






BRÓCOLI
Pudrición blanda de la cabeza por bacterias y hongos
CEBOLLA
La amenaza de patógenos oportunistas transmitidos por vectores hemípteros
TOMATE
Temperatura y humedad, factores decisivos en la severidad del moho gris
PIÑA
Marchitamiento ocasionado por la cochinilla algodonosa, de etiología complicada
SOLANÁCEAS
Tizón tardío, enfermedad destructora de cultivos alimenticios de gran importancia
PEPINO
Prácticas culturales esenciales para obtener rendimientos adecuados
AGUACATE
Un fruto tropical que es sumamente apreciado a nivel mundial
TOMATE DE CÁSCARA
La variabilidad genética no suele garantizar un rendimiento constante
CÍTRICOS
La introducción de genotipos exóticos es causa de la presencia de clorosis variegada
HIGO
Frutal que se adapta bien a la sequía y altas temperaturas
CAMELIA SIMESIS
Enfermedades de hojas, tallo y raíz debidas a hongos patógenos en la planta de té
FERTILIZACIÓN
Influencia de los nutrientes sobre la resistencia de la planta a enfermedades
POLINIZACIÓN
Proceso esencial en la producción de cosechas de aliemntos
CUCURBITÁCEAS
Marchitamiento o pudrición de raíz y tallo por Pythium aphanidermatum
MANZANA
Impacto acumulativo del mildiú polvoriento en el rendimeinto de un año a otro




Temperatura y humedad en el desarrollo de infecciones por Pseudocercospora fuligena
El impacto negativo en la salud humana y el medio ambiente de la producción de alimentos
Factores que limitan el uso de plaguicidas convencionales

El riego foliar podría ser una fuente importante de contaminación bacteriana en productos frescos
Rendimientos mayores y mínimo desperdicio de agua, beneficios de los métodos modernos de riego
Programación del riego en función del monitoreo del suelo, la planta y el microclima
94 Congreso de Brócoli 2025 Una oportunidad de negocios en Puebla


El 2025 ha comenzado con un panorama complejo para la agricultura en México. La crisis hídrica que atraviesan diversas regiones del país no es una amenaza futura, sino una realidad que está impactando directamente en la producción agrícola. Estados como Chihuahua y Sinaloa enfrentan niveles críticamente bajos en sus presas y acuíferos, poniendo en jaque la producción de cultivos esenciales como maíz, frijol y trigo. La pregunta ya no es si la escasez de agua afectará al campo, sino cómo lograremos que el campo sobreviva a esta crisis.
Frente a este escenario, los productores han sido los primeros en reaccionar, buscando alternativas para mantener sus cultivos a flote. Desde la implementación de sistemas de riego tecnificado hasta la selección de variedades más resistentes a la sequía, los agricultores están demostrando una vez más su capacidad de adaptación. Sin embargo, estas soluciones deben ir acompañadas de políticas públicas eficientes y sostenibles. La perforación de pozos profundos y la reforestación de cuencas hidrográficas pueden ser paliativos, pero el reto real es la gestión inteligente del agua a nivel nacional.
El otro gran problema que enfrenta el sector es la caída en la producción de granos y la creciente dependencia de importaciones. En 2024, la producción nacional de
maíz se redujo en cuatro millones de toneladas, mientras que las importaciones superaron los 24 millones de toneladas. Esta tendencia no solo compromete la seguridad alimentaria del país, sino que también evidencia la urgencia de replantear la manera en que producimos nuestros alimentos. No podemos seguir dependiendo de insumos extranjeros mientras nuestro potencial productivo se ve limitado por la falta de infraestructura, financiamiento y acceso a tecnología.
A estos desafíos se suma la inseguridad en el campo, el alza en los costos de producción y las tensiones comerciales en torno al maíz transgénico. Todo esto conforma un panorama que, lejos de desmoralizar a los productores, debe servir como un llamado a la acción. Es momento de que el sector agrícola adopte estrategias de largo plazo: modernización del riego, diversificación de cultivos, uso de tecnologías de precisión y una mayor vinculación con centros de investigación para desarrollar soluciones viables ante la crisis climática.
El futuro del campo mexicano dependerá de nuestra capacidad para adaptarnos y transformar los desafíos en oportunidades. No podemos seguir viendo al agua como un recurso inagotable; es momento de gestionarla con inteligencia, innovación y responsabilidad. La agricultura no puede esperar más. Es hora de actuar.
CONSEJO EDITORIAL
Dr. ADALBERTO BENAVIDES MENDOZA, M.C. MARTÍN VALENCIA ACEVES, Ing. MANUEL VILLAREAL
Dr. JESÚS MARTÍNEZ DE LA CERDA, Ing. CARLOS DE LIÑÁN CARRAL
Edición 138 Febrero - Marzo, 2025
EDITOR
JAVIER BOLAÑOS CARREÑO javierbolcar@prodigy.net.mx
PUBLISHER
MARIBEL JARILLO OLGUÍN maribeljarillo@yahoo.com.mx
IDEA ORIGINAL DE REVISTA EDITORIAL DERIEGO, S.A. DE C.V. DISEÑO
MARICELA FLORES FLORES diseno.editorialderiego@gmail.com
CORRECCIÓN DE ESTILO ROSALBA TURNER rslbturner@hotmail.co.uk
PROYECTOS ESPECIALES
GERARDO POLANCO ARCE ventas.editorialderiego@gmail.com
SUSCRIPCIONES suscripciones.editorialderiego@gmail.com
COBERTURAS ESPECIALES NANCY HERNÁNDEZ nhmontes@gmail.com
FINANZAS
LUCÍA MUÑOZ PÉREZ lumupe3@hotmail.com
Suscripciones y Ventas de Publicidad Tel.: +52 (55) 2596 2850 suscripciones.editorialderiego@gmail.com
Escríbenos a: Revista deRiego Apdo. Postal 86-053, Ciudad de México, C.P. 14391, México.
deRiego, Año 24 Nº 136, Octubre - Noviembre, 2024, es una publicación especializada, editada por EDITORIAL DERIEGO, S.A. DE C.V., enfocada al sector agrícola. Se encarga de difundir las más avanzadas tecnologías de riego, nutrición y protección para la producción de hortalizas y frutas. deRiego se publica bimestralmente en los meses de diciembre, febrero, abril, junio, agosto y octubre. El costo del ejemplar es de $60.00 MXN, y la suscripción por 1 año es de $380.00 MXN / $95.00 USD. Tiraje de 12 mil ejemplares, distribuidos y editados para productores activos, profesionales, investigadores y académicos involucrados directamente en el sector; e instituciones oficiales y privadas. Certificado de reserva de derechos: 04-2011-072210295800-102. Certificado de Título y Contenido 15802. Registro SEPOMEX: PP09-1923. Los artículos publicados son responsabilidad de cada autor. deRiego no tiene injerencia en su contenido. Queda prohibida la total o parcial reproducción del contenido sin previa autorización por escrito del Director General.

La pudrición blanda bacteriana del brócoli aparece primero como una lesión empapada de agua, seguida de maceración y pudrición blanda del tejido. Ocurre principalmente durante períodos de temperaturas moderadas y alta humedad relativa o precipitaciones y se encuentra presente en todas las zonas de producción de brócoli en todo el mundo afectando tanto a los tejidos de la flor como a los del tallo.
Describiendo un grupo de enfermedades que causan más pérdidas de cultivos en todo el mundo que cualquier otra enfermedad bacteriana, las pudriciones blandas bacterianas dañan las partes suculentas de las plantas, como frutos, tubérculos, tallos y bulbos de plantas en casi todas las familias de plantas. Las podredumbres
blandas suelen afectar a hortalizas como la papa, la zanahoria, el tomate, las cucurbitáceas --pepinos, melones, calabazas, calabazas-- y los cultivos crucíferos --brócoli, col, coliflor, bok choy--. Estas enfermedades pueden ocurrir en los cultivos en el campo, así como en los cultivos cosechados en el almacenamiento. La putrefacción puede ocurrir en un amplio rango de temperaturas, con la peor descomposición entre 21 y 26° C y es particularmente grave cuando el oxígeno es limitado.
La necrosis o pudrición de floretes individuales o agrupados de brócoli se conoce como pudrición de la cabeza. La pudrición de la cabeza puede ser causada por varios patógenos, incluidas varias especies de bacterias y algunas especies de hongos. Los síntomas causados por estos patógenos pueden ser similares y puede ser difícil
La infección del tejido vegetal ocurre cuando hay agua libre.
distinguirlos. Sin embargo, la correcta identificación del agente causal es importante para garantizar que se utilicen los procedimientos de control adecuados.
Las pudriciones blandas son causadas por varias bacterias, más comúnmente especies de Pectobacterium, particularmente Pectobacterium carotovorum --anteriormente llamado Erwinia carotovora--, especies de Dickeya, particularmente Dickeya dadantii, anteriormente llamado Erwinia chrysanthemi, y ciertas especies de Pseudomonas, Bacillus y Clostridium. Estas bacterias pueden ingresar a las plantas a través de heridas causadas por herramientas, insectos y condiciones climáticas adversas como el granizo, así como a través de aberturas naturales. Las bacterias pueden propagarse de una planta a otra por insectos, en herramientas contami-


nadas o por el movimiento de restos de plantas infestadas, tierra o agua contaminada. Las pudriciones blandas bacterianas tienden a ser más problemáticas durante el clima húmedo y pueden ser más graves cuando las plantas carecen de suficiente calcio.
Las bacterias que causan la putrefacción blanda suelen entrar en el cuerpo de la planta a través de heridas y multiplicarse, inicialmente, en los espacios intercelulares. Las lesiones por congelación, las magulladuras de la cosecha y los pinchazos de insectos suelen ser las puertas abiertas de entrada. Las bacterias nunca pueden penetrar en la epidermis intacta, ni normalmente, en las aberturas naturales. Sin embargo, en cámaras de almacenamiento húmedas o en suelos frecuentemente húmedos, las raíces y los órganos de almacenamiento no lesionados pueden infectarse a través de las lenticelas. En cualquier caso, se
requiere una humedad muy alta, probablemente superior al 90%, para un progreso exitoso y continuo en las primeras etapas de la enfermedad.
Una vez que las bacterias han entrado y las temperaturas son cálidas, la humedad es alta, comienza la multiplicación en los espacios intercelulares. Tenga en cuenta que las bacterias se dividen y duplican su número tan rápidamente como cada 30 minutos. Las bacterias nunca son capaces de penetrar en las células vivas, pero producen enzimas pectolíticas que se difunden antes que las bacterias y disuelven la lámina media. Esta sustancia, compuesta de pectatos, cementa las células. La destrucción de esta sustancia cementante provoca la separación de las células. Una vez privadas de continuidad, y posiblemente envenenadas por las enzimas pectolíticas y otros productos metabólicos de las bacterias y las células vegeta-
les moribundas, las células mueren. y se desintegran. Las células vegetales muertas y en desintegración proporcionan alimento para siempre· hordas crecientes de bacterias. Una cucharadita de papa podrida puede contener millones de bacterias. Aunque son altamente patógenas y capaces de causar la muerte y la destrucción, estas bacterias no son parásitos muy buenos, sino saprobios que viven de los desechos de las células muertas.
Debido a que las medidas de control químico no han sido consistentemente efectivas y no se han identificado cultivares resistentes bien adaptados a nuestro clima local y aceptables para los procesadores, otras estrategias culturales para el control de enfermedades justificaron la investigación. Sin embargo, el riego por aspersión
también puede favorecer el desarrollo de enfermedades al modificar el microclima del cultivo. Bajo riego por aspersión, se ha medido un aumento de la humedad relativa, una disminución de las temperaturas y períodos de rocío diurno más largos en el dosel del cultivo. Estos cambios favorecen el crecimiento bacteriano y el desarrollo de enfermedades. Por lo tanto, los ajustes en las prácticas de riego durante el espigado del brócoli pueden proporcionar cierto control de las enfermedades. El riego también debe ser adecuado durante el inicio y el crecimiento de la cabeza para obtener rendimientos satisfactorios de brócoli.
Los síntomas de la pudrición bacteriana de la espiga comienzan como áreas empapadas de agua en pequeños grupos de flores sin abrir en cabezas inmaduras. Estas flores infectadas se vuelven necróticas y se vuelven de color marrón a negro. Los síntomas comienzan a extenderse a los floretes circundantes y otras áreas de la cabeza, afectando eventualmente a grandes sectores de la cabeza. Se desarrolla una podredumbre blanda en los tejidos infectados, lo que significa que los tejidos se vuelven blandos y viscosos. Un mal olor a menudo se asocia con esta putrefacción. Después de la infección inicial, los patógenos bacterianos y fúngicos secundarios pueden invadir, causando una mayor putrefacción del tejido.
El manejo de la pudrición bacteriana de la cabeza puede ser difícil. Existen

diferencias varietales en la susceptibilidad a esta enfermedad. Las variedades que producen cabezas en forma de cúpula tienden a ser menos susceptibles que las variedades con cabezas planas, ya que arrojan agua más fácilmente.
• Una rotación de tres a cinco años a cultivos no brassica y la eliminación de las pilas de descarte en el campo pueden ayudar a reducir la abundancia de inóculo en el campo
• Seleccionar campos con buen drenaje, plantar en camas elevadas y aumentar el espacio entre plantas ayudará a mantener las plantas secas y reducirá el número y la duración de los períodos de infección. Retrasar la siembra hasta que las temperaturas comiencen a calentarse también ayudará a reducir la incidencia de la pudrición bacteriana de la cabeza. Evite el uso de riego por aspersión
La pudrición blanda de los tallos ocurre comúnmente en plantas como el tabaco, las coles, el brócoli y los tomates.
o programe riegos en la mañana para momentos en que las plantas se sequen rápidamente
• Evite la fertilización excesiva con nitrógeno (N) en variedades susceptibles.
Sin embargo, las altas tasas de N no parecen afectar a las variedades resistentes a las enfermedades en la misma medida que a las variedades susceptibles. No se ha demostrado que las aplicaciones de bactericidas sean efectivas para controlar la pudrición bacteriana de la cabeza. Los tensioactivos presentes en muchos productos insecticidas (o ayudas de tratamiento recomendadas) pueden aumentar el nivel de pudrición bacteriana de la cabeza, por lo que los insecticidas deben aplicarse antes de la aparición de esta enfermedad en áreas donde ocurre comúnmente.
Las enfermedades de la pudrición blanda se pueden reducir utilizando






Nuestra tecnología Proteomic Plug, incrementa el grosor de los tallos, asegura el correcto crecimiento de pella, favorece el desarrollo radicular y elimina el problema de tallo hueco con la correcta nutrición







@AFLAGRO @aflagro
• Plantar en terrenos bien drenados o proporcionar un drenaje adecuado
• Evite las poblaciones excesivas de plantas para que las plantas se sequen más rápido
• En el caso de las patatas, compre semillas libres de enfermedades que no tengan len tícolas agrandadas
• Los cuchillos de corte para trozos de semillas de papa deben estar limpios. Si las piezas de semillas enfermas se cortan inadvertidamente, limpie los cuchillos con alcohol isopropí-
Plante piezas de semilla sube rizadas que se traten con un fungicida recomendado para piezas de semilla. La suberización ocurre más rápido cuando las piezas de semilla se almacenan a alta humedad y temperaturas cálidas. Nunca se debe permitir que los trozos de semilla de papa se mojen. Las áreas de almacenamiento deben estar sombreadas y bien ventiladas
• Las plantas deben estar secas al cosechar, podar, retoñar, atar o desmochar
• Pode con prontitud, ya que la eliminación de los chupones
más grandes da como resultado heridas más grandes que son más propensas a
• Cuando cubra el tabaco, use un cuchillo afilado y corte el tallo en ángulo para minimizar la acumulación de agua en el tallo expuesto. Limpie el cuchillo con frecuencia con alcohol isopropílico y limpie el cuchillo inmediatamente si se corta una planta infectada. No rompa los tallos con la mano. Mejor aún, no toque las plantas infectadas
• Cultive variedades resistentes cuando sea posible.

La amenaza de patógenos oportunistas transmitidos por vectores hemípteros
POR PEDRO BEDOLLA CONTRERAS
La pudrición del bulbo producida por patógenos fúngicos o bacterianos es una enfermedad capaz de causar pérdidas de cebolla en almacenamiento del orden de 16 millones de dólares cada año tan solo en los Estados Unidos.
Los tejidos internos de las plantas suelen estar habitados por comunidades microbianas endófitas que interactúan con las plantas huésped como simbiontes, comensales o patógenos. Las plantas y sus endófitos suelen existir en un estado de equilibrio pero si la relación se ve perturbada significativamente por factores externos o internos, la interacción puede interrumpirse, lo que resulta en un cambio de un estilo de vida mutualista a uno patógeno del endófito. Estos patógenos se consideran oportunistas. En el caso de la pudrición de los bulbos de cebolla, la alta temperatura típicamente asociada con el curado en el campo en regiones cálidas o con el
curado poscosecha es propicia para el crecimiento y desarrollo de bacterias termófilas que son patógenos de los bulbos de cebolla.
Responsables de algunas de las pérdidas más devastadoras en cultivos económicamente importantes, la alta especificidad y patogenicidad de las bacterias, les permiten colonizar plantas hospedadoras específicas y han evolucionado para aprovechar los insectos como vectores para una diseminación efectiva a las plantas hospedadoras preferidas. La mayoría de ellos están limitados por floemas, con algunas bacterias limitadas por xilemas que proliferan en los elementos del tamiz e incluyen miembros de los géneros fitoplasmas, Spiroplasmas, liberibacters y Xyllela, que son transmitidos por hemípteros, sobre todo chicharritas y psílidos. Una vez ingeridos por el insecto vector, estos patógenos se desplazan por todo el cuerpo del insecto antes de transmitirse a una nueva planta huésped en un proceso conocido como transmi-
Los microbios se encuentran en comunidades complejas dentro de las plantas como endófitos y establecen una red de interacciones que pueden influir positiva o negativamente en la salud de las plantas
sión circulativa. Esto es beneficioso para las bacterias para una rápida propagación y transmisión a nuevos huéspedes de plantas. Los patógenos circulantes de las plantas comparten regulaciones moleculares generales similares, aunque el mecanismo exacto del movimiento del patógeno puede variar de una especie de bacteria a otra. Una mayor comprensión de la supervivencia del patógeno en el vector y la manipulación del vector es fundamental para controlar las enfermedades bacterianas transmitidas por vectores. Esta revisión se centra en las diferentes rutas de transmisión bacteriana y la participación de diferentes factores moleculares bacterianos necesarios para la supervivencia y diseminación dentro del insecto huésped.
La cebolla, Allium cepa L., es una hortaliza de gran importancia económica y uno de los cultivos más valiosos del mundo. La producción mundial actual se estima en 106,6 millones de toneladas, habiendo aumentado casi un 25%

en los últimos 10 años. Los mayores productores son India, 25.99%; China, 22.67%; Egipto, 3.11%; Estados Unidos, 2.91%; y Turquía, 2.55%. Estados Unidos produjo casi 3.1 millones de toneladas de bulbos de cebolla en 2022, con un valor económico de 1,630 millones de dólares. Aproximadamente el 68% de la cosecha anual producida se destina al almacenamiento, según datos del Departamento de Agricultura de los Estados Unidos, de 2017.
DIAGNÓSTICO MODERNO DE ENFERMEDADES BACTERIANAS EN HORTALIZAS
Nuevas tecnologías y la actualización taxonómica de patógenos, permiten que la identificación de los diferentes agentes causales se logre de manera más precisa. La co-ocurrencia de múltiples patógenos en una planta huésped puede aumentar la gravedad de algunas enfermedades de las plantas. Por ejemplo, varias especies de Pantoea pueden coexistir y coexpresar virulencia en los tejidos de la cebolla. La coinfección también puede ocurrir entre especies lejanamente relacionadas. Por ejemplo, el principal patógeno causante del nudo del olivo, Pseudomonas savastanoi pv. savastanoi, causa nudos más grandes cuando las plantas son cocolonizadas por las especies no patógenas Erwinia oleae, Erwinia toletana y Pantoea agglomerans.
Los principales patógenos de la pudrición del bulbo de la cebolla son las especies Burkholderia, Enterobacter, Pantoea y Pectobacterium. Sin embar-
go, algunos patógenos bacterianos oportunistas también se han asociado con la pudrición de los bulbos de cebolla, incluidos Bacillus amyloliquefaciens y especies de Dickeya, Pseudomonas, Rahnella y Serratia. Algunas de estas especies infectan las plantas de cebolla durante la temporada de campo y pueden causar síntomas foliares, desde los cuales las infecciones progresan por el cuello hasta el bulbo en desarrollo. Las semillas cosechadas de plantas sintomáticas y asintomáticas en cultivos de semillas de cebolla pueden albergar algunos de estos patógenos bacterianos de la cebolla.
Los bacteriófagos son ubicuos en la naturaleza y se ha demostrado que influyen en la estructura, función y evolución de las comunidades microbianas y que actúan como agentes de transferencia lateral de genes. En los estudios del microbioma humano, por ejemplo, los miembros dominantes de la microbiota comensal son los bacteriófagos que forman lo que se conoce como fageoma. La metagenómica viral de plantas centrada en el descubrimiento viral ha revelado una multitud de relaciones patógenas y mutualistas entre virus y plantas). Debido a la similitud en complejidad entre el microbioma intestinal y la rizosfera, los fagos podrían influir en la composición y la funcionalidad del microbioma de la rizosfera, incluidas las interacciones con la planta huésped.
Algunas de las enfermedades en plantas pueden brindar dinamismo de la

Las interacciones positivas pueden ser sinérgicas y conferir tolerancia al estrés abiótico y biótico
enfermedad en el hospedero. Estas asociaciones de microorganismos son diversas y pueden ocurrir entre bacterias y virus, bacterias y hongos y entre especies diferentes de bacterias, las cuales pueden llegar a producir cambios en la sintomatología y también a generar diagnósticos errados. Las enfermedades bacterianas pueden afectar plantas en cualquier estado de desarrollo y se incrementa conforme se desarrolla el sistema radical. Los síntomas usualmente se notan a los 60 días después de la siembra y difieren de acuerdo al estado de desarrollo de la planta y la duración de las condiciones favorables en el suelo, principalmente la temperatura.
El primer síntoma coincide con el período de bulbificación y se presenta como un amarillamiento general, continuado por muerte descendente de las hojas más externas y retardo del crecimiento. El deterioro gradual se da por varios días o semanas hasta concluir con el colapso final de las hojas y una pudrición basal seca o semiacuosa. Simultáneamente, en las raíces y hojas inferiores hay abundancia de micelio blanco, lanoso y superficial que pronto produce esclerocios negros y esféricos sobre la superficie o dentro de los tejidos enfermos. En este grado de desarrollo de la enfermedad las plantas afectadas son fácilmente arrancadas del suelo.


POR CESAR GARRIDO LÓPEZ
El moho gris del tomate es una enfermedad devastadora causada por B. cinerea y que resulta en la pudrición del tomate, causando un daño del 20 al 40% o que incluso puede alcanzar del 50 al 70% durante los años epidémicos. La temperatura y la humedad afectan en gran medida el desarrollo del moho gris del tomate. Por lo tanto, es de gran importancia explorar la mitigación del moho gris mediante la regulación de la temperatura y la humedad ambientales.
La mayoría de los estudios de laboratorio utilizaron la exposición a largo plazo al estrés por calor; por ejemplo, temperaturas ligeramente elevadas de 28° C durante un día. Los cambios intermitentes de temperatura y humedad pueden tener diferentes efectos
en las interacciones entre plantas y patógenos. En el presente estudio se establecieron diferentes combinaciones de temperatura y humedad para estudiar la influencia de la regulación de la temperatura y la humedad en el desarrollo del moho gris del tomate. Proporcionamos una cierta base teórica y una importancia práctica para suprimir las enfermedades de las plantas mediante el cambio del entorno del invernadero.
La reducción de las enfermedades de las plantas mediante el control de la temperatura y la humedad del invernadero es cada vez más importante en el campo del control ecológico. Entre los factores ambientales, la temperatura y la humedad son los que más afectan a las interacciones entre

plantas y patógenos. La alta humedad a menudo se asocia con brotes de enfermedades, y los cambios de temperatura afectan considerablemente el crecimiento, la germinación, la supervivencia y la virulencia de los patógenos. Para la mayoría de las enfermedades de las plantas causadas por patógenos necrótrofos, la disminución de la humedad o el aumento de la temperatura pueden inhibir en gran medida el desarrollo de la enfermedad.
La temperatura y la humedad afectan notablemente el proceso de infección de patógenos y la respuesta de defensa de las plantas. Las plantas son más susceptibles a los patógenos en condiciones de alta humedad. Con el aumento de la humedad del aire, la virulencia de Sclerotinia spp. aumenta y, cuando la humedad supera el 80%, la incidencia de lechuga es más alta. Los conidios de Botrytis cinerea no pueden germinar y la infección se inhibe cuando la humedad relativa (HR) es inferior al 80%.
El aumento de la temperatura promueve la necrosis del tejido vegetal y facilita la colonización de patógenos necrótrofos. La alta temperatura de 35 ° C aumenta la virulencia de Pectobacterium artosepticum, y la virulencia se correlaciona significativamente con las enzimas que degradan la pared celular de la planta. Las altas temperaturas inhiben la formación de pilus y la expresión de genes de virulencia de las cepas de Agrobacterium , lo que resulta en el fracaso de la infección.
La temperatura y la humedad juegan un papel importante en las interacciones entre plantas y patógenos

En King Seeds encontrarás un amplio catálogo de variedades de semillas de hortalizas. Pregunta por las opciones que tenemos para ti.








