Genetische structuur en herkomst van patrijs in Vlaanderen Io Deflem, Caroline Mouton, Nico De Regge, An Van Breusegem, Sabrina Neyrinck en Joachim Mergeay

Auteurs: Io Deflem , Caroline Mouton , Nico De Regge , An Van Breusegem , Sabrina Neyrinck , Joachim Mergeay Instituut voor Natuur- en Bosonderzoek Reviewers: Thomas Scheppers Het INBO is het onafhankelijk onderzoeksinstituut van de Vlaamse overheid dat via toegepast wetenschappelijk onderzoek, data- en kennisontsluiting het biodiversiteitsbeleid en -beheer onderbouwt en evalueert. Vestiging: INBO vlaanderen.be/inboGaverstraatGeraardsbergen4,9500Geraardsbergen e-mail: io.deflem@inbo.be Wijze van citeren: Deflem I, Mouton C, De Regge N, Van Breusegem A, Neyrinck S, Mergeay J (2021). Genetische structuur en herkomst van patrijs (Perdix perdix) in Vlaanderen. Rapporten van het Instituut voor Natuur- en Bosonderzoek 2021 (57). Instituut voor Natuur- en Bosonderzoek, Brussel. DOI: doi.org/10.21436/inbor.62125143 Depotnr: RapportenD/2021/3241/362vanhetInstituut voor Natuur- en Bosonderzoek 2021 (57) ISSN: Verantwoordelijke1782-9054 uitgever: Maurice Hoffmann Foto cover: © Nico De Regge Dit onderzoek werd uitgevoerd in opdracht van: Het Agentschap voor Natuur en Bos Dit werk valt onder een Creative Commons Naamsvermelding GelijkDelen 4.0 Internationaal licentie.
www.vlaanderen.be/inbo Io Deflem, Caroline Mouton, Nico De Regge, An Van Breusegem, Sabrina Neyrinck, Joachim Mergeay doi.org/10.21436/inbor.62125143 GENETISCHE STRUCTUUR EN HERKOMST VAN PATRIJS (PERDIX PERDIX) IN VLAANDEREN


///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 2 van57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo Dankwoord We bedanken Hubertus VerenigingVlaanderen, de lokale jachtrechthouders, grondeigenaars, Natuurinspectie, Didier Vangeluwe en de ringers van het Koninklijk Belgisch Instituut voor Natuurwetenschappen voor de enthousiaste medewerking bij het inzamelen van de stalen. Dank ook aan Ariane Staelens van het Instituut voor Landbouw , Visserij en Voedingsonderzoek en David Halfmaerten voorde hulp bij hetlabowerk. Ten slotte bedanken we Thomas Scheppers en Yasmine Verzelen voor hun inzicht en input bij de interpretatie van de Ditresultaten.onderzoek werd gefinancierd via het Jachtfonds van het Agentschap voor Natuur en Bos onder overeenkomst.
Ondanks de lage genetische differentiatie tussen gekweekte en wilde patrijzen, konden we een duidelijk onderscheid maken tussen beidetypes. Metuitzondering van individuen uit één kweekpopulatie, lijken alle kweekindividuen afkomstig te zijn van éénzelfde (al dan niet ancestrale) bronpopulatie. Hierdoor kunnen we dus geen uitspraken doen over de exacte herkomst van gekweekte individuen per kwekerij. Wanneer we vervolgens kijken naar de genetische structuur van enkel de wilde populaties in Vlaanderen, onderscheiden we twee genetische clusters. Patrijzen afkomstig uit de het Noorden van de provincie Antwerpen vormen een duidelijk aparte groep ten opzichte van alle andere individuen. Verder lijkt de geobserveerde genetische structuur deels verklaard te worden door verbreiding over korte afstanden, wat zich vertaalt in een duidelijke west oost gradiënt van genetische structuur. Wanneer we focussen op de detectie van uitzettingen, zien we dat de gevolgde methode toelaat om zuivere wilde individuen te onderscheiden van zuivere individuen van kweekherkomst en van kruisingen tussen beiden. In gevangenschap gekweekte patrijzen werden vooral waargenomen in de provincies Vlaams Brabanten Limburg;ditzijn regio’s waar ook de densiteiten van wilde patrijzen lagerzijn. Bovendien was een deel van deze individuen meerjarig en detecteerden we nakomeling van gekweekte en wilde individuen, wat doet vermoeden datsommige uitgezetteindividuen zich ook succesvol kunnen voortplanten. In een latere fase van het onderzoek zullen we een subset van merkers (circa 300) kiezen die in de
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 3 van 57 Samenvatting
Sinds hetJachtdecreetvan juli 1991 is hetin Vlaanderen wettelijk verboden om jachtwild uitte zetten. Ondanks dit verbod zijn er toch vermoedens dat patrijzen (Perdix perdix) op regelmatige basis worden uitgezet. Door de verschillende geografische herkomst en onvermijdelijke adaptatie aan kweekomstandigheden, verwachten we dat kweekindividuen genetisch duidelijk verschillen van wilde patrijzen. Vermenging van beide kan bovendien nadelig zijn voor natuurlijke populaties, zeker in het kader van de recente achteruitgang van patrijs in Vlaanderen. Om zulke illegale uitzettingen beter te kunnen opsporen, gingen we in deze studie na of het mogelijk is om met genetische technieken patrijzen afkomstig uit gevangenschap maarvrijgelaten in het wild te onderscheiden van wilde vogels. De bekomen resultaten geven ons ook een beter zicht op de genetische diversiteit en structuur van patrijs in Vlaanderen. Dat kan van belang zijn bij het beheer van patrijzen. In het najaar van 2020 en het voorjaar van 2021 werden verspreid over Vlaanderen stalen verzameld van 495 in hetwild voorkomendepatrijzen. Daarnaastwerden stalen genomen van 120 in gevangenschap gekweekte patrijzen afkomstig uit vijf kwekerijen uit Vlaanderen, Nederland en Frankrijk. We genotypeerden een totaal van 546 patrijzen op meer dan 30 000 genetische merkers. Op basis van deze genetische merkers werden verschillende parameters berekend. De genetische diversiteit was nietverschillend tussen kweek en wilde populaties. Bovendien was de diversiteitin geen enkeleregio laagen observeerden we in geen enkele regio tekenen van inteelt. We leiden hieruitaf dater in het kadervan genetische diversiteit geen noodzaak is tot bijplaatsingen, herintroducties of andere translocaties van patrijs in Vlaanderen.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 4 van57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo toekomsteen snelle en doeltreffende screeningvan patrijzen zal toelaten in het kader van de detectie van illegale uitzettingen.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 5 van 57 Aanbevelingen voor beheer en/of beleid
Hieronder
De genetischestructuurdie we waarnemen is gebaseerd op vrijwillige medewerking van wildbeheereenheden, en biedt daardoor geen volledig beeld van de praktijk van uitzettingen van patrijzen. We raden aan om de opening van de jacht op patrijs per wildbeheereenheid vanaf 2022 te verbinden aan een verplichting tot inzamelen van DNA van elk gestrektexemplaar. Dit zal ons toelaten om een meer objectief beeld te krijgen van de genetische structuur van patrijs in Vlaanderen, en van het voorkomen van uitgezette patrijzen.
Dit project had als doel om een methode te ontwikkelen die het mogelijk maakt om op een snelle en routinematige wijze illegale uitzettingen van patrijs te detecteren. Hiervoor bestudeerden we de genetische structuur en diversiteit van wilde patrijzen in Vlaanderen. vatten we alle aanbevelingen, die meer uitgebreid in de discussie van dit rapport worden besproken, samen. Genetische diversiteit verschiltnietsterk tussen wilde patrijzenpopulaties. Bovendien werd er nergens inteelt vastgesteld. Bijplaatsingen en/of translocaties zijn dus niet nodig met oog op het verhogen van de genetische diversiteit en om inteelt tegen te gaan. Op basis van de genetische structuur zien we dat patrijzen uit het noorden van Antwerpen een aparte cluster vormen. We suggereren deze ook als dusdanig als aparte beheereenheid te behandelen.
Due to differences in origin and strong adaptation to breeding conditions, we expect that farmed partridges differ genetically from their wild conspecifics. Mixing the two might be detrimental to natural populations, especially given the strong decline of grey partridge in Flanders. In order to detect such illegal releases, we explored the possibility to genetically distinguish partridges reared in captivity, but released into the wild, from wild birds. Moreover, the obtained results will also give us a better insight into the genetic diversity and structure of the grey partridge in Flanders, which can be important forfuture conservation and Inmanagementtheautumn of 2020 and the spring of 2021, samples were collected from 495 wild partridges distributed across Flanders. In addition, samples were taken from 120 captive bred partridges from five farms in Flanders, the Netherlands and France. We genotyped a total of 546 partridges on more than 30,000 genetic markers.
Focusing on the detection of illegal releases, we are able to detect both recent (pure captive bred individuals) and historical (introgressed individuals) releases. Captive bred partridges were mainly observed in the provinces of Flemish Brabant and Limburg, regions with low partridge densities. In addition, some of these individuals were adults (>1y old) and the detection of introgressed individuals indicates successful reproduction of some captive bred individuals in the past. We will further select a panel of circa 300 genetic markers for routine monitoring, which will allow a fast and effective screening of partridges in the future.
Releasing game has been forbidden in Flanders since 1991. Despite this ban, captive bred individuals of the grey partridge (Perdix perdix) are still illegally released on a regular basis.
Genetic diversity did not differ between farmed and wild populations. In addition, diversity was not reduced in any of the wild populations. This suggests thatintroducing or translocating individuals is not needed for the genetic conservation of the grey partridge in Flanders.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 6 van57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo English abstract
Despite the low genetic differentiation, we were able to distinguish captive bred partridges from wild birds. With the exception of individuals from one farm, all captive bred individuals appeared to originate from the same source population. As a result, we cannot make statements aboutthe exactorigin of illegallyreleased individuals On the basis of the genetic structure of the wild populations in Flanders, we observed that partridges from the north of the province of Antwerp form a clearly separate group from all otherindividuals. Furthermore, the observed genetic structure appears to be partly explained by short distance dispersal, which translates into a clear west east gradient of genetic structure.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 7 van 57 Inhoudstafel Dankwoord 2 Samenvatting 3 Aanbevelingen voor beheeren/of beleid...............................................................................5 English abstract 6 1 Inleiding en doelstellingen.............................................................................................9 1.1 Patrijs in Vlaanderen en Europa..............................................................................9 1.1.1 De achteruitgang van patrijs 10 1.1.2 De jacht op patrijs in Vlaanderen 11 1.2 Populatie en conservatiegenetica........................................................................13 1.2.1 Genetische aanpassingen aan gevangenschap 13 1.2.2 Genetische aanpassingen aan gevangenschap in patrijs 14 1.2.3 Genetische identificatie van (illegale) introducties..........................................15 1.3 Doelstellingen 16 2 Werkwijze 17 2.1 2.22.1.22.1.1Staalname...........................................................................................................17Wildvangstpatrijzen17Kwekerijen17Genetischeanalyses............................................................................................202.2.1Sequencing202.2.2Bioinformatischedataanalyse212.2.3Dataanalyse................................................................................................22 3 Resultaten 24 3.1 Stap 1: Genetische diversiteit en structuur van alle individuen 24 3.2 Stap 2: Genetische diversiteit en structuur van autochtone populaties ...................30 4 Discussie 37 4.1 Evaluatie van de genetische diversiteit 37 4.2 Evaluatie van de genetische structuur...................................................................38 4.2.1 Genetische afstand tussen wilde en kweekpopulaties 38 4.2.2 Genetische structuur van wilde populaties 39 4.3 Recente en minder recente uitzettingen van patrijs in Vlaanderen .........................39 4.3.1 Toelichtingen validatie van methodiek 39 4.3.2 Onderscheid tussen recente en minder recente uitzettingen 40 4.3.3 Uitzettingen in Vlaanderen ...........................................................................41 4.4 Vervolgonderzoek................................................................................................42 4.5 Aanbevelingen voor beleid 42
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 8 van57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo Referenties ........................................................................................................................44 BijlageBijlageBijlageBijlage................................................................................................................................511:Supplementairefiguren512Overzichtgenetischestructuuralleindividuen(perWBE)........................................543Overzichtgenetischestructuurwildeindividuen(perWBE).....................................57
Figuur 1. Voorlopige schattingen van het aantal broedvogels van patrijs in 2020 2022 (Vlaamse Vogelatlas 2021).
De gewone patrijs (Perdix perdix, Phasianidae) is een akkervogel die verspreid voorkomt in Europa. Patrijzen zijn typische grondbroedende vogels van open agrarische gebieden (Birkan 1988; Carroll 1993; McGowan & Kirwan 2013). Op Europees niveau bevindt, met 644 000 tot 1 266 000 broedparen, ongeveer 50% van de Europese patrijzenpopulatie zich binnen de landsgrenzen van Frankrijk. Hoge aantallen komen ook voor in Rusland, Polen en Roemenië (respectievelijk 14%, 9% en 8% van de totale patrijzenpopulatie; BirdLife International 2015). De totale Belgische patrijzenpopulatie bestaat uit een geschatte 5400 tot 13 600 paren (BirdLife International 2015, 2017), wat overeenkomt met minder dan 1% van de volledige Europese populatie(Bijlage 1;BirdLife International 2015). De Vlaamse populatie op zijn beurt zou zo’n 5000 broedparen tellen (periode 2013 2018, Vermeersch et al. 2020). In Vlaanderen zijn densiteiten het hoogst in de provincie West Vlaanderen en nabij de gewestgrens met Wallonië in de Vlaamse Ardennen. In de provincies Antwerpen en Limburg is de verspreiding het meest versnipperd, met hogere densiteiten rond Hoogstraten, Zuidoost Brabant en Haspengouw (Figuur1; Devos 2004; Vlaamse Vogelatlas 2021). In Wallonië worden de meeste patrijzen waargenomen in het noordwesten waar de grootste concentraties zich tegen de Franse grens bevinden in Henegouwen en Waals Brabant (Bijlage 1; AVES 2020).
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 9 van 57 1 INLEIDING EN DOELSTELLINGEN 1.1 PATRIJS IN VLAANDEREN EN EUROPA
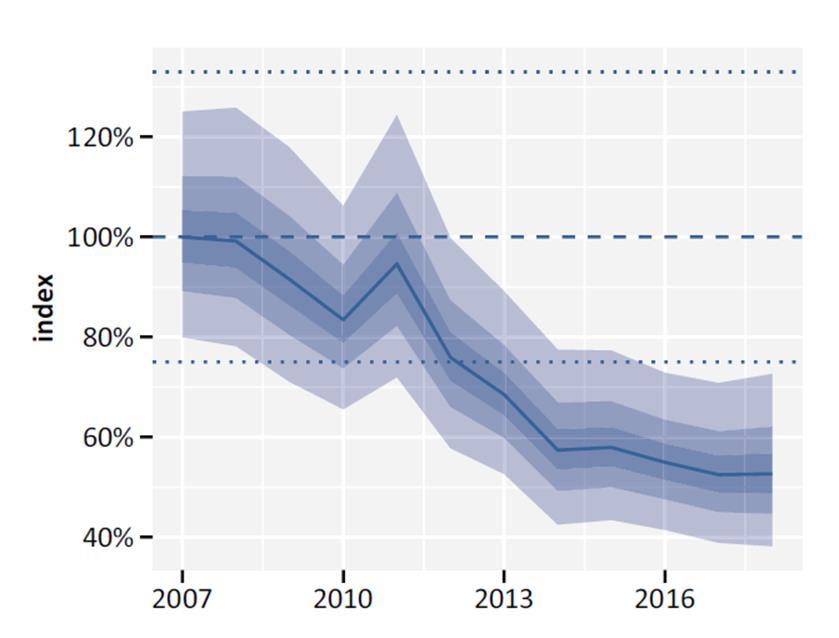
Figuur 2. Trend van de patrijs (index) in Vlaanderen tijdens de periode 2007 2018 waarbij het referentiejaar 2007 op 100% werd gezet (Vermeersch et al. 2020). Indexen voor 2019 2020 werden nog niet toegevoegd. 1.1.1 De achteruitgang van patrijs De achteruitgang van de patrijs wordt doorgaans toegewezen aan een verminderde habitatkwaliteit en geschiktheid ten gevolge van de intensivering van de landbouw. De moderniseringen de daarbij behorende nieuwe landbouwtechnieken resulteren vaak in het verwijderen van smalle hagen en verwilderde grasstroken waar diverse grassen en kruiden
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 10 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo Ondanks deze schijnbaar hoge aantallen, gaat de patrijs sinds enkele tientallen jaren sterk achteruit. Na een lange geschiedenis van fluctuaties in populatieaantallen vertoont de Europese en Vlaamse patrijzenpopulatie sinds enkeledecenniaeen dalendetrend (o.a. BirdLife International 2017; Devos et al. 2016a, 2016b; Kuijper et al. 2009; Vermeersch et al. 2020, 2021). Bovendien voorspellen populatiemodellen, op zowel korte als lange termijn, een verdere achteruitgang (Bijlage 1; BirdLife International 2015). Op Europese schaal onderging de soort sinds 1980 een erg steile daling van maar liefst 94% (PECBMS in Buner & Gottschalk 2021). Op Vlaams niveau kende de patrijzenpopulatie een sterke afname van 47% over de periode 2007 2020 (Onkelinx et al. 2021, Vermeersch et al. 2021). Figuur 2 toont deze trend, aangeduid als index, in Vlaanderen voor patrijs voor de periode 2007 2018 (Vermeersch et al. 2020). Een gelijkaardige dalingwordtook waargenomen in de afschotstatistieken bijgehouden door de Vlaamse wildbeheereenheden (Scheppers et al. 2019). Deze trends in de populatiedynamiek van patrijs zorgden er voor dat de soort werd opgenomen in de rode lijst van bedreigdeEuropese vogelsoorten en dat de status op Vlaams niveau in de meest recente versie van de IUCN rode lijst van de Vlaamse broedvogels als kwetsbaar werd aangeduid (Devos et al. 2016a).
