

Arteaga Martínez • García Peláez


Arteaga Martínez • García Peláez
Prefacio XI
Manuel Arteaga Martínez, Ma. Isabel García Peláez
SECCIÓN 1
1 2 3 4 5 6 7 8

Fertilización. El inicio de una nueva vida 63
Carlos E. de la Garza González, Josué Raúl Cortés Arellano
Desarrollo embrionario presomítico: la primera semana
Manuel Arteaga Martínez, Isabel García Peláez, Carlos E. de la Garza González, Anacleto de Jesús Merlín Pérez
Desarrollo embrionario presomítico: la segunda semana
Manuel Arteaga Martínez, Isabel García Peláez, Teófilo Toledo Hiray
75
83
Desarrollo embrionario presomítico: la tercera semana 93
Isabel García Peláez, Manuel Arteaga Martínez, Adriana González Villalva
Desarrollo embrionario somítico: de la tercera a la octava semana (etapa de organogénesis) 107
Manuel Arteaga Martínez, Isabel García Peláez
Desarrollo fetal: de la novena semana al nacimiento
Esteban Cruz Arenas, Patricia Aguilar Fuentes, Manuel Arteaga Martínez
Desarrollo del sistema muscular
Esteban Cruz Arenas, Isabel García Peláez, Manuel Arteaga Martínez
Desarrollo de los miembros
Manuel Arteaga Martínez, Isabel García Peláez, Laura G. Flores Peña, Adolfo Leyva Rendón, Teófilo Toledo Hiray
Desarrollo del sistema digestivo
Dora Virginia Chávez Corral
275
285
299
Desarrollo del sistema respiratorio 327
Ma. Guadalupe Treviño Alanís, Sergio Salazar Marioni, Teresa Fortoul van der Goes, Jorge Alberto Martínez Dávila, Manuel Arteaga Martínez
Desarrollo del sistema cardiovascular 341
Manuel Arteaga Martínez, Isabel García Peláez, Concepción Sánchez Gómez †
Desarrollo del sistema urogenital Índice analítico 551
Conceptos básicos de la señalización molecular en el desarrollo
Isabel García Peláez y Manuel Arteaga Martínez
OBJETIVOS
• Conocer los procesos implicados en el desarrollo para entender los eventos que participan en la embriogénesis y el desarrollo fetal.
• Identificar y analizar los procesos del desarrollo para integrarlos en la morfogénesis.
INTRODUCCIÓN
Durante el desarrollo prenatal a partir de una célula (el cigoto) se forma un organismo multicelular complejo con una anatomía particular que incluye un conjunto de órganos y estructuras formados por células especializadas que cumplen diferentes funciones.
La morfogénesis es el proceso por el que un organismo adopta su forma mediante la creación de tejidos, órganos y estructuras. Los mecanismos por los que, a partir del cigoto, se compone un organismo multicelular con una anatomía particular son los llamados que incluyen el crecimiento, la diferenciación celular, el cambio en la forma celular, la muerte celular, el movimiento celular y la afinidad celular diferencial, que les confiere a las células la posibilidad de unirse de manera selectiva durante la morfogénesis. A continuación, se muestran los principales procesos básicos del desarrollo.
CRECIMIENTO
El crecimiento da lugar al aumento en el tamaño, la configuración de la morfología y el cambio en las proporciones anatómicas.
El crecimiento es un proceso por el que se incrementa el tamaño de tejidos, órganos o estructuras como resultado de los siguientes fenómenos:
• El incremento en el número de células por proliferación celular, a partir de la división controlada de las células. Las moléculas que regulan el crecimiento son proteínas extracelulares como los mitógenos, los cuales estimulan la división celular mediante el control del ciclo celular.
• El aumento en el tamaño de las células, que se produce durante la fase G1 del ciclo celular, que está regulado por factores de crecimiento que promueven la síntesis
de proteínas y otras macromoléculas, así como el incre
La presencia de factores de supervivencia que impiden
Estos patrones de crecimiento se pueden combinar y también presentarse en una determinada región o dirección. El tamaño es regulado por señales extracelulares que activan o inhiben la división celular, el crecimiento celular o la apoptosis.