El moho gris es una enfermedad que prospera a bajas temperaturas y alta humedad. La temperatura óptima para la infección por B. cinerea es de 23° C, y los ambientes de alta humedad con HR > 80% son propicios para la aparición de la enfermedad.
Las temperaturas superiores a 30 °C pueden reducir la aparición de moho gris. El aumento de la temperatura de la región de la raíz del tomate a 28 °C puede mejorar la expresión de genes de defensa y la erupción de especies reactivas de oxígeno (ROS), inducir inmunidad al tomate y mejorar la resistencia del tomate a B. cinerea.
SÍNTOMAS VISIBLES DE LA INFECCIÓN DE B. CINEREA EN EL CULTIVO
El moho gris afecta a todas las partes aéreas de la planta de tomate, in-


cuando la infección se produce en el borde. A medida que la enfermedad progresa, todo el folíolo se marchita y se quema con el tiempo. Las flores marchitas son muy susceptibles al moho gris y pueden servir como punto de partida para la colonización de los folíolos cuando caen sobre las hojas. Las cicatrices de las hojas y las heridas de poda son entradas comunes de infección del tallo. Los síntomas en los tallos aparecen como lesiones de color marrón oscuro y cancros que pueden causar el anillamiento de los tallos y el marchitamiento por encima del área infectada de la planta. En la fruta, los
una podredumbre blanda empapada en agua. Eventualmente, toda la fruta infectada se pudre en el campo o durante el transporte poscosecha. Cuando los pequeños frutos verdes se infectan directamente por esporas en el aire, los síntomas aparecen como manchas blancas y circulares llamadas "manchas fantasma". Aunque pueden afectar la comerciabilidad de la fruta, las manchas fantasma generalmente no provocan la pudrición de la fruta. En condiciones húmedas, el hongo forma moho gris difuso en la superficie de las hojas, flores, tallos y frutos enfermos, que es un signo ca-

racterístico de la enfermedad y una característica diagnóstica para distinguir el moho gris de otras enfermedades bióticas y abióticas del tomate.
El agente causante del moho gris, Botrytis cinerea, es un patógeno fúngico débil que tiene una gama de huéspedes extremadamente amplia con más de 200 especies reportadas en vegetales y plantas ornamentales. El hongo sobrevive como micelios y/o esclerocios en plantas perennes, en el suelo y en restos de plantas entre estaciones. Las esporas de hongos se transportan fácilmente por el aire y pueden ser transportadas de un campo a otro. Las esporas germinan y penetran en los tejidos de las plantas a través de heridas causadas por insectos o lesiones físicas. La germinación e infección de las esporas no requiere un período prolongado de alta humedad. De hecho, las condiciones húmedas dentro del dosel del tomate por la noche suelen ser suficientes para la

infección. Por lo tanto, las plantaciones densas favorecen el desarrollo de enfermedades debido al movimiento limitado del aire y la alta humedad relativa dentro del dosel. Las condiciones relativamente frescas y húmedas a principios de primavera y finales de otoño son favorables para el desarrollo de la enfermedad.
EMPLEO DE BIOAGENTES MICROBIANOS
CONTRA LA AMENAZA DEL HONGO
En décadas anteriores, el control de B. cinerea ha dependido principalmente de fungicidas químicos. Sin embargo, la evidencia revela que B. cinerea ha desarrollado cierta resistencia a muchos fungicidas, incluidos los anilinopirimidinas, las dicarboximidas, la benzamida, la fenhexamida, el diethofencarb, la procimidona, el pirimetanil y la hidroxianiilida. Además, una cepa de B. cinerea tiene múltiples tipos de resistencia a fungicidas que se pueden encontrar en las tierras de cultivo. Hasta la fecha, el abuso y el uso excesivo de fungicidas han causado contaminación ambiental y alteración de los ecosistemas ecológicos, además de representar un peligro para la salud humana debido al alto efecto residual de los fungicidas en la fruta del tomate. Por lo tanto, existe una demanda urgente de medidas sustitutivas que puedan controlar de manera segura y eficiente el moho gris del tomate.
El control biológico es, sin duda, una buena opción. En general, los bioagentes microbianos, por ejemplo, hongos, bacterias y levaduras, pueden controlar eficazmente las enfermedades de las plantas a través de la antibiosis, la competencia por lugares y
Las epidemias graves de la enfermedad pueden causar daños significativos en las hojas, cancro del tallo y pudrición de la fruta, lo que resulta en una grave reducción del rendimiento durante la producción y la poscosecha
nutrientes, los compuestos volátiles, el parasitismo y el inicio de la defensa de las plantas. M. anisopliae es un hongo entomopatógeno ampliamente distribuido con una inmensa eficacia contra diversas plagas de insectos como langostas, trips, moscas blancas y garrapatas. Recientemente, Meterhizium spp. también mostró múltiples roles en el ecosistema como endófitos, promotores del crecimiento de las plantas y antagonistas de patógenos de las plantas. M. anisopliae reveló expresión diferencial de genes que codifican enzimas hidrolíticas, péptidos antimicrobianos, toxinas, proteínas de la pared celular, así como otras moléculas. Los metabolitos secundarios generados por este hongo exhiben diversas propiedades insecticidas, anticancerígenas, antioxidantes y antimicrobianas. También se han sugerido como potenciales sustitutos para la mejora de nuevos agentes bioactivos. Si una cepa fúngica conserva acciones duales contra las plagas de insectos y también contra los patógenos de las plantas, esto sería una potente ventaja para la protección de los cultivos. Por lo tanto, el hongo entomopatógeno en el estudio actual, M. anisopliae, fue inspeccionado por sus actividades antifúngicas contra B. cinerea.
Pocos estudios previos han presentado el efecto inhibidor de M. anisopliae sobre B. cinerea. Sin embargo, aún se desconoce la eficiencia del biocontrol in vivo y el posible mecanismo del proceso de inhibición. El presente estudio evaluó los efectos antagónicos de M. anisopliae sobre B. cinerea utilizando filtrado de cultivo, extracto crudo y compuestos orgánicos volátiles (COV), además de explorar los mecanismos de inhibición. Además, se evaluó la eficacia de M. anisopliae para la reducción de la gravedad de la enfermedad mediante la evaluación en hojas desprendidas, plantas de tomate enteras, así como frutos maduros después de la cosecha.

La piña, Ananas comosus, es una monocotiledónea xerófita, planta tropical frutal perenne que pertenece a la familia Bromeliaceae la cual cuenta con más de 2 mil 500 especies. Es una fruta reconocida como una excelente fuente de minerales y vitaminas; produce una gran cantidad de calcio, potasio, glucosa, la enzima digestora de proteínas bromelina, fibra, vitamina A, B y C

Marchitamiento ocasionado por la cochinilla algodonosa, de etiología complicada
POR PABLO ESCUDERO SÁNCHEZ
En el cultivo de la piña, los materiales de siembra utilizados para la producción comercial suelen ser la corona desde la parte superior de los frutos. Las coronas plantadas, conocidas como cultivo de plantas, producirán cada una sola fruta de piña de 18 a 20 meses después de la siembra. Después de la cosecha de cultivos vegetales, se desarrolla un cultivo de sotones a partir de retoños producidos en el tallo de las plantas de cultivo. El fruto de la cosecha de soca se cosecha en aproximadamente 12 a 15 meses.
La piña se siembra típicamente en densidades de 60 mil a 90 mil plantas/ ha y requiere de 15 a 20 meses para producir el primer fruto. Después del cultivo de la planta, la planta madre que produjo el fruto declina y da lugar a una o dos socas. La primera cosecha de soca producirá frutos en 12 a 15 meses. Los cultivos de segunda y tercera soca no son comunes debido a la acumulación de nematodos, enfermedades fúngicas y bacterianas, y MWP. Las plantas de piña infectadas con PMWaVs pueden ser asintomáticas. Por lo tanto, son necesarios ensayos específicos de detección de virus para detectar infecciones latentes.
El ciclo de producción relativamente largo desde la siembra de las espigas o coronas hasta la cosecha de la fruta hace que el cultivo sea vulnerable a la infestación de cochinillas, la propagación de virus y el MWP. El material de propagación utilizado para establecer nuevos campos se recoge normalmente de los campos existentes. Esta práctica contribuye a un aumento de la incidencia del virus a lo largo del tiempo. En la actualidad, la indexación viral de la gran cantidad de deslizamientos y coronas necesarios para establecer los campos no es práctica; por lo tanto, se necesitan estrategias alternativas para minimizar el aumento de los PMWaV.
La piña puede verse afectada por muchas plagas y patógenos diferentes. Entre los más graves se encuentran los nematodos fitoparásitos y el complejo de closterovirus asociados con la marchitez de la piña (MWP). Rotylenchus reniformis y Meloidogyne javanica son los principales nematodos fitoparásitos que se encuentran en las áreas de cultivo de piña. R. reniformis reduce el rendimiento de los cultivos vegetales y devasta los primeros cultivos de soca. Los efectos de los nematodos se mitigan mediante un barbecho entre ciclos, fumigación del suelo antes de la siembra y aplicaciones de nematicidas posteriores a la planta.
Ejemplo del riego que esta enfermedad supone, la cochinilla casi destruyó la industria de la piña hawaiana a principios de 1900 y continúa representando una amenaza para los rendimientos. Las cochinillas son vectores del virus y las hormigas juegan como agentes para propagar el vector en las granjas de piña. Las cochinillas proporcionan una fuente de alimento llamada melaza a las

hormigas, y las hormigas brindan protección contra los enemigos naturales y el transporte a este vector. Se han identificado varias especies de virus distintas que están asociadas con esta enfermedad de marchitez en la piña. Se han encontrado dos closterovirus distintos en plantas de piña de Hawái y de todo el mundo.
SÍNTOMAS DE LAS MARCHITEZ LENTA Y RÁPIDA
En general, los síntomas de la enfermedad de la cochinilla de la piña (PMW) se pueden clasificar en dos tipos, que son la marchitez lenta y la marchitez rápida. Lo más probable es que el síntoma de marchitez sea el resultado de la destrucción del tejido de las hojas
Dado que las piñas se cultivan en fase vegetativa, es probable que los virus de este importante cultivo estén muy extendidos
como resultado de la alimentación de un gran número de cochinillas. Mientras tanto, la marchitez rápida se observa aproximadamente dos meses después de un período repentino de alimentación por muchas cochinillas. Los síntomas de la marchitez rápida en plantas de hasta seis meses de edad incluyen el cambio de color en las hojas internas, ya sea demasiado amarillas o rosadas (varía según las variedades), pérdida de turgencia, pardeamiento de las puntas de las hojas (muerte regresiva), enroscamiento y desecación. Tanto en la marchitez rápida como en la marchitez lenta, la planta puede morir, no producir frutos o producir una forma malformada o frutos más pequeños que los frutos normales si la planta no se recupera. En el caso de la planta recuperada de las enfermedades, las nuevas hojas pueden crecer fuera del centro de la planta. En una infección grave, la enfermedad se propaga gradualmente de las hojas a las raíces, lo que provoca el colapso de la raíz.
La incidencia del virus asociado a la marchitez de la cochinilla de la piña1 (PMWaV-1) varía del 20 al 100% en apariencia saludable. Las incidencias de infección por PMWaV-2 son más
bajas, oscilando entre el 0 y el 36% en las mismas selecciones. Las coronas recolectadas de estas selecciones se utilizan para establecer cultivos de plantas y, si están infectadas con PMWaVs, sirven como fuente principal de virus en el campo. Las infecciones se han perpetuado a través de esta propagación clonal de la piña durante décadas. Tanto la PMWaV-1 como la PMWaV-2 se transmiten por la cochinilla. Tanto el PMWaV-2 como las cochinillas desempeñan un papel en la inducción de los síntomas de la MWP, mientras que el PMWaV-1 no parece ser necesario para el desarrollo de los síntomas. Aunque la infección por PMWaV-1 puede no estar directamente involucrada en el MWP, su presencia

Un complejo de virus asociados a la marchitez es la cochinilla de la piña, PMWaV, culpable de la producción de rendimientos reducidos y marchitez de la planta
se correlaciona con la reducción del crecimiento en el cultivo de plantas y reduce el rendimiento de la fruta en el cultivo de soca. Los nematodos y la infección por virus afectan la rentabilidad y el cultivo continuo de la piña.
En la actualidad, las plantas de piña se clasifican como "sanas" o "enfermas", sobre la base de una evaluación de los síntomas visuales de la enfermedad del marchitamiento de la cochinilla. En el futuro, si las investigaciones adicionales sobre la etiología de la cochinilla arrojan un ensayo definitivo, puede ser posible hacer una clasificación similar sobre una base más rigurosa. Sin embargo, dada la naturaleza sistémica de la marchitez de la cochinilla, parece probable que cualquier método de evaluación alternativo que se avecina, al igual que el presente, proporcione datos sobre la incidencia.
Los impactos de las interacciones entre el estrés de las plantas no son infrecuentes. Se ha documentado que las interacciones de los nematodos con hongos, bacterias y virus provocan sinérgicamente ciertas enfermedades en algunas especies. Por ejemplo, Meloidogyne y Fusarium juntos pueden exacerbar el marchitamiento del algodón y el tabaco, y Criconemella y Pseudomonas syringae pv.


Tizón tardío, enfermedad destructora de cultivos alimenticios de gran importancia
POR MÓNICA GUZMÁN LARA
Una de las enfermedades más devastadoras en la agricultura y, sin duda, la enfermedad más devastadora en papa y muchas otras solanáceas, es el tizón tardío causado por Phytophthora infestans. En el caso de la papa, se ha estimado que la enfermedad causa más de 6 mil millones de dólares en pérdidas y costos de manejo anualmente. No solo se destruye el follaje sino que los tubérculos de la papa también pueden infectarse. La enfermedad es al menos tan destructiva en los tomates como en la papa. Es capaz de destruir las plantas muchas veces en cuestión de horas. Las plantas de tomate han sido destruidas casi por completo por el tizón tardío, así como muchas plantas vegetales que se ven afectadas por esta devastadora enfermedad.
Los ciclos reproductivos asexuales son responsables de epidemias devastadoras. Como oomiceto, P. infestans produce esporangios que pueden germinar directamente para producir un tubo germinativo, o indirectamente para producir zoosporas. Después de un corto período de motilidad, minutos a horas, las zoosporas se enquistan y germinan a través de un tubo germinativo. Si las zoosporas están en el tejido del huésped, el tubo germinativo puede penetrar en el huésped e iniciar infecciones. La esporulación se produce a partir de lesiones y es estimulada por condiciones húmedas a temperaturas moderadas (15 a 22° C). Una sola lesión puede producir varios cientos de miles de esporangios, que se dispersan por el aire. La reproducción asexual también puede conducir al desarrollo de linajes clonales. Los individuos de un linaje clonal se derivan de un solo
evento de recombinación y difieren entre sí solo por mutación o recombinación mitótica. Los miembros de un mismo linaje clonal son generalmente fenotípicamente iguales entre sí.
Dado el potencial devastador de este patógeno, es fácil entender la atención que recibe. Sin embargo, el patógeno y la enfermedad han surgido y resurgido tantas veces que podría ser lógico concluir que no se puede decir nada nuevo sobre ellas, por ejemplo desde la tragedia histórica asociada a esta enfermedad como lo fue la hambruna irlandesa de la papa. La tecnología actual permite a los investigadores responder a algunas preguntas de importancia histórica. La devastación causada por el patógeno continúa apareciendo en nuevos lugares sorprendentes o con una intensidad sorprendentemente nueva. Las poblaciones de P. infestans en todo


el mundo están en constante cambio, con cambios que tienen importantes implicaciones para el manejo de enfermedades. La revolución de la genómica ha permitido a los investigadores hacer enormes progresos en términos de comprensión de las interacciones huésped-patógeno.
La enfermedad apareció muy repentinamente a mediados del siglo XIX. La suposición inicial fue que P. infestans se originó en los Andes de América del Sur junto con la papa. Sin embargo, había algunos escépticos, entre ellos estaba Donald Reddick de la Universidad de Cornell (Reddick 1928). Reddick pensó que si P. infestans hubiera sido endémico de América del Sur, habría sido observado allí por botánicos europeos, pero no encontró tales informes. Once años más tarde y basándose en parte en el hecho de que las especies nativas de Solanum en México son en gran me-
dida resistentes a P. infestans, Reddick estaba dispuesto a sugerir que México era el centro de origen de P. infestans. La idea de que el centro de México podría ser el centro de origen de P. infestans cobró mucho impulso cuando se descubrió en la década de 1950 que la población de P. infestans en el Valle de Toluca, en el centro de México, era sexual, conteniendo los tipos de apareamiento A1 y A2. Antes de ese momento, P. Se pensaba que los infestans eran exclusivamente asexuales. La demostración de que la población en el centro de México era muy diversa genotípicamente consolidó aún más la opinión de que el centro de México era el centro de origen de esta especie.
Esta hipótesis prevaleció hasta principios del siglo XXI, cuando un estudio de Gómez-Alpizar et al. (2007) sobre genealogías de genes mitocondria-
les y nucleares de aislados de varias localidades del mundo llevó a estos autores a concluir que P. infestans tenía un origen sudamericano. Su informe reavivó la controversia, pero un estudio posterior, utilizando un conjunto más amplio de aislados e incluyendo parientes más cercanos de P. infestans, nuevamente llevó a la conclusión de que las tierras altas del centro de México son el centro de origen. Este último estudio concilia observaciones previas sobre la diversidad genética, el rango de huéspedes y la historia natural del patógeno.
En 2009, una pandemia del tizón tardío del tomate en el este de los Estados Unidos desafortunadamente introdujo a muchos productores orgánicos y jardineros domésticos a la enferme-

dad del tizón tardío. Parece probable al menos para los Estados Unidos que esta epidemia haya introducido a más patólogos no vegetales a la enfermedad y al patógeno que cualquier otro evento reciente.
Las plantas son capaces de detectar la presencia de patógenos en su superficie, percibiendo señales químicas y físicas del origen del patógeno y reaccionan rápidamente al intento de infección. Las reacciones tempranas de las células epidérmicas localizadas incluyen reordenamientos importantes dentro del citoplasma
a la invasión de oomicetos, que van desde células invadidas sin respuesta visible a reacciones más localizadas que impiden la formación de estructuras intracelulares de patógenos, hasta HR y muerte celular programada, que ocurre solo después de que el crecimiento del patógeno haya procedido a la formación de un haustorio claramente identificable.
La línea principal de defensa inducida en las plantas de tomate contra P. infestans es la resistencia contra la penetración en la superficie de la hoja; esta es una estrategia de defensa altamente efectiva movilizada rápidamente por las plantas hospederas para intentar penetrar. Estudios recientes han demostrado la participación directa de las proteínas receptoras unidas a la membrana plasmática en el reconocimiento de elicitores de patógenos apoplásticos y la activación de las defensas del huésped.

Durante la fase temprana de crecimiento biotrófico en P. infestans, los nutrientes se obtienen de células vegetales vivas
Para proteger los cultivos de papa y tomate, se han llegado a utilizar más de 2 mil 700 toneladas de fungicidas para suprimir esta enfermedad
El tomate es susceptible a muchas enfermedades, como las causadas por Phytophthora infestans (Mont.) de Bary, Alternaria solani (Ell. & Mart.) Jones & Grout y Ralstonia solanacearum Smith, que son algunos de los patógenos más conocidos, por las pérdidas entre el 40 y 70% que ocasionan en las solanáceas. El tizón tardío es causado por el Chromista Phytophthora infestans, común en zonas con temperaturas entre 15 y 22° C y humedad relativa mayor al 80%. El patógeno, se transmite a través de semillas de tomate y puede sobrevivir en forma de micelio en otras plantas cultivadas o arvenses de la familia de las solanáceas, o en residuos de cosecha, que permanecen en el suelo. La infección en el fruto puede oscilar del 41 al 100% en los campos sin aplicación de fungicidas y del 12 al 65%, en parcelas protegidas con fungicidas sistémicos. La resistencia genética es la capacidad que tiene el hospedante de contrarrestar la acción de un patógeno; esta es la estrategia más ampliamente recomendada y estudiada en el manejo integrado de plagas. Los parientes silvestres son una fuente vital de genes de resistencia contra enfermedades, como el tizón tardío, lo cual, ayuda a asegurar que exista una adecuada diversidad genética en el patrimonio genético, de un cultivo en particular.


Los conidios permanecen viables durante 6 meses en condiciones secas en un invernadero. Los conidios en equipos agrícolas sirven como inóculo principal en invernaderos comerciales
La temperatura óptima para el crecimiento micelial del moho de la hoja negra por Pseudocercospora fuligena, suele ser de 26° C, mientras que a 34° C, según quedó establecido en estudios realizados, no se observó crecimiento abundante. Por otra parte, se produjeron abundantes conidios entre 18° C y 22° C en medio de agar de cocción de tomate, pero fueron escasos a 28° C y ausentes a ≥30° C.
Se ha encontrado que los conidios germinan en agua libre y alta hume-
dad relativa de 91.0% pero no al 84.5% de HR o menos. La tasa de germinación de conidios en placas de PDA o agar agua es significativamente mayor que en agua esterilizada o en agua libre en hojas de tomate. Los conidios expuestos a 50° C durante 5 min o más no germinan. Los conidios germinan con mayor frecuencia a partir de células apicales y basales. El crecimiento micelial es muy lento en todos los medios, incluidos el PDA, el agar jugo V8, el agar zanahoria y el extracto de hoja de tomate mezclado con extracto de avena (TOA). La temperatura óptima para el crecimiento micelial en
medio sólido y en medio líquido es de 28° C, y no se produce crecimiento a 36° C. El mejor crecimiento micelial de P. fuligena en cultivo se produce con glucosa o almidón como fuente de carbono, con DL-valina como fuente de nitrógeno. Un fotoperiodo de 12 h al día mejora significativamente la esporulación en cultivos en placa TOA incubados a 28° C.
La concentración óptima de conidios para la inoculación en condiciones controladas es de 5 mil conidios/ml. Cuando las plantas inoculadas se mantienen bajo alta humedad durante

más de 4 días, la gravedad de la enfermedad aumenta significativamente. La germinación de los conidios en hojas de tomate desprendidas fue de 71 y 99% después de 6 y 48 h de incubación, respectivamente. La germinación fue más rápida dentro de 5-10 h de exposición. Los tubos germinativos del 10 y 31% de los conidios germinados penetraron en los estomas después de 72 y 120 h de incubación, respectivamente. Las lesiones fueron visibles por primera vez 6 días después de la inoculación. Los conidióforos de color gris claro emergieron de las aberturas estomáticas 11 días después de la inoculación. Los conidióforos y conidios maduros se encontraron a los 12 días después de la inoculación. Los nuevos conidios están disponibles para la inoculación de ciclos secundarios de la enfermedad.
El moho de la hoja negra se desarrolla en condiciones de temperaturas cálidas, alta humedad relativa y largos períodos de humedad de las hojas, condiciones que son típicas de los trópicos húmedos y del cultivo protegido en climas tropicales y subtropicales. En condiciones naturales de campo, la presencia de humedad libre del rocío, la lluvia y la niebla puede proporcionar condiciones favorables para la germinación, penetración, infección y esporulación de P. fuligena. Durante la estación lluviosa y los meses más fríos del año, cabe esperar que la enfermedad se agrave. Otro estudio indicó que los tomates frescos del mercado plantados durante temperaturas cálidas en invernaderos de 50 mallas y expuestos a períodos prolongados de humedad son muy propensos a la infección por moho de la hoja negra en sus etapas jóvenes de crecimiento.
Los conidios podían sobrevivir hasta 6 meses en hojas secas almacenadas en macetas de barro en el interior. En un estudio reciente, se encontraron conidios viables en equipos de invernaderos que tuvieron un gran brote de moho negro el año anterior. Los conidios en el equipo agrícola sirven como inóculo primario, que la infección primaria y secundaria ocurren al mismo tiempo durante grandes brotes y que el inóculo primario tiene un papel importante en el ciclo de la enfermedad del moho de la hoja negra durante largos períodos en invernaderos comerciales.
P. fuligena probablemente sobrevive en los desechos de plantas enfermas y produce esporas en estos desechos al comienzo de la temporada de
crecimiento. Las esporas pueden ser salpicadas por la lluvia, lavadas con agua corriente o transportadas por los trabajadores en sus ropas, herramientas, equipos e implementos; También pueden ser diseminados a largas distancias por los vientos.
Las hojas de tomate infectadas con moho negro secadas al aire se almacenaron en bolsas de plástico a 4, 12, 20, 28 y 32° C y a temperatura ambiente para estudios de supervivencia de los conidios. Se colocaron submuestras en el campo en bolsas de malla de nylon, una suspendida sobre la superficie del suelo y otra enterrada a 5 cm por debajo de la superficie del suelo. Algo de germinación (<1%) ocurrió después de 18 meses con conidios de muestras mantenidas a 4, 12 y 20° C, y temperatura ambiente. No hubo germinación con conidios a 28° C almacenados después de 17 meses, a 32° C o más después de 13 meses, a partir de muestras de campo suspendidas después de 7 meses, o muestras enterradas después de 4 meses P. fuligena probablemente sobrevive en los desechos de plantas enfermas y produce esporas en estos desechos al comienzo de la temporada de crecimiento. Las esporas pueden ser salpicadas por la lluvia, lavadas con agua corriente o transportadas por los trabajadores en sus ropas, herramientas, equipos e implementos; También pueden ser diseminados a largas distancias por los vientos.
La enfermedad se presenta en las hojas de Solanum lycopersicum.

