
Harnessing GLP-1 Through the Gut Microbiome Bundle
Everything you need to know about incorporating targeted probiotic strains to support natural GLP-1 production.


Everything you need to know about incorporating targeted probiotic strains to support natural GLP-1 production.
GLP-1 Probiotic is formulated with beneficial bacterial strains that naturally increase GLP-1*, a satiety hormone that helps maintain a healthy weight.
This dietary supplement naturally boosts GLP-1* production. It does not contain GLP-1 and is not a GLP-1 agonist drug. No prescription
91% reported reduced overall food cravings**
Enhances natural GLP-1 production*
Serving size: 1 capsule
Servings per container: 30
Amount per capsule
Probiotic blend
Clostridium butyricum WB-STR-0006
Akkermansia muciniphila WB-STR-0001
Bifidobacterium infantis
Chicory inulin
Helps maintain a healthy weight
Helps maintain a healthy weight
500 million AFU 211mg %DV † †
† Daily Value (DV) not established.
Other ingredients: hypromellose (vegetarian capsule), microcrystalline cellulose, silica
Directions: Take 1 daily with food

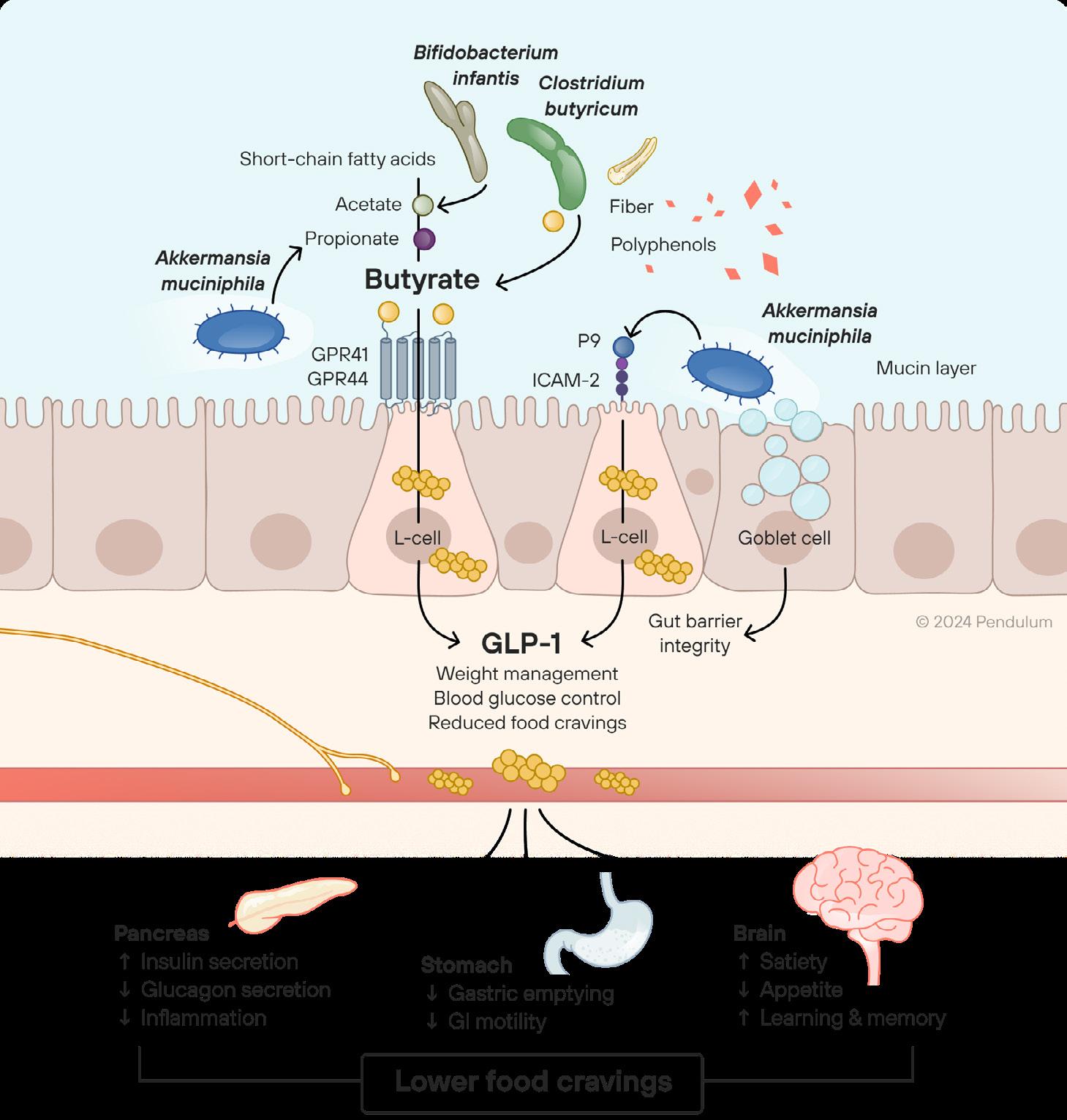
GLP-1 Probiotic is formulated with the beneficial bacterial strains that help produce more GLP-1.*
Secretes the postbiotics propionate and p9 which induce GLP-1 production.*
Produces the postbiotic butyrate which stimulates GLP-1.*
Produces the postbiotics acetate and lactate which feed bacteria such as Akkermansia and Butyricum, promoting an ecosystem in the gut that supports a healthy gut lining and naturally boosts GLP-1.*
GLP-1 Probiotic is formulated with the beneficial bacterial strains that help produce more GLP-1.*
As someone eats, metabolic processes are triggered in the body 1
In the gut microbiome (distal colon), the beneficial bacteria… 2
Metabolize fibers into butyrate, acetate, and propionate
Secrete key proteins such as p9
Stimulate mucin rejuvenation through consumption of “old” and rebuilding new mucin
These signals reduce appetite, helping to maintain a healthy weight
Butyrate and p9 stimulate GLP-1, the hormone that tells the pancreas to produce insulin, slowing down the emptying of the stomach and signaling to the brain that the person is full 3

GLP-1 Probiotic is a multi-strain formula that naturally increases GLP-1*, an “un-hunger” hormone that sends a message to your brain that you’re full. Your body already makes this hormone, but now you’ve got the power to boost it* to help curb your cravings and appetite.