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 11 van 57 gedijen (Southwood & Cross 1969). Deze landschapselementen zijn cruciaal voor het voortbestaan van de patrijs, dewelke hierin hun voedsel en toevlucht vinden tijdens de broedperiodeen terbeschermingdienen tegen predatoren. Hieraan samenhangend mogen de potentiële gevaren van ongelukkig gekozen maaimomenten niet vergeten worden, daar het uitmaaien van nesten en kuikens zware gevolgen kan hebben voorpopulaties van akkervogels (ANB Bovendien2020).leidthetintensiveren van landbouw vaak tot een verhoogd gebruik van pesticiden en herbiciden. Verschillende studies (o.a. Kuijper et al. 2009; Southwood & Cross 1969) toonden intussen aan dat dit in direct verband staat met een veranderend en verminderd voedselaanbod in verschillende kritieke periodes van het jaar. Volwassen individuen consumeren voornamelijk zaden, granen, klaver, groeitoppen van gewassen en insecten (McGowan & Kirwan 2013). Hierbij foerageren patrijzen tussen gewassen binnen een straal van 500m verwijderd van de nestplaats (Green 1984). Kuikens daarentegen zijn in de vroegste fases van hun leven voornamelijk aangewezen op het eten van verschillende insecten (Southwood &Cross 1969). De overlevingvan de kuikens speelteen sleutelrol in de algemene populatiedynamiek bij patrijs (Blank et al. 1967; Kuijper et al. 2009; Manly 1977 in Bro et al. 2000; Southwood &Cross 1967, 1969). Wanneerermindervoedsel voorhanden is, in dit geval door een lage densiteit aan insecten door pesticidengebruik en landbouwingrepen zoals het ploegen van grond waarin insectenlarven en poppen gedijen, moeten kuikens vaak langere afstanden afleggen om geschiktvoedselte vinden. Ditmaaktde kans op sterfte doorhongerof door een verhoogde blootstellingaan predatoren groter. Voorhetvoortbestaan van de patrijs is het dus cruciaal om de negatieve impact van het pesticidengebruik en landbouwbeleid op het voedselaanbod te verminderen (Kuijper et al. 2009). 1.1.2 De jacht op patrijs in Vlaanderen De achteruitgang van de patrijs heeft onder andere geleid tot een verstrenging van de jachtwetgeving. De jachtop de patrijs is momenteel geopend van 15 september tot en met 14 november (BVR 09/10/20151). Deze verstrenging houdt ook in dat er enkel gejaagd mag worden op patrijs binnen een erkende wildbeheereenheid waarvan uit het faunabeheerplan blijkt dat er een patrijsvriendelijk beheer wordt uitgevoerd en, wanneer er gedurende drie voorafgaande jaren een gemiddelde voorjaarsstand van drie koppels per 100 ha open ruimte geteld werden (BVR 25/04/20142). Voor een meer uitgebreide informatie inzake de voorwaarden waaraan voldaan moet worden om de jacht op patrijs te laten doorgaan, verwijzen we naarde rapporten van o. a. Scheppers et al. (2019) en Verzelen et al. (2020). Het bejaagbaar zijn van een soort die sterk achteruitgaat leidt echter tot heel wat controverse. Overbejaging kan nefast zijn wanneer een soort al onder druk staat door veranderingen in omgeving en habitatkwaliteit, zeker in gebieden met lage populatiedensiteiten. Onderzoek
Art. 22. Besluit van de Vlaamse Regering houdende vaststelling van de voorwaarden waaronder de jacht kan worden uitgeoefend [citeeropschrift:"het Jachtvoorwaardenbesluit van 25 april 2014"] (B.S. 2014)
Art. 4. § 1. Besluit van de Vlaamse Regering betreffende de jachtopeningstijden in het Vlaamse Gewest [citeeropschrift: "het jachtopeningsbesluit"] (B.S. 2015)
2
1
toont bovendien aan dat de voorjaarsstand van patrijzen lager is in situaties waar jacht toegelaten is en datdeze verhoogdemortaliteitamperwordtgecompenseerd door een daling in natuurlijke mortaliteit (Scheppers et al. 2019). Uitzettingen van allerlei klein jachtwild, in functie van de jacht, komen nog steeds frequent voor in grote delen van Europa. In Frankrijk is het vrijlaten van in gevangenschap grootgebrachte dieren in functie van de jacht (en soortherstel) volledig legaal. Deze uitzettingen worden onderandere gereguleerd door het Franse voedselagentschap. Volgens het Syndicat National des Producteurs de Gibier de Chasse (SNPGC) zouden jaarlijks een geschatte 5 miljoen patrijzen (zowelgewoneals rode patrijzen, Alectoris rufa) uitgezetworden en zo’n 30 tot 40 miljoen eieren (fazant, rode en gewone patrijs) geproduceerd worden, waarvan een groot deel bedoeld is voor export binnen Europa Met ongeveer 300 gespecialiseerde kwekerijen zijn uitzettingen ten behoeve van de jacht ook in het Vereni gd Koninkrijk alomtegenwoordig. Jaarlijks zouden hiergemiddeld 200 000 uitzettingen van patrijs plaatsvinden (Aebischer 2019 in Buner & Gottschalk 2020). Zo zou 83% van het afschot in het Verenigd Koninkrijk afhankelijk zijn van geïntroduceerde individuen afkomstig uit kwekerijen (The Game Farmers’ Association 2021).
In Vlaanderen daarentegen is hetsinds hetJachtdecreetvan 24 juli 1991, verboden om wild uit te zetten. In Wallonië is sinds 2021 de uitzetting van patrijzen voor de jacht sensu stricto ook verboden. Enkel in functie van bijplaatsingen of herintroducties is de uitzetting nog onder specifieke voorwaarden mogelijk. Ondanks hetuitzetverbod in Vlaanderen werd in het najaar van 2020 een lokaal jachtverbod uitgevaardigd nadat uitgezette patrijzen werden teruggevonden in de regio rond Veurne (Schauvliege & De Bruyn 2020). Na dit voorval rees opnieuw de vraag welke gevolgen dergelijke introducties kunnen teweeg brengen. Het uitzetten van gekweektjachtwild blijktzelden te leiden tot duurzame populaties omwille van de zeer beperkte overlevingskansen van de uitgezette dieren (Homberger et al. 2021) Daarnaast verhoogt de kans dat wilde patrijzen mee geschoten worden met de gekweekte dieren, hetgeen overbejaging van de wilde patrijzen in de hand werkt. Bovendien kunnen uitzettingen ook nadelige gevolgen hebben voor natuurlijke populaties, wanneer genetische en/of gedragsmatige verschillen tussen wilde en gekweekte individuen te groot zijn en er kruisingen optreden (Karlsson et al. 2016; Lavretsky et al. 2020; Putaala & Hissa 1998). Om na te gaan watde genetischeeffecten zijn van uitzettingen is hetdaarom nuttigom de genetische diversiteit en structuur van overblijvende populaties in kaart te brengen.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 12 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo
1.2.1 Genetische aanpassingen aan gevangenschap De mogelijke genetische problemen van hetuitzetten van in gevangenschap gekweektedieren zithem in de afkomsten achtergrond van deze individuen. In hetalgemeen worden individuen afkomstiguitkwekerijen blootgesteld aan condities die sterk verschillen van een natuurlijke omgeving. Bovendien zijn effectieve populatiegroottes van kweekpopulaties vaak laag. Hierdoorkan het kweken van individuen in gevangenschap een sterke invloed hebben op de genetische achtergrond van individuen en populaties, wat kan resulteren in een verlies aan genetische diversiteit, inteelt, accumulatie van nadelige mutaties (‘inbreeding load’) en adaptatie aan gevangenschap (Frankham et al. 2002; Frankham 2008; Laikre et al. 2010). Adaptatie aan gevangenschap wordtvaak bestudeerd bij vissen en amfibieën, maar is ook een welbekend fenomeen bij vogels en zoogdieren (Frankham 2007) en kan al optreden binnen enkele generaties (Araki et al. 2007; Christie et al. 2012; Laikre et al. 2010). Deze snelle aanpassingen zijn vaak hetgevolgvan de hoge selectiedruk die individuen in gevangenschap ervaren. Voorbeelden van zulke aanpassingen zijn veranderingen in gedrag, een stijging in het aantal nakomelingen en veranderde timing in reproductie (Gilligan & Frankham 2003).
Het bestuderen van de populatiegenetica van patrijs in Vlaanderen kan ons belangrijke informatie verschaffen omtrentde genetische populatiestructuur, de invloed van uitzettingen en de huidige status van de soort. Voldoende genetische diversiteit is essentieel voor het behoud van het adaptief potentieel van populaties, zeker wanneer men kijkt naar de snelle veranderingvan de omgeving(Frankham etal. 2010). Indien blijktdatde genetischediversiteit laag is in bepaalde regio’s kunnen bovendien bepaaldemaatregelen, zoals hetlegaal uitzetten of het verplaatsen van individuen, genomen worden. Het bestuderen van de genetische structuurdaarentegen geefteen beeld overde genetische verschillen tussen populaties, en de connectiviteit en dispersie afstanden van patrijzen. De genetische structuur van patrijs in Vlaanderen kan echter beïnvloed zijn door (al dan niet recente) uitzettingen van in gevangenschap gekweekte individuen.
Wanneergekweekte individuen vervolgens worden vrijgelaten in een natuurlijke omgeving, blijkt hun overlevings en voortplantingssucces over het algemeen vrij laag (Buner & Schaub 2008; Champagnon et al. 2012; Parish & Sotherton 2007; Sokos et al. 2008). Dit wordt vaak toegewezen aan de afwezigheid van anti predatiegedrag (Kraaijeveld Smit et al. 2006), lage stresstolerantie, suboptimale habitatselectie (Brittas et al. 1992; Sage et al. 2003) en lage reproductieveoutputvan gekweekteindividuen in een natuurlijke omgeving(Araki et al. 2007; Frankham et al. 2002). Naast de lage overlevingskansen onder andere ten gevolge van maladaptatie, kunnen gekweekte individuen ook een negatieve invloed hebben op de genetische integriteit van wilde individuen en populaties. Bij kruisingen tussen beide (introgressie) kunnen maladaptieve genvarianten zich verspreiden, wat leidt tot fitnessverlies door deze genetische vermenging (McGinnity et al. 2003; Muñoz Fuentes et al. 2007; Randi 2008).
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 13 van 57 1.2 POPULATIE- EN CONSERVATIEGENETICA
overlevingskansen
Bovendien toont een groot aantal studies uit verschillende Europese landen aan dat de overlevings en voorplantingskansen van gekweekte patrijzen erg laag zijn Deze lage overlevingis vaak hetresultaatvan de gebrekkige habitatselectie en de afwezigheid van anti predatiegedrag in gekweekte individuen. Andere verschillen die de overlevingskansen van gekweekte individuen verlagen zijn verminderde waakzaamheid, een gebrek aan correcte voedingsstrategie en vaak gebrekkige resistentie tegen ziektes. De patrijzen die wel langere tijd overleven, planten zich zelden succesvol voort (Buner et al. 2005; 2011; Morris & Gilroy 2008; Parish & Sotherton 2007; Putaala et al. 2001; Rantanen et al. 2010; Rymešová et al. Slechts2012) een beperkt aantal studies heeft de genetische structuur van patrijs onderzocht (Andersen & Kahlert 2012; Bech et al. 2012; 2014; 2020; Liukkonen Anttila et al. 2012). Van deze studies bestudeerde enkel Bech et al. (2020) de genetische verschillen tussen wilde en gekweekteindividuen, en meteen zeer beperkte genetische resolutie. Uit deze studie bleek dat wilde en gekweekte patrijzen in hetnoorden van Frankrijk nietsterk van elkaarverschillen. Bovendien toontgeen enkele studie aan dathybriden frequentvoorkomen, in tegenstellingtot bij rode patrijzen (Barbanera et al. 2010; Blanco Aguiar et al. 2008; Casas et al. 2011). Deze afwezigheid of lage densiteit van hybriden kan worden verklaard door de lage van uitgezette patrijzen, zoals eerder vermeld, door de lage van gekruiste individuen, of door het gebrek aan statistische kracht om kruisingen te detecteren (Bech et al. 2020).
overlevingskansen
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 14 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo 1.2.2 Genetische aanpassingen aan gevangenschap in patrijs In functie van de jacht, worden patrijzen afkomstig uit gevangenschap al decennia lang uitgezet in verschillende Europese landen. Doorgaans worden patrijzen gekweekt in grootschalige industriële complexen met behulp van broedmachines. Meer conservatieve kwekerijen daarentegen proberen meer natuurlijke condities na te bootsen en proberen inteelt tegen te gaan door inmenging van individuen uit het wild (Bech et al. 2020; Bro et al. 2017; Soko et al. 2008). De sterk verschillende condities in kwekerijen ten opzichte van een natuurlijke omgeving uiting zich onder meer in een onbeperkte toegang tot kunstmatige voeding, de hoge densiteitaan pathogenen en het gebruik van anti parasitaire middelen, het gebrek aan natuurlijke omstandigheden en mogelijkheid tot sociaal gedrag, en de incubatie van een onbeperktaantal eieren. Dit leidt verder tot het verbreken van de ruilfunctie tussen tussen legselgrootteen de kans op hetsuccesvol uitbroeden van alle eieren, aanpassingen aan een onnatuurlijk dieet, verlaagde vluchtrespons en verhoogde stresstole rantie. Sommige producenten gebruiken al tientallen generaties bepaalde kweeklijnen, watonvermijdelijk leidt tot sterke adaptatie aan gevangenschap (Gilligan & Frankham 2003).
Genetische verschillen tussen gekweekte en wilde patrijzen, die we kunnen verwachten ten gevolge van een verschil in herkomsten adaptaties aan gevangenschap,geven de mogelijkheid om illegale uitzettingen te detecteren. Er zijn verschillende complementaire methodes waarmee we in principe kunnen achterhalen of en op welke schaal individuen gekweekt in gevangenschap worden uitgezet.
Wahlundeffect. Wanneerpopulaties meteen verschillende genetische samenstelling vermengd worden (bv. een autochtone populatie waar dieren van een andere herkomst worden aan toegevoegd), is de waargenomen heterozygositeit lager dan verwacht onder random paarvorming tussen de ouders van de bemonsterde dieren. Zo’n afwijking is een eerste indicatie van een illegale uitzetting. We verwachten dit echter enkel te kunnen waarnemen wanneer het aandeel van uitgezette en autochtone dieren van dezelfde grootteorde is. Multivariate ordinatie. De genetische samenstelling van alle dieren laat toe om genetische afstanden te bepalen tussen genotypes. Die genetische afstanden gebruiken we om met een Principiële Componenten Analyse (PCA) de belangrijkste genetische structuren te onderscheiden. Genotypes die sterk op elkaar gelijken zijn nauw verwantals gevolgvan een gemeenschappelijke oorsprongen groeperen samen op een diagram. Wanneer in eenzelfde gebied patrijzen voorkomen die in een PCA diagram niet samen clusteren, geeft dit aan dat er recent twee of meer populaties vermengd werden met elkaar.. Discriminant analyses, clustering en toewijzingen. Aan de hand van de gekende oorsprong van stalen (bv. gekweekt versus wild) is het mogelijk om na te gaan hoe goed elk dier via de genetische samenstelling wordt toegewezen aan zijn referentiegroep. Dieren die als mogelijk wild werden geïdentificeerd (afschot, ringvangst, verkeersslachtoffer …) maar die toch worden toegewezen aan een referentiepopulatie van gekweekte dieren hebben met hoge zekerheid een herkomst uitgevangenschap. Complementairhieraan kunnen Bayesiaanse clustering methodes toelaten om zonder voorkennis groepen van elkaar te onderscheiden. De combinatie van bovenstaande methodes laat ons bovendien toe om een onderscheid te maken tussen recente (geen introgressie) en oudere introducties (introgressie).
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 15 van 57 1.2.3
Genetische identificatie van (illegale) introducties
Dit project heeft als doel het ontwikkelen van een genetische methode die toelaat om te detecteren waar en of patrijzen illegaal uitgezet worden in Vlaanderen. Om dit na te gaan zullen we genetischeverschillen tussen wilde populaties onderlingen tussen wilde populaties en kweekpopulaties onderzoeken. Algemeen zal deze studieons een beterinzichtgeven in het genetische verleden, heden en toekomstvan patrijzen in Vlaanderen. Verdere conclusies over de populatiestructuurkunnen hieruitgetrokken worden. Meer concreet hebben we volgende doelstellingen: Nagaan of en waar er illegaal patrijzen werden uitgezet, met uitrol van een eenvoudige genetische identificatiemethode om routinematig, via steekproeven of gerichte controle, patrijzen toe te wijzen aan referentiegroepen. De genetische diversiteit, structuur en connectiviteit van patrijs in Vlaanderen bepalen, met het oog op evaluatie van de noodzaak tot bijplaatsingen
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 16 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo 1.3 DOELSTELLINGEN
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 17 van 57 2 WERKWIJZE 2.1 STAALNAME 2.1.1 Wildvangst patrijzen Patrijzen ‘in de vrije wildbaan’ werden tijdens twee fases bemonsterd. Beide staalname campagnes gebeurden met medewerking van de lokale jagers. Er zijn dus geen stalen afkomstig uit gebieden waar de lokale jagers niet wilden meewerken aan dit onderzoek.