No se conocen restricciones de cuarentena para P. fuligena, debido a su diseminación en el aire
Cinco especies de tomate, Solanum cheesmaniae, S. habrochaites, S. peruvianum, S. pimpinellifolium y S. lycopersicum x S. pimpinellifolium, fueron reportadas como hospederas de P. fuligena por Hartman y Wang (1993). La solanácea negra (Solanum nigrum) es el principal hospedero de P. fuligena, también se ha reportado berenjena (S. melongena) como huésped de P. fuligena. Un total de 137 accesiones que representan 26 especies y cinco géneros de plantas solanáceas fueron inoculadas con P. fuligena en condiciones controladas y en campo. Veinte de las 26 especies desarrollaron síntomas después de la inoculación. Ocho especies de Solanum (S. chilense, S. chmielewskii, S. corneliomuelleri, S. neorickii, S. pennellii, S. aethiopicum, S. lasiocarpum (S. indicum) y S. macrocarpon, y cuatro
especies de Capsicum (C. annuum, C. baccatum, C. chinense y C. frutescens) fueron reportados como hospederos de P. fuligena. La enfermedad del moho de la mancha negra de la hoja, causada por P. fuligena, se informó en Withania somnifera (ashwagandha y ginseng indio) en 2014.
Los síntomas iniciales del moho negro de la hoja aparecen como diminutas decoloraciones de color amarillo pálido a lesiones de color verde claro, de 1 a 20 mm de diámetro. Las lesiones más antiguas pueden tener centros marrones rodeados por zonas de color verde pálido a amarillo. Las lesiones en la superficie inferior de la hoja se cubren inicialmente con micelio blanco que se vuelve gris a negro a medida que el hongo esporula. Las lesiones no tienen márgenes definidos ni en la superficie superior ni en la inferior de las hojas. En ambas superficies se produce una fructificación negra y hollín. Un gran número de estas áreas aparecen en cada hoja y pueden fusionarse para formar grandes parches. Las hojas infectadas pueden retorcerse, marchitarse, secarse con la edad, quebradizas y, por lo general, permanecer colgando de la planta con una apariencia cubierta de hollín; pero de vez en cuando también se puede observar defoliación. Las infecciones también ocurren en pecíolos, tallos y pedúnculos de la fruta, pero no hay informes de síntomas que ocurran en la fruta. La incubación nocturna de las lesiones en las hojas de tomate a 95-100% HR y 26-28° C dará como resultado una producción abundante de

conidios y conidióforos para su identificación bajo el microscopio.
El hongo puede aislarse mediante la esterilización superficial del tejido foliar infectado y la siembra en agar o mediante el aislamiento de una sola espora en agar. La esporulación de los cultivos se puede lograr en placas de agar tomate y avena. Los cultivos deben incubarse a 28° C con 12 h de luz al día. La identidad del patógeno puede confirmarse aún más a nivel molecular utilizando cebadores universales como ITS1/ITS4 a través de PCR.
El moho de la hoja negra del tomate causado por Pseudocercospora fuligena (Roldan) Deighton, es una enfermedad que se encuentra en regiones tropicales y subtropicales de Asia, África, Oceanía, partes de América del Norte y América del Sur. Causa daños económicos en países como Brasil, India, Japón, Filipinas, Taiwán y Tailandia. Se han reportado reducciones de rendimiento del 30% al 34% atribuibles a la enfermedad.
El microorganismo causante de esta enfermedad tiene micelio interno de color marrón oliva, hialino a subhialino. Los esporodoquios no estromáticos forman penachos de conidióforos geniculados a través de los estomas en ambas superficies (anfigénicos). Los conidióforos son de olivaceo pálido a marrón pálido, rectos a sinuosos, punta redonda o truncada, simples no ramificados, fasciculados, de 0 a 4 septados y de 14.0-74.5 x 3.0-5.5 μm.
Los conidios son subhialinas a oliváceas pálidas, cilíndricos a cilíndricos obclavados y rectos a ligeramente curvados. Las células de la punta son redondeadas con células basales, obcónicas largas a truncadas obcónicas largas, 2-14 septadas. Los conidios miden 9.0-137.0 x 3.5-6.5 μm.

El número de septos por conidio oscila entre 2 y 27. El hilio (cicatriz) no está engrosado. Algunos conidios son ramificados o bifurcados en la célula de la punta.
Los síntomas aparecieron inicialmente como manchas foliares, amarillas en la superficie de los folíolos adaxiales,
correspondientes a un crecimiento fuliginoso gris oscuro en la superficie abaxial, evolucionando a manchas necróticas irregulares. A medida que se desarrollaba la enfermedad, las manchas foliares se unían.

Factores que limitan el uso de co nv
POR OSCAR OJEDA BUENROSTRO
Las desventajas del uso de plaguicidas en la agricultura a nivel mundial son ampliamente reconocidas en términos de costo, peligros para la salud y el medio ambiente además de su escasa durabilidad. Por otra parte, no sólo muchos agricultores pueden enfrentar dificultades para acceder a plaguicidas seguros sino que las condiciones climáticas locales de sus tierras pueden ser tales que dificulten el uso seguro de dichas sustancias como por ejemplo, la simple necesidad de usar ropa protectora. Reconociendo esto, los fitopatólogos internacionales del sector público con una orientación hacia los pequeños agricultores se han centrado generalmente en el desarrollo de variedades
de cultivos resistentes a enfermedades y opciones de control biológico.
Muchos plaguicidas, incluidos los fungicidas e insecticidas que se utilizan para controlar los vectores de enfermedades, se están volviendo ineficaces a medida que las plagas evolucionan para superar las presiones de selección que les ejercen los insumos químicos. Además, los cambios regulatorios están reduciendo la disponibilidad de opciones de plaguicidas, ya que los peligros ecológicos y para la salud de los productos químicos son cada vez más preocupantes. La cuota de mercado de la producción ecológica está aumentando rápidamente: un número cada vez mayor de consumidores no quiere que sus alimentos sean
rociados con plaguicidas. La riqueza de la resistencia genética que se ha identificado para muchos cultivos a través de los sistemas de investigación internacionales es una ventaja para los fitopatólogos, genetistas y mejoradores de todo el mundo.
A pesar de los considerables avances en la comprensión de la biología y la etiología de los patógenos de las plantas, el manejo de las enfermedades de las plantas, que definimos como el despliegue integrado de variedades resistentes a las enfermedades, la evitación, el saneamiento y los fungicidas (incluidos los naturales, sintéticos y biológicos), no ha cambiado significativamente en los últimos 50 años, ya que depende principalmente de los fungicidas. Al igual que muchas otras

disciplinas, la fitopatología adolece de una incapacidad prolongada para aplicar los descubrimientos en bioquímica, fisiología y genética de plantas y hongos al problema práctico de proteger a las plantas contra los patógenos y aumentar el rendimiento de los cultivos. Con un 20 a 40% de la producción potencial de cultivos del mundo que se pierde anualmente debido a malezas, plagas de artrópodos y patógenos, las intervenciones más exitosas para mitigar estas pérdidas son los pesticidas, y los fungicidas continúan desempeñando un papel principal en el manejo de las enfermedades de las plantas.
Sin embargo, debido a las condiciones climáticas cambiantes, se prevé que las pérdidas de cultivos alcancen entre el 50 y el 80%, duplicando las pérdidas actuales y, por lo tanto, aumentando la dependencia de todos los pesticidas, no solo los fungicidas, para toda la producción de plantas. Dado que la mayoría de las tierras de cultivo del mundo no producen su potencial
ciones para la seguridad alimentaria mundial, lo que subraya la urgencia de reducir las pérdidas relacionadas con enfermedades con fungicidas efectivos. Es evidente que el uso de fungicidas tiene sus raíces en la necesidad de manejar los patógenos de las plantas, reduciendo así las pérdidas tanto por rendimiento como por deterioro poscosecha.
PROTOCOLOS REGULATORIOS NO FAVORECEN FUNGICIDAS DE AMPLIO ESPECTRO
Las tecnologías de péptidos pueden ofrecer soluciones innovadoras para la protección de las plantas y la salud de los cultivos
Esto enfatiza aún más que el papel de la industria de protección de cultivos en la mejora de la seguridad alimentaria se basa en la ganancia, no en el altruismo, y que este interés está sujeto a los márgenes de ganancia y pérdida. Aunque las grandes empresas multinacionales son las impulsoras del descubrimiento de fungicidas (junto con la evaluación, el desarrollo, la producción y la comercialización), los reguladores gubernamentales evalúan y aprueban todos los pesticidas para su uso. En el paso final del proceso, los fungicidas se comercializan a los productores, quienes finalmente deciden qué fungicida se compra, junto con dónde, cuándo, cómo y cuánto aplicar, según lo prescrito por la etiqueta del fungicida.

Los productos químicos introducidos en la década de 1970 proporcionaron a los productores fungicidas que podían usarse en dosis más bajas y también moverse sistémicamente dentro de la planta y proporcionar un control curativo de los patógenos que ya estaban allí. Convencer a los productores de que adoptaran estos productos mejorados requirió poco esfuerzo: su eficacia superior era obvia y brindó a los productores la oportunidad de realizar un tratamiento correctivo de las plantas infectadas. Además de los claros beneficios (ampliación de las opciones, mejor momento de aplicación, flexibilidad y control de enfermedades de espectro intrínsecamente más amplio en comparación con los fungicidas más antiguos), estos fungicidas proporcionaron a los productores un mejor retorno de la inversión. Como resultado, la dependencia de los productos químicos más antiguos disminuyó y los productos químicos más nuevos, en particular los inhibidores de la biosíntesis de esteroles se adoptaron rápidamente.
El desarrollo de fungicidas de amplio espectro con múltiples modos de acción para las necesidades gemelas de manejo de enfermedades y resistencia a fungicidas parece poco probable en esta coyuntura. Por diversas razones, los protocolos regulatorios modernos no favorecen los fungicidas de amplio espectro con modos de acción inespecíficos. A pesar de la necesidad de estos productos para el manejo de la resistencia a los fungicidas, es muy poco probable que se desarrolle y subsiguiente se apruebe un fungicida con un amplio rango de objetivos junto con un modo de acción no específico y toxicidad no objetivo para mejorar la seguridad y el perfil de uso. Sin un reemplazo potencial para el captan, el cobre, el clorotalonilo o el mancozeb para reducir la dependencia de estos productos, su uso continuará e incluso aumentará, particularmente a la luz del hecho de que brindan estos beneficios a un bajo costo.
IMPLICACIONES DEL CAMBIO CLIMÁTICO SOBRE LA AGRICULTURA MUNDIAL
A medida que la agricultura se ve afectada por el cambio ambiental global, la fitopatología juega un papel más importante para el manejo de las enfermedades de las plantas. En acuerdos de colaboración internacionales, el trabajo de esta especialidad mejora la estabilidad y la productividad de los sistemas de producción tanto en
FARAON
Tomate indeterminado con adaptabilidad en distintas zonas de producción (Bajío, Oaxaca, Puebla, Hidalgo, Estado de Méxicoy Altiplano).
Características de la planta
Produce racimos muy uniformes con 8 frutos en promedio y fácil amarre. Frutos de excelente calidad para mercado fresco y exportación. Planta uniforme de porte vigoroso que soporta la producción, tamaño y calidad a lo largo del ciclo.
Color: rojo intenso externo e interno.
Forma: oval , alta firmeza y sobremadurez.
Tolerancia: TSWV - TYLCV
Resistencias: TMV - V- Fol 2 – N

SANBA
Híbrido con buena adaptación a las zonas productoras de Bajío, Altiplano, Occidente, Sinaloa y Centro - Sur de México. Madurez relativa para corte: 90-95 días concentrando primero y segundo set para corte. Buena adaptación para producción en macro túnel. En campo abierto se recomienda, al menos una linea (rafia) de soporte. Fruto: Color verde oscuro. Grosor de pared 0.7-.8 mm promedio. Llenado completo de placenta. Peso optimo por fruto de 75 grs. Longitud de fruto mínimo 4.5 hasta 6 pulgadas manteniendo los tamaños. Pungencia. IntermediaAlta.
HR: BLS 1-3 Planta con vigor intermedio alto con buena estructura que permite el desarrollo adecuado de los frutos.
ORION

Lechuga tipo greenleaf de color verde intenso y uniforme con volumen alto definido por la cantidad de hojas que desarrolla. Crecimiento en V que facilita la manipulación para cosecha en bolsa, fresco y proceso. Vigor alto Madurez ciclo precoz a intermedio (35-50 días) dependiendo de la temporada.
Costilla fina. Tolerancia alta a floración (bolting) Tolerancia en campo a enfermedades ocasionadas por hongos


campo abierto como en la llamada agricultura protegida así como la calidad de sus productos. Dado que los patógenos de las plantas migran y evolucionan para presentar nuevos desafíos dentro de los sistemas existentes, el trabajo de los patólogos es inherentemente dinámico.
A pesar de la mejor producción de alimentos en las últimas décadas, miles de personas carecen de una nutrición adecuada. Haya o no margen para aumentar la producción de alimentos en muchas zonas, es de vital importancia asegurar la cosecha de los rendimientos reales y potenciales de los cultivos existentes. Por lo tanto, existe un creciente interés mundial en mejorar la seguridad alimentaria mediante la reducción de las pérdidas y el desperdicio. Las enfermedades de las plantas son una fuente importante de pérdidas de cultivos: se estima que entre el 10 y el 16% de los cultivos se pierden a causa de las enfermedades.
Entre 1960 y 2000 se lograron enormes aumentos en la producción de cereales gracias a la “Revolución Verde”. La producción mundial de
cereales casi se triplicó durante este período gracias a una combinación de fitomejoramiento, mayor uso de insumos como fertilizantes, plaguicidas y riego así como mecanización con el uso de tractores para la labranza y la plantación y cosechadoras. Algunos de los factores que influyen en la intensidad de las enfermedades y las pérdidas de los cultivos, como el tiempo y el cambio climático y la migración de patógenos, escapan en gran medida al control de los agricultores y de los servicios de investigación y extensión que se esfuerzan por apoyarlos. Otros factores se manejan a nivel de finca y paisaje, como los insumos agrícolas que reducen o aumentan la presión de patógenos, otras prácticas agronómicas que influyen en el microclima y la dispersión de propágulos, y la diversidad de especies y variedades en el tiempo y el espacio.
Se espera que las pérdidas de plagas de insectos aumenten con el cambio climático global. Se espera que las condiciones más probables que impulsen el aumento de las pérdidas ocurran principalmente en las regiones templadas, que actualmente albergan
No obstante los avances tanto en biología de hongos así como etiología de las enfermedades de las plantas, el manejo de éstas no ha cambiado mucho en los últimos 50 años
los niveles más altos de producción de cultivos. No está tan claro cómo afectará el cambio climático a las enfermedades de los cultivos. El cambio climático interactúa con las enfermedades virales de maneras complejas, a través de efectos sobre el crecimiento de las plantas, la abundancia y biología de los vectores y la biología de los virus. Es probable que los cambios generales en la temperatura y las precipitaciones cambien los patrones de incidencia y gravedad de las enfermedades en cada lugar, y es probable que el aumento de la variabilidad del tiempo y los eventos relacionados con el clima aumente la gama de problemas de enfermedades con los que los agricultores (y los fitopatólogos y fitomejoradores) se ven obligados a lidiar.

POR BERENICE TERRAZAS ACEVEDO
El proceso de producción de alimentos requiere insumos sustanciales de tierra, agua, fertilizantes, plaguicidas y energía. La conversión de tierras naturales en tierras agrícolas, especialmente a través de la deforestación, conduce a la pérdida de biodiversidad, escorrentía de nutrientes, erosión del suelo y emisiones de gases de efecto invernadero.
En los Estados Unidos, alrededor del 80% del uso de agua dulce, incluidas las aguas superficiales y subterrá-
neas, se destina a la agricultura de regadío, y en algunas partes del país ha contribuido a la escasez de agua y a los caudales ambientales inadecuados. El uso de fertilizantes puede provocar pérdidas de nutrientes en las vías fluviales, las aguas subterráneas y la atmósfera. Una vez en el medio ambiente, el exceso de nitrógeno puede provocar la acidificación del agua, la eutrofización, el cambio climático y la pérdida de biodiversidad, así como impactos en la salud humana. La agricultura también requiere energía; La quema de combustibles fósiles para la producción de fertilizantes, la agricultura mecanizada y el transpor-
La producción de hortalizas de invernadero periurbanas ha desempeñado un papel importante en el suministro local
te emite gases de efecto invernadero y contaminantes como el dióxido de carbono y los óxidos de nitrógeno. Además, la fermentación entérica del ganado es un importante contribuyente de metano. Se espera que el sector ganadero de la producción de alimentos por sí solo supere las contribuciones sostenibles de la humanidad al cambio climático global y al nitrógeno para 2050, lo que requiere una evaluación continua de las tendencias actuales y proyectadas de consumo de alimentos.
La huella de nitrógeno de un producto alimenticio se refiere a la cantidad



total de nitrógeno reactivo, es decir todas las especies de nitrógeno excepto N2, liberado al medio ambiente por la producción y el consumo de un producto alimenticio. La creación de nitrógeno reactivo como fertilizante sintético ha apoyado a una población creciente. Sin embargo, la alteración humana relacionada con el ciclo del nitrógeno ha llevado a la acumulación de un exceso de nitrógeno reactivo en el medio ambiente. Una vez liberado al medio ambiente, el nitrógeno reactivo puede contribuir a una serie de impactos humanos y ambientales negativos, como el smog, la lluvia ácida, la pérdida de biodiversidad, el cambio climático y la eutrofización.
El nitrógeno desempeña un papel insustituible en los sistemas agrícolas. La cantidad de fertilizante nitrogenado aplicado ha excedido el límite
seguro del ecosistema agrícola, y el excedente de Nr, siendo Nr todas las especies denitrógeno excepto N2, se ha perdido en el medio ambiente y ha causado consecuencias negativas para el suelo, el agua y el aire. El Nr en el ecosistema tiene impactos adversos tanto en el medio ambiente como en la salud humana. Por ejemplo, el N2O aumenta los gases de efecto invernadero en la atmósfera y destruye el ozono, y el NO3 causa problemas de contaminación de las aguas subterráneas. Los nitratos que ingresan a los ríos con las aguas subterráneas pueden causar eutrofización (floraciones de algas y muerte de peces) y degradación de la calidad del agua. El NH3, como gas alcalino, puede reaccionar con gases ácidos como el NO2 en la atmósfera para formar aerosoles de sal de amonio, que es la principal forma de formar neblina en el aire y tiene un
impacto negativo en la calidad del aire e incluso representa una grave amenaza para la salud humana.
Además, la aplicación excesiva de fertilizantes ha causado enormes pérdidas económicas. La pérdida económica directa causada por la fertilización excesiva de las tierras de cultivo en todo el mundo es muy alta y el costo supera los miles de millones de dólares por año. Aunque el uso de fertilizantes nitrogenados ha mejorado el rendimiento y la seguridad alimentaria en diferentes países, las pérdidas económicas del uso excesivo de fertilizantes nitrogenados están aumentando progresivamente con el crecimiento de los precios de la energía y los fertilizantes.
ESENCIAL LOGRAR LA OPTIMIZACIÓN DEL USO DE FERTILIZANTES NITROGENADOS
El uso excesivo de fertilizantes nitrogenados es más frecuente en la producción de hortalizas de invernadero en comparación con los sistemas agrícolas tradicionales, principalmente porque las hortalizas de invernadero pueden funcionar independientemente de las condiciones naturales y las limitaciones que enfrentan los sistemas agrícolas tradicionales. La tasa promedio de aplicación denitrógenode las hortalizas de invernadero supera los 2,000 kg denitrógenoha-1 año-1, que es aproximadamente 4 veces mayor que la del trigo y el maíz y de 3 a 6 veces mayor que la de las hortalizas europeas y australianas.
Se han implementado muchas investigaciones para mitigar las pérdidas de Nr y los riesgos ambientales relacionados con los sistemas de producción agrícola, por ejemplo, las estrategias de administración de nutrientes de las 4R (momento correcto, lugar correcto, tasa correcta, fuente correcta) para optimizar el aporte de fertilizantes denitrógenode acuerdo con los requisitos del cultivo y la heterogeneidad
Es crucial minimizar las pérdidas de Nr y las emisiones de GEI de los sistemas intensivos de cultivo de hortalizas a través de medidas de gestión agrícola optimizadas
de los nutrientes del suelo; y la mejora de la tecnología de fertilizantes, en particular los fertilizantes de mayor eficiencia, como los fertilizantes de liberación controlada y los inhibidores, que pueden mejorar la eficiencia agronómica y económica de los fertilizantes. Mientras tanto, también existen barreras, desafíos y costos de implementación para que los agricultores adopten las nuevas prácticas y tecnologías de manejo. Por lo tanto, es necesario tener en cuenta la accesibilidad y la eficacia en función de los costos de las prácticas de gestión y las estrategias de mitigación propuestas.
En las últimas décadas, se han implementado cada vez más estudios sobre la huella denitrógeno(NF) y el análisis costo-beneficio para evaluar la sostenibilidad ambiental y económica del uso delnitrógenopara la producción de cultivos. La huella denitrógeno(NF) se ha propuesto como un indicador


para evaluar cómo los individuos, las comunidades, las organizaciones o los países contribuyen a la contaminación por Nr a través de su consumo. Proporciona una medida completa del impacto humano en la contaminación relacionada con el N.
Limitar el impacto negativo de las prácticas agrícolas en el medio ambiente y, al mismo tiempo, preservar el rendimiento sostenible de los cultivos es uno de los principales retos a los que se enfrenta la agricultura en los próximos años. Como elemento esencial para la nutrición de las plantas, el nitrógeno representa un ejemplo paradigmático de este desafío. Además, debido al efecto combinado de una elevada solubilidad y poca retención en los suelos, la falta de este elemento es y será uno de los principales factores limitantes del rendimiento en todo el mundo. Al mismo tiempo, la aplicación de fertilizantes nitrogenados sintéticos es, en muchos sistemas agrícolas, un enfoque de baja eficiencia que se ha relacionado con la degradación de los recursos naturales. Una de las estrategias adoptadas para limitar la huella económica y ambiental de la producción de cultivos y mantener al mismo tiempo un rendimiento sostenible es el “reciclaje” de productos biodegradables y ricos en minerales de los sectores ganadero y agrícola.


El nitrógeno aplicado al suelo, excepto para la absorción por los cultivos, se pierde principalmente a través de la volatilización, fijación, nitrificación-desnitrificación, escorrentía o lixiviación del amoníaco (NH3). Esto ha dado lugar a una serie de problemas ambientales como la contaminación de las aguas subterráneas, la acumulación de nitratos y la acidificación del suelo. Además, el óxido nitroso (N2O), como gas de efecto invernadero (GEI) con un potencial de calentamiento a 100 años 273 veces mayor que el dióxido de carbono (CO2), contribuye significativamente al calentamiento global.