Durante el desarrollo embrionario, las diferentes estructuras y regiones del cuerpo crecen a distinta velocidad, lo cual se denomina crecimiento alométrico. Así, el embrión va modelando su morfología externa e interna a la vez que aumenta su tamaño. En la etapa fetal y posnatal, el crecimiento continúa siendo alométrico, por lo que las proporciones corporales cambian con la edad (Figura 5-1).

La diferenciación celular se desarrollan diferentes tipos celulares, como eritrocitos, neuronas, células musculares, etcé tera. Se efectúa mediante la expresión de genes específicos.
El inicio del desarrollo comienza con una única célula denominada cigoto, como resultado de la fertilización del ovocito por el espermatozoide. A partir del cigoto, por la diferenciación, se originan todos los distintos tipos celu lares del organismo con diferente morfología y función. En la diferenciación se expresan genes específicos que dan como resultado proteínas concretas que dan lugar a un cambio en la forma y la función de la célula (
Especificación de la diferenciación
Los cambios en la forma y función celular van prece didos por un proceso que compromete a la célula a su destino de diferenciación final, lo cual está determinado por la regulación irreversible en la expresión de sus genes. En los vertebrados, el destino de la diferenciación está especificado por las interacciones de las células, sus posiciones relativas y las cantidades específicas de moléculas secretadas por otras células, que se denominan morfógenos, como las proteínas morfogenéticas óseas (BMP, bone morphogenetic proteins), los factores de crecimiento transformantes (TGF, transforming growth factor) o los factores de crecimiento fibroblástico (FGF, fibroblast growth factor ). Asimismo, se ha comprobado la regulación de la diferenciación por medio de otras vías de señalización mediante ligandos como el ácido retinoico, Sonic hedgehog (SHH) y NOTCH.
La expresión y represión de genes específicos se mantiene en las células diferenciadas y sus generaciones posteriores gracias a mecanismos epigenéticos que son fundamentales. Estos establecen modificaciones en la cromatina y el DNA, ya sea a través de procesos de metilación, fosforilación, acetilación y otros, o mediante la interacción con RNA no codificantes que cambian los patrones de
Uno de los principios derivados de la especificación condicélulas madre, son aquellas que se dividen indefinidamente para generar más células madre semejantes y también otras células que se pueden especializar. Son primordiales para las poblaciones celulares que sobreviven períodos largos y que tienen que ser renovadas. A continuación se presentan los diferentes tipos de células madre:
. Son capaces de generar todas las estructuras de un embrión y sus anexos, como el cigoto y los blastómeros.
• Células madre pluripotenciales. Pueden diferenciarse en ectodermo, mesodermo y endodermo, es decir, en las células del embrión, pero no en las de los anexos. Estas células son las del embrioblasto del blastocisto.
• Células madre multipotenciales o comprometidas Son capaces de diferenciarse en una determinada población celular, como las células mesenquimatosas, que se diferencian en fibroblastos, adipocitos, condrocitos, osteocitos, miocitos, etcétera.
La restricción del potencial de diferenciación de las células madre es progresiva y está determinada por su
DIFERENCIACIÓN CELULAR
que determina la inicia con la es el
que produce que originan la
entorno. Una vez que las células están comprometidas, no cambian su destino final de diferenciación. Las células progenitoras o precursoras no son células madre, ya que sus divisiones no dan células progenitoras similares, sino células que continúan su diferenciación.
CAMBIOS EN LA FORMA CELULAR
El cambio en la forma de las células las capacita para su migración o para la formación de surcos, tubos, vesículas y otras estructuras.
La forma de las células es el resultado del equilibrio entre las fuerzas intrínsecas del citoplasma sobre la membrana celular y las fuerzas extrínsecas del medio extracelular. Las fuerzas intrínsecas se generan por la presión osmótica y la disposición del citoesqueleto, mientras que las fuerzas extrínsecas son consecuencia de las uniones entre las células o de las uniones de las células con la matriz extracelular.
Existen señales externas que pueden regular el citoesqueleto a través de una familia de proteínas llamadas Rho, Rac y Cdc42. La activación de las Rho GTPasas permite la activación de la polimerización de la actina, lo
que cambia la forma celular. Señales como las de las fuerzas de presión o cambios en la adhesividad a la matriz extracelular pueden activar Rho, de esta manera el citoesqueleto recibe señales extracelulares que regulan su forma celular.