Tijdens de eerste fase, in oktober 2020, werd via Hubertus Vereniging Vlaanderen (HVV) aan alle wildbeheereenheden (WBE’s) gevraagd om van geschoten patrijzen een poot of teen te bewaren bij 20 °C. Op deze manier werden 326 stalen ingezameld uit 28 WBE’s (Figuur 3, Tabel 1). Om ook stalen te verzamelen van gebieden waarwe geen of onvoldoende stalen van hadden ontvangen, werd er tijdens een tweede fase, van 31 januari tot en met 16 april 2021, een staalnamecampagne georganiseerd om patrijzen in het veld te bemonsteren in samenwerking met HVV en de lokale WBE’s. Deze campagne werd goedgekeurd door de Ethische CommissieDierenwelzijn (ECINBO13). In samenwerkingmetlokale jachtrechthouders werd er een lokgeluid afgespeeld op geschikte locaties. Solitaire hanen, dewelke hierop reageerden, werden vervolgens gevangen. Van de gevangen vogels werd een bloedstaal genomen. De bloedstalen werden vervolgens bewaard in Queen’s Lysis buffer met een verhouding 100 µL bloed in 1000 µL buffer. Het nemen van een bloedstaal werd in dit genetisch onderzoek geprefereerd doorde hoge kwaliteit en kwantiteit aan DNA die op deze manier verzameld kan worden (Guidelines for Taking Samples from Birds and Mammals for DNA Extraction, n.d.). Elk gevangen individu kreegvervolgens een wetenschappelijke metalen ringen werd levend terugvrijgelaten op de locatie van vangst. Op deze manier werden er 169 stalen (168 hanen, 1 hen) uit 30 WBE’s toegevoegd (Figuur 3, Tabel 1). Op basis van het ruipatroon en de slijtage van de twee buitenste handpennen werd de leeftijdsklasse geschat. Van de 169 individuen waren 45 vogels met zekerheid ouder dan 1 jaar, 114 vogels waren jonger dan 1 jaar, en van 10 individuen was de leeftijdsbepaling onzeker. 2.1.2 Kwekerijen Eendagskuikens afkomstiguit een Franse, een Vlaamse en een Nederlandse/Duitse kwekerij werden geëuthanaseerd, alvorens ereen bloed en weefselstaalwerd genomen. Individuen uit Kwekerij 1werden opgedeeld in tweegroepen (Kwekerij 1A en Kwekerij 1B), waarbij patrijzen uit Kwekerij 1A volgens de leverancier speciaal worden gekweekt voor conservatietranslocaties/populatieherstel. Verder werden er op 6 november en 30 oktober 2020, respectievelijk 7en 23 volwassen patrijzen, afkomstiguiteen nietnadergekendeen een gekende Franse kwekerij, in beslag genomen door Natuurinspectie West van het Agentschap voor Natuur en Bos (Tabel 2).
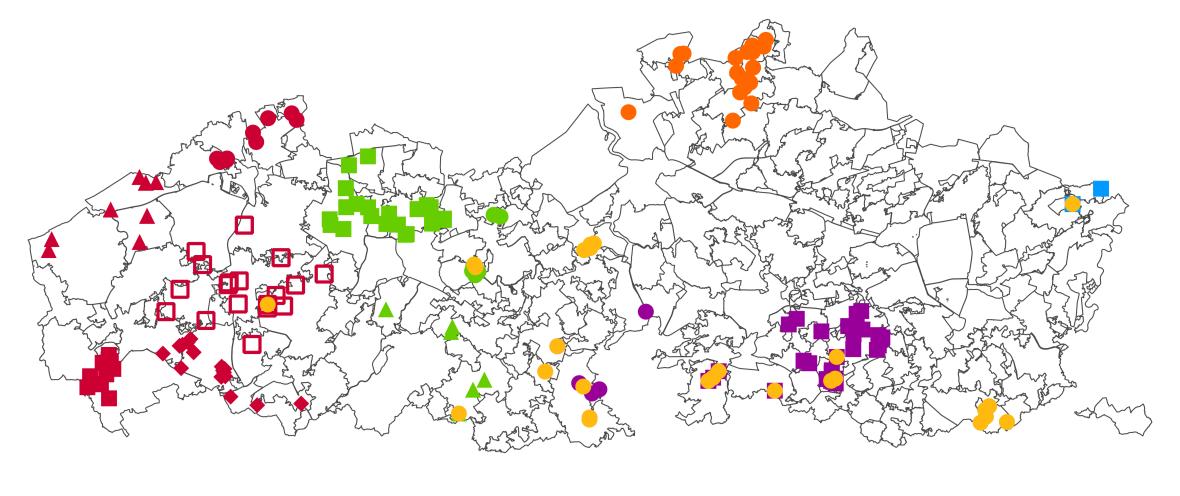
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 18 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo Figuur 3. Overzichtskaarten van alle gevangen patrijs (tijdens beide fases) met de grenzen van de wildbeheereenheden in Vlaanderen. De wildbeheereenheden aangeduid in het grijs hebben in 2020 (a) en 2021 (b) op patrijs gejaagd. ((a)b)




///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 19 van 57 Tabel 1. Overzicht van het aantal wildvangst stalen verzameld door de wildbeheereenheden (WBE’s, aantal jachtstalen) en tijdens de staalnamecampagne (aantal INBO stalen). Alsook het totale aantal wildvangst stalen en het aantal stalen dat uiteindelijk mee werd genomen in de genetische analyses per WBE. De stalenmeegenomen inde genetische analyses zijnstalenwaarvande kwaliteit voldoende hoog was en die vervolgens ook succesvol gegenotypeerd werden. Per WBE werd aangeduid aan welke regio de stalen werden toegewezen in functie van de verdere analyses. WBE Regio Jacht INBO Totaal Geanalyseerd Antwerpse Polders Noord Noorderkempen 1 0 1 1 De Beneden Marck Noorderkempen 4 0 4 4 Essen Noorderkempen 0 3 4 2 Groot Brecht Noorderkempen 57 0 57 13 Maxburg Noorderkempen 14 6 20 17 Noorderkempen Noorderkempen 0 0 1 1 Kinrooi Limburg Noord 0 4 4 3 Molenbeersel Limburg Noord 0 1 1 1 Jekervallei Limburg Zuid 0 2 2 2 Patrijzenkuilen Limburg Zuid 0 4 4 4 Havengebied Gent OostVl Noord 0 14 14 13 Langs de Hoge Kale OostVl Noord 14 5 19 17 Meetjesland Noord OostVl Noord 9 0 9 8 Moerstuiver Damkeuken OostVl Noord 6 0 6 2 Rond Kale, Lieve & Burggrave OostVl Noord 15 0 15 10 Etbos OostVl Oost 8 0 8 7 Scheldevallei OostVl Oost 0 18 18 18 Klaverbeek OostVl Oost 0 9 9 9 De Zavelputten OostVl Zuid 7 0 7 6 Dendervallei OostVl Zuid 21 0 21 19 Scheldeoord vzw OostVl Zuid 4 0 4 4 De Bekkevoortse Oost Brabant 0 5 5 4 Meerdaal Honsem Oost Brabant 0 1 1 1 ROW Oost Brabant 17 6 23 14 Tussen Voer & Yse Oost Brabant 0 5 5 5 Velpedal Oost Brabant 16 4 20 16 Walenbos Oost Brabant 0 2 2 2 Land vanAalst Pajottenland 4 0 4 4 Dendervallei West Pajottenland Pajottenland 0 1 1 1 Nederbrabant Pajottenland 0 2 2 2 Oost Pajottenland Pajottenland 0 12 12 12 De Kemmelbeek Ieper 4 4 8 7 Hoppeland Ieper 0 1 1 1 InFlanders Fields Ieper 10 15 25 24 Achterhaven Zeebrugge Kust Oost 0 2 2 2 Damme Oostkust Kust Oost 11 0 11 7 Paddegat Kust Oost 0 12 12 10 De Middenkustvallei Kust West 22 8 30 25 Tussen Ijzer en Houtland Kust West 9 0 9 6 Westhoek Kust West 0 2 2 2
In een eerste stap werd DNA van alle stalen geëxtraheerd met de DNeasy Blood & Tissue Kit (Qiagen). De concentratie van het geëxtraheerde DNA werd bepaald met de Quant iT PicoGreen dsDNA kit (Thermo Fisher Scientific Inc.). DNA kwaliteit werd gecontroleerd via Agarose gel elektroforese (1% agarose in TAE, 30 min/100 V). De kwaliteit van 67 stalen bleek onvoldoende. Het DNA van deze stalen werd opnieuw geëxtraheerd met de QIAamp DNA micro kit (Qiagen). Van 47 stalen was de kwaliteit na deze nieuwe extractie nog steeds niet voldoende. Al deze stalen waren tenen of pootjes afkomstig van de jacht. Van deze locaties werden wel nogandere stalen meegenomen waarvan de kwaliteitwel goed was. Op basis van de kwaliteit en aantal stalen per WBE, werden uiteindelijk 546 stalen gebruikt voor verdere genetische analyses (Tabel 1, Tabel 2). Deze stalen werden willekeurigverdeeld overzes pools van 90 individuen. Aan elke pool werden telkens zes replicaten toegevoegd.
Als genetische merker werd er gekozen om met Single Nucleotide Polymorphisms (SNPs) te werken. Bijvoorbeeld, in een populatie kan op een bepaalde plaats in het genoom de DNA sequentie 5’ CACTATACCGAG 3’ voorkomen, maar ook 5’ CACTATGCCGAG 3’. De variabele positie is hierA of G (in hetvetaangeduid). In sommige populaties komt de ene variant meer voor dan de andere. Wanneerzulke informatie op honderden tot tienduizenden plaatsen op hetgenoom wordtgecombineerd, krijgje een beeld van de genoomwijdegenetische structuur
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 20 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo ‘t Veld WestVl Centraal 6 1 7 6 Baekeland WestVl Centraal 4 0 4 4 De Hazebeek WestVl Centraal 3 0 3 2 Driekoningen WestVl Centraal 10 0 10 6 Houtland WestVl Centraal 0 3 3 3 Mandelvallei WestVl Centraal 26 0 26 20 De Harelbeek en Heulebeekvallei WestVl Zuid 16 15 31 26 Ieperlee WestVl Zuid 3 0 3 3 Leie en Schelde kouters WestVl Zuid 5 1 6 2 Totaal 326 169 495 378 Tabel 2. Overzicht van het totaal aantal patrijzen per kwekerij en het aantal dat werd meegenomen in de genetische analyses. De stalen meegenomen in de genetische analyses zijn stalen waarvan de kwaliteit voldoende hoog was en die vervolgens ook succesvol gegenotypeerd werden. Afkomst Totaal Geanalyseerd Kwekerij 1A (Frans) 20 19 Kwekerij 1B (Frans) 20 19 Kwekerij 2 (Frans) 7 7 Kwekerij 3 (Belgisch) 30 28 Kwekerij 4 (Nederlands/Duits) 20 13 Kwekerij 5 (Onbekend) 23 18 Totaal 120 104 2.2 GENETISCHE ANALYSES 2.2.1 Sequencing
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 21 van 57 binnen en tussen populaties. Voor het identificeren van SNPs volgden we een Doubl e Digest Genotyping by Sequencing(GBS) protocol, gebaseerd op methodes beschreven in Elshireet al. (2011) en Poland et al. (2012). Van elk individueel staal werd 100 ng DNA gefragmenteerd met behulp van de restrictie enzymen PstI en MseI gedurende 2 uur op 37°C. Vervolgens werden per pool aan elk individueel staal een uniekebarcoded (PstI) en gemeenschappelijke (MseI) adapter geligeerd met behulp van T4 DNA ligase op een temperatuur van 22°C gedurende 60 minuten, gevolgd door 30 minuten op 65°C. Alle stalen werden na ligatie opgezuiverd met behulp van Ampure XP beads (1:1 verhouding). Hiernawerd hetgeknipte DNA geamplificeerd (16 cycli van 30s op 96°C, 20s op 65°C en 30s op 68°C), waarna de stalen opnieuw werden opgezuiverd metbehulp van Ampure XP beads (1:0.8 verhouding). De finale concentratieDNA in elk staal werd bepaald methetQuantiFluorOne ds DNA system (Promega). Gebaseerd op deze concentratie werd van elk van de 96 stalen voor elk van de zes pools 10 ng samengevoegd. Vervolgens werden er voor elk van de zes pools, DNA fragmenten geselecteerd met een grootte tussen 310 en 460 baseparen (zonder adapters) met een 2% agarose cassette op een Blue Pippin toestel (Sage Science). De keuze van de enzymen en de selectie van de fragmenten werd gebaseerd op in silico genome digestion in SimRAD v0.96 in R (Lepais & Weir 2014). De zes pools, die elk 96 stalen (90 individuen + 6 controlestalen) bevatten, werden vervolgens 150 bp paired end gesequenced op een Illumina HiSeq 2500 platform door Admera Health (New Jersey, Verenigde Staten).
2.2.2 Bio informatische data analyse Ruwe DNA sequenties werden geleverd in twee FastQ bestanden per pool. Eerst werd de kwaliteit van de sequenties gecontroleerd met behulp van FastQC software versie 0.11.9 (Andrews 2015). De gepoolde sequenties werden vervolgens gescheiden op basis van de unieke barcodes toegevoegd aan elk individueel staal tijdens de ligatie via de process_radtags module van Stacks versie 2.54 (Catchen et al. 2011, 2013; Rochette et al. 2019). Op deze manierwerden de sequenties opnieuw verzameld per individu. Reads met een uncalled base en lage kwaliteit (Phred score < 10) werden verwijderd. Vervolgens werden de adapter sequenties en overblijfselen van de RE cutsites verwijderd metbehulp van Cutadapt versie 3.5 (Martin 2011). De sequenties werden hierna gemapt ten opzicht van het referentiegenoom van kip (Gallus gallus domesticus), fazant(Phasianus colchicus), rode patrijs (Alectoris rufa) en kalkoen (Meleagris gallopavo) met bowtie2 versie 2.4.1 (Langmead & Salzberg 2012). Beste alignmentrates werden bekomen op basis van hetgenoom van fazant, dewelkewerd gebruikt voor de volgende analyses. De alignment rate varieert tussen 1.39 en 83.80%, met een gemiddelde van 78.66%. Individuen met lage alignment rate (<70%) werden verwijderd. De aanwezigheid van SNPs werd bepaald met de gstacks module van Stacks versie 2.54 en vervolgens een eerstekeergefilterd metde populations module (Paris et al. 2017; Rochette & Catchen 2017; Rochette etal. 2019). Verdere filteringvan individuen en SNPs werd uitgevoerd metVCFtools (Danecek etal. 2011). Individuen metmeerdan 20% afwezige genotypes werden verwijderd. Vervolgens werden enkel SNPs met MAF groter dan 0.01, MAC groter dan 3, missingness (uitval van loci) lager dan 20% en sequencing depth van minstens 8 behouden.
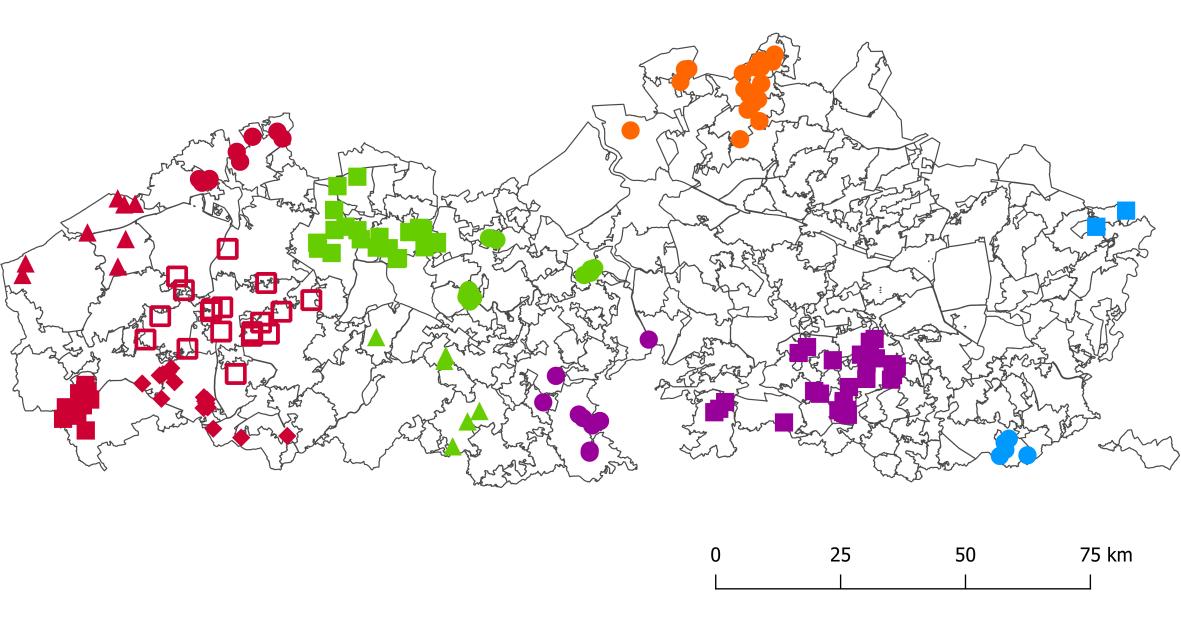
De data analyse werd uitgevoerd in twee stappen. Alle individuen gevangen in de ‘vrije wildbaan’ (zie 2.1.1), vanaf nu aangeduid als wildvangst individuen, werden opgedeeld op basis van regio, zoals weergegeven in Figuur 4 en Tabel 1. In de eerste stap werden alle wildvangst en kweekindividuen samen geanalyseerd. De term weekindividuen refereert naar de stalen genomen uitkwekerijen (zie 2.1.2). In een tweede stap werden vervolgens, op basis van de resultaten en de toewijzingen, de stalen geïdentificeerd als wildvangst maar afkomstig uitkweek en de kweekindividuen verwijderd Alle analyses in deze stap werden vervolgens uitgevoerd voordeze autochtone groep. Alle analyses werden uitgevoerd in R versie 4.0.2 (R Core Team 2021). Figuur 4. Overzicht van alle wildvangst (geschoten en eigen vangst) patrijzen. Kleuren en symbolen geven deopdeling in regio’s, zoals gebruikt voor de verdere analyses, weer.
