influence hair physiology.
Hair follicles have an abundant blood supply. Vascularization is pronounced in the upper and lower portions of the follicle, with vessels penetrating the papilla.
THE FOLLICULAR UNIT
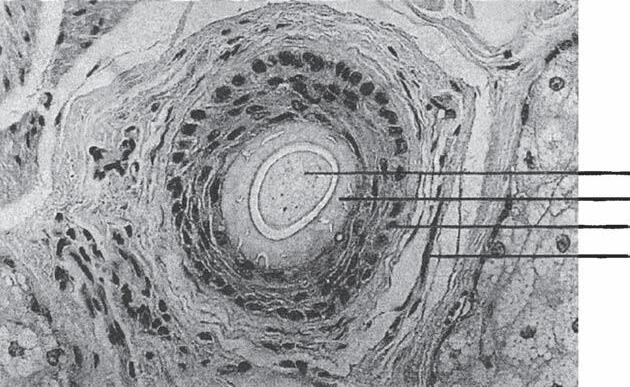
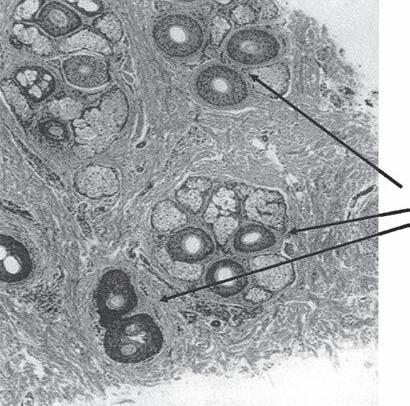
In 1984, Headington published a paper describing the transverse anatomy of hair follicles (20). He demonstrated that hairs at the level of the sebaceous duct exist in groupings, which he termed FUs. Headington defined an FU as consisting of two to four terminal follicles and one or more vellus hairs, as well as the associated sebaceous glands, insertion of the AP muscle, and the surrounding perifollicular dermal sheath (Fig. 1A-7a–c). These groupings are obvious from the surface to the level of the AP muscle insertion. Below this level the individual follicles tend to separate in the fatty layer making the groupings less obvious.
Recent three-dimensional reconstructions of the FU by Poblet et al. provide evidence and support the idea that a single AP muscle inserts into every follicle in an FU (21). Additionally, the AP muscle was found to be concave at its base, near the point of insertion. The concavity is believed to provide structural support to sebaceous lobules that fill the space, between the hair follicle and the AP muscle. There is also evidence for the role of the AP muscle as a tie that binds all the individual hair follicles in an FU together at the level of the isthmus.
FU density on the scalp is currently believed to average between 75 and 100 FU/cm2 although it may range higher or lower. An FU can contain from one to five terminal hairs but
Figure 1A-5 Cross-sectional photograph of bulb isthmus region.
Papilla
Hair bulb
Outer root sheath
Inner root sheath
Hair shaft
Figure 1A-6 The hair shaft, inner root sheath, and outer root sheath visualized as concentric cylinders embedded within each other.
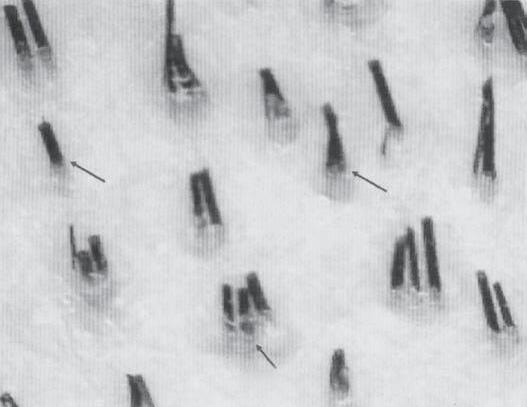
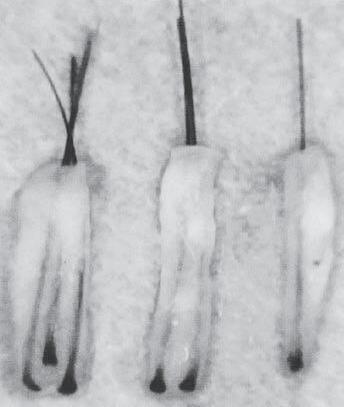
Figure 1A-7 Follicular unit (FU) groupings seen in: (a) a cross-sectional histological view through the scalp; (b) a close-up photograph of the surface of the scalp. The three-haired FU is the “power-house” of follicular unit transplantation. (c) One, two, and three-hair follicular unit grafts.
the majority (~80%) contains one to three terminal hairs. Variations in density exist at different locations on the scalp and in different ethnic groups.
VELLUS VS. TERMINAL HAIR
Hairs may be termed vellus or terminal depending on a number of characteristics including length, diameter, pigmentation, and location of the root.
Vellus Hairs
Vellus hairs are small, soft hairs with a diameter less than 0.03 mm and a length less than 1 cm. They have no pigment or medullary cavity and are often rooted superficially in the upper dermis. Depigmented hairs that are less than 0.03 mm and have been miniaturized by androgenic alopecia, or any
other cause, can be classified as “vellus-like hairs.” The term vellus hairs is often used to include both true vellus hairs and vellus-like hairs. At puberty, vellus hairs are replaced by terminal hairs in certain areas of the body (axilla, pubis, and face), and this change occurs by means of androgenic hormones.
Terminal Hairs
Terminal hairs are larger hairs with a diameter exceeding 0.06 mm and a length greater than 1 cm. They are often pigmented and medullated. They are rooted in the subcutaneous or deep dermis layer.
It is sometimes useful to differentiate intermediate hairs, between 0.03 and 0.06 mm in diameter, which may represent early miniaturization of terminal hairs, as seen in androgenic alopecia. The normal ratio of terminal to vellus hairs in a normal scalp is 7:1.
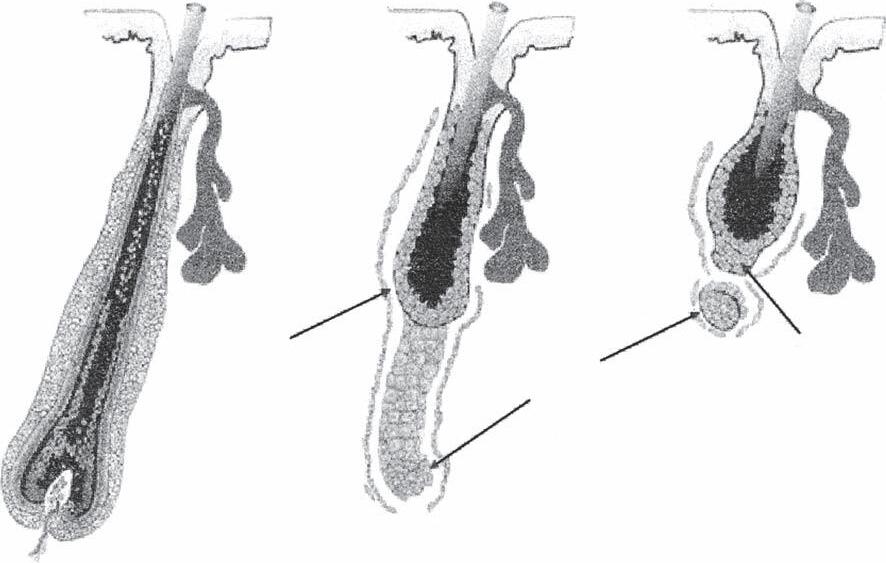
HAIR CYCLE
Each hair follicle goes through the hair cycle 10 to 20 times in a lifetime, every cycle recapitulating much of the embryological development (22). The hair cycle is classically composed of three distinct phases: anagen (growth) phase, catagen (involution phase), and telogen (resting) phase (Fig. 1A-8). A fourth exogen stage has been described recently, although it remains a source of controversy as to whether it truly represents a separate stage in the hair cycle. Anagen generally lasts from three to ten years, catagen two to three weeks, and telogen three to four months. In the human scalp, the cycle is asynchronous. At any one time, an average of 13% of the hair is in telogen, although it can range from 4% to 24%. Only 1% is in the catagen phase. Neighboring hairs within an FU may be in different stages (Box 1A-3).