En la morfogénesis se ha podido determinar que los cambios en la forma de las células están implicados en la migración de células de forma individual o en grupos. Para la migración celular individual, la célula cambia su forma y emite procesos o prolongaciones celulares como filopodios y lamelopodios, que le permiten avanzar por las matrices extracelulares. En la migración de células en grupo, las células permanecen unidas, y por los cambios en la forma de las células se desplazan o se forman surcos, vesículas o tubos.
La muerte celular programada contribuye a la eliminación de órganos y teji dos transitorios, a la remodelación de órganos o estructuras y al control del número de células en tejidos específicos.
Las dos formas de muerte celular programada que se observan en el desarrollo son la muerte celular progra mada por apoptosis y la muerte celular programada por autofagia (Figura 5-3
Apoptosis
En la apoptosis, también denominada gramada tipo I, los cambios morfológicos que se observan en las células son su disminución de tamaño, la conden sación del citoplasma, la condensación marginal de la cromatina, la integridad de los organelos que permanecen intactos (ya que no hay autólisis) y finalmente la frag mentación celular sin desintegración de la membrana. Los fragmentos celulares o cuerpos apoptóticos son eli minados por células fagocíticas. A nivel molecular, se ha informado que BMP, FGF y SHH participan activamente en la regulación de la apoptosis en el desarrollo del cerebro o las extremidades, mediante la activación de caspasas que son proteasas que a su vez activan enzimas que degradan el DNA, elementos del citoesqueleto y otros sustratos celulares que llevan a la célula a morir.
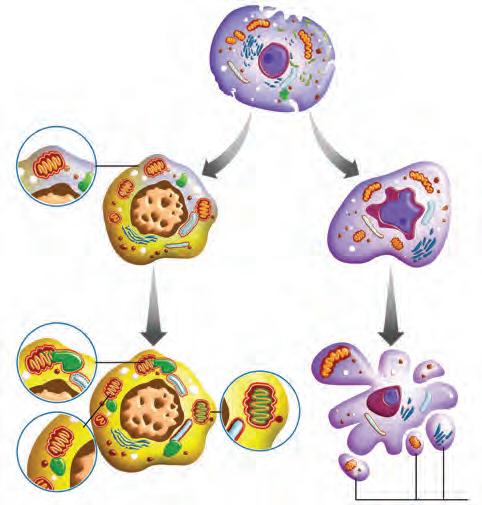
Tipos de muerte celular programada: autofarodeado por una membrana Fusión del autofagosoma con un lisoEl lisosoma vierte las enzimas hidrolíticas en el Degradación del contenido del orgánulo en el autofagosoma por las enzimas hidrolíticas. *** Cuer
Autofagia
En la autofagia, también denominada muerte celular programada tipo II, se forman autofagosomas, que son
estructuras membranosas que envuelven organelos. Es por esto que, morfológicamente, se observan en el citoplasma celular como vesículas de diferentes tamaños. Los autofagosomas se fusionan con los lisosomas y las enzimas lisosómicas degradan los organelos. La autofagia es importante para mantener a la célula saludable, dado que así se eliminan organelos dañados o envejecidos y permite la supervivencia de la célula en condiciones adversas tales como la privación de nutrientes, aunque cada vez hay más evidencia que sugiere que también puede mediar en la muerte celular durante el desarrollo y la patogénesis.
De la regulación molecular de la autofagia se piensa que la pérdida del estímulo por parte de factores de crecimiento o la disminución del transporte de nutrientes conduce a la activación del complejo serina/treonina proteína cinasa (ULK1) que conduce a activar múltiples señales que llevan a la formación de los autofagosomas. También se sabe que hay un grupo de genes llamados autophagy-related genes (ATG) que regulan este proceso, pero no se sabe cómo la autofagia puede llevar a la muerte de la célula. Se piensa que la activación de los lisosomas actúa en paralelo con las caspasas, llevando a la célula a su destrucción final.
