PESTS & DISEASES
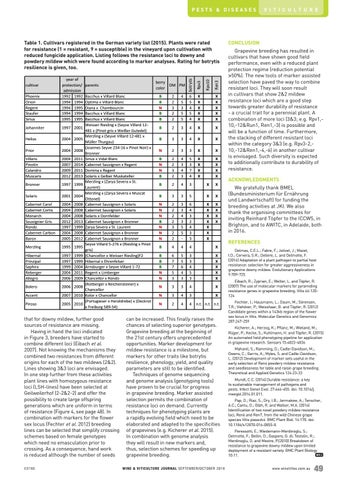
Helios
2004 2005
Prior
2004 2008
Villaris Pinotin Calandro Muscaris
2004 2007 2009 2012
Bronner
1997 1999
Solaris
2001 2004
Cabernet Carol Cabernet Cortis Monarch Souvignier Gris Rondo Cabernet Carbon Baron
2004 2004 2004 2012 1997 2004 2005
Merzling
1995 1995
Hibernal Prinzipal Saphira Reberger Allegro
1997 1997 1999 2004 2006
Bolero
2006 2008
Accent
2007 2010
Piroso
2005 2010
2011 2014 2011 2013
2008 2008 2008 2013 1999 2008 2012
1999 1999 2004 2011 2009
Bacchus x Villard Blanc Optima x Villard Blanc Diana x Chambourcin Bacchus x Villard Blanc Bacchus x Villard Blanc Weisser Riesling x (Seyve Villard 12481 x (Pinot gris x Weißer Gutedel) Merzling x (Seyve Villard 12-481 x Müller Thurgau) (Joannes Seyve 234-16 x Pinot Noir) x Bronner Sirius x Vidal Blanc Cabernet Sauvignon x Regent Domina x Regent Solaris x Gelber Muskateller Merzling x (Zarya Severa x St. Laurent) Merzling x (Zarya Severa x Muscat Ottonel) Cabernet Sauvignon x Solaris Cabernet Sauvignon x Solaris Solaris x Dornfelder Cabernet Sauvignon x Bronner Zarya Severa x St. Laurent Cabernet Sauvignon x Bronner Cabernet Sauvignon x Bronner Seyve Villard 5-276 x (Riesling x Pinot gris) (Chancellor x Weisser Riesling)F2 Hibernal x Ehrenfelser Arnsburger x Seyve Villard 1-72 Regent x Limberger Chancellor x Rondo (Rotberger x Reichensteiner) x Chancellor Kolor x Chancellor (Portugieser x Heroldrebe) x (Deckrot x Freiburg 589-54)
that for downy mildew, further good sources of resistance are missing. Having in hand the loci indicated in Figure 3, breeders have started to combine different loci (Eibach et al. 2007). Not knowing the mechanisms they combined two resistances from different origins for each of the two mildews (2&2). Lines showing 3&3 loci are envisaged. In one step further from these activities plant lines with homozygous resistance loci (LSH-lines) have been selected at Geilweilerhof (2-2&2-2) and offer the possibility to create large offspring generations which are uniform in terms of resistance (Figure 4, see page 48). In combination with markers for the flower sex locus (Fechter et al. 2012) breeding lines can be selected that simplify crossing schemes based on female genotypes which need no emasculation prior to crossing. As a consequence, hand work is reduced although the number of seeds V3 1N 5
2 2 3 2 2
4 5 3 5 5
6 5 4 5 4
X X X X X
X X X X X
B
2
3
4
X
X
B
3
3
4
X
X
N
2
3
3
X
X
B N N B
2 2 3 2
4 3 4 3
5 3 7 4
X X X X
X X X X
B
2
4
3
X
X
B
3
3
5
X
X
N N N B N N N
2 2 2 2 3 2 2
3 3 4 3 5 5 -
6 4 3 2 4 3 3
X X X X X X X
X X X X
B
4
4
4
X
B B B N N
6 7 7 5 3
5 5 6 4 3
3 3 4 5 3
X X X X X
N
3
3
4
X
N
3
4
3
N
2
4
4
Ren3
1997 2001
B B N B B
berry DM PM color
parents
Rpv10
Johanniter
Rpv3
Phoenix Orion Regent Staufer Sirius
year of protection/ admission 1992 1992 1994 1994 1994 1995 1994 1994 1995 1995
cultivar
botrytis
Table 1. Cultivars registered in the German variety list (2015). Plants were rated for resistance (1 = resistant, 9 = susceptible) in the vineyard upon cultivation with reduced fungicide application. Listing follows the resistance loci to downy and powdery mildew which were found according to marker analyses. Rating for botrytis resilience is given, too.