Anagen to Catagen
At the end of the anagen (growth) phase, the hair follicle receives an unknown signal to enter the catagen phase. The matrix cells cease proliferating, and the production of melanin terminates. The dermal papilla contracts and is released from the follicle. The middle portion of the follicle constricts and the lower portion expands to become the club hair. This lower portion retracts up to the level of the AP muscle leaving behind a thin band (streamer) of undifferentiated epithelial cells. The mechanism of retraction is felt, in part, to be due to apoptosis (controlled cell death) (23). Although the exact signal for the induction of catagen is not known, factor 5 (FGF5) has been implicated; in animal models without this factor, catagen is delayed (24).
Box 1A-3 Hair Cycle Duration
Anagen: 3–10 years
Catagen: 2–3 weeks
Telogen: 3–4 months
Exogen: weeks to months
Catagen to Telogen
The catagen phase is generally less than four weeks in duration and, therefore, it is difficult to distinguish it from an early telogen phase. Specific molecular signaling aiding in this transition has not yet been characterized.
Telogen to Anagen
During telogen, the band of epithelial cells (streamer) left behind during catagen moves upward and forms a short projection underneath the club hair called the secondary hair germ. This path upwards is followed by the dermal papilla, whose size is greatly reduced at this stage. At some point anagen is initiated by complex molecular signaling occurring between the dermal papilla, secondary hair germ, and stem cells from the bulge area. This signaling triggers stem cells to become transcriptionally active and migrate to the hair germ (24–26). The dermal papilla enlarges and the hair germ exhibits proliferation and downward growth eventually enveloping the enlarging dermal papilla. A new hair bulb is formed and anagen progresses.
Current evidence suggests that regulation and transition into anagen phase depend on the careful interplay of BMP, FGF7, and Wnt/β-catenin pathways (15).
Anagen
Club hair
Remnant dermal papilla
New germ
Catagen
Telogen
Figure 1A-8 Human hair cycle.
Exogen
The exogen phase begins with the completion of a club hair and ends with its release. Actual loss of the club fiber signals the end of the exogen phase (27). The term teloptosis has been used to describe the actual release of the club fiber, and the term kenogen has been used to denote the period of telogen without a hair fiber in the follicle (28). The exogen phase is considered to be a closely regulated, independent, cycle phase, although the exact molecular mechanism has not yet been described. On the receipt of an unknown signal, proteolytic enzymes may convert the previously tightly held hair shaft within the bulbous base of the follicle into a slenderized, more spear-shaped exogen bulb, which facilitates shedding of the hair.
ACKNOWLEDGMENT
The author would like to thank Patrick Tafoya for his illustrations.
1B1. ANATOMY OF THE SCALP
Matt L Leavitt and Paul T Rose
INTRODUCTION
The practice of hair transplant surgery is facilitated by a thorough understanding of the anatomical and histological structure of the scalp.
LAYERS OF THE SCALP
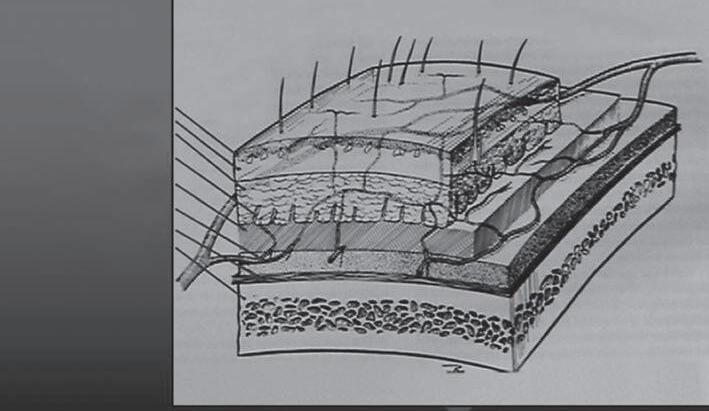
The scalp is composed of five distinct layers (Fig. 1B1-1). These layers are often referred to by the acronym SCALP, taken from the first letter of each level: skin, subcutaneous layer, aponeurotica, loose connective tissue, and pericranium. Specifically, these levels of the scalp are best characterized as follows.
Skin: The skin is the most superficial layer of the scalp • and consists of the epidermis (superficial layer) and the dermis (deep layer). It varies in thickness, ranging from 3 to 8 mm. The hair follicle and adnexal structures course through these layers. The bulb of hair follicles may be found in the upper subcutaneous layer before passing through the dermis and epidermis. The follicle is surrounded by a rich, end-organ network of nerves and capillaries.
The angularity of hair growth cannot be determined without specific examination. Although the direction of this angle tends to be the same in contiguous areas of the body, both the angle and the direction of the hair shaft can change within 2 cm or less. It is important to note that the angle of the hair at the surface is usually slightly different at the level of the fat and dermis. Therefore, great care is needed when harvesting donor tissue so that as few follicles as possible are transected, since the hair shaft angles vary.
The skin provides a relatively stable environment for the hair follicle. The dermis contains the arrector pili muscles. These muscles insert into the follicular sheath. Their action is to elevate the hair in response to cold, to fightor-flight reflex, or to other conditions that direct blood away from the skin. When one is cold or afraid, these muscles
contract, causing the hair to stand erect and compress the sebaceous glands, producing cutis anserina, or goose flesh. The sebum secreted from these glands lubricates the hair shaft and carries away debris generated in the follicle. Sweat ducts also pass through the two layers of skin from sweat glands in the subcutaneous layer.
Subcutaneous layer: The subcutaneous layer of the scalp • is a layer of superficial fascia that is located in between the skin (above) and the galea aponeurotica (below). It consists primarily of fat and a network of fibrous septa. This fibrous network creates stability between the top three layers of the scalp and allows them to move together as a unit over the deeper subgaleal layer of loose connective tissue. The main arteries, veins, lymphatics, and nerves of the scalp are located at the bottom of this layer just above the galea.