Epiblasto Línea primitiva
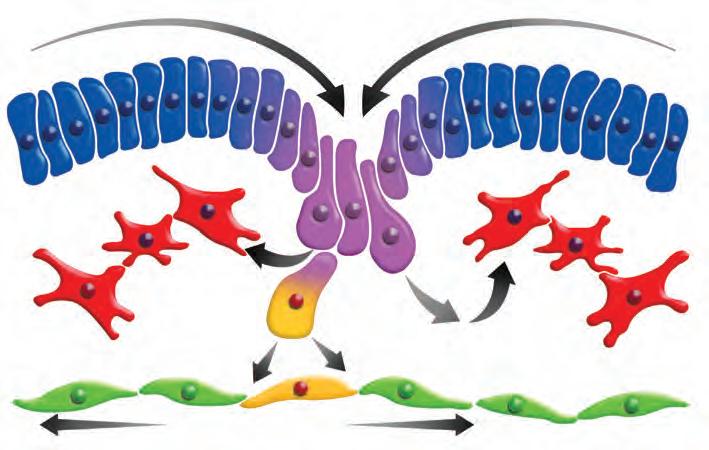
El movimiento de las células durante el desarrollo es fundamental para establecer la estructura del embrión. Durante el desarrollo embrionario se realizan dos tipos diferentes de movimiento celular: la migración celular colec tiva y la migración celular individual.
Durante la morfogénesis se llevan a cabo reorganizaciones celulares para producir tejidos, órganos y estructuras. En el desarrollo de los órganos se incorporan células que tienen que desplazarse desde su origen hasta su ubicación definitiva. Un ejemplo son las células germinales, aque llas que se diferencian en el ovocito o el espermatozoide, que migran desde el saco vitelino y se introducen en el embrión hasta alcanzar las gónadas que están en desarrollo. Los dos tipos de movimiento que se observan en el desarrollo son la migración de células unidas que forman grupos celulares y la migración de células de forma individual.
de este tipo de movimiento es el que se produce durante la gastrulación, en la que las células del epiblasto, que están unidas para formar un epitelio, se desplazan hacia la línea primitiva situada en la línea media del embrión. Las células del epiblasto se introducen entre este y el hipoblasto Figura 5-4).
Se ha mostrado que la expresión del factor de crecimiento en el epiblasto regula su desplazamiento, ya que , se altera la línea primitiva y hay una reducción en la formación del mesodermo. Para la migración en grupo, las células se polarizan debido a señales externas que determinan la dirección de la migración. Las señales activan receptores del FGF y el factor de crecimiento derivado de las plaquetas (PDGF, platelet) situados al frente de las células y en consecuencia emiten proyecciones transitorias como filopodios y lamelopodios que se unen a la matriz extracelular y así pueden desplazarse en conjunto, ya que se mantienen juntas mediante uniones celulares. Se ha comprobado que la molécula de adhesión celular cadherina E ejerce un papel importante en la migración colectiva durante la gastrulación y el cierre del tubo neural.
Migración celular colectiva
La migración celular colectiva consiste en el desplazamiento de células unidas que forman un tejido, el cual se mueve de manera coordinada, ya que estas células mantienen sus uniones celulares durante el movimiento. Un ejemplo
Migración celular individual
La migración celular individual requiere la pérdida de contactos intercelulares, ya que las células migran de forma independiente a través de la matriz extracelular. La