POR PABLO JARAMILLO VILLEGAS
En el cultivo comercial de pepino en invernadero, el objetivo principal es lograr una distribución de luz solar uniforme en todo la estructura para conseguir una producción óptima. En otras palabras, es importante lograr un equilibrio entre el crecimiento vegetativo y la carga de frutos durante todo el ciclo de crecimiento de la planta.
De acuerdo con su experiencia y preferencia, los productores suelen adoptar varios métodos de “enrejado” del cultivo., con un sistema de cuerda o alambre. Asimismo, es necesaria la poda continua de brotes, follaje, frutos y flores. Si hay demasiados frutos, una gran proporción puede estar abortada, malformada o mal coloreada, porque la planta puede no tener suficientes asimilados. La situación se deteriora aún más en condiciones de poca luz. Generalmente, para los cultivares de frutos largos, solo se debe permitir el desarrollo de un fruto por eje de hoja, aunque con cultivares vigorosos a veces puede madurar más de un fruto en un nudo. Los tipos midi cortos pueden soportar varios frutos por nudo y dar buenos rendimientos, con un mínimo de tres a cuatro frutos cosechados en cada nudo.
Las prácticas culturales tienen como objetivo proporcionar un medio de raíz apropiado, un crecimiento equilibrado
de raíces/hojas, un desarrollo equilibrado de fuentes/sumideros y una buena tasa de cosecha de frutos de alta calidad. Las prácticas de cultivo más importantes son el control del microclima, la fertirrigación y la formación. El pepino es muy sensible tanto al estrés abiótico como al biótico, y pueden surgir serios problemas en caso de un manejo inadecuado del cultivo.
Los pepinos se cultivan en campo o invernaderos. Las plantas de pepino cultivadas en el campo generalmente se inician como semillas y se plantan mecánicamente o a mano. Muchas operaciones comerciales entrenan a sus plantas para que crezcan en postes o enrejados para mantener la fruta suspendida. Se utilizan varios sistemas de entrenamiento para el cultivo en espaldera, pero el sistema de paraguas es el más común. En el sistema de paraguas, todas las ramas laterales se eliminan a medida que aparecen hasta que el tallo principal alcanza una altura predeterminada. Luego, se permite que la planta crezca más libremente para que la planta pueda concentrarse en el crecimiento de la fruta, en lugar de en la altura. Algunos productores plantan variedades tipo arbusto y permiten que la fruta se extienda por el suelo.
El control de malezas y plagas también son prácticas de manejo importantes para garantizar una producción óptima. El control de malezas en la producción de pepinos se logra a través de una variedad de métodos que incluyen el uso de cultivos de cobertura y mantillos, el cultivo y el deshierbe manual, y la aplicación de herbicidas dirigidos a los tipos específicos de malezas comunes en un campo en particular. Las plantas de pepino son susceptibles a una variedad de infecciones por insectos, bacterias, hongos y nematodos. La identificación temprana de tales infecciones o infestaciones es clave para métodos de control apropiados

y rápidos. Las estrategias de prevención de enfermedades incluyen la rotación de cultivos, la selección cuidadosa del campo, el saneamiento, el tratamiento del suelo y la selección adecuada de semillas. Es una práctica común utilizar mantillos de suelo para controlar las malezas y protegerse de los insectos, así
como para modificar la temperatura del suelo, conservar el agua y controlar la erosión. Los mantillos pueden incluir turba u otro material orgánico. Muchas operaciones comerciales utilizan cubiertas de plástico como mantillo para proteger los cultivos. Dependiendo del efecto deseado, los productores
pueden emplear colores como claro, negro, blanco o aluminio. Todos estos colores tienen un efecto de calentamiento en el suelo por la noche, pero pueden calentar, enfriar o no tener ningún efecto durante el día.
La temperatura del aire influye en el crecimiento vegetativo, el inicio de la floración, el crecimiento del fruto y la calidad del fruto. La tasa de crecimiento del pepino depende de la temperatura promedio de 24 horas: cuanto mayor sea la temperatura promedio (≤ 25 ° C), más rápido será el crecimiento. La temperatura óptima del aire depende del período de crecimiento. En el momento de la germinación, la temperatura óptima es de 25-35° C y, con buena humedad, las semillas tardan 2-3 días en germinar. Por el contrario, a 12 °C, las semillas necesitan entre 12 y 20 días para germinar y hay muchas pérdidas. El crecimiento de los brotes no se produce a temperaturas del aire de < de 13 a 15 ° C, mientras que la temperatura máxima para el crecimiento vegetativo es de aproximadamente 38 a 40 ° C.
Durante la primera semana después de la siembra, la temperatura ideal del aire es de 22 a 24 °C; a partir de entonces, las temperaturas deben mantenerse entre 20 y 22 °C hasta el
comienzo de la cosecha. Durante el clima cálido, a finales de primavera y principios de otoño y en la cosecha, se recomienda reducir la temperatura del aire en ≤ 2 °C para favorecer el crecimiento vegetativo, especialmente por la noche. A temperaturas del aire más altas, las frutas crecen rápidamente y compiten por la asimilación. Puede comenzar una pausa de cosecha más larga, especialmente después de cosechar pepinos del tallo principal. Esto puede anular la ventaja de un inicio más temprano de la cosecha; Además, las plantas envejecen antes. Para una buena calidad de la fruta, las temperaturas deben ser de 22 a 24 ° C; a temperaturas de < 18° C, los frutos tienden a ser más cortos. Se recomienda una diferencia de temperatura entre el día y la noche solo para el cultivo en invierno y principios de primavera. El rendimiento del crecimiento depende de la temperatura media de 24 horas durante los días largos y las noches cortas de primavera/verano. La disminución de las temperaturas nocturnas en este período no es una ventaja fisiológica para la planta; Sin embargo, podría hacerse para ahorrar energía.
En pepino, la frecuencia de riego es generalmente alta debido a que sus requerimientos de agua son relativamente altos durante su cultivo
El control de la temperatura debe considerarse en el contexto de la intensidad de la luz. La radiación afecta el área total de hojas de la planta, la producción de carbohidratos y, en consecuencia, la productividad. Durante el invierno, el suministro de carbohidratos es bajo y la productividad se reduce, lo que resulta en muchos frutos abortados. La luz también influye directamente en la calidad de la fruta. Por ejemplo, las frutas cultivadas en condiciones de poca luz tienen menos materia seca, son generalmente de color verde claro en la cosecha y se vuelven amarillas fácilmente en el estante. Los frutos jóvenes suelen ser más sensibles a la baja intensidad de luz que los frutos más viejos de la misma planta.
Los pepinos son muy sensibles a la alta salinidad. Las plantas cultivadas en condiciones salinas están sujetas a serios problemas que resultan en un rendimiento insatisfactorio. La alta salinidad provoca un retraso en el crecimiento de las plantas, entrenudos cortos y una reducción del área foliar. Las hojas suelen ser de color verde oscuro y opacas. En condiciones extremas, los tejidos necróticos pueden estar presentes en las hojas más viejas. La salinidad puede deberse a un alto nivel de sales en el agua subterránea, el agua de riego, el suelo o el medio de crecimiento, o a la aplicación excesiva de fertilizantes.



CARACTERÍSTICAS DE LAS VARIEDADES
MÁS AMPLIAMENTE CONSUMIDAS
Los pepinos a menudo se comen como verdura, pero científicamente se consideran una fruta ya que contienen semillas encerradas y se desarrollan a partir de una flor. Los pepinos, al igual que los melones, la calabaza, las calabazas y las sandías, son miembros de la familia de las cucurbitáceas. Son cultivos de vid y se pueden cultivar en el suelo o en postes o espalderas para suspender el fruto. Los pepinos vienen en tres tipos distintos: sin semillas, sin semillas y mini. Hay cerca de 100 variedades, pero las más comunes son la inglesa, la de jardín, la persa, la mini y la de limón. El pepino inglés es el más largo, es estrecho y, a menudo, se comercializa en una envoltura de plástico. La piel de los pepinos ingleses es delgada y, a menudo, no requiere pelado. Por el contrario, el pepino de jardín tiene una piel cerosa oscura. Normalmente, los consumido-
res eliminan la piel debido a su sabor amargo. Los pepinos persas se llaman sin eructos porque tienden a ser más pequeños, más dulces y sin semillas.
La piel es más lisa, más fina y, al igual que la variedad inglesa, no requiere descamación. Estos pepinos tienden a ser más suaves y fáciles para el sistema digestivo. Los pepinos Kirby son los más pequeños. Estos mini pepinos se están volviendo populares en el mercado debido a las preferencias de los consumidores. Tienen una amplia variedad de colores de piel que van desde el amarillo hasta el verde oscuro. Los pepinos limón son redondos y amarillos, parecidos a los limones, pero son dulces, tienen piel fina y contienen semillas. Las variedades tradicionales de pepino tienen flores masculinas y femeninas y requieren polinización para producir frutos sanos con semillas y espinas blancas. Cuando los pepinos no polinizan correctamente, los frutos están deformes y poco desarrollados,
Con el fin de prevenir la fertilización excesiva o insuficiente, es vital realizar análisis frecuentes del contenido de nutrientes del suelo y el agua utilizados
especialmente en el extremo de la flor. Los tipos de pepinos más populares que se cultivan actualmente en invernaderos son cultivares híbridos largos y sin semillas, a menudo denominados pepinos “europeos” u “holandeses”. Estas variedades son ginosas y producen solo flores femeninas. Los frutos son partenocárpicos y no hay necesidad de polinización. El fruto tiene una piel delgada, comestible, lisa y verde, a veces con costillas longitudinales tenues. Entre ellos se encuentran los populares cultivares partenocárpicos ‘Beit Alpha’, que están adaptados para el enrejado, tienen entrenudos más cortos y producen múltiples frutos en un racimo
Hay tres usos distintos para los pepinos en el mercado: frescos enteros, frescos en rodajas y en escabeche. Ya sea de la variedad inglesa, de jardín, persa, mini o limón, los pepinos enteros frescos se cultivan para la venta minorista al consumidor. Los pepinos frescos en rodajas suelen ser de jardín y se cultivan para el sector de la restauración, que requiere rodajas de tamaño uniforme para las ensaladas empaquetadas y las barras de ensaladas de las cadenas de restaurantes. Los pepinos encurtidos tienden a ser más pequeños y gruesos. La variedad más conocida es el pepinillo de piel rugosa.


popularidad del aguacate en los mercados ha crecido casi de manera abrumadora en la última década

Un fruto tropical que es sumamente apreciado a nivel mundial
Los factores ambientales que pueden afectar en mayor grado el rendimiento de la producción de aguacate son el estrés hídrico, la fotosfera y la temperatura, además del nivel de sal en el suelo. Las investigaciones han demostrado que de 20 a 60 semanas, las condiciones ambientales y las prácticas culturales pueden determinar el éxito productivo de la explotación.
No se permite que el aguacate madure en el árbol, es decie que la fruta se cosecha en la madurez hortícola. Después de cosechar los frutos de aguacate, se lavan, se enceran, se clasifican y se empaquetan. Por lo tanto, en la producción de un aguacate de la mejor calidad, el cuidado
adecuado y un sistema de gestión del huerto es vital para el retorno económico. Las operaciones de poscosecha deben centrarse en la gama ampliada de almacenamiento de la fruta, ralentizando el proceso de senescencia y controlando la maduración de la fruta cuando se cosecha en una etapa madura o en la etapa inmadura. Toda la cosecha, la manipulación y el transporte a las plantas de envasado y todas las demás operaciones deben realizarse con cuidado, es decir, sin posibilidad de pérdidas mecánicas. Los daños o lesiones pueden acelerar la maduración de la fruta y pueden afectar negativamente la apariencia de la cáscara de la fruta, causar pardeamiento e imperfecciones después y antes del almacenamiento. Comúnmente, los aguacates son muy sensibles a estropearse durante el ablandamiento, luego deben transportarse con cuidado para exhibirlos en los mercados. Durante el ablandamiento, la calidad también puede reducirse, por lo que es esencial controlar la maduración del aguacate con regularidad y consumir primero la fruta madura. Si es posible, toda la fruta madura o casi madura cosechada debe colocarse a una temperatura

más baja de 1 a 6°C para evitar la descomposición.
A medida que el número de divisiones celulares se divide rápidamente, el tamaño de los frutos también aumenta, y la división celular depende del metabolismo de los carbohidratos, la biosíntesis de isoprenoides y el apoyo de las actividades hormonales de las plantas. También se observan algunos nutrientes importantes para una producción de fruta de mejor calidad, como el calcio, el magnesio, el boro y el zinc. El nitrógeno es también el elemento manipulador esencial que se utiliza para controlar la fuerza del árbol y debe gestionarse adecuadamente, ya que una fuerza inmoderada del árbol es contraproducente para obtener una fruta de buena calidad.
El estudio reciente sobre la composición fitoquímica y nutricional de
las frutas de aguacate muestra que aproximadamente la mitad de un aguacate consiste en fitoquímicos pesados y nutrientes de los alimentos que contienen los siguientes nutrientes esenciales; 4.6 gramos dietéticos, 0.2 gramos azúcar total; 345 miligramos potasio, 5.5 miligramos sodio, 19.5 miligramos magnesio, 5.0 μg RAE vitamina A, 6.0 miligramos vitamina C, 1.3 miligramos vitamina E, 14 μg vitamina K1, 60 miligramos ácido fólico, 0.2 miligramos vitamina B-6, 1.3 miligramos niacina, 1.0 miligramos ácido pantoténico, 1.0 miligramos riboflavina, 10 miligramos colina, 185 μg luteína/ zeaxantina, 18.5 μg criptoxantina, 57 miligramos fitoesteroles, 6.7 gramos ácidos grasos monoinsaturados altos y alrededor de 114 kilocalorías de fibra dietética insoluble, esto puede respaldar una amplia gama de efectos sobre la salud potencial. Las frutas de aguacate son la forma pura de fruta
pesada y densa en energía porque contiene aproximadamente el 80 por ciento del agua de la parte comestible total de la fruta y aproximadamente el 6.8 por ciento de fibra dietética. Las investigaciones muestran que la fruta del aguacate tiene los mismos efectos en el control de peso que las frutas y verduras menos grasas.
FORMAS Y TAMAÑOS Y PARTICULARIDADES DE SU CULTIVO
Persea americana es una especie tropical cuyos frutos --originarias de México y América Central-- son altamente apreciados hoy en día en todo el mundo. Pertenece a la familia de las lauráceas. El origen, el clima y la nutrición de las plantas son los factores que determinan el crecimiento y la calidad del aguacate. El aguacate pertenece a la familia Lauraceae. Otros miembros incluyen alcanfor, laurel de California,

canela y sasafrás. Persea es el género, y americana es la especie. El género y la especie de aguacate relatan su origen en el Nuevo Mundo, el primer científico que explicó el aguacate en 1974 (Miller).
En general, las especies de aguacate se han dividido en tres razas: mexicana, guatemalteca y antillana. Algunas clasificaciones dividieron a la raza mexicana en su especie, P. drymifolia. no eran especies separadas, pero la raza mexicana debería clasificarse como una subespecie. el guatemalteco en su especie y P. nubigena, sacando a las otras dos razas en P. Americana. El árbol de aguacate es de hoja perenne durante todo el año. Su altura alcanza unos 12-30 metros del suelo y tiene muchas ramas largas. La forma de las hojas es redonda, ovalada y ovada, y su longitud es de 9 a 30 centímetros. El tamaño de la flor del aguacate es pequeño y verdoso. Las flores tienen partes masculinas

Siendo el aguacate rico en vitaminas, grasas y minerales, ayuda a mantener el cuerpo funcionando normalmente, evitando incluso enfermedades tóxicas
y femeninas. La forma del fruto del aguacate puede ser redonda, ovada o en forma de pera y la piel del fruto es diferente en color y apariencia en todas las verdades. La piel puede tener más flexibilidad, de lisa a áspera y de aspecto amarillo verdoso, rojo violáceo o negro. El color de la pulpa de la fruta del aguacate es de amarillo verdoso a amarillento brillante, y tiene una textura aceitosa cuando se madura, pero la superficie interna será fibrosa.
La fruta del aguacate contiene una semilla grande, es decir, de forma ovalada u ovalada, y hace que el peso de la fruta sea de aproximadamente el 10 al 25 por ciento del peso total. La cantidad de aceite y el contenido de humedad son diferentes en todas las variedades de aguacate y aproximadamente un máximo del 30 por ciento de aceite está presente en una fruta. El peso promedio oscila entre 450 g y 1.3 kg. La verdad Quintal es el cruce de dos razas, es decir, antillana y guatemalteca. Los tamaños de estos híbridos son grandes, es decir, de 500 a 800 g. Estos frutos tienen forma piriforme, piel verde lisa, pulpa amarilla y tienen una semilla grande, comúnmente suelta dentro de la fruta. Esta es la variedad más consumida en el mundo, con una proporción de lípidos de aproximadamente el 63% de ácidos grasos omega-9. La variedad de aguacate más transportada a nivel mundial es la Hass y con un promedio de 180-300 g de tamaños.
Las frutas de aguacate tienen diferentes formas. Incluían elipsoide, clavado, esferoide, obviado y estrechamente obviado. Además, algunas variedades diferentes incluyen alto obovado y esferoide. Alrededor del 54,4 % de los frutos tienen crestas parciales y completamente envueltos con crestas son aproximadamente el 17 por ciento. Alrededor del 28.3 por ciento de las frutas no tienen crestas. Alrededor del 50.9 por ciento tiene un tallo central y el 49.1 por ciento tiene una posición simétrica del tallo en los frutos. Rango de longitud del tallo: 0,68-1,5 cm. Alrededor del 96,2 por ciento de los frutos tienen una longitud de tallo superior a 1,5 cm. Alrededor del 35.8 por ciento de las frutas tienen un brillo fuerte en la piel, alrededor del 35.8 por ciento tienen un brillo medio y alrededor del 28.3 por ciento tienen un brillo débil.
La longitud promedio de la fruta del aguacate oscila entre 11.10.10 ± 1.52 con una longitud promedio entre 7 19 cm. longitudes entre 10-13 cm son aproximadamente 49.1 por ciento, 34 por ciento tienen hasta 10 cm y más de 13 cm es 17 por ciento. Alrededor del 81.1 por ciento de los frutos de aguacate tienen el mismo diámetro, entre 7 y 9 cm. Alrededor del 1.9 por ciento tienen una longitud que oscila entre 9 y 11 cm. Una proporción relativamente pequeña de alrededor del 17 por ciento tiene una longitud de entre 5 y 7 cm. La fruta del aguacate tiene un peso diferente. Alrededor del 58.6 por ciento pesa entre 220 y 370 gramos. Alrededor del 18.9 por ciento tiene un peso superior a los 420 gramos, sin embargo, el 11.3 por ciento pesa entre 170 y 220 gramos y el resto tiene un peso entre 370 y 420 gramos. Se estudió una amplia gama de longitudes de pedúnculo de los frutos. La mayoría del 83 por ciento de los frutos tenían longitudes de pedúnculo entre 2 y 6 cm. Alrededor del 47 por ciento de los frutos tenían longitudes de pedúnculo de 4 a 6 cm y el 7.6 por ciento tenían longitudes de pedúnculo de más de 8 cm de largo.


PLa nutrición vegetal pretende establecer el papel que juega cada uno de los elementos esenciales en el desarrollo de la planta
arte importante y costosa de la producción de cosechas, es la semilla y con ello, naturalmente su selección debe ser cuidadosa para garantizar la obtención del producto con la calidad requerida en el mercado. Por tal motivo, son de gran interés científico-técnico los trabajos encaminados a estimular y mantener los niveles de germinación al conservar las semillas, para poder elevar la productividad de los cultivos de forma sostenible y enfrentar los cambios en el entorno de manera más apropiada. En la producción comercial de semillas, la calidad está determinada por un conjunto de atributos, donde la calidad genética, física, sanitaria y fisiológica juega un papel importante.
La calidad fisiológica implica la integridad de las estructuras y procesos
fisiológicos, siendo los principales indicadores: la viabilidad, germinación y vigor, que dependen del genotipo. Entre los factores que pueden tener efecto en la calidad de la semilla están el grado de madurez y tiempo de maduración de la semilla después de la cosecha, el desarrollo y maduración de la semilla son aspectos importantes que deben ser considerados en la producción de semillas.
Los factores ambientales prevalecientes en la etapa de floración y formación de fruto determinan en gran medida la calidad de la semilla. Se debe cosechar cuando la semilla logre alcanzar su máxima calidad fisiológica. Durante la producción es primordial que exista una adecuada disponibilidad de agua y de nutrientes para que la formación de semilla sea la adecuada. Es interesante conocer otros parámetros que permitan detectar la maduración

fisiológica, correlacionándola con características morfológicas de la planta, de los frutos y/o semillas. El desarrollo de la semilla esta normalmente acompañada por el desarrollo del fruto y por una serie de importantes estados ontogénicos desde la fertilización en la acumulación de nutrimentos, hasta la madurez de ésta. La escasa germinación de semillas inmaduras ha sido atribuida a bajos niveles de nutrientes entre otras causas.
Para hortalizas de frutos carnosos, como pimiento o tomate, la maduración de las semillas generalmente coincide con el inicio de cambio de coloración de los frutos, es decir, frutos verdes con manchas rojizas. Es importante destacar que no siempre es necesario esperar la maduración completa de los frutos para retirar las semillas. Muchas veces, semillas provenientes de frutos en etapa de
maduración ya alcanzaron la maduración fisiológica. El fruto de la especie Physalis ixocarpa (Brot, ex Hornem.) es conocido como tomatilllo, tomate de cáscara, miltomate, tomate de fresadilla, tomate de hoja, tomate milpero, es una solanácea originaria de México. A México se le reconoce como centro de origen, diversidad y domesticación del género Physalis ya que está representado por 50 especies.
El tomate de cáscara o tomatillo es una planta anual con tallo erecto y ramificado, de 0.9 a 1.2 m de altura. Su tallo es glabro o casi glabro, herbáceo o ligeramente leñoso en la base. Las hojas son delgadas ovaladas o lanceoladas, entre 5 y 7.5 cm de largo, dentadas y con peciolos largos de textura suave. Las flores son grandes y abiertas, de 1.8 cm de diámetro, con bordes amarillos brillantes, monopétalas con corola, presentan cinco
manchas de color pardo negruzco, las anteras son purpuras; por lo general las flores están sobre pedicelos axilares o extraxilares, es cáliz es pentadentado, tiene cinco estambres; el estilo es delgado; el estigma casi bilobulado el fruto es una baya amarilla o verdosa algo viscosa, mide de 1 a 5 cm de diámetro, globoso, liso, pegajoso, algo acido, cubierto por cáliz avejigado.
A pesar de la amplia variabilidad genética, tanto en el tomate silvestre como en el domesticado, el rendimiento medio nacional se considera bajo, de acuerdo al rendimiento potencial de la especie 40 t ha-1 esto debido a una actividad agronómica limitada por el uso de variedades nativas de bajo rendimiento y sistemas de producción ineficientes, escasez de agua de insumos agrícolas y de semilla de baja calidad. Se han realizado numerosos esfuerzos para mejorar el

rendimiento y la calidad funcional y nutrimental de este fruto como cruzas dialélicas, selección masal visual estratificada, cruzas planta x planta y más recientemente inducción de poliploídia entre otros.
VIGOR Y POTENCIAL GERMINATIVO DE LA SEMILLA
Tras la madures fisiológica inicia un proceso continuo e irreversible de deterioro hasta perder su capacidad germinativa. También se ha establecido que madurez fisiológica de la semilla se obtiene cuando el fruto toma una coloración amarillenta y la semilla tiene un color parduzco y café. El proceso consiste en extraer la semilla del fruto, lavarla con agua, eliminar las que flotan y se dejan a secar en la sombra, aquellas que quedaron al
La dinámica agrícola del cultivo demanda la generación de cultivares mejorados que cubran las necesidades del mercado nacional e internacional
fondo se vuelven a seleccionar para obtener semillas de alto porcentaje de germinación.
La calidad de la semilla puede ser afectada por diversos factores, entre los cuales cabe mencionar las condiciones climáticas como nutricionales, durante el desarrollo y formación de estas, los métodos de cosecha, contenido de humedad y condiciones de almacenamiento. La nutrición durante el crecimiento del fruto constituye uno de los factores más importantes que afecta su calidad y comportamiento poscosecha. Los cultivos hortofrutícolas, requieren grandes cantidades de potasio acumulado en los frutos para incrementar su calidad. Aun cuando la calidad del fruto está definida por varios atributos que dependen del manejo cultural y del genotipo, la nu-

trición potásica constituye un aspecto de manejo agronómico que permite uniformizar la maduración y lograr un mejor fruto.
El potasio es considerado un fabricador de calidad, aumenta la floración, influye en la precocidad, coloración y calidad de fruto. El potasio es requerido intensamente durante los estados fisiológicos de producción, así como en el cuajado y llenado del fruto. Este elemento cumple un papel vital en el llenado de frutas, granos y semillas. Asimismo, el potasio incrementa el rendimiento y calidad, por lo cual en esta investigación se plantearon los siguientes objetivos e hipótesis.
El acondicionamiento de semilla es obtener, de la cantidad total de semilla disponible, solamente aquel volumen que cumpla con las normas de calidad vigentes. En este sentido después de realizar la molienda de los frutos en la despulpadora, las semillas útiles deben ser secadas a la sombra para que alcancen 11% de humedad. A lo que sigue el empleo de una cribadora con una malla de perforaciones redondas de 1.5 mm, con lo cual se obtiene semilla de tamaño medio que tiene mejor calidad que aquella que logra pasar la misma criba y que finalmente se clasifica como pequeña; cuando el primer grupo de semillas es sometido a una clasificación por peso en una mesa de gravedad, en la que se eliminan las semillas medianas de peso ligero, también aumenta la calidad de semilla del lote, ya que se mejora el vigor.
El uso de maquinaria para el acondicionamiento de semilla permite hacer más eficiente este proceso y mejora la calidad de los lotes; además de la limpieza, la clasificación de la semilla incrementa la calidad fisiológica.

es causa de la presencia de clorosis variegada
Ala fecha se reconoce que la enfermedad denominada clorosis variegada de los cítricos suele ser consecuencia de las interacciones entre el patógeno Xylella fastidiosa, la planta misma y el medio ambiente. Sin embargo, qué proporción de contacto entre X. fastidiosa y las plantas hospederas en el medio natural resulta en enfermedad es aún un factor que demanda mayor investihación.
Dicho hecho deja abierta la posibilidad de que la enfermedad pueda representar una proporción relativamente

pequeña de estas interacciones, lo que lleva a la sugerencia de que X. fastidiosa puede considerarse principalmente un endófito en lugar de un patógeno. Puede ser notable que para X. fastidiosa, la señalización célula-célula regula la virulencia limitada a las plantas, al tiempo que promueve la transmisión vectorial de planta a planta. Las tasas de transmisión también pueden ser moduladas por la respuesta del vector a los síntomas de la enfermedad. En dos sistemas de enfermedades inducidos por X. fastidiosa estudiados, las chicharritas no evitaron las plantas infectadas pero asintomáticas o las plantas sanas



pintadas para imitar los síntomas de la enfermedad, sino que discriminaron entre las plantas infectadas y sintomáticas. Este comportamiento puede ser ventajoso para estos insectos: las plantas estresadas por el agua e infectadas por X. fastidiosa tienen algunas características fisiológicas compartidas, de las cuales la savia del xilema a alta tensión es de suma relevancia. El aumento de la tensión en una columna de agua conduce a una fuente de alimento que es energéticamente costosa para los insectos, lo que resulta en la ingestión de menos savia de xilema y promueve el movimiento de vectores a otro huésped.