**Based on a consumer survey of 274 people Enhances natural
*Based on preclinical studies
Follow these 5 tips to nourish your gut
Take GLP-1 Probiotic once a day with food
Increase your intake of fiber-filled foods including whole grains, nuts, seeds, fruits, and vegetables
Build healthy eating habits by balancing food groups (carbohydrates, proteins and fats), limiting intake of added sugars, and eating fewer simple carbohydrates such as bread and pasta
Boost your microbiome with polyphenols, antioxidants, fermented foods, and prebiotics
Prioritize movement—research shows that 20 minutes of moderate exercise a day reduces your risk of metabolic disease

GLP-1 Probiotic is formulated with bacterial strains that boost this process naturally.*
As you eat, metabolic processes are triggered in your body
In the gut microbiome, the beneficial bacteria–such as Akkermansia and Butyricum–convert the fiber in your food into butyrate, and Akkermansia secretes a protein called p9
Butyrate and p9 stimulate GLP-1, the hormone that tells your pancreas to produce insulin, slowing down the emptying of your stomach
This sends a signal to your brain that you’re full, reducing your appetite, helping to maintain a healthy weight
GLP-1 Probiotic is formulated with the beneficial bacterial strains that help you produce more GLP-1.*
Akkermansia muciniphila
Secretes the postbiotics propionate and P9 which induce GLP-1 production.
Clostridium butyricum
Produces the postbiotic butyrate which stimulates GLP-1.
Bifidobacterium infantis
Produces the postbiotics acetate and lactate which feed bacteria such as Akkermansia and Butyricum, promoting an ecosystem in the gut that supports a healthy gut lining and naturally boosts GLP-1.

Developed in collaboration with Dr. Kara Fitzgerald, ND, IFMCP
Disclaimer: The Pendulum team has created these protocols in collaboration with practitioners to help healthcare partners make decisions when building treatment plans When using this protocol, you understand and accept that the recommendations in the protocol are for educational guidance only This protocol is not a substitute for medical advice, and is not intended to diagnose, treat or prevent any medical disease Please consult with your qualified medical professional to determine if this protocol is appropriate for you
Description
Understanding the physiological mechanisms of metabolic health and obesity is essential for developing new and targeted approaches for clinical management This area of study concerns the connection between metabolic function and gut health especially the gut microbiota, the collection of bacteria and other microorganisms inhabiting the digestive tract, with the greatest concentration in the colon. Cross-sectional population studies profiling the gut microbiome have repeatedly demonstrated decreased diversity in individuals exhibiting obesity and/or type 2 diabetes 1 With growing evidence suggesting that gut microbiota contribute to many aspects of metabolism, the field is moving closer to specific gut-focused solutions for managing metabolic health and body weight 2
When fermentable carbohydrates are digested by certain bacteria in the gut, the bacteria release metabolites (e.g. short-chain fatty acids, or SCFAs) that trigger the secretion of GLP-1 and PYY, helping regulate energy balance and glucose homeostasis 3 Butyrate is a key SCFA that is mechanistically linked with metabolic benefits in numerous preclinical studies; bacterial strains such as Clostridium butyricum and Faecalibacterium prausnitzii, which possess genes to specifically increase butyrate production, can be of use to trigger host secretion of GLP-1 and PYY Additionally, Akkermansia muciniphila, a keystone bacterial strain, secretes a P9 protein which stimulates GLP-1 production.4
Evidence indicates that lifestyle interventions (i e diet and exercise) for weight loss and metabolic health may depend on the baseline gut microbiota. A real-world challenge is that the gut microbiomes of individuals in developed countries tend to be depleted through many factors such as frequent antibiotic use, a Western diet, physical inactivity, and chronic stress A promising clinical approach is thus to normalize an individual’s gut microbiota so it is more like that found in healthy subjects Precise approaches can be envisioned: namely, using precise probiotic strains with relevant mechanisms of action to target metabolic health improvements Replenishment of specific microbes repeatedly shown to be diminished in patients with metabolic disorders, e.g., Akkermansia muciniphila and butyrate producers have the potential to support and amplify the beneficial effects of lifestyle interventions Demonstratively, in a preclinical study, administration of a three-strain probiotic formulation containing A muciniphila, C butyricum, and B infantis, 91% of participants demonstrated reduced overall food cravings.5
● Supplement with Pendulum’s GLP-1 Probiotic: 1 capsule, once daily with food
● Encourage intake of polyphenols such as pomegranate, green tea, and grape seed6
○ Supplement as necessary with Pendulum’s Polyphenol Booster to boost Akkermansia which may further enhance the benefits
● Encourage intake of prebiotic dietary fibers such as onion, chicory, garlic, asparagus, banana, and artichokes7
● Emphasize a dietary pattern with sufficient protein and fat to trigger satiety
● Consider supplementation with DHA/EPA to promote Akkermansia abundance8
● Consider a classic 5R protocol
1. Huda MN, Kim M, Bennett BJ. Modulating the Microbiota as a Therapeutic Intervention for Type 2 Diabetes. Front Endocrinol (Lausanne). 2021 Apr7;12:632335. doi: 10.3389/fendo.2021.632335. PMID: 33897618; PMCID: PMC8060771.