Aponeurotica (epicranium, galea): The galea is the muscu- • loaponeurotic layer of the scalp. It is a strong tendon-like structure interposed between the frontal muscle in the forehead and the occipital muscle in the neck and is the skull’s deep fascia. The galea represents the continuation of the frontalis muscle anteriorly and the occipitalis muscle posteriorly. Collectively, the galea and these two muscles are termed the epicranius muscle.
The aponeurotica is bounded anteriorly by the supraorbital margin along the frontal process of zygoma and the upper border of the zygomatic arch (Fig. 1B1-2). Posteriorly, it is bounded by the superior nuchal line. It wraps around posteriorly above the insertion of the sternocleidomastoid muscle and the origin of the trapezoid. Laterally, its upper border is the auricular muscles and it extends to the external auditory meatus along the mastoid process (1).
The tensile strength of this layer makes it useful when suturing various types of scalp reduction and flap procedures. However, this same tensile strength can sometimes become an obstacle in scalp surgery, making it difficult to close scalp defects without tension. The use of scalp expanders and scalp extenders under the galea can help alleviate this problem.
Loose connective tissue (subgaleal fascia): This level of • scalp is thin and (relatively) avascular except for the emissary veins that connect the scalp with the intracranial venous system (2). The loose connections in this layer allow for the free movement of the scalp. The avascular nature affords ease of undermining with minimal bleeding.
Although rare, scalp infections can occur. These infections can arise at the level of the loose connective tissue. At this level, the infection can progress via the emissary veins that course through the parietal foramina. From this location, the infection can involve the meninges. Additionally, because the frontalis muscle attaches to the skin rather than the underlying bone, space exists that allows for the spread of infection or other fluids into periorbital area.
Pericranium: This is the periosteum that is attached to the • skull and is the deepest layer of the scalp. Like the periosteum of other bones, it adheres to the underlying skull. In the region of the cranial sutures, it adheres because strong fibrous attachments to the dura mater exist.
Epidermis
Dermis
Subcutaneous layer
Fibrous septum
Galea aponeurotica
Subgaleal fascia
Pericranium
Cranium
1B1-1 Cross-section of layers of scalp tissue often referred to by the acronym SCALP (skin, subcutaneous layer, aponeurotica, loose connective tissue, pericranium).
Figure 1B1-2 Diagram of muscles in the scalp. Anterior: supraorbital margin along the frontal muscle (F.m.) process zygoma. Lateral: auricular muscles (A.m.m.)—the upper border external auditory meatus along the mastoid process. Posterior: superior nuchal line wraps around the insertion of sternocleidomastoid muscle and the origin of the trapezoid. G.a. refers to the galea aponeurotica and O.m. refers to the occipital muscle.
ARTERIAL SUPPLY
The scalp is one of the most vascular areas of the body (Fig. 1B1-3). Understanding the arterial supply is of critical importance to the hair restoration process, especially when performing the more complex scalp reduction and flap
Figure 1B1-3 Diagram of the arterial network in the scalp. Fivepaired arteries travel in the subcutaneous layer. In the lateral scalp: external carotid artery (E.c.a.) (three branches): (i ) superficial temporary artery frontal (S.t.a.) (transverse facial), parietal branch (P.b.) (middle temporal); (ii ) postauricular artery (P.a.a.); (iii ) occipital artery (Occ.a), internal carotid artery (I.c.a.); (iv ) supratrochlear (St.a); (v ) supraorbital (So.a), ophthalmic artery (Oph.a.).
procedures. Knowledge of the blood supply helps the physician avoid injury to major vessels and to unwanted intra-operative blood loss. Compromising the blood supply by overtransplantation can lead to poor growth or even necrosis. Remarkably, experience has shown that the scalp can remain viable even when only two of the arterial branches remain intact.
Figure
G.a.
O.m.
A.m.m.
F.m.
S.t.a.
Occ.a.
P.b. Occ.a.
I.c.a.
E.c.a.
St.a.
So.a.
Oph.a.
P.a.a.
The vascular supply to the scalp is derived from fivepaired arteries that arise from the external and internal carotid systems.
The External Carotid System
The external carotid system provides the blood supply primarily to the lateral and posterior scalp via three branches: (i) the superficial temporal artery, (ii) the posterior auricular artery, and (iii) the occipital artery.
The superficial temporal artery is easily found in the tem- 1. poral region, and travels in the superficial layers of the galea, just deep to the subcutaneous fat. It is a terminal branch of the external carotid. It usually branches into the transverse facial and temporal artery as it extends superior to the zygoma; however, its branching pattern may be quite variable. This artery anastomoses with the supraorbital and supratrochlear arteries anteriorly as well as the retroauricular and occipital arteries posteriorly.
The posterior auricular artery comes off the external carotid 2. artery superior to the digastric and stylohyoid muscles and courses along the retroauricular groove adherent to the periosteum of the mastoid process. Superiorly, the artery travels deep to the posterior auricular muscle as the auricular artery. It supplies the cranial surface of the ear and finally anastomoses with branches at the superficial temporal artery. In performing scalp reduction the surgeon may inadvertently injure this artery when trying to extend the dissection down to the ear.
The occipital artery originates from the external carotid at 3. the inferior margin of the digastric muscle. It divides into an ascending branch, a descending branch, and a transverse branch at the nuchal border of the splenius capitas and sternocleidomastoid attachments. These branches provide blood supply to the lateral half of the posterior scalp. It is important to note that lack of proper visualization and/or inadequate attention to the depth of the incision while donor harvesting is taking place may result in injury to the occipital artery or one of its branches. If the occipital artery is transected during surgery, it should be closed with either electrocautery or suturing of the vessel (3).
The Internal Carotid System
The internal carotid system provides the blood supply to the frontal scalp via two end branches of the ophthalmic artery: (i) the supratrochlear artery and (ii) the supraorbital artery.
The supratrochlear artery arises at the medial eyebrow 1. approximately 2 cm from the midline. The supraorbital artery exits lateral to the supratrochlear 2. and ascends into a similar subcutaneous plane. These vessels anastomose with each other as well as the superficial temporal and auricular arteries to form a rich vascular bed in the frontal scalp.
All main arteries travel just superior to the galea in the subcutaneous layer. The subcutaneous layer of the scalp is a particularly poor plane for dissection because of the presence of these large vessels and fibrous septa, which interfere with blood vessel constriction.
The existence and course of these main arteries, as well as their rich anastomoses, explain why scalp flaps often survive well (2). This high degree of vascularity also accounts for the
extremely low incidence of infection associated with scalp surgery. An infection in hair transplantation is almost nonexistent. The high degree of vascularity can, however, lead to profuse blood loss. Bleeding from lacerations of the scalp can be difficult to manage because the vessel does not tend to retract. Rather, the dense connective tissue surrounding vessels in the subcutaneous layer acts to hold vessels open. In the case of scalp reduction, and especially scalp lifting procedures, large amounts of blood can be lost quickly and the surgeon must be prepared to control the bleeding.