Xylella fastidiosa es una bacteria gram negativa y limitada al xilema, que causa numerosas enfermedades en muchos árboles frutales y de sombra urbanos de importancia económica, así como en la vid


están fuertemente colonizadas por X. fastidiosa, evitar las plantas infectadas por vectores puede reducir la tasa de transmisión cuando la incidencia de la enfermedad es baja y seleccionar la disminución de la virulencia bacteriana. Este efecto podría ser importante cuando las tasa de transmisión es baja. Sin embargo, si los vectores son comunes y las tasas de transmisión altas, se puede favorecer el rápido crecimiento bacteriano que conduce a una mayor virulencia, un patrón que se observa a menudo en enfermedades que se transmiten entre vectores.
La vía más común que conduce a las epidemias de X. fastidiosa es la introducción de genotipos exóticos en ambientes que son ecológicamente propensos al mantenimiento de la bacteria en la comunidad vegetal, principalmente debido a la disponibilidad de insectos vectores endémicos y especies de plantas huésped susceptibles a infecciones persistentes. Aunque la introducción de insectos vectores portadores de X. fastidiosa representa una vía potencial, sólo una especie de vector se considera invasora (Ho-
malodisca vitripennis, Cicadellidae, un saltahojas francotirador), y otra se distribuye más allá de su región de origen (Philaneus spumarius, Aphrophoridae, un salivazo). La expansión en el rango geográfico de estas especies no se ha asociado con la propagación de X. fastidiosa, por lo tanto, consideramos que esta es una ruta poco probable. La principal vía de dispersión sería entonces el movimiento de material vegetal infectado, y potencialmente asintomático, desde las áreas donde se produce el patógeno.
En 2002 algunos investigadores determinaros que este patógeno podía ser causante de algunas de las más recientes amenazas de enfermedades más significativas en el continente americano. Fue de gran interés, asimismo, determinar cómo un patógeno de plantas relativamente oscuro se convirtió en un par de años en el primer patógeno bacteriano de plantas en tener su genoma secuenciado. Diez años más tarde, la bacteria X. fastidiosa ha emergido de la oscuridad y han surgido nuevas enfermedades y las enfermedades que causa se han convertido en
amenazas mundiales. Además, hasta principios de la década de 2000 solo se publicaban anualmente un puñado de manuscritos revisados por pares sobre X. fastidiosa, y eso ha cambiado significativamente durante la última década. Por lo tanto, nuestra comprensión de X. fastidiosa y las enfermedades que causa ha mejorado sustancialmente.
LA ENFERMEDAD DEL CÍTRICO AMARILLO DE TAMAÑO
La clorosis variegada de los cítricos puede ser transmitida por insectos francotiradores de la familia Cicadellidae, yemas de plantas contaminadas e injertos de raíces naturales. Es una enfermedad que se ha extendido rápidamente a todas las áreas productoras de cítricos en Brasil, alcanzando una incidencia promedio del 35% en el estado de São Paulo. Una enfermedad similar conocida como "pecosita" fue encontrada en algunas regiones de Argentina. Aunque los naranjos dulces afectados por la enfermedad no mueren, la fruta de los árboles puede ser de tamaño muy

inferior, dura y carecer de jugo. Dicha fruta es inaceptable tanto para el jugo como para el mercado fresco.
El nombre en inglés de la enfermedad de los cítricos, clorosis variegada de los cítricos, proviene de la llamativa variegación clorótica inducida en las hojas de naranja dulce por el patógeno. Sin embargo, la enfermedad también se conoce como la enfermedad del pequeño cítrico amarillo, nombre que enfatiza la importancia de los síntomas de la enfermedad en la fruta, que incluyen coloración prematura y falta de llenado hasta el tamaño normal. Los síntomas típicos de la enfermedad se pueden reconocer fácilmente desde la distancia. Debido a que estos síntomas de la fruta son fáciles de reconocer para los productores y es el tamaño más pequeño de la fruta lo que causa pérdidas económicas, la mayoría de los productores de cítricos brasileños prefieren el nombre amarelinho de citros para la enfermedad.
La gravedad del problema de la enfermedad clorosis variegada de los cítricos contribuyó a la selección de una cepa cítrica de X. fastidiosa como la primera bacteria fitopatógena en tener su genoma completo secuenciado. La cepa seleccionada fue la 9a5c, obtenida de plantas de naranja dulce infectadas con clorosis variegada de los cítricos en Brasil. Se han logrado muchos avances impor-
La clorosis variegada de los cítricos una enfermedad destructiva de la naranja dulce, Citrus sinensis, descubierta por primera vez en São Paulo, Brasil, en 1987. Posteriormente se demostró que el agente causante es nuevas cepas de X. fastidiosa
tantes en la investigación con las cepas cítricas de X. fastidiosa, incluidas pruebas de patogenicidad mejoradas, identificación de huéspedes experimentales alternativos, transformación genética y métodos moleculares para la diferenciación de cepas. Se ha prestado una atención similar a la cepa de la enfermedad de Pierce. Al menos algunas cepas de X. fastidiosa, incluidas las cepas de melocotón y cítricos, pueden colonizar todo el sistema vascular de la planta, incluidas las raíces. Sin embargo, la presencia y distribución de la bacteria en frutos y semillas ha recibido poca atención. En un informe, los insectos vectores de X. fastidiosa no pudieron transmitir el patógeno después de alimentarse de racimos de uvas cosechados de vides con la enfermedad de Pierce. Sin embargo, en este experimento, la mayor parte de la alimentación de los insectos se produjo en los tallos del racimo de frutos en lugar de en los frutos mismos. No se abordó la posible presencia de X. fastidiosa en la semilla de uva de dicha fruta.
Aunque se encontró X. fastidiosa en pedúnculos de naranjas dulces cuando la bacteria se asoció por primera vez con la enfermedad en 1990, la colonización de la fruta y la semilla de la naranja dulce por el patógeno no ha recibido atención. Los parientes más cercanos de X. fastidiosa pertenecen al género Xanthomonas. Se sabe
que algunas xantomonas infectan y se transmiten por semillas infectadas. Sin embargo, el hecho de que X. fastidiosa esté limitada en las plantas a los elementos vasculares del xilema ha persuadido a muchos investigadores de que la transmisión de X. fastidiosa por semilla es imposible.
Detectamos la bacteria mediante PCR en el pedúnculo, así como en los haces axial, dorsal y marginal, y septos de frutos enfermos de los tres cultivares de naranja dulce analizados.
También se detectó X. fastidiosa en la capa externa o testa de las semillas, así como en los embriones. La alta tasa de infección de la semilla es consistente con la diseminación de X. fastidiosa a través del sistema vascular desde el árbol infectado hasta la fruta y hacia la semilla. No fue posible separar completamente la cubierta interna de la semilla, tegumento, de los embriones --incluidos los cotiledones-- o la cubierta externa de la semilla, por lo que la cubierta interna de la semilla estaba presente en partes tanto de la cubierta externa de la semilla como en las muestras de embriones. Incluso con esta advertencia, nuestros resultados mostraron claramente infección de la cubierta de la semilla, testa más tegumento, con X. fastidiosa.

La colonización, supervivencia y proliferación de patógenos bacterianos exógenos puede verse fuertemente influenciada por la microbiota productora

podría ser una fuente importante de contaminación bacteriana
POR
Un hecho preocupante en la producción agrícola es que el agua de riego puede convertirse en una fuente de patógenos transmisibles a las cosechas. El agua utilizada puede estar contaminado por desbordes de aguas residuales, escorrentía de aguas pluviales contaminadas y escorrentía agrícola. Muchos patógenos bacterianos son capaces de sobrevivir o incluso proliferar durante un tiempo prolongado en el agua de riego contaminada, dependiendo de sus condiciones de aireación.
Los patógenos transmitidos por los alimentos pueden ser dispersados por el agua de riego contaminada y adherirse a la superficie de la planta. Pueden entrar a través de estomas y hidatodos, y luego persistir o proliferar durante la producción y el posterior almacenamiento/distribución. Debido al empeoramiento de la escasez de agua dulce y al aumento de la demanda mundial de alimentos, es necesario explorar fuentes alternativas de agua para uso agrícola. La calidad del agua de riego incide sobre la productividad
de los cultivos, la presencia de microorganismos en ella puede significar un efecto benéfico o uno perjudicial. Un sistema de recuperación biológico natural, para la contaminación de las aguas, se da a través de las microalgas, éstas son microorganismos benéficos en el agua, y cuando los ríos o las lagunas tienen grandes cantidades, éstas se alimentan generalmente de bacterias y utilizan la energía solar para fijar CO2, proceso que oxigena el agua y ayuda a la eliminación de microorganismos contaminantes. Naturalmente, la producción agrícola y el agua siempre han estado incondicionalmente relacionadas. Al ser una fuente natural renovable, los recursos hídricos pueden agotarse por el uso inadecuado y la demanda en la producción agrícola. En muchas regiones del planeta la disponibilidad del agua se limita por los cambios climáticos que conllevan a largas temporadas de sequía.
También existen plantas como las macrofitas que ayudan en la depuración de contaminantes de aguas residuales. En otros casos hay bacterias, hongos y parásitos que están contaminando estas aguas

y pasan a los cultivos, de tal forma que pueden generar algún tipo de fitopatología dependiendo de la susceptibilidad de este a la acción del patógeno. La filosfera de los productos agrícolas alberga muchas comunidades microbianas grandes y diversas, donde las bacterias suelen ser los habitantes más abundantes. Aunque la mayoría de las bacterias residenciales son comensales o simbióticas, los patógenos de plantas y, a veces, los patógenos humanos transmitidos por los alimentos pueden ser miembros de estas comunidades microbianas. Los brotes recientes y pasados de enfermedades transmitidas por los alimentos asociadas con los productos frescos han suscitado preocupaciones sobre la contaminación por patógenos transmitidos por los alimentos durante la producción y distribución de productos frescos. La microbiota de los productos frescos
puede tener un profundo efecto en la supervivencia y la persistencia de los patógenos contaminantes transmitidos por los alimentos.
Comprender el impacto del riego con fuentes de agua alternativas en la composición y la población de la microbiota en la filosfera de productos puede proporcionar conocimientos para evaluar y prevenir los riesgos potenciales de infección por patógenos de plantas y contaminación de patógenos transmitidos por alimentos durante el riego. Los avances recientes en la secuenciación de ADN de alto rendimiento y los análisis de secuencias facilitan en gran medida la identificación y el análisis de comunidades bacterianas complejas. La transmisión de enfermedades transmitidas por los alimentos por patógenos presentes en los alimentos de origen animal, como la carne, las aves de corral y la leche
cruda, está bien establecida, pero cada vez hay más conciencia de que las frutas y verduras frescas o mínimamente procesadas también pueden ser fuentes de bacterias patógenas, protozoos, virus y helmintos.
OTRAS FUENTES DE CONTAMINANTES PARA LAS COSECHAS DE CONSUMO EN FRESCO
Las bacterias patógenas incluyen Campylobacter spp., Clostridium botulinum, Clostridium perfringens, Bacillus cereus enterotoxigénico, Escherichia coli O157:H7 y otras E. coli productoras de toxina Shiga, Listeria monocytogenes, Salmonella spp., Shigella spp., enterotoxigénicas Staphylococcus aureus, Vibrio cholerae y Yersinia enterocolitica. Los protozoos patógenos incluyen Cyclospora cayetanensis, Cryptosporidium parvum, Giardia lamblia y Entamoeba histolytica. Los virus pató-
genos incluyen hepatitis A, enterovirus, echovirus, rotavirus y virus similares a Norwalk. El nematodo (gusano redondo) Ascaris lumbricoides es un ejemplo de patógeno helmintoico que puede transmitirse a través de los alimentos. Aunque algunos patógenos, como B. cereus, S. aureus, C. perfringens, C. botulinum y L. monocytogenes, se encuentran comúnmente en el medio ambiente, la presencia de la mayoría de estos patógenos es indicativa de contaminación fecal humana o animal reciente.
Las frutas y verduras pueden contaminarse con microorganismos patógenos por contacto con el suelo o el estiércol compostado incorrectamente, el riego o el lavado poscosecha con agua contaminada, o el contacto con manipuladores de alimentos infectados. Las fuentes de agua de riego incluyen aguas subterráneas, aguas superficiales y aguas residuales humanas. El agua subterránea se encuentra en los acuíferos debajo de la superficie de la tierra.
La evidencia de que el agua de riego contaminada podría ser una fuente de patógenos transmitidos por los

alimentos en frutas y verduras se puede encontrar en investigaciones epidemiológicas de brotes de intoxicación alimentaria, estudios experimentales que examinaron la contaminación por E. coli O157:H7 de lechuga y observaciones de una mayor incidencia de enfermedades en áreas que practican el riego de aguas residuales con poco o ningún tratamiento de aguas residuales. El agua de riego estuvo implicada en brotes de infección por E. coli O157:H7 por lechuga contaminada e infección por C. cayetanensis por lechuga y frambuesas contaminadas.
El agua de riego se implica como una fuente de E. coli detectada en plán-
tulas de repollo regadas con agua contaminada con aguas residuales porque no se encontró ninguna en las plántulas en un campo adyacente regado con agua municipal. Los estudios experimentales que examinaron la contaminación de la lechuga con E. coli O157:H7 demostraron que el agua de riego podía transmitir efectivamente E. coli a las plantas de lechuga.
Riesgos potenciales en la aplicación de fuentes de agua no convencionales en la agricultura, incluida la posible contaminación de patógenos transmitidos por los alimentos


La comprensión de las relaciones interactivas entre el suelo, las plantas y el clima con respecto a la disposición y utilización ideales del agua continúa evolucionando







POR FEDERICO SALAS CAMACHO
Objetivo importante en el diseño de los sistemas de riego agrícola es el de administrar y optimizar la ubicación y el momento de las aplicaciones con el fin de promover el crecimiento y el rendimiento de las cosechas, al tiempo que protegen contra la erosión del suelo, la salinización, la degradación de la calidad del agua u otros impactos ambientales perjudiciales.
Desafortunadamente no existe una respuesta única para diseñar, desarrollar y administrar un sistema de




riego dado que las circunstancias físicas y las condiciones socioeconómicas son específicas de cada sitio. En todas las circunstancias, sin embargo, los factores y principios involucrados son universales. La práctica del riego ha evolucionado gradualmente hacia un mejor control sobre las plantas, el suelo e incluso las variables climáticas. El grado de control posible hoy en día es todavía sólo parcial debido a los extremos impredecibles del clima; además, con la continua pérdida de tierras y agua adecuadas y la creciente demanda de productos agrícolas, la búsqueda de nuevos conocimientos sobre cómo mejorar el riego y la
necesidad de aplicar estos nuevos conocimientos se han vuelto cada vez más urgentes. Cualquier intento de riego debe basarse en una comprensión profunda de las relaciones suelo-agua-planta.
El movimiento del agua, una vez aplicada, consiste en una secuencia de procesos dinámicos que comienzan con la entrada de agua en el suelo, llamada infiltración. La tasa de infiltración se rige por la velocidad a la que se aplica el agua a la superficie del suelo, si la tasa de aplicación no excede la capacidad del suelo para absorberla. Un criterio importante para un siste-

ma de aspersión o microirrigación es entregar agua a una tasa que evite el encharcamiento, la escorrentía y la erosión. Después de la infiltración, el agua normalmente continúa moviéndose debido a la gravedad y los gradientes hidráulicos en el suelo. El agua se mueve hacia abajo y, con algunos sistemas de riego, lateralmente en un proceso llamado redistribución. En este proceso, la zona relativamente seca y profunda del perfil del suelo absorbe el agua que drena de las zonas más húmedas de arriba. A los pocos días, dependiendo del sistema de riego y de la gestión, el caudal se vuelve tan bajo que es insignificante. El contenido de agua de la zona mojada a medida que el flujo se vuelve insignificante se denomina capacidad de campo y representa el límite superior de la capacidad del suelo para almacenar agua. La capacidad de campo
es normalmente mayor en suelos arcillosos que en arenosos.
Cualquier agua que drene por debajo de la zona de las raíces generalmente se considera una pérdida desde el punto de vista del uso inmediato del agua de la planta. Sin embargo, no es necesariamente una pérdida final. Si el área está subyacida por un acuífero explotable, el agua que se filtra por debajo de la zona de raíces puede eventualmente recargar el acuífero y recuperarse mediante bombeo. Alguna percolación profunda puede regresar más tarde a los arroyos o sistemas de drenaje. Esta cantidad de agua más la escorrentía superficial de la agricultura de regadío se denomina flujo de retorno. Cuando el nivel freático está cerca de la superficie del suelo, parte del agua puede entrar en la zona de las raíces por ascenso
capilar desde la zona saturada por debajo del nivel freático y suministrar una parte de las necesidades hídricas del cultivo. Este proceso de subirrigación, sin embargo, puede infundir sales en la zona de las raíces. El agua que fluye a través de la zona de las raíces puede lixiviar sales solubles o nutrientes de los cultivos y degradar la calidad del agua subterránea.
Los métodos modernos de riego, diseñados y gestionados adecuadamente, pueden aumentar el rendimiento de los cultivos, evitando al mismo tiempo el desperdicio, reduciendo el drenaje y promoviendo la integración del riego con las operaciones esenciales de gestión de cultivos concurrentes. El uso del agua degradada se ha vuelto más factible, y los suelos de textura gruesa, las tierras con pendiente pronunciada y los suelos pedregosos, que antes se

consideraban no irrigables, ahora son productivos. Tales avances y sus consecuencias eran imprevistos hace solo unas décadas.
PAPEL DEL RIEGO AGRÍCOLA EN LA PRODUCCIÓN ESTABLE DE ALIMENTOS
El riego es el suministro de agua a los cultivos por medios artificiales. Está diseñado para permitir el crecimiento deseado de las plantas en las regiones áridas y para compensar la sequía en las regiones semiáridas o subhúmedas. Incluso en zonas donde la precipitación media estacional puede parecer abundante, las lluvias

El riego moderno es una operación sofisticada que implica el monitoreo y la manipulación de numerosos factores que afectan la producción de cultivos
suelen estar distribuidas de manera desigual o los suelos tienen una baja capacidad de retención de agua, por lo que la agricultura tradicional de secano es una empresa de alto riesgo. En algunas áreas, el riego prolonga la temporada de crecimiento efectiva. Con la seguridad que proporciona el riego, los insumos adicionales, como las variedades de mayor producción, los fertilizantes adicionales, un mejor control de plagas y una mejor labranza, se vuelven económicamente viables. El riego reduce el riesgo de que estos costosos insumos se desperdicien por la sequía. A escala mundial, el riego tiene un profundo impacto en el suministro de agua dulce, la producción mundial de alimentos y la estética y el valor de los paisajes. Un tercio de los alimentos del mundo proviene del 21% de la superficie cultivada del mundo que se riega.
El riego ha convertido muchas de las tierras más secas y fértiles de la tierra en importantes regiones productoras de cultivos. Por ejemplo, Egipto no podía cultivar prácticamente ningún alimento sin agua extraída del Nilo o
de los acuíferos subterráneos. El Valle Central de California y la cuenca del Mar de Aral, las cestas de frutas y verduras de los Estados Unidos y la antigua Unión Soviética, producirían poco sin riego. Las principales áreas productoras de granos del mundo son el norte de China, el noroeste de la India y los EE. UU. Las Grandes Llanuras disminuirían entre un tercio y la mitad sin riego para complementar las precipitaciones. El riego desempeña un papel clave en la alimentación de una población mundial en expansión y parece destinado a desempeñar un papel aún mayor en el futuro.
Sin embargo, tal como se practica en muchos lugares, el riego todavía se basa en gran medida en métodos tradicionales que no logran medir y optimizar el suministro de agua para satisfacer las demandas de agua de las plantas. El riego no medido tiende a desperdiciar agua, nutrientes y energía, y puede causar la degradación del suelo por anegamiento, erosión y salinización. La tarea vital de asegurar una producción mundial adecuada de alimentos debe incluir un esfuerzo concertado para modernizar los sistemas de riego y mejorar la gestión de los recursos hídricos. Estas técnicas mejoradas ayudarán a lograr una producción sostenible y eficiente,

al tiempo que protegen el medio ambiente. Los nuevos sistemas deben basarse en principios y diseños sólidos para optimizar el riego en relación con los insumos y las operaciones esenciales, garantizando al mismo tiempo la sostenibilidad de la agricultura de regadío. El agua y el suelo deben ser reconocidos como recursos vitales, preciosos y vulnerables, y gestionados en consecuencia.
CANALES DE RIEGO Y PUNTOS DE DESVÍO DEL AGUA DE CIVILIZACIONES TEMPRANAS
Durante miles de años, el riego ha contribuido sustancialmente a la producción mundial de alimentos. Los historiadores señalan que el riego fue una de las primeras modificaciones del entorno natural emprendidas por las primeras civilizaciones. Hace varios milenios, la irrigación permitió
a las tribus nómadas establecerse en comunidades más estables con la garantía de la productividad anual de los cultivos. Los primeros intentos de irrigación fueron rudimentarios, consistiendo en encharcar el agua en cuencas encerradas por diques bajos de tierra.
Las primeras sociedades que dependieron con éxito de la irrigación se encontraban en cuatro cuencas fluviales principales: el Nilo en Egipto alrededor del año 6.000 a.E.C., el Tigris y el Éufrates en Mesopotamia alrededor del 4.000 a.E.C., el río Amarillo en China alrededor del 3.000 a.E.C., y el Indo en la India aproximadamente 2.500 a.E.C. En México y América del Sur, las civilizaciones maya e inca practicaron el riego hace más de 2.000 años. En Irán, los ganats, túneles de 3.000 años de antigüedad para llevar agua de las montañas al valle, se utilizan hasta el










día de hoy. Las presas de tierra para almacenar agua superficial se construyeron por primera vez en los siglos II y III en Japón para regar el arroz.
En Europa central, la irrigación se documentó ya en el siglo III E.C. En América del Norte, se sabe que la irrigación existió entre los nativos americanos del suroeste ya en el año 1200 a.C. Los primeros exploradores españoles encontraron evidencia de canales de irrigación y puntos de desvío a lo largo de los ríos. Los españoles introdujeron nuevos métodos de irrigación y regaron cultivos como uvas, frutas, verduras, olivos, trigo y cebada. Al igual que en otras áreas del mundo, la irrigación hizo posible que los nativos americanos desarrollaran asentamientos y disfrutaran de una fuente de alimentos más segura.












La modernización asociada con el control, el rendimiento, la flexibilidad y, sobre todo, el ahorro de agua se ha vuelto importante para garantizar la estabilidad y la seguridad de la producción de alimentos, así como para promover un mayor consciente uso del agua en tiempos de escasez
suelo, la planta y el microclima
POR URIEL PANTOJA SÁNCHEZ
Un sistema de unificación en el que todos los procesos involucrados en la producción de cosechas agrícolas son interdependientes es el denominado relación suelo-planta-atmósfera y es precisamente en esta relación en la cual la gestión moderna del riego se ve fundamentada.
Cabe recordar que la disponibilidad de agua del suelo no es una propiedad privativa del mismo sino una función de la planta, el suelo y el medio ambiente. La tasa de absorción de agua por parte de la planta depende de la capacidad de la raíz para absorber agua del suelo, la capacidad del suelo para transmitir agua hacia las raíces y la demanda de evaporación de la atmósfera. Estos, a su vez,




dependen de características propias de la planta como lo son la densidad de enraizamiento, la profundidad de enraizamiento, la tasa de crecimiento de las raíces y la capacidad de la planta para mantener sus funciones vitales bajo estrés hídrico. Además, propiedades del suelo como la conductividad hidráulica, la densidad aparente del suelo, la textura del suelo, las capas del suelo, la retención de agua y la capacidad de agua disponible y por último las condiciones climáticas que dictan la tasa de transpiración del cultivo y la evaporación del suelo. Estos componentes del continuo se presentarán y discutirán para el uso del agua del suelo y del agua de las plantas.
El manejo adecuado del agua de riego es uno de los problemas más complejos y urgentes que enfrenta el mundo debido a diversos retos del mundo actual: económicos, ambientales y sociales, donde la creciente
competencia entre los municipios, la industria y la agricultura debe ser abordada debido al interés e importancia de la sociedad. En la agricultura, la programación del riego es el término que describe el procedimiento mediante el cual un regante determina el momento y la cantidad de la aplicación de agua. Es posible programar riegos en función del monitoreo del suelo, la planta y/o el microclima. Al monitorear la humedad del suelo, la idea es medir la reserva de agua dentro de la zona de las raíces del cultivo a medida que disminuye después de cada riego para determinar cuándo el agua del suelo se ha agotado a un nivel mínimo prescrito. La detección del estado hídrico de la planta es un segundo método para detectar el comienzo del estrés hídrico de la planta. Hay muchas técnicas de detección de plantas disponibles hoy en día para medir o inferir el estado del agua de las plantas,
Conociendo la capacidad del suelo para almacenar agua --o monitoreando el estado hídrico de la planta-- es como se establece el momento óptimo del riego
que van desde equipos especializados hasta observación visual. A pesar de lo importante que es la detección temprana del estrés hídrico de las plantas, no proporciona información sobre la cantidad de agua que se debe aplicar. Una tercera técnica consiste en el seguimiento de las condiciones meteorológicas que imponen la demanda evapotranspiracional sobre el cultivo. Acumulando la cantidad de agua perdida a la atmósfera por el cultivo se estimará la cantidad de agua a aplicar.
Los problemas de calidad del agua incluyen tanto las aguas subterráneas como las superficiales. El riego a menudo resulta en una percolación profunda, lo que resulta en la lixiviación de fertilizantes solubles u otros productos químicos. En algunas zonas, la lixiviación de nitratos ha dado lugar a concentraciones de nitratos en las aguas subterráneas superiores a la concentración máxima permitida para el consumo humano. La percolación profunda se puede minimizar con una buena programación del riego. Los escurrimientos del riego pueden contener nutrientes, pesticidas y sedimentos. Esto puede ser particularmente un problema en los sistemas de riego superficial donde tales escurrimientos se recolectan y reutilizan en otros campos de cultivo.