Na kwaliteitscontrole varieerde het aantal reads per pool tussen 537 951 948 (Pool 1) en 713 315 417 (Pool 4). Het aantal reads per individu varieerde tussen 10 156 en 28 206 569, met een gemiddeldevan 6 059 954 reads. Negen individuen metminderdan 500 000 reads werden verwijderd. Vervolgens werden 20individuen metalignmentrate lagerdan 70% verwijderd om dat deze in een volgende fase de analyses bemoeilijken. Van de zes controlestalen, die aan elke pool werden toegevoegd, werd telkens hetstaal methet meeste aantal reads behouden; 30 stalen werden dus verwijderd. Vervolgens werden ernog 35 individuen verwijderd waarvan meer dan 20% van de SNPs niet gegenotypeerd werden. De finale dataset bedraagt 30 194 SNPs gesequeneerd in 482 individuen afkomstiguit49 WBE’s en vijf kwekerijen (Tabel 1, Tabel 2). Gemiddelde read depth per individu is 14.431x (range = 5.289 60.796). Gemiddelde missingness is 3.8% per individu (range = 1.3% 23.6%). 2.2.3 Data-analyse
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 22 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo



Om een beeld te krijgen van de invloed van geografische afstand op genetische afstand tussen wilde individuen, werd een Mantel correlogram opgesteld met behulp van R package Vegan versie 2.3 5 (Oksanen etal. 2007). In een correlogram wordtde correlatie tussen genetische en geografische afstand weergegeven voor een bepaald aantal afstandsklassen. In deze dataset werd gekozen voor afstandsklassen van 2 km. Toewijzingen
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 23 van 57
Om individuen toe te wijzen aan groepen, werd een Discriminant Analysis of Principal Components (DAPC) uitgevoerd. DAPC transformeert de data eerst met behulp van een PCA, waarna het aantal clusters en toewijzingen van individuen aan deze clusters wordt bepaald met behulp van Discriminant Analysis (DA). Deze analyse werd uitgevoerd zonder priorinformatie overhetaantal groepen op basis van het K means algoritme. Hierbij wordthet optimaal aantal clusters (K) geschat op basis van het Bayesian Information Criterion (BIC). Vervolgens werd elk individu toegewezen tot één van de clusters (Jombart & Collins 2015). Aansluitend werd een STRUCTURE analyse uitgevoerd met R package ParallelStructure versie 1.0 (Besnier&Glover 2013; Falush et al. 2003; Pritchard et al. 2000). Net als bij een DAPC gaat het STRUCTURE algoritme op zoek naar genetische clusters (K) in de dataset. Dit kan zowel zonderals metprior informatie overhetaantal groepen. Vervolgens wordtvoorelk individu de afkomstvan elk van deze groepen bepaald. STRUCTURE laat beter dan DAPC toe om (recente en minderrecente) genetische vermengingtussen populaties te kwantificeren. De STRUCTURE analyse werd drie keer uitgevoerd. Hierbij gebruikten we een admixture model zonder prior informatie overhetaantal groepen (burn in = 10 000, iteraties = 100 000). Het optimaal aantal clusters K werd daarna bepaald op basis van DeltaK en loglikelihood metbehulp van Structure Harvester v0.6.94 (Earl & vonHoldt 2012).
Genetische diversiteit De genetische diversiteit werd uitgedrukt aan de hand van de gemiddelde waargenomen heterozygositeit (HO), die per locus 0 of 1 kan zijn (het locus is wel of niet variabel binnen dat individu). Op niveau van een groep van individuen (“populatie”) werd de HE berekend (hetaandeel verwachte heterozygotegenotypes onder Hardy Weinberg evenwicht) en de gemiddelde waargenomen heterozygositeit (HO). Dit werd berekend met behulp van R package Hierfstat versie 0.5 7 (Goudet 2005). HO en HE kunnen waardes aannemen tussen 0 (geen genetische variatie, alle loci of individuen zijn homozygoot) tot 1 (maximale genetische variatie, alle loci of individuen zijn heterozygoot). Onder normale omstandigheden is de maximale gemiddelde waarde van HE = 0.5 vooreen SNP locus mettwee allelen, wanneer de twee allelen een gelijke frequentie hebben in de populatie. Genetische structuur De genetische structuurwerd visueel weergegeven metbehulp van een PCA plot(Principal ComponentAnalysis) metRpackage ade4 versie 1.7 18. PCA is een techniek die toelaatom de hoogdimensionelecomplexedatate reduceren totenkeledimensies die het gros van de structuur in de data weergeven. Op een PCA plot clusteren individuen met een gelijkaardige genetische samenstellingsamen. Vervolgens werd de fixatie index(FST), een maat voor genetische afstand tussen groepen, berekend met R package Hierfstat versie 0.5 7 (Goudet 2005).
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 24 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo 3 RESULTATEN 3.1 STAP 1: GENETISCHE DIVERSITEIT EN STRUCTUUR VAN ALLE INDIVIDUEN In deze eerste stap werden alle 482 (zowel wildvangst als kweek) individuen, die succesvol gegenotypeerd werden, opgenomen in de analyse. Genetische diversiteit De waargenomen heterozygositeit HO varieerde tussen 0.231 (Regio Noorderkempen) en 0.247 (Regio Limburg Zuid) voor de wildvangst populaties, en tussen 0.239 (Kwekerij 4) en 0.250 (Kwekerij 5) voor de kweekpopulaties (Tabel 3). Verwachte heterozygositeitHE verschilde nietsignificantvan waargenomen heterozygositeit HO (Tabel 3). Tabel 3. Waargenomen en verwachte heterozygositeit (HO en HE) en het aantal individuen (n) voor elk van de wildvangst groepen en de kweekpopulaties. Regio n HO HE Noorderkempen 38 0.231 0.224 Limburg Noord 4 0.238 0.235 Limburg Zuid 6 0.247 0.238 OostVl Noord 50 0.230 0.226 OostVl Oost 34 0.237 0.236 OostVl Zuid 29 0.231 0.225 Pajottenland 19 0.237 0.233 Oost Brabant 42 0.242 0.234 Ieper 32 0.234 0.229 Kust Oost 19 0.229 0.223 Kust West 33 0.232 0.224 WestVl Centraal 41 0.235 0.227 WestVl Zuid 31 0.232 0.228 Kwekerij 1A (Frans) 19 0.244 0.238 Kwekerij 1B (Frans) 19 0.244 0.236 Kwekerij 2 (Frans) 7 0.248 0.235 Kwekerij 3 (Belgisch) 28 0.243 0.237 Kwekerij 4 (Nederlands/Duits) 13 0.239 0.231 Kwekerij 5 (Onbekend) 18 0.250 0.237 Genetische structuur De kweekindividuen clusteren samen en worden duidelijk gescheiden van de autochtone individuen op basis van PC1 (Figuur5a). Binnen deze groep clusteren enkel de stalen afkomstig uit Kwekerij 1A licht apart van de andere kweekindividuen Wildvangst stalen afkomstiguitregio Noorderkempen clusteren vervolgens apart op basis van PC2 van de andere wildvangst en kweekstalen en lijken een genetische groep te vormen. Op basis van Figuur 5b kunnen we duidelijk zien welke wildvangst individuen clusteren met de kweekindividuen. Dit zijn 47 individuen afkomstig uit de regio’s Oost Brabant (n = 20),
Figuur 5. Ordinatieplot (PCA) waarbij de symbolen de verschillende patrijzenindividuen voorstellen. De kleuren en symbolen geven aan tot welke groepen deze individuen behoren. Hoe dichter de afstand tussen individuen op de plot, hoe meer gelijkend deze zijn qua genetische samenstelling. (a) Overzicht van alle (wildvangst en kweek) individuen. (b) Detail van de ordinatie, gefocust op dat deel dat overeenkomt met de kweekpopulaties (a) (b)
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 25 van 57 Pajottenland (n = 7), Noord en Zuid Limburg (n = 2 en 6) en het Oosten en Zuiden van Oost Vlaanderen (n = 9 en 2). De exacte afkomst van deze individuen is toegevoegd als bijlage (Bijlage 2). Gelijkaardige resultaten worden gevonden op basis van genetische differentiatie (gemeten als paarsgewijze FST) tussen de regio’s (Tabel 4). Overhetalgemeen is de genetische differentiatie laag (overall FST = 0.022, range = 0 0.042).



///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 26 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo Tabel 4. Paarsgewijze genetische afstand (op basis van FST) tussen de verschillende wildvangst en kweekpopulaties. Noorderkempen Limburg-Noord LimburgZuid OostVlNoord OostVlOost OostVlZuid Pajottenland BrabantOost- Ieper OostKust WestKust CentraalWestVl WestVlZuid 1AKwekerij 1BKwekerij 2Kwekerij 3Kwekerij 4Kwekerij 5Kwekerij Noorderkempen 0.026 0.040 0.033 0.030 0.039 0.033 0.031 0.035 0.037 0.040 0.035 0.034 0.036 0.042 0.043 0.041 0.040 0.040 Limburg - Noord 0.026 - 0.003 0.020 0.007 0.023 0.010 0.008 0.021 0.025 0.030 0.022 0.019 0.007 0.004 0.005 0.004 0.004 0.006 Limburg Zuid 0.040 0.003 0.033 0.015 0.035 0.018 0.014 0.033 0.037 0.042 0.032 0.030 0.008 0.001 0.000 0.002 0.000 0.001 OostVl - Noord 0.033 0.020 0.033 0.016 0.024 0.021 0.024 0.019 0.016 0.024 0.017 0.019 0.029 0.035 0.035 0.035 0.034 0.033 OostVl Oost 0.030 0.007 0.015 0.016 0.021 0.009 0.013 0.019 0.021 0.026 0.019 0.017 0.015 0.017 0.018 0.018 0.017 0.016 OostVl - Zuid 0.039 0.023 0.035 0.024 0.021 0.023 0.027 0.026 0.028 0.032 0.025 0.025 0.031 0.036 0.036 0.036 0.035 0.034 Pajottenland 0.033 0.010 0.018 0.021 0.009 0.023 0.016 0.022 0.025 0.030 0.023 0.021 0.018 0.019 0.019 0.019 0.018 0.018 Oost-Brabant 0.031 0.008 0.014 0.024 0.013 0.027 0.016 0.025 0.027 0.032 0.025 0.022 0.014 0.015 0.016 0.016 0.015 0.015 Ieper 0.035 0.021 0.033 0.019 0.019 0.026 0.022 0.025 0.021 0.024 0.018 0.014 0.029 0.035 0.035 0.035 0.034 0.033 Kust Oost 0.037 0.025 0.037 0.016 0.021 0.028 0.025 0.027 0.021 0.021 0.019 0.020 0.031 0.039 0.039 0.039 0.038 0.037 Kust - West 0.040 0.030 0.042 0.024 0.026 0.032 0.030 0.032 0.024 0.021 0.022 0.024 0.038 0.043 0.043 0.043 0.042 0.041 WVl - Centraal 0.035 0.022 0.032 0.017 0.019 0.025 0.023 0.025 0.018 0.019 0.022 0.016 0.028 0.035 0.035 0.034 0.034 0.033 WestVl - Zuid 0.034 0.019 0.030 0.019 0.017 0.025 0.021 0.022 0.014 0.020 0.024 0.016 0.025 0.032 0.031 0.032 0.030 0.030 Kwekerij 1A 0.036 0.007 0.008 0.029 0.015 0.031 0.018 0.014 0.029 0.031 0.038 0.028 0.025 0.012 0.011 0.011 0.011 0.011 Kwekerij 1B 0.042 0.004 0.001 0.035 0.017 0.036 0.019 0.015 0.035 0.039 0.043 0.035 0.032 0.012 0.002 0.001 0.001 0.000 Kwekerij 2 0.043 0.005 0.000 0.035 0.018 0.036 0.019 0.016 0.035 0.039 0.043 0.035 0.031 0.011 0.002 0.001 0.002 0.001 Kwekerij 3 0.041 0.004 0.002 0.035 0.018 0.036 0.019 0.016 0.035 0.039 0.043 0.034 0.032 0.011 0.001 0.001 0.001 0.001 Kwekerij 4 0.040 0.004 0.000 0.034 0.017 0.035 0.018 0.015 0.034 0.038 0.042 0.034 0.030 0.011 0.001 0.002 0.001 0.002 Kwekerij 5 0.040 0.006 -0.001 0.033 0.016 0.034 0.018 0.015 0.033 0.037 0.041 0.033 0.030 0.011 0.000 0.001 0.001 0.002 -
clusters.