2. Fan Y, Pedersen O Gut microbiota in human metabolic health and disease. Nat Rev Microbiol. 2021 Jan;19(1):55-71. doi:10.1038/s41579-020-0433-9. Epub 2020 Sep 4.PMID: 32887946.
3. Everard A, Cani PD Gut microbiota and GLP-1. Rev Endocr Metab Disord. 2014 Sep;15(3):189-96. doi: 10.1007/s11154-014-9288-6. PMID: 24789701.
4. Yoon, H.S., Cho, C.H., Yun, M.S. et al. Akkermansia muciniphila secretes a glucagon-like peptide-1-inducing protein that improves glucose homeostasis and ameliorates metabolic disease in mice. Nat Microbiol 6, 563–573 (2021). https://doi.org/10.1038/s41564-021-00880-5
5. Based on a consumer study of 274 people
6. Anhê FF, Pilon G, Roy D, Desjardins Y, Levy E, Marette A. TriggeringAkkermansiawith dietary polyphenols: A new weapon to combat the metabolic syndrome? Gut Microbes. 2016;7(2):146-153. doi:10.1080/19490976.2016.1142036
7. Zhou K. Strategies to promote abundance of Akkermansia muciniphila, an emerging probiotics in the gut, evidence from dietary intervention studies. Journal of Functional Foods. 2017;33:194-201. doi:10.1016/j.jff.2017.03.045
8. Costantini L, Molinari R, Farinon B, Merendino N. Impact of omega-3 fatty acids on the gut microbiota. International Journal of Molecular Sciences. 2017;18(12):2645. doi:10.3390/ijms18122645
Orville Kolterman, MD
Adam Perlman, MD, MPH, FACP
Only 12% of Americans are metabolically healthy.1 This represents a significant public health concern, given the increased risk of various diseases that comes with poor metabolic health, as well as increased mortality. Metabolic diseases include type 2 diabetes, obesity, atherosclerotic heart disease, stroke, liver, and kidney disease in addition to a number of cancers such as breast and colon cancer. This represents an opportunity for the entire healthcare community to work together to improve the population’s metabolic health.
Glucagon-like peptide 1 (GLP-1) is a well-studied hormone produced by L-cells within the mucosa of the human gut, performing several functions critical to human metabolic health. It augments insulin secretion in response to orally ingested nutrients while enhancing satiety, leading to decreased energy intake.2 GLP-1 function is altered in type 2 diabetes (T2D), obesity, and other metabolic conditions due primarily to decreased secretion.3 In some instances, GLP-1 action is reduced compared with healthy individuals (hormone resistance). In those instances, higher GLP-1 levels are required to achieve an equivalent insulinotropic response and decrease blood glucose levels.4
GLP-1 has long been of interest for therapeutic use. However, for many years, its potential use as a drug was hindered because it is rapidly degraded in the body. Once formulations were developed that overcame the challenge of rapid degradation, GLP-1 was successfully leveraged to improve metabolic health through improved glucose control and weight loss.5 However, these drug therapies have a prolonged circulating half-life, contributing to plasma concentrations of the hormone/agonists that are much higher than would occur naturally in the body, and side effects that are troublesome to many patients. Recent research has demonstrated the complex role of the gut microbial ecosystem in inducing the release of GLP-1 in response to a meal.6 In the gut, specific bacterial taxa, notably Akkermansia muciniphila and Clostridium butyricum, participate in the physiological stimulation of GLP-1 production in response to the presence of nutrients within the gut lumen. Assuring adequate levels of these key species supports the natural release of GLP-1 and enhances normal concentration and functions in the human body, resulting in a more natural regulation of metabolic health.
GLP-1 is a 30 amino acid peptide hormone that is produced in response to nutrient ingestion, through cleavage of the prohormone proglucagon within the enteroendocrine L-cells located within the epithelial lining of the intestine – predominantly in the colon and terminal ileum. It is a member of the incretin hormone family known to augment β-cell glucose-dependent insulin secretion in response to nutrient ingestion. Since this stimulation of insulin secretion is glucose-dependent, insulin levels decrease in parallel with the decline in the prevailing plasma glucose concentration. This feature blunts the likelihood of inducing hypoglycemia and its attendant risks, an attribute that has significant clinical benefit.7
While researchers initially demonstrated that GLP-1 stimulated glucose-dependent insulin secretion following nutrient ingestion, GLP-1 has proved to be a multifaceted hormone with effects throughout the body, impacting metabolic control and numerous other functions.8 In addition to stimulating postprandial insulin secretion, GLP-1 suppresses the secretion of glucagon, the pancreatic hormone that opposes the effects of insulin to guard against hypoglycemia by increasing the release of glucose by the liver. GLP-1 also reduces the rate at which nutrients pass through the stomach (gastric emptying), thus contributing to sustained satiety and restraining the absorption rate of ingested nutrients into the peripheral circulation.9
Importantly, GLP-1 also exerts effects within the central nervous system to promote satiety.10 These appetite modulating effects are mediated in part by afferent signaling via the vagus nerve triggered by GLP-1 bearing neurons within the gut epithelium combined with the systemic effects of GLP-1. Circulating GLP-1 hormone crosses the blood brain barrier through fenestrations near the hindbrain region of the area postrema where interactions with neuronal GLP-1 receptors within the brain activate well established pathways that are involved in appetite control. GLP-1 produced within the brain also contributes to overall satiety.
As shown in Figure 1, GLP-1 has multiple other beneficial effects.
Figure 1. Production and functions of glucagon-like peptide 1 (GLP-1) inside the digestive tract. The hormone is produced by the L-cells in the epithelial layer and exerts multiple effects throughout the body.