An artery can be located by palpation or by using a Doppler flowmeter. Arteries with triphasic or even biphasic waveforms are adequate to support flaps. The use of the Doppler flowmeter is particularly indicated in patients who may have aberrant vasculartiy (i.e., history of extensive trauma, previous scalp surgeries, or radiation therapy). Particular care must be taken when designing and creating flaps on a scarred scalp because neovascularity through a scar may be minimal. Thus, scalp flaps traversed by scars should not generally be used because their blood supply may be compromised.
VENOUS SYSTEM
Most scalp veins follow the arterial network and drain into the jugular system (Fig. 1B1-4). Emissary veins perforate the skull and drain into the dural venous sinuses.
INNERVATION OF THE SCALP
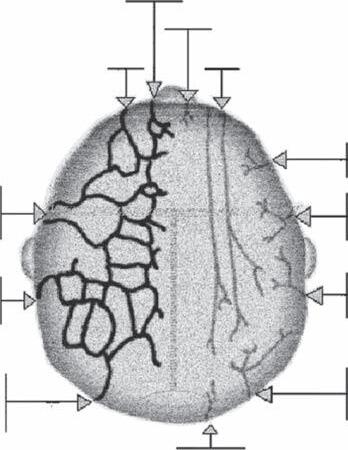
The innervations of the scalp are both sensory and motor (Fig. 1B1-5) (2). The sensory input is primarily derived from the three divisions of the trigeminal nerve: ophthalmic, maxillary,
Figure 1B1-4 The vein network in the scalp. Veins usually follow arteries and most often drain into the jugular veins. Emissary veins perforate the skull bone (and branches of the ophthalmic artery) and drain into intercranial veins. Abbreviations: S.t.v., superficial temporal vein frontal (transverse facial); A.a.v., anterior auricular vein; P.a.v., postauricular vein; Occ.v., occipital vein; St.v., supratrochlear vein; So.v., supraorbital vein; I.j.v., internal jugular vein; E.j.v, external jugular vein.
So.v.
St.v.
and mandibular. The motor innervations of the scalp are primarily supplied by branches of the facial nerve. It is particularly important for the hair restoration surgeon to understand the sensory pathways of the scalp because procedures are often performed under local anesthesia with field or nerve blocks. The sensory innervations are such that a caudal block creates a field of anesthesia cranially (3).
Sensory Innervations
Sensory branches have a distribution similar to that of the arterial network. The major innervations of the scalp are described as follows:
Supraorbital and supratrochlear nerves: These nerves sup- • ply the sensory innervations to the forehead and frontal scalp as far back as the crown. They are terminal sensory branches of the ophthalmic nerve, which is the (superior) division of the trigeminal nerve. The supraorbital nerve exits the scalp through the supraorbital foramen. The nerve is easily palpated by feeling for the depression at the orbital rim, which represents the exit point of the nerve. It is usually within 2.5 to 3.0 cm lateral to the midline of the brow. The supraorbital nerve supplies a greater degree of innervations to the scalp than the supratrochlear nerve. These nerves are common sites of nerve blocks.
Zygomatic nerve: This nerve is a branch of the maxillary • (middle) division of the trigeminal nerve, which exits the skull through the infraorbital foramen. The zygomatic nerve quickly divides into two branches named the zygomaticofacial and zygomaticotemporal nerves. The zygomaticotemporal nerve provides the sensory innervations for the anterior temporal scalp. The zygomaticofacial nerve supplies the innervations for the malar area.
Figure 1B1-5 Innervation of the scalp. Distribution similar to that of the arterial network. Sensory ability is provided by all three branches of the cranial nerve and upper cervical nerves. Abbreviations: So.n., supraorbital nerve; St.n., supratrochlear nerve; Zt.b.z.n., zygomaticotemporal branch nerve; L.o.n., lesser occipital nerve; G.o.n., greater occipital nerve; P.a.n., posterior auricular; P.a.b.f.n., posterior auricular branch facial nerve; F.n., facial nerve; T.b.f.n., temporal branch of facial nerve; At.n., auriculotemporal nerve; G.a.n., greater auricular nerve.
Auriculotemporal nerve: This nerve supplies the sensory
• innervations to the temporal and temporal parietal scalp as well as part of the anterior ear. It is a branch of the mandibular (inferior) division of the trigeminal nerve. It is located adjacent and deep to the superficial artery and vein.
Greater auricular nerve: This nerve supplies sensory inner-
• vations to the postauricular area as well as part of the occipital area. It is an extension of the cervical plexus, which is formed by the anterior rami of the second, third, and fourth cervical nerves.
Lesser occipital nerve: This nerve supplies sensory inner-
• vations to the majority of the posterior occipital area. It is also a branch of the cervical plexus.
Motor Innervations
The facial nerve (seventh cranial nerve) arises from the pons and supplies most of the muscles of facial expression. It has five major branches: temporal, zygomatic, buccal, mandibular, and cervical. The temporal branch of the facial nerve supplies motor innervations to the frontalis muscle, upper portion of the obicularis oris muscle, and corrugators supervilli. It is considered one of the most vulnerable branches of the facial nerve because of its location. It exists under the parotid gland and crosses the zygomatic arch before innervating the frontalis muscle. The posterior auricular muscles and the occipital portion of the epicranius muscle are also supplied by branches of the facial nerve. The temporalis muscles receive innervations through the anterior and posterior deep temporal branches of the mandibular division of the trigeminal nerve. During donor harvesting and scalp reduction, superficial sensory nerves of the scalp may be partially severed, but usually without permanent consequence. The motor nerves in the occipital muscles are rather deep and are rarely injured. Major sensory nerves are usually not injured during the course of hair transplantation or midline scalp reduction because they are not ordinarily undertaken near them. The situation is different with extensive scalp lifting or flap procedures during which greater care needs to be taken to locate and preserve neurovascular bundles.
The occurrence of hypoesthesia, lasting as long as a few months after scalp surgery, is common, and patients should be so warned. This numbness is presumably a direct result of surgical trauma. A few postoperative patients may have areas of permanent numbness or paresthesia, but the condition creates no significant problem for them. Inducing tumescence in the occipital area before donor harvesting, elevates the skin away from the deeper nerves and vessels and may decrease the risk of injury.
In addition to the neural network in the scalp, a fine lattice of sensory nerves envelops the entire length of the follicle. Nerve fibers from the deep dermis approach from all quadrants and converge on the hair follicle at the level of the subcutaneous ducts. This perifollicular network of nerves and end organs greatly extends the range of cutaneous sensory perception.
LYMPHATIC SYSTEM
The lymphatic fluid from the scalp drains into the pericervical complex of lymphatics. This so-called ring consists of the submental, submandibular, mastoid, parotid, and occipital nodes. The drainage from those nodes enters into the deep cervical lymph nodes.
At.n.
P.a.b.f.n. P.a.n.