QUIMIOTAXIS
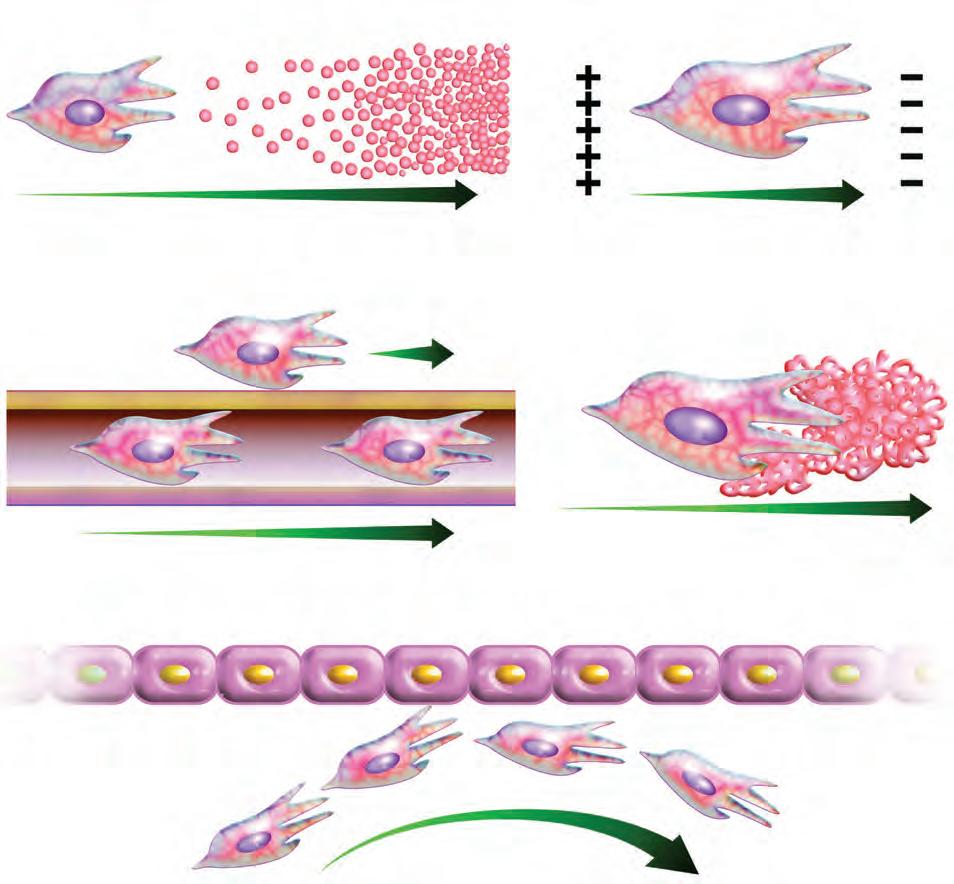
5-5. Diferentes mecanismos que regulan la dirección de la migración celular.
GALVANOTAXIS
son generar la señal, detectar la señal, transmitir la señal y ejecutar la señal.
Generación, detección y transmisión de la señal
A continuación, se describen algunos de los mecanismos para la generación, detección y transmisión de señales que usan las células en el proceso de migración (Figura 5-5).
• Quimiotaxis. Se refiere a la migración dirigida por un gradiente de señales difusibles o quimioatrayentes como péptidos, productos de la cascada del complemento, metabolitos de fosfolípidos, quimiocinas y factores de migración es un mecanismo conservado evolutivamente que participa en diferentes acontecimientos de la morfogénesis. Un ejemplo ilustrativo es la migración de las células de la cresta neural. Estas células se desprenden del tubo neural y siguen diferentes vías de migración, diferenciándose en distintas líneas y estructuras celulares. En un inicio, en las células, se activa la motilidad aleatoria a través de señales genéticas. Esta consiste en la formación de pequeñas proyecciones celulares de corta duración y en todas direcciones con el fin de dispersarse y explorar. Posteriormente, en respuesta a señales externas, migran en una trayectoria determinada hasta llegar a su destino definitivo. Para este tipo de migración se han descrito los “cuatro pilares de la migración dirigida” que
crecimiento de células endoteliales y estromales. Un ejemplo es la recepción de la señal a través de receptores transmembranales como los de tirosinacinasa (RTK, receptor tyrosine-kinase) que interactúa con el factor de crecimiento endotelial (EGF, endothelial growth factor) y el PDGF. La activación del receptor desencadena una cascada de señalización celular que conduce a cambios en el citoesqueleto, por lo que la célula se polariza para que pueda migrar.
• Hapoptasis. Es la migración a través de señales ligadas a la matriz extracelular, como fibronectina, laminina y diferentes tipos de colágeno, que hacen que la matriz sea un sustrato de migración estable y duradero mar cando así las rutas para la migración. Las señales se detectan mediante receptores en la membrana celular, como las integrinas, que al unirse a las señales en la matriz extracelular conducen a la activación de cina sas. Esto causa la reestructuración del citoesqueleto, lo que permite la formación de las proyecciones celulares que se fijan al sustrato para la migración de la célula.