The observation in the mid to late 1990s that GLP-1 lowered both fasting and postprandial glucose concentrations in rodents and humans11-12 led to immediate interest in harnessing this phenomenon to improve management of hyperglycemia in patients with T2D9. This interest intensified with subsequent demonstrations that the hormone also decreased glucagon following meals, restrained gastric emptying, and increased satiety. In contrast with the hypoglycemic risk that accompanies exogenous insulin and sulfonylurea therapy, GLP-1 achieved these benefits
without increasing the risk for hypoglycemia since the stimulation of insulin secretion is glucose-dependent with the stimulatory effect dissipating in parallel with the subsequent fall of plasma glucose concentrations towards normal. This attribute of GLP-1 allows the reduction of plasma glucose concentrations to plateau within the normal range without the occurrence of clinically meaningful hypoglycemia. The safety margin is further enhanced by the suppression of glucagon secretion and restraint of gastric emptying also receding as plasma glucose concentrations enter the normal range, allowing endogenous glucagon secretion and orally administered glucose to reverse hypoglycemia should it occur.13
Glucose-dependent stimulation of insulin secretion, coupled with suppression of glucagon and restraint of the rate of nutrient absorption, provides excellent control of the rise in plasma glucose concentrations following meals.5 These attributes of GLP-1, as well as the ability to lower fasting glucose concentrations with a low risk of hypoglycemia, were first demonstrated through intravenous infusion of the native peptide, showing the potential of a GLP-1 based therapy for the management of T2D.11-12
However, naturally-produced GLP-1 has a circulating half-life that is measured in minutes due to its susceptibility to enzymatic degradation by dipeptidyl peptidase IV (DPP-IV), which is ubiquitous throughout the body. Thus, molecular entities with longer half-lives which retained the conformational requirements for binding to the GLP-1 receptor were necessary. This was achieved through two routes. First, exendin 4, a naturally occurring peptide in the salivary secretions of the gila monster lizard (Heloderma suspectum), was found to be naturally resistant to degradation by DPP-IV with a circulating half-life that delivers therapeutic efficacy with twice daily injections. Synthetic exendin 4, known as exenatide, was developed and received FDA approval as the first GLP-1 receptor agonist in 2005.14 The second approach consisted of conjugating a 16-carbon fatty acid moiety to native GLP-1 to foster non-covalent binding to circulating plasma proteins, primarily albumin, creating a “slow-release depot” with a pharmacokinetic profile that supports once-daily administration by injection.9 Liraglutide was the first FDA approved drug employing this approach, with several additional products following shortly thereafter that utilized a similar approach. Further pharmaceutical development leveraging newer approaches to injectable drug delivery resulted in products that provided the desired efficacy with once-weekly administration. More recently, an oral formulation of semaglutide has entered the market, but its uptake has been restrained by the requirement for more complicated dosing algorithms.
The benefits of such drugs are now shown to go well beyond T2D therapeutic efficacy. GLP-1 agonism is cardioprotective, as demonstrated by a reduction in major adverse cardiovascular events (MACE) in patients with T2D during long-term exposure to various GLP-1 receptor agonists.15 Emerging evidence indicates that GLP-1 receptor agonist medications also reduce the risk for heart attacks and stroke in individuals without diabetes and have benefits for congestive heart failure, renal disease, neurodegenerative disease, as well as addictive behaviors. The therapeutic attractiveness of GLP-1 receptor agonists is further augmented by the limited risk for hypoglycemia.14
Subsequent to use for T2D, GLP-1 receptor agonists have shown distinct efficacy for weight loss in obesity.16 As noted above, GLP-1 analogues stand apart from most other antidiabetic agents in that they do not drive weight gain. In fact, their therapeutic use is usually associated with weight loss. This observation led to interest in the potential for these agents to be used in weight control programs. Exploration of higher doses of liraglutide and semaglutide
for this purpose produced impressive results (>15% reduction in body weight in subjects with BMI>30 kg/m2) with subsequent approval of those formulations for use in the management of obesity.17 Tirzepatide, which combines a GLP-1 and GIP (glucose dependent insulinotropic polypeptide) agonists in a single molecule, has demonstrated robust weight loss (>20% reduction from baseline weight in obese subjects without diabetes).18 This compound was granted Fast Track Designation by the FDA for use in obesity and approved in late 2023.19 Driven by these pharmacological agents, analysts predict obesity drug sales could reach $100 billion by the year 2030.20
All approved GLP-1 agonists have so far demonstrated a good short-term safety profile even though they produce circulating levels of GLP-1 that are approximately ten times greater than the physiological GLP-1 concentrations seen in healthy subjects. Side effects with these agents are primarily related to gastrointestinal tolerability, with nausea and/or vomiting occurring in over 25% of subjects in most studies.21 Most afflicted individuals find that these side effects dissipate or cease during the initial month of therapy. After considering the benefits and risks of these agents, only ~5% of patients elect to discontinue therapy. However, some healthcare professionals are concerned about the growing popularity of these drugs, since the mechanisms by which they achieve substantial weight loss are not completely understood. The long-term effects of high circulating levels of GLP-1 are still unknown, and significant longer-term side effects have emerged in the scientific literature. According to a systematic review and meta-analysis of 76 randomized clinical trials, use of GLP-1 receptor agonist drugs was linked to an increased risk of gallbladder and biliary tract diseases. This association was stronger with higher doses and longer durations of the medication use, and was particularly strong when the drugs were used for weight loss (although the incidence of these diseases are already higher in those with obesity and in those who experience rapid weight loss).22 Moreover, the American Society of Anesthesiologists have raised concerns about case reports that GLP-1 agonists can increase the risk of regurgitation and pulmonary aspiration of stomach contents during anesthesia, which carries an increased risk of death.23
Given these potential risks, approaches to increased GLP-1 agonism that do not result in high circulating concentrations of GLP-1 are worth investigating. Endogenous GLP-1 stimulation may be a significantly safer way to benefit from the metabolic effects of GLP-1.