1B2. SURGICAL RAMIFICATIONS OF SCALP ANATOMY
Gerard E Seery
ANATOMY
The scalp is a five-layered tissue composed of collagen, elastin, blood vessels, nerve fibers, and lymphatics with mucopolysaccharide ground substance, tissue fluid, hair follicles, and sebaceous and sweat glands (Fig. 1B2-1).
Skin
The scalp skin, the first layer, is thicker than skin elsewhere on the body, ranging from 3 (vertex) to 8 mm (occiput).a It contains hair follicles, sebaceous and sweat glands, and is firmly bound to the deeper tissues.a
Subcutaneous
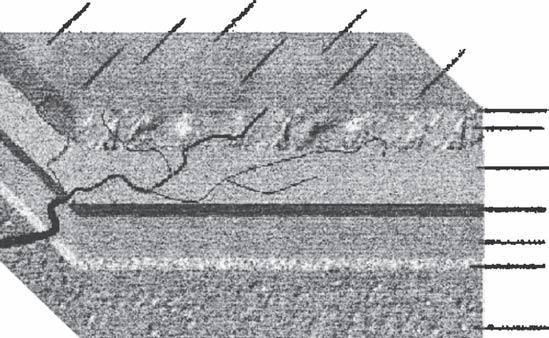
The second, or subcutaneous, layer is notable for its rich network of anastomosing arteries, veins, and lymphatics (Fig. 1B2-2). Subcutaneous fat is divided into multiple small compartments by fibrous septa. The arteries are attached to the deep layers of the dermis, and the profuse hemorrhaging often seen in superficial wounds of the scalp is attributed to a relative inability of the arteries to retract because of their attachment to the dermis and fibrous septa (1,2). The practical application of this is that manual compression and immediate suture, without attempting to grasp bleeding points with hemostats, may be the most effective way of controlling bleeding from superficial scalp lacerations.
Vascular Considerations
The scalp is supplied by a system of anastomoses between branches of the external and internal carotid arteries (Fig. 1B2-3). The blood supply is centripetal, that is, the larger trunks run medially and centrally from the periphery, becoming smaller as they enter a system of free anastomoses with their fellows. This and their spatial or depth location are of critical importance to the surgeon. The arteries are cutaneous and classically described as running in the subcutaneous
tissues. The fact that scalp is not supplied by musculocutaneous perforators (3) is of enormous practical importance for surgeons, as surgical transection of subcutaneous vasculature, and particularly of larger trunks in the periphery of the scalp, has dire consequences for the blood supply of tissues both adjacent to and distant from the transection site. The importance of this will be discussed later. In all likelihood, scalp also derives at least some minimal blood supply from bone perforators deriving from meningeal vessels. This would explain the survival of areas of surgically circumscribed scalp. The belief that scalp has a superabundant blood supply and consequently is very forgiving of surgical indiscretions may need to be reconsidered. This has particular relevance to hair restoration surgery.
In 1980, Klemp and colleagues (4) evaluated scalp subcutaneous blood supply in subjects with male pattern baldness (MPB) and concluded that blood supply is reduced in bald tissues relative to controls. In 1990, Toshitani and associates (5), using Doppler flow meter and thermography studies, demonstrated relatively reduced flow in the central scalp of MPB patients. In 1996, Goldman (6) reported details of a study that measured scalp transcutaneous pO2 in subjects with MPB. Significant microvascular insufficiency in regions of the scalp that lose hair and an associated relative tissue hypoxia were found. These findings have particular relevance to hair restoration surgery, with the possibility that relative local hypoxia in bald areas may play a part in the pathophysiology by which age, genetics, and androgens interact to cause MPB. It may also be a factor in explaining why the diameters of transplanted hair become significantly reduced in the post-transplant state (7).
Neurological Considerations
The sensory nerve supply of the scalp is similar to the vascular anatomy in that it is centripetal, subcutaneous, and of a similar distribution. Incisions in the peripheral scalp that transect vital larger vascular and nerve trunks result in extensive, hypesthetic, poorly vascularized tissues and should be avoided. Where avoidance is not possible because of the specific goals of the surgery, a knowledge of the exact anatomical location of the major neurovascular trunks is imperative
Epidermis
Dermis
Subcutaneous Layer
Galea aponeurotica
Subgaleal space
Pericranium
Cranium
Figure 1B2-1 Cross-section of the scalp. Note the location of the subcutaneous vascular plexus.
(as is magnification) if these structures are to be spared during surgery. A corollary of the above is that incisions in the central scalp are associated with minimal neurovascular trauma, making their use the preferred choice where a choice exists.
Galea Aponeurotica
The galea aponeurotica (Fig. 1B2-4) (8), or third layer of the scalp, should be considered only when taken in conjunction
Supratrochlear artery
Supraorbital artery
Superficial temporal artery
Posterior auricular artery
Occipital artery
Supratrochlear nerve
Supraorbital nerve
Zygomaticotemporal nerve
Auricolotemporal nerve
Lesser occipital nerve
Greater occipital nerve
Third occipital nerve
with the occipitofrontalis muscle, which has occipital and frontal bellies separated by the aponeurosis into which the muscle bellies are inserted. The occipitalis bellies arise from the lateral three fourths of the highest nuchal lines and the mastoid and pass forward into the aponeurosis. The galea, also called the epicranial aponeurosis, lies over the vertex between the bellies of the occipitofrontalis muscle. It fades out laterally by blending with the temporal fascia. The anterior frontalis muscle arises from the front of the aponeurosis and is inserted into the upper part of the orbicularis oculi and the overlying skin of the eyebrow. Scalp skin is firmly bound down to the muscles and aponeurosis. The occipitalis muscle is supplied by the posterior auricular nerve and the frontalis by the superior zygomatic branch of the facial nerve. The occipital belly pulls the scalp back in certain individuals, but usually only anchors the aponeurosis. Contraction of the frontalis elevates the eyebrows and produces the horizontal wrinkles in the forehead.
The subaponeurotic space extends beneath these muscles and aponeurosis over the vault of the skull. It is limited behind by the attachments of the occipitalis to the highest nuchal lines and at the sides by blending with the temporal fascia. In front, the space extends down beneath the orbicularis oculi into the eyelids. Bleeding may track down in this space and produce the ubiquitous periorbital hematoma, or “black eye.”
The galea is a dense, inelastic membranous or fibrous sheet normally 1 to 2 mm in thickness. It is better developed in some individuals than others, and the occasional case of
Figure 1B2-3 The location and extent of the galea aponeurotica.
Figure 1B2-2 Neurovascular anatomy of the scalp.
Figure 1B2-4 Diagrammatic representation of the lines of minimum tension (Langer’s lines).
congenital absence has been reported (Stough D, personal communication). Its surface is densely adherent to the subcutaneous tissues, thus making dissection in this plane difficult and bloody.
The galea is significantly resistant to stretching and is believed by some surgeons to be the main factor standing in the way of meaningful scalp resection. Although this conclusion is understandable, the argument is made here that the galea is enormously beneficial in that it limits the harmful effects of surgical overstretching. Consequently, it is likely that it is actually protective and facilitates complication-free scalp surgery.