• Durotaxis. Es la migración dirigida por la rigidez del sustrato y la preferencia de las células para migrar en estos sustratos o alejarse de ellos. Las enzimas lisil entrecruzan fibras de colágeno u otros componentes del sustrato y lo endurecen; por el contrario, enzimas como las metaloproteinasas de la matriz relajan o suavizan el sustrato. Los sensores o receptores son mecánicamente sensibles y están a ambos lados de la membrana celular o en el citoplasma de la célula acoplados mecánicamente al sustrato. El mecanismo por el que se transducen las señales se desconoce, pero se ha propuesto que enzimas y proteínas mecánicamente sensibles pueden generar segundos mensajeros, como cambios en el calcio intrace lular. Asimismo, se ha considerado que proteínas como la vinculina y la talina, que unen a las integrinas con los filamentos de actina, aumentan en respuesta a fuerzas mecánicas y activan la polimerización y la polarización de la actina.
• Topotaxis. Se refiere a la migración dirigida por señales biofísicas determinadas por las característi cas topográficas del sustrato que permiten y orientan la migración celular. Las señales pueden ser mediante fibras de colágeno alineadas, fibras musculares, fibras nerviosas, vasos o túneles formados en la matriz extracelular. Recientemente, se ha identificado la deformación local o curvatura de la membrana celular como un sensor que lleva a la reorganización del citoesqueleto celular para la migración. Se especula que las señales involucran el incremento del calcio que conduce a la polimerización de los filamentos de actina.
Anterior
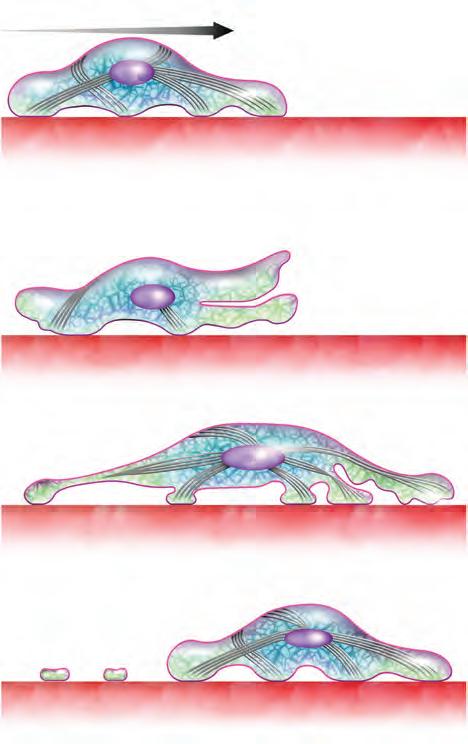
Haces contráctiles de actina y miosina
Formación de lamelopodios
En la migración celular, las señales conducen a una cascada molecular compleja que produce la reorganización interna de las células para que se polaricen y puedan desplazarse en la orientación correcta. La locomoción de las células sucede mediante un proceso cíclico que se repite y que incluye los siguientes pasos (Figura 5-6):
• Galvanotaxis. Es la migración dirigida por campos eléctricos que producen la despolarización y electromigración de proteínas de la membrana. Esto activa vías de señalización intracelular que contribuyen a la migración.
1. Polarización. Es un proceso complejo y regulado por el que, en la célula, en función de la dirección de la migración, se determina cuál va a ser su borde anterior, o frontal, y cuál va a ser su borde posterior. Para esto la célula reorganiza su citoplasma mediante el movimiento de organelos dirigidos por los microtúbulos y la reorganización de otros elementos del citoesqueleto como la actina y la miosina II, que son los motores fundamentales para el desplazamiento.
2. Protrusión. Es la formación de procesos o prolongaciones celulares al frente de la célula. Las prolongaciones pueden ser lamelopodios o filopodios. Se forman por la polimerización de los filamentos de actina que se extienden hacia el frente de la célula y que hacen que la membrana se protruya, formándose así las prolongaciones.
3. Adhesión. Una vez que se ha formado la prolongación celular, esta se adhiere al sustrato, matriz extracelular, para poder impulsarse. La unión se realiza a través de las integrinas, proteínas de la membrana celular, que a su vez están unidas en el citoplasma a la actina y en la matriz extracelular a distintas mo fibronectina o la laminina. La interacción de la actina con la miosina forma haces paralelos contráctiles que desempeñan un papel importante en la creación y el mantenimiento de las fuerzas de tracción que permi ten a la célula unirse e impulsarse sobre el sustrato.
4. Retracción del borde posterior. célula se mueve hacia adelante, el borde posterior debe perder su contacto con el sustrato y retraerse para que la célula pueda avanzar. La retracción se efectúa por la fuerza ejercida por la interacción de la actina con la miosina. Una vez que la célula se retrae, vuelve a emitir una nueva prolongación repitiendo los pasos del ciclo.