Toewijzingen Hetoptimaal aantal clusters K dat wordt waargenomen, wordt op basis van de DAPC resultaten in deze stap geschat op twee. Hierbij worden de kweekpopulaties duidelijk onderscheiden van de autochtone populaties. De membership probability is voor elk individu hoog(> 99%) en 47 wildvangst individuen uit Noord en Zuid Limburg (n = 2 en 6), het Oosten en Zuiden van Oost Vlaanderen (n = 9 en 2), Oost Brabant (n = 20) en Pajottenland (n = 7) worden methoge waarschijnlijkheid toegewezen aan de groep metkweekindividuen (Tabel 5). Deze individuen zijn dezelfde als in de PCA plotclusteren metde kweekpopulaties (Bijlage 2). Tabel 5. Totaal aantal individuen (n) per regio in de genetische dataset, en het aantal wildvangst individuen dat geïdentificeerd werd als afkomstig uit kweek door de PCA, DAPC en STRUCTURE analyse. In de STRUCTURE analyse maken we een onderscheid tussen zuivere kweekindividuen en een (gekruiste) nakomeling. Tussen haakjes wordt het percentage weergegeven van het aantal individuen geïdentificeerd als afkomstig uit een kwekerij ten opzichte van het totaal geeftaan wat de op basis van de waargenomen Wanneerdeze waarde voormeerdan 70% tot de cluster met kweekindividuen behoort, gaan we ervan uit datdit individu een rechtstreekse afstammeling is van een kweekpopulatie. De drempel van 70% werd gekozen omdat dit de gemiddelde toewijzingswaarde is die wordt waargenomen bij de kweekindividuen van kwekerij1A aan de andere kweekpopulatie Verder is er voor alle individuen een beetje statistische ruis in de toewijzing aan groepen als gevolg van regionale genetisch verschillen. Afstamming uit kweek stelden we hier vast voor 35 individuen, dewelke ook eerdergeïdentificeerd worden doorde PCA en DAPC als afkomstig uit een kwekerij (Tabel 5). Een waarde tussen 50 en 70% geeft dan weer aan dat een individu waarschijnlijk een nakomeling is van een gekweekte patrijs. In totaal werden 13 individuen geïdentificeerd als afstammeling in regio’s Oost Brabant (n = 11) en Pajottenland (n = 2). Dit
oorsprongvan elk individu is
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 27 van 57
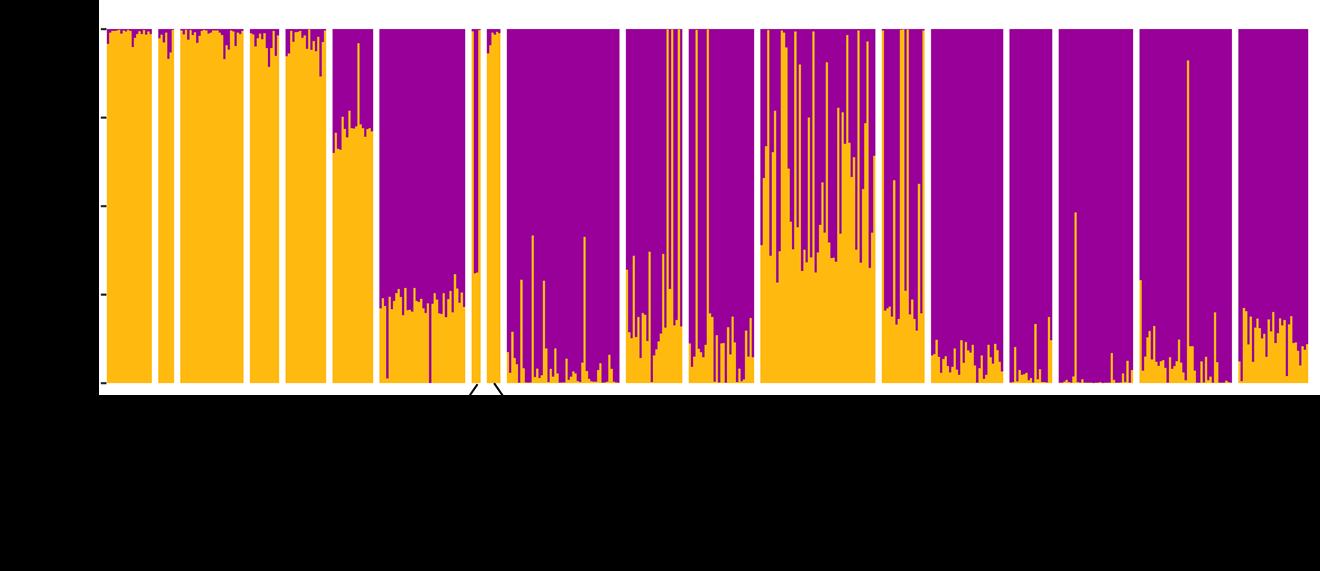
aantal individuen in die regio Regio n PCA DAPC STRUCTUREZuiver NakomelingenSTRUCTURE Noorderkempen 38 0 (0%) 0 (0%) 0 (0%) 0 (0%) Limburg Noord 4 2 (50%) 2 (50%) 2 (50%) 0 (0%) Limburg Zuid 6 6 (100%) 6 (100%) 6 (100%) 0 (0%) OostVl Noord 50 0 (0%) 0 (0%) 0 (0%) 0 (0%) OostVl Oost 34 9 (26.5%) 9 (26.5%) 9 (26.5%) 0 (0%) OostVl Zuid 29 2 (6.9%) 2 (6.9%) 2 (6.9%) 0 (0%) Pajottenland 19 7 (36.8%) 7 (36.8%) 5 (26.3%) 2 (10.5%) Oost Brabant 42 20 (47.6%) 20 (47.6%) 10 (23.8%) 11 (26.2%) Ieper 32 0 (0%) 0 (0%) 0 (0%) 0 (0%) Kust Oost 19 0 (0%) 0 (0%) 0 (0%) 0 (0%) Kust West 33 0 (0%) 0 (0%) 0 (0%) 0 (0%) WestVl Centraal 41 1 (2.4%) 1 (2.4%) 1 (2.4%) 0 (0%) WestVl Zuid 31 0 (0%) 0 (0%) 0 (0%) 0 (0%) Totaal 378 47 (12.4%) 47 (12.4%) 35 (9.3%) 13 (3.4%) STRUCTURE identificeerde, net als DAPC, twee genetische clusters (Figuur 6), waarbij één clusteralle kweekindividuen bevat en de andere cluster de meeste autochtone stalen. De Q waarde
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 28 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo
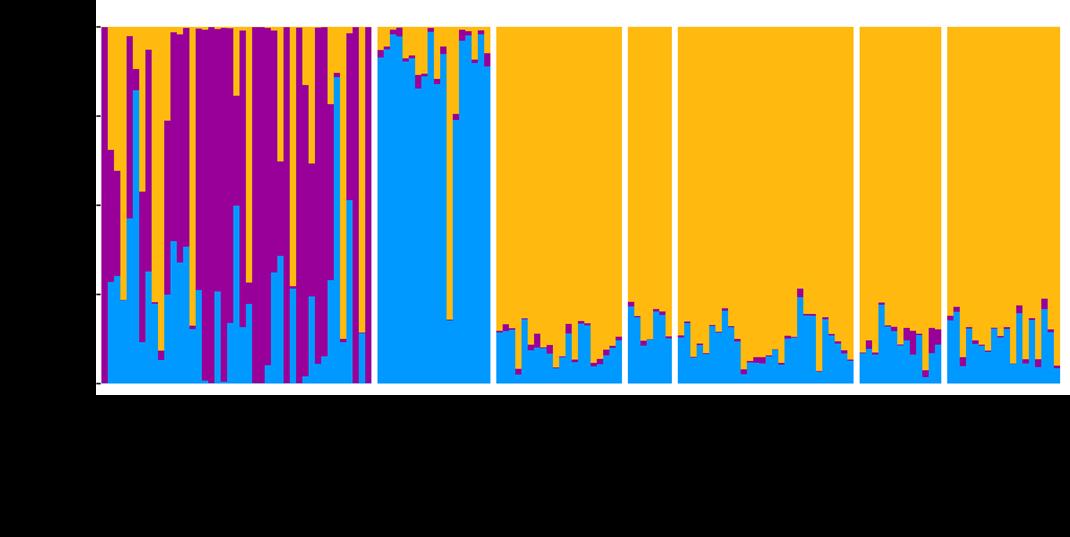
Om een beterzicht te krijgen op de afkomst van de wildvangst stalen in Vlaams Brabant, werd een aparte STRUCTUREanalyse uitgevoerd metde wildvangst patrijzen uit Vlaams Brabant en de kweekpopulaties. Deze analyse onderscheidt wel individuen uit Kwekerij 1A als een andere herkomst (op één exemplaar na dat toch afkomstig lijkt uit de andere kwekerij), en we zien verder ook dat individuen uit Vlaams Brabant toegewezen worden aan (1) Kwekerij 1A, (2) aan andere kwekerijen, (3) aan de autochtone populatie, of (4) aan een mengsel van deze drie herkomsten. Ditgeeftnietnoodzakelijk weer dat er frequente hybridisatie optreedt tussen verschillende kweekstammen in de vrije wildbaan; het is ook mogelijk dat sommige kwekers (niet door ons bemonsterd) zelf een mengsel gebruiken van deze twee herkomsten.
Figuur 6. Q waarde berekend voor elk individu (weergegeven door één staaf) met behulp van STRUCTURE software. Deze waarde geeft de admixture proportie van elk individu weer. STRUCTURE identificeerde twee genetische clusters (geel = kweek, paars = wild). Een Q waarde van meer dan 70% wordt geïnterpreteerd als een zuivere afstammeling van de cluster.
Figuur 7. Q waarde berekend voor elk individu (weergegeven door één staaf) voor de wildvangst stalen uit Vlaams Brabant en de kweekpopulaties. STRUCTURE identificeerde drie genetische clusters, weergegeven door de drie kleuren.
zijn waarschijnlijk afkomstig van genetische vermenging tussen autochtone wilde en gekweektepatrijzen.
Van deze 13 individuen zijn er12 die eerderwerden toegewezen aan de kweekpopulatie door PCA en DAPC (Tabel 5).
Figuur 8. Overzicht van alle wildvangst patrijzen met in het geel aangeduid welke individuen werden geïdentificeerd als (a) zuivere kweekindividu en (b) nakomeling van een kweekindividu.
Wildvangst individuen (n = 35) die door elke methode (PCA, DAPC, STRUCTURE) geïdentificeerd werden als afkomstig uit een gekweekte populatie worden weergegeven in Figuur 8a. Bovendien werden er door STRUCTURE 13 individuen als nakomelingen van gekweekteen wilde individuen gedetecteerd regio’s Oost Brabanten Pajottenland (Figuur8b).
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 29 van 57
((a)b)


Genetische diversiteit Waargenomen heterozygositeitvan de autochtone populaties varieert tussen 0.229 (Regio Kust Oost) en 0.242 (Oost Brabant) en verschilt niet significant van verwachte heterozygositeit (Tabel 6). Heterozygositeit is over het algemeen lager na het verwijderen van kweekindividuen en wildvangstindividuen die in voorgaande analyses werden toegewezen als afkomstiguitkweekpopulaties, maardit effect is verwaarloosbaar (range = 0 0.007). Tabel 6. Waargenomen en verwachte heterozygositeit (HO en HE), en het aantal individuen (n) per groep voor alle individuen en voor alle autochtone individuen (exclusief kweekindividuen wildvangst individuen die in voorgaande analyses werden toegewezen als afkomstig uit kweekpopulaties). In het vet staat aangeduid uit welke groepen individuen afkomstig uit kweek werden geïdentificeerd.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 30 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo 3.2 STAP 2: GENETISCHE DIVERSITEIT EN STRUCTUUR VAN AUTOCHTONE POPULATIES
Totaal Autochtoon Regio n HO HE n HO HE Noorderkempen 38 0.231 0.224 38 0.231 0.224 Limburg Noord 4 0.238 0.235 2 0.231 0.223 Limburg Zuid 6 0.247 0.238 0 OostVl Noord 50 0.230 0.226 50 0.230 0.226 OostVl Oost 34 0.237 0.236 25 0.233 0.231 OostVl Zuid 29 0.231 0.225 27 0.230 0.224 Pajottenland 19 0.237 0.233 12 0.232 0.226 Oost Brabant 42 0.242 0.234 21 0.242 0.229 Ieper 32 0.234 0.229 32 0.234 0.229 Kust Oost 19 0.229 0.223 19 0.229 0.223 Kust West 33 0.232 0.224 33 0.232 0.224 WestVl - Centraal 41 0.235 0.227 40 0.235 0.227 WestVl Zuid 31 0.232 0.228 31 0.232 0.228 Genetische structuur De autochtone individuen uitde regio Noorderkempen vormen zoals in de vorige analyses een duidelijke aparte cluster(Figuur9). Verderclusteren enkele individuen uit de regio centraal West Vlaanderen apart. Al deze individuen zijn afkomstig uit één WBE (Bijlage 3). FST waardes variëren tussen 0.014 en 0.042 (overall FST = 0.026). Genetische differentiatie is het grootst tussen de Noorderkempen en Oost Vlaanderen Zuid (Tabel 7). Zowel DAPC als STRUCTURE (Figuur10, Figuur11) bevestigen dit resultaat en geven aan de er
In hettweede deelvan de analyses werden alle individuen die gevangen werden in het wild, maar in voorgaande analyses werden toegewezen als afkomstig uit kweekpopulaties (n = 48) verwijderd om zo een beeld te krijgen van de populatiegenetische structuurvan de autochtone patrijzen in Vlaanderen. Metuitzonderingvan Noord en Zuid Limburg(n = 2 en 0), bleven er in elke regio nog voldoende individuen over. Ook de kweekindividuen (n = 104) werden verwijderd uitdeze dataset. Uiteindelijk bleven er autochtone 330 individuen over (Tabel 6).
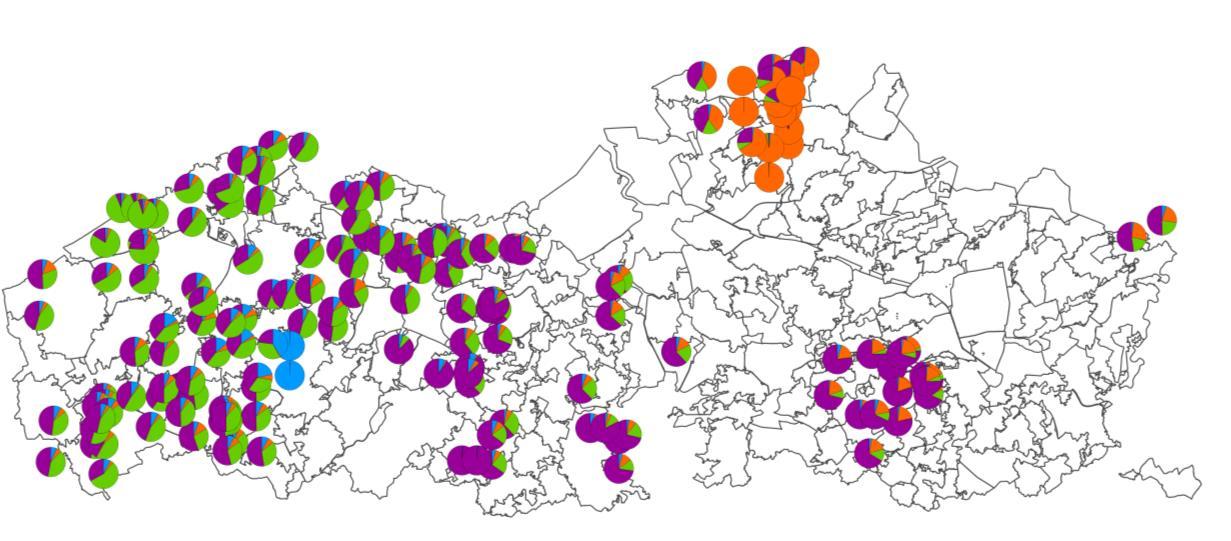
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 31 van 57 twee of drie genetischeclusters zijn:de Noorderkempen, een clustermetenkele individuen uit West Vlaanderen en alle andere regio’s. Genetische en geografische afstand tussen individuen is het sterkst gecorreleerd bij korte afstanden (<2 km). Deze correlatie is significantmaar veel minder sterk voor afstanden tot 20 km (Figuur 12).
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 32 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo Tabel 7. Paarsgewijzegenetische afstand(opbasis vanFST)tussen de verschillendeautochtone groepen. Noorderkempen LimburgNoord OostVlNoord OostVl-Oost OostVlZuid Pajottenland BrabantOost- Ieper OostKust WestKust- CentraalWestVl WestVlZuid Noorderkempen 0.029 0.033 0.034 0.042 0.041 0.036 0.035 0.037 0.040 0.036 0.034 Limburg - Noord 0.029 0.024 0.022 0.033 0.030 0.028 0.026 0.030 0.035 0.028 0.024 OostVl Noord 0.033 0.024 0.018 0.026 0.027 0.029 0.019 0.016 0.024 0.018 0.019 OostVl - Oost 0.034 0.022 0.018 0.026 0.021 0.025 0.022 0.024 0.029 0.023 0.020 OostVl Zuid 0.042 0.033 0.026 0.026 0.032 0.036 0.028 0.030 0.034 0.027 0.026 Pajottenland 0.041 0.030 0.027 0.021 0.032 - 0.033 0.028 0.032 0.036 0.030 0.028 Oost Brabant 0.036 0.028 0.029 0.025 0.036 0.033 0.03 0.032 0.037 0.031 0.027 Ieper 0.035 0.026 0.019 0.022 0.028 0.028 0.03 0.021 0.024 0.019 0.014 Kust Oost 0.037 0.030 0.016 0.024 0.030 0.032 0.032 0.021 0.021 0.020 0.020 Kust - West 0.040 0.035 0.024 0.029 0.034 0.036 0.037 0.024 0.021 - 0.023 0.024 WestVl - Centraal 0.036 0.028 0.018 0.023 0.027 0.030 0.031 0.019 0.020 0.023 0.016 WestVl - Zuid 0.034 0.024 0.019 0.020 0.026 0.028 0.027 0.014 0.020 0.024 0.016
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 33 van 57 Figuur 9 Ordinatie plot (PCA) waarbij de symbolen de autochtone individuen voorstellen. De kleuren en symbolen geven aan tot welke groepen deze individuen behoren. Hoe dichter de afstand tussen individuen,hoe meer gelijkendqua genetische samenstelling.(a)Overzichtvanalleindividuen.(b)Detail van de cluster waartoe de meeste individuen behoren. (a) (b)



///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 34 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo Figuur 10. Q waardeberekend voor elk autochtoon individu(weergegeven door één streep) met behulp vanSTRUCTURE software. Deze waarde geeft de admixture proportie vanelk individuweer voor (a) K = 2, (b) K = 3 (c)en K = 4. (a) (b) (c)


///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 35 van 57 Figuur 11. Q waarde berekend voor elk autochtoon individu (weergegeven door één taartdiagram per individu) met behulp van STRUCTURE software. Deze waarde geeft de admixture proportie van elk individu weer voor (a) K = 2, (b) K = 3 (c) en K = 4. (a) (b) (c)



///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 36 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo Figuur 12. Spatiële autocorrelatie tussen genotypes. Dit geeft de correlatie weer tussen genetische en geografische afstanden tussen de wilde individuen voor afstandsklassen die telkens 2 km van elkaar verschillen. Volle vierkantjes geven een significante (positieve of negatieve) correlatie tussen beide weer, holle vierkantjes geven een niet significante waarde weer.

Bovendien verschilde de geobserveerdeheterozygositeit(HO) niet van de heterozygositeit die men verwacht onder Hardy Weinberg evenwicht (i.e. verwachte heterozygositeit, HE). Geobserveerde heterozygositeitkan lager zijn dan verwachte heterozygositeit als een gevolg van populatiestructuur (non random paarvorming binnen het beschouwde staal), inteelt, recente bottlenecks, kleine populatiegroottes en/of een Wahlund effect (i.e. wanneer populaties met een verschillende genetische samenstelling vermengd worden).
Geobserveerde en verwachte heterozygositeit wordt vaak gebruikt als maat voor de genetische diversiteitvan populaties (Frankham etal. 2002). Een individu is heterozygoot voor een bepaald locus als voor dit specifieke locus twee verschillende allelen (of varianten) aanwezig zijn. In dit geval wordt elk van de > 30 000 geanalyseerde SNPs beschouwd als een locus. De geobserveerde heterozygositeitwordtdan berekend als het percentage van loci dat heterozygoot zijn in een bepaalde populatie of regio. Voor de wilde patrijzen varieerde de geobserveerde heterozygositeit tussen 0.229 (Regio Kust Oost) en 0.242 (Oost Brabant). De verschillen in genetische diversiteit tussen de verschillende regio’s zijn verwaarloosbaar (grootste verschil = 0.013). Genetische diversiteit was bovendien maar een fractie hoger (maximale verschilvan 0.018) in kweekpopulaties ten opzichte van de autochtone populaties.