Cross-sectional population studies profiling the gut microbiome have repeatedly demonstrated decreased diversity in individuals exhibiting obesity and/or T2D.24 When fecal transplants from these subjects to germ free mice are performed, a similar microbiome is established in the rodent recipients, along with features of the human donor’s phenotype, i.e., development of obesity25 and/or diabetes. Salient characteristics of these microbiomes include a marked decrease or absence of Akkermansia muciniphila as well as butyrate-producing strains, compared to healthy subjects. Subsequent reversal of those changes in the rodent models’ microbiomes attenuate or reverse the metabolic abnormalities. Similar short-term improvements in metabolic parameters have been observed in human subjects with T2D following both single and multiple fecal transplants.26
The fecal transplant data demonstrates that constituents of fecal material from healthy subjects contain elements capable of improving metabolic control. Characterization of the gut microbiome profiles of obese subjects with or without T2D have repeatedly shown decreased diversity with significant decreases in both Akkermansia muciniphila and strains known to produce short-chain fatty acids (SCFAs) by the fermentation of dietary fiber. Akkermansia muciniphila stimulates GLP-1 secretion by enteroendocrine cells via three distinct mechanisms: the release of a
protein referred to as P9, a component of its outer wall identified as Amuc_1100, and the release of extracellular vesicles.27 With regard to P9, researchers found that mice treated with P9 alone recapitulated the reduced weight gain seen with Akkermansia muciniphila administration, and a one-time injection of P9 triggered an increase in GLP-1 secretion.28 Independent Pendulum investigations show that P9 stimulates GLP-1 expression and also triggers an increase in the concentration of GLP-1 in cultured human L-cells (Figure 2, internal data). This dose dependent GLP-1 secretion was observed in an established human L-cell line, when exposed to media containing the isolated P9 protein alone. As shown in Figure 2, the bacterial product strongly stimulates an increase in GLP-1 concentrations.
Interestingly, Akkermansia muciniphila that has been pasteurized prior to administration retains the ability to stimulate GLP-1 secretion to some degree.29 However, the administration of killed bacteria limits the stimulatory capacity to those present in the administered product whereas, in theory, administration of live bacteria allows for amplification of the stimulus via continuous production within the gut.
Figure 2. Concentrations of GLP-1 secreted by L-cells in intestinal cell culture, in control media and with the addition of P9 protein, purified from Akkermansia muciniphila sequence.30 (Pendulum Internal Data, 2024)
In addition, multiple studies have demonstrated the ability of butyrate producing gut microbes to increase GLP-1 secretion by L-cells in rodent models.31 Bacteria from another species, Clostridium butyricum, are adept at producing the SCFAs acetate and butyrate32, thereby stimulating GLP-1 secretion by the intestinal L-cells.28,33 Bacterial supernatant that contains these fatty acids, compared to media alone, induced significantly more GLP-1 expression in intestinal cell culture. See Figure 3, which demonstrates a dose-response effect, where higher concentrations of Clostridium butyricum supernatant lead to higher GLP-1 secretion.30 Clostridium butyricum has the ability to also synthesize ursodeoxycholate (UDCA) from the primary bile acid chenodeoxycholate during butyrogenic growth.35 UDCA has been shown to directly increase GLP-1 levels in healthy subjects36 and improve metabolic state parameters in both preclinical and clinical studies.37,38
Figure 3. Concentrations of GLP-1 secreted by human intestinal L-cells cultured in the presence of bacterial culture supernatant. Bacterial supernatant was diluted in Krebs buffer + 50mM glucose and used to treat L-cells at the following concentrations: 50%, 25%, and 12.5%. The Krebs buffer is present as a negative control, and PMA (Phorbol 12-myristate 13-acetate), a known inducer of direct GLP-1 secretion, is used as a positive control.30 (Pendulum Internal Data, 2024)
These observations raised the possibility that a targeted probiotic containing Akkermansia muciniphila along with UDCA and butyrate-producing specialist bacteria such as Clostridium butyricum could help improve metabolic control via increased GLP-1 activity. A recently published human study confirmed this by demonstrating that administration of a five-strain probiotic containing Akkermansia muciniphila and three butyrate-producing strains (including Clostridium butyricum) for twelve weeks led to significant improvement in metabolic control (reduction in plasma glucose concentrations following ingestion of a mixed meal accompanied by reduction in A1c, the gold standard for long-term glucose control) in subjects with T2D treated with metformin.34 These improvements in metabolic control were accompanied by increased fecal and plasma butyrate concentrations.35 Investigators also saw an increase in plasma levels of the secondary bile acid ursodeoxycholate, which has also shown the ability to stimulate GLP-1 secretion from L-cells.36
Together, these data suggest a natural way to harness GLP-1 in the digestive tract: using targeted probiotic formulations in the gut to stimulate physiologic GLP-1 production. Such an approach would avoid the high circulating levels of GLP-1 induced by current drug therapies and potentially reduce side effects, mimicking more closely GLP1’s normal concentration and functions in the human body.
Emerging evidence in humans supports the idea that individuals without metabolic disease may derive benefits from ingesting bacterial strains known to stimulate GLP-1 naturally within the gut. In a consumer survey that included 274 individuals from the general population, the participants ingested Pendulum’s GLP-1 Probiotic product containing 500 million Active Fluorescent Units (AFU) of probiotic bacteria, consisting of:
Clostridium butyricum WB-STR-0006
Akkermansia muciniphila WB-STR-0001
Bifidobacterium infantis Bi-26TM
The product also contains 211 mg of chicory inulin. After consuming the product daily for six weeks, participants showed reduced scores (p < 0.001)39 with the Food-Craving Inventory II, a validated questionnaire40, with 91% of participants experiencing a reduction in their total food cravings39. See Figure 4.