The belief that the relative inability of galea to stretch limits the extent of surgical resection and has led to the surgical procedure of galeotomy, or linear galeal transection, done in an attempt to surgically limit its restrictive effects. Raposio and associates (9), in a study designed to measure the compliance of flaps in sagittal scalp reduction, describe making three relaxing incisions in the galea, each 12 cm in length and parallel to a sagittal scalp reduction incision. The compliance of the flaps to stepwise loading was measured by dynamometer. The aim of the study was to assess the quantitative effects of galeotomies on the biomechanical properties of a scalp flap to quantify the surgery-related benefits provided by the procedure. It was concluded that the mean gain in length of the flap per galeotomy was 1.67 mm. This corresponds to a mean 40% reduction of the closing tension obtained with each galeotomy.
This confirms the clinical impression of the usefulness of galeotomy for obtaining wound closure in cases where it would otherwise be difficult or impossible. The procedure, however, is not without a downside and is prone to traumatize subcutaneous structures and produce troublesome bleeding. It is associated with an increased incidence of hematoma and infection. By and large, galeotomy has fallen into disuse.
Subgaleal Layer/Space
The subgaleal compartment, the fourth layer of the scalp, is bounded by galea above and has pericranium as its floor. It is largely avascular, containing only a filmy layer of loose fibroareolar tissue. It is of enormous surgical importance and is a space that readily lends itself to surgical dissection.
The scalp may be considered as being composed of two anatomically different parts: the part that overlies the subgaleal space and the part that does not. The former extends from the superior nuchal lines (to which the occipitofrontalis muscle and galea aponeurotica attach) and proceeds cephalad into the forehead. The lateral limit of this space ends where the galea blends with the temporalis fascia. That part of the scalp overlying the subgaleal space has the standard five layers: skin, subcutaneous tissue, galea, subgaleal loose areolar layer, and pericranium. The remainder of the scalp has three layers: skin, subcutaneous tissues, and deep fascia (e.g., the deep fascia that overlies the trapezius and sternomastoid muscles).
The anatomic difference between the two areas has great practical importance, as it is this that largely determines the widths of tissue amenable to surgical removal. Generally speaking, where five layers of scalp exist, wide excisions are possible; where three layers exist, the width of tissue amenable to removal is relatively restricted. The five-layered scalp is
notable for having the subgaleal fibroareolar layer, which allows the scalp to slide or glide on the pericranium. It is this property that allows enhanced excision relative to the remainder of the scalp.
The orientation of the lines of minimum tension (Langer’s lines) also play a major part in determining the width of the excision (Fig. 1B2-5). These topics are discussed in some detail later. It must also be remembered that some scalps have relatively poorly developed fibroareolar layers (i.e., “tight scalps,” in which the gliding phenomenon is minimal). The extent of this is easily determined by simply placing the pulps of the examining fingers on the scalp and moving it on the underlying pericranium.
As was mentioned earlier, the blood supply to the scalp is cutaneous; that is, the vessels enter from the periphery and run medially in the subcutaneous tissues. This means that if surgical dissection is confined to the subgaleal plane during reconstructive procedures, the blood supply to the tissues is not compromised in any way. This in turn means that large flaps can be safely raised without delaying procedures (10) (remember, however, that incisions in the peripheral scalp that transect arteries may grossly limit blood supply to areas both adjacent to and remote from the locus of transection. This is in contrast to tissues supplied by musculocutaneous perforators). The ideal scalp incision is midline because this allows access to the subgaleal space while inflicting minimal arterial and nerve trauma.
Pericranium
The pericranium is the fifth and deepest layer of the scalp. The pericranium is not well described in textbooks of anatomy. The following description is based on more than
The pericranium raised as an intact membranous sheet. Note the comparative thickness of the pericranium and galea aponeurotica.
Figure 1B2-5
700 pericranial dissections I have performed (11). The pericranium is a dense membranous or fibrous sheet (Fig. 1B2-5) loosely fused on its outer aspect to the galea aponeurotica, from which it is readily separable via the subgaleal space. It is usually of similar thickness to the galea and is easily bluntly dissected from the outer table of the skull as an intact sheet (contrary to descriptions in textbooks, which state it to be bound down and densely adherent at the skull suture lines).
Pericranial dissection from bone is bloodless other than the occasional, easily controlled, minimal bleeding from the rare bone perforator. Subperiosteal stripping is simple, easily accomplished, and safe.
Over the course of the work, multiple periosteal flaps of various sizes were raised in different locations, thereby denuding the outer table of the skull of periosteal cover. On re-exploration months later, the denuded areas were covered with tissue indistinguishable from pericranium.
Pericranial thickness varied from one individual to another and from one skull location to another in the same individual, but the impression is that the pericranium in the frontal areas may be slightly thinner than on the crown.
The pericranium is capable of retaining sutures even when subjected to significant tension. Incorporating a pericranial flap in deep wound closure seemingly contributed to the production of fine linear scalp scars. It was apparent that the pericranium is a very serviceable tissue and probably is underutilized in reconstructive and cosmetic craniofacial surgery.
DISCUSSION
The scalp is subject to a multiplicity of conditions amenable to surgical treatment, including benign lesions (sebaceous cysts, nevi, keratoses, hemangiomas, and others), malignant lesions (basal cell and squamous cell carcinoma, melanoma), burns, congenital malformations, hair restoration, and a variety of miscellaneous conditions including neurofibromas, aneurysms, turban tumors, and others. The list is quite extensive. It is reported that 72% of automobile accident victims sustain head injuries, making scalp trauma a leading cause of emergency room visits (12). Because of this array of conditions requiring scalp surgery, its lineage can be traced to general surgery, plastic surgery, neurosurgery, dermatologic surgery, craniofacial surgery, and hair restoration surgery—none of which seem to claim or want dominion.
Neurosurgeons and craniofacial surgeons may see the scalp as a barrier that must be breached in order to gain access to the brain and craniofacial skeleton. To plastic surgeons, the scalp is a tissue in which incisions may be hidden and flaps developed. Hair transplantation surgeons are inclined to view it as a passive tissue in which hair is surgically transplanted and/or rearranged. General and dermatologic surgeons often make little distinction between hairless skin and scalp in their choice of surgical methodology.
This fragmentation of scalp surgery through the various specialties and an element of tunnel vision on the part of some surgeons who, interested only in scalp surgery as it relates to their own particular specialty, may contribute to the high incidence of complications that are a feature of present-day scalp surgery. Such complications include stretched unsightly hairless scars, distorted hair patterns, hair loss, insensate scalp,
and plasticized or stretch-atrophied tissues, virtually all of which are avoidable if certain simple parameters of surgery are observed (13).
Incisions/Excisions
Lines of cleavage in the skin were first described by Langer in 1881 and reinvestigated by Cox in 1941 (14). The lines are believed to be due to collagen bundles arranged in parallel in the dermis, although this has not been confirmed by electron microscopy (15). Where crease lines exist, their direction generally coincides with the lines of minimum tension. These tend to be longitudinal in the limbs and scalp, and circumferential in the neck and trunk. The lines of minimum tension are surgically important for two main reasons: elliptical excisions achieve the greatest width of tissue removal when made parallel to the lines, and incisions made along or parallel to the lines heal with a minimum of scarring, whereas incisions made across them heal less well.