Deze resultaten tonen aan dat diversiteit in geen enkele regio lager is en dat Vlaamse autochtone patrijzenpopulaties niet meetbaar te lijden hebben onder inteelt, ook niet in regio’s waardensiteiten lagerzijn. Translocaties of gecontroleerde bijplaatsingen met als doel de nadelige invloeden van lage genetische diversiteit en inteelt tegen te gaan, lijken dus niet nodig te zijn voor de instandhouding van patrijzenpopulaties in Vlaanderen (Weeks et al. 2011)
Het doel van ditrapport was om enerzijds een methodiek op punt te stellen die het mogelijk maakt om wilde patrijzen van gekweekte individuen te onderscheiden, en anderzijds om de populatie genetische structuurvan patrijs in Vlaanderen te bestuderen (met oog op evaluatie van de noodzaak tot translocaties). In de discussie overlopen we eerst patronen van genetische diversiteit in populaties van autochtone, wildvangst en kweekindividuen, en de noodzaak vooreventuelemaatregelen. Vervolgens bespreken we de genetische structuur van autochtone, wildvangst en kweekindividuen. Tot slot focussen we op de mogelijkheden om gekweekte van autochtone individuen te onderscheiden en de ontwikkeling van een genetische methodeom ditonderscheid in de toekomstop een routinematige wijze te kunnen maken. EVALUATIE VAN DE GENETISCHE DIVERSITEIT
4.1
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 37 van 57 4 DISCUSSIE
Met een gemiddeldeFST waarde van 0.022 voorzowel alle wildvangst en kweekindividuen, en een gemiddelde FST waarde van 0.026 voor de autochtone populaties, lijkt de genetische differentiatie van patrijs over het algemeen vrij laag in Vlaanderen. F ST (de Fixatie index van subpopulaties ten opzichte van de totale populatie) wordt gebruikt als maat voor genetische differentiatie tussen populaties en varieert tussen 0 (één grote homogene populatie) en 1 (populaties verschillen volledig). 4.2.1 Genetische afstand tussen wilde en kweekpopulaties Overhet algemeen kan ereen duidelijk onderscheid gemaaktworden tussen kweekindividuen en autochtone individuen. Hetverschil tussen beide is zichtbaarop de PCA plotdoordat er een afzonderlijkeclustermetkweekindividuen aanwezig is. Bovendien is genetische afstand over het algemeen groter tussen de wilde (zowel autochtone als wildvangst) populaties en de kweekpopulaties, dan tussen wilde populaties onderling (zowelautochtoon als wildvangst). De gemiddelde genetischedifferentiatietussen de kweekpopulaties en wildvangst individuen is het grootst in de regio’s Noorderkempen (0.040), Kust West (0.042), Kust Oost (0.037), Ieper (0.034), West Vlaanderen Centraal (0.033) en Zuid (0.030), Oost Vlaanderen Noord (0.034) en Zuid (0.035). Dit verschil is beduidend kleiner in regio Oost Vlaanderen Oost (0.017), regio Pajottenland (0.019), regio Oost Brabant (0.015) en nagenoeg afwezig in regio’s Limburg Noord (0.005) en Zuid (0.001). In dit laatste geval is dit omdat nagenoeg alle individuen afkomstig lijken te zijn van uitzettingen vanuit kweekpopulaties. In Noord Franse populaties van patrijs wordt een genetische differentiatie van gemiddeld 0.025 gerapporteerd tussen kweek en wilde populaties (Bech et al. 2020). In deze studie was het verschil bovendien groter tussen wilde populaties onderling, dan tussen wilde en kweekpopulaties. Deze observatie valt eenvoudig te verklaren door de geografische afstand die veel groteris tussen wildepopulaties in Frankrijk. De broeddieren die gebruikt worden in Franse kwekerijen zijn afkomstig van wilde populaties uit Noord en West Frankrijk. Dit zou ook deels kunnen verklaren waarom de genetische afstand met Vlaamse patrijzen, ondanks dat deze duidelijk aanwezig is, eerder beperkt is.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 38 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo 4.2 EVALUATIE VAN DE GENETISCHE STRUCTUUR
Alle kweekpopulaties, met uitzondering van individuen uit Kwekerij 1A, clusteren duidelijk samen en vormen één homogene genetische groep, zoals bevestigd door de F ST waardes. Hieruit kunnen we afleiden dat zowel de Franse als de Nederlandse/Duitse en Belgische kweekindividuen waarschijnlijk dezelfde oorspronghebben. Eén van de mogelijke verklaringen hiervoor kan liggen bij het feit dat enkele grote commerciële kwekerijen uit het westen van Frankrijk eieren exporteren naar verschillende kleinere kwekerijen binnen, maar ook buiten Frankrijk. Een andere mogelijkeverklaring kan zijn dat meerdere kwekerijen gebruik hebben gemaaktmaken van eenzelfde ancestrale setbroeddieren, afkomstigvan wildepatrijzen in het noorden van Frankrijk. Hierdoor kunnen we over het algemeen geen onderscheid maken tussen individuen afkomstig uit verschillende kwekerijen en kunnen we enkel aantonen wanneer een wildvangst individu, afkomstig is uit een kweekpopulatie. Deze studie had als
4.3.1 Toelichting en validatie van methodiek Frankrijk is binnen Europa de belangrijkste leverancier van gekweekte patrijzen. Hiervoor worden grotendeels of uitsluitend dieren van Franse oorspronggebruikt (Bech et al. 2020). We kunnen dan ook genetische verschillen verwachten metBelgischepatrijzenpopulaties, op basis van een gebrek aan genmigratie over grote geografische afstanden. Het kweken van
4.3 RECENTE EN MINDER RECENTE UITZETTINGEN VAN PATRIJS IN VLAANDEREN
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 39 van 57 doel om te kijken of het mogelijk is een onderscheid te maken tussen autochtone en kweekpopulaties, maarnietnoodzakelijk om gekweekte individuen toe te wijzen aan bepaalde kwekerijen. In een latere fase is het mogelijk om individuen uit andere kwekerijen te genotyperen om te onderzoeken of dit mogelijk is. Individuen uit kwekerij 1A zijn genetisch licht verschillend van individuen uit andere kwekerijen. Deze individuen zijn netals individuen uitkwekerij1B, afkomstigvan een kwekerij uithet noorden van Frankrijk. Bovendien lijken deze individuen genetisch sterker op Vlaamse autochtone patrijzen. Deze kweekindividuen werden verkocht als zijnde ideaal voor de aanvulling van natuurlijke patrijzenpopulaties. Waarschijnlijk wordt voor deze genetische kweeklijn gebruik gemaaktvan een andere setvan broeddieren. De kleine genetische afstand tot de wilde patrijzen uit Oost Brabant, waar overigens het meest kweekindividuen werden geïdentificeerd, kan verschillende oorzaken hebben. Waarschijnlijk werden er recent en/of minderrecent patrijzen van deze kweeklijn uitgezet in deze regio, heeft er een grootschalige uitzetting plaatsgevonden, waardoor meer dieren overleven, of hebben deze dieren een grotere overlevings en voortplantingskans in het wild. 4.2.2 Genetische structuur van wilde populaties Wanneerwe kijken naar de genetischestructuur van de autochtone patrijzenpopulaties, zien we twee genetischeclusters. Belangrijke kanttekeninghierbij is datnagenoegalle patrijzen uit Limburg (8 van de 10 individuen) een herkomst uit kweek hebben, waardoor we over deze regio weinig conclusies kunnen trekken. Patrijzen uit de regio Noorderkempen lijken een aparte genetische groep te vormen. Binnen de tweede groep die alle andere regio’s omvat, zien we een subtiele genetische structuur die vooral een west oost gradiënt en een kleinschalige ruimtelijke structuur lijkt te weerspiegelen. Over het algemeen is de dispersieafstand van patrijzen beperkt tot enkele kilometers (Buner et al. 2005; Potts 2005). Dit lijktbevestigd te worden dooronze resultaten, aangezien de correlatie tussen genetische afstand en euclidische afstand na enkele km zeer sterk afneemt. Bovendien is er geen significante correlatie meer bij afstanden groter dan 20 km. Dit geeft aan dat de directe en indirecte invloed van genmigratie tussen gebieden over meerdere generaties op grotere afstanden niet meer waarneembaar is met onze gegevens.
Een Wahlund effect wordt geobserveerd wanneer waargenomen heterozygositeit lager is dan verwacht als een gevolg van een vermenging van populaties met een verschillendegenetische samenstelling. Zoals eerdervermeld verschilde waargenomen en verwachte heterozygositeitin geen enkel van de regio’s. Dit geeft in het geval van patrijzen in Vlaanderen dus geen indicatie over uitzettingen. Via multivariate ordinatie, op basis van een PCA in deze studie, kunnen we de genetische afstand tussen individuen visueel weergeven op een diagram. Individuen die genetisch gelijkend zijn clusteren hierbij samen. Zoals eerder beschreven zien we een duidelijke groep met alle kweekindividuen en ook 47 wildvangst individuen die samen met deze kweekdieren clusteren. Individuen die binnen deze eerste cluster vallen zijn waarschijnlijk afkomstigvan een kwekerij. Hierbij zien we ook duidelijk dat er individuen binnen een bepaalde regio samen clusteren met zowel de wilde populaties als de kweekpopulaties.
Viadiscriminantanalyses, clustering en toewijzingen kunnen we individuen toewijzen aan bepaalde genetische clusters. Via DAPC analyses werden een aantal wilde individuen duidelijk toegewezen aan dezelfde cluster als de kweekpopulaties Bovendien toonde deze analyse aan dat er twee genetische clusters zijn, een cluster met gekweekte individuen en een cluster met wilde individuen. Op basis van de multivariateordinatie en de toewijzingen hebben we 47wildvangst individuen kunnen identificeren als afkomstigvan een kwekerij. Dezelfde individuen werden steeds met beide methodes geïdentificeerd. Verder geven de FST waardes een goed beeld over regio’s waar individuen uitgezet zijn. Dit zijn opnieuw dezelfde regio’s waarvan individuen bij de multivariate ordinatieen toewijzingen werden geïdentificeerd als een individu afkomstig uit een kwekerij. Zoals eerder aangehaald is het, waarschijnlijk door gelijkaardige herkomst van individuen, niet mogelijk om een onderscheid te maken tussen verschillende kwekerijen. 4.3.2 Onderscheid tussen recente en minder recente uitzettingen In een DAPC analyses worden individuen met een zeer hoge waarschijnlijkheid toegewezen aan de verschillende genetische groepen. Een STRUCTURE analyse daarentegen geeft meer genuanceerde resultaten en houdt rekening met de mogelijke aanwezigheid van genetische vermenging tussen groepen. Op basis van de STRUCTURE resultaten kunnen we betere conclusies maken over recente en minder recente uitzettingen. Bij een recente uitzetting verwachten we hoge toewijzingsscores op basis van STRUCTURE, aangezien er nog geen genetische vermenging heeft plaatsgevonden. Dit is anders bij minder recente uitzettingen, waarbij nageslacht tussen kweek en wilde individuen deels worden toegewezen aan beide
individuen in gevangenschap kan bovendien een sterke invloed hebben op de genetische achtergrond van individuen en populaties, waardoor we een verschil in genetische samenstelling verwachten tussen autochtone en kweekindividuen. Zoals toegelicht in de inleiding van dit rapport zijn er verschillende genetische analysetechnieken beschikbaar om uitzettingen te detecteren in wildvangst populaties.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 40 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo
Wildvangstpatrijzen meteen herkomstuitkweekpopulaties detecteerden we nagenoeg in alle regio’s, behalvede Noorderkempen en de meeste West Vlaamse regio’s. Metuitzonderingvan Zuid Limburg, waren er in elk van deze regio’s telkens ook autochtone individuen aanwezig. Van de uitgezetteindividuen waren stalen aanwezigdie zowelafkomstigwaren van de jacht (n = 15 op een totaal van 216 individuen, jachtseizoen 2020) als van onze eigen staalnamecampagne (n = 33 op een totaal van 160 individuen, voorjaar 2021).
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 41 van 57 groepen. Hierbij verwachten we assignmentscores tussen 50en 70%. Van de 48 individuen die werden geïdentificeerd als afkomstig uit een kwekerij, zijn er 13 waarschijnlijk kruisingen tussen wilde en kweekindividuen. 4.3.3 Uitzettingen in Vlaanderen
lijken uitzettingen een groteraandeel te hebben op de totale populatie in Oost Brabant (21 van de 42 individuen), waar ook een aanzienlijk deel kruisingen werd gedetecteerd (11van de 21 individuen), en Limburg (8 van de 10 individuen). Een belangrijke nuance die we hierbij moeten maken, is datin de provincie Limburgslechts 11 stalen bekomen werden, afkomstiguitvierWBE’s. Dit ondanks hetfeitdat er in deze provincie in 2020 nog 739 patrijzen verdeeld over 11 WBE’s geschoten werden (Scheppers 2021). Van de WBE’s waar uitzettingen gedetecteerd zijn, zijn erdrie die volgens hun gerapporteerde afschotcijfers in het najaar van 2020 nietop patrijs hebben gejaagd (Figuur 3). Hoewel het uitzetten van patrijzen in functie van jacht in Wallonië actueel ter discussie staat, met recente wijzigingen in de regelgevinghieromtrent, was hetin 2020 nog toegestaan om in Wallonië gekweekte patrijzen uitte zetten. Ditis vooral relevantvoorgebieden dichtbijde Waalse grens (vb. Zuid Limburg). Hierbij moet rekening gehouden worden met de (beperkte) dispersieafstand van patrijzen. Sinds 2020 is het uitzetten van patrijzen aan strikte voorwaarden verbonden in Wallonië. Uitzettingen zijn enkelnogmogelijk in hetkadervan bijplaatsingen voor soortherstel, waarna er gedurende drie jaren een jachtverbod bestaat. Bovendien moeten alle uitgezette dieren voorzien zijn van een wetenschappelijke ring. Dit verkleint drastisch de kans dat in de toekomst een in Wallonië(legaal) uitgezettepatrijs foutiefgedetecteerd zou worden als een in Vlaanderen uitgezet exemplaar. Van de individuen diegevangen werden, werd ook telkens de leeftijdsklasse (<1j, >1 j) geschat in het veld op basis van het ruipatroon. Hieruit bleek dat 28% van de gevangen patrijzen
Overhetalgemeen
We benadrukken datditgeen representatief beeld geeft van de omvang van uitzettingen van patrijzen. De staalnames waren beperkt tot gebieden waar lokale wildbeheereenheden zich bereid hadden getoond totsamenwerking, zowel voorhetaanleveren van stalen van gestrekte patrijzen als voor hulp bij de vangst van levende patrijzen. Dit maakt dat we zeker voor geschoten patrijzen relatief weinig uitgezette dieren verwachtten. Aangezien het overlevingssucces van uitgezettepatrijzen doorgaans lager wordt ingeschat dan dat van wilde exemplaren (Buner et al. 2005, 2011; Parish & Sotherton 2007; Rymešová et al. 2012), verwachtten we ook een onderschatting van het aandeel uitgezette patrijzen bij de ringvangsten in de periode februari tot april 2021.
Aangezien de stalenreeks die hier geanalyseerd werd geen willekeurige weergave is van het totaal van wilde en gekweekte patrijzen, raden we aan om routinematig DNA in te zamelen van het gerealiseerde afschot van patrijs. Enkel dit zal toelaten om te bepalen hoe de verhoudingtussen wilde en gekweekte vogels in werkelijkheid is. We adviseren dan ook om aan het afschot een verplichting te koppelen tot het bewaren van een achterteen van elke gestrekte patrijs voorverdere DNA analyse. Een vergelijkbare regeling is reeds van kracht bij ree en everzwijn, waarbij de linker onderkaak bewaard dient te worden en ter beschikking
De methodes gebruikt in deze studie laten toe om zowel recente als minder recente uitzettingen van in gevangenschap gekweekte patrijzen te detecteren in Vlaanderen. De methode die we gebruikten is zeer krachtig, maar arbeidsintensief. Met de genotypes op 30 000 genetischemerkers zullen we in 2022 een selectievan enkele honderden merkers maken waardoorwe met een veel kleinere inspanning het onderscheid tussen gekweekte en wilde patrijzen kunnen maken. Hiervoorgaan we gebruik maken van een Genotyping in Thousands by sequencing methode (GT seq, Campbell et al. 2015; Schmidt et al. 2020). Deze methode laat toe om efficiënt, snelen relatief eenvoudigde afkomst van een groot aantal individuen te bepalen (Campbell et al. 2015). In parallel zullen we ook nagaan in welke mate eradaptievegenetische verschillen zijn tussen patrijzen afkomstig uit kweek en autochtone patrijzen. Mogelijk heeft het lagere reproductiesucces van uitgezette patrijzen in het wild een genetische basis die gelinkt kan worden aan aanpassingen aan omstandigheden in gevangenschap. Dit kan een bijkomende reden vormen om uitzettingen van patrijs te ontmoedigen, ook in landen en regio’s waar dit nog wel toegelaten is. 4.5 AANBEVELINGEN VOOR BELEID
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 42 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo minstens 1 j oud was, en waarvan ook enkele dieren uit kweek afkomstig waren. De gemiddelde leeftijd van patrijzen in hetvoorjaaris 1.8 jaar, met een maximalegerapporteerde leeftijd van 5 jaar (Robinson 2005). We kunnen echter geen conclusies maken over overlevingssucces, aangezien we geen beeld hebben over het aantal individuen dat initieel werd uitgezet. Bovendien detecteerden we in bepaalde regio’s (vnl. Oost Brabant) een aanzienlijk aandeel kruisingen tussen autochtone en kweekindividuen, wat aangeeft dat sommige gekweekte individuen zich succesvolvoortplanten. Ditondanks de verwachtingen dat dit eerder zeldzaam is (vb. Buner et al. 2005; 2011; Morris & Gilroy 2008; Parish & Sotherton 2007; Putaala et al. 2001; Rantanen et al. 2010; Rymešová et al. 2012). Belangrijk om hier te vermelden is dat een succesvolle overleving en voortplanting van kweekindividuen niet noodzakelijk leidt tot duurzame populaties (Homberger et al. 2021). 4.4 VERVOLGONDERZOEK
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 43 van 57 gesteld dientte worden van hetANB of het INBO (BVR 25/04/20143). Dit kan gebruikt worden voor monitoringvan de genetische structuuroverde jaren heen, waarbijplotse verschuivingen ook kunnen wijzen op uitzettingen. Deze gebiedsdekkende staalname laat het beleid ook toe om jaarlijks een selectie te maken van stalen voor de herkomstbepaling van gestrekte patrijzen. 3 Art. 20. Besluit vande Vlaamse Regering houdende vaststelling van de voorwaarden waaronder de jacht kan worden uitgeoefend [citeeropschrift:"het Jachtvoorwaardenbesluit van 25 april 2014"] (B.S. 2014)
AVES (2020). L’Atlas des Oiseaux Nicheurs: Explications sur les cartes (unpublished data). Barbanera F., Pergams O.R., Guerrini M., Forcina G., Panayides P.& Dini F. (2010). Genetic consequences of intensive management in game birds. Biological Conservation, 143(5), 1259 1268. https://doi.org/10.1016/j.biocon.2010.02.035.