Figure 4. Total food cravings were reduced in 91% of subjects after six weeks of GLP-1 Probiotic use 39 (Pendulum Internal Data, 2024)
Individuals who scored higher on total cravings at baseline had a greater reduction in scores at six weeks (p < 0.001)39
See Figure 5.
Figure 5. Participant food craving scores at baseline and six weeks, grouped by baseline scores. Individuals with higher total cravings at baseline showed a greater reduction in their cravings at six weeks 39 (Pendulum Internal Data, 2024)
When the specific food subcategories were analyzed separately, mean food craving scores were significantly reduced from baseline to six weeks in all four subcategories (p < 0.001)39. See Figure 6. The percentage of individuals showing improved food cravings for each food subcategory is as follows:
Sweets – 88% of individuals
Carbs/starches – 87% of individuals
Fast Food Fats – 85% of individuals
High Fats – 82% of individuals
These observations are consistent with the concept that a general GLP-1 related mechanism (affecting overall appetite control) is involved.
Carb/Starch, Fast Food Fats, High Fats, Sweets
In conclusion, this consumer survey provides evidence that a probiotic targeting GLP-1 production decreases food cravings and has the potential to moderate food intake in the general population.39
GLP-1 is shown to play an important role in maintaining metabolic homeostasis. Its therapeutic utility was only realized after the development of approaches to prevent its rapid degradation by the ubiquitous DPP-IV enzyme. Once that challenge was met, the expectations for leveraging its activity to improve glucose control and induce weight loss were realized. Increasing knowledge of the gut microbiome, however, has revealed that Akkermansia muciniphila and microbial strains that produce butyrate (for example, Clostridium butyricum) can augment GLP1 production through natural stimulation of the L-cells. This path to increased GLP-1 production introduces the hormone into the body via its normal pathway, allowing its effects as a hormone, its communications with the intestinal immune system, and its interactions with both the peripheral and central nervous systems to be exerted via the natural physiological processes. This approach may be an effective alternative solution in allowing moderate benefits of GLP-1 to be realized without the need for the exaggerated circulating concentration required to achieve the efficacy of the synthetic analogs – and perhaps avoiding the negative side effects of these GLP-1 drugs. Given the complexity of maintaining metabolic homeostasis in the body and the important contributions of GLP-1 to overall metabolic health, efforts to approximate the hormone’s natural functioning in the human gut are highly worthwhile.
Orville Kolterman, MD is Emeritus Chief Medical Officer at Pendulum Therapeutics and Adjunct Clinical Professor of Medicine at the University of California, San Diego. Dr. Kolterman is a past Chief Medical Officer of Amylin Pharmaceuticals and led the team that developed the first FDA approved and marketed GLP-1 receptor agonist for the treatment of type 2 diabetes. He is a past President of the California Affiliate of the ADA and currently serves as member of the Board of Directors for the diaTribe Foundation. He received his M.D. from Stanford University and also completed his fellowship in Endocrinology and Metabolism at Stanford University School of Medicine.
Adam Perlman, MD, MPH, FACP is Chief Medical Officer at Pendulum Therapeutics. Dr Perlman is board-certified in Internal Medicine and specializes in both integrative and functional medicine. With more than 25 years of clinical, entrepreneurial, and leadership experience, he most recently served as the Director of Integrative Health and Wellbeing at the Mayo Clinic. Prior to that, he served as the Associate Vice President for Health and Wellness for the Duke University Health System. He received his BA from Tufts University, his MD from the Boston University School of Medicine, and his MPH from the Boston University School of Public Health.
1. Araújo J, Jianwen C, June S. Prevalence of Optimal Metabolic Health in American Adults: National Health and Nutrition Examination Survey 2009–2016. Metabolic Syndrome and Related Disorders, 2018; DOI: 10.1089/met.2018.0105
2. Shah M, Vella A. Effects of GLP-1 on appetite and weight. Rev Endocr Metab Disord. 2014 Sep;15(3):181-7. doi: 10.1007/s11154-014-9289-5.
3. Boer GA, Holst JJ. Incretin Hormones and Type 2 Diabetes-Mechanistic Insights and Therapeutic Approaches. Biology (Basel). 2020 Dec 16;9(12):473. doi: 10.3390/biology9120473.
4. Kjems LL, Holst JJ, Vølund A, Madsbad S. The influence of GLP-1 on glucose-stimulated insulin secretion: effects on beta-cell sensitivity in type 2 and nondiabetic subjects. Diabetes. 2003 Feb;52(2):380-6. doi: 10.2337/diabetes.52.2.380. PMID: 12540611.
5. Drucker DJ. Mechanisms of Action and Therapeutic Application of Glucagon-like Peptide-1. Cell Metab. 2018 Apr 3;27(4):740-756. doi: 10.1016/j. cmet.2018.03.001.
6. Abdalqadir N, Adeli K. GLP-1 and GLP-2 Orchestrate Intestine Integrity, Gut Microbiota, and Immune System Crosstalk. Microorganisms. 2022 Oct 19;10(10):2061. Doi: 10.3390/microorganisms10102061
7. Holst JJ, Gasbjerg LS, Rosenkilde MM. The Role of Incretins on Insulin Function and Glucose Homeostasis. Endocrinology. 2021 Jul 1;162(7):bqab065. doi: 10.1210/endocr/bqab065.
8. Drucker DJ, Holst JJ. The expanding incretin universe: from basic biology to clinical translation. Diabetologia. 2023 Oct;66(10):1765-1779. doi: 10.1007/s00125023-05906-7. Epub 2023 Mar 28.
9. Drucker DJ, Habener JF, Holst JJ. Discovery, characterization, and clinical development of the glucagon-like peptides. J Clin Invest. 2017 Dec 1;127(12):42174227. doi: 10.1172/JCI97233. Epub 2017 Dec 1.