Movimientos morfogenéticos
Los movimientos morfogenéticos son los patrones típicos de desplazamiento de los grupos específicos de células que migran:
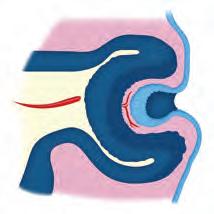
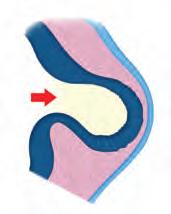
• Invaginación. Hundimiento de un grupo de células que penetran en otro tejido o en la luz de una estruc tura para formar el esbozo de un tejido. Por ejemplo: el movimiento que presenta el tejido de la lente (crista lino) a partir del ectodermo superficial hacia la cavidad de la copa óptica (
• Evaginación. Formación de una saliente a partir de un grupo de células que se proyectan hacia el exterior de una estructura u órgano. Por ejemplo: la formación de las vesículas ópticas (Figura 5-7B).
• Delaminación. Un grupo sólido de células se reacomoda para formar hojas o láminas separadas por un espacio o cavidad. Por ejemplo: la delaminación del mesodermo lateral (Figura 5-7E).
Síndrome de microdeleción 22q11.2
• Convergencia. Migración de las células para reunirse en un sitio determinado (Figura 5-7C).
• Divergencia. Migración de las células alejándose de un punto determinado para dispersarse o reunirse en otro ( Figura 5-7C ). Por ejemplo: los movimientos celulares durante la gastrulación.
• Cavitación. Formación de un espacio entre las células que en principio formaban estructuras sólidas. Por ejemplo: la formación de la cavidad amniótica primitiva (Figura 5-7D).
es causado por una pérdida de material genético en el brazo largo del cromosoma 22. Tiene una incidencia de 1 caso por cada 000 recién nacidos y no existen diferencias en cuanto a etnia o sexo. Engloba una serie de síndromes descritos previamente en los que se ha identificado esta etiología común, como los síndromes de DiGeorge, velocardiofacial o de Shprintzen, la anomalía facioconotruncal y el síndrome cardiofacial de Cayler. Las manifestaciones clínicas más frecuentes son malformaciones cardíacas, microcefalia y facies característica: pabellones auriculares pequeños, fisuras palpebrales antimongoloides y telecanto, disfunción velopalatina, con o sin paladar hendido, y labio hendido. También puede presentarse hipotiroidismo e
Se ha propuesto que las células de la cresta neural craneal desempeñan un papel crítico en la patogenia de este síndrome, ya que estas migran al corazón en desarrollo y a los arcos faríngeos de los que se van a formar las estructuras y órganos afectados. La alteración en la migración o pérdida de las células de la cresta neural, así como las anomalías en su función, son un factor importante en la patogenia de las dismorfias, puesto que las células de la cresta neural craneal son un componente fundamental para el desarrollo de la
La afinidad celular diferencial es el reconocimiento y la unión celular específica para formar tejidos, construir órganos, reconocer el destino final en la migración celular y coordinar el crecimiento, entre otras funciones.
La morfogénesis implica la interacción entre diferentes tejidos y, por lo tanto, la interacción entre distintas células. Cada tipo celular tiene en su membrana diferentes proteínas que son responsables de la unión específica de las células, lo cual determina la formación de estructuras, tejidos y órganos. Un ejemplo es un experimento que se realizó en el que se disgregaban el ectodermo, el
A Vesícula de la lente
Capa externa de la copa óptica intrarretiniano
Vesícula óptica
Convergencia
Divergencia
Figura 5-7. Movimientos morfogenéticos. Invaginación. En el desarrollo del ojo, la formación de la lente a partir del ectodermo superficial que se introduce en la concavidad de la copa óptica. Evaginación. También en el desarrollo del ojo, pero en etapa más temprana, la protrusión que realiza la vesícula óptica hacia el exterior. Convergencia y divergencia. Durante la gastrulación, dirección de la migración de las células en dirección medial o lateral. Cavitación. Durante el desarrollo de la blástula, la formación de la cavidad amniótica primitiva entre el epiblasto y el trofoblasto. Delaminación. Durante el proceso de segmentación del mesodermo, la separación que tiene el mesodermo lateral para formar las capas somática y esplácnica, y entre ellas el celoma intraembrionario.