Andrews8312.2011.01833.x.S.(2015).FastQC:
Perdix perdix (Grey Partridge) European Red List of Birds. Supplementary Material. Luxembourg: Office for Official Publications of the European Communities. BirdLifeInternational (2017).European birds of conservation concern: populations, trends and national responsibilities. Cambridge, UK. Birkan M. (1971). Réussite des lâchers de perdrix grise (Perdix perdix) et de perdrix rouge (Alectoris rufa) d’élevage. Bulletin du Conseil Supérieur dela Chasse(15):44 56.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 44 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo Referenties
Bech N. (2010). Génétique de la Conservation et du Paysage: Étude des Populations de Lagopède alpin et de Perdrix grise en France. Sciences del’environnement, UniversitédePerpignan. Bech N., Novoa C., Allienne J.F. & Boissier J. (2010). Transferability of microsatellite markers among economically and ecologically important galliform birds. Genetics and Molecular Research 9 (2): 1121 1129. https://doi.org/10.4238/vol9 2gmr760. Bech N., Novoa C., Bro E. & Boissier J. (2012). Conservation de la diversité génétique des populations naturelles de perdrix grise: la question des lâchers d’oiseaux d’élevage. FauneSauvage297:10 14. Bech N., Novoa C., Allienne J.F., Boissier J. & Bro E. (2020). Quantifying genetic distance between wild and captive strains of the grey partridge Perdix perdix in France: conservation implications. Biodiversity and Conservation, 29(2), 609 624. https://doi.org/10.1007/s10531 019 01901 w. Besnier F. & Glover K.A. (2013). ParallelStructure: AR package to distribute parallel runs of the population genetics program STRUCTURE on multi core computers. PLoS One, 8(7), e70651. BirdLifehttps://doi.org/10.1371/journal.pone.0070651.International(2015).
ANB (2020). Subsidie bescherming nesten en jongen broedvogels op landbouwpercelen 2020 Aebischer N.J. (2019). Fifty year trends in UK hunting bags of birds and mammals, and calibrated estimation of national bag size, using GWCT’s National Gamebag Census. European Journal of Wildlife Research 65 (4). https://doi.org/10.1007/s10344 019 1299 x. Andersen L.W. & Kahlert J. (2012). Genetic indications of translocated and stocked grey partridges (Perdix perdix): does the indigenous Danish grey partridge still exist? Biological journal of the Linnean Society 105 (3): 694 710. https://doi.org/https://doi.org/10.1111/j.1095 A quality control tool for high throughput sequence data [WWW document]. https://www.bioinformatics.babraham.ac.uk/projects/fastqc/. Araki H., Cooper B. & Blouin M.S. (2007). Genetic effects of captive breeding cause a rapid, cumulative fitness declineinthe wild.Science 318 (5847): 100 103.https://doi.org/10.1126/science.1145621.
Buner F., Jenny M., Zbinden N. & Naef daenzer B. (2005). Ecologically enhanced areas a key habitat structure for re introduced grey partridges Perdix perdix. 124: 373 381. Bunerhttps://doi.org/10.1016/j.biocon.2005.01.043.F.&SchaubM.(2008).Howdodifferentreleasing techniques affect the survival of reintroduced grey partridges Perdix perdix? Wildlife Biology 14 (1). Campbell N.R., Harmon S.A. & Narum S.R. (2015). Genotyping‐in‐Thousands by sequencing (GT‐seq): A cost effective SNP genotyping method based on custom amplicon sequencing. Molecular ecology resources, 15(4), 855 867. https://doi.org/10.1111/1755 0998.12357. Carroll J.P. (1993). The birds of North America. Life histories for the 21st Century. The Academy of Natural Sciences, The American Ornithologists’ Union, Philadelphia, Washington DC, 1 20 p. Casas F., Mougeot F., Sánchez Barbudo I., Dávila J.A. & Viñuela J. (2012). Fitness consequences of anthropogenic hybridizationinwildred leggedpartridge (Alectoris rufa, Phasianidae)populations. Biological Invasions, 14(2), 295 305. https://doi.org/10.1007/s10530 011 0062 3. Catchen J., Hohenlohe P.A., Bassham S., Amores A. & Cresko W.A. (2013). Stacks: an analysis tool set for populationgenomics.Molecular Ecology 22 (11): 3124 3140. https://doi.org/10.1111/mec.12354.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 45 van 57 Birkan M. & Jacob M. (1988). La Perdrix Grise. Hatier, Paris, France. Blanco‐Aguiar J.A., González‐Jara P., Ferrero M.E., Sánchez‐Barbudo I., Virgós E., VillafuerteR. & Dávila J.A. (2008).Assessment of game restockingcontributions to anthropogenic hybridization: the case of the Iberian red‐legged partridge. Animal Conservation, 11(6), 535 545. Blank T.H., Southwood T.R.E. & Cross D.J. (1967). The Ecology of the Partridge : I. Outlineof Population Processes withParticularReference to ChickMortalityandNest Density. Journal of Animal Ecology 36 (3): 549 556. Brittas R., Marcström V., Kenward R.E. & Karlbom M. (1992). Survival and breeding success of reared and wild ring necked pheasants in Sweden. The Journal of wildlife management, 368 376. https://doi.org/10.2307/3808836. Bro E., Sarrazin F., Clobert J. & Reitz F. (2000). Demography and the decline of the grey partridge Perdix perdix in France. Journal of Applied Ecology 37 (3): 432 448. https://doi.org/10.1046/j.1365 2664.2000.00511.x. Bro E., Sautereau L., Bouteiller R. et al. (2017) Opérations de repeuplement et de renforcement de populations de faisan commun et de perdrix grise. Rôle des conservatoires de l’ONCFS et du collectif Perdrix. Faune Sauvag 317:53 57. Buner F. & Gottschalk E. (2020). Grey Partridge. In: Keller, V., Herrando, S., Voříšek, P., Franch, M., Kipson, M., Milanesi, P., Martí, D., Anton, M., Klvaňová, A., Kalyakin, M.V., Bauer, H. G. & Foppen, R.P.B. (2020).European Breeding BirdAtlas 2: Distribution,Abundance and Change. European Bird Census & Lynx Edicions, Barcelona. Buner F., Browne S. & Aebischer N.J. (2011). Experimental assessment of release methods for the re establishment of a red listed galliform, the grey partridge (Perdix perdix). Biological Conservation 144 (1): 593 601. https://doi.org/https://doi.org/10.1016/j.biocon.2010.10.017.
Catchen J.M., Amores A., Hohenlohe P., Cresko W. & Postlethwait J.H. (2011). Stacks : Building and Genotyping Loci De Novo From Short Read Sequences. G3 Genes|Genomes|Genetics 1 (3): 171 182. https://doi.org/10.1534/g3.111.000240.
https://doi.org/10.1371/journal.pone.0019379.
Champagnon J., Elmberg J., Guillemain M., Gauthier Clerc M. & Lebreton J.D. (2012). Conspecifics can be aliens too: a review of effects of restocking practices in vertebrates. Journal for Nature Conservation, 20(4), 231 241. https://doi.org/10.1016/j.jnc.2012.02.002. Christie M.R., Marine M.L., French R.A. & Blouin M.S. (2012). Genetic adaptation to captivity can occur in a single generation. Proceedings of the National Academy of Sciences of the United States of America 109 (1): 238 242. https://doi.org/10.1073/pnas.1111073109.
https://doi.org/10.1111/j.1471
https://doi.org/10.1111/j.1365
Commissie voor Leefmilieu Natuur Ruimtelijke Ordening Energie en Dierenwelzijn (2021). Vraag om uitleg over de illegale uitzettingen van fazanten en patrijzen [WWW document]. werk/comhttps://www.vlaamsparlement.be/nl/parlementairmissies/commissievergaderingen/1079443/verslag/1080934.Danecek P., Auton A., Abecasis G., Albers C.A., Banks E., DePristo M.A., Hndsaker, Robert E., Lunter G., Marth G.T., Sherry S.T., McVean G. & DurbinR. (2011). The variantcallformatandVCFtools. Bioinformatics 27 (15): 2156 2158. https://doi.org/https://doi.org/10.1093/bioinformatics/btr330.
/////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////
Pagina 46 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo
Devos K. (2004). Patrijs. In: Vermeersch G., Anselin A., Devos K., Herremans M., Stevens J., Gabriëls J. & Van Der Krieken B. (2004). Atlas van de Vlaamse broedvogels 2000 2002. Mededelingen van het Instituut voor Natuurbehoud 23, Brussel. Devos K., Anselin A., Driessens G., Herremans M., Onkelinx T., Spanoghe G., Stienen E.W.M., T’jollyn F., Vermeersch G. & Maes D. (2016a). De IUCN Rode Lijst van de broedvogels in Vlaanderen (2016). Rapporten van het Instituut voor Natuur en Bosonderzoek jaar (11485739) (DOI: dx.doi.org/10.21436/inbor.11485739): 52. Devos K., Vermeersch G., Onkelinx T., T’Jollyn F. & Lewylle I. (2016b) HetprojectAlgemeneBroedvogels Vlaanderen (ABV): een nieuwe update van populatietrends (2007 2016). Vogelnieuws, ornithologische nieuwsbrief van het INBO, (27) 28 31, Brussel. ecopedia.be (2021). Patrijs | Ecopedia [WWW document]. https://www.ecopedia.be/dieren/patrijs. Earl D.A. & VonHoldt B.M. (2012). STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conservation genetics resources, 4(2), 359 361. Elshire R.J., Glaubitz J.C., Sun Q., Poland J.A., Kawamoto K., Buckler E.S. & Mitchell S.E. (2011). A Robust, Simple Genotyping by Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 6 (5): e19379. Falush D., Stephens M. & Pritchard J. K. (2007). Inference of population structure using multilocus genotype data: dominant markers and null alleles. Molecular ecology notes, 7(4), 574 578. 8286.2007.01758.x. Frankham R., Ballou J. & Briscoe D.A. (2002). "Genetic management of captive populations." Introduction to Conservation Genetics: 419 446. Frankham R. (2008). Genetic adaptation to captivity in species conservation programs. (17): 325 333. 294X.2007.03399.x. Frankham R., Balloux J.D. & Briscoe D.A. (2010). Introduction to Conservation Genetics. Cambridge University Press, Cambridge, UK, 618 p. Goudet J. (2004). HIERFSTAT, a package for R to compute and test hierarchical F statistics. Molecular Ecology Notes 5 (1): 184 186. https://doi.org/doi.org/10.1111/j.1471 8286.2004.00828.x.
Kraaijeveldhttps://doi.org/10.1093/icesjms/fsw121.SmitF.J.,GriffithsR.A.,MooreR.D.&
Homberger B., Jenni L., Duplain J., Lanz M. & Schaub M. (2021). Strong effects of radio tags, social group and release date on survival of reintroduced grey partridges. Animal Conservation. Huyghebaerthttps://doi.org/10.1111/acv.12673.P.(2020).Lokaaljachtverbod op patrijs, haas en fazant nadat dieren werden uitgezet om te bejagen: “Verboden en ontoelaatbaar” *WWW document+. In: vrt.be. https://www.vrt.be/vrtnws/nl/2020/10/04/lokaal jachtverbod nadat patrijzen werden uitgezet om te bejagen/ (accessed October 4, 2020). Jombart T. & Collins C.(2015).A tutorial for discriminantanalysis of principal components (DAPC) using adegenet 2.0. 0. London: Imperial College London, MRC Centre for Outbreak Analysis and KarlssonModelling.S.,DiserudO.H., Fiske P., Hindar K. (2016).Widespreadgenetic introgressionofescapedfarmed Atlantic salmon in wild salmon populations. ICES Journal of Marine Science, 73: 2488 2498.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 47 van 57 Green R.E. (1984). The feeding ecology and survival of partridge chciks (Alectoris rufa and Perdix perdix) on arable farmland in East Anglia. Journal of Applied Ecology (21): 817 830. Guidelines for taking samples from birds and mammals for DNA extraction (s.d.). Facility Protocols Sheffield Molecular Genetics. http://library1.nida.ac.th/termpaper6/sd/2554/19755.pdf.
Beebee, T.J. (2006). Captive breeding and the fitness of reintroduced species: a test of the responses to predators in a threatened amphibian. Journal of Applied Ecology, 360 365. Kuijper D.P.J., Oosterveld E. & Wymenga E. (2009). Decline and potential recovery of the European grey partridge (Perdix perdix) population a review. European Journal of Wildlife Research 55 (5): 455 463. https://doi.org/10.1007/s10344 009 0311 2. Laikre L., Schwartz M.K., Waples R.S. & Ryman N. (2010). Compromising genetic diversity in the wild: unmonitored large scale release of plants and animals. Trends in Ecology and Evolution 25: 520 529. https://doi.org/10.1016/j.tree.2010.06.013. Langmead B. & Salzberg S.L. (2012). Fast gapped read alignment with Bowtie2. Nature Methods 9: 357 Lavretsky359.P., McInerney N.R., Mohl J.E., Brown J.I., James H.F., McCracken K.G. & Fleischer R.C. (2020). Assessing changes in genomic divergence following a century of human‐mediated secondary contact among wild and captive‐bred ducks. Molecular Ecology: 29(3), 578 595. Lepaishttps://doi.org/10.1111/mec.15343O.&WeirJ.T.(2014).SimRAD:anR packagefor simulation basedpredictionofthe number of loci expected in RADseq and similar genotyping by sequencing approaches. Molecular Ecology Resources 14 (6): 1314 1321. https://doi.org/10.1111/1755 0998.12273. Liukkonen T., Kvist L. & Mykrä S. (2012). Microsatellite markers show distinctiveness of released and wild grey partridges in Finland. Animal Biodiversity and Conservation 35 (2): 419 428. Manlyhttps://doi.org/10.32800/abc.2012.35.0419.B.F.J.(1977).Thedeterminationofkeyfactors from life table data. Oecologia (31): 111 117. Martin J., Novoa C., Blanc Manel S. & Tabarlet P. (2003). Les populations de perdrix grise des Pyrénéés (Perdix perdix hispaniensis) ont elles subi une introgression génétique à partir d’individus
/////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////
McGowan P.J. & Kirwan G.M. (2013). Grey partridge (Perdix perdix). In: del Hoyo J., Elliott A., Sargatal J., Christie D.A. & de Juana E. (eds.). Handbook of the Birds of the World Alive. Lynx Edicions, MeriggiBarcelona.A.&Della
Mcginnity P., Prodöhl P., Ferguson A., Hynes R., Maoiléidigh N., Baker N., Cotter D., Hea B.O., Cooke D., Rogan G., Taggart J., Cross T., Mcginnity P., Prodo P., Ferguson A., Hynes R., Baker N., Cotter D., Hea B.O., Cooke D., Rogan G., Taggart J. & Cross T. (2003). Fitness reduction and potential extinction of wild populations of Atlantic salmon, Salmo salar, as a result of interactions with escaped farm salmon. Proceedings of the Royal Society of London. Series B: Biological Sciences, 270(1532), 2443 2450. https://doi.org/10.1098/rspb.2003.2520.
Muñoz Fuentes V., Vilà C., Green A. J., Negro J.J. & Sorenson M.D. (2007). Hybridization between white headed ducks and introduced ruddy ducks in Spain. Molecular Ecology (16): 629 638. https://doi.org/10.1111/j.1365 294X.2006.03170.x. Onkelinx T., Vermeersch G. & Devos K. (2021). Patrijs. In: Trends op basis van de Algemene Broedvogelmonitoring Vlaanderen (ABV). Technisch achtergrondrapport voor de periode 2007‐2020. Rapporten van het Instituutvoor Natuur‐ en Bosonderzoek 2021 (14). Instituutvoor Natuur‐en Bosonderzoek, Brussel. https://doi.org/10.21436/inbor.34162521
Poland J.A., Brown P.J., Sorrells M.E. & Jannink J. L. (2012). Development of High Density Genetic Maps for Barley and Wheat Using a Novel Two Enzyme Genotyping by Sequencing Approach. PLoS ONE 7 (2): e32253. https://doi.org/10.1371/journal.pone.0032253. Pritchard J K., Stephens M. & Donnelly, P. (2000). Inference of population structure using multilocus genotype data. Genetics, 155(2), 945 959. https://doi.org/10.1093/genetics/155.2.945
Stella R.M. (2004). Dynamics of a reintroduced population of red legged partridges Alectoris rufa in central Italy. Wildlife Biology 10 (1): 1 9. https://doi.org/10.2981/wlb.2004.002. Morris A.J. & Gilroy J.J. (2008). Close to the edge: predationrisks for two declining farmland passerines. Ibis 150 (1): 168 177. https://doi.org/https://doi.org/10.1111/j.1474 919X.2008.00857.x.