10. Richards P, Parker HE, Adriaenssens AE, Hodgson JM, Cork SC, Trapp S, Gribble FM, Reimann F. Identification and characterization of GLP-1 receptor-
expressing cells using a new transgenic mouse model. Diabetes. 2014 Apr;63(4):1224-33. doi: 10.2337/db13-1440. Epub 2013 Dec 2.
11. Nauck MA, Kleine N, Orskov C, Holst JJ, Willms B, Creutzfeldt W. Normalization of fasting hyperglycaemia by exogenous glucagon-like peptide 1 (7-36 amide) in type 2 (non-insulin-dependent) diabetic patients. Diabetologia. 1993 Aug;36(8):741-4. doi: 10.1007/BF00401145.
12. Nauck MA, Sauerwald A, Ritzel R, Holst JJ, Schmiegel W. Influence of glucagon-like peptide 1 on fasting glycemia in type 2 diabetic patients treated with insulin after sulfonylurea secondary failure. Diabetes Care. 1998 Nov;21(11):1925-31. doi: 10.2337/diacare.21.11.1925.
13. Degn KB, Brock B, Juhl CB, Djurhuus CB, Grubert J, Kim D, Han J, Taylor K, Fineman M, Schmitz O. Effect of intravenous infusion of exenatide (synthetic exendin-4) on glucose-dependent insulin secretion and counterregulation during hypoglycemia. Diabetes. 2004 Sep;53(9):2397-403. doi:10.2337/ diabetes.53.9.2397.
14. McBrayer DN, Tal-Gan Y. Recent Advances in GLP-1 Receptor Agonists for Use in Diabetes Mellitus. Drug Dev Res. 2017 Sep;78(6):292-299. doi: 10.1002/ ddr.21404. Epub 2017 Aug 8.
15. Kristensen SL, Rørth R, Jhund PS, Docherty KF, Sattar N, Preiss D, Køber L, Petrie MC, McMurray JJV. Cardiovascular, mortality, and kidney outcomes with GLP-1 receptor agonists in patients with type 2 diabetes: a systematic review and meta-analysis of cardiovascular outcome trials. Lancet Diabetes Endocrinol. 2019 Oct;7(10):776-785. doi: 10.1016/S2213-8587(19)30249-9. Epub 2019 Aug 14. Erratum in: Lancet Diabetes Endocrinol. 2020 Mar;8(3):e2.
16. Wang JY, Wang QW, Yang XY, Yang W, Li DR, Jin JY, Zhang HC, Zhang XF. GLP-1 receptor agonists for the treatment of obesity: Role as a promising approach. Front Endocrinol (Lausanne). 2023 Feb 1;14:1085799. doi: 10.3389/fendo.2023.1085799.
17. Moore PW, Malone K, VanValkenburg D, Rando LL, Williams BC, Matejowsky HG, Ahmadzadeh S, Shekoohi S, Cornett EM, Kaye AD. GLP-1 Agonists for Weight Loss: Pharmacology and Clinical Implications. Adv Ther. 2023 Mar;40(3):723-742. doi: 10.1007/s12325-022-02394-w. Epub 2022 Dec 24.
18. Jastreboff AM, Aronne LJ, Ahmad NN, Wharton S, Connery L, Alves B, Kiyosue A, Zhang S, Liu B, Bunck MC, Stefanski A; SURMOUNT-1 Investigators. Tirzepatide Once Weekly for the Treatment of Obesity. N Engl J Med. 2022 Jul 21;387(3):205-216. doi: 10.1056/NEJMoa2206038. Epub 2022 Jun 4.
19. FDA, 2023. FDA Approves New Medication for Chronic Weight Management. Available online: https://www.fda.gov/news-events/press-announcements/ fda-approves-new-medication-chronic-weight-management
20. Goldman Sachs Research, 2023. Why the anti-obesity drug market could grow to $100 billion by 2030. Available online: https://www.goldmansachs.com/ intelligence/pages/anti-obesity-drug-market.html
21. Tran KL, Park YI, Pandya S, Muliyil NJ, Jensen BD, Huynh K, Nguyen QT. Overview of Glucagon-Like Peptide-1 Receptor Agonists for the Treatment of Patients with Type 2 Diabetes. Am Health Drug Benefits. 2017 Jun;10(4):178-188.
22. He L, Wang J, Ping F, Yang N, Huang J, Li Y, Xu L, Li W, Zhang H. Association of Glucagon-Like Peptide-1 Receptor Agonist Use With Risk of Gallbladder and Biliary Diseases: A Systematic Review and Meta-analysis of Randomized Clinical Trials. JAMA Intern Med. 2022 May 1;182(5):513-519. doi: 10.1001/ jamainternmed.2022.0338.
23. Joshi G, Abdelmalak B, Weigel W, Soriano S, Harbell M, Kuo C, Strickler P, Domino K, American Society of Anesthesiologists (ASA) Task Force on Preoperative Fasting. (2023, June 29). American Society of Anesthesiologists Consensus-Based Guidance on Preoperative Management of Patients (Adults and Children) on Glucagon-Like Peptide-1 (GLP-1) Receptor Agonists. ASAHQ. https://www.asahq.org/about-asa/newsroom/news-releases/2023/06/american-societyof-anesthesiologists-consensus-based-guidance-on-preoperative
24. Huda MN, Kim M, Bennett BJ. Modulating the Microbiota as a Therapeutic Intervention for Type 2 Diabetes. Front Endocrinol (Lausanne). 2021 Apr 7;12:632335. doi: 10.3389/fendo.2021.632335.