These findings have led to the concept of collagen transection scarring, which is the unsatisfactory scarring that results when incisions are made across collagen bundles. Consequently, knowledge of the orientation of collagen in the skin is clearly to the surgeon’s advantage. It is likely, however, that present knowledge of collagen orientation may still be rudimentary. In light of this, the argument is made that, where possible, surgical incisions in scalp tissue should be made in a vertical axis, except in the occipital hair transplantation donor, for whom a horizontal axis paralleling crease lines are preferred.
Although much has been made of the problems of collagen transection, it is probably as important not to transect elastic fibers. Fortunately, as collagen and elastin generally run in parallel, incisions that spare one spare the other. Ideally, incisions should also parallel the directional orientation of hair in order to avoid hair follicle transection.
Scalp Laxity
The extent of surgical scalp excision possible is predominantly a function of scalp laxity. The more lax the tissue, the greater the surgical excision.
Scalp laxity has two very distinct components. First is the ability of the scalp to slide or glide on the underlying pericranium. This is facilitated by the loose fibroareolar tissue in the subgaleal compartment, which allows the scalp to be moved on the cranium, often for a distance of several centimeters. This has nothing to do with tissue extensibility or stretching and is due to simple mechanical movement of the scalp on the pericranium (16). In a scalp with a high capacity to slide/glide, an excision width of 5 cm or more may be made and closure obtained without any undermining or stretching. Operations that take advantage of the scalp’s capacity to glide rather than stretch are virtually complication free and result in negligible topographic distortion of the tissues. The analogy of pulling a carpet over a polished floor comes to mind. The carpet and the furniture are moved but their topographic relationships to each other are neither changed nor are the physical components of the carpet altered. Scalp surgery that utilizes the scalp’s facility to glide is highly effective and minimally traumatic to tissues.
The second component of scalp laxity is its elasticity or ability to elongate (loosely termed, “stretch”). It is
reiterated that this is independent of the sliding phenomenon. Some scalps are highly elasticized, and even in the presence of the relatively inelastic galea are capable of reasonably significant elongation. Lines of minimum tension (Langer’s lines) are largely vertically disposed in scalp overlying the vault of the skull and allow generous excision of tissue cut in a vertical axis. Conversely, there is an associated relative limitation in excision widths in the horizontal axis (because here Langer’s lines are crosscut), but this is more than offset by the laxity provided by the fibroareolar layer in the five-layered scalp.
As Langer’s lines proceed downward or caudad into the occipital scalp, they increasingly assume a horizontal orientation, and in the lower part of the occipital (hair transplantation donor area) are entirely horizontal. This facilitates a relatively wider excision in this area than would otherwise be the case, but this does not nearly compensate for the absence of the “gliding” subgaleal fibroareolar layer present in the upper donor area. This has particular relevance to the donor area in hair transplantation. Here the upper donor area (i.e., above the superior nuchal lines) has five layers and allows relatively wide excision strips, whereas the lower part (below the superior nuchal line) has three layers only and excision is relatively restricted (Fig. 1B2-6). Maximum-width excisions are possible when made parallel to the lines of minimum tension and where five layers of scalp are present.
Biomechanics
The ability of skin to recover from stretch resides in its elastin component. When skin stretches, the elastic fibers elongate in
the direction of the stretching force, allowing the convolutions in collagen to straighten out. The resulting elongation is a function of progressive displacement of ground substance and tissue fluid that accompanies collagen realignment. This continues until there is a structure of parallel collagen fibers that resists further extension provided the elastic limit is not exceeded (17).
The elastic limit of skin is that point at which the components commence to rupture and the stress–strain ratios no longer apply.
It is accompanied by adverse tissues changes. The elastic limit for skin elastin is generally about 100% and for collagen 10%; stated differently—when skin elongates more than 100% of its resting length, the elastic fibers rupture. The impaired elastin is no longer able to return the collagen to its normal resting state, even when stress is removed. This results in permanent irreversible adverse consequences for the tissues called plasticization, better known to surgeons as stretch atrophy (thin, dry, brittle, poorly vascularized skin). This is commonly seen after serial scalp excision, particularly if traction closures have been done. Stretch-atrophied tissues are relatively unsatisfactory for subsequent surgery (18). This is particularly the case in hair transplantation.
Skin stretching also attenuates blood vessels, thus decreasing tissue perfusion. If allowed to continue unchecked, it will ultimately exceed the critical closing pressure and stop perfusion. Lesser degrees of stretch reduce circulation. Elongation of the nerves and lymphatics causes pain and edema, respectively. Nonundermined skin is better able to withstand the ill effects of tension stretching than undermined skin (19).
Undermining
The question about the advisability or necessity of undermining inevitably arises. It is believed that modest judicious undermining is usually indispensable for optimal wound closure. Extensive undermining, however, may be ineffectual and associated with problems. Undermining, particularly when extensive and blind, is potentially harmful as it opens tissue planes to infection, has the potential for traumatizing blood vessels and nerves, results in extensive scar formation throughout the undermined area, and allows tension forces to be conducted into areas remote from the wound. It is also ineffectual, as explained later.
I have found that in very lax scalps, it is possible to remove 5 cm widths at the midsagittal ellipse level and yet easily close the wound without any undermining. Conversely, in tight scalps, removing 5 cm and achieving closure is impossible regardless of the extent of undermining. This led to the conclusion that the more important factor, by far, in determining the extent of tissue amenable to excision is not the extent of undermining, but the degree of laxity. It was also concluded that the extent of undermining and tissue amenable to removal are not linearly related.
In a clinical research study (11), two groups of identical operations were described, one with undermining of 15 cm and the other with only 5 cm of undermining. The excision width in each group was identical—39 mm. There was no significant difference in stretch-back. This suggested that the extra undermining of 10 cm bilaterally contributed nothing in terms of increase of tissue excision amounts.
Figure 1B2-6 Schematic of a hair transplantation donor area. Lower deeply shaded area has three scalp layers only. Lightly shaded area above has five layers.
This conclusion is scientifically supported by Raposio and colleagues (9), who in an excellent article on tensiometric measurements in serial scalp reduction, reported “the benefits of an extensive (15 cm) undermining were minimal as compared with those obtained with 5-cm undermining.”
PRACTICAL CONSIDERATIONS
At its simplest, integumental surgery comprises excision and suturing. The tension generated at wound closure is manifested maximally at the wound site and adjacent tissues. The more tension, the greater the deleterious effect on wound healing and the viability of tissues.
In order to effect wound closure, it is necessary to counteract the tension vector forces tending to keep the wound open. Although closure may be possible by overpowering the tissues with tension clamps, big needles, heavy sutures, large “bites,” and muscular exertion, the price paid for this in terms of tissue viability may be prohibitive. It is believed that the modalities of wound closure, passed down from one generation to another, are in dire need of rethinking.