Putaala A. & Hissa R. (1998). Breeding dispersal and Demography of wild and Hand Reared Grey Partridges (Perdix perdix) in Finland. Wildlife Biology 4: 137 145. Putaala A., Turtola A. & Hissa R. (2001). Mortality of wild and released hand reared grey partridges (Perdix perdix) in Finland. Game and Wildlife Science (France). R Core Team (2021). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R project.org/
Paris J.R., Stevens J.R. & Catchen J.M. (2017). Lost in parameter space: a road map for stacks. Methods in Ecology and Evolution 8 (10): 1360 1373. Parish D.M.B. & Sotherton N.W. (2007). The fate of released captive reared grey partridges Perdix perdix: Implications for reintroduction programmes. Wildlife Biology 13 (2): 140 149. https://doi.org/10.2981/0909 6396(2007)13[140:TFORCG]2.0.CO;2.
Pagina 48 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo d’élevage? Analyse du polymorphisme de 1’ADN mitochondrial. Les Actes du BRG (4): 115 126. https://www.yumpu.com/fr/document/read/34851708/les populations de perdrix grise des pyrenees bureau des Martin M. (2011). Cutadapt removes adapter sequences from high throughput sequencing reads. EMBnet.journal 17 (1): 10. https://doi.org/10.14806/ej.17.1.200.
Southwood T.R.E. & Cross D.J. (1969). The Ecology of the Partridge: III. Breeding success and the abundance of insects in natural habitats. Journal of Animal Ecology 38 (3): 497 509.
Robinson R.A. (2005) BirdFacts: profiles of birds occurring in Britain & Ireland. BTO, Thetford (http://www.bto.org/birdfacts, accessed on 21 December 2021). Rochette N.C. & Catchen J.M. (2017). Deriving genotypes from RAD seq short read data using Stacks. Nature Protocols 12: 2640 2659. Rochette N.C., Rivera‐Colón A.G. & Catchen J.M. (2019). Stacks 2: Analytical methods for paired‐end sequencing improve RADseq‐based population genomics. Molecular Ecology 28 (21):4737 4754. https://doi.org/10.1111/mec.15253.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 49 van 57 Randi E. (2008). Detecting hybridization between wild species and their domesticated relatives. Molecular ecology, 17(1), 285 293. https://doi.org/10.1111/j.1365 294X.2007.03417.x. Rantanen E.M., Buner F., Riordan P., Sotherton N. & Macdonald D.W. (2010). Habitat preferences and survival in wildlife reintroductions: An ecological trap in reintroduced grey partridges. Journal of Applied Ecology 47 (6): 1357 1364. https://doi.org/10.1111/j.1365 2664.2010.01867.x.
Rymešová D., Tomášek O. & Šálek M. (2013). Differences in mortality rates, dispersal distances and breeding success of commercially reared and wild grey partridges in the Czech agricultural landscape.EuropeanJournal of Wildlife Research (59): 147 158. https://doi.org/10.1007/s10344 012 0659 6. Sage R. B., Putaala A., Pradell Ruiz V., Greenall T.L., Woodburn M.I. & Draycott R.A. (2003). Incubation success ofreleasedhand reared pheasants Phasianus colchicus compared with wild ones. Wildlife Biology, 9(4), 179 184. https://doi.org/10.2981/wlb.2003.048.
Schmidt D.A., Campbell N.R., Govindarajulu P., Larsen K.W. & Russello M.A. (2020). Genotyping‐in‐Thousands by sequencing (GT‐seq) panel developmentand application to minimally invasiveDNA samples to support studies in molecular ecology. Molecular Ecology Resources, 20(1), 114 124. https://doi.org/10.1111/1755 0998.13090. Sokos C.K., Birtsas P.K. & Tsachalidis E.P. (2008). The aims of galliformes release and choice of techniques. Wildlife Biology 14 (4): 412 422. https://doi.org/10.2981/0909 6396 14.4.412. Southwood T.R.E. & Cross D.J. (1967). The Ecology of the Partridge: II. The role of pre hatching influences. Journal of Animal Ecology 38 (3): 497. https://doi.org/10.2307/3030.
The Game Farmers’ Association (2021). Game Farming in the UK *WWW document+. https://www.gfa.org.uk/game farming in the uk.html.
Schauvliege M. & De Bruyn P. (2020). Verslag van de hoorzitting namens de Commissie voor Leefmilieu, Natuur, Ruimtelijke Ordening en Energie: over het voorstel van decreet tot wijziging van het Jachtdecreet van 24 juli 1991, wat de schrapping van de patrijs als bejaagbare soort betreft. Vlaams Parlement. Scheppers T. (2021). Afschotstatistieken 2020. Instituut voor Natuur en Bosonderzoek, 2 pp. Scheppers T., Verzelen Y., Devos K., Huysentruyt F., Casaer J., Adriaens T., De Bruyn L., Van Den Berge K., Van Daele T. & Vermeersch G. (2019). De impact van jacht op patrijzenpopulaties: Wat kunnen populatiemodellen ons leren? Rapporten van het Instituut voor Natuur en Bosonderzoek 2019 (29). Instituut voor Natuur en Bosonderzoek, Brussel. doi.org/10.21436/inbor.15402520.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 50 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo Vermeersch G. & Devos K. (2020). Patrijs. In: Vermeersch, G., Devos, K., Driessens G., Everaert J., Feys, S., Herremans M., Onkelinx T., Stienen E.W.M. & T’Jollyn F. (2020). Broedvogels in Vlaanderen 2013 2018. Recente status en trends van in Vlaanderen broedende vogelsoorten. Mededelingen van het Instituut voor Natuur en Bosonderzoek 2020 (1) Brussel, 228p. DOI: Vermeerschdoi.org/10.21436/inbor.18794135G.,OnkelinxT.&Driessens G. (2021). Resultaten van het project Algemene Broedvogels Vlaanderen (ABV) tot en met 2020. In: INBO Vogelnieuws 34. p. 25 29. Verzelen Y., Scheppers T., Onkelinx T., Devos K., Huysentruyt F., Boone N. & Maurice H. (2020). Advies over de toestand van de patrijs. 25: 1 14. Vlaamse Vogelatlas (2021). Vogelatlas.be De vogelatlas van Vlaanderen [WWW document]. Weekshttps://www.vogelatlas.be/atlasbe/soorten/soort/3670.A.R.,SgroC.M.,YoungA.G.,FrankhamR,MitchellN.J., Miller K. A., ... & Hoffmann A. A. (2011). Assessingthe benefits andrisks oftranslocationsin changing environments: a genetic perspective. Evolutionary Applications, 4(6), 709 725. https://doi.org/10.1111/j.1752 4571.2011.00192.x.
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 51 van 57 Bijlage Bijlage 1: Supplementaire figuren Figuur 13 Verspreidingskaart van patrijs in Wallonië gebaseerd op recente waarnemingen. Het zwaartepunt van het voorkomen van patrijs lijkt zich voornamelijk in de lager gelegen delen van Wallonië,boven de (Samber )Maaslijn, tebevinden met duidelijke‘hotspots’ nabij degewestgrens en de landsgrens met Frankrijk. (AVES 2020).

///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 52 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo Figuur 14. Populatietrends voor Europese patrijzenpopulaties op zowel korte (boven) als lange (onder) termijn. Op de beide kaarten werden zowel de groottes van de broedpopulaties per land en de verwachte populatietrends voor beide termijnen bepaald Op basis van de grootteorde, het kleur en de richting duiden de pijlen aan welke trend teruggevonden wordt per land waar patrijs aanwezig is. De grootte van de pijl geeft de proportie, overeenkomend met het aandeel van de Europese populatie dat per land teruggevonden kan worden, weer (≥10%, 1 9% en <1%). Hoe donkerder de kleur, hoe hoger het


///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 53 van 57 stijgings of dalingspercentage geschatwordt(donker rood =daling van ≥50%, rood =daling tussen 20 49%, roze = <20% en een lege pijl staat voor een daling van ongekende grootte). Enkel in Finland, Ierland en Moldavië worden op korte termijn grote stijgingen teruggevonden; op lange termijn worden respectievelijk een grote, ongekende en gemiddelde stijging teruggevonden. Volle rechthoekjes staan voor een fluctuerende of stabiele populatie, in landen met een leeg rechthoekje zijn de toekomstige trends niet bekend (BirdLife International 2015)
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 54 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo Bijlage 2 Overzicht genetische structuur alle individuen (per WBE) (a) (b)
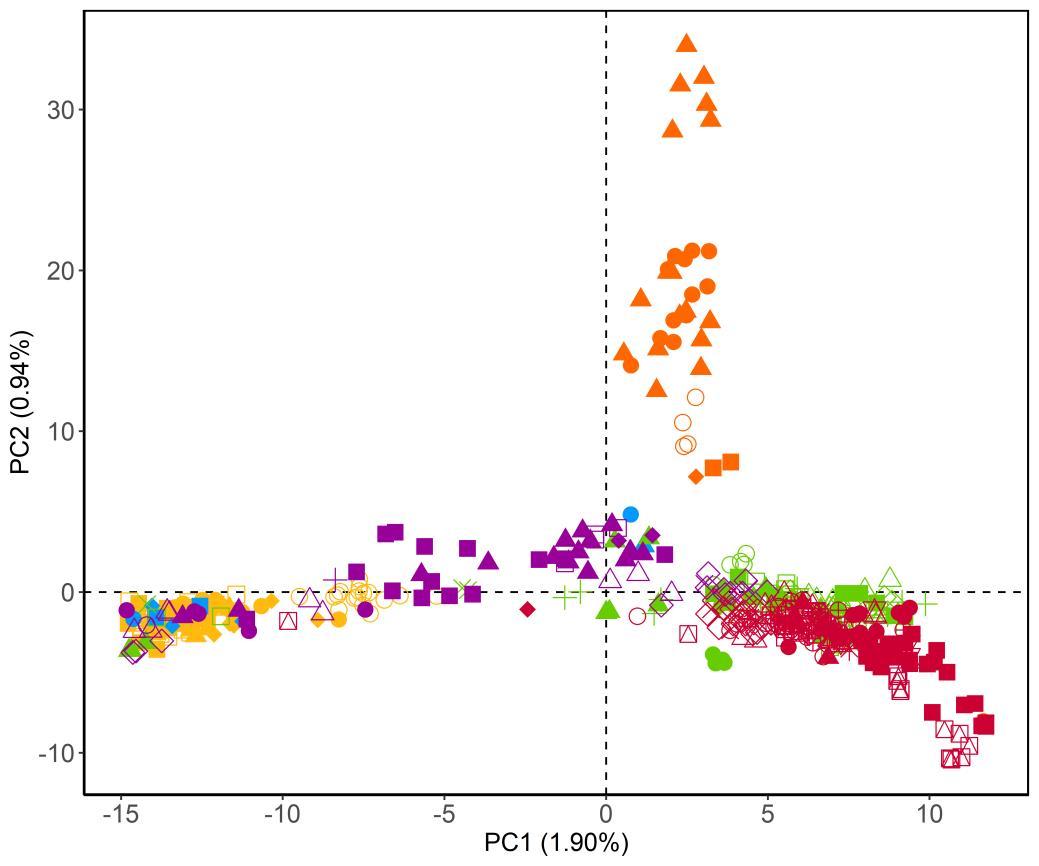
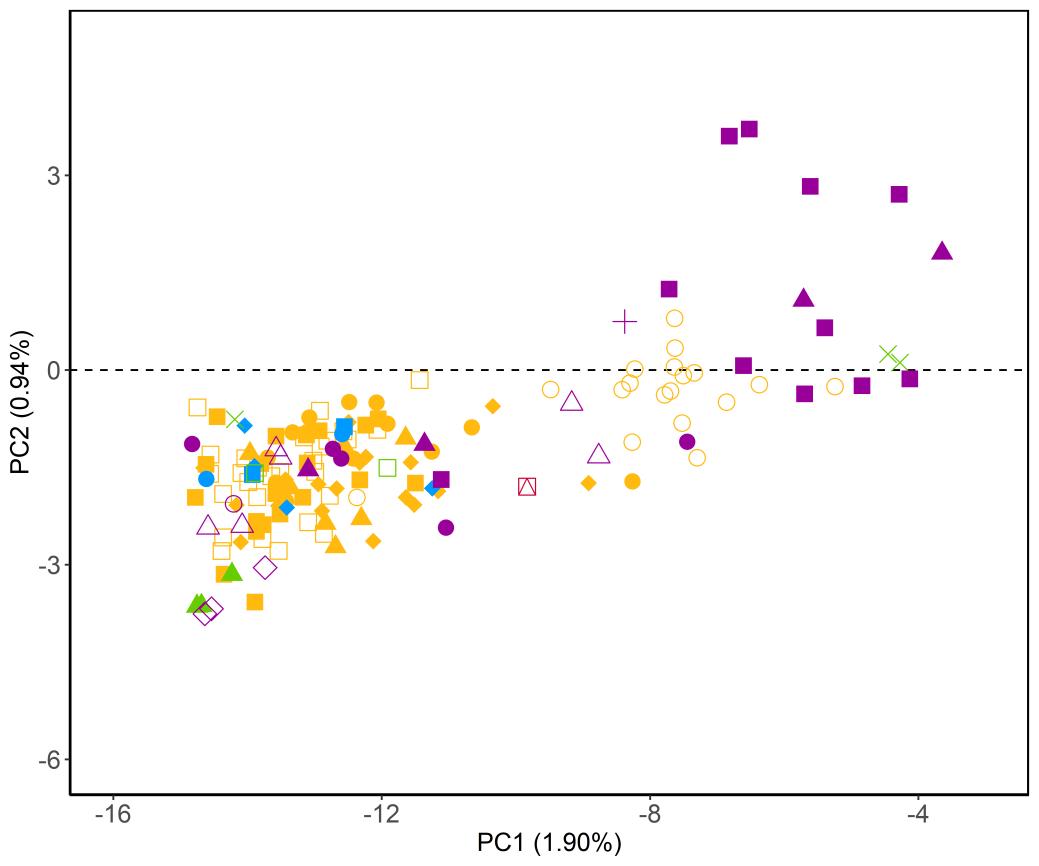
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 55 van 57 Figuur 15. Ordinatieplot(PCA) waarbij desymbolen de verschillendepatrijzenindividuenvoorstellen.De kleuren en symbolengeven aantot welke wildbeheereenheden (Tabel 8) deze individuenbehoren. Hoe dichter de afstandtussenindividuenopde plot, hoe meer gelijkenddeze zijnqua genetische samenstelling.(a)Overzichtvan alleindividuen.(b)Detail vande ordinatie,gefocust op dat deel dat overeenkomt met patrijzenafkomstiguitgevangenschap.

///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// Pagina 56 van 57 doi.org/10.21436/inbor.62125143 www.vlaanderen.be/inbo Tabel 8. Afkortingen gebruikt voor de wildbeheereenheden in Figuur 15 en 16. WBE Afkorting WBE Afkorting ‘t Veld Veld Klaverbeek KlBe Achterhaven Zeebrugge HaZe Land vanAalst LaAa Antwerpse Polders Noord AnPo Langs de Hoge Kale HoKa Baekeland BaLa Leie en Schelde kouters LSKo Damme Oostkust DaOo Mandelvallei MaVa De Bekkevoortse BeVo Maxburg MaBu De Beneden Marck BeMa Meerdaal Honsem MeHo De Harelbeek Heulebeekvalleien HaHe Meetjesland Noord MeNo De Hazebeek HaBe Moerstuiver Damkeuken MoDa De Kemmelbeek KeBe Molenbeersel MoBe De Middenkustvallei MiVa Nederbrabant NeBr De Zavelputten ZaPu Noorderkempen NoKe Dendervallei DeVa Oost Pajottenland OoPa Dendervallei West Pajottenland DeWP Paddegat PaGa Driekoningen DrKo Patrijzenkuilen PaKu Essen Essen Rond Kale, Lieve & Burggrave RKLB Etbos Etbos ROW ROW Groot Brecht GrBr Scheldeoord vzw ScOO Havengebied Gent HaGe Scheldevallei ScVa Hoppeland HopLa Tussen Ijzer en houtland IJHo Houtland HouLa Tussen Voer & Yse TuVY Ieperlee IeLe Velpedal VeDa InFlanders Fields IFFi Walenbos WaBo Jekervallei JeVa Westhoek WeHo Kinrooi Kin
///////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////////// www.vlaanderen.be/inbo doi.org/10.21436/inbor.62125143 Pagina 57 van 57 Bijlage 3 Overzicht genetische structuur wilde individuen (per WBE) Figuur 16 Ordinatie plot (PCA) voor PC1 2 waarbij de symbolen de verschillende patrijzenindividuen voorstellen. De kleuren en symbolen geven aan tot welke wildbeheereenheden (Tabel 8) deze individuen behoren. Hoe dichter de afstand tussen individuen op de plot, hoe meer gelijkend deze zijn qua genetische samenstelling.