mesodermo y el endodermo, que forman el embrión en la etapa de gastrulación, y se mezclaban sus células. Se observó poco tiempo después que se generaban, de forma espontánea, agregados celulares en los que las células del ectodermo recubrían la superficie externa, las del endodermo quedaba en el interior y entre ambos grupos celulares se dispusieron las del mesodermo, es decir con la misma relación que se observa en el disco embrionario. Esto significa que las células se reconocieron y se volvieron a agrupar de forma diferencial. Hay diferentes moléculas involucradas en los complejos de unión celular: Las principales son las cadherinas, que son proteínas transmembranales que se unen a otras cadherinas semejantes que están situadas en la membrana de otra célula, de manera que estas quedan
unidas. Dicha unión depende del calcio y tiene lugar entre cadherinas semejantes, por lo que es una unión homofílica: una célula que tenga en su membrana cadherina E se une a otra célula con cadherina E, y de esta forma se establece la unión específica y diferencial. Para que la unión sea resistente, las cadherinas se unen a las cateninas, situadas en el citoplasma, y las cateninas se unen a los filamentos de actina que forman parte del citoesqueleto de la célula.
Los principales tipos de cadherinas que se han identificado en el desarrollo de los mamíferos son los siguientes:
• Cadherina E. También se denomina cadherina epitelial o L-CAM y se expresa en todas las células de los embriones tempranos de mamíferos, incluso desde el estadio de cigoto. Posteriormente, queda restringida a las células epiteliales de los embriones y los adultos.
• Cadherina P. Se denomina cadherina placentaria, ya que se expresa en las células epiteliales de la superficie del útero y en el trofoblasto, por lo que se considera que facilita la unión del embrión al útero para su implantación y el desarrollo de la placenta.
• Cadherina N. Es la cadherina neural y se ha visto que en la gastrulación las células del epiblasto pier
den la cadherina E, que las mantiene unidas, y expresan la cadherina N cuando se transforman en mesodermo. Esta cadherina se expresa también en el sistema nervioso central en desarrollo.
• Cadherina EP. Es la cadherina C y es crítica para los movimientos celulares durante la gastrulación.
• Protocadherinas. Son moléculas de adhesión celular que dependen del calcio como las cadherinas; sin embargo, no no establecen uniones al citoesqueleto a través de las cateninas. Son muy importantes en la separación de la notocorda de otros componentes
RESUMEN
• Los procesos del desarrollo son el conjunto de mecanismos biológicos por los que, a partir de una célula, se forma un organismo multicelular con una morfología específica. Estos procesos interactúan dinámicamente durante la morfogénesis. Los principales procesos del desarrollo se resumen en los siguientes puntos.
• El crecimiento, que es el aumento del tamaño por el incremento en el número de células, en el tamaño de las células o en los componentes extracelulares.
• La diferenciación celular, que es la formación de diferentes tipos celulares por la expresión de genes específicos. Las células se diferencian a partir de células madre con diferente potencial de diferenciación.
• El cambio en la forma celular, el cual consiste en la transformación de la morfología celular que posibilita el desplazamiento de la célula o, junto con otras células, forma surcos, vesículas, tubos, etcétera, contribuyendo así a la morfogénesis.
• La muerte celular programada, que es la muerte celular regulada por factores internos o externos con la que se eliminan células para la morfogénesis o para mantener el número adecuado de células en los tejidos. Los dos tipos de muerte celular programada durante el desarrollo son la apoptosis y la autofagia.
• El movimiento celular, el cual permite la migración de células en grupo o de forma individual para la formación de tejidos, órganos o estructuras. Para la migración, las células emiten prolongaciones celulares y se desplazan guiadas por diferentes mecanismos que las llevan a su destino final.
• La afinidad celular diferencial, que consiste en la unión celular específica que permite que determinadas células se unan para formar tejidos, estructuras u órganos. El reconocimiento y la unión específica entre las células ocurre por proteínas situadas en la membrana celular.
AUTOEVALUACIÓN
ACTIVIDADES
EMBRIOFETOTECA VIRTUAL

LECTURAS