Rancho Grande No. 142, Col. Santa Cecilia, Coyoacán, 04930, Ciudad de Mexico www.comiisa.com.mx

(55)554267064 contacto@comiisa.com.mx
La quimigación debe gestionarse bien para evitar la aplicación de productos químicos en los cuerpos de agua superficiales o la contaminación de las aguas subterráneas a través del reflujo del sistema de riego. Por último, la calidad del suelo también es motivo de preocupación. Los sistemas de riego que provocan escorrentía también pueden desencadenar la erosión del suelo. En las regiones áridas, la acumulación de sal en el suelo puede ser una preocupación importante dependiendo de la salinidad del agua de riego. En estas situaciones, a menudo es necesario incluir drenaje subterráneo junto con el sistema de riego. Una buena gestión del riego debe tratar de aumentar la producción de alimentos --y los beneficios de las explotaciones agrícolas-- y minimizar los impactos negativos sobre los recursos hídricos y el medio ambiente. En muchas situaciones, las mejores







EFICIENCIA OPERATIVA Y DE USO DE LOS RECURSOS Y RECUPERACIÓN DE COSTOS DE SISTEMAS DE RIEGO
El riego moderno es una palabra sinónimo de riego por aspersión y goteo el cual es el método de riego que se usa generalmente en nuevas tierras desérticas con suelo arenoso común. Al mismo tiempo el riego tradicional o por inundación es el método de riego habitualmente utilizado en las tierras antiguas, concretamente en suelos arcillosos comunes. Mientras tanto, el riego moderno se denomina riego presurizado, ya que requiere bombear agua a través de la red de tuberías en comparación con el riego por inundación o gravedad.
operativa, recuperación de costos, externalidades ambientales y sostenibilidad financiera, en lugar de solo actividades operativas humanas. En cualquier caso, el uso eficiente y ambientalmente sostenible del agua de riego en el sistema para el cultivo de cultivos en una cantidad precisa en un momento adecuado a lo largo de todo el ciclo del cultivo es utilizar prácticas de gestión de sistemas de riego. Por lo tanto, la capacidad de gestionar el sistema de riego es esencial por sus efectos directos en la calidad y la productividad de los cultivos, especialmente en el contexto de la política y el entorno jurídico prevalecientes para establecer normas para el uso del agua de riego, como los protocolos de calidad del agua y los métodos agrícolas. El diseño del sistema de rie-
Febrero - Marzo 2025
go implica la selección de diferentes tipos de niveles de riego de las fuentes de agua, sistemas de regulación, niveles de automatización, considerando las restricciones hidráulicas. La gestión del sistema de riego está relacionada con los sistemas de monitoreo, mapeo y apoyo a la toma de decisiones para las actividades operativas y de mantenimiento para evitar pérdidas, daños o desperdicios de agua.
Los sistemas de riego por goteo son uno de los principales objetivos de la modernización de los sistemas de riego tradicionales es transformarlos en otros más eficientes y automatizados que utilicen menos agua, por lo que la estructura física y las modificaciones de los antiguos sistemas de riego han innovado a lo largo del tiempo en el marco de la gestión integrada del agua. La modernización asociada con el control, el rendimiento, la flexibilidad y, sobre todo, el ahorro de agua se ha vuelto importante para garantizar la estabilidad y la seguridad de la producción de alimentos, así como para promover un mayor uso del agua en tiempos de escasez de agua. Cuanto más rápido un país pueda modernizar sus sistemas de riego, más soluciones beneficiosas para los alimentos se pueden experimentar en el sector agrícola. De los métodos y técnicas de riego, uno que está desempeñando un papel cada vez más importante en la modernización del rendimiento de los sistemas de riego es la microirrigación. Entre las técnicas de microirrigación,
En la búsqueda por mantener la sostenibilidad el manejo de los riegos enfrenta muchos retos
las subterráneas son las más prometedoras debido a su menor pérdida de agua y a las condiciones más favorables de clima y cultivo.
El advenimiento de esas tecnologías avanzadas no solo ayuda a redefinir el entorno virtual de la gestión de sistemas agrícolas o agrícolas de precisión, sino que también mejora el ámbito de la gestión de sistemas de riego. Los sistemas de riego convencionales o modernos ya sean irrigadores de barra o sistemas de goteo, generalmente vienen en combinación con tecnologías de agricultura de precisión para monitorear el progreso del riego, optimizar la programación del riego, llevar a cabo un riego de tasa variable y diagnosticar ayudantes de fitohidratación a través de la gestión de datos. Operando bajo un comando de sistema de riego neutro, el sistema de riego convencional generalmente aplica un riego uniforme a la área designada de un determinado cultivo. Esto se amplió aún más a través de los servicios
convencionales del sistema de riego basados en sistemas informáticos como las imágenes satelitales.
Antes de la llegada de la tecnología digital avanzada, la gestión de precisión del riego de cultivos se realizaba mediante la programación manual del riego. La gestión del riego se lleva a cabo sobre la base de índices observables del estado hídrico de las plantas o del desarrollo fisiológico. A continuación, se utilizaron modelos interrelacionados de agua de cultivo como base para la programación de las necesidades de riego. A medida que el advenimiento de tecnologías digitales avanzadas se generaliza en los sistemas agrícolas, las tecnologías convencionales siempre estuvieron equipadas con tecnología de visión por computadora, Internet de las cosas, sensores remotos e Inteligencia Artificial, aprendizaje automático, entre ellos, equipos de intervención y automatización, con el objetivo de soluciones de agricultura económica de precisión. Las soluciones para estanques bajo servicios básicos de calidad pueden relacionarse con los sistemas convencionales de gestión de riego que utilizan visión artificial, Internet de las cosas, sensores remotos, inteligencia artificial y aprendizaje automático. Las soluciones basadas en software entran en el dominio de la agricultura de precisión que se descubrió que tiene un impacto positivo en los beneficios económicos de diversas prácticas agrícolas de riego.


POR BEATRIZ CARRANCO LINO
El higo es un árbol frutal que se puede cultivar en una amplia gama de suelos, incluidas arcillas pesadas, margas y arenas ligeras, siempre y cuado exista un buen drenaje. Es una especie que no suele verse seriamente afectada por plagas excepto en las zonas de alta precipitación. Debe recogerse cuando esté ligeramente inmaduro para que esté lo suficientemente firme para el procesamiento.
La higuera es tolerante a una amplia gama de condiciones ambientales, tiene un bajo requerimiento de enfria
miento, soportará algunas heladas y es tolerante a la sequía, aunque crece más vigorosamente con abundante agua, pero para la producción de fruta fresca es esencial un clima seco con lluvias ligeras porque las lluvias intensas durante la fructificación y la maduración son perjudiciales.
El higo común es una especie vegetal ginodioica con dos géneros diferentes: árboles femeninos que producen siconos con flores femeninas que se convertirán en higos comestibles con semillas (sicono con múltiples frutos de una sola semilla o drupas) y caprifigs que producen siconos con flores masculinas y femeninas que presen
Sus frutos son una buena fuente natural de nutrientes, minerales y fitoquímicos, que pueden mejorar la salud humana
tan un estilo más corto que el fruto de los árboles femeninos. El polen solo es producido por caprífigas, por lo que el sistema reproductivo es funcionalmente dioico. El fruto deniminado breba nace en brotes viejos y son más grandes y jugosos que los higos de la cosecha principal, que se producen por las axilas de las hojas.
Estas frutas son muy nutritivas y se consumen frescas o secas en todo el mundo. Estos frutos son apreciados por su tamaño, color claro y dulzura. Además, son una excelente fuente de minerales, vitaminas, fibra y polifenoles, especialmente antocianinas y otros flavonoides que actúan co-

mo antioxidantes y pueden proteger contra varias enfermedades degenerativas comunes. En consecuencia, el consumo de brebas e higos tiene un papel importante en la contribución positiva a la salud humana y la protección contra varias enfermedades humanas.
En las condiciones de cultivo adecuadas, las higueras producen una o dos cosechas al año, dependiendo del cultivar. Anualmente, se cosechan más de 1 millón de toneladas de higos frescos en 308,460 hectáreas en todo el mundo. Los países mediterráneos son los principales productores de higos. Argelia produce el 12.54% de la cosecha mundial total de higos. Los trabajadores deben usar guantes y ropa protectora debido al látex. Las frutas cosechadas se extienden a la sombra durante un día para que el látex se seque un poco. Se trata de una fruta compuesta llamada ''siconio''. El fruto maduro del higo comestible tiene una piel algo dura, una corteza interior blanquecina y una pulpa dulce y gelatinosa compuesta por las drupas maduras individuales. Las semillas dentro de las drupas van desde prácticamente inexistentes hasta sutilmente crujientes. Las higueras son propensas al ataque de nematodos del género Meloidogyne, pero el control es posible con la aplicación adecuada de nematicidas.
Además, un problema común es la roya de la hoja, la mancha foliar y el mosaico de higueras. El higo comestible es una fruta dietética muy importante para la salud humana debido a su alto valor nutritivo, contenido de fibra y peculiaridad laxante. Reduce el colesterol, controla la presión arterial, ayuda en la pérdida de peso, previene el estreñimiento, aumenta la densidad ósea, previene el cáncer, es bueno para quienes sufren de anemia y previene los ataques de asma. Por lo tanto, se consume en forma fresca, seca, enlatada y conservada en todo el mundo. El valor nutricional de los higos frescos es comparable al de muchas otras

A nivel comercial son tres los tipos comestibles de higo hembra que se cultivan principalmente; el tipo de higo común que desarrolla frutos partenocárpicamente, el tipo Smyrna que necesita polinización de caprífigos --caprificación-- para desarrollar frutos, y el tipo San Pedro que produce una primera cosecha partenocárpica y una segunda o principal cosecha solo después de la caprificación. Los higos de tipo común pueden producir una, tipos unifera, o dos cosechas, tipos bifera
frutas. Es probable que la importancia económica de la producción de higos continúe en el futuro. En el mercado mundial, existe una creciente demanda de higos frescos y una demanda estable de higos secos. Por ejemplo, el higo comestible es una nueva fruta de Bangladesh. Bangladesh tiene un clima excelente y tierras fértiles, también agua adecuada para el riego y una amplia variedad de suelos, así como un recurso humano dinámico. Estas características favorables permiten el cultivo de una amplia gama de árboles frutales.
Estos frutos tienen formas, colores, sabores, propiedades tecnológicas y terapéuticas que difieren entre las variedades y generalmente se nombran de acuerdo con su forma, color y la
región en la que más se cultiva. En 1850, se introdujeron en Argelia más de 70 variedades de higos. Sin embargo, estos no fueron adoptados por los agricultores locales, que continuaron cultivando las higueras que les eran familiares. Recientemente, el Instituto Técnico de Arboricultura Frutícola de Argelia describió 40 variedades, incluidas las variedades comestibles y los tipos caprifig. Sin embargo, es probable que el número de cultivares cultivados en Argelia sea mucho mayor que estas 40 variedades.
Además de ser una excelente fuente de minerales, vitaminas y fibra dietética, son libres de grasas y colesterol y contienen una alta cantidad de aminoácidos. Al igual que otras especies frutales, los higos contienen azúcares y ácidos orgánicos que influyen en su calidad. Se han realizado varios estudios sobre los compuestos fenólicos de los higos como ácidos fenólicos, flavonoides y antocianinas con capacidad antioxidante en los frutos de higo. Los compuestos antioxidantes eliminan los radicales libres, inhibiendo así los mecanismos oxidativos que conducen a las enfermedades degenerativas.
El higo cultivado pertenece a la familia Moraceae, es claramente de la mayor importancia como fuente de alimento humano. El higo probablemente se originó en Asia occidental y se extendió al Mediterráneo. Hoy en día, el higo es un cultivo frutal de importancia moderada en todo el mundo. El género Ficus incluye más de 1,400 especies clasificadas en unos 40 géneros, la mayoría de los cuales se encuentran en los trópicos o subtrópicos y solo un puñado con frutos considerados comestibles. La higuera común es un árbol originario del suroeste de Asia y de la región del Mediterráneo oriental, perteneciente a la familia botánica Moraceae. La higuera se cultiva por su fruto en climas cálidos y secos, y la fruta seca ha sido un alimento familiar para los humanos desde el año 3000 a.C.

Enfermedades de hojas, tallo y raíz debidas a hongos patógenos en la planta de té
POR CAMILO ARTEAGA NAVARRO
Apesar de la naturaleza resistente de la planta de té, Camellia sinensis es vulnerable a enfermedades destructivas causadas por patógenos microbianos. Entre las enfermedades de las hojas, el tizón de la ampolla causado por el hongo Exobasidium vexans Massee, Basidiomycetes, es una enfermedad de importancia económica en los principales países productores de té. Con su ciclo de infección policíclico, el hongo prospera en alta humedad, >80%, y temperaturas suaves de 15 a 25 ° C, especialmente durante los monzones. Infecta selectivamente los brotes tiernos cosechables de la planta de té, lo que provoca importantes pérdidas de rendimiento. Según estudios anteriores, las pérdidas estimadas de cultivos causadas por tizón de la ampolla oscilan entre el 20 y el 50% en Sri Lanka, India e Indonesia, y entre el 40 y el 45% en China.
Las hojas tiernas infectadas desarrollan manchas translúcidas de color amarillo pálido de 8 a 10 días después de la infección. Estas manchas translúcidas se convierten gradualmente en ampollas esporulantes, después

El hongo E. vexans ataca los brotes jóvenes cosechables y causa una pérdida del 20 al 50% de los cultivos. en última instancia conduce a tallos distorsionados y muerte regresiva de los brotes. La muerte regresiva de los brotes, particularmente en arbustos que se están recuperando de la poda, puede retrasar significativamente el proceso de recuperación, extendiendo el período no cosechable. Además, el mal manejo de la enfermedad puede conducir a una eventual muerte regresiva de los arbustos.
de aproximadamente 15 a 19 días. Las ampollas esporulantes generalmente aparecen como una superficie blanca y aterciopelada (himenio) en la lámina abaxial de la hoja causada por millones de esporas que atraviesan la epidermis de la hoja. El himenio en la lámina foliar abaxial y adaxial es común cuando la infección ocurre en las venas. La esporulación y la secreción de esporas son seguidas por el secado del sitio de infección. A medida que la ampolla se seca, proporciona un hábitat adecuado para varios hongos saprófitos, que a menudo atraen a otros patógenos del té, como Pestalotiopsis spp., patógeno del tizón gris, y Colletotrichum spp., patógeno del tizón marrón.
Además de las hojas, los tallos tiernos también son propensos a la infección y producen síntomas similares, lo que
ESTRATEGIAS DE MANEJO INTEGRADO DE LA ENFERMEDAD
El desarrollo y cultivo de variedades de cultivos con resistencia inherente es una estrategia clave a largo plazo para el manejo de enfermedades dentro del manejo integrado. Diferentes cultivares de té exhiben diferentes respuestas a la infección por tizón de la ampolla, demostrando diversos grados de resistencia al patógeno. Debido a las opciones limitadas y los posibles efectos perjudiciales de los fungicidas sintéticos en la salud humana y el medio ambiente, los principales países productores de té encabezan la investigación para aprovechar
la resistencia a las enfermedades de las plantas de té, allanando el camino para soluciones sostenibles en el manejo de las enfermedades del té. Como resultado, se ha generado una amplia base de conocimientos. Este artículo recopila y resume información actualizada sobre el arsenal de defensa de la planta de té contra tizón de la ampolla, explora cómo el patógeno supera la resistencia de la planta y profundiza en aspectos inexplorados de la interacción entre la planta de té y E. vexans para facilitar futuras investigaciones.
Exobasidium vexans es un hongo biotrófico obligado que requiere un huésped para completar su ciclo de vida. E. vexans se ubica bajo el orden Exobasidiales y la familia Exobasidiaceae. El patógeno produce basidiosporas en el aire como propágulos, las corrientes de aire facilitan el transporte y la deposición de esporas ligeras en la superficie de la hoja. Bajo condiciones climáticas favorables, como alta humedad relativa (>80%), disponibilidad de humedad y temperatura suave (15 a 25 ° C), las esporas que se depositan en una superficie foliar húmeda germinan y producen tubos germinativos seguidos de la formación de apresores. Posteriormente, se forma un delgado tubo de infección o clavija de penetración que penetra gradualmente en la hoja, pasando a través de la epidermis superior. Luego, el hongo prolifera en los tejidos de las hojas, formando haustorios para absorber nutrientes. Finalmente, el hongo llega a la epidermis inferior, forma una capa fructífera que se extiende a través de las células epidérmicas y comienza a liberar esporas para nuevas infecciones.
Las plantas de té exhiben diversos grados de resistencia a la infección por tizón de la ampolla, pero ningún cultivar o genotipo posee inmunidad completa. Los estudios han indicado que la resistencia a tizón de la ampolla en la planta de té está controlada por numerosos genes. La diversidad de
cultivares de té en términos de morfología, anatomía y perfiles químicos contribuye notablemente a la resistencia preformada e inducida contra tizón de la ampolla. Las siguientes secciones exploran exhaustivamente los mecanismos empleados por las plantas de té para resistir la infección por tizón de la ampolla.
Por otra parte, las características estructurales de las plantas de té dificultan la penetración directa del hongo E. vexans. Varias características morfológicas y anatómicas se han correlacionado con una mayor resistencia a tizón de la ampolla. Aunque los estudios se han centrado principalmente en las defensas estructurales preexistentes, según nuestra revisión de la literatura, no ha habido investigaciones sobre los mecanismos de defensa estructural inducidos contra tizón de la ampolla. Las hojas de los cultivares resistentes generalmente exhiben una mayor densidad de pubescencia, cera epicuticular, relación longitud-ancho y grosor de cutícula en comparación con las de los cultivares susceptibles. Además, los cultivares resistentes poseen hojas más gruesas y mantienen un ángulo más grande entre la hoja y la ramita. Por el contrario, la densidad estomática es menor en los cultivares resistentes.
SIEMBRA Y CARACTERÍSTICAS GENERALES DEL CULTIVO
Camellia sinensis es el único cultivo de bebidas de la familia Theaceae. Se clasifica en tres variedades taxonómicas: C. sinensis var. sinensis, C. sinensis var. assamica, y C. sinensis var. Lasiocalyx, aunque muchos creen que este último es un híbrido de los dos primeros. El té es una planta con flores bisexuales, autoincompatibles, que exhiben alogamia. En consecuencia, las plantas de té son altamente heterocigóticas y heterogéneas, lo que dificulta la obtención de homocigotos. es un cultivo perenne ampliamente cultivado en países tropicales y subtropicales. La
tradición de beber té tiene una historia de casi 5 mil años y hoy en día sigue siendo la segunda bebida más popular del mundo después del agua. El té se cultiva comercialmente en más de 50 países. Entre ellos, China, India, Kenia, Sri Lanka, Turquía, Vietnam e Indonesia son los mayores productores. Se estima que el valor de mercado de la industria mundial del té alcanzará los 93,200 millones de dólares en 2031, según las previsiones de la industria. Además, la industria del té proporciona un medio de vida a unos 13 millones de personas en todo el mundo.
La adaptabilidad ambiental del té es limitada. Las condiciones ideales para el crecimiento y el desarrollo incluyen elevaciones de hasta 2,000 m sobre el nivel del mar, pendientes que oscilan entre 0.5 y 10 grados, precipitaciones anuales espaciadas uniformemente de al menos 1.250 mm, temperaturas entre 10 y 30° C y suelos ácidos bien drenados con un pH de 4.5 a 5.5.
El té se cultiva predominantemente en monocultivo. El material de siembra (semillas y/o esquejes vegetativos) se prepara en viveros antes de plantar en los campos, con 1.2 m entre hileras y 0.6 m entre plantas. Los árboles de té crecen naturalmente hasta aproximadamente 10 m de altura, pero a menudo se podan para facilitar el crecimiento horizontal, mejorar el acceso a la cosecha y fomentar el crecimiento de nuevos brotes. La parte cosechable del té son los brotes tiernos de té. Esto generalmente incluye una yema apical y dos o tres hojas inmaduras. Una vez cosechada, se libera la dominancia apical y la yema axilar comienza a crecer produciendo un nuevo brote. En los países tropicales, la cosecha de té se lleva a cabo durante todo el año a intervalos de 7 a 10 días. El té es muy sensible a las altas temperaturas. Por lo tanto, los árboles de sombra se cultivan para proporcionar la sombra necesaria y proteger las plantas de té de la radiación solar directa.
en México con
ING.JUAN MANUEL BARRIENTOS
La agricultura mexicana está en constante evolución, adoptando tecnologías innovadoras que impulsan la productividad y sostenibilidad de los cultivos. En este contexto, los bioestimulantes se posicionan como una herramienta revolucionaria, capaces de optimizar el rendimiento agrícola y enfrentar los desafíos del cambio climático y la degradación de los suelos. DUCOR, una empresa líder en biotecnología agrícola, ha desarrollado una línea de productos de bioestimulación que redefine las prácticas en el campo.
Con el propósito de demostrar el impacto de estas tecnologías, DUCOR realizó un ensayo en el cultivo de jitomate, empleando estrategias de manejo innovadoras respaldadas por su avanzada línea de bioestimulantes. Este cultivo, esencial para la economía y alimentación de México, demanda soluciones efectivas frente a los retos actuales como la escasez de agua, temperaturas extremas y la necesidad de mantener altos estándares de calidad para mercados nacionales e internacionales.
El ensayo destacó por integrar tecnologías de bioestimulación que potencian el metabolismo de la planta, promoviendo un desarrollo radicular robusto, resistencia a factores de estrés y un aumento significativo en el rendimiento y calidad del fruto. Estos resultados no solo mejoran la rentabilidad del productor, sino que también contribuyen a la sostenibilidad del agro mexicano, reduciendo el impacto ambiental de las prácticas tradicionales.
La implementación de productos DUCOR representa un paso adelante en la adopción de biotecnología en el campo mexicano, sentando un precedente para la agricultura moderna. En este artículo exploramos los detalles del ensayo y los beneficios que estas tecnologías ofrecen para transformar la forma en que cultivamos, impulsando una agricultura más eficiente, innovadora y respetuosa con el medio ambiente.
By
Este estudio se realizó en un invernadero recientemente establecido en el municipio de Tanhuato de Guerrero, Michoacán, con el objetivo de inducir la floración para optimizar la cosecha. El experimento se llevó a cabo en seis surcos bajo macrotúneles, mientras que el área testigo tendrá la misma extensión y número de surcos para una comparación adecuada.
El enfoque es inducir la floración de manera controlada para mejorar la cantidad y calidad de la cosecha. Ambos tratamientos (experimental y testigo) estarán expuestos a las mismas condiciones ambientales y agronómicas, permitiendo evaluar el impacto de la inducción sobre la producción final.
Productos a destacar: TYTANITSUPERFRUTO-ALGAFIX24F-BIOTIC. Todo producto será aplicado de manera foliar.
Productos testigo:

El protocolo consistió en un total de seis aplicaciones de productos específicos, alternándose semanalmente entre dos combinaciones para evaluar sus efectos sobre la inducción de la floración y la mejora de la cosecha. Cada aplicación se realizó siguiendo el siguiente esquema:
1. Primera semana: Se aplicó el primer paquete, compuesto por Biotic y Superfruto, con el objetivo de promover el inicio de la floración.
2. Segunda semana: Se aplicó el segundo paquete, conformado por Tytanit y Algafix 24 F, para reforzar la respuesta de la planta y optimizar el desarrollo de la cosecha.
Este ciclo de aplicaciones se repitió de manera alternada a lo largo del estudio, permitiendo una evaluación precisa de la interacción entre ambos tratamientos y su impacto en la productividad y calidad del cultivo.
La primera evaluación se realizó al finalizar la cuarta semana de aplicación del producto. (21/ octubre/ 2024)
A lo largo de las semanas de aplicación, se observó de manera contundente una uniformidad en el desarrollo y una clara tendencia positiva en el crecimiento de los frutos. La floración presentó un mayor vigor, mientras que los frutos experimentaron un crecimiento significativo, evidenciando una mejora notable en su desarrollo y calidad.
La última evaluación consistió en un análisis de savia a partir del extracto de pecíolo. Los resultados mostraron que, en el tratamiento con Ducor, se obtuvieron niveles más bajos de nitratos y potasio. Sin embargo, es importante destacar que estos valores no indican una deficiencia nutricional en la planta. Por el contrario, estos resultados sugieren que la planta está asimilando los nutrientes de manera más eficiente, optimizando su utilización y transformándolos en aminoácidos esenciales, lo cual favorece su desarrollo y salud general.
Se evaluó el pesaje del fruto, comparando cantidad con peso.
Resaltando que con el tratamiento Ducor, se observó que, a pesar de una menor cantidad de piezas, los frutos presentaron un mayor peso promedio.
Tras realizar la medición de cosecha, se obtuvo un total de 27 cajas por macro túnel tanto en el grupo de testigo como en el de tratamiento. Cada hectárea comprende aproximadamente 20 macro túneles, lo que representa un incremento total de 540 kilogramos gracias al manejo con la bioestimulación Ducor.
La Segunda evaluación se realizó al finalizar la sexta aplicación del producto. (11/noviembre/2024)
Una evaluación más se realizó al finalizar las aplicaciones, en donde el objetivo era observar el cuajado del fruto. (13/noviembre/2024).
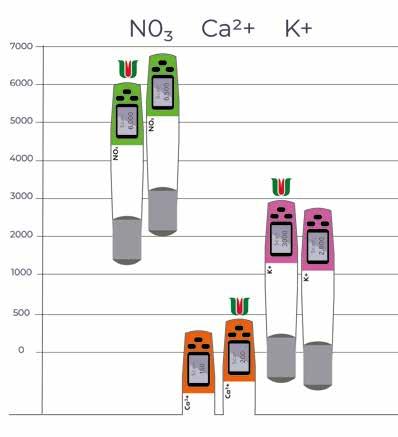




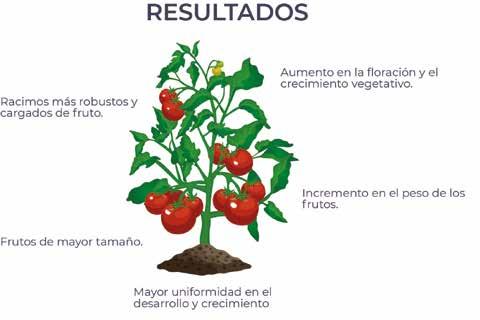
Como resultado del manejo adecuado de la bionutrición y el asesoramiento especializado, se logró un incremento del 4% en la cosecha, lo que se traduce en 3.6 toneladas adicionales al final del ciclo. Este incremento es especialmente significativo, considerando que los predios en cuestión alcanzan un rendimiento promedio de 90 toneladas por ciclo, lo que hace que la práctica sea altamente rentable.
POR JOSÉ BOBADILLA A.
El contenido de nutrientes minerales afecta la capacidad de las plantas para resistir el estrés abiótico y biótico a través de alteraciones en las vías metabólicas, efectos variados sobre la actividad enzimática, la síntesis y permeabilidad de las paredes y membranas celulares y la síntesis de polifenoles, azúcares y aminoácidos. Es probable que el efecto de cualquier elemento nutricional particular de la planta sobre la resistencia a las enfermedades de la planta sea específico del elemento aplicado, la planta huésped y el patógeno. Los minerales son los componentes básicos de las moléculas, los orgánulos ce-
lulares y los órganos de las plantas, y sirven como componente principal de muchos sistemas vegetales. La nutrición equilibrada de las plantas influye en el desarrollo y crecimiento de las plantas cultivadas.
Las plantas requieren al menos 14 elementos esenciales llamados "nutrientes minerales" para mantener la función vital y completar su ciclo de vida, además de carbono en forma de CO2, oxígeno e hidrógeno en forma de agua. Algunas plantas tienen requisitos específicos para elementos adicionales. La adquisición a través de las raíces y el uso de estos elementos son el tema de la nutrición de las plantas. Los nutrientes son absorbidos
Una nutrición equilibrada es fundamental para la salud de las plantas y la capacidad de resistir el estrés biótico
por las raíces a través del transporte activo o pasivo a través de las membranas, y viajan desde el suelo a granel hasta las raíces a través de la difusión o el flujo de masa. Sin embargo, para acceder a todos los nutrientes disponibles, las plantas han desarrollado sistemas de raíces dinámicos y plásticos que exploran el suelo para obtener la máxima absorción de nutrientes. En las monocotiledóneas, las raíces laterales crecen en el volumen del suelo entre las raíces seminales.
Al responder a las señales y gradientes del suelo, el sistema radicular puede maximizar el crecimiento en los parches de nutrientes locales y minimizar el crecimiento en las áreas de

deficiencia. Esto es extremadamente importante para la supervivencia de las plantas, particularmente en suelos deficientes o marginales. El crecimiento eficiente de las raíces también es un factor importante para maximizar el rendimiento con menos aplicaciones de fertilizantes, ya que el crecimiento "desperdiciado" de las raíces cuesta energía que, de otro modo, podría invertirse en el cultivo de interés (ya sean semillas, hojas, tallos o tubérculos). Por esta razón, la comprensión de las respuestas del entorno radicular y el mejoramiento de cultivos con sistemas radiculares eficientes para las condiciones de interés son actualmente áreas muy activas de investigación agronómica.
Aunque las cantidades absolutas de nutrientes necesarios varían según las especies vegetales, los genotipos y los entornos de crecimiento, los nutrientes esenciales se clasifican en los llamados macronutrientes (N, K, Ca, P, Mg, S) que las plantas requieren en mayores cantidades, y micronutrientes (Fe, Cl, Mn, B, Zn, Cu, Mo, Ni) que se necesitan en pequeñas cantidades. Los elementos beneficiosos adicionales incluyen Si (para las gramíneas) y Na (para muchas plantas de la orilla del mar).
PAPEL ESPECÍFICO DE LOS MACRO Y MICRONUTRIENTES
Los macronutrientes forman los componentes estructurales de las proteínas, las paredes celulares, las membranas, los nucleótidos y la clorofila, y desempeñan un papel en el mantenimiento de la energía y el agua. El macronutriente potasio tiene una función especial en la regulación del potencial osmótico de las células vegetales. En condiciones salinas o secas, el Cl (y para algunas plantas el Na) es importante en las relaciones hídricas de las plantas.

Se ha demostrado que el tratamiento con elementos esenciales induce resistencia a la enfermedad en ciertos casos
Los cationes potasio (K+), calcio (Ca2+) y magnesio (Mg2+) son importantes en la vida de las plantas. El potasio (K) es el catión más abundante en los tejidos vegetales, representando hasta el 6% del peso total de la planta seca. El K+ desempeña un papel en las reacciones enzimáticas, afecta el proceso de fotosíntesis y síntesis de proteínas, y controla la acidez del citosol y los cloroplastos. El K+ regula múltiples respuestas al estrés de las plantas, incluidas las especies reactivas de oxígeno (ROS), junto con el ácido jasmónico (JA), el etileno (ET) y las auxinas. Se sabe que el K reduce la incidencia de enfermedades causadas por varios patógenos, entre ellos Botrytis cinerea (moho gris), Sclerotinia sclerotiorum (moho blanco), mildiú velloso Peronospora plantaginis y P. belbahrii, mildiú polvoriento de cereales [Blumeria (syn. Erysiphe) graminis], Podosphaera xanthi (mildiú polvoriento del pepino) y Septoria tritici (mancha septoriana del trigo).
El calcio tiene un papel vital en las funciones celulares, por ejemplo, en la reducción del reblandecimiento y la senescencia de las frutas. El Ca2+ también está implicado en las respuestas de defensa de las plantas, regulando la expresión de genes relacionados con la defensa y la respuesta de hipersensibilidad. El Ca2+ confiere resistencia a patógenos como las especies de Pythium, S. sclerotiorum, B. cinerea, marchitez por Fusarium, mildiú velloso (Sclerospora graminicola) y P. xanthii en el pepino. Además, hay evidencia de que el calcio es antagonista de la antracnosis causada por Colletotrichum gloeosporioides y que el calcio puede afectar la resistencia de la enfermedad a la marchitez bacteriana por Ralstonia.
El magnesio es un elemento mineral esencial para las plantas y puede tener efectos directos e indirectos sobre la resistencia de las plantas a diversas enfermedades. Los niveles de Mg2+ pueden afectar a una amplia gama de funciones fisiológicas, como la transformación de la energía celular, el metabolismo de los carbohidratos, la síntesis de ADN, ARN y proteínas, y la fosforilación de proteínas. El Mg2+ antagoniza con otros minerales, lo que resulta en diversos efectos sobre las
respuestas a las enfermedades. Los patógenos de la marchitez por Fusarium tienden a ser menos infecciosos cuando el Mg2+ está disponible en niveles óptimos. Sin embargo, los altos niveles de Mg2+ interfieren con la absorción de Ca2+ y pueden promover la incidencia de mancha bacteriana (Xanthomonas campestris pv. vesicatoria) en tomate y pimiento, o de podredumbre de vaina de maní. Se ha demostrado que el Mg2+ reduce la gravedad de la enfermedad del mildiú velloso en la amapola, el tabaco, la albahaca dulce y el pepino causada por P. arborescens, P. tabacina, P. belbahrii y P. xanthii, respectivamente. Además de K+, Ca2+ y Mg2+, varios informes también han investigado el uso de cloruro de sodio (NaCl) en las plantas como estrategia de manejo de enfermedades. Se observó una reducción en las enfermedades de la raíz, como el marchitamiento de Rhizoctonia y la pudrición de la corona y la raíz por Fusarium, tras las aplicaciones de NaCl y se informó anteriormente.
Los nutrientes esenciales pueden afectar la gravedad de la enfermedad en varios sistemas de plantas-patógenos. Sin embargo, se informó que los tratamientos con elementos nutritivos específicos disminuyen la gravedad de la enfermedad de un patógeno, pero aumentan la gravedad de otros patógenos. También se informó que diferentes elementos tienen diferentes efectos sobre la resistencia a las enfermedades de las plantas en diversas condiciones ambientales. Aunque se han reportado efectos supresores de la enfermedad de los tratamientos con elementos nutritivos para diferentes fitopatógenos, no se examinaron los mecanismos moleculares que impulsan este fenómeno y los regímenes exactos de tratamiento de elementos nutritivos específicos del huésped necesarios para controlar la enfermedad.
La supresión del mildiú velloso de la albahaca dulce causado por P. belbahrii y el mildiú polvoriento del pepino causado por P. xanthii por

K, Ca o Mg podría estar relacionado con efectos de resistencia inducidos, pero no proporcionaron evidencia molecular o fisiológica para esto. La efectividad de K, Ca, Mg y Na, en forma de sales de cloruro (Cl), como agentes de control de enfermedades en tomate. Estos elementos esenciales mejoran los resultados de la infección con el patógeno fúngico del tomate B. cinerea y el patógeno bacteriano X. euvesicatoria. Los tratamientos con elementos esenciales fueron suficientes para activar las respuestas inmunitarias, induciendo la expresión génica de defensa, la fuga celular y la producción de ROS. Nuestros resultados sugieren que los mecanismos genéticos que están controlados por estos elementos esenciales pueden ser explotados en las prácticas agrícolas para promover la resistencia a las enfermedades de las plantas.
Con la meta de lograr la sostenibilidad ecológica, económica y social actual y futura, se emplean cada vez más técnicas de gestión agrícola ecológicamente racionales, como la diversificación de cultivos, la recuperación de nutrientes de las plantas y el control biológico. No obstante, la actividad agrícola sostenible enfrenta desafíos, algunos de los principales incluye el aumento de la demanda de tierras y recursos agrícolas debido a la rápida expansión de la población, la reducción de la tierra cultivable y la resistencia de plagas y patógenos a los pesticidas y fungicidas

POR LUIS ANTONIO BERNAL
El proceso mediante el cual se posibilita la transferencia de polen desde la parte masculina de una flor hasta la parte femenina de la misma u otra flor, es esencial para el mantenimiento de la viabilidad y la diversidad genética de las plantas con flor, además de mejorar la calidad y cantidad de semillas y frutos, así como
de las características de la descendencia. Puede ser realizada de forma abiótica, mediante el transporte del polen por el viento o el agua, o biótica, empleando para ello a animales como vectores en el transporte. Diversos grupos, tales como invertebrados, aves o mamíferos, pueden actuar como polinizadores, movidos por la necesidad de encontrar recursos para su alimentación, desarrollo o reproducción.
Alrededor del 78% de las especies de plantas con flor en climas templados y del 94% en climas tropicales se benefician del proceso de la polinización mediada por animales, lo que equivale a más del 87% de todas las especies de angiospermas conocidas. En cultivos tropicales, el 70% de las 1,330 especies cultivadas se ve favorecido por estos polinizadores, mientras que en cultivos europeos el 84% de las 264 especies

El polinizador por excelencia
La introducción de la abeja de la miel o abeja melífera en casi todos los países del mundo, y su domesticación durante más de 400 años, podrían considerarse como uno de los mayores experimentos incontrolados inducidos por el ser humano, dada la facilidad de manejo y cría de esta especie, y la asunción acerca de su gran potencial como polinizador y de sus múltiples efectos beneficiosos sobre las cosechas.
Las abejas de la miel son recolectores masivos de polen y néctar que focalizan toda su actividad en pequeños parches de flores, agotando por completo todos los recursos que estas proveen y desplazando a otros polinizadores de sus fuentes de alimento. El proceso de extracción de recursos depende de la densidad de abejas presentes, naturalmente. Curiosamente, los polinizadores también muestran preferencias por las inflorescencias frente a las flores solitarias y por las flores simétricas frente a las asimétricas
cultivadas dependen del proceso de polinización animal.
A nivel global, el 87% de las especies cultivadas, que representan un 35% del suministro global de alimentos, se ven beneficiadas por este proceso, traduciéndose estos datos en un valor económico de la polinización que asciende a más de 153 billones de euros, o al 9.5% del total de la producción agrícola. Por otra parte, se estima que, dentro del 90% de la polinización que ocurre en plantas con flor en todo el mundo, un 67% es llevado a cabo por insectos, constituyéndose como el grupo de polinizadores más importante, tanto para especies de plantas silvestres como cultivadas.
El papel de los insectos polinizadores, y fundamentalmente el de las abejas, ha sido ampliamente demostrado para todo tipo cultivos, siendo cada vez mayor el número de investigaciones que se preocupan por la pérdida de estos organismos y de los servicios que proveen, mientras que son pocos los estudios que tratan de evaluar o explicar el porqué de su importancia y la necesidad de conservar el servicio de la polinización.
El 35% de la producción agrícola mundial depende prácticamente de un agente polinizador. Cuando se habla de agente polinizador pueden ser aves o insectos quirópteros, himenópteros, dípteros, lepidópteros e incluso coleópteros. Las abejas (Insecta: Hymenoptera) son quienes por excelencia se encargan de esta labor. Adicional a esto, la funcionalidad de un ecosistema no solo depende de su diversidad sino de la interacción entre aquellos que lo conforman, en ese sentido la polinización juega un papel primordial en dicha dinámica. Teniendo en cuenta lo anterior, no se puede dejar por fuera la polinización cuando se discute sobre seguridad alimentaria, pues cultivos como el cacao, tomate, aguacate, fríjol, café, maracuyá, mango, manzana, entre
La calidad y cantidad de polen, así como la forma en la que es depositado, puede afectar la producción de frutos y semillas y las características de la descendencia
otros, dependen de este servicio ecosistémico.
La polinización está estrechamente relacionada con la diversidad funcional de polinizadores, basada, a su vez, en las características especie específicas que dirigen el uso complementario de los recursos florales. Es decir, los polinizadores responden a un conjunto de estímulos y recompensas que caracterizan a las flores, y su forrajeo está determinado por rasgos tales como la morfología de la flor y su color, por los que se ven atraídos desde grandes distancias, o por su olor, siendo éste el estímulo predominante en distancias cortas.
EVALUACIÓN DE LA POLINIZACIÓN
DESDE DOS PERSPECTIVAS
Desde el punto de vista inicial de la comunidad de polinizadores y sus interacciones con el cultivo, y desde el punto de vista final de la producción de frutos y semillas, teniendo en cuenta que las primeras medidas se relacionan más directa y cuantitativamente con la polinización, mientras que las segundas son menos sensitivas, debido a la influencia de los numerosos factores abióticos que también intervienen en el proceso polinizador y de producción.
Los polinizadores, tanto silvestres como manejados, se ven muy influidos por los cambios en el entorno que les rodea, debido a su gran dependencia por los recursos florales como sustento y hábitats de nidificación adecuados.

Si el hábitat de nidificación de un polinizador sufre alguna modificación, éste se ve incapacitado para continuar viviendo en ese lugar, aunque en la zona haya recursos alimenticios en abundancia, siendo esta una de las causas más importantes en el declive de las poblaciones de polinizadores nativos.
La alteración, modificación o destrucción del hábitat de un polinizador puede deberse a múltiples causas naturales, tales como la escasez de recursos florales, la presencia estacional de especies vegetales más o menos ricas en polen y néctar, o las condiciones climáticas predominantes. Sin embargo, en la mayoría de las situaciones, el declive a gran escala de sus poblaciones es consecuencia de cambios antropogénicos en el paisaje.
Actividades humanas, tales como la intensificación en los usos del suelo, la fragmentación, degradación y pérdida del hábitat, o el uso de agroquímicos, tienen efectos adversos sobre la diversidad de polinizadores, habiéndose desarrollado algunos estudios que tratan de valorar estos efectos desde el punto de vista de la respuesta de las abejas a los distintos sistemas de manejo del cultivo, a la pérdida de hábitat natural o al incremento del
aislamiento entre parches de hábitat, y sus variaciones según el grado de especialización de una especie a los recursos florales, a lo largo del ciclo de vida de un polinizador o entre taxones.
Los insectos son los polinizadores más conocidos y son considerados como los animales más antiguos y más numerosos que realizan la polinización. Estos son las abejas, avispas y hormigas (himenópteros), las moscas (dípteros), las mariposas y polillas (lepidópteros) y los escarabajos (coleópteros). Las abejas nativas de cada región geográfica son consieradas los polinizadores de mayor importancia. Además, las abejas son bioindicadores de la conservación biológica en los ecosistemas. En estos sistemas
naturales, las abejas polinizan numerosas especies de plantas y también en los cultivos, especialmente si éstos se encuentran cercanos a áreas con vegetación conservada. Por ello, se recomienda conservar fragmentos de ecosistemas lo más grande posible para proteger a las abejas nativas y asegurar la polinización.
De esta forma, las abejas polinizan varios cultivos de importancia económica, como los de calabaza, melón, pepino y sandía (cucurbitáceas), tomate, chile, aguacate, naranja, limón, entre muchos más. Las abejas son nativas de cada zona o región y algunas son especialistas, por ejemplo, las abejas (Peponapis spp) que polinizan las calabazas, y otras





abejas son generalistas y pueden polinizar varios cultivos. Las abejas melíferas (Apis mellifera L.) son abejas exóticas y no son polinizadores eficientes por lo que se recomienda evitar su introducción. Si bien estas abejas contribuyen a la polinización, no son las más importantes ya que son abejas introducidas en varias partes del mundo, como en el continente americano, y algunas veces pueden ser invasoras y desplazar de los recursos florales a las abejas nativas.

La forma de comunicación entre la planta y su polinizador, e incluso sus depredadores, ocurre gracias a los compuestos volátiles expuestos por las flores


problema grave que puede observarse en cucurbitáceas sembradas directamente en el campo y en las bandejas de propagación en invernadero donde las semillas se han sembrado en el suelo infestado. Los síntomas primarios incluyen empapamiento de agua y anillamiento del hipocótilo y el tallo. El marchitamiento se observa más comúnmente en condiciones de suelo saturado.
Las especies de Pythium, hongo patógeno y agente causal de estas pudriciones, tienen una amplia gama de huéspedes y pueden causar pudrición de la raíz y marchitamiento antes y después de la emergencia en una variedad de especies. Muchos cultivos huéspedes comunes incluyen además de las cucurbitáceas, el algodón, soya, maíz, tabaco, otros cultivos de horta-

La prevención es la mejor cura para el marchitamiento y la pudrición de la raíz causada por Pythium o Phytophthora
Marchitamiento o pudrición de raíz y tallo Pythium aphanidermatum
curbitáceas se cultivan en sistemas basados en el suelo y en varios tipos de técnicas de cultivo sin suelo en condiciones de invernadero.
La infección por Pythium también conduce a la pudrición de la raíz tanto en plantas jóvenes como maduras, causando pardeamiento, necrosis y desprendimiento de la corteza de la raíz. El remanente filiforme del tejido vascular en el centro de la raíz es un buen síntoma de diagnóstico de campo para identificar la infección por Pythium. La enfermedad puede progresar hasta el tallo inferior de la planta, donde el tejido se oscurece, se ciñe y se empapa de agua. La destrucción progresiva del sistema radicular y del tejido vascular inferior del tallo da lugar a síntomas aéreos, como marchitamiento de las hojas,
clorosis de las hojas y retraso en el crecimiento. Los síntomas parecen similares en todos los cultivos de cucurbitáceas. Por lo general, las plantas más jóvenes se ven más afectadas negativamente por el Pythium que las plantas maduras. Las pérdidas típicas de los rodales de pepino y sandía son del 10 al 30%. Hasta el 90% de las plántulas o trasplantes de cultivares susceptibles pueden morir.
Los signos de infección por Pythium incluyen la producción de un micelio blanco y algodonoso en la superficie y oosporas incrustadas dentro del tejido necrótico del huésped. Otro importante patógeno de oomicetos transmitido por el suelo que afecta a las cucurbitáceas es Phytophthora capsici, organismo causante de la pudrición de la corona y la raíz por Phytophthora, cuyos síntomas podrían confundirse con los causados por Pythium spp. En las plantas más viejas que han pasado la etapa de plántula,
la pudrición de la corona y la raíz de Phytophthora se puede observar con más frecuencia que la pudrición de la raíz y el tallo de Pythium. Phytophthora capsici debe considerarse como un posible agente causal cuando las partes aéreas de las plantas colapsan rápidamente. Mientras que Pythium spp. puede infectar el tejido principal del tallo inferior y causar anillamiento, Phytophthora capsici puede causar anillamiento en tallos y pecíolos en otras partes de la planta debido al inóculo salpicado.
Se han identificado huéspedes de cucurbitáceas en 10 géneros de Cucurbitaceae. Es probable que todas las especies de cucurbitáceas sean susceptibles, ya que no se ha reportado una resistencia completa. Sin embargo, los cultivares de calabaza de botella y calabaza híbrida interespecífica (Cucurbita máxima & C. moschata) utilizados como portainjertos de cucurbitáceas son mucho menos susceptibles que varios cultivares de sandía y pepino injertados en ellos. En general, los géneros Cucumis y Citrullus parecen ser más susceptibles que Cucurbita y Lagenaria. Se ha de-

mostrado que los aislados de Pythium recuperados de un huésped de cucurbitáceas son patógenos en huéspedes de cucurbitáceas de diferentes géneros con inoculación artificial.
EL ESTRÉS, FACTOR PREDISPONENTE AL DESARROLLO DE LA ENFERMEDAD
El marchitamiento es una enfermedad epidémica grave que afecta a la producción de pepino. Puede causar más del 75% de mortalidad en las plántulas de pepino. Esta enfermedad es causada por varios hongos patógenos, los más importantes de los cuales son Pythium spp. Estudios previos indicaron la asociación de Pythium aphanidermatum, P. spinosum y P. splendens con pepino de invernadero. Entre ellas, P. aphanidermatum es la especie más dominante. Los patógenos fúngicos se introducen en los invernaderos a través de varias fuentes, como el suelo, los medios para macetas y los fertilizantes.
La raíz de las plantas maduras también puede ser atacada. Las enfermedades de marchitamiento inducidas por Pythium spp. ocurren principalmente en suelos fríos y húmedos en el campo, donde las plántulas jóvenes de cultivos de siembra directa pueden morir antes o poco después de emerger. P. aphanidermatum prospera en condiciones
cálidas, mientras que P. ultimum y P. irregulare atacan una variedad de cultivos en condiciones frías.
La infección de las raíces de las plántulas ocurre poco después de la emergencia de la radícula cuando los suelos están húmedos y las condiciones ambientales, como la niebla y el clima húmedo, reducen el crecimiento normal de las plantas. Las plantas maduras muestran síntomas en las raíces alimentadoras, las raíces laterales, las raíces pivotantes y los hipocótilos. Dentro de los 2 días posteriores al riego, Pythium spp. se puede recuperar de las raíces alimentadoras en cualquier momento durante la temporada. Las infecciones laterales y primarias de la raíz se observan generalmente después de los riegos después del cuajado de los frutos. En Israel y California, el estrés predisponente es un factor que contribuye al desarrollo de enfermedades que conducen al colapso de las plantas. Si bien los resultados en Israel implican al virus del mosaico del pepino como el factor de estrés, los virus no parecen estar involucrados en California. Sin embargo, el estrés de la carga de frutos, además del estrés por déficit de agua al final de la temporada, seguido por el riego antes de la cosecha, a menudo resulta en un marchitamiento repentino severo en California.
Este hongo patógeno desafortunadamente cuenta con una amplia gama de huéspedes. Puede causar graves pérdidas económicas cuando las condiciones le son favorables; aunque se dispone de fungicidas para su control, sería muy deseable que se utilizara cualquier tecnología que entrañara poco riesgo para la contaminación del medio ambiente o para la salud humana
Las especies de Pythium pueden ser responsables del marchitamiento tanto antes como después de la emergencia. El marchitamiento previo a la emergencia ocurre cuando la infección antes de la germinación de la semilla. La infección poco después de la germinación puede provocar el debilitamiento del tallo en desarrollo y el eventual colapso de la planta. Las infecciones posteriores pueden provocar la pudrición de la raíz y la corona. Los síntomas pueden incluir retraso en el crecimiento general de la planta, marchitamiento y amarilleo y caída de las hojas. Cuando las plantas
se arrancan de raíz, a menudo se pueden ver lesiones y necrosis en las raíces.
En las pruebas de invernadero con P. ultimum y P. aphanidermatum, se observó el colapso de las plantas solo cuando las plantas que habían dado frutos fueron sometidas a un breve período de estrés hídrico seguido de inundaciones durante un período de 24 o 48 h. Cuando las plantas se mantuvieron infructuosas o no se sometieron a estrés hídrico, no se produjeron síntomas de marchitez repentina. Sin embargo, pruebas similares con P. myriotylum mostraron que este patógeno es capaz de causar una rápida pudrición de la raíz y marchitamiento antes del cuajado de la fruta y sin estrés por déficit de agua. También se ha demostrado que el estrés por salinidad exacerba el desarrollo de enfermedades.
La pudrición de la raíz causada por Phytophthora drechsleri se ha observado en el melón y en la Cucurbita pepo, donde las plantas parecían relativamente normales hasta justo antes de la cosecha, momento en el que colapsaron. Esto sugiere que el estrés de la carga frutal puede estar implicado con este patógeno. Phytophthora capsici causa marchitamiento antes y después de la emergencia en las cucurbitáceas en condiciones de suelo cálido y húmedo. En las plántulas, se desarrolla una podredumbre acuosa en los hipocótilos en o cerca de la línea del suelo. La muerte de la planta después de la emergencia es precedida por el marchitamiento de la planta: un marchitamiento repentino y permanente de la planta sin un cambio en el color del follaje.