25. Ridaura VK, Faith JJ, Rey FE, Cheng J, Duncan AE, Kau AL, Griffin NW, Lombard V, Henrissat B, Bain JR, Muehlbauer MJ, Ilkayeva O, Semenkovich CF, Funai K, Hayashi DK, Lyle BJ, Martini MC, Ursell LK, Clemente JC, Van Treuren W, Walters WA, Knight R, Newgard CB, Heath AC, Gordon JI. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science. 2013 Sep 6;341(6150):1241214. doi:10.1126/science.1241214.
26. Hanssen NMJ, de Vos WM, Nieuwdorp M. Fecal microbiota transplantation in human metabolic diseases: From a murky past to a bright future? Cell Metab. 2021 Jun 1;33(6):1098-1110. doi: 10.1016/j.cmet.2021.05.005.
27. Keshavarz Azizi Raftar S, Ashrafian F, Yadegar A, Lari A, Moradi HR, Shahriary A, Azimirad M, Alavifard H, Mohsenifar Z, Davari M, Vaziri F, Moshiri A, Siadat SD, Zali MR. The Protective Effects of Live and Pasteurized Akkermansia muciniphila and Its Extracellular Vesicles against HFD/CCl4-Induced Liver Injury. Microbiol Spectr. 2021 Oct 31;9(2):e0048421. doi: 10.1128/Spectrum.00484-21. Epub 2021 Sep 22.
28. Yoon HS, Cho CH, Yun MS, Jang SJ, You HJ, Kim JH, Han D, Cha KH, Moon SH, Lee K, Kim YJ, Lee SJ, Nam TW, Ko G. Akkermansia muciniphila secretes a glucagon-like peptide-1-inducing protein that improves glucose homeostasis and ameliorates metabolic disease in mice. Nat Microbiol. 2021 May;6(5):563573. doi: 10.1038/s41564-021-00880-5. Epub 2021 Apr 5. PMID: 33820962.
29. Depommier C, Everard A, Druart C, Plovier H, Van Hul M, Vieira-Silva S, Falony G, Raes J, Maiter D, Delzenne NM, de Barsy M, Loumaye A, Hermans MP, Thissen JP, de Vos WM, Cani PD. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study. Nat Med. 2019 Jul;25(7):1096-1103. doi: 10.1038/s41591-019-0495-2. Epub 2019 Jul 1.
30. Pendulum Internal Data, 2024. Cell culture data from human intestinal L-cell lines. Data demonstrating dose dependent release of GLP-1 as a result of bacterial product administration.
31. Yadav H, Lee JH, Lloyd J, Walter P, Rane SG. Beneficial metabolic effects of a probiotic via butyrate-induced GLP-1 hormone secretion. J Biol Chem. 2013 Aug 30;288(35):25088-25097. doi: 10.1074/jbc.M113.452516. Epub 2013 Jul 8.
32. Tolhurst G, Heffron H, Lam YS, Parker HE,Habib AM, Diakogiannaki E, Cameron J, Grosse J, Reimann F, Gribble FM. Short-chain fatty acids stimulate glucagonlike peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes. 2012 Feb;61(2):364-71. doi: 10.2337/db11-1019. Epub 2011 Dec 21.
33. Stoeva MK, Garcia-So J, Justice N, Myers J, Tyagi S, Nemchek M, McMurdie PJ, Kolterman O, Eid J. Butyrate-producing human gut symbiont, Clostridium butyricum, and its role in health and disease. Gut Microbes. 2021 Jan-Dec;13(1):1-28. doi: 10.1080/19490976.2021.1907272.
34. Perraudeau F, McMurdie P, Bullard J, Cheng A, Cutcliffe C, Deo A, Eid J, Gines J, Iyer M, Justice N, Loo WT, Nemchek M, Schicklberger M, Souza M, Stoneburner B, Tyagi S, Kolterman O. Improvements to postprandial glucose control in subjects with type 2 diabetes: a multicenter, double blind, randomized placebo-controlled trial of a novel probiotic formulation. BMJ Open Diabetes Res Care. 2020 Jul;8(1):e001319. doi: 10.1136/bmjdrc-2020-001319.
35. McMurdie PJ, Stoeva MK, Justice N, Nemchek M, Sieber CMK, Tyagi S, Gines J, Skennerton CT, Souza M, Kolterman O, Eid J. Increased circulating butyrate and ursodeoxycholate during probiotic intervention in humans with type 2 diabetes. BMC Microbiol. 2022 Jan 8;22(1):19. doi: 10.1186/s12866-021-02415-8.
36. Murakami M, Une N, Nishizawa M, Suzuki S, Ito H, Horiuchi T. Incretin secretion stimulated by ursodeoxycholic acid in healthy subjects. Springerplus. 2013 Dec;2(1):20. doi: 10.1186/2193-1801-2-20. Epub 2013 Jan 22.
37. Mahmoud AAA, Elshazly SM (2014) Ursodeoxycholic Acid Ameliorates Fructose-Induced Metabolic Syndrome in Rats. PLOS ONE 9(9): e106993. https://doi. org/10.1371/journal.pone.0106993
38. She J, Tuerhongjiang G, Guo M, Liu J, Hao X, Guo L, Liu N, Xi W, Zheng T, Du B, Lou B, Gao X, Yuan X, Yu Y, Zhang Y, Gao F, Zhuo X, Xiong Y, Zhang X, Yu J, Yuan Z, Wu Y. Statins aggravate insulin resistance through reduced blood glucagon-like peptide-1 levels in a microbiota-dependent manner. Cell Metab. 2024 Feb 6;36(2):408-421.e5. doi: 10.1016/j.cmet.2023.12.027. PMID: 38325336.
39. Pendulum Internal Data, 2024. GLP-1 Daily–Hunger, Fullness, and Cravings Consumer Perception Study.
40. White MA, Whisenhunt BL, Williamson DA, Greenway FL, Netemeyer RG. Development and validation of the food-craving inventory. Obes Res. 2002 Feb;10(2):107-14. doi: 10.1038/oby.2002.17.