Simple skin edge to skin edge suturing ensures that tension is borne by the skin alone. Layered closure is helpful, to some extent, in fractionating tension into increments of force evenly distributed throughout the wound. A layered skin closure, however, does not lessen the total force on the tissues necessary to effect closure.
In 1994, I embarked on a clinical research project (20) to determine how deleterious effects of tension in wounds could be ameliorated or prevented. This yielded information that went beyond the initial goals of the study. Data were derived pertaining to the adverse effects of widespread undermining; the benefits of directing tension forces into nonundermined tissues; the practical utility of maximally using the scalp’s capacity of gliding on the pericranium rather than stretching; and perhaps most important of all, the concept of deep plane fixation (which facilitates removal of greater amounts of tissue while allowing reduced tension wound closure). Deep plane fixation is explained in a series of articles published in Dermatologic Surgery (11,13,19,21).
CONCLUSION
The features of surgical anatomy of the scalp deemed most relevant to facilitating complication-free surgery are detailed and discussed. It is believed that optimal scalp surgery is consistently possible if the simple parameters of surgical practice detailed in this section are respected.
1C. NEW SOFTWARE FOR MAKING DENSITY AND SURFACE AREA MEASUREMENTS IN HAIR TRANSPLANTATION
Jean Devroye
INTRODUCTION
In hair transplantation it is useful, some would argue essential, to accurately quantify hair density, follicular unit (FU) density, hair diameter, the proportion of hairs in telogen and anagen, and surface area. Defining these characteristics
is indispensable in carrying out research and evaluating results. Furthermore, as Dr. Ron Shapiro has eloquently pointed out on innumerable occasions, planning in hair transplantation is in many respects a mathematical problem: except in rare cases, the available donor supply is less than what is required to fully restore density to an area of alopecia, and, therefore, hair restoration surgeons must effectively ration. This can only be accomplished if they have an accurate means of quantifying both the available donor supply, and the size of the current and anticipated recipient region.
EXISTING MEASUREMENT TOOLS IN HAIR TRANSPLANTATION
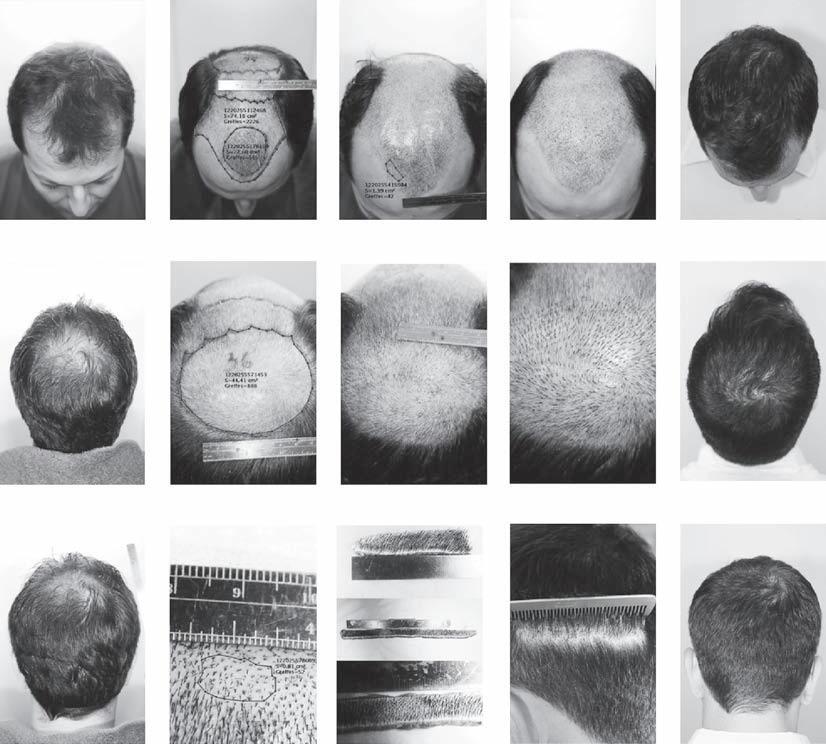
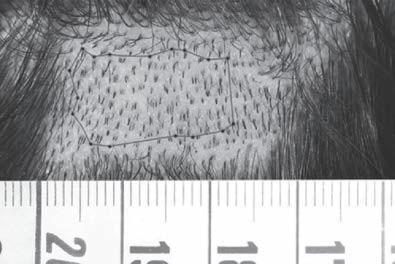
Over the years, a number of individuals have developed instruments designed to measure these variables of interest: some are simple but functional, while others are technologically complicated. Chang, for example, developed a basic but effective method for determining the size of the recipient area that involved superimposing tracings from the scalp onto grids (Fig. 9A1-20). Other practitioners have relied on macrophotography and microphotography, using either digital or plain film cameras, and either manual or automated counts and measurements (see Chapter 9A1 for additional information on the different types of devices).
A NEW MEASUREMENT TOOL
Added to this group of instruments is a new measurement tool. With the aid of a French programmer, Alexandre Duvallet, who previously developed software to analyze aerial photographs, the author created the software program Hairmes. In digital photographs that include a calibrating marker, such as a straight ruler or metric ribbon, it calculates information on surface area, FU density, expected donor yield, and incisional density. In addition, it corrects parallax errors caused by the curvature of the scalp. It has also a counter. It’s possible to tag the follicles and to obtain an abstract with the number of FU1, FU2, FU3 per cm2. An example of a complete photographic analysis can be seen in Fig. 1C-1.
Donor Area Measurements
In the donor area, the hair is clipped short prior to digital photography. Once the zone of interest is digitally traced, the software will calculate the FU density (Fig. 1C-2). Donor area density varies among locations; thus the author recommends that the density is calculated using an average of three measurements taken from the occipital, parietal, and temporal donor regions.
This information, combined with a measurement of the surface area of the donor strip, can be used to produce the expected yield of a surgical session. To compute surface area, practitioners must account for tissue elasticity, since both the width and length of donor strips decrease approximately 10% once removed from the scalp. Therefore, the width and length of the strip should be measured both before and after strip harvesting. A comparison of these values will permit calculation of a global corrective factor that can be applied to the surface area measurements (Fig. 1C-3). Once the donor density and the surface area of the donor strip are both known, the program will compute the expected donor yield, expressed as an absolute number of FUs.
Patient 34 years oldDonor area
3289 grafts 3384 FU 8423 hairs
Density 90 FU/cm2
Strip area measured 35.6 cm2
Number FU estimated 3205
Recipient site
Frontal area a 51.6 cm2 * 35 = 1805 FU
Frontal area b 22.6 cm2 * 30 = 678 FU
Vertex 44.4 cm2 * 20 = 888 FU
Total : 3371 FU
Figure 1C-1 An example of a complete surgical analysis using Hairmes. Abbreviation: FU, follicular unit.
S=1.73 cm2 Nbre grafts=121 Density=69.9
S=1.62 cm2 Nbre grafts=119 Density=73
Figure 1C-2 Measurement of follicular unit density in the donor region.