Pythium spp. es un problema importante en los cultivos de invernadero en diferentes partes del mundo
Se han utilizado varias técnicas de manejo para manejar las enfermedades inducidas por Pythium spp. en invernaderos. Una técnica común utilizada para reducir el inóculo de Pythium en invernaderos es la solarización y los fungicidas: a saber, mefenoxam, himexazol y propamocarb. Se ha encontrado que la aplicación de los mismos fungicidas en grandes
cantidades durante un largo período de tiempo fomenta la acumulación de resistencia en los aislados de hongos a los fungicidas. Estudios previos evaluaron la resistencia de Pythium spp. al mefenoxam. Sin embargo, no se hizo ningún intento de evaluar la resistencia al segundo fungicida más utilizado, el himexazol.
Impacto acumulativo del mildiú polvoriento en el rendimiento de un año a otro
POR MARCOS CISNEROS CARDENAS
Las pérdidas económicas anuales incurridas por el mildiú polvoriento de la manzana pueden aumentar sustancialmente en las próximas décadas, donde el cambio climático cambia las condiciones ambientales para favorecer el desarrollo de enfermedades.
De hecho, se espera que el cambio climático tenga implicaciones con respecto a muchos patógenos del oídio en todo el mundo. Investigaciones recientes de varias especies de hongos causantes de mildiú polvoriento en cultivos económicamente valiosos como la uva, Erysiphe necator; las cucurbitáceas, Podosphaera xanthii; el roble, Erysiphe alphitoides, y el trigo, Blumeria graminis f. sp. Tritici, han demostrado que el aumento de las temperaturas y los niveles elevados de dióxido de carbono favorecen el desarrollo del oídio ayudando a su propagación más allá de las regiones establecidas. Como se ha sugerido para el mildiú polvoriento de la uva, el cambio climático también
puede alterar la dinámica de sincronía huésped/patógeno entre P. leucotricha y su huésped manzana.
El oídio es una enfermedad fúngica endémica de las regiones productoras de manzanas de todo el mundo, causada por el biótrofo obligado Podosphaera leucotricha del orden Erysiphales. La historia de vida de este hongo está estrechamente sincronizada con la fenología de su huésped perenne, la manzana (Malus × domestica Borkh.) y es el foco de la mayoría de las investigaciones académicas. Además de la manzana, P. leucotricha también puede infectar la almendra, la pera, el membrillo, la cereza africana, el fruto del melocotón y el arbusto ornamental de hoja perenne photinia. A pesar de la prevalencia del mildiú polvoriento en la mayoría de las regiones productoras de manzanas del mundo, faltan investigaciones para cuantificar las pérdidas económicas de rendimiento y los impactos en el vigor y la productividad de los árboles. Las evaluaciones económicas de pérdidas y lesiones varían debido a las
diferencias en la susceptibilidad de los cultivares y los enfoques de manejo de enfermedades. Se sabe que el impacto en el rendimiento es acumulativo de un año a otro, y si no se aborda el mildiú polvoriento en una temporada de crecimiento se produce una mayor pérdida de rendimiento en la temporada siguiente. Si no se trata, se sabe que el mildiú polvoriento reduce la producción fotosintética de un árbol y el contenido de nutrientes de sus hojas, pero aún no se ha definido el impacto cuantitativo de la pérdida total de fotosintato en el rendimiento de los cultivos. Dado que las decisiones de control de enfermedades generalmente sopesan el equilibrio entre el rendimiento de los cultivos y el costo de control, un umbral de manejo debe guiar las prácticas de manejo.
Estudiar el desarrollo y la propagación de un patógeno puede arrojar luz sobre las mejores prácticas para su manejo. El desarrollo de P. leucotricha está estrechamente relacionado con la fenología del huésped y las condiciones ambientales. Se han desarrollado

El desarrollo y la esporulación de las colonias de P. leucotricha se rigen por factores ambientales similares de precipitación y temperatura

varios modelos para establecer relaciones funcionales con el progreso de la enfermedad a lo largo del tiempo. El ciclo de la enfermedad del oídio puede interrumpirse en varias etapas específicas con prácticas de manejo: fungicidas, selección de huéspedes resistentes y poda mecánica.
FACTORES MEDIOAMBIENTALES QUE
PUEDEN AGUDIZAR EL PROBLEMA
TEMPERATURA
Un factor clave que influye en la liberación y germinación de conidios en P. leucotricha es la temperatura, siendo las óptimas para la liberación de conidios entre 15 y 28° C, con una germinación óptima a 22° C. Por debajo de 4 a 10° C y por encima de 30° C, los conidios no pueden germinar de manera efectiva. La dispersión de conidios es diurna, con una gran liberación a primera hora de la tarde y un
pico más pequeño justo antes o justo después del anochecer. La concentración de conidios en el aire también se ha correlacionado positivamente con la velocidad del viento, la temperatura y la radiación solar y negativamente con la humedad relativa y la humedad de las hojas.
HUMEDAD
La humedad ambiental no afecta en gran medida la germinación de los conidios de P. leucotricha, aunque el agua libre es inhibidora. Se ha demostrado que P. leucotricha germina con éxito en hojas de manzano a una humedad relativa que oscila entre el 40 y el 100%. La dispersión, sin embargo, se ve afectada negativamente por la alta humedad relativa, mayor a >80%. Las lluvias intensas, superiores a los 3 mm o los eventos de precipitación prolongados, hasta 3 h, causaron una
marcada disminución en la dispersión de conidios.
La exposición a la luz en las hojas de manzano aumentó la actividad fotosintética del huésped al tiempo que redujo el desarrollo de moho. Es probable que el sombreado no sea un beneficio directo para el desarrollo de P. leucotricha. En cambio, puede conducir a una reducción del tiempo de secado para la humedad de las hojas o un aumento de la humedad. El sombreado incurre en un potencial de reducción fotosintética en el huésped (y el cambio subsiguiente en la relación fuente-suministro de los fotosintatos) que conduce al estrés del huésped y puede beneficiar al patógeno indirectamente al alterar la susceptibilidad del tejido del huésped. Sin embargo, se necesita más investigación para determinar si el control de la sombra tiene
algún papel como un medio confiable de manejo de la enfermedad.
La evaluación de la intensidad de la enfermedad es una consideración importante para comprender los impactos en el rendimiento y tomar decisiones de manejo. La evaluación de la enfermedad puede ser un proceso laborioso, pero es esencial para el manejo racional de la enfermedad. Se necesitan evaluaciones rápidas, sencillas, precisas y exactas para tomar decisiones de gestión. La intensidad de la enfermedad puede medirse tanto como la incidencia (número de unidades vegetales visiblemente enfermas) como la gravedad (área/ volumen de tejido vegetal que está enfermo en relación con el área/volumen total). La gravedad de la enfermedad también puede denominarse densidad de la enfermedad cuando es la medida del número de colonias (o lesiones) por unidad de planta.
La incidencia es más fácil y rápida de evaluar que la gravedad, pero la gravedad puede ser más útil para las decisiones de tratamiento.
DEL MILDIÚ POLVORIENTO
Disminuir o erradicar las fuentes primarias de inóculo y, a su vez, la propagación de la enfermedad, son los objetivos principales. Debido a que P. leucotricha es un patógeno obligado cuyo micelio pasa el invierno dentro de los tejidos del huésped, la poda ha demostrado ser una práctica de saneamiento efectiva para reducir el


La disponibilidad de luz puede tener un marcado impacto en el desarrollo del oídio del manzano
inóculo, aunque el costo de la mano de obra para operaciones más grandes puede hacer que el esfuerzo sea poco práctico. Los beneficios de la poda mejoran en gran medida cuando se usa en combinación con un programa de fungicidas.
La eliminación de los brotes aparentemente infectados durante la temporada de latencia redujo la incidencia de la enfermedad primaria del oídio en la primavera siguiente; Sin embargo, esto es difícil porque no hay hojas que señalen fácilmente la presencia de la enfermedad y, en cambio, se basa en un efecto visual “rayado” dejado en el tejido del brote por el micelio restante de la temporada de crecimiento anterior. La poda regular también tiene el beneficio adicional de abrir el dosel del árbol, lo que conduce a una mejor deposición de fungicidas y reduce la humedad relativa que se encuentra dentro del microclima de la rama del brote, lo que impide aún más el desarrollo de patógenos foliares.
El azufre elemental se ha utilizado ampliamente para proteger contra el mildiú polvoriento en la producción de manzanas y continúa sirviendo como un fungicida central de amplio espectro en los programas de manejo de la enfermedad del mildiú polvoriento convencionales y orgánicos en todo el mundo. El azufre es una opción fun-
gicida económica que es eficaz para controlar el mildiú polvoriento cuando se usa de manera profiláctica y frecuente, en intervalos de aproximadamente 5 a 7 días. Es posible que el azufre no controle eficazmente la enfermedad en condiciones de alta presión de la enfermedad y tiene problemas de fitotoxicidad cuando se aplica a temperaturas superiores a 28° C.
En áreas donde ambas enfermedades son prevalentes, el manejo químico del mildiú polvoriento de la manzana generalmente se maneja simultáneamente con la sarna del manzano, Venturia inaequalis, ya que los dos patógenos fúngicos a menudo cohabitan en los huertos. En la producción convencional de manzanas, los inhibidores de la desmetilación, DMI; los inhibidores externos de la quinona, QoI; y los inhibidores de la succinato deshidrogenasa, SDHI, comprenden los tres grupos de fungicidas comúnmente utilizados para controlar el mildiú polvoriento. Los DMI, grupo FRAC 3, son uno de los fungicidas más efectivos y ampliamente utilizados para el manejo. Con el aumento de la resistencia a los fungicidas DMI en las poblaciones de V. inaequalis, se buscaron productos químicos alternativos a los fungicidas para controlar con éxito tanto la sarna del manzano como el mildiú polvoriento. QoIs --grupo FRAC 11-- ofreció un modo de acción reportado como efectivo contra el mildiú polvoriento en ensayos de campo, a menudo cuando se usa en premezclas de fungicidas. En comparación, los SDHI --grupo FRAC 7-- se han registrado para la “supresión” del mildiú polvoriento de la manzana y son los menos efectivos de los tres productos químicos. Las anilinopirimidinas, grupo FRAC 9, también se utilizan para el tratamiento del mildiú polvoriento del manzano, pero parecen haber dejado de ser de uso comercial generalizado. Múltiples estudios han explorado el desarrollo de resistencia a fungicidas en poblaciones de P. leucotricha.



Puebla se galardonó con la presencia de más de 200 asistentes al Congreso de Brócoli celebrado los días 5, 6 y 7 de febrero, en donde productores, técnicos, asesores y comercializadores del cultivo compartieron impresiones y conocimientos sobre la mejora de la producción. Ponentes nacionales e internacionales nos brindaron la oportunidad de conocerlos y escuchar sus experiencias e investigaciones para hacer frente a los nuevos retos que tiene la producción, nutrición y manejo post cosecha del cultivo.
A medida que el mercado crece, así como la conquista de nuevos nichos de mercado, el productor se ve
obligado a establecer medidas más específicas para una producción sustentable. Asimismo, con los cambios en la economía, el ambiente y el gusto del consumidor. La asistencia al congreso del brócoli se convierte en un clara opción para aprovechar la ocurrencia de tecnología de última, información y mesas de negocios bajo un mismo techo, algo que hoy en día puede resultar complejo si no se establecen alianzas para obtener mejores oportunidades. El valor de un producto se incrementa a medida que se conoce más sobre él, esto lo han comprendido los productores y demás cadenas involucradas con el brócoli, siendo pieza clave la actualización de sus conocimientos y
la ampliación de sus expectativas de producción y mercado.
Contar con la asistencia de integrantes de toda la cadena de producción y comercialización del cultivo de brócoli, así como de nuestros panelistas expertos, nos brinda gran satisfacción; en primer lugar, por el hecho de haber cumplido con las expectativas del establecimiento de interrelaciones, y en segundo lugar porque sabemos que el campo se fortalece con conocimiento. El congreso del brócoli destacó que cada día son más los interesados en conocer más a fondo cada uno de los eslabones de un sistema de producción y de conservación; cuidar lo que se produce en campo es algo funda-
mental, un descuido puede repercutir en pérdida total de las ganancias, la falta de sanidad o un mal enfriamiento en el cultivo de brócoli ocasionan que el producto pierda valor.
Durante el primer día del evento Nobuhiro Nagata destacado genetista de SAKATA, abrió el panel de expertos recalcando la importancia que tiene la ingeniería genética para obtener variedades que se adaptan a las nuevas condiciones y retos que el mercado y el consumidor final exigen. No se trata de investigar por investigar, un sistema de mejoramiento requiere de tiempo y las demandas cambian constantemente, por ello, es que cada variedad en el mercado debe estar dos o tres pasos delante de las necesidades del productor. Gracias a la ingeniería genética hoy en día se pueden obtener materiales con mejores características en menor tiempo. Por su parte el Dr. Concepción Rodríguez del Colegio de Posgraduados, hizo énfasis en que el producto final no solo debe tener buenas características organolépticas y visuales, se requiere de buena sanidad sin dejar de lado la responsabilidad social. El cliente requiere de productos sanos e inocuos, pero también, de productos que le hagan sentir emociones desde su selección hasta su consumo. Uno de los principales problemas presentes en el cultivo de brócoli continúa siendo la palomilla dorso diamante y su control requiere de estrategias mejor planteadas y diversificadas, que no solo involucren el uso de agroquímicos, sino que sean más integrales y organizadas. El caso de la palomilla dorso diamante, no solo afecta a los cultivos en campo, su mayor problema se presenta en el inicio del proceso de post cosecha, la alta incidencia de larvas en la cabezuela limita su comercialización y no poder realizar manejos inmediatos a la cosecha por el riesgo de toxicidad se convierten en casos sin solución. De ahí la importancia de crear planes de manejo regional, no solo individuales y locales, se requiere de una participa-









ción colectiva para bajar la incidencia de dicha plaga. Así mismo, el Dr. Concepción invita a todos los productores a implementar políticas de responsabilidad social sobre el desperdicio de alimentos.
En esta ocasión, Matt Linder especialista de semillas Sakata nos presentó el reto que se tiene con el uso de semillas de calidad, destacando que no se define por el precio, sino por el mercado final al que se desea integrar. Semillas de calidad brindará mayor seguridad al productor, la ingeniería genética se expresará en mayor o menor medida en función con las condiciones climáticas y cuidados dados, sin embargo, Sakata posee un plus en sus semillas, la genética de ellas está adaptada a condiciones adversas, lo que les permite subsistir aun en cambios repentinos de manejo y clima sin perder calidad.
Por su parte, el maestro Alejandro Rivera mostró los resultados obtenidos en el estado de Puebla sobre un plan de manejo sustentable, el uso de biofertilizantes favoreció un desarrollo radicular mayor en comparación con el tradicional, complementó con el uso de bioprotectores en cada una de las etapas del cultivo y del suelo, demostrando que con un plan integral se puede obtener una calidad de fruto excepcional y un menor desgaste al
suelo, enriqueciendo su microfauna. Al cierre del día Adriana García y Peyton Merriam hicieron un recuento de lo importante de las labores precosecha para tener una buena calidad post cosecha y con ello aumentar la rentabilidad para el ingreso en el mercado de exportación. Los canales de distribución no siempre son los mejores ni los más cortos para la comercialización de productos frescos, por ello, es importante tener los cuidados necesarios en la cosecha y su traslado al proceso, mucha de la calidad obtenida en campo puede llegar a perderse por un mal manejo de producto durante los traslados, en producto congelado se tienen menos riesgos de perdida, sin embargo, aquí si tiene mucho que ver el sistema de enfriamiento elegido y la variedad empleada.
Durante el segundo día, el congreso estuvo enfocado a presentar estrategias que favorezcan una producción más amigable, que permita al productor tener finanzas sanas. El Dr. Fernando Ramos hizo mucho hincapié en que el monitoreo es la herramienta más importante que posee el productor para eficientizar su sistema de manejo, tener varias vistas y una amplia perspectiva del cultivo favorecerá tomar decisiones más acertadas, hoy existen drones que nos permiten tomar tomas aéreas que facilitan la detección de







problemas que a nivel de parcela pueden parecer invisibles. La visibilidad de que brinda un dron en mucho más enfocada en el comparativo total de la parcela, detectando fugas de agua, errores de aplicación de productos fitosanitarios, problemas de germinación, entre otros, que permitirán una mejor programación de las acciones a implementar que a su vez disminuirán costos en la producción.
ACCIONES PREVENTIVAS EN EQUILIBRIO CON EL CUIDADO DEL AMBIENTE
PARA FAVORECER LOS CICLOS DE PRODUCCIÓN
El MC. Mauricio Navarro subrayó la importancia de crear planes de manejo nutricional adecuados a la medida de cada productor, basándose en la salud del suelo y el avance del cultivo ya que no es suficiente conocer las necesidades de un cultivo sino que es importante conocer cómo está trabajando la microbiología del suelo y estar al tanto de la disponibilidad de los nutrientes aplicados y de la respuesta de la planta. Los factores ambientales juegan un papel importante en el desarrollo, un mal manejo de los diferentes estrés que sufre una planta, encarecerán un ciclo de producción, lo que se verá reflejado en una disminución de las ganancias del productor si se desea conservar la calidad, aunado a esto, el cuidar de los suelo y realizar planes más armonizados con el planeta favorecerán la conservación del recurso más vital para la agricultura: el agua.
En complemento a un manejo equilibrado, el Ing. Tránsito Sánchez comentó a los asistentes cuán importante es la realización de la correcta identificación de las plagas en el cultivo del brócoli, no todos los insectos que llegan causan el mismo daño y requieren de manejos distintos; hoy ya no funcionan las técnicas de receta, el suelo y el ambiente han sufrido grandes cambios, la falta de alimento para la fauna benéfica hacen que las condiciones de control sean más
complejas. Existen plagas de gran importancia por su rápido crecimiento o por un daño voraz, sin embargo, no se les brinda la importancia necesaria por su desconocimiento.
En cuanto a la aplicación del correcto manejo sanitario la Dra. Ma. Fernanda Ramírez, explicó a los productores que la correcta identificación de una plaga y enfermedad ayuda a crear planes más integrados, pero si la aplicación y sobre todo la rotación de productos no se realiza de forma correcta, el problema puede agravarse. No basta con cambiar el ingrediente activo en cada aplicación, es importante verificar el grupo al que pertenecen, esto forma una herramienta más asertiva en el manejo de plagas. Conservar la calidad es un reto importante para el brócoli, si las condiciones de traslado, tratamiento post cosecha y almacenaje no son correctos, aquella calidad en campo se ve mermada, tal es así que una planta bien nutrida puede hacerles frente a las adversidades, la Dra. Nayelli Solano destacó la importancia de tratar a las plantas como plantas, es decir, permitir que expresen sus características genéticas dadas, si bien los planes de nutrición y sanidad ayudan a tener cosechas de calidad, es importante que la planta exprese todo su potencial, de esa manera se pueden obtener brócolis con mejores características, “una planta floja, brinda frutos flojos”.
EL RECORRIDO POR LAS PARCELAS DEMOSTRATIVAS
Además de la experiencia que los ponentes compartieron con cada uno de los asistentes, en el tercer día de evento, fueron partícipes de un recorrido de campo, en dónde ellos pudieron apreciar in situ las características de los materiales ofertados por SAKATA. Revista DeRiego no solo lleva información especializada, busca llevar la experiencia teórica a una experiencia práctica, en esta ocasión, acompañados de Sakata establecieron un campo demostrativo en donde

lograron apreciar las características genéticas expresadas en un ambiente real, de los materiales descritos por la empresa y del manejo nutricional y sanitario en el manejo de cada una de ellas.
Ubicados en las instalaciones de la agroexpo los Reyes, en los Reyes, Puebla, Semillas Genesis y SAKATA dieron cita los asistentes del congreso, quienes acompañados por el personal técnico de Sakata escucharon y vieron las características de cada uno de los materiales sembrados, se presentaron variedades precoces, intermedias y tardías, todas con un gran catálogo de resistencias a diversas patologías importantes. Destacando entre ellas a Purple Magic, variedad de color morado de reciente lanzamiento en México, es una variedad con un alto valor nutricional y un alto contenido de taninos que la hace exclusiva para el consumo en fresco, actualmente se encuentra en busca de productores interesados en incluirla dentro del catálogo de orgánicos, sus características físicas la hacen muy apetecible al paladar.
Armando fue el encargado de la planeación y organización del día de campo, quien expresó que los trabajos comenzaron con meses de anticipación, se establecieron las variedades que los productores locales






ya conocen, pero incluimos variedades que pueden ser otra opción para los productores en estaciones del año que les es difícil competir con las ya conocidas, queremos diversificar las variedades para aumentar las oportunidades de mercado para los productores el centro de México. Las nuevas generaciones de brócoli ya están listas para enfrentar los retos del cambio climático, las demandas del mercado y el gusto del productor, Sakata siempre vanguardista e innovando para el futuro.
Agradecemos a todas las empresas participantes SAKATA, Harvest Hold Fresh, Terra Seeds, Compo Expert, Simbiosis agrícola, El Tomate de Oro, Greenhow, Agroscience, Keithly-Williams Seeds, Cohisa, Greenforce, FMC, AFL-Agro, CAJINSA, GVC, DUCOR, Semillas Alfaomega, Confiagro, Fotosíntesis; quienes nos acompañaron en este congreso de brócoli, nos vemos en su próxima edición.





BRISEIDA GARCÍA
Contabilidad & Servicio al Cliente (686) 221-7714
briseida.garcia@westarseeds.com
FERNANDA VALDEZ
Servicio al Cliente (686) 225-7640
fernanda.valdez@westarseeds.com
ELIEL AGUIRRE
Responsable México (462) 199-1730 eliel@westarseeds.com
JOSUÉ CRUZ
Baja California (686) 119-3508
josue.cruz@westarseeds.com
JOSÉ CABRERA
Noreste (686) 570-8839
jose.cabrera@westarseeds.com
MARIO BELTRÁN
Sinaloa (686) 354-1595
mario.beltran@westarseeds.com
MANUEL LUQUE
Sonora (686) 119-7544
manuel.luque@westarseeds.com
OMAR PEREDO
Altiplano (686) 123-5432
omar.peredo@westarseeds.com
Grey Zucchini