X n.t. n.t. n.t.
can be increased. This finally raises the chances of selecting superior genotypes. Grapevine breeding at the beginning of the 21st century offers unprecedented opportunities. Marker development for mildew resistance is a milestone, but markers for other traits like botrytis resilience, phenology, yield, and quality parameters are still to be identified. Techniques of genome sequencing and genome analysis (genotyping tools) have proven to be crucial for progress in grapevine breeding. Marker assisted selection permits the combination of resistance loci on demand. Currently techniques for phenotyping plants are a rapidly evolving field which need to be elaborated and adapted to the specificities of grapevines (e.g. Kicherer et al. 2015). In combination with genome analysis they will result in new markers and, thus, selection schemes for speeding up grapevine breeding. W I N E & V I T I C ULTUR E JO UR NA L SEPTEMBER/O C TO BER 2016
V V II T T II C C U U L LT T U U R R E E
CONCLUSION Grapevine breeding has resulted in cultivars that have shown good field performance, even with a reduced plant protection regime (reduction potential >50%). The new tools of marker assisted selection have paved the way to combine resistant loci. They will soon result in cultivars that show 2&2 mildew resistance loci which are a good step towards greater durability of resistance - a crucial trait for a perennial plant. A combination of more loci (3&3; e.g. Rpv1,10,-12&Run1, Ren1,-3) is possible and will be a function of time. Furthermore, the stacking of different resistant loci within the category 3&3 (e.g. Rpv3-2,10,-12&Ren1,-4,-6) in another cultivar is envisaged. Such diversity is expected to additionally contribute to durability of resistance. ACKNOWLEDGMENTS We gratefully thank BMEL (Bundesministerium für Ernährung und Landwirtschaft) for funding the breeding activities at JKI. We also thank the organising committees for inviting Reinhard Töpfer to the ICCWS, in Brighton, and to AWITC, in Adelaide, both in 2016. REFERENCES Delmas, C.E.L.; Fabre, F.; Jolivet, J.; Mazet, I.D.; Cervera, S.R.; Deliere, L. and Delmotte, F. (2016) Adaptation of a plant pathogen to partial host resistance: selection for greater aggressiveness in grapevine downy mildew. Evolutionary Applications 9:709-725 Eibach, R.; Zyprian, E.; Welter, L. and Töpfer, R. (2007) The use of molecular markers for pyramiding resistance genes in grapevine breeding. Vitis 46:120124 Fechter, I.; Hausmann, L.; Daum, M.; Sörensen, T.R.; Viehöver, P.; Weisshaar, B. and Töpfer, R. (2012) Candidate genes within a 143kb region of the flower sex locus in Vitis. Molecular Genetics and Genomics 287:247-259 Kicherer, A.; Herzog, K.; Pflanz, M.; Wieland, M.; Rüger, P.; Kecke, S.; Kuhlmann, H. and Töpfer, R. (2015) An automated field phenotyping pipeline for application in grapevine research. Sensors 15:4823-4836 Mahanil, S.; Ramming, D.; Cadle-Davidson, M.; Owens, C.; Garris, A.; Myles, S. and Cadle-Davidson, L. (2012) Development of marker sets useful in the early selection of Ren4 powdery mildew resistance and seedlessness for table and raisin grape breeding. Theoretical and Applied Genetics 124:23-33 Mundt, C.C. (2014) Durable resistance: a key to sustainable management of pathogens and pests. Infect Genet Evol. 27:446-455. doi: 10.1016/j. meegid.2014.01.011. Pap, D.; Riaz, S.; Dry, I.B.; Jermakow, A.; Tenscher, A.C.; Cantu, D.; Oláh, R. and Walker, M.A. (2016) Identification of two novel powdery mildew resistance loci, Ren6 and Ren7, from the wild Chinese grape species Vitis piasezkii. BMC Plant Biol. 16:170. doi: 10.1186/s12870-016-0855-8. Peressotti, E.; Wiedemann-Merdinoglu, S.; Delmotte, F.; Bellin, D.; Gaspero, G. di; Testolin, R.; Merdinoglu, D. and Mestre, P.(2010) Breakdown of resistance to grapevine downy mildew upon limited deployment of a resistant variety. BMC Plant Biology WVJ 10:11. www.winetitles. com . au
49